Родственные заявки
Настоящая заявка является непредварительной заявкой, поданной в соответствии с 37 CFR 1.53 (b)(1), по которой заявляется приоритет в соответствии с 35 USC 119(e) по предварительной заявке номер 60/671 610, поданной 15 апреля 2005 года, содержание которой полностью включено в настоящее описание в качестве ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение относится, в основном, к области молекулярной биологии и регуляции факторами роста. Более конкретно настоящее изобретение относится к модуляторам сигнального пути HGF/c-met и к использованию указанных модуляторов.
Предпосылки создания изобретения
Фактор роста гепатоцитов (HGF), также известный как скаттер-фактор, или фактор рассеивания (SF), представляет собой лиганд для Met (Bottaro et al., 1991), рецептора тирозинкиназы, который кодируется протоонкогеном c-met (Cooper et al., 1984a & b). Связывание HGF с Met индуцирует фосфорилирование внутриклеточного домена киназы, что приводит к активации сложного комплекса внутриклеточных путей, а это, в свою очередь, ведет к росту, дифференцировке и миграции клеток разных типов; в ряде обзоров последних лет дается исчерпывающее описание этих процессов (Birchmeier et al., 2003; Trusolino and Comoglio, 2002; Maulik et al., 2002). Было показано, что сигнальный путь HGF/Met, имеющий фундаментальное значение в эмбриональном развитии и регенерации тканей, вовлекается также в инвазивный опухолевый рост и метастазирование и, в качестве такового, представляет собой интересную терапевтическую мишень (Birchmeier et al., 2003; Trusolino and Comoglio, 2002; Danilkovich-Miagkova and Zbar, 2002; Ma et al., 2003).
HGF принадлежит к семейству плазминоген-родственного фактора роста и включает α-цепь размером 69 кДа, содержащую N-концевой фингер-домен (N) и четыре крингл-домена (Kringle; K1-K4), и β-цепь размером 34 кДа, которая обладает большим сходством с доменами протеазы химотрипсин-подобных сериновых протеаз из семейства Clan PA(S)/FamilyS1 (Nakamura et al., 1989; Donate et al., 1994; Rawlings et al., 2002). Подобно плазминогену и другим зимогенам сериновых протеаз HGF секретируется в виде одноцепочечного предшественника (scHGF). Далее scHGF связывается с гепаран-сульфатными протеогликанами, такими как синдекан-1 (Derksen et al., 2002), на клеточных поверхностях или во внеклеточном матриксе. Гепаран-сульфатные протеогликаны связываются с N-доменом (Hartmann et al., 1998), который вносит определенный вклад в высокую аффинность связывания Met с аминокислотами, расположенными в K1 (Lokker et al., 1994). Хотя scHGF способен связываться с Met с высокой афинностью, он не может активировать рецептор (Lokker et al., 1992; Hartmann et al., 1992). Приобретение HGF сигнальной активности происходит при протеолитическом расщеплении (активации) scHGF в положении Arg494-Val495, что приводит к образованию зрелого HGF, гетеромера α/β, связанного дисульфидной связью (Lokker et al., 1992; Hartmann et al., 1992; Naldini et al., 1992). Протеазоподобный домен HGF (β-цепь HGF) не обладает каталитической активностью, поскольку он не содержит требуемую каталитическую триаду Asp[c102]-His[c57]-Ser[c195] (в тексте всего описания используется стандартная нумерация химотрипсиногена с включением скобок), обнаруженную во всех сериновых протеазах (Perona and Craik, 1995; Hedstrom, 2002), включая Gln534[c57] и Tyr673[c195].
В связи с важностью сигнальной активности HGF данный процесс должен жестко контролироваться HGF-превращающими ферментами и их соответствующими физиологическими ингибиторами. Активация scHGF опосредована in vitro химотрипсин-подобными сериновыми протеазами, включающими активатор фактора роста гепатоцитов (HGFA) (Miyazawa et al., 1993), матриптазу/MT-SP1 (Takеuchi et al., 1999; Lin et al., 1999), активатор плазминогена урокиназного типа (Naldini et al., 1992), фактор XIIa (Shimomura et al., 1995), фактор XIa (Peek et al., 2002) и калликреин плазмы (Peek et al., 2002). Как и scHGF, указанные протеазы продуцируются в виде неактивных предшественников; их ферментативная активность также жестко регулируется другими активирующими протеазами и ингибиторами обоих типов, как Куниц-типа (Kunitz), так и серпинового типа.
Сериновые протеазы и процесс их активации описаны в литературе (Donate et al., 1994). При активации сериновых протеаз происходит расщепление зимогена, с последующей конформационной перегруппировкой так называемого «активирующего домена», что приводит к образованию соответствующим образом сформированного активного сайта и области взаимодействия субстрат/ингибитор. Активирующий домен включает три поверхностных петли, обозначенные как [c140]-, [c180]- и [c220]-петли, и содержит инсерцию из вновь сформированного N-конца в гидрофобном кармане (Huber and Bode, 1978). В гомологичной паре лиганд/рецептор для белка, стимулирующего макрофаги (MSP)/Ron), β-цепь MSP, подобного сериновой протеазе, обеспечивает основную энергию для связывания с рецептором (Wang et al., 1997; Miller and Leonard, 1998). Указанный процесс представляет собой реверсию системы HGF/Met, где сайт связывания высокоаффинного рецептора с остатками Met находится в α-цепи HGF (Lokker et al., 1994; Okigaki et al., 1992).
Важность сигнальной оси HGF/Met в процессах клеточного функционирования в условиях нормы и в этиологии клинических нарушений указывает на ее возможную значимость при разработке высокоэффективных терапевтических средств, основанных на модуляции данной оси. Однако сложность указанного пути, частично из-за недостаточно изученного механизма взаимодействия HGF-HGF и HGF/Met, замедляет прогресс в этом направлении и подводит к необходимости разработки подходов, основанных на лучшем понимании механизма взаимодействия HGF-HGF и HGF/Met. Приведенное ниже описание отвечает данной потребности и обеспечивает другие преимущества.
Все цитируемые источники, в том числе патентные заявки и публикации, полностью включены в настоящее описание в качестве ссылок.
Описание изобретения
Фактор роста гепатоцитов (HGF), плазминоген-родственный фактор роста, связывается с рецептором тирозинкиназы Met (также называемым как С-Met, с-Met или с-met), которая вовлекается в процессы развития, регенерации тканей и инвазивного опухолевого роста. Сама β-цепь HGF типа сериновой протеазы связывается с Met. Кроме того, что происходит связывание с Met, не ясно, какие области и какие конкретные остатки в β-цепи HGF необходимы для осуществления соответствующей сигнальной функции по пути HGF/Met. Авторы полагают, что определенные области/положения в β-цепи вносят важный вклад в соответствующую функциональную активность HGF, тогда как указанный вклад может вовлекать, а может и не вовлекать, связывание β-цепи HGF со своим соответствующим рецептором. Приведенные в настоящем описании результаты подтверждают тот факт, что мутации N-концевого участка и/или участка димеризации β-цепи HGF могут нарушать биологическую функцию HGF/Met, при наличии или в отсутствие существенного нарушения связывания HGF (в частности, β-цепи HGF) с С-Met. В основном, но не обязательно, указанные мутации не вовлекают те положения, которые, как считается, содержат «активирующий домен» или «область активного сайта» HGF дикого типа.
Мутационный анализ, приведенный в настоящем описании, обеспечивает основу для разработки множества мутантов HGF, способных ингибировать взаимодействие HGF/HGF и HGF/с-met дикого типа по широкому спектру функций. Примеры таких мутантов приведены в настоящем описании. Указанные мутанты способны конкурировать с HGF дикого типа за связывание с c-met, демонстрируя при этом сниженную способность воздействовать на c-met-ассоциированные биологические функции. Это особенно важно в том случае, когда нежелательно полное или существенное ингибирование оси HGF/c-met; указанная возможность весьма существенна, поскольку HGF и c-met широко экспрессируются в нормальных клетках и тканях. Такого рода мутанты также могут использоваться как полезные терапевтические агенты для лечения патологических состояний, где желательно снижение, но не полное отсутствие биологической активности HGF/c-met. Способы и композиции согласно настоящему изобретению основаны, по меньшей мере частично, на тех фактах, которые были открыты авторами и которые ниже описаны более подробно.
В одном аспекте настоящее изобретение относится к молекуле-антагонисту HGF/С-Met, включающей мутант HGF, который содержит мутацию в N-концевом участке β-цепи HGF и/или в участке димеризации β-цепи HGF.
Мутация в N-концевом участке β-цепи HGF может представлять собой любую мутацию, которая нарушает вставку N-конца β-цепи в HGF-связывающий карман. В одном варианте полученный мутант по β-цепи HGF связывается с С-Met со сниженной связывающей аффинностью по сравнению с β-цепью HGF дикого типа. В одном варианте полученный мутант по β-цепи HGF связывается с С-Met с по существу эквивалентной аффинностью, как и в случае β-цепи HGF дикого типа. В одном варианте полученный полноразмерный HGF, содержащий мутированную β-цепь HGF, связывается с С-Met со сниженной связывающей аффинностью в сравнении с полноразмерным HGF дикого типа. В одном варианте полученный полноразмерный HGF, содержащий мутированную β-цепь HGF, связывается с С-Met с по существу эквивалентной аффинностью, как и полноразмерный HGF дикого типа. В одном варианте мутация находится в P1' положении (т.е. в положении 495[c16]) или в прилегающем положении, где указанная мутация приводит к получению расщепляемого HGF мутанта, и где N-конец β-цепи HGF не встраивается в активный сайт/карман связывания. Примеры неспособности встраиваться в активный сайт/карман связывания включают, без ограничения, конфигурации, в случае которых мутант является дефектным либо по одному, либо по обоим (i) гидрофобным взаимодействиям и (ii) по образованию солевого мостика, соединяющего N-конец с Asp672[c]194, то есть, где N-конец содержит мутацию, например включает положительно заряженный замещенный или встроенный аминокислотный остаток. В одном варианте сигнальная функция через данный мутант нарушается. В одном варианте мутация находится в одном или нескольких положениях P1', P2', Р3' и P4' или рядом с ними.
Мутация в домене димеризации β-цепи HGF может быть любой мутацией, которая, как ожидается, будет нарушать контактирование между двумя β-цепями HGF, так что димеризация двух цепей (и, таким образом, двух HGF молекул) нарушается. Возможность таких мутаций определяется аминокислотной структурой HGF комплексов, например, как описано в работе Kirchhofer et al., J. Biol. Сhem. (2004), 279(38):39915-24. Релевантные аминокислотные положения включают, без ограничения, приведенные в настоящем описании положения. В одном варианте полученный мутант HGF обладает сниженной способностью димеризоваться с другой β-цепью HGF. В одном варианте мутация в области димеризации β-цепи HGF существенно не нарушает связывания полученного HGF мутанта с С-Met.
Домен димеризации относится к участку β-цепи HGF, который взаимодействует с другой β-цепью HGF с образованием димера (например, в комплексе активации HGF/Met). При расщеплении proHGF β-цепь HGF подвергается конформационному изменению. N-концевой остаток 495 в β-цепи HGF образует солевой мостик с остатком Asp 672. В некоторых вариантах участок димеризации β-цепи HGF включает аминокислотные остатки, по существу состоит из аминокислотных остатков или состоит по меньшей мере из одного аминокислотного остатка (вплоть до всех аминокислотных остатков), соответствующих остаткам β-цепи HGF на участке от примерно 495 до примерно 502 аминокислоты [петля c140], включая Y619, T620, G621, [c180] петли, включающей участок 662-665, или их смеси. В одном варианте домен димеризации включает положения, локализованные вплотную/вблизи одного или нескольких положений, приведенных выше, и будет, таким образом, как предполагается, влиять на указанные одно или несколько положений. Например, в одном варианте домен димеризации может также включать положения 622 и 626.
В одном аспекте молекула антагониста HGF/Met согласно настоящему изобретению включает мутацию в N-концевом участке β-цепи HGF, где указанная мутация находится в положении V495, G498, R502 плюс T503 и/или D672. Мутация может быть представлена в любой форме, которая меняет первичную, вторичную и/или третичную структуру N-концевого участка β-цепи HGF. Например, в одном варианте мутация N-концевого участка β-цепи HGF представляет собой замещение, вставку и/или делецию, такие как V495G, V495A, G498I, G498P, G498V, R502del плюс T503del или D672N. В другом варианте мутация в N-концевом участке β-цепи HGF представляет собой делецию V495. Мутация, которая изменяет первичную, вторичную и/или третичную структуру N-концевого участка β-цепи HGF, может находиться также в положении любой аминокислоты, которая сама не находится в N-концевом участке β-цепи HGF. Например, мутация D672, которая устраняет образование солевого мостика (например, D672N) с N-концом β-цепи HGF, как ожидается, изменит первичную, вторичную и/или третичную структуру N-концевого участка β-цепи HGF. Таким образом, мутации в N-концевом участке β-цепи HGF и в участке димеризации β-цепи HGF необязательно должны быть взаимоисключающими. Например, как будет описано в настоящем тексте и как показано на фиг.1, мутации некоторых положений, как ожидается, будут влиять и на N-концевой домен, и на домен димеризации в β-цепи HGF.
В одном аспекте молекула-антагонист HGF/Met согласно настоящему изобретению включает мутацию в домене димеризации β-цепи HGF, где указанная мутация находится в положении N497, G498, P500 или рядом с T501 и R502 или R502. Мутация может иметь любую форму, которая изменяет первичную, вторичную и/или третичную структуру участка димеризации β-цепи HGF. Примеры мутаций, которые меняют структуру участка димеризации β-цепи HGF, включают мутации, вводящие заряженный остаток или остаток с крупной боковой цепью (например, объемную), в последовательность дикого типа, где заряженный остаток может приводить к взаимодействиям по типу отталкивания, а крупная боковая цепь может сказываться на возникновении неблагоприятных стерических взаимодействий. Кроме того, цистеиновые мутации (например, L622C, I664C, P500C и N497C) также могут быть введены таким образом, чтобы они стали доступными для модификации специфическими тиол-алкилирующими реагентами, такими как агенты, содержащие малеимидные и галогенацетильные группы. В одном варианте мутация в участке димеризации β-цепи HGF представляет собой замещение, инсерцию и/или делецию, такую как N497R или K; G498A или S; P500W, H или E; инсерцию между T501 и R502 (например, инсерцию R и/или S); или R502del. В одном варианте мутация в положении N497 не может быть N497F, A или E. В одном варианте мутация находится в одном или нескольких положениях на участке 495-503, где такая мутация может менять димеризацию β-цепи HGF и/или связывание с рецептором. В другом варианте мутации, которые затрагивают домен димеризации, могут быть объединены с мутацией в одном или нескольких положениях за пределами домена димеризации, например, это может быть мутация в сайте расщепления 494-495 или вблизи него. Например, в случае мутанта, который, как ожидается, будет нерасщепляемым (например, двойной мутант R494E:V495G) и который также содержит мутацию в домене димеризации, такой мутант будет тем не менее демонстрировать нарушенную биологическую функцию, даже если он подвергнется расщеплению in vivo.
В некоторых вариантах молекула-антагонист HGF/Met согласно настоящему изобретению будет включать аминокислоты дикого типа в положении 534, 578, 619, 673, 692, 693, 694, 695, 696, 699 и/или 702. В некоторых вариантах молекула-антагонист HGF/Met согласно настоящему изобретению включает мутации в положении L622 (например, L622C или K); I623 (например, I623C); D626 (например, D626K); L622 плюс D626 (например, L622K плюс D626K); K663 (например, K663C); I664 (например, I664C); R502 (например, 502C); P500 (например, P500C); N497 (например, N497C); R494 плюс I623 (например, R494E плюс I623C); N497 плюс G498 (например, N497R плюс G498A или N497K плюс G498A); N497 плюс P500 (например, N497R плюс P500H или N497K плюс P500H); G498 плюс P500 (например, G498A плюс P500H); N497 плюс G498 плюс P500 (например, N497R плюс G498A плюс P500Н или N497K плюс G498A плюс P500H); N497 плюс L622 (например, N497R плюс L622K или N497K плюс L622K); N497 плюс D626 (например, N497R плюс D626K или N497K плюс D626K); N497 плюс L622 плюс D626 (например, N497R плюс L622K плюс D626K или N497K плюс L622K плюс D626K).
В одном варианте молекула-антагонист HGF/Met согласно настоящему изобретению включает мутацию в активном сайте HGF, одну или в сочетании с одной или несколькими мутациями, приведенными в настоящем описании. Мутации в активном сайте включают мутации в положении 667 и/или 704. Подходящие мутации включают замещение одного или обоих из этих положений на С или W.
В основном, молекула-антагонист HGF/Met согласно настоящему изобретению включает молекулу HGF, содержащую мутацию в β-цепи HGF, которая снижает одну или несколько биологических характеристик, в норме ассоциированных с HGF дикого типа. Например, в одном варианте молекула обладает сниженной С-Met сигнальной способностью (например, фосфорилированием Met) по сравнению с HGF дикого типа. В другом варианте молекула обладает сниженной способность стимулировать миграцию клеток по сравнению с HGF дикого типа. В другом варианте молекула обладает сниженной способностью стимулировать пролиферацию клеток по сравнению с HGF дикого типа. В другом варианте молекула обладает сниженной способностью стимулировать ангиогенез по сравнению с HGF дикого типа. Молекула-антагонист HGF/Met согласно настоящему изобретению в основном включает по меньшей мере часть цепи HGF, которая вовлечена в связывание с Met, присоединенную к мутированной β-цепи HGF, согласно настоящему описанию.
