Данное изобретение относится к области разработки антител и, более конкретно, к способу скрининга антител и/или модуляции агонистической/антагонистической активности антител. Более конкретно изобретение касается способа модуляции антагонистической активности моноклонального антитела, или двухвалентного функционального фрагмента, или его производного с помощью генной инженерии. Изобретение также относится к полипептидам, используемым для такого способа модуляции, и к полученным антителам.
Термины «антитело», «антитела» или «иммуноглобулин» используются как синонимы в самом широком смысле и включают моноклональные антитела (например, моноклональные антитела полной длины или интактные), поликлональные антитела, мультивалентные антитела или мультиспецифические антитела (например, биспецифические антитела до тех пор, пока они проявляют требуемую биологическую активность).
В частности, такая молекула является гликопротеином, содержащим по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, связанные друг с другом дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области (или домена) тяжелой цепи (сокращенной здесь как HCVR или VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенной здесь как LCVR или VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL также можно разделить на области гипервариабельности, называемые областями, определяющими комплементарность (CDR), которые перемежаются более консервативными областями, называемыми каркасными областями (framework regions, FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, в том числе с различными клетками иммунной системы (например, эффекторными клетками) и первым компонентом (C1q) классической системы комплемента.
Тяжелые цепи иммуноглобулинов можно разделить на три функциональные области: область Fd, шарнирную область, область Fc (фрагмент, способный к кристаллизации), которые соединены гибкой шарнирной областью. Область Fd включает домены VH и СН1 и, в сочетании с легкой цепи, образует Fab - антиген-связывающий фрагмент. Fc-фрагмент отвечает за эффекторные функции иммуноглобулина, к которым относятся, например, связывание комплемента и связывание с родственными Fc-рецепторами эффекторных клеток. Шарнирная область, имеющаяся в иммуноглобулинах классов IgG, IgA и IgD, выступает в качестве гибкого разделителя, который позволяет Fab-части свободно перемещаться в пространстве по отношению к Fc-области. В отличие от константных областей, шарнирный домен может быть разнообразным по структуре, различаясь среди классов и подклассов иммуноглобулинов и по последовательности, и по длине.
По данным кристаллографических исследований шарнирную область иммуноглобулина можно также разделить структурно и функционально на три области: верхнюю, ядерную и нижнюю (Shin et al., Immunological Reviews 130: 87, 1992). Верхняя шарнирная область включает аминокислоты с карбоксильного конца СН1 до первого остатка шарнирной области, который ограничивает подвижность, как правило, первого цистеинового остатка, который формирует межцепочечную дисульфидную связь между двумя тяжелыми цепями. Длина верхней шарнирной области коррелирует с сегментарной гибкостью антитела. Ядро шарнирной области содержит дисульфидные мостики между тяжелыми цепями. Нижняя шарнирная область присоединяется к амино-концу домена СН2 и включает его остатки. Ядро шарнирной области человеческого IgG1 содержит последовательность Cys-Pro-Pro-Cys, что при димеризации путем образования дисульфидных связей приводит к образованию циклического октапептида, который, по-видимому, выступает в качестве точки опоры, таким образом обеспечивая подвижность. Конформационные изменения, разрешенные структурой и подвижностью полипептидной последовательности шарнирной области иммуноглобулина, могут влиять на эффекторные функции Fc-части антитела.
Как правило, для получения моноклональных антител мышиного происхождения или их функциональных фрагментов можно сослаться на способы, которые описаны, в частности, в руководстве «Antibodies» (Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor NY, pp.726, 1988), или на методики получения из гибридом, описанные Kohler и Milstein (Nature, 256: 495-497, 1975). Затем моноклональные антитела могут, например, быть очищены на аффинных колонках, на которых был заранее иммобилизован рецептор, представляющий интерес, или один из его фрагментов, содержащих эпитоп, который специфически распознается указанными моноклональными антителами. В частности, указанные моноклональные антитела могут быть очищены с помощью хроматографии на белке А и/или G, после чего следует или не следует ионообменная хроматография, направленная на ликвидацию остаточных загрязнений белком, а также ДНК и ЛПС, следует или не следует эксклюзионная хроматография на сефарозном геле с целью устранения потенциальных агрегатов в связи с наличием димеров или других мультимеров. Еще более предпочтительным образом все эти методики могут быть использованы одновременно или последовательно.
Под функциональным фрагментом антитела в соответствии с изобретением понимается, в частности, фрагмент антитела, такой как фрагменты Fv, scFv (sc обозначает одну цепь), Fab, F(ab')2, Fab', scFv-Fc, или димеры антител или любой фрагмент, время полужизни которого было увеличено в результате химической модификации, такой как добавление поли(алкилен)гликоля, такого как полиэтиленгликоль («пегилирование») (пегилированные фрагменты называются Fv-ПЭГ, scFv-ПЭГ, Fab-ПЭГ, F(ab')2-ПЭГ или Fab'-ПЭГ) («ПЭГ» означает полиэтиленгликоль), либо путем включения в липосому; при этом указанный фрагмент имеет по меньшей мере один характерный CDR исходного антитела.
Предпочтительно этими функциональными фрагментами будут фрагменты типа Fv, scFv, Fab, F(ab')2, F(ab'), scFv-Fc или димерные антитела, которые обычно имеют такую же специфичность связывания, как и антитело, из которого они получены. В соответствии с данным изобретением фрагменты антитела изобретения могут быть получены из антител описанными выше способами, такими как расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных мостиков путем химического восстановления. В другом случае фрагменты антитела могут быть получены с помощью методик генетической рекомбинации, также известных специалистам в данной области, или путем пептидного синтеза, например с помощью автоматических пептидных синтезаторов, например поставляемых компанией Applera, и т.д.
Термин «антагонист», используемый в данном документе, относится к молекуле, которая способна ингибировать одну или более биологическую активность молекулы-мишени, например внеклеточного или трансмембранного рецептора. Антагонисты могут действовать, влияя на связывание рецептора с лигандом и, наоборот, путем уменьшения фосфорилирования рецептора и/или путем выведения или уничтожения клеток, которые были активированы лигандом. Антагонист может полностью блокировать взаимодействия рецептор-лиганд, или могут существенно снизить такие взаимодействия за счет конкуренции, изменения конформации, выпадения или понижающей регуляции. Все такие точки воздействия антагониста рассматриваются как эквивалент для целей данного изобретения.
Термин «агонист», используемый в данном документе, относится к любому соединению, в том числе белку, полипептиду, пептиду, антителу, фрагменту антитела, конъюгату, большой молекуле, небольшой молекуле, способной активировать одну или более среди биологических активностей молекулы-мишени.
В исследовании терапевтических антител часто ожидается присутствие антител в качестве антагонистов, если это возможно.
Классическими примерами антагонистических антител являются антитела герцептин, пертузумаб, цетуксимаб, анти-VEGFR или анти-IGF-1R.
В качестве частного примера можно отметить анти-с-Met-антитело 5D5, созданное Genentech [WO 96/38557], которое проявляет себя на различных моделях как мощный агонист при введении его в одиночку. Для того чтобы решить эту техническую проблему, это антитело было разработано как Fab-фрагмент или как моновалентное антитело (5D5 с одной ветвью), имеющее антагонистическую активность. Как следствие, такое антитело может рассматриваться не как антитело, но как фрагмент, и оно не представляет все преимущества формата «полного антитела» (нет эффекторных функций, снижен клиренс и период полужизни [в два раза быстрее, чем традиционные двухвалентные антитела, описанные на плакате 411 на 20-м симпозиуме EORTC-NCI-AACR, Женева, 21-24 октября, 2008]).
Специалисту в данной области будет понятно, что эффекторные функции включают, например, C1q-связывание; комплемент-зависимую цитотоксичность (complement dependent cytotoxicity, CDC), Fc-рецепторное связывание; антитело-зависимую клеточную цитотоксичность (antibody-dependent cell-mediated cytotoxicity, ADCC); фагоцитоз; и отрицательную регуляцию рецепторов клеточной поверхности (например, В-клеточный рецептор, BCR) и удлинение периода полужизни посредством включения «лиганда связывания спасательного рецептора» (FcRn), описанного, например, в патенте США №5739277 от 14 апреля 1998.
Одним из предлагаемых аспектов данного изобретения является решение таких технических проблем, т.е. усиление антагонистической активности антитела при сохранении «полного двухвалентного» формата.
