Область техники
[0001] Настоящее изобретение относится к агенту для улучшения качества индуцированных плюрипотентных стволовых клеток (далее, ИПСК), способу получения клеток ИПСК, клеткам ИПСК, полученным таким способом, и составу для получения клеток ИПСК.
Уровень техники
[0002] Индуцированные плюрипотентные стволовые клетки (ИПСК) могут быть получены из соматических клеток посредством введения Oct3/4, Sox2, Klf4 и c-Myc (непатентный документ 1, патентный документ 1). Это может быть достигнуто посредством перепрограммирования транскрипционной сети родительской соматической клетки и эпигенетических подписей. Клетки ИПСК привносят различные преимущества для фундаментальных исследований, инноваций в области разработки лекарственных средств и регенеративной медицины. Однако серьезной проблемой все еще остается то, что клеточная популяция полученных клеток ИПСК более гетерогенна в плане качества по сравнению с популяциями эмбриональных стволовых клеток (ЭСК). Например, ЭСК обладают малой дисперсией свойств среди клеток, и по существу любая клетка может быть дифференцирована в требуемую клетку, но при этом клетки ИПСК обладают значительной дисперсией свойств среди клеток, и часто эти клетки не могут быть дифференцированы в требуемые клетки. Для фундаментальных исследований и клинических целей важно, чтобы любая клетка ИПСК демонстрировала высокое качество без дисперсии.
[0003] Множество попыток было предпринято, чтобы решить проблему, состоящую в гетерогенности популяций клеток ИПСК в плане качества. Например, в патентном документе 2 раскрыто, что эффективность и стабильность получения клеток ИПСК может быть улучшена посредством заранее определенного количества введений заранее определенного количества генов Oct3/4, Klf4, c-Myc и Sox2 в соматические клетки. Кроме того, в патентном документе 3 раскрыто, что индуцированные плюрипотентные стволовые клетки (клетки ИПСК) превосходного качества могут быть эффективно получены за короткий период времени посредством введения в соматические клетки гена Prdm14 или продукта этого гена, гена Esrrb или продукта этого гена, и гена Sall4a или продукта этого гена в добавление к гену Oct3/4 или продукту этого гена, гену Sox2 или продукту этого гена, гену Klf4 или продукту этого гена, или гену c-Myc или продукту этого гена. Кроме того, в патентном документе 4 раскрыто, что индуцированные плюрипотентные стволовые клетки (ИПСК) превосходного качества могут быть эффективно получены за короткий период времени посредством введения в соматические клетки мутантного гена Jarid2 или продукта этого гена в добавление к гену Oct3/4 или продукту этого гена, гену Sox2 или продукту этого гена, гену Klf4 или продукту этого гена, или гену c-Myc или продукту этого гена. Однако все еще остается пространство для улучшения качества клеток ИПСК. Таким образом, потребовалась разработка способа для получения высококачественных клеток ИПСК с меньшей дисперсией качества.
[0004] Линкерный гистон семейства H1 соединяется с линкерной ДНК и создает высокоупорядоченные хроматиновые структуры для контроля экспрессии гена. Элементы семейства H1 линкерных гистонов включают в себя гистоны H1a, H1b, H1c, H1d, H1e, H1foo, H1x, H1.0, H1t, H1T2 и HILS1. Большая часть элементов семейства линкерных гистонов - это соматические линкерные гистоны, которые компактируют хроматин. Соответственно, такие структуры в целом подавляют общую активность транскрипции гена (непатентные документы 2 и 3).
Документы по известному уровню техники
Патентные документы
[0005]
Патентный документ 1: Патент Японии №4183742
Патентный документ 2: Японская не прошедшая экспертизу патентная заявка, публикация №2011-004674
Патентный документ 3: Японская не прошедшая экспертизу патентная заявка, публикация №2014-217344
Патентный документ 4: Японская не прошедшая экспертизу патентная заявка, публикация №2014-217345
Непатентные документы
[0006] Непатентный документ 1: К. Такахаши, С. Яманака. «Индуцирование плюрипотентных стволовых клеток из фибробластных культур эмбрионов и взрослых особей мышей посредством определенных факторов». «Cell», №126, 663-676 (2006 г.)
Непатентный документ 2: О.К. Штайнбах, А.П. Вольффе, Р.А. Рупп «Соматические линкерные гистоны приводят к утере мезодермальной компетенции у шпорцевых лягушек». «Nature», №389, 395-399 (1997 г.)
Непатентный документ 3: П.Б. Хеббар, Т.К. Арчер «Изменение стехиометрии гистона H1 и отсутствие позиционирования нуклеосом на трансфицированной ДНК». «The Journal of Biological Chemistry», №283, 4595-4601 (2008 г.).
Сущность изобретения
Проблема, которую должно решить изобретение
[0007] Цель настоящего изобретения состоит в том, чтобы предложить агент для улучшения качества клеток ИПСК, способ получения клеток ИПСК, клетки ИПСК, полученные таким способом, и состав для получения клеток ИПСК.
Средства решения указанных проблем
[0008] Авторы настоящего изобретения провели тщательные исследования с целью улучшения качества клеток ИПСК и в результате обнаружили, что в способе получения клеток ИПСК посредством введения вещества для перепрограммирования ядра в соматическую клетку, введения не только вещества для перепрограммирования ядра, но и «гена H1foo или продукта этого гена» в соматическую клетку позволяет получить высококачественные клетки ИПСК с меньшей дисперсией качества, тем самым завершая настоящее изобретение. Для специалистов в данной области техники стало неожиданностью то, что сочетание «гена H1foo или продукта этого гена» и вещества для перепрограммирования ядра позволило получить высококачественные клетки ИПСК с меньшей дисперсией качества.
[0009] Соответственно, в изобретении предложен:
(1) Агент для улучшения качества клеток ИПСК, содержащий ген H1foo или продукт этого гена; и
(2) Агент для улучшения качества клеток ИПСК по п. (1), в котором агент содержит экспрессионный вектор, содержащий ген H1foo.
[0010] Кроме того, в настоящем изобретении предложен:
(3) Способ получения клеток ИПСК, содержащий шаг введения (а) вещества для перепрограммирования ядра и шаг введения (b) гена H1foo или продукта этого гена в соматическую клетку;
(4) Способ получения клеток ИПСК по п. (3), в котором вещество для перепрограммирования ядра содержит по меньшей мере один ген, выбранный из группы, содержащей ген из семейства генов Oct, ген из семейства генов Sox, ген из семейства генов Klf, ген из семейства генов Myc, ген из семейства генов Lin, ген Nanog и продукты таких генов;
(5) Способ получения клеток ИПСК по п. (3) или (4), в котором вещество для перепрограммирования ядра содержит ген из семейства генов Oct или продукт этого гена, ген из семейства генов Sox или продукт этого гена и ген из семейства Klf или продукт этого гена;
(6) Способ получения клеток ИПСК по любому из пп. (3)-(5), в котором вещество для перепрограммирования ядра содержит ген Oct3/4 или продукт этого гена, ген Sox2 или продукт этого гена и ген Klf4 или продукт этого гена; и
(7) Способ получения клеток ИПСК в по любому из пп. (3)-(5), в котором вещество для перепрограммирования ядра состоит из гена Oct3/4 или продукта этого гена, гена Sox2 или продукта этого гена, гена Klf4 или продукта этого гена и гена L-Myc или продукта этого гена.
[0011] Далее, в изобретении предложен(а):
(8) Клетки ИПСК, полученные посредством способа получения клеток ИПСК по любому из п.п. (1)-(7); и
(9) Состав для получения клеток ИПСК, содержащий (а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена.
Преимущества изобретения
[0012] В настоящем изобретении предложен агент для улучшения качества клеток ИПСК, способ получения клеток ИПСК, клетки ИПСК, полученные таким способом, и состав для получения клеток ИПСК. В соответствии с настоящим изобретением могут быть получены высококачественные клетки ИПСК с меньшей дисперсией качества.
Краткое описание чертежей
[0013] [Фигура 1] Фигура 1 - представляет собой совокупность изображений мышиных хвостовых фибробластов (далее - «мышиные фибробласты») мышей C57BL/6J, экспрессирующих экзогенный (мышиный) H1foo. Изображения «BF», «DAPI (ДАФИ)», «H1foo» и «MERGE (НАЛОЖЕНИЕ)» на Фигуре 1а представляют собой фазово-контрастное изображение, изображение, полученное с флуоресцентным красителем ДАФИ, изображение H1foo, полученное с окрашиванием флуоресцентным красителем, и наложение 3 изображений друг на друга. Левое и правое изображение на Фигуре 1b представляют собой 2-мерное и 2,5 мерное (изометрическое) изображения H1foo с окрашиванием флуоресцентным красителем соответственно. Левое и правое изображение на Фигуре 1с представляют собой изображения контрольных клеток (мышиных фибробластов) и мышиных фибробластов, экспрессирующих экзогенный H1foo соответственно, полученные посредством электронной микроскопии.
[Фигура 2] На Фигуре 2а показаны результаты введения 3 типов веществ для перепрограммирования ядра (мышиные гены Oct3/4, Sox2 и Klf4, [ген OSK]), генов OSK и гена линкерного гистона H1 (гены OSK+H1c) и генов OSK и гена H1foo (гены OSK+H1foo) в мышиные фибробласты и анализ эффективности колонизации ЭСК-подобных клеток (клеток ИПСК) с положительной активностью щелочной фосфатазы. На Фигуре 2b показаны результаты анализа экспрессии 3 типов плюрипотентных маркеров (Oct3/4, Nanog и SSEA1) в клетках ИПСК, полученных посредством введения генов OSK или генов OSK+H1foo (далее - «клетки OSK-ИПСК» и «клетки OSK+H1foo-ИПСК» соответственно). На Фигуре 2с показаны результаты анализа экспрессии эндогенного H1foo через 1-5 дней после введения генов OSK или генов OSK и гена c-Myc (гены OSKM). «МЭФ+H1foo» на Фигуре 2с показывает результаты анализа мышиных эмбриональных фибробластов (МЭФ), в которые введен ген H1foo.
[Фигура 3] На Фигуре 3 показаны результаты введения генов OSKM, генов OSKM и гена H1foo (гены OSKM+H1foo), генов OSK, или генов OSK+H1foo в фибробласты, экспрессирующие Nanog-GFP и анализ процентного количества Nanog-GFP-положительных ЭСК-подобных клеток (клеток ИПСК).
[Фигура 4] На Фигуре 4а показаны результаты сравнения глобальных профилей транскриптома гена клеток OSK+H1foo-ИПСК и ЭСК. На Фигуре 4b показаны результаты сравнения глобальных профилей транскриптома гена клеток OSK+H1foo-ИПСК и клеток OSK-ИПСК.
[Фигура 5] На Фигуре 5 показаны результаты анализа уровней метилирования ДНК в межгенной дифференциально-метилированной области (МГ ДМО) (Фигура 5а) и Gtl2-ДМО (Фигура 5b) в клетках OSK-ИПСК и клетках OSK+H1foo-ИПСК. «ЭСК» и «МЭФ» на фигуре указывают на результаты анализа ЭСК и МЭФ соответственно.
[Фигура 6] На Фигуре 6 показаны результаты анализа морфологии эмбриоидных телец (ЭТ), образованных из клеток OSK-ИПСК и OSK+H1foo-ИПСК.
[Фигура 7] На Фигуре 7 показаны результаты анализа процентного количества апоптических клеток в ЭТ, образованных из клеток OSK-ИПСК и OSK+H1foo-ИПСК.
[Фигура 8] На Фигуре 8 показаны результаты анализа экспрессии 2-х маркеров пролиферации клеток (Ki67 и PCNA) в ЭТ, образованных из клеток OSK-ИПСК и клеток OSK+H1foo-ИПСК.
[Фигура 9] На Фигуре 9 показаны результаты анализа химерной компетенции клеток OSK-ИПСК и клеток OSK+H1foo-ИПСК.
[Фигура 10] На Фигуре 10 показаны результаты анализа потенциала трансмиссии зародышевой линии химерных мышей, полученных из клеток OSK-iPS и клеток OSK+H1foo-ИПСК.
[Фигура 11] На Фигуре 11 показаны результаты количественного анализа с использованием полимеразной цепной реакции с обратной транскрипцией (RT-PCR) гена SRF (Фигура 11а) и гена ACTG2 (Фигура 11b) в клетках OSKL+H1foo-ИПСК (ИПСК получены посредством введения генов Oct3/4, Sox2, Klf4, L-Myc и H1foo) и клетках OSKL-ИПСК (ИПСК получены посредством введения генов Oct3/4, Sox2, Klf4 и L-Myc). Номера 1-4 на оси абсцисс графика указывают на номера клонов.
[Фигура 12] На Фигуре 12 показаны результаты количественного анализа с использованием полимеразной цепной реакции с обратной транскрипцией (RT-PCR) гена Oct3/4 в клетках OSKL+H1foo-ИПСК и клетках OSKL-ИПСК. Номера 1-4 на оси абсцисс графика указывают на номера клонов.
[Фигура 13] На Фигуре 13 показано количество жизнеспособных клеток через 5 дней культивирования клеток OSKL+H1foo-ИПСК и клеток OSKL-ИПСК с индукцией дифференциации. Номера 1-4 на оси абсцисс графика указывают на номера клонов.
[Фигура 14] На Фигуре 14 показаны результаты количественного анализа с использованием полимеразной цепной реакции с обратной транскрипцией (RT-PCR) гена Oct3/4 после культивирования клеток OSKL+H1foo-ИПСК и клеток OSKL-ИПСК в течение 5 дней с индукцией дифференциации. Номера 1-4 на оси абсцисс графика указывают на номера клонов.
Примеры осуществления изобретения
[0014] Способ получения клеток ИПСК
Способ получения клеток ИПСК в соответствии настоящим изобретением содержит по меньшей мере шаг введения (а) вещества для программирования ядра и (b) гена H1foo или продукта этого гена в соматическую клетку (далее - «шаг введения»), и дополнительно может содержать другой шаг, по мере необходимости.
[0015] <Шаг введения>
Шаг введения представляет собой, по меньшей мере, шаг введения (а) вещества для программирования ядра и шаг введения (b) гена H1foo или продукта этого гена в соматическую клетку. Посредством введения не только вещества для программирования клетки, но и гена H1foo или продукта этого гена в соматическую клетку, может быть получено более высокое количество высококачественных клеток ИПСК. Используемый в настоящем документе продукт гена обозначает матричную РНК (мРНК), транскрибированную из гена, и/или протеина, транслированного из мРНК. Ген H1foo обозначает полинуклеотид, кодирующий белок H1foo. Ген H1foo или продукт этого гена может быть использован в качестве агента для улучшения качества ИПСК. Кроме того, вектор, содержащий ген H1foo, описание которого приведено далее, может быть также использован в качестве агента для улучшения качества ИПСК.
[0016] (Ген H1foo)
Источник гена H1foo конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя млекопитающих, таких как человек, мышь, крыса, корова, овца, лошадь и обезьяна. Информация о последовательности гена H1foo может быть получена из общедоступной базы данных и может быть получена, например, с идентификатором ВС047943 (человек), AY158091 (человек) или ВС137916 (мышь) в базе данных «GenBank». Нуклеотидная последовательность человеческого гена H1foo (ВС047943) представлена в идентификаторе последовательности SEQ ID №1, и аминокислотная последовательность человеческого белка H1foo (белок, кодируемый геном H1foo ВС047943) представлена в идентификаторе последовательности SEQ ID №2, Нуклеотидная последовательность человеческого гена H1foo (AY158091) представлена в идентификаторе последовательности SEQ ID №59, и аминокислотная последовательность человеческого белка H1foo (белок, кодируемый геном H1foo AY158091) представлена в идентификаторе последовательности SEQ ID №60. Нуклеотидная последовательность мышиного гена H1foo (ВС137916) представлена в идентификаторе последовательности SEQ ID №3, и аминокислотная последовательность мышиного белка H1foo (белок, кодируемый геном H1foo ВС137916) представлена в идентификаторе последовательности SEQ ID №4.
[0017] Нуклеотидные последовательности генов H1foo и нуклеотидные последовательности их мРНК могут быть такими же, как и нуклеотидные последовательности генов H1foo дикого типа и нуклеотидными последовательностями их мРНК, или же могут содержать мутацию. Примеры такой нуклеотидной последовательности, содержащей мутацию, включают в себя «нуклеотидную последовательность, модифицированную из нуклеотидной последовательности гена H1foo дикого типа (например, нуклеотидная последовательность, представленная в идентификаторе последовательности SEQ ID №1 или SEQ ID №59, или SEQ ID №3), или нуклеотидную последовательность его мРНК посредством удаления, замены, вставки или добавления 1-30, в более предпочтительном варианте 1-20, в более предпочтительном варианте 1-15, в более предпочтительном варианте 1-10, в более предпочтительном варианте 1-5, в более предпочтительном варианте 1-3 нуклеотидов, и кодирующую белок с активностью H1foo», и «нуклеотидную последовательность, которая содержит, в части, транслируемой в белок, нуклеотидную последовательность с 70% или более, в предпочтительном варианте 80% или более, в более предпочтительном варианте 85% или более, в более предпочтительном варианте 90% или более, в более предпочтительном варианте 95% или более, в более предпочтительном варианте 98% или более идентичности последовательности с нуклеотидной последовательностью гена H1foo дикого типа (например, нуклеотидная последовательность, представленная в идентификаторе последовательности SEQ ID №1 или SEQ ID №59, или SEQ ID №3) или нуклеотидную последовательность его мРНК, и кодирующую белок с активностью H1foo».
