Изобретение относится к биотехнологии, а именно к получению плюрипотентных клеток из клеток, выделенных из пуповинно-плацентарного комплекса млекопитающего. Полученные в результате «репрограммирования» плюрипотентные клетки могут найти широкое применение в медицинской практике и в изучении механизмов «репрограммирования» и «трансдифференцировки».
В 2006 г. в лаборатории Шинья Яманаки (Киото, Япония) в терминально дифференцированные клетки мыши (фибробласты) с помощью ретровирусов были введены гены четырех транскрипционных факторов (1). Через 16 дней исследователи обнаружили, что фибробласты меняют свою морфологию, меняется и поведение клеток в культуре. В результате проведенного отбора остались клетки, которые по своим свойствам и характеристикам напоминали ЭСК мыши. Они были способны дифференцироваться в клетки взрослого организма, а также колонизировать ткани животных после их введения в бластоцисту. По своему внешнему виду, свойствам и генетическому портрету они были близки, но не идентичны ЭСК, полученным естественным путем, поэтому они получили название iPS (induced pluripotency stem) стволовые клетки с индуцированной плюрипотентностью. Из 24 генов-кандидатов наилучшие результаты дала комбинация генов Oct3/4, Sox2, c-Myc и Klf4. Гены Oct3/4 и Sox2 кодируют транскрипционные факторы, которые функционируют на ранних стадиях эмбриогенеза, а в клетках взрослого организма они не работают. Ген c-Myc тоже кодирует транскрипционный фактор, но в отличие от двух предыдущих он не является специфичным, а участвует в контроле клеточного цикла и деления любой клетки и является протоонкогеном. Последний ген Klf4 также не имеет определенной временной или тканевой специфичности и кодирует транскрипционный фактор, может играть как роль активатора, так и репрессора транскрипции, однако функции его известны чрезвычайно мало. Если первые два гена являются весьма специфичными для ранних стадий эмбриогенеза, то два последних играют большую роль в процессах образования опухолей.
Использование протоонкогена c-Myc привело к повышенной частоте опухолеобразования у полученных животных, а эффективность процедуры получения изначальных клеток была очень низка.
В настоящее время уже получены iPS клетки из десятка специализированных тканей организма, в том числе и человека. Кроме эмбриональных и кожных фибробластов, в iPS были превращены нейрональные предшественники, эпителиальные клетки кишечника, гематопоэтические, мезенхимальные стволовые клетки, мышечные, кератиноциты, гепатоциты, герминальные и другие (2).
Несмотря на воспроизведение сходных экспериментов в различных лабораториях, наиболее работоспособной оказывается генетическая система первооткрывателя (1), все остальные не столь эффективны.
Сильное мутагенное воздействие (вирусы + гены транскрипционных факторов + онкогены), да еще отбор на среде для культивирования ЭСК приводит к тому, что появляются единичные клетки, которые преодолели процесс селекции. В исходных работах эффективность образования iPS составляла сотые доли процента. Скорее всего, именно по этой причине существует большой разброс, наблюдаемый для «репрограммированных» клонов. На самом деле эти рассуждения говорят в пользу того, что и во взрослом организме могут потенциально существовать клетки, которые ближе по своим свойствам к плюрипотентным, чем остальное большинство.
К недостаткам всех известных методов репрограммирования соматических клеток можно отнести следующее.