Здесь следует отметить, что изобретение можно применить к модуляции агонистической/антагонистической активности человеческих антител, полученных путем иммунизации «очеловеченных мышей» (генетически модифицированных мышей, которые продуцируют иммуноглобулины человека) или с использованием методик фагового дисплея для создания целых антител из выбранных scFv, Fab или любых других эквивалентных фрагментов.
Другая классическая техническая проблема может быть решена в ходе химеризации и/или гуманизации мышиного антитела. Специалистам в данной области хорошо известно, что если процесс химеризации и/или гуманизации мышиного антитела в теории очень прост, то не так легко управлять химеризацией и/или гуманизацией такого мышиного антитела без потери всех или части первоначальных свойств. Химерное или гуманизированное антитело может потерять часть его ADCC, CDC, антагонистической/агонистической, связывающей, (ТВС) … активности. Данное изобретение касается, в частности, модификации агонистической/антагонистической активности мышиного антитела после процесса химеризации и/или гуманизации.
В качестве конкретного примера приведен ряд анти-cMet антител, описанных ниже как 224G11, 2274Н1 и 11Е1, которые ведут себя как мощные антагонистические мышиные антитела и которые стали частичными агонистами после химеризации по формату человеческого IgG1. Этот переход от мощных антагонистов к частичным агонистам привел к полной потере активности in vivo на моделях с ксенотрансплантатом.
Данное изобретение предназначено для решения этих проблем и относится, в частности, к способу повышения антагонистической активности моноклонального антитела, направленного против специфической молекулы-мишени, или двухвалентного функционального фрагмента или его производного, при этом указанное антитело способно ингибировать одну или более биологическую активность указанной молекулы-мишени, где указанный способ включает стадию реконфигурации шарнирной области, состоящую в модификации аминокислотной последовательности указанной шарнирной области путем удаления, добавления или замены по меньшей мере одной аминокислоты.
Понятно, что выражение «усиление антагонистической активности» следует понимать в его самом широком смысле, т.е. как желаемый результат. Механистически, такой результат может быть получен путем усиления внутренней антагонистической активности и/или снижения внутренней агонистической активности антитела.
В частности, основные определения терминов в количественной фармакологии основаны на обновленных рекомендациях, предоставленных Комитетом по номенклатуре рецепторов Международного союза фармакологии (IUPHAR) (см. Neubig et al., 2003).
Термин «агонист» обозначает лиганд (любой тип молекул), который связывается с рецептором и изменяет состояние рецептора, что приводит к стимулированию или повышенной биологической реакции. Агонисты могут действовать как полные агонисты или как частичные агонисты:
- Полный агонист: когда рецепторный стимул, индуцированный агонистом, достигает максимального реагирования системы, то он вызывает системную максимальную реакцию и становится полным агонистом в этой системе. Несколько агонистов могут вызвать такой же максимальный ответ, все они являются полными агонистами в этой экспериментальной системе.
- Частичный агонист: молекула, которая в той или иной ткани при определенных условиях не может вызвать такой большой эффект (даже тогда, когда применяется в высокой концентрации, так что все рецепторы должны быть заняты), какой может вызвать полный агонист, воздействующий через те же рецепторы в той же системе. Частичные агонисты, как правило, являются также частичными антагонистами, так как в одновременном присутствии полного агониста они снижают максимальную реакцию указанного полного агониста до их собственного максимального ответа. Это обозначение полного и частичного агониста зависит от системы, и полный агонист для одной системы или измерения может быть частичным агонистом для другой.
Термин «антагонист» обозначает молекулу, которая уменьшает действие другого лекарственного препарата, как правило, агониста. Многие антагонисты действуют на ту же рецепторную макромолекулу, что и агонист.
- Эффективность антагонизма может быть полным антагонизмом, где реакция системы при одновременном присутствии антагониста и агониста соответствует базальной (без какого-либо лиганда) активности системы.
- Антагонист может действовать как частичный антагонист, когда максимальное ингибирование (даже тогда, когда применяется в высокой концентрации, так что все рецепторы должны быть заняты антагонистом), вызванное одновременным присутствием антагониста и агониста, выше базальной активности системы.
- Антагонизм может быть конкурентным, когда связывание агониста и антагониста является взаимоисключающим. Это может быть из-за того, что агонист и антагонист конкурируют за один и тот же сайт связывания или объединяются со смежными сайтами, которые перекрываются. Третья возможность состоит в том, что участвуют различные сайты, но они влияют на рецепторные макромолекулы таким образом, что агонистические и антагонистические молекулы не могут быть связаны одновременно.
- Неконкурентный антагонизм наблюдается, когда агонист и антагонист могут быть связаны с рецептором одновременно; связывание с антагонистом уменьшает или предотвращает действие агониста с или без какого-либо влияния на связывание агониста.
Удаление, добавление или замена может быть проведена классически любым способом, известным специалистам.
Некоторые способы могут быть применены специалистом для выполнения добавления, удаления или вставки в данную последовательность ДНК. Без ограничений можно отметить частичное расщепление ДНК поджелудочной ДНКазой I, частичное расщепление ДНК ферментами рестрикции, основанные на линкере инсерционные мутанты, вложенные наборы делеционных мутантов с помощью нуклеазы BAL31, ДНКазы I или экзонуклеазы III. Эти способы широко описаны в лабораторных пособиях, таких как Molecular Cloning, A laboratory manual (Sambrook, Fritsch and Maniatis). Для выполнения удаления, вставки или сайт-направленного мутагенеза в молекуле ДНК также может быть использовано несколько основанных на ПЦР способов, таких как ПЦР с перекрывающимися праймерами (overlap extension PCR) (Wurch et al., 1998), но не ограничиваясь им. Для выполнения сайт-направленного мутагенеза могут быть использованы некоторые другие методики, например, но не ограничиваясь ими, можно упомянуть мутагенез с помощью олигонуклеотидов на основании одного или двух праймеров, способ Кункеля, основанный на встраивании урацила (Kunkel, 1985). Эти способы широко описаны в лабораторных пособиях, таких как Molecular Cloning, A laboratory manual (Sambrook, Fritsch and Maniatis).
В качестве неограничивающего примера добавления можно отметить добавление пролина в шарнирную область или рядом с ней.
В предпочтительном воплощении способа изобретения указанная модификация выбрана среди:
i) удаления по меньшей мере одной аминокислоты из указанной аминокислотной последовательности шарнирной области, и/или
ii) добавления по меньшей мере одного дисульфидного мостика в указанную шарнирную область.
Для того чтобы внести ясность в изобретение, вначале будет подробно описан первый аспект (i), а затем второй аспект (ii). Следует понимать, что это упорядочение сделано только для написания данной заявки, и что оба эти аспекта, как будет очевидно далее, имеют одинаковую важность.
В частном воплощении способ изменения аминокислотной последовательности шарнирной области будет состоять в удалении не более 2, 3 или 4 аминокислот из указанной аминокислотной последовательности шарнирной области.
Частным аспектом изобретения является то, что указанное моноклональное антитело является двухвалентным антителом. Действительно, как показано ниже, можно модулировать агонистическую/антагонистическую активность антитела, изменяя структуру указанного антитела. В первый раз изобретатели представляют оригинальный способ модуляции такой агонистической/антагонистической активности при сохранении двухвалентной формы антител, направленный на сохранение благоприятных свойств, таких как длительный период полужизни или эффекторные функции.
Здесь также можно упомянуть, что если об изменении шарнирой области моноклонального антитела с целью повышения эффекторных функций уже сообщалось на данном уровне техники, то, напротив, никогда не сообщалось, что такое изменение в шарнирной области может представлять интерес для модуляции агонистической/антагонистической активности моноклонального антитела. Это, несомненно, является предметом данного изобретения, который является новым и изобретенным в отношении существующего уровня техники.
В качестве аспекта в соответствии со способом изобретения моноклональное антитело является химерным антителом.
Под «химерным антителом» понимается антитело, которое содержит природный вариабельный участок (легкой цепи и тяжелой цепи), полученный из антитела данного вида, в сочетании с константными участками легкой цепи и тяжелой цепи антитела вида, гетерологичного указанному данному виду (например, мыши, лошади, кролика, собаки, коровы, курицы и т.д.).
