Настоящее изобретения относится к комбинации онколитического вируса и ингибитора CDK4; к применению такой комбинации в лечении такого заболевания, как опухоль; к онколитическому вирусу, предпочтительно онколитическому аденовирусу для применения в лечении такого заболевания, как опухоль, совместно с ингибитором CDK4/6; и к ингибитору CDK4/6 для применения в лечении такого заболевания, как опухоль, совместно с онколитическим вирусом, предпочтительно онколитическим аденовирусом.
В настоящее время в лечении опухолей используется ряд терапевтических концепций. Помимо хирургического вмешательства, преобладают химиотерапия и лучевая терапия. Однако все эти методы связаны со значительными побочными эффектами. Использование онколитических вирусов, селективных к репликации, предоставляет новую идейную основу для лечения опухолей. В связи с этим инициируется селективная репликация вирусного агента внутри опухоли, что приводит к репликации вируса, лизису инфекционной опухолевой клетки и распространению вируса на смежные опухолевые клетки. Поскольку способность вируса к репликации вируса ограничена опухолевыми клетками, то нормальная ткань защищена от репликации и, таким образом, от лизиса вирусом.
Задача, лежащая в основе настоящего изобретения, заключается в получении средств повышения эффективности терапии опухолей на основе онколитических вирусов, и в частности, аденовируса.
Эти и другие задачи решаются с помощью объекта изобретения, описанного в прилагаемых независимых пунктах формулы изобретения; предпочтительные варианты осуществления могут быть взяты из прилагаемых зависимых пунктов формулы изобретения.
Задача, лежащая в основе настоящего изобретения, также решается в первом аспекте, который также является первым вариантом осуществления такого первого аспекта, посредством комбинации, содержащей аденовирус, ингибитор CDK4/6 и по меньшей мере один дополнительный агент, выбранный из группы, включающей ингибитор PARP, ингибитор бромодомена и нутлин или производное нутлина.
Далее раскрываются дополнительные варианты осуществления такого первого аспекта.
Вариант осуществления 2: Комбинация по варианту осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Комбинация по любому из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Комбинация по варианту осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Комбинация по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Комбинация по варианту осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Комбинация по варианту осуществления 6, где белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Комбинация по варианту осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Комбинация по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Комбинация по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Комбинация по варианту осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Комбинация по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Комбинация по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Комбинация по варианту осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Комбинация по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Комбинация по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Комбинация по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Комбинация по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Комбинация по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Комбинация по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Комбинация по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающий XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы, не содержащие экспрессированный вирусный онкоген, который способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Комбинация по варианту осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Комбинация по варианту осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Комбинация по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Комбинация по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Комбинация по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Комбинация по варианту осуществления 26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Комбинация по варианту осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Комбинация по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Комбинация по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Комбинация по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Комбинация по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Комбинация по любому из вариантов осуществления 1-32, где композиция дополнительно содержит ингибитор PARP.
Вариант осуществления 34: Комбинация по варианту осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Комбинация по любому из вариантов осуществления 1-32, где композиция дополнительно содержит ингибитор бромодомена.
Вариант осуществления 36: Комбинация по варианту осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Комбинация по любому из вариантов осуществления 1-36, где композиция дополнительно содержит нутлин или его производное.
Вариант осуществления 38: Композиция по варианту осуществления 37, где нутлин представляет собой нутлин-3.
Вариант осуществления 39: Композиция по любому из вариантов осуществления 37 и 38, где производное нутлина выбрано из группы, включающей NVP-HDM201, идазанутлин, AM-8553, SAR405838, нутлин-3a, AMG232.
Вариант осуществления 40: Комбинация по любому из вариантов осуществления 1-39, где компоненты комбинации предназначены для раздельного введения.
Вариант осуществления 41: Комбинация по любому из вариантов осуществления 1-39, где компоненты комбинации предназначены для комбинированного введения.
Задача, лежащая в основе настоящего изобретения, также решается во втором аспекте, который также является первым вариантом осуществления такого второго аспекта, посредством комбинации в соответствии с первым аспектом, включая любые ее варианты осуществления, для применения в лечении заболеваний, более предпочтительно, опухоли или злокачественного новообразования, содержащей аденовирус и ингибитор CDK4/6.
1-я группа Далее раскрываются дополнительные варианты осуществления такого второго аспекта.
Вариант осуществления 1: Комбинация, содержащая аденовирус и ингибитор CDK4/6 для применения в способе лечения и/или предупреждения заболевания, предпочтительно, опухоли или злокачественного новообразования.
Вариант осуществления 2: Комбинация для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Комбинация для применения в соответствии с одним из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Комбинация для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Комбинация для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Комбинация для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Комбинация для применения в соответствии с вариантом осуществления 6, где белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Комбинация для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Комбинация для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Комбинация для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Комбинация для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Комбинация для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Комбинация для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Комбинация для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Комбинация для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Комбинация для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Комбинация для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Комбинация для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Комбинация для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Комбинация для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Комбинация для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающий XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Комбинация для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Комбинация для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Комбинация для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Комбинация для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Комбинация для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Комбинация для применения в соответствии с вариантом осуществления 26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Комбинация для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Комбинация для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Комбинация для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Комбинация для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Комбинация для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Комбинация для применения по любому из вариантов осуществления 1-32, где композиция дополнительно содержит ингибитор PARP.
Вариант осуществления 34: Комбинация для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Комбинация для применения по любому из вариантов осуществления 1-32, где композиция дополнительно содержит ингибитор бромодомена.
Вариант осуществления 36: Комбинация для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Комбинация для применения по любому из вариантов осуществления 1-36, где компоненты комбинации предназначены для отдельного введения.
Вариант осуществления 38: Комбинация для применения по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Комбинация для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Комбинация для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Комбинация для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Комбинация для применения в соответствии с вариантом осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Комбинация для применения в соответствии с вариантом осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44. Комбинация для применения в соответствии с вариантом осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45. Комбинация для применения по любому из вариантов осуществления 1-44, где клетки опухолевых клеток обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии или нечувствительны к ним.
Вариант осуществления 46: Комбинация для применения в соответствии с вариантом осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Комбинация для применения по п.46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Комбинация для применения по п.47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Комбинация для применения по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Комбинация для применения по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Комбинация для применения по любому из вариантов осуществления 1-50, где клетки опухоли содержат YB-1 в ядре.
Вариант осуществления 52: Комбинация для применения по любому из вариантов осуществления 1-51, где клетки опухоли содержат YB-1 в ядре после индукции.
Вариант осуществления 53: Комбинация для применения в соответствии с вариантом осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Комбинация для применения в соответствии с вариантом осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Комбинация для применения по любому из пп.1-54, где опухоль выбрана из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
Задача, лежащая в основе настоящего изобретения, также решается в третьем аспекте, который также является первым вариантом осуществления такого третьего аспекта, посредством аденовируса для применения в лечении и/или профилактике заболеваний у субъекта, более предпочтительно, опухоли или злокачественного новообразования, где способ включает введение субъекту аденовируса и ингибитора CDK4/6.
2-я группа Далее раскрываются дополнительные варианты осуществления такого третьего аспекта.
Вариант осуществления 2: Аденовирус для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Аденовирус для применения в соответствии с одним из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Аденовирус для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Аденовирус для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Аденовирус для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Аденовирус для применения в соответствии с вариантом осуществления 6, где белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Аденовирус для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Аденовирус для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Аденовирус для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Аденовирус для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Аденовирус для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Аденовирус для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Аденовирус для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Аденовирус для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Аденовирус для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Аденовирус для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Аденовирус для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Аденовирус для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Аденовирус для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Аденовирус для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающий XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Аденовирус для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Аденовирус для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Аденовирус для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Аденовирус для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Аденовирус для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Аденовирус для применения в соответствии с вариантом осуществления 26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Аденовирус для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Аденовирус для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Аденовирус для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Аденовирус для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Аденовирус для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Аденовирус для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP.
Вариант осуществления 34: Аденовирус для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Аденовирус для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Аденовирус для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Аденовирус для применения по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP и/или ингибитор бромодомена вводятся субъекту отдельно или в виде комбинации.
Вариант осуществления 38: Аденовирус для применения по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Аденовирус для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Аденовирус для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Аденовирус для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Аденовирус для применения в соответствии с вариантом осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Аденовирус для применения в соответствии с вариантом осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44. Аденовирус для применения в соответствии с вариантом осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45. Аденовирус для применения по любому из вариантов осуществления 1-44, где клетки опухолевых клеток обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии или нечувствительны к ним.
Вариант осуществления 46: Аденовирус для применения в соответствии с вариантом осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Аденовирус для применения по п.46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Аденовирус для применения по п.47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Аденовирус для применения по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Аденовирус для применения по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Аденовирус для применения по любому из вариантов осуществления 1-50, где клетки опухоли содержат YB-1 в ядре.
Вариант осуществления 52: Аденовирус для применения по любому из вариантов осуществления 1-51, где клетки опухоли содержат YB-1 в ядре после индукции.
Вариант осуществления 53: Аденовирус для применения в соответствии с вариантом осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Аденовирус для применения в соответствии с вариантом осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Аденовирус для применения по любому из пп.1-54, где опухоль выбрана из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
Задача, лежащая в основе настоящего изобретения, также решается в четвертом аспекте, который также является первым вариантом осуществления такого четвертого аспекта, посредством ингибитора CDK4/6 для применения в лечении и/или профилактике заболеваний у субъекта, более предпочтительно, опухоли или злокачественного новообразования, где способ включает введение субъекту аденовируса и ингибитора CDK4/6.
3-я группа Далее раскрываются дополнительные варианты осуществления такого четвертого аспекта.
Вариант осуществления 2: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Ингибитор CDK4/6 для применения в соответствии с одним из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 6, где белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающий XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP.
Вариант осуществления 34: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP и/или ингибитор бромодомена вводят субъекту отдельно или в виде любой комбинации.
Вариант осуществления 38: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44. Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45. Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-44, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
Вариант осуществления 46: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Ингибитор CDK4/6 для применения по п.46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Ингибитор CDK4/6 для применения по п.47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-50, где клетки опухоли содержат в ядре YB-1.
Вариант осуществления 52: Ингибитор CDK4/6 для применения по любому из вариантов осуществления 1-51, где клетки опухоли после индукции содержат в ядре YB-1.
Вариант осуществления 53: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Ингибитор CDK4/6 для применения в соответствии с вариантом осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Ингибитор CDK4/6 для применения по любому из пп.1-54, где опухоль выбран из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
Задача, лежащая в основе настоящего изобретения, решается в пятом аспекте, который также является первым вариантом осуществления такого пятого аспекта посредством ингибитора PARP для применения в лечении и/или профилактике заболеваний у субъекта, более предпочтительно, опухоли или злокачественного новообразования, где способ включает введение субъекту аденовируса, ингибитора CDK4/6 и ингибитора PARP.
Далее раскрываются дополнительные варианты осуществления такого пятого аспекта.
Вариант осуществления 2: Ингибитор PARP для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Ингибитор PARP для применения по любому из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Ингибитор PARP для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Ингибитор PARP для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Ингибитор PARP для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Ингибитор PARP для применения в соответствии с вариантом осуществления 6, белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Ингибитор PARP для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Ингибитор PARP для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Ингибитор PARP для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Ингибитор PARP для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Ингибитор PARP для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Ингибитор PARP для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Ингибитор PARP для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Ингибитор PARP для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Ингибитор PARP для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Ингибитор PARP для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Ингибитор PARP для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Ингибитор PARP для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Ингибитор PARP для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Ингибитор PARP для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающей XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Ингибитор PARP для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Ингибитор PARP для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Ингибитор PARP для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Ингибитор PARP для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Ингибитор PARP для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Ингибитор PARP для применения в соответствии с вариантом осуществления26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Ингибитор PARP для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Ингибитор PARP для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Ингибитор PARP для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Ингибитор PARP для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Ингибитор PARP для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Ингибитор PARP для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP
Вариант осуществления 34: The PARP для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Ингибитор PARP для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Ингибитор PARP для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ-1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Ингибитор PARP для применения по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP и/или ингибитор бромодомена вводят субъекту отдельно или в виде любой комбинации.
Вариант осуществления 38: Ингибитор PARP для применения по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Ингибитор PARP для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Ингибитор PARP для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Ингибитор PARP для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Ингибитор PARP для применения в соответствии с вариантом осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Ингибитор PARP для применения в соответствии с вариантом осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44. Ингибитор PARP для применения в соответствии с вариантом осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45. Ингибитор PARP для применения по любому из вариантов осуществления 1-44, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
Вариант осуществления 46: Ингибитор PARP для применения в соответствии с вариантом осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Ингибитор PARP для применения по п.46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Ингибитор PARP для применения по п.47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Ингибитор PARP для применения по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Ингибитор PARP для применения по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Ингибитор PARP для применения по любому из вариантов осуществления 1-50, где клетки опухоли содержат в ядре YB-1.
Вариант осуществления 52: Ингибитор PARP для применения по любому из вариантов осуществления 1-51, где клетки опухоли после индукции содержат в ядре YB-1.
Вариант осуществления 53: Ингибитор PARP для применения в соответствии с вариантом осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Ингибитор PARP для применения в соответствии с вариантом осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Ингибитор PARP для применения по любому из пп.1-54, где опухоль выбран из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
Задача, лежащая в основе настоящего изобретения, решается в шестом аспекте, который также является первым вариантом осуществления такого шестого аспекта, посредством ингибитора бромодомена для применения в лечении и/или профилактике заболеваний у субъекта, более предпочтительно, опухоли или злокачественного новообразования, где способ включает введение субъекту аденовируса, ингибитора CDK4/6 и ингибитора бромодомена.
Далее раскрываются дополнительные варианты осуществления такого шестого аспекта.
Вариант осуществления 2: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Ингибитор бромодомена для применения по любому из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Ингибитор бромодомена для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 6, белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Ингибитор бромодомена для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Ингибитор бромодомена для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Ингибитор бромодомена для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Ингибитор бромодомена для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающей XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Ингибитор бромодомена для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Ингибитор бромодомена для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Ингибитор бромодомена для применения в соответствии с вариантом осуществления26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP
Вариант осуществления 34: Бромодомен для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ-1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP и/или ингибитор бромодомена вводят субъекту отдельно или в виде любой комбинации.
Вариант осуществления 38: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44. Ингибитор бромодомена для применения в соответствии с вариантом осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45. Ингибитор бромодомена для применения по любому из вариантов осуществления 1-44, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
Вариант осуществления 46: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Ингибитор бромодомена для применения по п.46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Ингибитор бромодомена для применения по п.47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Ингибитор бромодомена для применения по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-50, где клетки опухоли содержат в ядре YB-1.
Вариант осуществления 52: Ингибитор бромодомена для применения по любому из вариантов осуществления 1-51, где клетки опухоли после индукции содержат в ядре YB-1.
Вариант осуществления 53: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Ингибитор бромодомена для применения по любому из пп.1-54, где опухоль выбран из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
Задача, лежащая в основе настоящего изобретения, решается в седьмом аспекте, который также является первым вариантом осуществления такого седьмого аспекта посредством нутлина или производного нутлина для применения в лечении и/или профилактике заболеваний у субъекта, более предпочтительно, опухоли или злокачественного новообразования, где способ включает введение субъекту аденовируса, ингибитора CDK4/6 и ингибитора бромодомена.
Далее раскрываются дополнительные варианты осуществления такого седьмого аспекта.
Вариант осуществления 2: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Нутлин или производное нутлина для применения по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 6, белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Нутлин или производное нутлина для применения по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Нутлин или производное нутлина для применения по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Нутлин или производное нутлина для применения по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Нутлин или производное нутлина для применения по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающей XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Нутлин или производное нутлина для применения по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Нутлин или производное нутлина для применения по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP
Вариант осуществления 34: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб, талазопариб и BMN673.
Вариант осуществления 35: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ-1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP, ингибитор бромодомена или нутлин или производное нутлина вводят субъекту отдельно или в виде любой комбинации.
Вариант осуществления 38: Нутлин или производное нутлина для применения по любому из пп.1-37, где производное нутлина выбрано из группы, включающей NVP-HDM201, идазанутлин, AM-8553, SAR405838, нутлин-3a и AMG232.
Вариант осуществления 39: Нутлин или производное нутлина для применения по любому из пп.1-37, где производное нутлина отличается от нутлина-3a.
Вариант осуществления 40: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-39, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 41: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-40, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 42: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-40, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 43: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-40, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 44: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 43, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 45: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 43, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 46. Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 45, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 47. Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-46, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
Вариант осуществления 48: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 47, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 49: Нутлин или производное нутлина для применения по п.48, где резистентность опосредована транспортером ABC.
Вариант осуществления 50: Нутлин или производное нутлина для применения по п.49, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 51: Нутлин или производное нутлина для применения по любому из вариантов осуществления 47-50, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 52: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-51, где клетки опухоли являются Rb-отрицательными.
Вариант осуществления 53: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-52, где клетки опухоли являются Rb-положительными.
Вариант осуществления 54: Нутлин или производное нутлина для применения по любому из вариантов осуществления 1-53, где клетки опухоли содержат в ядре YB-1.
Вариант осуществления 55: Нутлин или производное нутлина или для применения по любому из вариантов осуществления 1-54, где клетки опухоли после индукции содержат в ядре YB-1 .
Вариант осуществления 56: Нутлин или производное нутлина для применения в соответствии с вариантом осуществления 55, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 57: Ингибитор бромодомена для применения в соответствии с вариантом осуществления 56, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Проблема, лежащая в основе настоящего изобретения, решается в восьмом аспекте, который также является первым вариантом осуществления такого седьмого аспекта, посредством комбинированной терапии, при этом такая комбинированная терапия включает введение нуждающемуся в этом субъекту, среди прочего,
a) аденовируса, в частности такой, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, и ингибитор PARP, в частности такой, как определено в настоящем документе в соответствии с четвертым аспектом;
b) аденовируса, в частности, такого, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, и ингибитора бромодомена, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом;
c) аденовируса, в частности, такого, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, и нутлина или производного нутлина, в частности, такого, как определено в настоящем документе в соответствии с шестым аспектом;
d) аденовируса, в частности, такого, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, ингибитора бромодомена, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом; и нутлина или производного нутлина, в частности, такого, как определено в настоящем документе в соответствии с шестым аспектом;
e) аденовируса, в частности, такого, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, ингибитора бромодомена, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом; и ингибитора PARP, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом; и
f) аденовируса, в частности, такого, как определено в настоящем документе в соответствии с третьим аспектом, включая любой его вариант осуществления; ингибитора CDK4/6, в частности, такого, как определено в настоящем документе в соответствии с четвертым аспектом, включая любой его вариант осуществления, ингибитора бромодомена, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом; ингибитора PARP, в частности, такого, как определено в настоящем документе в соответствии с пятым аспектом; и нутлина или производного нутлина, в частности, такого, как определено в настоящем документе в соответствии с шестым аспектом.
В рамках настоящего изобретения различные варианты осуществления, описанные в связи с третьим аспектом, включая любой его вариант осуществления, с четвертым аспектом, включая любой его вариант осуществления, с пятым аспектом, включая любой его вариант осуществления, и с шестым аспектом, включая любой его вариант осуществления, могут быть вариантами комбинированной терапии в соответствии с восьмым аспектом в ее различных формах, в частности, как определено выше как a), b), c), d), e) и f).
Задача, лежащая в основе настоящего изобретения, решается в девятом аспекте, который также является первым вариантом осуществления такого седьмого аспекта, посредством способа лечения и/или профилактики заболевания у субъекта, более предпочтительно, опухоли или рака, где способ включает введение субъекту аденовируса и ингибитора CDK4/6.
Далее описаны дополнительные варианты осуществления такого девятого аспекта.
Вариант осуществления 2: Способ по варианту осуществления 1, где аденовирусом является онколитический аденовирус.
Вариант осуществления 3: Способ по любому из вариантов осуществления 1 и 2, где аденовирус реплицируется YB-1-зависимым образом.
Вариант осуществления 4: Способ по варианту осуществления 3, где аденовирус является дефицитным по репликации в клетках, не содержащих YB-1 в ядре, но реплицируется в клетках, которые содержат YB-1 в ядре.
Вариант осуществления 5: Способ по любому из вариантов осуществления 2-4, где аденовирус кодирует онкогенный белок, где онкогенный белок трансактивирует по меньшей мере один аденовирусный ген, причем аденовирусный ген выбран из группы, включающей E1B55kDa, E4orf6, E4orf3 и E3ADP.
Вариант осуществления 6: Способ по варианту осуществления 5, где онкогенным белком является белок E1A.
Вариант осуществления 7: Способ по варианту осуществления 6, белок E1A способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 8: Способ по варианту осуществления 6, где белок E1A не способен связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 9: Способ по любому из вариантов осуществления 6-8, где белок E1A не вызывает локализацию YB-1 в ядре.
Вариант осуществления 10: Способ по любому из вариантов осуществления 5-9, где онкогенный белок демонстрирует одну или несколько мутаций или делеций по сравнению с белком онкогена E1A дикого типа.
Вариант осуществления 11: Способ по варианту осуществления 10, где делеция выбрана из группы, включающей делеции фрагментов CR3 и делеции N-конца и делеции C-конца.
Вариант осуществления 12: Способ по любому из вариантов осуществления 6-11, где белок E1A способен связываться с Rb.
Вариант осуществления 13: Способ по любому из вариантов осуществления 6-12, где белок E1A содержит одну или несколько мутаций или делеций по сравнению с онкогенным белком дикого типа, причем делеция, предпочтительно, является делецией в области CR1 и/или области CR2.
Вариант осуществления 14: Способ по варианту осуществления 13, где белок E1A не способен связываться с Rb.
Вариант осуществления 15: Способ по любому из вариантов осуществления 1-14, где вирусом является аденовирус, экспрессирующий белок E1A12S.
Вариант осуществления 16: Способ по любому из вариантов осуществления 1-15, где вирусом является аденовирус, не экспрессирующий белок E1A13S.
Вариант осуществления 17: Способ по любому из вариантов осуществления 1-16, где вирусом является аденовирус, не содержащий функционально активную аденовирусную область E3.
Вариант осуществления 18: Способ по любому из вариантов осуществления 1-17, где вирусом является аденовирус, не экспрессирующий белок E1B 19 кДа.
Вариант осуществления 19: Способ по любому из вариантов осуществления 1-18, где вирусом является аденовирус, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 20: Способ по любому из вариантов осуществления 1-19, где вирусом является аденовирус серотипа 5.
Вариант осуществления 21: Способ по любому из вариантов осуществления 1-20, где аденовирус выбран из группы, включающей XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Вариант осуществления 22: Способ по варианту осуществления 21, где аденовирусом является XVir-N-31.
Вариант осуществления 23: Способ по варианту осуществления 21, где аденовирусом является dl520, и где аденовирусная область E3 является функционально неактивной.
Вариант осуществления 24: Способ по любому из вариантов осуществления 21-23, где аденовирусом является dl520, и где в dl520 отсутствует экспрессия белка E1B 19 кДа.
Вариант осуществления 25: Способ по любому из вариантов осуществления 21-24, где аденовирусом является dl520, экспрессирующий RGD-мотив на волокне.
Вариант осуществления 26: Способ по любому из вариантов осуществления 1-25, где вирус кодирует YB-1.
Вариант осуществления 27: Способ по варианту осуществления26, где ген, кодирующий YB-1, находится под контролем тканеспецифического промотора, опухолеспецифического промотора и/или YB-1 зависимого промотора.
Вариант осуществления 28: Способ по варианту осуществления 27, где YB-1 зависимым промотором является аденовирусный Е2-поздний промотор.
Вариант осуществления 29: Способ по любому из вариантов осуществления 1-28, где ингибитор CDK4/6 представляет собой соединение, которое снижает фосфорилирование Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 30: Способ по любому из вариантов осуществления 1-29, где ингибитор CDK4/6 представляет собой соединение, которое уменьшает экспрессию Rb в клетке, предпочтительно, в опухолевой клетке.
Вариант осуществления 31: Способ по любому из вариантов осуществления 1-30, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
Вариант осуществления 32: Способ по любому из вариантов осуществления 1-31, где ингибитор CDK4/6 вызывает арест G1 в клетке и ингибирует E2F1.
Вариант осуществления 33: Способ по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора PARP.
Вариант осуществления 34: Способ по варианту осуществления 33, где ингибитор PARP выбран из группы, включающей олапариб, велипариб, рукапариб и BMN673.
Вариант осуществления 35: Способ по любому из вариантов осуществления 1-32, где способ дополнительно включает введение субъекту ингибитора бромодомена.
Вариант осуществления 36: Способ по варианту осуществления 35, где ингибитор бромодомена выбран из группы, включающей JQ-1, OTX-015, I-BET151, CPI-0610, I-BET762, CPI203, PFI-1 и MS 436.
Вариант осуществления 37: Способ по любому из вариантов осуществления 1-36, где аденовирус, ингибитор CDK4/6, ингибитор PARP и/или ингибитор бромодомена вводят субъекту отдельно или в виде любой комбинации.
Вариант осуществления 38: Способ по любому из вариантов осуществления 1-37, где клетки опухоли имеют нарушение сигнального пути CDK4/6.
Вариант осуществления 39: Способ по любому из вариантов осуществления 1-38, где клетки опухоли имеют неконтролируемый переход G1-S клеточного цикла.
Вариант осуществления 40: Способ по любому из вариантов осуществления 1-38, где клетки опухоли имеют потерю функцию мутации или делеции в гене, выбранном из группы, включающей ген RB1, ген CDKN2A и ген CDKN2B.
Вариант осуществления 41: Способ по любому из вариантов осуществления 1-38, где клетки опухоли имеют амплификацию гена и/или активирующую мутацию гена.
Вариант осуществления 42: Способ по варианту осуществления 41, где ген выбран из группы, включающей CCND1, E2F1, E2F2, E2F3, CDK4 и CDK6.
Вариант осуществления 43: Способ по варианту осуществления 41, где ген является геном, кодирующим компонент митогенного сигнального пути.
Вариант осуществления 44: Способ по варианту осуществления 43, где митогенный сигнальный путь выбран из группы, включающей сигнальный путь PI3K и сигнальный путь MAPK.
Вариант осуществления 45: Способ по любому из вариантов осуществления 1-44, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
Вариант осуществления 46: Способ по варианту осуществления 45, где фармацевтически активный агент является цитостатическим.
Вариант осуществления 47: Способ по пункту 46, где резистентность опосредована транспортером ABC.
Вариант осуществления 48: Способ по пункту 47, где транспортер ABC выбран из группы, включающей MRP и MDR, в частности, MDR-1.
Вариант осуществления 49: Способ по любому из вариантов осуществления 45-48, где резистентность представляет собой множественную резистентность или полирезистентность, в частности, мульти- или полирезистентность в отношении цитостатиков и/или лучевой терапии.
Вариант осуществления 50: Способ по любому из вариантов осуществления 1-49, где клетки опухоли являются Rb-положительными.
Вариант осуществления 51: Способ по любому из вариантов осуществления 1-50, где клетки опухоли содержат в ядре YB-1.
Вариант осуществления 52: Способ по любому из вариантов осуществления 1-51, где клетки опухоли после индукции содержат в ядре YB-1 .
Вариант осуществления 53: Способ по варианту осуществления 52, где транспорт YB-1 в ядре запускается по меньшей мере одним действием, выбранным из группы, включающей облучение, введение цитостатиков и гипертермию.
Вариант осуществления 54: Способ по варианту осуществления 53, где действие применяется к клетке, органу или организму, предпочтительно, к организму, который в этом нуждается, более предпочтительно, к организму, страдающему опухолью.
Вариант осуществления 55: Способ по пунктам 1-54, где опухоль выбран из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
В десятом аспекте настоящее изобретение также относится к применению композиции при изготовлении лекарственного средства, где композиция представляет собой композицию, описанную в связи с первым аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи со вторым аспектом настоящего изобретения, включая любой его вариант осуществления.
В одиннадцатом аспекте настоящее изобретение относится также к применению аденовируса при изготовлении лекарственного средства, где аденовирусом является аденовирус, описанный в связи с третьим аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи с третьим аспектом настоящего изобретения, включая любой его вариант осуществления.
В двенадцатом аспекте настоящее изобретение относится также к применению ингибитора CDK4/6 при изготовлении лекарственного средства, где ингибитор CDK4/6 представляет собой ингибитор CDK4/6, как описано в связи с четвертым аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи с четвертым аспектом настоящего изобретения, включая любой его вариант осуществления.
В 13ом аспекте настоящее изобретение относится также к применению ингибитора PARP при изготовлении лекарственного средства, где ингибитор PARP представляет собой ингибитор PARP, описанный в связи с пятым аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи с пятым аспектом настоящего изобретения, включая любой его вариант осуществления.
В 14ом аспекте настоящее изобретение относится также к применению ингибитора бромодомена при изготовлении лекарственного средства, где ингибитор бромодомена представляет собой ингибитор бромодомена, описанный в связи с шестым аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи с шестым аспектом настоящего изобретения, включая любой его вариант осуществления.
В 15ом аспекте настоящее изобретение относится также к применению нутлина или его производного при изготовлении лекарственного средства, где нутлин или производное нутлина представляет собой нутлин или производное нутлина, описанный в связи с седьмым аспектом настоящего изобретения, включая любой его вариант осуществления, и лекарственное средство предназначено для лечения и/или профилактики заболевания, указанного в связи с седьмым аспектом настоящего изобретения, включая любой его вариант осуществления.
В варианте осуществления каждого и любого аспекта, раскрытого в настоящем документе, включая любой его вариант осуществления, ингибитор CDK4/6 представляет собой агент, который ингибирует или способен ингибировать CDK4/6, вызывая, таким образом, задержку соответствующим образом обработанной клетки в фазе G1.
В варианте осуществления каждого и любого аспекта настоящего изобретения, включая любой его вариант осуществления, ингибитор бромодомена представляет собой двухвалентный ингибитор бромодомена, более предпочтительно, ингибитор бромодомена представляет собой AZD5153 следующей формулы:
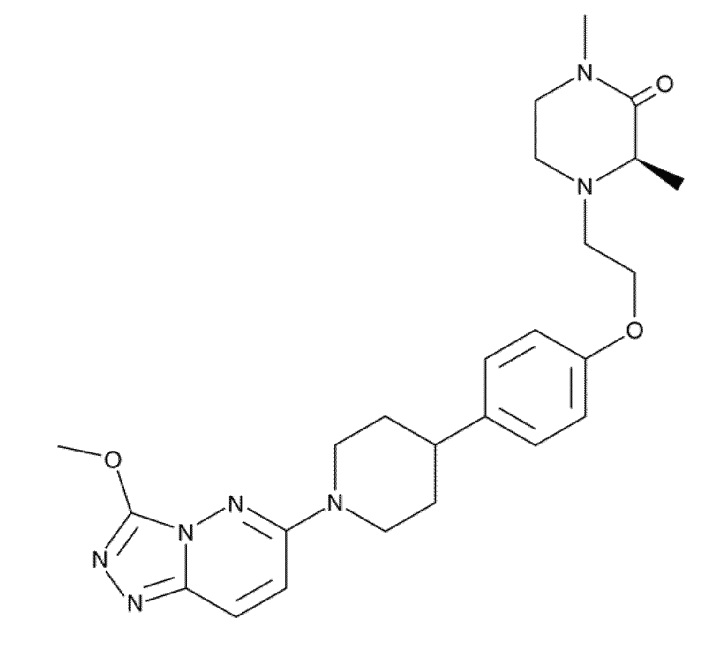
В одном из вариантов осуществления каждого и любого аспекта настоящего изобретения, включая любой его вариант, ингибитор бромодомена представляет собой деструктор BET, предпочтительно, ингибитор бромодомена выбирают из группы, включающей dBET6 следующей формулы и ARV771 следующей формулы:
(см., например, Scheepstra M et al. (Computational and Structural Biotechnology Journal 17(2019) 160-179).
Специалисту в данной области техники понятно, что каждый и любой вариант осуществления одного аспекта настоящего изобретения также является вариантом осуществления каждого и любого из других аспектов настоящего изобретения, включая любой его вариант осуществления.
Не желая связывать себя какой-либо теорией, авторы настоящего изобретения неожиданно обнаружили, что комбинация онколитического вируса, предпочтительно, онколитического аденовируса, с ингибитором CDK4/6 увеличивает эффективность терапии опухолей на основе такого онколитического аденовируса. Более конкретно, предполагается, что ингибитор CDK4/6 ингибирует E2F-1 (также упоминаемым как E2F-1 в настоящем документе) таким образом снижая его эффективную концентрацию, предпочтительно, в опухолевых клетках, и синхронизирует арест G1 в клетках. Благодаря этому большее количество инфицированных клеток может завершить весь жизненный цикл вируса.
Основываясь на доказательствах и выводах, представленных в настоящем документе, специалист в данной области поймет, что для использования при практическом осуществлении настоящего изобретения подходит любой - мутант - аденовирус, что дает по меньшей мере всего 10%, 20% или 30% экспрессии дикого типа и, соответственно, таким аденовирусом достигается активность E1B55K и E4orf6. Специалисту в данной области будет понятно, что такой мутантный аденовирус может быть получен путем модификации E1A. Примерами мутантных аденовирусов являются аденовирус XVir-N-31, dl520, AdΔ24, AdΔ24-RGD, dl922-947, E1Ad/01/07, dl1119/1131, CB 016, VCN-01, E1Adl1107, E1Adl1101, ORCA-010, Enadenotucirev и вирусы без экспрессированного вирусного онкогена, способный связывать функциональный продукт гена-супрессора опухоли Rb.
Эпонимом аденовирусов является первое выделение вируса в миндалинах и аденоидной ткани человека в 1953 г., осуществленное Уоллесом П. Роу и Робертом Дж. Хюбнером (Rowe et al., 1953). Семейство Adenoviridae включает пять родов, а именно мастаденовирусы, авиаденовирусы, сиаденовирусы, атаденовирусы и ихтаденовирусы (Modrow, 2013). По онкогенности для новорожденных грызунов их можно разделить на семь подгрупп от HAdV-A до HAdV-G (Boulanger и Blair, 1991), всего 62 серотипа. Таким образом, исследования онколитической виротерапии в основном сосредоточены на мастаденовирусе типа C серотипа 5.
Непокрытый икосаэдрический капсид размером от 80 до 110 нм состоит из 252 капсомеров, которые состоят из 12 пентонов, собранных из пентонной основы и шипообразных белковых структур, называемых волокнами, на вершинах капсида и 249 граней, называемых гексонами (Modrow, 2013). Весь жизненный цикл аденовирусов можно подразделить на раннюю фазу с проникновением в клетку, ядерную транслокацию вирусного генома, транскрипцию и трансляцию ранних генов и позднюю фазу с транскрипцией и трансляцией поздних генов. Таким образом, за сборку структурных белков и созревание вирионов в основном ответственны поздние белки (Russell, 2000). В пермиссивных клетках ранняя фаза занимает около 6-8 часов с последующей поздней фазой около 4-6 часов. Присоединение происходит за счет взаимодействия утолщенной структуры, которая присутствует на каждом конце волокнистых структур, с рецептором на клетках-мишенях, по крайней мере, в случае аденовирусов HAdV-A, -C, -E и -F. Поскольку этот рецептор был обнаружен как тот же самый, который отвечает за адсорбцию вируса Коксаки B, этот рецептор получил название рецептора вируса Коксаки и аденовируса (CAR) (Bergelson, 1997). Кроме того, связывание на поверхности клетки-мишени поддерживается «мостиковыми молекулами», растворимыми белками в жидкостях организма, такими как факторы свертывания крови VII и X, которые опосредуют связывание белков волокон определенных типов аденовирусов (Modrow, 2013).
После этой стадии адсорбции RGD-мотив (аргинин-глицин-аспарагиновая кислота) в основании пентона взаимодействует с гетеродимерными интегринами αvβ3 или αvβ5, которые действуют в этом процессе как корецепторы. Это взаимодействие приводит к интернализации вируса (Wickham et al., 1993). Впоследствии происходит эндоцитоз через клатрин-опосредованную интернализацию в цитоплазматической мембране, и вирус находится в эндосомах. После подкисления эндоцитарных пузырьков белок вирусного волокна меняет свою конформацию, что приводит к разрушению эндосомальной мембраны (Greber et al., 1996). Теперь вирусные частицы в цитоплазме свободны. За счет связывания остаточных частиц с динеинами микротрубочек вирусный геном переносится в ядро (Modrow, 2013).
Геном аденовирусов состоит из двухцепочечной линейной ДНК длиной 36-38 т.п.н. Путем взаимодействия двух концевых молекул белка (TP), ковалентно связанных с обоими 5'-концами, образуется квазициклическое состояние (Modrow, 2013). В целом, пять кодирующих областей аденовирусного генома можно подразделить на ранние гены E1-E4, активные в основном в ранней фазе инфекции, и поздние гены (L1-L5), которые кодируют белки, в основном необходимые для образования вирусных частиц (Modrow, 2013).
Репликация аденовирусов особенно зависит от экспрессии раннего вирусного гена E2, который сильно индуцируется большим белком E1A (E1A13S). Первым вирусным геном, который будет транскрибироваться после инфицирования, является ранняя область 1A (E1A). Первичный транскрипт E1A обрабатывается с помощью дифференциального сплайсинга с получением пяти различных сообщений с коэффициентами седиментации 13S, 12S, 11S, 10S и 9S. мРНК 13S и 12S наиболее многочисленны на ранних этапах инфицирования, тогда как мРНК 9S наиболее распространена на поздних этапах. мРНК 11S и 10S представляют собой второстепенные виды, которые становятся более многочисленными на поздних сроках после инфицирования. мРНК 13S, 12S, 11S, 10S и 9S E1A кодируют 289 остатков (R), 243R, 217R, 171R и 55R белков, соответственно, все из которых обнаруживаются in vivo, за исключением продукта 9S, который был обнаружен только in vitro. Обычно экспрессия аденовирусного гена сильно регулируется в ходе инфекции с высокой степенью сложности. Таким образом, транскрипция генов E2, продукты которых кодируют вирусную ДНК-полимеразу и другие белки, необходимые для эффективной репликации вируса, находится под контролем двух промоторов, E2-раннего и E2-позднего промотора.
Благодаря двум перекрывающимся областям контроля транскрипции, E2-ранний промотор может быть подразделен на главный промотор, начинающийся в положении +1, и минорный промотор, начинающийся в положении -26, которые оба содержат мотив TATA (Swaminathan и Thimmapaya, 1996). Эти мотивы служат сайтами связывания для белков, связывающих ТАТА-бокс (TBP). Кроме того, один сайт связывания активирующего транскрипционного фактора (ATF) между положениями -68 и -77 и два сайта связывания E2F1/DP-1 (TTTCGCGC), выровненные в перевернутой ориентации по отношению друг к другу, расположены в положении -35 и -63 основного E2-раннего промотора (Swaminathan and Thimmapaya, 1996). Активация раннего промотора E2 через E1A в основном зависит от двух сайтов связывания E2F1, локализованных в основной части промотора.
На промежуточных стадиях инфицирования, примерно через 6 hpi (часов после инфицирования), экспрессия генов E2 контролируется E2-поздним промотором. В положениях от -33 до -22 его последовательности из 157 п.н. находится ТАТА-бокс, который может связываться и активироваться клеточным ТВР (Swaminathan и Thimmapaya, 1996). Более того, для E2-позднего промотора характерны два сайта узнавания SP1 и три бокса CCAAT.
Поскольку было показано, что клеточный фактор YB-1 способен связываться с инвертированными боксами CCAAT, было исследовано взаимодействие между связывающим белком 1 Y-бокса (YB-1) и E2-поздним промотором. Holm et al. показали в 2002 г., что на самом деле существует специфическое взаимодействие YB-1 с Y-боксами (инвертированные CCAAT-боксы), присутствующими в E2-позднем промоторе, и способными контролировать активность этого промотора (Holm et al., 2002). Чтобы проявить свою трансактивирующую активность, YB-1 должен быть перемещен в ядро через аденовирусный комплекс E1B-55k/E4-orf6. Эти ранние вирусные гены экспрессируются после трансактивации E1A-13S (Frisch и Mymryk, 2002).
Клеточный фактор YB-1, кодируемый геном YBX1, представляет собой домен холодового шока, несущий ДНК-связывающий белок с множеством функций в транскрипции, сплайсинге, контроле трансляции и восстановлении повреждений ДНК (Kohno et al., 2003). Более того, он играет важную роль в устойчивости к лекарствам из-за активации генов MDR1 и MRP1, которые участвуют в развитии фенотипа множественной лекарственной устойчивости в раковых клетках (Mantwill et al., 2006). Экспрессия YB-1 индуцируется последующим переносом ядер посредством воздействия внешних стрессовых факторов, таких как аденовирусная инфекция, химиотерапия или УФ-излучение (Mantwill et al., 2006).
Активация транскрипции ранних и поздних генов аденовируса имеет решающее значение для жизненного цикла вируса. Вкратце, жизненный цикл вируса начинается с активации транскрипции E1A, за которой следует каскад активации генов E2, E3 и E4. Наконец, главный поздний промотор (MLP) активируется, чтобы координировать экспрессию капсида и дополнительных белков, в значительной степени участвующих в инкапсидации генома (Turner et al 2015). Чтобы преодолеть блокировку репликации вирусной ДНК, присутствующей в непролиферирующих клетках, вирус экспрессирует ранние белки 1A (E1A). Эти немедленные ранние белки переводят клетки в S-фазу и вызывают экспрессию всех других вирусных ранних генов. В процессе инфицирования несколько изоформ E1A экспрессируются с белками из 289, 243, 217, 171 и 55 остатков, представленных для аденовируса человека типа 5. В контексте инфекции основным драйвером экспрессии вирусного гена является большой белок (Radko et al 2015).
После инфицирования экспрессия аденовирусного белка E1A способствует прогрессированию клеточного цикла из фазы G0/G1 в S-фазу и репликации вируса даже в терминально дифференцированных эпителиальных клетках, основной мишени аденовирусов человека. Этот процесс считается необходимым для жизненного цикла аденовирусов.
Аденовирусы были разработаны для инфицирования, репликации и уничтожения раковых клеток, сохраняя при этом нормальные клетки. После инфицирования и репликации в опухолевых клетках онколитические вирусы убивают клетки, высвобождая вирионы для последующих циклов амплификации. Для достижения репликации только в опухолевых клетках были проделаны два вида генетических модификаций, в результате чего были сконструированы три подкласса онколитического аденовируса (также называемого в настоящем документе как CRAd), все из которых могут быть использованы при практическом осуществлении настоящего изобретения. Кроме того, в WO 2003/099859 описаны онколитические аденовирусы, подходящие для использования при практическом осуществлении настоящего изобретения, среди прочего.
CRAd типа I характеризуются мутациями или делециями в области E1 генома, препятствующими инактивации регуляторов клеточного цикла, таких как p53 и белок ретинобластомы (Rb). Как следствие, CRAd типа I реплицируются в активно делящихся опухолевых клетках. Например, Onyx-015, также известный как dl1520, который не может экспрессировать белок E1B-55 кДа, не может инактивировать p53 и избежать ареста клеточного цикла, вызванного p53. Некоторые исследования объясняют молекулярную основу избирательности Onyx-015 отсутствием экспрессии p53 или одного из генов, участвующих в пути p53. Однако O'Shea et al. показали, что поздний экспорт вирусной РНК, а не инактивация p53, определяет селективность вируса Onyx-015. Другие CRAd типа I с делециями в области E1A неспособны связывать Rb и запускать вхождение в S-фазу. Например, dl922-947 и Δ24 содержат делецию 24 нуклеотида в домене CR2 области E1A, что исключает взаимодействие E1A-Rb. В результате эти вирусы реплицируются в основном в опухолевых клетках, где доступен свободный, несвязанный E2F.
Другим способом ограничить репликацию аденовируса опухолевыми клетками является регулирование транскрипции вирусных генов, необходимых для репликации вируса. В CRAd типа II геном находится под контролем опухолеспецифического промотора. Эти промоторы произошли от генов, которые, как известно, преимущественно экспрессируются в некоторых опухолях по сравнению с нормальной тканью (например, теломераза или циклооксигеназа II); или которые сверхэкспрессируются в опухолях (например, специфический антиген предстательной железы, PSA или α-фетопротеин, AFP) по сравнению с нормальными тканями. В CRAd типа III, таких как XVir-N-31 (Ad-Delo3-RGD), характерна делеция домена CR3 трансактивации в белке E1A13S. XVir-N-31 представляет собой аденовирусы с дефектом репликации в нормальных клетках. XVir-N-31 восстанавливает свою репликационную способность за счет присутствия в ядре клеточного многофункционального белка YB-1. Соответственно, CRAd способны реплицироваться только в опухолевых клетках и, таким образом, в конечном итоге лизировать их. Ни мутации p53, ни ras, ни RB не эффективны для восполнения дефицита репликации XVir-N-31. В XVir-N-31 отсутствует E1A13S, следовательно, белок E1B55k и белок E4orf6 не экспрессируются. Этот дефицит дополняется наличием YB-1 в ядре опухолевых клеток, который запускает экспрессию E1B55k и E4orf6 независимо от E1A13S. После индуцирования присутствием YB-1 в ядре E1B55k и E4orf6 переносят дальнейший клеточный YB-1 в ядро, вызывая репликацию вируса.
Клеточный цикл последовательно проходит через стадии промежутка 1 (G1), синтеза (S), промежутка 2 (G2) и митоза (M). Этот прогресс регулируется сложной сигнальной сетью. Белки CDK (циклин-зависимая киназа), CDK1, CDK2, CDK4 и CDK6, являются основными регуляторами развития клеточного цикла при образовании комплекса со специфическими белками циклинов. Конститутивная экспрессия CDK и временной контроль различных циклинов делает возможным регулирование конкретных фаз клеточного цикла с помощью различных комплексов циклин-CDK. Активность CDK негативно регулируется несколькими ингибирующими белками. Различные аспекты биологии и функции CDK были ранее всесторонне рассмотрены.
CDK4 и CDK6, которые демонстрируют структурную и функциональную гомологию, регулируют переход покоящихся клеток в фазе G1 в фазу S при образовании комплекса с белками циклина D. Белки циклина D имеют три подтипа, циклины D1-3, и накапливаются в присутствии митогенных стимулов. Отрицательные регуляторы CDK4/6 включают ингибиторы белков CDK4 (INK4), p16INK4A, p15INK4B, p18INK4C и p19INK4D, которые ингибируют активность CDK4/6, либо уменьшая их связывание с циклином D1, либо непосредственно занимая их каталитические домены.
Киназная активность CDK4/6 приводит к фосфорилированию членов семейства белков ретинобластомы (Rb), включая Rb, p107 и p130, что приводит к их функциональной инактивации. В покоящихся клетках активный гипофосфорилированный Rb связывается с членами семейства факторов транскрипции E2F1, которые образуют комплекс с DP-1/2 вместе с другими корепрессорами и подавляют функцию E2F1 (Rubin et al 2005). При фосфорилировании Rb диссоциирует из этого комплекса и позволяет транскрипцию генов-мишеней E2F, включая циклин A, циклин E и DHFR, среди прочих, которые необходимы для перехода клеточного цикла в S-фазу. Следовательно, ингибирование активности CDK4/6 приводит к дефосфорилированию Rb и подавлению активности E2F, что способствует аресту G0/G1. Это стимулировало разработку ингибиторов CDK4/6 в качестве таргетной терапии раковых клеток.
Нарушение сигнального пути CDK4/6-Rb и неконтролируемый переход G1-S клеточного цикла является общей чертой раковых клеток. Это может происходить из-за различных молекулярных изменений, включая мутации потери функции или делеции гена RB1 (кодирование Rb), CDKN2A (кодирование p16INK4A и p14ARF) или CDKN2B (кодирование p15INK4B). Такая дерегуляция также может быть результатом амплификации или активации мутаций в CCND1 (кодирующем циклин D1), E2F1-3, CDK4, CDK6 или компонентах различных путей передачи митогенных сигналов, таких как пути PI3K или MAPK.
Было разработано несколько АТФ-конкурентных низкомолекулярных ингибиторов CDK. Однако ингибиторы первого поколения, такие как флавопиридол, не селективны и могут подавлять ряд CDK, что может привести к ограниченной эффективности и высокой токсичности. Ингибиторы CDK4/6 следующего поколения обладают высокой селективностью и включают палбоциклиб (PD-0332991 от компании Pfizer), абемациклиб (LY-2835219 от компании Eli Lilly) и рибоциклиб (LEE011 от компании Novartis) и трилациклиб (G1T28). Эти ингибиторы CDK4/6 были протестированы доклинически на моделях in vitro и in vivo нескольких видов рака, включая лейкоз, рак молочной железы, меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легких, саркому, рак яичников, рак почек, предстательной железы и метастатический рак молочной железы (mBC). В большинстве исследований они продемонстрировали постоянный молекулярный и функциональный фенотип с устойчивый молекулярный и функциональный фенотип с дозозависимым снижением фосфорилирования Rb, экспрессии белка и транскрипции генов-мишеней E2F, что коррелирует с арестом G0/G1 и ингибированием клеточной пролиферации. Кроме того, все эти сообщения демонстрируют, что экспрессия Rb является предпосылкой для чувствительности к этим ингибиторам.
Ингибиторы CDK4/6, такие как PD-0332991, приводят к дозозависимому снижению общего белка Rb, которое коррелирует со снижением фосфорилированного Rb. Это снижение общего Rb частично коррелирует со снижением уровней транскриптов RB1 и транскрипции генов-мишеней E2F1 CCNA2 и CCNE2. Кроме того, уровень экспрессии E2F1 значительно снижен.
Ингибиторы CDK4/6, подходящие для использования при практическом осуществлении настоящего изобретения, представлены на фиг. 25.
Как видно из экспериментальной части, ингибитор CDK4/6 подходит для использования в комбинации с вирусом, предпочтительно, аденовирусом и, более предпочтительно, онколитическим аденовирусом, в результате чего ингибитор CDK4/6 вызывает арест G1 клеток и ингибирует E2F1, более конкретно, активность E2F1.
Специалисту в данной области будет понятно, что ингибитор CDK4/6 используется в терапевтически эффективной концентрации.
PARP1 является белком, который важен для восстановления одноцепочечных разрывов («зазубрины» в ДНК). У млекопитающих обнаружено 17 членов семейства PARP, и только 6 из них синтезируют поли-АДФ-рибозу (pADPr). PARP1, PARP2 и PARP3 играют роль в репарации ДНК. PARP1 связывается с ДНК, которая пострадала от одноцепочечных разрывов (SSB) и двухцепочечных разрывов (DSB). Затем PARP1 претерпевает конформационное изменение, которое выравнивает ключевые аминокислотные остатки в активном сайте, тем самым увеличивая его активность. Как только PARP1 активируется, он синтезирует pADPr, который связывается с белками и изменяет их функцию. pADPr быстро разрушается гликогидролазой pADPr для гарантии того, что уровни присутствующего pADPr отражают повреждение ДНК и что ответ на pADPr прекращается после репарации ДНК.
Ингибируя пути репарации ДНК, ингибиторы PARP1 вызывают увеличение одноцепочечных разрывов в ДНК. Это повреждение ДНК не восстанавливается и переносится в дочерние клетки после репликации, поскольку BER больше не возникает. Это приводит к увеличению DSB в опухолях, которые имеют мутации BRCA1 и BRCA2 (Scott et al. 2015, J Clin Oncol., 33(12): 1397-140). Химические структуры ингибиторов PARP, включая препараты-кандидаты PARP рукапариб, велипариб и олапариб, показаны на фиг. 26 и описаны Antolin и Mestres 2014, Oncotarget, 30;5(10):3023-8, включая бензамидный фрагмент, который характеризует все структуры ингибиторов PARP.
Кроме того, хорошо известно, что YB-1 усиливает активность PARP и снижает эффективность ингибиторов PARP1 (Alemasova et al., 2018, Oncotarget, 34, 23349-65), что позволяет предположить, что YB-1 зависит от онколитического аденовируса в сочетании с обоими CDK. Ингибиторы 4/6 и ингибиторы PARP действуют синергетически при уничтожении раковых клеток. Примерами ингибиторов PARP являются олапариб и BMN673 (талазолариб, разработанный Pfizer, США, Clin Cancer Res. 2013, 15; 19 (18): 5003-15).
Специалисту в данной области будет понятно, что ингибитор PARP используется в терапевтически эффективной концентрации.
Ингибиторы CDK4/6, подходящие для использования при практическом осуществлении настоящего изобретения, представлены на фиг. 25.