[0018] Аминокислотная последовательность белков H1foo может быть такой же, как и аминокислотные последовательности белков H1foo дикого типа (например, аминокислотная последовательность, представленная в идентификаторе последовательности SEQ ID №2 или SEQ ID №60, или SEQ ID №4), или может содержать мутацию. Примеры такого белка, содержащего мутацию, включают в себя «белок, содержащий аминокислотную последовательность, модифицированную из аминокислотной последовательности белка H1foo дикого типа (например, аминокислотная последовательность, представленная в идентификаторе последовательности SEQ ID №2 или SEQ ID №60, ИЛИ SEQ ID №4) посредством удаления, замены, вставки или добавления 1-30, в предпочтительном варианте 1-20, в более предпочтительном варианте 1-15, в более предпочтительном варианте 1-10, в более предпочтительном варианте 1-5, в более предпочтительном варианте 1-3 аминокислот, и с активностью H1foo» и «белок, содержащий аминокислотную последовательность с 70% или более, в предпочтительном варианте 80% или более, в более предпочтительном варианте 85% или более, в более предпочтительном варианте 90% или более, в более предпочтительном варианте 95% или более, в более предпочтительном варианте 98% или более идентичной последовательности с аминокислотной последовательностью белка H1foo дикого типа (например, аминокислотная последовательность, представленная в идентификаторе последовательности SEQ ID №2 или SEQ ID №60, или SEQ ID №4), и с активностью H1foo». Используемый в настоящем документе «белок с активностью H1foo» обозначает белок, который позволяет получить большее количество высококачественных клеток ИПСК, когда его вводят в соматическую клетку с веществом для перепрограммирования ядра, по сравнению с количеством, когда в соматическую клетку вводят только вещество для перепрограммирования ядра.
[0019] (Вещество для перепрограммирования ядра)
Используемое в настоящем документе «вещество для перепрограммирования ядра» обозначает вещество (или группу веществ), которое позволяет осуществить индуцирование соматической клетки в клетку ИПСК посредством введения вещества в отдельности или введении сочетания вещества с другим веществом в соматическую клетку. Такое вещество для перепрограммирования ядра может быть веществом, таким как ген (в том числе в форме встраивания в экспрессионный вектор), или продукт такого гена, или низкомолекулярное соединение, при условии, что оно является веществом (или группой веществ), которое позволяет осуществить индуцирование клетки ИПСК из соматической клетки. Ген, являющийся веществом для перепрограммирования ядра, обозначает нуклеотид, кодирующий белок, являющийся веществом для кодирования ядра. Примеры вещества для программирования ядра, когда таким веществом является ген или продукт такого гена, включают в себя по меньшей мере одно из следующих веществ, выбранных из группы, содержащей ген из семейства генов Oct, ген из семейства генов Sox, ген из семейства генов Klf, ген из семейства генов Myc, ген из семейства генов Lin, и ген Nanog, а также продукты таких генов (WO 2007/69666, патент Японии №5696282; «Science», 2007 г., 318: 1917-1920); в частности, в предпочтительном варианте 2 или более, в более предпочтительном варианте 3 или более выбранных из этой группы, в предпочтитльном варианте из этой группы выбраны 2-4, и в более предпочтительном варианте 3-4 вещества. Конкретные примеры генов из этих семейств и их сочетаний приведены далее. Несмотря на то, что далее раскрыты только наименования генов, также включено и использование продуктов таких генов.
[0020] Примером может быть:
(a) 1 вещество для перепрограммирования ядра, состоящее из гена из семейства генов Oct;
(b) Сочетание 2 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена из семейства генов Sox;
(c) Сочетание 2 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена из семейства генов Klf;
(d) Сочетание 2 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена Nanog;
(e) Сочетание 3 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Sox и гена из семейства генов Klf;
(f) Сочетание 3 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Klf и гена из семейства генов Myc;
(е) Сочетание 4 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Sox, гена из семейства генов Klf и гена из семейства генов Myc;
(h) Сочетание 4 веществ для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Sox, гена из семейства генов Lin и гена Nanog;
или сочетание веществ для перепрограммирования ядра, представленных в п.п. (a)-(h) или их сочетания с дополнительным добавлением другого вещества для перепрограммирования ядра (ген или продукт этого гена).
Конкретные примеры включают в себя следующее:
(а') Сочетание веществ для перепрограммирования ядра, содержащее 1 вещество для перепрограммирования ядра, состоящее из гена из семейства генов Oct;
(b') Сочетание веществ для перепрограммирования ядра, содержащее 2 вещества для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена из семейства генов Sox;
(с') Сочетание веществ для перепрограммирования ядра, содержащее 2 вещества для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена из семейства генов Klf;
(d') Сочетание веществ для перепрограммирования ядра, содержащее 2 вещества для перепрограммирования ядра, состоящее из гена из семейства генов Oct и гена Nanog;
(e') Сочетание веществ для перепрограммирования ядра, содержащее 3 вещества для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Sox и гена из семейства генов Klf;
(f') Сочетание веществ для перепрограммирования ядра, содержащее 3 вещества для перепрограммирования ядра, состоящее из гена из семейства генов Oct, гена из семейства генов Klf и гена из семейства генов Myc;
(g') Сочетание веществ для перепрограммирования ядра, содержащее 4 вещества для перепрограммирования ядра, состоящее из гена из семейства Oct, гена из семейства генов Sox, гена из семейства генов Klf и гена из семейства генов Myc;
(h') Сочетание веществ для перепрограммирования ядра, содержащее 4 вещества для перепрограммирования ядра, состоящее из гена из семейства Oct, гена из семейства Sox, гена из семейства Lin и гена Nanog;
[0021] Более конкретные примеры включают в себя, помимо прочего, следующие сочетания.
(1) Ген Oct3/4, ген Klf4, ген c-Myc;
(2) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc (причем ген Sox2 может быть заменен на ген Sox1, ген Sox3, ген Sox15, ген Sox17 или ген Sox18; ген Klf4 может быть заменен на ген Klf1, ген Klf2 или ген Klf5; и ген c-Myc может быть заменен на ген Т58А (активная форма мутанта), ген N-Myc или ген L-Myc);
(3) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген Fbx15, ген Nanog, ген Eras, ген ЕСАТ15-2, ген TclI, β-катенин (активная форма мутанта S33Y);
(4) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, Большой Т-антиген вируса SV40 (далее - SV40LT);
(5) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, ген HPV16 Е6;
(6) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, ген HPV16 Е7;
(7) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, ген HPV6 Е6, ген HPV16 E7;
(8) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, ген Bmil,
(Для получения информации о сочетаниях (1)-(8) см. заявку WO 2007/069666 (однако для получения информации о замене гена Sox2 на ген Sox18 и замене гена Klf4 на ген Klf1 или ген Klf5 в сочетании (2), см. «Nature Biotechnology», №26, 101-106 (2008 г.)). Для получения информации о сочетании «ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc», также см. «Cell», №126, 663-676 (2006 г.), «Cell», №131, 861-872 (2007 г.) и т.п. Для получения информации о сочетании «ген Oct3/4, ген Sox2, ген Klf2 (или Klf5), ген c-Myc» также см. «Nat. Cell Biol.», №11, 197-203 (2009 г.). Для получения информации о сочетании «ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген hTERT, ген SV40LT» также см. «Nature», №451, 141-146 (2008 г.).)
(9) Ген Oct3/4, ген Sox2, ген Klf4 (см. «Nature Biotechnology», №26, 101-106 (2008 г.));
(10) Ген Oct3/4, ген Sox2, ген Nanog, ген Lin28 (см. «Science», №318, 1917-1920 (2007 г.));
(11) Ген Oct3/4, ген Sox2, ген Nanog, ген Lin28, ген hTERT, ген SV40LT (см. «Stem Cells», №26, 1998-2005 (2008 г.));
(12) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген Nanog, ген Lin28 (см. «Cell Research» (2008 г.) 600-603);
(13) Ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc, ген SV40LT (см. «Stem Cells», №26, 1998-2005 (2008 г.));
(14) Ген Oct3/4, ген Klf4 (см. «Nature», №454, 646-650 (2008 г.), «Cell Stem Cell», №2, 525-528 (2008 г.));
(15) Ген Oct3/4, ген c-Myc (см. «Nature», №454, 646-650 (2008 г.));
(16) Ген Oct3/4, ген Sox2 (см. «Nature», №451, 141-146 (2008 г.), WO 2008/118820);
(17) Ген Oct3/4, ген Sox2, ген Nanog (см. WO 2008/118820);
(18) Ген Oct3/4, ген Sox2, ген Lin28 (см. WO 2008/118820);
(19) Ген Oct3/4, ген Sox2, ген c-Myc, ген Esrrb (причем ген Essrrb может быть заменен на ген Esrrg. См. «Nat. Cell Biol.», №11, 197-203 (2009 г.));
(20) Ген Oct3/4, ген Sox2, ген Esrrb (см. Nat. Cell Biol.», №11, 197-203 (2009 г.));
(21) Ген Oct3/4, ген Klf4, ген L-Myc;
(22) Ген Oct3/4, ген Nanog;
(23) Ген Oct3/4;
(24) Ген Oct3/4, ген Klf4, ген c-Myc, ген Sox2, ген Nanog, ген Lin28, ген SV40LT (см. «Science», №324, 797-801 (2009 г.));
[0022] В сочетаниях (1)-(24) другие гены-элементы семейства генов Oct (например, Oct1A, Oct6) могут быть использованы вместо гена Oct3/4. Кроме того, другие гены-элементы семейства генов Sox (например, ген Sox7) могут быть использованы вместо гена Sox2 (или гена Sox1, гена Sox3, гена Sox15, гена Sox17, гена Sox18). Далее, другие гены-элементы семейства генов Lin (например, ген Lin28b) могут быть использованы вместо гена Lin28.
Кроме того, сочетание, не совпадающее с любым из сочетаний (1)-(24), но содержащее все компоненты какого-либо из сочетаний (1)-(24) и дополнительно содержащее любое другое вещество (в предпочтительном варианте вещество для перепрограммирования ядра) может быть включено в категорию «вещество для перепрограммирования ядра» в настоящем изобретении. Кроме того, при условиях, в которых соматические клетки, подлежащие перепрограммированию эндогенно, экспрессируют часть компонентов в любом из сочетаний (1)-(24) на уровне, достаточном для перепрограммирования ядра, сочетания оставшихся компонентов, за исключением экспрессированных компонентов, может также быть включено в категорию «вещество для перепрограммирования ядра» в настоящем изобретении.
[0023] Далее, в добавление к веществам для перепрограммирования ядра, одно или более веществ для перепрограммирования ядра, выбранных из группы, состоящей из гена Fbx15, гена ERas, гена ЕСАТ15-2, гена Tcl1 и гена β-катенина могут быть объединены, и/или одно или более вещество для перепрограммирования ядра, выбранных из группы, состоящей из гена ЕСАТ1, гена Esg1, гена Dnmt3L, гена ЕСАТ8, гена Gdf3, гена Mybl2, гена ЕСАТ15-1, гена Fthl17, гена Sall4, гена Rex1, гена UTF1, гена Stella, гена Stat3 и гена Grb2 могут быть объединены. Эти сочетания конкретно раскрыты в заявке WO 2007/69666.
[0024] Примеры предпочтительных вариантов веществ для перепрограммирования ядра включают в себя ген Oct3/4, ген Sox2, ген Klf4, ген c-Myc (или ген L-Myc), ген Lin28 и ген Nanog, и по меньшей мере 1, в предпочтительном варианте 2 или более, в более предпочтительном варианте 3 или более выбранных из группы, состоящей из продуктов этих генов. Примеры особенно предпочтительных сочетаний веществ для перепрограммирования ядра включают в себя: (1) Ген Oct3/4 или продукт этого гена, ген Sox2 или продукт этого гена и ген Klf4 или продукт этого гена; (2) Ген Oct3/4 или продукт этого гена, ген Sox2 или продукт этого гена, ген Klf4 или продукт этого гена, ген c-Myc или продукт этого гена; и (3) Ген Oct3/4 или продукт этого гена, ген Sox2 или продукт этого гена, ген Klf4 или продукт этого гена, ген L-Myc или продукт этого гена; в частности, предпочтительные примеры включают в себя сочетание гена Oct3/4 или продукта этого гена, гена Sox2 или продукта этого гена и гена Klf4 или продукта этого гена; и сочетания гена Oct3/4 или продукт этого гена, гена Sox2 или продукта этого гена, гена Klf4 или продукта этого гена, гена L-Myc или продукта этого гена; и, в частности, предпочтительные примеры включают в себя сочетание гена Oct3/4, гена Sox2 и гена Klf4; и сочетание гена Oct3/4, гена Sox2, гена Klf4 и гена L-Myc.
[0025] В настоящем изобретении ген c-Myc или продукт этого гена может быть использован в качестве вещества для перепрограммирования ядра в, но предпочтительно не использовать ген c-Myc или продукт этого гена. Это обусловлено тем, что согласно полученным данным ген c-Myc снижает процентное количество Nanog-ЗБФ-положительных колоний и увеличивает канцерогенность клеток (М. Накагава и др., «Поколение индуцированных плюрипотентных стволовых клеток без гена Myc из мышиных и человеческих фибробластов». «Nature Biotechnology», №26, 101-106 (2008 г.)).
[0026] Когда вещество для перепрограммирования ядра представляет собой ген или продукт этого гена, источник такого гена не ограничен, но он может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя человека, мышь, крысу, корову, овцу, лошадь и обезьяну.
[0027] Информация о последовательности мышиной и человеческой кДНК веществ для перепрограммирования ядра может быть получена с использованием идентификаторов базы данных «GenBank», см. заявку WO 2007/069666. Ген Nanog описан под обозначением «ЕСАТ4» в проспекте. Кроме того, далее приведено описание информации о последовательности мышиных и человеческих кДНК особенно предпочтительных 3-х генов (ген Oct3/4, ген Sox2, ген Klf4) среди веществ для перепрограммирования ядра.


Последовательность кДНК человеческого гена Oct3/4 представлена в идентификаторе последовательности SEQ ID №47, аминокислотная последовательность человеческого белка Oct3/4 представлена в идентификаторе последовательности SEQ ID №48, последовательность кДНК человеческого гена Sox2 представлена в идентификаторе последовательности SEQ ID №49, аминокислотная последовательность человеческого белка Sox2 представлена в идентификаторе последовательности SEQ ID №50, последовательность кДНК человеческого гена Klf4 представлена в идентификаторе последовательности SEQ ID №51, аминокислотная последовательность человеческого белка Klf4 представлена в идентификаторе последовательности SEQ ID №52, последовательность кДНК человеческого гена L-Myc представлена в идентификаторе последовательности SEQ ID №53, аминокислотная последовательность человеческого белка L-Myc представлена в идентификаторе последовательности SEQ ID №54.
[0028] Среди веществ для перепрограммирования ядра информация о последовательности мышиных и человеческих кДНК генов, для которых идентификаторы в базе данных «GenBank» не указаны в заявке WO 2007/069666, приведена далее.

[0029] Специалисты в данной области техники могут с легкостью изолировать кДНК этих веществ для перепрограммирования ядра на основе информации о последовательности мышиных и человеческих кДНК веществ для перепрограммирования ядра.
[0030] Если вещества для перепрограммирования ядра представляют собой гены или мРНК таких генов, их нуклеотидные последовательности, в частности, не ограничены, при условии, что технический результат настоящего изобретения не нарушен, и они могут быть лишь частично транслированы в белок в нуклеотидных последовательностях генов или содержать другие части. Кроме того, нуклеотидные последовательности генов и нуклеотидные последовательности их мРНК могут быть такими же, как и нуклеотидные последовательности генов дикого типа или нуклеотидные последовательности их мРНК, или же могут содержать мутацию. Примеры такой нуклеотидной последовательности, содержащей мутацию, включают в себя «нуклеотидную последовательность, модифицированную из нуклеотидной последовательности гена дикого типа или нуклеотидной последовательности его мРНК посредством удаления, замены, вставки или добавления 1-30, в предпочтительном варианте 1-20, в более предпочтительном варианте 1-15, в более предпочтительном варианте 1-10, в более предпочтительном варианте 1-5, в более предпочтительном варианте 1-3 нуклеотидов и кодирования белка с эффектом перепрограммирования ядра продукта такого гена» и «нуклеотидную последовательность, содержащую 70% или более, в более предпочтительном варианте 80% или более, в более предпочтительном варианте 85% или более, в более предпочтительном варианте 90% или более, в более предпочтительном варианте 95% или более, в более предпочтительном варианте 98% или более идентичной последовательности с нуклеотидной последовательностью гена дикого типа или нуклеотидной последовательностью его мРНК в нуклеотидной последовательности в части, которая будет транслирована в белок, и кодирующую белок с эффектом перепрограммирования ядра продукта такого гена».
[0031] Если вещества для перепрограммирования ядра представляют собой белки, кодируемые генами (т.е. белки, транслированные из мРНК, транскрибированной из генов), их аминокислотные последовательности могут быть такими же, как и у белков, кодируемых генами дикого типа или могут содержать мутацию. Примеры такого белка, содержащего мутацию, включают в себя «белок, состоящий из аминокислотной последовательности, модифицированной из аминокислотной последовательности белка, кодируемого геном дикого типа, посредством удаления, замены, вставки или добавления 1-30, в более предпочтительном варианте 1-20, в более предпочтительном варианте 1-15, в более предпочтительном варианте 1-10, в более предпочтительном варианте 1-5, в более предпочтительном варианте 1-3 аминокислот, с эффектом перепрограммирования ядра» и «белок, состоящий из аминокислотной последовательности 70% или более, в более предпочтительном варианте 80% или более, в более предпочтительном варианте 85% или более, в более предпочтительном варианте 90% или более, в более предпочтительном варианте 95% или более, в более предпочтительном варианте 98% или более идентичной последовательности с аминокислотной последовательностью белка, кодируемого геном дикого типа, и с эффектом перепрограммирования ядра».