Как показано по результатам мутационного анализа, описанного в настоящем описании, некоторые участки β-цепи HGF, и в них конкретные положения аминокислот, играют важные роли в модуляции биологических функций HGF. Соответственно в одном аспекте настоящее изобретение относится к модуляторам HGF/Met, которые специфически направлены на указанные участки. Такие модуляторы включают нуклеиновые кислоты, такие как аптамеры, и полипептиды, такие как связывающие пептиды и антитела.
В контексте настоящего описания буква, стоящая перед числовым показателем, означает соответствующую аминокислоту дикого типа, имеющуюся в положении аминокислоты, обозначаемой данным номером, в полипептиде HGF человека дикого типа, и одна или несколько букв (если они присутствуют) после числового показателя указывают тип мутации/аминокислоту (например, замещение аминокислоты, делецию (del) или инсерцию (ins)).
В одном аспекте настоящее изобретение относится к мутанту HGF, который обладает модулирующей активностью в отношении HGF/c-met, например к антагонисту активности HGF/c-met или к варианту HGF, демонстрирующему снижение, но не отсутствие биологической активности HGF (например, активность, стимулирующая клеточный рост). В одном варианте антагонист согласно настоящему изобретению способен ингибировать биологическую активность HGF дикого типа in vivo или in vitro (такая биологическая активность включает, без ограничения, фосфорилирование рецептора, стимуляцию клеточной пролиферации, повышение выживаемости клеток, ускорение ангиогенеза, индукцию/усиление клеточной миграции). В одном варианте мутант HGF обеспечивает сниженную активность, способствующую клеточному росту (например, клеточную пролиферацию, выживаемость клеток, ангиогенез, клеточную миграцию).
В одном варианте молекула-антагонист согласно настоящему изобретению конкурирует с HGF дикого типа за связывание с Met. В некоторых вариантах указанная молекула ингибирует мультимеризацию рецептора c-met (например, димеризацию). В некоторых вариантах указанная молекула включает вариант (мутант) β-цепи, обладающий сниженной способностью к взаимодействию (например, к мультимеризации/димеризации) с β-цепью другой молекулы. В некоторых вариантах указанная молекула ингибирует мультимеризацию β-цепи HGF (например, димеризацию). В некоторых вариантах указанная молекула связывается с c-met, но демонстрирует сниженную способность воздействовать на активацию c-met (например, как выявлено по сниженному уровню фосфорилирования c-met, фосфорилирования митоген-активированной протеинкиназы (MAPK) и/или по снижению зависимой от HGF/c-met миграции клеток, клеточной пролиферации, по выживаемости клеток, морфогенезу клеток, ангиогенезу и т.п.).
В любой молекуле согласно настоящему изобретению, где одно или несколько положений было мутировано относительно соответствующей последовательности дикого типа, такая мутация может быть представлена в любой форме, которая изменяет функциональный эффект соответствующего остатка дикого типа. Мутация может быть получена посредством любого способа, известного в данной области (и/или может определена эмпирически), например, путем замещения, инсерции, добавления и/или делеции. В некоторых вариантах указанная мутация включает неконсервативное замещение. Подходящие замещения включают, без ограничения, замещения, приведенные в настоящем описании (в частности, в указанных ниже примерах), например, аминокислотами, такими как аланин или серин.
В одном аспекте молекулу/вещество (например, модуляторы HGF/c-met, приведенные в настоящем описании) связывают с токсином, таким как цитотоксический агент. Указанные молекулы/вещества могут быть получены в составе композиции или могут быть введены в сочетании с дополнительным/усиливающим агентом, таким как облучение и/или химиотерапевтический агент.
Настоящее изобретение также относится к способам и композициям, применяемым для модуляции патологический состояний, ассоциированных с нарушением регуляции сигнальной оси HGF/c-met. Таким образом, настоящее изобретение в одном аспекте относится к способу модуляции активации c-met у субъекта, включающему введение субъекту молекулы-антагониста HGF/c-met согласно настоящему изобретению, посредством чего достигается модуляция активации c-met. В одном варианте указанная молекула представляет собой антагонист HGF/c-met, который ингибирует активность HGF/c-met. В одном варианте указанный антагонист ингибирует специфическое связывание β-цепи HGF дикого типа с c-met. В одном аспекте настоящее изобретение относится к способу лечения патологического состояния, ассоциированного с активацией c-met у субъекта, включающему введение субъекту антагониста c-met согласно настоящему изобретению, в результате чего ингибируется активация c-met.
Сигнальный путь HGF/c-met вовлечен во множественные биологические и физиологические функции, включающие, например, стимуляцию клеточного роста (например, пролиферацию клеток, выживаемость клеток, миграцию клеток, клеточный морфогенез) и ангиогенез. Таким образом, в другом аспекте настоящее изобретение относится к способу ингибирования клеточного роста, активируемого с-met (например, пролиферации и/или выживаемости), где указанный способ включает контактирование клетки или ткани с антагонистом согласно настоящему изобретению, в результате которого достигается ингибирование клеточной пролиферации, ассоциированной с активацией c-met. В еще одном аспекте настоящее изобретение относится к способу ингибирования ангиогенеза, включающему введение в клетку, ткань и/или в организм субъекта с патологическим состоянием, ассоциированным с аномальным ангиогенезом, антагониста HGF/c-met согласно настоящему изобретению, в результате чего достигается ингибирование ангиогенеза.
В одном аспекте настоящее изобретение относится к использованию антагониста согласно настоящему изобретению при изготовлении лекарственного препарата для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом. Указанный антагонист может иметь любую приведенную в настоящем описании форму, включая форму антитела, фрагмента антитела, полипептида (например, олигопептида, мутанта/варианта полипептида HGF), нуклеиновой кислоты (аптамера) или их сочетания.
В одном аспекте настоящее изобретение относится к применению нуклеиновой кислоты согласно настоящему изобретению при изготовлении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом.
В одном аспекте настоящее изобретение относится к применению вектора экспрессии согласно настоящему изобретению при изготовлении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом.
В одном аспекте настоящее изобретение относится к применению клетки-хозяина согласно настоящему изобретению при изготовлении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом.
В одном аспекте настоящее изобретение относится к применению изделия согласно настоящему изобретению при изготовлении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом.
В одном аспекте настоящее изобретение относится к применению набора согласно настоящему изобретению при изготовлении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное нарушение, иммунное нарушение (такое как аутоиммунное нарушение) и/или нарушение, связанное с ангиогенезом.
В одном аспекте настоящее изобретение относится к способу ингибирования клеточной пролиферации, активируемой c-met, включающему контактирование клетки или ткани с эффективным количеством антагониста согласно настоящему изобретению, посредством чего достигается ингибирование клеточной пролиферации, ассоциированной с активацией c-met.
В одном аспекте настоящее изобретение относится к способу лечения патологического состояния, ассоциированного с нарушенной регуляцией c-met-активации у субъекта, где указанный способ включает введение указанному субъекту эффективного количества антагониста согласно настоящему изобретению, в результате чего достигается лечение указанного состояния.
В одном аспекте настоящее изобретение относится к способу ингибирования роста клетки, которая экспрессирует c-met или фактор роста гепатоцитов, или оба из них, где указанный способ включает контактирование указанной клетки с антагонистом c-met согласно настоящему изобретению, что приводит к ингибированию роста указанной клетки. В одном варианте указанную клетку приводят в контакт с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта).
В одном аспекте настоящее изобретение относится к способу терапевтического лечения млекопитающего, имеющего раковую опухоль, которая включает клетку, экспрессирующую c-met или фактор роста гепатоцитов, или оба из них, где указанный способ включает введение указанному млекопитающему эффективного количества антагониста согласно настоящему изобретению, что приводит к эффективному лечению указанного млекопитающего. В одном варианте указанную клетку приводят в контакт с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта).
В одном аспекте настоящее изобретение относится к способу лечения или профилактики клеточно-пролиферативного нарушения, ассоциированного с повышенной экспрессией или активностью c-met или фактора роста гепатоцитов, или их обоих, где указанный способ включает введение субъекту, при наличии необходимости в таком лечении, эффективного количества антагониста согласно настоящему изобретению, что приводит к эффективному лечению или предупреждению указанного клеточно-пролиферативного нарушения. В одном варианте указанное пролиферативное нарушение представляет собой рак.
В одном аспекте настоящее изобретение относится к способу ингибирования роста клетки, где рост указанной клетки, по меньшей мере частично, зависит от рост-потенцирующего эффекта c-met или фактора роста гепатоцитов, или их обоих, где указанный способ включает контактирование указанной клетки с эффективным количеством антагониста согласно настоящему изобретению, в результате чего достигается ингибирование роста указанной клетки. В одном варианте клетку приводят в контакт с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта).
В одном аспекте настоящее изобретение относится к способу терапевтического воздействия на опухоль млекопитающего, где рост указанной опухоли, по меньшей мере частично, зависит от рост-потенцирующего эффекта c-met или фактора роста гепатоцитов, или их обоих, где указанный способ включает контактирование указанной клетки с эффективным количеством антагониста согласно настоящему изобретению, в результате чего достигается эффективное лечение данной опухоли. В одном варианте клетку приводят в контакт с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта).
Способы согласно настоящему изобретению могут использоваться для воздействия на любое подходящее патологическое состояние, например, клетки и/или ткани, ассоциированные с нарушенной регуляцией HGF/c-met сигнального пути. В одном варианте клетка, которая является мишенью в способе согласно настоящему изобретению, представляет собой раковую клетку. Например, раковая клетка может представлять собой клетку, выбранную из группы, состоящей из: раковой клетки молочной железы, клетки колоректального рака, клетки рака легкого, клетки папиллярной карциномы (например, клетки щитовидной железы), клетки рака ободочной кишки, клетки рака поджелудочной железы, клетки рака яичника, цервикальной раковой клетки, клетки рака центральной нервной системы, клетки остеогенной саркомы, клетки карциномы почки, клетки гепатоцеллюлярной карциномы, клетки рака мочевого пузыря, клетки рака желудка, клетки плоскоклеточной карциномы головы и шеи, клетки меланомы, клетки множественной меланомы и лейкозной клетки. В одном варианте клетка, которая является мишенью в способе согласно настоящему изобретению, представляет собой геперпролиферирующую и/или гиперплазирующую клетку. В одном варианте клетка, которая является мишенью в способе согласно настоящему изобретению, представляет собой дипластическую клетку. В еще одном варианте клетка, которая представляет собой мишень в способе согласно настоящему изобретению, является метастазирующей клеткой.
Способы согласно настоящему изобретению могут также включать дополнительные стадии лечения. Например, в одном варианте способ также включает стадию, при которой клетку-мишень и/или ткань-мишень (например, раковую клетку) подвергают облучению или воздействию химиотерапевтического агента. В одном варианте молекулу-антагонист HGF/Met по настоящему изобретению вводят субъекту с одним или несколькими другими терапевтическими агентами, например с эрлотинибом (TARCEVA®), пеметрекседом (ALIMTA®), бевацизумабом (AVASTIN®), гефитинибом (IRESSA®), трастузумабом (HERCEPTIN®) и ритуксимабом (RITUXAN®). Введение терапевтических агентов при проведении комбинированной терапии может осуществляться в одновременном или в последовательном режиме.
Как приведено в настоящем описании, c-met-активация представляет собой важный биологический процесс, нарушение регуляции которого ведет к развитию множества патологических состояний. Соответственно в одном варианте осуществления способов согласно настоящему изобретению клетка, которая является мишенью (например, раковая клетка), представляет собой такую клетку, в которой активация c-met повышена по сравнению с нормальной клеткой ткани того же происхождения. В одном варианте способ согласно настоящему изобретению вызывает гибель клетки-мишени. Например, контактирование с антагонистом согласно настоящему изобретению может привести к неспособности клетки проводить сигнал по пути c-met, что приводит к гибели клетки.
Нарушение регуляции c-met-активации (и соответственно сигнальной функции) может происходить из-за множества клеточных изменений, включающих, например, сверхэкспрессию HGF (когнатных c-met-лигандов) и/или самого c-met. Соответственно в некоторых вариантах способ согласно настоящему изобретению включает воздействие на клетку, в тех условиях, когда c-met, или фактор роста гепатоцитов, или они оба экспрессируются в усиленных количествах данной клеткой (например, раковой клеткой) по сравнению с нормальной клеткой в ткани того же происхождения. Клетка, экспрессирующая c-met, может подвергаться регуляции под действием HGF, происходящего из множества источников, например, аутокринным или паракринным способом. Например, в одном варианте осуществления способов согласно настоящему изобретению клетку-мишень приводят в контакт/связывают с фактором роста гепатоцитов, экспрессируемым в другой клетке (например, посредством паракринного эффекта). Такая другая клетка может происходить из той же самой ткани или из ткани другого происхождения. В одном варианте клетку-мишень приводят в контакт/связывают с HGF, экспрессирумым самой клеткой-мишенью (например, посредством аутокринного эффекта/петли).
В некоторых вариантах антагонисты HGF/Met согласно настоящему изобретению включают мутанты HGF, которые содержат модификации, усиливающие их ингибирующий и/или терапевтический эффект (включая, например, повышенную аффинность, улучшенные фармакокинетические свойства (такие как период полувыведения, стабильность, скорость клиренса) сниженную токсичность для субъекта). Подходящие модификации такого рода включают, например, процессы гликозилирования, пэгилирования, замещения неприродной, но функционально эквивалентной аминокислотой, связующими группами и т.п. Такие модификации известны в данной области и могут быть также определены эмпирически, при необходимости.
В одном аспекте настоящее изобретение относится к композициям, включающим один или несколько антагонистов HGF/c-met согласно настоящему изобретению и носитель. В одном варианте носитель является фармацевтически приемлемым.
В одном аспекте изобретение относится к нуклеиновым кислотам, кодирующим антагонист HGF/c-met согласно настоящему изобретению. В одном варианте нуклеиновая кислота согласно настоящему изобретению кодирует антагонист HGF/c-met, который является полипептидом или включает полипептид (например, мутант/вариант HGF). В одном варианте нуклеиновая кислота согласно настоящему изобретению кодирует антагонист HGF/c-met, который является антителом или его фрагментом или включает антитело или его фрагмент.
В одном аспекте настоящее изобретение относится к векторам, включающим нуклеиновую кислоту согласно настоящему изобретению.
В одном аспекте настоящее изобретение относится к клеткам-хозяевам, включающим нуклеиновую кислоту или вектор согласно настоящему изобретению. Вектор может быть любого типа, например, он может быть рекомбинантным вектором, таким как вектор экспрессии. Может использоваться любой представитель из множества известных клеток-хозяев. В одном варианте клетка-хозяин представляет собой прокариотическую клетку, например клетку E. coli. В одном варианте клетка-хозяин представляет собой эукариотическую клетку, например клетку млекопитающего, такую как клетка яичника китайского хомяка (CHO).
В одном аспекте настоящее изобретение относится к способам получения антагониста HGF/c-met согласно настоящему изобретению. Например, настоящее изобретение относится к способу получения антагониста, который представляет собой антитело (или его фрагмент) или включает антитело (или его фрагмент), где указанный способ включает экспрессию в подходящей клетке-хозяине рекомбинантного вектора согласно настоящему изобретению, кодирующего указанное антитело (или его фрагмент), и выделение указанного антитела. В другом примере настоящее изобретение относится к способу получения антагониста HGF/c-met, который представляет собой полипептид или включает полипептид (такой как мутант/вариант HGF), где указанный способ включает экспрессию в соответствующей клетке-хозяине рекомбинантного вектора согласно настоящему изобретению, кодирующего указанный полипептид, и выделение указанного полипептида.
В одном аспекте настоящее изобретение относится к изделию, включающему контейнер, а также композицию, содержащуюся в данном контейнере, где указанная композиция включает один или несколько антагонистов HGF/c-met согласно настоящему изобретению. В одном варианте композиция включает нуклеиновую кислоту согласно настоящему изобретению. В одном варианте композиция, включающая антагонист HGF/c-met, также включает носитель, который в некоторых вариантах является фармацевтически приемлемым. В одном варианте изделие согласно настоящему изобретению включает инструкции по введению композиции субъекту.
В одном аспекте настоящее изобретение относится к набору, включающему первый контейнер, содержащий композицию, которая включает один или несколько антагонистов HGF/c-met согласно настоящему изобретению; и второй контейнер, содержащий буфер. В одном варианте буфер является фармацевтически приемлемым. В одном варианте композиция, включающая антагонист HGF/c-met, также включает носитель, который в некоторых вариантах является фармацевтически приемлемым. В одном варианте набор также содержит инструкции по введению композиции субъекту.