Аберрации в эпигенетическом ландшафте являются признаком рака, а ацетилирование остатков лизина представляет собой посттрансляционную модификацию, имеющую большое значение для передачи сигналов в клетках и в биологии болезни. Ферменты, которые «записывают» (гистоновые ацетилтрансферазы, HAT) и «стирают» (гистоновые деацетилазы, HDAC) сайты ацетилирования, являются областью обширных исследований при разработке современных лекарственных средств. Рекрутирование белков в макромолекулярные комплексы с помощью ацетилированных остатков лизина опосредуется бромодоменами (BRD), которые являются эволюционно высококонсервативными модулями взаимодействия белков, распознающими мотивы ацетилирования α-N-лизина. Консервативная складка BRD содержит глубокий, в значительной степени гидрофобный сайт связывания ацетиллизина, который представляет собой привлекательный карман для развития небольших фармацевтически активных молекул. В развитие большого количества заболеваний вовлечены белки, содержащие BRD.
Недавно для двух высокоэффективных и селективных ингибиторов, нацеленных на BRD, из семейства BET (бромодомены и экстра-терминальный), предоставили убедительные данные, подтверждающие нацеливание этих BRD на злокачественное новообразование. Подсемейство BET (бромодомен и экстратерминальный домен) бромодоменных белков, состоящее из BRD2, BRD3, BRD4 и BRDT, выполняет различные роли в регуляции транскрипции с помощью РНК-полимеразы II (POLII) и представляет собой новый класс эпигенетических мишеней для лекарственных средств. Эти белки облегчают фазы инициации и элонгации транскрипции за счет связывания с активированным хроматином по ацетилированным остаткам лизина. Распознавание активированного хроматина этими так называемыми эпигенетическими «читателями» способствует привлечению комплекса РНК-полимеразы II к участкам активной транскрипции. Взаимодействие BRD4/P-TEFb важно для быстрой повторной инициации транскрипции после митоза (Muller et al., 2011, Expert Rev. Mol. Medicine, 13, e19). P-TEFb был идентифицирован и очищен как фактор, необходимый для генерации длинных транскриптов с использованием транскрипционной системы in vitro, полученной из клеток дрозофилы. Он представляет собой циклинзависимую киназу, содержащую каталитическую субъединицу Cdk9 и регуляторную субъединицу циклин Т у дрозофилы. У человека существует несколько форм P-TEFb, которые содержат Cdk9 и одну из нескольких субъединиц циклина, циклин T1, T2 и K. P-TEFb ассоциируется с другими факторами, включая белок бромодомена BRD4, и, как было обнаружено, связан с большим комплексом белков, который называется комплексом суперэлонгации (Yang Z, et al., 2005. Mol Cell; 19:535-45; Fu et al., 1999, J Biol Chem., 274:34527-30).
JQ-1 (тиено-триазоло-1-4-диазепин) является мощным ингибитором семейства бромодоменных белков BET, которое включает BRD2, BRD3, BRD4 (Filippakopoulos et al., 2010 Nature 468, 1067-1073). JQ-1 предотвращает взаимодействие между бромодоменом и ацетильной группой, вызывая подавление определенных генов. Были описаны другие бромодомены-ингибиторы BET, включая OTOX15, BAY1238097, GSK2820151, I-BET762 и PLX51107 (Perez-Salvia и Esteller 2017, EPIGENETICS, 12, 323-339; Brandt et al., 2015ACS Chem. Biol., 10, 22-39). JQ-1 структурно родственен бензодиазепинам. Его формулой является C23H25ClN4O2S.
Недавно было показано, что ингибитор BET JQ-1 облегчает аденовирусную инфекцию и опосредованную аденовирусным вектором доставку генов. Обработка клеток JQ-1 вызывает увеличение ассоциации BRD4 с CDK9, субъединицей P-TEFb элонгации транскрипции. Однако, как указано в документе, необходимы дальнейшие исследования, чтобы выяснить механизм, с помощью которого BED4 используется для регулирования аденовирусной инфекции и экспрессии трансгенов (Baojie Lv et al 2018, Scientific reports, 8, 11554). Важно отметить, что вирусная репликация и транскрипция вируса не исследовались. Однако известно, что CDK9 стимулирует высвобождение приостановленной полимеразы и активирует транскрипцию за счет увеличения количества транскрибирующих полимераз и, следовательно, количества синтеза мРНК за период времени (Gressel et al. 2017, eLife, 6, e29736). Кроме того, было показано, что устойчивость к ингибиторам BET можно преодолеть с помощью ингибиторов CDK 4/6 (Jin et al. 2018, Mol Cell;71(4):592-605). Недавно было продемонстрировано резкое усиление взаимодействия P-TEFb-Brd4 от позднего митоза до ранних фаз G1 клеточного цикла и активное привлечение P-TEFb в хромосомы с последующим инициированием транскрипции ключевых генов для прогрессирования G1. Важно отметить, что истощение Brd4 отменяет весь процесс за счет снижения транскрипции основных генов G1, что приводит к аресту клеточного цикла G1 и апоптозу (Yang et al., 2008, Mol Cell Biol., 28:967-976, Kohoutek, 2009, Cell Division, 4. 19).
Однако об использовании YB-1-зависимых онколитических аденовирусов в сочетании с ингибиторами CDK 4/6 и ингибиторами BET ничего не известно.
Следует принимать во внимание и в рамках настоящего изобретения, что другие бромодомены-ингибиторы будут в равной степени подходящими для использования в тройной терапии с использованием вируса, предпочтительно, аденовируса, более предпочтительно, онколитического аденовируса, такого как XVir-N-31, и ингибитора CDK4/6.
Специалисту в данной области будет понятно, что ингибитор бромодомена (Bet) используется в терапевтически эффективной концентрации.
Ингибиторы бромодомена, подходящие для использования при практическом осуществлении настоящего изобретения, представлены на фиг. 27.
Роль MDM2 на p53 и E2F1
Белки p53 и ретинобластомы (Rb) являются двумя ключевыми супрессорами опухолей. Мутации в одном или обоих обнаружены во всех раковых опухолях человека, и оба были тщательно изучены в качестве потенциальных терапевтических мишеней в программах разработки лекарств. Поскольку p53 и E2F1 являются ключевыми регуляторами клеточной пролиферации и жизнеспособности (гибель клеток), их численность и активность строго регулируются. Мышиный белок Double Minute 2 (MDM2), также известный как убиквитин-протеинлигаза E3, по-видимому, регулирует как MDM2-p53, так и путь Rb-E2F1. Поскольку Rb и p53 не способны взаимодействовать напрямую, предполагается, что MDM2 представляет собой мостик между Rb и p53 (Polager and Ginsberg, Nat Rev Cancer 2009, 9, 738-48). Кроме того, подтверждая роль MDM2 как онкогена, было показано, что несколько типов опухолей человека имеют повышенные уровни MDM2, включая саркомы мягких тканей и остеосаркомы, а также опухоли молочной железы. Онкобелок MDM2 убиквитинирует и противостоит р53, но может также выполнять независимые от р53 функции. MDM2 напрямую связывается с p53 и ингибирует его транскрипционную активность. Кроме того, в качестве р53-селективной убиквитинлигазы Е3 MDM2 способствует убиквитилированию р53 и нацеливает его на протеасомную деградацию (Eischem et al, Hum. Mutant. 20014, 35, 728-737). Однако существуют доказательства того, что MDM2 также необходим для выживания р53-дефицитных раковых клеток (Feeley et al, Cancer Res 2017, 77, 3823-3833).
В соответствии с положительным влиянием MDM2 на путь E2F1, MDM2 может физически взаимодействовать с Rb, E2F1 и гетеродимерным партнером E2F1, DP1, чтобы способствовать переходу клеточного цикла G1/S. Соответственно, взаимодействие MDM2 с E2F1 или DP1 может стимулировать транскрипцию генов-мишеней E2F1, которые участвуют в развитии клеточного цикла. Более того, нацеливание E2F1 на деградацию белком F-box SKP2 противодействует связыванию MDM2. Таким образом, существуют доказательства того, что связывание MDM2 увеличивает стабильность E2F1 (Zhang et al. Oncogene 2005, 24, 7238-7247), в то время как ингибирование MDM2 связано со снижением уровня белка E2F1. MDM2, как и многие другие онкопротеины, избирательно связывается с гипофосфорилированным Rb. Взаимодействие между MDM2 и Rb ингибирует образование комплексов Rb-E2F1, что приводит к подавлению функции Rb. Кроме того, взаимодействие между RB и MDM2 вызывает опосредованную MDM2 деградацию Rb. Нынешние факты, по-видимому, подтверждают, что MDM2 также (помимо p53) является ключевым негативным регулятором Rb (Shi и Gu, Genes Cancer. 2012, 3: 240-248; Yap et al., Oncogene 1999, 18, 7681-7689, Wu et al., JBC 2009, 284, 26315-26321).
Нутлин-3
Нутлины являются аналогами цис-имидазолина, которые ингибируют взаимодействие между MDM2 и опухолевым супрессором p53. Нутлин-3 - это соединение, наиболее часто используемое в исследованиях по борьбе с раком. Небольшие молекулы нутлина занимают p53-связывающий карман MDM2 и эффективно нарушают взаимодействие p53-MDM2, что приводит к активации пути p53 в клетках p53 дикого типа. В ответ на лечение нутлином-3 раковые клетки p53 подвергаются либо аресту клеточного цикла (G0/G1 или G2/S), либо апоптозу. Кроме того, нутлин-3 может индуцировать дифференцировку и клеточное старение. Было показано, что на исход лечения с помощью нутлина-3 влияет множество факторов, включая однонуклеотидный полиморфизм MDM2, MDM4, p73, ATM и E2F1. Активация p53 активирует p21 и MDM2, оба из которых являются важными регуляторами Rb. Недавно было показано, что нутлин-3 отрицательно влияет на уровни белков Rb и E2F1 и на фосфорилирование Rb, что значительно влияет на клеточные ответы на нутлин-3 (Du et al., JBC 2009, 284, 26315-26321).
Хотя предыдущие исследования предполагали, что канцерогенность MDM2 связана с его негативной регуляцией р53, р53-независимые взаимодействия могут быть столь же важными. Во время недавних исследований с использованием ингибиторов MDM2 было отмечено, что фактор транскрипции E2F 1 (E2F1) подавляется при ингибировании MDM2, независимо от статуса рака p53. В одной публикации описано использование антисмысловой РНК для ингибирования функции MDM2. Они пришли к выводу, что MDM2 играет роль в росте рака предстательной железы через p53-зависимые и p53-независимые механизмы. Кроме того, было показано, что уровни белков Bcl2, Rb, pRb и E2F1 были снижены, тогда как p21 повышен (Zhang et al 2003, PNAS 2003,100, 11636-11641). Однако было показано, что MDM2 продлевает время полужизни белка E2F1, ингибируя его убиквитинирование. MDM2 заменяет SCF (SKP2), лигазу E2F1 E3. Прямое связывание между MDM2 и E2F1 необходимо для негативных эффектов MDM2 на убиквитинирование E2F1, и делеция сигнала ядерной локализации MDM2 не приводит к потере способности повышать уровень белка E2F1. Понижающая регуляция E2F1 при ингибировании MDM2 не была связана ни с pRB, ни с p14 (Arf). Кроме того, E2F1 был ответственен, по крайней мере, за часть ингибирования клеточной пролиферации, вызванной нокдауном MDM2. В заключение, настоящее исследование предоставляет доказательства того, что стабилизация белка E2F1, вероятно, является еще одним p53-независимым компонентом опосредованного MDM2 онкогенеза (Zhang et al., Oncogene 2005, 24, 7238-7247).
Очевидно, что пути Rb-E2F1 и MDM2-p53, наряду с многогранными перекрестными помехами между ними, являются критическими регуляторами прогрессии и жизнеспособности клеточного цикла. Однако клинические исследования с использованием препаратов, нацеленных на MDM2, в настоящее время не оправдали первоначальных ожиданий, по крайней мере, при использовании в качестве отдельных препаратов. Это стимулировало поиск оптимизированных комбинаций ингибиторов MDM2 с другими противораковыми препаратами.
Было разработано несколько производных нутлина, которые прошли испытания на людях (фиг. 1). Считается, что эти соединения лучше всего работают с опухолями, которые содержат нормальный или «дикий» p53. Однако недавние результаты показывают, что транскрипционная активность E2F1 является критической детерминантой апоптоза, индуцированного антагонистом MDM2, а p73 важен для апоптоза, опосредованного E2F1, индуцированного нутлином-3 (Burgess et al., Frontiers in Oncology 2016, 6, article 7; Skalniak et al., 2019, 29, 151-170; Kitagawa et al., Oncogene 2008, 27, 5303-5314).
Нутлин и его производные (NVP-HDM201, идазанутлин, AM-8553, SAR405838, нутлин-3a, AMG232) показаны на фиг. 43.
Опухоли, которые, в частности, можно лечить с помощью вирусов и, таким образом, комбинаций по настоящему изобретению, описанные в настоящем документе, предпочтительно, представляют собой опухоли, выбранные из группы, включающей опухоли нервной системы, опухоли глаз, опухоли кожи, опухоли мягких тканей, опухоли желудочно-кишечного тракта, опухоли дыхательной системы, опухоли костей, опухоли эндокринной системы, опухоли женской половой системы, опухоли молочной железы, опухоли мужской половой системы, опухоли мочевыводящей системы, опухоли кроветворной системы, включая смешанные и эмбриональные опухоли, и лейкоз. В рамках настоящего изобретения эти опухоли являются, в частности, устойчивыми опухолями, в частности, определенным в настоящем документе.
Группа опухолей нервной системы предпочтительно включает:
1. Опухоли черепа, а также головного мозга (внутричерепные), предпочтительно, астроцитому, олигодендроглиому, менингиому, нейробластому, ганглионеврому, эпендимому, шванноглиому, нейрофиброму, гемангиобластому, липому, краниофарингиому, тератому и хордому;
2. Опухоли спинного мозга и позвоночного канала, предпочтительно, глиобластому, менингиому, нейробластому, нейрофиброму, остеосаркому, хондросаркому, гемангиосаркому, фибросаркому и множественную миелому; и
3. Опухоли периферических нервов, предпочтительно, шванноглиому, нейрофиброму, нейрофибросаркому и периневральную фибробластому.
Группа глазных опухолей предпочтительно включает:
1. Опухоли век и желез на веках, предпочтительно, аденому, аденокарциному, папиллому, гистиоцитому, опухоль тучных клеток, базальноклеточную опухоль, меланому, плоскоклеточную карциному, фиброму и фибросаркому;
2. Опухоли конъюнктивы и мигательной перепонки, предпочтительно, плоскоклеточный рак, гемангиому, гемангиосаркому, аденому, аденокарциному, фибросаркому, меланому и папиллому; и
3. Опухоли орбиты, зрительного нерва и глазного яблока, предпочтительно, ретинобластому, остеосаркому, тучноклеточную опухоль, менингиому, ретикулярно-клеточную опухоль, глиому, шванноглиому, хондрому, аденокарциному, плоскоклеточную карциному, плазматическую клеточную опухоль, лимфому, рабдомиосаркому и меланому.
Группа опухолей кожи предпочтительно включает:
опухоли гистиоцитому, липому, фибросаркому, фиброму, тучноклеточные опухоли, злокачественную меланому, папиллому, базальноклеточную опухоль, кератоакантому, гемангиоперицитому, опухоли волосяных фолликулов, опухоли потовых желез, опухоли сальных желез, гемангиому, гемангиосаркому, липому, липосаркому, злокачественную фиброзную гистиоцитому, плазмоцитому и лимфангиому.
Группа опухолей мягких тканей предпочтительно включает:
опухоли альвеолярной саркомы мягких тканей, эпителиоидноклеточную саркому, хондросаркому мягких тканей, остеосаркому мягких тканей, саркому Юинга мягких тканей, примитивные нейроэктодермальные опухоли (PNET), фибросаркому, фиброму, лейомиосаркому, лейомиому, липосаркому, злокачественную фиброзную гистиоцитому, злокачественную гемангиоперицитому, гемангиому, гемангиосаркому, злокачественную мезенхимому, злокачественную опухоль оболочки периферических нервов (MPNST), злокачественную шванноглиому, злокачественную меланоцитарную шванноглиому, рабдомиосаркому, синовиальную саркому, лимфангиому и лимфангиосаркому.
Группа опухолей желудочно-кишечного тракта предпочтительно включает:
1. Опухоли ротовой полости и языка, предпочтительно, плоскоклеточную карциному, фибросаркому, опухоль из клеток Меркеля, индуктивную фиброамелобластому, фиброму, фибросаркому, вирусный папилломатоз, идиопатический папилломатоз, полипы носоглотки, лейомиосаркому, миобластому и опухоль тучных клеток;
2. Опухоли слюнных желез, предпочтительно, аденокарциному;
3. Опухоли пищевода, предпочтительно, плоскоклеточную карциному, лейомиосаркому, фибросаркому, остеосаркому, карциному Барретта и параэзофагеальные опухоли;
4. Опухоли экзокринной части поджелудочной железы, предпочтительно, аденокарциному; и
5. Опухоли желудка, предпочтительно, аденокарциному, лейомиому, лейомиосаркому и фибросаркому.
Группа опухолей дыхательной системы предпочтительно включает:
1. Опухоли носа и носовой полости, гортани и трахеи, предпочтительно, плоскоклеточную карциному, фибросаркому, фиброму, лимфосаркому, лимфому, гемангиому, гемангиосаркому, меланому, опухоль тучных клеток, остеосаркому, хондросаркому, онкоцитому (рабдомиому), аденокарциному и миобластому; и
2. Опухоли легкого, предпочтительно, плоскоклеточную карциному, фибросаркому, фиброму, лимфосаркому, лимфому, гемангиому, гемангиосаркому, меланому, тучноклеточную опухоль, остеосаркому, хондросаркому, онкоцитому (рабдомиому), аденокарциному, миобластому, мелкоклеточную карциному, немелкоклеточную карциному, бронхиальную аденокарциному, бронхоальвеолярную аденокарциному и альвеолярную аденокарциному.
Группа опухолей костей предпочтительно включает:
остеосаркому, хондросаркому, паростальную остеосаркому, гемангиосаркому, синовиально-клеточную саркому, гемангиосаркому, фибросаркому, злокачественную мезенхимому, гигантоклеточную опухоль, остеому и многодолевую остеому.
Группа опухолей эндокринной системы предпочтительно включает:
1. Опухоли щитовидной железы/паращитовидной железы, предпочтительно, аденому и аденокарциному;
2. Опухоли надпочечников, предпочтительно, аденому, аденокарциному и феохромоцитому (медуллосупрареному);
3. Опухоли гипоталамуса/гипофиза, предпочтительно, аденому и аденокарциному;
4. Опухоли эндокринной поджелудочной железы, предпочтительно, инсулиному (бета-клеточная опухоль, APUDom) и синдром Золлингера-Эллисона (опухоль, выделяющая гастрин дельта-клетками поджелудочной железы); и
5. а также множественные эндокринные неоплазии (MEN) и хемодэктомы.
Группа опухолей женской половой системы предпочтительно включает:
1. Опухоли яичников, предпочтительно, аденому, аденокарциному, цистаденому и недифференцированную карциному;
2. Опухоли матки, предпочтительно, лейомиому, лейомиосаркому, аденому, аденокарциному, фиброму, фибросаркому и липому;
3. Опухоли шейки матки, предпочтительно, аденокарциному, аденому, лейомиосаркому и лейомиому;
4. Опухоли влагалища и вульвы, предпочтительно, лейомиому, лейомиосаркому, фибролейомиому, фиброму, фибросаркому, полипы и плоскоклеточную карциному.
Группа опухолей молочных желез предпочтительно включает:
фиброаденому, аденому, аденокарциному, мезенхимальную опухоль, карциному, карциносаркому.
Группа опухолей мужской половой системы предпочтительно включает:
1. Опухоли яичек, предпочтительно, семиному, интерстициально-клеточную опухоль и опухоль из клеток Сертоли;
2. Опухоли предстательной железы, предпочтительно, аденокарциному, недифференцированную карциному, плоскоклеточную карциному, лейомиосаркому и переходно-клеточную карциному; и
3. Опухоли полового члена и наружных половых органов, предпочтительно, опухоль тучных клеток и плоскоклеточную карциному.
Группа опухолей системы оттока мочи предпочтительно включает:
1. Опухоли почки, предпочтительно, аденокарциному, переходно-клеточную карциному (эпителиальные опухоли), фибросаркому, хондросаркому (мезенхимальные опухоли), опухоль Вильма, нефробластому и эмбриональную нефрому (эмбриональную плюрипотентную бластому);
2. Опухоли мочеточника, предпочтительно, лейомиому, лейомиосаркому, фибропапиллому, переходно-клеточную карциному;
3. Опухоли мочевого пузыря, предпочтительно, переходно-клеточную карциному, плоскоклеточную карциному, аденокарциному, ботриоидную (эмбриональную рабдомиосаркому), фиброму, фибросаркому, лейомиому, лейомиосаркому, папиллому и гемангиосаркому; и
4. Опухоли уретры, предпочтительно, переходно-клеточную карциному, плоскоклеточную карциному и лейомиосаркому.
Группа опухолей кроветворной системы предпочтительно включает:
1. Лимфому, лимфатический лейкоз, нелимфактический лейкоз, миелопролиферативный лейкоз, лимфому Ходжкина, неходжкинскую лимфому.
Группа смешанных и эмбриональных опухолей предпочтительно включает:
гемангиосаркому, тимому и мезотелиому.
Предпочтительно эти опухоли выбраны из группы, включающей рак молочной железы, карциному яичников, карциному предстательной железы, остеосаркому, глиобластому, меланому, мелкоклеточную карциному легкого и колоректальную карциному. Другие опухоли представляют собой опухоли, которые являются устойчивыми, как описано в данном документе, предпочтительно, опухоли с множественной устойчивостью, особенно опухоли из группы, описанной выше.
В рамках настоящего изобретения также идентифицируются и проверяются субъекты, которым должна быть введена комбинация по изобретению. Такая идентификация пациентов, которым может быть полезно настоящее изобретение в его разнообразных аспектах, основана на обнаружении YB-1 в ядре образца от субъекта.
В одном варианте осуществления исследование опухолевой ткани проводят с использованием агента, который выбран из группы, включающей антитела против YB-1, аптамеры против YB-1 и шпигельмеры против YB-1, а также антикалины против YB-1. В принципе, те же средства могут быть изготовлены для соответствующих маркеров и использоваться соответственно. Производство антител, в частности, моноклональных антител, известно специалистам в данной области. Дополнительным средством специфического обнаружения YB-1 или маркеров являются пептиды, которые связываются с высоким сродством с целевыми структурами, в данном случае с YB-1 или указанными маркерами. Из уровня техники известны такие методы, как фаговый дисплей, для получения таких пептидов. Обычно за отправную точку берут пептидную библиотеку, при этом отдельные пептиды имеют длину от 8 до 20 аминокислот, а размер библиотеки составляет примерно от 102 до 1018, предпочтительно, от 108 до 1015 различных пептидов. Особой формой полипептидов, связывающих молекулу-мишень, являются так называемые антикалины, которые, например, описаны в заявке на патент Германии DE 19742706.
Дополнительным средством для специфического связывания YB-1 или соответствующих маркеров, описанных в настоящем документе, и, таким образом, для обнаружения независимой от клеточного цикла локализации YB-1 в клеточном ядре, являются так называемые аптамеры, то есть D-нуклеиновые кислоты, присутствующие либо в виде РНК, либо в виде ДНК, либо в виде одной цепи, либо в виде двойной цепи и специфически связывающиеся с молекулой-мишенью. Получение аптамеров описано, например, в европейском патенте ЕР 0533838. Особой формой аптамеров являются так называемые аптазимы, которые, например, описаны Piganeau, N. et al. (2000), Angew. Chem. Int. Ed., 39, № 29, стр. 4369-4373. Это особые варианты аптамеров, поскольку они содержат, помимо аптамерной части, рибозимную часть и становятся каталитически активными при связывании или высвобождении молекулы-мишени, связывающейся с аптамерной частью и расщепляющей субстрат нуклеиновой кислоты, который сопровождается генерацией сигнала.
Другой формой аптамеров являются так называемые шпигельмеры, то есть нуклеиновые кислоты, связывающие молекулу-мишень, которые состоят из L-нуклеиновых кислот. Способ получения таких шпигельмеров описан, например, в WO 98/08856.
Образец опухолевой ткани может быть получен путем пункции или хирургического вмешательства. Оценка того, локализуется ли YB-1 в ядре независимо от клеточного цикла, часто проводится с использованием микроскопических методов и/или иммуно-гистоанализа, предпочтительно, с использованием антитела или любого из других вышеупомянутых средств. Дополнительные средства для обнаружения YB-1 в ядре и, в частности, для обнаружения того, что YB-1 находится там независимо от клеточного цикла, известны специалистам в данной области. Например, локализация YB-1 может быть легко обнаружена в окрашенных срезах ткани при их скрининге. Частота присутствия YB-1 в ядре уже указывает на то, что локализация не зависит от клеточного цикла. Другой вариант независимого от клеточного цикла обнаружения YB-1 в ядре заключается в окрашивании против YB-1 и определении того, локализуется ли YB-1 в ядре, и определении фазы клеток. Это, а также обнаружение YB-1 можно также выполнить с использованием вышеупомянутых средств, направленных против YB-1. Обнаружение средств осуществляется методами, известными специалистам в данной области. С помощью указанных агентов, специфически связывающихся с YB-1, а не с какими-либо другими структурами в анализируемом образце, в частности, с клетками, их локализацией, а также благодаря их специфическому связыванию с YB-1 также может быть обнаружена и установлена локализация YB-1 подходящей маркировкой средств. Способы маркировки указанных средств известны специалистам в данной области.
Далее настоящее изобретение будет дополнительно проиллюстрировано со ссылкой на фигуры и образцы, из которых могут быть выявлены новые особенности, варианты осуществления и преимущества.
Фиг. 1a представляет собой столбиковую диаграмму, показывающую относительную абсорбцию в качестве индикатора жизнеспособности клеток для XVir-N-31 (XVir), аденовируса дикого типа (WT) и контроля (Ctrl) при использовании в сочетании с ингибиторами CDK4/6 LY (LY-2835219), PD (PD-032991) или LEE (LEE011).
Фиг. 1b представляет собой столбиковую диаграмму, показывающую титр вируса для XVir-N-31 (XVir) и аденовируса дикого типа (WT) в сочетании с ингибиторами CDK4/6 LY (LY-2835219), PD (PD-032991) или LEE (LEE011).
Фиг. 1c представляет собой столбиковую диаграмму, показывающую сравнение волокон ДНК для XVir-N-31 (XVir) и аденовируса дикого типа (WT) в сочетании с ингибиторами CDK4/6 LY (LY-2835219), PD (PD-032991) или LEE (LEE011).
На фиг. 2 показаны результаты вестерн-блоттинга.
На фиг. 3a-d представлены столбчатые диаграммы.
На фиг. 4a-d представлены столбчатые диаграммы.
На фиг. 5 представлены столбчатые диаграммы.
Фиг. 6 представляет собой серию микрофотографий.
На фиг. 7 представлено флуоресцентное микроскопическое изображение клеток Т24, инфицированных аденовирусом с удаленным E1, экспрессирующим GFP, с обработкой палбоциклибом и без нее.
Фиг. 8 представляет собой столбиковую диаграмму, показывающую репликацию вирусной ДНК аденовируса dl703 через 48 часов с использованием соединений нутлин-3a, Lee, Cl1040 и росковитина.
На фиг. 9A-C показаны результаты вестерн-блоттинга клеток UMUC, обработанных указанными концентрациями нутлин-3a и LEE011 (рибоциклиб) (фиг.9A), росковитина (фиг.9B) и CI-1040 (фиг.9C); Rb означает белок ретинобластомы; phRB означает фосфорилированный белок ретинобластомы; E2F1 означает фактор транскрипции E2F1; и GAPDH служил контролем загрузки.
Фиг. 10 представляет собой столбиковую диаграмму, показывающую распределение клеточного цикла в клетках UMUC3, определенное через 48 часов после обработки, при этом концентрации ингибиторов CDK4/6 были следующими: росковитин: 10 мкМ, CI-1040: 1 мкМ, нутлин-3a: 10 мкМ и LEE011: 10 мкМ.
Фиг. 11 представляет собой панель микроскопических изображений, показывающих экспрессию гена гексона аденовируса с обработкой палбоциклибом и без нее.
Фиг. 12 представляет собой столбиковую диаграмму, показывающую результаты количественного определения активного вещества/компонента клеток Т24, подвергнутых воздействию только XVir-N-31, 15 нМ ингибитора PARP PARPi, 500 нМ PD (палбоциклиб) или комбинацией 15 нМ PARPi и 500 нМ PD, в виде процента выживаемости клеток, при этом клетки были либо не инфицированы (левый столбец), либо инфицированы MOI 10 (средний столбец) или MOI 50 (правый столбец).
Фиг. 13 представляет собой панель изображений, показывающих культуры клеток Т24, окрашенных SRB после обработки XVir-N-31 (20 MOI), XVir-N-31 и 15 нМ PARPi, XVir-N-31 и 500 нМ PD, и XVir-N-31, 15 нМ PARPi и 500 нМ PD, 1 дпи, 2 дпи, 3 дпи, 4 дпи, 5 дпи и 6 дпи.
Фиг. 14 представляет собой панель изображений, показывающих культуры клеток UMUC, окрашенных SRB, после обработки XVir-N-31 (10 MOI), XVir-N-31 и 160 нМ PARPi, XVir-N-31 и 400 нМ PD, и XVir-N-31, 160 нМ PARPi и 400 нМ PD, 1 дпи, 2 дпи, 3 дпи, 4 дпи, 5 дпи и 6 дпи.
Фиг. 15 представляет собой столбиковую диаграмму, показывающую результаты количественного определения активного вещества/компонента клеток Т24 через 5 дней после инфицирования XVir-N-31, ингибитором CDK 4/6 палбоциклибом и ингибитором бромодомена JQ-1; ось Y: выживаемость клеток в %.
Фиг. 16 представляет собой столбиковую диаграмму, показывающую результаты количественного определения активного вещества/компонента клеток SK-N-MC через 5 дней после воздействия только XVir-N-31, с 200 нМ абемациклибом, 500 нМ JQ-1 или комбинацией 200 нМ абемациклиба и 500 нМ JQ-1 в виде процента выживаемости клеток, при этом клетки либо не были инфицированы, либо инфицированы MOI 5, 10 или 20.
На фиг. 17 показаны результаты вестерн-блоттинга клеток SK-N-MC, обработанных указанными концентрациями ингибитора CDK 4/6 LY-2835219 (абемациклиб) и ингибитора Wee-MK-1775 (адавосертиб) через 24 и 48 часов после обработки; Rb означает белок ретинобластомы; phRB означает фосфорилированный белок ретинобластомы; E2F1 означает фактор транскрипции E2F1; и GAPDH служил контролем загрузки.
На фиг. 18 показаны результаты количественного определения активного вещества/компонента клеток SK-N-MC через 5 дней после инфицирования XVir-N-31, ингибитором CDK 4/6 абемациклибом и адавосертибом (ингибитор Wee MK-1775), выраженные в процентах живых клеток.
На фиг. 19 показано распределение клеточного цикла после обработки клеток SK-N-MC указанными ингибиторами.
Фиг. 20 представляет собой столбиковую диаграмму, показывающую влияние направленной на E2F1 миРНК на экспрессию E2F1 в различных клеточных линиях; ось Y: экспрессия E2F1, нормализованная по актину, в виде % от клеток, трансфицированных siCTRL.
Фиг. 21 представляет собой столбиковую диаграмму, показывающую, что ингибирование E2F1 вызывает повышенную экспрессию E2 на ранней стадии в клетках T24, обработанных миРНК-E2F1; ось Y: экспрессия аденовирусного гена, нормализованная по актину (в % от siCTRL).
Фиг. 22 представляет собой схему, показывающую расположение праймеров для определения экспрессии ранних E2 аденовируса.
На фиг. 23 представлена нуклеотидная последовательность E2-раннего промотора аденовируса дикого типа (вверху) и мутантного E2-раннего промотора, имеющего мутации в сайтах связывания E2F (внизу).
Фиг. 24 представляет собой столбиковую диаграмму, показывающую экспрессию РНК в инфицированных AdWT-RGD и AdE2Fm (также содержащих мотив RGD в волокне) клетках Т24, полученных с помощью RT-qPCR через 24 часа после инфицирования; экспрессия гена AD-WT была установлена на 100%.
На фиг. 25 показаны различные ингибиторы CDK4/6, подходящие для использования по настоящему изобретению.
На фиг. 26 показаны различные ингибиторы PARP, подходящие для использования по настоящему изобретению.
На фиг. 27 показаны различные ингибиторы Bet, подходящие для использования по настоящему изобретению.
На фиг. 28 показана структура WT-Ad5 и аденовируса dl520, который представляет собой онколитический аденовирус, экспрессирующий только белок E1A12 посредством делеции CR3-домена гена E1A.
На фиг. 29 показана структура XVir-N-31, которая характеризуется делецией белка E1B19K, делецией 2 т.п.н. в области E3, делецией белка E1A13S и введением мотива RGD в волоконный белок.
На фиг. 30 показана структура Ad-Delta 24 и Ad-Delta 24-RGD, которые также описаны Kleijn et al. (Kleijn et al., PLoS One. 2014; 9 (5): e97495) и характеризуются делецией CR2-домена гена E1A; они реплицируется только в опухолевых клетках с нарушенным регулированием пути ретинобластомы (Rb). Ad-Delta 24-RGD дополнительно содержит мотив RGD в утолщении волокна, как показано на XVir-N-31. Обращается внимание на то, что онколитический аденовирус dl922-947 похож на delta24, поскольку делеция в этом вирусе также находится в домене E1A-CR2 и влияет на связывание RB (белок ретинобластомы).
На фиг. 31 показана структура VCN-01, который представляет собой компетентный по репликации аденовирус, специально созданный для репликации в опухолях с дефектным путем RB, демонстрирует повышенную инфекционность за счет модифицированного волокна и улучшенное распределение за счет экспрессии растворимой гиалуронидазы (Pascual-Pasto et al. Sci Transl Med. 2019,11 476). Делеция в E1A в VCN-01 аналогична делеции в delta24 (делеция CR2-домена в E1A). Кроме того, экспрессия этого белка E1A регулируется путем введения сайтов связывания E2F в промотор E1A. Кроме того, он содержит мотив RGD в утолщении волокна и экспрессирует растворимую гиалуронидазу (Martínez-Vélez et al. 2016, Clin Cancer Res. 1; 22(9):2217-25. The Oncolytic Adenovirus VCN-01 as Therapeutic Approach Against Pediatric Osteosarcoma).
На фиг. 32 показана структура E1Adl1107 и E1Adl1101, в результате чего делеция этих двух онколитических аденовирусов влияет на связывание с p300 (гистонацетилтрансфераза p300, также известная как p300 HAT или связанный с E1A белок p300) или pRb (белок ретинобластомы) (Howe et al., MOLECULAR THERAPY 2000, 2, 485-495).
На фиг. 33 показана структура онколитического аденовируса CB016 и аденовируса 5 дикого типа (WT-Ad5), где делеция в домене E1A-CR2 аналогична Ad-Delta 24. Кроме того, CB016 содержит делецию в домене CR1. Кроме того, он содержит либо мотив RGD в волокне, либо волокно от серотипа 3 (LaRocca et al., Oral Oncol. 2016, 56, 25-31).
На фиг. 34 показана структура аденовируса ORCA-010, который содержит делецию E1AΔ24 в CR2-домене E1A, мутацию T1, повышающую эффективность в белке E3/19K, и модификацию RGD волокна, повышающую инфекционность (Dong et al., Hum Gene Ther. 1 октября 2014; 25(10): 897-904).
Фиг. 35 представляет собой столбиковую диаграмму, показывающую результаты анализа эффективности, выраженные как выживаемость клеток в %, для определения эффекта уничтожения клеток XVir-N-31 в отдельности или в комбинации с одним палбоциклибом, с одним талазопарибом или с обоими палбоциклибом и талазопарибом («комбинация ») в клетках UMUC-3 через 5 дней после заражения (5 dpi), при этом MOI XVir-N-31 составлял 10, 20 или 50.
Фиг. 36 представляет собой столбиковую диаграмму, показывающую результаты анализа эффективности, выраженные как выживаемость клеток в %, для определения эффекта уничтожения клеток XVir-N-31 в отдельности или в комбинации с одним палбоциклибом, с одним талазопарибом или с обоими палбоциклибом и талазопарибом («комбинация ») в клетках T24 через 4 дня после заражения (4 dpi), при этом MOI XVir-N-31 составлял 10, 50 или 100.
Фиг. 37 представляет собой столбиковую диаграмму, показывающую результаты анализа эффективности, выраженные в виде выживаемости клеток в %, для определения эффекта уничтожения клеток XVir-N-31 в отдельности или в комбинации с одним палбоциклибом, с одним талазопарибом или с обоими палбоциклибом и талазопарибом («комбинация») в клетках 253J через 5 дней после инфицирования (5 dpi), при этом MOI XVir-N-31 составлял 10, 20 или 50.
Фиг. 38 представляет собой столбиковую диаграмму, показывающую % клеток в фазах клеточного цикла G0/G1, S и G2 клеток T24 (слева) и клеток UMUC-3 (справа) через 48 часов после инфицирования одним талазопарибом, одним палбоциклибом или обоими талазопариб и палбоциклиб («комбинация»).
Фиг. 39 представляет собой гистограмму, показывающую процент выживших клеток в количественном определении активного вещества/компонента на клетках A673 через 4 дня после инфицирования XVir-N-31 при 5, 10 и 20 MOI, ингибиторе CDK 4/6 абемациклибе при 200 нМ и ингибиторе бромодомена JQ-1 при 200 нМ.
Фиг. 40 представляет собой панель изображений, показывающих культуры клеток Cal-33, окрашенных SRB, после обработки XVir-N-31 (1, 5 и 10 MOI), 100 нМ палбоциклиба отдельно, 100 нМ JQ-1 отдельно или в комбинации ( как палбоциклиб, так и JQ-1) через 4 дня после инфицирования.
Фиг. 41 представляет собой гистограмму, показывающую результат теста на эффективность, как показано на фиг. 40, выраженный как выживаемость клеток в % через 4 дня после инфицирования клеток Cal-33 при множественности заражения XVir-N-31, равной 5, с использованием 100 нМ палбоциклиба, 100 нм JQ-1 или комбинация палбоциклиба и JQ-1.
Фиг. 42 (A, B) представляет собой гистограммы, показывающие репликацию XVir-N-31, выраженную как «относительное волокно» в клетках Cal-33 через 24 часа (фиг. 42A) и 48 часов (фиг. 42B) после инфицирования. MOI составляла 10, концентрация палбоциклиба составляла 100 нМ, а концентрация JQ-1 была равной 100 нМ.
На фиг.43 показаны нутлин и его производные, а именно NVP-HDM201, идасанутлин, АМ-8553, SAR405838, нутлин-3а, AMG232.
Фиг. 44 представляет собой панель изображений, показывающих культуры клеток T24, окрашенных SRB, после обработки только XVir-N-31, XVir-N-1 с 30 мкМ нутлина-3a, XVir-N-31 с 500 нМ палбоциклиба и XVir-N-31 как с нутлином-3a, так и с палбоциклибом, при этом MOI XVir-N-31 составляет 0, 1, 5, 10, 20 или 30.
Фиг. 45 представляет собой столбиковую диаграмму, показывающую результат теста на активность, как показано на фиг. 44, выраженный в виде выживших клеток по отношению к контролю в %.
Фиг. 46 представляет собой панель изображений, показывающих культуры окрашенных SRB клеток T24shRb после обработки только XVir-N-31, XVir-N-1 с 30 мкМ нутлина-3a, XVir-N-31 с 500 нМ палбоциклиба и XVir- N-31 как с нутлином-3a, так и с палбоциклибом, при этом MOI XVir-N-31 составляет 0, 1, 5, 10, 20 или 30.
Фиг. 47 представляет собой столбиковую диаграмму, показывающую результат теста на активность, как показано на фиг. 46, выраженный в виде выживших клеток по отношению к контролю в %.
Фиг. 48 представляет собой панель изображений, показывающих культуры клеток T24, окрашенных SRB, после обработки только XVir-N-31, XVir-N-1 с 10 мкМ изаданутлина, XVir-N-31 с 500 нМ палбоциклиба и XVir-N-31 как с изаданутлином, так и с Палбоциклибом, при этом MOI XVir-N-31 составляет 0, 5, 10, 20, 40 и 60.
Фиг. 49 представляет собой столбиковую диаграмму, показывающую результат теста на активность, как показано на фиг. 48, выраженный как выжившие клетки по отношению к контролю в %.
Фиг. 50 представляет собой панель изображений, показывающих культуры окрашенных SRB клеток T24shRb после обработки только XVir-N-31, XVir-N-1 с 10 мкМ идасанутлином, XVir-N-31 с 500 нМ палбоциклиба и XVir-N-31 как с идасанутлином, так и с палбоциклибом, при этом MOI XVir-N-31 составляет 0, 5, 10, 20, 40 и 60.
Фиг. 51 представляет собой столбиковую диаграмму, показывающую результат теста на активность, как показано на фиг. 50, выраженный как количество выживших клеток по отношению к контролю в %.
Фиг. 52 представляет собой столбиковую диаграмму, показывающую репликацию XVir-N-31, выраженную как «относительное волокно» в клетках T24shRb (слева) и клетках T24 (справа) через 24 часа и 48 часов, соответственно. MOI составляла 20, концентрация палбоциклиба составляла 500 нМ, а концентрация нутлина-3а составляла 30 мкМ.
На рис. 53 показан результат вестерн-блоттинга.
Фиг. 54 представляет собой столбиковую диаграмму, показывающую относительное количество белка E2F1 при воздействии на клетки T24shRB палбоциклибом, нутлином-3а или их комбинацией.
Фиг. 55(A)-(D) представляют собой столбиковые диаграммы, показывающие % клеток в фазах клеточного цикла G0/G1, S и G2 клеток T24 (A), клеток T24shRb (B), клеток UMUC-3 (C) и клеток RT112. (D) при воздействии на клетки палбоциклибом, нутлином-3а или их комбинацией в указанных концентрациях.
Фиг. 56(A)-(D) представляют собой столбиковые диаграммы, показывающие % клеток в фазах клеточного цикла G0/G1, S и G2 клеток T24 (A), клеток T24shRb (B), клеток UMUC-3 (C) и клеток RT112 (D) при воздействии на клетки палбоциклибом, нутлином-3а или их комбинацией в указанных концентрациях.
Фиг. 57 представляет собой столбиковую диаграмму, показывающую относительную выживаемость клеток U87 после инфицирования XVir-N-31 100 нМ рибоциклиба (LEE, также упоминаемым как LEE011), 100 нМJQ1 и обоими 100 нМ рибоциклиба (LEE) и 100 нМJQ1, с МВД для XVir-N-31 в количестве 5.
Фиг. 58 представляет собой столбиковую диаграмму, показывающую относительную выживаемость клеток LN229 после инфицирования XVir-N-31 100 нМ рибоциклиба (LEE, также упоминаемым как LEE011), 200 нМJQ1 и обоими 100 нМ рибоциклиба (LEE) и 200 нМJQ1, с МВД для XVir-N-31 в количестве 20.
Фиг. 59 представляет собой столбиковую диаграмму, показывающую относительную выживаемость клеток T98G после инфицирования XVir-N-31 1 мкМ рибоциклиба (LEE, также упоминаемым как LEE011), 200 нМJQ1 и обоими 100 нМ рибоциклиба (LEE) и 200 нМJQ1, с МВД для XVir-N-31 в количестве 50.
Фиг. 60 представляет собой столбиковую диаграмму, показывающую относительное количество ДНК волокна аденовируса XVir-N-31 в клетках U87 при воздействии рибоциклиба (LEE) (500 нМ), JQ1 (50 нМ) или комбинации рибоциклиба (500 нМ) и JQ1. 24 (50 нМ) через 24 часа после инфицирования (hpi) XVir-N-31.
Фиг. 61 представляет собой столбиковую диаграмму, показывающую относительное количество ДНК волокна аденовируса XVir-N-31 в клетках LN229 при воздействии рибоциклиба (LEE) (500 нМ), JQ1 (100 нМ) или комбинации рибоциклиба (500 нМ) и JQ1. (100 нМ) через 48 часов после инфицирования (hpi) XVir-N-31.
Фиг. 62 представляет собой столбиковую диаграмму, показывающую относительное количество ДНК волокна аденовируса XVir-N-31 в клетках T98G при воздействии рибоциклиба (LEE) (1 мкМ), JQ1 (100 нМ) или комбинации рибоциклиба (1 мкМ) и JQ1. (100 нМ) через 48 часов после инфицирования (hpi) XVir-N-31.
На фиг.63 показан результат вестерн-блоттинга клеток LN229 через 24 часа после инфицирования XVir-N-31 (MOI 20) при воздействии 500 нМ LEE, 200 нМ JQ-1 или комбинации 500 нМ LEE и 200 нМ LEE. нм JQ-1.
На фиг. 64 показан результат вестерн-блоттинга клеток LN229 через 48 часов после инфицирования XVir-N-31 (MOI 20) при воздействии 500 нМ LEE, 200 нМ JQ-1 или комбинации 500 нМ LEE и 200 нМ LEE. нм JQ-1.
На фиг. 65 показан результат вестерн-блоттинга клеток LN229 через 72 часа после заражения XVir-N-31 (MOI 20) при воздействии 500 нМ LEE, 200 нМ JQ-1 или комбинации 500 нМ LEE и 200 нМ LEE. нм JQ-1.
На фиг. 66 показан результат вестерн-блоттинга клеток LN229 через 72 часа после инфицирования XVir-N-31 (MOI 20) при воздействии 500 нМ LEE, 200 нМ JQ-1 или комбинации 500 нМ LEE и 200 нМ LEE. нм JQ-1.
Фиг. 67 представляет собой иллюстрацию взаимодействующих партнеров E1A и расположение консервативной области CR1-CR4.
Фиг. 68 представляет собой столбиковую диаграмму, показывающую относительное увеличение ДНК аденовируса в клетках LN229 через 24 часа после инфицирования (PI) аденовирусами AdWT, dl1119, Ad delta 24, XVir-N-31 и AdWT/E2Fm (MOI 20) при JQ- 1 процедура (200 нм).
Фиг. 69 представляет собой столбиковую диаграмму, показывающую относительное увеличение ДНК аденовируса в клетках LN229 через 48 часов после инфицирования (PI) аденовирусами AdWT, dl1119, Ad delta 24, XVir-N-31 и AdWT/E2Fm (MOI 20) при JQ- 1 процедура (200 нм).
Фиг. 70 представляет собой диаграмму, показывающую количественную ДНК волокна в клетках UMUC-3 (выраженную как волокно/актин), нормализованную по отношению к волокну через 4 часа) через 24 часа после инфицирования XVir-N-31 после праймирования или одновременной обработки либо 100 нМ JQ. -1 (слева) или 500 нМ JQ-1 (справа).
Фиг. 71 представляет собой диаграмму, показывающую образование частиц XVir-N-31 (выраженное в БОЕ/мл) в клетках UMUC-3 через 39, 49, 62 или 72 часа после инфицирования (hpi) с 500 нМ JQ-1 и без него.
Фиг. 72 представляет собой панель светлопольных микрофотографий инфицированных XVir-N-31 клеток (MOI 10) после теста на титр гексонов через 39, 49, 62 или 72 часа после инфицирования (hpi) без (верхний ряд) и 500 нМ JQ-1 (нижний ряд).
Фиг. 73 представляет собой вестерн-блот-анализ клеток UMUC-3 после обработки XVir-N-31 с 500 нМ JQ-1 или без него через 12 часов после заражения, через 24 часа после заражения, через 36 часов после заражения и через 48 часов после заражения, иллюстрирующий количественная кинетика вирусной экспрессии.
Фиг. 74 представляет собой столбиковую диаграмму, показывающую процентное содержание клеток UMUC-3 (слева) и клеток RT112 (справа) в фазе G0/G1, фазе S и фазе G2 под влиянием 100 нМ JQ-1, 300 нМ JQ-1, 500 нМ палбоциклиба, комбинацию 100 нМ JQ-1 и 500 нМ палбоцилиба и комбинацию 300 нМ JQ-1 и 500 нМ палбоциклиба.
На фиг. 75 показан результат вестерн-блоттинга клеток UMUC-3 (слева) и клеток RT-112 (справа) при воздействии JQ-1 и/или палбоциклиба в дозах 0, 0,2 мкМ и 0,5 мкМ через 24 часа после обработки.
Фиг. 76 представляет собой столбиковую диаграмму, показывающую влияние XVir-N31 (MOI 5) на уничтожение клеток UMUC-3 при обработке 200 нМ JQ-1, 100 нМ палбоциклиба и комбинации 200 нМ JQ-1 и палбоциклиб в дозе 100 нМ каждые пять дней после инфицирования.
Фиг. 77 представляет собой столбиковую диаграмму, показывающую влияние XVir-N31 (MOI 40) на уничтожение клеток RT112 при обработке 200 нМ JQ-1, 300 нМ палбоциклиба и комбинации 200 нМ JQ-1 и 300 нМ палбоциклиба, каждые пять дней после заражения.
Фиг. 78 представляет собой столбиковую диаграмму, показывающую влияние XVir-N31 (MOI 40) на уничтожение клеток T24 при обработке 100 нМ JQ-1, 200 нМ палбоциклиба и комбинации 100 нМ JQ-1 и 200 нМ палбоциклиба, каждые пять дней после заражения.
Фиг. 79 представляет собой столбиковую диаграмму, показывающую репликацию XVir-N-31 в клетках UMUC-3 через 24 часа после инфицирования XVir-N-31 (MOI 10) после праймирования и одновременной обработки JQ-1 (300 нМ), палбоциклибом (100 нМ) или комбинацией JQ-1 (300 нМ) и палбоциклиба (100 нМ), при этом репликация XVir-N-31 количественно определяется как относительный уровень ДНК волокна.
Фиг. 80 представляет собой столбиковую диаграмму, показывающую репликацию XVir-N-31 в клетках T24 через 24 часа после инфицирования XVir-N-31 (MOI 50) после праймирования и одновременной обработки JQ-1 (100 нМ), палбоциклибом (200 нМ). нМ) или комбинацией JQ-1 (100 нМ) и палбоциклиба (200 нМ), при этом репликация XVir-N-31 количественно определяется как относительный уровень ДНК волокна.
Фиг. 81 представляет собой столбиковую диаграмму, показывающую репликацию XVir-N-31 в клетках RT112 через 24 часа после инфицирования XVir-N-31 (MOI 20) после праймирования и одновременной обработки JQ-1 (200 нМ), палбоциклибом (300 нМ). нМ) или комбинацией как JQ-1 (200 нМ), так и палбоциклиба (300 нМ), при этом репликация XVir-N-31 количественно оценивается как относительный уровень ДНК волокна.
Фиг. 82 представляет собой столбиковую диаграмму, иллюстрирующую среднее количественное продуцирование клеток UMUC-3, продуцирующих вирус, инфицированных XVir-N-31 (MOI 9) с 200 нМ JQ-1, с 500 нМ палбоциклиба или комбинацией обоих 200 нМ JQ-1 и 500 нМ палбоциклиба, при этом количественная оценка средней продукции указывается как количество окрашенных клеток в поле зрения (f.o.v).
Фиг. 83 представляет собой панель светлопольных микрофотографий инфицированных XVir-N-31 клеток UMUC-3 (MOI 9) после теста на титр гексонов, при этом клетки были инфицированы только XVir-N-31, комбинацией XVir-N-31 и 200 нМ JQ-1, комбинация XVir-N-31 и 500 нМ палбоциклиба или комбинацией XVir-N-31, 200 нМ JQ-1 и 500 нМ палбоциклиба.
Фиг. 84 представляет собой столбиковую диаграмму, показывающую относительную выживаемость клеток UMUC-3 после инфицирования XVir-N-31 (при множественности 0, 5 и 10) в комбинации с указанными ингибиторами BET OTX (300 нМ), AZD (5 нМ), dBet6 (50 нМ) и АРВ (50 нМ).
Фиг. 85 представляет собой столбиковую диаграмму, показывающую относительную выживаемость клеток RT112 после инфицирования XVir-N-31 (при множественности 0, 20 и 50) в комбинации с указанными ингибиторами BET OTX (130 нМ), AZD (10 нМ), dBet6 (150 нМ) и АРВ (10 нМ).