Антитела или их фрагменты химерного типа в соответствии с изобретением могут быть получены с помощью методик генетической рекомбинации. Например, химерное антитело может быть получено путем клонирования рекомбинантной ДНК, содержащей промотор и последовательность, кодирующую вариабельную область нечеловеческого, особенно мышиного, моноклонального антитела в соответствии с изобретением, и последовательность, кодирующую константный участок человеческого антитела. Химерное антитело изобретения, закодированное таким рекомбинантным геном, будет, например, химерой мыши и человека, при этом специфичность этого антитела будет определяться вариабельным участком, полученным из мышиной ДНК, а его изотип будет определяться константным участком, полученным из человеческой ДНК. Для способов получения химерных антител можно сослаться, например, на Verhoeyn et al. (BioEssays, 8: 74, 1988), Morrison et al. (Proc. Natl. Acad. Sci. USA 82: 6851-6855, 1984) или на патент США №4816567.
В качестве другого аспекта в соответствии со способом изобретения моноклональное антитело является гуманизированным антителом.
Под «гуманизированным антителом» понимается антитело, которое содержит CDR-участки, полученные из антитела нечеловеческого происхождения, при этом другие части молекулы антитела получены из одного (или нескольких) человеческих антител или зародышевых последовательностей. Кроме того, некоторые остатки сегментов скелета (называемые FR, каркасные участки) могут быть изменены в целях сохранения аффинности связывания (Jones et al., Nature, 321: 522-525, 1986; Verhoeyen et al., Science, 239: 1534-1536, 1988; Riechmann et al., Nature, 332: 323-327, 1988).
Гуманизированные антитела в соответствии с изобретением или их фрагменты могут быть получены с помощью методик, известных специалистам в данной области (например, таких, которые описаны в документах Singer et al., J. Immun. 150: 2844-2857, 1992; Mountain et al., Biotechnol. Genet. Eng. Rev., 10: 1-142, 1992; и Bebbington et al., Bio/Technology, 10: 169-175, 1992).
Другим способом гуманизации, также известным специалистам в данной области, является, например, способ «CDR-прививки», описанный Protein Design Lab (PDL) в патентных заявках ЕР 0451261, ЕР 0682040, ЕР 09127, ЕР 0566647 или US 5530101, US 6180370, US 5585089 и US 5693761. Также можно отметить следующие патентные заявки: US 5639641; US 6054297; US 5886152 и US 5877293.
В качестве другого аспекта в соответствии со способом изобретения моноклональное антитело является человеческим антителом.
Термин «человеческое антитело» включает в себя все антитела, которые имеют одну или более вариабельную и константную область, полученную из человеческих иммуноглобулиновых последовательностей. В предпочтительном воплощении все вариабельные и константные домены (или области) получены из человеческой иммуноглобулиновой последовательности (полного человеческого антитела). Другими словами, он включает любое антитело, в котором вариабельные и константные области (если имеются) получены из человеческих зародышевых иммуноглобулиновых последовательностей, т.е. которое обладает аминокислотной последовательностью, соответствующей последовательности антитела, продуцируемого человеком и/или сделанного с помощью любых методик получения человеческих антител, известных специалистам в данной области.
В одном воплощении человеческие моноклональные антитела продуцируются гибридомой, которая содержит В-клетку, полученную из трансгенного животного (не человека), например трансгенной мыши, имеющей геном с трансгеном человеческой тяжелой цепи и трансгеном легкой цепи, слитую с иммортализованной клеткой.
В качестве примера такой трансгенной мыши можно отметить XENOMOUSE™, которая является разработанной мышиной линией, которая состоит из крупных фрагментов человеческих иммуноглобулиновых локусов и является дефицитной по продукции мышиного антитела (Green at al., 1994, Nature Genetics, 7: 13-21). XENOMOUSE™ продуцирует набор полных антител взрослого человека, а также образует антиген-специфические человеческие моноклональные антитела. Второе поколение XENOMOUSE™ содержит примерно 80% репертуара антител человека (Green & Jakobovits, 1998, J. Exp. Med., 188: 483-495).
Для создания человеческого антитела в соответствии с изобретением также может быть использована любая другая методика, известная специалистам в данной области, такая как методика фагового дисплея.
Способ в соответствии с изобретением может быть использован для любого типа иммуноглобулина, содержащего шарнирную область, т.е. для IgA, IgD и IgG.
В качестве примера, для изотипа IgA шарнирная область IgA1 содержит аминокислотную последовательность PSTPPTPSPSTPPTPSPS (SEQ ID №8), а шарнирная область IgA2 содержит аминокислотную последовательность РРРРР (SEQ ID №9).
Таким же образом шарнирная область IgD содержит аминокислотную последовательность
SPKAQASSVPTAQPQAEGSLAKATTAPATTRNTRGGEEKKKEKEKEEQEERETKTP (SEQ ID №10).
В качестве частного воплощения изобретения предпочтительно применять IgG, в том числе, например, IgG1, IgG2, IgG3 или IgG4.
Аминокислотные последовательности, соответствующие шарнирным областям различных изотипов IgG, представляют собой:
PKSCDKTHTCPPCP (SEQ ID №11) для IgG1,
RKCCVECPPCP (SEQ ID №7) для IgG2,
LKTPLFTGDTTHTCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCPRCPEPKSCDTPPPCPRCP (SEQ ID №12) для IgG3, и
SKYGPPCPSCP (SEQ ID №13) для IgG4.
Еще более конкретно, предпочтительно использовать IgG1. Действительно, в области терапевтических антител и, в частности, в лечении рака предпочтительным является создание IgG1 для получения эффекторных функций, таких как ADCC и CDC, в дополнение к функциям, связанным со специфическим связыванием целевого антигена.
Способ изобретения характеризуется тем, что указанное моноклональное антитело представляет собой IgG1.
Понятие «молекула-мишень» в рамках изобретения относится к любой молекуле, с которой моноклональное антитело способно специфически связываться или модулировать активность. Как правило, такая молекула-мишень может быть названа «антигеном».
В качестве неограничивающего примера молекулы-мишени, которая может стать мишенью моноклонального антитела, можно упомянуть растворимые лиганды, рецепторы, такие как трансмембранные рецепторы, мембранные опухолевые маркеры и т.д.
В предпочтительном воплощении указанная молекула-мишень является трансмембранным рецептором.
Выражение «трансмембранный рецептор» относится к белку, который проходит через плазматическую мембрану клетки, с внеклеточным доменом белка, способным связываться с лигандом, и внутриклеточным доменом с активностью (такой как протеинкиназная), которая может измениться (увеличиться или уменьшиться) при связывании лиганда. Другими словами, трансмембранные рецепторы являются интегральными мембранными белками, которые находятся и обычно работают в клеточной плазматической мембране, а также в мембранах некоторых субклеточных компартментов и органелл. Связываясь с сигнальной молекулой или иногда с парой таких молекул с одной стороны мембраны, трансмембранные рецепторы вызывают ответ на другой стороне. Таким образом, они играют уникальную и важную роль в связывании клеток и передаче сигнала.
Многие трансмембранные рецепторы состоят из двух или более белковых субъединиц, которые действуют совместно и могут диссоциировать, отпадать, когда связывают лиганды или на другом этапе их циклов «активации». Они часто классифицируются на основе их молекулярной структуры или, из-за того что структура детально неизвестна для всех рецепторов, кроме нескольких, классифицируются на основе их предположительной (а иногда и экспериментально проверенной) мембранной топологии. Полипептидные цепи самых простых, вероятно, пересекают билипидный слой только один раз, тогда как другие пересекают его большее количество раз, например семь раз (так называемые рецепторы, связанные с G-белком, или GPCR) или более.
Как и любой интегральный мембранный белок, трансмембранный рецептор может быть разделен на три части или домена, внеклеточный домен, трансмембранный домен и внутриклеточный домен.
Внеклеточный домен является частью рецептора, которая торчит из мембраны на внешней стороне клетки или органеллы. Если полипептидная цепь рецептора пересекает бислой несколько раз, внешний домен может состоять из нескольких «петель», торчащих из мембраны. По определению, основной функцией рецептора является распознавание и ответ на специфический лиганд, например медиатор или гормон (хотя некоторые рецепторы отвечают также на изменения трансмембранного потенциала), и во многих рецепторах эти лиганды связываются с внеклеточным доменом.