1. Введение в клетку гена в составе разнообразных векторов, особенно вирусных, приводит к трансгенному клеточному материалу, в котором гены и векторы, их содержащие, интегрируют в разные места геномы, тем самым модифицируя организм, что может привести к нежелательным последствия в будущем, в том числе и онкогенезу. Кроме онкогенеза за счет встраивания вирусной ДНК в геном существует значительная опасность реактивации трансгенов (генов, введенных в составе вирусов) в процессе жизнедеятельности клеток, их последующей дифференцировке. Это приведет к тому, что в отдельно взятой клетке опять может произойти репрограммирование, она может приобрести свойства плюрипотентности. При этом, находясь в окружении уже дифференцированных клеток организма, эти случайно репрограммированные клетки могут дать начало другим тканям, что приведет к появлению хористий и гамартий. Такое неправильное развитие ткани в конечном итоге опять может привести к возникновению опухоли. Генетическая модификация с помощью ДНК имеет также ряд недостатков в плане эффективности производства кодируемых ДНК белков. Во-первых, с молекулы ДНК должна быть синтезирована РНК. Синтез РНК происходит в ядре, поэтому из цитоплазмы молекула ДНК должна проникнуть в ядро. Большинство систем доставки ДНК, исключая вирусную, доставляют ДНК только в цитоплазму, откуда она должна случайным образом проникнуть в ядро. При этом количество молекул, которое проникло в ядро, неизвестно, не является стандартным, а разброс может привести к негативным последствиям в случае практического использования. Во-вторых, для синтеза РНК в клетке необходимо наличие промотора (последовательности, обеспечивающей посадку транскрипционных факторов клетки). Разные типы клеток имеют различный набор транскрипционных факторов, поэтому трудно подобрать промоторную последовательность, которая будет обеспечивать стандартную экспрессию с введенной конструкции в клеточном ядре. Например, промотор EF1 альфа не работает в фибробластах. Наиболее универсальными промоторными последовательностями, которые работают в большинстве типов клеток, но с разной эффективностью, являются вирусные промоторы, такие как CMV, RSV, SV40 и другие. При этом не обеспечивается стандартность работы в различных типах клеток и вводится ДНК фрагмент вирусного генома, что представляет собой определенную опасность в связи с возможным онкогенезом в случае интеграции промоторной последовательности в геном клетки. В-третьих, синтезированная в клеточном ядре РНК должна мигрировать в цитоплазму. При эндогенном синтезе РНК с геномной последовательностью, молекулы несут интроны и другие последовательности, обеспечивающие стабильность РНК и ее направленный транспорт в цитоплазму. В случае использования трансгена в большинстве случаев РНК лишены этих последовательностей, поэтому эффективность транспорта РНК из ядра в цитоплазму низкая, а незащищенные последовательности РНК подвергаются гидролизу РНКазами. Это приводит к существенному снижению эффективности и отсутствию стандартизации синтеза белкового продукта в цитоплазме.
2. В процессе индивидуального развития организма (онтогенеза) клетки проходят путь от состояния тотипотентности (зигота), через плюрипотентность (внутренняя клеточная масса, эмбриональные стволовые клетки in vitro), к терминальной дифференцировке (специализированные ткани). Специализированные ткани оргназма и региональные стволовые клетки, поддерживающие регенерацию этих тканей, не существуют бесконечно долго. Происходит процесс старения, который сопровождается смертью организма. Процесс старения организма связывают с целом рядом внутриклеточных событий, к основным относится накопление клетками биологического «мусора» (неразлагаемые белки, липиды и др) и генетического «мусора» (мутации, накопившиеся в клетках в процессе онтогенеза). Таким образом, клетки взрослого организма в процессе онтогенеза накапливают те или иные необратимые изменения, которые выражаются в процессе старения и приводят к смерти организма. Изменение генетической программы клетки взрослого организма возможно с использованием ряда методов, как репрограммирование с помощью переноса ядра (Вильмут), генетическое репрограммирование с использованием генов (Weintraub, Yamanaka). Однако в этих случаях происходит репрограммирование генома и клетки, уже накопивших необратимые изменения, поэтому последующее использование такого материала для целей терапии может привести к негативным последствиям в виде преждевременного старения, смерти или в лучшем случае развитию опухоли. Более того, клеточный материал млекопитающих и человека для репрограммирования не всегда бывает доступен, а процедура его взятия является инвазивной.
В результате проведенных исследований нами установлено, что плюрипотентные клетки могут быть эффективно получены из клеток пуповинно-плацентарного комплекса путем введения в них РНК, содержащих в своем составе последовательности, обеспечивающие переход клеток к плюрипотентному состоянию.