Фиг. 86 представляет собой столбиковую диаграмму, показывающую репликацию вируса в клетках UMUC-3 через 24 часа после инфицирования XVir-N-31 (MOI 10) в комбинации с указанными ингибиторами BET (OTX: 50 нМ; AZD: 50 нМ, dBet: 50 нМ и АРВ: 50 нМ).
Фиг. 87 представляет собой столбиковую диаграмму, показывающую репликацию вируса в клетках RT112 через 24 часа после инфицирования XVir-N-31 (MOI 50) в комбинации с указанными ингибиторами BET (OTX: 40 нМ; AZD: 15 нМ, dBet: 25 нМ, и ARV: 15 нМ).
Фиг. 88 представляет собой иллюстрацию исследований на животных, показывающую дозировку и запланированное применение XVir-N-31 и рибоциклиба. Рибоциклиба сукцинат (LEE011) вводили ежедневно в дозе 200 мг/кг/массы тела через пероральный зонд в течение 5 дней (от X до X+4). Растворитель без LEE011 применяли только к животным PBS и XVir-N-31. XVir-N-31 дважды вводили в опухоль в день Х+1 и Х+3. Все контрольные животные, не получавшие XVir-N-31, получали i.t. инъекции PBS, соответственно.
Фиг. 89 представляет собой диаграмму, показывающую кривые роста объема для различных групп лечения (только PBS, LEE, XVir и комбинация); каждая точка данных показывает среднее±размер опухоли в указанные дни после начала лечения.
Фиг. 90 представляет собой ящиковую диаграмму, показывающую объем опухоли [мм3] для различных групп лечения (количество животных в PBS=5), LEE (количество животных в PBS=6), только XVir (количество животных в PBS=7) и комбинированных групп. (количество животных PBS=7)) на 12-21 день после начала лечения.
Фиг. 91 представляет собой столбиковую диаграмму, показывающую вирусные геномы в опухолях (выраженные как волокна/1000 актина) у репрезентативных животных, получавших комбинированное лечение, по сравнению с лечением только XVir-N-31; оценка через 2 дня после второго i.t. введение XVir-N31.
Пример 1: Материалы и методы
Культура клеток
Клеточные линии рака мочевого пузыря человека культивировали в субконфлюэнтных условиях в среде RPMI или DMEM (Biochrom AG) при 5% или 10% CO2, соответственно, дополненной 10% FBS (Biochrom AG) и 1% NEA (Biochrom AG). В зависимости от клеточной линии и условий эксперимента 0,2-1×106, 0,5-1×105, 0,25-0,5×105 и 500-700 клеток засевали в 10 см, 6-луночном, 12-луночном и 96-луночном форматах, соответственно.
Клеточные линии
HeLaP
Клетки HeLa P (ATCC CCL-2) представляют собой эпителиальные клетки аденокарциномы шейки матки, названные в честь пациентки Генриетты Лакс. Эта клеточная линия является наиболее широко распространенной и самой старой клеточной линией (Rahbari et al., 2009), поскольку это была первая постоянная клеточная линия, созданная в 1951 году (Gey et al., 1952). Культивирование происходило в среде DMEM (10% FBS, 1% PS) в условиях 10% CO2 при 37°C.
HeLaRDB
HeLaRDB представляет собой субклеточную линию линии клеток HeLaP с устойчивостью к даунобластину на основе сверхэкспрессии гликопротеина Р. Устойчивость была достигнута путем культивирования на среде, содержащей этот антрациклин. Этот цитостатический агент внедряется в последовательности двухцепочечной ДНК и ингибирует клеточную транскрипцию и репликацию (Mizuno et al., 1975). В результате стрессовых реакций, вызванных обработкой даунобластином, клеточный фактор YB-1 показывает более высокую ядерную локализацию по сравнению с родительской клеточной линией (Holm et al., 2004). Для поддержания устойчивости к даунобластину каждые 14 дней клетки культивировали в среде DMEM (10% FBS, 1% PS), содержащей 0,25 мкг/мл даунобластина, в условиях 10% CO2 при 37°C.
A549
Клетки A549 (ATCC CCL-185) были выделены в 1972 г. из аденокарциномы базального альвеолярного отростка человека (Giard et al., 1973). Культивирование происходило в MEM Дульбекко (10% FBS и 1% PS) при 37°C и 10% CO2.
T24
Клетки T24 (ATCC HTB-4) получены в 1970 г. из первичной карциномы мочевого пузыря человека (Bubenik, Baresovà et al., 1973). Из-за точечной мутации в гене HRAS (Reddy et al., 1982) активируются пути MAPK и PI3K. Более того, в этой линии клеток присутствует дополнительная мутация в локусе гена супрессора опухолей р53 (Pinto-Leite et al., 2014). Клетки культивировали с RPMI, содержащим 10% FCS, 1% PS и 1% заменимых аминокислот, при 37°C в условиях 5% CO2.
HEK293
Клетки HEK293 (ATCC CRL-1573) представляют собой человеческие эмбриональные клетки почек, выделенные в 1973 году. Благодаря стабильной трансфекции части генома размером 4,5 т.п.н. аденовируса серотипа 5, который включает всю область E1 (Graham и Smiley, 1977), эта клеточная линия используется для производства Е1-дефицитных аденовирусов и для измерения титра вируса.
Invitrogen
Invitrogen
Metabion
Metabion
Metabion
Metabion
Metabion
Life Techno
Life Technology
Life Technology
MWG
Eurofins
Eurofins
Eurofins
Eurofins
Вирус Характеристики
Ad-WT +AdWT-RGD Мастаденовирус дикого типа, тип C, серотип 5 и ADWT с дополнительным мотивом RGD-волокна
AdWT-E2F1mut Мастаденовирус, тип C, серотип 5, мутации в обоих сайтах связывания E2F E2-раннего промотора с дополнительным мотивом RGD-волокна и делецией размером 2,7 т.п.н. в области E3 (ΔE3)
XVir-N-31 Мастаденовирус, серотип 5 с делециями в области E1B (1716-1915, 200 п.н.), области E3 (28132-30813) и делецией 12 оснований в области E1A. Реплицируется в раковых клетках, демонстрируя только ядерную экспрессию YB-1
XVir-N-31/E2F1M Мастаденовирус, тип C, серотип 5 с делециями в E1B-области (1716-1915, 200 п.н.), E3-области (28132-30813) и делецией 12 оснований в E1A-области. Реплицируется в раковых клетках, демонстрируя только ядерную экспрессию YB-1. Мутации в обоих сайтах связывания E2F E2-раннего промотора с дополнительным мотивом RGD-волокна и делецией размером 2,7 т.п.н. в области E3 (ΔE3)
Методы
Трансфекция миРНК
Подавление определенных генов осуществляли с помощью трансфекции миРНК. Таким образом, 5 мкл реагента Lipofectamin RNAiMAX (Thermo Fischer) добавляли к 150 мкл Opti-MEM в одной пробирке и 36 пмоль миРНК объединяли с 150 мкл Opti-MEM в другой пробирке. После объединения содержимого обеих пробирок и кратковременного встряхивания раствор инкубировали в течение 5 минут при комнатной температуре. Затем 250 мкл комплекса миРНК-липид добавляли к 250000-1000000 клеток, засеянных в 6-луночные планшеты в предыдущий день, без изменения среды, до достижения конечной концентрации миРНК 30 пмоль на лунку. После 48 часов инкубации при 37°C в условиях 10% CO2 имело место инфицирование или лизирование.
Количественное определение РНК в комбинации с миРНК
РНК также количественно определяли в клетках, в которых вирус объединяли с трансфекцией миРНК. При этом 125000 клеток были засеяны и на следующий день трансфицированы 30 пмоль миРНК-конструкции Ctrl-, YB-1- и E2F1-миРНК. Через 48 часов инкубации имело место инфицирование, и через 24 часа после инфицирования произошел лизис. Лизаты хранили при -20°C.
Выделение РНК
Клетки промывали PBS и лизировали лизисным буфером (набор для выделения miRNA mirVana, Life Technologies) и переносили в реакционную пробирку на 1,5 мл. К лизатам добавляли 50 мкл добавки гомогената (набор для выделения miRNA mirVana, Life Technologies), ресуспендировали и инкубировали в течение 10 минут на льду. Добавляли 500 мкл смеси кислота-фенол-хлороформ, встряхивали в течение примерно 30 секунд и инкубировали в течение 2 минут на льду. После центрифугирования в течение 5 минут при комнатной температуре при 14000 g водную и органическую фазы разделяют. Верхнюю водную фазу переносили в новую защелкивающуюся крышку, объединяли и инвертировали с равным количеством изопропанола. После инкубации в течение 10 минут при комнатной температуре образцы центрифугировали при температуре 4°C и 14000 g в течение 30 минут. Затем супернатант удаляли, а осадок РНК промывали 1 мл 75% этанола. Образцы ненадолго центрифугировали при 7500 g в течение 5 минут при температуре 4°C. После удаления супернатанта высушенный на воздухе осадок растворяли в 20 мкл воды, не содержащей нуклеазы, и инкубировали в течение 10 минут при 55°C и 500 об/мин в термомиксере. Далее определяли концентрации РНК спектрофотометрическим методом. Чтобы избежать амплификации колей ДНК, проводили расщепление ДНКазой. При этом использовалась деоксирибонуклеаза I, набор для амплификации Deoxyribonuclease I, Amplification Grade-Kit от компании Invitrogen by life technologies. К 1 мкг РНК добавляли 1 мкл 10× реакционного буфера ДНКазы I и 1 мкл ДНКазы I, заполняли водой, обработанной DEPC, до конечного объема 10 мкл и инкубировали ровно 15 минут при комнатной температуре. При добавлении 1 мкл 25 мМ раствора ЭДТА ДНКаза I инактивируется и тем самым останавливается процесс переваривания ДНКазой. Образцы инкубировали в течение 10 минут при 65°C, а затем использовали для обратной транскрипции.
Обратная транскрипция
Для перезаписи РНК в кДНК использовали набор для обратной транскрипции кДНК высокой емкости (Thermo Scientific). 2 мкг РНК расщепленных образцов ДНК добавляли к Mastermix, содержащей транскрипционный буфер, 100 мМ dNTP и ингибитор РНКазы, в мягких пробирках для ПЦР. При этом необходимо учитывать, что РНК, транскрибируемая через E2-ранний и E2-поздний промоторы, не может быть перезаписана случайными праймерами, обычно используемыми для обратной транскрипции, потому что эти случайные праймеры будут связываться с обеими цепями двухцепочечного аденовирусного генома. При этом перезапись с РНК на кДНК для образцов, используемых для количественного определения E2-раннего и E2-позднего, выполнялась с использованием специфического E2-раннего обратного праймера (табл. 1). Для гена «домашнего хозяйства» актина, который использовали для нормализации результатов, использовали случайный праймер.
Анализ репликации ДНК
Для исследования репликации вируса в инфицированных клетках был проведен анализ репликации ДНК. 125000 клеток высевали в 6-луночные планшеты и инфицировали 10-20 MOI. Через 2, соответственно, 8, 12, 24, 36 и 48 часов после инфицирования происходил лизис. При этом среду удаляли и прилипшие клетки промывали 1 мл PBS. После добавления 200 мкл буфера для лизиса ДНК прилипшие клетки отделяли от планшета с помощью скребка для клеток. Затем лизат переносили в защелкивающийся колпачок. Добавляли 3 мкл фермента протеиназы К и инкубировали при 56°C и 550 об/мин в термомиксере в течение ночи. На следующий день проводили выделение ДНК.
Выделение ДНК
Для очистки ДНК к лизату добавляли 200 мкл смеси фенол-хлороформ-изоамиловый спирт. После встряхивания и последующей инкубации на льду в течение 5 минут проводили разделение фаз центрифугированием в течение 3 минут при 16430 g при температуре 4°C. Верхнюю водную фазу переносили в новую защелкивающуюся крышку, содержащую 200 мкл хлороформа и 20 мкл крезолового красного в 10 мМ TrisCl для лучшей визуализации фаз. После встряхивания и инкубации в течение 5 минут в течение 5 минут на льду центрифугировали в течение 3 минут при 16430 g при температуре 4°C. Снова верхнюю водную фазу объединяли с 800 мкл этанола и 50 мкл 3М раствора ацетата натрия. Добавляли 2 мкл гликогена для достижения лучшего осаждения. После непродолжительного переворачивания пробирки раствор центрифугировали 30 минут при 16430 g при температуре 4°C. После этого осадок ДНК покрывали 400 мкл 70% этанола и инкубировали в течение 10 минут при комнатной температуре. После центрифугирования в течение 7 минут при 4760 g при комнатной температуре осадок ДНК сушили в течение примерно 5-10 минут при 37°C. Затем гранулы растворяли в 100 мкл 0,1×TE-буфера и встряхивали при 40°C при 400 об/мин в течение приблизительно 3 часов. Когда ДНК полностью растворялась, определяли концентрацию ДНК с помощью спектрофотометра, используя 2 мкл раствора ДНК для измерения и 0,1×TE-буфер в качестве холостого раствора. Затем ДНК хранили при температуре 4°C.
qPCR
Для дальнейшего количественного определения использовали количественную ПЦР в реальном времени. Использовали 5 мкл матричной ДНК или кДНК в конечной концентрации 10 нг/мкл. qPCR выполняли с использованием 10 мкл Mastermix GoTaq qPCR (Promega Corporation) (7,5 мкл Mastermix, 1,5 мкл праймера, 1 мкл H2O) и 5 мкл матрицы ДНК в 96-луночном планшете, пипетированных в виде дубликатов. Относительную количественную оценку выполняли с использованием метода сравнительной КТ с двумя генами-нормализаторами. Планшет закрывали фольгой и центрифугировали при комнатной температуре в течение 2 минут при 220 g. Затем планшет инкубировали в соответствии с определенной температурно-временной программой в термоциклере. Используемые праймеры перечислены в табл. 1. Реакции проводили на системе обнаружения ПЦР в реальном времени CFX96 (Bio-Rad Laboratories).
Условия циклирования qPCR
Волокно: 94°C в течение 2 минут, 94°C в течение 15 секунд, 60°C в течение 15 секунд и 72°C в течение 15 секунд, в течение 45 циклов.
Другие вирусные гены: 94°C в течение 1,5 минут, 94°C в течение 15 секунд, 58°C в течение 15 секунд и 72°C в течение 15 секунд, в течение 45 циклов.
Rb: 94°C в течение 2 минут, 94°C в течение 15 секунд, 60°C в течение 30 секунд и 72°C в течение 1 минуты, для 44 циклов
E2Fs: 95°C в течение 2 минут, 95°C в течение 15 секунд, 60°C в течение 30 секунд и 72°C в течение 30 секунд, для 40 циклов
Выделение белка
Клетки лизировали с использованием 1% SDS-буфера, чтобы добиться разрушения ядерной мембраны. Чтобы избежать денатурации белков, весь процесс проводился на льду. После отсасывания среды клетки дважды промывали холодным PBS. Прилипшие клетки одной лунки дублированного подхода лизировали 200 мкл 1% буфера SDS и соскабливали с помощью скребка для клеток. Затем лизат переносили в другую лунку дублированного подхода и снова соскребали. Затем лизат из обеих лунок переносили в пробирку с защелкивающимся колпачком. Затем лизаты обрабатывали шприцем для разрушения вязкой ДНК и центрифугировали в течение 30 минут при температуре 4°C со скоростью 31000 об/мин. Поскольку в супернатанте присутствовали белки, супернатант переносили в новую пробирку с защелкивающимся колпачком и использовали на следуюших стадиях.
Количественное определение белка
Чтобы количественно определить долю белка, был проведен анализ бицинхониновой кислоты (BCA) с помощью набора Pierce TM BCA Protein Kit. При этом 112,5 мкл раствора BCA A+B (50:1) и 12,5 мкл образца добавляли в одну лунку 96-луночного планшета и инкубировали в течение 30 минут при температуре 37°C. В зависимости от концентрации белка происходило окрашивание раствора. С помощью стандартной серии с известными концентрациями белка фотометрическим измерением при 562 нм в считывающем устройстве для микропланшетов определяли концентрации белка в образцах.
SDS-гель-электрофорез
Для разделения белков при последующем электрофорезе в полиакриламидном геле с додецилсульфатом натрия рассчитанные количества лизата и лизисного буфера смешивали с 15 мкл смеси загрузочный буфер-DDT (6:1). Затем вещества, нагружающие белок, готовили в течение пяти минут при температуре 100°C. Затем в гель загружали 5 мкл стандарта окрашенного белка и 40 мкл образцов. Для разделения белков с обнаружением вирусных белков использовали 10% гель. Для разделения белков с обнаружением вирусных белков использовали 10% гель. Для изучения генов с пониженной регуляцией через миРНК использовали 12% гели. Состав разделяющего и концентрирующего гелей указан в разделе «Буферы и растворы». В течение примерно 20 минут гель работал в TGS-буфере при 90 В для концентрации всех белков в одной полосе. Затем гели работали в течение приблизительно 60 минут при 150 В буфере TGS для разделения белков по размеру.
Вестерн-блоттинг
Для переноса белков из геля на мембрану ее подвергали блоттингу методом вестерн-блоттинга. Чтобы активировать гидрофобную PVDF-мембрану ее инкубировали в течение примерно 2 минут в метаноле. Затем мембрану вместе с губчатыми веществами, фильтровальной бумагой и гелем осаждали в буфере для блоттинга. Посредством электрофореза в течение приблизительно двух часов при 100 В при температуре 4°С белки переносили на мембрану в буфере для блоттинга. Чтобы избежать неспецифического связывания антител мембрану блокировали вращением в течение одного часа при комнатной температуре в 10 мл 5% сухого молока в TBST для анализа клеточных белков, соответственно, в 5 мл 5% BSA-TBST для последующего использования антител, обнаруживающих вирусные белки. После пятикратной промывки мембраны в TBST в течение пяти минут каждую мембрану инкубировали с раствором первичных антител при температуре 4°С, вращая в течение ночи. Для антител GAPDH, E1A, E1B55K, E2A и E4orf6 эту стадию осуществляли в течение одного часа при комнатной температуре. При этом антитела разбавляли различными факторами в 5% BSA в TBST с 0,02% азидом натрия. После дополнительных пяти стадий промывки мембрану инкубировали при вращении в течение 30 минут при комнатной температуре при разведении вторичного антитела 1:10000. Вторичные антитела (антимышиные) к вирусным антителам разводили в 5% BSA-TBST, все остальные в 5% сухом молоке в TBST. Эти вторичные антитела были конъюгированы с пероксидазой хрена. После пяти заключительных стадий промывки мембрану инкубировали пять минут в растворе с усиленной хемилюминесценцией (ECL) для визуализации сигнала пероксидазы. Для мембран, инкубированных с первичными антителами DP-1 и E2F1, использовался реагент Amersham ECL Prime Western Blotting Detection Reagent от GE-Healthcare для достижения более ярких сигналов, для всех остальных использовались растворы ECL, полученные в лаборатории. Состав ECL A и ECL B, которые смешивали незадолго до использования в соотношении 1:1, перечислены в разделе «Буферы и растворы». Наконец, белки могут быть обнаружены посредством проявления сигнала на пленке.
Антитела:
Киназа контрольной точки 1 (sc-377231, Santa Cruz Biotechnology)
общий RB (554136, BD Biosciences)
фосфо RB Ser 780 (8180, Cell Signaling Technology)
E2F1 (sc-251, Santa Cruz Biotechnology)
E2F2 (ab138515, abcam)
E2F3 (PG37, Thermo Fisher Scientific)
E2F4 (WUF10, Thermo Fisher Scientific)
E2F5 (sc-999, Santa Cruz Biotechnology)
циклин D1 (92G2, Cell Signaling Technology)
циклин E2 (4132, Cell Signaling Technology)
CDK2 (78B2, Cell Signaling Technology)
GAPDH (14C10, Cell Signaling Technology)
актин (A2066, Sigma-Aldrich Chemie GmbH)
E1A (sc-25, Santa Cruz Biotechnology)
E1B55k (любезно предоставлено компанией M. Dobbelstein)
E4orf6 (любезно предоставлено компанией M. Dobbelstein
E2A (DBP, любезно предоставлено компанией M. Dobbelstein)
Гексон (ABIN2686029, Antibodies online)
Обработка низкомолекулярными ингибиторами
Изетионат PD-0332991 (палбоциклиб, Sigma-Aldrich Chemie GmbH) и LY-2835219 (абемациклиб, Selleck Chemicals) растворяли в стерильной воде в виде 10 мМ исходного раствора. LEE011 (рибоциклиб, MedChem Express) и нутлин-3a (Sigma) растворяли в ДМСО в виде 10 мМ и 5 мкМ маточного раствора, соответственно. Рабочие концентрации были приготовлены для немедленного использования.
Инфицирование вирусом и комбинированная обработка
Для определения индуцированного вирусом уничтожения клеток клетки высевали в 12-луночные планшеты. Для комбинированной обработки PD-033299, LY-2835219 и LEE011 клетки предварительно обрабатывали ингибиторами в течение 24 часов. Клетки инфицировали указанными вирусами при указанной MOI в 200-400 мкл среды без FBS. При 1 hpi к клеткам добавляли полную среду с низкомолекулярными ингибиторами или без них.
Жизнеспособность клеток (анализ SRB)
Клетки фиксировали 10% TCA в течение 1 часа при температуре 4°С и окрашивали 0,5% сульфородамином B (SRB, Sigma-Aldrich Chemie GmbH) в 1% уксусной кислоте в течение 30 минут при комнатной температуре с последующей промывкой 1% уксусной кислотой чтобы удалить излишки СРБ. Высушенный SRB растворяли в 10 мМ Трис-буфере, и количественное определение выполняли фотометрическим измерением при 590 нм.
Титровальный тест
Для определения продукции инфекционных вирусных частиц инфицированные клетки и супернатант собирали в трех точках на дюйм с использованием скребков для клеток. Вирус высвобождался из интактных клеток с помощью нескольких циклов замораживания-оттаивания с последующим центрифугированием при 1600 rcf (относительное ускорение центрифуги). Супернатанты клеточных лизатов тестировали на продуцирование вирусных частиц с использованием клеток Hek293, как описано в руководстве по набору AdEasy Viral Titer Kit (972500). Использовали следующие реагенты: козье анти-гексон антитело (1056, Chemicon), кроличье антикозье антитело (P0449, Dako), раствор DAB (Dako).
Пример 2: Влияние ингибитора CDK4/6 PD0332991 на репликацию E1-минус аденовируса
Было показано, что аденовирус с удаленным E1 реплицируется в раковых клетках, хотя и с очень низкой эффективностью. Клетки Т24 инфицировали 100 MOI Е1-минус аденовируса, экспрессирующего зеленый флуоресцентный белок (Ad-GFP), и обрабатывали 500 нМ PD0332991 за день до инфицирования и во время инкубации. В таких условиях наблюдалось увеличение экспрессии GFP, что указывает на E1A-независимую вирусную репликацию и экспрессию гена, опосредованную активацией E2-раннего промотора аденовируса.
Пример 3: Комбинированное применение аденовируса дикого типа или XVir-N-31 вместе с различными ингибиторами CDK4/6
На основании результатов с использованием аденовируса Ad-WT/E2M с ранней мутацией E2 и Ad-GFP в комбинации с PD0332991, были проведены эксперименты с использованием различных ингибиторов CDK4/6 в комбинации с аденовирусом Ad-WT дикого типа или XVir-N-31. Поскольку эти агенты делают арест клеток в фазе G1, было неожиданным обнаружить, что все ингибиторы способны поддерживать репликацию вируса.
Далее было исследовано, может ли обработка клеток тремя клинически продвинутыми ингибиторами CDK4/6 PD-033299, LY-2835219 и LEE011 влиять на воздействие инфицирования на жизнеспособность клеток, репликацию вируса и продукцию титра вируса.
После обработки все три ингибитора проявляли сходные эффекты на уровень экспрессии и фосфорилирования RB, которые были описаны ранее в многочисленных публикациях. После почти полного дефосфорилирования, а также подавления общего белка через 24 часа, уровень фосфорилирования частично восстанавливался с течением времени. Уровень CDK2 повышался после обработки, а уровень циклина D2, а также циклина E2 снижался.
Пример 4: Синергетические эффекты, возникающие при комбинации ингибиторов CDK4/6 и онколитических аденовирусов
Ингибиторы CDK4/6 PD-033299, LY-2835219 и LEE011 были объединены с инфицированием клеток аденовирусом. Инфицирование клеток происходило через 24 часа после обработки, поскольку последующее воздействие на молекулы-мишени можно обнаружить только между 8 и 24 часами после обработки.
Результаты показаны на фиг. 1.
Ингибиторы CDK4/6 индуцировали синергетический эффект на жизнеспособность клеток, репликацию вируса и титр вируса. (a) Клетки предварительно обрабатывали тремя ингибиторами CDK4/6 PD-033299, LY-2835219 и LEE011 в течение 24 часов и инфицировали XVir-N-31 (Moi 60) или аденовирусом дикого типа (Moi 80). Через четыре дня после инфицирования жизнеспособность клеток измеряли с помощью анализа SRB. На графиках показаны средние значения минимум трех независимых экспериментов. (b) Через три дня после инфицирования из клеток готовили лизаты и проводили титровальный тест на клетках HEK293. Титр вируса показан как кратное изменение относительно контроля. (c) ДНК экстрагировали из инфицированных клеток через 4, 24, 36 и 48 hpi и анализировали на вирусную репликацию с помощью кПЦР для кДНК волокна. Значения нормализованы к GAPDH при 4 hpi. На графиках представлены как минимум два независимых эксперимента. Планки погрешностей представляют собой стандартную ошибку.
Как видно из фиг. 1, все три ингибитора CDK4/6 наглядно поддерживали лизис клеток (фиг. 1a), репликацию внутри клеток (фиг. 18) и образование вирусных частиц (фиг. 1b).
Пример 5: Влияние ингибитора CDK4/6 палбоциклиба (PD-033299) на уровень экспрессии выбранных вирусных белков
Чтобы проанализировать эти эффекты более подробно, уровень экспрессии выбранных вирусных белков определяли в обработанных или необработанных клетках. Для этого эксперимента в качестве типичного ингибитора CDK4/6 использовали ингибитор палбоциклиб (PD-033299). Клетки инфицировали с MOI, равным 15. Обработка PD 500 нМ происходила в течение 24 часов и до тех пор, пока не происходило выделение белка. Через 12, 24 и 36 часов осуществляли выделение белка с использованием 1% SDS-буфера. Актин был включен в качестве положительного контроля. Поскольку контроль загрузки показывает одинаковые уровни белка клеточного актина во всех линиях, обеспечивается надлежащее сравнение между линиями. hpi: часы после инфицирования.
Результаты показаны на фиг. 2, где представлены результаты экспрессии вирусного белка клеток Т24, инфицированных Ad-WT и XVir-N-31, в комбинации с ингибитором CDK4/6 PD0332991 (PD). Все вирусные белки, исследованные в этом эксперименте (E1A, E1B-55k, DBP (E2A) и гексон), были экспрессированы на более высоком уровне в клетках, обработанных ингибитором CDK4/6 PD-0332991, по сравнению с вирусом аденовируса дикого типа. Этот эффект можно было наблюдать уже через 12 hpi для E1A и 24 hpi для других белков.
Пример 6: Специфичность эффектов, опосредованных ингибиторами CDK4/6
Класс ингибиторов CDK4/6 согласно примеру 5 требует экспрессии RB. Поэтому использовали три RB-положительные и две RB-отрицательные клеточные линии, происходящие от рака мочевого пузыря, и клетки, обработанные комбинированными препаратами. Клеточные линии предварительно обрабатывали в течение 24 часов PD-0332991 (T24: 500 нМ, RT112: 2000 нМ, 253J: 100 нМ) с концентрацией IC50 и инфицировали XVir-N-31(T24 MOI50, 253J MOI 25, RT112 MOI450). Значения представляют собой среднее значение минимум 2 независимых экспериментов. Планки погрешностей показывают стандартную ошибку. Четыре точки на дюйм, жизнеспособность клеток измеряли с помощью тестов SRB (a, c). (b, d) Лизаты клеток готовили с разрешением 3 точки на дюйм, и титровальный тест проводили на клетках Hek293. Титр вируса показан как кратное изменение относительно контроля.
Результаты показаны на фиг. 3.
Как видно из фиг. 3, только линии клеток, положительные по RB, показали значительное снижение роста и жизнеспособности клеток, соответственно (фиг. 3 a, c). Кроме того, образование вирусных частиц увеличивалось только в RB-положительных клеточных линиях после обработки PD-0332991 (фиг. 3b, d).
Пример 7: Эффект комбинированной обработки ингибитора CDK4/6 PD-0332991 с XVir-N-31
Чтобы исследовать влияние PD-0332991 на репликацию вирусов в RB-положительных клеточных линиях, проводили относительную количественную оценку копий ДНК волокна с помощью кПЦР. Клеточные линии рака мочевого пузыря предварительно обрабатывали в течение 24 часов и инфицировали XVir-N-31 (T24 MOI 40, UMUC3 и 253J MOI 20, RT112 MOI 400). ДНК экстрагировали 24-48 hpi и анализировали на вирусные волокна с помощью КПЦР. Значения нормализовали к GAPDH. Данные являются характерными как минимум для двух независимых экспериментов; планки погрешностей S.D.
Результаты показаны на фиг. 4.
Как можно понять из фиг. 4, комбинированная обработка ингибитором CDK4/6 PD-0332991 с XVir-N-31 резко увеличивает репликацию вируса.
Пример 8: Временная кинетика ингибиторов CDK4/6
Временная кинетика ингибиторов CDK4/6 на дефосфорилирование и деградацию RB составляет около 10 часов после обработки клеток. Кроме того, представленные выше результаты показали частичное восстановление целей RB в направлении 3ʼ с течением времени (фиг. 1). Это наблюдение подразумевает, что временная кинетика ингибитора CDK4/6 и его влияние на вызванную вирусом гибель клеток являются важным параметром для этой комбинированной терапии, как показано в примере 7. Для применения комбинированной терапии были протестированы различные временные точки для предварительной обработки клеток. В соответствии с этим клетки обрабатывали либо до (день/час до инфицирования, день/час), либо через 1 час после инфицирования, и рост клеток определяли с использованием анализа SRB. Планки погрешностей представляют S.E., а значения являются средним значением трех независимых экспериментов.
Результаты показаны на фиг. 5.
Как видно из фиг. 5, для увеличения гибели клеток уже было достаточно параллельной обработки.
Пример 9: Комбинированная обработка различных аденовирусов ингибитором CDK4/6 PD0332991
Этот пример был выполнен, чтобы предоставить экспериментальные доказательства того, что для уничтожения могут использоваться клеток различные онколитические аденовирусы вместе с ингибиторами CDK4/6, такими как PD0332991, и что наблюдаемое увеличение репликации вирусов и уничтожение клеток не ограничивалось XVir-N-31. В соответствии с этим раковые клетки Т24 с Ad-Delta24 и Onyx-015 были следующими: клетки рака мочевого пузыря Т24 были инфицированы 20 MOI указанных онколитических аденовирусов. Обработка 500 нМ ингибитора CDK4/6 PD0332991 проводилась за один день до инфицирования и в течение 4 дней после инфицирования. Фотографии были сделаны через 4 дня после инфицирования. Возникновение цитопатического эффекта (ЦПЭ) указывает на репликацию вируса и гибель клеток.
Результаты показаны на фиг. 6.
Как видно из фиг. 6, ингибитор CDK4/6 PD0332991 в качестве характерного примера ингибиторов CDK4/6, снижающих фосфорилирование RB, увеличивает гибель клеток в сочетании с другими онколитическими аденовирусами, такими как Ad-Delta24 и Onyx-015.
Пример 10: Инфицирование клеток Т24 рекомбинантным аденовирусом с удаленным E1, экспрессирующим GFP (Ad-минус/GFP), в комбинации с палбоциклибом вызывает повышение экспрессии GFP
100000 клеток Т24 на лунку высевали в 6-луночные планшеты и выращивали в среде RPMI, содержащей 10% FCS при 5% CO2 при 37°C. Клетки Т24 обрабатывали 500 нМ палбоциклиба за 24 часа до и опять через 1 час после инфицирования. Инфицирование аденовирусом с делецией E1, экспрессирующим GFP (Ad-минус/GFP), происходило в 400 мкл среды без сыворотки. Снимки были сделаны через 48 часов после инфицирования с использованием флуоресцентного микроскопа с 10-кратным увеличением.
Результат флуоресцентного микроскопического анализа экспрессии GFP с обработкой палбоциклибом и без нее показан на фиг. 7.
Результат показывает, что обработка клеток Т24 палбоциклибом вызвала сильное увеличение экспрессии GFP, которое опосредовано репликацией вирусной ДНК, индуцированной палбоциклибом.
Пример 11: E1A-независимая репликация вируса в клетках UMUC, обработанных различными ингибиторами клеточного цикла
Чтобы исследовать различия в репликации dl703 (Mantwill et al. 2013, Journal of Translational Medicine, 11, 216) при различных условиях обработки был проведен анализ репликации ДНК. 100000 клеток UMUC высевали в 6-луночные планшеты и выращивали в среде DMEM, содержащей 10% FCS в условиях 5% CO2 при 37°C. Через 24 часа после посева клетки обрабатывали в течение 24 часов 10 мкМ Lee (рибоциклиб), 1 мкМ CI-1040, 10 мкМ нутлин-3a и 10 мкМ росковитин, а затем снова после инфицирования добавляли в среду соответствующее количество ингибиторов. Инфицирование 50 MOI dl703 (мастаденовирус, тип C, серотип 5 с делецией размером 3,2 т.п.н. в области E1) происходило через 24 часа после обработки. Через 4 и 48 часов после инфицирования выделяли ДНК и проводили qPCR с использованием специфических праймеров для гена вирусного волокна. Волокно fwd. 5'-AAGCTAGCCCTGCAAACATCA-3' (SEQ ID NO: 17); волокно rev. 5'-CCCAAGCTACCAGTGGCAGTA-3' (SEQ ID NO: 18).
Результаты показаны на фиг. 8.
Как видно из фиг. 8, обработка клеток UMUC ингибитором CDK 4/6 LEE011 (рибоциклиб) вызывала резкое увеличение репликации вирусной ДНК E1-минус аденовируса dl703 (почти в 100 раз). Это увеличение убедительно свидетельствует о том, что специфически индуцированный G1-арест рибоциклибом в сочетании с ингибированием экспрессии E2F1 способствует независимой от E1 репликации аденовирусов. Как следствие, не только вирусы со специфическими делециями в гене E1A демонстрируют усиленную репликацию аденовирусной ДНК при обработке CDK 4/6, но даже аденовирусы с полной делецией гена E1A демонстрируют усиление репликации вирусной ДНК.
Хотя Mek-ингибитор GI-1040 продемонстрировал аналогичные свойства в отношении ингибирования экспрессии E2F1 и ареста G1, репликация была намного ниже по сравнению с клетками, обработанными рибоциклибом. Это может быть связано с тем, что одновременно ингибируются другие важные пути, связанные с клеточным циклом, такие как MEK/ERK, который необходим для репликации вируса. Кроме того, было показано, что ингибирование пути MEK/ERK снижает образование частиц более чем в 100 раз, что делает его непригодным в клинических условиях для комбинированной терапии с репликацией онколитического аденовируса (Schümann и Doppelstein 2016, Cancer Research, 66, 1282-1288).
Пример 12: Вестерн-блот-анализ клеток UMUC, обработанных указанными ингибиторами клеточного цикла
Вестерн-блоттинг клеток UMUC, обработанных указанными концентрациями CI-1040, росковитина, нутлина-3а и LEE011 (рибоциклиб). 1×106 клеток высевали в чашки диаметром 10 см. Через 24 часа после обработки белки выделяли с использованием 1% SDS-буфера, чтобы добиться разрушения ядерной мембраны. Все образцы набирали несколько раз в шприц для разрушения ДНК и затем центрифугировали при 30000 об/мин при температуре 4°C в течение 30 минут. Супернатант переносили в новую реакционную пробирку и непосредственно использовали для дальнейших стадий или хранили при температуре -80°C. Для разделения белков проводили электрофорез в полиакриламидном геле с додецилсульфатом натрия. Посредством электрофореза в течение приблизительно двух часов при 100 В при температуре 4°C загружали 40 мкг общих белков и зондировали их против специфических указанных антител.
Результаты показаны на фиг. 9A, 9B и 9C.
Как видно из фиг. 9, в то время как росковитин и нутлин-3a не оказывали выраженного эффекта на экспрессию Rb, phRB и E2F1, LEE-011 (рибоциклиб) при 10 мкМ и MI-1040 при 1 мкМ индуцировали ингибирование E2F-1, а также экспрессию Rb и phRb.
Пример 13: Анализ ингибиторов CDK 4/6 на репликацию вирусной ДНК аденовируса dl703, дефектного по репликации с удаленной E1
Для анализа клеточного цикла клетки высевали в 6-луночные планшеты (2,5×10E4 с/лунку). За 8 часов до инфицирования dl703 клетки обрабатывали указанной концентрацией ингибиторов клеточного цикла. После инфицирования 10 MOI клетки dl703 снова обрабатывали в течение 48 часов. Контролем служили необработанные клетки и клетки, инфицированные только dl703. Через 48 часов после инфицирования клетки собирали трипсинизацией и фиксировали 80% этанолом при встряхивании. Чтобы исследовать статус клеточного цикла, фиксированные клетки центрифугировали 5 мин при комнатной температуре и 300 g и аспирировали этанол. Клетки ресуспендировали и промывали 1% BSA-PBS (бычий сывороточный альбумин) и снова центрифугировали. Клетки окрашивали EDU и анализ клеточного цикла выполняли с использованием наборов для проточной цитометрии Click-iT™ Plus EdU, каталожные номера C10632 от компании Thermo Fischer. Кроме того, после трехкратной промывки 1% BSA/PBS клетки окрашивали PI (пропидиум йод, 50 мкг/мл). Измерение проводили непосредственно после окрашивания с помощью системы проточной цитометрии FACScalibur. Данные были проанализированы с помощью программного обеспечения FlowJo.
Характеристики ингибиторов CDK4/6
CI1040: двойная специфическая треонин/тирозинкиназа, киназа митоген-активированная протеинкиназы (MEK), является ключевым компонентом пути передачи сигналов RAS/RAF/MEK/ERK, который часто активируется в опухолях человека. CI-1040 представляет собой бензгидроксаматное соединение, которое сильно ингибирует MEK (Allen et al. 2003, Semin Oncol. (5 Suppl 16):105-16) и вызывает арест G1.
Нутлин-3a: Нутлин-3, низкомолекулярный антагонист MDM2, эффективно восстанавливает функцию p53 как в нормальной экспрессии MDM2, так и в линиях клеток со сверхэкспрессией MDM2 с p53 дикого типа, что приводит к аресту клеточного цикла и апоптозу (Wang et al 2012, Acta Biochimica et Biophysica Sinica, том 44, пункт 8, 1 августа 2012, стр. 685-691).
Росковитин (селициклиб или CYC202) является экспериментальным лекарственным средством-кандидатом в семействе фармакологических ингибиторов циклин-зависимой киназы (CDK), которые предпочтительно ингибируют несколько ферментных мишеней, включая CDK2, CDK7 и CDK9, которые изменяют фазу роста или состояние в пределах клеточного цикла обработанных клеток (Whitaker et al. 2004, Cancer Research 64, 262-272).
LEE011 (рибоциклиб; торговое название Kisqali]) является ингибитором циклина D1/CDK4 и CDK6 и используется для лечения некоторых видов рака молочной железы. Ингибирование CDK 4/6 вызывает арест клеточного цикла G1 и ингибирование экспрессии E2F1 (Kim S. et al, Oncotarget. 16 октября 2018; 9(81):35226-35240; Yang C et al., Oncogene (2017) 36, 2255-2264).
Результаты показаны на фиг. 10.
Ингибиторы CDK 4/6 LEE011 (рибоциклиб) и CI-1040 вызывали явный арест G1. Обработка росковитином показала небольшое увеличение количества арестованных клеток G2/m. Нутлин-3a в используемой концентрации оказывал незначительное влияние на клеточный цикл или не оказывал никакого влияния на него. Инфицирование клеток UMUC рекомбинантным E1-удаленным (не имеющим белка E1A) аденовирусом dl703 существенно не меняло распределение клеточного цикла.
Пример 14: Палбоциклиб увеличивал окрашивание гексоном аденовируса in vitro после обработки
Линии клеток мочевого пузыря RT112, T24 и UMUC высевали в 6-луночные планшеты (2×105 клеток/лунку). Через сутки после посева клетки обрабатывали 500 нМ палбоциклиба в течение 24 часов до и опять через 1 час после инфицирования. Инфицирование указанными MOI AD-WT происходило в 400 мкл среды DMEM, не содержащей сыворотки. Окрашивание гексоном проводили в соответствии с инструкциями производителя с использованием титровального набора Adeasy Viral Titer Kit от компании Agilent (номер по каталогу 972500) через два дня после инфицирования.
Результаты показаны на фиг. 11.
Как видно из фиг. 11, обработка палбоциклибом (500 нМ) в качестве иллюстративного ингибитора CDK4/6 значительно увеличивала количество гексон-положительных клеток в клетках, обработанных палбоциклибом, через 48 часов после инфицирования, что показано коричневым/красным цветом. Следует сделать вывод, что большее количество клеток, подвергшихся обработке палбоциклибом, способно продуцировать вирусные частицы и показывать усиление репликации вирусной ДНК, поскольку экспрессия гексона аденовируса происходит исключительно в начале репликации вируса.
Из результатов, приведенных в примерах 10-14, очевидно, что только ингибиторы CDK4/6, но никакие другие ингибиторы клеточного цикла не способны увеличивать репликацию и экспрессию генов дефектного по репликации аденовируса (dl703, не содержащего генов E1) и Ad-GFP. Кроме того, ингибитор CDK4/6 для обеспечения такой повышенной репликации вируса и экспрессии гена должен вызывать арест G1 (инфицированных) клеток и ингибирование экспрессии F2F1.
Пример 15: Обработка клеток Т24 с использованием тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP
Чтобы продемонстрировать эффективность тройной терапии клеток T24 с использованием тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP (BMN673 (талазолариб)), был проведено количественное определение активного вещества/компонента.
12500 клеток Т24 высевали на лунку в 12-луночные планшеты и выращивали в течение ночи в среде RPMI, содержащей 10% FCS, при температуре 37°C. Обработка клеток ингибитором происходила через 24 часа после посева клеток и опять через 1 час после инфицирования путем добавления указанной концентрации в среду. Инфицирование клеток происходило через 24 часа после обработки ингибитором в 250 мкл среды без сыворотки. Фиксацию и окрашивание SRB проводили через 4 дня после инфицирования. PD, палбоциклиб; PARPi: BMN673.
Для окрашивания SRB среду удаляли аспирацией. Клетки фиксировали 1 мл (на лунку) 10% холодной TCA при температуре 4°C в течение 1 часа. TCA удаляли путем аспирации, а слои клеток промывали 4 раза водопроводной водой. Клетки окрашивали 1 мл (на лунку) 0,5% SRB (сульфородамин B) в 1% уксусной кислоте в течение 30 мин. Несвязанный SRB удаляли за пять стадий промывки 1 мл 1% уксусной кислоты на лунку; после каждой стадии промывки уксусную кислоту удаляли аспирацией. Планшеты сушили на воздухе в течение 2 часов. Для солюбилизации окрашенных SRB клеток в каждую лунку добавляли 200 мкл 10 мМ трис-основания. После этого 20 мкл соответственно распределяли в лунки 96-луночного планшета. 96-луночный планшет загружали в устройство для чтения планшетов для ELISA и измеряли поглощение образцов при 560 нм. Макетные обработанные клетки имели 100% выживаемость клеток.
Результаты показаны на фиг. 12.
Результаты, показанные на фиг. 12, ясно демонстрируют, что тройная терапия, состоящая из палбоциклиба, BMN673 и XVir-N-31, показала превосходные характеристики по сравнению с моно- или комбинированной терапией в отношении уничтожения клеток. Почти 90% уничтожения клеток может быть достигнуто с использованием 10 MOI XVir-N-31 в сочетании с ингибитором PARP PARPi (BMN673) и ингибитором CDK4/6 палбоциклибом (PD). Комбинация PARPi и палбоциклиба, не содержащая XVir-N-31, убила только 65% клеток. Клетки T24 и клетки UMUC чувствительны к CDK 4/6-ингибиторам (обеспечивая арест G1 с подавлением E2F-1).
Пример 16: Кинетика тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP
Чтобы продемонстрировать кинетику тройной терапии клеток Т24 с использованием тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP (BMN673 (талазолариб)), был проведено количественное определение активного вещества/компонента , и эффективность оценивалась в разные моменты времени.
3000 клеток Т24 высевали на лунку в 12-луночные планшеты и выращивали в течение ночи в среде RPMI, содержащей 10% FCS, при температуре 37°C. Обработка клеток ингибитором происходила через 24 часа после посева клеток и опять через 1 час после инфицирования путем добавления указанной концентрации в среду. Инфицирование клеток происходило через 24 часа после обработки ингибитором в 250 мкл среды без сыворотки. Фиксацию и окрашивание SRB проводили через 1-5 дней после инфицирования (dpi: дни после инфицирования). 15 нМ PARPi соответствуют значению IC-80 в клетках Т24.
Результаты показаны на фиг. 13.
Как видно из фиг. 13, тройная терапия с использованием помимо XVir-N-31 ингибитора CDK4/6 (палбоциклиба (PD)) и PARP ингибитора PARPI (BMN673) также с кинетической точки зрения также намного более эффективна, чем монотерапия с использованием только XVir-N-31 или комбинированная терапия с использованием XVir-N-31 либо ингибитора PARP, либо ингибитора CDK4/6. Важно отметить, что повторный рост опухолевых клеток был значительно снижен на 4 и 5 день в линиях клеток, чувствительных к CDK 4/6, UMUC и T24 (dpi: дни после инфицирования).
Пример 17: Кинетика тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP
Чтобы продемонстрировать кинетику тройной терапии клеток UMUC с использованием тройной терапии, включающей XVir-N-31, палбоциклиб и ингибитор PARP (BMN673 (талазолариб)), был проведен анализ эффективности, и эффективность оценивалась в разные моменты времени.
Посев UMUC-3: 3000 клеток высевали на лунку в 12-луночные планшеты и выращивали в течение ночи в среде DMEM, содержащей 10% FCS, при температуре 37°C. Обработка клеток ингибитором происходила через 24 часа после посева и опять через 1 час после инфицирования путем добавления указанной концентрации в среду. Инфицирование клеток происходило через 24 часа после обработки ингибитором. Фиксацию и окрашивание SRB проводили через 1-6 дней после инфицирования (dpi: дни после инфицирования). 160 нМ PARPi соответствуют значению IC-80 в ячейках UMUC3.
Результаты показаны на фиг. 14.
Результаты, показанные на фиг. 14, ясно демонстрируют, что тройная терапия, состоящая из палбоциклиба, BMN673 и XVir-N-31, показала превосходные характеристики по сравнению с моно- или комбинированной терапией. Важно отметить, что повторный рост опухолевых клеток был значительно снижен на 4 и 5 день в линиях клеток, чувствительных к CDK 4/6, UMUC и T24 (dpi: дни после инфицирования).
Пример 18: Тройная терапия, включающая XVir-N-31, ингибитор CDK4/6 и ингибитор бромодомена
5000 клеток Т24 высевали в 12-луночные планшеты и выращивали в 1 мл среды RPMI, содержащей 10% FCS. На следующий день клетки обрабатывали 500 нМ палбоциклиба и 300 нМ JQ-1. Через 24 часа после обработки клетки инфицировали указанными MOI XVir-N-31 в 200 мкл среды RPMI, не содержащей FCS. Через 1 час в каждую лунку добавляли 800 мкл среды RPMI, содержащей 10% FCS. Кроме того, в среду добавляли 500 нМ палбоциклиба и 300 нМ JQ-1. SRB-окрашивание проводили через 5 дней после инфицирования. Макетные обработанные клетки имели 100% выживаемость клеток.
Результаты показаны на фиг. 15.
Как видно из фиг. 15, ингибитор бромодомена JQ-1 увеличивал способность XVir-N-31 убивать клетки в комбинации с ингибитором CDK 4/6 палбоциклибом при низких значениях MOI. Световой микроскопический анализ через 48 часов после инфицирования показывал уже массовую гибель клеток в случае клеток, обработанных JQ-1/палбоциклиб/XVir-N-31. Следует сделать вывод, что JQ-1 увеличивал транскрипцию вируса и, следовательно, репликацию вируса в клетках, обработанных палбоциклибом, поскольку монотерапия только 300 нМ JQ-1 не увеличивала гибель клеток XVir-N-31 при 10 и 20 MOI.
Предпосылкой для наблюдаемого усиления JQ-1 в инфицированных аденовирусом раковых клетках является способность палбоциклиба вызывать арест G1. В клетках, устойчивых к палбоциклибу (см. пример 18, идентичная процедура обработки), не наблюдалось увеличения гибели клеток. Это наблюдение резко контрастировало с данными Baojie Lv et al 2018, Scientific reports, 8, 11554, где клетки обрабатывали концентрациями JQ-1, не вызывающими арест G1, и одновременно не использовали палбоциклиб.
Пример 19: Тройная терапия, включающая XVir-N-31, ингибитор CDK4/6 и ингибитор бромодомен
100000 клеток SK-N-MC на лунку высевали в 12-луночные планшеты и выращивали в среде RPMI, содержащей 10% FCS при 5% CO2 при температуре 37°C. Клетки обрабатывали 200 нМ абемациклиб+500 нМ JQ-1 за 24 часа до и опять через 1 час после инфицирования, добавляя соответствующее количество в среду. Инфицирование XVir-N-31 происходило в 500 мкл среды RPMI, не содержащей сыворотки. SRB-окрашивание проводили через 5 дней после инфицирования. Макетные обработанные клетки имели 100% выживаемость клеток.
Результаты показаны на фиг. 16.
Установлено, что клетки SK-N-MC устойчивы к ингибиторам CDK 4/6, которые, таким образом, не вызывают арест G1. Добавление JQ-1 не увеличивало гибель клеток SK-N-MC, устойчивых к CDK 4/6 (абемациклиб), что указывает на то, что арест G1, опосредованный CDK 4/6, является предпосылкой опосредованного JQ-1 эффекта на гибель клеток.
Таким образом, на фиг. 16 (а также на фиг. 15) показано, что ингибиторы бромодомена, нацеленные на BRD2, BRD3, BRD4, еще больше увеличивают эффект уничтожения клеток XVir-N-3 при условии, что ингибиторы CDK 4/6 вызывают арест G1 в обработанных клетках.
Пример 20: Вестерн-блоттинг клеток SK-N-MC, обработанных ингибитором CDK 4/6 LY-2835219 (абемациклиб) и ингибитором Wee MK-1775 (адавосертиб)
1×106 клеток высевали в чашки диаметром 10 см. Через 24 часа после обработки выделяли белки с использованием 1% SDS-буфера, чтобы добиться разрушения ядерной мембраны. Все образцы набирали несколько раз в шприц для разрушения ДНК и затем центрифугировали при 30000 об/мин при температуре 4°C в течение 30 минут. Супернатант переносили в новую реакционную пробирку и непосредственно использовали для дальнейших стадий или хранили при температуре -80°C. Для разделения белков проводили электрофорез в полиакриламидном геле с додецилсульфатом натрия. Посредством электрофореза в течение приблизительно двух часов при 100 В при температуре 4°C загружали 40 мкг общих белков и зондировали их против специфических указанных антител.
Результаты показаны на фиг. 17.
Известно, что клетки SK-N-MC устойчивы к обработке абенациклибом (Dowless M et al., 2018, Clin Cancer Res: 24, 6028-6039). Wee1 является критическим компонентом контроля контрольной точки клеточного цикла G2/M и опосредует арест клеточного цикла, регулируя фосфорилирование CDC2. Сообщалось, что ингибирование Wee1 с помощью MK1775 усиливает цитотоксический эффект агентов, повреждающих ДНК, при различных типах карцином. В ряде исследований было продемонстрировано, что фармакологическое ингибирование Wee1 низкомолекулярным ингибитором киназы MK-1775 приводит к удалению фосфорилирования CDC2 по Tyr15 в опухолевых клетках (Kreahling et al 2013, PLoS One.8 (3), e 57523). Хотя при комбинированной обработке наблюдается сильный арест G1, изменений в экспрессии Rb и E2F1 не наблюдается.