Краткое описание чертежей
Фиг.1 (А) иллюстрирует характеристики разных мутантов HGF. Приведены примеры мутантов по N-концевой инсерции в β-цепи HGF и мутантов по участку димеризации β-цепи HGF. Данные по «клеточной миграции» относятся к миграции клеток MDA-MB435 в присутствии полноразмерного HGF, содержащего указанную мутацию(мутации), где указанные данные выражены в виде процента относительно миграции в присутствии HGF дикого типа. Данные по «связыванию HGF β/MetIgG» относятся к связыванию β-цепи HGF (содержащей указанную мутацию(мутации) с MetIgG, где приведенные данные выражены в виде соотношений IC50 (для мутанта) к IC50 (для молекулы дикого типа), где указанные данные получены в конкурентном тесте на связывание. В приведенных данных "дикий тип" относится к мутанту С604S; мутанты HGF β также содержат данную мутацию. Следует отметить, что в приведенных материалах мутации указываются жирным шрифтом, если они, как ожидается, будут нарушать потенциальное взаимодействие по типу β-цепь - β-цепь, и мутации выделяются курсивом и подчеркиванием (жирный или нежирный шрифт), если они, как ожидается, будут нарушать инсерцию N-конца. Указанные предположения основаны на доминирующем эффекте, наблюдаемом или ожидаемом для соответствующей мутации, то есть определяются воздействием либо на взаимодействие по типу β-цепь - β-цепь, либо на способность N-конца β-цепи встраиваться в активный сайт/карман связывания. При этом любой практикующий специалист может легко определить, оказывает ли конкретная мутация один или оба из указанных эффектов, независимо от того, указана она в качестве таковой или нет, на фиг.1А. Например, в некоторых случаях мутация может влиять и на взаимодействие по типу β-цепь - β-цепь, и на инсерцию N-конца, или в некоторых случаях мутация, показанная на фиг.1А, которая, как ожидается, будет оказывать эффект на взаимодействие по типу β-цепь - β-цепь, будет также, как это эмпирически может быть продемонстрировано, влиять на инсерцию N-конца. Аналогично, величины явно «нарушенного» связывания Met указаны курсивом, а величины явно «нормального» связывания указаны жирным шрифтом, хотя степень «нарушения» и «нормальности» являются относительными понятиями.
(B) Связывание полноразмерного HGF, содержащего указанную мутацию (мутации) с Met, по результатом оценки в тесте на конкурентное связывание. Данные выражены в виде соотношения IC50 (для мутанта) к IC50 (для молекулы дикого типа).
(С) Ингибирование миграции клеток и пролиферация полноразмерным HGF, содержащим указанную мутацию (мутации). Уровень клеточной миграции и пролиферативной активности соответственно в присутствии мутантного HGF и HGF дикого типа (1 нМ - для оценки миграции в присутствии HGF дикого типа; 0,25 нМ - для оценки пролиферации, в присутствии HGF дикого типа) выражают в виде процента относительно активности, наблюдаемой в присутствии одного HGF дикого типа.
Фиг.2 (А) Показано ингибирование HGF-зависимого фосфорилирования Met в клетках А549 HGF мутантами; обозначение R494A:R494E относится к одноцепочечному HGF. Уровень фосфорилирования Met указан в виде RLU (относительные световые единицы). (B) Показано ингибирование HGF-зависимого фосфорилирования Met в клетках карциномы легкого А549 мутантами HGF. Уровень фосфорилирования Met указан в виде процента относительно контроля (который представляет собой количество, наблюдаемое в присутствии 0,5 нМ HGF дикого типа).
Фиг.3 (А) и (B) Фосфорилирование Met в клетках А549 в присутствии HGF дикого типа и мутанта HGF. Уровень фосфорилирования Met показан в виде процента относительно максимального фосфорилирования, наблюдаемого в присутствии HGF дикого типа для каждой соответствующей концентрации HGF дикого типа.
Фиг.4 Ангиогенная активность в присутствии мутанта HGF. Уровень ангиогенеза показан как количество проростков/гранул в присутствии мутантов HGF.
Способы осуществления настоящего изобретения
Настоящее изобретение относится к способам, композициям, наборам и изделиям, применяемым для модуляции сигнального пути HGF/c-met.
Ниже приведено детальное описание, относящееся к данным способам, композициям, наборам и изделиям.
Основные методики
В практике осуществления настоящего изобретения могут использоваться, если особо не указано иное, стандартные методы молекулярной биологии (включая рекомбинантные методики), микробиологии, клеточной биологии, биохимии и иммунологии, которые известны специалистам в данной области. Такие методики хорошо описаны в литературе, в частности, в таких работах как «Molecular Cloning: A Laboratory Manual», второе издание (Sambrook et al., 1989); «Oligonucleotide Synthesis» (M.J. Gait, ed., 1984); «Animal Cell Culture» (R.I. Freshney, ed., 1987); «Methods in Enzymology» (Academic Press, Inc); «Current Protocols in Molecular Biology» (F.M.Ausubel et al., eds., 1987, включая периодические обновления); «PCR: The Polymerase Chain Reaction», (Mullis et al., ed., 1994); «A Practical Guide to Molecular Сloning» (Perbal Bernard V., 1988).
Определения
Ссылки на названия аминокислот, приведенные в настоящем описании, относятся к принятым в данной области обозначениям, в виде одной или нескольких форм, которые используются взаимозаменяемо в данном тексте: (I) полные наименования (например, триптофан, серин, глицин и т.п.), (ii) трехбуквенные сокращения (например, Trp, Ser, Gly и т.п.), и (iii) однобуквенные обозначения (например, W, S, G и т.п.).
Термин «процент (%) идентичности по аминокислотной последовательности» применительно к пептидной или полипептидной последовательности определяется как процент аминокислотных остатков в предполагаемой последовательности, которые идентичны аминокислотным остаткам в конкретной пептидной или полипептидной последовательности после выравнивания последовательностей и введения, при необходимости, разрывов, для достижения максимального процента идентичности по последовательности, при этом любые консервативные замещения не рассматриваются как часть идентичности по последовательности. Выравнивание с целью определения процента идентичности по аминокислотной последовательности может быть проведено различными способами, которые известны специалистам в данной области, например, с использованием широко доступного компьютерного программного обеспечения, такого как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры, необходимые для определения уровня идентичности при выравнивании, включая любые соответствующие алгоритмы, требуемые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для целей настоящего изобретения % идентичности по аминокислотной последовательности определяют с использованием компьютерной программы для сравнения последовательностей ALIGN-2, описанной в патенте США № 6828146.
В контексте настоящего описания термины «пептид» и «полипептид» используются взаимозаменяемо, за исключением того, что термин «пептид» в основном относится к полипептиду, включающему менее 200 непрерывных аминокислот. Термин «пептид» в основном относится к непрерывной и относительно короткой последовательности аминокислот, соединенных пептидными связями. В типичном случае, но необязательно, пептид имеет длину примерно из 2-50 аминокислот, 4-40 аминокислот или 10-30 аминокислот.
Термин «вектор» в контексте настоящего описания используется применительно к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, к которой они присоединена. Один тип вектора представляет собой «плазмиду», где данный термин относится к кольцевой двуцепочечной петлевой ДНК, внутрь которой могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Еще один тип вектора представлен вирусным вектором, где дополнительные сегменты ДНК могут быть лигированы в состав вирусного генома. Некоторые векторы способны к автономной репликации в клетке-хозяине, внутрь которой они введены (например, бактериальные векторы, содержащие бактериальный репликатор, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, так что при этом достигается совместная репликация с геномом-хозяином. Кроме того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы рассматриваются в настоящем описании как «рекомбинантные векторы экспрессии» (или просто «рекомбинантные векторы»). Чаще всего векторы экспрессии, используемые в методиках рекомбинантных ДНК, имеют форму плазмид. В тексте настоящего описания термины «плазмида» и «вектор» могут использоваться взаимозаменяемо, поскольку плазмида представляет собой чаще всего используемую форму вектора.
Термины «полинуклеотид» или «нуклеиновая кислота», используемые здесь взаимозаменяемо, относятся к полимерам нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги, или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой или в ходе синтетической реакции. Полинуклеотид может включать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. В случае наличия такая модификация в структуре нуклеотидов может быть введена до или после сборки полимера. Последовательность нуклеотидов может быть прервана компонентами не нуклеотидной природы. Полинуклеотид может также быть модифицирован после синтеза, например, путем конъюгации с меткой. Другие типы модификаций включают, например, «кэпирование», замещение одного или нескольких природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, модификации под действием вводимых незаряженных связей (например, с использованием метилфосфонатов, фосфотриэфиров, фосфоамидатов, карбаматов и т.п.), заряженных связей (например, с использованием фосфоротиоатов, фосфородитиоатов и т.п.), а также агентов, содержащих присоединенные фрагменты, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, ply-L-лизин и т.п.), интеркаляторов (например, акридина, псоралена и т.п.), хелатирующих агентов (например, металлов, радиоактивных металлов, бора, окислительных металлов и т.п.), алкилирующих агентов, в частности, содержащих модифицированные связи (например, альфа-аномерных нуклеиновых кислот и т.п.), а также с использованием немодифицированных форм одного или нескольких полинуклеотидов. Кроме того, любая из гидроксильных групп, первоначально присутствующих в сахарах, может быть замещена, например, фосфонатными группами, фосфатными группами, защищенными с использованием стандартных защитных групп, или активирована для получения дополнительных связей с дополнительными нуклеотидами, или они могут быть конъюгированы с твердыми или полутвердыми подложками. 5'- и 3'-Концевые OH могут быть фосфорилированы или замещены аминами или органическими кэппинг-группами, содержащими от 1 до 20 атомов углерода. Другие гидроксилы могут быть также дериватизированы с использованием стандартных защитных групп. Полинуклеотиды могут также содержать аналоги рибозы или дезоксирибозы, которые в основном известны в данной области и включают, например, 2'-О-метил-, 2'-О-аллил-, 2'-фтор- или 2'-азидорибозу-, карбоциклические аналоги сахаров, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и безосновные нуклеозидные аналоги, такие как метилрибозид. Одна или несколько фосфодиэфирных связей могут быть замещены альтернативными связующими группами. Указанные альтернативные связующие группы включают, без ограничения, варианты, в которых фосфат замещен P(O)S(«тиоатом»), P(S)S («дитиоатом»), «(O)NR2» («амидатом»), P(O)R, P(O)OR', CO или CH2 («формацеталем»), где каждый из R или R' независимо обозначают H или замещенный или незамещенный алкил (1-20С), необязательно содержащий эфирную (-О-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралкил. Необязательно, чтобы все связи в полинуклеотиде были идентичными. Указанное выше описание относится ко всем полинуклеотидам, приведенным в настоящем описании, включая РНК и ДНК.
Термин «олигонуклеотид» в контексте настоящего описания в основном относится к коротким, в основном одноцепочечным, в основным синтетическим полинуклеотидам, которые в основном, но необязательно, имеют длину менее чем примерно 200 нуклеотидов. Термин «олигонуклеотид» и «полинуклеотид» не являются взаимоисключающими. Описания, приведенные выше применительно к полинуклеотидам, равным образом и полностью применимы к олигонуклеотидам.
Термин «фактор роста гепатоцитов» или «HGF» в контексте настоящего описания, если особо не указано иное, относится к любому нативному или вариантному (нативному или синтетическому) полипептиду HGF, который способен активировать сигнальный путь HGF/c-met в условиях, позволяющих осуществляться такому процессу. Термин «HGF дикого типа» в основном относится к полипептиду, который включает аминокислотную последовательность природного белка HGF. Термин «последовательность HGF дикого типа» в основном относится к аминокислотной последовательности, обнаруженной в природном HGF.
Фразы «существенно не нарушает», «не снижает в существенной мере», «по существу аналогичны» или «по существу эквивалентны» и их вариации в контексте настоящего описания обозначают достаточно высокий уровень сходства между двумя числовыми значениями, так что специалист в данной области будет рассматривать различие между двумя величинами как имеющее небольшую биологическую значимость или вообще не имеющее биологической значимости, применительно к биологической характеристике, измеряемой данными показателями. Различие между двумя значениями составляют предпочтительно менее чем примерно 50%, предпочтительно менее чем примерно 40%, предпочтительно менее чем примерно 30%, предпочтительно менее чем примерно 20%, и предпочтительно менее, чем примерно 10%. Примеры «двух числовых значений» включают, в частности, значение, ассоциированное с белком дикого типа, и значение, ассоциированное с мутантной формой указанного белка.
Термины «антитело» и «иммуноглобулин» используются в настоящем описании взаимозаменяемо в самом широком смысле и включают моноклональные антитела (например, полноразмерные антитела или интактные моноклональные антитела), поликлональные антитела, поливалентные антитела, полиспецифические антитела (например, биспецифические антитела, при условии, что они демонстрируют желательную биологическую активность) и могут также включать некоторые фрагменты антител (как ниже будет описано более подробно). Антитело может быть человеческим, гуманизированным и/или созревшим по аффинности.
«Фрагменты антитела» включают только часть интактного антитела, где указанная часть предпочтительно сохраняет по меньшей мере одну и предпочтительно большую часть функций или все функции, нормально ассоциированные с данной частью, когда она присутствует в интактном антителе. В одном варианте фрагмент антитела включает антиген-связывающий сайт интактного антитела и, таким образом, сохраняет способность связываться с антигеном. В другом варианте фрагмент антитела представляет собой, например, фрагмент, который включает одну Fc-область и сохраняет по меньшей мере одну из биологических функций, в норме ассоциированных с Fc-областью, когда этот фрагмент присутствует в интактном антителе, такую как FcRn-связывание, модуляция периода полувыведения антитела, функция ADCC и связывание комплемента. В одном варианте фрагмент антитела представляет собой моновалентное антитело, которое имеет период полувыведения in vivo, по существу аналогичный таковому для интактного антитела. Например, такой фрагмент антитела может включать антиген-связывающее плечо, присоединенное к последовательности Fc, которое способно придавать стабильность in vivo указанному фрагменту.
Термин «моноклональное антитело», используемый в тексте настоящего изобретения, относится к антителу, полученному из популяции по существу гомогенных антител, то есть индивидуальных антител, составляющих популяцию, которые характеризуются идентичностью, за исключением возможных природных мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела являются высокоспецифическими и направлены против одного антигена. Кроме того, в отличие от препаратов поликлональных антител, которые в типичном случае включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене.
Приведенные в настоящем описании моноклональные антитела конкретно включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителе, полученном из конкретного вида или принадлежащего к конкретному классу или подклассу антител, тогда как оставшиеся части одной или нескольких цепей идентичны или гомологичны соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они демонстрируют желательную биологическую активность (патент США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
«Гуманизированные» формы антител, отличных от человеческих (например, мышиных антител), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, отличного от человеческого. По большей части гуманизированные антитела являются человеческими иммуноглобулинами (реципиентное антитело), в которых остатки из гипервариабельного участка реципиента замещены остатками из гипервариабельного участка молекулы, отличной от человеческой (донорное антитело), такой как молекула мыши, крысы, кролика или примата, отличного от человека, имеющей желательную специфичность, аффинность и способность. В некоторых вариантах остатки каркасного участка (FR) человеческого иммуноглобулина замещаются соответствующими остатками из молекулы, отличной от человеческой. Кроме того, гуманизированные антитела могут включать остатки, которые не присутствуют ни в реципиентном антителе, ни в донорном антителе. Такие модификации создают с целью улучшения характеристик антитела. В основном, гуманизированное антитело включает по существу все из по меньшей мере одного и в типичном случае двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют гипервариабельным петлям в иммуноглобулине молекулы, отличной от человеческой, и все или по существу все FR представляют собой FR из последовательности человеческого иммуноглобулина. Гуманизированное антитело необязательно также включает по меньшей мере часть константной области иммуноглобулина (Fc), в типичном случае человеческого иммуноглобулина. Дополнительные детали описаны в Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). См. также следующие обзорные статьи и содержащиеся в них ссылки: Vaswani and Hamilton, Ann. Allegry, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
«Человеческое антитело» представляет собой такое антитело, которое обладает аминокислотной последовательностью, соответствующей таковой в антителе, продуцируемом в организме человека и/или получаемом с использованием любой из методик создания человеческих антител, приведенных в настоящем описании. Такое определение человеческого антитела специфически исключает гуманизированное антитело, включающее антиген-связывающие остатки из молекулы, отличной от человеческой.
Антитело «созревшее по аффинности» представляет собой такое антитело, которое содержит одно или несколько изменений в одном или нескольких участках CDR, которые приводят к повышению аффинности антитела для антигена в сравнению с исходным антителом, не содержащем такого изменения (изменений). Предпочтительные созревшие по аффинности антитела обладают аффинностями в диапазоне наномолярных или даже пикомолярных концентраций в отношении антигена-мишени.
Зрелые по аффинности антитела получают по процедурам, известным в данной области. Так, в Marks et al. Bio/Technology 10:779-783 (1992) описывается процесс созревания по аффинности путем перестановки VН и VL доменов. Может также использоваться случайный мутагенез CDR и/или остатков каркасной области, как описано в Barbas et al., Proc. Nat. Acad. Sci, USA 91:3809-3813 (1994); Schier et al., Gene 169:147-155 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995) и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
«Блокирующее» антитело или антитело-«антагонист» представляет собой такое антитело, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается. Предпочтительные блокирующие антитела или антитела-антагонисты по существу или полностью ингибируют биологическую активность антигена.