В большинстве рецепторов, для которых доказана структура, трансмембранные альфа-спирали составляют большую часть трансмембранного домена. В некоторых рецепторах, таких как никотиновый ацетилхолиновый рецептор, трансмембранный домен формирует связанную с белком пору через мембрану или ионный канал. После активации внеклеточного домена путем связывания соответствующего лиганда пора становится доступной для ионов, которые затем проходят через нее. В других рецепторах трансмембранные домены предположительно подвергаются конформационному изменению при связывании, что оказывает влияние внутриклеточно. В некоторых рецепторах, таких как члены надсемейства 7ТМ, трансмембранный домен может содержать лиганд-связывающий карман.
Внутриклеточный (или цитоплазматический) домен рецептора взаимодействует с внутренней частью клетки или органеллы, ретранслируя сигнал. Есть два принципиально разных способа этого взаимодействия: а) Внутриклеточный домен осуществляет связь посредством специфических белок-белковых взаимодействий с эффекторными белками, которые в свою очередь посылают сигнал по сигнальной цепи к месту назначения; и б) у фермент-связанных рецепторов внутриклеточный домен имеет ферментативную активность. Часто это тирозинкиназная активность. Ферментативная активность также может быть расположена на ферменте, связанном с внутриклеточным доменом.
У клетки есть несколько способов регулировать активность трансмембранного рецептора. Большинство из них работает через внутриклеточный домен. Наиболее важными способами являются фосфорилирование и интернализация (см. убиквитин) или активация каскадов вторичных посредников, таких как цАМФ, IP, Ca2+ или цГМФ.
Все мембранные белки, демонстрирующие ферментативную активность, также могут быть мишенью для антител с модификацией, описанной в данном изобретении. Можно упомянуть в качестве примеров, но без ограничений, семейство матричных металлопротеиназ (ММР), семейство «дезинтегринов в комплексе с металлопротеазамипротеазными доменами» (ADAM), аденилатциклазы, …
Все мембранные белки, действующие как ионные каналы, поры и переносчики, также могут быть мишенью для антител с модификацией, описанной в данном изобретении. Можно упомянуть в качестве примеров, но без ограничений, семейство натриевых каналов, семейство калиевых каналов, семейство никотиновых ацетилхолиновых рецепторов, сигма-рецепторы, семейство транспортеров моноаминов.
В более широком смысле все мембранные белки, идентифицированные как специфические маркеры для данного заболевания, также могут быть мишенью для лечения антителом, при этом антитело также может быть улучшено путем модификаций, описанных в данном изобретении.
В предпочтительном воплощении изобретения указанный трансмембранный рецептор выбран из группы, состоящей из тирозинкиназных рецепторов, тетраспанина и GPCR.
В более предпочтительном воплощении указанный трансмембранный рецептор представляет собой тирозинкиназный рецептор, выбранный преимущественно из группы, состоящей из IGF-1R, c-Met, RON, AxI, VEGF, VEGFR, Ee-2neu, гомодимеров и гетеродимеров семейства ErbB и т.д.
В данной заявке и, в частности, в следующем описании последовательности будут определены в соответствии с определением IMGT. Уникальная нумерация IMGT была создана для сравнения вариабельных доменов независимо от антигенного рецептора, типа цепи или вида [Lefranc M.-P., Immunology Today 18, 509 (1997) / Lefranc M.-P., The Immunologist, 7, 132-136 (1999) / Lefranc, M.-P., Pommie, С., Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Comp. Immunol., 27, 55-77 (2003)]. В уникальной нумерации IMGT консервативные аминокислоты всегда занимают одну и ту же позицию, например цистеин 23 (1-CYS), триптофан 41 (CONSERVED-TRP), гидрофобная аминокислота 89, цистеин 104 (2-CYS), фенилаланин или триптофан 118 (J-PHE или J-TRP). Уникальная нумерация IMGT предусматривает стандартизированное разграничение каркасных участков (FR1-IMGT: позиции 1-26, FR2-IMGT: 39-55, FR3-IMGT: 66-104 и FR4-IMGT: 118-128) и участков, определяющих комплементарность: CDR1-IMGT: 27-38, CDR2-IMGT: 56-65 и CDR3-IMGT: 105-117. Т.к. разрывы представляют собой незанятые позиции, то длины CDR-IMGT (показаны в скобках и разделены точками, например [8.8.13]) становятся важной информацией. Уникальная нумерация IMGT используется в 20-графическом представлении, обозначенном как IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)], и в 3D-структурах в IMGT/3Dstructure-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., T cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)].
Для специалиста в данной области будет легко перевести изобретение, описанное в соответствии с системой IMGT, в любую другую систему нумерации, такую как, например, система нумерации Кабата.
Уникальная нумерация IMGT для всех IG и TR V-областей всех видов основывается на высокой консервативности структуры вариабельной области. Эта нумерация, созданная после выравнивания более 5000 последовательностей, учитывает и сочетает определение каркасной (FR) и определяющей комплементарность областей (CDR), структурные данные исследований рентгеновской дифракции и характеристику гипервариабельных петель. Было проведено установление границ областей FR-IMGT и CDR-IMGT. Точно так же уникальная нумерация IMGT была применена для С-домена и позволила точно разграничить Ig-подобные домены. С-домен соответствует полной С-области, большей части С-области или только части С-области в зависимости от типа иммуноглобулина (IG).
Нумерация IMGT для С-домена (IG и TR) была получена как уникальная нумерация IMGT для V-области по принципам уникальной нумерации IMGT для V-области до позиции 104. Таким образом, можно легко сравнить аминокислотные позиции в С-домене и V-домене.
Чтобы точно определить шарнирную область, нумерация IMGT С-домена была применена для точной локализации доменов СН1 и СН2. Шарнирная область включает все аминокислотные остатки между последним остатком IMGT-CH1 и первым остатком IMGT-CH2.
Все остальные схемы нумерации иммуноглобулинов, например Кабата или А.Онеггера, для того же шарнирного домена, включены в данное изобретение.
В качестве предпочтительного примера изобретения, основанного на описанной выше системе нумерации IMGT, аминокислотная последовательность шарнирной области IgG1 включает остатки Н1-Н14 с сегментом Н1-Н9, соответствующим верхней шарнирной области, и с сегментом Н10-Н14, соответствующим ядру шарнирной области. Более конкретно, человеческая IgG1 шарнирная область состоит из аминокислотной последовательности PKSCDKTHTCPPCP (SEQ ID №11), а мышиная IgG1 шарнирная область состоит из аминокислотной последовательности PRDCGCKPCICT (SEQ ID №14).
В предпочтительном воплощении изобретения рассматривается модификация, направленная на уменьшение длины белковой последовательности, кодирующей шарнирную область двухвалентного антитела. В частности, способ в соответствии с изобретением включает этап удаления по меньшей мере одной аминокислоты в шарнирной области.
Как упоминалось ранее, предпочтительным является удаление не более двух аминокислот указанной шарнирной области.
Как упоминалось ранее, предпочтительным является удаление не более трех аминокислот указанной шарнирной области.
Как упоминалось ранее, предпочтительным является удаление не более четырех аминокислот указанной шарнирной области.
В частном применении способа изобретения модификация состоит по меньшей мере в удалении аминокислоты, выбранной среди аминокислот в позициях Н1, Н2, Н3, Н5, Н6, Н7, Н8, Н9, Н11, Н12 или Н14.
Более конкретно, авторы изобретения продемонстрировали включение конкретного остатка, и конкретный аспект изобретения состоит в выборе определенных остатков.
В предпочтительном случае IgG1 аминокислота в позиции Н1 является пролином; аминокислота в позиции Н2 является лизином в человеческой версии и аргинином в мышиной версии; аминокислота в позиции Н3 является серином в человеческой версии и аспартатом в мышиной версии; аминокислота в положении Н5 является аспартатом в человеческой версии и глицином в мышиной версии; аминокислота в позиции Н6 является лизином; аминокислота в позиции Н8 является гистидином в человеческой версии и лизином в мышиной версии; аминокислота в позиции Н9 является треонином в человеческой версии и пролином в мышиной версии; аминокислота в позиции Н11 является пролином в человеческой версии и изолейцином в мышиной версии; и аминокислота в позиции Н12 является пролином в человеческом версии.
В соответствии с предпочтительным воплощением это удаление должно быть сделано в «верхней шарнирной» области.
В более предпочтительном воплощении это удаление является частью «верхней шарнирной» области, состоящей, например для IgG1, из аминокислот Н1-Н9 в сравнении с «ядерной шарнирной» областью, состоящей из аминокислот Н10-Н14.