Предлагаемый нами метод репрограммирования соматических клеток основан на введении в клетку РНК, полученной в системе транскрипции in vitro, т.е. вне клетки. При этом используются безопасные прокариотические промоторные последовательности, например, SP6, Т7, Т3 и другие и соответствующие им полимеразы SP6, Т7, Т3. При этом синтезированная in vitro РНК перед введением в клетку очищается современными методами ионнобменной хроматографии от чужеродных белков и чужеродной ДНК. Получаемые in vitro количества РНК могут измеряться милиграммами и даже граммами, а система in vitro транскрипции может быть стандартизована. В этом случае можно с точность до молекулы рассчитать какое количество РНК, кодирующей тот или иной белковый продукт, может проникнуть в клетку млекопитающих in vitro или in vivo. Для доставки РНК в клетку могут быть использованы такие методы, как химическая трансфекция с использованием химических веществ, например дендример, биологическая трансфекция с использованием липидов и белков, например липосомы, электромеханическая трансфекция, например электорпорация. Также РНК можно вводить в составе бактерий, катионных, анионных полимеров, липосомальных комплексов, под действием электрического поля, инертных частиц или других способов, пригодных для введения нуклеиновых кислот в клетку. При этом РНК попадает в цитоплазму клетки, минуя стадию выхода из ядра и разрушения. Таким образом, с нее в цитоплазме сразу может начаться синтез соответствующих белков. Количество РНК, с которой будет происходить синтез, может быть при этом четко определено. Среднее время полураспада молекулы РНК в клетке составляет несколько часов (3-6), а вводимая молекула лишена последовательностей, с которых может произойти синтез ДНК. Таким образом, при введении РНК не происходит генетической модификации клетки, не изменяется клеточный геном и в клетку не попадают генетические элементы вирусов, которые могут привести к онкогенезу. Более того, не требуется дополнительного времени для попадания молекул в ядро, на синтез РНК и ее транспорт обратно в цитоплазму, а количество вводимой РНК может быть стандартизовано.
Также основным отличием от известных способов получения плюрипотентных клеток является то, что использованы клетки пуповинно-плацентарного комплекса млекопитающего, в которые вводят известные последовательности, обеспечивающие переход клеток к плюрипотентному состоянию, в составе РНК.
Во время рождения млекопитающего (человека) происходит отделение пуповинно-плацентарного комплекса, содержащего ряд клеточных популяций, например фибробласты, кератиноциты, эндотелиоциты, гемопоэтические и мезенхимальные клетки, которые могут быть изолированы, размножены и сохранены. Эти клетки являются самыми «молодыми» клетками организма, они еще не были экспонированы в окружающей среде, а получение их на этой стадии не инвазивно. Таким образом, все типы клеток, содержащиеся в пуповинно-плацентарном комплексе, являются наиболее доступными и безопасными для индивидуального репрограммирования, поскольку имеют минимально возможный набор дефектов, приобретаемых в процессе онтогенеза. Весь набор клеток, выделяемый из пуповинно-плацентарного комплекса, может быть использован для введения в них рибонуклеиновых кислот определенного состава для их перехода в плюрипотентное состояние.
В качестве последовательностей, обеспечивающих переход клеток к плюрипотентному состоянию, могут быть использованы (Reprogramming Factors), соответствующие белкам: Oct4 (POU5Fl),Sox2, HNF3b, PDX1, HNF6, ngn3, PAX4. NKX 2.2,FoxB3,HNF4a, Nkx2.5, Nkx2.2, Nkx6.1 и другие семейства Nkx, Pax4, Klf4, Ngn3, Pdxl, Mafa или другие, приводящие к желаемым изменениям свойств культуры. Они могут быть введены в клетки в составе РНК совместно или поодиночке.