«Антитело-агонист» в контексте настоящего описания обозначает антитело, которое имитирует по меньшей мере одну из функциональных активностей полипептида, представляющего интерес.
«Нарушение» или «патологическое состояние» представляет собой любое состояние, для которого может быть полезно лечение с использованием вещества/молекулы или способа согласно настоящему изобретению. Такое состояние включает хронические и острые нарушения или заболевания, включающие те патологические состояния, при которых организм млекопитающего предрасположен к рассматриваемому нарушению. Неограничивающие примеры нарушений, подлежащих лечению согласно настоящему изобретению, включает злокачественные и доброкачественные опухоли или виды рака; нелейкозные и лимфоидные злокачественные новообразования; нейрональные, глиальные, астроцитальные, гипоталамусные и другие гландулярные, макрофагальные, эпителиальные, стромальные и бластоцельные нарушения; а также воспалительные, иммунологические, нейродегенеративные нарушения, нарушения, связанные с ангиогенезом, и нарушения, связанные с дефектами обмена в митохондриях или метаболических путей.
Термины «клеточно-пролиферативное нарушение» и «пролиферативное нарушение» относятся к нарушениям, которые ассоциированы с аномальной в некоторой степени пролиферацией клеток. В одном варианте клеточно-пролиферативное нарушение представляет собой рак.
Термин «опухоль» в контексте настоящего описания относится к росту и пролиферации всех клеток неоплазмы, независимо от того, имеет она злокачественный или доброкачественный характер, а также включает все предраковые и раковые клетки и ткани. Термины «рак», «раковый», «клеточно-пролиферативное нарушение», «пролиферативное нарушение» и «опухоль» взаимно не исключают друг друга в настоящем описании.
Термины «рак» и «раковый» относятся или описывают физиологическое состояние у млекопитающих, которое в типичном случае характеризуется нерегулируемым ростом/пролиферацией клеток. Примеры рака включают, без ограничения, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры таких видов рака включают множественную миелому, плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, гепатоцеллюлярный рак, рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнной железы, рак почки (например, карциному почечных клеток), рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, печеночную карциному и различные типы рака головы и шеи.
В контексте настоящего описания термин «лечение» относится к процедуре клинического воздействия, предпринимаемого с целью изменить естественный ход процесса у индивидуума или в клетках, подлежащих лечению, и может проводиться либо с профилактической целью, либо в ходе клинической патологии. Желательные эффекты лечения включают предупреждение появления или повторения заболевания, облегчение симптомов, снижение любых прямых или опосредованных патологических последствий заболевания, предупреждение метастазирования, снижение скорости прогрессирования заболевания, облегчение или ослабление болезненного состояния и ремиссию или улучшение прогноза. В некоторых вариантах антитела согласно настоящему изобретению используют для задержки развития заболевания или нарушения.
Термин «эффективное количество» относится к количеству, которое эффективно в тех дозировках и в течение того периода времени, которое необходимо для достижения желательного терапевтического или профилактического результата.
«Терапевтически эффективное количество» вещества/молекулы согласно настоящему изобретению, агониста или антагониста, может варьировать, в зависимости от ряда факторов, таких как природа болезненного состояния, возраст, пол и вес индивидуума, и от способности вещества/молекулы агониста или антагониста вызывать в организме индивидуума желательный ответ. Терапевтически эффективное количество также представляет собой количество, в случае которого любые токсические или другие вредные эффекты данных вещества/молекулы агониста или антагониста перевешиваются терапевтически благоприятными эффектами. «Профилактически эффективное количество» относится к тому количеству, которое эффективно, в соответствующих дозировках и в течение необходимого периода времени, для достижения желательного профилактического результата. В типичном случае, но необязательно, в случае использования профилактической дозы субъектами до начала заболевания или на его ранних стадиях, профилактически эффективное количество будет меньше, чем терапевтическое количество.
Термин «цитотоксический агент» в контексте настоящего описания относится к веществу, которое ингибирует или предотвращает определенную функцию клеток и/или вызывает деструкцию клеток. Данный термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические агенты, например метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как малые молекулы токсинов или энзиматически активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты, и различные противоопухолевые и противораковые агенты, приведенные далее в данном тексте. Другие цитотоксические агенты также описываются в настоящем тексте. Агент, уничтожающий опухолевые клетки, вызывает деструкцию опухолевых клеток.
«Химиотерапевтический агент» представляет собой химическое вещество, используемое при лечении рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включающие альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в особенности, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелецин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлоретамина; мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в особенности калихеамицин гамма II и калихеамицин омега II (см., например, Agnew, Chem Intl. Ed. Engl., 33:183-186 (1994)); динемицин, включая динемицин А; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарциностатиновый хромофор и родственные хромопротеину энедииновые антибиотические хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицин, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин ADRIAMYCIN® (включая морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловая кислота, ногаламицин, оливомицин, пепломицин, потфиромицин, пуромицин, келамицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадреналиновые агенты, такие как аминоглютетимид, митотан, трилостан; восполнители фолиевой кислоты, такие как фролиновая кислота; ацеглатон; альдофосфамидный гликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазикон; элфорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтансиноиды, такие как майтансин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновая кислота; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазикон; 2,2',2''-трихлортриэтиламин; трихотецены (в особенности, Т-2 токсин, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-C»); циклофосфамид; тиотепа; таксоиды, например, паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), полученная из альбумина композиция на основе наночастиц паклитаксела, не содержащая кремофор ABRAXANETM (American Pharmaceutical Partners, Schaumberg, Illinois) и доксетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлоранбуцил; гемцитабин GEMZAR®; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин NAVELBINE®; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; CPT-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; и фармацевтически приемлемые соли, кислоты или производные указанных выше веществ.
В определение приведенного выше термина «химиотерапевтический агент» включаются также антигормональные агенты, которые действуют в направлении регуляции или ингибирования действия гормонов на опухоли, такие как антиэстрогены и селективные модуляторы рецептора эстрогена (SERM), включающие, например, тамоксифен (включая тамоксифен NOLVADEX®), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен FARESTON; ингибиторы ароматазы, которые ингибируют фермент ароматазы, регулирующей продукцию эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглютетимид, ацетат мегестрола MEGASE®, эксеместан AROMASIN®; форместанин, фадрозол, ворозол RIVISOR®; летрозол FEMARA® и анастрозол ARIMIDEX®; и антиадрогены, такие как флутамид, нилутамид, бикалутамид, леупролид и гозерелин; а также троксацитабин (1,3-диоксолановый аналог цитозинового нуклеозида); антисмысловые олигонуклеотиды, в частности, те из них, которые ингибируют экспрессию генов сигнального пути, вовлекаемые в аберрантную клеточную пролиферацию, такие как, например, PKC-альфа, Ralf и H-Ras; рибозимы, такие как ингибитор экспрессии VEGF (например, рибозим ANGIOZIME®) и ингибиторы экспрессии HER2; вакцины, такие как вакцины, применяемые при генной терапии, например вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; PROLEUKIN® rIL-2; ингибитор топоизомеразы 1 LURTOTECAN®; ABARELIX® rmRH; и фармацевтически приемлемые соли, кислоты или производные указанных выше соединений.
Используемый здесь термин «агент, ингибирующий рост» относится к соединению или композиции, которые ингибируют рост клетки, в том случае, когда рост клетки зависит от активации HGF/c-met, in vitro или in vivo. Таким образом, агентом, ингибирующим рост, может быть любой агент, который снижает в существенной мере процент HGF/c-met-зависимых клеток в S-фазе. Примеры агентов, ингибирующих рост, включают агенты, которые блокируют прогрессию клеточного цикла (в точке, отличной от S-фазы), такие как агенты, которые индуцируют остановку на G1- и остановку на М-фазе. Классические блокаторы М-фазы включают алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Агенты, которые блокируют G1-фазу, также оказывают определенный эффект по блокировке S-фазы, и включают, например, ДНК-алкилирующие агенты, такие как тамоксифен, преднизон, дакарбазин, меклоретамин, цисплатин, метотрексат, 5-фторурацил и аrа-С. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, озаглавленной как «Cell cycle regulation, oncogenes, and antineoplastic drugs» от Murakami et al. (WB Saunders: Philadelphia, 1995, в особенности на стр.13). Таксаны (паклитаксел и доцетаксел) представляют собой противораковые лекарственные средства, полученные из дерева тис. Доцетаксел TAXOTERE®, Phone-Poulenc Rorer), получаемый из европейского тиса, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел ускоряют сборку микротрубочек из тубулиновых димеров и стабилизируют микротрубочки за счет предупреждения деполимеризации, что проводит к ингибированию митоза в клетках.
Доксорубицин представляет собой антрациклиновый антибиотик. Полное химическое название доксорубицина (8S-цис)-10-[(3-амино-2,3,6-тридезокси-α-L-ликсогексапиранозил)окси]-7,8,9,10-тетрагидро-6,8,11-тригидрокси-8-(гидроксиацетил)-1-метокси-5,12-нафтаценедион.
Антагонисты HGF/Met-пептиды/полипептиды (включая антитела)
Один аспект настоящего изобретения относится к выделенному пептиду/полипептиду и к модуляторам антител взаимодействия β-цепи - β-цепи HGF и взаимодействия HGF-Met. В одном варианте модуляторы (такие как пептиды/полипептиды и антитела) могут быть выделены из клеток или тканей, в качестве их источников, с использованием подходящей для этого системы очистки, включающей стандартные методики очистки белков. В другом варианте модуляторы получают в рамках методик рекомбинантных ДНК. В качестве альтернативы рекомбинантной экспрессии модуляторы могут быть синтезированы химически с использованием стандартных методик пептидного синтеза.
Молекулы-антагонисты HGF/Met согласно настоящему изобретению включают молекулы, проиллюстрированные на фиг.1. Настоящее изобретение также относится к мутантному или вариантному белку, в котором любой из остатков может быть изменен на соответствующие остатки из указанных пептидов/полипептидов, где все еще поддерживается характер кодирования пептида/полипептида, который сохраняет свою модулирующую активность. В одном варианте вариант антагониста пептида/полипептида обладает по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% идентичностью по аминокислотной последовательности с последовательностью эталонного антагониста пептида/полипептида. В основном, указанный вариант демонстрирует по существу ту же самую или более высокую связывающую аффинность, что и эталонный антагонист пептида/полипептида по связыванию, которая составляет, например, по меньшей мере 0,74Х, 0,8X, 0,9X, 1,0X, 1,25X или 1,5X от связывающей аффинности эталонного связующего пептида/полипептида/лиганда, при оценке результатов теста на связывание по стандартной количественной методике оценки в системе единиц/метрической системе, и при этом сохраняется желательная степень антагонистической активности.
В основном, варианты согласно настоящему изобретению включают те варианты, в которых остатки в конкретном положении последовательности заменены другими аминокислотами, и также включают возможность вставки дополнительного одного или нескольких остатков между двумя остатками исходного белка/пептида, а также возможность делеции одного или нескольких остатков из исходной последовательности или добавление одного или нескольких остатков к исходной последовательности. Любые аминокислотные замещения, инсерции или делеции входят в объем настоящего изобретения. В благоприятных условиях замещение представляют собой консервативное замещение, описанное в настоящем тексте.
«Выделенный» или «очищенный» пептид, полипептид, белок или биологически активный фрагмент отделяют и/или восстанавливают от/из компонентов его естественной среды. Контаминантные компоненты включают материалы, которые в типичном случае препятствует диагностическому или терапевтическому использованию полипептида, и могут включать ферменты, гормоны и другие белковые и небелковые материалы. Препараты, содержащие предпочтительно менее 30%, по сухому весу, нежелательного загрязняющего материала (контаминантов), предпочтительно менее 20%, 10% и предпочтительно менее 5% контаминантов, рассматриваются как по существу изолированные. Изолированные, полученные рекомбинантными способами пептиды/полипептиды или их биологически активная часть предпочтительно по существу свободны от культуральной среды, то есть культуральная среда составляет предпочтительно менее 20%, предпочтительно менее примерно 10% и предпочтительно менее примерно 5% от объема препарата пептида/полипептида. Примеры контаминантов включают осколки клеток, остатки культуральной среды и вещества, используемые и продуцируемые в ходе in vitro синтеза пептида/полипептида.
Консервативные замещения пептидов/полипептидов показаны в таблице А в столбце, озаглавленном «Предпочтительные замещения». Если такие замещения приводят к изменению биологической активности, то в продукты могут вводиться более существенные изменения, обозначенные как «Репрезентативные замещения» в таблице А, или описанные далее в тексте применительно к различным классам аминокислот, и далее такие продукты подвергают скринингу.
Выраженные модификации биологических свойств пептида/полипептида сопровождаются отбором замещений, которые существенно различаются по своему эффекту на подержание (а) структуры полипептидного скелета в области замещения, например, в виде пластинчатой или спиральной конформации, (b) заряженности или гидрофобности молекулы в целевом сайте или (с) объемности боковой цепи. Природные остатки подразделяются на группы на основании общих свойств их боковой цепи:
(1) гидрофобные остатки: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные остатки: cys, ser, thr;
(3) кислотные остатки: asp, glu;
(4) основные остатки: asn, gln, his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические остатки: trp, tyr, phe.
Неконсервативные замещения относятся к замене представителя одного из этих классов на представителя другого класса.
Варианты модуляторов антител, которые не оказывают существенного влияния на активность антитела, могут быть также созданы на основании известной информации. Например, варианты антител могут характеризоваться тем, что по меньшей мере один аминокислотный остаток в молекуле такого антитела замещен другим остатком. В случае антител сайты, представляющие наибольший интерес с точки зрения заместительного мутагенеза, в основном включают гипервариабельные участки, но также могут рассматриваться изменения в каркасной области (FR).
В случае антител один тип варианта по замещению включает замещение одного или нескольких остатков гипервариабельного участка в исходном антителе (например, в случае гуманизированного или человеческого антитела). В основном, получаемые один или несколько вариантов, отбираемые для дальнейшей разработки, обладают улучшенными биологическими свойствами относительно исходного антитела, из которого они были получены. Удобный способ создания таких вариантов по замещению включает способ «созревания» по аффинности с использованием методики фагового представления. Вкратце, метод состоит в том, что несколько сайтов в гипервариабельном участке (например, 6-7 сайтов) подвергают мутированию с созданием всех возможных аминокислотных замещений по каждому сайту. Полученные таким образом антитела анализируют с использованием частиц нитевидных фагов, представляющих слияние с продуктом гена III из М13, упакованных в каждой частице. Варианты с проявлением фага далее подвергают скринированию для оценки их биологической активности (например, связывающей аффинности), как будет описано далее в описании. С целью идентификации возможных сайтов гипервариабельной области, подходящих для модификации, может быть проведен мутагенез по методу аланинового сканирования для идентификации остатков гипервариабельного участка, оказывающих значимое воздействие на связывание с антигеном. Альтернативно или дополнительно, может быть полезно проанализировать кристаллическую структуру комплекса антиген-антитело для идентификации контактных точек между антителом и антигеном. Такие контактные остатки и прилегающие остатки являются кандидатами для замещения, в соответствии с методиками, приведенными в настоящем описании. При получении таких вариантов панель данных вариантов подвергают скринингу в соответствии с настоящим описанием, и антитела с улучшенными свойствами, продемонстрированными в одном или нескольких релевантных тестах, могут быть отобраны для дальнейшей разработки.
Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности антитела, получают с использованием множества известных в данной области методик. Такие методики включают, без ограничения, выделение из природного источника (в случае природных вариантов аминокислотной последовательности) или получение путем олигонуклеотидного мутагенеза, путем ПЦР-мутагенеза и кассетного мутагенеза ранее полученного варианта или невариантной версии антитела.
Может быть желательно встроить одну или несколько аминокислотных модификаций в Fc-участок иммуноглобулиновых полипептидов согласно настоящему изобретению с созданием варианта Fc-участка. Вариант Fc-участка может включать последовательность человеческого Fc-участка (например, участка человеческого IgG1, IgG2, IgG3 или IgG4), включающую аминокислотную модификацию (например, замещение) по одной или нескольким аминокислотным позициям, включая цистеин шарнирной области.
В одном варианте вариант Fc-участка может демонстрировать измененную связывающую аффинность с неонатальным рецептором Fc (FcRn). Такие варианты Fc-участков могут включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 436, 439 или 447 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата. Варианты Fc-участка со сниженной связывающей способностью с FcRn могут включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 252, 253, 254, 255, 288, 309, 386, 388, 400, 415, 433, 435, 436, 439 или 447 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата. Указанные выше варианты Fc-участка могут в альтернативном случае демонстрировать повышенное связывание с FcRN и включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата.
Варианты Fc-участка со сниженной способностью к связыванию с FcγR могут включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 239, 248, 249, 252, 254, 265, 268, 269, 270, 272, 278, 289, 292, 293, 294, 295, 296, 298, 301, 303, 322, 324, 327, 329, 333, 335, 338, 340, 373, 376, 382, 388, 389, 414, 416, 419, 434, 435, 437, 438 или 439 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата.
Например, вариант Fc-участка может демонстрировать сниженное связывание с FcγRI и включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 265, 269, 270, 327 или 329 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата.
