Настоящее изобретение относится к новому бивалентному антителу, способному к специфичному связыванию с рецептором c-Met человека и/или способному к специфичному ингибированию тирозинкиназной активности данного рецептора, предпочтительно как лиганд-зависимым, так и лиганд-независимым путем, а также к аминокислотной и нуклеиново-кислотной последовательности, кодирующей данное антитело. Более предпочтительно данное антитело содержит модифицированную шарнирную область и проявляет улучшенную антагонистическую активность. Более конкретно антитело в соответствии с изобретением способно к ингибированию димеризации c-Met. Изобретение также включает применение данного антитела в качестве лекарства для профилактического и/или терапевтического лечения раков, предпочтительно рака, характеризующегося лиганд-независимой активацией c-Met, или любой патологии, связанной с гиперэкспрессией данного рецептора, а также способы и наборы для диагностики заболевания, связанного с гиперэкспрессией c-Met. Изобретение, наконец, включает препараты и/или композиции, содержащие такое антитело в комбинации с другими антителами и/или химическими соединениями, направленными против других факторов роста, вовлеченных в прогрессирование или метастаз опухоли, и/или соединениями и/или противораковыми агентами или агентами, конъюгированными с токсинами, и их применение для предупреждения и/или лечения определенных раков.
Агенты, направленные на рецепторные тирозинкиназы (RTK), такие как ингибиторы трастузумаб, цетуксимаб, бевацизумаб, иматиниб и гефитиниб, показали интерес к направленности данного класса белков на лечение избранных раков.
c-Met является прототипным членом подсемейства RTK, которое также включает RON и SEA. Семейство c-Met RTK структурно отличается от других семейств RTK и является единственным известным высоко аффинным рецептором к фактору роста гепатоцитов (HGF), также называемому рассеивающим фактором (SF) [D.P. Bottaro et al., Science 1991, 251:802-804; L. Naldini et al., Eur. Mol. Biol. Org. J. 1991, 10:2867-2878]. c-Met и HGF распространенно экспрессируются в ряде тканей, и их экспрессия в норме ограничена клетками эпителиального и мезенхимального происхождения соответственно [M.F. Di Renzo et al., Oncogene 1991, 6:1997-2003; E. Sonnenberg et al., J. Cell. Biol. 1993, 123:223-235]. Оба они требуются для нормального развития млекопитающих, и показано, что они особенно важны при миграции клеток, морфогенетической дифференциации и организации трехмерных тубулярных структур, а также при росте и ангиогенезе [F. Baldt et al., Nature 1995, 376:768-771; С. Schmidt et al., Nature. 1995:373:699-702; Tsarfaty et al., Science 1994, 263:98-101]. Хотя показано, что контролируемая регуляция c-Met и HGF важна при развитии млекопитающих, поддержании и репарации тканей [Nagayama Т., Nagayama M., Kohara S., Kamiguchi H., Shibuya M., Katoh Y., Itoh J., Shinohara Y., Brain Res. 2004, 5;999(2):155-66; Tahara Y, Ido A., Yamamoto S., Miyata Y., Uto H., Hori Т., Hayashi K., Tsubouchi H., J Pharmacol Exp Ther. 2003, 307(1):146-51], нарушение их регуляции считают причиной прогрессирования раков.
Аберрантная передача сигнала, направляемая неправильной активацией с-Met, является одной из наиболее частых аберраций, наблюдаемых при раках человека, и играет критическую роль в канцерогенезе и метастазах [Birchmeier et al., Nat. Rev. Mol. Cell Biol. 2003, 4:915-925; L. Trusolino and Comoglio P. M., Nat Rev. Cancer. 2002, 2(4):289-300].
Неправильная активация c-Met может возникать в результате лиганд-зависимых и независимых механизмов, которые включают гиперэкспрессию c-Met и/или паракринную или аутокринную активацию, или посредством прироста функциональных мутаций [J.G. Christensen, Burrows J. and Salgia R., Cancer Latters. 2005, 226:1-26]. Однако олигомеризация рецептора c-Met в присутствии или в отсутствие лиганда необходима для регуляции связывающего сродства и кинетики связывания киназы с АТФ и содержащими тирозин пептидными субстратами [Hays JL, Watowich SJ, Biochemistry, 2004 Aug 17, 43:10570-8]. Активированный c-Met рекрутирует сигнальные эффекторы к его сайту множественной стыковки, расположенному в цитоплазматическом домене, приводя в результате к активации нескольких ключевых путей передачи сигнала, включая Ras-MAPK, PI3K, Src и Stat3 [Gao CF, Vande Woude GF, Cell Res. 2005, 15(1):49-51; Furge KA, Zhang YW, Vande Woude GF, Oncogene. 2000, 19(49):5582-9]. Эти пути существенны для пролиферации опухолевых клеток, инвазии и ангиогенеза и для уклонения от апоптоза [Furge KA, Zhang YW, Vande Woude GF, Oncogene, 2000, 19(49):5582-9; Gu H., Neel BG, Trends Cell Biol. 2003 Mar, 13(3):122-30; Fan S., Ma YX, Wang JA, Yuan RQ, Meng Q, Cao Y., Laterra JJ, Goldberg ID, Rosen EM, Oncogene. 2000 Apr 27, 19(18):2212-23]. Кроме того, уникальным аспектом передачи сигнала c-Met относительно других RTK является его описанное взаимодействие с фокальными адгезионными комплексами и не киназными партнерами связывания, такими как интегрины α6β4 [Trusolino L., Bertotti A., Comoglio PM, Cell. 2001, 107:643-54], CD44v6 [Van der Voort R., Taher ТЕ, Wielenga VJ, Spaargaren M., Prevo R., Smit L., David G., Hartmann G., Gherardi E., Pals ST, J. Biol. Chem. 1999, 274(10):6499-506], плексин B1 или семафорины [Giordano S., Corso S., Conrotto P., Artigiani S., Gilestro G., Barberis D., Tamagnone L., Comoglio PM, Nat Cell Biol. 2002, 4(9):720-4; Conrotto P., Valdembri D., Corso S., Serini G, Tamagnone L., Comoglio PM, Bussolino F., Giordano S., Blood. 2005, 105(11):4321-9; Conrotto P., Corso S., Gamberini S., Comoglio PM, Giordano S., Oncogene. 2004, 23:5131-7], что может дополнительно способствовать комплексности регуляции клеточной функции этим рецептором. Наконец, последние данные демонстрируют, что c-Met может быть вовлечен в устойчивость опухоли к гефитинибу или эрлотинибу, что позволяет предположить, что комбинация соединения, направленная как на EGFR, так и на c-Met, могла бы представлять значительный интерес [Engelman JA et al., Science, 2007, 316:1039-43].
В последние несколько лет множество различных стратегий разработано для аттенуации передачи сигнала c-Met в раковых клеточных линиях. Эти стратегии включают i) нейтрализующие антитела против c-Met или HGF/SF [Сао В., Su Y., Oskarsson M., Zhao P., Kort EJ, Fisher RJ, Wang LM, Vande Woude GF, Proc Natl Acad Sci USA. 2001, 98(13)7443-8; Martens Т., Schmidt NO, Eckerich C, Fillbrandt R., Merchant M., Schwall R., Westphal M., Lamszus K., Clin Cancer Res. 2006, 12(20):6144-52] или применение антагониста HGF/SF NK4 для предотвращения связывания лиганда с c-Met [Kuba K., Matsumoto K., Date K., Shimura H., Tanaka M., Nakamura Т., Cancer Res., 2000, 60:6737-43], ii) низкомолекулярные ингибиторы сайта связывания АТФ с c-Met, которые блокируют киназную активность [Christensen JG, Schreck R., Burrows J., Kuruganti P., Chan E, Le P., Chen J., Wang X., Ruslim L., Blake R., Lipson KE, Ramphal J., Do S., Cui JJ, Cherrington JM, Mendel DB, Cancer Res. 2003, 63:7345-55], iii) сконструированный полипептид домена SH2, который препятствует доступу к множественному докинг-сайту и РНКи или рибозим, которые снижают экспрессию рецептора или лиганда. Большинство этих подходов проявляют селективное ингибирование c-Met, приводящее в результате к ингибированию опухоли и показывают, что c-Met может представлять интерес для терапевтического вмешательства в рак.
Среди молекул, созданных для направления на c-Met, некоторые являются антителами. Наиболее широко описанным является антитело против c-Met 5D5, полученное фирмой Genentech [WO 96/38557], которое ведет себя как сильный агонист при отдельном добавлении в различных моделях и как антагонист при использовании в виде Fab фрагмента. Моновалентная сконструированная форма этого антитела, описанная как 5D5 с одним плечом (OA5D5) и продуцированная в виде рекомбинантного белка в Е.coli, также является объектом заявки на патент [WO 2006/015371] фирмы Genentech. Однако данная молекула, которую нельзя рассматривать как антитело в связи с ее особым каркасом, также проявляет мутации, которые могли бы быть иммуногенными у людей. Что касается активности, эта не гликозилированная молекула не обладает эффекторными функциями, и, наконец, нет четких данных, демонстрирующих, что OA5D5 ингибирует димеризацию c-Met. Кроме того, при тестировании в модели G55 in vivo на клеточной линии глиобластомы, которая экспрессирует c-Met, но не экспрессирует мРНК и белок HGF, и которая растет независимо от лиганда, анти-c-Met с одним плечом не обладает значимым эффектом на рост опухоли G55, что позволяет предположить, что OA5D5 действует, прежде всего, посредством блокирования связывания HGF и неспособен к направленности на опухоли, активированные независимо от HGF [Martens Т. et al., Clin. Cancer Res., 2006, 12(20):6144-6152].
Другое антитело, направленное на c-Met, описано фирмой Pfizer как антитело, действующее "преимущественно как антагонист c-Met и в некоторых случаях как агонист c-Met" [WO 2005/016382]. Никаких данных, показывающих какой-либо эффект антител Pfizer на димеризацию c-Met, в данной заявке не описано.
Одним из новаторских аспектов настоящего изобретения является создание химерного и/или гуманизированного моноклонального антитела без собственной агонистической активности, и ингибирующего димеризацию c-Met. Более конкретно новаторским аспектом настоящего изобретения является создание химерного и/или гуманизированного моноклонального антитела с антагонистической активностью и ингибированием димеризации c-Met.
В дополнение к направленности на лиганд-зависимые опухоли, данный подход будет также нарушать лиганд-независимые активации c-Met вследствие его гиперэкспрессии или мутаций внутриклеточных доменов, которые остаются зависимыми от олигомеризации для передачи сигнала. Другим аспектом активности этого антитела могло бы быть стерическое затруднение взаимодействия c-Met с его партнерами, которое приведет в результате к нарушению функций c-Met. Это антитело является гуманизированным и сконструировано преимущественно, но не ограниченно этим, как IgG1 человека с приобретением эффекторных функций, таких как АЗКЦ и КЗЦ, в дополнение к функциям, связанным со специфичной блокадой рецептора c-Met.
Авторы изобретения неожиданно впервые смогли создать химерное и/или гуманизированное моноклональное антагонистическое антитело, способное к связыванию с c-Met, но также способное к ингибированию димеризации c-Met, где данное моноклональное антитело является бивалентным в противоположность существующим антагонистическим антителам, направленным против c-Met. Если верно, что, как иногда предполагали на предшествующем уровне техники, антитело, способное к ингибированию димеризации c-Met его партнерами, может представлять интерес, антитело, способное осуществлять это, никогда не было ни описано, ни четко предложено. Кроме того, что касается специфичности антитела, было вовсе неочевидно достижение успеха в создании такого активного бивалентного антитела.
Как было объяснено выше, ингибирование димеризации c-Met является главным аспектом изобретения, поскольку такие антитела представляют действительный интерес для более широкой группы пациентов. Не только лиганд-зависимый, c-Met-активированный рак, как это было до настоящего изобретения, но также лиганд-независимый, c-Met-активированный рак можно лечить антителами, полученными способом по настоящему изобретению.
Антитела оценивали с помощью анализа BRET на клетках, экспрессирующих оба гена c-Met-RLuc/c-Met-YFP, и отбирали на их способность ингибировать по меньшей мере 40%, предпочтительно 45%, 50%, 55% и наиболее предпочтительно 60% сигнала BRET.
Технология BRET известна как репрезентативная для димеризации белков [Angers et al., PNAS, 2000, 97:3684-89].
Технология BRET хорошо известна специалистам в данной области техники и подробно описана в приведенных ниже примерах. Более конкретно BRET (резонансный перенос энергии биолюминесценции, Bioluminescence Resonance Energy Transfer) представляет собой нерадиоактивный перенос энергии, происходящий между биолюминесцентным донором (люциферазой Renilla (Rluc)) и флуоресцентным акцептором, мутантом GFP (зеленого флуоресцентного белка), или YFP (желтым флуоресцентным белком). В настоящем случае использовали EYFP (усиленный желтый флуоресцентный белок). Эффективность переноса зависит от ориентации и расстояния между донором и акцептором. Поэтому перенос энергии может происходить только тогда, когда две молекулы находятся в тесной приближенности (1-10 нм). Это свойство используют для создания анализов взаимодействия между белками. Действительно, чтобы исследовать взаимодействие между двумя партнерами, первый генетическим путем сливают с люциферазой Renilla, а второй с желтым мутантом GFP. Слитые белки обычно но не обязательно экспрессируют в клетках млекопитающих. В присутствии его субстрата, проникающего через мембрану (целентеразина) Rluc испускает синий свет.Если мутант GFP находится ближе, чем на 10 нм от Rluc, может происходить перенос энергии, и можно обнаружить дополнительный желтый сигнал. Сигнал BRET измеряют как отношение между светом, испускаемым акцептором, и светом, испускаемым донором. Таким образом, сигнал BRET возрастает, когда два слитых белка приводят в близкий контакт, либо если конформационное изменение приводит Rluc и мутант GFP в более близкий контакт.
Если анализ BRET составляет предпочтительную форму осуществления, любой способ, известный специалистам в данной области техники, можно использовать для измерения димеризации c-Met. Без ограничения можно упомянуть следующие технологии: FRET (резонансный перенос энергии флуоресценции), HTRF (гомогенная времяпролетная флуоресценция), FLIM (микроскопия на основе измерения времени жизни флуоресценции) или SW-FCCS (одноволновая спектроскопия кросс-корреляции флуоресценции).
Можно также использовать другие классические технологии, такие как совместная иммунопреципитация, альфа-скрининг, химическое сшивание, двойная гибридизация, аффинная хроматография, ELISA или Far Вестерн-блоттинг.
Термины "антитело", "антитела" или "иммуноглобулин" используют взаимозаменяемо в самом широком смысле, и они включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, поливалентные антитела или мультиспецифические антитела (например, биспецифические антитела, если они проявляют желаемую биологическую активность).
Более конкретно такую молекулу составляет гликопротеин, содержащий по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, взаимно соединенные дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области (или домена) тяжелой цепи (в данной заявке сокращенно HCVR или VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (в данной заявке сокращенно LCVR или VL) и константной области легкой цепи. Константная область легкой цепи состоит из двух доменов CL. Области VH и VL можно дополнительно разделить на гипервариабельные участки, называемые участками определения комплементарности (CDR), чередующиеся с участками, которые являются более консервативными, называемыми каркасными участками (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, который связывается с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Тяжелые цепи иммуноглобулинов можно разделить на три функциональных области: область Fd, шарнирную область, и область Fc (кристаллизуемый фрагмент). Область Fd содержит домены VH и СН1 ив комбинации с легкой цепью образует Fab - антигенсвязывающий фрагмент. Фрагмент Fc ответственен за эффекторные функции иммуноглобулина, которые включают, например, фиксацию комплемента и связывание с распознаванием рецепторов Fc эффекторных клеток. Шарнирная область, обнаруженная в иммуноглобулинах классов IgG, IgA и IgD, действует как гибкий спейсер, который дает возможность участку Fab свободно перемещаться в пространстве относительно области Fc. Шарнирные домены являются структурно различными, варьируя как по последовательности, так и по длине среди классов и подклассов иммуноглобулинов.
Согласно кристаллографическим исследованиям шарнирную область иммуноглобулина можно дополнительно подразделить структурно и функционально на три области: верхнюю шарнирную, сердцевину и нижнюю шарнирную (Shin et al., Immunological Reviews 130:87, 1992). Верхняя шарнирная область включает аминокислоты от карбокси-конца СН1 до первого остатка в шарнирной области, который ограничивает движение, обычно до первого остатка цистеина, который образует межцепочечную дисульфидную связь между двумя тяжелыми цепями. Длина верхней шарнирной области коррелирует с сегментной гибкостью антитела. Сердцевинная шарнирная область содержит дисульфидные мостики между тяжелыми цепями. Нижняя шарнирная область соединяет амино-конец домена СН2 и включает его остатки. Сердцевинная шарнирная область IgG1 человека содержит последовательность Cys-Pro-Pro-Cys, которая при димеризации вследствие образования дисульфидной связи приводит в результате к циклическому октапептиду, который считают действующим в качестве поворотной точки, таким образом, придавая гибкость. Конформационные изменения, допустимые структурой, и гибкость полипептидной последовательности шарнирной области иммуноглобулина могут влиять на эффекторные функции участка Fc антитела.
Термин "моноклональное антитело" используют в соответствии с его обычным значением для обозначения антитела, полученного из популяции по существу однородных антител, то есть индивидуальные антитела, составляющие эту популяцию, идентичны за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в минорных количествах. Иными словами, моноклональное антитело состоит из однородного антитела, полученного в результате пролиферации единственного клона клеток (например, клеток гибридомы, эукариотических клеток-хозяев, трансфицированных ДНК, кодирующей однородное антитело, прокариотических клеток-хозяев, трансфицированных ДНК, кодирующей однородное антитело, и т.д.), которое, как правило, характеризуется тяжелыми цепями одного класса и подкласса и легкими цепями одного типа. Моноклональные антитела высоко специфичны, поскольку направлены против одного антигена. Кроме того, в противоположность препаратам поликлональных антител, которые типично включают различные антитела, направленные против различных детерминантов или эпитопов, каждое моноклональное антитело направлено против единственного детерминанта на антигене.
В настоящем описании термины полипептиды, полипептидные последовательности, аминокислотные последовательности, пептиды и белки, присоединенные к соединениям антитела или к их последовательности, взаимозаменяемы.
Изобретение относится к моноклональному антителу или к бивалентному функциональному фрагменту или к его производному, способному ингибировать димеризацию c-Met, и включающему тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 5, 6 или 7, где данное антитело дополнительно характеризуется тем, что оно также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 56.
В предпочтительной форме осуществления настоящее изобретение направлено на моноклональное антитело или его бивалентный функциональный фрагмент или производное, способное ингибировать димеризацию c-Met, где данное антитело включает тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7, где данное антитело также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 56; для применения для предупреждения или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся лиганд-независимой активацией с-Met, предпочтительно раком, также характеризующимся гиперэкспрессией c-Met, где гиперэкспрессия c-Met более предпочтительно является результатом генной амплификации c-Met, и, также более предпочтительно, результатом лиганд-независимой активации c-Met.
В данном аспекте настоящее изобретение включает способ применения для предупреждения или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся лиганд-независимой активацией c-Met, предпочтительно раком, также характеризующимся гиперэкспрессией c-Met, где гиперэкспрессия c-Met более предпочтительно является результатом генной амплификации c-Met, и, также более предпочтительно, результатом лиганд-независимой активации c-Met, где данный способ включает стадию введения композиции, содержащей моноклональное антитело или бивалентный функциональный фрагмент или его производное, способное ингибировать димеризацию c-Met где данное антитело включает тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7, где данное антитело также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 56.
В данном конкретном аспекте и в более предпочтительной форме осуществления рак, характеризующийся лиганд-независимой активацией c-Met, и предпочтительно также характеризующийся гиперэкспрессией c-Met, более предпочтительно являющейся результатом генной амплификации c-Met, и, также более предпочтительно, результатом лиганд-независимой активации c-Met, выбран из группы, состоящей из почечно-клеточной карциномы и рака ЖКТ.
Более конкретно изобретение относится к моноклональному антителу или к бивалентному функциональному фрагменту или к его производному, как описано выше, характеризующемуся тем, что шарнирная область содержит аминокислотную последовательность SEQ ID No. 57.
Иными словами, изобретение относится к моноклональному антителу или к бивалентному функциональному фрагменту или к его производному, способному ингибировать димеризацию c-Met и включающему тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 5, 6 или 7, где данное антитело дополнительно характеризуется тем, что оно также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 57.
Более конкретно изобретение относится к моноклональному антителу или к бивалентному функциональному фрагменту или к его производному, как описано выше, характеризующемуся тем, что оно также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 21.
Иными словами, изобретение относится к моноклональному антителу или к бивалентному функциональному фрагменту или к его производному, способному ингибировать димеризацию c-Met и включающему тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7 или последовательностью, обладающей по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 5, 6 или 7, где данное антитело дополнительно характеризуется тем, что оно также включает шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 21.
Как будет очевидно специалисту в данной области техники, консенсус-последовательности SEQ ID No. 57 и 21 включены в консенсус-последовательность SEQ ID No. 56.
Выражение "функциональные фрагменты и производные" подробно определено ниже в настоящем описании.
Под участками CDR или CDR(s) подразумевают указывать гипервариабельные участки тяжелых и легких цепей иммуноглобулинов, которые определены с помощью IMGT.
Уникальная нумерация IMGT определена для сравнения вариабельных доменов любых рецепторов антигенов, типов цепи или видов [Lefranc M.-P., Immunology Today 18, 509 (1997); Lefranc M.-P., The Immunologist, 7, 132-136 (1999); Lefranc, M.-P., Pommie, С, Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Сотр. Immunol, 27, 55-77 (2003)]. В уникальной нумерации IMGT консервативные аминокислоты всегда имеют одно и то же положение, например, цистеин 23 (1st-CYS), триптофан 41 (CONSERVED-TRP), гидрофобная аминокислота 89, цистеин 104 (2nd-CYS), фенилаланин или триптофан 118 (J-PHE или J-TRP). Уникальная нумерация IMGT обеспечивает стандартизованное определение границ каркасных областей (FR1-IMGT: положения 1-26, FR2-IMGT: 39-55, FR3-IMGT: 66-104 и FR4-IMGT: 118-128) и областей определения комплементарности: CDR1-IMGT: 27-38, CDR2-IMGT: 56-65 и CDR3-IMGT: 105-117. Поскольку гэпы представляют собой незанятые положения, длины CDR-IMGT (показанные между скобками и определенные точками, например, [8.8.13]) становятся критической информацией. Уникальную нумерацию IMGT используют в 2D графических представлениях, обозначенных как IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002); Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)], и в 3D структурах в IMGT/3Dstructure-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., Т cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)].