[0032] (Соматическая клетка)
Соматическая клетка конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели, и ее примеры включают в себя эмбриональные соматические клетки и зрелые соматические клетки. Конкретные примеры зрелых соматических клеток включают в себя стволовые клетки тканей (соматические стволовые клетки), такие как мезенхимальные стволовые клетки, гемопоэтические стволовые клетки, стромальные клетки жировой ткани, нейрональные стволовые клетки и сперматстенные стволовые клетки; прогениторные клетки тканей, дифференцированные клетки, такие как фибробласты, эпителиальные клетки, лимфоциты и клетки мышц.
[0033] Вид организма, из которого получена соматическая клетка, конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и примеры видов включают в себя любых млекопитающих, таких как человек, мышь, крыса, корова, овца, лошадь и обезьяна. Кроме того, виды организмов, из которых получены соматические клетки, и виды организмов, из которых ген введен в полученную соматическую клетку не обязательно представляют собой один и тот же вид, но в предпочтительном варианте - это один и тот же вид.
[0034] Индивидуум, из которого получают соматическую клетку, конкретно не ограничен и может быть выбран соответствующим образом в зависимости от цели, если полученные клетки ИПСК будут использованы в регенеративной медицине, в предпочтительном варианте, с точки зрения отторжения, используют индивидуума, подвергаемого лечению, или другого индивидуума с таким же или по существу с таким же типом главного комплекса гистосовместимости (МНС/ГКГС). По существу, такой же тип ГКГС обозначает тип ГКГС, совпадающий в степени, позволяющей приживлять трансплантируемые клетки с использованием иммуноподавляющего вещества и т.п. , когда клетки, полученные индуцированием дифференциации из клетки ИПСК, полученных из соматических клеток, трансплантируют в индивидуум.
[0035] Соматическая клетка может быть рекомбинантом, чтобы способствовать селекции клетки ИПСК. Конкретные примеры рекомбинантных соматических клеток включают в себя рекомбинантную соматическую клетку, в которой по меньшей мере или репортерный ген, или ген лекарственной резистентности генного локуса высоко экспрессирован в частности в плюрипотентных клетках. Примеры высоко экспрессируемого гена в частности в плюрипотентных клетках включают в себя ген Fbx15, ген Nanog и ген Oct3/4. Примеры репортерного гена включают в себя ген ЗФБ (зеленый флуоресцентный белок), ген люциферазы, ген бета-галактозидазы. Примеры генов лекарственной резистентности включают в себя ген резистентности к бластицидину, ген резистентности к гигромицину, ген резистентности к пуромицину, ген резистентности к неомицину.
[0036] В частности, условия культивирования соматических клеток не ограничены, но могут быть выбраны соответствующим образом в зависимости от цели, их примеры включают в себя температуру культивирования приблизительно 37 град. С и концентрацию CO2 приблизительно 2-5%. Среда, используемая для культивирования соматических клеток, конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели, ее примеры включают в себя минимальную необходимую среду (MEM), модифицированную по способу Дюльбекко среду Игла (DMEM), среду RPMI1640, среду 199, среду F12, содержащую 5-20% масс. сыворотки.
[0037] (Способ введения (а) вещества для перепрограммирования ядра и введения (b) гена H1foo или продукта этого гена в соматическую клетку)
Способ введения (а) вещества для перепрограммирования ядра и введения (b) гена H1foo или продукта этого гена в соматическую клетку реально не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя способ с использованием экспрессионного вектора, способ с использованием мРНК и способ с использованием рекомбинантного белка. С учетом простоты введения в соматическую клетку, более предпочтительно введение вещества для перепрограммирования в соматическую клетку в форме гена или в форме мРНК гена, а не в форме белка, и в более предпочтительном варианте введение в соматическую клетку в форме гена. Таким геном может быть ДНК или РНК, или химера ДНК/РНК, но с точки зрения стабильности предпочтительно использование ДНК. Ген может быть двухцепочечным или одноцепочечным, предпочтительно использование двухцепочечного гена. Примеры предпочтительного вещества для перепрограммирования ядра включают в себя кДНК гена и, в частности, в предпочтительном варианте включают в себя двухцепочечную кДНК гена.
[0038] Аналогичным образом, с учетом простоты введения в соматическую клетку, более предпочтительно введение «гена H1foo, или продукта этого гена» в соматическую клетку в форме гена или в форме мРНК гена, а не в форме белка, в более предпочтительном варианте введение в соматическую клетку в форме гена. Таким геном H1foo может быть ДНК или РНК, или химера ДНК/РНК, но с точки зрения стабильности предпочтительно использование ДНК. Ген H1foo может быть двухцепочечным или одноцепочечным, но предпочтительно использование двухцепочечного гена. Предпочтительные аспекты «гена H1foo или продукта этого гена» включают в себя кДНК гена H1foo и, в частности, в предпочтительном варианте включают в себя двухцепочечную кДНК гена H1foo.
[0039] (Экспрессионный вектор)
При вводе вещества для перепрограммирования ядра в соматическую клетку в форме гена, или когда ген H1foo вводят в соматическую клетку, в предпочтительном варианте может быть использован экспрессионный вектор, полученный посредством инкорпорации вещества для перепрограммирования ядра или гена H1foo в подходящий экспрессионный вектор, содержащий промотор, который способен функционировать в соматической клетке-носителе. Экспрессионный вектор, в который будет инкорпорировано вещество для перепрограммирования ядра или ген H1foo, конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, его примеры включают в себя эписомный вектор, плазмидный вектор и вирусный вектор.
[0040] Примеры промотора, используемого в экспрессионном векторе, включают в себя промотор SR α, ранний промотор SV40, длинные концевые повторы (LTR/ДКП) ретровирусов, цитомегаловирус (CMV/ЦМВ), вирус саркомы Рауса (RSV ВСР), тимидинкиназу симплексного вируса герпеса (HSV-TK ТКСВГ), промотор EF1α, металлотионеин и тепловой шок. Кроме того, энхансер непосредственно раннего человеческого гена CMV может быть использован в качестве промотора. В качестве примера может быть использован промотор CAG (содержащий энхансер CMV, промотор куриный β-актин и поли(А)сигнал гена β-глобина).
[0041] Экспрессионный вектор может содержать, в случае необходимости, энхансер, дополнительный поли(А)сигнал, маркерный ген, точку начала репликации, и ген, кодирующий белок, связанный с точкой начала репликации и контролирующий репродукцию, помимо промотора. Маркерный ген относится к гену, который позволяет сортировать селекцию клетки посредством введения маркерного гена в клетку. Конкретные примеры маркерного гена включают в себя ген лекарственной резистентности, ген флуоресцентного белка, ген люминесцентного энзима и ген хромогенного энзима. Они могут быть использованы по отдельности или в сочетании двух или более. Конкретные примеры гена лекарственной резистентности включают в себя ген резистентности к неомицину, ген резистентности к тетрациклину, ген резистентности к канамицину, ген резистентности к зеоцину и ген резистеностности к гигромицину. Конкретные примеры гена флуоресцентного белка включают в себя ген зеленого флуоресцентного белка (ЗФБ), ген желтого флуоресцентного белка (ЖФБ) и ген красного флуоресцентного белка (КФБ). Конкретные примеры гена люминесцентного энзима включат в себя ген люциферазы. Конкретные примеры гена хромогенного энзима включают в себя ген β-галактозидазы, ген β-глюкуронидазы и ген щелочной фосфатазы.
[0042] Эписомный вектор представляет собой вектор, обладающий способностью внехромосомной автономной репликации. Конкретные средства использования эписомного вектора раскрыты у Юй и др., «Science», №324, 797-801 (2009 г.). В особенно предпочтительном варианте осуществления настоящего изобретения может быть использован эписомный вектор, в котором последовательности loxP размещены в одном и том же направлении в 5' и 3' элементов вектора, необходимых для репликации эписомного вектора. Поскольку эписомные векторы обладают способностью внехромосомной автономной репликации, они могут обеспечить стабильную экспрессию в клетках-носителях без встраивания в геном. Однако желательно удаление вектора сразу после получения клетки ИПСК. С 2 последовательностями loxP, фланкирующими элементы вектора, необходимыми для репликации эписомного вектора, и рекомбиназой Cre, действующей на него для вырезки элементов вектора, компетенция автономной репликации эписомного вектора может быть утеряна и вектор может быть удален из клетки ИПСК на раннем этапе.
[0043] Примеры эписомного вектора, используемого в настоящем изобретении, включают в себя вектор, содержащий последовательности, необходимые для автономной репликации, полученные из вируса EBV, вируса SV40 и т.п. в качестве элементов вектора. Элементы вектора, необходимые для автономной репликации представляют собой, в частности, точку начала репликации и ген, кодирующий белок, который присоединяется к точке начала репликации и контролирует репликацию, и его примеры включают в себя точку начала репликации oriP и ген EBNA-1 в вирусе EBV, и точку начала репликации ori и ген SV40LT в вирусе SV40.
[0044] Кроме того, примеры вектора на основе искусственной хромосомы включают в себя вектор на основе искусственной хромосомы дрожжей (YAC), вектор на основе искусственной хромосомы бактерии (ВАС) и вектор на основе искусственной хромосомы из Р1 (РАС).
[0045] Кроме того, в частности, плазмидный вектор не ограничен, при условии, что он является плазмидным вектором, который может быть экспрессирован в соматической клетке, в которую его помещают, и его примеры включают в себя плазмидный вектор для экспрессии в клетках животных, например рА1-11, pXT1, pRc/CMV, pRc/RSV и pcDNAI/Neo, когда соматическая клетка представляет собой - клетку млекопитающего.
[0046] Примеры вирусного вектора включают в себя вектор на основе ретровируса (в том числе лентивируса), векторы на основе аденовируса, вектор на основе адено-ассоциированного вируса, вектор на основе вируса Сендай (Sendai), вектор на основе вируса герпеса, вектор на основе вируса осповакцины, вектор на основе поксвируса, вектор на основе полиовируса, вектор на основе вируса Синдбис (Sindbis), вектор на основе рабдовируса, вектор на основе парамиксовируса и вектор на основе ортомиксовируса.
[0047] Способ для введения экспрессионного вектора в соматическую клетку конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя липофекцию, микроинъекцию, способ с использованием диэтиламиноэтилдекстрана, способ с использованием генной пушки, электропорацию и способ с использованием фосфата кальция.
[0048] Когда вирусный вектор используют в качестве экспрессионного вектора для введения в соматическую клетку, могут быть использованы вирусные частицы, полученные посредством использования упаковывающей клетки. Такая упаковывающая клетка представляет собой клетку, в которую введены гены, кодирующие структурные белки вируса, и которая, когда рекомбинантный, имеющий вирусный вектор с инкорпорированным в нее целевым геном, вводят в клетку, производит рекомбинантные вирусные частицы, в которые инкорпорирован целевой ген. Упаковывающая клетка конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели, и ее примеры включают в себя упаковывающую клетку на основе клетки HEK293, полученной из человеческой почки, и клетки NIH3T3, полученной из мышиного фибробласта, клетку PLAT-E, которая разработана для экспрессии оболочечных гликопротеинов, полученных из экотропного вируса, клетку PLAT-A, разработанную для экспрессии оболочечных гликопротеинов, полученных из амфотропного вируса, и клетку PLAT-GP, разработанную для экспрессии оболочечных гликопротеинов, полученных из вируса везикулярного стоматита. В частности, клетка PLAT-A и клетка PLAT-GP предпочтительны в плане тропизма, когда вектор на основе рекомбинантного вируса вводят в человеческую соматическую клетку. Способ введения вирусного вектора в упаковывающую клетку конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя липофекцию, электропорацию и способ с использованием фосфата кальция. Способ инфицирования соматической клетки полученными вирусными частицами конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя способ с использованием полибрена.
[0049] Когда вещество для перепрограммирования ядра или ген H1foo вводят в соматическую клетку с использованием экспрессионного вектора, один ген или 2 или более генов могут быть инкорпорированы в один экспрессионный вектор. Инкорпорирование 2-х или более генов в один вектор обеспечивает одновременную экспрессию (далее - «коэкспрессия») 2-х или более генов. Кроме того, все вещества для перепрограммирования ядра, которые будут введены в соматическую клетку, и ген H1foo могут быть инкорпорированы в один экспрессионный вектор.
[0050] Способ инкорпорирования 2-х или более генов в один вектор конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, однако предпочтительным вариантом является инкорпорирование 2-х или более генов с линкерной последовательностью. Такая линкерная последовательность конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели, и ее примеры включают в себя последовательность гена, кодирующего пептид 2А, полученный из вируса ящура (Picornaviridae Aphthovirus) и участка внутренней посадки рибосомы (IRES).
[0051] Способ получения клеток ИПСК в соответствии с настоящим изобретением может включать в себя введение (а) вещества для перепрограммирования ядра и введение (b) продукта гена H1foo в форме мРНК в соматическую клетку. Способ введения мРНК (матричная РНК) в соматические клетки конкретно не ограничен, и известный способ может быть выбран и использован по мере необходимости. Например, доступный на рынке реагент для трансфекции РНК, такой как Lipofectamine® MessengerMAX (производитель - компания «Life Technologies Corporation») и т.п.
[0052] Способ получения клеток ИПСК в соответствии с настоящим изобретением может включать в себя введение (а) вещества для перепрограммирования ядра и введение (b) продукта гена H1foo в форме белка в соматическую клетку. Способ введения такого белка в соматические клетки конкретно не ограничен, и известный способ может быть выбран и использован по мере необходимости. Примеры такого способа включают в себя способ с использованием реагента для трансфекции белка, способ с использованием белка слияния домена белковой трансдукции (ДБТ) и микроинъекция. Доступные на рынке реагенты для белковой трансфекции включают в себя катионные реагенты на основе липидов BioPOTER® Protein Delivery Reagent (производитель - компания «Gene Therapy Systems, Inc.») и Pro-Ject™ Protein Transfection Reagent (производитель - компания «PIERCE»), реагент на основе липидов Profect-1 (производитель - компания «Targeting Systems»), мембранопроницаемые реагенты на основе пептидов Penetratin Peptide (производитель - компания «Q biogene Inc.») и Chariot Kit (производитель - компания «Active Motif»), и GenomONE (производитель - компания «Ishihara Sangyo Kaisha, Ltd.»), в котором используется оболочка ГВЯ (неактивированный вирус Сендай). Введение может быть выполнено в соответствии с протоколами, приложенными к этим реагентам, однако общая процедура следующая: «(а) вещество для перепрограммирования» и/или «(b) ген H1foo или продукт этого гена» в форме белка растворяют в соответствующем растворителе (например, буферный раствор, такой как PBS или HEPES) и инкубируют при комнатной температуре в течение 5-15 мин. после добавления реагента для трансфекции для образования комплекса. Его добавляют в клетки, помещенные в среду без сыворотки, и получаемая культура может быть инкубирована при температуре 37 град. С в течение 1 или более часов. Затем среда может быть удалена и заменена на среду, содержащую сыворотку.
[0053] Примеры PTD (фосфатидилсерин) включают в себя препараты, разработанные с использованием домена клеточной проницаемости белка, например AntP, полученный из Drosophila, TAT, полученный из вируса иммунодефицита человека (ВИЧ), или VP22, полученный симплексного вируса герпеса. Такой PTD может быть использован для введения посредством конструкции экспрессионного фактора на основе белка слияния, в который встроены кДНК «(а) вещества для перепрограммирования ядра» и/или (b) продукты гена H1foo» и последовательность PTD, экспрессируя их рекомбинантно, и сборка белка слияния. Введение такого белка слияния может быть выполнено, как указано выше, за исключением того, что реагент для трансфекции белка не добавляют.
[0054] Микроинъекция представляет собой способ, включающий в себя помещение белкового раствора в стеклянную иглу с диаметром наконечника приблизительно 1 мкм и прокалывание клетки для введения раствора в клетку, способ обеспечивает надежное введение белка в клетку. Способы получения клеток ИПСК посредством введения вещества для перепрограммирования ядра в форме белка с проникающим через клетку пептидом (СРР ПКП), таким как полиаргинин или ТАТ, разработаны на мышах и на человеке, и эти техники также могут быть использованы («Cell Stem Cell», №4, 381-384 (2009 г.)).
[0055] В способе получения клеток ИПСК в соответствии с настоящим изобретением, введение «(а) вещества для перепрограммирования ядра» и введение «(b) гена H1foo или продукта этого гена» в соматическую клетку может быть однократным или многократным. Время введения конкретно не ограничено, но может быть выбрано соответствующим образом в зависимости от цели, и «(а) вещество для перепрограммирования ядра» и «(b) ген H1foo или продукт этого гена» могут быть введены за один и тот же период времени, или они частично или полностью могут введены в разные периоды времени. «Ген H1foo или продукт этого гена» могут быть объектом, в котором использован только ген, объектом, в котором использован только продукт этого гена, или объектом, в котором использован как ген, так и продукт этого гена. Если вещество для перепрограммирования ядра представляет собой ген или продукт этого гена, это может быть объектом, в котором использован только ген, объектом, в котором использован только продукт такого гена, или объектом, в котором использован как ген, так и продукт этого гена. Кроме того, вещество для перепрограммирования ядра представляет собой ген(ы) или продукт таких генов, и 2 или более «генов или продуктов таких генов» используются в сочетании, то это сочетание может быть объектом, в котором используются как определенные гены и продукты этих генов, так и другие гены и продкты этх других генов.
[0056] Количество вводимого гена H1foo или продукта такого гена в соматическую клетку конкретно не ограничено, при условии, что высококачественные клетки ИПСК с меньшей дисперсией качества могут быть получены, когда (а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена вводят в соматическую клетку. Кроме того, когда веществом для перепрограммирования ядра представляет собой ген(ы) или продукт(ы) такого гена, количество(а) вводимых генов или продуктов таких генов в соматическую клетку конкретно не ограничено, при условии, что ядро такой соматической клетки может быть перепрограммировано, и все используемые гены или продукты таких генов могут быть введены в одинаковом количестве или в разных количествах. В примере, в котором используемый ген или продукт такого гена представляет собой ген, ген Oct3/4 предлочтительно вводят в большом количестве, например, приблизительно в 3 раза превышающем количество гена Sox2, гена Klf4 или гена c-Myc («PNAS», №106 (31), 12759-12764 (2009 г.); «J. Biol. Chem.», №287(43), 36273-36282 (2012 г.)).