Вариант Fc-участка может демонстрировать сниженное связывание с FcγRII и включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 265, 269, 270, 292, 294, 295, 298, 303, 324, 327, 329, 333, 335, 338, 373, 376, 414, 416, 419, 435, 438 или 439 Fc-участка, где нумерация остатков Fc-участка соответствует индексу EU, как приведено у Кабата.
Вариант Fc-участка может демонстрировать сниженное связывание с FcγRIII и включать модификацию аминокислоты по любой одной или нескольким аминокислотам в положениях 238, 239, 248, 249, 252, 254, 265, 268, 269, 270, 272, 278, 289, 293, 294, 295, 296, 301, 303, 322, 327, 329, 338, 340, 373, 376, 382, 388, 389, 416, 434, 435 или 437 Fc-участка, где нумерация остатков Fc-участка соответствует индексу Eu, как приведено у Кабата.
Варианты Fc-участка с измененным (например, повышенным или сниженным) уровнем связывания с C1q и/или комплемент-зависимой цитотоксичностью (CDC) описаны в WO99/51642. Такие варианты могут включать замещение аминокислоты по любой, одной или нескольким аминокислотам в положениях 270, 322, 326, 327, 329, 331, 333 или 334 Fc-участка. (См. также Duncan & Winter Nature 322:738-40 (1988); патент США № 5648260; патент США № 5624821; и WO94/29351, где указанные работы относятся к вариантам Fc-участка).
Конструирование вектора
Приведенные в настоящем описании полинуклеотидные последовательности, кодирующие рассматриваемые в настоящем изобретении пептиды/полипептиды, могут быть получены с использованием стандартных рекомбинантных методик. Желательные полинуклеотидные последовательности могут быть выделены из соответствующих клеток-источников и далее секвенированы. Клетки-источники для получения антител включают антитело-продуцирующие клетки, такие как гибридомные клетки. Альтернативно, полинуклеотиды могут быть синтезированы с использованием нуклеотидного синтезатора или в рамках методики ПЦР. Последовательности, кодирующие иммуноглобулины, после их получения, встраивают в рекомбинантный вектор, способный к репликации и экспрессии гетерологичных полинуклеотидов в клетке-хозяине. Многие векторы, которые доступны и известны в данной области, могут использоваться в целях настоящего изобретения. Выбор соответствующего вектора определяется, в основном, размером нуклеиновой кислоты, встраиваемой в вектор, и конкретной клеткой-хозяином, трансформируемой данным вектором. Каждый вектор содержит различные компоненты, определяемые его функцией (амплификация или экспрессия гетерологичного полинуклеотида или обе функции вместе) и его совместимостью с конкретной клеткой-хозяином, в которой он будет находиться. Векторные компоненты в основном включают, без ограничения, репликатор (в особенности, в том случае, когда вектор встраивается в прокариотическую клетку), селектируемый ген-маркер, промотор, сайт связывания с рибосомой (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
В основном, плазмидные векторы, содержащие репликон и контрольные последовательности, получаемые из видов, совместимых с клеткой-хозяином, используют в сочетании с такими клетками-хозяевами. Вектор обычно содержит сайт репликации, а также маркирующие последовательности, которые способны обеспечивать фенотипический отбор трансформированных клеток. Например, Е. coli в типичном случае трансформируют с использованием pBR322, плазмиды, получаемой из E. coli. pBR322 содержит гены, кодирующие резистентность к ампициллину (Amp) и тетрациклину (Тet), и, таким образом, обеспечивает легкий способ идентификации трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаг могут также содержать соответствующие промоторы, или могут быть модифицированы, с тем, чтобы включать промоторы, которые могут использоваться микробным организмом для экспрессии эндогенных белков.
Дополнительно, фаговые векторы, содержащие репликон и контрольные последовательности, совместимые с микроорганизмом-хозяином, могут использоваться в качестве трансформирующих векторов, в сочетании с указанными клетками-хозяевами. Например, бактериофаг, такой как αGEM.TM.-11, может применяться при создании рекомбинантного вектора, который далее может быть использован для трансформации чувствительных клеток-хозяев, таких как E. coli LE392.
В соответствии с настоящим изобретением могут использоваться либо конститутивные, либо индуцибельные промоторы, в зависимости от конкретной ситуации, и их потребность может определить любой специалист со средним уровнем знаний в данной области. В данной области известно множество промоторов, распознаваемых различными потенциальными клетками-хозяевами. Отобранный промотор может быть далее функционально связан с цистроном ДНК, кодирующей полипептид согласно настоящему описанию, путем удаления промотора из источника ДНК за счет расщепления рестрикционным ферментом и с последующим встраиванием выделенной промоторной последовательности в выбранный вектор. Как последовательность нативного промотора, так и последовательность многих гетерологичных промоторов, может быть использована для осуществления амплификации и/или экспрессии целевых генов. Однако гетерологичные промоторы предпочтительны, поскольку они в основном позволяют достичь более высокого уровня транскрипции и более высокого выхода экспрессированного целевого гена по сравнению с нативным промотором целевого полипептида.
Промоторы, подходящие для использования в прокариотических клетках-хозяевах, включают промотор PhoA, промоторные системы β-галактамазы и лактозы, промоторную систему триптофана (trp) и гибридные промоторы, такие как tac или trc промотор. Однако приемлемы и другие промоторы, которые функционируют в бактериях (такие как другие известные бактериальные и фаговые промоторы). Их нуклеотидные последовательности известны и опубликованы, что позволяет специалисту в данной области провести их оперативное лигирование с цистронами, кодирующими целевую легкую и тяжелую цепи (Siebenlist et al., (1980) Cell 20:269)), с использованием соответствующих линкеров или адаптеров для создания любых требуемых сайтов рестрикции.
В некоторых вариантах каждый цистрон в составе рекомбинантного вектора включает последовательность сигнала секреции, где данный компонент направляет транспортировку экспрессированных полипептидов через мембрану. В основном, сигнальная последовательность может представлять собой компонент вектора или может представлять собой часть ДНК целевого полипептида, в которую встраивается вектор. Сигнальная последовательность, выбранная для целей настоящего изобретения, должна быть такой последовательностью, которая распознается и процессируется (то есть расщепляется сигнальной пептидазой) клеткой-хозяином. В случае прокариотических клеток-хозяев, которые не распознают и не процессируют сигнальные последовательности, нативные для гетерологичных полипептидов, такая сигнальная последовательность замещается прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Ipp или термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP.
Прокариотические клетки-хозяева, подходящие для экспрессии полипептидов, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры используемых бактерий включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, виды Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia mercescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. Предпочтительно используют грам-отрицательные клетки. Предпочтительно клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и желательно включение дополнительных ингибиторов протеаз в клеточную культуру.
Продукция пептида/полипептида
Клетки-хозяева подвергают трансформации или трансфекции указанными выше векторами экспрессии и культивируют в стандартных питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих желательные последовательности.
Термин «трансфекция» относится к процессу поступления вектора экспрессии в клетку-хозяин, независимо от того, происходит ли фактически экспрессия кодирующих последовательностей. Специалистам известны различные методики трансфекции, например метод осаждения с использованием CaPO4 и метод электропорации. Считается, что трансфекция произошла успешно, если в клетке-хозяине отмечаются признаки действия данного вектора.
Термин «трансформация» означает встраивание ДНК в прокариотический организм-хозяин, так что указанная ДНК становится реплицируемой либо в виде экстра-хромосомального элемента, либо в интегрированном с хромосомой состоянии. В зависимости от типа используемой клетки-хозяина трансформацию осуществляют с использованием тех стандартных методик, которые подходят для таких клеток. Метод кальциевой обработки, в котором используется хлорид кальция, в основном применим для бактериальных клеток, которые содержат трудные для преодоления барьеры в виде клеточных стенок. Другой метод трансформации включает использование полиэтиленгликоля/ДМСО. Еще одним методом является электропорация.
Прокариотические клетки, используемые для получения пептидов/полупептидов, согласно настоящему изобретению растят в средах, известных специалистам в данной области, которые подходят для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают бульон Луриа ((LB) плюс необходимые питательные добавки. В предпочтительных вариантах среда также содержит селектируемый агент, выбранный с учетом конструкции вектора экспрессии, позволяющий добиться селективного роста прокариотических клеток, содержащих вектор экспрессии. Например, добавляют ампициллин к средам для роста клеток, экспрессирующих ампициллин-резистентный ген.
Могут быть также включены любые необходимые добавки, в дополнение к источникам углерода, азота и неорганического фосфата, в соответствующих концентрациях, которые вводятся либо поодиночке, либо в смеси с другими компонентами или средой, такой как комплексный источник азота. Необязательно культуральная среда может содержать один или несколько восстановителей, выбранных из группы, состоящей из глютатиона, цистеина, цистамина, тиогликоллята, дитиоэритритола и дитиотрейтола.
Прокариотические клетки-хозяева культивируют при подходящих температурах. Для роста E. coli, например, предпочтительные диапазоны температур составляют от примерно 20°С до примерно 39°С, более предпочтительно от примерно 25°С до примерно 37°С, и еще более предпочтительно примерно при 30°С. Среда может иметь любой pH, варьирующий от примерно 5 до примерно 9, главным образом, в зависимости от используемого организма-хозяина. В случае E.coli, pH предпочтительно составляет от примерно 8 до примерно 7,4 и более предпочтительно составляет примерно 7,0.
Если в векторе экспрессии используется индуцибельный промотор, экспрессию белка индуцируют в условиях, подходящих для активации промотора. Так, например, если для контроля транскрипции используют промотор PhoA, то трансформированные клетки могут культивироваться в среде с ограниченным количеством фосфата, для индукции. Может использоваться множество других индукторов, определяемых используемой конструкцией вектора, которые известны в данной области.
Приведенные в настоящем описании пептиды/полипепиды, экспрессируемые в микроорганизме, могут секретироваться и восстанавливаться из периплазмы клеток-хозяев. Процесс восстановления белка в типичном случае вовлекает разрушение микроорганизма с использованием, в основном, таких способов, как осмотический шок, озвучивание или лизис. После разрешения клеток удаляют клеточные осколки или целые клетки центрифугированием или фильтрованием. Белки могут быть далее очищены, например, с использованием аффинной хроматографии на смоле. Альтернативно, белки могут быть перенесены в культуральную среду и выделены из нее. Клетки могут быть удалены из культуры и далее культуральный супернатант фильтруют и концентрируют с целью дальнейшей очистки полученных белков. Экспрессированные полипептиды могут быть также выделены и идентифицированы с использованием широко известных методов, таких как фракционирование на колонках для иммуноаффинной хроматографии или на ионообменных колонках; с использованием метода осаждения этанолом; метода ВЭЖХ с обращением фазы, хроматография на силикагеле или на катионообменной смоле, такой как DEAE; хроматофокусирование; электрофорез SDC-PAGE; осаждение сульфатом аммония; гель-фильтрация с использованием, например, Сефадекс G-75; аффинная хроматография на смолах, основанная на гидрофобном взаимодействии; хроматография, основанная на аффинности к лиганду, с использованием соответствующего антигена, иммобилизованного на матрице и вестерн-блот анализ.
Кроме прокариотических клеток-хозяев, эукариотические системы клеток-хозяев также могут быть установлены методами, известными в данном области. Подходящие организмы-хозяева включают клеточные линии млекопитающих, такие как CHO, и клетки насекомых, такие как клетки, описанные ниже.
Очистка пептида/полипептида
Полученные пептиды/полипептиды могут быть далее очищены с целью получения препаратов, по существу гомогенных для целей дальнейшего тестирования и использования. Стандартные методы очистки белка известны в данной области и могут использоваться соответствующим образом. Приведенные ниже процедуры являются лишь примерами подходящих процедур очистки: фракционирование на иммуноаффинных колонках или на ионообменных колонках, метод осаждения этанолом, метод ВЭЖХ с обращением фазы, хроматография на силикагеле или на катионообменной смоле, такой как DEAE; хроматофокусирование; электрофорез SDC-PAGE; осаждение сульфатом аммония и гель фильтрация с использованием, например, Сефадекс G-75.
Способы настоящего изобретения
Настоящее изобретение относится к различным способам, основанным на том факте, что мутация в некоторых участках β-цепи HGF приводит к модификации биологических активностей молекулы, так что такие мутантные молекулы демонстрируют антагонистические эффекты на модуляции пути HGF/Met.
Могут использоваться различные вещества или молекулы (включая пептиды, полипептиды и т.п.) в качестве терапевтических агентов в соответствии со способами настоящего изобретения. Указанные вещества могут быть изготовлены по известным способам получения фармацевтически полезных композиций, где такой продукт в составе соответствующей смеси с фармацевтически приемлемым носителем. Терапевтические композиции, подходящие для хранения, получают путем смешивания активного ингредиента, обладающего желательной степенью чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Science 16th edition, Osol, A. Ed. (1980)), с получением продуктов в виде лиофилизированных композиций или водных растворов. Используют при этом приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в применяемых дозировках и концентрациях, и включают буферы, такие как фосфат, цитрат, и буферы на основе других органических кислот, антиоксиданты, включающие аскорбиновую кислоту; низкомолекулярные полипептиды (содержащие менее 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон, аминокислоты, такие как глицин, глютамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или ПЭГ.
Композиции, используемые для введения in vivo, должны быть стерильными. Стерильность легко достигается путем фильтрования через стерильные фильтрующие мембраны перед лиофилизацией и восстановлением или после них.
Приведенные в настоящем описании терапевтические композиции помещают в контейнер, имеющий стерильный порт доступа, например в пакет или флакон с внутривенным раствором, имеющим пробку, прокалываемую гиподермной иглой для инъекций.
Введение может быть осуществлено в соответствии с известными методами, например, путем инъекции или инфузии в режиме внутривенного, внутрибрюшинного, интрацеребрального, внутримышечного, внутриглазного, внутриартериального способов введения или введения внутрь повреждения, с помощью местного введения или с использованием систем с пролонгированным высвобождением.
Дозировки и желательные концентрации лекарственного средства в фармацевтических композициях согласно настоящему изобретению могут варьировать, в зависимости от рассматриваемого конкретного способа введения. Определение соответствующей дозировки или способа введения в состоянии осуществить любой практикующий врач. Результаты экспериментов на животных дают надежную основу для определения эффективных доз при лечении людей. Межвидовой анализ эффективных доз может быть проведен с использованием принципов, установленных Морденти и Шаппеллем (Mordenti, J. and Chappel, W. «The use of interspecies scaling in toxicokinetics» In Toxicokinetics and New Drug Development, Yacobi et al., Eds., Pergamon Press, New York 1989, pp.42-96).
В том случае, когда практикуют введение вещества или молекулу согласно настоящему изобретению in vivo, нормальные количества в дозировке могут варьировать от примерно 10 нг/кг до 100 мг/кг веса тела млекопитающего или более в день, предпочтительно примерно 1 мкг/кг/день - 10 мг/кг/день, в зависимости от способа введения. В литературе имеется руководство, касающееся конкретных дозировок и способов доставки. См., например, патенты США №№ 4657760; 5206344 или 5225212. Ожидается, что разные композиции будут эффективны для разных соединений, применяемых для лечения, и разных нарушений, поскольку введение направлено на один орган или ткань, например, может вызвать необходимость доставки по способу, отличному от доставки, направленной на другой орган или ткань.
Когда желательно длительное введение вещества или молекулы в составе композиции с пролонгируемым высвобождением, где характеристики высвобождения приемлемы для лечения конкретного заболевания или нарушения, требующего введения данного вещества или молекулы, рассматривается возможность микроинкапсулирования данного вещества или молекулы. Микроинкапсулирование рекомбинантных белков для достижения пролонгированного высвобождения было успешно осуществлено для человеческого гормона роста (rhGH), интерферона (rhIFN-), интерлейкина-2, и MN rgp 120 (Johnson et al., Nat. Med., 2:795-799; Yasuda, Biomed. Ther. 27:1221-1223 (1993); Hora et al., Bio/Technology, 8:755-758 (1990); Clelan, «Design and Production of Single Immunization Vaccines Using Polylactid Polyglycolid Microsphere Systems», in Vaccines Design: The Subunit and Adjuvant Approach, Powell and Newman, eds. (Plenum Press: New York, 1995), pp.439-463; WO 97/03692, WO 96/40072, WO 96/07399; и патент США № 5654010).
Композиции указанных белков с пролонгированным высвобождением были разработаны с использованием полимера полимолочной-когликолевой кислоты (PLGA), благодаря его биодоступности и широкому диапазону биодеградируемых свойств. Продукты биодеградации PLGA, молочная и гликолевая кислоты, могут быть быстро выведены из организма человека. Кроме того, способность к деградации данного полимера может быть откорректирована до достижения периодов, составляющих от нескольких месяцев до нескольких лет, в зависимости от его молекулярного веса и состава (Lewis, «Controlled release of bioactive agents from lactide/glycolidе polymer» in: M. Chasin and R. Langer (Eds.), Biodegradable Polymers as Drug Delivery Systems (Marcel Dekker: New York, 1990), pp.1-41).