Существует три CDR тяжелой цепи и 3 CDR легкой цепи. Термин CDR используют в данной заявке с целью указания, в соответствии с ситуацией, одного из трех участков или нескольких, либо даже всех этих участков, которые содержат большинство аминокислотных остатков, ответственных за связывание по сродству антитела к антигену или к эпитопу, который оно распознает.
Под "процентом идентичности" между двумя нуклеиново-кислотными или аминокислотными последовательностями в смысле настоящего изобретения подразумевают указывать процент нуклеотидов или идентичных аминокислотных остатков между двумя сравниваемыми последовательностями, полученный после наилучшего выравнивания (оптимального выравнивания), где этот процент является исключительно статистическим, и различия между двумя последовательностями распределены случайно и по всей их длине. Сравнения последовательностей между двумя нуклеиново-кислотными или аминокислотными последовательностями традиционно осуществляют путем сравнения этих последовательностей после того, как они оптимально выровнены, где данное сравнение можно проводить по сегментам или с помощью "окна сравнения". Оптимальное выравнивание последовательностей для сравнения можно осуществлять, в дополнение к ручному способу, с помощью алгоритма локальной гомологии Smith and Waterman (1981) [Ad. App. Math. 2:482], с помощью алгоритма локальной гомологии Neddleman and Wunsch (1970) [J. Mol. Biol. 48:443], с помощью метода поиска подобий Pearson and Lipman (1988) [Proc. Natl. Acad. Sci. USA 85:2444), с помощью компьютерного программного обеспечения, использующего эти алгоритмы (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI, или еще с помощью программного обеспечения BLAST N или BLAST Р).
Процент идентичности между двумя нуклеиново-кислотными или аминокислотными последовательностями определяют путем сравнения этих двух последовательностей, выровненных оптимально, и в которых сравниваемая нуклеиново-кислотная или аминокислотная последовательность может содержать добавления или делеции по отношению к референсной последовательности для оптимального выравнивания между двумя последовательностями. Процент идентичности вычисляют путем определения числа идентичных положений, для которых нуклеотид или аминокислотный остаток идентичен между двумя последовательностями, деления этого числа идентичных положений на общее число положений в окне сравнения и умножения полученного результата на 100, чтобы получить процент идентичности между двумя последовательностями.
Например, возможно использовать программу BLAST, "BLAST 2 sequences" (Tatusova et al., "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol Lett. 174:247-250), доступную на сайте http://www.ncbi.nlm.nih.gov/ gorf/bl2.html, где используемыми параметрами являются параметры по умолчанию (в частности, параметры "штраф на открытие гэпа": 5 и "штраф на удлинение гэпа": 2; где выбранная матрица представляет собой, например, матрицу "BLOSUM 62", предложенную программой), где процент идентичности между двумя сравниваемыми последовательностями вычисляется непосредственно программой.
В качестве аминокислотной последовательности, обладающей по меньшей мере 80%, предпочтительно 85%, 90%, 95% и 98% идентичностью с референсной аминокислотной последовательностью, предпочтительны те, которые имеют относительно референсной последовательности определенные модификации, в частности, делецию, добавление или замену по меньшей мере одной аминокислоты, укорочение или удлинение. В случае замены одной или более чем одной последовательной или не последовательной аминокислоты предпочтительны замены, при которых замененные аминокислоты заменены "эквивалентными" аминокислотами. Выражение "эквивалентные аминокислоты" в данной заявке предназначено для указания любой аминокислоты, которая может быть заменена любой из аминокислот базовой структуры, но без существенного модифицирования биологических активностей соответствующих антител, и как будет определено ниже, в частности, в примерах. Эти эквивалентные аминокислоты могут быть определены, основываясь либо на их структурной гомологии с аминокислотами, которые они заменяют, либо на результатах сравнительных исследований биологической активности между различными антителами, которые можно осуществить.
Для примера можно упомянуть возможности замены, которую можно осуществить, не приводя в результате к значительному модифицированию биологической активности соответствующего модифицированного антитела.
В качестве не ограничивающего примера в приведенной ниже таблице 2 даны возможности замены, согласованной с сохранением биологической активности модифицированного антитела. Обратные замены, конечно, также возможны при тех же условиях.
В данной заявке должно быть понятно, что изобретение не относится к антителам в природной форме, то есть, что они не находятся в их природном окружении, но что они могут быть выделены или получены путем очистки из природных источников, или, кроме того, получены путем генетической рекомбинации или путем химического синтеза, и что они поэтому могут содержать неприродные аминокислоты, как описано далее.
Должно быть также понятно, как упомянуто выше, что изобретение относится более конкретно к химерному и/или к гуманизированному бивалентному антителу или к любому бивалентному функциональному фрагменту или производному с антагонистической активностью. Бивалентные антитела предшествующего уровня техники являются агонистами или частичными агонистами. Моноклональное антитело по изобретению, включающее модифицированную шарнирную область, как описано выше, то есть включающее шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 56, 57 или 21, является новым и представляет такую особенность, что оно обладает улучшенной антагонистической активностью по сравнению с химерным или гуманизированным антителом 224G11 без такой модифицированной шарнирной области, как будет очевидно из приведенных ниже примеров.
В противоположность предшествующему уровню техники, авторы изобретения получили улучшенную антагонистическую активность без модификации формата антитела. Действительно, на ближайшем уровне техники, представленном антителом 5D5, было необходимо разработать моновалентный фрагмент антитела, чтобы получить антагонистическую активность. В настоящей заявке за счет использования шарнирной области по изобретению возможно впервые получить полноразмерное бивалентное антитело с повышенной антагонистической активностью, и это противоречит общим знаниям.
В предпочтительной форме осуществления антитело по изобретению включает каркасную область, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 22-28 и 58-72, или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностями SEQ ID No. 22-28 и 58-72.
Для лучшей ясности в приведенных ниже таблицах 3 и 4 перегруппированы аминокислотные последовательности и нуклеотидные последовательности различных предпочтительных шарнирных областей по изобретению.
В соответствии с первым подходом антитело будет определено последовательностью его тяжелой цепи. Более конкретно антитело по изобретению или любое из его функциональных фрагментов или производных характеризуется тем, что оно включает тяжелую цепь, содержащую по меньшей мере один CDR, выбранный из CDR, содержащих аминокислотные последовательности SEQ ID No. 1-3.
Упомянутые последовательности приведены ниже:
SEQ ID No. 1: GYIFTAYT
SEQ ID No. 2: IKPNNGLA
SEQ ID No. 3: ARSEITTEFDY
В соответствии с предпочтительным аспектом антитело по изобретению или любое из его функциональных фрагментов или производных включает тяжелую цепь, содержащую по меньшей мере один, предпочтительно два и более предпочтительно три CDR, выбранных из CDR-H1, CDR-H2 и CDR-H3, где:
- CDR-H1 содержит аминокислотную последовательность SEQ ID No. 1
- CDR-H2 содержит аминокислотную последовательность SEQ ID No. 2
- CDR-H3 содержит аминокислотную последовательность SEQ ID No. 3.
При втором подходе антитело теперь будет определено последовательностью его легкой цепи. Более конкретно в соответствии со вторым конкретным аспектом изобретения антитело по изобретению или любое из его функциональных фрагментов или производных характеризуется тем, что оно включает легкую цепь, содержащую по меньшей мере один CDR, выбранный из CDR, содержащих аминокислотную последовательность SEQ ID No. 5-7.
Упомянутые последовательности являются следующими:
SEQ ID No. 5: ESVDSYANSF
SEQ ID No. 6: RAS
SEQ ID No. 7: QQSKEDPLT
В соответствии с другим предпочтительным аспектом антитело по изобретению или любое из его функциональных фрагментов или производных включает тяжелую цепь, содержащую по меньшей мере один, предпочтительно два и более предпочтительно три CDR, выбранных из CDR-L1, CDR-L2 и CDR-L3, где:
- CDR-L1 содержит аминокислотную последовательность SEQ ID No. 5,
- CDR-L2 содержит аминокислотную последовательность SEQ ID No. 6,
- CDR-L3 содержит аминокислотную последовательность SEQ ID No. 7.
Гибридома мыши, способная секретировать моноклональные антитела в соответствии с настоящим изобретением, в частности, гибридома мышиного происхождения, была депонирована в CNCM (Институт Пастера, Париж, Франция) 03/14/2007 под номером CNCM 1-3731.
В настоящем изобретении IgG1 предпочтителен для получения эффекторных функций, и наиболее предпочтительно АЗКЦ и КЗЦ.
Специалисту в данной области техники известно, что эффекторные функции включают, например, связывание Clq; комплементзависимую цитотоксичность (КЗЦ); связывание рецептора Fc; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; и понижающая регуляция рецепторов клеточной поверхности (например, рецептора В-клеток; BCR).
Антитела в соответствии с настоящим изобретением предпочтительно представляют собой специфичные моноклональные антитела, в частности, мышиного, химерного или гуманизированного происхождения, которые могут быть получены в соответствии со стандартными способами, хорошо известными специалистам в данной области техники.
Как правило, для получения моноклональных антител или их функциональных фрагментов или производных, в частности, мышиного происхождения, возможно сделать ссылку на методы, которые описаны, в частности, в руководстве "Antibodies" (Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor NY, pp.726, 1988), или на метод получения из гибридом, описанный Kohler and Milstein (Nature, 256:495-497, 1975).
Моноклональные антитела в соответствии с изобретением могут быть получены, например, из животной клетки, иммунизированной против c-Met или одного из его фрагментов, содержащих эпитоп, специфично распознаваемый моноклональными антителами в соответствии с изобретением. c-Met или одно из его фрагментов можно, в частности, продуцировать в соответствии с обычными рабочими способами, путем генетической рекомбинации, начиная с нуклеиново-кислотной последовательности, содержащейся в последовательности кДНК, кодирующей c-Met, или путем пептидного синтеза, начиная с последовательности аминокислот, содержащейся в пептидной последовательности c-Met.
Моноклональные антитела в соответствии с изобретением могут быть очищены, например, на аффинной колонке, на которой предварительно иммобилизован c-Met или одно из его фрагментов, содержащий эпитоп, специфично распознаваемый моноклональными антителами в соответствии с изобретением. Более конкретно моноклональные антитела могут быть очищены с помощью хроматографии на белке А и/или G, за которой следует или не следует ионообменная хроматография, целью которой является удаление остаточных белковых загрязнений, а также ДНК и ЛПС, за которой следует или не следует эксклюзионная хроматография на геле Сефароза™ с целью удаления потенциальных агрегатов вследствие присутствия димеров или других мультимеров. При более предпочтительном способе можно даже использовать все эти методы одновременно или последовательно.
Антитело по изобретению или его бивалентный функциональный фрагмент или производное предпочтительно составляет химерное антитело.
Под химерным антителом подразумевают указание антитела, которое содержит природную вариабельную область (легкой цепи и тяжелой цепи), имеющую происхождение от антитела данного вида, в комбинации с константными областями тяжелой цепи и легкой цепи антитела вида, гетерологичного по отношению к данному виду (например, мыши, лошади, кролика, собаки, коровы, курицы и т.д.).
Антитела или их фрагменты химерного типа в соответствии с изобретением могут быть получены путем использования методов генетической рекомбинации. Например, химерное антитело можно продуцировать путем клонирования рекомбинантной ДНК, содержащей промотор и последовательность, кодирующую вариабельную область вида, отличного от человека, в частности, мыши, моноклональное антитело в соответствии с изобретением и последовательность, кодирующую константную область антитела человека. Химерное антитело по изобретению, кодируемое таким рекомбинантным геном, будет, например, представлять собой химеру мышь-человек, где специфичность этого антитела определяется вариабельной областью, имеющей происхождение от ДНК мыши, а его изотип определяется константной областью, имеющей происхождение от ДНК человека. Для способов получения химерных антител возможно, например, сделать ссылку на документы Verhoeyn et al. (BioEssays, 8:74, 1988), Morrison et al. (Proc. Natl. Acad. Sci. USA 82:6851-6855, 1984) или US 4816567.
Более конкретно антитело или его функциональный фрагмент или производное содержит химерный вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 46 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 46.
SEQ ID No. 46: EVQLQQSGPELVKPGASVKISCKTSGYIFTAYTMHWVRQSLGESLDWIGGIKPNNGLANYNQKFKGKATLTVDKSSSTAYMDLRSLTSEDSAVYYCARSEITTEFDYWGQGTALTVSS
Более конкретно антитело или его функциональный фрагмент или производное содержит химерный вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 47 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No.47.
SEQ ID No. 47: DIVLTQSPASLAVSLGQRATISCRASESVDSYANSFMHWYQQKPGQPPKLLIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQSKEDPLTFGSGTKLEMKR
Более конкретно предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [IgG2chim], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No.22.
В настоящей заявке использование квадратных скобок не является необходимым, и, в качестве примера, ссылка [224G11] [IgG2chim] должна рассматриваться как идентичная 224G11IgG2chim. Таким же образом, чтобы указать, что антитело представляет собой мышиное антитело, можно добавить выражение murine или букву m; чтобы указать, что антитело представляет собой химерное антитело, можно добавить выражение chim или букву с; и, чтобы указать, что антитело представляет собой гуманизированное антитело, можно добавить выражение hum, hz, Hz или букву h. В качестве примера, химерное антитело 224G11gG2 может быть обозначено как c224G11IgG2, c224G11[IgG2], c[224G11]IgG2, c[224G11][IgG2], 224G11IgG2chim, 224G11[IgG2chim], [224G11]IgG2chim или [224G11][IgG2chim].
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [TH7chim], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
В настоящей заявке ссылка ТН7 должна рассматриваться как идентичная С7Δ6-9 или ТН7С7Δ6-9. Символ Δ означает делецию.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [MHchim], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 23.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [MUP9Hchim], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 26.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [MMCHchim], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 24
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С1], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 58.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С2], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 59.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С3], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 60.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С5], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 61.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С6], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 62.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С7], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 63.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С9], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 64.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [Δ1-3], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 65.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С7Δ6], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 66.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С6Δ9], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 67.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С2Δ5-7], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 68.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С5Δ2-6], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 69.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [С9Δ2-7], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 70.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [Δ5-6-7-8], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 71.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [IgG1/IgG2], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 46, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 47, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 72.
Антитело по изобретению или его бивалентный функциональный фрагмент или производное предпочтительно составляет антитело человека.
Термин "антитело человека" включает все антитела, которые имеют одну или более чем одну вариабельную и константную область, имеющую происхождение от последовательностей иммуноглобулинов человека. В предпочтительной форме осуществления все вариабельные и константные домены (или области) имеют происхождение от последовательности иммуноглобулина человека (полноразмерного антитела человека). Иными словами, оно включает любое антитело, которое имеет вариабельные и константные области (если они присутствуют), имеющие происхождение от иммуноглобулина зародышевой линии человека, то есть, которые имеют аминокислотную последовательность, соответствующую последовательности антитела, продуцируемого человеком, и/или получены с использованием любых методов получения антител человека, известных специалисту в данной области техники.
В одной форме осуществления моноклональные антитела человека получают с помощью гибридомы, которая включает В клетку, полученную от трансгенного животного, не являющегося человеком, например, трансгенной мыши, имеющей геном, содержащий трансген тяжелой цепи и трансген легкой цепи человека, где В клетку сливают с иммортализованной клеткой.
В качестве примера такой трансгенной мыши можно упомянуть XENOMOUSE™, которая представляет собой сконструированную линию мыши, которая содержит большие фрагменты локусов иммуноглобулина человека и дефицитна по продуцированию мышиных антител (Green at al., 1994, Nature Genetics, 7:13-21). Линия XENOMOUSE™ продуцирует спектр, подобный спектру взрослого человека, полноразмерных антител и вырабатывает антиген-специфичные моноклональные антитела человека. Второе поколение линии XENOMOUSE™ содержит примерно 80% спектра антител человека (Green & Jakobovits, 1998, J. Exp. Med., 188:483-495).
Любой другой метод, известный специалисту в данной области техники, такой как метод фагового дисплея, можно также использовать для создания антитела человека в соответствии с изобретением.
Антитело по изобретению или его бивалентный функциональный фрагмент или производное предпочтительно составляет гуманизированное антитело.
Под выражением "гуманизированное антитело" подразумевают указывать антитело, которое содержит участки CDR, имеющие происхождение от антитела не человеческого происхождения, где другие части молекулы антитела имеют происхождение от одного (или от нескольких) антител человека. Кроме того, некоторые из остатков сегментов каркаса (названных FR) могут быть модифицированы с целью сохранения сродства связывания (Jones et al., Nature, 321:522-525, 1986; Verhoeyen et al., Science, 239:1534-1536, 1988; Riechmann et al., Nature, 332:323-327, 1988).
Гуманизированные антитела в соответствии с изобретением или их фрагменты могут быть получены методами, известными специалисту в данной области техники (такими как, например, описаны в документах Singer et al., J. Immun. 150:2844-2857, 1992; Mountain et al., Biotechnol. Genet. Eng. Rev., 10: 1-142, 1992; или Bebbington et al., Bio/Technology, 10:169-175, 1992).
Специалисту в данной области техники известен другой способ гуманизации, такой как, например, способ "прививания CDR", описанный Protein Design Lab (PDL) в заявках на патенты ЕР 0451261, ЕР 0682040, ЕР 09127, ЕР 0566647 или US 5530101, US 6180370, US 5585089 и US 5693761. Можно также упомянуть приведенные ниже заявки на патенты: US 5639641; US 6054297; US 5886152 и US 5877293.
Более конкретно антитело или его функциональный фрагмент или производное содержит гуманизированный вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 4. SEQ ID No. 4: QVQLVQSGAEVKKPGASVKVSCKASGYIFTAYTMHWVRQAPGQGLEWMGWIKPNNGLANYAQKFQGRVTMTRDTSISTAYMELSRLRSDDTAVYYCARSEITTEFDYWGQGTLVTVSS
Более конкретно антитело или его функциональный фрагмент или производное содержит гуманизированный вариабельный домен легкой цепи, выбранный из группы последовательностей, содержащих аминокислотную последовательность SEQ ID No. 8, 9 или 10 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 8, 9 или 10.
SEQ ID No. 8: DIVLTQSPDSLAVSLGERATINCKSSESVDSYANSFMHWYQQKPGQPPKLLIYRASTRESGVPDRFSGSGSRTDFTLTISSLQAEDVAVYYCQQSKEDPLTFGGGTKVEIKR
SEQ ID No. 9: DIVMTQSPDSLAVSLGERATINCKSSESVDSYANSFMHWYQQKPGQPPKLLIYRASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQSKEDPLTFGGGTKVEIKR
SEQ ID No. 10: DIVMTQSPDSLAVSLGERATINCKSSESVDSYANSFLHWYQQKPGQPPKLLIYRASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQSKEDPLTFGGGTKVEIKR
Более конкретно предпочтительное антитело или его бивалентный функциональный фрагмент или производное, в соответствии с изобретением, названное [224G11] [IgG2Hz1], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 8, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное, в соответствии с изобретением, названное [224G11] [IgG2Hz2], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 9, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное, в соответствии с изобретением, названное [224G11] [IgG2Hz3], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 10, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
В другом аспекте предпочтительное антитело, или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [TH7Hz1], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 8, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное, в соответствии с изобретением, названное [224G11] [TH7z2], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 9, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
В другом аспекте предпочтительное антитело или его бивалентный функциональный фрагмент или производное в соответствии с изобретением, названное [224G11] [TH7Hz3], содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No. 4, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID No. 10, и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
В другом аспекте антитела по изобретению могут быть описаны на основании их суммарных тяжелых и легких цепей, соответственно.
В качестве примера, антитело [224G11] [IgG2chim] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 50 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 50, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [TH7chim] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 51 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 51, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С1] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 88 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 88, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С2] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 89 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 89, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С3] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 90 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 90, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С5] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 91 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 91, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С6] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 92 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 92, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С7] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 93 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 93, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С9] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 94 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 94, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [Δ1-3] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 95 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 95, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С7Δ6] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 96 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 96, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С6Δ9] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 97 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 97, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С2Δ5-7] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 98 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 98, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С5Δ2-6] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 99 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 99, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [С9Δ2-7] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 100 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 100, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [Δ5-6-7-8] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 101 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 101, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера антитело [224G11] [IgG1/IgG2] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 102 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 102, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 52 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 52.
В качестве другого примера, антитело [224G11] [IgG2Hz1] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 36 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 36, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 38 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 38.
В качестве другого примера антитело [224G11] [IgG2Hz2] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 36 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 36, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 39 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 39.
В качестве другого примера антитело [224G11] [IgG2Hz3] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 36 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 36, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 40 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 40.
В качестве другого примера антитело [224G11] [TH7Hz1] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 37 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 37, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 38 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 38.
В качестве другого примера, антитело [224G11] [TH7Hz2] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 37 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 37, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 39 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 39.
В качестве другого примера антитело [224G11] [TH7Hz3] по изобретению содержит полноразмерную тяжелую цепь, содержащую аминокислотную последовательность SEQ ID No. 37 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 37, и полноразмерную легкую цепь, содержащую аминокислотную последовательность SEQ ID No. 40 или последовательность, обладающую по меньшей мере 80% идентичностью после оптимального выравнивания с последовательностью SEQ ID No. 40.
Другие примеры антител или их производных в соответствии с изобретением включают полноразмерные тяжелые цепи, содержащие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 88-102 (соответствующими нуклеотидными последовательностями являются SEQ ID No. 103-117).
Под "функциональным фрагментом" антитела в соответствии с изобретением подразумевают указывать, в частности, фрагмент антитела, такой как фрагменты или диатела Fv, scFv (sc для одной цепи), Fab, F(ab’)2, Fab’, scFv-Fc, или любой фрагмент, время полувыведения которого было бы увеличено в результате химической модификации, такой как присоединение поли(алкилен)гликоля, такого как поли(этилен)гликоль ("ПЭГилирование") (пэгилированные фрагменты, называемые Fv-PEG, scFv-PEG, Fab-PEG, F(ab’)2-PEG или Fab’-PEG) ("PEG" означает поли(этилен)гликоль), либо в результате включения в липосому, где эти фрагменты имеют по меньшей мере один из характеристических CDR последовательности SEQ ID No. 1-3 и 5-7 в соответствии с изобретением, и, в частности, способны в целом проявлять даже частичную активность антитела, от которого они получены, такую как, в частности, способность распознавать и связывать c-Met, и, если необходимо, ингибировать активность c-Met.
Предпочтительно эти функциональные фрагменты будут состоять из частичной последовательности или включать частичную последовательность вариабельной области тяжелой или легкой цепи антитела, от которого они образованы, где эта частичная последовательность достаточна для сохранения той же специфичности связывания, как для антитела, от которого она образована, и достаточного сродства, предпочтительно по меньшей мере равного 1/100, более предпочтительно по меньшей мере 1/10, от сродства антитела, от которого она образована, в отношении c-Met. Такой функциональный фрагмент будет содержать как минимум 5 аминокислот, предпочтительно 6, 7, 8, 9, 10, 12, 15, 25, 50 и 100 последовательных аминокислот последовательности антитела, от которого она имеет происхождение.