[0057] Если вещество для перепрограммирования ядра, которое будет использовано в способе получения клеток ИПСК в соответствии с настоящим изобретением представляет собой соединение с низким молекулярным весом, такое соединение с низким молекулярным весом может быть добавлено к соматической клетке посредством разбавления соединения с низким молекулярным весом в водном или безводном растворителе с подходящей концентрацией, добавления раствора соединения с низким молекулярным весом в среду, подходящую для культивирования соматических клеток (например, минимальная необходимая среда (MEM), модифицированная по способу Дюльбекко среда Игла (DMEM), среда RPMI1640, среда 199, среда F12, содержащая 5-20% масс. эмбриональной телячьей сыворотки), таким образом, чтобы достичь достаточной концентрации соединения с низким молекулярным весом для того, чтобы вызвать перепрограммирование ядра в соматической клетке, и в диапазоне, в котором не достигается цитотоксичность, и культивирования клетки в течение определенного периода времени. Концентрация соединения с низким молекулярным весом, являющимся веществом для перепрограммирования ядра, зависит от типа используемого соединения с низким молекулярным весом, но может быть выбрана соответствующим образом в диапазоне приблизительно 0,1-100 нМ (моль/л). Продолжительность контакта конкретно не ограничена, при условии, что период времени контакта достаточно продолжительный, чтобы вызвать перепрограммирование ядра ячейки, однако вещества могут быть оставлены вместе в среде до появления положительной колонии.
[0058] <Другой шаг>
Как указано выше, способ получения клеток ИПСК в соответствии с настоящим изобретением содержит по меньшей мере шаг введения (а) вещества для программирования ядра и шаг введения (b) гена H1foo или продукта этого гена в соматическую клетку («шаг введения»), и далее может содержать другой шаг(и), по мере необходимости. Другой шаг конкретно не ограничен, при условии, что эффект настоящего изобретения не нарушен, но может быть выбран соответствующим образом в зависимости от цели, и его примеры включают в себя шаг культивирования соматической клетки, в котором (а) вещество перепрограммирования ядра и (b) ген H1foo или продукт этого гена введен (далее - «трансфицированная клетка») (далее - «шаг культивирования трансфицированной клетки»).
[0059] (Шаг культивирования трансфицированной клетки)
Шаг культивирования трансфицированной клетки - это шаг культивирования соматической клетки, в которую вводят (а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена. Условия культивирования трансфицированной клетки конкретно не ограничены, и их примеры включат в себя условия, подходящие для культивирования клеток ЭСК. Примеры таких условий включают в себя температуру культивирования приблизительно 37 град. С и концентрацию CO2 приблизительно 2-5%. Кроме того, среда, используемая для культивирования трансфицированной клетки, конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели. Мышиные клетки культивируют в нормальной среде, в которую добавлен фактор, ингибирующий лейкемию (LIF), в качестве дифференциального ингибирующего фактора. Для человеческих клеток желательно добавление базового фактора роста фибробластов (bFGF) и/или фактора стволовых клеток (SCF) вместо LIF. Кроме того, обычно клетки культивируют в присутствии мышиных эмбриональных фибробластов (MEF), обработанных радиацией или антибиотиком, чтобы исключить деление клеток, в качестве питающих клеток. В качестве MEF часто используют клетки STO, но также и клетки SNL (А.П. МакМахон и А. Брэдли, «Cell», №62, 1073-1085 (1990 г.)) часто используют для индуцирования клеток ИПСК. Сокультивирование с питающими клетками может быть начато до введения (а) вещества для перепрограммирования ядра и (b) гена H1foo или продукта этого гена, во время введения или после введения (например, через 1-10 дней).
[0060] Продолжительность шага культивирования трансфицированной клетки конкретно не ограничена, но может быть выбрана соответствующим образом в зависимости от цели.
[0061] 2. Клетки ИПСК
Клетка ИПСК в соответствии с настоящим изобретением, приготовленная этим способом получения клеток ИПСК, обладает плюрипотентностью и самообновлением. Плюрипотентность означает способность дифференцироваться во все три зародышевых листка. Способность к самообновлению означает способность репликации, сохраняя недифференцированное состояние.
[0062] Способ подтверждения того, что клетки, полученные этим методом получения клеток ИПСК представляют собой клетки ИПСК, конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели. Например, если клетка, используемая в качестве соматической клетки представляет собой рекомбинантную соматическую клетку, в которой по меньшей мере или репортерный ген, или ген лекарственной резистентности инкорпорирован в локус гена с высокой экспрессией, в частности в плюрипотентных клетках (например, Fbx15, Nanog или Oct3/4, предпочтительно Nanog или Oct3/4), подтверждение может быть осуществлено с использованием репортерного гена или гена лекарственной резистентности. Конкретные примеры, когда ген, используемый в качестве репортерного гена представляет собой зеленый флуоресцентный белок (GFP), включают в себя способ с использованием подтверждающей GFP-положительной клетки в проточном цитометре, и если ген, использованный в качестве гена лекарственной резистентности представляет собой ген резистентности к пуромицину, подтверждение может быть выполнено добавлением пуромицина к клетке.
[0063] Используемый в настоящем описании термин «высококачественные клетки ИПСК» обозначает клетки ИПСК, качество которых выше, чем клеток ИПСК, полученные таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку. Кроме того, используемый в настоящем описании термин «клетка ИПСК с меньшей дисперсией качества» обозначает клетки ИПСК с дисперсией качества меньшей, чем дисперсия качества клеток ИПСК, полученных таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку. Их качество может быть одного типа или 2 или более типов. Конкретные примеры «высококачественных клеток ИПСК» или «клеток ИПСК с меньшей дисперсией качества» включат в себя клетки ИПСК с одним или более свойствами, выбранными из нижеследующих пп. (a)-(q).
(а) клетки ИПСК, образующие эмбриоидные тельца, в увеличенном количестве на 5% или более, в предпочтительном варианте на 10% или более по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки «OSKL-ИПСК» в примерах, раскрытых ниже), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку при культивировании в течение 5 дней способом [получения эмбриоидных телец (ЕВ)], раскрытым далее.
(b) клетки ИПСК, образующие 78 или более, в предпочтительном варианте 80 или более эмбриоидных телец при культивировании в течение 5 дней способом [получения эмбриоидных телец (ЕВ)], раскрытым далее.
(с) клетки ИПСК, образующие эмбриоидные тельца с дисперсией (σ2) размера (μм2), уменьшенной на 25% или более, в предпочтительном варианте на 40% или более по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки «OSKL-ИПСК» в примерах, раскрытых ниже), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку при культивировании в течение 5 дней способом [получения эмбриоидных телец (ЕВ)], раскрытым далее.
(d) клетки ИПСК, образующие эмбриоидные тельца с дисперсией (σ2) размера (μм2) 10000 (μм2) или менее, в предпочтительном варианте 8000 (μм2) или менее при культивировании в течение 5 дней способом [получения эмбриоидных телец (ЕВ)], раскрытым далее.
(e) клетки ИПСК, в которых процентное количество жизнеспособных клеток (аннексин V(-)/ИП(-)-клетки), согласно измерениям по способу [Анализ апоптоза], раскрытому далее, увеличено в 1,1 раза или более, в предпочтительном варианте в 1,2 раза или более по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки OSKL-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(f) клетки ИПСК, в которых процентное количество жизнеспособных клеток (аннексин V(-)/ИП(-)-клетки), согласно измерениям по способу [Анализа апоптоза], раскрытого далее, составляет 68% или более, в предпочтительном варианте 73% или более.
(g) клетки ИПСК, в которых процентное количество апоптических клеток (сумма аннексин V(-)/ИП(-)-клеток и аннексин V(+)/ИП(+)-клеток), согласно измерениям по способу [Анализ апоптоза], раскрытому далее, уменьшено до 0,75 или менее, в предпочтительном варианте до 0,67 или менее по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки OSKL-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(h) клетки ИПСК, в которых процентное количество апоптических клеток (сумма аннексин V(+)/ИП(-)-клеток и аннексин V(+)/ИП(+)-клеток), согласно измерениям по способу [Анализ апоптоза], раскрытому далее, составляет 30% или менее, в предпочтительном варианте 25% или менее.
(i) клетки ИПСК, в которых уровень экспрессии гена Ki67 или гена PCNA (оба гена известны как маркеры пролиферации клеток), измеренный способом, раскрытым далее в разделе [Количественный RT-PCR-анализ], увеличен в 1,7 раза, в предпочтительном варианте в 2,2 раза по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки OSKL-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку
(j) клетки ИПСК, в которых уровень экспрессии гена Ki67 или гена PCNA (оба гена известны как маркеры пролиферации клеток), измеренный способом, раскрытым далее в разделе [Количественный RT-PCR-анализ], составляет 0,65 или более, в предпочтительном варианте 0,73 или более по сравнению с ЭСК.
(k) клетки ИПСК с более высокой химерной компетенцией по сравнению с клетками ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки «OSKL-iPS» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(l) клетки ИПСК с более высоким потенциалом трансмиссии зародышевой линии по сравнению с клетеами ИПСК (предпочтительно «клетки OSK-ИПСК» или «клетки «OSKL-iPS» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(m) клетки ИПСК, в которых уровень экспрессии гена SRF (ген SRF известен как маркер хромосомной аберрации, уровень экспрессии которого уменьшается при наличии хромосомной аберрации), измеренный способом, раскрытым далее в разделе [Количественный RT-PCR-анализ], увеличен в 1,1 раза, в предпочтительном варианте в 1,2 раза по сравнению с клетками ИПСК (предпочтительно «клетки OSKL-ИПСК» или «клетки OSK-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(n) клетки ИПСК, в которых уровень экспрессии гена ACTG2 (ген ACTG2 известен как маркер хромосомной аберрации, уровень экспрессии которого уменьшается при наличии хромосомной аберрации), измеренный способом, раскрытым далее в разделе [Количественный RT-PCR-анализ], увеличен в 1,2 раза, в предпочтительном варианте в 1,5 раза по сравнению с клетками ИПСК (предпочтительно «клетки OSKL-ИПСК» или «клетки OSK-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(о) клетки ИПСК, в которых дисперсия (σ2) уровней экспрессии гена Oct3/4 (маркер недифференцированного состояния стволовых клеток), измеренная способом, раскрытым далее в разделе [Количественный RT-PCR-анализ], уменьшена на 10% или более, в предпочтительном варианте на 20% или более по сравнению с клетками ИПСК (предпочтительно «клетки OSKL-ИПСК» или «OSK-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(р) клетки ИПСК, в которых число жизнеспособных клеток, измеренное способом, раскрытым далее в разделе [Сравнение выживаемости клеток на раннем этапе после индукции дифференциации], увеличено в 1,2 раза, в предпочтительном варианте в 1,5 раза по сравнению с клетками ИПСК (предпочтительно «клетки OSKL-ИПСК» или «клетки OSK-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
(q) клетки ИПСК, в которых дисперсия (σ2) уровней экспрессии гена Oct3/4 (маркер недифференцированного состояния стволовых клеток), согласно измерениям по способу, раскрытому далее в разделе [Сравнение выживаемости клеток на раннем этапе после индукции дифференциации ИПСК], уменьшена на 10% или более, в предпочтительном варианте на 20% или более по сравнению с клетками ИПСК (предпочтительно «клетки OSKL-ИПСК» или «OSK-ИПСК» в примерах, раскрытых далее), полученными таким же образом, но без введения гена H1foo или продукта этого гена в соматическую клетку.
[0064] Вид организма, из которого получены клетки ИПСК, конкретно не ограничен, но может быть выбран соответствующим образом в зависимости от цели, и примеры видов включают в себя любых млекопитающих, таких как человек, мышь, крыса, корова, овца, лошадь и обезьяна.
[0065] 3. Агент для улучшения качества клеток ИПСК
Агент для улучшения качества клеток ИПСК в соответствии с настоящим изобретением включает в себя по меньшей мере ген H1foo или продукт этого гена, и может дополнительно содержать другую конфигурацию, по мере необходимости. Ген H1foo или продукт этого гена является сходным с раскрытым для способа получения клеток ИПСК. Кроме того, ген H1foo или продукт этого гена может содержать сходную мутацию, раскрытую для способа получения клеток ИПСК. Кроме того, может быть использован ген H1foo, встроенный в экспрессионный вектор, содержащий промотор, который может функционировать в соматической клетке-носителе.
[0066] 4. Состав для получения клеток ИПСК
Состав для получения клеток ИПСК в соответствии с настоящим изобретением содержит по меньшей мере (а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена, и может дополнительно содержать другую конфигурацию, по мере необходимости.
[0067] <(а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена>
(а) Вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена сходны с теми, что раскрыты для способа получения клеток ИПСК. Кроме того, ген H1foo или продукт этого гена и ген или продукт этого гена, являющийся веществом для перепрограммирования ядра, может содержать сходную мутацию, раскрытую для способа получения клеток ИПСК.
[0068] Примеры предпочтительной формы вещества для перепрограммирования ядра в составе для получения клеток ИПСК включают в себя форму гена, встроенного в вектор, форму синтетической мРНК и форму белка (предпочтительно рекомбинантного). Примеры вектора включают в себя примеры, сходные с раскрытыми для способа получения клеток ИПСК. Синтетическая мРНК и белок (предпочтительно рекомбинантный) могут быть получены известным способом.
[0069] Состав для получения клеток ИПСК может быть упакован так, что (а) вещество для перепрограммирования ядра и (b) ген H1foo или продукт этого гена находятся раздельно в индивидуальных контейнерах или вместе в одном контейнере, или в группе в контейнере с любым количеством на контейнер.
[0070] Количество (а) вещества для перепрограммирования ядра и (b) гена H1foo или продукта этого гена в составе для получения клеток ИПСК конкретно не ограничено, и все вещества для перепрограммирования ядра и/или ген H1foo или продукт этого гена могут содержаться в одинаковых количествах или в разных количествах.
[0071] Состав для получения клеток ИПСК может содержать, помимо (а) вещества для перепрограммирования ядра и (b) гена H1foo или продукта этого гена, ген или продукт этого гена отличных от них. Кроме того, когда используют вектор на основе вируса, состав может содержать, например, упаковывающую клетку. Примеры такого гена или продукта такого гена, отличного от генов или продуктов таких генов, и упаковывающей клетки включают в себя примеры, раскрытые для способа получения клеток ИПСК.
[0072] Настоящее изобретение также содержит, в качестве иных объектов,
«Способ улучшения качества клеток ИПСК, содержащий шаг введения гена H1foo или продукта этого гена в соматическую клетку»,
«Использование гена H1foo или продукта этого гена в производстве агента для улучшения качества клеток ИПСК», и
«Использование (а) вещества для перепрограммирования ядра и (b) гена H1foo или продукта этого гена в производстве состава для получения клеток ИПСК».
[0073] Шаг введения гена H1foo или продукта этого гена в соматическую клетку в «способе улучшения качества клеток ИПСК» тот же самый, что и шаг в способе получения клеток ИПСК в соответствии с настоящим изобретением. Включение такого шага позволяет улучшить качество клеток ИПСК и получить высококачественные клетки ИПСК.
[0074] Настоящее изобретение будет раскрыто далее более подробно с использованием примеров, но настоящее изобретение не ограничено этими примерами. Все эксперименты, раскрытые ниже, проведены в соответствии с Руководством по экспериментированию на животных и ДНК Университета Кэйо, утвержденным Комитетом по этике Университета Кэйо, и основаны на Руководстве о заботе и использовании лабораторных животных Национального института здравоохранения.
Примеры
[0075] 1. Материалы и способ
[Плазмидная конструкция]
Группы H1foo cDNA и H1c cDNA (Т. Тераниши и др., «Быстрая замена соматических линкерных гистонов на ооцит-специфичные линкерные гистоны H1foo в ядерном транспорте». «Developmental Biology», №266, 76-86 (2004 г.).) были вставлены соответственно в рестриктивный фермент BamH1-Sal1 и EcoR1-Sal1 в плазмиде pMXs, и было подтверждено секвенированием ДНК, что H1foo cDNA и H1c cDNA вставлены.
[0076] [Получение мышиных клеток ИПСК и способ их культивирования]
1) Получение мышиных клеток ИПСК проведено в соответствии с протоколом, описанным в литературе (К. Такахаши, К. Окита, М. Накагава и С. Яманака, «Индукция плюрипотентных стволовых клеток из фибробластных культур», «Nature Protocols», №2, 3081-3089 (2007 г.)). Однако в настоящем исследовании ген H1foo также использовали в добавление к генам, раскрытым в этом описании. В частности, клетки ИПСК были получены из мышиных фибробластов или хвостовых фибробластов (далее - «Nanog-GFP-экспрессирующие фибробласты») Nanog-GFP - IRES-puro трансгенных мышей (К. Окита, Т. Ичисака и С. Яманака, «Получение компетентной зародышевой линии плюрипотентных стволовых клеток», «Nature», №448, 313-317 (2007 г.)) с использованием вектора на основе ретровируса pMXs, содержащего гены Oct3/4, Sox2, Klf4 и H1foo (нуклеотидная последовательность указана в идентификаторе последовательности №3: ВС137916). В качестве контроля ген DsRed был использован вместо гена H1foo. Работа с лабораторными мышами выполнялась в соответствии с Руководством по экспериментированию на животных и ДНК Университета Кэйо.
[0077] 2) Мышиные ЭСК (B6J-23UTR) (И. Танимото, и др., «Эмбриональные стволовые клетки, полученные из мышей C57BL/6J и C57BL/6N», «Comparative Medicine», №58, 347-352 (2008 г.)) были получены из Центра ресурсов лабораторных животных при Университете Цукубы и использованы в соответствии с Руководством по распространению и утилизации человеческих эмбриональных стволовых клеток Министерства образования, культуры, спорта, науки и технологии Японии.