Идентификация участков внутри β-цепи HGF, которые являются решающими для функции HGF в сигнальном пути HGF/Met, указывает те сайты в β-цепи HGF, на которые может быть направлено действие антагонистов. Примеры потенциальных антагонистов включают олигонуклеотид (который может быть аптамером), который связывается с N-концевым участком и/или участком димеризации β-цепи HGF, и в частности антитела, включающие, без ограничения, поли- и моноклональные антитела и фрагменты антител, одноцепочечные антитела, антиидиотипические антитела, и химерные или гуманизированные версии таких антител или фрагментов, а также человеческие антитела и фрагменты таких антител. Аптамеры представляют собой молекулы нуклеиновой кислоты, которые способны связываться с целевой молекулой. Получение и терапевтическое использование аптамеров хорошо известно в настоящей области (См., например, патент США № 5475096, в котором также описывается терапевтическая эффективность Macugen® (Eyetech, New York) при лечении возрастной дегенерации желтого пятна).
Как показано в настоящем описании, HGF/Met антагонист-вещество/молекула согласно настоящему изобретению представляет собой пептид или полипептид (включая антитело). Способы получения таких пептидов и полипептидов известны в данной области и включают скрининг пептидных и полипептидных библиотек на наличие связывания с соответствующим антигеном-мишенью. В одном варианте подходящие антигены-мишени будут включать β-цепь HGF (или ее часть, которая включает N-концевой участок и/или участок димеризации), ниже описанные более подробно. В одном варианте подходящие антигены-мишени будут включать Met, например внеклеточный домен Met. Библиотеки пептидов и полипептидов хорошо известны в данной области и могут быть также получены известными в данной области методами (см., например, Clark et al., патенты США №№ 6121416; и Garrard et al., 5750373, 5733743, 5837242, 5969108, 6172197, 5580717 и 5658727). Библиотеки пептидов и полипептидов, присоединенные к гетерологичному белковому компоненту, такому как белок фаговой оболочки, известны в данной области и описаны, например, в Clark et al. и Garrard et al., выше. Варианты первого выбранного связывающегося пептида или полипептида могут быть получены при скрининге мутантов пептида или полипептида с достижением интересующих характеристик (например, с повышенной аффинностью по связыванию с мишенью, с улучшенной фармакокинетикой, сниженной токсичностью, улучшенным терапевтическим индексом и т.п.). Например, интересующая характеристика может представлять способность связываться с Met, но при наличии сниженной способности активировать HGF-родственные биологические активности, такие как клеточная пролиферация, Met-фосфорилирование, миграция клеток и ангиогенез. Методики мутагенеза известны в данной области, и в настоящем описании описаны участки внутри β-цепи HGF, подходящие для мутации, в частности позиции, ассоциированные с N-концевой инсерцией на β-цепи HGF и/или с димеризацией по типу β-цепь - β-цепь. Кроме того, методы сканирующего мутагенеза (такие, как методы, основанные на аланиновом сканировании) могут быть особенно полезны для оценки структурной и/или функциональной значимости отдельных аминокислотных остатков в составе пептида или полипептида.
Определение способности потенциального вещества/молекулы согласно настоящему изобретению модулировать сигнальную активность HGF/c-met и/или биологическую активность, ассоциированную с указанным сигнальным механизмом, может быть осуществлена при тестировании модулирующей способности данного вещества/молекулы в тестах in vitro или in vivo, которые известны в настоящей области и описаны, например, в следующих работах: Okigaki et al., выше; Matsumoto et al., выше; Date et al., FEBS let. (1997), 420:1-6; Lokkert et al., выше; Hartman et al., выше; Kirchhofer et al., J. Biol. Chem. (2004), 279:39915-24; Stamos et al. (2004) EMBO J. 23:2325-35; Kirhhofer et al., FEBS Lett. (2005) 579:1945-50; и Nakatsu et al. (Microvasc. Res. 66:102, 2003).
Антитела против β-цепи HGF
Настоящее изобретение относится к способам, включающим использование антител. Типичные антитела включают поликлональные, моноклональные, гуманизированные, биспецифические и гетероконъюгированные антитела.
1. Поликлональные антитела
Антитела могут включать поликлональные антитела. Способы получения поликлональных антител известны специалистам в данной области. Поликлональные антитела могут быть получены у млекопитающего, например, путем одной или нескольких инъекций иммунизирующего агента и, при желании, адъюванта. В типичном случае иммунизирующий агент и/или адъювант вводят млекопитающему путем множественных подкожных или внутрибрюшинных инъекций. Иммунизирующий агент может включать β-цепь HGF (или ее часть) или их слитый белок. Может быть также полезно провести конъюгацию иммунизирующего агента с белком, в отношении которого известно, что он является иммуногенным для млекопитающего, подлежащего иммунизации. Примеры таких иммуногенных белков включают, без ограничения, гемоцианин улитки, сывороточный альбумин, бычий тиреоглобулин и ингибитор соевого трипсина. Примеры адъювантов, которые могут при этом использоваться, включают полный адъювант Фрейнда и адъювант MPL-TDM (монофосфорил-липид А, синтетический дикориномиколат трегалозы). Протокол проведения иммунизации может быть выбран специалистами в данной области без затруднительных экспериментов.
2. Моноклональные антитела
В альтернативном варианте антитела могут представлять собой моноклональные антитела. Моноклональные антитела могут быть получены по методам, включающим использование гибридом, описанных, например, Колером и Мильштейном (Kohler and Milstein, Nature, 256:495 (1997)). В соответствии с данным гибридомным методом проводят типичную иммунизацию мыши, хомяка или другого соответствующего организма-хозяина иммунизирующим агентом для проявления действия лимфоцитов, продуцирующих или способных продуцировать антитела, которые, в свою очередь, специфически связываются с иммунизирующим агентом. Альтернативно, лимфоциты могут быть иммунизированы in vitro.
Иммунизирующий агент в типичном случае включает β-цепь HGF (или ее часть) или белок на основе их слияния. В основном, используют либо лимфоциты периферической крови (PBL), если желательны клетки человеческого происхождения, либо клетки селезенки или клетки лимфатического узла, если желательны источники в виде организмов млекопитающих, отличных от человека. Далее лимфоциты сливают с иммортализованной клеточной линией с использованием подходящего агента для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки [Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp.59-103]. Иммортализованные клеточные линии обычно представляют собой трансформированные клетки млекопитающих, в частности миеломные клетки грызунов, быка и клетки человеческого происхождения. Обычно используют миеломные клеточные линии крысы или мыши. Гибридомные клетки могут культивироваться в соответствующей культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживаемость неслитых иммортализованных клеток. Например, если родительские клетки не содержат фермент гипоксантингуанинфосфорибозилтрансферазу (HGPRT или HPRT), то культуральная среда для гибридом в типичном случае включает гипоксантин, аминоптерин и тимидин («среда HAT»), где данные вещества препятствуют росту HGPRT-дефицитных клеток.
Предпочтительные иммортализованные клеточные линии представляют собой такие линии клеток, которые способны эффективно сливаться, поддерживать стабильный высокий уровень экспрессии антитела выбранными антитело-продуцирующими клетками и которые чувствительны к среде, такой как HAT среда. Более предпочтительные иммортализованные клеточные линии представляют собой миеломные линии мыши, которые могут быть получены, например, от Salk Institute Cell Distribution Center, San Diego, California и American Type Culture Collection, Manassas, Virginia. Человеческие миеломные клеточные линии и гетеромиеломные клеточные линии мыши-человека также описаны для целей получения человеческих моноклональных антител [Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Мonoclonal Antibodies Production Techniques and Application, Marcel Dekker, Inc., New York, (1987) pp.51-63].
Культуральная среда, в которой культивируют гибридомные клетки, может быть далее проанализирована на наличие моноклональных антител, направленных против β-цепи HGF. Предпочтительно, связывающую специфичность моноклональных антител, продуцируемых гибридомными клетками, определяют путем иммунопреципитации или при проведении теста на связывание in vitro, такого как радиоиммуноанализ (RIA) или иммуноферментный твердофазный анализ (ELISA). Такие методики и тесты известны в данной области. Связывающая аффинность моноклонального антитела может быть, например, определена по методике Скетчерда (Scatchard) (Munson and Pollard, Anal. Biochem., 107:220 (1980)).
После идентификации желательных гибридомных клеток соответствующие клоны могут быть подвержены субклонированию по процедурам предельного разведения, с дальнейшим их выращиванием по стандартным методикам [Goding, выше]. Подходящие культуральные среды для данной цели включают, например, среду Игла в модификации Дульбекко и среду RPMI1640. Альтернативно, гибридомные клетки могут быть выращены in vivo в виде асцитов в организме млекопитающего.
Моноклональные антитела, секретируемые субклонами, могут быть выделены или далее очищены из культуральной среды или асцитной жидкости с использованием стандартных методик очистки иммуноглобулинов, таких как, например, хроматография с использованием белка А-Сефарозы, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела могут быть также получены с использованием методов рекомбинантной ДНК, таких как методы, описанные в патенте США № 4816567. ДНК, кодирующие моноклональные антитела согласно настоящему изобретению, могут быть легко выделены и секвенированы с использованием стандартных процедур (например, путем использования олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи мышиных антител). Гибридомные клетки согласно настоящему изобретению служат в качестве предпочтительного источника таких ДНК. После выделения ДНК может быть введена в векторы экспрессии, которые затем используют для трансфекции клеток-хозяев, таких как COS клетки обезьяны, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые в ином случае не продуцируют иммуноглобулиновый белок, с достижением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК может быть также модифицирована путем замещения кодирующей последовательности для константных доменов тяжелой и легкой цепи человеческой молекулы гомологичными мышиными последовательностями [патент США № 4819567, Morrison et al., выше] или путем ковалентного присоединения к кодирующей последовательности иммуноглобулина полной или части кодирующей последовательности для полипептида, отличного от иммуноглобулина. Такой не иммуноглобулиновый полипептид может замещать константные домены антитела согласно настоящему изобретению или может замещать вариабельные домены антиген-объединенного сайта антитела согласно настоящему изобретению с образованием химерного бивалентного антитела.
Антитела могут представлять собой моновалентные антитела. Способы получения моновалентных антител известны в настоящее время. Например, один из таких способов включает рекомбинантную экспрессию легкой цепи иммуноглоубулина и модифицированной тяжелой цепи иммуноглобулина. Тяжелая цепь может быть, в основном, усечена по любой точке в Fc-участке, с тем чтобы предотвратить сшивку тяжелой цепи. Альтернативно, проводят замещение релевантных цистеиновых остатков другим остатком аминокислоты или делетируют, что также предупреждает сшивку.
Могут также использоваться методы in vitro для получения моновалентных антител. Расщепление антител с получением их фрагментов, в особенности Fab-фрагментов, может быть осуществлено с использованием стандартных методик, известных в данной области.
Антитела могут быть также получены путем скрининга фаговых библиотек на наличие антител или фрагментов антител, которые связываются с подходящей/желательной афинностью с β-цепью HGF (или их эквивалент). Такие методики известны в данной области и описаны, например, в патентах США №№ 5750373; 5780279; 5821047; 6040136; 5427908; 5580717 и содержащихся в них ссылках.
3. Человеческие и гуманизированные антитела
Антитела с β-цепью HGF согласно настоящему изобретению могут также включать гуманизированные антитела или человеческие антитела. Гуманизированные формы антител, отличных от человеческих (например, мышиных), представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антиген-связывающие последовательности антител), которые содержат минимальную последовательность, полученную из иммуноглобулина, отличного от человеческого. Гуманизированные антитела включают человеческие иммуноглобулины (реципиентное антитело), в которых остатки из участка, определяющего комплементарность (CDR) реципиента замещены остатками из CDR того организма, который относится к виду, отличному от человеческого (донорское антитело), например мыши, крысы или кролика, обладающих желательной специфичностью, аффинностью и способностью. В некоторых случаях остатки каркасной области Fv человеческого иммуноглобулина замещают соответствующими остатками из молекулы, отличной от человеческой молекулы. Гуманизированные антитела могут также включать остатки, которые не найдены ни в реципиентном антителе, ни в импортированных CDR или последовательностях каркасной области. В основном, гуманизированное антитело включает по существу все или по меньшей мере один, и в типичном случае два вариабельных домена, в которых все или по существу все CDR участки соответствуют таковым в иммуноглобулине, отличном от человеческого, и все или по существу все FR области относятся к консенсусной последовательности человеческого иммуноглобулина. Гуманизированное антитело в оптимальном варианте также включает по меньшей мере часть константной области иммуноглобулина (Fc), в типичном случае человеческого иммуноглобулина [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol., 2:593-596 (1992)].
Способы гуманизации антител, отличных от человеческих, известны в данной области. В основном гуманизированное антитело содержит один или несколько аминокислотных остатков, встроенных в него из источника, который отличается от человеческого. Указанные аминокислотные остатки, не относящиеся к человеческой молекуле, также называют «импортные» остатки, которые обычно берут из «импортного» вариабельного домена. Гуманизация может быть по существу проведена по методу Винтера с соавторами (Winter and co-workers) [Jones et al., Nature, 321, 522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)] путем замещения одной или несколькими последовательностями CDR грызунов соответствующих последовательностей в человеческом антителе. Соответственно такие «гуманизированные» антитела представляют собой химерные антитела (патент США № 4816567), где по существу менее чем интактный человеческий вариабельный домен был замещен соответствующей последовательностью из молекулы, отличной от человеческой. Практически, гуманизированные антитела в типичном случае представляют собой человеческие антитела, в которых некоторые CDR остатки и, возможно, FR остатки замещены остатками из аналогичных сайтов антител грызунов.
Человеческие антитела могут быть также получены с использованием различных методик, известных в настоящей области, включающих использование фаговых библиотек [Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991)]. Методики Кула с соавторами и Бернера с соавторами (Cole et al., and Boerner et al.) также применимы для получения человеческих моноклональных антител (Cole et al., Monoclonal antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985) и Boerner et al., J. Immunol., 147(1):86-95 (1991). Аналогично, человеческие антитела могут быть получены путем встраивания локусов человеческого иммуноглобулина в трансгенных животных, например мышей, где эндогенные иммуноглобулиновые гены были частично или полностью инактивированы. При провокации наблюдается продукция человеческого антитела, которая во всех отношениях очень близко отражает то, что имеется в организме человека, включая перегруппировку генов, их сборку и антительный репертуар. Такой подход описан, например, в патентах США №№ 5545807; 5545806; 5569825; 5625126; 5633425; 5661016 и в следующих научных публикациях: Marks et al., Bio/Technology 10, 779-783 (1992); Lonberg et al., Nature, 368, 856-859 (1994); Morrison, Nature, 368, 812-13 (1994); Fishwild et al., Nature Biotechnology 14, 845-51 (1996); Neuberger, Nature Biotechnology 14, 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13, 65-93 (1995).
Антитела могут быть подвернуты «созреванию» по аффинности с использованием известных методов селекции и/или мутагенеза, описанных выше. Предпочтительные «созревшие» по аффинности антитела имеют аффинность, которая в пять раз, более предпочтительно в 10 раз, еще более предпочтительно в 20 или 30 раз больше, чем активность исходного антитела (в основном мышиного, гуманизированного или человеческого антитела), из которого было получено «созревшее» антитело.
4. Биспецифические антитела
Биспецифические антитела представляют собой моноклональные, предпочтительно человеческие или гуманизированные антитела, которые обладают связывающими специфичностями по меньшей мере для двух разных антигенов. В данном случае одна из связывающих специфичностей относится к β-цепи HGF, а другая - к любому другому антигену и, предпочтительно, к белку клеточной поверхности, или рецептору, или субъединице рецептора.
Способы получения биспецифических антител известны в данной области. Стандартный рекомбинантный способ получения биспецифических антител основан на совместной экспрессии двух пар тяжелой цепи/легкой цепи иммуноглобулина, где две тяжелых цепи имеют разную специфичность [Molstein and Cuello, Nature, 305:537-539 (1983)]. В результате случайной перегруппировки тяжелой и легкой цепей иммуноглобулина указанные гибридомы (квадромы) продуцируют потенциальную смесь из десяти разных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Очистка правильной молекулы обычно производится с помощью нескольких стадий аффинной хроматографии. Аналогичные процедуры описаны в WO93/08829, опубликованном 13 мая 1993 года, и в Traunecker et al., EМBO J., 10:3655-3659 (1991).
Вариабельные домены антител с желательными связывающими специфичностями (объединенный сайты антитело-антиген) могут быть присоединены к последовательностям константного домена иммуноглобулина. Слияние предпочтительно осуществляют с константным доменом тяжелой цепи иммуноглобулина, включающим по меньшей мере часть шарнира, участки СН2 и CH3. Предпочтительно, чтобы константный участок первой тяжелой цепи (CH1), содержащий сайт, необходимый для связывания легкой цепи, присутствовал по меньшей мере в одном из слитых продуктов. ДНК, кодирующую слитые продукты на основе тяжелых цепей иммуноглобулина, и, при желательности, легкой цепи иммуноглобулина, встраивают в отдельный вектор экспрессии и подвергают совместной трансфекции в соответствующий организм-хозяин. Дополнительные детали создания биспецифических антител описаны, например, в работе Суреша с соавторами (Suresh et al., Methods in Enzymology, 121:210 (1986)).