Предпочтительно эти функциональные фрагменты будут представлять собой фрагменты Fv, scFv, Fab, F(ab’)2, F(ab’), scFv-Fc типа или диатела, которые, как правило, обладают той же специфичностью связывания, что и антитело, от которого они образованы. В более предпочтительной форме осуществления изобретения эти фрагменты выбраны из бивалентных фрагментов, таких как фрагменты F(ab’)2. Согласно настоящему изобретению, фрагменты антител по изобретению могут быть получены, начиная с антител, таких как описано выше, способами, такими как расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных мостиков путем химического восстановления. Другим способом фрагменты антител, включенные в настоящее изобретение, могут быть получены методами генетической рекомбинации, такими как известны специалистам в данной области техники, или, кроме того, путем пептидного синтеза с помощью, например, автоматических синтезаторов пептидов, таких как поставляются фирмой Applied Biosystems, и т.д.
Под "бивалентным фрагментом" должны подразумеваться любые фрагменты антител, содержащие два плеча, и более конкретно фрагменты F(ab’)2.
Под "производными" антитела в соответствии с изобретением подразумевают связывающий белок, содержащий белковый каркас и по меньшей мере один из CDR, выбранный из исходного антитела, с целью сохранения связывающей способности. Такие соединения хорошо известны специалистам в данной области техники и более подробно описаны в последующем описании.
Более конкретно антитело или одно из его функциональных фрагментов или производных в соответствии с изобретением характеризуется тем, что это производное составляет связывающий белок, содержащий каркас, на котором привит по меньшей мере один CDR для сохранения свойств паратопного распознавания исходного антитела.
Одна или несколько последовательностей из всех 6 последовательностей CDR, описанных в изобретении, может быть представлена на белковом каркасе. В этом случае белковый каркас воспроизводит белковую структуру с соответствующей укладкой привитого (привитых) CDR, таким образом, позволяя ему (или им) сохранять их свойства паратопного распознавания антигена.
Специалисту в данной области техники известно, как выбрать белковый каркас, на котором может быть привит по меньшей мере один CDR, выбранный из исходного антитела. Более конкретно известно, что такой каркас для выбора должен проявлять несколько признаков, которые описаны ниже (Skerra A., J. Mol. Recogn., 13, 2000, 167-187):
- хорошую филогенетическую консервативность,
- прочную структуру с хорошо известной трехмерной молекулярной организацией (например, на основании кристаллографии или ЯМР),
- малый размер,
- отсутствие или только низкую степень посттрансляционных модификаций,
- простоту продуцирования, экспрессии и очистки.
Такой белковый каркас может без ограничения представлять собой структуру, выбранную из группы, состоящей из фибронектина и предпочтительно десятого домена фибронектина типа III (FNfn10), липокалина, антикалина (Skerra А., J. Biotechnol, 2001, 74(4):257-75), производного белка Z из домена В стафилококкового белка А, тиоредоксина А или любого белка с повторяющимся доменом, таким как "анкириновый повтор" (Kohl et al., PNAS, 2003, vol. 100, No.4, 1700-1705), "повтор армадилло", "лейцин-богатый повтор" или "тетратрикопептидный повтор".
Можно также упомянуть каркасное производное от токсинов (таких как, например, токсины скорпионов, насекомых, растений или моллюсков) или белковых ингибиторов нейрональной синтазы оксида азота (PIN).
В качестве не ограничивающего примера таких гибридных конструкций можно упомянуть инсерцию CDR-H1 (тяжелой цепи) антитела анти-CD4, то есть антитела 13В8.2, в одну из доступных петель PIN. Связывающие свойства полученного связывающего белка остаются подобными исходному антителу (Bes et al., BBRC 343, 2006, 334-344). Можно также упомянуть прививание CDR-H3 (тяжелой цепи) антитела VHH против лизоцима на петлю неокарциностатина (Nicaise et al., 2004).
Как упомянуто выше, такой белковый каркас может содержать от 1 до 6 CDR из исходного антитела. В предпочтительной форме осуществления, но без какого-либо ограничения, специалист в данной области техники выберет по меньшей мере один CDR из тяжелой цепи, где известно, что эта тяжелая цепь особенно вовлечена в специфичность антитела. Выбор CDR, представляющих интерес, известным способом очевиден специалисту в данной области техники (BES et al., FEBS letters 508, 2001, 67-74).
Как очевидно, эти примеры не являются ограничивающими, и любой другой каркас, известный или описанный, должен быть включен в настоящее описание.
Согласно новому аспекту, настоящее изобретение относится к изолированной нуклеиновой кислоте, характеризующейся тем, что она выбрана из нижеследующих нуклеиновых кислот:
a) нуклеиновой кислоты, ДНК или РНК, кодирующей антитело или одно из его функциональных фрагментов или производных в соответствии с изобретением;
b) нуклеиново-кислотной последовательности, включающей последовательности SEQ ID No. 11, SEQ ID No. 12, SEQ ID No. 13 и последовательности SEQ ID No. 15, SEQ ID No. 16 и SEQ ID No. 17;
c) нуклеиново-кислотной последовательности, включающей последовательности SEQ ID No. 14 и SEQ ID No. 18, 19 или 20;
d) соответствующих РНК-нуклеиновых кислот из нуклеиновых кислот, определенных в b) или с);
e) нуклеиновых кислот, комплементарных нуклеиновым кислотам, определенным в а), b) и с); и
f) нуклеиновой кислоты по меньшей мере из 18 нуклеотидов, способной к гибридизации в условиях высокой жесткости по меньшей мере с одним из CDR последовательности SEQ ID No. 11-13 и 15-17.
В соответствии еще с одним другим аспектом настоящее изобретение относится к изолированной нуклеиновой кислоте, характеризующейся тем, что она выбрана из нижеследующих нуклеиновых кислот:
- нуклеиновой кислоты, ДНК или РНК, кодирующей антитело или одно из его функциональных фрагментов или производных в соответствии с настоящим изобретением, где нуклеиновая последовательность, кодирующая каркасную область данного антитела, содержит или имеет последовательность, выбранную из группы, состоящей из последовательностей SEQ ID No. 29-35 и SEQ ID No. 73-87.
Под нуклеиновой кислотой, нуклеиновой или нуклеиново-кислотной последовательностью, полинуклеотидом, олигонуклеотидом, полинуклеотидной последовательностью, нуклеотидной последовательностью, терминами, которые будут использованы в настоящем изобретении без различия, подразумевают указание на точную связь нуклеотидов, которые являются модифицированными или немодифицированными, позволяющую определить фрагмент или участок нуклеиновой кислоты, содержащую или не содержащую неприродные нуклеотиды, и способную соответствовать как двунитевой ДНК, так и однонитевой ДНК, а также продуктам транскрипции этих ДНК.
В данной заявке также должно быть понятно, что настоящее изобретение не относится к нуклеотидным последовательностям в их природном хромосомном окружении, то есть в естественном состоянии. Оно относится к последовательностям, которые изолированы и/или очищены, то есть они выбраны прямо или косвенно, например, путем копирования, из их окружения, являясь по меньшей мере частично модифицированными. Таким образом, они предназначены в данной заявке для указания изолированных нуклеиновых кислот, полученных путем генетической рекомбинации посредством, например, клеток-хозяев или полученных путем химического синтеза.
Гибридизация в условиях высокой жесткости означает, что температурные условия и условия ионной силы выбраны таким образом, что дают возможность поддержания гибридизации между двумя фрагментами комплементарной ДНК. В качестве иллюстрации, условия высокой жесткости стадии гибридизации в целях определения полинуклеотидных фрагментов, описанных выше, предпочтительно являются такими, как описано ниже.
Гибридизацию ДНК-ДНК или ДНК-РНК осуществляют в две стадии: (1) прегибридизация при 42°С в течение 3 часов в фосфатном буфере (20 мМ, рН 7,5), содержащем 5×SSC (1×SSC соответствует раствору 0,15 М NaCl + 0,015 М цитрата натрия), 50% формамида, 7% додецилсульфата натрия (ДСН), 10х раствор Денхардта, 5% декстрансульфата и 1% ДНК спермы лосося; (2) собственно гибридизация в течение 20 часов при температуре в зависимости от размера зонда (т.е.: 42°С для размера зонда>100 нуклеотидов) с последующими 2 отмывками по 20 минут при 20°С в 2×SSC+2% ДСН, 1 отмывкой 20 минут при 20°С в 0,1×SSC+0,1% ДСН. Последнюю отмывку проводят в 0,1×SSC+0,1% ДСН в течение 30 минут при 60°С для размера зонда >100 нуклеотидов. Условия гибридизации высокой жесткости, описанные выше для полинуклеотида определенного размера, могут быть адаптированы специалистом в данной области техники для олигонуклеотидов большего или меньшего размера, в соответствии с теорией Sambrook et al. (1989, Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor).
Подобным образом, изобретение относится к вектору, содержащему нуклеиновую кислоту в соответствии с настоящим изобретением.
Изобретение направлено, в частности, на клонирующие и/или экспрессионные векторы, которые содержат нуклеотидную последовательность в соответствии с изобретением.
Векторы в соответствии с изобретением предпочтительно содержат элементы, которые дают возможность экспрессии и/или секреции транслированных нуклеотидных последовательностей в определенной клетке-хозяине. Вектор должен, таким образом, содержать промотор, сигналы инициации и терминации трансляции, а также соответствующие участки регуляции транскрипции. Он должен обладать способностью стабильно поддерживаться в клетке-хозяине и может необязательно иметь конкретные сигналы, которые обозначают секрецию транслированного белка. Эти различные элементы выбирает и оптимизирует специалист в данной области техники в зависимости от используемой клетки-хозяина. С этой целью нуклеотидные последовательности в соответствии с изобретением могут быть встроены в автономно реплицирующиеся векторы в выбранном хозяине или представлять собой интегративные векторы выбранного хозяина.
Такие векторы получают способами, используемыми в настоящее время специалистами в данной области техники, и полученные в результате клоны можно вводить в соответствующего хозяина стандартными способами, такими как липофекция, электропорация, термошок или химические методы.
Векторы в соответствии с изобретением представляют собой, например, векторы плазмидного или вирусного происхождения. Они полезны для трансформации клеток-хозяев с целью клонирования или экспрессии нуклеотидных последовательностей в соответствии с изобретением.
Изобретение, таким образом, включает клетки-хозяева, трансформированные вектором в соответствии с изобретением или содержащие его.
Клетка-хозяин может быть выбрана из прокариотических или эукариотических систем, например, бактериальных клеток, но также дрожжевых клеток или животных клеток, в частности, клеток млекопитающих. Подобным образом, возможно использовать клетки насекомых или клетки растений.
Изобретение, таким образом, относится к животным, за исключением человека, которые содержат по меньшей мере одну клетку, трансформированную в соответствии с изобретением.
Согласно другому аспекту, объектом изобретения является способ получения антитела или одного из его функциональных фрагментов в соответствии с изобретением, характеризующийся тем, что он включает приведенные ниже стадии:
a) культивирование в среде и в соответствующих условиях культивирования клетки-хозяина в соответствии с изобретением; и
b) выделение антител или одного из их функциональных фрагментов, продуцированных таким путем, начиная с культуральной среды или культивируемых клеток.
Клетки, трансформированные в соответствии с изобретением, можно использовать в способах получения рекомбинантных полипептидов в соответствии с изобретением. Сами способы получения полипептида в соответствии с изобретением в рекомбинантной форме, характеризующиеся тем, что они используют вектор и/или клетку, трансформированную вектором в соответствии с изобретением, включены в настоящее изобретение. Предпочтительно клетку, трансформированную вектором в соответствии с изобретением, культивируют в условиях, которые дают возможность экспрессии этого полипептида, и выделяют рекомбинантный пептид.
Как указано, клетка-хозяин может быть выбрана из прокариотических или эукариотических систем. В частности, возможно идентифицировать нуклеотидные последовательности в соответствии с изобретением, способствующие секреции в такой прокариотической или эукариотической системе. Вектор в соответствии с изобретением, несущий такую последовательность, можно, следовательно, предпочтительно использовать для продуцирования рекомбинантных белков, предназначенных для секреции. В результате очистка этих представляющих интерес рекомбинантных белков будет облегчена за счет того факта, что они присутствуют в супернатанте клеточной культуры вероятнее, чем внутри клеток-хозяев.
Подобным образом, возможно получить полипептиды в соответствии с изобретением путем химического синтеза. Такой способ получения также является объектом изобретения. Специалисту в данной области техники известны способы химического синтеза, например, методы, в которых используют твердые фазы [Steward et al., 1984, Solid phase peptide synthesis, Pierce Chem. Company, Rockford, 111, 2nd ed., (1984)], или методы, использующие частичные твердые фазы, путем конденсации фрагментов или путем классического синтеза в растворе. Полипептиды, полученные путем химического синтеза и способные содержать соответствующие неприродные аминокислоты, также включены в изобретение.
Антитела или одно из их функциональных фрагментов или производных, которые могут быть получены способом в соответствии с изобретением, также включены в настоящее изобретение.
Изобретение также относится к антителу по изобретению в качестве лекарства.
Изобретение также относится к фармацевтической композиции, содержащей в качестве активного вещества соединение, состоящее из антитела или одного из его функциональных фрагментов в соответствии с изобретением, предпочтительно смешанное с эксципиентом и/или фармацевтически приемлемым носителем.
Другую дополнительную форму осуществления изобретения составляет композиция, как описано выше, которая содержит, кроме того, в виде комбинированного препарата для одновременного, отдельного или последовательного применения противоопухолевое антитело.
Наиболее предпочтительно второе противоопухолевое антитело может быть выбрано из антител против IGF-IR, против EGFR, против HER2/neu, против VEGFR, против VEGF и т.д., или из любых других противоопухолевых антител, известных специалисту в данной области техники. Очевидно, что применение в качестве второго антитела функциональных фрагментов или производных вышеупомянутых антител составляет часть изобретения.
В качестве наиболее предпочтительного антитела выбраны антитела против EGFR, как, например, антитело С225 (Эрбитукс).
"Одновременное применение" понимают как означающее введение двух соединений композиции в соответствии с изобретением в одной или в идентичной фармацевтической форме.
"Отдельное применение" понимают как означающее введение в одно и то же время двух соединений композиции в соответствии с изобретением в отдельных фармацевтических формах.
"Последовательное применение" понимают как означающее последовательное введение двух соединений композиции в соответствии с изобретением, каждое в отдельной фармацевтической форме.
Как правило, композиция в соответствии с изобретением значительно повышает эффективность лечения рака. Иными словами, терапевтический эффект антител против c-Met в соответствии с изобретением неожиданно потенцируется введением цитотоксического агента. Другое основное следующее преимущество, которое дает композиция в соответствии с изобретением, касается возможности применения более низких эффективных доз активного вещества, что позволяет избежать риска появления вторичных эффектов, в частности, эффектов цитотоксического агента, или уменьшить его.
Кроме того, данная композиция в соответствии с изобретением дала бы возможность получить желаемый терапевтический эффект быстрее.
Композиция по изобретению может также характеризоваться тем, что она содержит, кроме того, в виде комбинированного препарата для одновременного, отдельного или последовательного применения цитотоксический/цитостатический агент.
Под "противораковыми терапевтическими агентами" или "цитотоксическими/цитостатическими агентами" подразумевают вещество, которое при введении субъекту лечит или предупреждает развитие рака в организме субъекта. В качестве не ограничивающего примера таких агентов можно упомянуть алкилирующие агенты, антиметаболиты, противоопухолевые антибиотики, митотические ингибиторы, ингибиторы функции хроматина, антиангиогенные агенты, антиэстрогены, антиандрогены или иммуномодуляторы.
Такие агенты, например, указаны в 2001 издании VIDAL на странице, посвященной соединениям, относящимся к колонке канцерологии и гематологии "Цитотоксические агенты", где эти цитотоксические соединения, указанные со ссылкой на этот документ, указаны в данной заявке как предпочтительные цитотоксические агенты.
Более конкретно приведенные ниже агенты предпочтительны в соответствии с изобретением.
"Алкилирующий агент" относится к любому веществу, которое может сшивать или алкилировать любую молекулу, предпочтительно нуклеиновую кислоту (например, ДНК), внутри клетки. Примеры алкилирующих агентов включают азотистый иприт, такой как меклоретамин, хлорамбукол, мелфален, хлоридрат, пипобромен, преднимустин, динатрия фосфат или эстрамустин; оксазофорины, такие как циклофосфамид, алтретамин, трофосфамид, сульфофосфамид или ифосфамид; азиридины или иминэтилены, такие как тиотепа, триэтиленамин или алтретамин; нитрозомочевины, такие как кармустин, стрептозоцин, фотемустин или ломустин; алкилсульфонаты, такие как бусульфан, треосульфан или импросульфан; триазены, такие как дакарбазин; или комплексы платины, такие как цисплатин, оксалиплатин и карбоплатин.
"Антиметаболиты" относятся к веществам, которые блокируют клеточный рост и/или метаболизм посредством вмешательства в некоторые активности, обычно в синтез ДНК. Примеры антиметаболитов включают метотрексат, 5-фторурацил, флоксуридин, 5-фтордезоксиуридин, капецитабин, цитарабин, флударабин, цитозин арабинозид, 6-меркаптопурин (6-МР), 6-тиогуанин (6-TG), хлордезоксиаденозин, 5-азацитидин, гемцитабин, кладрибин, дезоксикоформицин и пентостатин.
"Противоопухолевые антибиотики" относятся к соединениям, которые могут предупреждать или ингибировать синтез ДНК, РНК и/или белка. Примеры противоопухолевых антибиотиков включают доксорубицин, даунорубицин, идарубицин, валрубицин, митоксантрон, дактиномицин, митрамицин, пликамицин, митомицин С, блеомицин и прокарбазин.
"Митотические ингибиторы" предотвращают нормальное прохождение клеточного цикла и митоза. Как правило, ингибиторы микротрубочек или таксоиды, такие как паклитаксел и доцетаксел, способны к ингибированию митоза. Винка-алкалоиды, такие как винбластин, винкристин, виндезин и винорельбин, также способны к ингибированию митоза.
"Ингибиторы функции хроматина" или "ингибиторы топоизомераз" относятся к веществам, которые ингибируют нормальную функцию белков моделирования хроматина, таких как топоизомераза I или топоизомераза II. Примеры ингибиторов функции хроматина включают для топоизомеразы I камптотецин и его производные, такие как топотекан или иринотекан, и для топоизомеразы II этопозид, этопозид фосфат и тенипозид.
"Антиангиогенный агент" относится к любому лекарству, соединению, веществу или агенту, которые ингибируют рост кровеносных сосудов. Примеры антиангиогенных агентов включают, но никоим образом не ограничены ими, разоксин, маримастат, батимастат, приномастат, таномастат, иломастат, CGS-27023А, галофунгинон, COL-3, неовастат, BMS-275291, талидомид, CDC 501, DMXAA, L-651582, скваламин, эндостатин, SU5416, SU6668, интерферон-альфа, EMD121974, интерлейкин-12, IM862, ангиостатин и витаксин.
"Антиэстроген" или "антиэстрогенный агент" относится к любому веществу, которое снижает, антагонизирует или ингибирует действие эстрогена. Примерами антиэстрогенных агентов являются тамоксифен, торемифен, ралоксифен, дролоксифен, йодоксифен, анастрозол, летрозол и экземестан.
"Антиандрогены" или "антиандрогенные агенты" относятся к любому веществу, которое снижает, антагонизирует или ингибирует действие андрогена. Примерами антиандрогенов являются флутамид, нилутамид, бикалутамид, спиронолактон, ципротерон ацетат, финастерид и цимитидин.
"Иммуномодуляторы" представляют собой вещества, которые стимулируют иммунную систему.
Примеры иммуномодуляторов включают интерферон, интерлейкин, такой как альдеслейкин, ОСТ-43, дерилейкин дифлитокс и интерлейкин-2, факторы некроза опухоли, такие как тазонермин, или другие иммуномодуляторы, такие как лентинан, сизофиран, роквинимекс, пидотимод, пегадемаза, тимопентин, поли I:С или левамизол в сочетании с 5-фторурацилом.
Для дополнительных подробностей специалист в данной области техники может сослаться на руководство, изданное "Association Francaise des Enseignants de Chimie Therapeutique" и озаглавленное "Traite de chimie therapeutique", т.6, Medicaments antitumoraux et perspectives dans le traitement des cancers, издательство ТЕС & DOC, 2003.
В качестве химических агентов или цитотоксических агентов можно также упомянуть все ингибиторы киназ, такие как, например, гефитиниб или эрлотиниб.
В особенно предпочтительной форме осуществления композиция в виде комбинированного препарата в соответствии с изобретением характеризуется тем, что цитотоксический агент сочетают химическим путем с антителом для одновременного применения.
Чтобы способствовать сочетанию между цитотоксическим агентом и антителом в соответствии с изобретением, в частности, возможно ввести спейсерные молекулы между двумя соединениями, которые нужно подвергать сочетанию, такие как поли(алкилен)гликоли, такие как полиэтиленгликоль, или, кроме того, аминокислоты, или в другой форме осуществления использовать активные производные цитотоксических агентов, в которые были бы введены функциональные группы, способные взаимодействовать с антителом в соответствии с изобретением. Эти методы сочетания хорошо известны специалистам в данной области техники и не будут широко описаны в настоящем описании.
Изобретение относится в другом аспекте к композиции, характеризующейся тем, что по меньшей мере одно из антител или одно из их функциональных фрагментов или производных конъюгируют с клеточным токсином и/или с радиоактивным элементом.
Предпочтительно токсин или радиоактивный элемент способен к ингибированию по меньшей мере одной клеточной активности клеток экспрессирующих c-Met, более предпочтительно способных к предотвращению роста или пролиферации этой клетки, в частности, к полной инактивации этой клетки.
Также предпочтительно токсин представляет собой энтеробактериальный токсин, в частности, экзотоксин A Pseudomonas.
Радиоактивные элементы (или радиоизотопы), предпочтительно конъюгированные с антителами, применяемыми для терапии, представляют собой радиоизотопы, которые испускают гамма-лучи, и предпочтительно йод131, иттрий90, золото199, палладий100, медь67, висмут217 и сурьму211. Радиоизотопы, которые испускают бета- и альфа-лучи, можно также использовать для терапии.
Под токсином или радиоактивным элементом, конъюгированным по меньшей мере с одним антителом или с одним из его функциональных фрагментов в соответствии с изобретением подразумевают указывать любые средства, дающие возможность токсину или радиоактивному элементу связаться по меньшей мере с одним антителом, в частности, посредством ковалентного сочетания между двумя соединениями, с введением или без введения линкерной молекулы.