В данной заявке нумерация аминокислот осуществляется по системе IMGT, описанной выше. Очевидно, что любая другая система нумерации с модификацией нумерации, но не природы замещенного остатка в шарнирной области, должна рассматриваться как эквивалентная. В качестве примера изменение нумерации идентифицированной аминокислотной части изобретения (в соответствии с системой IMGT) в системе Кабат должно рассматриваться как эквивалент.
Другой аспект изобретения основан на удалении по меньшей мере одного цистеина в «верхней шарнирной» области, предпочтительно находящегося в позиции Н4.
Другой аспект изобретения основан на добавлении по меньшей мере одного дисульфидного мостика в шарнирной области.
Более конкретно, способ изобретения характеризуется тем, что модификация состоит в введении по меньшей мере одного цистеина в «верхнюю шарнирную» область.
По словам изобретателей, правдоподобное объяснение основано на возможности «ужесточения» шарнира в результате или сокращения длины, и/или введения другого дисульфидного мостика. Такое «ужесточение» позволит поддерживать соответствующую пространственную конформацию антитела с повышенной, как следствие, антагонистической активностью.
Ясно, что любой способ, направленный на ужесточение шарнирной области, следует рассматривать как способ, эквивалентный способу в соответствии с данным изобретением.
Введение цистеина может быть сделано путем добавления такой аминокислоты, при этом указанное добавление проводится любым способом, известным специалисту в данной области.
Другой предпочтительный способ введения цистеина в шарнирную область состоит в замене по меньшей мере одной аминокислоты.
Более конкретно, предпочтительный способ введения цистеина в шарнирную область состоит в замене по меньшей мере аминокислоты, выбранной среди Н1-Н9. Такая замена может быть сделана любым способом, известным специалистам.
Более конкретно, способ изобретения включает замену треонина в позиции Н7 в «верхней шарнирной» области цистеином.
В другом воплощении способ изобретения включает замену лизина в позиции Н6 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену пролина в положении Н1 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену лизина в позиции Н2 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену серина в позиции Н3 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену аспартата в позиции Н5 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену гистидина в позиции Н8 в «верхней шарнирной» области цистеином.
В еще одном воплощении способ изобретения включает замену треонина в позиции Н9 в «верхней шарнирной» области цистеином.
В соответствии с другим воплощением также можно уменьшить длину шарнирной области и/или добавить дисульфидный мостик, изменяя всю аминокислотную последовательность, кодирующую шарнирную область.
В качестве предпочтительного примера, изменение способа изобретения включает замену аминокислот Н1-Н14 из шарнирной области IgG1 аминокислотами Н1-Н14 из шарнирной области IgG2, предпочтительно когда указанное моноклональное антитело, антагонистическую активность которого необходимо улучшить, является антителом IgG1.
В другом приложении изобретение относится к способу выбора антагонистического моноклонального антитела, направленного против специфической молекулы-мишени, или двухвалентного функционального фрагмента или его производного, при этом указанное антитело способно ингибировать одну или более биологическую активность указанной молекулы-мишени, где указанный способ включает этапы:
(а) выбора начального антитела с начальным уровнем ингибирования указанной одной или более биологической активности указанной молекулы-мишени,
(б) изменения аминокислотной последовательности шарнирной области указанного исходного антитела с помощью способа изобретения,
(в) оценки модифицированного антитела этапа (б) в плане его способности ингибировать указанную одну или более биологическую активность указанной молекулы-мишени, и
(г) выбора, как положительный результат, антитела этапа (в) с уровнем ингибирования указанной одной или более биологической активности указанной молекулы-мишени, более высоким, чем исходный уровень указанного ингибирования.
Исходные антитела могут быть выбраны среди существующих антител, таких как, но не ограничиваясь ими, антитела-антагонисты к IGF-1R, c-Met, RON, AxI, CD151, VEGF, VEGFR, Ee-2neu, гомодимеры и гетеродимеры семейства ErbB. В качестве неограничивающего предпочтительного примера указанные исходные антитела могут состоять из герцептина, пертузумаба, цетуксимаба, анти-VEGFR или анти-IGF-1R антител.
«Ингибирующий уровень» в отношении изобретения иллюстрирует антагонистическую активность антитела. Такой ингибирующий уровень может быть определен любым способом, известным специалисту в данной области, таким как, но не ограничиваясь им: а) прямым подсчетом клеток или с использованием 3[H]тимидина, солей тетразолия или любых других флуоресцентных средств для оценки пролиферации, б) вестерн-блоттингом, фосфо-ИФА или альфа-скрининговым анализом для контроля передачи сигнала, в) анализом BRET или FRET для анализа димеризации, г) микроскопией или флуоресцентными способами контроля миграции, инвазии, ангиогенеза или морфогенеза и е) измерением величины опухоли для оценки in vivo.
Этот скрининговый способ может быть использован для улучшения подтвержденных антител или как отборочный этап для научных или доклинических исследований антител.
Другой аспект в соответствии с изобретением относится к моноклональному антителу, направленному против специфической молекулы-мишени, или к двухвалентным функциональным фрагментам или их производным, получаемым с помощью способа изобретения, при этом указанное антитело характеризуется тем, что оно содержит аминокислотную последовательность шарнирной области, выбранную из группы, состоящей из SEQ ID №1 (PRDCGCKPCICT), SEQ ID №2 (PKSCGCKPCICT), SEQ ID №3 (PKSCGCKPCICP), SEQ ID №4 (PRDCGCKPCPPCP), SEQ ID №5 (PRDCGCHTCPPCP), SEQ ID №6 (PKSCDCHCPPCP), SEQ ID №7 (RKCCVECPPCP), SEQ ID №22 (CKSCDKTHTCPPCP), SEQ ID №23 (PCSCDKTHTCPPCP), SEQ ID №24 (PKCCDKTHTCPPCP), SEQ ID №25 (PKSCCKTHTCPPCP), SEQ ID №26 (PKSCDCTHTCPPCP), SEQ ID №27 (PKSCDKCHTCPPCP), SEQ ID №28 (PKSCDKTCTCPPCP), SEQ ID №29 (PKSCDKTHCCPPCP), SEQ ID №30 (PKSCDKTHTCCPCP), SEQ ID №31 (PKSCDKTHTCPCCP), SEQ ID №32 (PKSCDKTHTCPPCC), SEQ ID №33 (PSCDKTHTCPPCP), SEQ ID №34 (PKSCDTHTCPPCP), SEQ ID №35 (PKSCDKTHCPPCP), SEQ ID №36 (KCDKTHTCPPCP), SEQ ID №37 (PSCKTHTCPPCP), SEQ ID №38 (PKSCDTHCPPCP), SEQ ID №39 (PKSCTHTCPPCP), SEQ ID №40 (PKSCDKTTCPCP), SEQ ID №41 (PKSCDKTHCPPC), SEQ ID №42 (PKSCDCHTCPPCP), SEQ ID №43 (PKSCDCTHCPPCP), SEQ ID №44 (PCSCKHTCPPCP), SEQ ID №45 (PSCCTHTCPPCP), SEQ ID №46 (PSCDKHCCPPCP), SEQ ID №47 (PKSTHTCPPCP), SEQ ID №48 (PKSCTCPPCP) или SEQ ID №49 (PKSCDKCVECPPCP).
Предпочтительное моноклональное антитело, полученное при реализации способа изобретения, можно охарактеризовать тем, что оно содержит аминокислотную последовательность, выбранную из группы, состоящей SEQ ID №1 (PRDCGCKPCICT), SEQ ID №2 (PKSCGCKPCICT), SEQ ID №3 (PKSCGCKPCICP), SEQ ID №4 (PRDCGCKPCPPCP), SEQ ID №5 (PRDCGCHTCPPCP), SEQ ID №6 (PKSCDCHCPPCP), SEQ ID №7 (RKCCVECPPCP), SEQ ID №22 (CKSCDKTHTCPPCP), SEQ ID №23 (PCSCDKTHTCPPCP), SEQ ID №24 (PKCCDKTHTCPPCP), SEQ ID №25 (PKSCCKTHTCPPCP), SEQ ID №26 (PKSCDCTHTCPPCP), SEQ ID №27 (PKSCDKCHTCPPCP), SEQ ID №28 (PKSCDKTCTCPPCP), SEQ ID №29 (PKSCDKTHCCPPCP), SEQ ID №30 (PKSCDKTHTCCPCP), SEQ ID №31 (PKSCDKTHTCPCCP), SEQ ID №32 (PKSCDKTHTCPPCC), SEQ ID №33 (PSCDKTHTCPPCP), SEQ ID №34 (PKSCDTHTCPPCP), SEQ ID №35 (PKSCDKTHCPPCP), SEQ ID №36 (KCDKTHTCPPCP), SEQ ID №37 (PSCKTHTCPPCP), SEQ ID №38 (PKSCDTHCPPCP), SEQ ID №39 (PKSCTHTCPPCP), SEQ ID №40 (PKSCDKTTCPCP), SEQ ID №41 (PKSCDKTHCPPC), SEQ ID №42 (PKSCDCHTCPPCP), SEQ ID №43 (PKSCDCTHCPPCP), SEQ ID №44 (PCSCKHTCPPCP), SEQ ID №45 (PSCCTHTCPPCP), SEQ ID №46 (PSCDKHCCPPCP), SEQ ID №47 (PKSTHTCPPCP), SEQ ID №48 (PKSCTCPPCP) или SEQ ID №49 (PKSCDKCVECPPCP).