В соответствии с другим подходом, описанным в WO 96/27011, интерфейс между парой молекул антител может быть организован таким образом, чтобы максимизировать процент гетеродимеров, восстанавливаемых из культуры рекомбинантных клеток. Предпочтительный интерфейс включает по меньшей мере часть участка CH3 константного домена антитела. В соответствии с данным способом одну или несколько малых боковых цепей аминокислот из интерфейса первой антительной молекулы замещают более крупными боковыми цепями (например, боковыми цепями тирозина или триптофана). На поверхности второй молекулы антитела создают компенсаторные «полости» идентичного или близкого размера к одной или нескольким крупным боковым цепям, путем замещения крупных боковых цепей аминокислоты более мелкими (например, аланина или треонина). Это создает механизм повышения выхода гетеродимера относительно других нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела могут быть получены в виде полноразмерных антител или фрагментов антител (например, F(ab')2 биспецифических антител). Методики создания биспецифических антител из фрагментов антител описаны в научной литературе. Например, биспецифические антитела могут быть получены с использованием химического связывания. Бреннан с соавторами (Brenann et al., Science 229:81 (1985)) описывают процедуру, в соответствии с которой интактные антитела подвергают протеолитическому расщеплению с образованием фрагментов F(ab')2. Указанные фрагменты восстанавливают в присутствии арсенита натрия, в качестве агента, образующего комплекс с дитиолом, для стабилизации близлежащих дитиолов и предотвращения образования межмолекулярных дисульфидных связей. Полученные Fab' фрагменты далее превращают в производные тионитробензоата (TNB). Один из Fab'-TNB производных далее преобразуют до Fab'-тиола путем восстановления с использованием меркаптоэтиламина и смешивают с эквимолярным количеством другого Fab'-TNB производного с образованием биспецифического антитела. Полученные биспецифические антитела могут быть далее использованы в качестве агентов для селективной иммобилизации ферментов.
Fab'-фрагменты могут быть непосредственно выделены из клеток E. coli и далее химически связаны с образованием биспецифических антител. Шалаби с соавторами (Shalaby et al., J. Exp. Med. 175:217-225 (1992)) описывают получение полностью гуманизированных молекул биспецифического антитела F(ab')2. Fab'-фрагмент отдельно секретируется клетками E. coli, после чего его подвергают направленному химическому связыванию in vitro с образованием биспецифического антитела. Созданное таким образом биспецифическое антитело способно связываться с клетками, осуществляющими сверхэкспрессию рецептора ErbB2, и с нормальными человеческими T-клетками, а также запускать литическую активность человеческих цитотоксических лимфоцитов против опухолевых клеток молочной железы человека, в качестве мишени.
Описаны также различные методики получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получают с использованием лейциновых зипперов (Kostelny et al., J. Immunol. 148(5):1547-1553 (1992)). Лейциновые зипперные пептиды из белков Fos и Jun соединяют с Fab'-частями двух разных антител путем генного слияния. Гомодимеры антител восстанавливают по шарнирному участку с образованием мономеров и затем повторно окисляют с образованием антительных гетеродимеров. Данный способ может также использоваться для получения гомодимеров антител. Методика получения «диантител», описанная Холлингером с соавторами (Hollinger et al., Prоc. Natl. Acad. Sci USA 90:6444-6448 (1993)), предлагает альтернативным механизм создания фрагментов биспецифических антител. Указанные фрагменты включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменов легкой цепи (VL) с помощью линкера, который достаточно короткий, чтобы осуществить спаривание между двумя доменами на одной и той же цепи. Соответственно (VH) и (VL) домены из одного фрагмента заставляют спариваться с комплементарными (VL) и (VH) доменами другого фрагмента, что приводит к образованию двух антиген-связывающих сайтов. Описана также другая стратегия создания фрагментов биспецифических антител путем использования димеров одноцепочечных Fv (sFv). (См. Gruber et al., J. Immunol. 152:5368 (1994)).
Рассматриваются также антитела, обладающие более чем двумя валентностями. Например, могут быть получены триспецифические антитела (Tutt et al., J. Immunol. 147:60 (1991)).
Типичные биспецифические антитела могут связываться с двумя разными эпитопами на β-цепи HGF или с эпитопом на β-цепи HGF и с эпитопом на другом полипептиде (например, c-met или α-цепи HGF).
5. Гетероконъюгированные антитела
Гетероконъюгированные антитела также входят в объем настоящего изобретения. Гетероконъюгированные антитела состоят из двух ковалентно соединенных антител. Такие антитела, например, были предложены для того, чтобы направлять клетки иммунной системы на нежелательные клетки [патент США № 4676980] и для лечения ВИЧ-инфекции [WO 91/00369; WO 92/200372; EP 03089]. Такие антитела могут быть получены in vitro с использованием известных методик синтетической белковой химии, включая методики, использующие сшивающие агенты. Например, иммунотоксины могут быть сконструированы реакцией дисульфидного обмена или путем образования тиоэфирной связи. Примеры подходящих для данной цели реагентов включают иммунотиоат и метил-4-меркаптобутиримидат, а также агенты, описанные, например, в патенте США № 4676980.
6. Воздействие на эффекторную функцию с использованием методов генной инженерии
В антителе согласно настоящему изобретению может быть желательно модифицировать его эффекторную функцию, с тем чтобы повысить, например, эффективность антитела при лечении рака. Например, один или несколько цистеиновых остатков могут быть введены в Fc-участок, что создает возможность для образования межцепьевой дисульфидной связи в данном участке. Полученное таким образом гомодимерное антитело может обладать улучшенной способностью к интернализации и/или повышенной способностью к комплемент-зависимому лизису клеток и антитело-обусловленной клеточной токсичностью (ADCC) (См. Caron et al., J. Exp. Med., 176:1191-1195 (1992) и Shopes, J. Immunol., 148:2918-2922 (1992)). Гомодимерные антитела с повышенной противоопухолевой активностью могут быть также получены с использованием гетеробифункциональных сшивающих агентов, как описано, например, в Wolff et al., Cancer Research, 53:2560-2565 (1993).
Альтернативно, антитело может быть получено генно-инженерными методами, так что оно будет содержать двойные участки Fc и, таким образом, может обладать усиленными способностями к комплемент-зависимому лизису и к ADCC (См. Stevenson et al., Anti-Cancer Drug Design, 3:219-230 (1989)).
7. Иммуноконъюгаты
Настоящее изобретение также относится к иммуноконъюгатам, включающим антитело, конъюгированное с цитотоксическим агентом, таким как химиотерапевтический агент, токсин (например, энзиматически активный токсин бактериального, грибного, растительного или животного происхождения или их фрагменты) или с радиоактивным изотопом (то есть радиоконъюгатом).
Химиотерапевтические агенты, используемые при создании таких иммуноконъюгатов, были описаны выше. Энзиматически активные токсины и фрагменты, которые могут использоваться, включают А цепь дифтерии, несвязывающиеся активные фрагменты дифтерийного токсина, А цепь экзотоксина (из Pseudomonas aeruginosa), А цепь рицина, А цепь абрина, А цепь модеццина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Доступно множество радионуклидов для получения радиоконъюгированных антител. Соответствующие примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического агента создают с использованием большого числа бифункциональных белок-связывающих агентов, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глютарельдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), бис-диазониевые соединения (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толиен-2,6-диизоцианат) и бис-активные фтор-содержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, рициновый иммунотоксин может быть получен по методу, описанному в работе Вилетта с соавторами (Vitetta et al., Science, 238:1098 (1987)). Меченый по углероду-14 1-изотиоцианатобензил-3-метилдиэтилен-триаминпентауксусная кислота (MX-DTPA) представляет собой репрезентативный хелатирующий агент, применяемый для конъюгации радионуклида и антитела. См. WO94/11026.
В другом варианте антитело может быть конъюгировано с «рецептором» (таким как стрептавидин) для использования с целью предварительного нацеливания на опухоль, когда конъюгат антитело-рецептор вводят пациенту, с последующим удалением из кровотока несвязанного конъюгата с использованием выводящего агента, после чего вводят «лиганд» (например, авидин), который был конъюгирован с цитотоксическим агентом (например, с радионуклидом).
8. Иммунолипосомы
Приведенные в настоящем описании антитела могут быть также изготовлены в виде иммунолипосом. Липосомы, содержащие антитело, получают по известным в данной области методикам, таким как описанные в Epstein et al., Proc. Natl. Acad. Sci. USA, 82:3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77:4030 (1980); и патентах США №№ 4485045 и 4544545. Липосомы с повышенным временем пребывания в кровотоке описаны в патенте США № 5013556.
Особенно часто используемые липосомы могут быть получены по методу выпаривания с обращением фазы при использовании липидной композиции, включающей фосфатидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (PEG-PE). Липосомы подвергают экструзии через фильтры с определенным размером пор с получением липосом желательного диаметра. Fab'-фрагменты антител согласно настоящему изобретению могут быть конъюгированы с липосомами по методу, описанному Martin et al., J. Biol. Chem., 257:286-288 (1982) по процедуре реакции дисульфидного обмена. Химиотерапевтический агент (такой как доксорубицин) может необязательно содержаться в составе липосомы. См. Gabizon et al., J. National Cancer Inst., 81(19):1484 (1989).
9. Фармацевтические композиции антител
Антитела могут вводиться для лечения различных нарушений в виде фармацевтических композиций.
Если используют целые антитела в качестве ингибиторов, предпочтительны антитела, способные к интернализации. Однако липофектины или липосомы также могут использоваться для доставки антител согласно настоящему изобретению в клетки, если это желательно. В том случае, когда используют фрагменты антител, предпочтителен наименьший ингибирующий фрагмент. Например, на основе последовательностей вариабельного участка антитела могут быть созданы пептидные и полипептидные молекулы, которые сохраняют способность связывать β-цепь HGF и/или препятствовать взаимодействию между β-цепью HGF и c-met, мешать инсерции N-конца β-цепи HGF и/или препятствовать взаимодействию по типу β-цепь - β-цепь HGF. Такие пептиды и полипептиды могут быть синтезированы химически и/или могут быть получены методами рекомбинантной ДНК (см., например, Marasco et al., Proc. Natl. Acad. Sci. USA, 90:7889-7893 (1993)). Описанная в настоящей заявке композиция может содержать более одного активного соединения в соответствии с потребностью определенного показания, подлежащего лечению, предпочтительно с наличием дополнительных активностей, которые не оказывают друг на друга неблагоприятного воздействия. Альтернативно или дополнительно, композиция может включать агент, который повышает ее функцию, такой как, например, цитотоксический агент, цитокин, химиотерапевтический агент или агент, ингибирующий рост. Такие молекулы вводятся в сочетании, в тех количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты могут быть также введены в микрокапсулы, получаемые, например, путем коацервации или по методу межпоточной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы или поли-(метилметакрилатные) микрокапсулы, соответственно, в состав коллоидных систем доставки лекарственных средств (например, в состав липосом, альбуминовых микросфер, микроэмульсий, наночастиц и нанокапсул) или в состав макроэмульсий. Такие методики описаны в руководстве Ремингтона (Remington's Pharmaceutical Science, выше).
Композиции, подходящие для использования с целью введения in vivo, должны быть стерильными. Стерильность легко может быть достигнута при проведении фильтрования через мембраны для стерильного фильтрования.
Могут быть получены препараты с пролонгированным высвобождением. Подходящие примеры препаратов с пролонгированным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело, где указанная матрица имеет вид изделия определенной формы, например, имеет вид пленок или микрокапсул. Примеры матриц с пролонгированным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США № 3773919), сополимеры L-глютаминовой кислоты и γ-этил-L-глютамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата леупролида) и поли-D-(-)-3-гидроксимасляную кислоту. Тогда как полимеры, такие как этиленвинилацетат и сополимер молочной кислоты-гликолевой кислоты способны высвобождать молекулы в течение 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов времени. В том случае, когда инкапсулированные антитела остаются в организме в течение длительного времени, они могут денатурировать или образовывать агрегаты в результате воздействия влажности при температуре 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. В этой связи должны быть разработаны рациональные стратегии стабилизации, определяемые соответствующим функционирующим механизмом. Например, если показано, что механизм агрегации включает образование межмолекулярной S-S связи через тиодисульфидный обмен, то стабилизация может быть достигнута путем модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля содержания влаги, с использованием соответствующих добавок и путем разработки специфических полимерных матричных композиций.
Ниже приведены примеры способов и композиций согласно настоящему изобретению. Следует понимать, что в практике осуществления настоящего изобретения могут быть приемлемы другие варианты, тогда как приведенный выше текст дает лишь общее описание изобретения.
ПРИМЕРЫ
МАТЕРИАЛЫ И МЕТОДЫ
Мутанты HGF получают, по существу, по методике, описанной в Kirchhofer et al., J Biol. Chem. (2004), 279:39915-24; Stamos et al. (2004) EMBO J. 23:2325-35. Тесты на связывание с Met и на миграцию клеток MDA-MB435 проводят с использованием реагентов и способов, описанных в Kirchhofer et al., выше и Stamos et al., выше. Тест на фосфорилирование Met проводят с использованием клеток A549, по существу, как описано в Kirchhofer et al., выше. Аналогично, достигают ингибирования фосфорилирования Met, за исключением того, что мутанты HGF для оценки ингибирования фосфорилирования добавляют в зависимом от дозы режиме с использованием 50 нг/мл HGF. Ингибирование клеточной пролиферации проводят в тесте на BxPC3, по существу, как описано в Kirchhofer et al., FEBS Lett. (2005) 579:1945-50). Тест на ангиогенез in vitro проводят, по существу, как описано у Nakatsu et al., Microvasc. Res. 66:102, 2003). Мутанты HGF добавляют в культуральную среду в концентрации 10 мкг/мл через день в течение 6-дневного эксперимента. В конце эксперимента определяют число проростков на гранулу и выражают результат в виде среднего показателя ±SD для 4 независимых экспериментов.
Результаты
Мутанты β-цепи HGF (в виде одной β-цепи и в виде полноразмерного HGF), имеющие указанные мутации, оценивают на их способность связываться с MetIgG по сравнению с β-цепью HGF дикого типа (одна β-цепь и полноразмерный HGF), используя тест на основе конкурентного ELISA. Для минимизации любого возможного образования димера, связанного дисульфидной связью, β-цепь HGF дикого типа и мутант β-цепи HGF анализируют на фоне C604S; таким образом, β HGF дикого типа фактически является HGF β C604S. Кроме того, оценивают выбранные полноразмерные двухцепочечные мутанты HGF в тесте на клеточную миграцию для определения эффектов, если таковые имеются, на биологическую функцию в результате мутации в β-цепи. Полученные данные показаны на фиг.1A, B, C. Мутант HGF G498I ингибирует HGF-зависимое фосфорилирование Met, зависимым от дозы образом, как показано на фиг.2; приведены также результаты для HGF мутанта R424A:R494E (одноцепочечный HGF). Мутанты HGF G498I и G498P активируют Met в значительно меньшей степени, чем HGF дикого типа, по результатам теста на фосфорилирование Met, как показано на фиг.3; приведены также результаты для HGF мутанта R424A:R494E. HGF-зависимое фосфорилирование Met модулируется зависимым от дозы образом. Мутанты HGF G498I и G498P также ингибируют пролиферацию клеток BxPC3, характеризующихся уровнем активности 2,5% и 56% относительно активности HGF дикого типа. Кроме того, выбранные полноразмерные мутанты HGF в концентрации 10 мкг/мл (D672N, V495G, G498I, R424A:R494E) ингибируют ангиогенез в тесте in vitro (фиг.4), дополнительно подтверждая значимость β-цепи HGF (и выбранных мутаций) для осуществления полной биологической функции HGF.