Пример 21: Тройная терапия, включающая XVir-N-31, ингибитор CDK4/6 абемациклиб и адавосертиб (ингибитор Wee MK-1775)
100000 клеток SK-N-MC на лунку высевали в 12-луночные планшеты и выращивали в среде RPMI, содержащей 10% FCS при 5% CO2 при температуре 37°C. Клетки обрабатывали 200 нМ абемациклиба за 24 часа до и опять через 1 час после инфицирования, добавляя соответствующее количество в среду. Инфицирование XVir-N-31 происходило в 500 мкл среды RPMI, не содержащей сыворотку. SRB-окрашивание проводили через 5 дней после инфицирования. Макетные обработанные клетки имели 100% выживаемость клеток.
Результаты показаны на фиг. 18.
На фиг. 17 (и 18) продемонстрировано, что комбинация ингибитора CDK 4/6 абемациклиба и ингибитора Wee-MK-1775 индуцировала арест G1 без ингибирования E2F1. Количественное определение активного вещества/компонента на фиг. 18 показывает, что эта комбинация не усиливает эффект уничтожения клеток онколитического аденовируса XVir-N-31. Эти результаты ясно демонстрируют, что индуцированный арест G1 комбинацией ингибитора CDK 4/6 абемациклиба и ингибитора Wee MK-1775 не способствовал способности уничтожать клетки XVir-N-31. Таким образом, ингибирование экспрессии E2F1 является дополнительным требованием для усиления вирусного онколиза.
Пример 22: Арест G1 в комбинации с ингибированием E2F1 является предпосылкой для усиленного уничтожения клеток с помощью XVir-N-31 в сочетании с ингибиторами CDK 4/6
Через 48 часов после обработки клетки дважды промывали PBS (содержащим РНКазу A, 100 Ед/мл). Клетки трипсинизировали и центрифугировали при 1500 об/мин и температуре 4°C в течение 5 мин. Клетки фиксировали, медленно добавляя по каплям к осадку 1 мл ледяного 80% этанола и инкубируя в течение ночи. Окрашивание проводили, добавляя к клеткам 1 мл окрашивающего раствора (пропидиум йодид, 50 мкг/мл) и инкубируя 30-60 мин при комнатной температуре с осторожным покачиванием. MK: MK-1775; LY: LY-2835219.
Результаты показаны на фиг. 19.
Как видно из фиг. 19, обработка клеток SK-M-NC LY (абемациклиб) не влияла на клеточный цикл. Обработка только MK-1775 вызывала на 500 нМ увеличение G2/M клеток. Комбинация обоих вызывала сильный арест G1.
Пример 23: Роль экспрессии E2F1 в репликации вирусной ДНК
I
2×105 клеток T24, A549 и HeLa высевали на лунку в 6-луночный планшет и выращивали в 1,5 мл среды RPMI 1640, содержащей (или среду DMEM) 10% FBS, пенициллин/стрептомицин и заменимые аминокислоты. На следующий день 30 пмоль миРНК - будь то миРНК отрицательного контроля (Qiagen # 1022076) или siE2F1 (Sigma # NM_005225, миРНК ID SASI_Hs01_00162220) разводили в 150 мкл среды Opti-MEM и получали 9 мкл липофектамина RNAiMAX в 150 мкл Opti-MEM. Раствор миРНК и раствор липофектамина RNAiMAX смешивали и инкубировали в течение 5 минут. Смесь по каплям добавляли к клеткам. Через 48 часов РНК выделяли и проводили RT-qPCR.
Результаты показаны на фиг. 20. Как явствует из фиг. 20, E2-ранняя экспрессия снижена.
II
Для каждой лунки 6-луночного планшета 2×105 клеток Т24 высевали в 1,5 мл среды RPMI 1640, содержащей 10% FBS, пенициллин/стрептомицин и заменимые аминокислоты. На следующий день 30 пмоль миРНК - будь то миРНК отрицательного контроля (Qiagen # 1022076) или siE2F1 (Sigma # NM_005225, миРНК ID SASI_Hs01_00162220) разводили в 150 мкл среды Opti-MEM и получали 9 мкл липофектамина RNAiMAX в 150 мкл Opti-MEM. Раствор миРНК и раствор липофектамина RNAiMAX смешивали и инкубировали в течение 5 минут. Смесь по каплям добавляли к клеткам Т24. Инфицирование происходило через 48 часов во время инкубации клеток с 10 MOI ADWTRGD в 400 мкл бессывороточной среды и покачивания планшета каждые 10-15 минут. Через 1 час добавляли 1,6 мл полной среды. Выделение РНК проводили через 24 часа после инфицирования.
Результаты показаны на фиг. 21.
III
Клетки промывали холодным PBS и разрушали добавлением 500 мкл лизирующего буфера из набора MirVana, номер по каталогу AM1560 компании Thermo Fisher, лизаты собирали шпателем и переносили пипеткой в пробирку на 1,5 мл. Для органической экстракции добавляли 50 мкл добавки гомогената и образцы инкубировали на льду в течение 10 мин. Добавляли 500 мкл смеси кислота-фенол:хлороформ, образцы встряхивали в течение 60 секунд и инкубировали на льду в течение 2 минут. Образцы центрифугировали при 14000 g при комнатной температуре в течение 5 минут для разделения водной и органической фаз. Верхнюю фазу осторожно переносили в новую пробирку и добавляли равное количество изопропанола. После инкубации при комнатной температуре в течение 10 минут РНК осаждали (14000×g, 4°C, 30 минут) и дважды промывали 1 мл 75% этанола (центрифуга 7500×g, 4°C, 5 минут). РНК сушили на воздухе в течение 5-10 минут и ресуспендировали в 20-50 мкл воды, не содержащей РНКазы, и растворяли встряхиванием при 500 об/мин, 55°C в течение 10 минут и измеряли с помощью Nanodrop. После расщепления ДНК (дезоксирибонуклеаза I, Invitrogen, номер по каталогу 18068-015) с использованием 1 мкг образца РНК и 1 мкл 10-кратного реакционного буфера ДНКазы I, воды, не содержащей нуклеазу, до объема 9 мкл, 1 мкл ДНКазы I (1 Ед/мкл), инкубация 15 мин при комнатной температуре, инактивация ДНКазы I путем добавления 1 мкл 25 мМ раствора EDTA, нагревание в течение 10 мин при температуре 65°C. Обратную транскрипцию выполняли с использованием набора для обратной транскрипции кДНК высокой емкости (номер по каталогу Thermo Fisher/Applied Biosystems™: 4368814). С использованием случайного гексамера для транскрипции для ПЦР волокон и актина и с использованием раннего праймера E2 для транскрипции для E2Early-ПЦР.
Используемые праймеры и миРНК
E2 Earlyfw: CCGTCATCTCTACAGCCCAT (SEQ ID NO: 19)
E2 Earlyrev: GGGCTTTGTCAGAGTCTTGC (SEQ ID NO: 20)
fiberfw: AAGCTAGCCCTGCAAACATCA (SEQ ID NO: 21)
fiberrev: CCCAAGCTACCAGTGGCAGTA (SEQ ID NO: 22)
Actinfw: TCACCCACACTGTGCCCATCTACG (SEQ ID NO: 23)
Actinrev: CAGCGGAACCGCTCATTGCCAATGG (SEQ ID NO: 24)
E2F1 fw: CATCCCAGGAGGTCACTTCTG (SEQ ID NO: 25)
E2F1 rev: GACAACAGCGGTTCTTGCTC (SEQ ID NO: 26)
Контрольная миРНК
Смысловой: UUCUCCGAACGUGUCACGUdTdT (SEQ ID NO: 27)
Антисмысловой: ACGUGACACGUUCGGAGAAdTdT (SEQ ID NO: 28)
E2F1 миРНК
CUGAGGAGUUCAUCAGCCU[dT][dT] (SEQ ID NO: 29)
AGGCUGAUGAACUCCUCAG[dT][dT] (SEQ ID NO: 30)
Чтобы доказать роль E2-ранней экспрессии с помощью RT-qPCR, абсолютно необходимо выбрать правильный праймер. Местоположение праймера должно быть между E2-ранним и E2-поздним промоторами. В противном случае E2-поздний промотор сильно повлияет на результаты. Расположение праймеров показано на фиг. 22.
Как видно из фиг. 20 и 21, подавление E2F1 с помощью миРНК вызывает повышение E2-ранней экспрессии. Это можно объяснить только репрессивной ролью E2F1 в E2-ранней экспрессии. Если бы E2F1 был активатором, следствием этого было бы снижение E2-ранней экспрессии. Кроме того, миРНК против E2F-1 имитирует эффект ингибиторов CDK 4/6, который также ингибирует экспрессию E2F-1 (Yang C et al., Oncogene 2017, 36,2255-2264).
Пример 24: Рекомбинантный аденовирус с мутациями двух сайтов связывания E2F1 в аденовирусном E2-раннем промоторе показывает повышенную экспрессию раннего E2
Был получен мутантный аденовирус, имеющий мутации в двух E2F1-связывающих сайтах аденовирусного E2-раннего промотора. Промотор как E2-раннего промотора дикого типа, так и мутантного E2-раннего промотора показан на фиг. 23 (SEQ ID NO 44, 45, 46 и 47).
Анализ экспрессии РНК проводили в клетках Т24, инфицированных AdWT-RGD и AdE2Fm (содержащих также мотив RGD), полученных с помощью RT-qPCR через 24 часа после инфицирования. Экспрессия гена AD-WT была установлена на 100%. Метод был идентичен методу, описанному в разделе III примера 23.
Результаты показаны на фиг. 24.
Как явствует из фиг. 24, экспрессия в случае экспрессии E2-раннего гена была выше в клетках, инфицированных AdE2Fm, по сравнению с клетками, инфицированными AD-WT. Следовательно, следует сделать вывод, что E2F-1 играет репрессивную роль в активации E2-раннего промотора. Это резко контрастирует с нынешним пониманием, согласно которому E2F1 постулируется как активатор (DeCaprio JA, Virology. 20 февраля 2009; 384(2):274-84.
Хорошо известно, что структура E2-области во всех известных в настоящее время онколитических аденовирусах построена, как показано на фиг. 22. Таким образом, механизм действия E2F1 идентичен описанному в настоящем документе. Следовательно, все они, то есть все онколитические аденовирусы, могут использоваться в комбинации с ингибиторами CDK 4/6, включая ColoAd1 и Delta-24-RGD.
ColoAd1 можно охарактеризовать следующим образом:
Enadenotucirev (ранее ColoAd1) представляет собой опухолелективный химерный аденовирус с продемонстрированной доклинической активностью. Капсид ColoAd1 происходит от Ad11p, серотипа с ограниченной серологической распространенностью у людей. EnAd инфицирует клетки путем связывания с CD46 и/или десмоглеином 2,6, которые широко экспрессируются на многих клетках карциномы. Большая часть генома EnAd происходит от Ad11p с большой делецией в E3 и меньшей делецией в E4. Кроме того, область E2B состоит из химеры последовательностей Ad11p и Ad3. Делеция E4 в EnAd находится в E4ORF4, который в Ad5 кодирует белок, который инактивирует протеин-фосфатазу 2A и, таким образом, активирует аппарат трансляции белка, а также регулирует активность белка E1A в петле ингибирования обратной связи. Эти делеции, возможно в сочетании с химерным участком E2B, вероятно, способствуют поразительной избирательной репликации EnAd в отношении злокачественного новообразования (Deyer et al., Mol Ther Oncolytics. 2017, 16; 5: 62-74)
Delta-24-RGD (DNX-2401) можно охарактеризовать следующим образом:
Delta-24-RGD (DNX-2401) представляет собой онколитический вирус, способный к условной репликации, сконструированный для преимущественной репликации и лизиса опухолевых клеток с аномалиями пути p16/RB/E2F. Fueyo et al., Oncogene. 2000 Jan 6;19(1):2-12. Мутантный онколитический аденовирус, нацеленный на путь Rb, оказывает антиглиомный эффект in vivo; Dai B. et al. Mol Cancer Therapy. 2017 Apr;16(4):662-670.
Пример 25: Тройная терапия, содержащая XVir-N-31, ингибитор CDK4/6 палбоциклиб и ингибитор PARP талазопариб и FACS-анализ клеток T24 и UMUC-3
Анализ активности/окрашивание SRB
Эффект индуцированного вирусом уничтожения клеток отдельно и в сочетании с низкомолекулярными ингибиторами палбоциклибом и талазопарибом анализировали в 12-луночных планшетах. Таким образом, 12,5×103 клеток T24 или 6,3×103 клеток UMUC-3/253J высевали и через 24 часа инфицировали увеличивающейся концентрацией (множественность заражения, MOI) XVir-N-31. Для комбинированного лечения указанными низкомолекулярными ингибиторами клетки обрабатывали соответствующими концентрациями талазопариба, палбоциклиба или комбинации обоих препаратов за 24 часа до заражения и через 1 час после заражения (hpi). Клетки заражали в трехкратной повторности указанным вирусом в 200 мкл среды без эмбриональной бычьей сыворотки (FBS). При 1 hpi, к клеткам добавляли полную среду с низкомолекулярными ингибиторами или без них. Через четыре (T24) или пять (UMUC-3/253J) дней после инфицирования (dpi) клетки фиксировали 10% трихлоруксусной кислотой (TCA) в течение 1 часа при температуре 4°C и окрашивали сульфородамином B (SRB) в течение 30 минут при комнатной температуре с последующей промывкой 1% уксусной кислотой для удаления избытка SRB. Высушенный SRB растворяли в 10 мМ трис-основе и проводили количественную оценку с помощью фотометрического измерения с использованием планшет-ридера с несколькими метками (PerkinElmer Victor X3) при 562 нм.
Результаты показаны на фиг. 35, 36 и 37.
FACS-анализ
Влияние применяемых низкомолекулярных ингибиторов в указанных концентрациях на клеточный цикл клеточных линий рака мочевого пузыря T24 и UMUC-3 анализировали методом проточной цитометрии после окрашивания ДНК. 5×104 клеток высевали в 6-луночные планшеты и через 24 часа обрабатывали соответствующими ингибиторами. Через два дня при приблизительно 80%-ной конфлюэнтности клетки промывали PBS, трипсинизировали, снова промывали и фиксировали охлажденным на льду 80%-ным этанолом. Для анализа клеточного цикла образцы инкубировали с интеркалирующим ДНК красителем 7-аминоактиномицином D (7-AAD) и измеряли с помощью анализа FACS. Оценку измеренных данных проводили с помощью программного обеспечения FlowJo.
Результат показан на фиг. 38.
Результаты, показанные на рис. 35, 36 и 37 ясно демонстрирует, что тройная терапия, состоящая из палбоциклиба, талазопариба и XVir-N-31, продемонстрировала превосходную эффективность по сравнению с монотерапией или комбинированной терапией в отношении уничтожения клеток.
Анализ FACS обработанных клеток мочевого пузыря T24 и UMUC-3, представленный на фиг. 38, показывает, что на остановку G1, необходимую для повышенной репликации вируса, не влияет добавление ингибитора PARP талазопариба.
Пример 26: Тройная терапия, включающая XVir-N-31, ингибитор CDK4/6 палбоциклиб и ингибитор бромодомена JQ-1
Количественное определение активного вещества/компонента
Эффект индуцированного вирусом уничтожения клеток с использованием одного вируса и в комбинации с низкомолекулярными ингибиторами анализировали в 12-луночных планшетах. Для этого высевали 2×104 клеток (Cal-33) и через 24 часа инфицировали 5 инфекционными вирусными частицами на клетку (множественность заражения, MOI) XVir-N-31. Для комбинированного лечения указанными низкомолекулярными ингибиторами клетки обрабатывали 100 нМ палбоциклиба, 100 нМ JQ-1 или комбинацией обоих препаратов за 24 часа до заражения и через 1 час после заражения (hpi). Клетки заражали в трех повторностях указанным вирусом в 200 мкл среды без FBS. При 1 hpi к клеткам добавляли полную среду с низкомолекулярными ингибиторами или без них. Через четыре дня после инфицирования (dpi) клетки фиксировали 10% трихлоруксусной кислотой (TCA) в течение 1 часа при температуре 4°C и окрашивали сульфородамином B (SRB) в течение 30 минут при комнатной температуре с последующим промыванием 1% уксусной кислотой до удалить излишки SRB. Высушенный SRB растворяли в 10 мМ трис-основе и проводили количественную оценку с помощью фотометрического измерения с использованием планшет-ридера с несколькими метками (PerkinElmer Victor X3) при 562 нм.
Анализ репликации аденовирусов
Волоконная qPCR
Выделение ДНК
Для очистки ДНК среду отсасывали, в лунки добавляли по 200 мкл лизиса-распылителя. После добавления в раствор протеиназы К инкубировали при 56°С в течение 10 минут. К лизату вирусных клеток добавляли 200 мкл фенол-хлороформ-изоамилового спирта. После встряхивания и последующей инкубации в течение 5 минут на льду фазовое разделение достигалось центрифугированием в течение 3 минут при 16430g при 4°C. Верхнюю водную фазу переносили в новую защелкивающуюся крышку, содержащую 200 мкл хлороформа и 20 мкл крезолового красного в 10 мМ TrisCl для лучшей визуализации фаз. После встряхивания и инкубации в течение 5 минут на льду проводили центрифугирование в течение 3 минут при 16430 g при 4°С. Снова верхнюю водную фазу объединяли с 800 мкл этанола и 50 мкл 3М раствора ацетата натрия. Добавляли 2 мкл раствора гликогена для лучшего осаждения. После кратковременного переворачивания пробирки раствор центрифугировали 30 мин при 16430 g при 4°С. Затем осадок ДНК покрывали 400 мкл 70% этанола и инкубировали в течение 10 минут при комнатной температуре. После центрифугирования в течение 7 минут при 4760 g при комнатной температуре осадки ДНК сушили в течение примерно 5-10 минут при 37°С. Затем осадки растворяли в 100 мкл 0,1xTE-буфера и встряхивали при 40°С при 400 об/мин в течение примерно 3 часов. Когда ДНК полностью растворялась, концентрацию ДНК измеряли с помощью спектрофотометра, используя 2 мкл раствора ДНК для измерения и 0,1xTE-буфер в качестве контрольного раствора. Затем ДНК хранили при 4°C.
PCR в реальном времени
Вирусную репликацию анализировали с помощью количественной ПЦР, специфичной для аденовирусов (qPCR). Используя метод ΔΔCT, репликацию XVir-N-31 рассчитывали по копиям вирусной ДНК на клетку (нормализация к клеточному β-актину) по отношению к уровню проникновения вируса в клетки (значение 4 hpi). Клетки Cal-33 высевали в 6-луночные планшеты (1,5×105 на лунку) и предварительно обрабатывали указанными ингибиторами в течение 24 часов. На следующий день клетки инфицировали 10 MOI XVir-N-31 в 400 мкл среды без FBS. При 1 hpi к клеткам добавляли полную среду с низкомолекулярными ингибиторами или без них. Для выделения ДНК клетки собирали при 4, 24 и 48 hpi. Затем ДНК выделяли методом экстракции фенолом и хлороформом. Определенные с помощью qPCR значения кратности изменений контрольных образцов (10 MOI XVir-N-31 без дополнительного низкомолекулярного ингибитора) устанавливали равными 1,0, и относительно них наносили кратность изменений других образцов. Количественную PCR проводили на 96-луночных планшетах с SYBR Green Mastermix (Eurogentec) при следующих условиях циклирования: 95°С в течение 2 мин и 45 циклов 94°С в течение 15 с, 60°С в течение 15 с, 72°С в течение 15 с. Экспрессию генов рассчитывали с помощью ΔΔCT-метода.
Для выявления вирусной ДНК использовали следующие специфические праймеры:
Актин прям. 5'-TAAGTAGGTGCACAGTAGGTCTG-3' (SEQ ID NO: 31)
Волокно обратн. 5'-AAAGTGCAAAGAACACGGCTAAG-3' (SEQ ID NO: 32)
Волокно прям. 5'-AAGCTAGCCCTGCAAACATCA-3' (SEQ ID NO: 33)
Волокно обратн. 5'-CCCAAGCTACCAGTGGCAGTA-3' (SEQ ID NO: 34)
Клетки A673 (ATCC CRL 1598): клеточная линия саркомы Юинга, созданная в 1973 г. Martínez-Ramirez et al. 2003. Характеристика клеточной линии А673 (опухоль Юинга) методами молекулярной цитогенетики. Cancer Genet Cytogenet. Mar;141(2):138-42.
Клетки Cal-33: плоскоклеточный рак языка. CAL 33 представляет собой широко используемую клеточную линию плоскоклеточного рака головы и шеи (HNSCC) для тестирования терапевтических агентов.
Результаты
Результат количественного определения активного вещества/компонента клеточной линии саркомы Юинга A673 с использованием только Xvir-N-31, в комбинации с ингибитором CDK4/6 абемациклибом, в комбинации с ингибитором бромодомена JQ-1 или в комбинации с ингибитором CDK4/6 абемациклибом и ингибитором бромодоменома JQ-1 с MOI 5, 10 и 20 показан на фиг. 39.
Результат количественного определения активного вещества/компонента клеток плоскоклеточного рака головы и шеи (HNSCC) Cal-33 показан на фиг. 40 и 41.
QPCR проводили для анализа репликации вируса при следующих условиях обработки: XVir-N-31 MOI:10; 100 нМ палбоциклиб, 100 нМ JQ-1; клеточная линия: Cal-33; анализ через 24 часа после заражения и через 48 часов после заражения). Инфицирование происходило с помощью 10 MOI XVir-N-31 через 24 часа после обработки JQ-1, палбоциклибом или обоими (комбинацией). Через 4, 24 и 48 часов выделяли ДНК и проводили PCR в реальном времени для определения репликации вируса. Полученные результаты нормализовали по β-актину и 4-часовым значениям.
Результаты показаны на фиг. 42А и фиг. 42В. Во всех клеточных линиях тройная терапия превосходила монотерапию или любую двойную терапию, т.е. XVir-N-31+ингибитор CDK4/6; XVir-N-31+ингибитор бромодомена; и ингибитор CDK4/6+ингибитор бромодомена.
Пример 27: Тройная терапия, включающая XVir-N-31, ингибитор CDK4/6 палбоциклиб и нутлин и производные нутлина
Пример 27.1 Методы и материалы
Анализ активности/окрашивание SRB
Гибель клеток с использованием только XVir-N-31 и в комбинации с низкомолекулярными ингибиторами анализировали в 12-луночных планшетах. Таким образом, 20 000 клеток T24 или 25 000 клеток UMUC-3 высевали и через 24 часа инфицировали увеличивающейся концентрацией (множественность заражения, MOI) XVir-N-31. Для комбинированного лечения указанными низкомолекулярными ингибиторами клетки обрабатывали соответствующими концентрациями нутлина-3a или идазанутлина, палбоциклиба и комбинацией обоих препаратов за 24 часа до инфицирования. Клетки заражали в трехкратной повторности указанным вирусом в 250 мкл среды без FBS. При 1 hpi к клеткам, предварительно обработанным палбоциклибом, добавляли полную среду с палбоциклибом. К контролю, а также к клеткам, обработанным нутлином, добавляли полную среду без ингибиторов. Через четыре дня после заражения (dpi) клетки фиксировали 10% трихлоруксусной кислотой (TCA) в течение 1 часа при 4°C и окрашивали 0,05% сульфородамина B (SRB) в 1% уксусной кислоте в течение не менее 30 минут при комнатной температуре с последующей промывкой 1% уксусной кислотой для удаления избытка SRB. После фотографирования планшетов высушенный SRB растворяли в 10 мМ трис-основе и проводили количественный анализ с помощью фотометрического измерения с использованием многофункционального планшетного ридера (PerkinElmer Victor X3) при 562 нм.
FACS-анализ
Влияние применяемых низкомолекулярных ингибиторов в указанных концентрациях на клеточный цикл клеточных линий рака мочевого пузыря T24, T24shRb, UMUC-3 и RT112 анализировали методом проточной цитометрии после окрашивания ДНК. Для этого 5×104 клеток высевали в 6-луночные планшеты и через 24 часа обрабатывали соответствующими ингибиторами. Через два дня при приблизительно 80%-ной конфлюэнтности клетки промывали PBS, трипсинизировали, снова промывали и фиксировали охлажденным на льду 80%-ным этанолом. Для анализа клеточного цикла образцы инкубировали с проподиевым йодом (PI) и измеряли с помощью анализа FACS. Оценку измеренных данных проводили с помощью программного обеспечения FlowJo.
Аденовирусная репликация
Волоконная qPCR
Выделение ДНК
Для очистки ДНК среду отсасывали, в лунки добавляли по 200 мкл лизиса-распылителя. После добавления в раствор протеиназы К инкубировали при 56°С в течение 10 минут. К лизату вирусных клеток добавляли 200 мкл фенол-хлороформ-изоамилового спирта. После встряхивания и последующей инкубации в течение 5 минут на льду фазовое разделение достигалось центрифугированием в течение 3 минут при 16430 g при 4°C. Верхнюю водную фазу переносили в новую защелкивающуюся крышку, содержащую 200 мкл хлороформа и 20 мкл крезолового красного в 10 мМ TrisCl для лучшей визуализации фаз. После встряхивания и инкубации в течение 5 минут на льду проводили центрифугирование в течение 3 минут при 16430 g при 4°C. Снова верхнюю водную фазу объединяли с 800 мкл этанола и 50 мкл 3 М раствора ацетата натрия. Добавляли 2 мкл раствора гликогена для лучшего осаждения. После кратковременного переворачивания пробирки раствор центрифугировали 30 мин при 16430 g при 4°С. Затем осадок ДНК покрывали 400 мкл 70% этанола и инкубировали в течение 10 минут при комнатной температуре. После центрифугирования в течение 7 минут при 4760 g при комнатной температуре осадки ДНК сушили в течение примерно 5-10 минут при 37°C. Затем осадки растворяли в 100 мкл 0,1xTE-буфера и встряхивали при 40°С при 400 об/мин в течение примерно 3 часов. Когда ДНК полностью растворялась, концентрацию ДНК измеряли с помощью спектрофотометра, используя 2 мкл раствора ДНК для измерения и 0,1×TE-буфер в качестве контрольного раствора. Затем ДНК хранили при 4°C.
PCR в реальном времени
Вирусную репликацию анализировали с помощью количественной PCR, специфичной для аденовирусов (qPCR). Используя метод ΔΔCT, репликацию XVir-N-31 рассчитывали по копиям вирусной ДНК на клетку (нормализация к клеточному β-актину) по отношению к уровню проникновения вируса в клетки (значение 4 hpi). Клетки T24 и T24shRb высевали в 6-луночные планшеты (5×105 на лунку) и предварительно обрабатывали указанными ингибиторами в течение 24 часов. На следующий день клетки инфицировали 20 MOI XVir-N-31 в 400 мкл среды без FBS. При 1 hpi полную среду с палбоциклибом добавляли к клеткам, уже предварительно обработанным палбоциклибом. К контролям и клеткам, обработанным Нутлином, добавляли полную среду без ингибиторов. Для выделения ДНК клетки собирали при 4 и 48 hpi. Затем ДНК выделяли методом экстракции фенолом и хлороформом. Определенные с помощью qPCR значения кратности изменений контрольных образцов (20 MOI XVir-N-31 без дополнительного низкомолекулярного ингибитора) устанавливали равными 1,0, и относительно них наносили кратность изменений других образцов. Количественную PCR проводили на 96-луночных планшетах с SYBR Green Mastermix (Eurogentec) при следующих условиях циклирования: 95°С в течение 2 мин и 45 циклов 94°С в течение 15 с, 60°С в течение 15 с, 72°С в течение 15 с. Экспрессию гена рассчитывали с использованием ΔΔCT-метода.
Для выявления вирусных копий использовали следующие специфические праймеры:
Актин прям. 5'-TAAGTAGGTGCACAGTAGGTCTG-3' (SEQ ID NO: 35)
Волокно обратн. 5'-AAAGTGCAAAGAACACGGCTAAG-3' (SEQ ID NO: 36)
Волокно прям. 5'-AAGCTAGCCCTGCAAACATCA-3' (SEQ ID NO: 37)
Волокно обратн. 5'-CCCAAGCTACCAGTGGCAGTA-3' (SEQ ID NO: 38)
Пример 27.2 Тройная терапия, включающая XVir-N-31, нутлин-3a и палбоциклиб на клетках T24
Анализ эффективности с использованием клеток T24 был проведен для определения эффекта уничтожения клеток только XVir-N-31, XVir-N-31 с нутлином-3a, XVir-N-31 с палбоциклибом и XVir-N-31 с обоими нутлином-3а и палбоциклибом.
Результат показан на фиг. 44 и 45.
Как видно из фиг. 44 и 45, предварительная обработка клеток Т24 палбоциклибом увеличивала гибель клеток с помощью XVir-N-31. Комбинированное лечение палбоциклибом и нутлином-3а усиливало этот эффект. Кроме того, обработка нутлином-3a усиливала гибель клеток, наблюдаемую для комбинации XVir-N-31 и палбоциклиба.
Пример 27.3 Тройная терапия, включающая XVir-N-31, нутлин-3a и палбоциклиб на клетках T24 shRB
Тот же анализ эффективности, что и в примере 27.2, проводили с использованием клеток T24shRB. Клетки T24shRB представляют собой клетки T24 без экспрессии Rb. Такая недостаточная экспрессия Rb была получена с помощью лентивирусной трансфекции, при этом лентивирус кодировал shРНК, направленную на Rb. Используемый лентивирусный вектор представлял собой pLKO-RB1-shRNA19 (полученный от Addgene, Watertown, MA 02472, USA), последовательность shRNA была следующей: CAGAGATCGTGTATTGAGATTCTCGAGAATCTCAATACACGATCTCTG (SEQ ID NO: 39). Процедура также описана Michaud K et al. (Cancer Res. 2010 Apr 15. 70(8):3228-38).
Результат показан на фиг. 46 и 47.
Как видно из фиг. 46 и 47, лечение нутлином-3а или палбоциклибом по отдельности не было эффективным в уничтожении Rb-отрицательных клеток (T24shRb) с помощью XVir-N-31. Только комбинация палбоциклиба и нутлина-3а показала увеличение опосредованной XVir-N-31 гибели клеток. Таким образом, нутлин-3a усиливал онколитический эффект двойной комбинированной терапии с использованием XVir-N-31 и палбоциклиба, обработанных против Rb-негативных клеток.
Пример 27.4 Тройная терапия, включающая XVir-N-31, идазанутлин и палбоциклиб на клетках T24 и клетках T24shRb
Анализ эффективности с использованием клеток T24 был проведен для определения эффекта уничтожения клеток XVir-N-31, XVir-N-31 с идасанутлином, XVir-N-31 и палбоциклибом, а также XVir-N-31, идасанутлином и палбоциклибом.
Результат показан на фиг. 48 и 49.
Аналогичный анализ эффективности с использованием клеток T24shRb был проведен для определения эффекта уничтожения клеток XVir-N-31, XVir-N-31 с идасанутлином, XVir-N-31 и палбоциклибом, а также XVir-N-31, идасанутлином и палбоциклибом.
Результат показан на фиг. 50 и 51.
Как видно на фиг. с 48 по 51, идасанутлин оказывал аналогичные эффекты, как и нутлин-3а, при тройной терапии с использованием XVir-N-31 и ингибитора CDK4/6.
Пример 27.5 Оценка репликации XVir-N-31 в условиях, описанных в примерах 27.2-27.4
Репликацию XVir-N-31 в клетках T24shRb и клетках T24 оценивали путем определения вирусной ДНК методом ΔΔCT.
Экспериментальная установка была следующей:
Репликация вируса определялась через 24 и 48 часов. Инфицирование происходило с 20 MOI XVir-N-31 через 24 часа после обработки нутлибом-3a, палбоциклибом или обоими (комбинация). Через 4 и 48 часов выделяли ДНК и проводили ПЦР в реальном времени для определения репликации вируса. Полученные результаты нормализовали к β-актину и 4-часовым значениям в соответствии с методом ΔΔCT.
Результат показан на фиг. 52.
Как видно на фиг. 52, нутлин-3a показал увеличение вирусной ДНК в сочетании с палбоциклибом. В то время как относительная ДНК волокна уже была высокой в клетках T24, обработанных палбоциклибом, комбинация с Nutlin-3a показала значение почти в 10 раз выше, чем в необработанном контроле. В Rb-негативных клетках T24shRb эффект проявлялся только при комбинировании нутлином-3a, палбоциклибом и XVir-N-31. При этом палбоциклиб сам по себе оказывал менее выраженное влияние на вирусную ДНК по сравнению с контролем.
Вестерн-блот-анализ проводили на клетках T24shRb, обработанных нутлином-3a (5 мкМ, 10 мкМ или 30 мкМ), палбоциклибом (500 мкМ) или обоими (в комбинации). Результат показан на фиг. 53.
Уровни белка E2F1, показанные на фиг. 53, после лечения палбоциклибом и/или нутлином-3а (нутлин-3а (5 мкМ, 10 мкМ или 30 мкМ), палбоциклибом (500 мкМ) или обоими (в комбинации)) показаны на фиг. 54. Уровень белка E2F1 в необработанных клетках был установлен на «1».
Как видно на фиг. 53 и 54, комбинированное лечение 30 мкМ нутлина-3a и 500 нМ палбоциклиба снижало относительное количество E2F1 по сравнению с монотерапией.
Пример 27.6 FACS-анализ клеток, полученных в результате обработки по примеру 27.5
Клетки, обработанные, как описано в примере 27.5, подвергали FACS-анализу. Кроме того, клетки UMUC и клетки RT112 подвергали такому же режиму обработки, как описано в примере 27.5.
Результаты показаны на фиг. 55 (А) для клеток T24, фиг. 55 (В) для клеток T24shRb, фиг. 55 (С) для клеток UMUC-3 и фиг. 55 (D) для клеток RT112.
Результат дальнейшего анализа результатов, представленных на фиг. 55(A)-(D) с акцентом на процентное содержание клеток, в G1 показано на фиг. 56 (A) для клеток T24, на фиг. 56 (B) для клеток T24shRb, на фиг. 56(C) для клеток UNUC-3 и фиг. 56 (D) для клеток RT112.
Как видно из указанных фиг. 55(А)-(D) и фиг. 56(A)-(D), влияние на арест G0/G1 во всех клеточных линиях было наиболее выраженным при обработке нутлином-3a и палбоциклибом в комбинации.
Пример 28: Комбинированная терапия, включающая ингибитор бромодомена JQ1 и XVir-N-31, в лечении глиобластомы
2×104 клеток (клетки U87, клетки LN229 или клетки T98G) высевали в 12-луночные планшеты в среде DMEM и на следующий день обрабатывали ингибитором. Через 48 часов после посева клетки инфицировали XVir-N-31. Через 4 дня клетки фиксировали 10% ТХУ. Планшеты окрашивали 0,1% раствором SRB. Избыток удаляли и планшеты промывали 1% уксусной кислотой. После удаления уксусной кислоты планшеты сушили на воздухе в течение ночи. SRB растворяли в 1 мл 10 мМ раствора Tris Base и измеряли разведение 1:10 с использованием фотометра при 560 нм.
Результаты показаны на фиг. 57, 58 и 59.
Как видно на фиг. 57, 58 и 59, комбинация of JQ1 и XVir-N-31 приводит к сильно усиленному уничтожению клеток. Использование ингибитора CDK4/6 LEE011 может еще больше увеличить гибель клеток во всех трех клеточных линиях глиобластомы.
Пример 29: Репликация XVir-N-31 при воздействии комбинации JQ1 и рибоциклиба
1×105 клеток высевали в 6-луночные планшеты в среде DMEM. На следующий день клетки обрабатывали 500 нМ LEE011 и 50 нМ JQ1. На следующее утро клетки инфицировали 20 MOI для U87 и LN229 и 50 MOI для T98G, соответственно. Через час после инфицирования к клеткам добавляли среду DMEM, содержащую ингибиторы. После каждой временной точки (4, 24, 48 часов) клетки один раз промывали PBS и добавляли 200 мкл SDS буфера для лизиса ДНК. Клетки инкубировали с протеиназой К в течение 1 часа, затем очищали фенол-хлороформ-изоамиловым спиртом. ДНК разводили в буфере 1×TE и определяли концентрацию ДНК с помощью Nanodrop. Для каждого образца готовили раствор 10 нг/мкл и проводили RT-qPCR с использованием 50 нг ДНК и смеси GoTaq Master. Использовались прямые и обратные праймеры для конкретных волокон, а также для расчета значения Δct использовались специфические для актина праймеры. Для каждого образца измеряли 3 значения волокна и 2 значения актина. Расчет был выполнен в Excel, и каждые 24 и 48 часов рассчитывались на основе его значения за 4 часа (начальное проникновение вируса до репликации).
Результаты показаны на фиг. 60, 61 и 62.
Как видно на фиг. 60, 61 и 62, XVir-N-31 показал сильное увеличение репликации при воздействии JQ1. В двух клеточных линиях этот эффект был намного выше по сравнению с использованием ингибитора CDK4/6 LEE011 (LEE). Во всех трех клеточных линиях увеличение репликации вируса было самым высоким при использовании комбинации как JQ1, так и LEE011 (LEE).
Пример 30: Вестерн-блоттинг инфицированных XVir-N-31 раковых клеток
2,5×105 клеток LN229 высевали в чашки диаметром 10 см в среде DMEM. На следующий день клетки обрабатывали 500 нМ LEE011 и 200 нМ JQ1 или их комбинацией. На следующее утро клетки инфицировали XVir-N-31 при 20 MOI. Через час после инфицирования к клеткам добавляли среду DMEM, содержащую ингибиторы. После каждой временной точки (24, 48, 72 часа) клетки помещали на лед, дважды промывали охлажденным льдом PBS и добавляли 300 мкл буфера для лизиса белка SDS. Клетки переносили в реакционные пробирки, ДНК разрезали шприцем и иглой, а супернатант переносили для дальнейшего использования после центрифугирования. Концентрацию белка измеряли с помощью анализа BCA, и образцы с 4x Lämmli:DTT 6:1 и концентрацией 20 мкг получали и переносили в гель с 10% SDS. Гель-электрофорез начинали при 90 В и повышали до 150 В после того, как начал отделяться лэддер. Белки на геле переносили на мембрану на 2 часа при 100 В и блокировали в 5% молоке в TBS-T на 1 час при комнатной температуре. После промывки в TBS-T первое антитело добавляли в соответствии с информацией производителя в течение ночи при 4°C. Используемые антитела были направлены на GAPDH, E1A, DBP, E2F1, Rb, pRb для всех временных точек, а также на каспазу 3 и полноразмерную и расщепленную версию PARP дополнительно для 72-часовых образцов. На следующий день промывку повторяли и добавляли вторичное антитело на 1 час при комнатной температуре. После промывки использовали ECL для визуализации хемилюминесценции с помощью системы визуализации ChemiDocTM MP Imaging.
Результаты показаны на фиг. 63, 64, 65, и 66.
Как показано на фиг. 63, вирусные белки E1A и DBP увеличивались при комбинированной обработке XVir-N-31 с 500 нМ LEE011 и/или 200 нМ JQ-1. Клеточные белки Rb и pRb в этих образцах оставались ингибированными (дорожки 5, 6 и 7).
Как показано на фиг. 64, вирусные белки E1A и DBP увеличивались при комбинированной обработке XVir-N-31 с LEE011 и/или JQ-1. Клеточные белки Rb и pRb в этих образцах оставались ингибированными, а E2F-1 стабилизировался в этих образцах (дорожки 5, 6 и 7).
Как показано на фиг. 65, вирусные белки E1A и DBP увеличивались при комбинированной обработке XVir-N-31 с LEE011 и/или JQ-1, в то время как клеточный белок Rb уменьшался, а E2F1 увеличивался в этих образцах. Более того, были видны продукты расщепления Rb с молекулярной массой 48 кДа, что указывало на апоптоз (Fattman, Cheryl L.; Delach, Scott M.; Dou, Qing Ping; Johnson, Daniel E. (2001): Sequential two-step cleavage of the retinoblastoma protein by caspase-3/-7 during etoposide-induced apoptosis. In: Oncogene 20 (23), S. 2918-2926).
Как показано на фиг. 66, каспаза 3 и PARP присутствовали во всех клеточных лизатах, даже в контроле. Расщепленный PARP обнаруживали не только по расщепленному PARP-антителу, но и по полноразмерному PARP-антителу. Расщепленный PARP был виден только в инфицированных клетках, и его присутствие увеличивалось при обработке LEE011 и JQ1. Это расщепление было признаком апоптоза (Chaitanya et al.: PARP-1 cleavage fragments: signatures of cell-death proteases in neurodegeneration. Cell Communication и Signaling 2010 8:31).
Пример 31: Влияние JQ1 на различные аденовирусы при глиобластоме
Аденовирусный E1A состоит из различных консервативных областей (CR), которые связывают различные клеточные белки и нарушают их функцию. CR1 связывает p300/CBP, который также может связываться с CR3. Другие области CR3 могут связывать различные субъединицы медиаторного комплекса, такие как MED23, и, следовательно, играть роль в трансактивации других вирусных ранних генов с помощью E1A. Район аденовируса CR1, а также CR2 способен связывать RB. Это взаимодействие приводит к высвобождению E2F и последующему развитию клеточного цикла. Более того, E2F связывает ранний промотор аденовирусного E2 и активирует его транскрипцию.
Взаимодействующие партнеры E1A и расположение консервативной области CR1-CR4 показаны на фиг. 67.
Чтобы выяснить, какие консервативные области Е1А необходимы для действия JQ1 на усиление репликации вируса, использовали аденовирусы с различными делециями и мутациями. Аденовирус дикого типа, содержащий мотив RGD для лучшей инфекционности и делецию E3 для ненарушенного противовирусного ответа, использовали в качестве контроля. Другими вирусами были dl119, у которого делетированы N-конец и CR1 и CR2, и Ad delta24, который находится под контролем промотора CMV и лишен CR2. В качестве вируса с делецией CR3 использовали XVir-N-31, который также содержит мотив RGD и делецию E3, а также делецию E1B-19k для усиления апоптоза. Другим используемым вирусом был ADWT/E2Fm. Этот вирус содержит мутации в сайтах связывания E2F на раннем промоторе E2, которые блокируют связывание E2F. Эти характеристики снова резюмируются в следующем:
ADWT: дикий тип E1A
XVir-N-31: ρCR3 E1A
Dl119: ρCR1 и ρCR2 E1A
Addelta24: ρCR2 E1A
ADWT/E2Fm: дикий тип E1A
1×105 клеток LN229 высевали в 6-луночные планшеты в среде DMEM. На следующий день клетки инфицировали 20 MOI различных вирусов (AdWt, dl119, Addelta24, XVir-N-31, AdWt/E2Fm). Через один час после инфицирования к клеткам добавляли среду DMEM или среду DMEM, содержащую 200 нМ JQ1, так что клетки были инфицированы с обработкой JQ1 или без нее. После каждой временной точки (4, 24, 48 часов) клетки один раз промывали PBS и добавляли 200 мкл SDS буфера для лизиса ДНК. Клетки инкубировали с протеиназой К в течение 15 мин при 56°С перед очисткой фенол-хлороформ-изоамиловым спиртом. ДНК разводили в буфере AE и определяли концентрацию ДНК с помощью Nanodrop. Для каждого образца готовили раствор 10 нг/мкл и проводили количественную ПЦР с использованием 50 нг ДНК и смеси GoTaq Master. Использовались прямые и обратные праймеры для конкретных волокон, а также для расчета значения Δct использовались специфические для актина праймеры. Для каждого образца измеряли 3 значения волокна и 3 значения актина. Расчет был выполнен в Excel, и каждые 24 и 48 часов рассчитывались на основе его значения за 4 часа (начальное проникновение вируса до репликации) и увеличения репликации через JQ1 по отношению к необработанному инфицированному образцу.
Нуклеотидная последовательность различных праймеров была следующей:
Результаты показаны на фиг. 68 и 69.
Среди различных вирусов XVir-N-31 продемонстрировал самое высокое увеличение вирусной репликации под влиянием JQ1 через 48 часов, что указывает на то, что вирус с делецией CR3 получает наибольшую пользу от обработки JQ1.
Пример 32: Двойная терапия XVir-N-31 и JQ-1 при раке мочевого пузыря
В качестве первого аспекта оценивали репликацию XVir-N-31.
2,5×104 клеток UMUC-3 высевали в 12-луночные планшеты в среде DMEM. На следующий день клетки обрабатывали средой JQ-1 (праймирование) или средой DMEM. На следующее утро клетки инфицировали 10 MOI XVir-N-31. Через один час после инфицирования к клеткам добавляли среду DMEM, содержащую JQ-1 (одновременная обработка). Через каждые 4 и 24 часа клетки один раз промывали PBS и добавляли 200 мкл буфера для лизиса ДНК SDS. Клетки инкубировали с протеиназой К в течение по меньшей мере 1 часа перед очисткой фенол-хлороформ-изоамиловым спиртом. ДНК разводили в буфере 1xTE и определяли концентрацию ДНК с помощью Nanodrop. Для каждого образца готовили раствор 15 нг/мкл и проводили RT-qPCR с использованием 75 нг ДНК и смеси GoTaq Master. Использовались специфические прямые и обратные праймеры для волокон, а также для расчета значения Δct использовались специфические актиновые праймеры. Для каждого образца измеряли 3 значения волокна и 3 значения актина. Расчет был выполнен в Excel, и каждое 24-часовое значение рассчитывалось на основе его 4-часового значения (начальное проникновение вируса до репликации).
Результаты показаны на фиг. 70.
Как видно на фиг. 70, праймирование per se не оказывает благотворного влияния на репликацию XVir-N-31. Незначительные положительные эффекты праймирования на изображенном рисунке возникли из-за недостаточного вымывания в экспериментальной установке. Параллельное лечение с JQ-1 имело решающее значение для успеха двойной терапии, тогда как предварительное добавление JQ-1 (праймирование) приводило к снижению эффективности по сравнению с одновременным лечением.
В качестве второго аспекта оценивали образование частиц XVir-N-31.
Продуцирование вирусных частиц анализировали с помощью иммуногистохимического окрашивания белков гексонов (тест Hexon Titer). Таким образом, 2×105 прикрепленных клеток HEK293 на лунку высевали в 24-луночные планшеты. Образцы клеточных культур, собранные после указанных моментов времени после заражения и указанной совместной обработки JQ-1, предварительно обрабатывали тремя циклами оттаивания-замораживания для высвобождения вирусных частиц из клеток, а затем серийно разбавляли. Клетки HEK293 инфицировали в объемах либо 50 мкл, либо 10 мкл на образец. Затем планшеты инкубировали в течение 40 ч при 37°C и 10% CO2 перед проверкой обнаруживаемых цитопатических эффектов. Среду отсасывали, а планшеты сушили в течение 5-10 мин перед фиксацией клеток добавлением ледяного метанола в течение 10 мин при -20°С. Лунки дважды промывали PBS+1% BSA. Добавляли первичное антитело (козье анти-гексон, 1:500) и планшеты инкубировали в течение 1 ч при 37°C перед двукратным промыванием PBS+1% BSA. Затем добавляли вторичное антитело (конъюгат Rabbit Anti-Goat HRP, 1:1000) и инкубировали в течение 1 ч при 37°C. Опять же, лунки дважды промывали PBS, содержащим 1% BSA, перед добавлением раствора DAB. После 30-минутной инкубации под 20-кратным объективом микроскопа подсчитывали 10 различных случайных полей зрения (FOV) на лунку. Итерации подсчета усредняли и по этой формуле рассчитывали инфекционных частиц на мл:

Результаты показаны на фиг. 71 и 72.
Как видно на фиг. 71, образование частиц XVir-N-31 значительно увеличивалось в клетках UMUC-3 при добавлении 500 нМ JQ-1. Кроме того, кинетика образования частиц XVir-N-31 значительно ускоряется при одновременной обработке 500 нМ JQ-1.
В качестве третьего аспекта был проведен Вестерн-блот-анализ.
1×106 клеток UMUC-3 высевали в чашки диаметром 10 см в среде DMEM и через 24 часа после посева инфицировали 10 MOI XVir-N-31. Через 1 час после инфицирования среду для культивирования либо повторно заполняли DMEM, либо добавляли DMEM с добавлением 500 нМ JQ-1. 12, 24, 36 и 48 hpi, лизаты общих белков брали с использованием SDS-буфера. Анализ BCA применяли для корректировки эквивалентных концентраций белка. Затем к образцам белка добавляли буфер Лэммли и разделяли с помощью гель-электрофореза с SDS-PA на 10% PA-гелях. Образцы наносили на мембраны PVDF с использованием системы BioRAD Wet Tank System. Эти мембраны блокировали буфером TBS-Tween, содержащим 5% сухого обезжиренного молока, в течение 1 часа при комнатной температуре. Инкубацию с первичными антителами проводили в течение ночи при 4°С, а с вторичными антителами, конъюгированными с пероксидазой хрена, в течение 1 часа при комнатной температуре. Хемилюминесценцию детектировали с помощью ECL prime (Pierce) с системой визуализации ChemiDoc (BioRad). Рисунки были разработаны в Adobe® Illustrator, а оценка была выполнена с помощью ImageLab.
Результаты показаны на фиг. 73.
Как видно на фиг. 73, абсолютная экспрессия вирусного белка и, прежде всего, кинетика вирусных белков XVir-N-31 (E1A, E1B55k, DBP и E40rf6) значительно ускорялись при одновременном лечении JQ-1. Благодаря лечению JQ-1, особенно более поздние вирусные белки, такие как Hexon, экспрессировались на высоких уровнях в ранние сроки после заражения XVir-N-31.
Пример 33: Тройная терапия рака мочевого пузыря с использованием XVir-N-31, палбоциклиба и JQ-1
В качестве первого аспекта влияние применяемых низкомолекулярных ингибиторов в их указанных концентрациях на клеточный цикл клеточных линий рака мочевого пузыря UMUC-3 и RT112 анализировали с помощью анализа проточной цитометрии после окрашивания ДНК.
Для этой цели 5×104 клеток UMUC-3 и RT112, соответственно, высевали в 6-луночные планшеты и через 24 часа обрабатывали соответствующими ингибиторами. Через 1 день при примерно 80% слиянии клетки промывали PBS, трипсинизировали, снова промывали и фиксировали охлажденным на льду 80% этанолом. Для анализа клеточного цикла образцы инкубировали с интеркалирующим ДНК красителем 7-аминоактиномицином D (7-AAD) и измеряли с помощью анализа FACS. Оценку измеренных данных проводили с помощью программного обеспечения FlowJo.
Результаты показаны на фиг. 74.
Как видно на фиг. 74, палбоциклиб индуцировал сильный арест в G1-фазе клеточного цикла. JQ-1 не оказывал значительного влияния на клеточный цикл ни в качестве монотерапии, ни в комбинации с палбоциклибом. Примечательно, что JQ-1 применяли в относительно низких концентрациях, которые все же достаточны для сильного усиления онколитического потенциала XVir-N-31. В опубликованной научной литературе были обнаружены неубедительные данные о более высоких дозах JQ-1, вызывающих арест G1 или G2. Однако низкие дозы JQ-1 не влияли на клеточный цикл.
В качестве второго аспекта был количественно определен уровень экспрессии специфических клеточных белков-мишеней после обработки как JQ-1, так и палбоциклибом.
1-2×106 клеток UMUC-3 или RT-112 высевали в чашки диаметром 10 см в среде DMEM и через 24 часа после посева обрабатывали соответствующими низкомолекулярными ингибиторами. Через 24 часа после обработки лизаты общих белков брали с использованием SDS-буфера. Анализ BCA применяли для корректировки эквивалентных концентраций белка. Затем к образцам белка добавляли буфер Лэммли и разделяли с помощью гель-электрофореза с SDS-PA на 10% PA-гелях. Образцы наносили на мембраны PVDF с использованием системы BioRAD Wet Tank System. Эти мембраны блокировали буфером TBS-Tween, содержащим 5% сухого обезжиренного молока, в течение 1 часа при комнатной температуре. Инкубацию с первичными антителами проводили в течение ночи при 4°С, а с вторичными антителами, конъюгированными с пероксидазой хрена, в течение 1 часа при комнатной температуре. Хемилюминесценцию детектировали с помощью ECL prime (Pierce) с системой визуализации ChemiDoc (BioRad). Фигуры были разработаны в Adobe® Illustrator, а оценка была выполнена с помощью ImageLab.
Результаты показаны на фиг. 75.
Как видно на фиг. 75, JQ-1 не оказывал в указанных концентрациях никакого влияния на уровень регуляторов клеточного цикла RB, фосфо-RB или белка E2F-1 ни при монотерапии, ни в комбинации с палбоциклибом. Репрессор РНК-полимеразы II Hexim1 подавляется после лечения JQ-1, но восстанавливается в сочетании с палбоциклибом. Таким образом, белки клеточного цикла RB и E2F-1 не ответственны за увеличение репликационной способности XVir-N-31 при обработке JQ-1.