Среди агентов, дающих возможность химического (ковалентного), электростатического или нековалентного связывания всех или части компонентов конъюгата, можно, в частности, упомянуть бензохинон, карбодиимид и более конкретно ЭДК (1-этил-3-[3-диметиламинопропил]-карбодиимида гидрохлорид), дималеимид, дитиобис-нитробензойную кислоту (ДТНВ), N-сукцинимидил-S-ацетилтиоацетат (САТА), агенты, образующие мостиковую связь, имеющие одну или более чем одну фенилазидную группу, взаимодействующую с ультрафиолетовым светом (УФ), и предпочтительно N-[-4-(азидосалициламино)бутил]-3’-(2’-пиридилдитио)-пропионамид (АПДП), N-сукцинимидил-3-(2-пиридилтиопропионат (СПДП), 6-гидразиноникотинамид (HYNIC).
Другую форму сочетания, особенно для радиоактивных элементов, может составлять использование бифункционального ионного хелатирующего агента.
Среди этих хелатирующих агентов возможно упомянуть хелатирующие агенты, образованные из ЭДТА (этилендиаминтетрауксусной кислоты) или из ДТПА (диэтилентриаминпентауксусной кислоты), которые разработаны для связывания металлов, в частности, радиоактивных металлов, и иммуноглобулинов. Так, ДПТА и ее производные могут быть замещены различными группами на углеродной цепи с целью повышения стабильности и прочности комплекса лиганд-металл (Krejcarek et al. (1977); Brechbiel et al. (1991); Gansow (1991); патент США 4831175).
Например, диэтилентриаминпентауксусная кислота (ДТПА) и ее производные, которые широко использовались в медицине и в биологии в течение длительного времени либо в их свободной форме, либо в форме комплекса с ионом металла, обладают примечательным характерным свойством образования стабильных хелатов с ионами металлов и возможности сочетания с белками, представляющими терапевтический или диагностический интерес, такими как антитела, для разработки радиоиммуноконъюгатов в терапии рака (Meases et al., 1984; Gansow et al., 1990).
Также предпочтительно по меньшей мере одно антитело, образующее конъюгат в соответствии с изобретением, выбрано из его функциональных фрагментов, в частности, фрагментов, укороченных по их Fc компоненту, таких как фрагменты scFv.
Как уже упомянуто, в предпочтительной форме осуществления изобретения цитотоксический/цитостатический агент токсина и/или радиоактивного элемента подвергают сочетанию химическим путем по меньшей мере с одним из элементов композиции для одновременного применения.
Настоящее изобретение включает описанную композицию в качестве лекарства.
Настоящее изобретение, кроме того, включает применение композиции в соответствии с изобретением для получения лекарства.
В другом аспекте изобретение относится к применению антитела или одного из его функциональных фрагментов или производных и/или композиции, как описано выше, для получения лекарства, предназначенного для ингибирования роста и/или пролиферации опухолевых клеток.
Другой аспект изобретения состоит в применении антитела или одного из его функциональных фрагментов или производных и/или композиции, как описано выше или в вышеупомянутом применении, для получения лекарства, предназначенного для предупреждения или для лечения рака.
В настоящее изобретение также включен способ, предназначенный для ингибирования роста и/или пролиферации опухолевых клеток у пациента, включающий введение пациенту, нуждающемуся в этом, антитела или одного из его функциональных фрагментов или производных в соответствии с изобретением, антитела, продуцированного гибридомой в соответствии с изобретением или композиции в соответствии с изобретением.
Настоящее изобретение, кроме того, включает способ предупреждения или лечения рака у пациента, нуждающегося в этом, включающий введение пациенту антитела или одного из его функциональных фрагментов или производных в соответствии с изобретением, антитела, продуцированного гибридомой в соответствии с изобретением или композиции в соответствии с изобретением.
В особенно предпочтительном аспекте рак представляет собой рак, выбранный из рака простаты, остеосарком, рака легкого, рака молочной железы, рака эндометрия, глиобластомы или рака ободочной кишки.
Как объяснено выше, преимущество изобретения состоит в том, что оно дает возможность лечения HGF-зависимых и независимых раков, связанных с активацией Met.
В конкретном аспекте изобретение включает применение антитела или одного из его функциональных фрагментов или производных и/или композиции, как описано выше, или применение, упомянутое выше, для получения лекарства, предназначенного для предупреждения или для лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся гиперэкспрессией c-Met (например, вследствие генной амплификации c-Met), приводящей в результате к лиганд-независимой конститутивной активации рецептора c-Met.
Предпочтительно этот рак характеризуется генной амплификацией c-Met, приводящей в результате к лиганд-независимой конститутивной активации рецептора c-Met, выбран из группы, состоящей из почечно-клеточной карциномы и рака ЖКТ.
Изобретение еще в одном другом аспекте охватывает способ диагностики in vitro заболевания, индуцированного гиперэкспрессией или пониженной экспрессией рецептора c-Met, начиная с биологического образца, в котором подозревают аномальное присутствие рецептора c-Met, где этот способ характеризуется тем, что он включает стадию, где биологический образец приводят в контакт с антителом по изобретению, где для данного антитела существует возможность, если необходимо, мечения.
Предпочтительно заболевание, связанное с аномальным присутствием рецептора c-Met при данном способе диагностики представляет собой раки.
Антитело или один из его функциональных фрагментов может присутствовать в форме иммуноконъюгата или меченого антитела, так чтобы получить обнаружимый и/или количественно определяемый сигнал.
Антитела, меченые в соответствии с изобретением, или их функциональные фрагменты включают, например, антитела, названные иммуноконъюгатами, которые могут быть конъюгированы, например, с ферментами, такими как пероксидаза, щелочная фосфатаза, бета-D-галактозидаза, глюкозооксидаза, глюкозоамилаза, карбоновая ангидраза, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо-6-фосфатдегидрогеназа, или с молекулой, такой как биотин, дигоксигенин или 5-бромдезоксиуридин. Флуоресцентные метки можно подобным образом конъюгировать с антителами или с их функциональными фрагментами в соответствии с изобретением, и они, в частности, включают флуоресцеин и его производные, флуорохром, родамин и его производные, GFP (GFP означает "зеленый флуоресцентный белок"), дансил, умбеллиферон и т.д. В таких конъюгатах антитела по изобретению или их функциональные фрагменты могут быть получены способами, известными специалистам в данной области техники. Их можно подвергать сочетанию с ферментами или с флуоресцентными метками непосредственно или при посредничестве спейсерной группы или связывающей группы, например, полиальдегида, такого как глутаральдегид, этилендиаминтетрауксусной кислоты (ЭДТА), диэтилентриаминпентауксусной кислоты (ДПТА) или в присутствии агентов сочетания, таких как упомянутые выше для терапевтических конъюгатов. Конъюгаты, содержащие метки флуоресцеинового типа, могут быть получены путем взаимодействия с изотиоцианатом.
Другие конъюгаты могут, подобным образом, включать хемилюминесцентные метки, такие как люминол и диоксетаны, биолюминесцентные метки, такие как люцифераза и люциферин, или, кроме того, радиоактивные метки, такие как йод123, йод125, йод126, йод133, бром77, технеций99m, индий111, индий113m, галлий67, галлий68, рутений95, рутений97, рутений103, рутений105, ртуть107, ртуть203, рений99m, рений101, рений105, скандий47, теллур121m, теллур122m, теллур125m, тулий165, тулий167, тулий168, фтор18, иттрий199, йод131. Способы, известные специалистам в данной области техники, существующие для сочетания терапевтических радиоактивных изотопов с антителами, либо непосредственно, либо посредством хелатирующего агента, такого как ЭДТА, ДТПА, упомянутые выше, можно использовать для радиоактивных элементов, которые можно применять при диагностике. Подобным образом, возможно упомянуть мечение Na[I125] методом хлорамина Т [Hunter W.M. and Greenwood F.C. (1962) Nature 194:495] или, кроме того, технецием99m методом Crockford et al. (патент США 4424200) или присоединеным через ДТПА, как описано Hnatowich (патент США 4479930).
Таким образом, антитело или его функциональный фрагмент или производное в соответствии с изобретением можно применять в способе обнаружения и/или количественного определения гиперэкспрессии или сниженной экспрессии, предпочтительно гиперэкспрессии, рецептора c-Met в биологическом образце, характеризующемся тем, что он включает приведенные ниже стадии:
а) приведение биологического образца в контакт с антителом или его функциональным фрагментом или производным в соответствии с изобретением; и
b) демонстрация возможно образовавшегося комплекса c-Met/антитело.
В конкретной форме осуществления антитело или его функциональный фрагмент или производное в соответствии с изобретением можно применять в способе обнаружения и/или количественного определения рецептора c-Met в биологическом образце для мониторинга эффективности профилактического и/или терапевтического лечения c-Met-зависимого рака.
Более обобщенно, антитело или его функциональный фрагмент или производное в соответствии с изобретением можно предпочтительно применять в любой ситуации, где экспрессию рецептора c-Met необходимо наблюдать качественно и/или количественно.
Предпочтительно биологический образец образован биологической жидкостью, такой как сыворотка, цельная кровь, клетки, образец ткани или биопсии человеческого происхождения.
Любую методику или общепринятый тест можно использовать с целью проведения такого обнаружения и/или дозирования. Этот тест может представлять собой конкурентный или сэндвич-тест или любой тест, известный специалисту в данной области техники, в зависимости от образования иммунного комплекса типа антитело-антиген. Следуя применениям в соответствии с изобретением, антитело или его функциональный фрагмент или производное может быть иммобилизованным или меченым. Эту иммобилизацию можно осуществлять на различных подложках, известных специалисту в данной области техники. Эти подложки могут, в частности, включать стекло, полистирол, полипропилен, полиэтилен, декстран, нейлон, либо природные или модифицированные клетки. Эти подложки могут быть либо растворимыми, либо нерастворимыми.
В качестве примера, в предпочтительном способе задействованы иммуноферментные способы в соответствии с методом ELISA, способы иммунофлуоресценции или методы радиоиммунологического анализа (РИА), либо эквивалентные.
Таким образом, настоящее изобретение также включает наборы или комплекты, необходимые для осуществления способа диагностики заболеваний, индуцированных гиперэкспрессией или сниженной экспрессией рецептора c-Met, или для осуществления способа обнаружения и/или количественного определения гиперэкспрессии или сниженной экспрессии рецептора c-Met в биологическом образце, предпочтительно гиперэкспрессии этого рецептора, характеризующиеся тем, что данный набор или комплект включает приведенные ниже элементы:
а) антитело или его функциональный фрагмент или производное в соответствии с изобретением;
b) необязательно реагенты для образования среды, благоприятной для иммунологической реакции;
c) необязательно реагенты, дающие возможность демонстрации комплексов c-Met/антитело, полученных в результате иммунологической реакции.
Объектом изобретения также является применение антитела или композиции в соответствии с изобретением для получения лекарства, предназначенного для специфичной направленности биологически активного соединения в клетки, экспрессирующие или гиперэкспрессирующие рецептор c-Met.
Под биологически активным соединением в данной заявке подразумевают указание на любое соединение, способное к модулированию, в частности, к ингибированию активности клеток, в частности, их роста, их пролиферации, транскрипции или трансляции генов.
Объектом изобретения также является реагент для диагностики in vivo, включающий антитело в соответствии с изобретением или его функциональный фрагмент или его производное, предпочтительно меченое, в частности, радиоактивно меченое, и его применение в медицинской визуализации, в частности, для обнаружения рака, связанного с экспрессией или гиперэкспрессией клеткой рецептора c-Met.
Изобретение также относится к композиции в виде комбинированного препарата или к конъюгату антитела против c-Met/токсина или радиоактивного элемента в соответствии с изобретением в качестве лекарства.
Предпочтительно композицию в виде комбинированного препарата или конъюгат в соответствии с изобретением будут смешивать с эксципиентом и/или с фармацевтически приемлемым носителем.
В настоящем описании фармацевтически приемлемый носитель предназначен для указания соединения или комбинации соединений, входящих в фармацевтическую композицию, не вызывающих вторичных реакций, которые дают возможность, например, облегчения введения активного соединения (соединений), повышения его времени жизни и/или его эффективности в организме, повышения его растворимости в растворе или, кроме того, улучшение его консервации. Эти фармацевтически приемлемые носители хорошо известны и будут адаптированы специалистом в данной области техники в зависимости от природы и от выбранного режима введения активного соединения (соединений).
Предпочтительно эти соединения будут вводить системным путем, в частности, внутривенным путем, внутримышечным, внутрикожным, внутрибрюшинным или подкожным путем, либо пероральным путем. Более предпочтительно композицию, содержащую антитела в соответствии с изобретением, будут вводить несколько раз последовательным способом.
Режимы их введения, дозировки и оптимальные фармацевтические формы могут быть определены в соответствии с критериями, обычно учитываемыми при установлении терапии, адаптированной для пациента, такими как, например, возраст или масса тела пациента, серьезность его/ее общего состояния, переносимость лечения и отмеченные вторичные эффекты.
Другие характеристики и преимущества изобретения видны в продолжении описания с помощью примеров и графических материалов, где:
Фиг.1: Эффект нерелевантных IgG1 Mab мышиного и человеческого происхождения и ФСБ на фосфорилирование рецептора c-Met на клетках А549.
Фиг.2А и 2В: Эффект мышиного и гуманизированного 224G11 Mab, продуцируемого как IgG1 человека/изотип каппа, на фосфорилирование рецептора c-Met на клетках А549.
Фиг.2А: агонистический эффект, вычисленный в виде процента против максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.2В: антагонистический эффект, вычисленный в виде процента ингибирования максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.3А и 3В: Сравнение между мышиным 224G11 Mab и химерными 224G11 Mab, содержащими различные сконструированные шарнирные области, на фосфорилирование рецептора c-Met на клетках А549.
Фиг.3А: агонистический эффект, вычисленный в виде процента против максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.3В: антагонистический эффект, вычисленный в виде процента ингибирования максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.4А и 4В: Сравнение между мышиным 224G11 Mab и химерным и гуманизированным 224G11 Mab, продуцируемыми как IgG2 человека/изотип каппа, на фосфорилирование рецептора c-Met на клетках А549.
Фиг.4А: агонистический эффект, вычисленный в виде процента против максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.4В: антагонистический эффект, вычисленный в виде процента ингибирования максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.5А и 5В: Сравнение между мышиным 224G11 Mab и химерным и гуманизированным 224G11 Mab, полученными как сконструированный мутант шарнирной области TH7IgG1/каппа на фосфорилирование рецептора c-Met на клетках А549.
Фиг.5А: агонистический эффект, вычисленный в виде процента против максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.5В: антагонистический эффект, вычисленный в виде процента ингибирования максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.6А и 6В, Фиг.7А и 7В, Фиг.8А и 8В, Фиг.9А и 9В, Фиг.10А и 10В: модели BRET, где Фиг. А: модель димеризации c-Met; и Фиг. В: модель активации c-Met.
Фиг.11: распознавание c-Met химерной и гуманизированной формой 224G11.
Фиг.12: Эффект мышиных и химерных антител на индуцированную HGF пролиферацию клеток NCI-H441 in vitro. Клетки NCI-H441 высевали в среду без сыворотки. Через 24 часа после высевания добавляли m224G11 и [224G11]chim либо в отсутствие, либо в присутствии HGF. Черными стрелками указаны лунки, засеянные одними клетками либо в отсутствие ↙, либо в присутствии ↘ HGF. IgG1 (mIgG1) вводили в качестве контроля изотипа.
Фиг.13: Сравнение in vivo мышиного и химерного с IgG1 224G11 Mab на модели ксенотрансплантата NCI-H441.
Фиг.14А и 14В: Эффект мышиного 224G11 Mab и различных химерных и гуманизированных вариантов этого антитела на индуцированную HGF пролиферацию клеток NCI-H441 in vitro. Клетки NCI-H441 высевали в среду без сыворотки. Через двадцать четыре часа после высевания добавляли тестируемое антитело либо в отсутствие, либо в присутствии HGF. На панели (Фиг.14А) показаны мышиный m224G11, химерный IgG1 [224G11]chim, гуманизированные IgG1 [224G11] [Hz1], [224G11] [Hz2], [224G11] [Hz3] варианты. На панели (Фиг.14В) представлены мышиная m224G11 и различные химерные формы IgG1 ([224G11] chim, [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [ММСН chim], [224G11] [ТН7 chim]. Черными стрелками указаны лунки, засеянные только клетками, либо в отсутствие ↙, либо в присутствии ↘ HGF. Мышиный IgG1 вводили в качестве отрицательного контроля на агонистическую активность. m5D5 использовали в качестве дозозависимого полного агонистического контроля.
Фиг.15: Эффект мышиного 224G11 Mab и различных химерных и гуманизированных вариантов этого антитела на индуцированную HGF пролиферацию клеток NCI-H441 in vitro. Клетки NCI-H441 высевали в среду без сыворотки. Через двадцать четыре часа после высевания добавляли тестируемое антитело либо в отсутствие, либо в присутствии HGF. Представлены мышиная m224G11, химерные [224G11] chim, [224G11] [ТН7 chim]) формы IgG1 и [224G11] [ТН7 Hz1], [224G11] [ТН7 Hz3]. Черными стрелками указаны лунки, засеянные только клетками, либо в отсутствие ↙, либо в присутствии ↘ HGF. Мышиный IgG1 вводили в качестве отрицательного контроля на агонистическую активность. m5D5 использовали в качестве дозозависимого полного агонистического контроля.
Фиг.16, А-С: Сравнение in vivo мышиных, химерных и гуманизированных 224G11 Мао на модели ксенотрансплантата NCI-H441.
Фиг.17А: агонистический эффект, вычисленный в виде процента против максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.17В: антагонистический эффект, вычисленный в виде процента ингибирования максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.18: Модели BRET с моделью активации c-Met.
Фиг.19, А-В: Эффект m224G11 и h224G11 на распад c-Met на клетках А549. А) Среднее 4 независимых экспериментов +/- СКО (средняя квадратичная ошибка). В) изображение Вестерн-блоттинга 4 независимых проведенных экспериментов.
Фиг.20, А-В: Эффект m224G11 и h224G11 на распад c-Met на клетках NCI-H441. А) Среднее 4 независимых экспериментов +/- СКО. В) изображение Вестерн-блоттинга 4 независимых проведенных экспериментов.
Фиг.21: Постановка ELISA для оценки шеддинга c-Met.
Фиг.22: Оценка in vitro шеддинга c-Met на клетках NCI-H441, обработанных в течение 5 суток m224G11. mIgG1 является нерелевантным антителом, используемым в качестве контроля изотипа.
Фиг.23: Оценка in vitro шеддинга c-Met на амплифицированных клеточных линиях Hs746T, MKN45 и ЕВС-1, обработанных в течение 5 суток m224G11. mIgG1 является нерелевантным антителом, используемым в качестве контроля изотипа. РМА является индуктором шеддинга, используемым в качестве положительного контроля.
Фиг.24: Оценка in vitro шеддинга c-Met на NCI-H441 и амплифицированных клеточных линиях Hs746T, MKN45 и ЕВС-1, обработанных в течение 5 суток m224G11. mIgG1 является нерелевантным антителом, используемым в качестве контроля изотипа. РМА является индуктором шеддинга, используемым в качестве положительного контроля.
Фиг.25: Исследование собственного фосфорилирования D224G11 на клеточной линии Hs746T.
Фиг.26, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии NCI-H441. А) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.27, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии Hs578T. A) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.28, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии NCI-H125. А) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.29, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии T98G. А) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.30, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии MDA-MB-231. А) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.31, А-В: Исследование собственного фосфорилирования h224G11 на клеточной линии РС3. А) фосфо-ELISA и В) анализ с помощью Вестерн-блоттинга.
Фиг.32: Исследование собственного фосфорилирования h224G11 на клетках HUVEC.
Фиг.33: Сравнение in vivo мышиного антитела 224G11 дикого типа с 224G11[C2D5-7] Mab со сконструированной шарнирной областью на модели ксенотрансплантата NCI-H441.
Фиг.34, А-В: Индукция АЗКЦ h224G11 как на клетках Hs746T, так и на клетках NCI-H441. 51Cr-меченые клетки Hs746T (А) или клетки NCI-H441 (В), с нанесенным (сплошные квадраты) или не нанесенным (пустые квадраты) h224G11 смешивали в различном соотношении с клетками человека NK и инкубировали в течение 4 ч. Клетки собирали и считали имп/мин 51Cr, высвобожденного в результате лизиса. Результаты наносили на график в виде процента лизиса против соотношения эффекторных клеток/клеток-мишеней. NL для клеток без антитела.
Фиг.35, А-С: Окрашивание h224G11 в ксенотрансплантате опухоли, который экспрессировал различные уровни c-Met (A: Hs746T амплифицированная клеточная линия для c-Met, В: NCI-H441 высокий уровень экспрессии c-Met и С: MCF-7 низкий уровень c-Met).
Фиг.36, A-D: Активность in vivo h224G11 в модели ксенотрансплантата Hs746T. А) Анализ с помощью FACS (флуоресцентной сортировки клеток), показывающий экспрессию c-Met на клетках Hs746T по сравнению с другими клеточными линиями, экспрессирующими c-Met. В) Q-OT-ПЦР, показывающая амплификацию c-Met. С) Фосфорилирование c-Met в отсутствие или в присутствии HGF. D) Модель in vivo.
Фиг.37, A-D: Активность in vivo h224G11 в модели ксенотрансплантата MKN-45. А) Анализ с помощью FACS, показывающий экспрессию c-Met на клетках MKN45 по сравнению с другими клеточными линиями, экспрессирующими c-Met. В) Q-OT-ПЦР, показывающая амплификацию c-Met. С) Фосфорилирование c-Met в отсутствие или в присутствии HGF. D) Модель in vivo.
Фиг.38, A-D: Активность in vivo h224G11 в модели ксенотрансплантата ЕВС-1. А) Анализ с помощью FACS, показывающий экспрессию c-Met на клетках ЕВС-1 по сравнению с другими клеточными линиями, экспрессирующими c-Met. В) Q-OT-ПЦР, показывающая амплификацию c-Met. С) Фосфорилирование c-Met в отсутствие или в присутствии HGF. D) Модель in vivo.
Фиг.39, А-Н: Исследование с помощью ИГХ опухолей Hs746T у мышей, обработанных h224G11 Mab, по сравнению с опухолями контрольной обработки. Панели (А) и (В) соответствуют антителам контроля изотипа. На (С) и (D) показана экспрессия c-Met на срезах опухолей от мышей, обработанных ФСБ или h224G11, соответственно, демонстрирующая, что хроническая обработка h224G11 индуцировала резкую понижающую регуляцию c-Met. Окрашивание Ki-67 значительно снижено на обработанных животных (F) по сравнению с контрольными образцами ФСБ (Е). Значительный апоптоз наблюдали в опухоли, обработанной h224G11 (Н), тогда как в контрольных опухолях никакого апоптоза не было отмечено(G).