ЧАСТИЧНЫЙ ПЕРЕЧЕНЬ ЦИТИРОВАННЫХ РАБОТ





| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТАГОНИСТЫ, НАПРАВЛЕННЫЕ ПРОТИВ c-met | 2005 |
|
RU2398777C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДУЛЯЦИИ АКТИВАЦИИ ФАКТОРА РОСТА ГЕПАТОЦИТОВ | 2005 |
|
RU2405041C2 |
| СПОСОБ ЛЕЧЕНИЯ ОПУХОЛИ У СУБЪЕКТА | 2006 |
|
RU2404193C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ АНТАГОНИСТАМИ С-МЕТ И EGFR | 2009 |
|
RU2601892C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2552161C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ с-МЕТ | 2011 |
|
RU2607377C2 |
| СПОСОБ МОДУЛИРОВАНИЯ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТИ МОНОКЛОНАЛЬНОГО АНТИТЕЛА | 2009 |
|
RU2575600C2 |
| МОДУЛЯТОРЫ АКТИВАТОРА ФАКТОРА РОСТА ГЕПАТОЦИТОВ | 2005 |
|
RU2418004C2 |
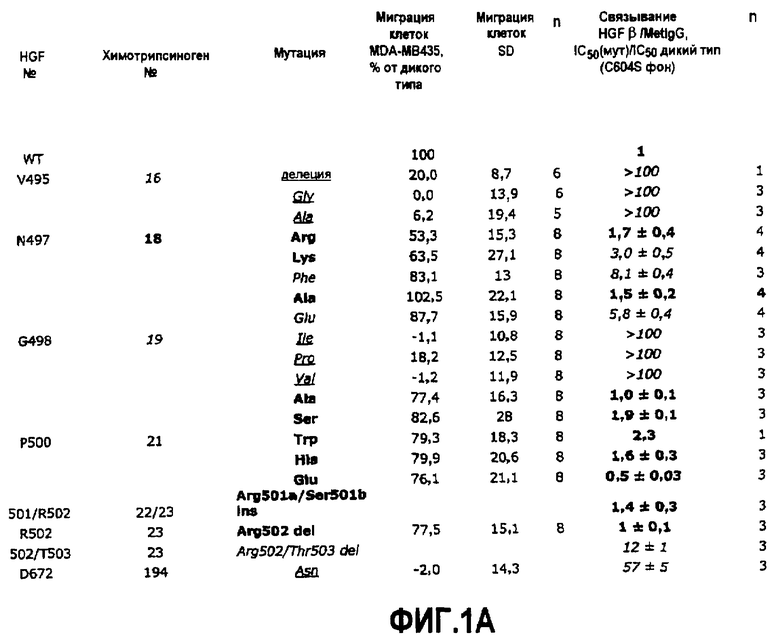

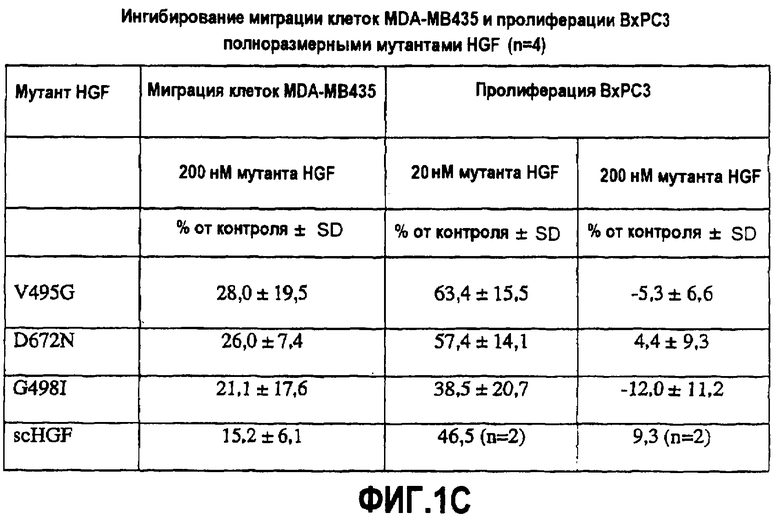
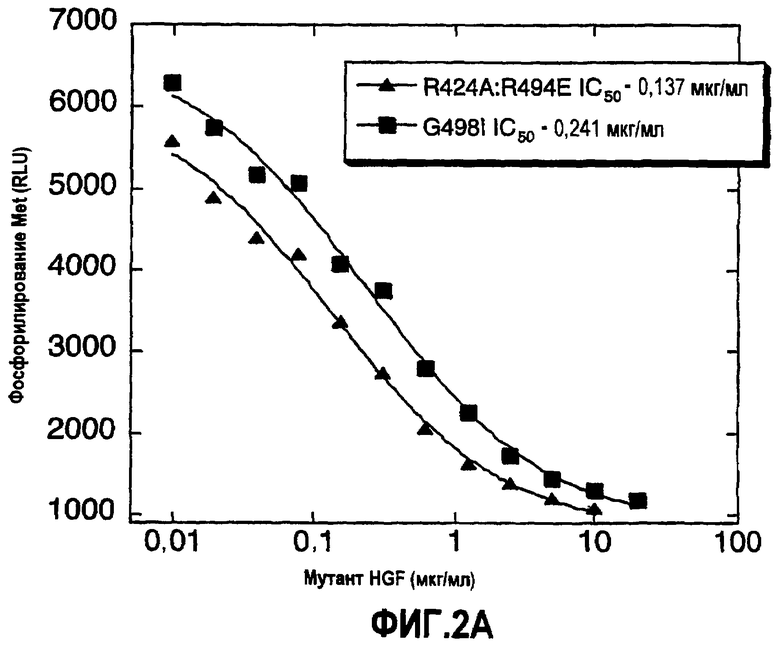
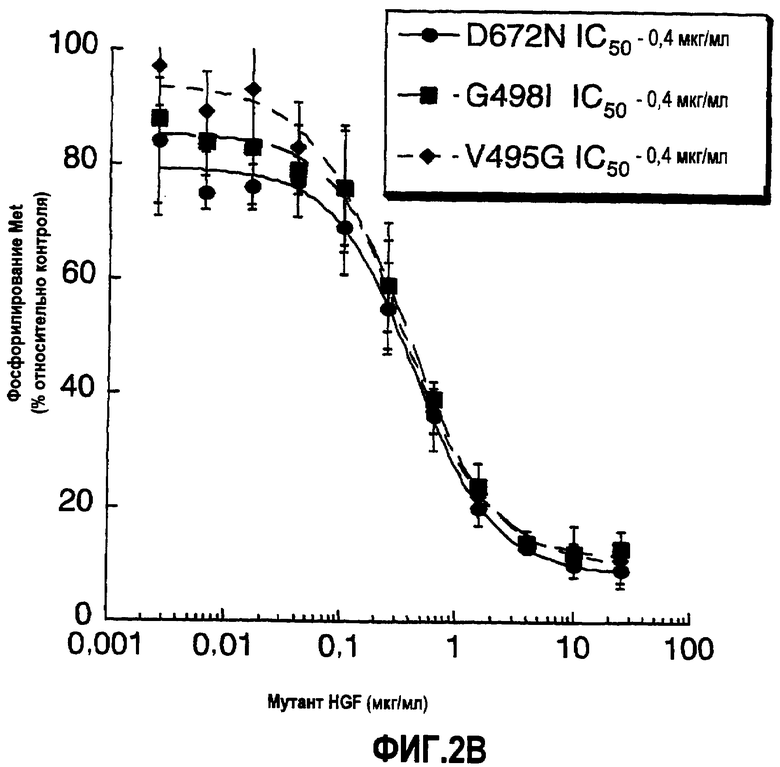
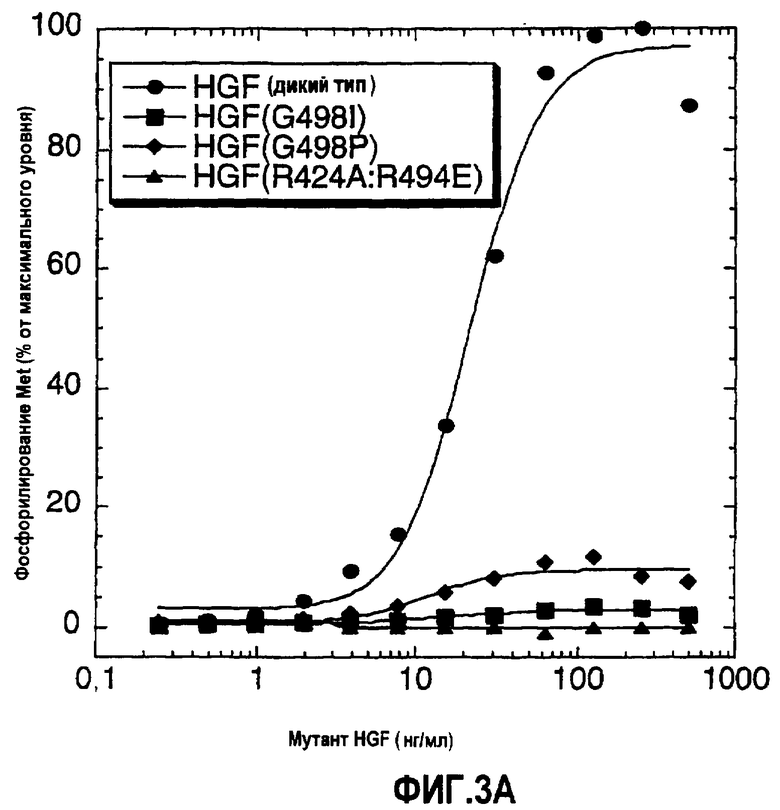
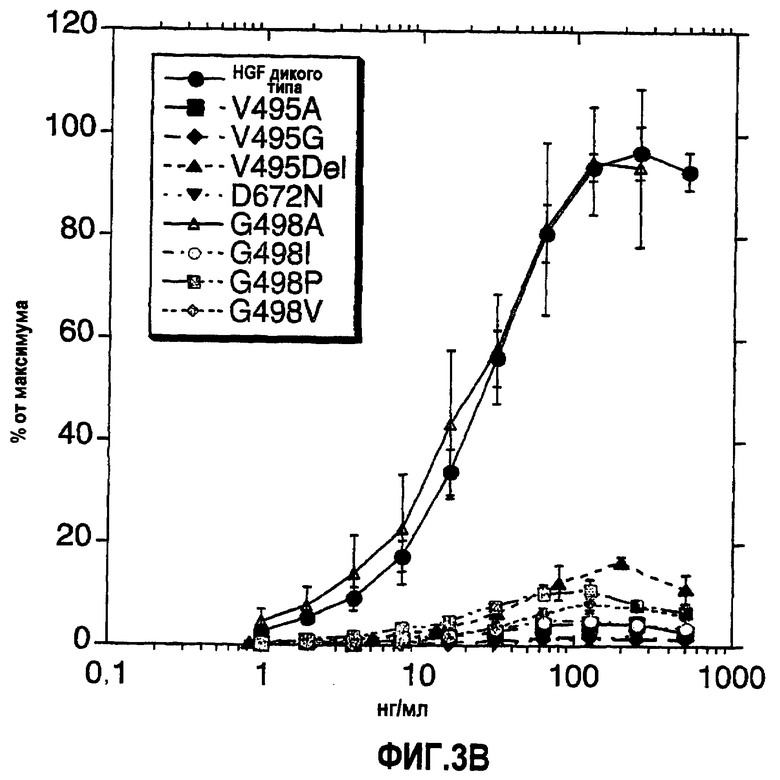
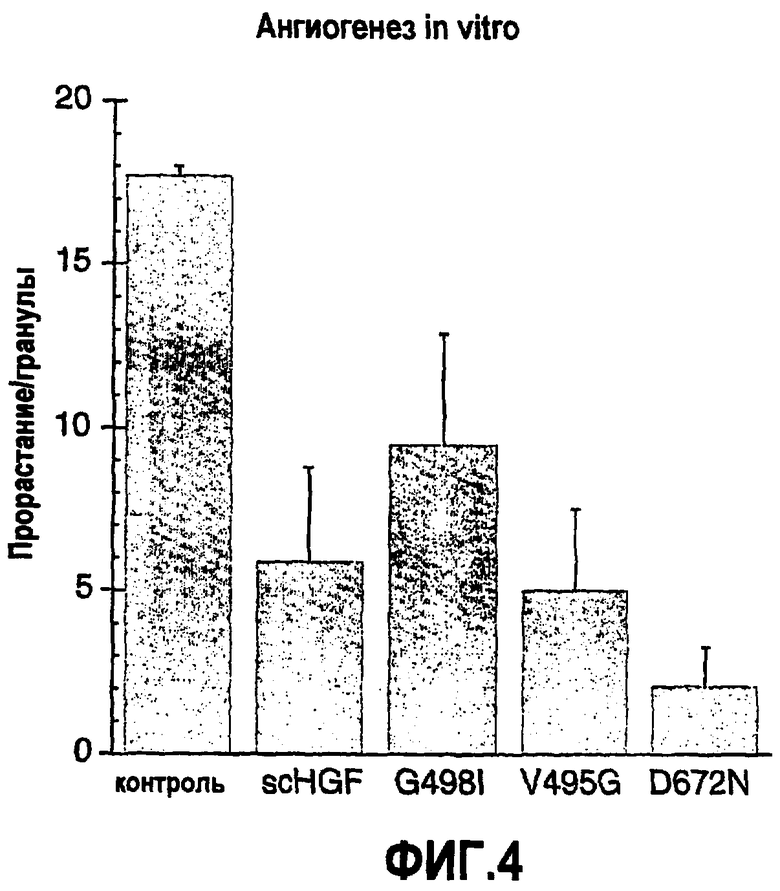
Настоящее изобретение относится к области молекулярной биологии и может быть использовано при разработке средств и методов для модулирования функций организма, связанных с сигнальным путем HGF/c-met. Предложены полипептиды-антагонисты HGF/c-met, представляющие собой мутантные формы HGF, которые содержат мутацию в N-концевом участке β-цепи и/или в участке ее димеризации. Полипептиды по изобретению обладают пониженной в сравнении с HGF дикого типа биологической активностью и могут быть использованы для модуляции активации c-met, клеточной пролиферации, клеточной миграции и ангиогенной активности клетки. Описан способ получения новых мутеинов HGF с помощью технологии рекомбинантных ДНК и средства, необходимые для его осуществления. 8 н. и 14 з.п.ф-лы, 8 ил., 1 табл.
1. Полипептид-антагонист HGF/C-Met, представляющий собой мутант HGF, содержащий мутацию в N-концевом участке β-цепи HGF и/или в участке димеризации β-цепи HGF, где мутация в N-концевом участке β-цепи HGF представляет собой V495G, V495A, G498I, G498P, G498V, R502del + T503del или D672N и где мутация в участке димеризации (3-цепи HGF представляет собой N497R, N497K, G498A, G498S, P500W, Р500Н, Р500Е, инсерцию, например, R и S, между Т501 и R502, или R502del.
2. Полипептид-антагонист по п.1, отличающийся тем, что мутация в N-концевом участке β-цепи HGF нарушает вставку N-конца β-цепи HGF в HGF-связывающий карман и где полученный мутант HGF обладает значительно сниженной биологической функцией по сравнению с HGF дикого типа.
3. Полипептид-антагонист по п.2, отличающийся тем, что биологическая функция представляет клеточную пролиферацию, миграцию клеток, фосфорилирование Met или ангиогенез.
4. Полипептид-антагонист по любому из предшествующих пунктов, отличающийся тем, что мутация в N-концевом участке β-цепи HGF нарушает вставку N-конца β-цепи HGF в HGF-связывающий карман и где полученный мутант HGF связывается с C-Met со значительно сниженной связывающей аффинностью по сравнению с HGF дикого типа.
5. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что мутация в N-концевом участке β-цепи HGF нарушает вставку N-конца β-цепи HGF в HGF-связывающий карман и где полученный мутант HGF связывается с C-Met с по существу эквивалентной аффинностью, как и HGF дикого типа.
6. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что молекула включает мутацию в участке димеризации β-цепи HGF, где полученный мутант HGF обладает значительно сниженной биологической функцией по сравнению с HGF дикого типа.
7. Полипептид-антагонист по п.6, отличающийся тем, что биологическая функция представляет собой клеточную пролиферацию, миграцию клеток, фосфорилирование Met или ангиогенез.
8. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что полипептид включает мутацию в участке димеризации β-цепи HGF, где полученный мутант HGF обладает сниженной способностью к димеризации с другой β-цепью HGF.
9. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что мутация в участке димеризации цепи HGF, по существу, не нарушает связывания полученного мутанта HGF с C-Met.
10. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что мутация в положении N497 не является мутацией N497F, А или Е.
11. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что молекула включает аминокислоты дикого типа в положениях 534, 578, 619, 673, 692, 693, 694, 695, 696, 699 и/или 702.
12. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что полипептид обладает сниженной C-Met-сигнальной способностью по сравнению с HGF дикого типа.
13. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что полипептид обладает сниженной способностью стимулировать миграцию клеток по сравнению с HGF дикого типа.
14. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что полипептид обладает сниженной способностью стимулировать клеточную пролиферацию по сравнению с HGF дикого типа.
15. Полипептид-антагонист по любому из пп.1-3, отличающийся тем, что полипептид обладает сниженной способностью стимулировать ангиогенез по сравнению с HGF дикого типа.
16. Способ модуляции c-met-активации in vitro, включающий введение полипептида-антагониста HGF/c-met по любому из пп.1-15, посредством чего достигается модуляция активации c-met.
17. Способ модуляции клеточной пролиферации in vitro, включающий введение полипептида-антагониста HGF/c-met по любому из пп.1-15, посредством чего достигается модуляция клеточной пролиферации.
18. Способ модуляции клеточной миграции in vitro, включающий введение полипептида-антагониста HGF/c-met по любому из пп.1-15, посредством чего достигается модуляция клеточной миграции.
19. Способ модуляции ангиогенной активности клетки in vitro, включающий введение полипептида-антагониста HGF/c-met по любому из пп.1-15, посредством чего достигается модуляция ангиогенной активности клетки.
20. Нуклеиновая кислота, кодирующая полипептид-антагонист HGF/c-met по любому из пп.1-15.
21. Клетка-хозяин, включающая нуклеиновую кислоту по п.20, для экспрессии полипептида-антагониста HGF/c-met по любому из пп.1-15.
22. Способ получения полипептида-антагониста HGF/c-met по любому из пп.1-15, включающий экспрессию в клетке-хозяине нуклеиновой кислоты, кодирующей полипептид-антагонист, и выделение полипептида-антагониста из клетки.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| STAMOS J | |||
| ET AL | |||
| EMBO J, 23, 2325-2335, 2004 | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| LOKKER N | |||
| ET AL | |||
| EMBO J., 11(7), 2503-2510, 1992. | |||