В качестве третьего аспекта способность тройной терапии к уничтожению клеток определяли с помощью количественного определения активного вещества/компонента .
1-3×104 клеток высевали в 12-луночные планшеты в среде DMEM или RPMI и на следующий день обрабатывали ингибитором. Через 48 часов после посева клетки инфицировали XVir-N-31. Через 4 дня клетки фиксировали 10% TCA. Планшеты окрашивали 0,1% раствором SRB. Избыток удаляли и планшеты промывали 1% уксусной кислотой. После удаления уксусной кислоты планшеты сушили на воздухе в течение ночи. SRB растворяли в 1 мл 10 мМ раствора Tris Base и измеряли разведение 1:10 с использованием фотометра при 560 нм.
Результаты показаны на фиг. 76, 77 и 78.
Как видно на фиг. 76, 77 и 78 лечение вместе с низкими дозами палбоциклиба и JQ-1 позволяет XVir-N-31, даже при низком MOI, полностью ликвидировать выживаемость раковых клеток мочевого пузыря in vitro, что иллюстрируется соответствующими эффектами, наблюдаемыми в каждом и любом из клетки рака мочевого пузыря UMUC-3, RT112 и T24.
В качестве четвертого аспекта оценивалось влияние этой тройной терапии на репликацию вируса.
2-5×104 клеток UMUC-3, T24 или RT112 высевали в 12-луночные планшеты в среде DMEM или RPMI. На следующий день клетки обрабатывали JQ1, палбоциклибом (праймирование) или средой. На следующий день клетки инфицировали 10, 20 или 50 MOI XVir-N-31. Через час после инфицирования к клеткам добавляли среду, содержащую JQ-1 или палбоциклиб. Через 4 и 24 часа клетки один раз промывали PBS и добавляли 200 мкл SDS буфера для лизиса ДНК. Клетки инкубировали с протеиназой К в течение по меньшей мере 1 часа перед очисткой фенол-хлороформ-изоамиловым спиртом. ДНК разводили в буфере 1×TE и определяли концентрацию ДНК с помощью Nanodrop. Для каждого образца готовили раствор 15 нг/мкл и проводили RT-qPCR с использованием 75 нг ДНК и смеси GoTaq Master. Использовались прямые и обратные праймеры для конкретных волокон, а также для расчета значения Δct использовались специфические для актина праймеры. Для каждого образца измеряли 3 значения волокна и 3 значения актина. Расчет был выполнен в Excel, и каждое 24-часовое значение рассчитывалось на основе его 4-часового значения (начальное проникновение вируса до репликации).
Результаты показаны на фиг. 79, 80 и 81.
Как видно на фиг. 79, 80 и 81, дополнительное лечение низкими дозами палбоциклиба и JQ-1 значительно ускоряет репликацию XVir-N-31 до 200 раз.
В качестве пятого аспекта оценивалось влияние этой тройной терапии на продуктивно инфицированные опухолевые клетки.
Количество продуктивно инфицированных опухолевых клеток анализировали с помощью прямого иммуногистохимического окрашивания белков гексонов инфицированных клеток UMUC-3 (тест на псевдо-гексон-титр). Для этого 5×105 клеток UMUC-3 на лунку высевали в 12-луночные планшеты. На следующий день клетки праймировали низкими дозами JQ-1 и палбоциклиба. Затем через 48 часов после посева клетки инфицировали XVir-N-31 (9 MOI) и через один час одновременно обрабатывали соответствующими ингибиторами. Затем планшеты инкубировали в течение 40 ч при 37°C и 10% CO2 перед проверкой обнаруживаемых цитопатических эффектов. Среду отсасывали, а планшеты сушили в течение 5-10 мин перед фиксацией клеток добавлением ледяного метанола в течение 10 мин при -20°С. Лунки дважды промывали PBS+1% BSA. Добавляли первичное антитело (Goat Anti-hexon, 1:500) и планшеты инкубировали в течение 1 ч при 37°C, затем дважды промывали PBS+1% BSA. Затем добавляли вторичное антитело (конъюгат Rabbit Anti-Goat HRP, 1:1000) и инкубировали в течение 1 ч при 37°C. Опять же, лунки дважды промывали PBS, содержащим 1% BSA, перед добавлением раствора DAB. После 30-минутной инкубации под 20-кратным объективом микроскопа подсчитывали 10 различных случайных полей зрения (f.o.v.) на лунку.
Результаты показаны на фиг. 82 и 83.
Как видно на фиг. 82 и 83, дополнительное лечение низкими дозами палбоциклиба и JQ-1 значительно ускоряет количество опухолевых клеток, проходящих через продуктивную инфекцию XVir-N-31.
Пример 34: Влияние различных ингибиторов BET и деструктора BET в комбинации с XVir-N-31 на клеточные линии UMUC3- и RT112
Цель исследования состояла в том, чтобы определить эффективность XVir-N-31 в сочетании с различными ингибиторами BET (бромодомен и экстратерминальный мотив), включая деструкторы. Ингибитор BET (BETi): OTX015 (моновалентный), AZD5153 (двухвалентный) конкурентно ингибируют BET; деградатор BET (BETd): dBet6, ARV55 приводят к деградации BET (см., например, Rhyasen GW, et al. AZD5153: A Novel Bivalent BET Bromodomain Inhibitor Highly Active against Hematologic Malignancies. Mol Cancer Ther. 2016 Nov;15(11):2563-2574; J. Kay Noel, et al. Abstract C244: Development of the BET bromodomain inhibitor OTX015. Mol Cancer Ther. November 2013 12; C244; Winter GE, et al.: BET Bromodomain Proteins Function as Master Transcription Elongation Factors Independent of CDK9 Recruitment. Mol Cell. 2017 Jul 6;67(1):5-18.e19). Raina K, et al.: PROTAC-induced BET protein degradation as a therapy for castration-resistant prostate cancer. Proc Natl Acad Sci U S A. 2016 Jun 28;113(26):7124-9; описание которых включено в настоящий документ в качестве ссылки).
В первом аспекте выживание клеток UMUC3 и RT112 после инфицирования XVir-N-31 в сочетании с обработкой BETi и BETd анализировали с помощью количественного определения активного вещества/компонента .
1,3×104 клеток UMUC-3 и 3×104 клеток RT112 высевали в 12-луночные планшеты в среде DMEM и RT112. На следующий день клетки предварительно обрабатывали указанными концентрациями BETi и BETd. Через 48 часов после посева клетки инфицировали онколитическим аденовирусом XVir-N-31 и обрабатывали BETi и BETd. Через 5 дней клетки фиксировали 10% ТХУ. Планшеты окрашивали 0,05% раствором SRB. Избыток удаляли и планшеты промывали 1% уксусной кислотой. После удаления уксусной кислоты планшеты сушили на воздухе в течение ночи. SRB растворяли в 500 мкл 10 мМ раствора Tris Base и измеряли разведение 1:10 с использованием фотометра при 562 нМ.
Результаты показаны на фиг. 84 и 85.
Как видно на фиг. 84 и 85, комбинация BETi и BETd с вирусом XVir-N-31 проявляла синергизм в обеих клеточных линиях и приводила к сильному снижению выживаемости клеток.
Во втором аспекте влияние BETi и BETd на вирусную репликацию ДНК волокон в клеточных линиях UMUC-3 и RT112 анализировали с помощью qPCR.