В предпочтительном воплощении указанное моноклональное антитело является человеческим антителом, более предпочтительно оно является антителом IgG1.
Изобретение также относится к изолированной нуклеиновой кислоте, кодирующей описанное выше моноклональное антитело, т.е. содержащей аминокислотную последовательность шарнирной области, выбранную из группы, состоящей из SEQ ID №1 (PRDCGCKPCICT), SEQ ID №2 (PKSCGCKPCICT), SEQ ID №3 (PKSCGCKPCICP), SEQ ID №4 (PRDCGCKPCPPCP), SEQ ID №5 (PRDCGCHTCPPCP), SEQ ID №6 (PKSCDCHCPPCP), SEQ ID №7 (RKCCVECPPCP), SEQ ID №22 (CKSCDKTHTCPPCP), SEQ ID №23 (PCSCDKTHTCPPCP), SEQ ID №24 (PKCCDKTHTCPPCP), SEQ ID №25 (PKSCCKTHTCPPCP), SEQ ID №26 (PKSCDCTHTCPPCP), SEQ ID №27 (PKSCDKCHTCPPCP), SEQ ID №28 (PKSCDKTCTCPPCP), SEQ ID №29 (PKSCDKTHCCPPCP), SEQ ID №30 (PKSCDKTHTCCPCP), SEQ ID №31 (PKSCDKTHTCPCCP), SEQ ID №32 (PKSCDKTHTCPPCC), SEQ ID №33 (PSCDKTHTCPPCP), SEQ ID №34 (PKSCDTHTCPPCP), SEQ ID №35 (PKSCDKTHCPPCP), SEQ ID №36 (KCDKTHTCPPCP), SEQ ID №37 (PSCKTHTCPPCP), SEQ ID №38 (PKSCDTHCPPCP), SEQ ID №39 (PKSCTHTCPPCP), SEQ ID №40 (PKSCDKTTCPCP), SEQ ID №41 (PKSCDKTHCPPC), SEQ ID №42 (PKSCDCHTCPPCP), SEQ ID №43 (PKSCDCTHCPPCP), SEQ ID №44 (PCSCKHTCPPCP), SEQ ID №45 (PSCCTHTCPPCP), SEQ ID №46 (PSCDKHCCPPCP), SEQ ID №47 (PKSTHTCPPCP), SEQ ID №48 (PKSCTCPPCP) или SEQ ID №49 (PKSCDKCVECPPCP).
В соответствии с еще одним аспектом данное изобретение относится к изолированной нуклеиновой кислоте, характеризующейся тем, что она выбрана среди следующих нуклеиновых кислот:
а) нуклеиновой кислоты, ДНК или РНК, кодирующей искусственную шарнирную область в соответствии с изобретением, соответствующей РНК или комплементарной последовательности;
б) изолированной нуклеиновокислотной последовательности, содержащей нуклеиновую последовательность, выбранную из группы, включающей от SEQ ID №15 до SEQ ID №21, от SEQ ID №50 до SEQ ID №77, соответствующей РНК и комплементарной последовательности; и
в) нуклеиновой кислоты по меньшей мере с 18 нуклеотидами, способной к гибридизации в условиях высокой жесткости по меньшей мере с одной из последовательностей от SEQ ID №15 до 21 и от 50 до 77.
Предпочтительно изобретение включает изолированную нуклеиновую кислоту с нуклеиновой последовательностью, выбранной из группы, включающей от SEQ ID №15 до SEQ ID №21 и от SEQ ID №50 до SEQ ID №77.
Другая часть изобретения включает вектор экспрессии или трансформированную принимающую клетку (клетку-хозяина), содержащую описанную выше изолированную нуклеиновую кислоту и, в частности, изолированную нуклеиновую кислоту с нуклеиновой последовательностью, выбранной из группы, включающей от SEQ ID №15 до SEQ ID №21 и от SEQ ID №50 до SEQ ID №77, соответствующую РНК и комплементарную последовательность.
Под нуклеиновой кислотой, нуклеиновой или нуклеиновокислотной последовательностью, полинуклеотидом, олигонуклеотидом, полинуклеотидной последовательностью, нуклеотидной последовательностью, терминами, которые равнозначно используются в данном изобретении, понимается четкая последовательность нуклеотидов, модифицированных или не модифицированных, определяющая фрагмент или область нуклеиновой кислоты, содержащая или не содержащая неприродные нуклеотиды и являющаяся либо двуцепочечной ДНК, либо одноцепочечной ДНК, либо продуктами транскрипции указанных ДНК.
Здесь также следует упомянуть, что данное изобретение не относится к нуклеотидным последовательностям в их природной хромосомной среде, т.е. в природном состоянии. Оно касается последовательностей, которые были выделены и/или очищены, т.е. были взяты прямо или косвенно, например путем копирования, при этом их среда была по меньшей мере частично модифицирована. Таким образом, также здесь следует подразумевать выделенные нуклеиновые кислоты, полученные путем генетической рекомбинации, например с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
Гибридизация в условиях высокой жесткости означает, что температурные условия и условия ионной силы выбирают таким образом, чтобы они позволяли сохранять гибридизацию двух комплементарных фрагментов ДНК. В качестве примера, условия высокой жесткости этапа гибридизации для определения описанных выше полинуклеотидных фрагментов преимущественно являются следующими.
ДНК-ДНК- или ДНК-РНК-гибридизацию осуществляют в два этапа: (1) прегибридизация при 42°С в течение трех часов в фосфатном буфере (20 мМ, рН 7,5), содержащем 5× SSC (цитратно-солевой буфер) (1× SSC соответствует раствору 0,15 М NaCl+0,015 М цитрата натрия), 50% формамида, 7% додецилсульфата натрия (SDS), 10× раствор Денхардта, 5% сульфата декстрана и 1% ДНК спермы лосося; (2) собственно гибридизация в течение 20 часов при температуре в зависимости от длины зонда (например: 42°С для зонда длиной более 100 нуклеотидов), а затем два 20-минутных промывания при температуре 20°С в 2× SSC+2% SDS, одно 20-минутное промывание при 20°С в 0,1× SSC+0,1% SDS. Последнее промывание осуществляют в 0,1× SSC+0,1% SDS в течение 30 минут при температуре 60°С для зонда длиной более 100 нуклеотидов. Гибридизационные условия высокой жесткости, описанные выше для полинуклеотида определенного размера, специалист в данной области может адаптировать для олигонуклеотидов большей или меньшей длины в соответствии с описанием в Sambrook, et al. (Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory; 3rd edition, 2001).
Изобретение также относится к вектору, содержащему нуклеиновую кислоту в соответствии с данным изобретением.
Изобретение особенно нацелено на клонирующие и/или экспрессионные векторы, которые содержат нуклеотидную последовательность в соответствии с изобретением.
Векторы в соответствии с изобретением предпочтительно содержат элементы, которые позволяют экспрессировать и/или секретировать транслированные нуклеотидные последовательности в определенной принимающей клетке (клетке-хозяине). Таким образом, вектор должен содержать промотор, сигналы инициации и терминации трансляции, а также подходящие участки регуляции транскрипции. Он должен быть способен поддерживать в стабильном состоянии принимающую клетку и дополнительно может иметь специфические сигналы, которые обуславливают секрецию транслированного белка. Специалисты в данной области выбирают и оптимизируют эти различные элементы в соответствии с функцией используемой принимающей клетки. Для этого нуклеотидные последовательности в соответствии с изобретением могут быть введены в векторах автономной репликации в выбранного хозяина или могут быть интегративными векторами выбранного хозяина.
