ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к новой рекомбинантной генной последовательности, гибридному белку, экспрессируемому с этого гена, и вакцине, полученной с этим геном, а также различным генным вакцинам, получаемым заменой гена антигена в рекомбинантной последовательности, или применению продуктов экспрессии в качестве вакцин гибридных пептидов.
УРОВЕНЬ ТЕХНИКИ
С момента открытия вторичного лимфоидного хемокина человека (SLC, от англ. secondary lymphoid tissue chemokine) ему было уделено очень много внимания. SLC регулирует секрецию цитокинов, изменяет иммуносупрессорные статусы микроокружения опухоли и способствует развитию Th1. Таким образом, SLC полезен в противоопухолевом иммунитете. Далее, SLC оказывает влияние на привлечение лимфоцитов и дендритных клеток (DC, от dendritic cells). Внутриопухолевая инъекция индуцирует местный и системный противоопухолевые иммунные ответы. SLC может также контролировать супрессию ангиогенеза в опухолях. Таким образом, SLC весьма полезен в иммунотерапии рака. Однако применение белка SLC самого по себе или трансгенный метод не дали удовлетворительных результатов в отношении противоопухолевого иммунитета. Предполагалось (Cancer Research 61, 197-205, January 1, 2001), что противоопухолевая иммунная реакция может быть усилена с помощью иммунизации мышей дендритными клетками, которые трансфицированы рекомбинантным ретровирусным вектором, несущим гены, кодирующие фрагмент IgG Fc и антиген опухоли. Механизм заключается в том, что гибридный белок, экспрессирующийся и секретирующийся из DC, повторно захватывается дендритными клетками через рецептор Fc (обозначен далее как FcR), и затем эпитоп гибридного белка презентируется Т-хелперным клеткам (Th) через молекулы МНС класса II и перекрестно презентируется цитотоксическим Т лимфоцитам (CTL) через молекулы МНС класса I, в результате чего индуцируется системно противоопухолевый гуморальный-и-клеточный иммунитет. Сообщалось (the Journal of Immunology, 1998, 161: 6059-6067), что способность DC no захвату антигенов с помощью FcR-опосредованного эндоцитоза в 10000 раз выше, чем с помощью пиноцитоза. Однако выделение, амплификация и трансфекция гена в DC увеличивает сложность производства вакцины. В этом случае дендритные клетки должны готовиться индивидуально, что затрудняет контроль качества и промышленное производство.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Чтобы преодолеть недостатки прототипов, целью изобретения является создание новой рекомбинантной генной конструкции для индукции иммунного ответа.
Другой целью данного изобретения является создание гибридного (слитого) белка, зкспрессируемого этой рекомбинантной генной конструкцией.
Другой целью данного изобретения является создание вакцины гибридного белка, кодируемого рекомбинантной генной конструкцией по изобретению, и вакцины рекомбинантного опухоль-хемотаксического антигена.
Для достижения целей изобретения предложены следующие технические решения.
Изобретение относится к рекомбинантной генной конструкции для индукции иммунного ответа, содержащей ген SLC человека, ген антигена и ген Fc-фрагмента IgG1, где ген SLC находится слева от гена антигена и ген Fc-фрагмента IgG1 находится справа от гена антигена. Указанный ген антигена содержит Her2/neu, P53, PSA, PAP, PSM, MAGE1, MAGE2, MAGE3, BAGE, GAGE1, GAGE2, CAG3, RAGE, NY-ESO-1, Тирозиназу, СЕА, Ig идиотип, gp100, melan A, gp75, TRP-1, TRP-2, CDK4, CASP-8, ras, bcr/abl, MUC-1 и гены, кодирующие белки, имеющие отношение к вирусам гепатита С, ВИЧ и патогенным микроорганизмам
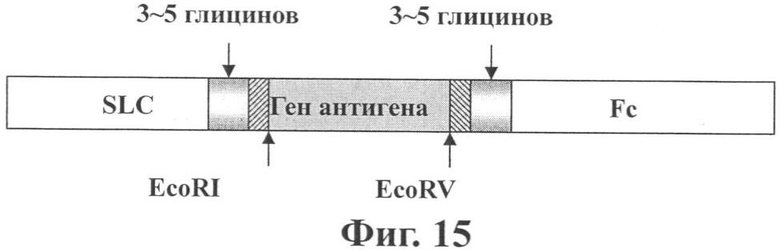
В рекомбинантной генной конструкции по изобретению сайты эндонуклеазы рестрикции EcoRI и гены трех глицинов находятся между геном SLC и геном антигена, а сайты эндонуклеазы рестрикции EcoRV и гены пяти глицинов находятся между геном антигена и геном Fc-фрагмента IgG1.
Рекомбинантная генная конструкция по изобретению содержит нуклеотидную последовательность SEQ ID NO:1 и SEQ ID NO:12.
Изобретение также относится к аминокислотной последовательности, кодируемой рекомбинантной генной конструкцией по изобретению. Эта аминокислотная последовательность содержит последовательность SEQ ID NO:2.
Изобретение также относится к аминокислотной последовательности, кодируемой SEQ ID NO:12. Эта аминокислотная последовательность представляет собой SEQ ID NO:13
Изобретение также относится к генной вакцине, содержащей рекомбинантную генную последовательность по изобретению.
Кроме того, изобретение также относится к вакцине. Предпочтительно вакцина содержит SEQ ID NO:1, SEQ ID NO:12 и соответствующие аминокислотные последовательности
Другими словами, изобретение относится к рекомбинантной генной конструкции, содержащей ген SLC человека, ген антигена и ген Fc-фрагмента IgG1, где ген SLC находится слева от гена антигена и ген Fc-фрагмента IgG1 находится справа от гена антигена. Указанный ген антигена содержит Her2/neu, Р53, PSA, PAP, PSM, MAGE1, MAGE2, MAGE3, BAGE, GAGE1, GAGE2, CAG3, RAGE, NY-ESO-1, Тирозиназу, CEA, Ig идиотип, gp100, melanA, gp75, TRP-1, TRP-2, CDK4, CASP-8, ras, bcr/abl, MUC-1 и другие гены, кодирующие белки, имеющие отношение к вирусам гепатита С, ВИЧ и другим патогенным микроорганизмам. Сайты эндонуклеазы рестрикции EcoRI и гены трех глицинов находятся между геном SLC и геном антигена, а сайты эндонуклеазы рестрикции EcoRV и гены пяти глицинов находятся между геном антигена и геном Fc-фрагмента IgG1.
Изобретение также относится к применению рекомбинантной генной конструкции по изобретению в приготовлении вакцин. А именно, вакцине, содержащей рекомбинантную генную конструкцию по изобретению, в частности последовательности нуклеотидов SEQ ID NO:1, SEQ ID NO:12 и соответствующие аминокислотные последовательности SEQ ID NO:2 и SEQ ID NO:13.
Другими словами изобретение относится к рекомбинантной генной конструкции, содержащей человеческий ген SLC, ген антигена и ген Fc-фрагмента IgG1 (см. Фиг. 13). Ген SLC помещен до гена антигена, и вставлены сайты эндонуклеазы рестрикции EcoRI и гены трех глицинов. Ген Fc-фрагмента IgG1 помещен после гена антигена, и вставлены сайты эндонуклеазы рестрикции EcoRV и гены пяти глицинов.