3×104 клеток UMUC-3 и 4×104 клеток RT112 высевали в 12-луночные планшеты в среде DMEM и RPMI. На следующий день клетки предварительно обрабатывали BETi и BETd. Через 48 часов после посева клетки инфицировали 10 MOI для UMUC-3 и 50 MOI для RT112 и обрабатывали указанными концентрациями BETi и BETd. После каждой временной точки (4, 24 часа) клетки один раз промывали PBS и добавляли 200 мкл SDS буфера для лизиса ДНК. Клетки инкубировали с протеиназой К перед очисткой фенол-хлороформ-изоамиловым спиртом. ДНК разбавляли водой, не содержащей ДНКазы, и определяли концентрацию ДНК с помощью Nanodrop. Для каждого образца готовили раствор 15 нг/мкл и проводили RT-qPCR с использованием 75 нг ДНК и смеси GoTaq Master. Использовались прямые и обратные праймеры для конкретных волокон, а также для расчета значения Δct использовались специфические для актина праймеры. Для каждого образца измеряли 3 значения волокна и 3 значения актина. Расчет был выполнен в Excel, и каждые 24 часа рассчитывались на основе его значения за 4 часа (начальное проникновение вируса до репликации).
Результаты показаны на фиг. 86 и 87.
Как видно на фиг. 86 и 87, обработка BETi и BETd сильно увеличивала репликацию XVir-N-31 через 24 часа после инфицирования. В обеих клеточных линиях увеличение репликации вируса было самым высоким при комбинации с двухвалентным BETi AZD5153, что указывает на то, что двухвалентные ингибиторы в целом более подходят для комбинации с XVir-N-31.
Пример 35: Комбинация ингибитора CDK 4/6 рибоциклиба с XVir-N-31 на модели голых мышей с ксенотрансплантатом саркомы
Методы
Исследования на животных
Для модели ксенотрансплантата саркомы человека мышам в возрасте 10-20 недель подкожно (п/к) в правый бок вводили 3×106 опухолевых клеток A673 в PBS. Размер опухоли измеряли каждые 2-3 дня, а объем опухоли рассчитывали по формуле: объем=0,5×длина×ширина2. После того, как объем опухоли превышал 100-150 мм3, мышей рандомизировали в указанные лечебные/контрольные группы: PBS (то есть 0,5% метилцеллюлоза без LEE011 и PBS внутри опухоли [i.t.]), LEE (то есть 0,5% метилцеллюлоза, содержащая LEE011 и PBS i.t.), только XVir (то есть 0,5% метилцеллюлозы без LEE011 и XVir-N-31) и комбо (т.е. 0,5% метилцеллюлозы, содержащей LEE011 и XVir-N-31). Затем соответствующие животные получали сукцинат рибоциклиба (LEE011) в дозе 200 мг/кг массы тела (растворенный в 0,5% метилцеллюлозе) в день X (DX) до дня X+4 (DX+4) или имитация контроля (0,5% метилцеллюлоза без LEE011) через пероральный желудочный зонд. 1×1011 VP XVir-N-31 или PBS (в 50 соответственно) вводили внутрибрюшинно. в день X+1 (DX+1) и день X+3 (DX+3). В день X+5 3 репрезентативных животных из групп лечения только XVir и комбинированного лечения были умерщвлены для гистопатологической оценки и количественного определения репликации вируса в эксплантированных опухолях. Размер опухоли оставшихся мышей измеряли до тех пор, пока объем опухоли не превышал 1000 мм3, и мышей умерщвляли.
Экспериментальный план исследований на животных показан на фиг. 88
Статистический анализ
Рост опухоли in vivo анализировали с помощью веб-инструмента открытого доступа TumGrowth (https://kroemerlab.shinyapps.io/TumGrowth). Вкратце, измеренные данные об объемах опухоли были подвергнуты линейному моделированию смешанного эффекта, что позволило сравнить продольный наклон роста опухоли, а также оценить ответ на лечение в желаемые моменты времени (анализ поперечного сечения). Значения P были рассчитаны с помощью программного обеспечения с использованием ANOVA типа II и выбранных попарных сравнений для продольного и поперечного анализов (с поправкой Холма, когда это указано). Кривые роста опухоли были созданы с помощью Prism 5 (GraphPad Software, San Diego, CA, USA). Кривые роста опухоли были построены с использованием среднего объема опухоли и стандартной ошибки среднего (sem). Значения P<0,05 считались статистически значимыми (*p<0,05; **p<0,005; ***p<0,0005).
Результаты - Эффект комбинированной терапии и монотерапии на модели голых мышей с ксенотрансплантатами
Чтобы выяснить, могут ли биологически значимые преимущества предложенной комбинированной стратегии также трансформироваться в возможные терапевтические преимущества, контроль над опухолью ксенотрансплантированных клеток саркомы A673 оценивали у бестимусных мышей с ослабленным иммунитетом. Применяя веб-инструмент открытого доступа TumGrowth, контроль роста опухоли был значительно улучшен у животных, получавших комбинированное лечение (combo), по сравнению с животными, получавшими монотерапию (только LEE или XVir) или контроль (PBS). Максимальные различия в ответе на лечение между группами наблюдались на 12-21 день после начала терапии. (Следует отметить, что репликация вируса была значительно увеличена в эксплантированных опухолях репрезентативных животных, получавших комбо, по сравнению сXVir-N-31.)
Результаты показаны на фиг. 89, 90 и 91. Выбранные попарные сравнения для продольного анализа показаны в следующей табл.:
Результаты, представленные на Фиг. 29, 30 и 31 ясно демонстрируют, что комбинированная терапия, включающая XVir-N-31 и рибоциклиб, является более эффективной, чем только XVir-N-31 или рибоциклиб.
Признаки изобретения, раскрытые в предыдущем описании, формуле изобретения, а также на чертежах, могут быть важны как по отдельности, так и в любой комбинации для реализации изобретения в его различных вариантах осуществления.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Klinikum rechts der Isar der Technischen Universität
München
<120> ЛЕЧЕНИЕ ОПУХОЛЕЙ КОМБИНАЦИЕЙ ОНКОЛИТИЧЕСКОГО АДЕНОВИРУСА, ИНГИБИТОРА CDK4/6 И ДОПОЛНИТЕЛЬНОГО ТЕРАПЕВТИЧЕСКОГО АГЕНТА ve agent
<130> H 10073 PCT
<150> EP 19 000 406.9
<151> 2019-09-12
<150> EP 19 000 396.2
<151> 2019-09-05
<160> 54
<170> PatentIn version 3.5
<210> 1
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 1
aagctagccc tgcaaacatc acccaagcta ccagtggcag ta 42
<210> 2
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 2
ccgtcatctc tacagcccat gggctttgtc agagtcttgc 40
<210> 3
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 3
cttcctagcg actttgtgcc gtcagagtgg taggcaaggt 40
<210> 4
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 4
cgacgaggat gaagtcctgt gtctgctcag gatagcaggc gccat 45
<210> 5
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 5
gaggatgaag tcctgtgtct caggatagca ggcgccat 38
<210> 6
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 6
tgtttgtcta cagtcctgtg tctgctcagg atagcaggcg ccat 44
<210> 7
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 7
ttgtctacag tcctgtgtct caggatagca ggcgccat 38
<210> 8
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 8
tccctcccaa cacacagagt gacaggaaac cgtgtggaat 40
<210> 9
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 9
agcaaccctc ctaaaccact tgtttgaggt atccatgcta tca 43
<210> 10
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 10
acgctatgag acctcactga atcctgggtc aacccctcaa g 41
<210> 11
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 11
cgtccctgag ttcccaaccg cgaagtgtca taccgagtct t 41