Пример 1: Образование антител против c-Met
Для образования антител против c-Met 8-недельных мышей BALB/c иммунизировали либо от 3 до 5 раз подкожно трансфицированной клеточной линией СНО, которая экспрессирует c-Met на плазматической мембране клеток (20×106 клеток/доза/мышь), либо от 2 до 3 раз слитым белком внеклеточного домена c-Met (10-15 мкг/доза/мышь) (R&D Systems, № по каталогу 358МТ), либо фрагментами этого рекомбинантного белка, смешивая с полным адъювантом Фрейнда для первой иммунизации и с неполным адъювантом Фрейнда для последующих. Также осуществляли смешанные протоколы, при которых мыши получали и клетки CHO-cMet, и рекомбинантные белки. За трое суток до слияния клеток мышам проводили бустер-инъекцию i.p. или i.v. рекомбинантным белком или фрагментами. Затем селезенки мышей собирали и сливали с клетками миеломы SP2/0-Ag14 (АТСС) и подвергали селекции на среде HAT (гипоксантин, аминоптерин и тимидин). Было проведено четыре слияния. В целом для получения моноклональных антител или их функциональных фрагментов, в частности, мышиного происхождения, возможно ссылаться на методы, которые описаны, в частности, в руководстве "Antibodies" (Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor NY, pp.726, 1988), или на метод получения гибридом, описанный Kofiler and Milstein (Nature, 256:495-497, 1975).
Полученные гибридомы сначала подвергали скринингу с помощью ELISA на рекомбинантный белок c-Met, а затем с помощью анализа FACS на клеточных линиях А549 NSCLC, ВхРС3 поджелудочной железы и U87-MG глиобластомы, чтобы убедиться, что продуцируемые антитела будут способны также распознавать нативный рецептор на опухолевых клетках. Положительные реакции на этих 2 тестах амплифицировали, клонировали, и серия гибридом была выделена, очищена и подвергнута скринингу на их способность ингибировать in vitro клеточную пролиферацию в модели ВхРС3. С этой целью 50000 клеток ВхРС3 высевали в 96-луночные планшеты в среде RPMI, 2 мМ L-глутамин, без SVF. Через 24 часа после высевания тестируемые антитела добавляли при конечной концентрации в диапазоне от 0,0097 до 40 мкг/мл за 60 мин до добавления 100 нг/мл hHGF. После 3 суток клетки метили импульсной меткой 0,5 мкКи [3H]тимидин в течение 16 часов. Количество [3H]тимидина, включенного в ДНК, нерастворимую в трихлоруксусной кислоте, определяли количественно с помощью жидкостного сцинтилляционного счета. Результаты выражали в виде необработанных данных, чтобы действительно выявить собственный агонистический эффект каждого Mab.
Затем антитела, ингибирующие по меньшей мере 50% клеточной пролиферации, оценивали на их активность в отношении димеризации c-Met и активации анализа BRET на трансфицированных клетках. Активность рецептора с-Met определяли количественно путем измерения рекрутмента сигнальной молекулы Gab1 на активированном c-Met. С этой целью были созданы стабильные клеточные линии СНО, экспрессирующие C-Met-Rluc или C-Met-Rluc и C-Met-K1100A-YFP для димеризации c-Met или C-Met-Rluc и мутированную форму Gab1 [Maroun et al., Mol. Cell. Biol, 1999, 19:1784-1799], слитого с YFP, для активации с-Met. Клетки распределяли в белые 96-луночные микропланшеты в культуральной среде DMEM-F12/ФСТ 5% за одни или за двое суток до экспериментов BRET. Сначала клетки культивировали при 37°C с CO2 5%, чтобы дать возможность прикрепления клеток к планшету. Затем клетки подвергали голоданию в 200 мкл DMEM/лунка в течение ночи. Непосредственно перед экспериментом DMEM удаляли, и клетки быстро промывали ФСБ. Клетки инкубировали в ФСБ в присутствии и в отсутствие тестируемых антител или референсных соединений 10 мин при 37°С перед добавлением целентеразина с HGF и без него в конечном объеме 50 мкл. После инкубации в течение еще 10 минут при 37°С начинали снятие данных испускания света при 485 нм и 530 нм, используя люминометр Mithras (Berthold) (1 с/длина волны/лунка, повторяли 15 раз).
Отношение BRET определено ранее [Angers et al., Proc. Natl. Acad. Sci. USA, 2000, 97:3684-3689] как: [(испускание при 530 нм) - (испускание при 485 нм) Х Cf] / (испускание при 485 нм), где Cf соответствует (испускание при 530 нм) / (испускание при 485 нм) для клеток, экспрессирующих только слитый белок Rluc в тех же условиях эксперимента. Упрощение этого уравнения показывает, что отношение BRET соответствует отношению 530/485 нм, полученному в присутствии двух партнеров, скорректированному по отношению 530/485 нм, полученному в тех же условиях эксперимента, когда только один партнер, слитый с люциферазой R. Reniformis, присутствует в анализе. В целях читаемости результаты выражали в единицах миллиВРЕТ (mBU); mBU соответствует отношению BRET, умноженному на 1000.
После этого второго теста in vitro было отобрано антитело 224G11 i) без собственной активности в виде целой молекулы в функциональном тесте пролиферации, ii) значительно ингибирующее пролиферацию ВхРС3 и iii) ингибирующее димеризацию c-Met. В экспериментах 5D5 Mab, полученное фирмой Genentech и доступное в АТСС, добавляли в качестве контроля на собственную агонистическую активность.
Пример 2: Способ гуманизации мышиного 224G11 Mab путем прививания CDR
1) Гуманизация вариабельного домена легкой цепи (VL)
В качестве предварительной стадии нуклеотидную последовательность 224G11 VL сравнивали с последовательностями генов зародышевой линии мыши, включенными в базу данных IMGT (http://imgt.cines.fr). Идентифицированы гены зародышевой линии мыши IGKV3-5*01 и IGKJ4*01, проявляющие идентичность последовательности 99,31% для V области и 94,28% для J области соответственно. С учетом этих высоких гомологии нуклеотидная последовательность 224G11VL использована непосредственно для поиска гомологии человека вместо соответствующих зародышевых генов мыши.
На второй стадии проведен поиск гена зародышевой линии человека, проявляющего наилучшую идентичность с 224G11VL, чтобы идентифицировать лучший человеческий кандидат на прививание CDR. Для оптимизации селекции провели выравнивания между аминокислотными последовательностями. Ген зародышевой линии человека IGKV4-1*01 дал идентичность последовательности 67,30%, но показал отличающуюся длину для CDR1 (10 аминокислот в 224G11 VL и 12 аминокислот в IGKV4-1*01). Для J области был отобран ген зародышевой линии человека IGKJ4*02 (идентичность последовательности 77,14%).
На следующей стадии участки CDR 224G11 VL были привиты на отобранные выше каркасные последовательности человека. Каждое положение аминокислоты анализировали на несколько критериев, таких как участие в пограничной поверхности VH/VL, в связывании антигена или в структуре CDR, локализация остатка в 3D структуре вариабельного домена, якори CDR, остатки, принадлежащие к зоне Верньера. Было сконструировано три гуманизированных варианта, соответствующих SEQ ID No. 8, SEQ ID No. 9 и SEQ ID No. 10, и они содержали соответственно четыре (4, 39, 40, 84), два (39, 40) или один (40) мышиный остаток в их FR областях и CDR участки, соответствующие 224G11 VL мыши.
2) Гуманизация вариабельного домена тяжелой цепи (VH)
В качестве предварительной стадии нуклеотидную последовательность 224G11 VH сравнивали с последовательностями генов зародышевой линии мыши, включенными в базу данных IMGT (http://imgt.cines.fr).
Идентифицированы гены зародышевой линии мыши IGHV1-18*01, IGHD2-4*01 и IGHJ2*01 с идентичностью последовательности 92,70% для V области, 75,00% для D области и 89,36% для J области соответственно. С учетом этих высоких гомологии нуклеотидная последовательность 224G11 VH использована непосредственно для поиска гомологии человека вместо соответствующих зародышевых генов мыши.
На второй стадии проведен поиск гена зародышевой линии человека, проявляющего наилучшую идентичность с 224G11 VH, чтобы идентифицировать лучший человеческий кандидат на прививание CDR. С этой целью проведено выравнивание нуклеотидной последовательности 224G11 VH с последовательностями генов зародышевой линии человека, принадлежащими к базе данных IMGT. Последовательность гена человека IGHV1-2*02 V проявляла идентичность последовательности 75,00% на нуклеотидном уровне и 64,30% на аминокислотном уровне. Поиск гомологии для J области привел к идентификации гена зародышевой линии человека IGHJ4*04 с идентичностью последовательности 78,72%.
На следующей стадии участки CDR 224G11 VH были привиты на отобранные выше каркасные последовательности человека. Каждое положение аминокислоты анализировали на несколько критериев, таких как участие в пограничной поверхности VH/VL, в связывании антигена или в структуре CDR, локализация остатка в 3D структуре вариабельного домена, якори CDR, остатки, принадлежащие к зоне Верньера. Была сконструирована одна полностью гуманизированная форма, соответствующая SEQ ID 4; она содержит исключительно человеческие остатки в ее FR областях и CDR, соответствующие 224G11 VH мыши.
Пример 3: Конструирование улучшенных мутантов шарнирной области
Специалисту в данной области техники хорошо известно, что шарнирная область принимает сильное участие в гибкости вариабельного домена иммуноглобулинов (см. Brekke et al., 1995; Roux et al., 1997). В процессе химеризации 224G11 Mab константный домен мыши IGHG1 был замещен эквивалентным участком IGHG1 человеческого происхождения. Поскольку аминокислотная последовательность шарнирной области была в высокой степени дивергентной, "муринизацию" шарнирной области осуществляли с целью сохранения ее длины и прочности. Поскольку шарнирная область IGHG2 человека соответствует ближайшему гомологу шарнирной области мыши IGHG1, эта последовательность была также рассмотрена. Была сконструирована серия из 7 различных шарнирных последовательностей (SEQ ID No. 22-28) путем включения участков шарнирных областей IGHG1 мыши и IGHG2 человека в участок шарнирной области IGHG1 человека.
Другая серия мутантов шарнирной области была разработана и сконструирована (SEQ ID No. 58-72) для оценки влияния дополнительного цистеина и его положения по длине шарнирного домена, делеции 1, 2, 3 или 4 аминокислот по длине шарнирного домена и комбинации этих двух параметров (добавления цистеина и делеции аминокислоты).
Пример 4: Продуцирование гуманизированных 224G11 Mab и форматов Mab со сконструированной шарнирной областью
Все вышеописанные формы Mab, содержащие либо химерные, либо гуманизированные и/или полученные генно-инженерным путем шарнирные области, продуцировали при транзитной трансфекции путем использования системы HEK293/EBNA с экспрессионным вектором рСЕР4 (InVitrogen, US).
Полноразмерные нуклеотидные последовательности, соответствующие гуманизированным вариантам легкой (SEQ ID No. 18, SEQ ID No. 19 и SEQ ID No. 20) и тяжелой (SEQ ID No. 14) цепи вариабельного домена 224G11 Mab синтезировали глобального синтеза генов (Genecust, Luxembourg). Их субклонировали в векторе рСЕР4 (InVitrogen, US), несущем полноразмерную кодирующую последовательность константного домена [CH1-Hinge-CH2-CH3] иммуноглобулина IgG1 или IgG2 человека. Модификацию шарнирной области осуществляли путем обмена фрагмента рестрикции {Nhell-Bcll} на эквивалентный участок, несущий желаемые модификации, где каждый соответствующий фрагмент {Nhel-Bcll} синтезировали путем глобального синтеза генов (Genecust, LU). Все стадии клонирования осуществляли в соответствии с общепринятыми методами молекулярной биологии, как описано в лабораторном руководстве (Sambrook and Russel, 2001), или в соответствии с инструкциями поставщика. Каждая генетическая конструкция была полностью подтверждена секвенированием нуклеотидов с использованием набора Big Dye для терминаторного цикла секвенирования (Applied Biosystems, US) и проанализирована с использованием генетического анализатора 3100 Genetic Analyzer (Applied Biosystems, US).
Адаптированные к росту в суспензии клетки HEK293 EBNA (InVitrogen, US) выращивали обычным путем во флаконах на 250 мл в 50 мл среды без сыворотки Excell 293 (SAFC Biosciences) с добавлением 6 мМ глутамина на орбитальном встряхивателе (скорость вращения 110 об/мин). Транзитную трансфекцию осуществляли с использованием 2.106 клеток/мл, используя линейный 25 кДа полиэтиленимин (ПЭИ) (Polysciences), приготовленный в воде при конечной концентрации 1 мг/мл, смешанный с плазмидной ДНК (конечная концентрация 1,25 мкг/мл для соотношения плазмид тяжелой и легкой цепи 1:1). Через 4 часа после трансфекции культуру разводили одним объемом свежей культуральной среды до достижения конечной плотности клеток 106 клеток/мл. Мониторинг процесса культивирования осуществляли на основе жизнеспособности клеток и продуцирования Mab. Типично культуры поддерживали в течение от 4 до 5 суток. Mab очищали, используя общепринятый хроматографический метод, на смоле с белком A (GE Healthcare, US).
Все различные формы Mab продуцировались на уровнях, пригодных для функциональных оценок. Уровни продуктивности типично находятся в интервале от 15 до 30 мг/л очищенных Mab.
Пример 5: Оценка состояния фосфорилирования c-Met с помощью фосфо-с-Met-специфичного анализа ELISA
Данный функциональный анализ дает возможность мониторинга модулирования состояния фосфорилирования c-Met либо одними Mab, либо при одновременном присутствии HGF.
Клетки А549 высевали в планшет 12MW в полную среду для роста [F12K + 10% ФСТ (фетальной сыворотки теленка)]. Клетки подвергали голоданию в течение 16 часов перед стимуляцией HGF [100 мг/мл], и каждое тестируемое Mab добавляли при его конечной концентрации 30 мкг/мл за 15 минут до стимуляции лиганда. Охлажденный во льду буфер для лизиса добавляли через 15 минут после добавления HGF, чтобы остановить реакцию фосфорилирования. Клетки соскабливали механическим путем, и клеточные лизаты собирали центрифугированием при 13000 об/мин в течение 10 мин при 4°С, что соответствует фазе супернатанта. Содержание белка количественно определяли, используя набор ВСА (Pierce), и хранили при -20°С до использования. Состояние фосфорилирования c-Met количественно определяли с помощью ELISA. Mab коза против c-Met (R&D, № по каталогу AF276) использовали в качестве иммобилизованного антитела (покрытие в течение ночи при 4°С), и после стадии насыщения с буфером TBS-BSA 5% (1 час при комнатной температуре (КТ)) в каждую лунку покрытого планшета 96MW добавляли 25 мкг белковых лизатов. После 90 минут инкубации при КТ планшеты промывали четыре раза и добавляли идентифицирующее антитело (Mab против фосфо-c-Met, направленное против фосфорилированных остатков Туг в положении 1230, 1234 и 1235). После дополнительной инкубации в течение 1 часа и 4 отмывок антитело против кролика, конъюгированное с HRP (Biosource), добавляли на 1 час при КТ, и обнаружение люминесценции осуществляли путем добавления люминола. Считывания люминесценции осуществляли на многорежимном считывающем устройстве для планшетов Mithras LB920 (Berthold).
Уровень как базового, так и индуцированного HGF [100 мг/мл] фосфорилирования рецептора c-Met не подвергался влиянию ни в результате обработки ФСБ, ни в результате добавления мышиных или человеческих Mab, которые не направлены на рецептор c-Met человека (Фиг.1). С другой стороны, мышиное (m) 224G11 Mab сильно ингибировало индуцированное HGF [100 нг/мл] фосфорилирование c-Met (Фиг.2В) без изменения собственного фосфорилирования рецептора (Фиг.2А). Удивительно, что химерная форма 224G11 Mab (224G11chim/IgG1), означающая вариабельный домен (VH+VL) от m224G11, объединенный с константным доменом человека IgG1/каппа, давала сильную (17% от максимального эффекта HGF, Фиг.2А) агонистическую активность, ассоциированную со сниженной антагонистической эффективностью (54% ингибирование максимального эффекта HGF по сравнению с m224G11, которое дает 75% ингибирование максимального эффекта HGF, Фиг.2В). Три гуманизированные формы 224G11 Mab, [224G11]Hz1/IgG1, [224G11]Hz2/IgG1 и [224G11]Hz3/IgG1, также сконструированные на каркасе IgG1/каппа человека, давали также сниженную антагонистическую активность и значительную агонистическую активность (от 11 до 24% максимального уровня HGF) по сравнению с мышиным 224G11 (Фиг.2А и 2В). Была сконструирована серия полученных генно-инженерным путем вариантов шарнирного домена тяжелой цепи и проанализирована в анализе фосфорилирования рецептора c-Met. Как показано на Фиг.3А, важное снижение агонистического эффекта, связанное с изотипом hIgG1/каппа, наблюдали как для конструкции на основе IgG2, так и для полученных генно-инженерным путем конструкций IgG1/каппа [МН, MUP9H и ТН7]. Сопутствующее повышение антагонистической эффективности было также получено. Мутант шарнирной области ТН7 на основе hIgG1/каппа с самой большой частью человеческой последовательности был отобран для осуществления процесса гуманизации. На следующей стадии было получено три гуманизированных варианта вариабельного домена 224G11 Mab путем комбинирования либо с IgG2/каппа человека, либо со сконструированным шарнирным константным доменом доменом ТН7 на основе IgG1/каппа. Для гуманизированных конструкций hIgG2/каппа гуманизированный вариант Hz3 давал сильный агонизм (Фиг.4А), и для всех трех гуманизированных вариантов антагонистическая эффективность была ниже, чем наблюдали с мышиным 224G11 Mab, и сравнима с химерным Mab на основе hIgG1 (56-57% ингибирование эффекта HGF, Фиг.4В). С другой стороны, комбинирование трех гуманизированных вариантов Hz1, Hz2 или Hz3 со сконструированным мутантом IgG1/TH7 наиболее полно восстанавливало свойства мышиного 224G11 Mab в отношении слабой агонистической активности (5-6% эффекта HGF) и сильной антагонистической эффективности (68-72% ингибирование эффекта HGF) фосфорилирования рецептора c-Met (Фиг.5А и 5В). Эти варианты были в высокой степени улучшены по сравнению с химерным 224G11 Mab на основе IgG1, но также и с гуманизированными формами на основе IgG2.
Вторая серия полученных генно-инженерным путем вариантов шарнирного домена тяжелой цепи была сконструирована и проанализирована в анализе фосфорилирования рецептора c-Met. Как показано на фиг.17А, все эти новые варианты (c224G11[C2], c224G11[C3], c224G11[C5], c224G11[C6], c224G11[C7], C224G11[Δ1-3], c224G11[С7Δ6], c224G11[C6Δ9], c224G11[C2Δ5-7], c224G11[C5Δ2-6], c224G11[С9Δ2-7] и c224G11[Δ5-6-7-8]) проявляли более слабый агонистический эффект, чем C224G11, поскольку их агонистические активности составляют от 6 до 14% эффекта HGF по сравнению с 23% для C224G11. Как и C224G11 [ТН7], все эти новые варианты проявляли сопутствующее повышение антагонистической эффективности [фиг.17В]. Эти результаты показали, что конструирование домена тяжелой цепи генно-инженерным путем за счет точечной мутации и/или делеции может модифицировать агонистические/антагонистические свойства антитела.
Пример 6: Анализ ВРЕТ
В первой серии экспериментов был поставлен контроль, что нерелевантный IgG1 мыши, IgG1 человека и IgG2 человека не обладают эффектом индуцированного HGF сигнала BRET в обеих моделях BRET (репрезентативный эксперимент из 12 независимых экспериментов; Фиг.6). Эти Mab далее указаны как контроли.
Оценивали эффект химерной формы IgG1 мышиного 224G11 Mab ([224G11]chim) на модели BRET как димеризации c-Met, так и активации c-Met. Если мышиное 224G11 Mab ингибировало 59,4% индуцированного HGF сигнала BRET на модели димеризации c-Met, [224G11]chim Mab ингибировало только 28,9% (Фиг.7А). Антитело [224G11]chim было также менее эффективным при ингибировании индуцированной HGF активации c-Met, так как антитела [224G11]chim и m224G11 ингибировали соответственно 34,5% и 56,4% индуцированного HGF сигнала BRET (Фиг.7В). Кроме того, одно m224G11 не обладало эффектом на активацию c-Met, тогда как [224G11]chim обладало частичным агонистическим эффектом на активацию c-Met, соответствующим 32,9% индуцированного HGF сигнала. Этот частичный агонистический эффект [224G11]chim также наблюдали в отношении модели BRET димеризации c-Met, поскольку одно [224G11]chim индуцировало повышение BRET, соответствующее 46,6% индуцированного HGF сигнала по сравнению с 21,3% для m224G11 (Фиг.7А).
На Фиг.8А и 8В химерные формы антитела 224G11 с мутированной шарнирной областью проявляли более высокий ингибиторный эффект в отношении индуцированного HGF сигнала BRET, чем [224G11]chim, поскольку они показали 59,7%, 64,4%, 53,2% и 73,8% ингибирования индуцированного HGF сигнала активации BRET (Фиг.8В) и 61,8%, 64,4% 52,5% и 64,4% ингибирования индуцированного HGF сигнала BRET димеризации c-Met (Фиг.8А) для [224G11][МН chim], [224G11][MUP9H chim], [224G11][ММСН chim] и [224G11][ТН7 chim] соответственно. В противоположность [224G11]chim, которое обладало частичным агонистическим эффектом на активацию c-Met, одни химерные формы антитела 224G11 с мутированной шарнирной областью не проявляли значительного эффекта на активацию c-Met (5,1%, 7,6%, -2,0% и -6,9% соответственно), как наблюдали для m224G11.
На Фиг.9В, подобно [224G11] [ТН7 chim], 3 гуманизированных варианта антитела 224G11 IgG1 с шарнирной областью ТН7 не индуцировали значительного повышения сигнала BRET в модели активации при тестировании их одних и проявляли сильное ингибирование индуцированного HGF сигнала BRET: 59,9%, 41,8% и 57,9% для форм Hz1, Hz2 и Hz3 соответственно. Кроме того, [224G11] [ТН7 Hz1], [224G11] [ТН7 Hz2] и [224G11] [ТН7 Hz3] ингибировали индуцированный HGF сигнал BRET на модели димеризации на 52,2%, 35,8% и 49,4% соответственно (фиг.9А).
В противоположность [224G11]chim, одна химерная форма антитела 224G11 IgG2 ([224G11] [IgG2 chim]) не проявляла частичного агонистического эффекта и ингибировала 66,3% эффекта HGF на модели активации c-Met (Фиг.10В). На модели димеризации c-Met [224G11] [IgG2 chim] ингибировало 62,4% индуцированного HGF сигнала BRET (Фиг.10А).