[0078] 3) Клеточные линии мышиных клеток ИПСК и клеток ЭСК были культивированы и хранились на мышиных эмбриональных фибробластах, облученных рентгеновским излучением (iMEF питающие клетки), полученных из дикого типа мышей ICR, в культуральной среде DMEM (производитель - компания «Sigma Aldrich Co. LLC.»), содержащей 20% среды KnockOut Strom Replacement (производитель - компания «Gibco»), в 1 мМ (миллимоль на литр) среды GlutaMAX (производитель - компания «Gibco»), 1 мМ заменимых аминокислот (производитель - компания «Sigma Aldrich Co. LLC.»), 0,1 мМ 2-меркаптоэтанола, 50 ед. пенициллина, 50 мг/мл стрептомицина (производитель - компания «Gibco») и мышиного фактора, ингибирующего лейкемию (LIF) (далее - «культуральная среда клеток ИПСК»). Культуральную среду мышиных клеток ИПСК меняли каждые 2-3 дня, и клетки были субкультивированы с использованием 0,5 мМ трипсина-EDTA (производитель - компания «Gibco») каждые 2-3 дня.
[0079] [Получение эмбриоидных телец (ЕВ)]
Получение ЭТ из ИПСК было выполнено посевом 5×104 ИПСК, полученных с 1 мг/мл коллагеназы IV, на низкий планшет 100 мм (производитель - компания «AGC Techno Glass Co. Ltd.») и культивированием клеток в течении 5 дней в присутствии минимальной необходимой среды Alpha Medium (производитель - компания «Gibco»), содержащей 20% ФБС (производитель - компания «Gibco»), 2 мМ GlutaMAX (производитель - компания «Gibco»), 0,1 мМ заменимых аминокислот (производитель - компания «Sigma Aldrich Co. LLC.»), 0,1 мМ 2-меркаптоэтанола, 50 ед./мл пенициллина и 50 мг/мл стрептомицина (производитель - компания «Gibco») (далее - «культуральная среда для получения ЕВ»). Культуральную среду для получения ЭТ меняли каждые 2-3 дня. В качестве контроля использовали клетки ЭСК.
[0080] [Получение тератомы]
Тератомообразующий потенциал клеток ИПСК был подтвержден гематоксилин-эозиновым (НЕ) окрашиванием после инъекции клеток ИПСК в семенники мышей SCID (производитель - компания «CLEA Japan, Inc.») с использованием анестезии в виде смеси кетамина (50 мг/кг), ксилазина (10 мг/кг) и хлорпромазина (1,25 мг/кг), с умерщвлением мышей смещением шейных позвонков приблизительно через 8 недель, фиксируя части ткани, в которые были инжектированы клетки ИПСК, в 10%-м параформальдегиде (ПФА) в течение суток, с встраиванием секций в парафине. Анестезия мышей была выполнена соответствующим образом с мониторингом пульса, расслабления мышц и чувственного отклика (т.е. отсутствие реакции при ущемлении хвоста мыши).
[0081] [Иммуногистохимическое окрашивание]
Клетки ИПСК и фибробласты, посеянные на стеклянную чашку (производитель - компания «AGC Techno Glass Co. Ltd»), были однократно промыты буфром PBS и зафиксированы в 4%-м параформальдегиде (производитель - компания «Muto Pure Chemicals Co., Ltd.») при температуре 4 град. С в течение 15 мин. Зафиксированные клетки были обработаны для пермеабилизации в 0,5%-м Triton Х-100 в PBS при комнатной температуре в течение 10 мин., затем заблокированы в блокирующем реагенте ImmunoBlock (производитель - компания «DS Pharma Biomedical Co. Ltd.») в течение 20 мин., инкубированы в присутствии 4 основных антител (анти-Oct3/4) [антитело Oct3/4 8629, производитель - компания «Santa Cruz Biotechnology, Inc.»], анти-Nanog [RCAB0001P, производитель - компания «ReproCELL Inc.»], анти-SSEA1 [sc-21702, производитель - компания «Santa Cruz Biotechnology, Inc.»] и анти-H1foo [HPA037992, производитель - компания «Sigma Aldrich Co. LLC.»]) при комнатной температуре в течение 60 мин., промыты реагентом ImmunoBlock (производитель - компания «DS Pharma Biomedical Co. Ltd.»), и инкубированы с вторичными антителами (кроличий анти-IgG и мышиный анти-IgG или антитела IgM, конъюгированные с Alexa Fluor 488 или Alexa Fluor 568 (производитель - компания «Life Technologies Corporation») в соответствии с первичными антителами при комнатной температуре в течение 60 мин. После окрашивания ядра 6-диамидин-2-фенилиндолом DAPI/ДАФИ, производитель - компания «Life Technologies Corporation»), наблюдение за флуоресценцией было выполнено с использованием флуоресцентного лазерного микроскопа с цветной камерой на основе прибора с зарядовой связью (BZ-9000, производитель - компания «Keyence Corporation»), и с использованием оптического микроскопа (IX71, производитель - компания «Olympus Corporation») и лазерного конфокального микроскопа (LSM 510 МЕТА, производитель - компания «Carl Zeiss Jena GmbH»).
[0082] [Количественный RT-PCR-анализ]
Все пробы РНК были изолированы с использованием реагента TRIZOL (производитель - компания «Life Technologies Corporation») в соответствии с руководством, приложенным к продукту. Концентрация и чистота проб РНК были измерены спектрофотометром ND-1000 (производитель - компания «Thermo Fisher Scientific Inc.»), и препарирование кДНК было выполнено с использованием ReverTra Асе qPCR RT Master Mix (производитель - компания «Toyobo Co., Ltd.»). Количественный анализ с использованием полимеразной цепной накции с обратной транскрипцией (RT-PCR-анализ) был выполнен на системе 7500 Real Time PCR (производитель - компания «Life Technologies Corporation») с использованием вещества SYBR Premix ExTaq (производитель - компания «Takara Bio Inc.»). Количество мРНК было нормализовано с количеством мРНК GAPDH. Нуклеотидные последовательности наборов праймеров, используемые для количественного RT-PCR-анализа, указаны далее в Таблице 1.
[0083]

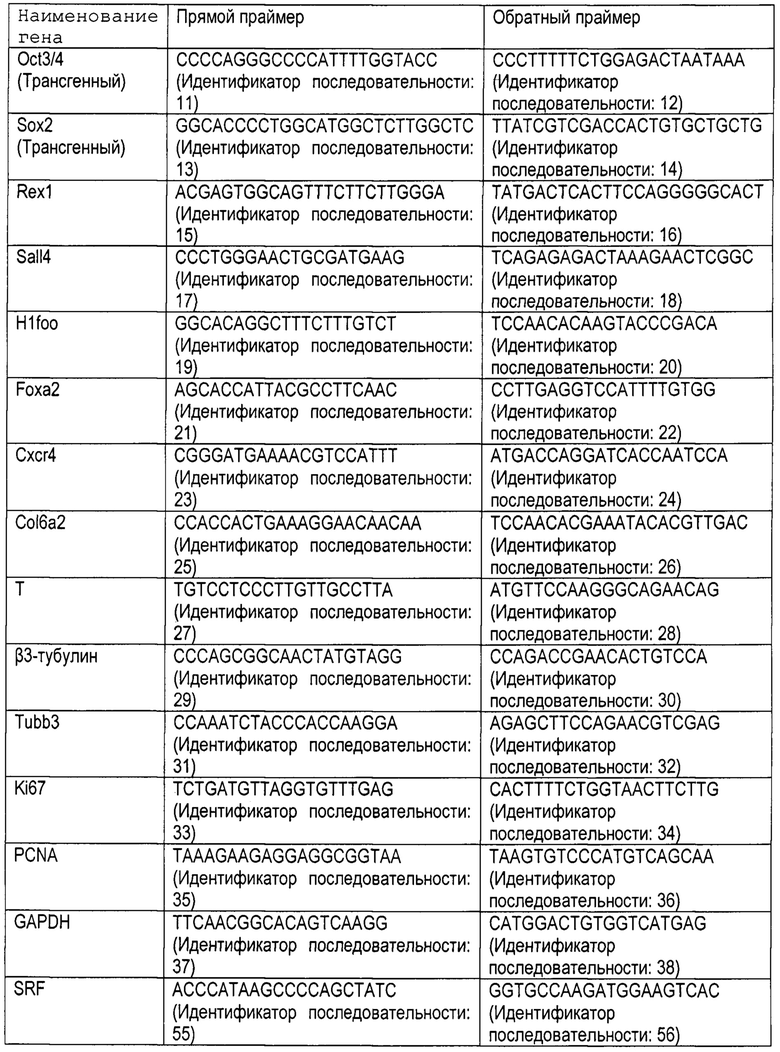
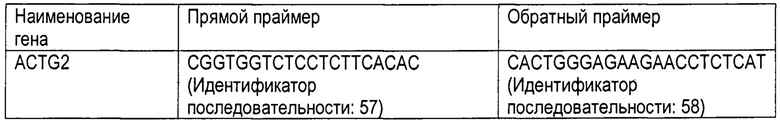
[0084] [Анализ апоптоза]
Клетки ИПСК через 1 день после начала культивирования в культуральной среде для получения ЕВ были трипсинизированы и суспендированы в буфере для связывания аннексина А5 (производитель - компания «Takara Bio Inc.»), затем ранние апоптические клетки и поздние апоптические клетки были окрашены аннексин-А5-FITC и иодированным пропидием (PI) (производитель - компания «Becton, Dickinson and Company») соответственно, согласно руководству, приложенному к этим продуктам. Клетки были профильтрованы через нейлонную мембрану с размером поры 70 мм, и флуоресцентно окрашенные клетки были проанализированы методом проточной цитометрии на приборе FACS Aria3 (производитель - компания «Becton, Dickinson and Company») с использованием программного обеспечения CellQuest (производитель - компания «Becton, Dickinson and Company»), чтобы определить процентное количество апоптических клеток, используя программное обеспечение FlowJo (производитель - компания «Tree Star, Inc.»).
[0085] [Анализ метилирования ДНК]
Чтобы проанализировать уровень метилирования ДНК в областях промотора генов Oct3/4 и Nanog с использованием бисульфитного секвенирования, геномная ДНК была изолирована и очищена от проб, используя очистительный комплект SV Genomic DNA Purification Kit (производитель - компания «Promega Corporation»). Неметилированный цитозин (С) в очищенной геномной ДНК был преобразован в урацил (U), используя комплект EZ DNA Methylation-Gold Kit (производитель - компания «ZYMO RESEARCH») в соответствии с руководством, приложенным к продукту, и была приготовлена входная ДНК для бисульфитной полимерной цепной реакции. Используя такую входную ДНК, праймеры и вещество TaKaRa EpiTaq HS (производитель - компания «TaKaRa Bio Inc.»), выполнили бисульфитную полимерную цепную реакцию (PCR) в условиях реакции PCR (1 цикл денатурирования при температуре 98 град. С в течение 10 мин.; 40 циклов амплификации при температуре 95 град. С в течение 20 с, при температуре 55 град. С в течение 30 с и при температуре 72 град. С в течение 60 с; и 1 цикл финального удлинения при температуре 72 град. С в течение 5 мин.) Нуклеотидные последовательности наборов праймеров, используемые для бисульфитного ПЦРОТ-анализа, указаны далее в Таблице 2.
[0086]
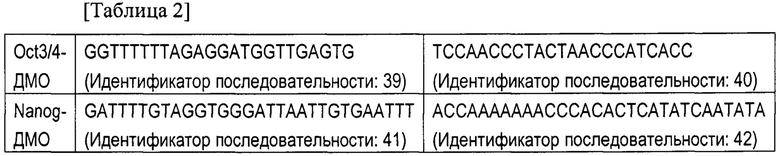
[0087] Продукты полимерной цепной реакции, были клонированы в Vector pGEM-T (производитель - компания «Promega Corporation») посредством ТА-клонирования после очистки и секвенирования. Чтобы проанализировать состояние метилирования ДНК в межгенной дифференциально-метилированной области (ДМО) (МГ ДМО), расположенной между генами Dlk1 и Dlk2, и ДМО Gtl2 (Gtl2-flMO), геномная ДНК была очищена, как указано выше, приготовлена входная ДНК для бисульфитной полимерной цепной реакции, и затем было выполнено пиросеквенирование с использованием прибора PyroMark Q24 (производитель - компания «QIAGEN») в соответствии с руководством, приложенным к изделию. Нуклеотидные последовательности праймеров, используемых для пиросеквенирования, приведены в Таблице 3 ниже.
[0088]

[0089] [Агрегация сокультуры ЭСК и ИПСК]
Эмбрионы на 2-клеточном этапе были собраны из суперовулированных и естественно спаренных самок мышей ICR (CD-1 (R)), и затем культивированы до этапа бластоцисты. Клетки ЭСК и ИПСК были диссоциированы 0,25%-м трипсином непосредственно перед агрегацией сокультуры, и 15-20 клеток были агрегированы с бластомерами на 8-клеточном этапе, из которых была удалена оболочка zonae pellucidae. Химерные эмбрионы на этапе бластоцисты были перенесены в рог матки псевдобеременных мышей ICR на 2,5 день после совокупления.
[0090] [Анализ глобальной экспрессии генов]
Общие РНК были изолированы из клеток ИПСК и ЭСК. Антисмысловая РНК, помеченная цианином, была амплифицирована с использованием комплекта Quick Amp Labeling Kit (производитель - компания «Agilent Technologies»), гибридизирована на Whole Mouse Genome Oligo Microarray (производитель - компания «Agilent Technologies») с комплектом гибридизации экспрессии генов, и подвергнута микроматричному анализу на сканере Agilent Microarray Scanner. Данные были проанализированы, используя программное обеспечение GX12.0 (производитель - компания «Agilent Technologies»).
[0091] [Статистический анализ]
Значения на гистограмме со столбцами погрешности указаны на фигурах как среднее ± стандартное отклонение (SME). Анализ данных был выполнен с использованием программного обеспечения StatView J-4.5. Сравнение 2 групп было выполнено, используя t-Критерий Стьюдента. Сравнение среди групп было выполнено с использованием однофакторного дисперсионного анализа и ретроспективного анализа Бонферрони. Значки «*» и «**» на фигурах указывают на значимость (Р<0,05 и Р<0,01 соответственно).
[0092] 2. Результаты
[Получение клеток ИПСК, используя экзогенный ген H1foo]
Исследовано влияние H1foo на перепрограммирование соматической клетки.
Во-первых, способ, раскрытый выше в разделе [Иммуногистохимическое окрашивание], был выполнен, чтобы подтвердить исключительную экспрессию экзогенного H1foo в ядре (Фигура 1а), и что он расположен, в основном, по периферии в ядре (Фигура 1b). Набухание ядра и уменьшение области с высокой плотностью электронов внутри мембраны ядра (гетерохроматин) за счет экспрессии экзогенного H1foo не наблюдалось (Фигура 1с).
[0093] Затем, в соответствии со способом, раскрытым выше в разделе [Получение мышиных клеток ИПСК и способ их культивирования], 3 вещества для перепрограммирования ядра (гены Oct3/4, Sox2 и Klf4 [гены OSK]), а также ген H1foo (гены OSK+H1foo) были введены в мышиные фибробласты, являющиеся соматическими клетками, используя вектор на основе ретровируса, и окрашивание щелочной фосфатазой (ALP) было выполнено с использованием комплекта TRACP & ALP Double-Stain Kit (производитель - компания «Takara Bio Inc.») на 20 день, чтобы определить число колоний, которые являются ALP-положительными и состоящими из ЭСК-подобных клеток. Щелочная фосфатаза (ALP) известна как маркер стволовых клеток. В качестве контроля также были проведены эксперименты с введением только генов OSK или введением только гена линкерного гистона H1 (гены OSK+H1c) вместо H1foo в генах OSK+H1foo. В результате, продемонстрировано, что введение генов OSK+H1foo в мышиные фибробласты приводит к отмечаемому увеличению числа колоний ALP-положительных ЭСК-подобных клеток (клеток ИПСК) по сравнению с введением генов OSK или генов OSK+H1c (Фигура 2а).
[0094] В соответствии со способом, раскрытым выше в разделе [Иммуногистохимическое окрашивание], проанализирована экспрессия 3 маркеров плюрипотентности (Oct3/4, Nanog и SSEA1). Анализ показал, что клетки OSK+H1foo-ИПСК экспрессируют плюрипотентные маркеры, как и клетки OSK-ИПСК (Фигура 2b). И наоборот, экспрессия эндогенного H1foo не наблюдалась ни во время процесса индукции клеток ИПСК, ни в полученных клетках ИПСК (Фигура 2b, с).
[0095] Кроме того, гены OSK+H1foo были введены в Nanog-GFP-экспрессирующие фибробласты, чтобы исследовать качество полученных клеток ИПСК (Фигура 3а), используя сигнал GFP в качестве индикатора. Nanog-GFP-положительные клетки известны в качестве маркера высококачественных стволовых клеток. Стоит отметить то, что введение гена H1foo в дополнение к генам OSK в Nanog-GFP-экспрессирующие фибробласты увеличивает число колоний Nanog-GFP-положительных ЭСК-подобных клеток (клеток ИПСК) приблизительно в 8 раз по сравнению с введением генов OSK (Фигура 3b). Кроме того, соотношение Nanog-ЗБФ-положительных ЭСК-подобных клеток (клеток ИПСК) к популяции полученных клеток ИПСК достигло 90% или более (Фигура 3c). И наоборот, показано, что введение генов OSK и гена c-Myc (гены OSKM) или введение гена H1foo, а также генов OSKM (гены OSKM+H1foo) в Nanog-GFP-экспрессирующие фибробласты приводит к увеличенному числу самих Nanog-GFP-положительных колоний (Фигура 3b), но соотношение Nanog-GFP-положительных клеток к популяции полученных клеток ИПСК низкое, по сравнению с введением генов OSK+H1foo (Фигура 3c). Эти результаты указывают на то, что введение генов OSK+H1foo в соматические клетки позволяет получить популяции клеток ИПСК более высокого качества по сравнению с введением генов OSK или введением генов OSKM в известном уровне техники.