<210> 12
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 12
tggcatggac tgtggtcatg agactggcgt cttcaccacc atgg 44
<210> 13
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 13
taagtaggtg cacagtaggt ctgaaaagtg caaagaacac ggctaag 47
<210> 14
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
gaaccagggc cgcccatact ggggctttgt cagagtcttg c 41
<210> 15
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
ccgttagccc aagagcaacc ggccgtgatg gtagagaag 39
<210> 16
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
ctgtggtact tcccagagac caggtgagtt ataccctgcc 40
<210> 17
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 17
aagctagccc tgcaaacatc a 21
<210> 18
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 18
cccaagctac cagtggcagt a 21
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 19
ccgtcatctc tacagcccat 20
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 20
gggctttgtc agagtcttgc 20
<210> 21
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 21
aagctagccc tgcaaacatc a 21
<210> 22
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 22
cccaagctac cagtggcagt a 21
<210> 23
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 23
tcacccacac tgtgcccatc tacg 24
<210> 24
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 24
cagcggaacc gctcattgcc aatgg 25
<210> 25
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 25
catcccagga ggtcacttct g 21
<210> 26
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 26
gacaacagcg gttcttgctc 20
<210> 27
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 27
uucuccgaac gugucacgut t 21
<210> 28
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 28
acgugacacg uucggagaat t 21
<210> 29
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 29
cugaggaguu caucagccut t 21
<210> 30
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 30
aggcugauga acuccucagt t 21
<210> 31
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 31
taagtaggtg cacagtaggt ctg 23
<210> 32
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 32
aaagtgcaaa gaacacggct aag 23
<210> 33
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 33
aagctagccc tgcaaacatc a 21
<210> 34
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 34
cccaagctac cagtggcagt a 21
<210> 35
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 35
taagtaggtg cacagtaggt ctg 23
<210> 36
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 36
aaagtgcaaa gaacacggct aag 23
<210> 37
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 37
aagctagccc tgcaaacatc a 21
<210> 38
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 38
cccaagctac cagtggcagt a 21
<210> 39
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 39
cagagatcgt gtattgagat tctcgagaat ctcaatacac gatctctg 48
<210> 40
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 40
taagtaggtg cacagtaggt ctga 24
<210> 41
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 41
aaagtgcaaa gaacacggct aag 23
<210> 42
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 42
aagctagccc tgcaaacatc a 21
<210> 43
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 43
cccaagctac cagtggcagt a 21
<210> 44
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 44
agttttcgcg cttaaatttg agaaagggcg cgaaactagt cctt 44
<210> 45
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 45
aaggactagt ttcgcgccct ttctcaaatt taagcgcgaa aact 44
<210> 46
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 46
aggagtcgga tttaaatttg agaaagtaat aggtactagt cctt 44
<210> 47
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 47
aaggactagt acctattact ttctcaaatt taaatccgac tcct 44
<210> 48
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 48
atcgatctta cctgccacga ggcttttcca 30
<210> 49
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 49
tacaaaagca g 11
<210> 50
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 50
acaactccat ct 12
<210> 51
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 51
atcgattttc ca 12
<210> 52
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 52
tacaaaaagc a 11
<210> 53
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 53
acaacttgtg actgccgcgg agactgtttc tgcccatct 39
<210> 54
<211> 12
<212> белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 54
Thr Thr Cys Asp Cys Arg Gly Asp Cys Phe Cys Ser
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ ОПУХОЛЕЙ КОМБИНАЦИЕЙ ОНКОЛИТИЧЕСКОГО АДЕНОВИРУСА И ИНГИБИТОРА CDK4/6 | 2019 |
|
RU2811278C2 |
| ОНКОЛИТИЧЕСКИЕ АДЕНОВИРУСЫ, КОДИРУЮЩИЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, А ТАКЖЕ СПОСОБЫ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2016 |
|
RU2725799C2 |
| СПОСОБЫ ЛЕЧЕНИЯ AR+ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2017 |
|
RU2769527C2 |
| СПОСОБЫ ЛЕЧЕНИЯ УСТОЙЧИВОГО К ИНГИБИТОРАМ CDK4/6 РАКА | 2019 |
|
RU2820478C2 |
| АДЕНОВИРУС С ОБРАЩЕННОЙ ГЕННОЙ ЭКСПРЕССИЕЙ И ЕГО ПРИМЕНЕНИЕ | 2003 |
|
RU2393221C2 |
| ЛЕЧЕНИЕ РАКА ГОЛОВНОГО МОЗГА ОНКОЛИТИЧЕСКИМ АДЕНОВИРУСОМ | 2014 |
|
RU2689553C2 |
| РАСШИРЕННАЯ АДОПТИВНАЯ КЛЕТОЧНАЯ ТЕРАПИЯ | 2014 |
|
RU2703438C2 |
| АДЕНОВИРУС, СОДЕРЖАЩИЙ АЛЬБУМИН-СВЯЗЫВАЮЩИЙ УЧАСТОК | 2015 |
|
RU2711371C2 |
| ИММОРТАЛИЗОВАННЫЕ ЛИНИИ КЛЕТОК ПТИЦ ДЛЯ ПОЛУЧЕНИЯ ВИРУСОВ | 2004 |
|
RU2359999C2 |
| СПОСОБЫ И КОМБИНАЦИИ ИНГИБИТОРОВ KAT6 ДЛЯ ЛЕЧЕНИЯ РАКА | 2021 |
|
RU2833058C1 |
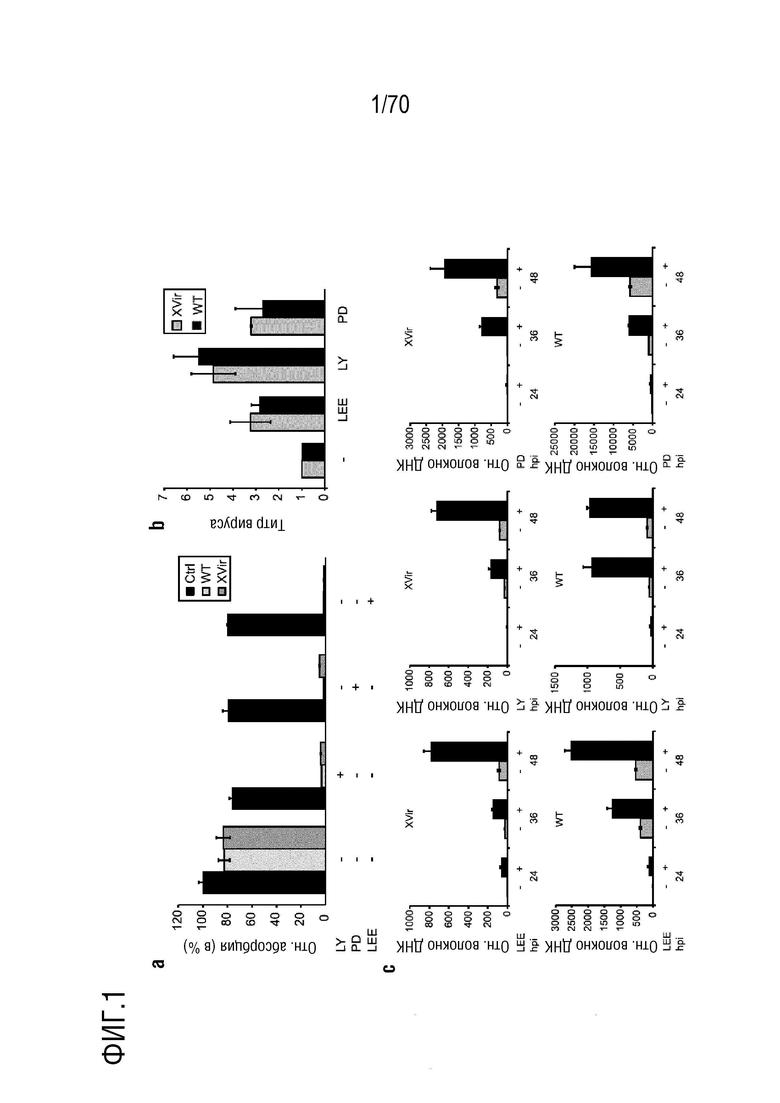

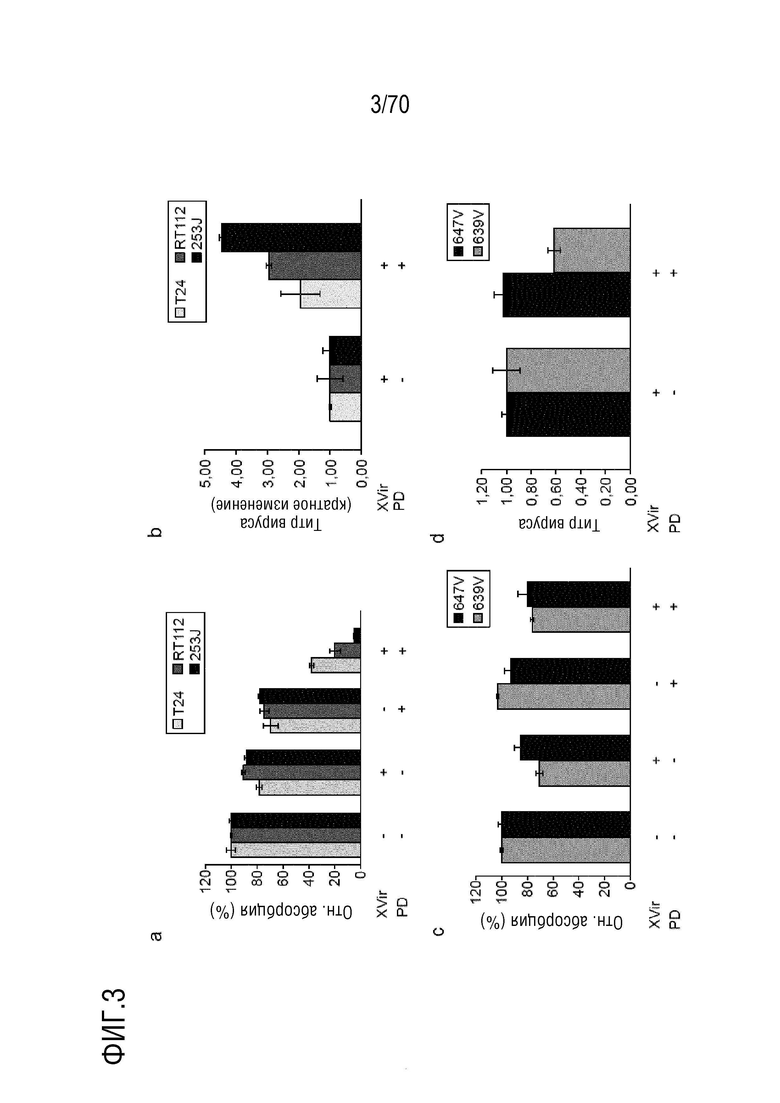
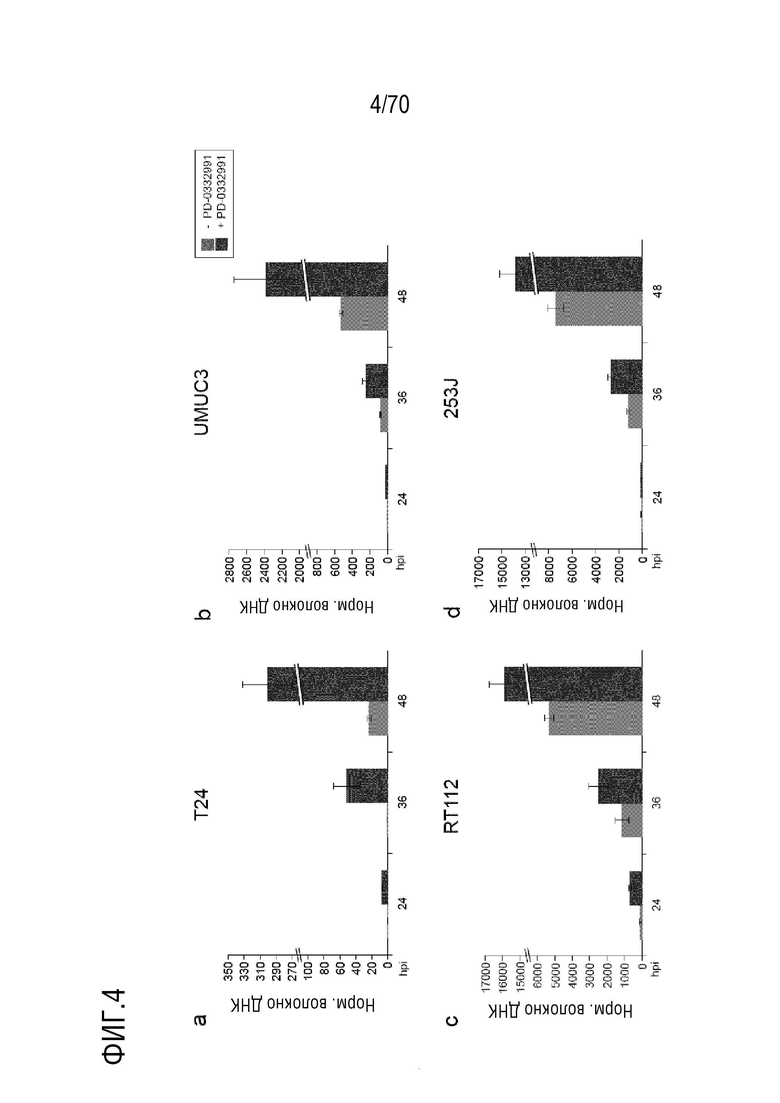
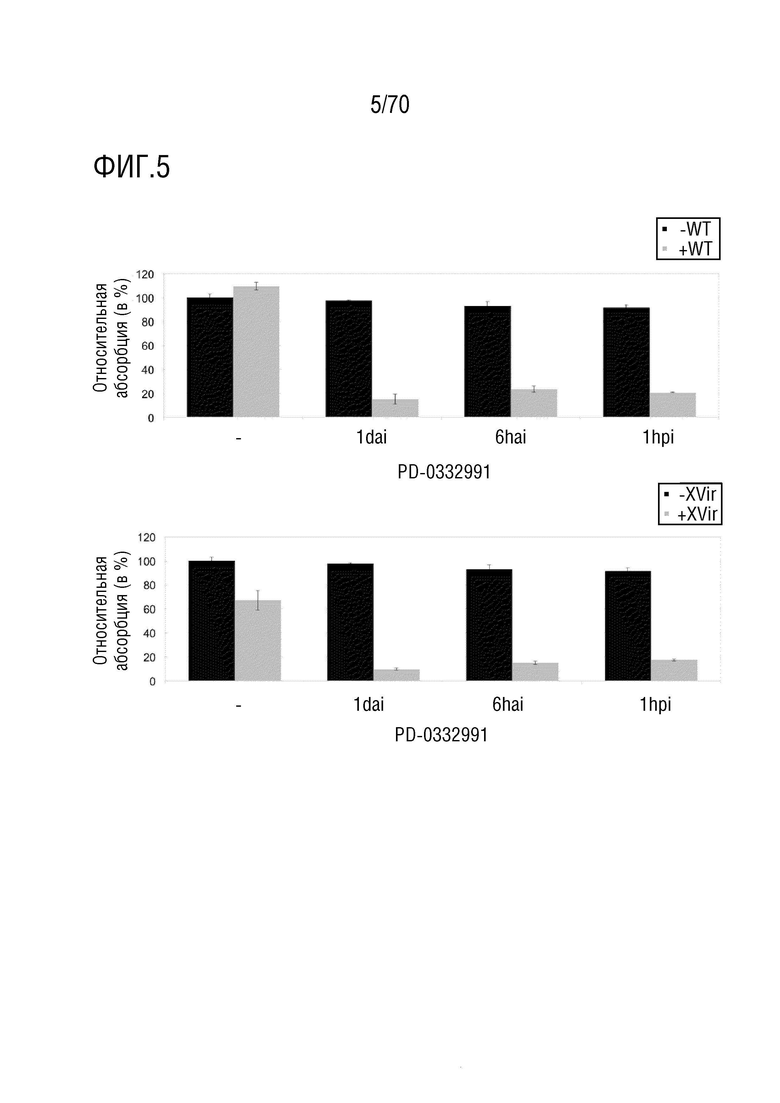
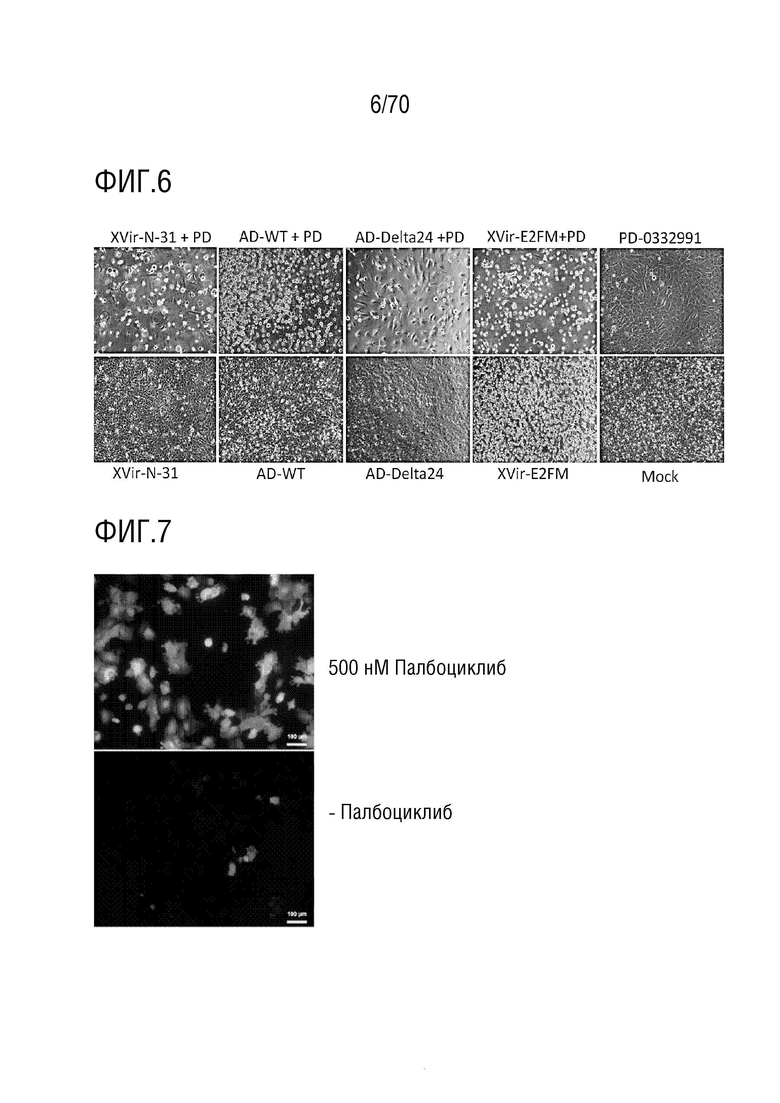
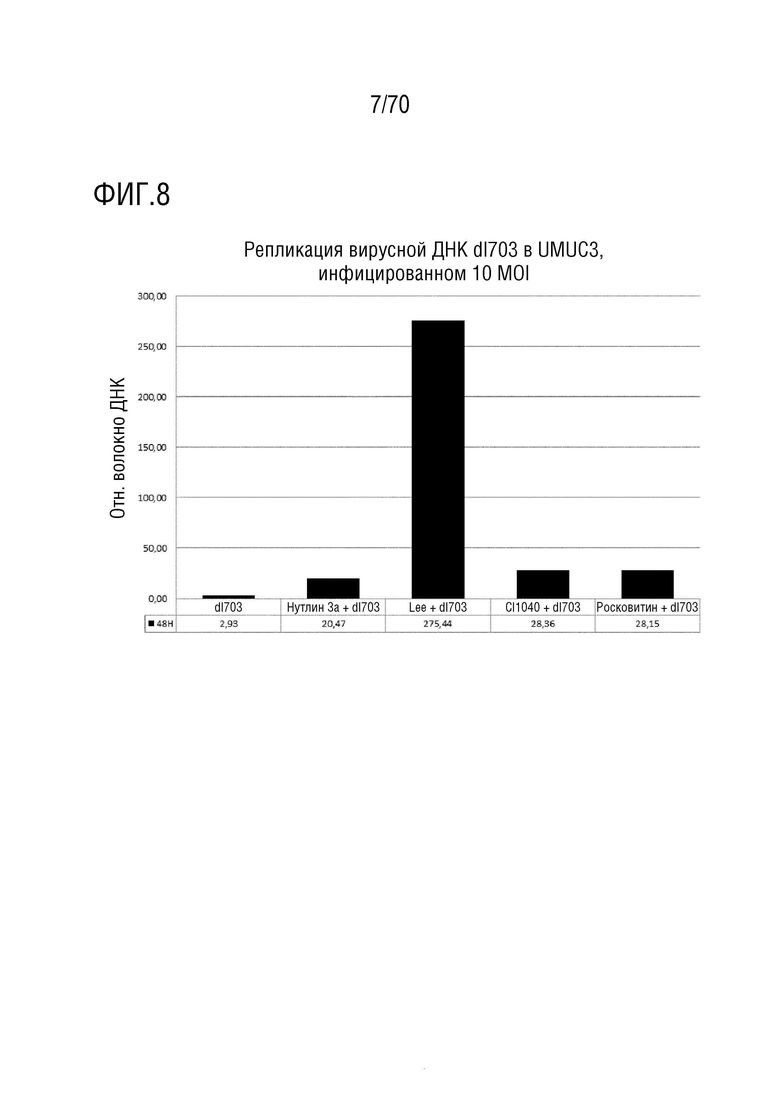
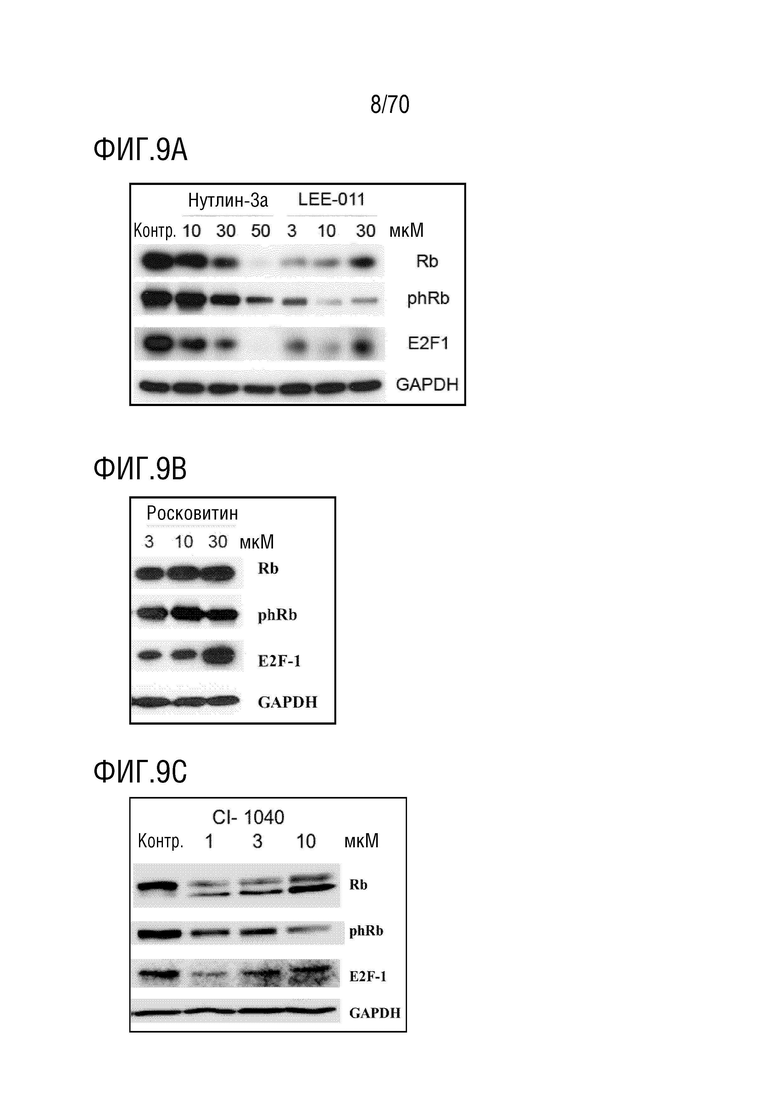
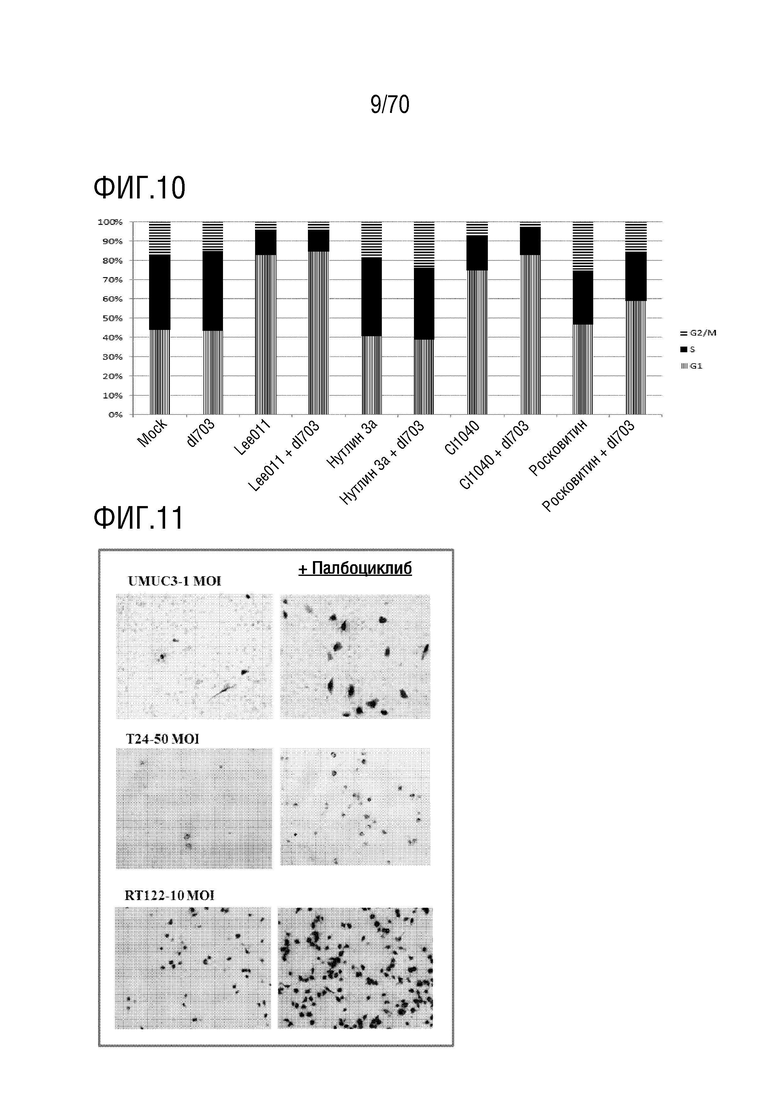
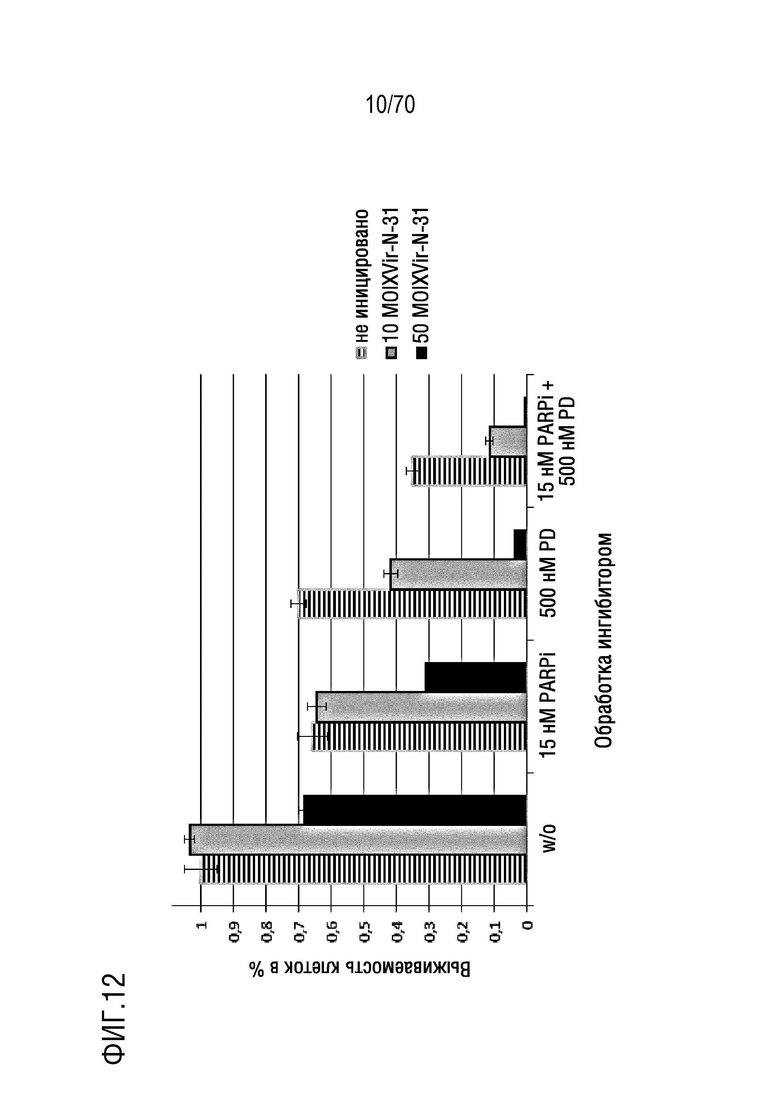
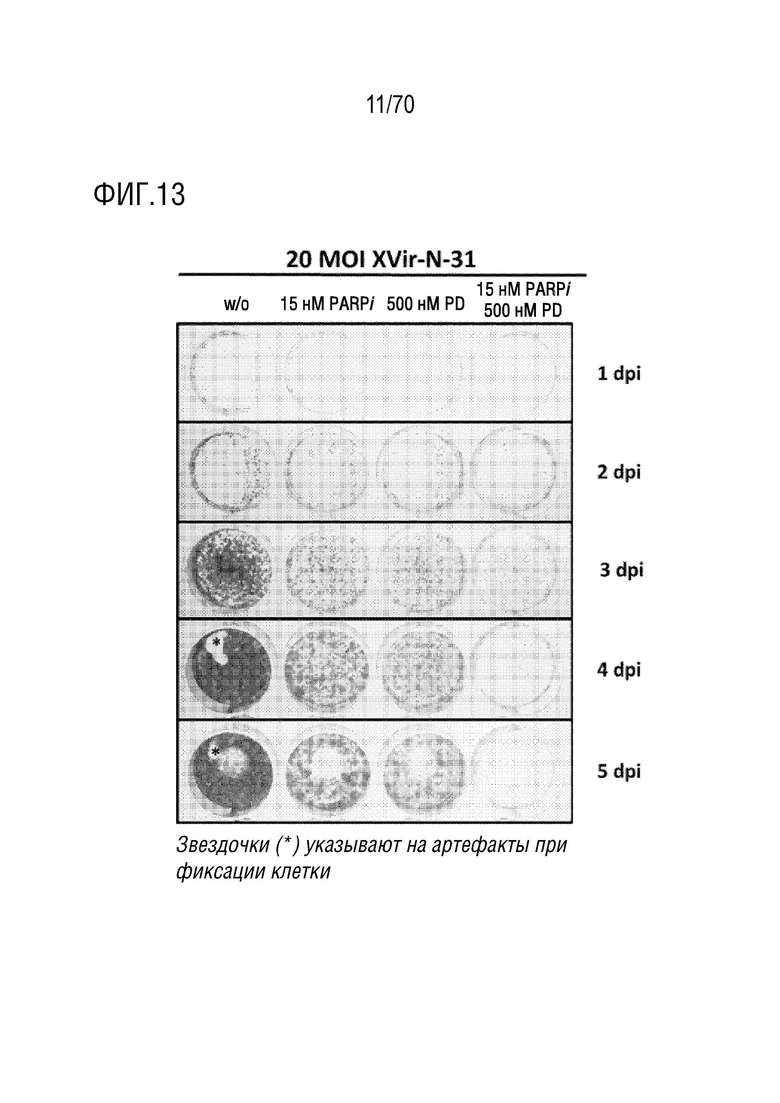
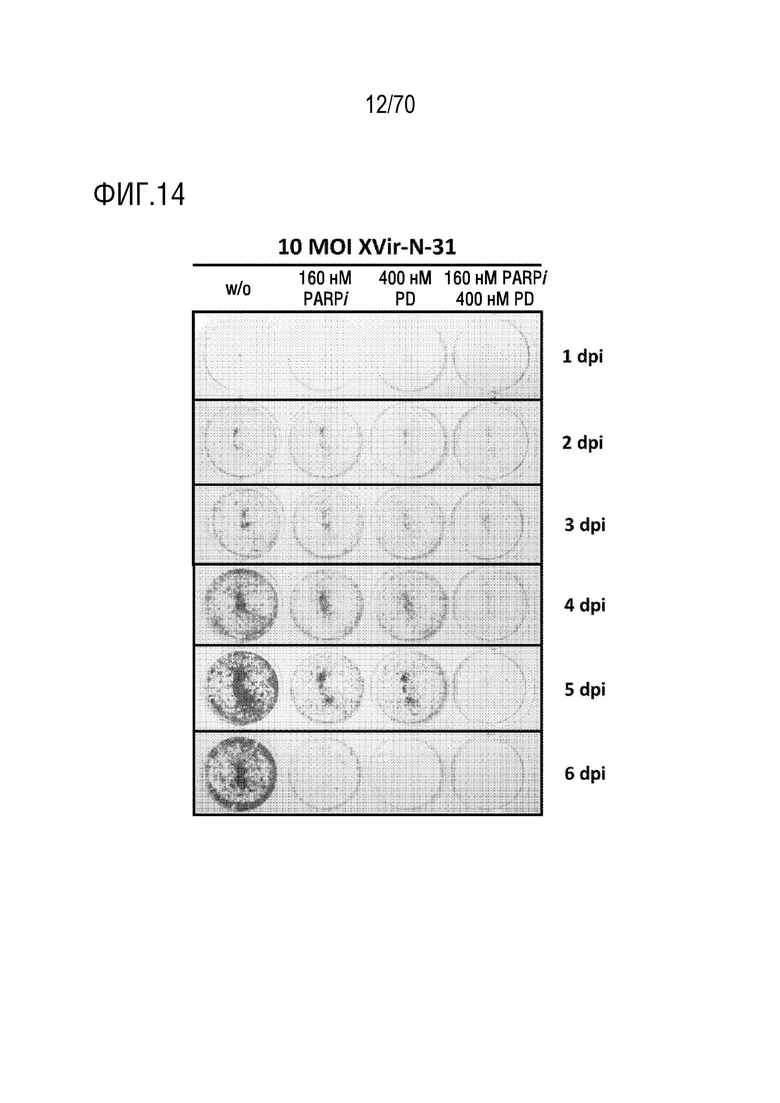
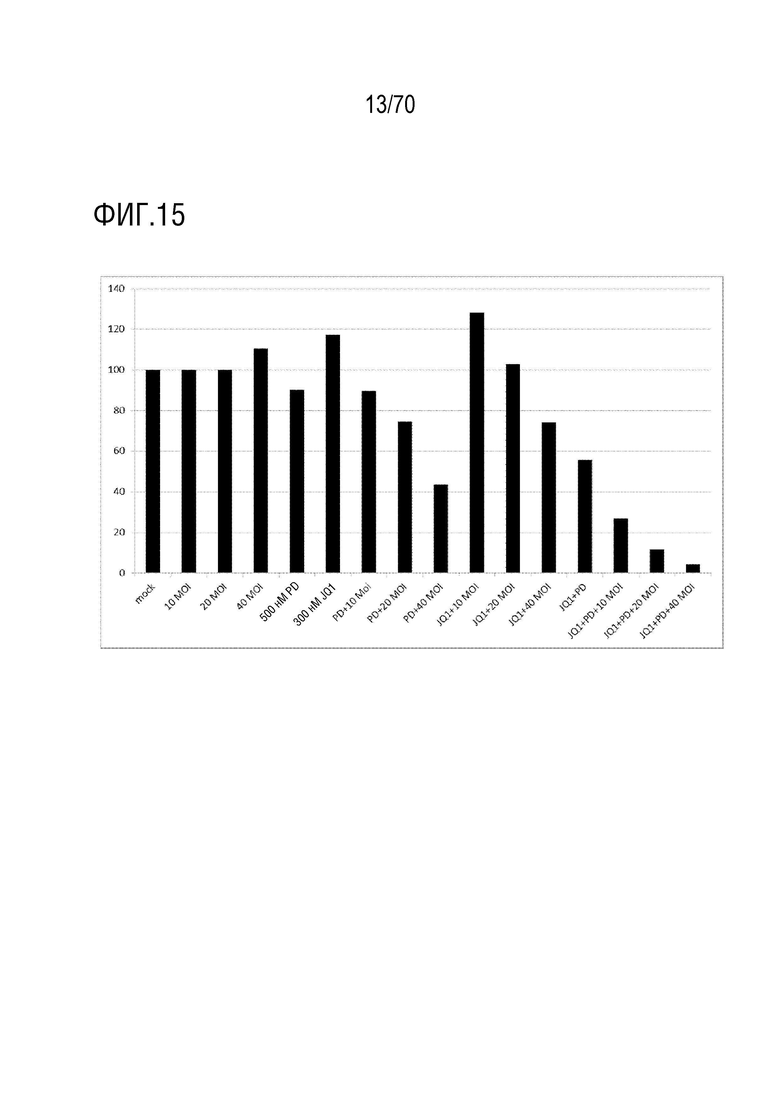
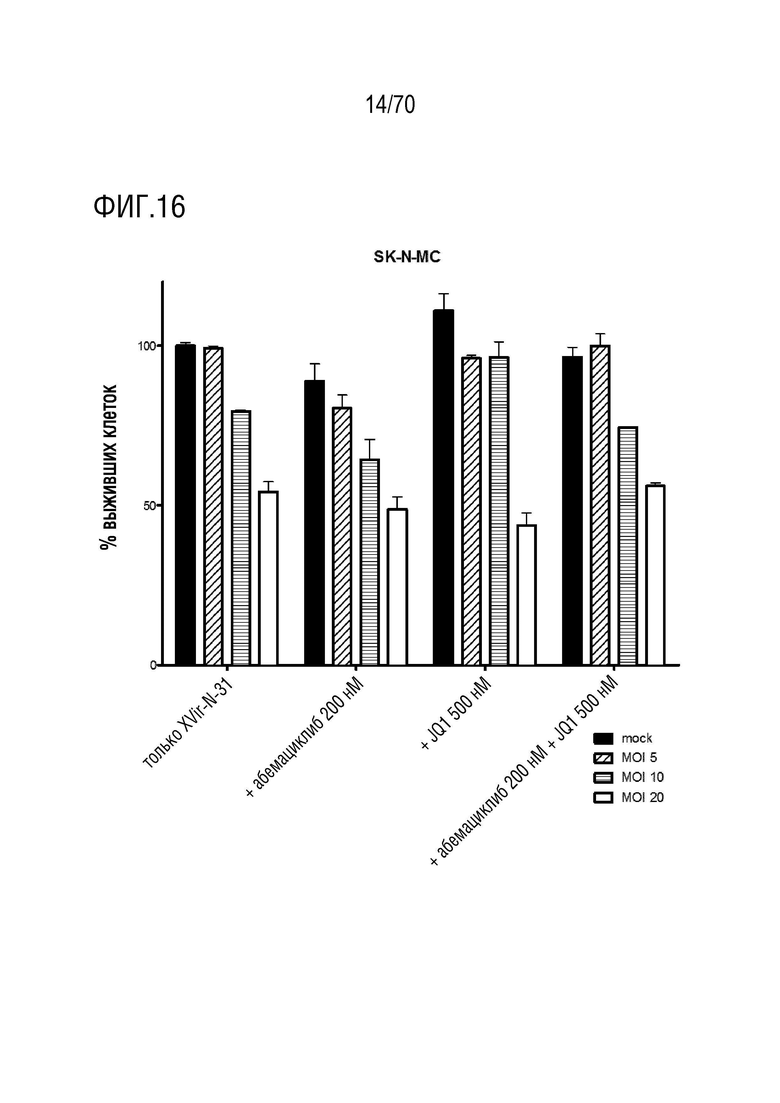
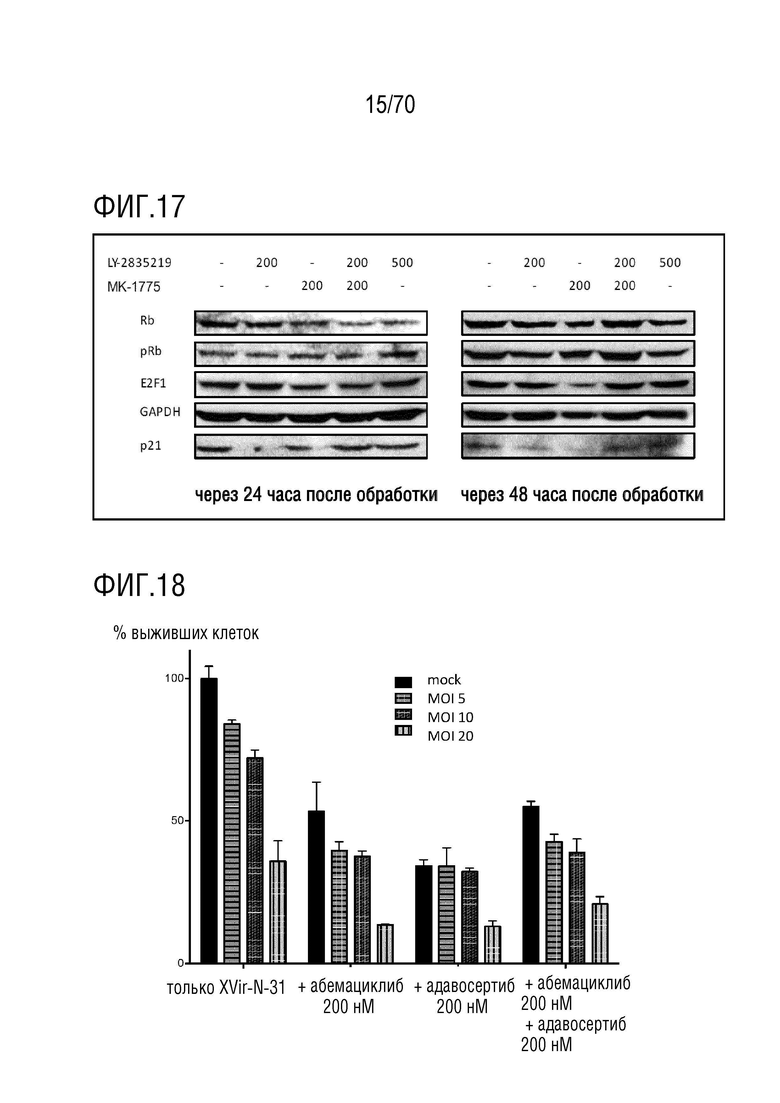
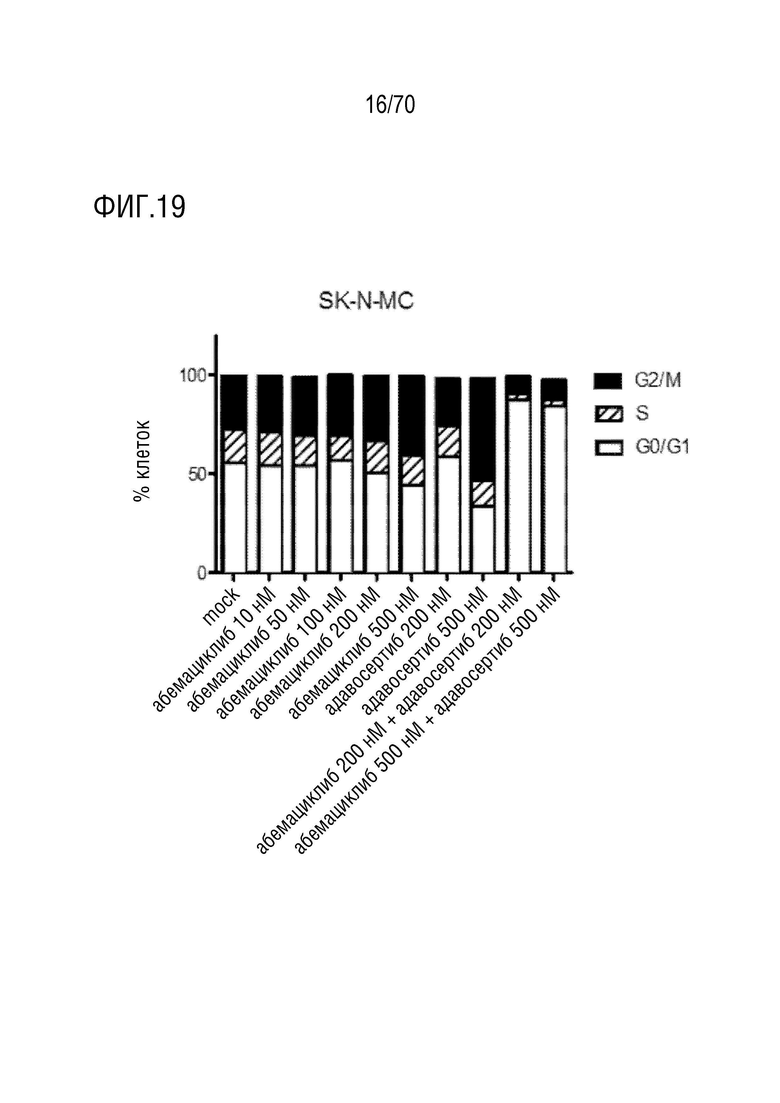
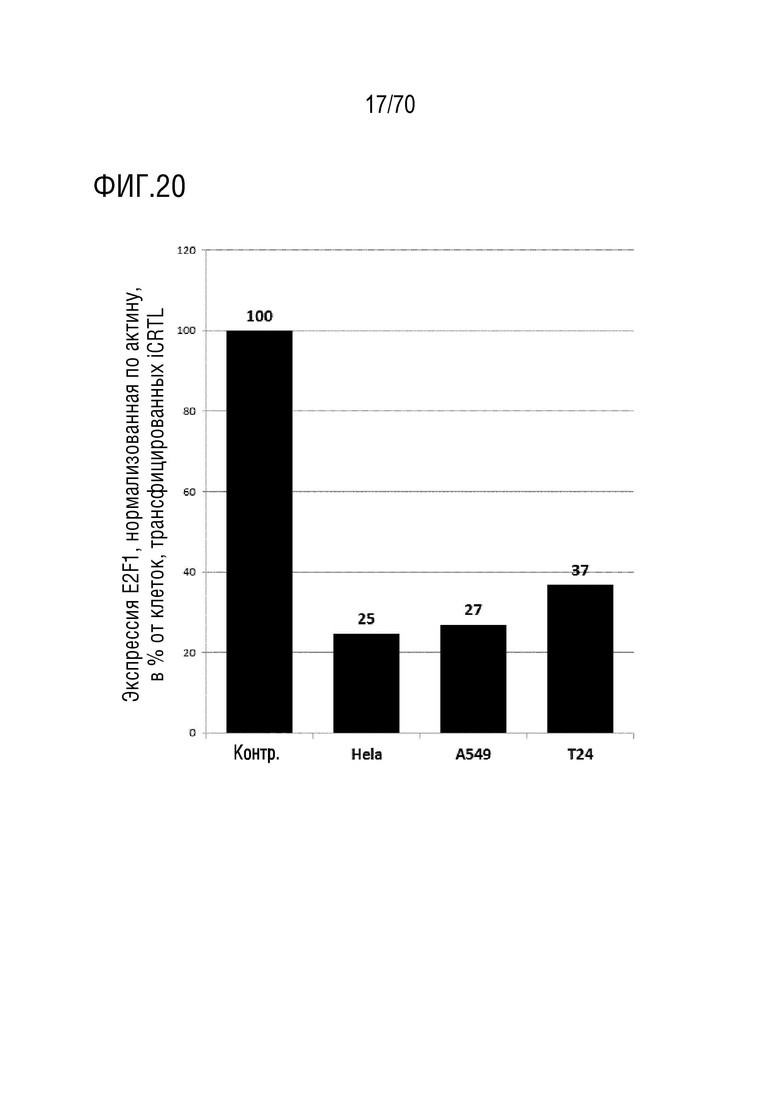
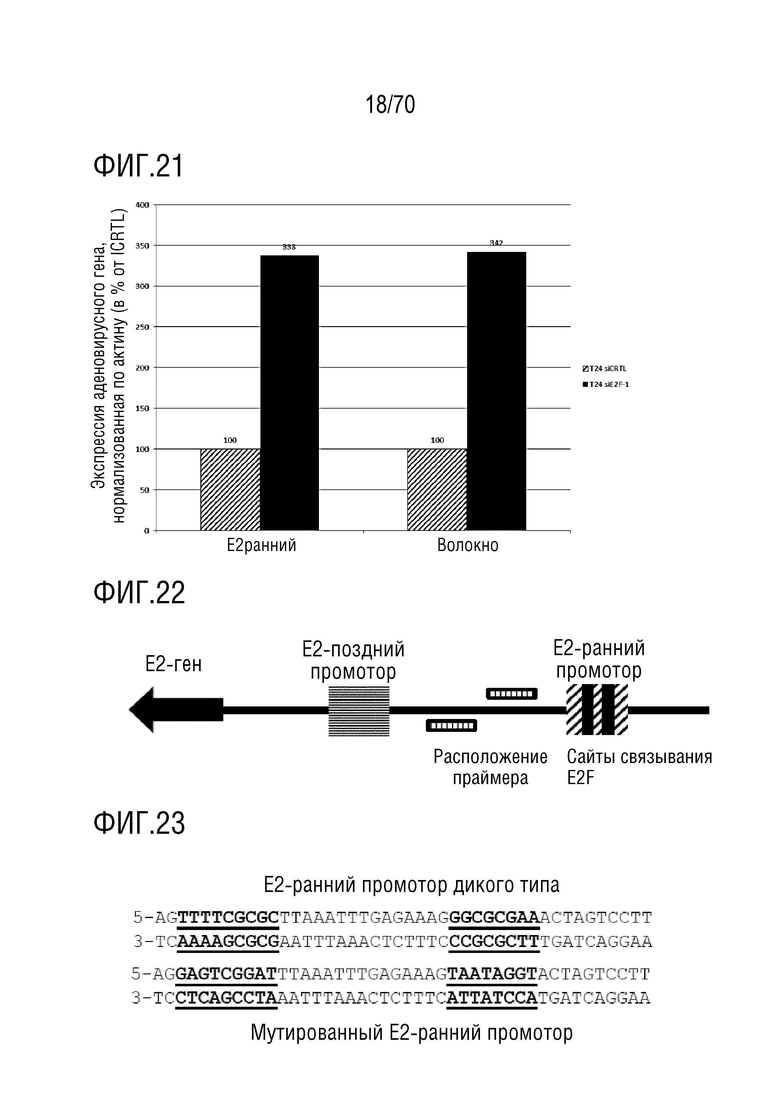
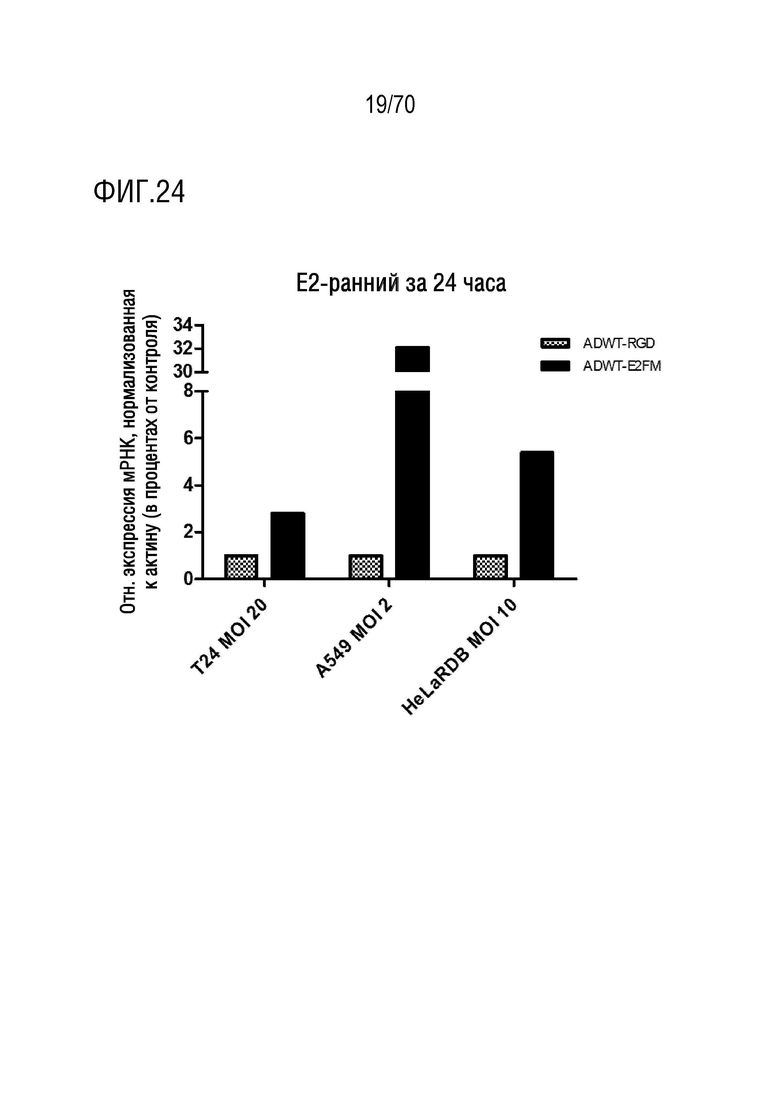
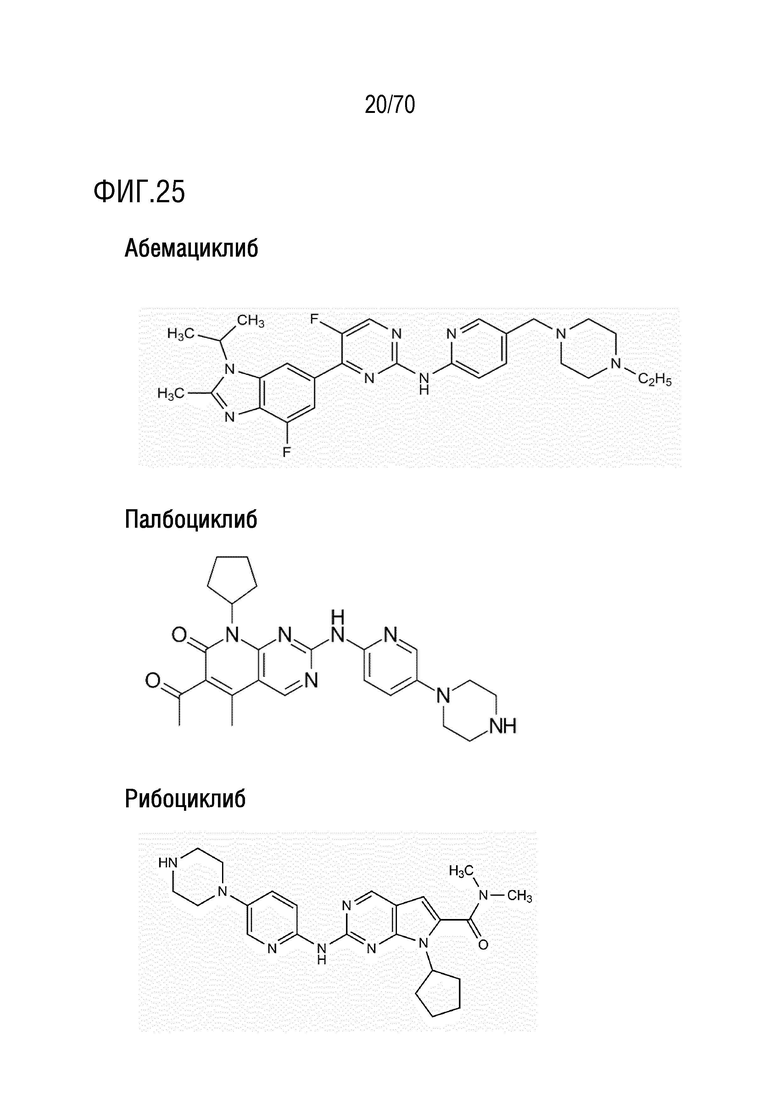
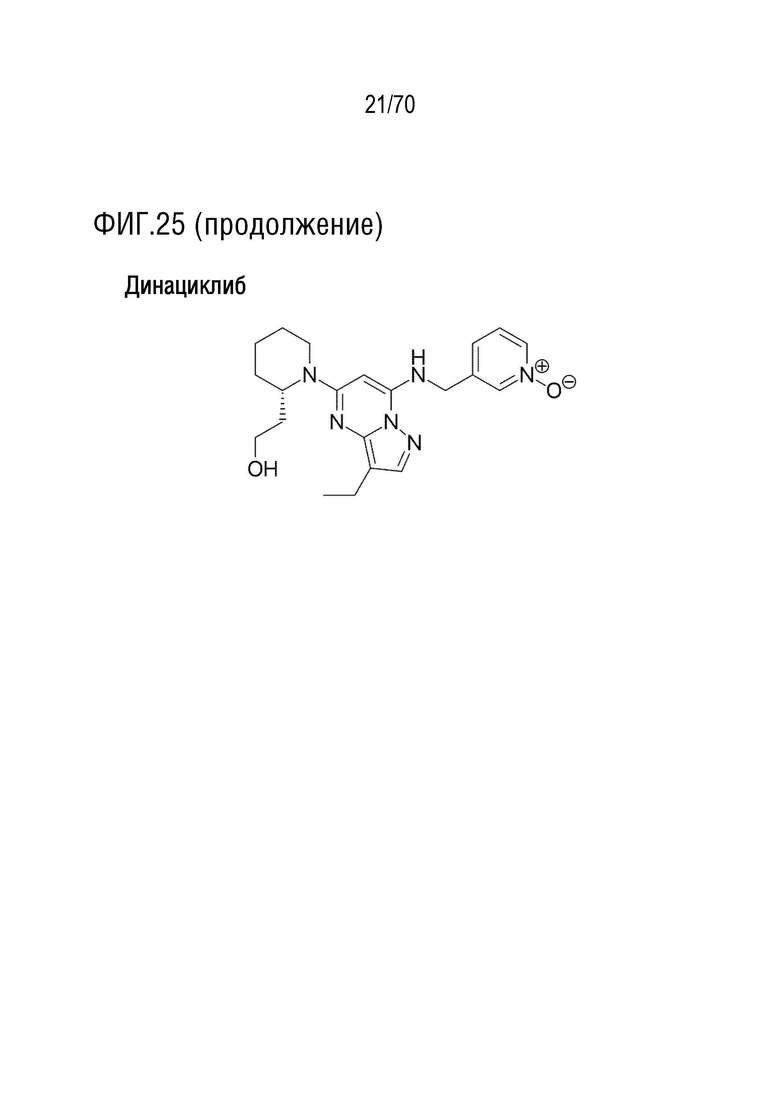
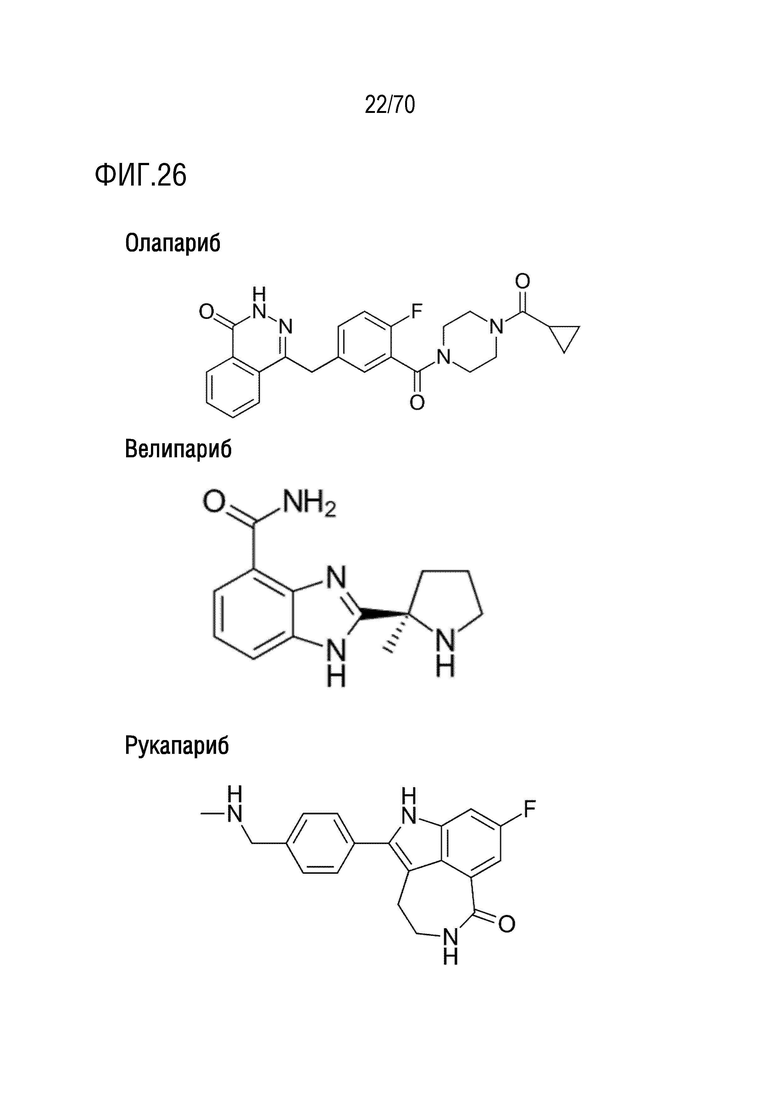
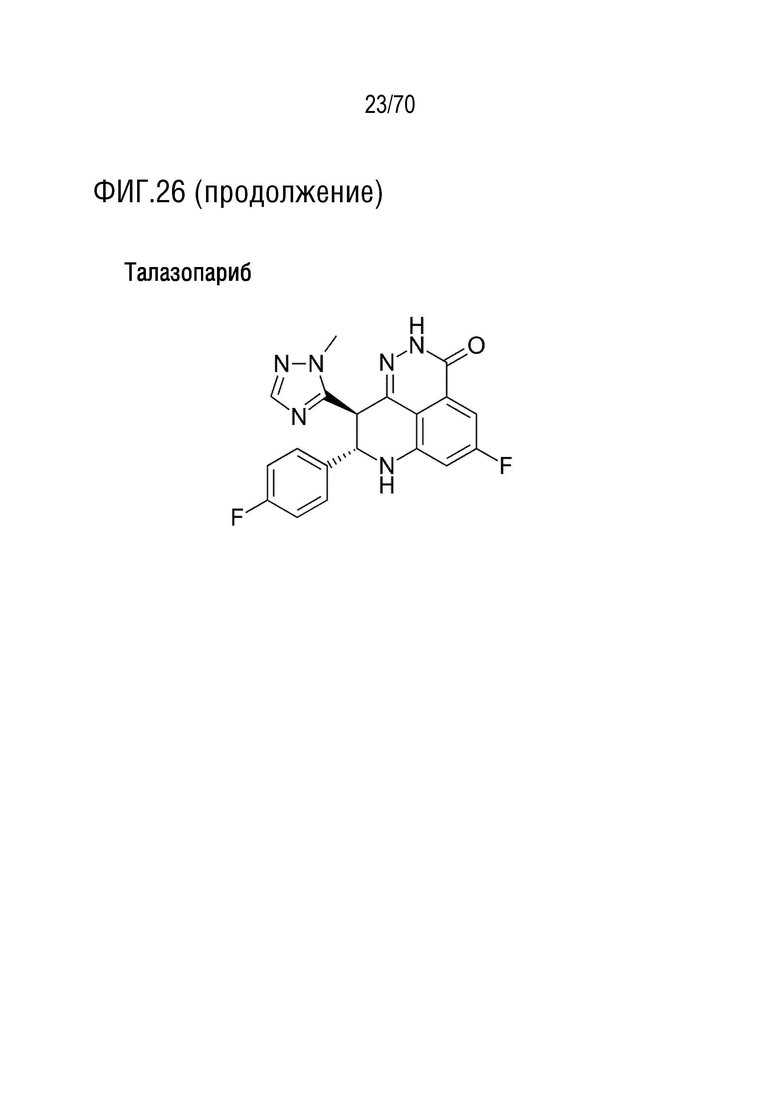
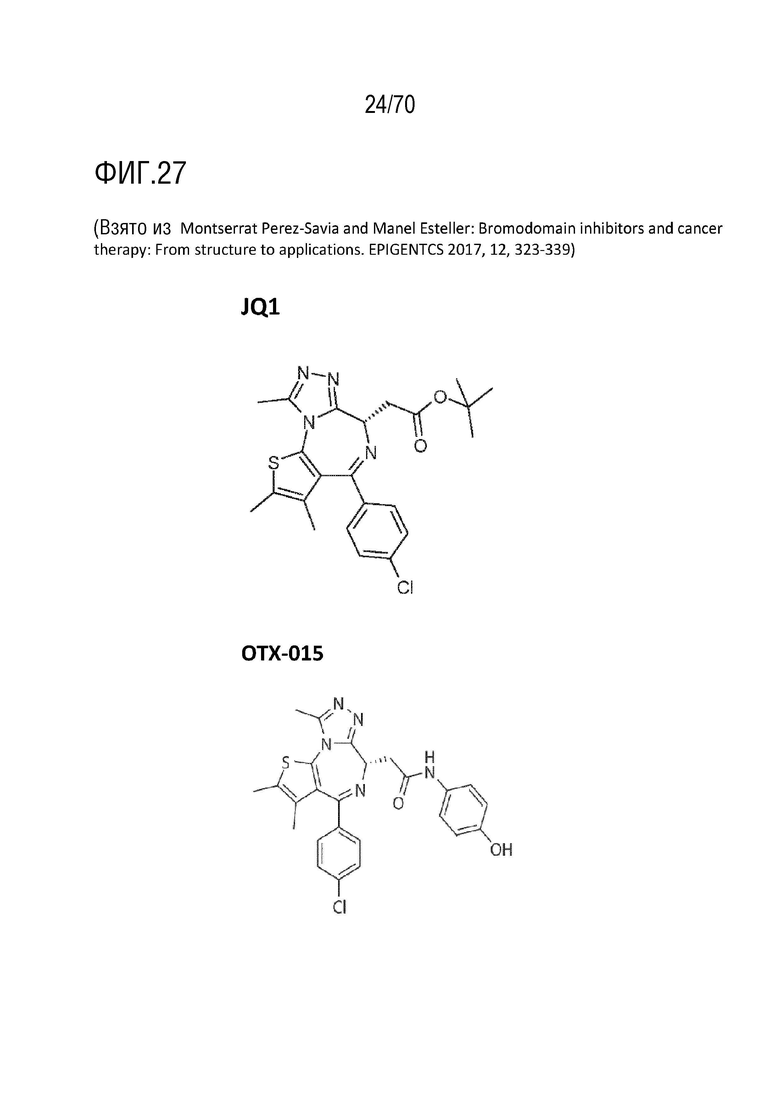
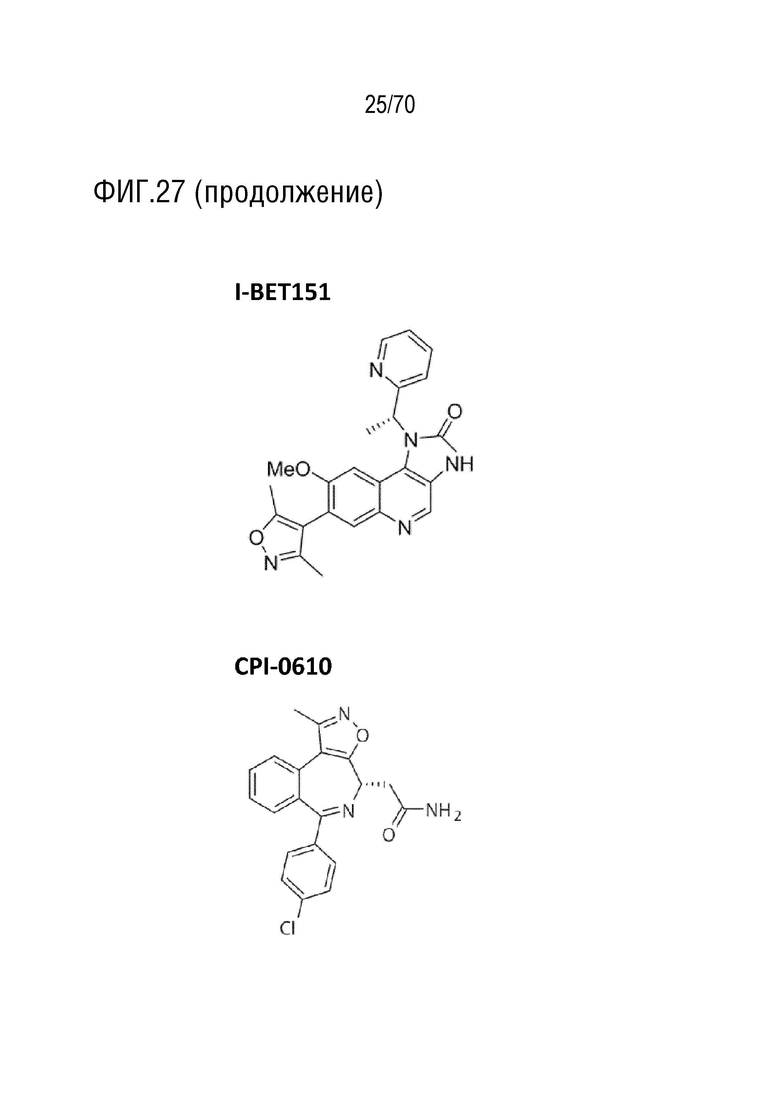
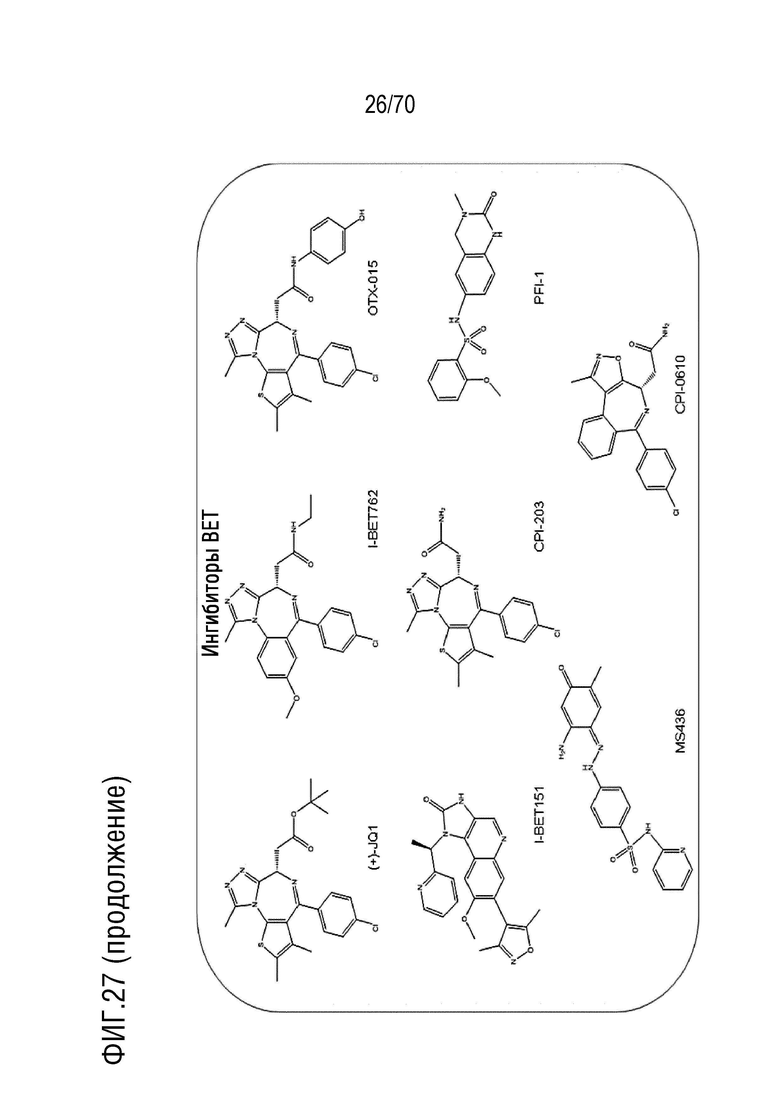
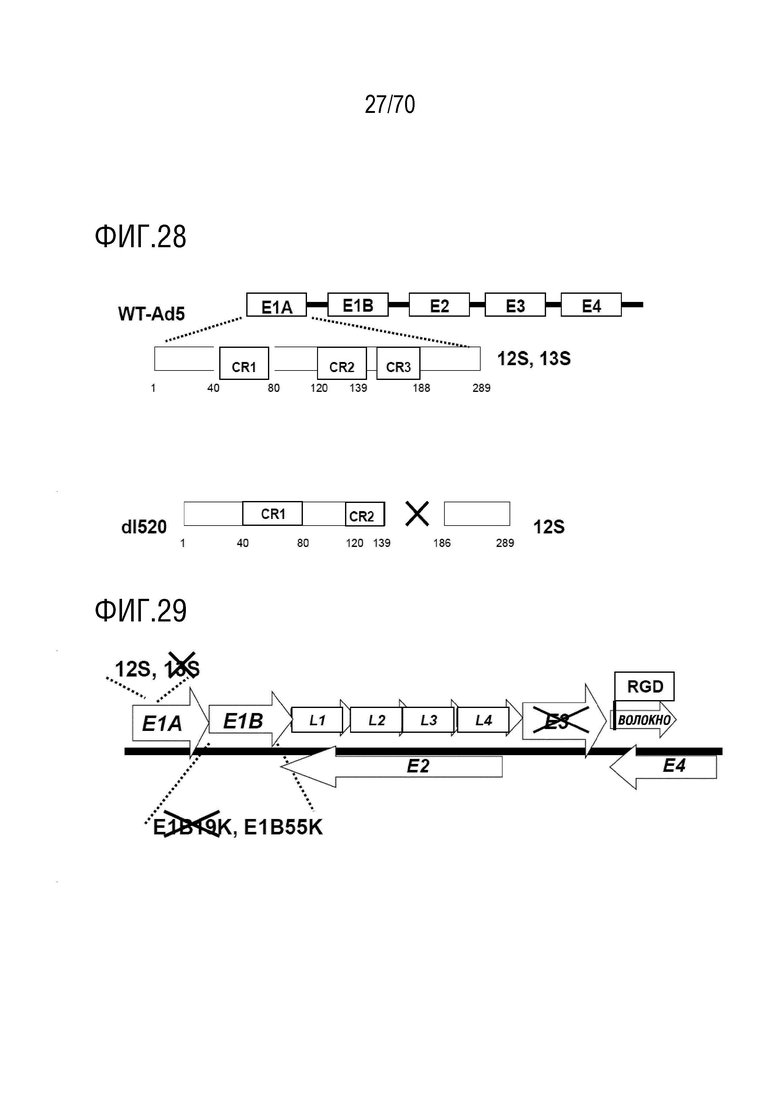
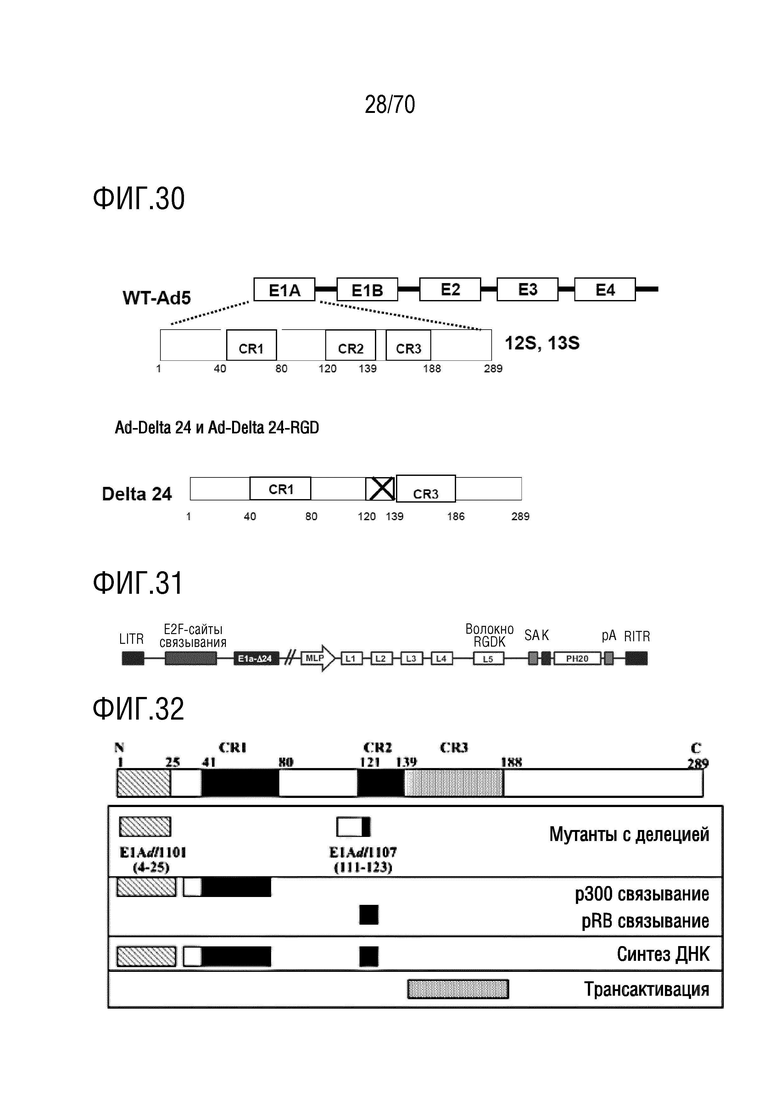
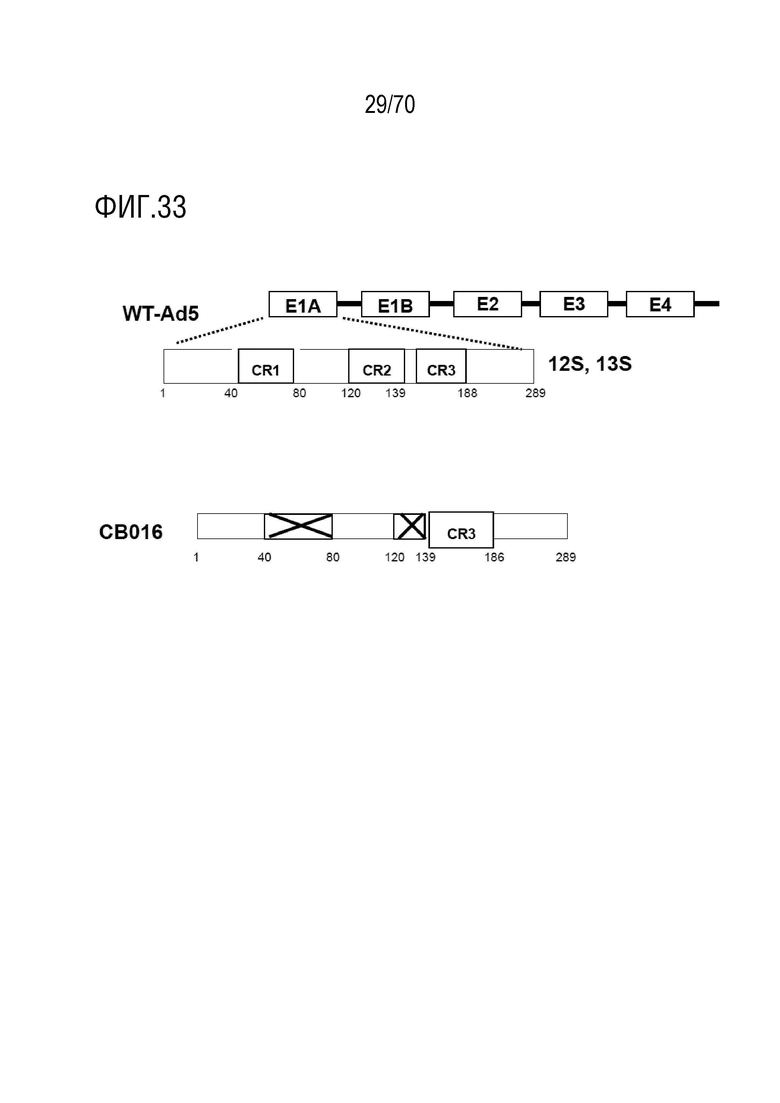
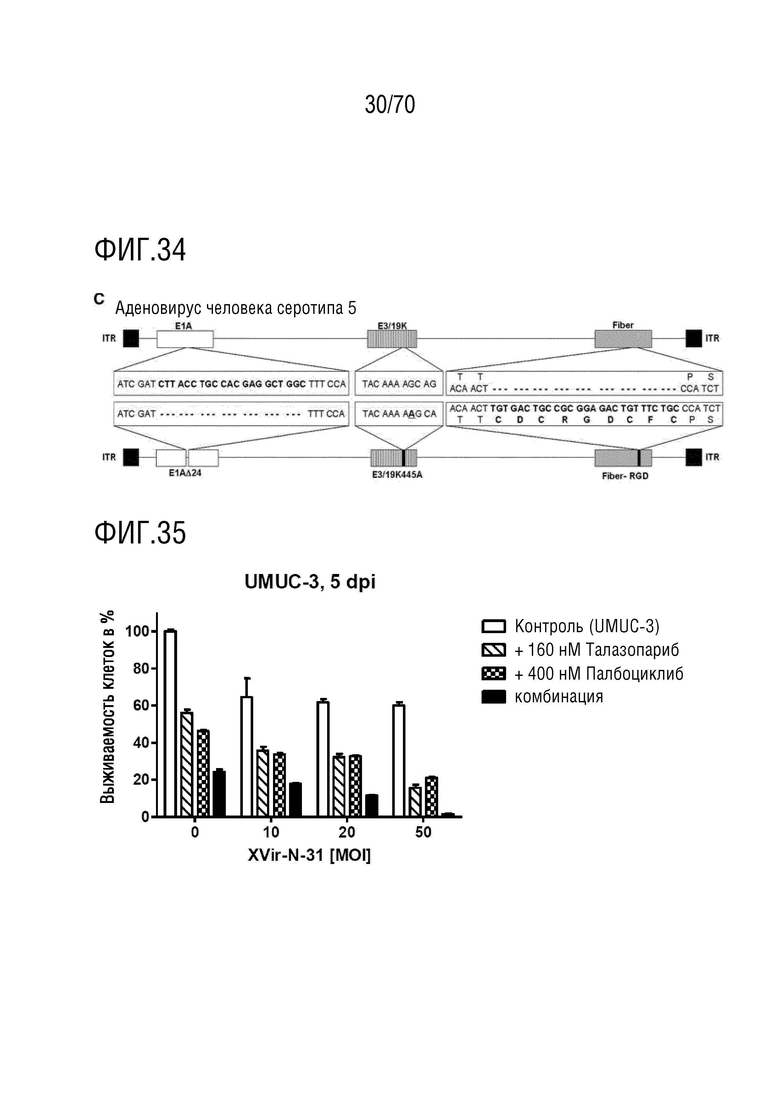
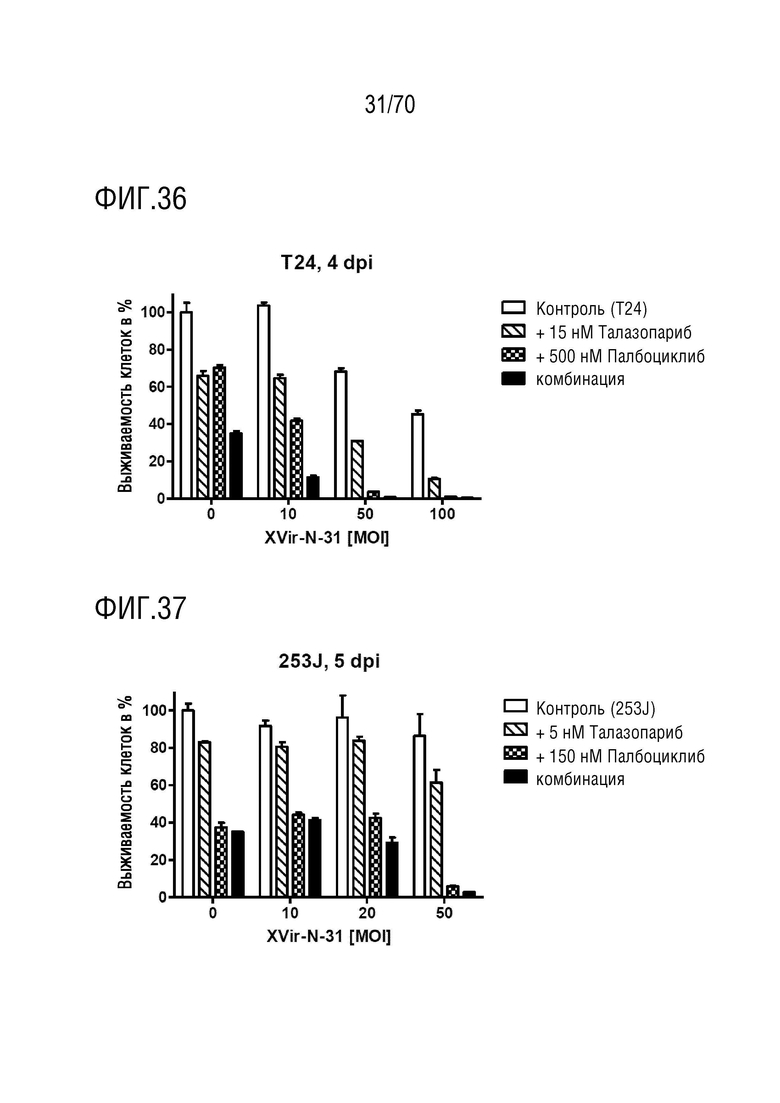
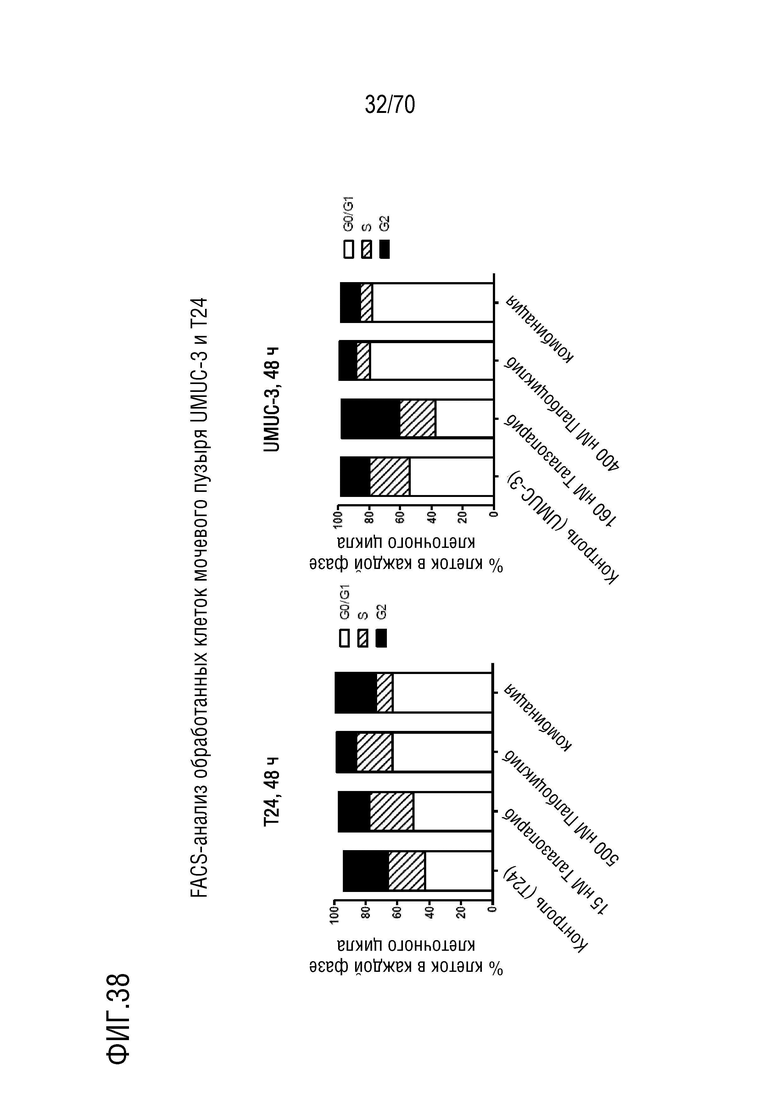
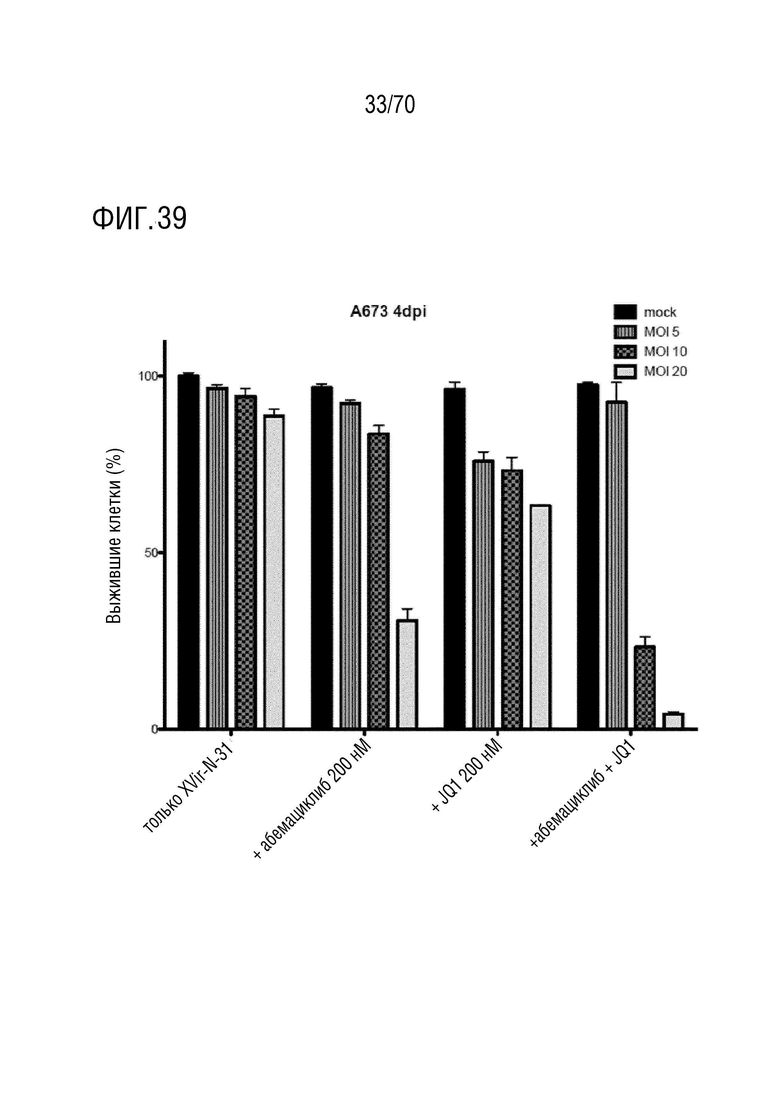
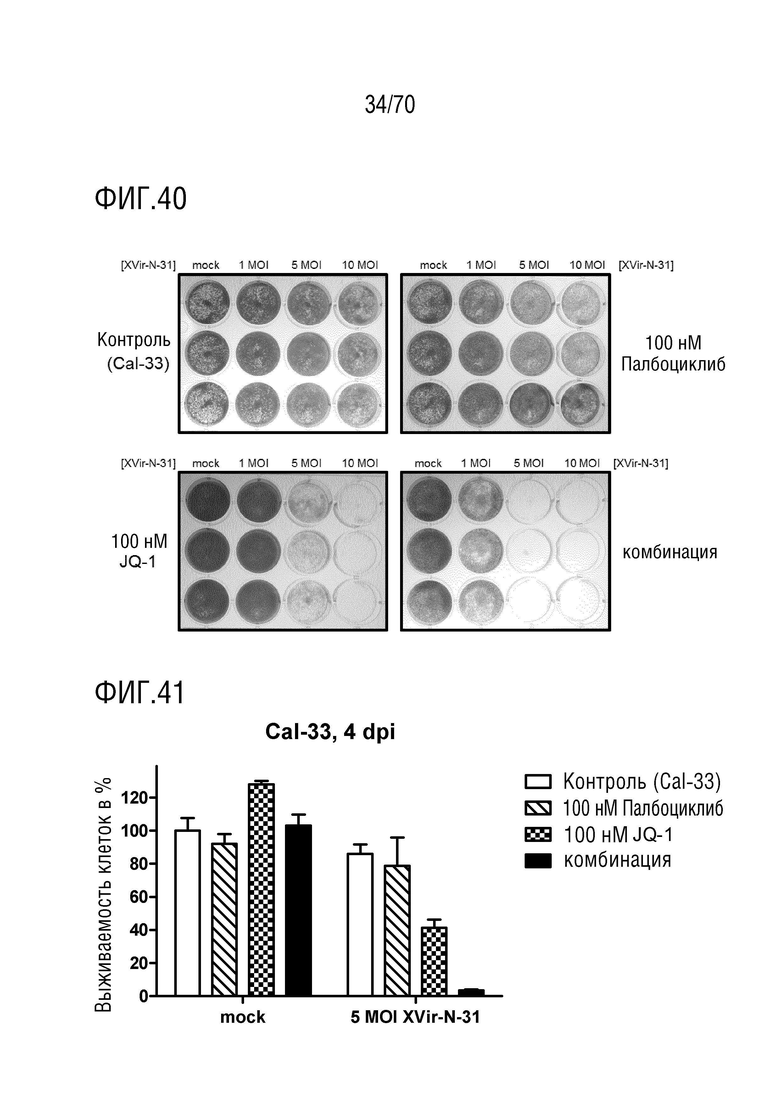
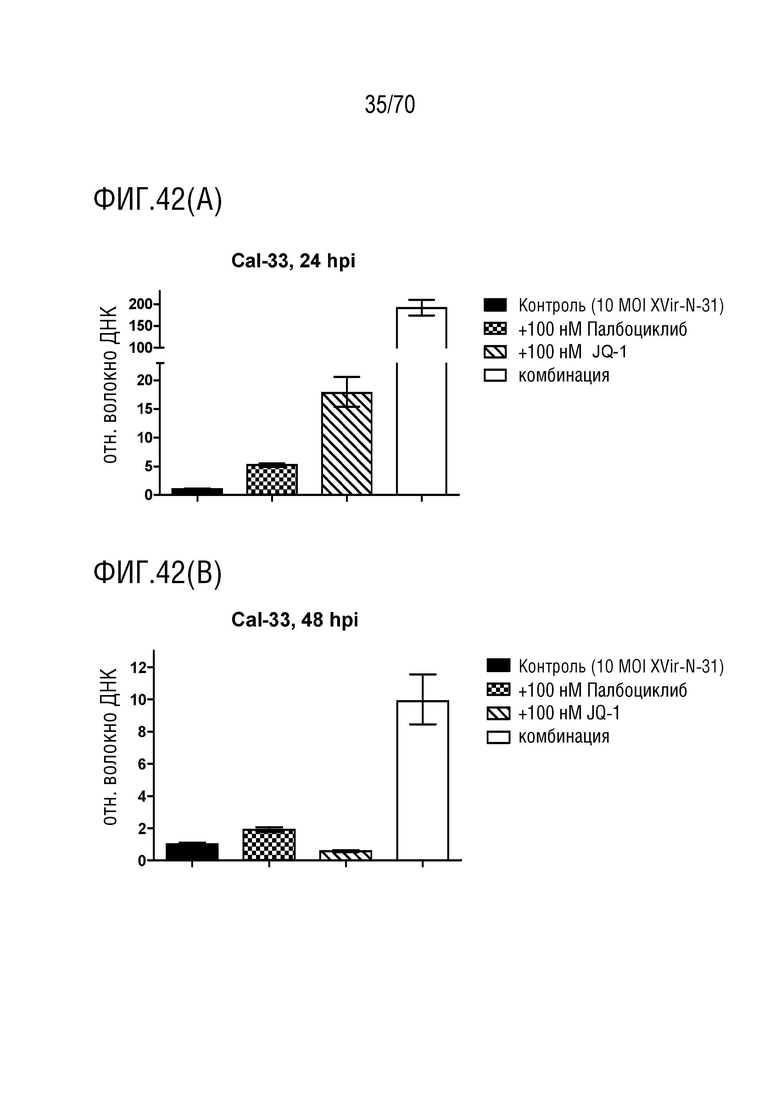