Таким образом, изобретение может быть охарактеризовано следующей совокупностью существенных признаков «Способ получения плюрипотентных клеток с помощью введения в клетки млекопитающего нуклеиновых кислот, имеющих в своем составе по меньшей мере одну последовательность, обеспечивающую переход клеток к плюрипотентному состоянию, отличающийся тем, что используют соматические клетки пуповинно-плацентарного комплекса, а в качестве нуклеиновых кислот вводят очищенную РНК, синтезированную in vitro, используя невирусные методы трансфекции».
Полученные таким образом плюрипотентные клетки для дальнейшего их использования могут быть селектированы через 6-10 дней с использованием сред, поддерживающих рост эмбриональных стволовых клеток млекопитающих и человека (KnockOut ДМЕМ, заменитель сыворотки, mTeSR или другие), в отсутствии или при наличии фидерного слоя клеток. Для получения желаемого фенотипа клеток могут быть использованы среды, поддерживающие рост мышечных клеток, например кардиомиоцитов, секреторных клеток, например инсулин-продуцирующих, фильтрующих клеток, например гепатоцитов, нейрональных, глиальных и других. При культивировании плюрипотентных клеток на этих средах через 10-20 дней формируются клетки желаемого фенотипа. Полученные клеточные типы могут быть размножены in vitro для увеличения количества. Определенные количества клеток могут быть использованы для дальнейшего применения с целью терапии заболеваний и скрининга субстанций.
Возможность осуществления изобретения подтверждается следующими примерами.
Пример 1.
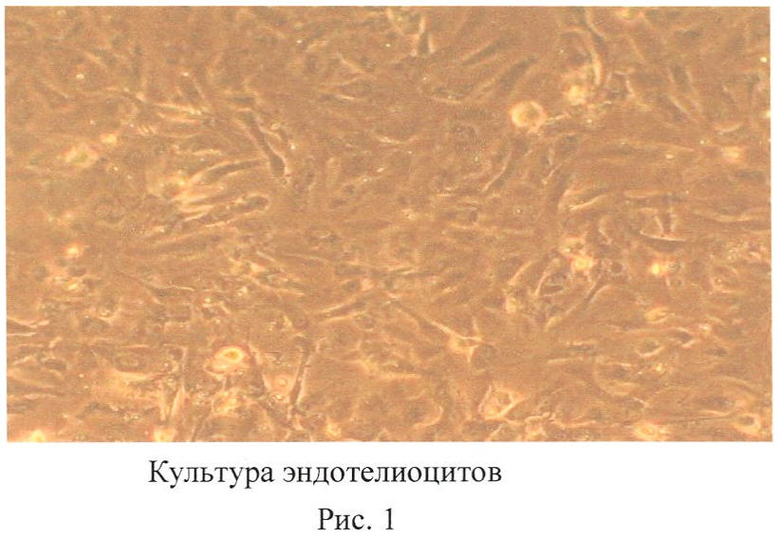
Эндотелиоциты, выделенные из пупочного канатика человека, культивировали в определенном составе среды, как Алфа MEM, 20% FBS, non-essensial aminiacids, 10-6 M гидрокортизона, IGF 5 нг/мл, bFGF 5 нг/мл, VEGF 10 нг/мл, EGF 5 нг/мл (фиг.1). На 2-3 пассаже культивирования проводили трансфекцию культуры эндотелиоцитов, синтезированной in vitro РНК. Для доставки использован реагент Turbofect (Fermentas) в соответствии с рекомендациями производителя. Были использованы РНК, содержащие последовательности, соответствующие белкам Oct4 (POU5F1), Sox2 и Klf4. Трансфекции проводились ежедневно на протяжении 3 дней. Через 6 дней после последней трансфекции клетки переносили на другие чашки, которые покрыты 0,1% желатином (Merck) и на которых находятся инактивированные эмбриональные фибробласты мыши в качестве фидера. При этом использовали культуральную среду следующего состава: 80% KnockOut DMEM, 20% FBS (ES qualified), 1 мМ глутамина, 1% заменимых аминокислот, 50 units/мл пенициллина, 50 µg/мл стрептомицина (все от Invitrogen), 0,1 мМ β-меркаптоэтанола (Sigma).