Агонистическую эффективность второй серии сконструированных вариантов шарнирного домена тяжелой цепи оценивали в модели BRET активации c-Met (Фиг.18). В противоположность c224G11, которое обладало частичным агонистическим эффектом на активацию c-Met, одни химерные формы антитела 224G11 с мутированной шарнирной областью c224G11 [С2], c224G11 [С3], c224G11 [С5], c224G11 [С6], c224G11 [С7], c224G11[Δ1-3], c224G11[C7Δ6], c224G11[C6Δ9], c224G11[C2Δ5-7], c224G11[C5Δ2-6], c224G11[C9Δ2-7] и c224G11[Δ5-6-7-8] не проявляли значительного эффекта на активацию c-Met.
Пример 7: Распознавание c-Met химерными и гуманизированными формами 224G11
Поставлен прямой анализ ELISA, чтобы определить связывающую способность различных химерных и гуманизированных форм в отношении рекомбинантного c-Met. Кратко, рекомбинантным димерным c-Met от R&D Systems покрывали при 1,25 мкг/мл 96-луночные планшеты Immunlon II. После инкубации в течение ночи при 4°С лунки насыщали раствором 0,5% желатин/ФСБ. Затем планшеты инкубировали в течение 1 часа при 37°С, после чего добавляли 2-кратные разведения тестируемых антител. Планшеты инкубировали дополнительно в течение часа, после чего добавляли антитело IgG коза против мыши HRP для обнаружения мышиного антитела и коза против легкой цепи каппа человека HRP для распознавания химерных и гуманизированных антител. Планшеты инкубировали в течение одного часа и добавляли субстрат пероксидазы ТМВ Uptima на 5 мин, после чего нейтрализовали H2SO4 1 М. Результаты, представленные на Фиг.11, показали, что все тестируемые формы сравнимы по распознаванию c-Met.
Пример 8: Эффект мышиного и химерных 224G11 на индуцированную HGF пролиферацию клеток NCI-H441 in vitro
Клетки NCI-H441 из АТСС культивировали обычным путем в среде RPMI 1640 (Invitrogen Corporation, Scotland, UK), 10% ФСТ (Invitrogen Corporation), 1% L-глутамин (Invitrogen corporation). Для анализов пролиферации клетки делили за 3 суток до использования, так, чтобы они находились в конфлюентной фазе роста перед высевом. Клетки NCI-H441 высевали в 96-луночные планшеты для тканевых культур при плотности 3,75×104 клеток/лунка в 200 мкл среды без сыворотки (среда RPMI 1640 с 1% L-глутамином). Через двадцать четыре часа после высевания тестируемые антитела добавляли к NCI-H441 и инкубировали при 37°С в течение тридцати минут перед добавлением HGF при конечной концентрации 400 нг/мл (5 нМ) еще на 142 дополнительных часа. Диапазон дозы, тестируемый для каждого антитела, составляет от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В данном эксперименте мышиное IgG1 Mab добавляли в качестве контроля изотипа мыши, и тестируемые антитела составляли следующие: m224G11 и его химерная форма IgG1 человека, идентифицированная как [224G11]chim. Лунки, засеянные одними клетками -/+ HGF, были также включены. Затем клетки метили импульсной меткой 0,25 мкКи [3H]тимидин (Amersham Biosciences AB, Uppsala, Sweden) в течение 7 часов и 30 минут. Количество [3H]тимидина, включенного в ДНК, нерастворимую в трихлоруксусной кислоте, определяли количественно с помощью жидкостного сцинтилляционного счета. Результаты выражены в виде не преобразованных данных имп/мин, чтобы лучше оценить потенциальную собственную агонистическую активность, которая может встречаться при добавлении одних антител Mab против c-Met к опухолевой клетке.
Результаты, описанные на Фиг.12, демонстрируют, что, как ожидали, мышиное антитело m224G11 не проявляло агонистического эффекта, когда его добавляют одно к раковым клеткам, независимо от тестируемой дозы. Значительного ингибирования индуцированной HGF пролиферации не наблюдали с контролем изотипа с учетом вариаций имп/мин, наблюдаемых для этого соединения в данном эксперименте. Когда его добавляют одно, антитело m224G11 не показало никакого агонистического эффекта по сравнению с контролем изотипа mIgG1 Mab или одними клетками. Дозозависимые антипролиферативные активности, достигающие 78%, наблюдали для m224G11 (вычисление % ингибирования: 100-[(имп/мин клетки + тестируемые Mab - среднее фоновое число имп/мин mIgG1) × 100 / (среднее число имп/мин клетки + HGF - среднее число имп/мин одних клеток)]). Неожиданно химерные формы 224G11 Mab индуцировали значительный дозозависимый агонистический эффект, когда их добавляют одни. Этот агонистический эффект оказывал влияние на ингибирование in vitro индуцированной HGF пролиферации, которая смещалась с 78% для мышиного 224G11 до 50% для его химерной формы. Чтобы определить, совместима ли такая "более низкая" собственная агонистическая активность in vitro с неизмененным эффектом in vivo, оба антитела m224G11 и [224G11]chim продуцировали для тестирования in vivo. Как в предшествующих исследованиях, доза 30 мкг/мышь продемонстрировала значительную активность in vivo, и эта доза была выбрана для оценки in vivo.
Пример 9: Сравнение in vivo мышиного и химерных 224G11 Mab на модели ксенотрансплантата NCI-H441
Клетки NCI-H441 имеют происхождение из папиллярной аденокарциномы легкого, экспрессируют высокие уровни c-Met и демонстрируют конститутивное фосфорилирование c-Met RTK.
Для оценки эффекта антител in vivo на модели ксенотрансплантата NCI-H441 шести-восьминедельных бестимусных мышей держали в стерилизованных клетках с фильтрующими насадками, поддерживали в стерильных условиях и проводили манипуляции с ними в соответствии с французскими и европейскими руководствами. Мышам инъецировали подкожно 9х106 клеток. Затем через шесть суток после имплантации клеток опухоли были измеримыми (примерно 100 мм3), животных делили на группы по 6 мышей со сравнимым размером опухоли и обрабатывали сначала насыщающей дозой 60 мкг антитела/мышь, а затем дважды в неделю 30 мкг/доза каждого тестируемого антитела. Мышей обследовали для наблюдения скорости роста ксенотрансплантата. Объем опухоли вычисляли по формуле: π (пи)/6 × длина × ширина × высота. Результаты, описанные на Фиг.13, демонстрируют, что мышиное Mab с отсутствующей агонистической активностью in vivo ведет себя, как ожидали, как сильный антагонист даже при низкой тестируемой дозе. В противоположность наблюдаемому для мышиного Mab, химерное антитело проявляло весьма преходящую активность in vivo, и опухоль полностью избежала обработки на сутки D20 после инъекции клеток. Этот эксперимент четко демонстрирует, что повышение агонистического эффекта in vitro, которое привело в результате к снижению антагонистической активности, также ответственно за значительную потерю антагонистической активности in vivo.
Пример 10: Эффект мышиного 224G11 Mab и различных химерных и гуманизированных вариантов этого антитела на индуцированную HGF пролиферацию клеток NCI-H441 in vitro
Клетки NCI-H441 из АТСС культивировали обычным путем в среде RPMI 1640 (Invitrogen Corporation, Scotland, UK), 10% ФСТ (Invitrogen Corporation), 1% L-глутамин (Invitrogen corporation). Для анализов пролиферации клетки делили за 3 суток до использования, так, чтобы они находились в конфлюентной фазе роста перед высевом. Клетки NCI-H441 высевали в 96-луночные планшеты для тканевых культур при плотности 3,75×104 клеток/лунка в 200 мкл среды без сыворотки (среда RPMI 1640 с 1% L-глутамином). Через двадцать четыре часа после высевания тестируемые антитела добавляли к NCI-H441 и инкубировали при 37°С в течение тридцати минут перед добавлением HGF при конечной концентрации 400 нг/мл (5 нМ) еще на 142 дополнительных часа. Диапазон дозы, тестируемый для каждого антитела, составляет от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В данном эксперименте мышиное IgG1 Mab добавляли в качестве контроля изотипа мыши и в качестве отрицательного контроля агониста. Тестируемые антитела составляли следующие: i) m224G11, ii) его химерные формы IgG1 человека, соответственно идентифицированные как [224G11] chim, [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [ММСН chim], [224G11] [ТН7 chim] iii) его гуманизированные формы IgG1, соответственно описанные как [224G11] [Hz1], [224G11] [Hz2], [224G11] [Hz3]. Лунки, засеянные одними клетками -/+ HGF, были также включены. Полное антитело 5D5 от Genentech, имеющееся в продаже от АТСС в виде гибридомной клеточной линии, вводили в качестве положительного контроля полного агониста, и далее его называют m5D5. Затем клетки метили импульсной меткой 0,25 мкКи [3H]тимидин (Amersham Biosciences AB, Uppsala, Sweden) в течение 7 часов и 30 минут. Количество [3Н]тимидина, включенного в ДНК, нерастворимую в трихлоруксусной кислоте, определяли количественно с помощью жидкостного сцинтилляционного счета. Результаты выражены в виде не преобразованных данных имп/мин, чтобы лучше оценить потенциальную собственную агонистическую активность, которая может встречаться при добавлении одних антител Mab против c-Met к опухолевой клетке.
Результаты, описанные на Фиг.14А, продемонстрировали, что, как ожидали, ни контроль изотипа, ни m224G11 не проявляли какой-либо агонистической активности в отношении пролиферации NCI-H441. Контроль изотипа не обладал никаким эффектом на индуцированную HGF пролиферацию, тогда как m224G11 показало 66% ингибирование при добавлении при конечной концентрации 10 мкг/мл. Антитело m5D5, используемое в качестве контроля агониста, показало, как ожидали, полностью дозозависимый агонистический эффект, когда его добавляют к клеткам одно. Как уже наблюдали, [224G11] chim Mab проявляло значительный дозозависимый агонистический эффект, и наблюдали сниженную ингибиторную активность этой формы антитела: 19% вместо 66% для мышиной формы. Когда их добавляли одни, 3 гуманизированных IgG1 Mab демонстрировали дозозависимые агонистические эффекты по сравнению с формой m224G11. [224G11] [Hz1], [224G11] [Hz2] и [224G11] [Hz3] обладали сравнимыми агонистическими активностями примерно 46, 30 и 35%. Эти активности значительно ниже, чем наблюдаемые для m224G11. На Фиг.14В тестировали различные химерные формы IgG1. По сравнению с формой [224G11] chim, которая проявляла дозозависимый агонистический эффект, когда ее добавляли одну к клеткам NCI-Н441, у форм [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [ММСН chim], [224G11] [ТН7 chim] отсутствовал значительный собственный агонистический эффект. Их антагонистическая активность была выше, чем активность, наблюдаемая для m224G11 Mab (57%), где ингибирования достигали 79, 78, 84 и 93% соответственно для [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [ММСН chim] и [224G11] [ТН7 chim].
Пример 11: Эффект in vitro различных гуманизированных форм IgG1 224G11 Mab
Клетки NCI-H441 из АТСС культивировали обычным путем в среде RPMI 1640 (Invitrogen Corporation, Scotland, UK), 10% ФСТ (Invitrogen Corporation), 1% L-глутамин (Invitrogen corporation). Для анализов пролиферации клетки делили за 3 суток до использования, так, чтобы они находились в конфлюентной фазе роста перед высевом. Клетки NCI-H441 высевали в 96-луночные планшеты для тканевых культур при плотности 3,75×104 клеток/лунка в 200 мкл среды без сыворотки (среда RPMI 1640 с 1% L-глутамином). Через двадцать четыре часа после высевания тестируемые антитела добавляли к NCI-H441 и инкубировали при 37°С в течение тридцати минут перед добавлением HGF при конечной концентрации 400 нг/мл (5 нМ) еще на 142 дополнительных часа. Диапазон дозы, тестируемый для каждого антитела, составляет от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В данном эксперименте мышиное IgG1 Mab добавляли в качестве отрицательного фонового контроля на агонистическую активность, и тестируемые антитела были следующими: i) m224G11, ii) его химерные формы IgG1 человека, соответственно идентифицированные как [224G11] chim, [224G11] [ТН7 chim] iii) его гуманизированные формы IgG1 соответственно описаны как [224G11] [ТН7 Hz1], [224G11] [ТН7 Hz3]. Лунки, засеянные одними клетками -/+ HGF, были также включены. Полное антитело 5D5 от Genentech, имеющееся в продаже от АТСС в виде гибридомной клеточной линии, вводили в качестве положительного контроля полного агониста, и далее его называют m5D5. Затем клетки метили импульсной меткой 0,25 мкКи [3H]тимидин (Amersham Biosciences AB, Uppsala, Sweden) в течение 7 часов и 30 минут. Количество [3Н]тимидина, включенного в ДНК, нерастворимую в трихлоруксусной кислоте, определяли количественно с помощью жидкостного сцинтилляционного счета. Результаты выражены в виде не преобразованных данных имп/мин, чтобы лучше оценить потенциальную собственную агонистическую активность, которая может встречаться при добавлении одних антител Mab против c-Met к опухолевой клетке.
На Фиг.15 показано, что m224G11 Mab проявляет обычный ингибиторный эффект (74% ингибирования). Химерная форма IgG1 [224G11] chim обладала, как ожидали, дозозависимым собственным агонистическим эффектом и более низким антагонистическим эффектом по сравнению с мышиной формой: 33% против 74% ингибирования. Антитело [224G11] [ТН7 chim] обладало очень слабой агонистической активностью в данном эксперименте. Однако оно проявляло высокий ингибиторный эффект (81%), близкий к отмеченному для мышиного Mab. Две гуманизированные формы не обладали собственным агонистическим эффектом и обладали антагонистической активностью, близкой к активностям, наблюдаемым для мышиного Mab или [224G11] [ТН7 chim] соответственно при 67 и 76% ингибирования для [224G11] [ТН7 Hz1] и [224G11] [ТН7 Hz3].
Пример 12: Сравнение in vivo мышиного, химерных и гуманизированных 224G11 Mab, несущих либо шарнирную область дикого типа, либо сконструированную шарнирную область ТН7 (модель ксенотрансплантата NCI-H441)
Клетки NCI-H441 имеют происхождение из папиллярной аденокарциномы легкого, экспрессируют высокие уровни c-Met и демонстрируют конститутивное фосфорилирование c-Met RTK.
Чтобы оценить необходимость конструирования шарнирной области генно-инженерным путем с сохранением активности in vivo мышиного антитела 224G11, шести-восьминедельных бестимусных мышей держали в стерилизованных клетках с фильтрующими насадками, поддерживали в стерильных условиях и проводили манипуляции с ними в соответствии с французскими и европейскими руководствами. Мышам инъецировали подкожно 9×106 клеток NCI-H441. Затем через шесть суток после имплантации клеток опухоли были измеримыми (примерно 100 мм3), животных делили на группы по 6 мышей со сравнимым размером опухоли и обрабатывали сначала насыщающей дозой 2 мг антитела/мышь, а затем дважды в неделю 1 мг/доза каждого тестируемого антитела. В данном эксперименте оценивали десять антител, включающих m224G11, химерную форму, проявляющую шарнирную область дикого типа (c224G11), химерную форму со сконструированной шарнирной областью ТН7 (224G11[TH7 chim]), три гуманизированных формы, несущих шарнирную область дикого типа (224G11[IgG1 Hz1], 224G11[IgG1 Hz2] и 224G11[IgG1 Hz3]) и три соответствующих формы со сконструированной шарнирной областью ТН7 (224G11[TH7 Hz1], 224G11[TH7 Hz2] и 224G11[TH7 Hz3]). Мышей обследовали для наблюдения скорости роста ксенотрансплантата.
Объем опухоли вычисляли по формуле: π (пи)/6 × длина × ширина × высота.
Результаты, описанные на Фиг.16, демонстрируют, что мышиное Мао с отсутствующей агонистической активностью in vitro ведет себя, как ожидали, как сильный антагонист in vivo. В противоположность тому, что наблюдали с мышиным Mab, как химерные, так и гуманизированные формы, несущие шарнирную область дикого типа, проявляли только весьма преходящую активность in vivo. В любом случае замена шарнирной области дикого типа сконструированной шарнирной областью ТН7 привела в результате к полному восстановлению активности in vivo, наблюдаемой с мышиными антителами. Данный эксперимент четко демонстрирует, что повышение агонистического эффекта in vitro, которое приводило в результате к снижению антагонистической активности, было также ответственно за значительную потерю антагонистической активности in vivo. Он также демонстрирует, что использование сконструированной области ТН7 вместо дикого типа необходимо для поддержания свойств in vivo мышиного Mab.
Пример 13: Эффект m224G11 и его гуманизированной формы h224G11 на понижающую регуляцию c-Met in vitro
В последующих примерах, во избежание сомнений, экспрессия h224G11 относится к гуманизированной форме 224G11 [ТН7 Hz3] антитела по изобретению.
Для направления активности антител против c-Met на распад рецептора с-Met выбраны две клеточные линии. А549 (№ НТВ-174) и NCI-H441 (№ CCL-185) представляют собой две клеточные линии NSCLC из коллекции АТСС. Клетки NCI-H441 высевали в RPMI 1640+1% L-глутамина + 10% термоинактивированной ФСТ при 3×104 клеток/см2 в шестилуночные планшеты на 24 ч при 37°С в атмосфере 5% CO2. Клетки А549 высевали в F12K + 10% термоинактивированной ФСТ при 2×104 клеток/см2 в шестилуночные планшеты на 24 ч при 37°С в атмосфере 5% CO2. Затем клетки дважды промывали фосфатно-солевым буферным раствором (ФСБ), после чего их подвергали голоданию по сыворотке дополнительно в течение 24 часов. Антитела против c-Met (10 мкг/мл), нерелевантное mIgG1 (10 мкг/мл) или HGF (400 нг/мл) добавляли в среду DMEM без сыворотки при 37°С. После либо 4 часов, либо 24 инкубации среду осторожно удаляли, и клетки дважды промывали холодным ФСБ. Клетки лизировали 500 мкл охлажденного во льду буфера для лизиса [50 мМ Трис-HCl (рН 7,5); 150 мМ NaCl; 1% Nonidet P40; 0,5% деоксихолат; и 1 таблетка полной смеси ингибиторов протеаз с 1% антифосфатаз]. Клеточные лизаты встряхивали в течение 90 мин при 4°С и осветляли при 15000 об/мин в течение 10 минут. На этой стадии клеточные лизаты можно было хранить при -20°С до потребности для Вестерн-блот-анализа. Концентрацию белка определяли количественно, используя ВСА. Цельные клеточные лизаты (5 мкг в 20 мкл) разделяли с помощью электрофореза в ДСН-ПААГ и переносили на нитроцеллюлозную мембрану. Мембраны насыщали в течение 1 ч при КГ TBS-Твин 20 0,1% (TBST); 5% обезжиренного сухого молока и гибридизовали с антителом против c-Met (разведение 1/1000) в течение ночи при 4°С в TBST-5% обезжиренного сухого молока. Антитела разводили в физиологическом растворе, забуференном Трис, с 0,1% Твин 20 (об/об) (TBST) с 1% обезжиренного сухого молока. Затем мембраны промывали TBST и инкубировали со вторым антителом, конъюгированным с пероксидазой (разведение 1:1000), в течение 1 ч при КГ. Иммунореактивные белки визуализировали с помощью ECL (Pierce # 32209). После визуализации c-Met мембраны еще раз промывали TBST и инкубировали в течение 1 ч при КТ с мышиным антителом против GAPDH (разведение 1/200000) в TBST-5% обезжиренном молоке. Затем мембраны промывали в TBST и инкубировали со вторыми антителами, конъюгированными с пероксидазой, в течение 1 ч при КТ. Мембраны промывали, и GAPDH проявляли, используя ECL. Интенсивность полос количественно определяли с помощью денситометрии.
Результаты, представленные на фиг.19,А и 20,А, продемонстрировали, что оба антитела, m224G11 и D224G11, способны дозозависимо осуществлять значительную понижающую регуляцию c-Met в обеих клеточных линиях А549 и NCI-H441. Эта понижающая регуляция значительна уже после 4-часового периода инкубации и еще возрастает к 24-му часу. Гистограммы, представленные на фиг.19,А и 20,А, соответствуют средним значениям или соответственно 4 и 3 независимым экспериментам. Изображения Вестерн-блоттинга, соответствующие одному значимому эксперименту, включены в фиг.19,В и 20,В.
Пример 14: Эффект m224G11 и его гуманизированных форм h224G11 на шеддинг c-Met in vitro
Растворимые формы рецептора c-Met, претерпевшего шеддинг, встречаются в естественных условиях в сыворотке мышей с привитым ксенотрансплантатом опухоли человека или в сыворотке пациента-человека, несущего опухоли, экспрессирующие c-Met. Кроме того, антитела, направленные против c-Met, такие как DN30 Mab, описаны как индукторы шеддинга c-Met в экспериментах in vitro. Чтобы определить, обладает ли m224G11 таким свойством, клетки высевали в шестилуночные планшеты со средой с 10% ФСТ. Когда они достигали примерно 80% конфлюентности, среду удаляли и добавляли свежую полную культуральную среду +/- тестируемые соединения. Клетки инкубировали дополнительно в течение 72 часов либо с m224G11, либо с контролем изотипа mIgG1, либо с ФСБ. ФМА (форболмиристоацетат) вводили в качестве индуктора шеддинга. HGF также тестировали на клетках, чтобы определить воздействие лиганда c-Met на естественно происходящий шеддинг. Затем супернатанты собирали и фильтровали на 0,2 мкм перед использованием в тесте ELISA, где эти растворимые формы c-Met были иммобилизованы антителом против c-Met, которое не распознает тот же эпитоп, что либо m224G11, либо с11Е1 (Фиг.21). Кроме того, клетки из каждой лунки промывали один раз ФСБ и лизировали, чтобы определить концентрацию белка. Для ELISA в качестве иммобилизующего антитела использовали 224D10, и после насыщения планшетов фильтрованные супернатанты из шестилуночных планшетов добавляли в тест ELISA. Мономерную форму c-Met использовали в качестве положительного контроля. После инкубации супернатанта планшеты промывали, чтобы удалить несвязанный c-Met, и с11Е1 использовали, чтобы обнаружить c-Met, иммобилизованный 224G11 Mab. Проявление теста, наконец, осуществляли путем добавления HRP-конъюгированного поликлонального антитела против hFc.