Согласно изобретению замена гена антигена в указанных генах по сайтам эндонуклеазы рестрикции может привести к различным гибридным генам хемотаксического антигена. В частности, это изобретение относится к рекомбинантному гену, содержащему последовательность нуклеиновой кислоты SEQ ID NO:1. Последовательность нуклеиновой кислоты SEQ ID NO:1 представляет собой искусственно лигированную последовательность SLC-Her2/P53-Fc, где ген SLC находится с положения 1 до положения 402 нуклеиновой кислоты; лигирующий ген Her2 находится с положения 418 до положения 711 нуклеиновой кислоты. Нуклеотиды с положения 418 до положения 444 в рекомбинантном гене соответствуют нуклеотидам с положения 1105 до положения 1131 в открытой рамке считывания (ОРС) гена Her2/neu. Нуклеотиды с положения 445 до положения 546 в рекомбинантном гене соответствуют нуклеотидам с положения 244 до положения 345 в открытой рамке считывания (ОРС) гена Her2/neu, куда включена мутация с G на С в положении 250. Нуклеотиды с положения 547 до положения 711 в рекомбинантном гене соответствуют нуклеотидам с положения 1333 до положения 1479 в ОРС гена Her2/neu. В рекомбинантном гене часть гена Р53 находится с положения 712 до положения 1113, соответствующим нуклеотидам с положения 475 до положения 876 ОРС р53. Ген IgG Fc (включая интроны) находится с положения 1147 до положения 2057 в рекомбинантном гене, причем два интрона находятся с положения 1189 до положения 1309 и с положения 1640 до положения 1739 соответственно. Последовательность нуклеиновой кислоты с положения 1147 до положения 1189 является последовательность. шарнира С-области. Последовательность нуклеиновой кислоты с положения 1310 до положения 1369 является последовательностью для СН2, последовательность нуклеиновой кислоты с положения 1740 до положения 2057 является последовательностью для СН3. Три гена глицинов и сайты EcoRI эндонуклеазы рестрикции вставлены перед фрагментом Неr2/Р53, и пять генов глицинов и сайты EcoRV эндонукпеазы рестрикции вставлены после Her2/Р53. Согласно этому изобретению, чтобы сделать нормальной биологическую функцию экспрессированных факторов, 3~5 генов глицинов вставлены перед и после (справа и слева от) гена антигена. Эффект генов глицина заключается в том, чтобы предотвратить интерференцию рекомбинантного антигена с трехмерной структурой.
Данное изобретение также относится к аминокислотной последовательности (612 а.к., зрел589а.к.) SEQ ID NO:2, кодируемой нуклеотидной последовательностью SEQ ID NO:1, где SEQ ID NO:1 представляет собой искусственно рекомбинантный SLC-Her2/P53-Fc ген. В указанной аминокислотной последовательности фрагмент SLC находится с положения 1 до положения 134, три глицина находятся с положения 135 до положения 137, и часть пептида Неr2 находится с положения 140 до положения 237. Аминокислоты с положения 140 до положения 148 в гибридном белке соответствуют аминокислотам с положения 369 до положения 377 в Her2/neu. Аминокислоты с положения 149 до положения 182 в гибридном белке соответствуют аминокислотам с положения 82 до положения 115 в Her2/neu, где имеется мутация с V на L в положении 84. Аминокислоты с положения 183 до положения 238 в гибридном белке соответствуют аминокислотам с положения 445 до положения 499 в Her2/neu. Часть пептида Р53 находится с положения 238 до положения 371 в гибридном белке (соответствуя аминокислотам с положения 159 до положения 292 в природной форме Р53). Пять глицинов находятся с положения 374 до положения 378. IgG Fc находится с положения 383 до положения 612, где последовательность с положения 383 до положения 396 представляет собой петлю С-области, последовательность с положения 397 до положения 506 является последовательностью для СН2, и последовательность с положения 507 до положения 612 является последовательностью для СН3.
Данное изобретение также относится к другому рекомбинантному гену для вакцины против рака простаты, последовательность которого показана в SEQ ID NO: 12. Нуклеотидная последовательность SEQ ID NO: 12 представляет собой последовательность искусственно лигированного SLC-PSM-mPAP-PSA-Fc (SLC-3Р-Fc) гена, куда введены только частичные последовательности PSM (human Prostate Specific Membrane Antigen), mPAP (mouse Prostatic Acid Phosphatase) и PSA (human Prostate Specific Antigen). В гибридом гене ген SLC находится с положения 1 до положения 402; часть гена PSM находится с положения 418 до положения 603, соответствуя последовательности с положения 1987 до положения 2172 в открытой рамке считывания (ОРС) PSM; ген mPAP находится с положения 604 до положения 759, соответствуя последовательности с положения 328 до положения 484 в mPAP ОРС; ген PSA находится с положения 760 до положения 981, соответствуя последовательности с положения 151 до положения 372 в PSA ОРС; ген IgG Fc (включая интроны) находится с положения 1003 до положения 1925, причем эти два интрона находятся с положения 1057 до положения 1177 и с положения 1508 до положения 1607 соответственно. Гены трех глицинов и сайты EcoRI эндонуклеазы рестрикции вставлены перед (слева от) 3P, и гены пяти глицинов и сайты EcoRV эндонуклеазы рестрикции вставлены после (справа от) 3P.
Данное изобретение также относится к аминокислотной последовательности SEQ ID NO: 13, кодируемой нуклеотидной последовательностью SEQ ID NO: 12. В частности, изобретение относится к аминокислотной последовательности (568 а.к.), соответствующей искусственно рекомбинантному гену SLC-3P-Fc. В этой гибридной аминокислотной последовательности SLC находится с положения 1 до положения 134, три глицина находятся с положения 135 до положения 137, PSM находится с положения 140 до положения 201, часть пептида mPAP находится с положения 202 до положения 253, часть пептида PSA находится с положения 254 до положения 327, пять глицинов находятся с положения 374 до положения 378, и IgG1-Fc находится с положения 335 до положения 568.
Данное изобретение также относится к применению рекомбинантной генной последовательности по изобретению, включая SEQ ID NO:1, в получении вакцин.
Как может быть видно из вышесказанного, стратегия для хемотаксического антигена по изобретению является следующей: присоединение хемокина SLC слева от антигена и присоединение Fc иммуноглобулина IgG справа от антигена; введение гибридного гена, состоящего из трех частей, непосредственно в здоровую или опухолевую ткань для экспрессии и секреции; образование состоящего из двух частей объединенного тела через шарнирную область Fc. Указанный рекомбинантный белок активно привлекает DC и Т клетки благодаря хемотаксической активности SLC in vivo. DC захватывает гибридный белок с высокой эффективностью с помощью пиноцитоза, особенно с помощью рецептора Fc и SLC рецептора CCR7. При этом DC несет антиген в клетку. После процессинга и обработки антиген представляется Th и CTL с помощью молекул MHC-II или MHC-I. И, наконец, полностью индуцируется специфический противоопухолевый гуморальный и клеточный иммунный ответ. Кроме того, DC может быть активирована комбинацией фрагмента Fc с Fc рецептором DC. SLC может активно извлекать DC и Т лимфоциты, а также способствовать секреции цитокинов, таких как ИЛ-12, интерферон-γ и интерферон-индуцируемый белок (IP-10), способствовать развитию Th1, подавлять продуцирование TGF-β и VEGF. Таким образом, SLC очень полезен для противоопухолевого иммунитета. По вышеописанным причинам такая комбинация обладает синергическим эффектом и может индуцировать более сильный противоопухолевый иммунный ответ (см. Фиг.1).
В данном изобретении использованы различные рекомбинантные гены, но ими это изобретение не ограничивается. Исследование контроля выявило, что и в форме плазмиды, и в форме гибридного белка рекомбинантный хемокиновый антиген может быть получен промышленным путем. Стадия приготовления DC в данной заявке была опущена. Однако DC могут быть с высокой эффективностью нацелены in vivo. Более того, введение сайтов эндонуклеазы рестрикции с двух сторон гена антигена делает возможной замену антигена для получения вакцин различных хемокиновых антигенов против опухолевых и неопухолевых заболеваний. Например, новая вакцина хемокинового антигена может быть получена заменой антигена HP в SLC-HP-Fc на антиген, относящийся к раку простаты.
Рекомбинантный ген может быть сконструирован в эукариотических векторах с использованием непосредственной инъекции плазмиды в тело или опухоль. Другие методики, такие как генная пушка или электрический импульс, могут увеличить степень трансфекции и эффективность экспрессии. Сконструированный рекомбинантный вирус, такой как ретровирус, аденовирус, аденоассоциированный вирус, вирус коровьей оспы, вирус простого герпеса и тому подобные, также могут быть использованы как вакцины для инъекции in vivo. Эукариотические клетки (например СНО) могут быть трансфицированы плазмидами и культивированы in vitro для экспрессии и секреции. Благодаря фрагменту Fc гибридный белок может быть легко и эффективно выделен и очищен методами аффинной хроматографии и использован в качестве белковой вакцины.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 показывает схему механизма генной вакцины хемотаксического антигена по изобретению.