Такие векторы получают с помощью способов, использующихся в настоящее время специалистами в данной области, и полученные клоны могут быть введены в подходящего хозяина с помощью стандартных способов, таких как липофекция, электропорация, тепловой шок или химические способы.
Векторы в соответствии с изобретением представляют собой, например, векторы плазмидного или вирусного происхождения. Они используются для трансформации принимающих клеток, чтобы клонировать или экспрессировать нуклеотидные последовательности в соответствии с изобретением.
Изобретение также включает принимающие клетки, трансформированные с помощью вектора в соответствии с изобретением или содержащие его.
Принимающая клетка может быть выбрана из прокариотических или эукариотических систем, например бактериальных клеток, а также дрожжевых клеток или клеток животных, в частности клеток млекопитающих. Можно также использовать клетки насекомых или растений.
Другие характеристики и преимущества изобретения появятся в продолжении описания с примерами и графическими материалами, где:
Фиг.1А и 1В: влияние серий мышиных и соответствующих химерных анти-с-Met MKA, продуцируемых как человеческий IgG1/каппа изотип, на фосфорилирование c-Met-рецептора на клетках А549.
Фиг.1А: агонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.1В: антагонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.2А и 2В: сравнение влияния мышиного MKA 224G11 и химерных MKA 224G11, содержащих различные разработанные шарнирные области, на фосфорилирование c-Met-рецептора на клетках А549.
Фиг.2А: агонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.2В: антагонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.3А и 3В, 4 и 5: BRET-модели c-Met-димеризации и активации.
Фиг.6А и 6В: распознавание c-Met химерными и гуманизированными формами 224G11.
Фиг.7А, 7В и 7С: влияние мышиных и химерных антител на HGF-индуцированную пролиферацию клеток NCl-H441 in vitro. Клетки NCI-H441 помещали в бессывороточную среду. Через двадцать четыре часа после посева добавляли (фиг.7А) m11E1 и [11E1]chim, (Фиг.7 В) m227H1 и [227H1]chim или (фиг.7) m224G11 и [224G11]chim в отсутствие или в присутствии HGF. Черные стрелки указывают лунки, покрытые только клетками либо в отсутствие (стрелка влево), либо в присутствии (стрелка вправо) HGF. Мышиный IgG1 (mIgG1) вводили в качестве изотипического контроля.
Фиг.8: сравнение in vivo мышиных и химерных MKA 224G11 на модели с ксенотрансплантатом NCl-H441.
Фиг.9А и 9В: влияние мышиного МКА 224G11 и различных химерных и гуманизированных версий этого антитела на HGF-индуцированную пролиферацию клеток NCl-H441 in vitro. Клетки NCl-H441 помещали в бессывороточную среду. Через двадцать четыре часа после посева добавляли тестируемые антитела в отсутствие или в присутствии HGF. На панели (фиг.9А) показаны мышиная m224G11, химерная IgG1 [224G11]chim, гуманизированная IgG1 [224G11] [Hz1], [224G11] [Hz2], [224G11] [Hz3] версии. На панели (фиг.9 В) представлены мышиная m224G11 и различные химерные IgG1 формы ([224G11]chim, [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [ММСН chim], [224G11] [ТН7 chim]). Черные стрелки указывают лунки с клетками только в отсутствие (стрелка влево) или в присутствии (стрелка вправо) HGF. Мышиный IgG1 (mIgG1) вводили в качестве отрицательного контроля агонистической активности. M5D5 использовали в качестве дозозависимого полного агонистического контроля.
Фиг.10: влияние мышиного МКА 224G11 и различных химерных и гуманизированных версий этого антитела на HGF-индуцированную пролиферацию клеток NCI-H441 in vitro. Клетки NCl-H441 помещали в бессывороточную среду. Через двадцать четыре часа после посева добавляли тестируемые антитела в отсутствие или в присутствии HGF. Представлены мышиная m224G11, химерные формы IgG1 [224G11]chim, [224G11] [TH7 chim]) и [224G11] [ТН7 Hz1], [224G11] [ТН7 Hz3]). Черные стрелки указывают на лунки с клетками только в отсутствие (стрелка влево) или в присутствии (стрелка вправо) HGF. Мышиный IgG1 (mIgG1) вводили в качестве отрицательного контроля агонистической активности. M5D5 использовали в качестве дозозависимого полного агонистического контроля.
Фиг.11А-11В и 12А-12В: влияние серий анти-c-Met-MKA изобретения с разработанной шарнирной областью на фосфорилирование c-Met-рецептора на клетках А549.
Фиг.11А и 11В: агонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.12А и 12В: антагонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.13А и 13В: BRET-модели c-Met-димеризации и активации.
Фиг.14А: агонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.14В: антагонистическое влияние, рассчитанное как процент от максимальной стимуляции фосфорилирования c-Met посредством HGF [100 нг/мл].
Фиг.15: BRET-модели c-Met-димеризации и активации.
Фиг.16: микроскопический анализ влияния различных форм МКА 214 В2 на адгезию клеток РС3.
Фиг.17: анализ влияния различных форм МКА 214 В2 на адгезию клеток РСЗ с помощью АТФ-анализа. В каждой лунке определяли число прикрепленных клеток с помощью стандартной кривой РС3, от 0 до 200000 клеток/лунка. Результаты представлены следующим образом: необработанные клетки взяты в качестве контроля (100%), а обработанные клетки представлены в виде % от контроля.
Пример 1: Конструирование химерных МКА и функциональная оценка состояния фосфорилирования рецептора c-Met
Несколько мышиных МКА, нацеленных на прототипный тирозинкиназный рецептор (c-Met-рецептор), были переделаны как химерные МКА с мышиными вариабельными доменами и человеческими константными доменами. Их внутренние активности были проанализированы на основании функционального мониторинга ингибирования лиганд(HGF)-зависимого фосфорилирования c-Met-рецептора.
После ПЦР-клонирования последовательностей мышиных вариабельных доменов (VH, VL) химерные МКА были сконструированы путем лигирования фрагмента рестрикции {Nhe1-Bcl1}, несущего мышиную VH или VL последовательность, с вектором рСЕР4 (Invitrogen, США), несущим всю кодирующую последовательность константного домена либо человеческой легкой цепи Ckappa, либо человеческой тяжелой цепи [СН1-шарнир-СН2-СН3] из иммуноглобулина IgG1. Все этапы клонирования выполняли в соответствии с общепринятыми методиками молекулярной биологии, описанными в Laboratory manual (Sambrook and Russel, 2001), или в соответствии с инструкциями производителя. Каждую генетическую конструкцию полностью подтверждали путем секвенирования нуклеотидных последовательностей с помощью набора Big Dye terminator cycle sequencing kit (Applied Biosystems, US) и анализировали с помощью анализатора 3100 Genetic Analyzer (Applied Biosystems, US). Продукцию соответствующих химерных МКА проводили с помощью адаптированных к суспензии клеток НЕК293 EBNA (Invitrogen, США), выращенных в бессывороточной среде Excel 293 (SAFC Biosciences) с добавлением 6 мМ глутамина. Транзиентную трансфекцию выполняли с помощь линейного 25 кДа полиэтиленимина (PEI) (Polysciences). Процесс культивации контролировали на основании жизнеспособности клеток и продукции МКА. МКА очищали с помощью обычного хроматографического подхода на смоле с белком A (GE Healthcare, США).
Все различные формы МКА были получены на уровне, подходящем для функциональной оценки. Уровни продуктивности, как правило, находились в диапазоне от 15 до 30 мг/л очищенного МКА.
Функциональные оценки проводили на клетках человеческого рака легкого А549. Состояние фосфорилирования рецептора c-Met контролировали на клеточных лизатах путем специфического ловушечного ИФА. Козлиное анти-c-Met-МКА (R&D, исх. №AF276) использовали в качестве иммобилизованного (захватывающего) антитела, в то время как антитело для выявления соответствовало анти-фосфо-c-Met-MKA (Biosource, №KHO0281). Показатели люминесценции регистрировали на многоканальном планшетном спектрофотометре Mithras LB920 (Berthold).