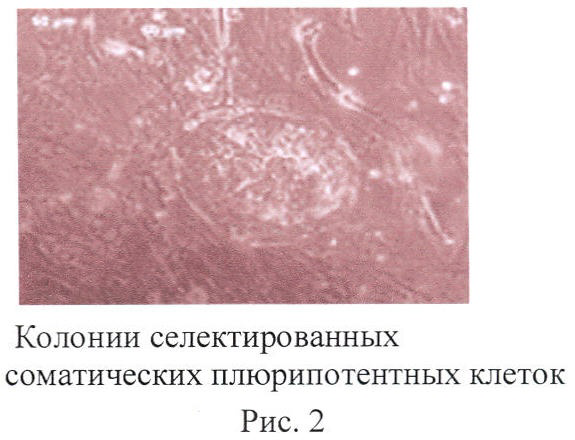
Через 17 дней сформировались колонии, идентичные тем, которые формируются эмбриональными стволовыми клетками человека (фиг.2).
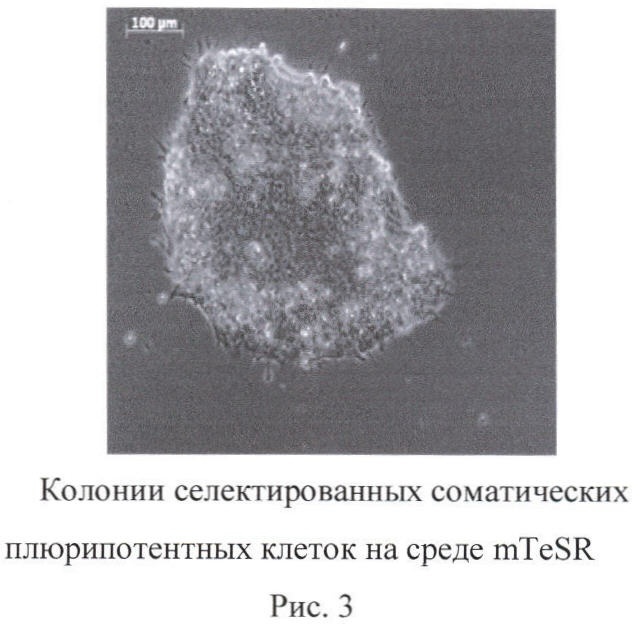
Формирование колоний происходит и при применении другой среды, поддерживающей рост эмбриональных стволовых клеток, например, mTeSR (фиг.3).
Эти клетки можно обозначить как ССПК - селектированные соматические плюрипотентные клетки.
Эффективность получения ССПК составляет более 0,001%. Полученные клеточные популяции могут быть в соответствующих условия культивированы и размножены. При этом они сохраняют нормальный кариотип, соответствующий исходным клеткам (фиг.4), но их фенотип и свойства меняются. Использование последовательностей Oct4 (POU5F1), Sox2 и Klf4 приводит к появлению маркеров SSEA4, TRA1-60, экспрессии гена Nanog, FoxD3 и других, что свойственно эмбриональным стволовым клеткам человека.