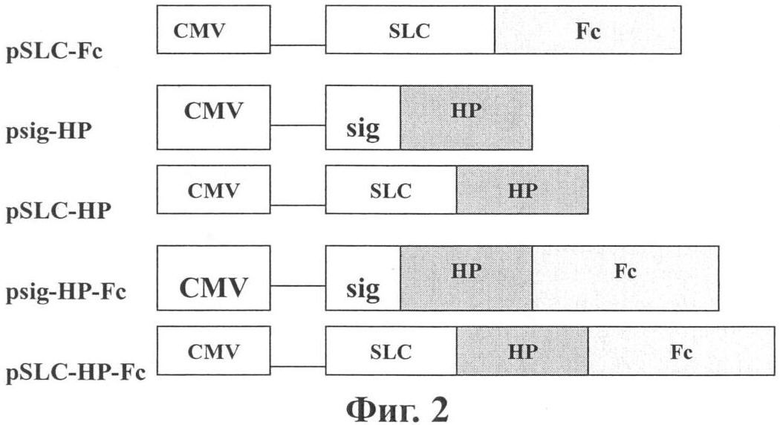
Фиг.2 показывает конструкции плазмид. HP обозначает Her2/neu-р53 гибридный ген; sig обозначает сигнальный пептид.
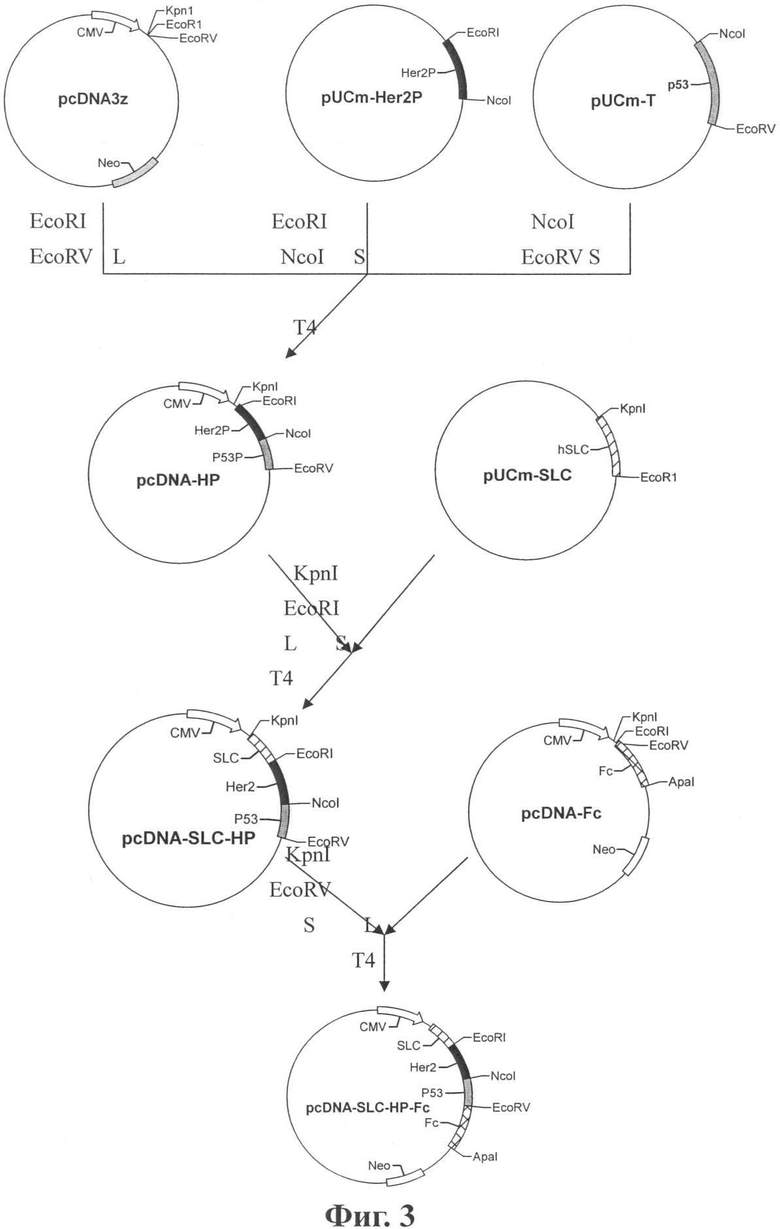
Фиг.3 показывает рекомбинацию SLC-Her2/neu-Fc по изобретению.
Фиг.4 показывает определение вестерн-блотингом клеточной экспрессии и секреции гибридного белка в клетках, трансфицированных генной вакциной pSLC-HP-Fc. А обозначает лизаты клеток, трансфицированных pSLC-HP-Fc; В обозначает супернатант культуры клеток, трансфицированных pSLC-HP-Fc; С обозначает супернатант культуры клеток, трансфицированных pcDNA.
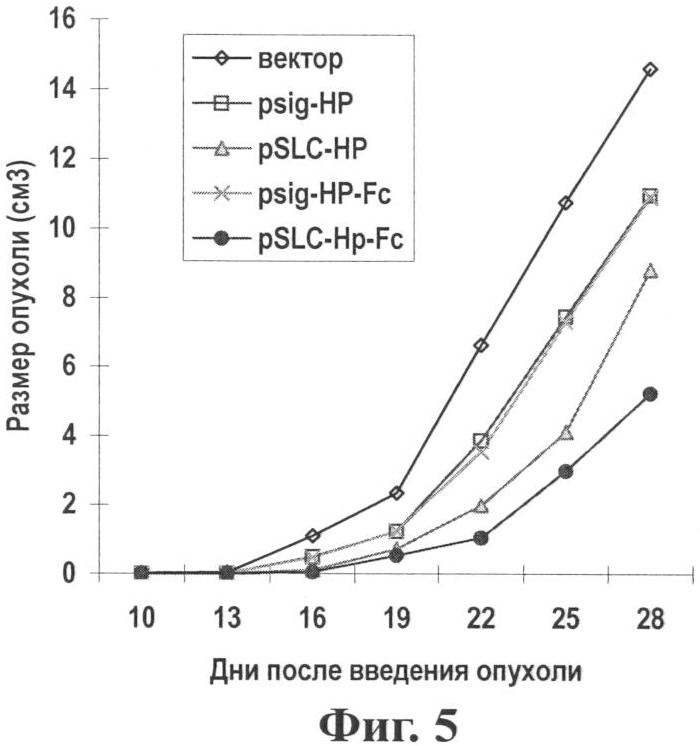
Фиг.5 показывает эффект по ингибированию опухоли in vivo с помощью предварительной вакцинации генной вакциной опухоль-хемотаксического антигена (pSLC-HP-Fc).
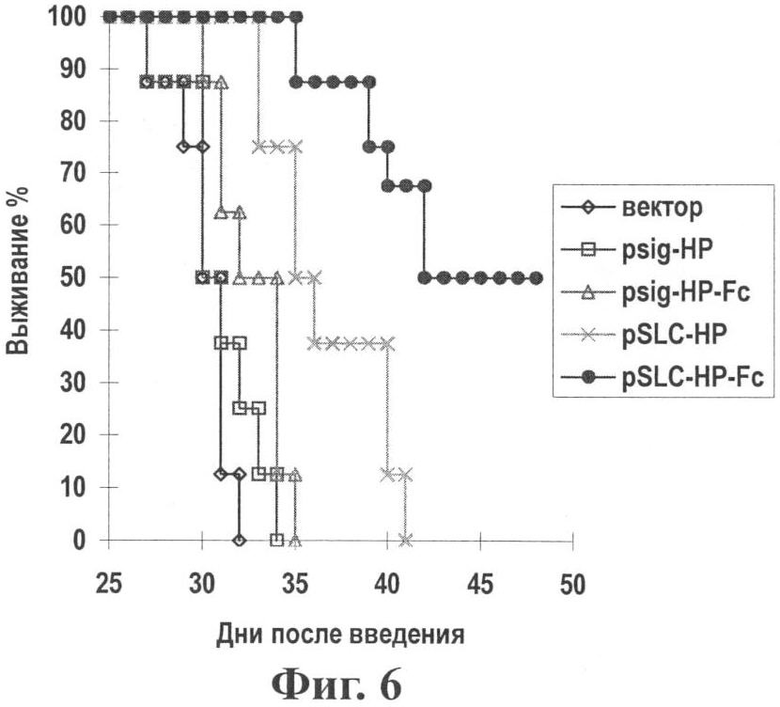
Фиг.6 показывает время выживания мышей, которым введена опухоль, с помощью предварительной вакцинации генной вакциной опухоль-хемотаксического антигена (pSLC-HP-Fc).