Результаты, показанные на фиг.22, указывают на то, что естественный шеддинг c-Met происходил, когда клетки культивировали в течение 72 часов in vitro. Эффекта mIgG1 не наблюдали. Однако добавление m224G11, по-видимому, ингибирует шеддинг c-Met. Эти результаты были подтверждены для 3 других клеточных линий (Hs746T, EBC1 и MKN45) на фиг.23. В этом втором эксперименте ФМА, добавленный в качестве положительного индуктора шеддинга, значительно повышал, как и ожидали, шеддинг c-Met по меньшей мере в 2 клеточных линиях (Hs746T и MKN45). Наконец, в третьем эксперименте (Фиг.24) HGF вводили в качестве контроля. Дополнительный шеддинг не был индуцирован HGF по сравнению с одними клетками или с клетками + mIgG1. Опять же, значительное ингибирование шеддинга c-Met наблюдали с m224G11.
Пример 15: Собственный эффект h224G11 Ab на различных клеточных линиях
В предшествующих экспериментах, описанных в данном патенте, продемонстрировано, что, в противоположность тому, что наблюдали с другими антителами, такими как 5D5, m224G11 и его гуманизированная форма h224G11 не проявляют значительную собственную активность на опухолевых клеточных линиях. Чтобы распространить это свойство на другие клеточные линии, были проведены эксперименты по Вестерн-блоттингу и фосфо-ELISA с одним антителом, добавленным в течение различных промежутков времени, на серии раковых клеточных линий с варьирующими уровнями экспрессии c-Met, включая Hs746T, NCI-H441, Hs578T, NCI-H125, T98G, MDA-MB-231, РС3. Тот же тест был также проведен на нормальных клетках: HUVEC.
Способ анализа фосфо-cMet ELISA был уже описан в примере 5 настоящей заявки на патент. Для Вестерн-анализа белковые лизаты получали из осажденных клеток путем инкубации в буфере для лизиса с ингибиторами протеаз и фосфатаз [10 нМ Трис (рН 7,4), 150 мМ NaCl, 1 мМ ЭДТА, 1 мМ ЭГТА, 0,5% Nonidet P40, 100 мМ фторид натрия, 10 мМ пирофосфат натрия, 2 мМ ортованадат натрия, 2 мМ ФМСФ (фенилметилсульфонилфторид), 10 мг/мл лейпептин, 10 мг/мл апротинин] при 4°С. Белковые лизаты осветляли от клеточного детрита путем центрифугирования, разделяли с помощью электрофореза на 8% ДСН-ПААГ гелях и подвергали электропереносу на нитроцеллюлозную мембрану. Для экспериментов с c-Met лизаты подвергали иммунопреципитации на специфичный интересующий белок перед электрофорезом и переносом.
Результаты, представленные на фиг.25-32, снова демонстрируют, что в тестируемых клетках никакой собственной активности антитела h224G11 не наблюдали.
Пример 16: Сравнение in vivo мышиного антитела 224G11 дикого типа с химерной формой 224G11 со сконструированной шарнирной областью, описанной как 224G11[C2D5-7] (модель ксенотрансплантата NCI-H441)
Клетки NCI-H441 имеют происхождение из папиллярной аденокарциномы легкого, экспрессируют высокие уровни c-Met и демонстрируют конститутивное фосфорилирование c-Met RTK.
Чтобы оценить необходимость конструирования шарнирной области генно-инженерным путем с сохранением активности in vivo мышиного антитела 224G11, шести-восьминедельных бестимусных мышей держали в стерилизованных клетках с фильтрующими насадками, поддерживали в стерильных условиях и проводили манипуляции с ними в соответствии с французскими и европейскими руководствами. Мышам инъецировали подкожно 9×106 клеток NCI-H441. Затем через шесть суток после имплантации клеток опухоли были измеримыми (примерно 100 мм3), животных делили на группы по 6 мышей со сравнимым размером опухоли и обрабатывали сначала насыщающей дозой 2 мг антитела/мышь, а затем дважды в неделю 1 мг/доза каждого тестируемого антитела. Мышей обследовали для наблюдения скорости роста ксенотрансплантата. Объем опухоли вычисляли по формуле: π (пи)/6 × длина × ширина × высота. Результаты, описанные на фиг.33, демонстрируют, что мышиное Mab без какой-либо агонистической активности in vitro ведет себя, как и ожидали, как сильный антагонист in vivo. Как предполагали на основании результатов, полученных in vitro в анализах фосфорилирования, антитело c224G11[C2D5-7] со сконструированной шарнирной областью, которое не проявляло значительного агонистического эффекта, демонстрирует сильную активность in vivo, сравнимую с активностью m224G11, на модели ксенотрансплантата NCI-H441.
Пример 17: Оценка h224G11 в тесте АЗКЦ
Поскольку h224G11 имеет изотип IgG1, АЗКЦ может составлять часть его эффективности in vivo у человека. Анализ цитотоксичности по высвобождению in vitro [51Cr] проводили, используя либо клетки Hs746T, либо клетки NCI-H441 в качестве клеток-мишеней и клетки NK, очищенные из мононуклеарных лимфоцитов периферической крови человека.
Кратко, один миллион клеток-мишеней Hs746T или NCI-H441 инкубировали в присутствии или в отсутствие 20 мкг h224G11 Ab в присутствии 100 мкКи 51хрома (Perkin Elmer) в течение 1 ч. Затем 4×103 клеток высевали с возрастающим числом естественных киллерных клеток (NK) человека, выделенных из мононуклеарных клеток периферической крови (РВМС) с использованием отрицательной селекции (Stemcell Technologies). Клетки инкубировали вместе дополнительно в течение 4 часов при 37°С. Процент лизиса клеток вычисляли, следуя формуле: [(экспериментальное высвобождение 51Cr - спонтанное высвобождение 51Cr)/(полное высвобождение 51Cr - спонтанное высвобождение 51Cr)]×100. Спонтанное высвобождение представляет число импульсов в минуту, полученное, когда клетки-мишени культивировали в отсутствие естественных киллерных клеток. Полное высвобождение представляет число импульсов в минуту, полученное, когда клетки-мишени лизировали 1% Тритон Х-100. h224G11 значительно усиливало лизис как клеток Hs746T (фиг.34,А), так и клеток NCI-H441 (фиг.34) на 62,9% и 63,2% соответственно в соотношении МК/клетки-мишени, равном 100.
Пример 18: Иммуногистохимические исследования (ИГХ)
Методики ИГХ окрашивания опухолей, заключенных в парафин: 8-12 мкм срезы замороженной опухоли немедленно фиксировали в предварительно охлажденном ацетоне при -20°С в течение 3 минут. Затем предметные стекла охлаждали при комнатной температуре в течение 30 минут - 1 часа. После 2 отмывок в ФСБ эндогенную пероксидазную активность блокировали, используя реагент для блокирования пероксидаз (Dako K4007), в течение пяти минут. Срезы промывали ФСБ и инкубировали в блокирующем реагенте авидин/биотин (Dako Х0590) непосредственно перед насыщением неспецифических сайтов ФСБ-БСА 4% в течение 30 минут при комнатной температуре. Затем стекла инкубировали с биотинилированным антителом h224G11 (от 50 до 10 мкг/мл) или с биотинилированным IgG1/каппа человека (от 50 до 10 мкг/мл, сайт связывания) в качестве отрицательного контроля в течение 2 часов при комнатной температуре.
Срезы промывали ФСБ и инкубировали с универсальным комплексом стрептавидин - пероксидаза (Dako K0679) в течение 30-45 минут. 3-Амино-9-этилкарбазол использовали для проявления продукта реакции красного цвета (Sigma). Стекла погружали в гематоксилин на 4 минуты для одновременного окрашивания (Dako S3309).
Результаты представлены на Фиг.35,А-С.
h224G11 дифференциально окрашивает клеточную мембрану различных типов опухоли. В данной иммуногистохимической методике продукт реакции красного цвета коррелирует с положительным окрашиванием клеточной мембраны, а отсутствие продукта реакции красного цвета коррелирует с отрицательным окрашиванием и отсутствием визуализации клеточной мембраны. Контроль IgG, IgG1/каппа человека, является контролем соответствия изотипу.
Пример 19: Активность in vivo Mab против c-Met на трех клеточных линиях, амплифицированных на c-Met, и ксенотрасплантированных мышам
Hs746T (АТСС) и MKN45 (JCRB), две клеточные линии карциномы желудка, и ЕВС-1 (JCRB), клеточная линия чешуйчато-клеточной карциномы легкого, являются тремя клеточными линиями, которые осуществляют гиперэкспрессию с-Met, как показано с помощью анализа FACS на фиг.36,А, 37,А и 38,А. Эти клеточные линии также описаны в литературе как амплифицированные клеточные линии на с-Met. Для подтверждения этого последнего утверждения амплификацию гена c-Met определяли с помощью количественной ПЦР в реальном времени, используя способ ΔΔCt Использовали пятьдесят нг геномной ДНК из каждой указанной клеточной линии, экстрагированной с использованием набора QIAamp DNA midi kit (Qiagen, Les Ulis, FR). Референсный ген представлял собой ген ДНК топоизомеразы III альфа, локализованный на хромосоме 17. Праймеры на ген с-Met были локализованы на экзоне 14. Соответствующие праймеры для обоих генов представляют собой праймеры, описанные Smolen et al. (PNAS, 2006, 103:2316-2321), и обеспечивающие сходные эффективности ПЦР. Протокол Q-ПЦР состоит в 10 мин цикле при 95°С, за которым следует 40 последовательных циклов 15 сек при 95°С/45 сек при 95°С. Сначала данные нормализовали по референсному гену, а затем каждую клеточную линию нормализовали по клеткам MCF-7, содержащим единственную копию гена c-Met на гаплоидный геном. Результаты, представленные на фиг.36,В, 37,В и 38,В, демонстрируют, что эти три клеточные линии проявляют значительную амплификацию c-Met, оцениваемую как от 5 до 10-кратной амплификации гена.