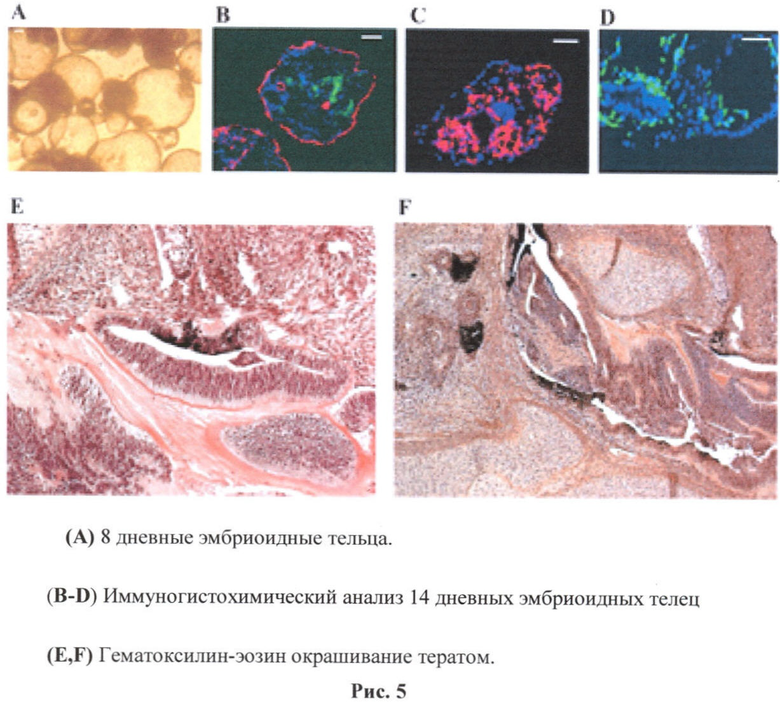
Получившиеся клетки по своему генотипу полностью соответствуют исходным, не исключая и комплекс иммуногистосовместимости 1 и 2 классов. Полученные плюрипотентные клетки могут формировать ткани всех трех зародышевых листков, что подтверждается формированием эмбриоидных телец (фиг.5А).
Эмбриоидные тела содержат клетки трех зародышевых листков, мезодерму (фиг.5В окрашивание антителами к CD 105 (зеленый) и МОС-31 (красный)), энтодерму (фиг.5В окрашивание антителами к альфа-фетапротенину), эктодерму (фиг.5С окрашивание антителами к десмину) и тератомы (фиг.5E, F гематоксилин-эозин окрашивание тератом).
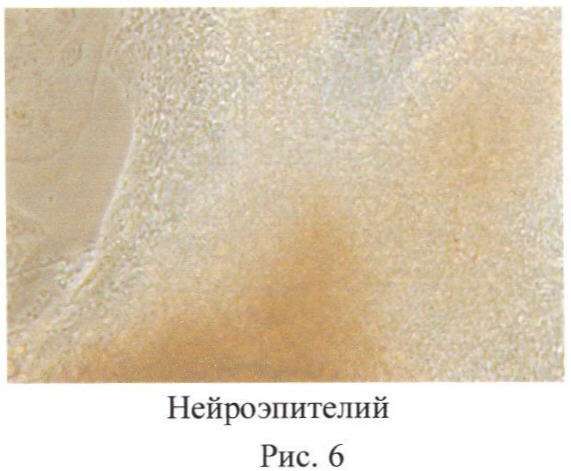
Таким образом, полученные из эндотелия клетки являются плюрипотентными и из них могут быть получены все типы тканей млекопитающих (человека). Например, на фиг.6 показаны нейроглиальные клетки, полученные из ССПК. Также могут быть получены клетки другой специализации, включая эндотелий, кардиомиоциты, гепатоциты, бета-клетки, кератиноциты, нейросенсорные, пигментированные и другие типы клеток, составляющие организм млекопитающих. При этом они являются генетически идентичными организму, от которого были получены исходно эндотелиоциты.
Пример 2.
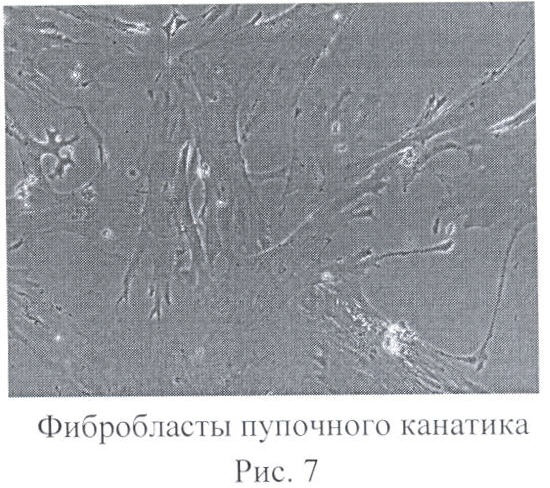
Получение клеток с индуцированной плюрипотентностью из фибробластов пупочного канатика.