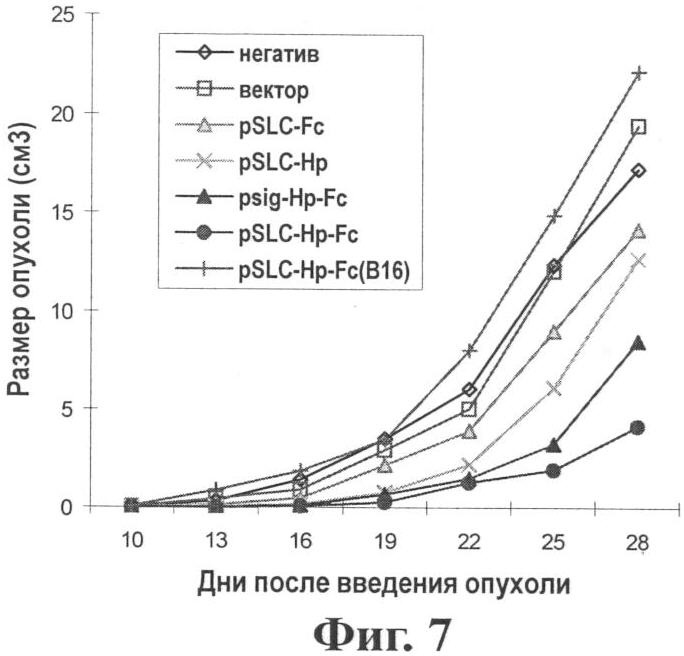
Фиг.7 показывает эффект по ингибированию опухоли in vivo при воздействии генной вакциной опухоль-хемотаксического антигена (pSLC-HP-Fc).
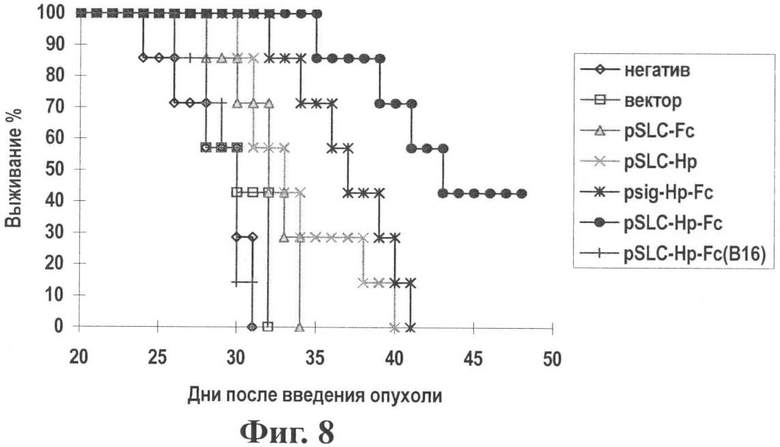
Фиг.8 показывает время выживания при воздействии генной вакциной опухоль-хемотаксического антигена (pSLC-HP-Fc).
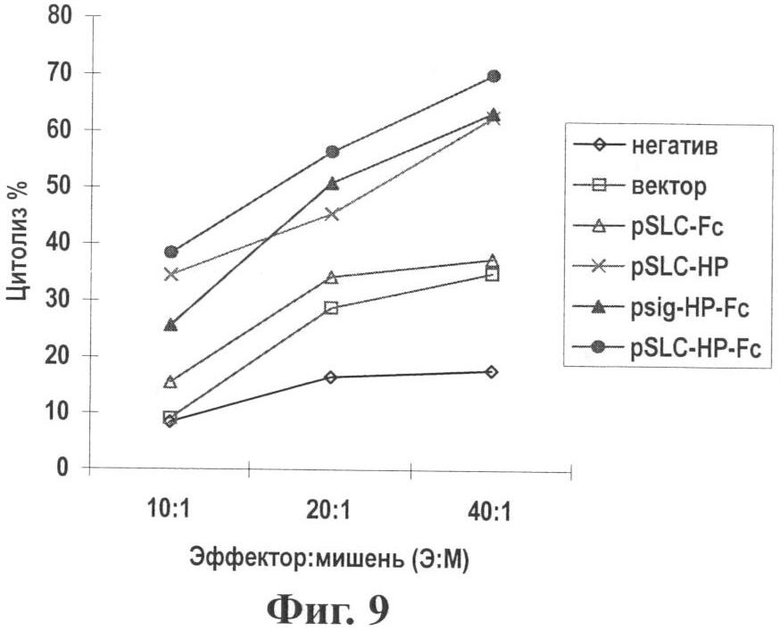
Фиг.9 показывает активность CTL после иммунизации генной вакциной опухоль-хемотаксического антигена (pSLC-HP-Fc).
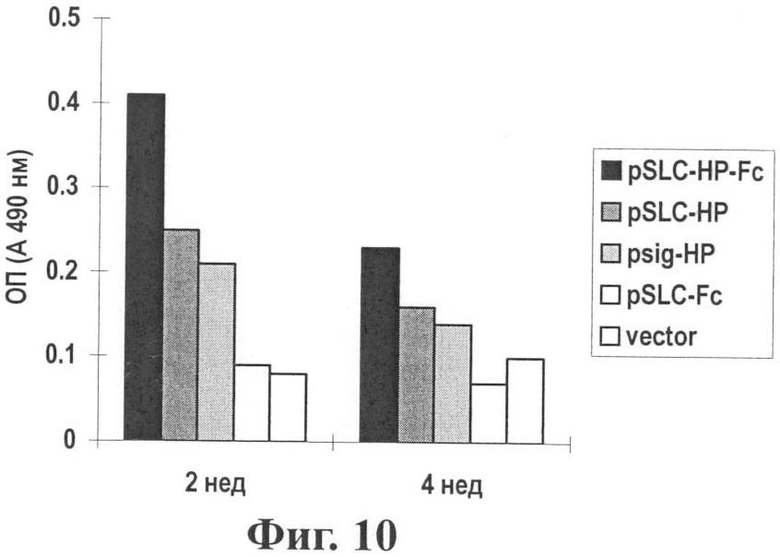
Фиг.10 показывает количества специфичных антител к HP в сыворотке через 2 и 4 недели после вакцинации. Сыворотку мышей разводили в отношении 1:100. Использовали анализ ELISA.
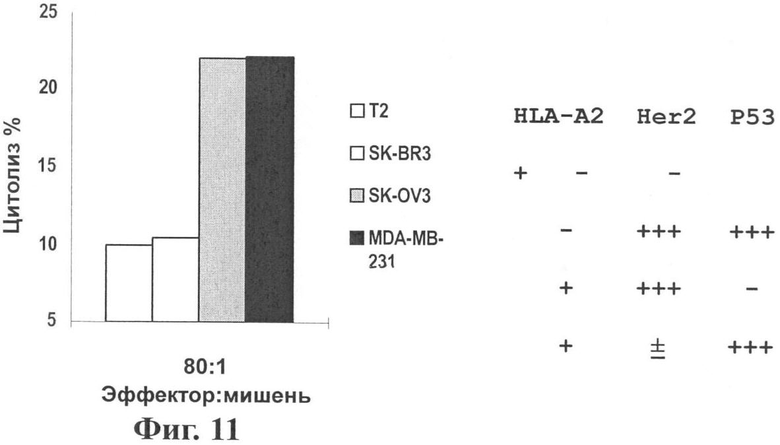
Фиг.11 показывает, что вакцина pSLC-HP-Fc может индуцировать CTL специфически против Her2/neu и р53 in vitro.
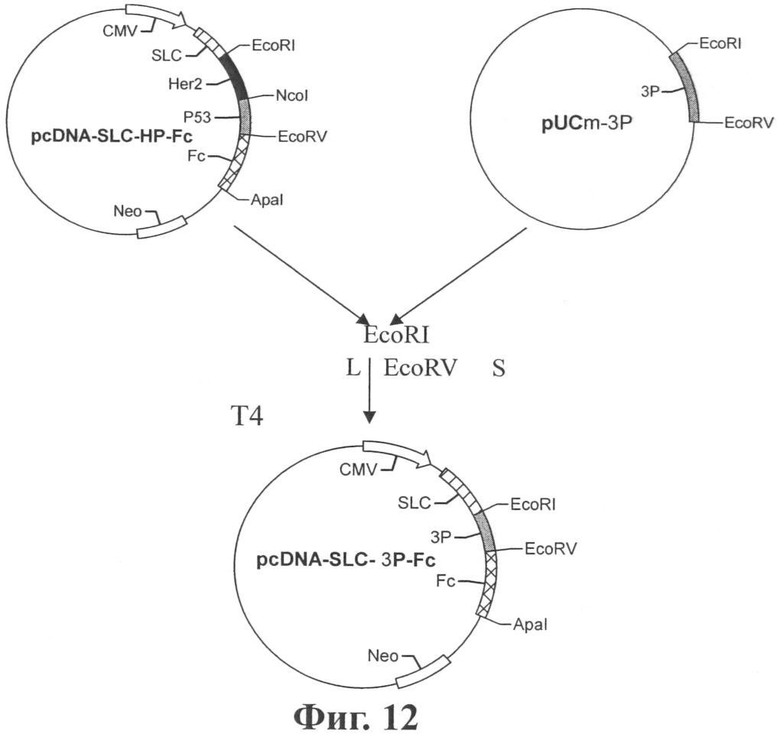
Фиг.12 показывает конструкцию плазмиды pSLC-3P-Fc.