[0096] [Свойства клеток OSK+H1foo-ИПСК]
Свойства клеток OSK+H1foo-ИПСК были подробно проанализированы. Анализ экспрессии 4-х маркеров плюрипотентности (Oct3/4, Sox2, Rex1 и Sall4) в соответствии со способом, раскрытым выше в разделе [Количественный RT-PCR анализ], показал, что клетки OSK+H1foo-ИПСК экспрессируют маркеры плюрипотентности, как и клетки OSK-ИПСК (Таблица 4). Экспрессия трансгенов, включая ген H1foo, была подавлена (Таблица 5). Также была проанализирована пролиферация клеток, и анализ показал, что уровень пролиферации клеток OSK+H1foo-ИПСК был эквивалентным клеткам OSK-ИПСК.
[0097]
[Таблица 4]
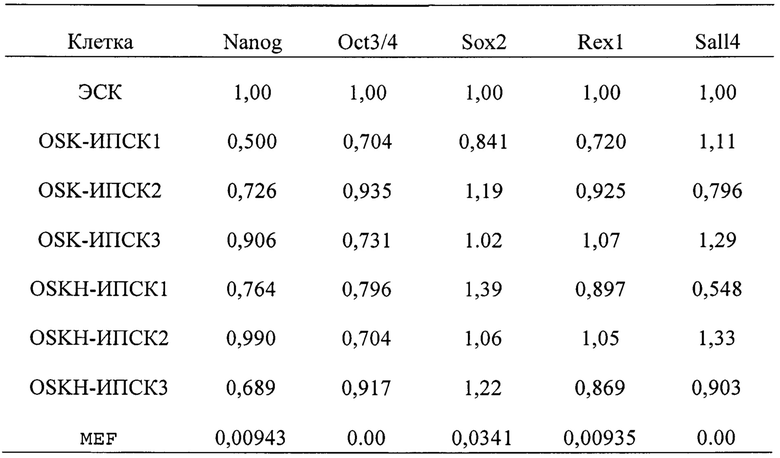
[0098] В Таблице 4 показаны результаты анализа экспрессии 5-ти маркеров плюрипотентности (Nanog, Oct3/4, Sox2, Rex1 и Sall4) в клетках ЭСК (ЭСК), 3-х клонах клеток OSK-ИПСК (OSK-ИПСК 1-3), 3-х клонах клеток OSK+H1foo-ИПСК (OSKH-ИПСК 1-3) и клетках MEF (MEF). Уровни экспрессии маркеров плюрипотентности выражены в качестве значений относительно результата для клеток ЭСК, который равен 1.
[0099]
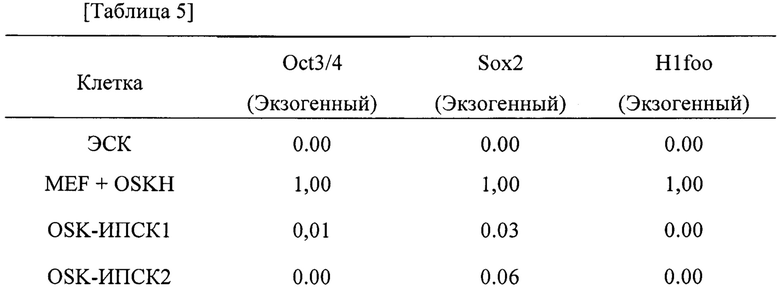

[0100] В Таблице 5 показаны результаты экспрессии 3-х экзогенных белков (Oct3/4, Sox2 и H1foo) в клетках ЭСК (ЭСК), OSKH-трансдуцированных MEF (MEF+OSKH), 3-х клонов клеток OSK-ИПСК (OSK-ИПСК 1-3), 3-х клонов клеток OSK+H1foo-ИПСК (OSKH-ИПСК 1-3) и клеток MEF (MEF). Уровни экспрессии маркеров плюрипотентности выражены в качестве значений относительно результата по клеткам OSKH-трансдуцированным MEF, который принят за 1 (единицу).
[0101] Сравнены глобальные профили транскриптома гена между клетками OSK+H1foo-ИПСК и клетками ЭСК или OSK-ИПСК (Фигуры 4а и b). В результате, продемонстрировано, что уровень экспрессии генов в клетках OSK+H1foo-ИПСК и уровень экспрессии генов в ЭСК и OSK-ИПСК очень сходны (коэффициент корреляции [R2] 0,99).
[0102] Анализ метилирования геномной ДНК был выполнен в соответствии со способом в разделе [Анализ метилирования ДНК]. Результаты показали, что ДНК в промоторных областях 2-х маркерных генов плюрипотентности (Oct3/4 и Nanog), IG-DMR и Gtl2-DMR были деметилированы в клетках OSK+H1foo-ИПСК, как и в клетках ЭСК и клетках OSK-ИПСК (Фигуры 5a и b). Уровень деметилирования оказался гетерогенным среди клонов (клеточных групп) клеток OSK-ИПСК, но такая гетерогенность оказалась меньшей среди клеток OSK+H1foo-ИПСК (Фигуры 5a и b).
[0103] [Свойства эмбриоидных телец (ЕВ), полученных из клеток OSK+H1foo-ИПСК]
Для индукции дифференциации из клеток ИПСК в различные клетки обычно сначала получают ЕВ, и затем индуцируют дифференциацию способом для индукции дифференциации в зависимости от типа клеток. Поскольку считается, что гомогенность числа полученных ЕВ и размер ЕВ влияет на эффективность последующей индукции дифференциации, клетки ИПСК, способные формировать большое количество ЕВ гомогенных размеров, считаются желательным вариантом. Поэтому был проанализирован потенциал клеток OSK+H1foo-ИПСК к формированию ЕВ.
[0104] При получении ЕВ из клеток OSK+H1foo-ИПСК в соответствии со способом, раскрытым выше в разделе [Получение эмбриональных телец (ЭТ)], количество ЕВ, полученных из таких клеток OSK+H1foo-ИПСК, оказывалось больше (увеличение примерно на 12%), а также размер ЕВ (μм2) по сравнению с ЕВ, полученными из клеток OSK-ИПСК (Фигуры 6а-с), при этом дисперсия (σ2) размера ЕВ (μм2) оказывалась меньше (приблизительно на 50%) (Фигуры 6d and е).
[0105] Анализ процентного количества апоптических клеток в соответствии с методом, раскрытым выше в разделе [Анализ апоптоза], показал, что ЕВ, полученные из клеток OSK+H1foo-ИПСК, содержат меньшее процентное количество ранних апоптических клеток (аннексин V-положительные(+)/ИП-отрицательные(-)-клетки) и поздних апоптических клеток (аннексин V(+)/ИП(+) и аннексин V(-)/ИП(+)), и более высокое процентное количество жизнеспособных клеток (аннексин V(-)/ИП(-)-клетки) по сравнению с ЕВ, полученными из клеток OSK-ИПСК (Фигуры 7а и b). ЕВ, полученные из клеток OSK+H1foo-ИПСК, содержат процентное количество жизнеспособных клеток (аннексин V(-)/ИП(-)-клетки), увеличенное приблизительно в 1,2 раза по сравнению с ЕВ, полученными из клеток OSK-ИПСК. Кроме того, ЕВ, полученные из клеток OSK+H1foo-ИПСК, содержат процентное количество апоптических клеток (сумма аннексин V(+)/ИП(-)-клеток и аннексин V(+)/PI(+)-клеток), уменьшенное приблизительно до 0,65 по сравнению с ЕВ, полученными из клеток OSK-ИПСК.
[0106] Кроме того, анализ клеток ИПСК на второй день после начала культивирования в культуральной среде для получения ЕВ на экспрессии 2-х маркеров клеточной пролиферации (Ki67 и PCNA) в соответствии со способом, раскрытым выше в разделе [Количественный ПЦРОТ-анализ], показал, что ЕВ, полученные из клеток OSK+H1foo-ИПСК, обладают более высоким уровнем экспрессии (увеличенным приблизительно до 2,4-2,6 раз) маркеров пролиферации по сравнению с ЕВ, полученными из клеток OSK-ИПСК (Фигуры 8а и b).
Вышеизложенные результаты указывают на то, что клетки OSK+H1foo-ИПСК образуют большее количество ЕВ с относительно сходными размерами, и могут формировать ЕВ с меньшим процентным количеством мертвых клеток (высококачественные ЕВ) по сравнению с клетками OSK.
[0107] [Плюрипотентность клеток OSK+H1foo-ИПСК]
Проанализирована плюрипотентность клеток OSK+H1foo-ИПСК. Химерные мыши, обладающие нормальным числом хромосом, были получены из 2-х клонов (OSKH1 и 3) клеток OSK+H1foo-ИПСК в соответствии со способом, раскрытым выше в разделе [Агрегация сокультуры ЭСК и ИПСК]. Кроме того, в качестве контроля, химерные мыши с нормальным числом хромосом были получены из 2-х клонов (OSK1 и 2) клеток OSK-ИПСК в соответствии с тем же самым способом. Поскольку клетки ИПСК и ЭСК, использованные в эксперименте, были приготовлены из соматических клеток черных мышей, более высокая химерная компетенция клеток ИПСК и ЭСК приводит к появлению поколения с большим процентным количеством черных мышей. В результате исследования поколения химерных мышей было показано, что 2 клона (OSKH1 и 3) клеток OSK+H1foo-ИПСК обладают химерной компетенцией, равной или превышающей таковую, чем 2 клона (OSK1 и 2) клеток OSK-ИПСК (Фигуры 9а и b). В частности, показано, что OSKH3 обладает химерной компетенцией на таком же уровне, как и клетки ЭСК (Фигуры 9а и b).
[0108] Потенциал трансмиссии зародышевой линии относится к потенциалу плюрипотентных стволовых клеток, таких как клетки ЭСК и ИПСК, чтобы дифференцироваться в зародышевые клетки и трансмиссии генов, полученных из плюрипотентных стволовых клеток, и передавать гены, полученные от плюрипотентных стволовых клеток, следующему поколению через химеру. Этот потенциал трансмиссии зародышевой линии представляет соообой один из самых строгих отличительных признаков плюрипотентных стволовых клеток, и один из признаков, указывающих на то, что плюрипотентные стволовые клетки обладают высоким качеством. Таким образом, потенциал трансмиссии зародышевой линии у 100% химерных мышей был исследован с использованием искусственного оплодотворения (IVF ИО). В результате, 4 химерных мыши (OSKH1, OSKH3-1, OSKH3-2 и OSKH3-3), полученные из клеток OSK+H1foo-ИПСК, продемонстрировали более высокий репродуктивный потенциал, чем 2 химерных мыши (OSK2-1 и OSK2-2), полученные из клеток OSK-ИПСК (Фигура 10а), и произвели множество мышат с разноцветным покровом (Фигуры 10b и с). У этих мышат не было обнаружено аномалий фенотипа.
Вышеуказанные результаты показали, что клетки OSK+H1foo-ИПСК обладают высокой плюрипотентностью.
[0109] 3. Способ
[Получение человеческих клеток ИПСК и способ их культивирования]
Человеческие клетки ИПСК были получены в соответствии с протоколом, описанным в литературе (Т. Секи, С. Юаса, К. Фукуда, «Получение индуцированных плюрипотентных стволовых клеток из небольшого количества человеческой периферической крови с использованием сочетания активированных Т-клеток и вируса Сендай», «Nature Protocols», №7, 718-728, 2012 г). Клетки ИПСК (клетки OSKL+H1foo-ИПСК) были получены из лимфоцитов человеческой периферической крови или из фибробластов человеческой кожи с использованием вектора на основе вируса Сендай, содержащего человеческие гены Oct3/4, Sox2, Klf4, L-Myc и H1foo (нуклеотидная последовательность представлена в идентификаторе последовательности №1: ВС047943). В качестве контроля, клетки ИПСК (клетки OSKL-ИПСК) были получены с использованием гена Azami-Green вместо гена H1foo. Этот эксперимент был проведен в соответствии с руководством по генному рекомбинантному экспериментированию Университета Кэйо.
[0110] Линии человеческих клеток ИПСК были культивированы и хранились на культуральных планшетах, покрытых веществом iMatrix-511 (производитель - компания «Nippi, Incorporated») в культуральной среде StemFit AK02N (производитель - компания «Ajinomoto Co., Inc.») (далее - «культуральная среда человеческих ИПСК»). Культуральную среду человеческих ИПСК меняли каждые 2 дня, и клетки субкультивировали с использованием вещества StemPro Accutase (производитель - компания «Gibco») каждые 5-7 дней.
[0111] 4. Результаты
[Сравнение уровня экспрессии генов, отражающих хромосомные аберрации в клетках ИПСК]
Чтобы изучить нестабильность генома полученных человеческих клеток ИПСК посредством сравнения в соответствии со способом, раскрытым выше в разделе [Получение человеческих клеток ИПСК и способ для их культивирования], уровней экспрессии генов SRF и ACTG2 (Н. Ламм, Б. Керем, «Геномная нестабильность человеческих плюрипотентных клеток возникает из репликативного стресса и дефектов конденсации хромосомы», «Cell Stem Cell», №18 (2), 253-261, 2016 г.), уровни экспрессии которых снижаются, когда в клетке присутствует хромосомная аберрация, были проведены исследования с использованием сравнения на основе количественного анализа с использованием полимеразной цепной реакции с обратной транскрипцией (RT-PCR). Количественный RT-PCR-анализ был выполнен в соответствии со способом, раскрытым выше в разделе [Количественный RT-PCR-анализ], с использованием 4-х клонов клеток OSKL+H1foo-ИПСК и 4-х клонов клеток OSKL-ИПСК (контроль). На Фигуре 11а показаны результаты количественного RT-PCR-анализа на гене SRF, и на Фигуре 11b показаны результаты количественного RT-PCR-анализа на гене ACTG2.
[0112] Согласно результатам на Фигурах 11а и b, популяции клеток OSKL+H1foo-ИПСК также продемонстрировали значительно более высокий уровень экспрессии как гена SRF, так и гена ACTG2 по сравнению с (контрольной) популяцией клеток OSKL-ИПСК. Эти результаты показали, что использование гена H1foo в дополнение к веществу для перепрограммирования ядра, подавляет хромосомную аберрацию, которая может возникнуть при получении клеток ИПСК и позволяет получить популяцию клеток ИПСК более высокого качества.
[0113] [Сравнение уровня экспрессии маркера недифференцированного состояния клеток ИПСК]
Чтобы изучить, сохраняют ли клетки ИПСК, полученные в соответствии со способом, раскрытым выше в разделе [Получение человеческих ИПСК и способ их культивирования], стабильное недифференцированное состояние, был исследован уровень экспрессии гена Oct3/4 (маркер недифференцированного состояния стволовых клеток) посредством сравнения на основе количественного RT-PCR-анализа. Количественный RT-PCR-анализ был выполнен с использованием 4-х клонов клеток OSKL+H1foo-ИПСК и 4-х клонов OSKL-ИПСК (контроль) в соответствии со способом, раскрытым в разделе [Количественный ПЦРОТ-анализ] выше. На Фигуре 12 показаны результаты количественного RT-PCR-анализа.
[0114] Как видно из результатов, показанных на Фигуре 12, популяция клеток OSKL+H1foo-ИПСК обладает значительно меньшей дисперсией уровня экспрессии гена Oct3/4 (маркер недифференцированного состояния в стволовых клетках) среди клонов по сравнению с популяцией клеток OSKL-ИПСК (контроль). Результат показал, что использование гена H1foo в добавление к веществу для перепрограммирования ядра приводит к сохранению более стабильного недифференцированного состояния и позволяет получать популяции клеток ИПСК с меньшей дисперсией качества.
[0115] [Сравнение выживаемости клеток на раннем этапе после индукции дифференциации]
Чтобы изучить количество жизнеспособных клеток на раннем этапе после индукции дифференциации человеческих клеток ИПСК, полученных в соответствии со способом, раскрытым выше в разделе [Получение человеческих ИПСК и способ их культивирования], определенное число клеток (1×106 клеток на лунку) посеяли, и затем среду сменили с культуральной среды клеток ИПСК на RPMI-1640 (производитель - компания «Sigma Aldrich Co. LLC.») + культуральная среда В27 Supplement (производитель - компания «Gibco»), клетки собрали на следующий день, и количество жизнеспособных клеток рассчитали с использованием трипанового синего, с последующим сравнением. На Фигуре 13 показаны результаты такого расчета количества зародышевых клеток, выполненного для 4-х клонов клеток OSKL+H1foo-ИПСК и 4-х клонов клеток OSKL-ИПСК (контроль).
[0116] Согласно результатам на Фигуре 13, популяция клеток OSKL+H1foo-ИПСК содержала значительно более высокое число жизнеспособных клеток после индукции дифференциации, и обладала склонностью к более высокой адаптивности к среде для индукции дифференциации по сравнению с (контрольной) популяцией клеток OSKL-ИПСК. Результат показал, что использование гена H1foo в дополнение к веществу для перепрограммирования ядра приводит к большему числу жизнеспособных клеток после индукции дифференциации, и позволяет получать популяции клеток ИПСК более высокого качества за счет более высокой адаптивности к среде для индукции дифференциации.