Фибробласты пупочного канатика культивировались в среде DMEM/F12 15% сыворотки и 10 нг/мл основной фактор роста фибробластов (Peprotech) (фиг.7).
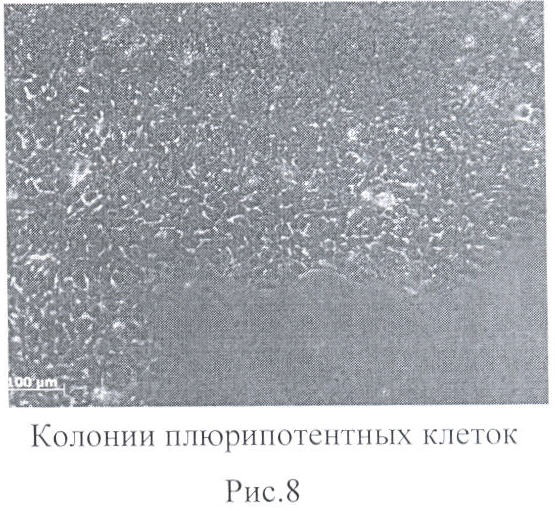
Для трансфекции нуклеиновыми кислотами использовался реагент Effecten (Qiagen) или XFect (Clontech), как описано производителем. Через 10 дней культивирования клетки пересевались на среду, как описано в примере 1. Через 28 дней культивирования в среде для ЭСК формировались колонии, аналогичные эмбриональным стволовым клеткам человека (фиг.8).
Проведенные иммуногистохимический и молекулярный анализы показали, что полученные колонии имеют стабильный кариотип и экспрессируют маркеры, характерные для эмбриональных стволовых клеток человека. Эффективность возникновения колоний не зависит от производителя трансфекционных реагентов.
Пример 3. Получение индуцированных плюрипотентных клеток из клеток крови пуповинно-плацентарного комплекса.
Пуповинная кровь, освобожденная от эритроцитарной массы центрифугированием, высевалась в питательную среду RPMI, 10% сыворотки. Клетки не разделялись на адгезионные или суспензионные (фиг.9). Трансфекция нуклеиновыми кислотами производилась с помощью реагентов Turbofect (Fermentas) или Effecten (Qiagen).
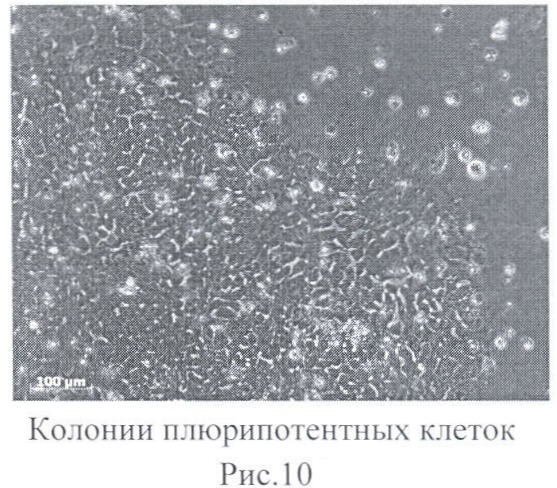
Через 7-10 дней после трансфекции клетки пересевались на среду для культивирования ЭСК, как описано в примере 1. Через 35 дней после пересева на Матригеле начинали формироваться колонии клеток, аналогичные по своей морфологии колониям ЭСК человека (фиг.10). Через 40 дней индивидуальные колонии можно было пересевать для последующего наращивания.
Проведенный молекулярно-генетический и иммуногистохимический анализ показал, что получившиеся культуры клеток имеют маркеры эмбриональных стволовых клеток человека, нормальный кариотип, а также формируют в культуре эмбриоидные тельца, что свидетельствует о функциональной плюрипотентности полученных клеток. При применении различных трансфекционных реагентов не отмечено разницы в эффективности формирования колоний, что свидетельствует о том, что для индукции плюрипотентного состояния может быть использован любой из липосомальных агентов трансфекции нуклеиновых кислот.