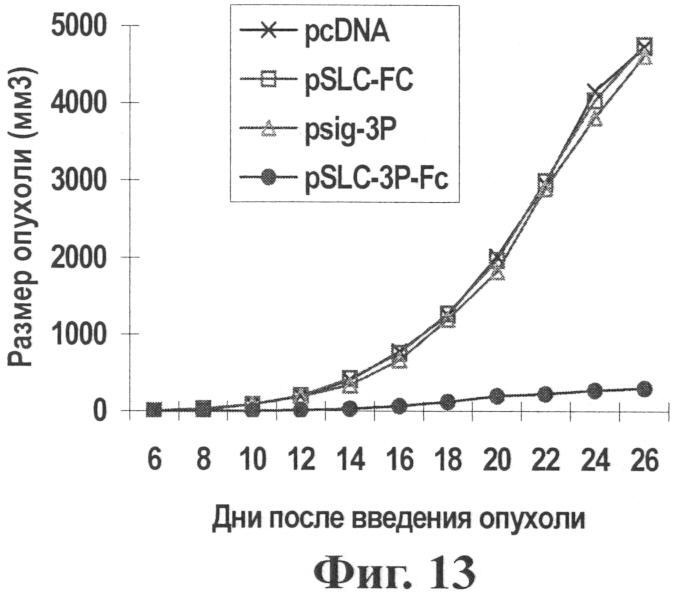
Фиг.13 показывает эффект по ингибированию опухоли in vivo после воздействия генной вакциной опухоль-хемотаксического антигена (pSLC-3P-Fc).
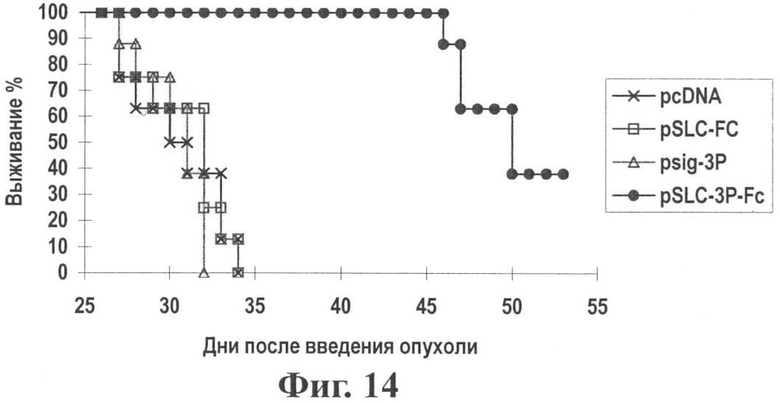
Фиг.14 показывает время выживания после воздействия генной вакциной опухоль-хемотаксического антигена (pSLC-3P-Fc).
Фиг.15 показывает принципиальную структуру рекомбинантного гена по изобретению.
ПОДРОБНОЕ РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Изобретение будет описано далее в деталях и проиллюстрировано. Необходимо понимать, что целью показанных воплощений является лишь пояснить, но никак не ограничить данное изобретение.
Пример 1. Клонирование человеческого гена SLC
мРНК экстрагировали из лимфатических узлов пациента, страдающего раком. С использованием методов, хорошо известных специалистам, сайт Kpnl эндонуклеазы рестрикции был добавлен к прямому праймеру SLC (5'-cggtaccacagacatggctcagtcac-3') и последовательность, кодирующая три глицина и сайт EcoRI эндонуклеазы рестрикции, была добавлена к обратному праймеру SLC (5'-taaattctcctcctcctggccctttagg-3'). Ген SLC был получен ОТ-ПЦР амплификацией. Полученный фрагмент был очищен и вставлен в вектор pUCmT для секвенирования (pUC-SLC). Результаты показывают, что последовательность полученного фрагмента соответствует запланированной последовательности (подчеркнутый фрагмент).
В данном изобретении секвенирование продуктов ПЦР осуществляли с использованием 377 ДНК секвенатора (фирма ABI) методом четырехцветного флуоресцентного терминаторного секвенирования, основанного на методе Сэнгера терминирующих дидезоксинуклеотидов. Последовательность с положения 36 до положения 464 представляет собой последовательность клонированного фрагмента по изобретению (ссылка на SEQ ID NO: 3).
Пример 2. Приготовление и соединение гена Her-2/neu
Сначала два фрагмента гена Her-2/neu клонировали методом ПЦР. Первый фрагмент получали следующим образом: использовался прямой праймер 5'-agaattcaagatctttgggagcctggcatttctgggctacctgctcatcgctcac-3', несущий сайт EcoRI эндонуклеазы рестрикции, и использовался обратный праймер 5'-gatgcccagcccttgca gggccagggcatagttgtc-3'. Плазмиду, содержащую внеклеточный сегмент гена neu, взяли в качестве матрицы и амплифицировали фрагмент, соответствующий аминокислотной последовательности с положения 82 до положения 115, где аминокислота V в положении 103 была заменена аминокислотой L. Второй фрагмент был получен следующим образом: использовали прямой праймер 5'-ctgcaagggctgggcatc-3' и обратный праймер, несущий сайт NcoI эндонуклеазы рестрикции 5'-tccatggcccggttggcagtgtggag-3'. Был амплифицирован фрагмент, соответствующий аминокислотной последовательности с положения 445 до положения 499.
Два продукта ПЦР собрали и соединили в ПЦР системе. Далее полученный фрагмент был использован в качестве матрицы, и с использованием прямого праймера первого фрагмента и обратного праймера второго фрагмента был амплифицирован гибридный ген размером 294 п.н. Затем полученный гибридный ген был вставлен в вектор pUCmT (pUC-Her2P) и секвенирован. Результаты показывают, что последовательность полученного гибридного гена соответствует запланированной последовательности (подчеркнутая последовательность).
Результат показан в SEQ ID NO: 4, где последовательность с положения 121 до положения 465 представляет собой последовательность клонированного фрагмента по изобретению.
Пример 3. Клонирование сегмента гена р53
Фрагмент гена 402 п.н., кодирующий аминокислоты с положения 156 до положения 289 белка р53, был клонирован с плазмиды, содержащей полноразмерный ген р53. Использовали следующие праймеры: прямой праймер с одним сайтом NcoI эндонуклеазы рестрикции представлял собой 5'-асс atg gcc atc tac aag cag tca cag сас atg ac-3'; обратный праймер с одним сайтом EcoRV эндонуклеазы рестрикции представлял собой 5'-tga tat ctt tct tgc gga gat tct ctt c-3'. Полученный фрагмент вставили в вектор pUCmT (pUC-p53P) для секвенирования. Результаты показывают, что последовательность полученного фрагмента соответствует ожидаемой последовательности (подчеркнутый сегмент).
Результат показан в SEQ ID NO: 5, где последовательность с положения 39 до положения 445 представляет собой последовательность фрагмента по изобретению.
Пример 4. Соединение фрагментов Her2/neu и р53
Как показано на Фиг.3, меньший фрагмент из pUC-Her2P, разрезанной с помощью EcoRI/NcoI, и меньший фрагмент из pUC-p53P, разрезанной с помощью NcoI/EcoRV, были лигированы с большим фрагментом из pcDNA3z, разрезанной с помощью EcoRI/EcoRV, чтобы получить плазмиду pcDNA-Her2/p53, называемую также pcDNA-HP. HP означает ген, лигированный из Her2/neu и р53. Искусственно лигированный фрагмент Her2/neu-p53 содержит гены, кодирующие пептид, связывающий молекулы HLA-I и HLA-II.
Пример 5. Конструкция PcDNA-Fc
Сначала химически синтезированный полилинкер был использован, чтобы заменить полилинкер pcDNA3. 1.
Был использован следующий способ конструирования.
Химически синтезировали два олигонуклеотида.