[0117] [Сравнение остаточных количеств маркера недифференцированного состояния на раннем этапе после индукции дифференциации клеток ИПСК]
На раннем этапе после индукции дифференциации человеческих клеток ИПСК, полученных в соответствии со способом, раскрытым выше в разделе [Получение человеческих клеток ИПСК и способ их культивирования], сравнены остаточные количества гена Oct3/4 (маркер недифференцированного состояния стволовых клеток). В частности, определенное количество клеток (1×106 клеток на лунку) посеяли в лунки, затем среду заменили со среды для культивирования клеток ИПСК на культуральную среду для индукции дифференциации [DMEM F12 НАМ (производитель - компания «Sigma Aldrich Co. LLC.») + FBS (производитель - компания «Gibco») + GlutaMAX (производитель - компания «Gibco») + PenStrep (производитель - компания «Gibco») + 2-меркаптоэтанол (производитель - компания «Sigma Aldrich Co. LLC.»)], и клетки были собраны на 5-й день после смены культуральной среды для сравнения уровня экспрессии гена Oct3/4, являющегося маркером недифференцированного состояния, на основе RT-PCR-анализа. Количественный RT-PCR-анализ был выполнен в соответствии со способом, раскрытым выше в разделе [Количественный RT-PCR-анализ], с использованием 4-х клонов клеток OSKL+H1foo-ИПСК и 4-х клонов OSKL-ИПСК (контроль). На Фигуре 14 показаны результаты количественного RT-PCR-анализа.
[0118] Согласно результатам на Фигуре 14, (контрольная) популяция клеток OSKL-ИПСК обладала клонами (OSKL-ИПСК 2 и OSKL-ИПСК 4), в которых уровень маркера недифференцированного состояния (Oct3/4) остается на высоком уровне, несмотря на культивирование в культуральной среде для индукции дифференциации в течение 5 дней, в то время как популяция клетки OSKL+H1foo-ИПСК обладала клонами, каждый из которых экспрессировал маркер недифференцированного состояния на низком уровне, и дисперсия экспрессии маркера недифференцированного состояния среди клонов была значительно меньше. Результаты показали, что экспрессия маркера недифференцированного состояния в клетках ИПСК, полученных с использованием гена H1foo в добавление к веществу для перепрограммирования ядра быстро исчезает, когда клетки ИПСК культивируют в культуральной среде для индукции дифференциации, и дисперсия исчезновения - малая. Таким образом, продемонстрировано, что использование гена H1foo в добавление к веществу для перепрограммирования ядра позволяет получать популяции клеток ИПСК с меньшей дисперсией качества.
Промышленная применимость
[0119] В настоящем изобретении предложен агент для улучшения качества клеток ИПСК, способ получения клеток ИПСК, клетки ИПСК, полученные таким способом, и состав для получения клеток ИПСК. В соответствии с настоящим изобретением могут быть получены клетки ИПСК более высокого качества с меньшей дисперсией качества.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕРСОНАЛИЗИРОВАННОЕ ПОЛУЧЕНИЕ БИОПРЕПАРАТОВ И СПОСОБ ПЕРЕПРОГРАММИРОВАНИЯ СТВОЛОВЫХ КЛЕТОК | 2012 |
|
RU2595390C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2009 |
|
RU2399667C1 |
| СТИМУЛЯЦИЯ ПУТИ Wnt ПРИ ПЕРЕПРОГРАММИРОВАНИИ СОМАТИЧЕСКИХ КЛЕТОК | 2008 |
|
RU2492232C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ИЗ ФИБРОБЛАСТОВ ПАЦИЕНТОВ С БОЛЕЗНЬЮ ХАНГИНГТОНА | 2011 |
|
RU2458983C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ПАЦИЕНТОВ С СИНДРОМОМ ДАУНА | 2012 |
|
RU2492233C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПАЦИЕНТ (ДОНОР)-СПЕЦИФИЧЕСКИХ ФИБРОБЛАСТОПОДОБНЫХ КЛЕТОК ИЗ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2014 |
|
RU2568059C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pSN-ZsGreen, КОДИРУЮЩАЯ БЕЛКИ SOX2 И NANOG ЧЕЛОВЕКА И ФЛУОРЕСЦЕНТНЫЙ БЕЛОК ZsGreen, ПРЕДНАЗНАЧЕННАЯ ДЛЯ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2011 |
|
RU2495124C2 |
| ТРАНЗИЕНТНОЕ КЛЕТОЧНОЕ ПЕРЕПРОГРАММИРОВАНИЕ ДЛЯ ОБРАЩЕНИЯ СТАРЕНИЯ КЛЕТКИ | 2019 |
|
RU2801316C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНА СТВОЛОВЫХ КЛЕТОК, ПРИГОДНОГО ДЛЯ ИНДУЦИРОВАНИЯ ДИФФЕРЕНЦИРОВКИ В СОМАТИЧЕСКИЕ КЛЕТКИ | 2016 |
|
RU2730034C2 |
| СПОСОБ ПОВЫШЕНИЯ ЭКСПРЕССИИ РНК В КЛЕТКЕ | 2018 |
|
RU2784654C2 |

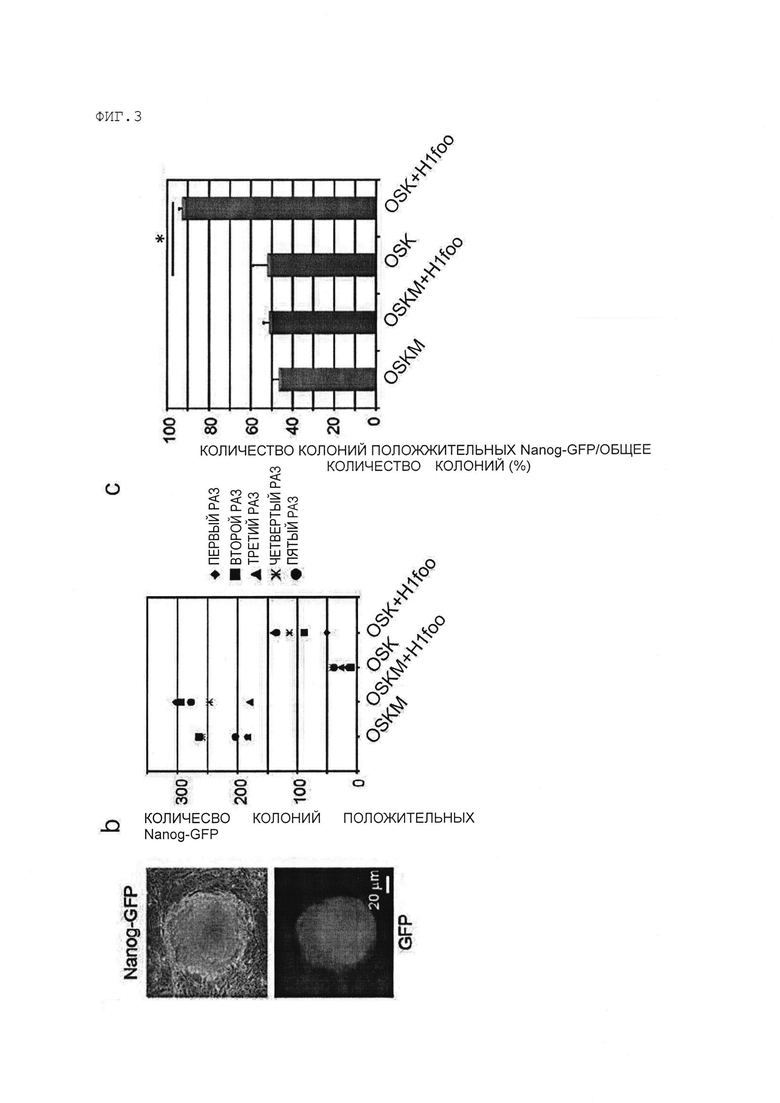
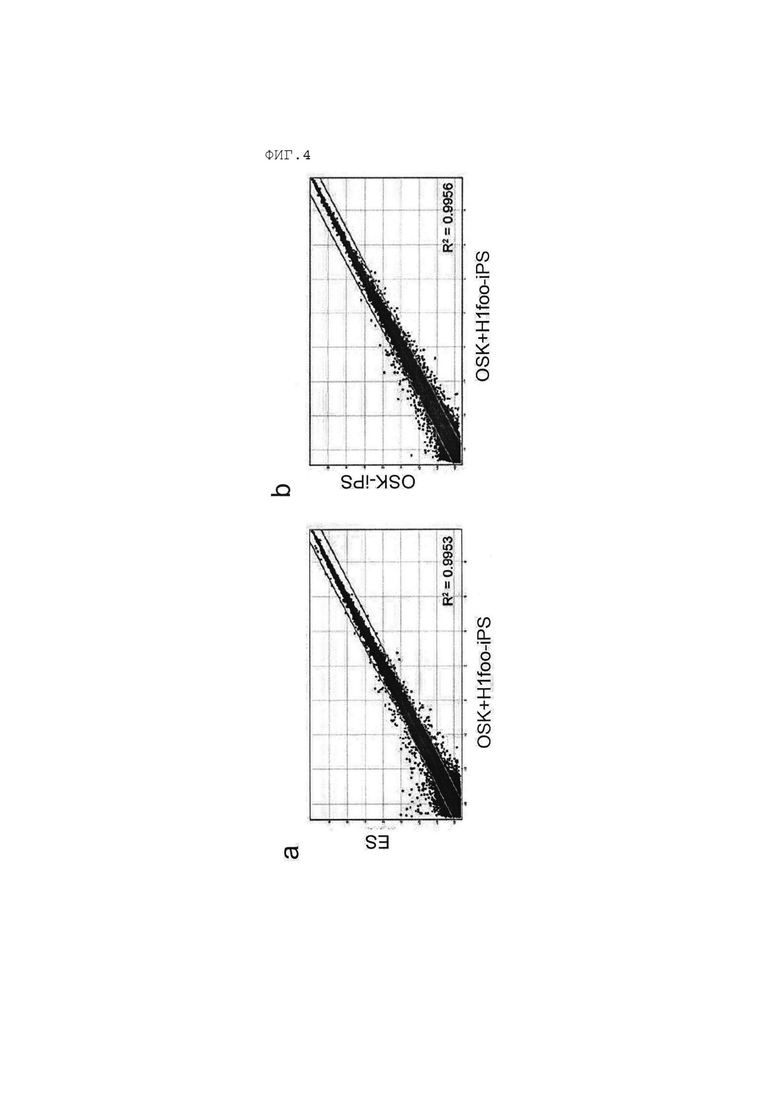
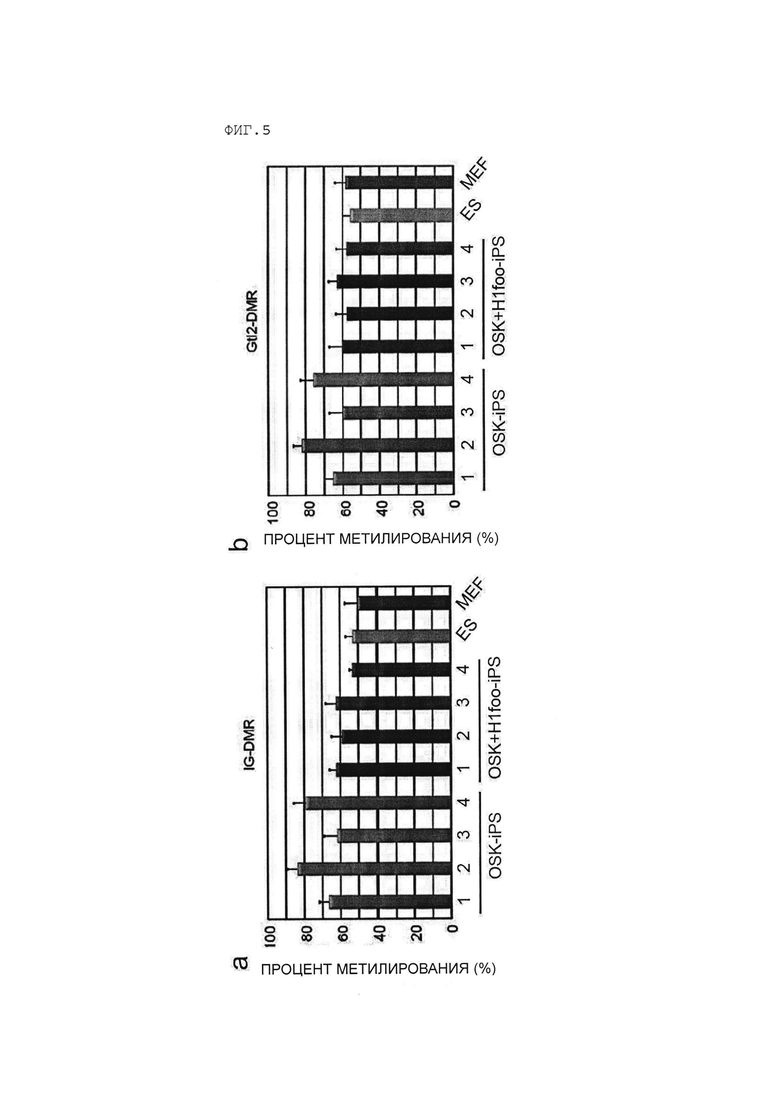
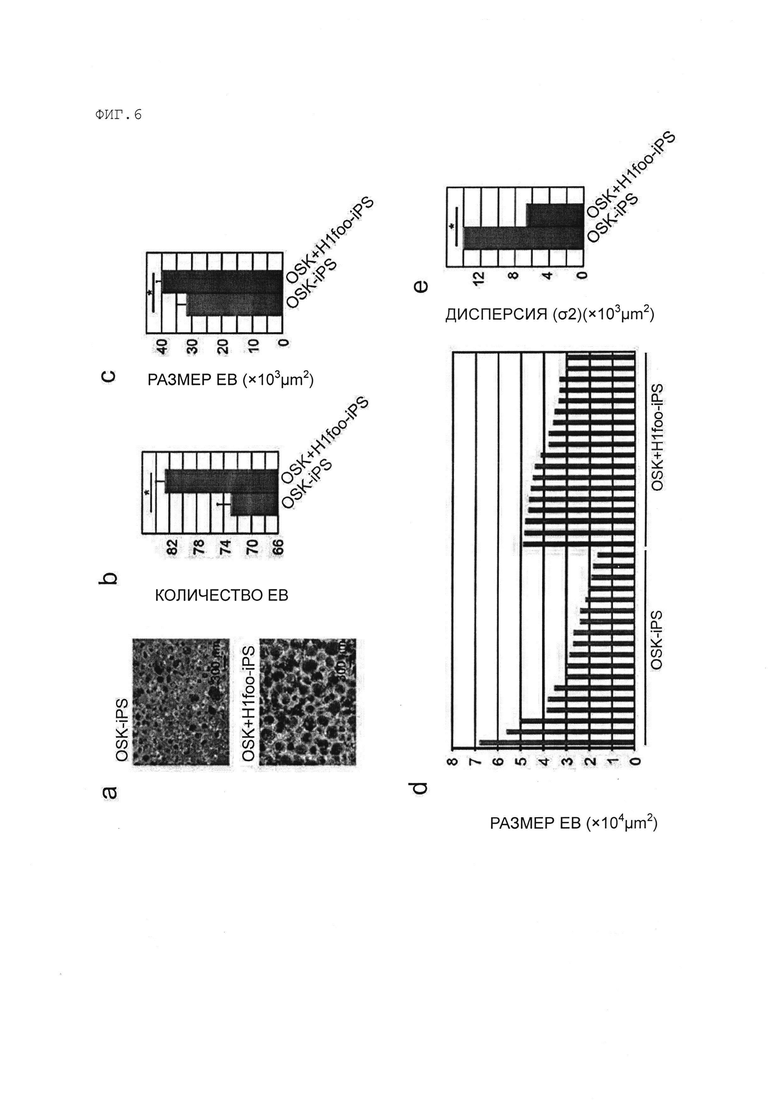

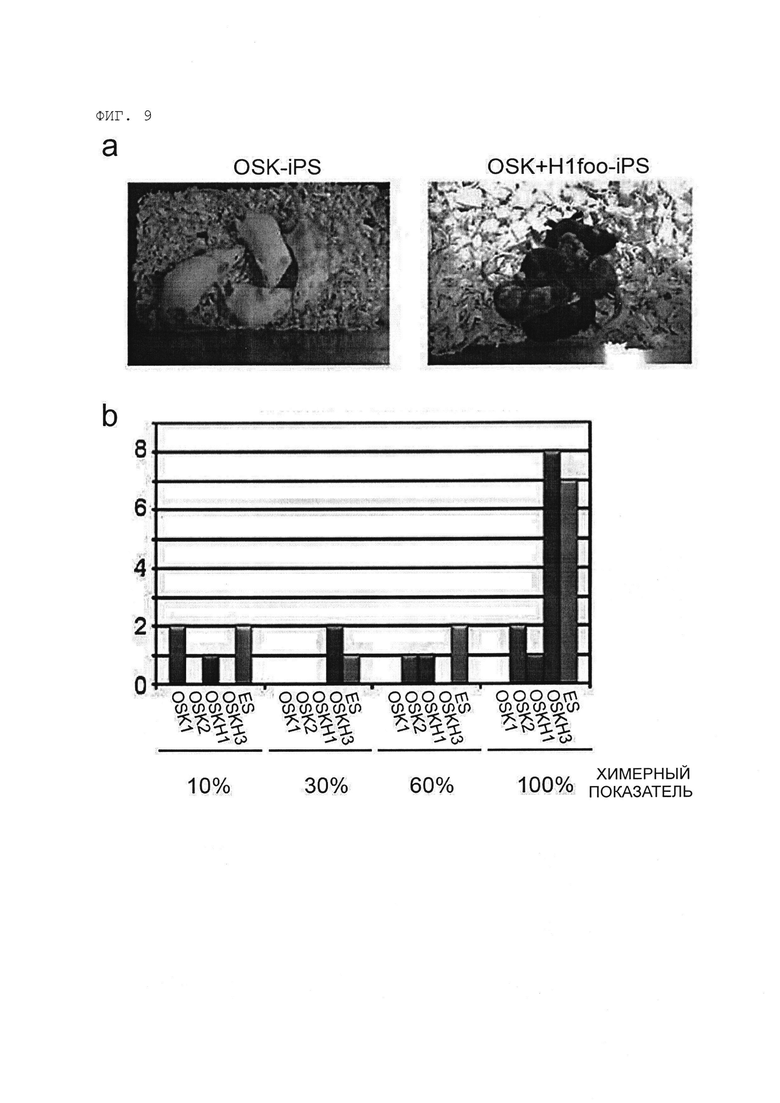
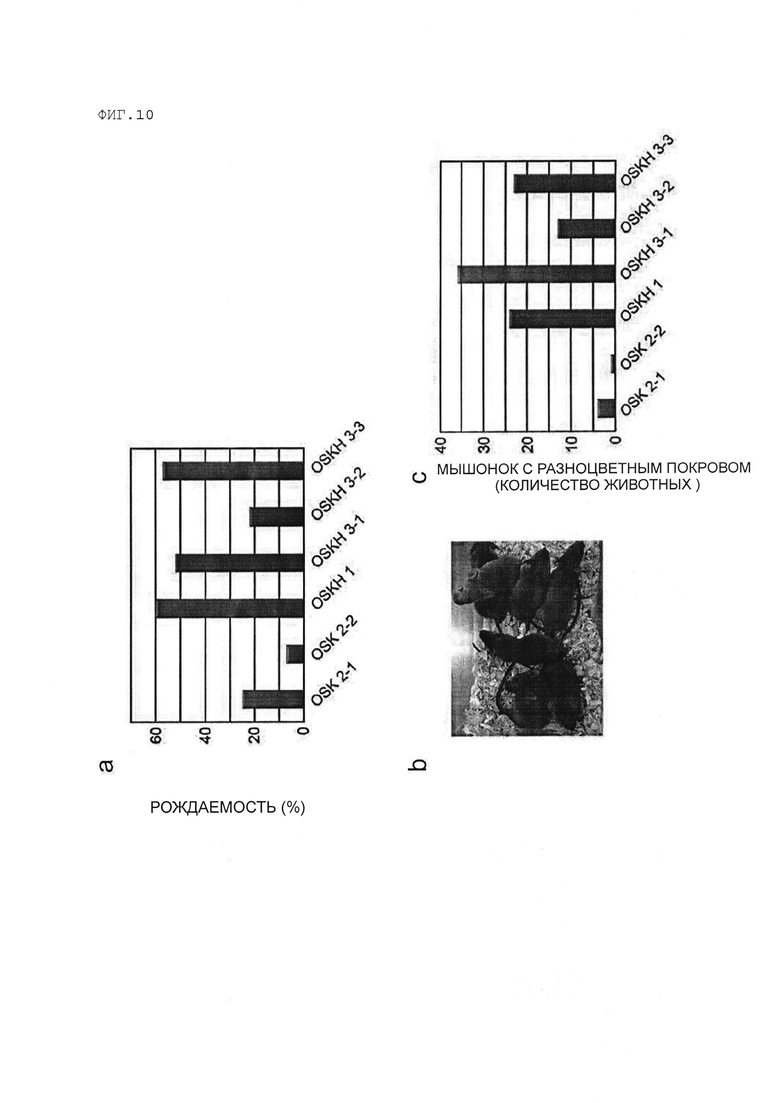
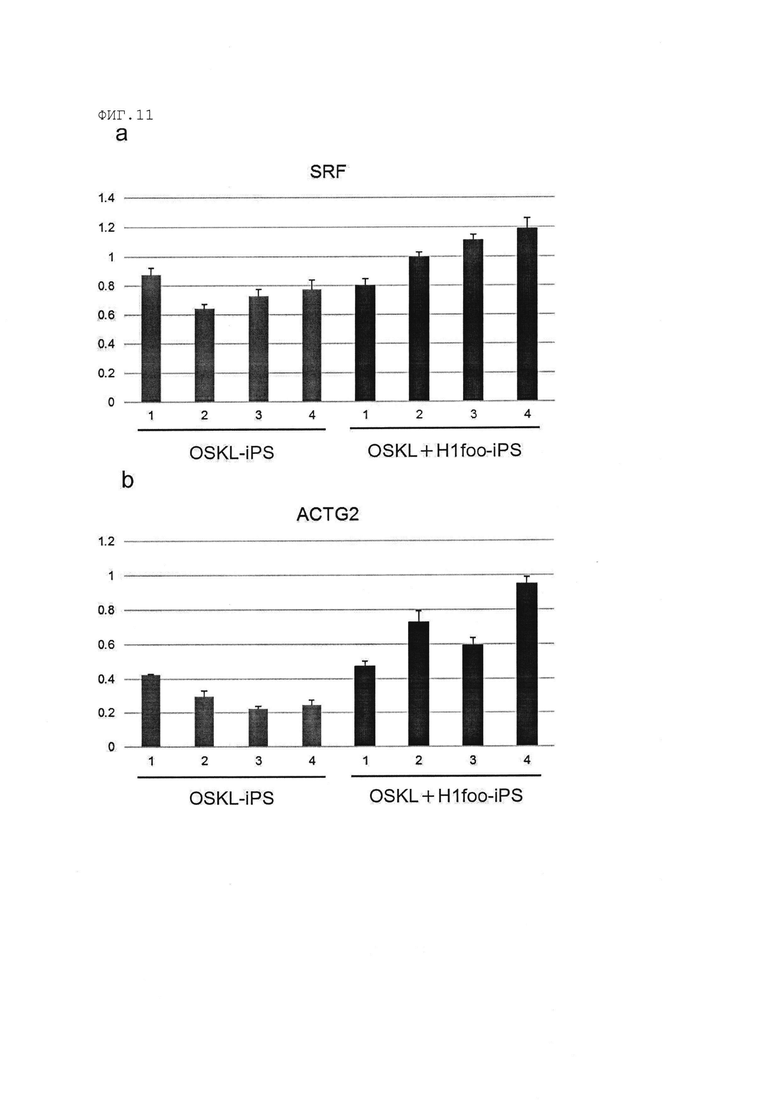
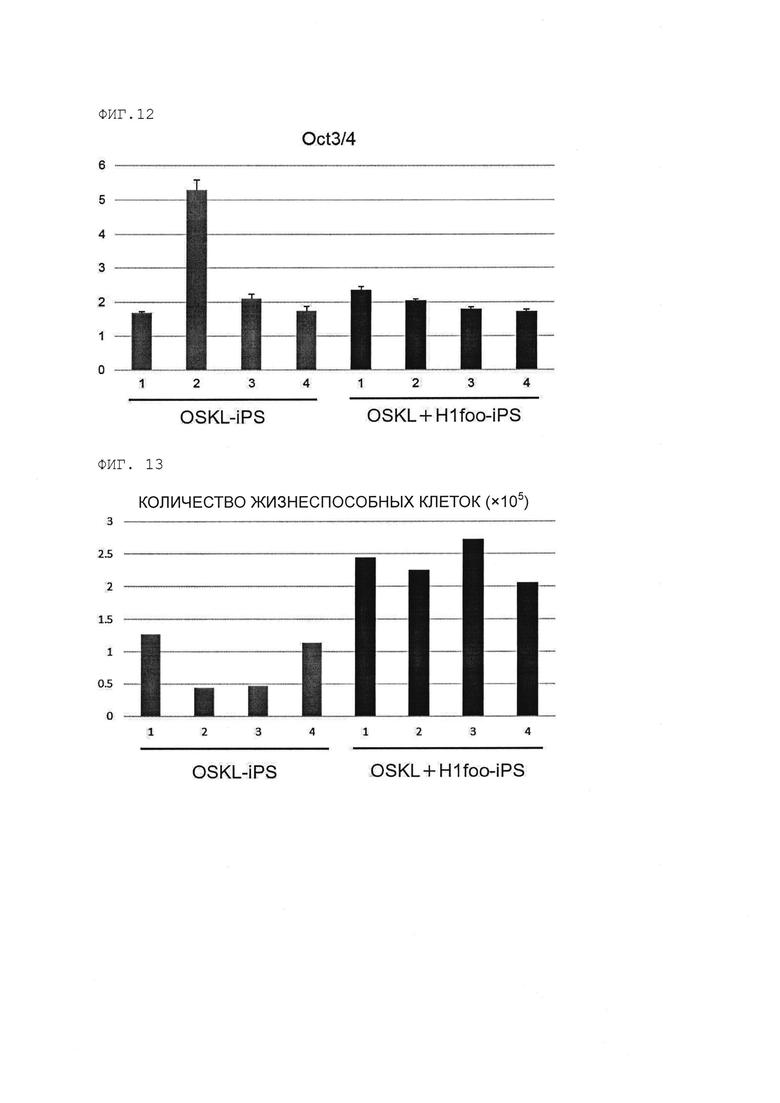
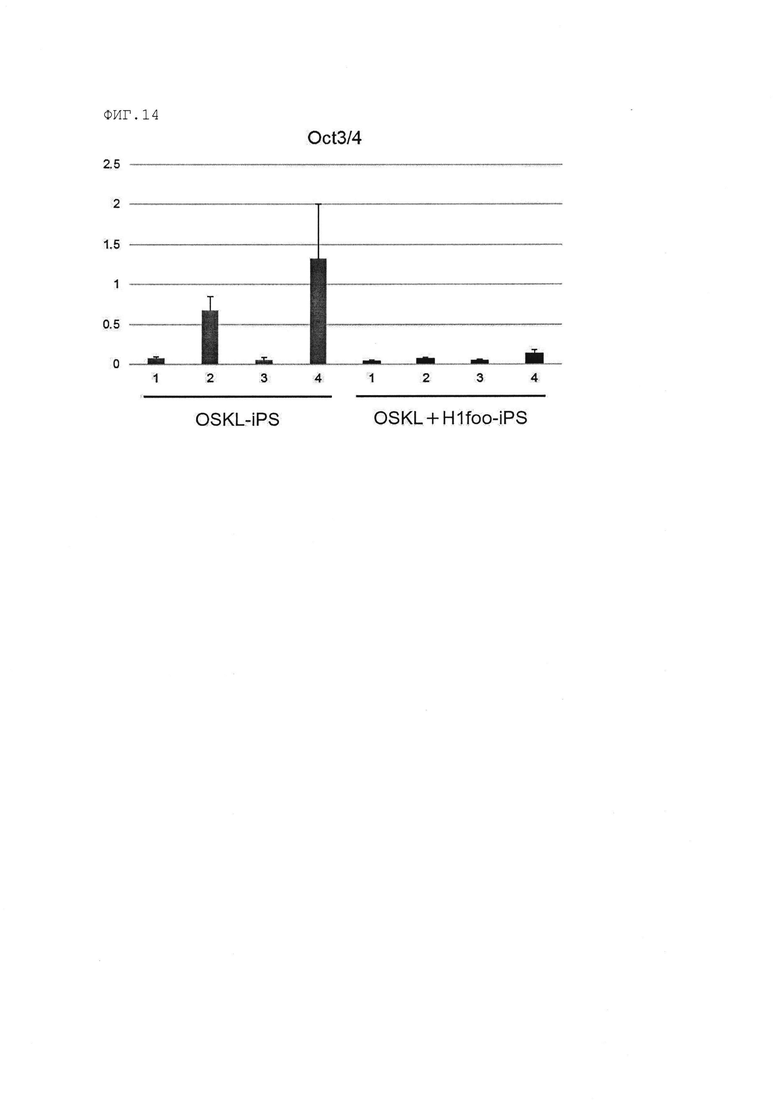
Изобретение относится к области биохимии, в частности к применению вещества, содержащего ген Н1foo, матричную РНК Н1foo или протеин Н1foo, для улучшения качества индуцированной плюрипотентной стволовой клетки. Также раскрыт способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества, содержащий шаг введения (а) вещества для перепрограммирования ядра и шаг введения (b) гена Н1foo или матричную РНК Н1foo или протеин Н1foo в соматическую клетку. Изобретение также относится к применению состава для получения индуцированной плюрипотентной стволовой клетки улучшенного качества, содержащего (а) вещество для перепрограммирования ядра и (b) ген Н1foo или матричную РНК Н1foo или протеин Н1foo. Изобретение позволяет эффективно получать индуцированную плюрипотентную стволовую клетку улучшенного качества. 3 н. и 5 з.п. ф-лы, 5 табл., 14 ил.
1. Применение вещества, содержащего ген Н1foo, матричную РНК Н1foo или протеин Н1foo, для улучшения качества индуцированной плюрипотентной стволовой клетки, при этом вещество является веществом, используемым для введения в соматическую клетку, когда индуцированную плюрипотентную стволовую клетку получают путем введения в соматическую клетку вещества для перепрограммирования ядра, причем улучшение качества индуцированной плюрипотентной стволовой клетки представляет собой два или более вариантов из следующих вариантов изменений с (1) по (11):