Данные примеры показывают только частный случай способа получения плюрипотентных клеток.
Предложенный способ позволяет эффективно получать плюрипотентные клетки из «молодых» клеток пуповинно-плацентарного комплекса, при этом снижается вероятность возникновения онкогенеза и других негативных последствий репрограммирования.
Источники информации

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ИЗ ФИБРОБЛАСТОВ ПАЦИЕНТОВ С БОЛЕЗНЬЮ ХАНГИНГТОНА | 2011 |
|
RU2458983C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ПАЦИЕНТОВ С СИНДРОМОМ ДАУНА | 2012 |
|
RU2492233C1 |
| СПОСОБ ПОВЫШЕНИЯ ЭКСПРЕССИИ РНК В КЛЕТКЕ | 2018 |
|
RU2784654C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКОКАЧЕСТВЕННЫХ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК (ИПСК) | 2016 |
|
RU2721266C1 |
| ПОЛУЧЕНИЕ ПЛЮРИПОТЕНТНЫХ КЛЕТОК DE NOVO | 2013 |
|
RU2696071C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА (ВАРИАНТЫ) | 2008 |
|
RU2359030C1 |
| СПОСОБ ОЦЕНКИ МОРФОФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ДИФЕРЕНЦИРОВАННЫХ В ДОФАМИНЕРГИЧЕСКИЕ НЕЙРОНЫ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК БОЛЬНЫХ ПАРКИНСОНИЗМОМ | 2012 |
|
RU2501853C1 |
| СПОСОБЫ И ПРОДУКТЫ ДЛЯ ЭКСПРЕССИИ БЕЛКОВ В КЛЕТКАХ | 2013 |
|
RU2711249C2 |
| СТИМУЛЯЦИЯ ПУТИ Wnt ПРИ ПЕРЕПРОГРАММИРОВАНИИ СОМАТИЧЕСКИХ КЛЕТОК | 2008 |
|
RU2492232C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pOK-DsRed2, КОДИРУЮЩАЯ БЕЛКИ ОСТ4 И KLF4 ЧЕЛОВЕКА И ФЛУОРЕСЦЕНТНЫЙ БЕЛОК DsRed2, ПРЕДНАЗНАЧЕНАЯ ДЛЯ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2011 |
|
RU2495125C2 |


Изобретение относится к биотехнологии, а именно к получению плюрипотентных клеток, и может быть использовано для получения всех типов тканей млекопитающих. В клетки пуповинно-плацентарного комплекса вводят очищенную РНК, синтезированную in vitro, используя невирусные методы трансфекции. В составе РНК имеется по меньшей мере одна последовательность, обеспечивающая переход клеток к плюрипотентному состоянию. Способ позволяет увеличить эффективность получения селектированных соматических плюрипотентных клеток до значения, превышающего 0,001%, при этом снижается вероятность возникновения онкогенеза и других негативных последствий репрограммирования клетки, а также исключается инвазивность на стадии получения клеточного материала. 10 ил.
Способ получения плюрипотентных клеток с помощью введения в клетки млекопитающего нуклеиновых кислот, имеющих в своем составе по меньшей мере одну последовательность, обеспечивающую переход клеток к плюрипотентному состоянию, отличающийся тем, что используют соматические клетки пуповинно-плацентарного комплекса, а в качестве нуклеиновых кислот вводят РНК, синтезированную in vitro, используя невирусные методы трансфекции.
| US 2009047263 A1, 19.02.2009 | |||
| KNUT WOLTJEN et al | |||
| Piggyback transposition reprograms fibroblast to induced pluropotent stem cells | |||
| Приспособление для отопления печей нефтью | 1922 |
|
SU458A1 |
| KEISUKE KAJI et al | |||
| Virus-free induction of pluripotency and subsequent excision of reprogramming factors | |||
| Приспособление для отопления печей нефтью | 1922 |
|
SU458A1 |