Прямой праймер:
5'-GCTAGCGAAGCTTTGGTACCGTAGGATCCACGAATTCAGTCCA-GGATATCGGCGGTGG-3'
Обратный праймер:
5'-GGTTTAAACGTTAACCCCGGGCCCTCGAGCTCTAGAGCCTCCT-CCACCGCCGATATC-3'
Последние 15 оснований на 3' концах каждого праймера комплементарны друг другу. Продукт ПЦР получали в системе со смесью двух праймеров в соотношении 1:1, ДНК полимеразой Taq и dNTP при 45°С для ренатурации и элонгации. Очищенный продукт вставляли в вектор pUCmT для секвенирования. Результаты показывают, что последовательность полученного продукта соответствует запланированной последовательности. Подчеркнутая последовательность показывает синтезированный сайт поликлонирования. Окончательный сайт поликлонирования представлял собой: «Nhel-HindIII-Kpn1-BamH1-EcoR1-EcoRV- глицин × 5 -Xbal- Xho1- Apa1-Sma1- Hpa1-Pme1». В этой последовательности небольшой фрагмент гена (GGCGGTGGAGGAGGC), кодирующий пять глицинов, был вставлен методу сайтам EcoRV и Xbal. Разрезали клонированную плазмиду Nhel/Pmel и получили фрагмент размером около 100 п.н. Затем этот фрагмент лигировали с большим фрагментом из pcDNA3. 1, разрезанной с помощью Nhel/Pmel, чтобы получить новый вектор, названный pcDNAf.
Результаты секвенирования химически синтезированного полилинкера показаны в SEQ ID NO: 6, где последовательность с положения 3 до положения 100 представляет собой последовательность фрагмента по изобретению.
Способ конструирования плазмиды PcDNA-Fc
Фрагмент Fc был вырезан из плазмиды, содержащей IgG1 Fc, с помощью XbaI/ApaI эндонуклеаз рестрикции и вставлен в вектор pcDNAf, который был обработан теми же самыми эндонуклеазами. Полученная конструкция была названа PcDNA-Fc. Последовательность IgG1 Fc была подтверждена секвенированием полученной плазмиды.
Секвенирование фрагмента Fc
Фрагмент Fc размером 900 п.н. был амплифицирован с плазмиды, содержащей ген IgG1 Fc, с помощью ПЦР и вставлен в вектор pUCmT для секвенирования. Для амплификации использовали следующие праймеры: прямой праймер: 5'-gaattcggagttaacgagcccaaatcttg-3'; обратный праймер: 5'-gggccctcatttacccggagac-3'. Результаты секвенирования показывают, что плазмида содержит шарнирную область и области СН2 и СН3 от IgG1. Это значит, что плазмида содержит Fc (см. подчеркнутую последовательность). Результат показан в SEQ ID NO:7, где последовательность с положения 78 до положения 991 представляет собой последовательность по изобретению.
Пример 6. Соединение гена SLC-Her2/neu-Fc
Как показано на Фиг.3, больший фрагмент из pcDNA-HP, разрезанной KpnI/EcoRI, был присоединен к меньшему фрагменту из pUC-SLC, разрезанной KpnI/EcoRI, для получения pcDNA-SLC-HP. Затем меньший фрагмент из pcDNA-SLC-HP, разрезанной KpnI/EcoRV, был присоединен к большему фрагменту из pcDNA-Fc, разрезанной KpnI/EcoRV, с получением конструкта pcDNA-SLC-HP-Fc (также названного pSLC-HP-Fc).
Конструкция контрольной плазмиды показана на Фиг.2
Пример 7. Определение вестерн-блотингом клеточного и секретируемого рекомбинантного белка в клетках, трансфицированных генной вакциной
Рекомбинантной плазмидой pSLC-HP-Fc были трансфицированы клетки B16-F10 методом липосом. Положительные клоны отобрали с помощью G418. Клетки культивировали в бессывороточной среде 1640 в течение 24 часов, и затем собрали и концентрировали супернатант. Метод вестерн-блотинга был использован для определения экспрессии гибридного белка. Использованное первичное антитело представляло собой поликлональное кроличье антитело к человеческому р53, вторичное антитело представляло собой конъюгированное с пероксидазой хрена козье анти-кроличье антитело. Блоты визуализируют с помощью набора для хемилюминисцентного определения ECL-Plus (Amersham Pharmacia Biotech). В заключение пленку подвергают действию рентгеновских лучей. Результаты показывают, что экспрессия гибридного белка может быть определена в супернатанте и клеточном лизате В16-Р10 клеток, трансфицированных вектором pSLC-HP-Fc. Молекулярный вес соответствует ожидаемому (см. Фиг.4). Было подтверждено, что последовательность гибридного белка представляет собой SEQ ID NO: 2.
Пример 8. Клонирование гена человеческого простат-специфичного мембранного антигена PSM (Prostate Specific Membrane Antigen)
Тотальная РНК была экстрагирована из клеточной линии человеческого рака простаты LNCaP. ОТ-ПЦР была осуществлена с использованием прямого праймера (5'-ACTCGAGAT GAAGACATACAGTGTATC-3') и обратного праймера (5'-TGATATCTTAGGCTAC TTCACTCAAAG-3'). После электрофореза была вырезана полоса hPSM размером 390 п.н. Фрагмент был очищен и извлечен методом Glass Milk. Очищенный фрагмент был лигирован в вектор pUCm-T с получением pUCm-hPSM. Положительный клон были взят и секвенирован. Результаты соответствуют ожидаемым. Если взглянуть на SEQ ID NO: 8, полученный фрагмент гена соответствует последовательности с положения 1864 до положения 2253 ОРС человеческого PSM. Последовательность с положения 229 до положения 618 в полученной последовательности является последовательностью по изобретению.
Пример 9. Клонирование фрагмента гена человеческого простат-специфичного антигена PSA (Prostate Specific Antigen)
Тотальная РНК была экстрагирована из клеточной линии человеческого рака простаты LNCaP. ОТ-ПЦР была осуществлена с использованием прямого праймера (5'-TCTCGAGGGC GGTGTTCTGGTGCA-3') и обратного праймера (5'-AGATATCATGTCC AGCGTCCAGCAC-3'). После электрофореза была вырезана полоса размером около 600 п.н. Фрагмент был очищен и извлечен методом Glass Milk. Полученный фрагмент был вставлен в вектор pUCm-T с получением pUCm-PSA. Положительный клон были взят и секвенирован. Результаты показаны в SEQ ID NO: 9. Полученный фрагмент гена соответствует последовательности с положения 151 до положения 609 ОРС человеческого PSA. Последовательность с положения 45 до положения 503 в полученной последовательности является последовательностью по изобретению.
Пример 10. Клонирование фрагмента гена мышиной PAP (Prostatic Acid Phosphatase)
Тотальная РНК была экстрагирована из ткани мышиной простаты. ОТ-ПЦР была осуществлена с использованием прямого праймера (5'-TCTAGATGAGAGCTGTTCCTCTG-3') и обратного праймера (5'-GGGCCCTTAATTCCGTCCTTGGTG-3'). После электрофореза была вырезана полоса размером 1146 п.н. Фрагмент был очищен и извлечен методом Glass Milk. Полученный фрагмент был вставлен в вектор pUCm-T с получением pUCm-mPAP. Положительный клон были взят и секвенирован. Результаты показаны в SEQ ID NO: 10. Последовательность с положения 48 до положения 1193 в полученной последовательности является последовательностью по изобретению.
Пример 11. Соединение фрагментов генов PSM, mPAP и PSA (3P)
Фрагмент гена hPSM был клонирован с плазмиды pUCm-hPSM с помощью ПЦР со следующими праймерами: прямой праймер, соответствующий последовательности с положения 1987 до положения 2007 ОРС hPSM (вставлен сайт EcoRI эндонуклеазы рестрикции) представлял собой 5'-AGAATTCATGATGAATGATCAACTCATG-3'; обратный праймер представлял собой 5'-AGCACTCATCAAAGTCCTGGCCTTGGAAGGGTCCAC-3' (подчеркнутая часть соответствует последовательности с положения 328 до положения 345 ОРС mPAP, остальная часть соответствует последовательности с положения 2155 до положения 2172 ОРС hPSM). После электрофореза полученный фрагмент hPSM был вырезан и очищен методом Glass Milk. Фрагмент гена mPAP был клонирован с плазмиды pUCm-mPAP с помощью ПЦР со следующими праймерами: прямой праймер, соответствующий последовательности с положения 328 до положения 348 OPC mPAP представлял собой 5'-AGGACTTTGATGAGTGCTATG-3'; обратный праймер, соответствующий последовательности с положения 463 до положения 483 OPC mPAP представлял собой 5'-AGGGCAGTCTCTGAAAGGCAG-3'. После электрофореза полученный фрагмент mPAP был вырезан и очищен методом Glass Milk. Фрагмент гена hPSA был клонирован с плазмиды pUCm-hPSA с помощью ПЦР со следующими праймерами: прямой праймер представлял собой 5'-CCTTTCAGAGACTGCCCTGGCGGTGTTC TGGTGCAC-3' (подчеркнутая часть соответствовала последовательности с положения 466 до положения 483 OPC mPAP, и оставшаяся часть соответствовала последовательности с положения 151 до положения 168 OPC hPSA); обратный праймер представлял собой 5'-AGATATCGAGCAGCATGAGGTCGT-3'. После электрофореза полученный фрагмент hPSA был вырезан и очищен методом Glass Milk.
Праймер 5'-AGAATTCATGATGAATGATCAACTCATG-3' был использован для амплификации hPSM, а праймер 5'-AGGGCAGTCTCTGAAAGGCAG-3' был использован для амплификации mPAP индивидуально в 25 мкл системы на 10 циклов, чтобы получить одиночные цепи. Затем две системы были смешаны для дальнейшей ПЦР в 50 мкл системы на 18 циклов. После электрофореза полученный фрагмент hPSM-mPAP размером 352 п.н. был вырезан и очищен методом Glass Milk.
Праймер 5'-AGGACTTTGATGAGTGCTATG-3' (соответствующий последовательности с положения 328 до положения 348 OPC mPAP) был использован для амплификации mPAP, и праймер 5'-AGATATCGAGCAGCATGAGGTCGTG-3' (соответствующий последовательности с положения 355 до положения 372 OPC PSA) был использован для амплификации hPSA индивидуально в 25 мкл системы на 10 циклов, чтобы получить одиночные цепи. Затем две системы были смешаны для дальнейшей ПЦР в 50 мкл системы на 18 циклов. После электрофореза полученный фрагмент mPAP-hPSA размером 378 п.н. был вырезан и очищен методом Glass Milk.
Праймер 5'-AGAATTCATGATGAATGATCAACTCATG-3' был использован для амплификации hPSM-mPAP, и праймер 5'-AGATATCGAGCAGCATGAGGTCGTG-3' был использован для амплификации mPAP-hPSA индивидуально в 25 мкл системы на 10 циклов, чтобы получить одиночные цепи. Затем две системы были смешаны для дальнейшей ПЦР в 50 мкл системы на 18 циклов. После электрофореза полученный фрагмент hPSM-mPAP-hPSA размером 564 п.н. был вырезан и очищен методом Glass Milk. Полученный фрагмент был вставлен в вектор pUCm-TV для секвенирования. Результаты соответствуют ожидаемыми. Смотри SEQ ID NO: 11, где последовательность с положения 54 до положения 629 является последовательностью по изобретению.
Пример 12. Рекомбинация гена хемокинового антигена SLC-3P-Рс
После секвенирования плазмида, содержащая корректную генную последовательность 3P (3P обозначает гибридный ген PSM-PAP-PSA размером 564 п.н., кодирующий 188 аминокислот) была разрезана EcoRI/EcoRV эндонуклеазами рестрикции. Полученный меньший фрагмент 3P был лигирован с разрезанным EcoRI/EcoRV вектором pSLC-HP-Fc с получением конструкции pSLC-3P-Рс (см. Фиг.12). Последовательность рекомбинантного гена SLC-3P-Fc показана в SEQ ID NO: 12, которая соответствует аминокислотной последовательности, показанной в SEQ ID NO: 13.
Тестовые примеры
Тест 1. Определение хемоатаксической активности гибридного белка в отношении иммунных клеток в камере Бойдена
Извлечение активности человеческого SLC in vitro:
А. Клеточная линия меланолмы B16F10 была трансфицирована очищенной плазмидой pcDNA3z-SLC-HP-Fc и контрольной плазмидой. Через 48 часов инкубации собрали культуральную среду и концентрировали с помощью PEG20000 (в качестве негативного контроля был использован супернатант нетрансфицированных клеток B16-F10).
Б. Клетки мононуклеаров периферической крови (РВМС, от peripheral blood mononuclear cells) были собраны из 5 мл здоровой цельной периферической крови с помощью среды для отделения лимофцитов (Lymphocyte Separation Medium), промыты дважды физиологическим раствором и ресуспендированы в бессывороточной среде RPMI-1640. РВМС были подсчитаны и разбавлены до концентрации 1×106/мл.
В. Приготовление камеры Бойдена: камера была приобретена у Neuro Probe Co. Нижние лунки камеры заполнили 27 мкл кондиционированной культуральной среды, а верхние - 50 мкл клеточной суспензии 1×106/мл РВМС. Нижние и верхние лунки были разделены поликарбонатным фильтром (Neuro Probe) с размером отверстий 5 мкм. Камеру инкубировали при 37°С с 5% СО2 в течение 4 ч.
Г. Фильтр выгрузили, и клетки соскоблили с верхней стороны фильтра, а затем промыли, фиксировали и окрасили фильтр. Мигрировавшие клетки были подсчитаны в пяти произвольно выбранных полях большой мощности (×200). Хемотаксический индекс Cl (Chemotactic Index) был подсчитан следующим образом: Cl = число клеток пяти полей большой мощности в исследованных лунках / число клеток пяти полей большой мощности в контрольных лунках. Величина Cl негативной группы была 1.
Результаты: Уровень хемоатаксической активности бессывороточного супернатанта клеток, трансфицированных pSLC-HP-Fc, значительно выше, чем у других, что означает, что после трансфекции плазмида может быть экспрессирована, и экспрессируемый белок активен. Уровень хемоатаксической способности клетки, трансфицированной pSLC-HP-Fc, аналогичен таковому у клетки, трансфицированной pSLC, что означает, что гибридный белок не повреждает активности SLC.
Тест 2. Противоопухолевый иммунный ответ на генную вакцину человеческого хемотаксического антигена у мышей
Частицы золота были нагружены плазмидами (голыми генами) и нанесены на внутреннюю поверхность трубки Tefzel, разрезанной на подходящие сегменты для пуль, чтобы доставить 1 мкг ДНК, согласно инструкции BioRad. Было выделено несколько контрольных групп: контрольный вектор, pSLC-Fc, psig-HP (плазмида, экспрессирующая HP, содержащий сигнальный пептид SLC), pSLC-HP и psig-HP-Fc.
Тест 3. Иммунный ответ против рака
За 14 дней и за 7 дней до введения опухоли мышей вакцинировали дважды pSLC-HP-Fc или контрольными векторами в кожу брюшного отдела. На день 0 каждой мыши ввели 5×104 опухолевых клеток B16F10 (B16F10 клетки были трансфицированы геном HP и отобраны на G418) в 0,2 мл раствора. Размер опухоли записывали 2-3 раза в неделю. Размер опухоли подсчитывали в виде длина (см) × ширина (см2).
Результаты: Вакцинация дважды с помощью pSLC-HP-Fc значительно улучшала эффект подавления опухоли (см. Фиг.5, 6), и среднее время выживания мыши значительно продлялось. Терапевтический эффект pSLC-HP-Fc значительно лучше, чем у других векторов (р<0,05), что указывает на синергическое иммунное действие фрагментов SLC и Fc.
Тест 4. Иммунотерапия рака
На день 0 каждой мыши ввели 5×104 опухолевых клеток B16F10 в 0,2 мл раствора. В дни 6, 12 и 18 мыши были вакцинированы с помощью системы генной пушки (Gene Gun System). Размер опухоли записывали 2-3 раза в неделю. Размер опухоли оценивали, как указано выше.
Результаты: Как показано на Фиг.7 и 8, вакцина pSLC-HP-Fc демонстрирует наиболее эффективное подавление роста опухоли. На день 28 разница между группами pSLC-HP-Fc и psig-HP-Fc является значительной (Р<0,05). Среднее время выживания мыши продлялось при иммунизации pSLC-HP-Fc. Мышам в группе В16 (pSLC-HP-Fc) вводили клетки В16 дикого типа. После трех иммунизаций pSLC-HP-Fc, группа В16 показала отсутствие эффекта супрессии опухоли, означая, что иммунный ответ, индуцированный pSLC-HP-Fc, является специфичным к HP антигену.