(1) увеличение процентного количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(2) увеличение количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(3) уменьшение дисперсии размера эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(4) увеличение процентного количества жизнеспособных клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(5) уменьшение процентного количества апоптических клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(6) увеличение уровня экспрессии гена Ki67 или гена PCNA при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(7) увеличение химерной компетенции индуцированных плюрипотентных стволовых клеток,
(8) увеличение потенциала трансмиссии зародышевой линии индуцированных плюрипотентных стволовых клеток,
(9) увеличение уровня экспрессии гена SRF в индуцированных плюрипотентных стволовых клетках,
(10) увеличение уровня экспрессии гена ACTG2 в индуцированных плюрипотентных стволовых клетках,
(11) увеличение дисперсии уровней экспрессии гена Oct3/4 в индуцированных плюрипотентных стволовых клетках.
2. Применение вещества по п. 1, при этом вещество содержит экспрессионный вектор, содержащий ген Н1foo.
3. Способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества, содержащий шаг введения (а) вещества для перепрограммирования ядра и шаг введения (b) гена Н1foo или матричную РНК Н1foo или протеин Н1foo в соматическую клетку, причем улучшение качества индуцированной плюрипотентной стволовой клетки представляет собой два или более вариантов из следующих вариантов изменений с (1) по (11):
(1) увеличение процентного количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(2) увеличение количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(3) уменьшение дисперсии размера эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(4) увеличение процентного количества жизнеспособных клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(5) уменьшение процентного количества апоптических клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(6) увеличение уровня экспрессии гена Ki67 или гена PCNA при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(7) увеличение химерной компетенции индуцированных плюрипотентных стволовых клеток,
(8) увеличение потенциала трансмиссии зародышевой линии индуцированных плюрипотентных стволовых клеток,
(9) увеличение уровня экспрессии гена SRF в индуцированных плюрипотентных стволовых клетках,
(10) увеличение уровня экспрессии гена ACTG2 в индуцированных плюрипотентных стволовых клетках,
(11) увеличение дисперсии уровней экспрессии гена Oct3/4 в индуцированных плюрипотентных стволовых клетках.
4. Способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества по п. 3, в котором вещество для перепрограммирования ядра содержит по меньшей мере один ген, выбранный из группы, содержащей ген из семейства генов Oct, ген из семейства генов Sox, ген из семейства генов Klf, ген из семейства генов Myc, ген из семейства генов Lin, ген Nanog и продукты таких генов.
5. Способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества по п. 3 или 4, в котором вещество для перепрограммирования ядра содержит ген из семейства генов Oct или продукт этого гена, ген из семейства генов Sox или продукт этого гена и ген из семейства Klf или продукт этого гена.
6. Способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества по любому из пп. 3-5, в котором вещество для перепрограммирования ядра содержит ген Oct3/4 или продукт этого гена, ген Sox2 или продукт этого гена и ген Klf4 или продукт этого гена.
7. Способ получения индуцированной плюрипотентной стволовой клетки улучшенного качества по любому из пп. 3-5, в котором вещество для перепрограммирования ядра состоит из гена Oct3/4 или продукта этого гена, гена Sox2 или продукта этого гена, гена Klf4 или продукта этого гена и гена L-Myc или продукта этого гена.
8. Применение состава для получения индуцированной плюрипотентной стволовой клетки улучшенного качества, содержащего (а) вещество для перепрограммирования ядра и (b) ген Н1foo или матричную РНК Н1foo или протеин Н1foo, при этом этот состав представляет собой состав, используемый путем введения в соматическую клетку при получении индуцированной плюрипотентной стволовой клетки, причем улучшение качества индуцированной плюрипотентной стволовой клетки представляет собой два или более вариантов из следующих вариантов изменений с (1) по (11):
(1) увеличение процентного количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(2) увеличение количества эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(3) уменьшение дисперсии размера эмбриоидных телец при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(4) увеличение процентного количества жизнеспособных клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(5) уменьшение процентного количества апоптических клеток при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(6) увеличение уровня экспрессии гена Ki67 или гена PCNA при культивировании индуцированных плюрипотентных стволовых клеток в культуральной среде для получения эмбриоидных телец,
(7) увеличение химерной компетенции индуцированных плюрипотентных стволовых клеток,
(8) увеличение потенциала трансмиссии зародышевой линии индуцированных плюрипотентных стволовых клеток,
(9) увеличение уровня экспрессии гена SRF в индуцированных плюрипотентных стволовых клетках,
(10) увеличение уровня экспрессии гена ACTG2 в индуцированных плюрипотентных стволовых клетках,
(11) увеличение дисперсии уровней экспрессии гена Oct3/4 в индуцированных плюрипотентных стволовых клетках.
| AWE J | |||
| P | |||
| et al., Identifying candidate oocyte reprogramming factors using cross-species global transcriptional analysis, CELLULAR REPROGRAMMING, vol | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Ударно-вращательная врубовая машина | 1922 |
|
SU126A1 |
| HAYAKAWA K | |||
| et al., Oocyte-specific linker histone H1foo is an epigenomic modulator that decondenses chromatin and impairs pluripotency, EPIGENETICS,vol | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |