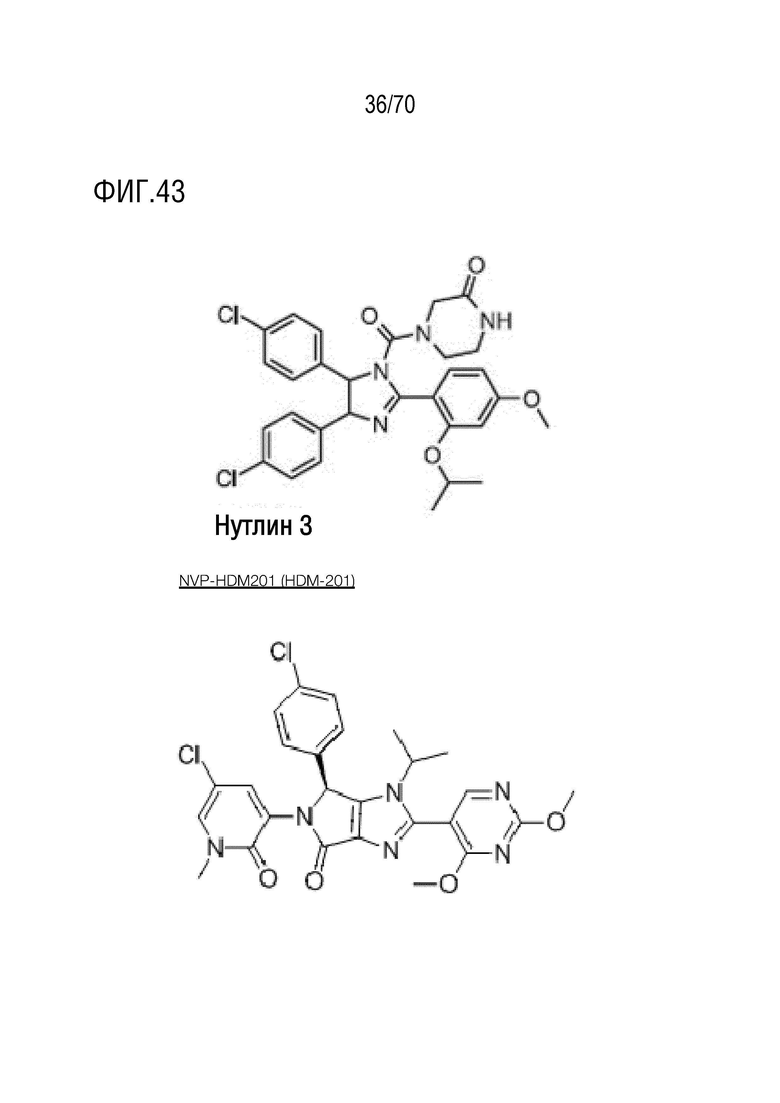
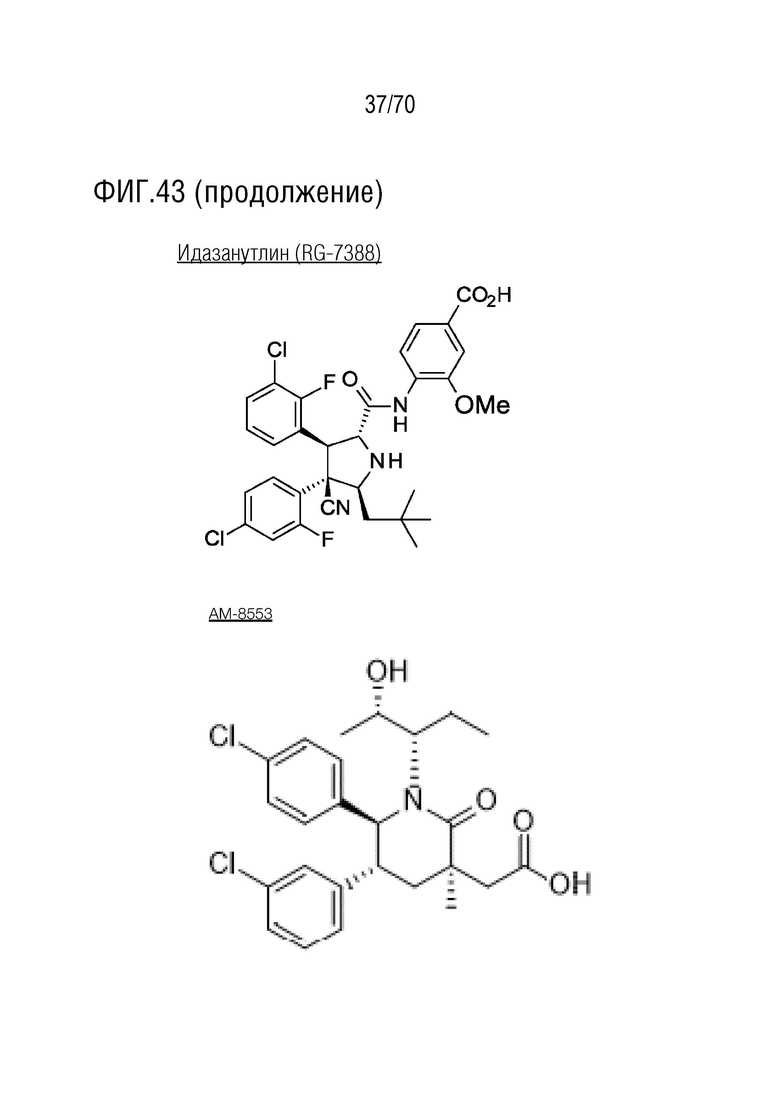
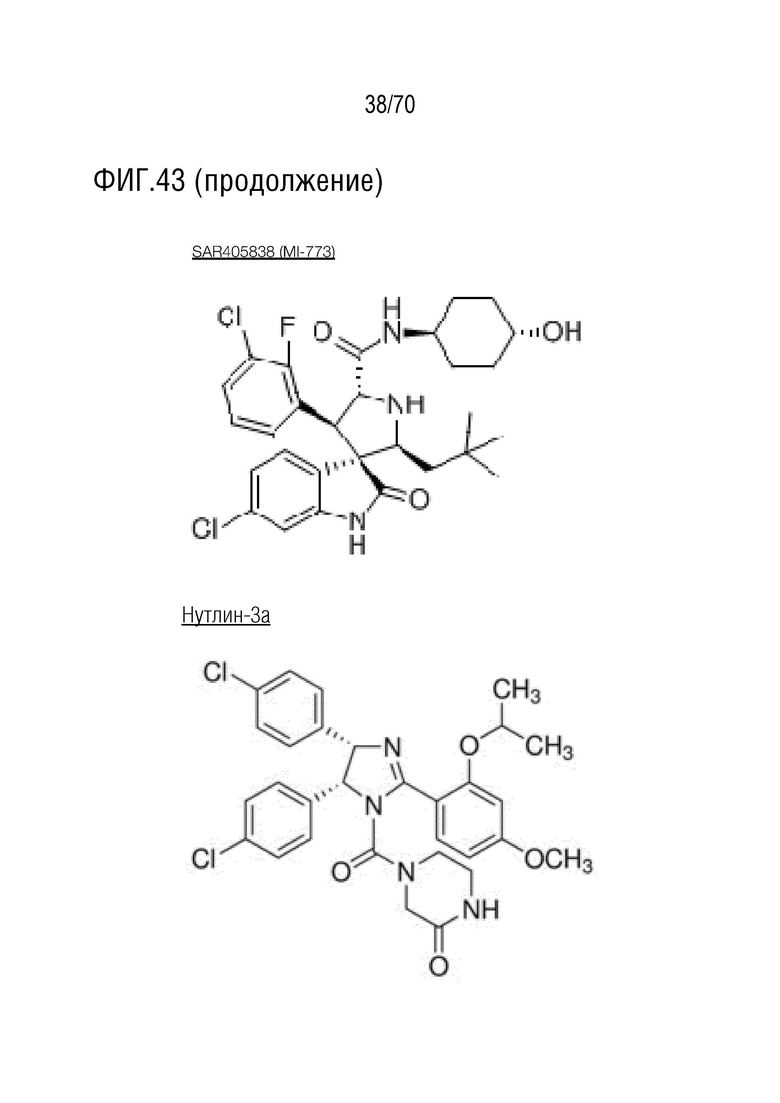
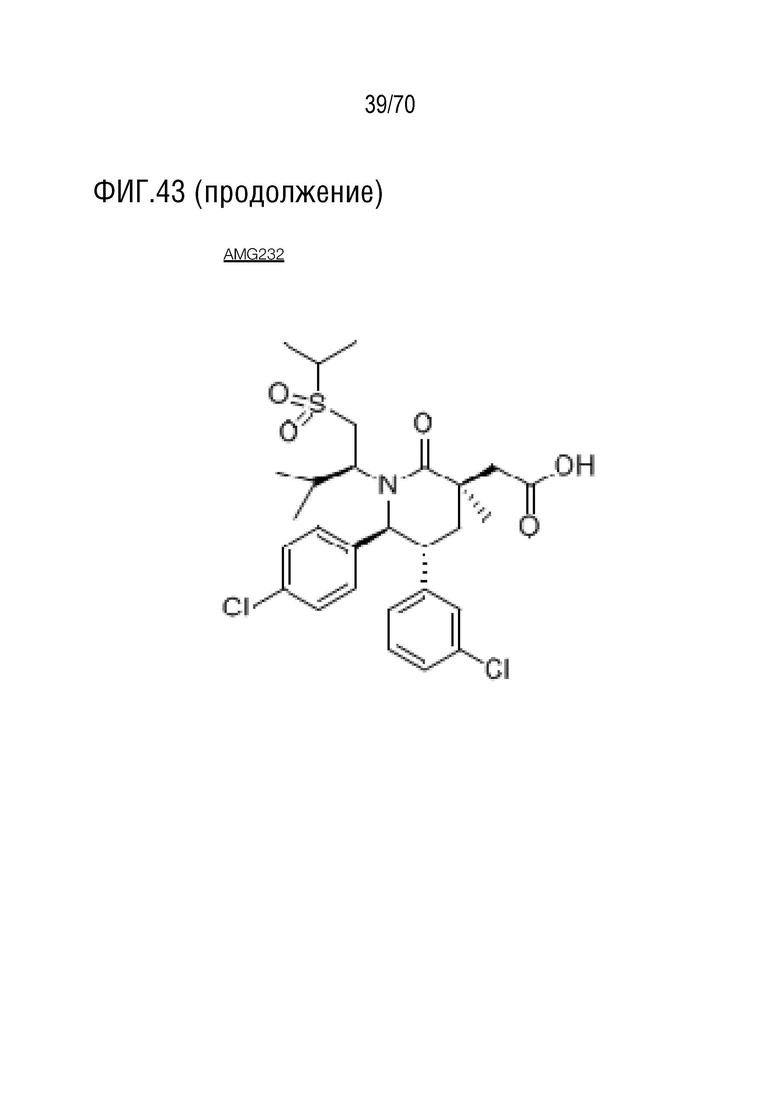
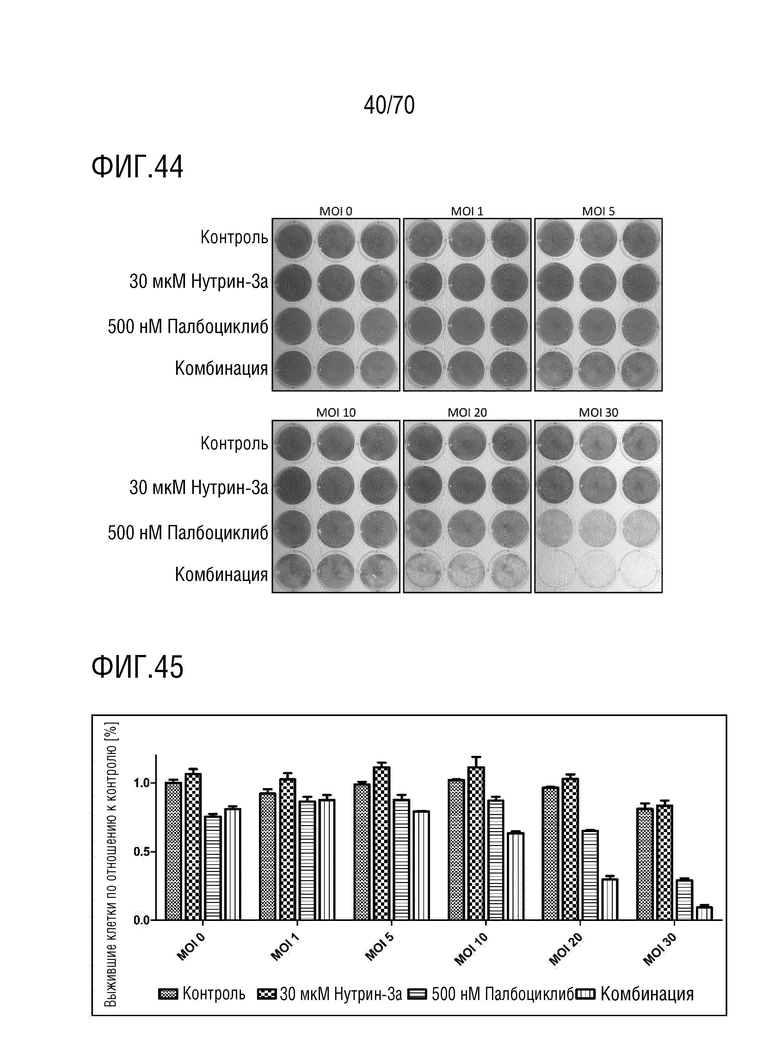
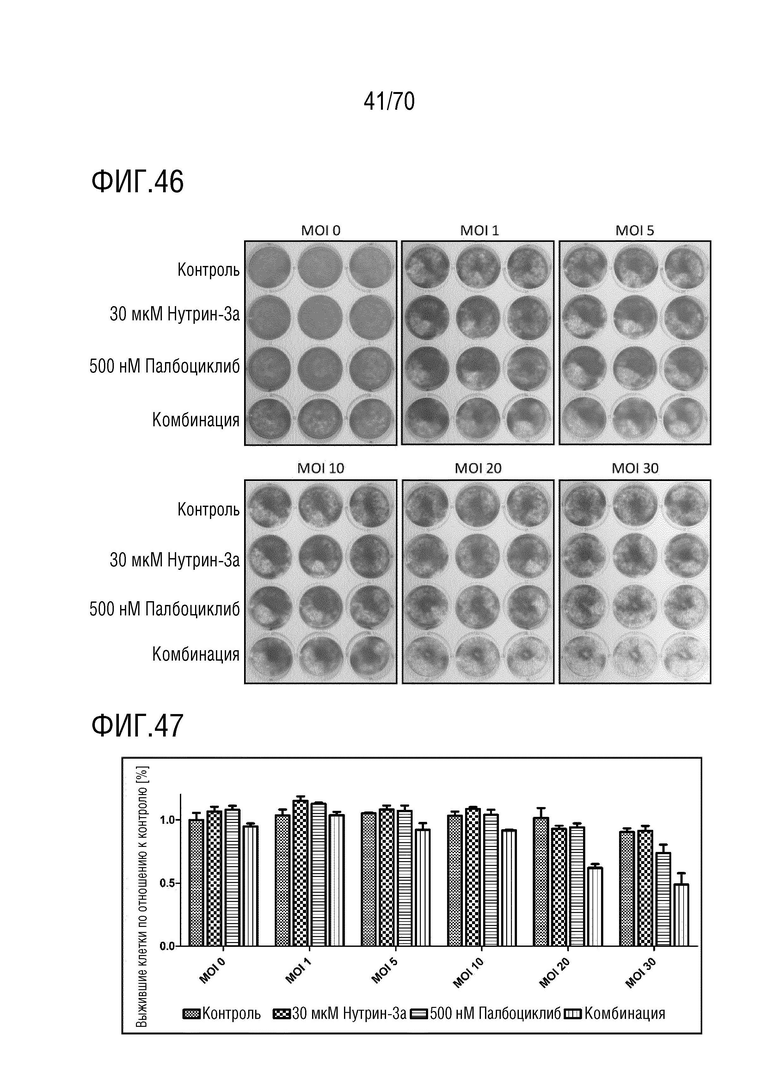
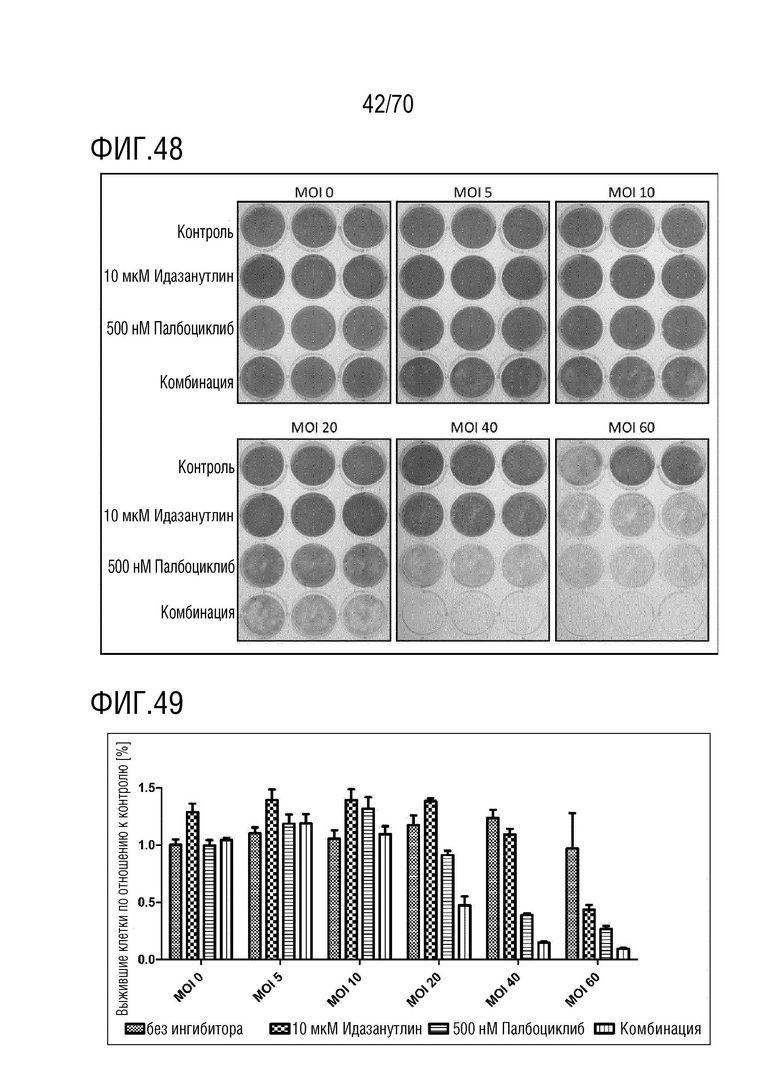
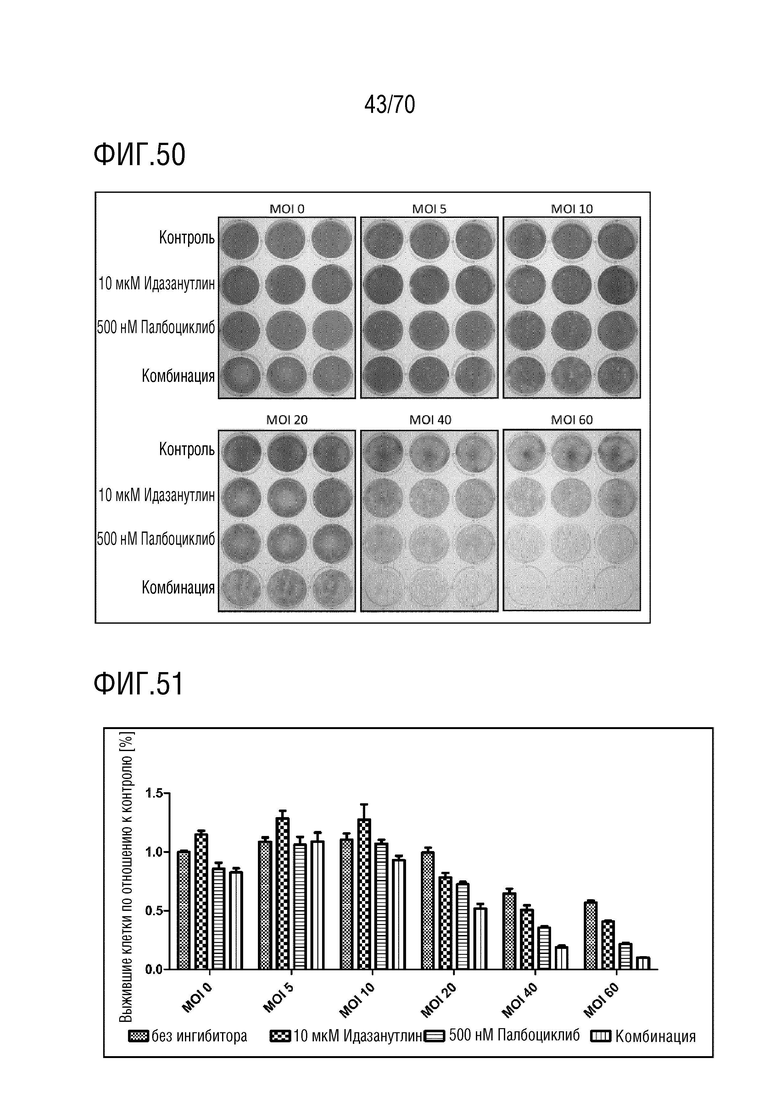
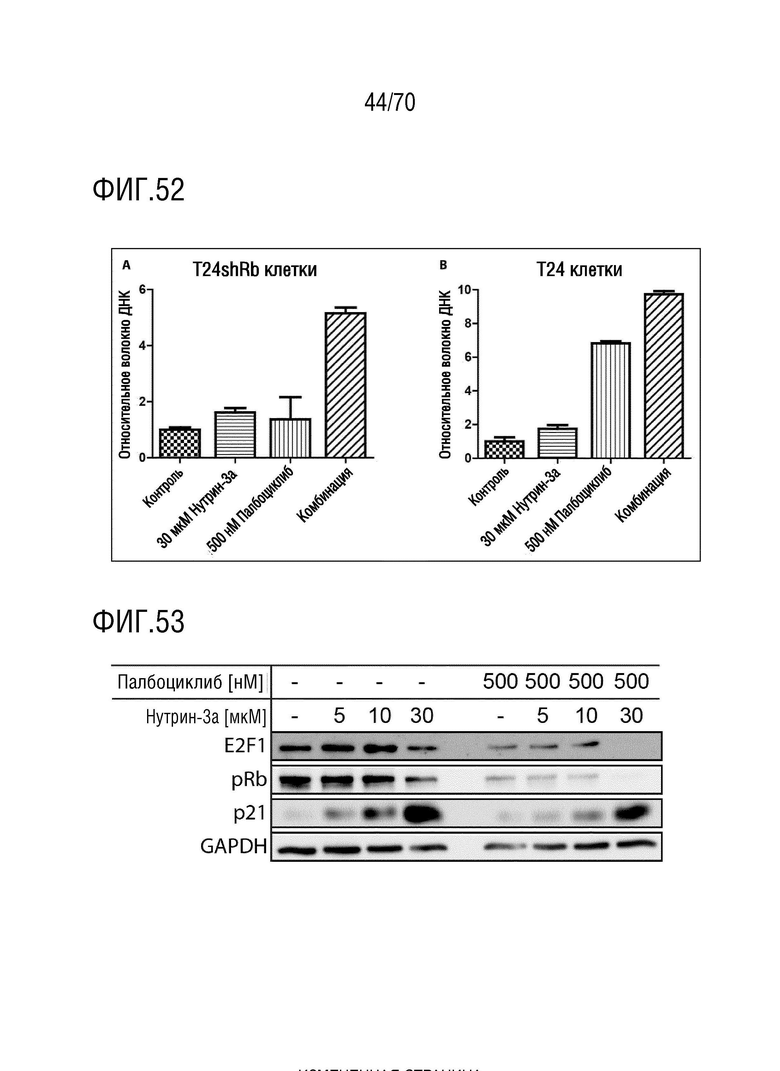
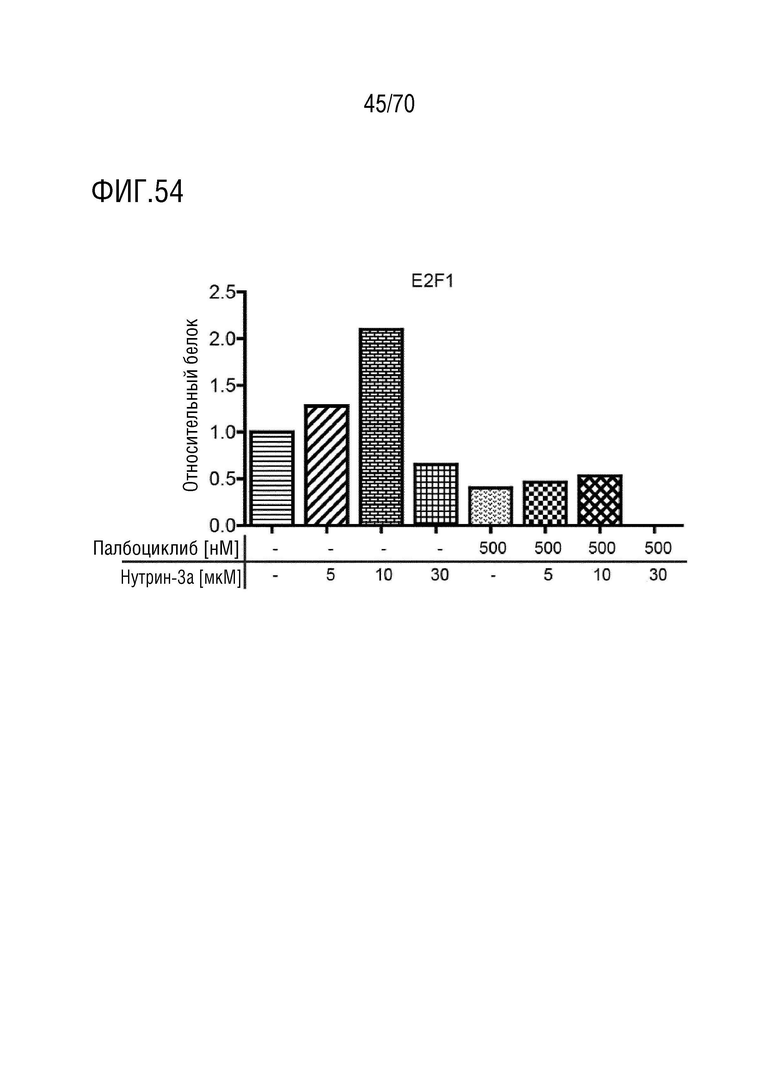
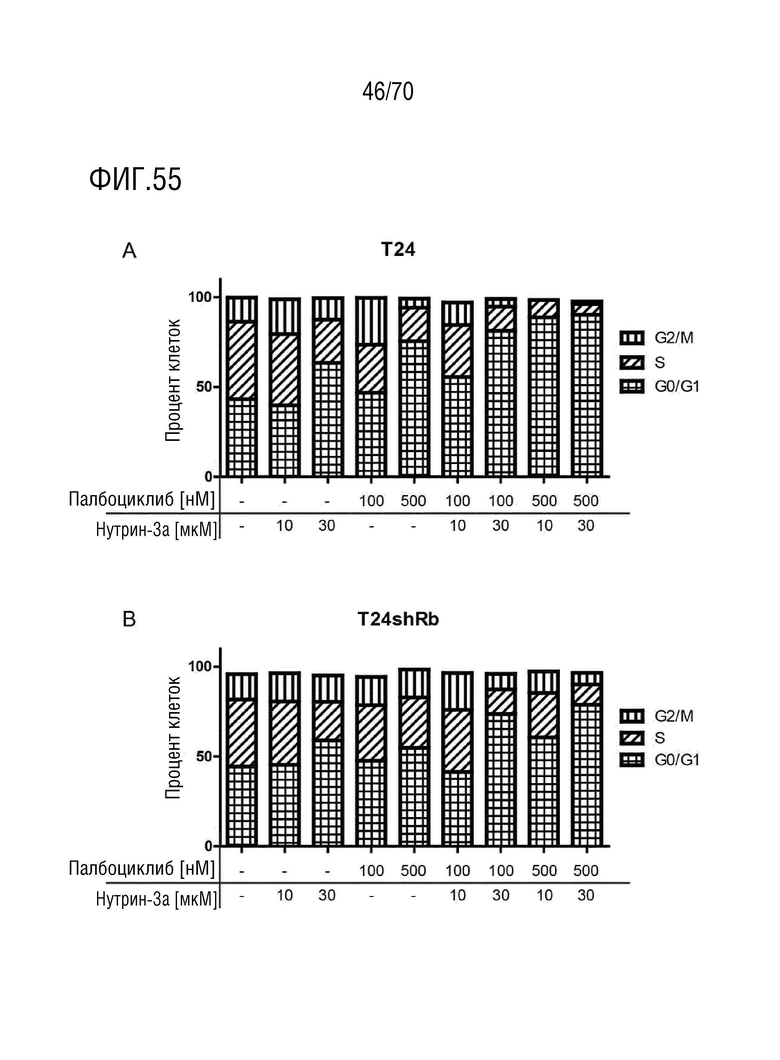
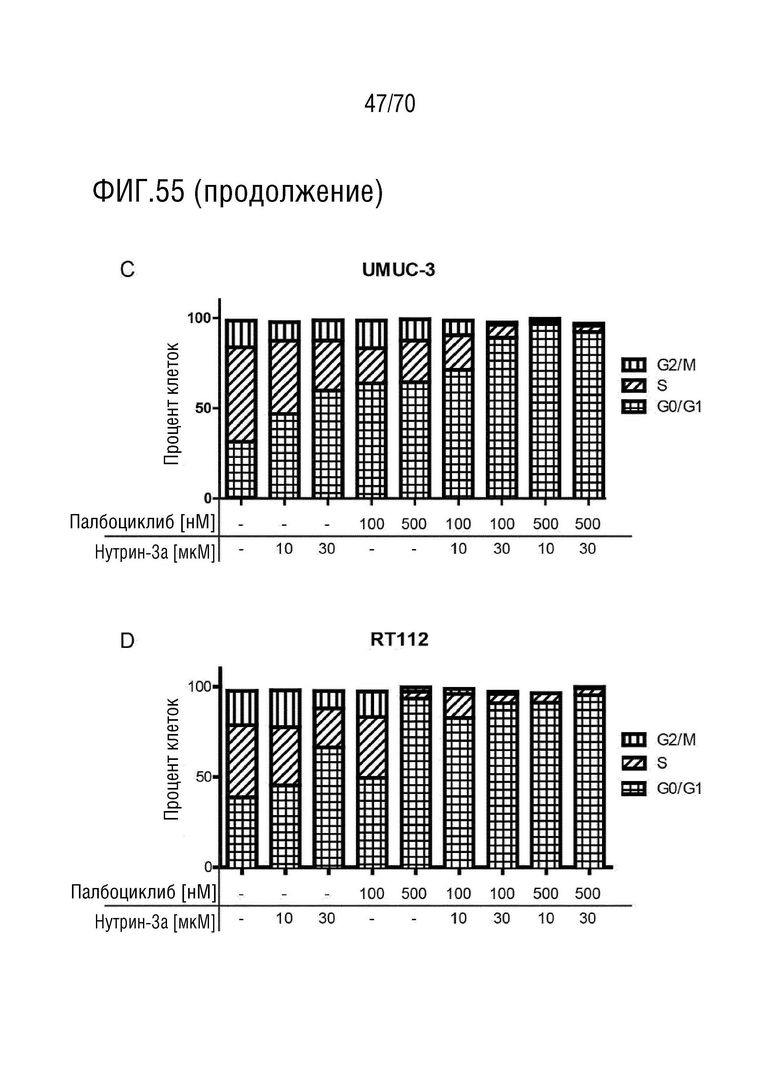
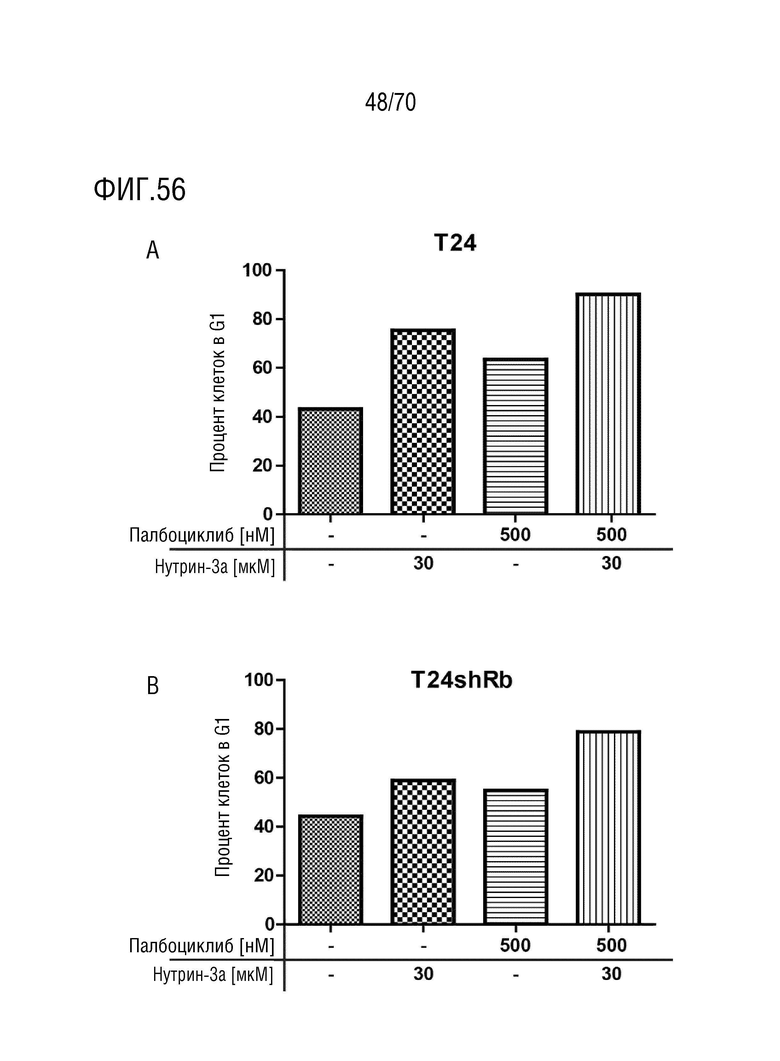
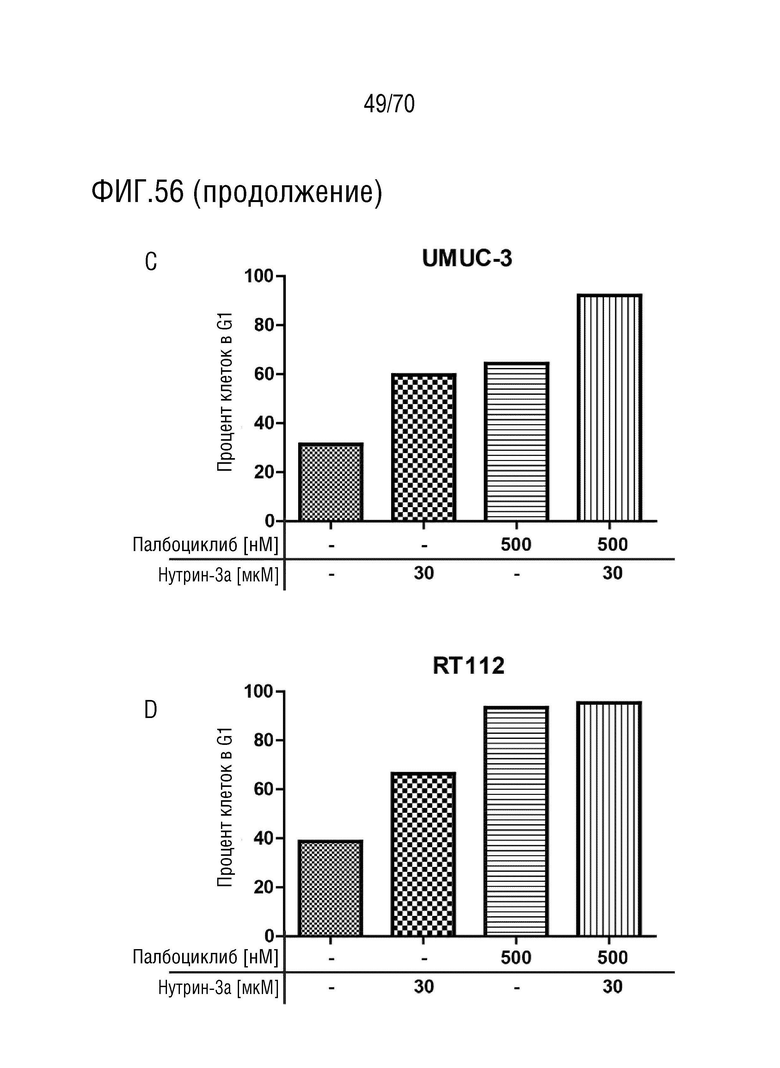
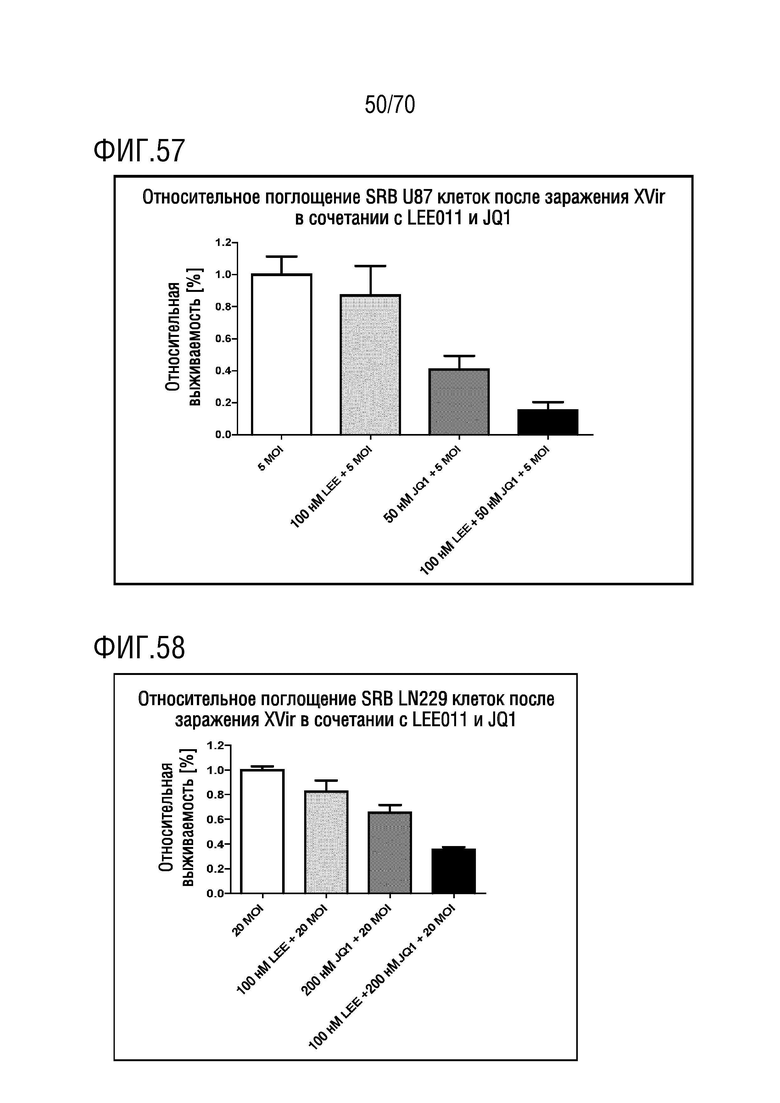
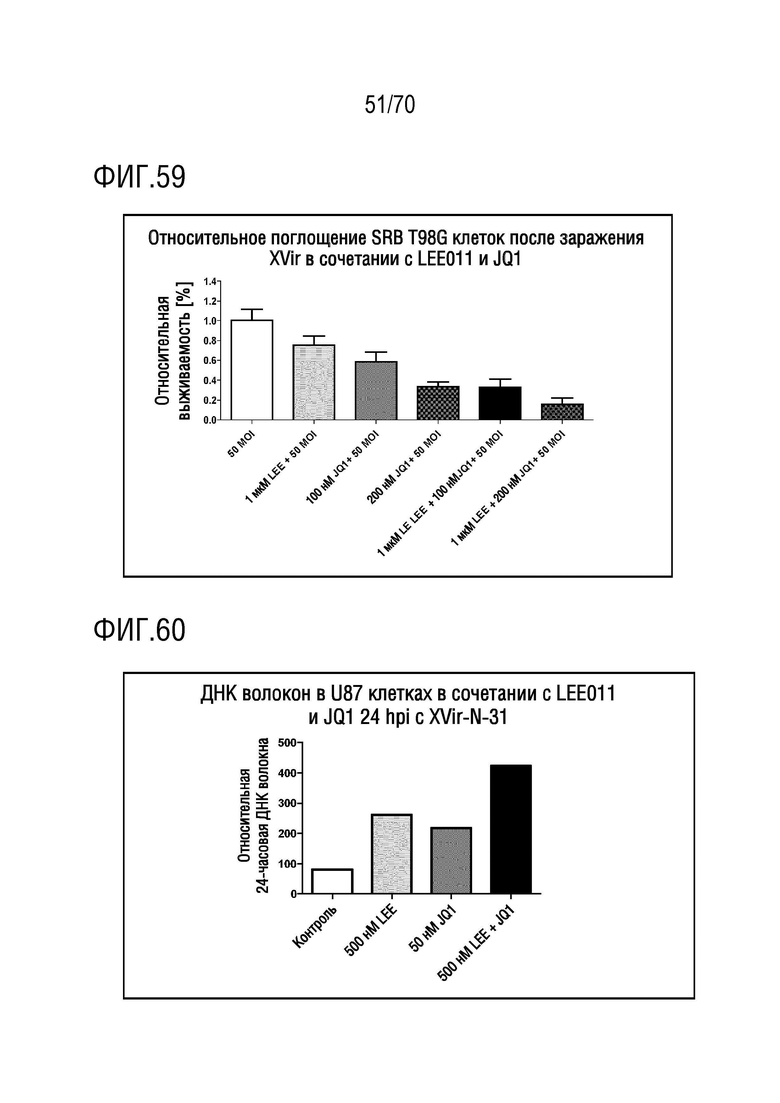
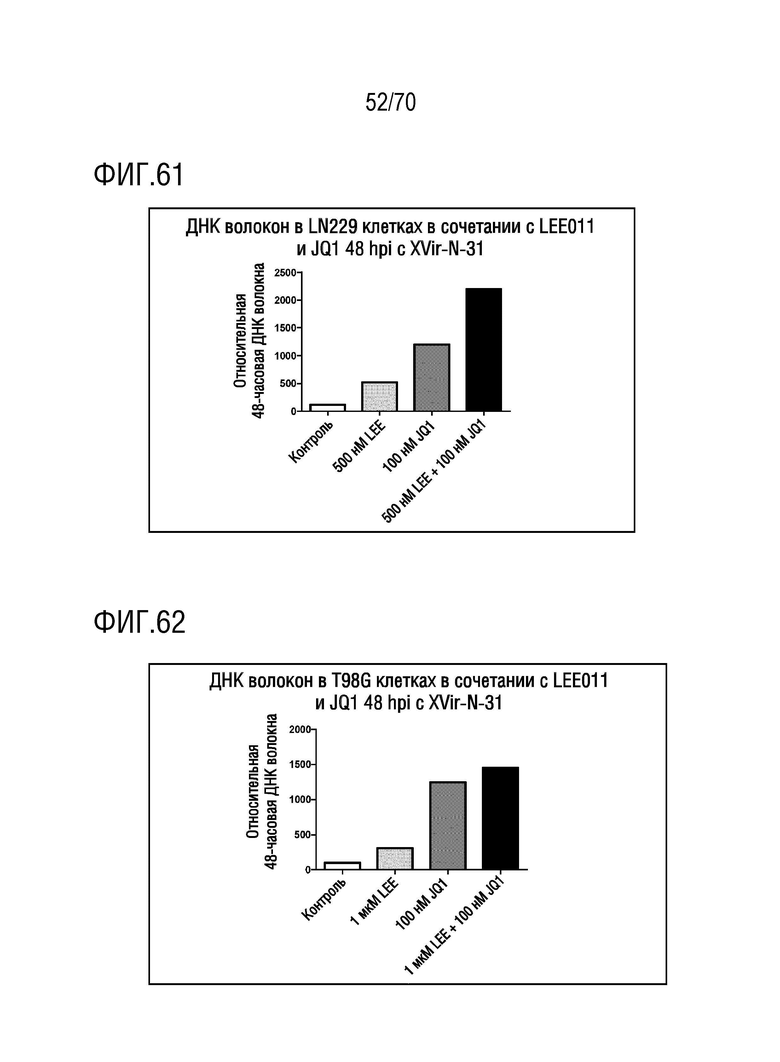
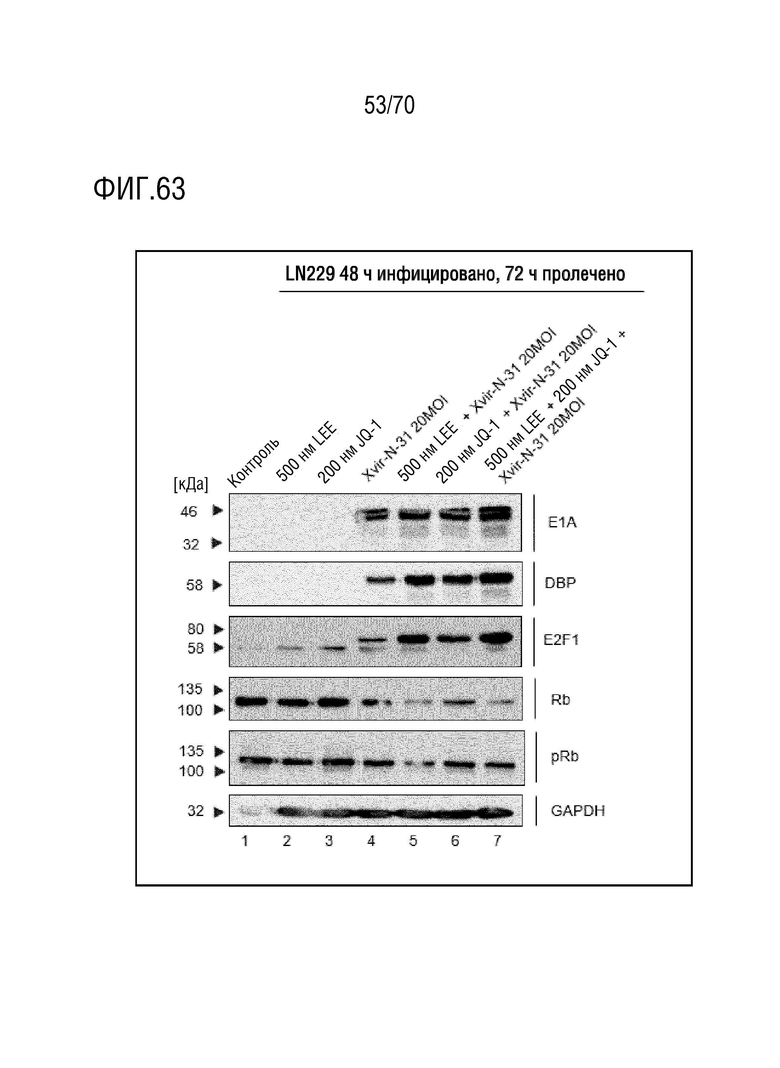
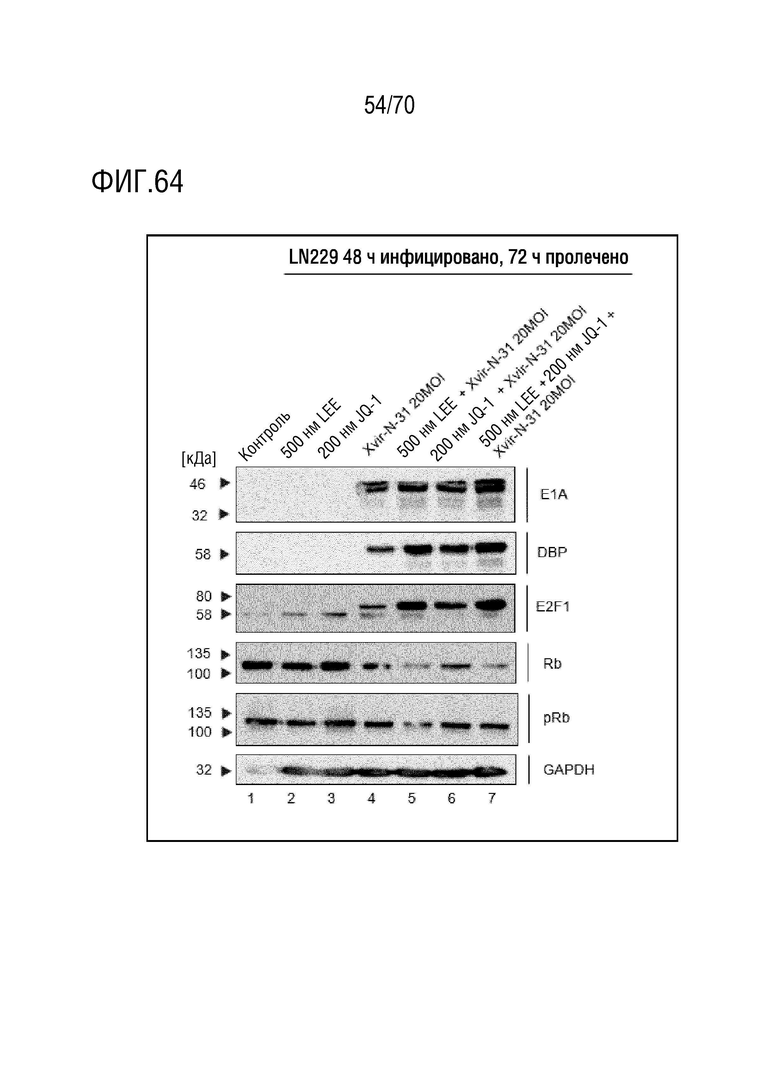
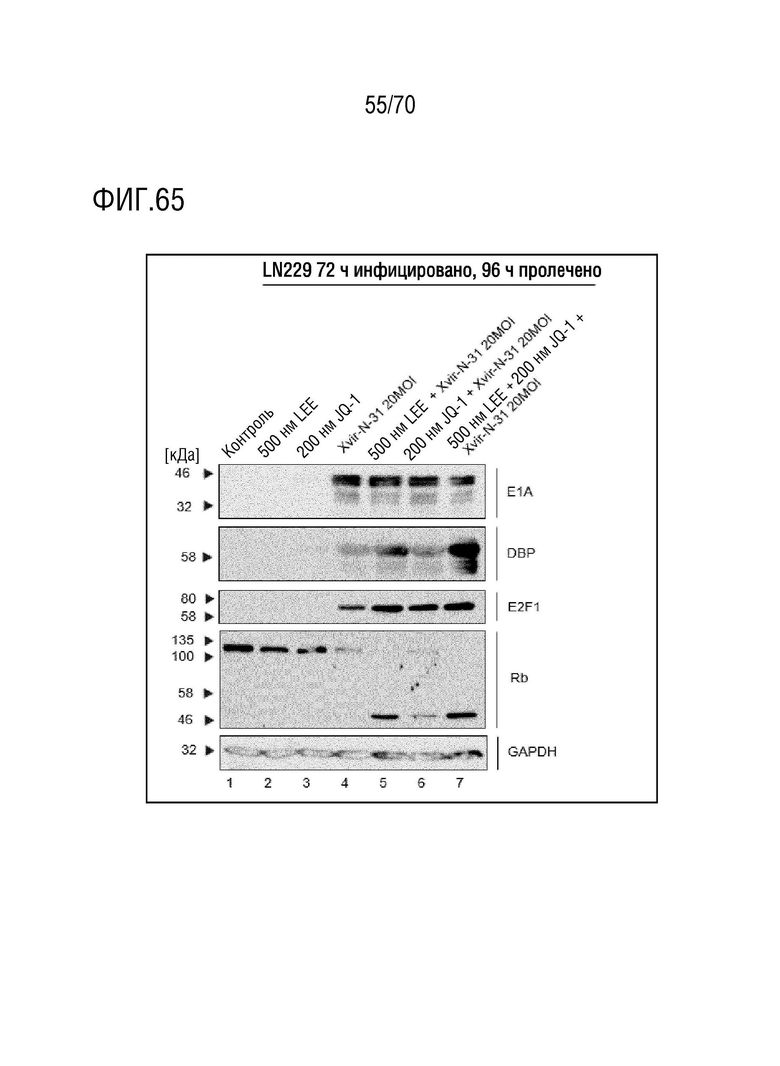
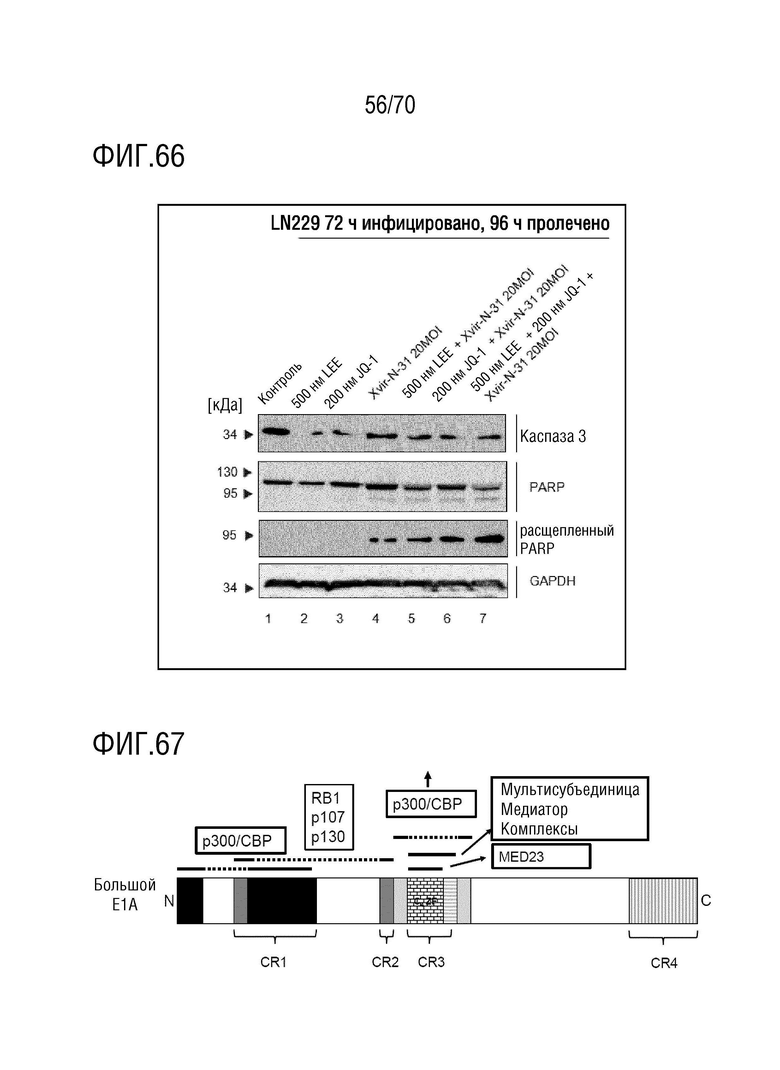
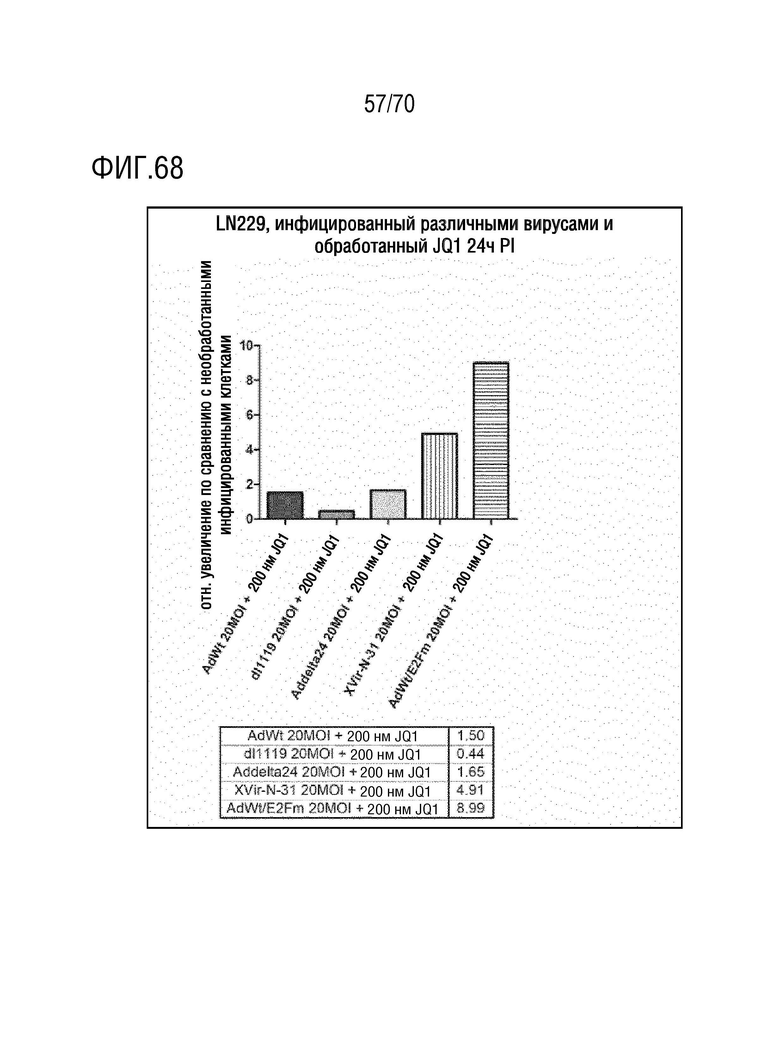
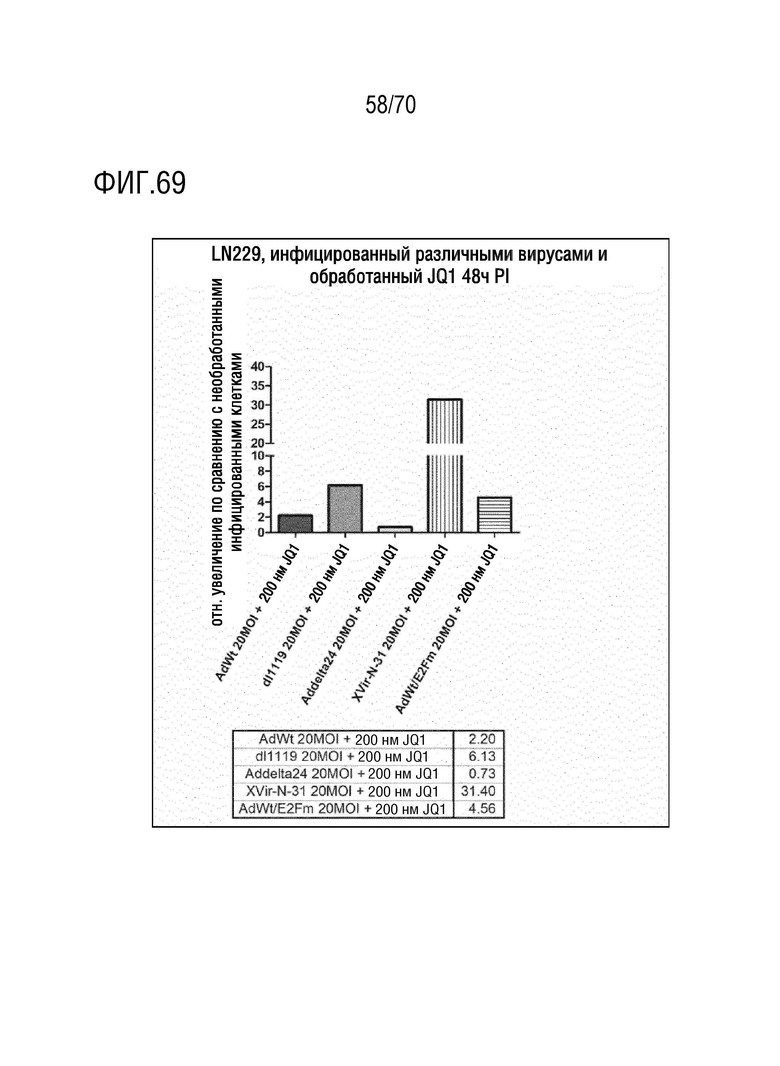
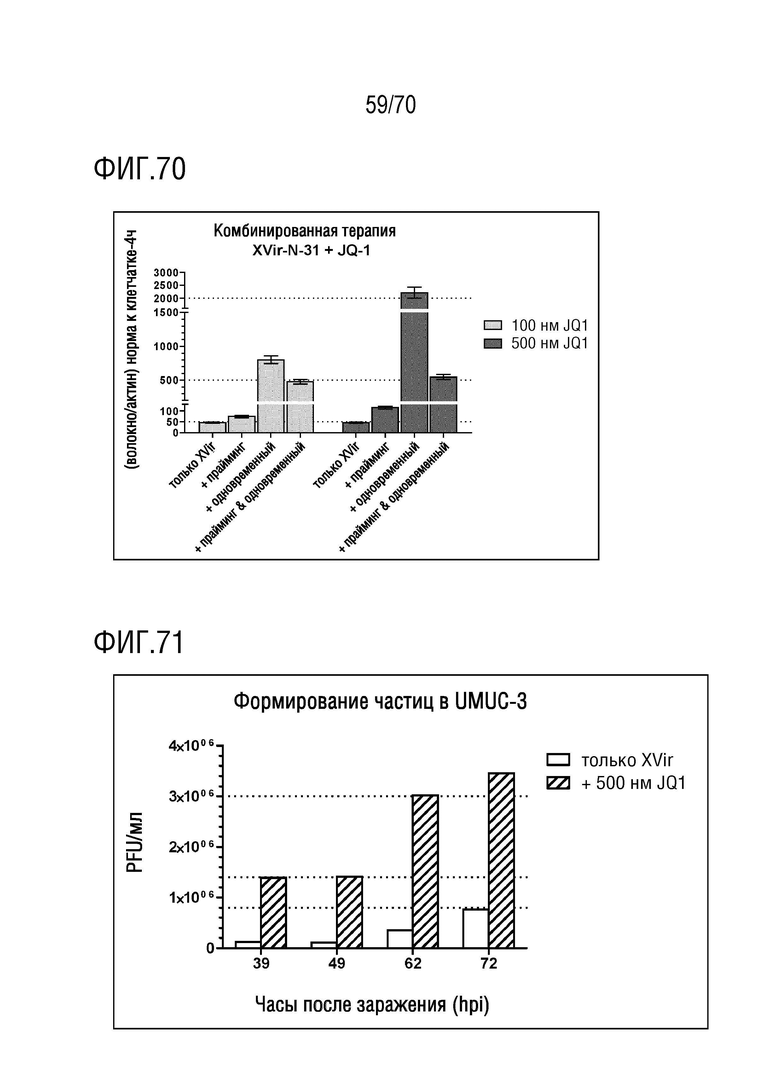
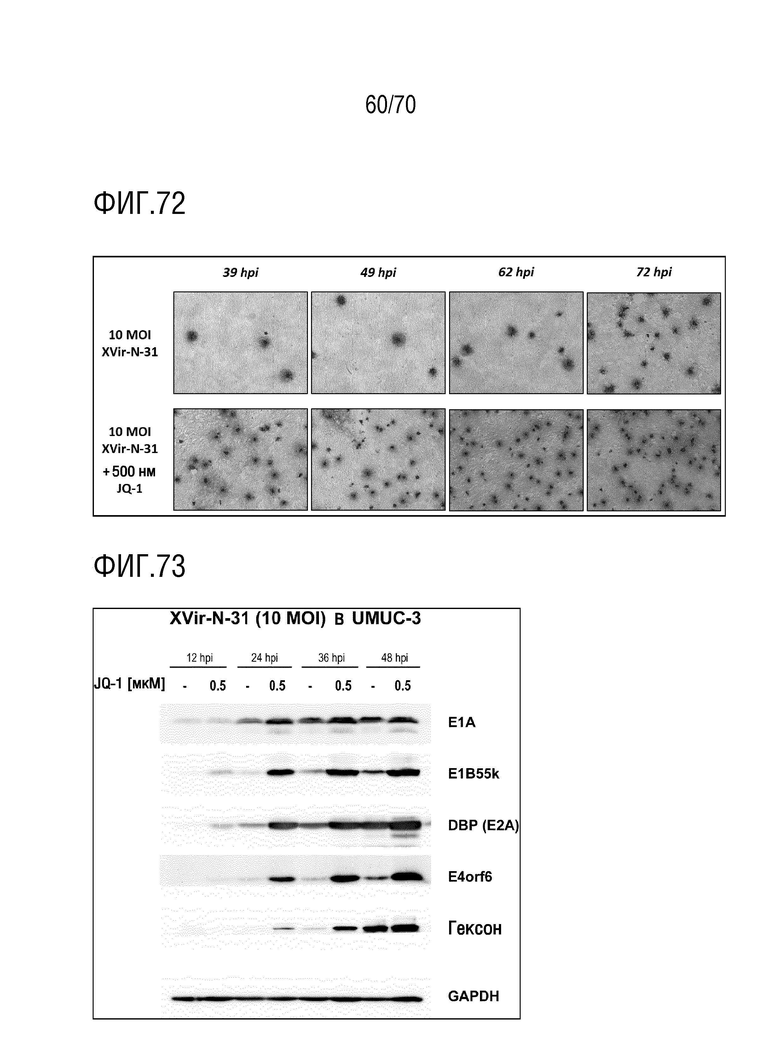
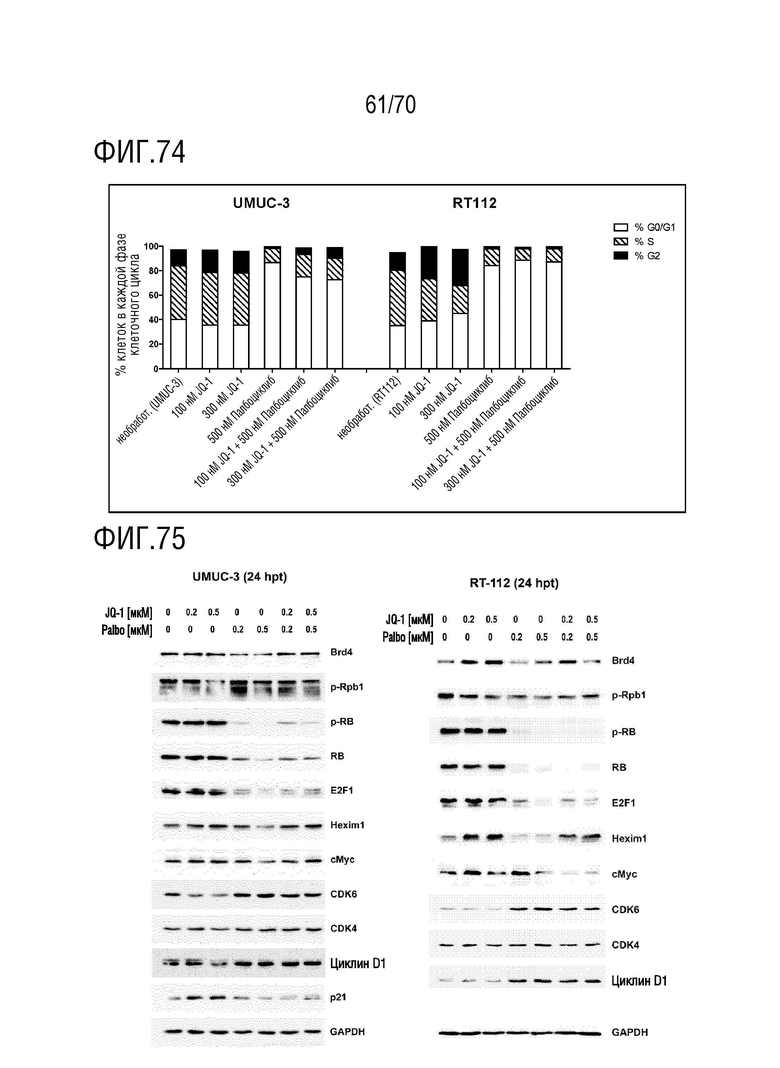
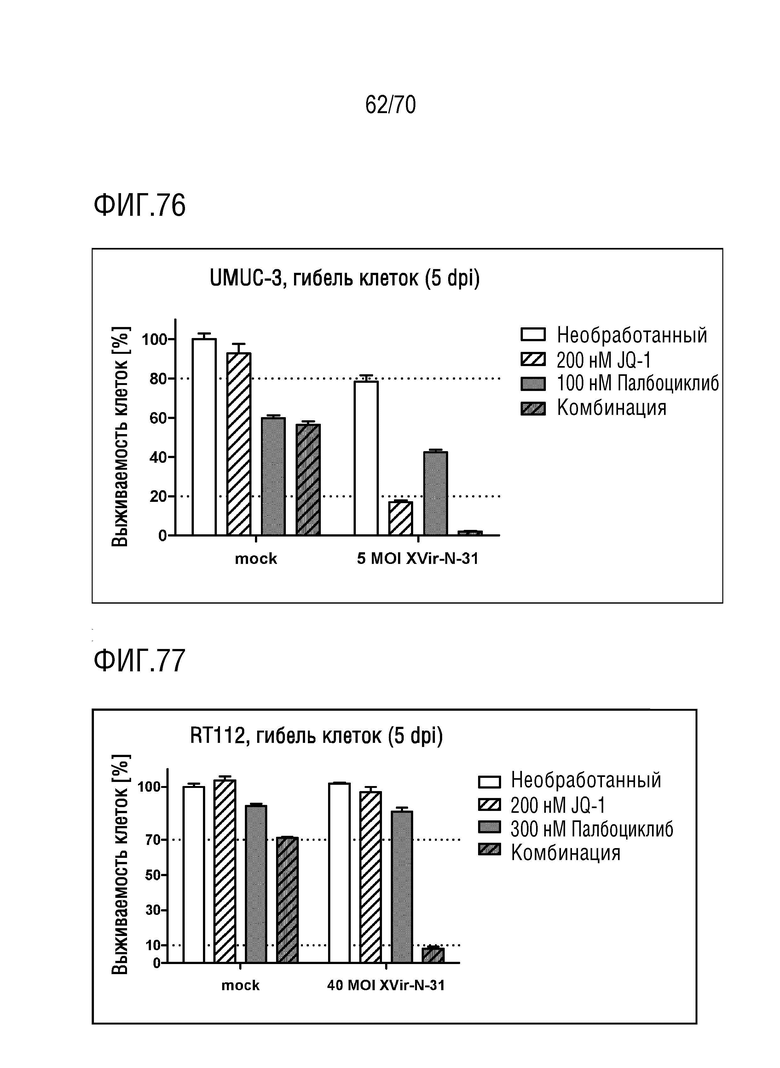
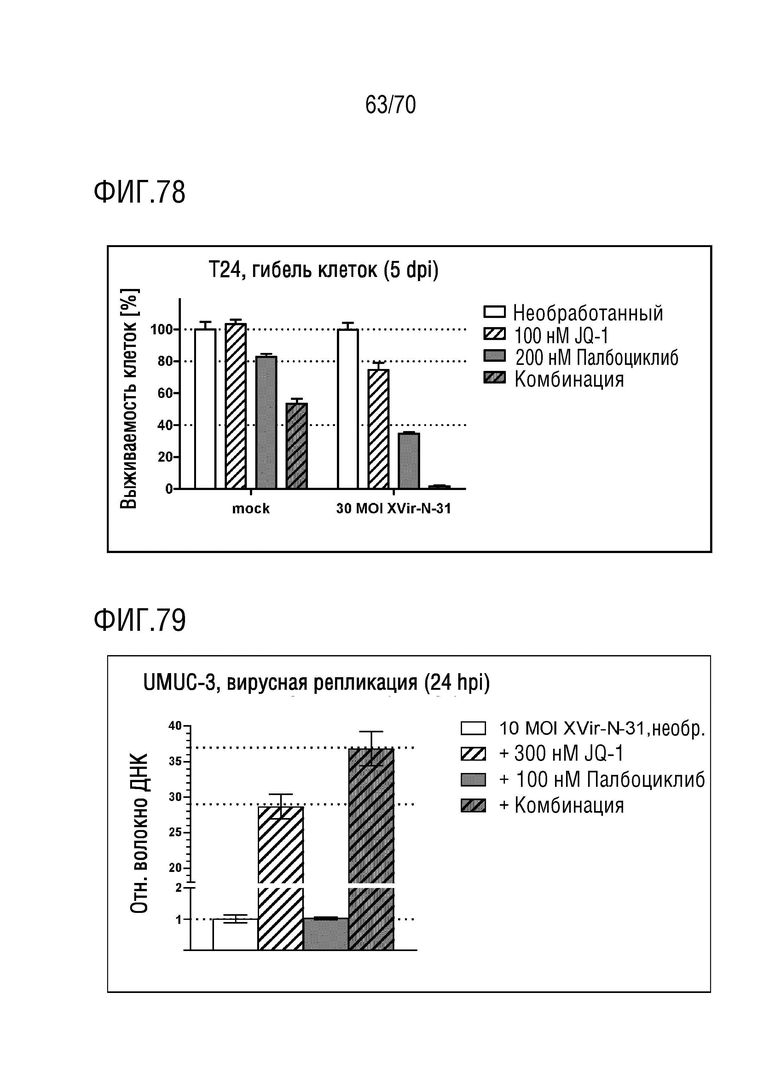
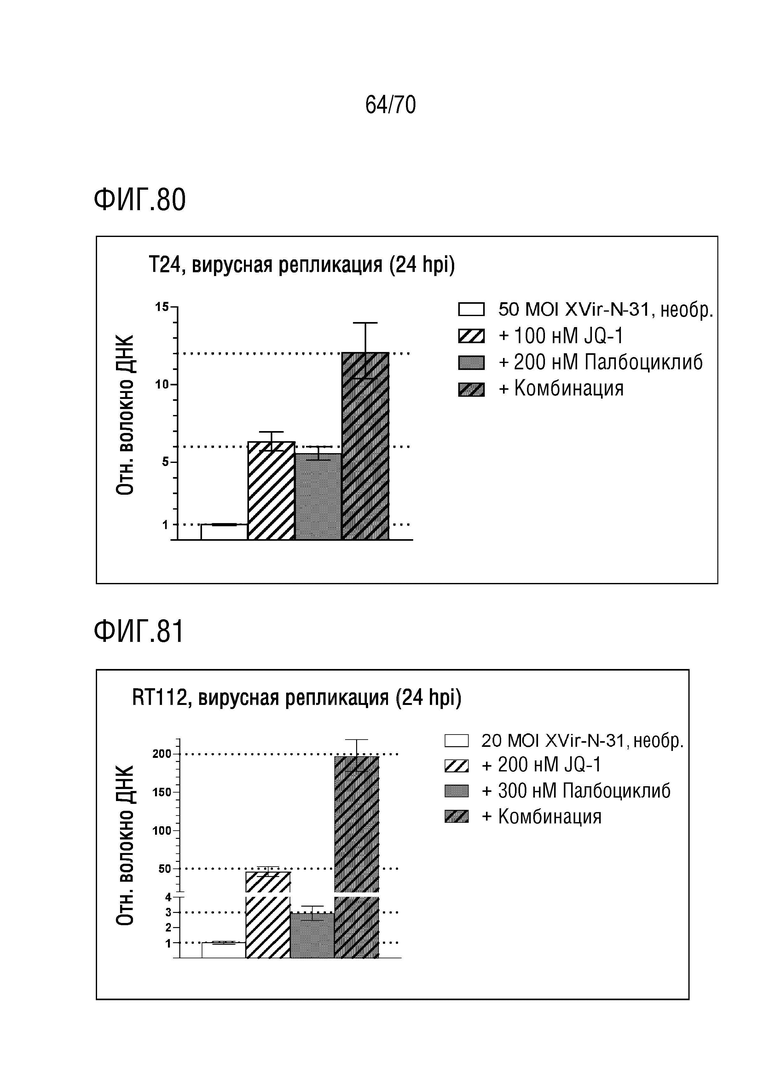
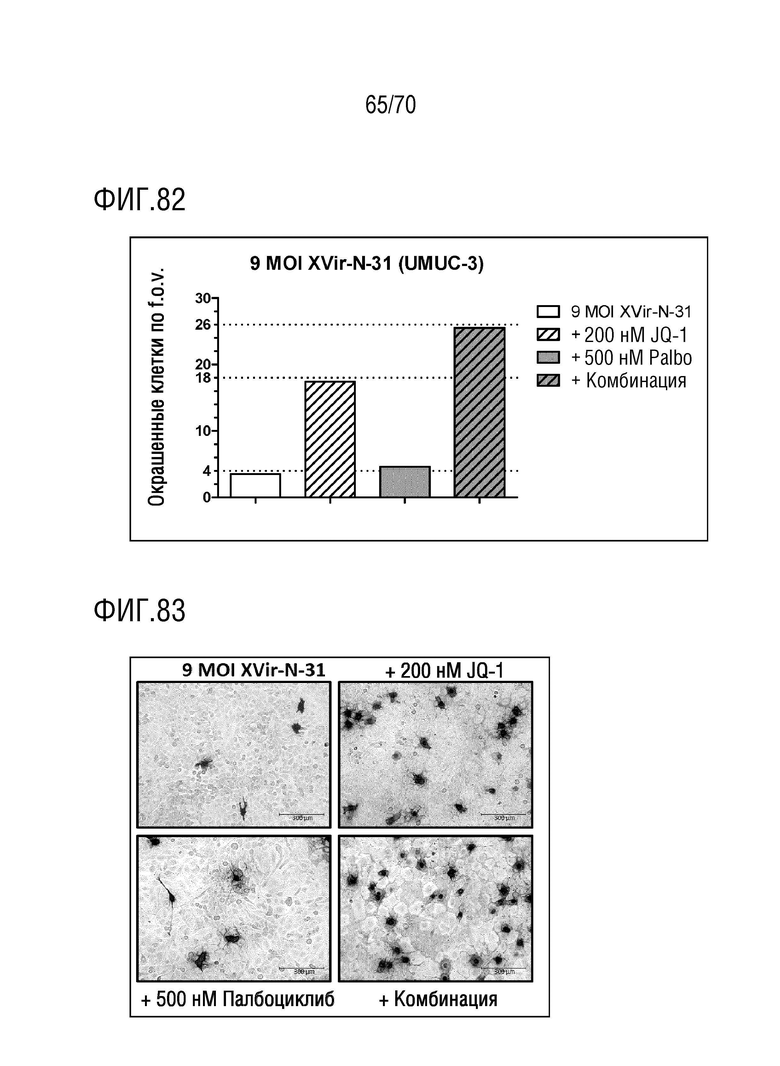
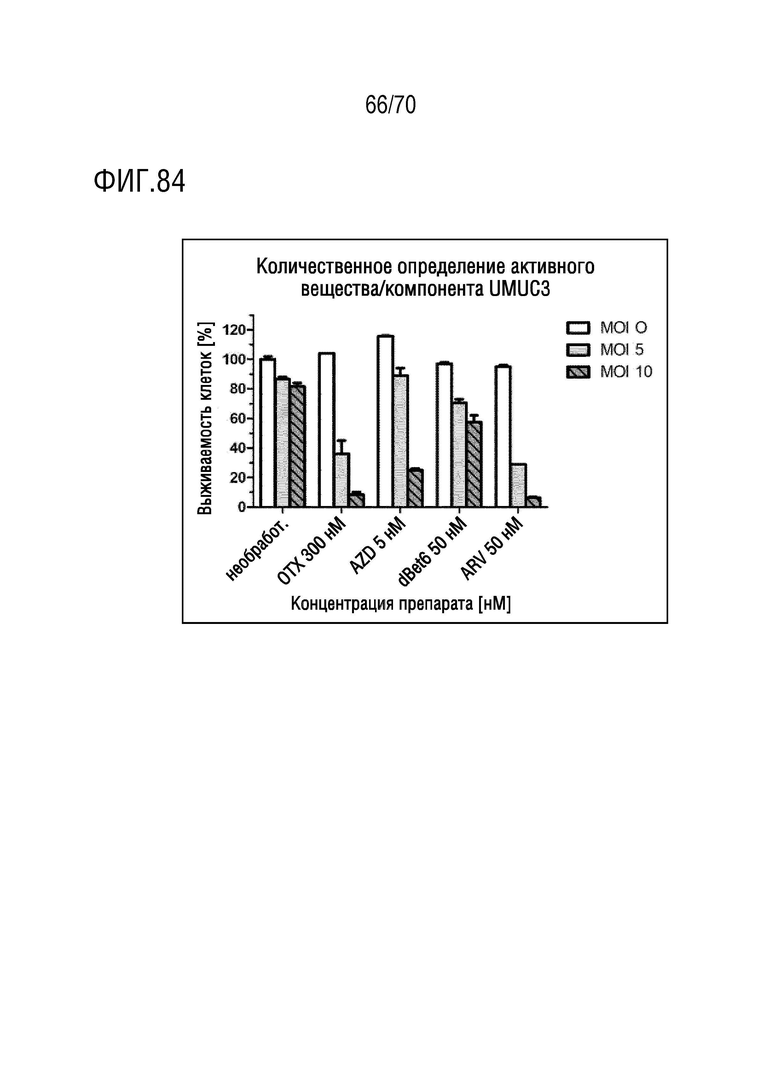
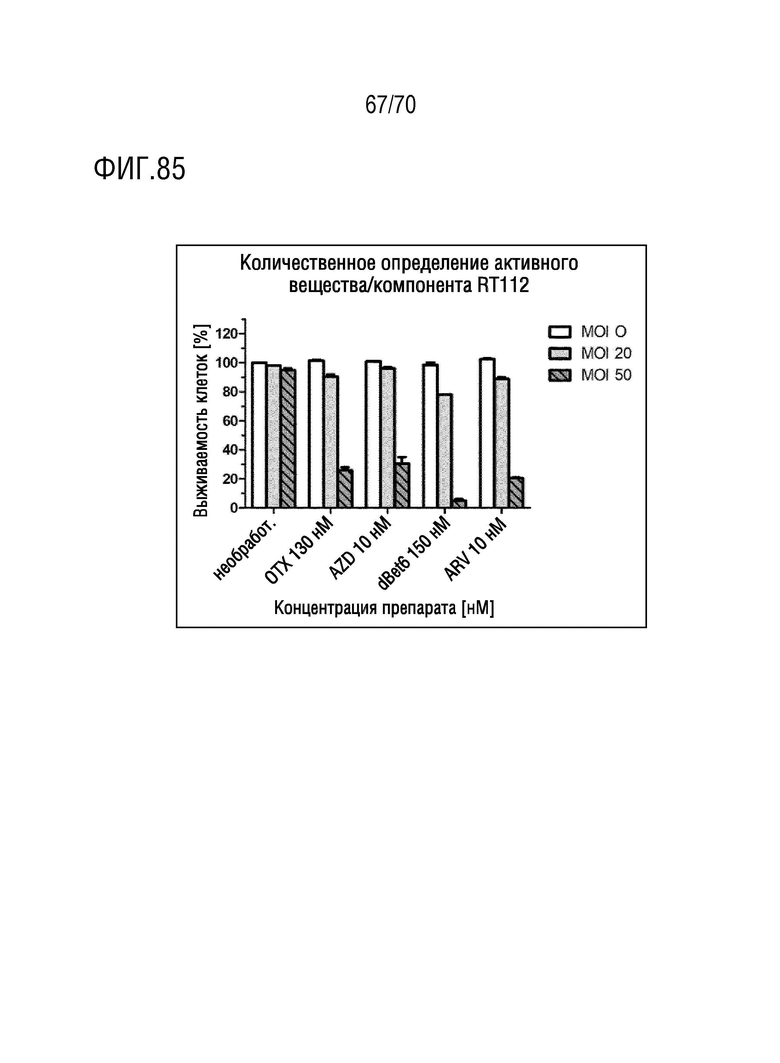
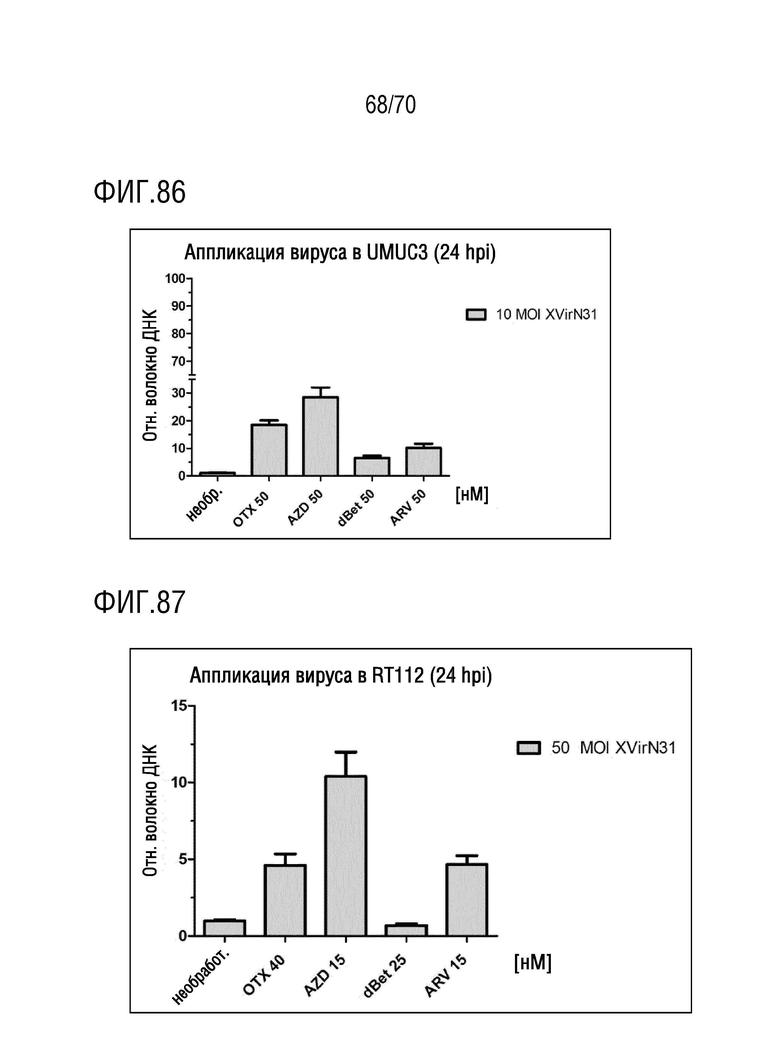
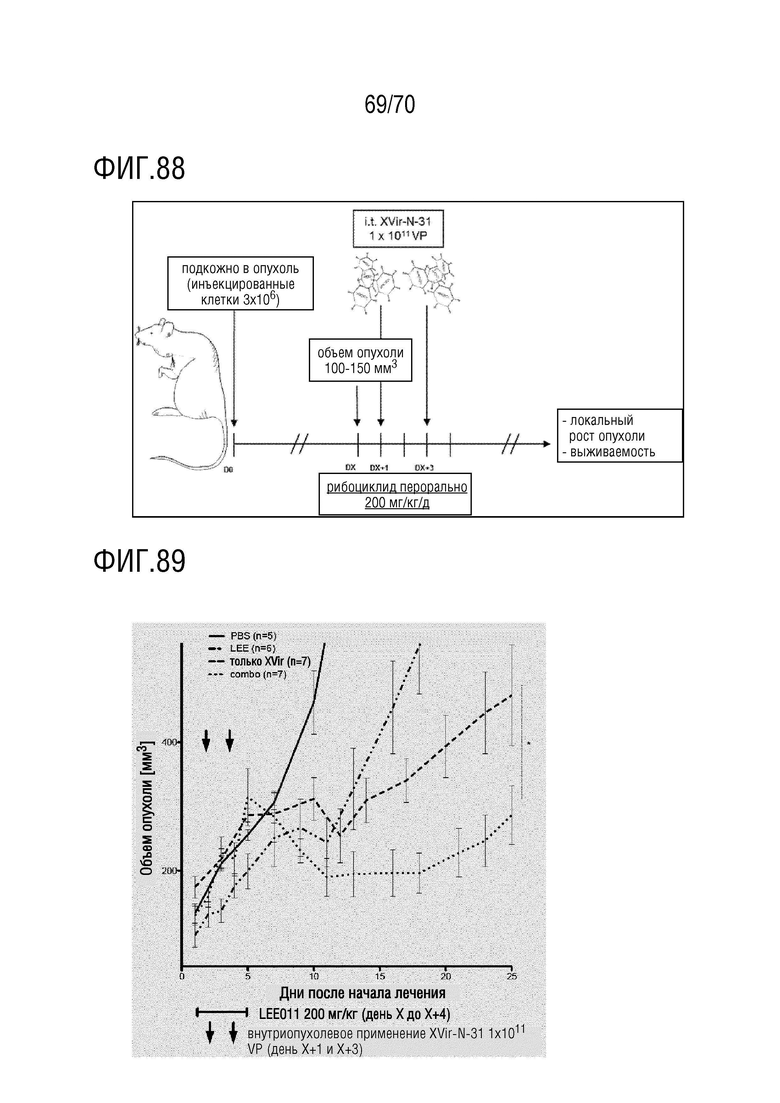
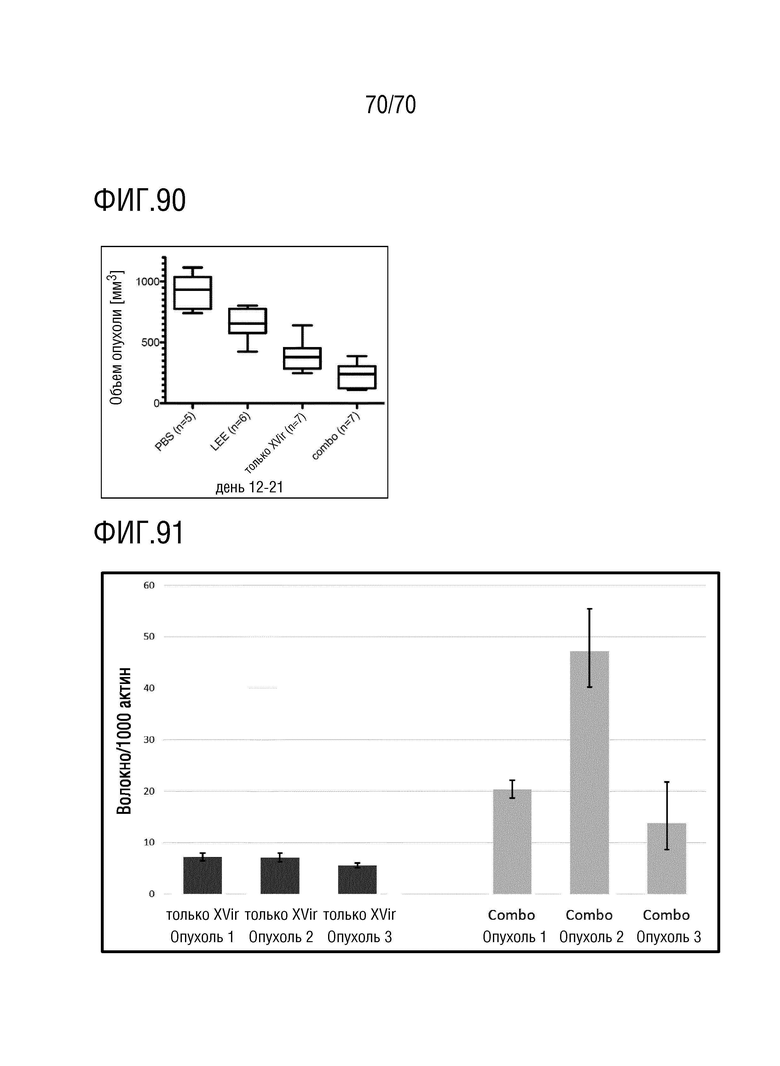
Изобретение относится к биотехнологии. Описана комбинация для лечения опухоли, где композиция содержит онколитический аденовирус, CDK4/6 ингибитор и нутлин или производное нутлина, причем онколитический аденовирус экспрессирует белок E1A12S и не экспрессирует белок E1A13S. Также описан способ лечения опухоли, согласно которому субъекту вводят онколитический аденовирус, ингибитор CDK4/6 и нутлин или производное нутлина, причем онколитический аденовирус экспрессирует белок E1A12S и не экспрессирует белок E1A13S. Изобретение расширяет арсенал средств для лечения опухоли. 2 н. и 30 з.п. ф-лы, 91 ил., 1 табл., 35 пр.
1. Комбинация для лечения опухоли, где композиция содержит онколитический аденовирус, CDK4/6 ингибитор и нутлин или производное нутлина, причем онколитический аденовирус экспрессирует белок E1A12S и не экспрессирует белок E1A13S.
2. Комбинация по п. 1, где аденовирус является дефектным по репликации в клетках, у которых отсутствует YB-1 в ядре, но реплицируется в клетках, которые имеют YB-1 в ядре.
3. Комбинация по любому из пп. 1 и 2, где аденовирус не экспрессирует белок E1B 19 кДа.
4. Комбинация по любому из пп. 1-3, где аденовирус представляет собой аденовирус, экспрессирующий RGD-мотив на волокне.
5. Комбинация по любому из пп. 1-4, где аденовирус представляет собой XVir-N-31.
6. Комбинация по любому из пп. 1-5, где ингибитор CDK4/6 представляет собой ингибитор CDK4/6, останавливающий клетки в фазе G1 и ингибирующий E2F1.
7. Комбинация по любому из пп. 1-6, где ингибитор CDK4/6 представляет собой АТФ-конкурентный ингибитор CDK4/6.
8. Комбинация по любому из пп. 1-7, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также называется PD 0332991, абемациклиб, который также называется LY-2835219, рибоциклиб, который также называется LEE011, трилациклиб, который также называется G1T28, и динациклиб.
9. Комбинация по любому из пп. 1-8, где комбинация дополнительно содержит ингибитор PARP.
10. Комбинация по любому из пп. 1-9, где комбинация дополнительно содержит ингибитор бромодомена.
11. Комбинация по любому из пп. 1-10, где опухоль экспрессирует Rb или является Rb-положительной.
12. Комбинация по любому из пп. 1-10, где опухоль является Rb-отрицательной.
13. Комбинация по любому из пп. 1-12, где опухолевые клетки обладают резистентностью или нечувствительностью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
14. Комбинация по любому из пп. 1-13, где опухолевые клетки имеют YB-1 в ядре.
15. Комбинация по любому из пп. 1-14, где опухолевые клетки имеют YB-1 в ядре клетки независимо от клеточного цикла.
16. Комбинация по любому из пп. 1-15, где опухоль выбрана из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легких, саркому, рак яичников, рак почки, рак предстательной железы и лейкоз.
17. Способ лечения опухоли, согласно которому субъекту вводят онколитический аденовирус, ингибитор CDK4/6 и нутлин или производное нутлина, причем онколитический аденовирус экспрессирует белок E1A12S и не экспрессирует белок E1A13S.
18. Способ по п. 17, где аденовирус является дефектным по репликации в клетках, у которых отсутствует YB-1 в ядре, но реплицируется в клетках, которые имеют YB-1 в ядре.
19. Способ по любому из пп. 17 и 18, где аденовирус не экспрессирует белок E1B 19 кДа.
20. Способ по любому из пп. 17-19, где аденовирус представляет собой аденовирус, экспрессирующий RGD-мотив на волокне.
21. Способ по любому из пп. 17-20, где аденовирусом является XVir-N-31.
22. Способ по любому из пп. 17-21, где ингибитор CDK4/6 представляет собой ингибитор CDK4/6, задерживающий клетки в фазе G1 и ингибирующий E2F1.
23. Способ по любому из пп. 17-22, где ингибитор CDK4/6 представляет собой АТФ-конкурентный ингибитор CDK4/6.
24. Способ по любому из пп. 17-23, где ингибитор CDK4/6 выбран из группы, включающей палбоциклиб, который также упоминается как PD 0332991, абемациклиб, который также упоминается как LY-2835219, рибоциклиб, который также упоминается как LEE011, трилациклиб, который также упоминается как G1T28 и динациклиб.
25. Способ по любому из пп. 17-24, в котором субъекту вводят ингибитор PARP.
26. Способ по любому из пп. 17-25, в котором субъекту вводят ингибитор бромодомена.
27. Способ по любому из пп. 17-26, где опухоль экспрессирует Rb или является Rb-положительной.
28. Способ по любому из пп. 17-27, где опухоль является Rb-отрицательной.
29. Способ по любому из пп. 17-28, где опухолевые клетки обладают устойчивостью к одному или нескольким фармацевтически активным агентам и/или к лучевой терапии.
30. Способ по любому из пп. 17-29, где опухолевые клетки имеют YB-1 в ядре.
31. Способ по любому из пп. 17-20, где опухолевые клетки имеют YB-1 в ядре клетки независимо от клеточного цикла.
32. Способ по любому из пп. 17-31, где опухоль выбрана из группы, включающей рак мочевого пузыря, рак молочной железы, метастатический рак молочной железы (mBC), меланому, глиому, рак поджелудочной железы, гепатоцеллюлярную карциному, аденокарциному легкого, саркому, рак яичников, рак почек, рак предстательной железы и лейкоз.
| РЕЖИМ ВВЕДЕНИЯ И СОСТАВЫ ДЛЯ АДЕНОВИРУСОВ ТИПА В | 2014 |
|
RU2671558C2 |
| RU 2016142896 A, 07.05.2018 | |||
| RU 2016110874 A, 04.10.2017 | |||
| WO 2018195450 A1, 25.10.2018 | |||
| US 20150328214 A1, 19.11.2015. | |||