Тест 5. CTL активность, индуцированная генной вакциной опухоль-хемотаксического антигена
Через две недели после трехкратной вакцинации системой генной пушки несущие опухоль мыши были убиты, и анализ на цитотоксичность CTL был проведен с использованием клеток селезенки с помощью LDH Kits (Promega). Отношение клеток селезенки и клеток-мишеней (отношение эффектор:мишень, Э:М) составило 40:1, 20:1 и 10:1 соответственно.
Результаты: Лимфоциты, происходящие из мышей, иммунизированных вакциной хемокинового нуклеотида, показали значительный цитотоксический эффект на клетки B16F10-HP (см. Фиг.9). Когда отношения Э:М были 40:1 или 20:1, имелась все еще значительная разница между исследуемой вакциной и контрольными векторами (Р<0,05), что подтверждает, что вакцина является более сильной в отношении индукции CTL активности.
Тест 6. Определение специфических антител в сыворотке иммунизированных мышей
Количество HP-специфичных антител в сыворотке группы иммунизированных мышей было определено с помощью ELISA через 2 и 4 недели после иммунизации (pSLC-HP-Fc). Вкратце, микротитрационные планшеты, покрытые рекомбинантным белком р53, были инкубированы в течение ночи, блокированы 20% телячьей сывороткой и инкубированы с первичными антителами (серийно разведенной мышиной сывороткой) и, в свою очередь, вторичными антителами (конъюгированными с пероксидазой хрена козьими антимышиными IgG). После промывания и визуализации с помощью O-фенилендиамина, величина ОП измерялась на 490 нм. Моноклональное антитело против р53 было использовано в качестве позитивного контроля, и нормальная мышиная сыворотка была использована в качестве негативного контроля.
Результаты: Уровень антитела р53 у мышей, иммунизированных SLC-HP-Fc, является наивысшим и значительно выше, чем уровень при других векторах (см. Фиг.10).
Тест 7. Специфические человеческие CTL против Her-2/neu или р53, индуцированные иммунизацией pSLC-HP-Fc, in vitro
С помощью центрифугирования с использованием среды для отделения лимфоцитов были собраны клетки мононуклеаров (MNCs, mononuclear cells) из цельной периферической крови здоровых доноров HLA-A2+. Половина MNCs была трансфицирована плазмидой pSLC-HP-Fc с использованием липосом и инкубирована с необработанными MNCs в отношении 1:1 при 37°С с 5% CO2. На день 6 были подсчитаны жизнеспособные клетки, ресуспендированы и культивированы с различными клетками-мишенями в различных соотношениях Э:М в 96-луночных микропланшетах. Анализ LDH (Promega Kits) был использован для анализа цитотоксичности.
Результаты: После смешения культур трансфицированные pSLC-HP-Fc MNC индуцировали специфичный CTL, чтобы убить клетки опухоли, которые сверхэкспрессируют Her2/neu или р53 МНС-рестриктирующим образом (см. Фиг.11). То есть только специфичные CTL в значительной степени убивают HLA-A2+ клетки опухоли, которые экспрессируют Her2/neu или р53, указывая на специфичность CTL, индуцированных генной вакциной хемотаксического антигена.
Тест 8. Противодействие раку простаты генной вакцины хемокинового антигена pSLC-3P-Рс in vivo
5×104 клеток рака В16-3P (трансфицированные pcDNA-3P клетки рака В16, отобранные на G418) были введены в бок мышам C57BL/6 (восемь мышей в каждой группе). Мыши были иммунизированы на третий, восьмой и тринадцатый день после введения клеток опухоли. В контрольных группах были использованы контрольные векторы pSLC-Fc и psig-3P (плазмида, содержащая ген сигнального пептида SLC слева от гена 3P). Наблюдали за размерами опухоли и временем выживания.
Результаты: Как показано на Фиг.13 и 14, генная вакцина хемокинового антигена (pSLC-3P-Fc) значительно ингибирует рост опухоли (по сравнению с контрольными группами Р<0,05) и значительно удлиняет время выживания (по сравнению с контрольными группами Р<0,05), означая, что изменение антигена в генной вакцине хемотаксического антигена может дать другие вакцины против соответствующего антигена.














Изобретение относится к области биотехнологии, конкретно к получению генно-инженерных вакцин, и может быть использовано в медицине. Получают рекомбинантную генную конструкцию, содержащую последовательно расположенные ген SLC человека, ген антигена и ген Fc-фрагмента IgG1. Изобретение позволяет значительно повысить эффективность иммунного ответа организма на вводимый антиген, по сравнению с известными антигенными конструкциями. 4 н. и 6 з.п. ф-лы, 1 табл., 15 ил.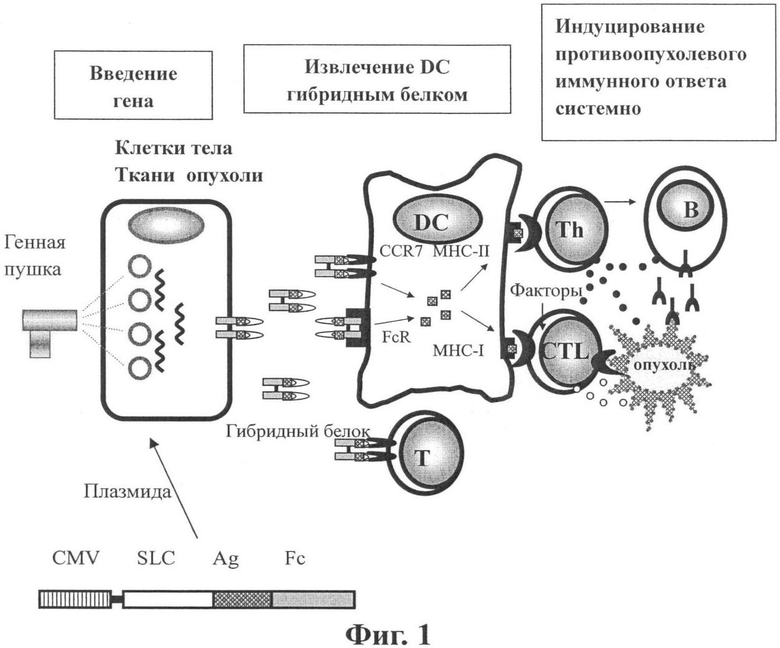
1. Рекомбинантная генная конструкция для индукции иммунного ответа, содержащая ген SLC человека, ген антигена и ген Fc-фрагмента IgG1, где ген SLC находится слева от гена антигена и ген Fc-фрагмента IgG1 находится справа от гена антигена.
2. Рекомбинантная генная конструкция по п.1, где указанный ген антигена включает Her2/neu, P53, PSA, PAP, PSM.
3. Рекомбинантная генная конструкция по п.1 или 2, где сайты эндонуклеазы рестрикции EcoRI и гены трех глицинов находятся между геном SLC и геном антигена, а сайты эндонуклеазы рестрикции EcoRV и гены пяти глицинов находятся между геном антигена и геном Fc-фрагмента IgG1.
4. Рекомбинантная генная конструкция по п.1, которая имеет последовательность SEQ ID NO:1.
5. Рекомбинантная генная конструкция по п.1, которая имеет последовательность SEQ ID NO:12.
6. Рекомбинантный полипептид для индукции иммунного ответа, характеризующийся аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:1.
7. Полипептид по п.6, где аминокислотная последовательность представляет собой SEQ ID NO:2.
8. Рекомбинантный полипептид для индукции иммунного ответа, характеризующийся аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:12, где аминокислотная последовательность представляет собой SEQ ID NO:13.
9. Вакцина для индукции иммунного ответа, содержащая рекомбинантную генную конструкцию по п.1 или соответствующий рекомбинантный полипептид.
10. Вакцина по п.9, где указанная генная конструкция представлена SEQ ID NO:1, SEQ ID NO:12, или их соответствующей аминокислотной последовательностью.
| WO 02058723 А2, 01.08.2002 | |||
| ЕО S.K | |||
| et aL, Immunopotentiation of DNA vaccine against herpes simplex virus via co-delivery of plasmid DNA expressing CCR7 ligands, Vaccine, 2001, v.19, n.32, p.4685-4693 | |||
| YOU Z | |||
| et al., Targeting dendritic cells to enhance DNA vaccine potency, Cancer Res., 2001, v.61, n.9, p.3704-3711 | |||
| KELLERMANN S.A | |||
| et al., The CC |