Поскольку описано, что такие клетки конститутивно активируются на c-Met, был проведен Вестерн-блот-анализ, чтобы определить уровень фосфорилирования c-Met каждой клеточной линии, либо в отсутствие, либо в присутствии HGF (50 нг/мл). Для этой цели клетки культивировали в течение ночи в среде без сыворотки, а затем обрабатывали или не обрабатывали 50 нг/мл HGF в течение 2 мин. Клетки подвергали лизису буфером для лизиса с добавлением ингибиторов протеаз. Клеточные лизаты инкубировали при 4°С в течение 90 мин, затем центрифугировали, и супернатанты собирали. Концентрации белка определяли, используя анализ белка ВСА (Pierce), и образцы иммобилизовали. Используемые антитела включали антитело против c-Met, клон 25Н2 от фирмы Cell Signaling и против P-c-Met Y1230, 1234, 1235 от фирмы Biosource. Иммуноблоттинг с фосфо-специфичным антителом против c-Met против Y1230/1234/1235 показал сильное базовое фосфорилирование рецептора в каждой амплифицированной клеточной линии (фиг.36,С, 37,С и 38,С). При добавлении HGF к этим клеткам дополнительного фосфорилирования не наблюдали. Данное последнее наблюдение подтверждает, что c-Met был активирован HGF-независимым путем в клеточных линиях Hs746T, MKN45 и ЕВС-1.
С целью тестирования антител против c-Met, созданных авторами изобретения, in vivo на лиганд-независимых клеточных линиях клетки Hs746T MKN45 и ЕВС-1 культивировали обычным способом в полной среде DMEM, RPMI 1640 или ЕМЕМ соответственно. Клетки разделяли за двое суток до прививания, так, чтобы они находились в экспоненциальной фазе роста. Семь миллионов клеток прививали каждой бестимусной мыши SCID (для Hs746T и ЕВС-1) или CD1 Swiss (для MKN45). Через пять-семь суток после имплантации опухоли были измеримыми, и животных делили на группы по 6 мышей со сравнимым размером опухоли. Мышей обрабатывали i.p. насыщающей дозой 2 мг h224G11 Mab /мышь, а затем дважды в неделю 1 мг антитела/мышь. Объем опухоли измеряли дважды в неделю и вычисляли по формуле: π (пи)/6 × длина × ширина × высота. Статистический анализ проводили в каждый момент измерения, используя критерий Манна-Уитни.
Данный эксперимент показывает, что h224G11 значимо снижает рост опухоли Hs746T, MKN45 и ЕВС1 (фиг.36,D, 37,D и 38,D), что указывает на то, что гуманизированное антитело способно ингибировать рост опухоли ЖКТ и легкого лиганд-независимых клеточных линий. Прежде, чем были получены эти результаты, не было очевидно, как повлияет присутствие модифицированной шарнирной области в данном гуманизированном антителе на его лиганд-независимую инактивацию c-Met, поскольку измененная шарнирная область также влияла на его агонистические/антагонистические свойства.
Пример 20: Анализ ex vivo активности h224G11 на модели ксенотрансплантата Hs746T у мышей SCID
Чтобы точнее определить механизмы действия, которые привели в результате к ингибированию ксенотрансплантата опухоли Hs746T, оценивали эффект трех i.p. инъекций h224G11 на митотический индекс опухоли (Ki67), апоптоз (активированная каспаза 3) и экспрессию c-Met, используя способы иммуногистохимии (ИГХ). Анализировали трех мышей на каждое условие. Наблюдали значительную понижающую регуляцию c-Met в отношении обработанных опухолей по сравнению с контрольными (фиг.39,С и D соответственно). Панели 39А и В соответствует контролям изотипа. Кроме значительного снижения уровней Ki67 (фиг.39,Е и F), было отмечено усиленное окрашивание активированной каспазы 3 (фиг.39,G и Н) на всех обработанных опухолях, что указывает на то, что h224G11 способно значительно ингибировать клеточную пролиферацию и индуцировать апоптоз клеток внутри опухоли.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| СПОСОБ МОДУЛИРОВАНИЯ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТИ МОНОКЛОНАЛЬНОГО АНТИТЕЛА | 2009 |
|
RU2575600C2 |
| НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2552161C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| КОМБИНАЦИЯ АНТАГОНИСТА С-МЕТ И АМИНОГЕТЕРОАРИЛА ДЛЯ ЛЕЧЕНИЯ РАКА | 2009 |
|
RU2526171C2 |
| АНТИ-МЕТ АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2765267C2 |
| ГУМАНИЗИРОВАННЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ФАКТОРУ РОСТА ГЕПАТОЦИТОВ | 2007 |
|
RU2461570C2 |
| КОНЪЮГАТЫ АНТИТЕЛА К cMet И ЛЕКАРСТВЕННОГО СРЕДСТВА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2740996C2 |
| НОВОЕ АНТИТЕЛО ДЛЯ ДИАГНОСТИКИ И (ИЛИ) ПРОГНОЗИРОВАНИЯ РАКА | 2011 |
|
RU2651513C2 |
| АНТИТЕЛО ПРОТИВ cMet И ЕГО ПРИМЕНЕНИЕ ДЛЯ ОПРЕДЕЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2010 |
|
RU2582265C2 |









































































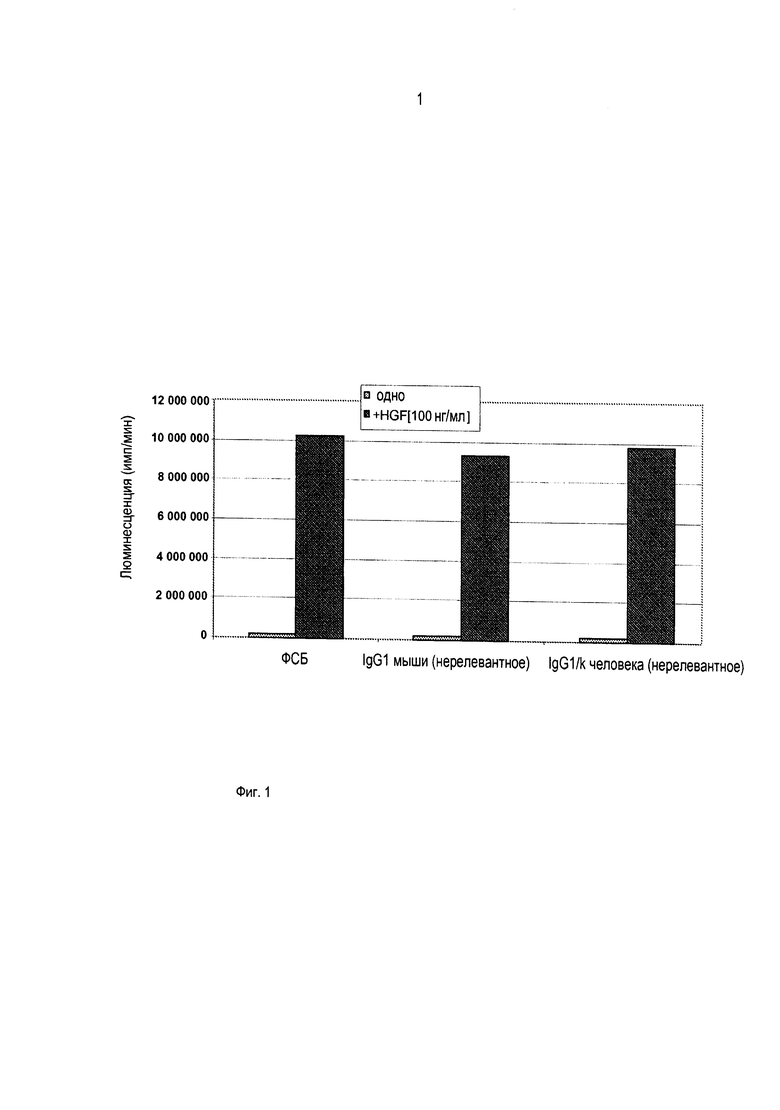

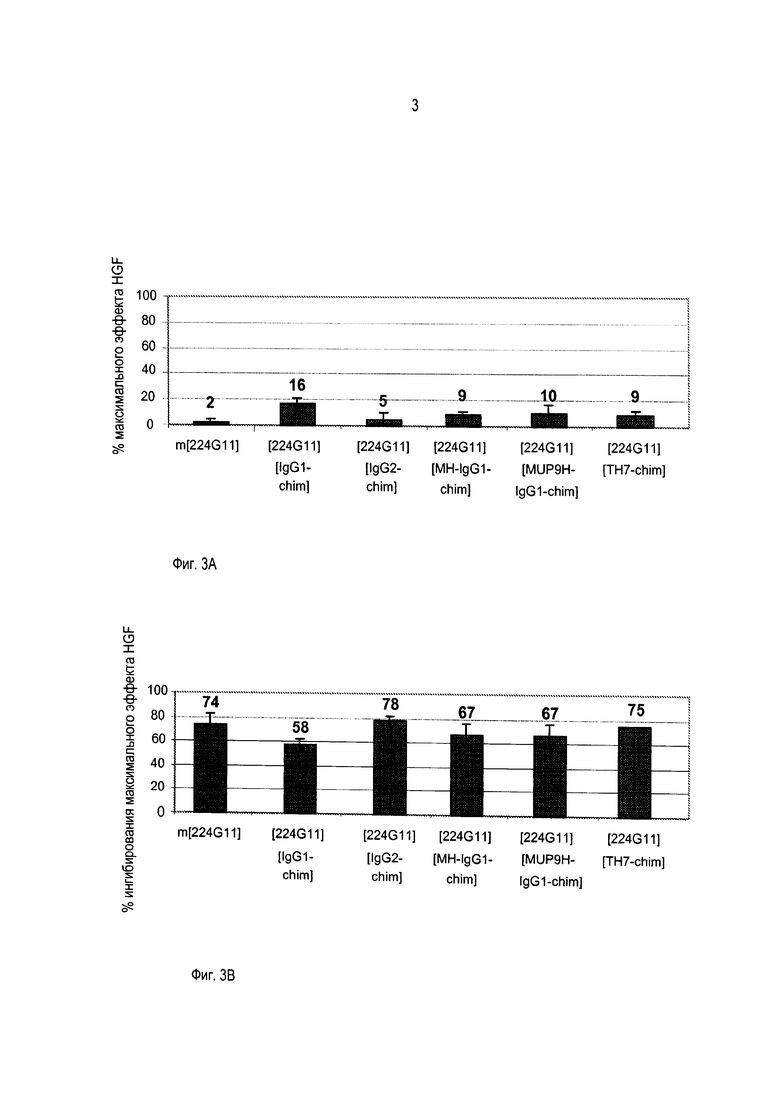
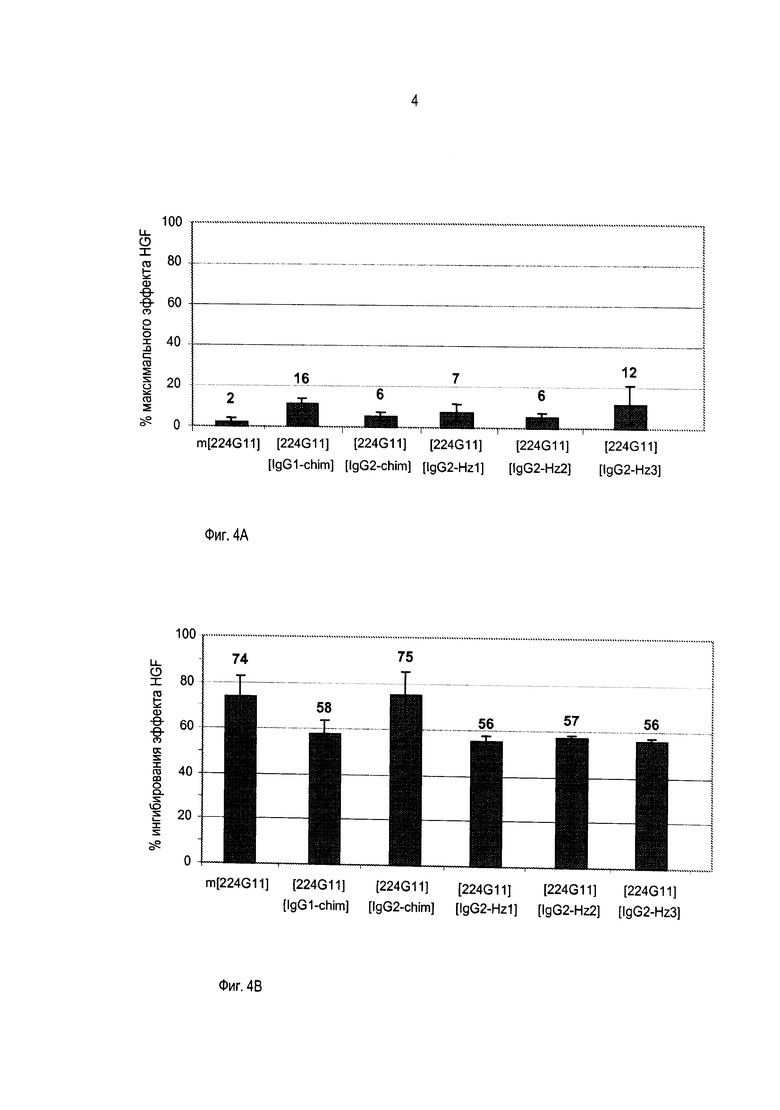
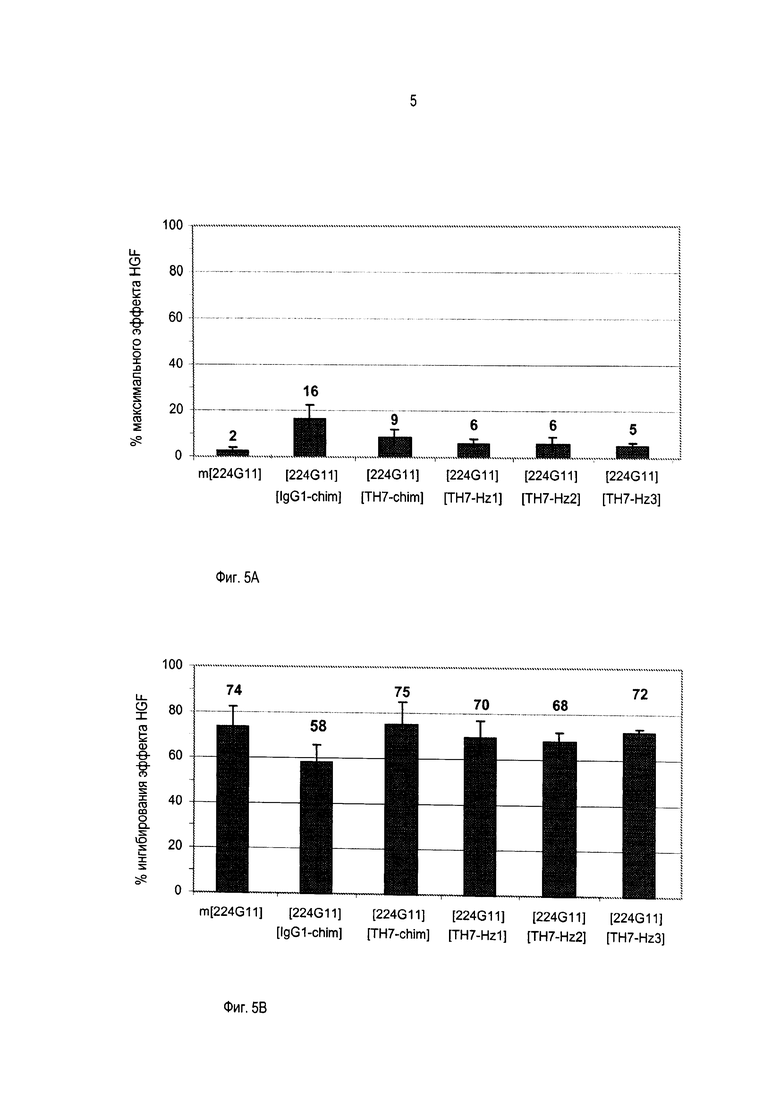











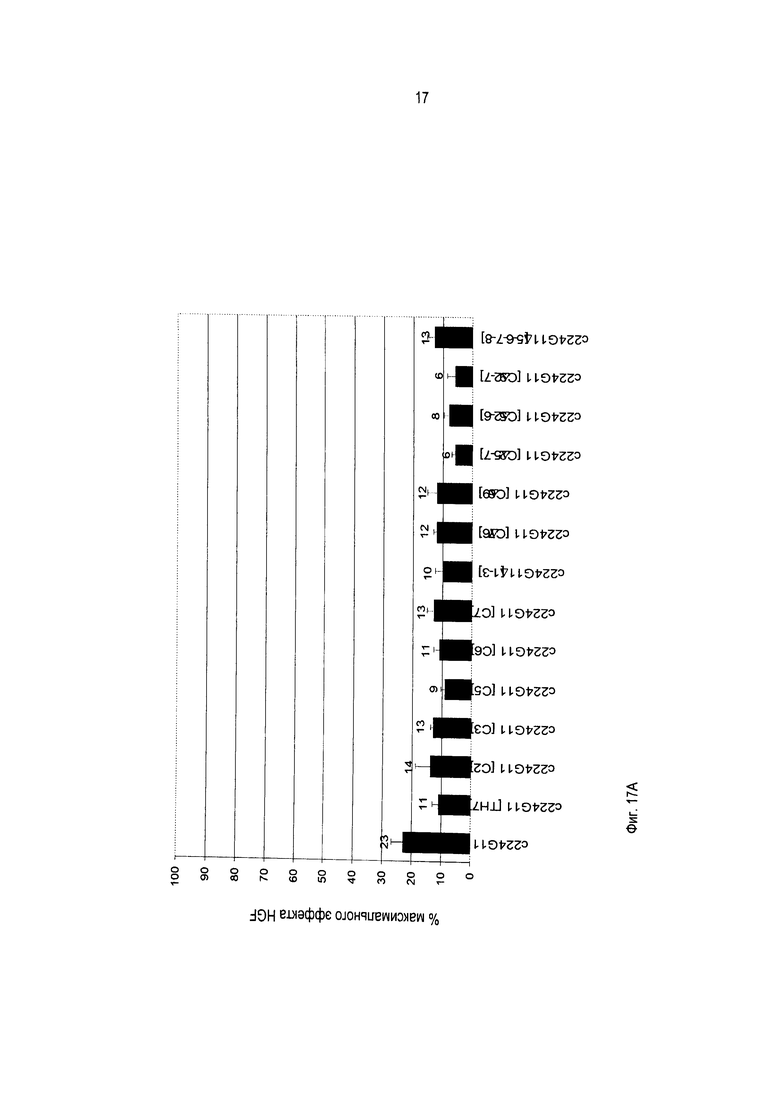


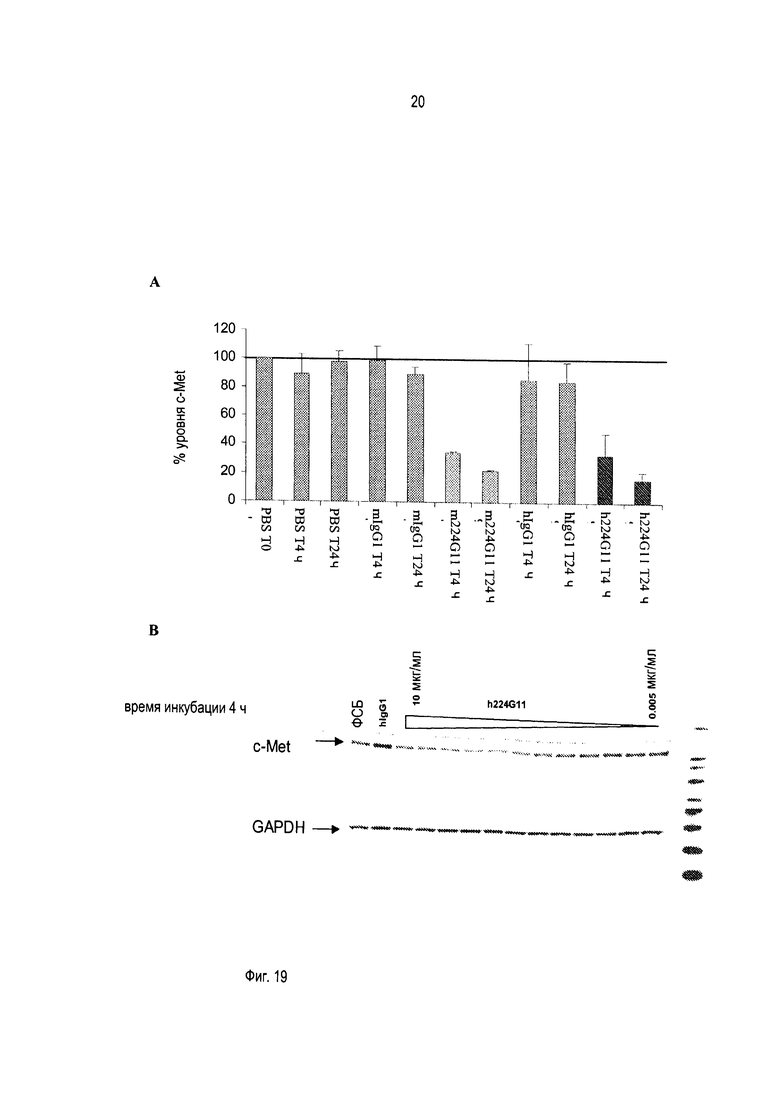





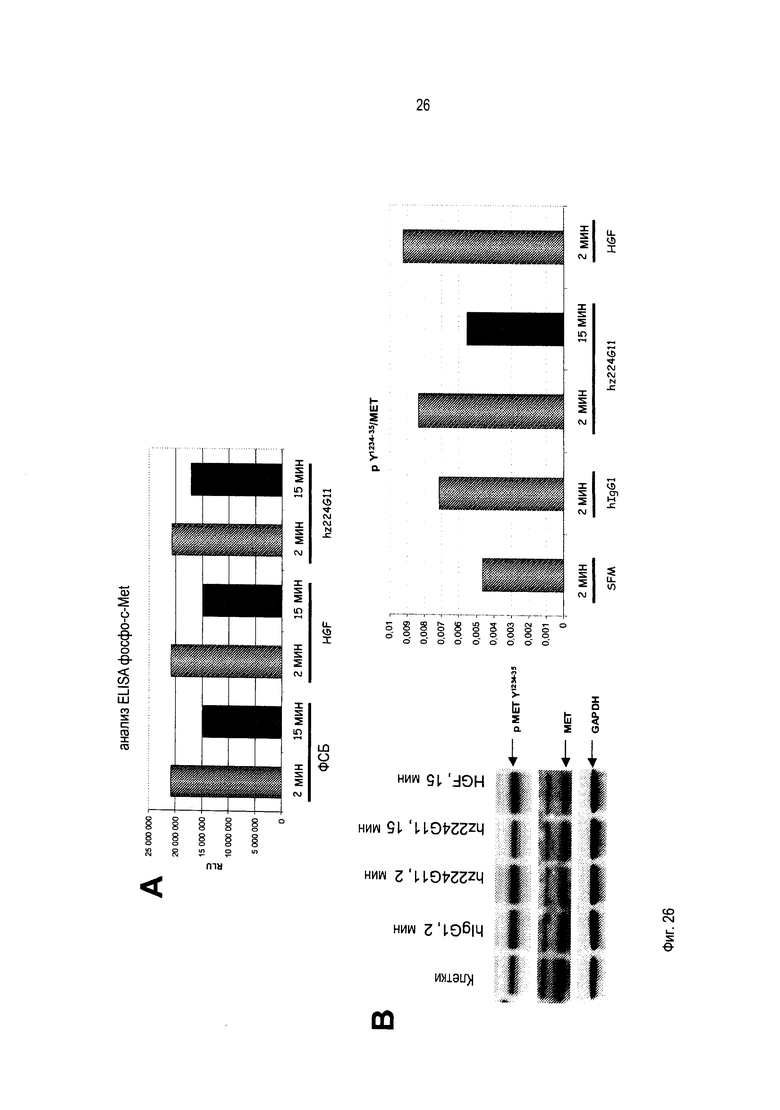





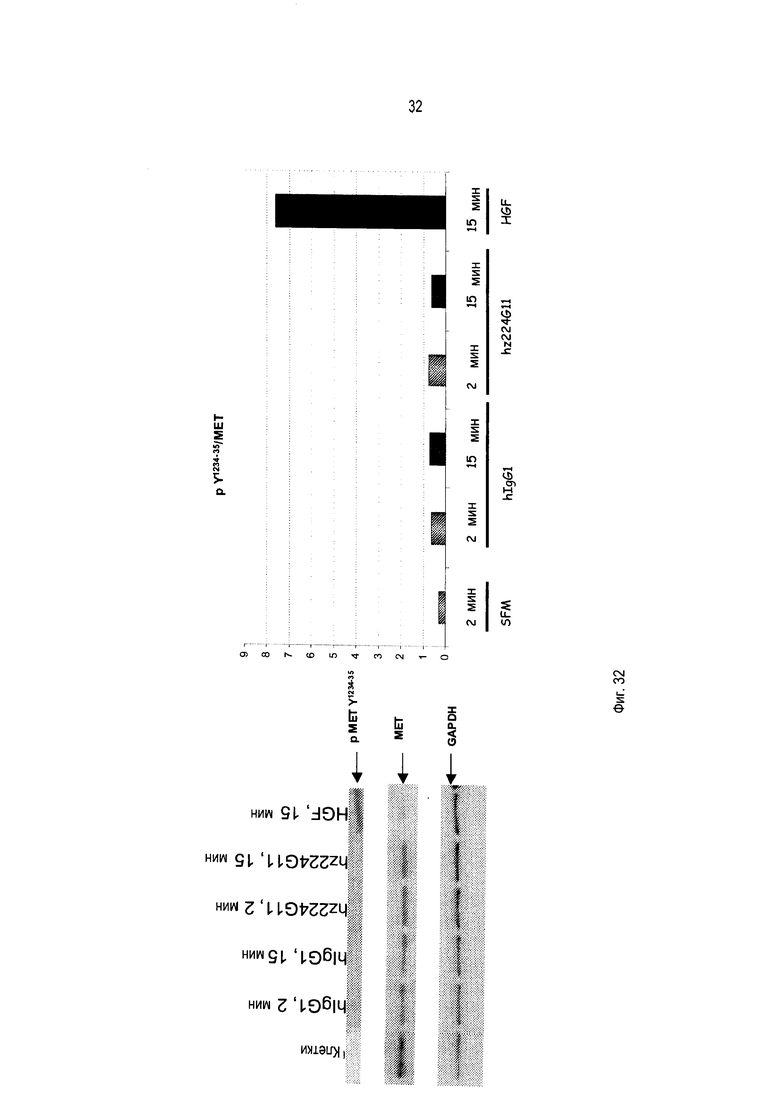
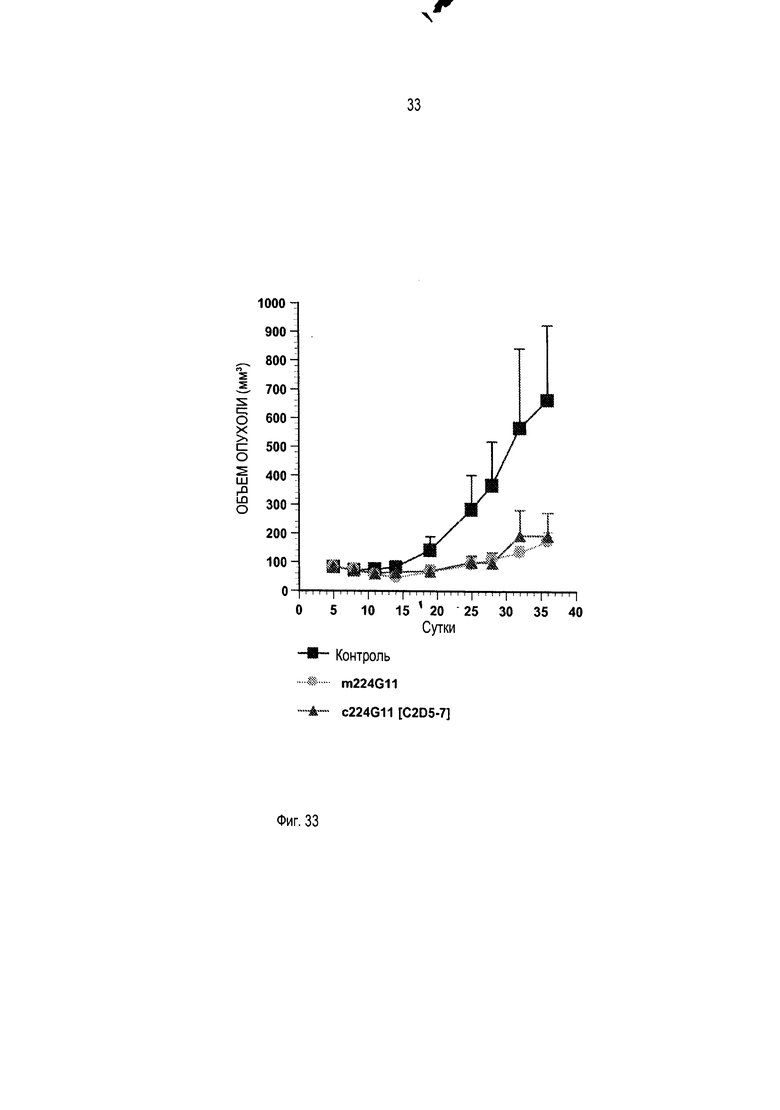
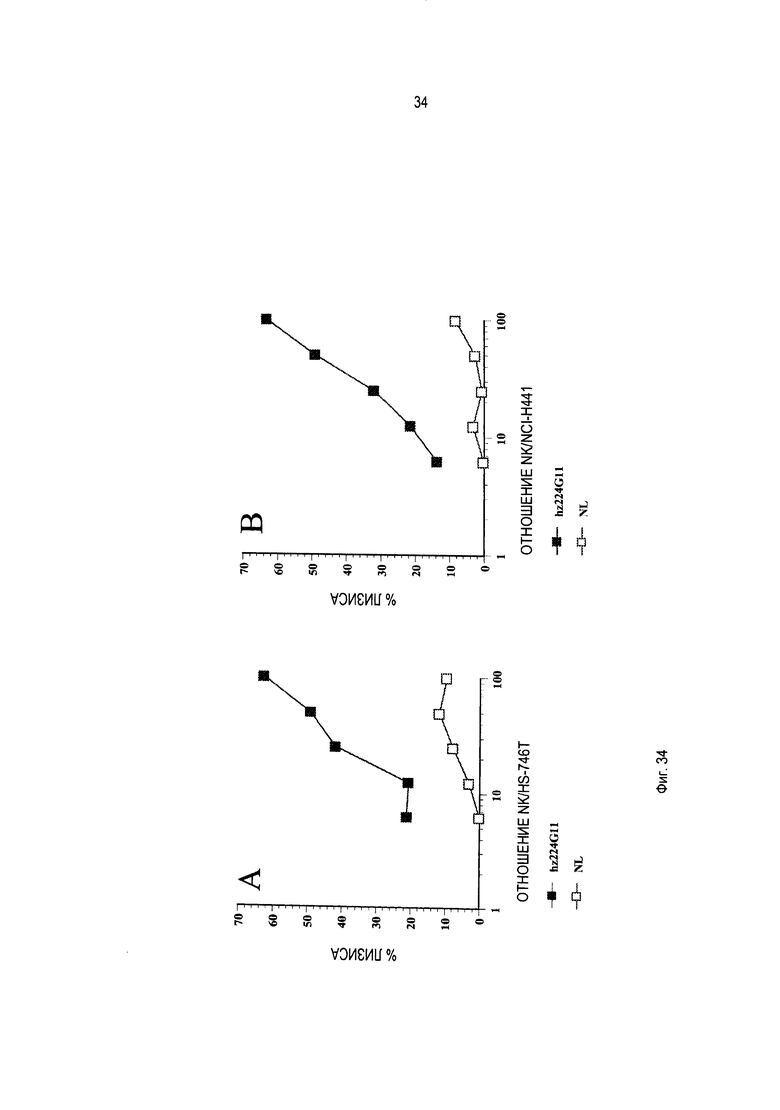





Изобретение относится к области биохимии, в частности к моноклональному антителу, которое способно ингибировать димеризацию c-Met. Также раскрыта композиция для профилактики и лечения рака, ассоциированного с c-Met, содержащая указанное антитело. Раскрыто применение указанных антитела и композиции для получения лекарственного средства против рака, ассоциированного с c-Met. Изобретение обладает способностью ингибировать димеризацию c-Met, что позволяет эффективно лечить рак, характеризующийся лиганд-независимой активацией c-Met. 3 н. и 16 з.п. ф-лы, 39 ил., 4 табл., 20 пр.
1. Моноклональное антитело, способное ингибировать димеризацию c-Met, где данное антитело включает тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3 с соответствующими аминокислотными последовательностями SEQ ID No. 1, 2 и 3; и легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3 с соответствующими аминокислотными последовательностями SEQ ID No. 5, 6 и 7, где данное антитело дополнительно характеризуется тем, что также включает шарнирную область, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 22-28 и SEQ ID No. 58-72;
для профилактики или лечения пациента, страдающего раком, характеризующимся лиганд-независимой активацией c-Met.
2. Антитело по п. 1 для применения для профилактики или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся гиперэкспрессией c-Met.
3. Антитело по п. 1 для применения для профилактики или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся гиперэкспрессией c-Met, являющейся результатом генной амплификации c-Met.
4. Антитело по п. 3 для применения для профилактики или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся гиперэкспрессией c-Met, являющейся результатом генной амплификации c-Met и являющейся результатом лиганд-независимой активации c-Met.
5. Антитело по п. 3 для применения для профилактики или лечения пациента, нуждающегося в этом, страдающего раком, характеризующимся гиперэкспрессией c-Met, являющейся результатом генной амплификации c-Met, где рак выбран из группы, состоящей из почечно-клеточной карциномы и рака ЖКТ.
6. Антитело по п. 1, характеризующееся тем, что оно составляет химерное антитело.
7. Антитело по п. 1, характеризующееся тем, что оно составляет антитело человека.
8. Антитело по п. 1, характеризующееся тем, что оно составляет гуманизированное антитело.
9. Антитело по п. 8, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; и вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 8, 9 или 10.
10. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 8; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
11. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 9; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
12. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 10; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 22.
13. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 8; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
14. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 9; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
15. Антитело по п. 9, характеризующееся тем, что оно включает вариабельный домен тяжелой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 4; вариабельный домен легкой цепи последовательности, содержащей аминокислотную последовательность SEQ ID No. 10; и шарнирную область, содержащую аминокислотную последовательность SEQ ID No. 28.
16. Антитело по пп. 1-15 в качестве лекарственного средства.
17. Композиция для профилактики или лечения пациента, страдающего раком, характеризующимся лиганд-независимой активацией c-Met, содержащая эффективное количество антитела по любому из пп. 1-16 в качестве активного ингредиента.
18. Композиция по п.17 в качестве лекарственного средства.
19. Применение антитела по пп. 1-16 или композиции по п. 17 или 18 для получения лекарственного средства для профилактики или лечения пациента, страдающего раком, характеризующимся лиганд-независимой активацией c-Met.
| WO 20090007427, от 15.01.2009 | |||
| WO 2006043892 A1, от 27.04.2006 | |||
| ЭЛАСТОМЕРНЫЕ СОПОЛИМЕРЫ ЭТИЛЕНА И СПОСОБ ИХ ПОЛУЧЕНИЯ | 1994 |
|
RU2143441C1 |
| Переставитель стеклоизделий | 1991 |
|
SU1773885A1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ФАКТОРУ РОСТА ГЕПАТОЦИТОВ | 2004 |
|
RU2361879C2 |