Все три мышиных МКА 11Е1, 224G11 и 227Н1 показывали сопоставимую внутреннюю активность в отношении фосфорилирования рецептора c-Met: почти полное отсутствие их собственной агонистической активности (менее 5% от эффекта HGF, фиг.1А) и сильное ингибирование HGF[100 нг/мл]-индуцированного фосфорилирования рецептора c-Met (>70% ингибирование эффекта HGF, фиг.1В). Очень удивительно, что при изменении только константного домена МКА для переключения мышиного IgGI/kappa на человеческий IgG1 kappa наблюдалось полное изменение внутренней активности полученных химерных МКА (фиг.1А-1В). Действительно, наблюдался сильный агонизм (достигающий 20% от эффекта HGF для с11Е1, фиг.1А), связанный со значительным уменьшением эффективности антагониста (только оставшееся 60% ингибирование эффекта HGF для C224G11, фиг.1В). Этот эффект не зависит от вариабельного домена антитела, так как одно и то же явление наблюдается для трех исследованных МКА (моноклональные антитела 11Е1, 224G11 и 227Н1, секретируемые гибридомами, сохраненными в Национальной коллекции культур микроорганизмов (CNCM, National Collection of Microorganism Cultures) (Институт Пастера, Париж, Франция) 03/14/2007 под номерами CNCM 1-3724 (соответствует 11Е1), 1-3731 (соответствует 224G11) и I-3732 (соответствует 227Н1) (см. патентную заявку РСТ, опубликованную под номером WO 2009/007427).
Пример 2: Разработка, клонирование и продукция разработанных версий шарнирной области
На основе проведенных выше наблюдений можно предположить, что нарушение фармакологического профиля, наблюдаемое при переформатировании мышиного IgG1 в человеческие МКА IgG1, было вызвано человеческим IgG1 доменом.
С одной стороны, из литературы известно, что активация c-Met-рецептора была связана с его димеризацией и что ингибирование димеризации c-Met-рецептора может ингибировать фосфорилирование c-Met-рецептора и снизить передачу сигнала.
С другой стороны, МКА по своей сущности являются двухвалентными молекулами из-за присущей им структурной основы, и, таким образом, они могут выступать в качестве индукторов димеризации c-Met-рецептора.
Таким образом, можно предположить, что за счет ограничения конформационной гибкости химерных МКА, например поворотов, изгибов или виляний (см. Roux Roux et al., 1997), можно было бы восстановить внутреннюю активность, представляющую интерес (сильный антагонизм и слабый агонизм) родительских мышиных МКА. Эта гипотеза подкрепляется анализом соответствующих последовательностей мышиных и человеческих IgG1 шарнирных областей (также упоминаемых как IgG1 Н-области):
мышиная IgG1 Н-область PRDCGCKPCICT (SEQ ID №1)
человеческая IgG1 Н-область PKSCDKTHTCPPCP (SEQ ID №11)
человеческая IgG2 Н-область RKCCVECPPCP (SEQ ID №7)
Это выравнивание показывает, что мышиная IgG1 Н-область короче и содержит один дополнительный дисульфидный мостик (Cys) по сравнению с человеческой IgG1 Н-область. Оно также показывает, что человеческая IgG2 Н-область имеет сходство с мышиной IgG1 Н-областью как по длине (11 АА), так и по количеству дисульфидных мостиков (4).
В связи с этим предположили, что повышенная жесткость человеческой IgG1 Н-области может быть получена путем введения стабилизирующих мутаций, таких как Cys-остатки, и/или путем сокращения данного конкретного сегмента. Это предполагаемая повышенная жесткость Н-области может быть связана с улучшением функциональных свойств разработанного человеческого IgG1 МКА.
Первую серию из 7 разработанных версий получили (табл.2), создавая химерные Н-области путем замены N-концевой или С-концевой части шарнира между мышиной и человеческой последовательностями. Также было выполнено конструирование эквивалента IgG2.
Дополнительные серии из 28 мутантов в Н-области были разработаны и сконструированы для оценки влияния либо введения одного дополнительного остатка цистеина в Н-область, либо удаления по меньшей мере одной аминокислоты, либо одновременного введения одного цистеина и удаления по меньшей мере одной аминокислоты в Н-области.
Эти новые серии шарнирных мутантов описаны в таблице 3.
В качестве примера разработки шарнира был выбран вариабельный домен (тяжелая и легкая цепи) мышиного анти-c-Met-MKA, называемого 224G11.
Эти мышиные последовательности были слиты в первую очередь с человеческими константными доменами [Ckappa] для легкой цепи и [CH1-шарнир-СН2-СН3] для тяжелой цепи человеческого IgG1. Модификация шарнирной области была проведена путем замены рестрикционного фрагмента {Nhe1-Bcl1} на эквивалентную часть, несущую желаемые модификации, при этом каждый соответствующий {Nhe1-Bcl1} фрагмент был синтезирован путем глобального синтеза гена (Genecust, Люксембург). Все новые мутанты шарнира были сконструированы на той же основе.
Все этапы клонирования выполняли в соответствии с общепринятыми методиками молекулярной биологии, описанными в Laboratory manual (Sambrook and Russel, 2001), или в соответствии с инструкциями производителя. Каждую генетическую конструкцию полностью подтверждали путем секвенирования нуклеотидных последовательностей с помощью набора Big Dye terminator cycle sequencing kit (Applied Biosystems, US) и анализировали с помощью анализатора 3100 Genetic Analyzer (Applied Biosystems, США).
Адаптированные к суспензии клетки НЕК293 EBNA (Invitrogen, США) выращивали стандартным способом в 250 мл колбах в 50 мл бессывороточной среды Excel 293 (SAFC Biosciences) с добавлением 6 мМ глутамина на орбитальном шейкере (со скоростью вращения 110 об/мин). Транзиентную трансфекцию выполняли с использованием 2×106 клеток/мл, линейного 25 кДа полиэтиленимина (PEI) (Polysciences), приготовленного в воде с конечной концентрацией 1 мг/мл, и плазмидной ДНК (конечная концентрация 1,25 мкг/мл с соотношением плазмид с тяжелой и легкой цепью 1:1). Через 4 часа после трансфекции культуры разводили одним объемом свежей культуральной среды для достижения конечной плотности клеток 106 клеток/мл. Процесс культивации контролировали на основании жизнеспособности клеток и продукции МКА. Как правило, культуры сохраняли в течение 4-5 дней. МКА очищали с помощью обычного хроматографического подхода на смоле с белком A (GE Healthcare, США).
Все различные формы МКА были получены на уровне, подходящем для функциональной оценки. Уровни продуктивности, как правило, находились в диапазоне от 15 до 30 мг/л очищенных МКА.
Пример 3: Оценка разработанных МКА в фосфо-с-Met-специфическом анализе ИФА
Клетки А549 высевали в 12 многолуночных (MW) планшетах в полной ростовой среде [F12K+10% FCS (эмбриональной телячьей сыворотки)]. Клетки оставляли в течение 16 часов до стимуляции HGF [100 нг/мл], и каждое МКА для тестирования добавляли в конечной концентрации 30 мкг/мл за 15 минут до стимуляции лигандом. Через 15 минут после добавления HGF добавляли ледяной литический буфер, чтобы остановить реакцию фосфорилирования. Клетки механически разрушали и клеточные лизаты собирали путем центрифугирования при 13000 об/мин в течение 10 мин при 4°С и соответственно фазе супернатанта. Содержание белка количественно оценивали с помощью набора ВСА kit (Pierce) и хранили при температуре -20°С до момента использования. Состояние фосфорилирования c-Met количественно оценивали с помощью ИФА. Козлиное анти-c-Met-MKA (R&D, №AF276) использовали в качестве иммобилизованного (захватывающего) антитела (посадка в течение ночи при температуре 4°С), и после этапа насыщения с 5% буфером TBS-BSA (1 час при комнатной температуре) добавляли 25 мкг белковых лизатов в каждую лунку покрытого 96-луночного микротитровального планшета. Через 90 минут инкубации при комнатной температуре планшеты промывали четыре раза и добавляли антитело для выявления (анти-фосфо-c-Met-MKA, направленное против фосфорилированных остатков Tyr в позициях 1230, 1234 и 1235). После дополнительной инкубации в течение 1 часа и четырех промываний добавляли антикроличье антитело, связанное с HRP (Biosource) на 1 час при комнатной температуре, и выявляли люминесценцию путем добавления люминола. Люминесценцию регистрировали на многоканальном планшетном спектрофотометре Mithras LB920 (Berthold).
Конструировали серии разработанных версий шарнирного домена тяжелой цепи и анализировали в анализе фосфорилирования c-Met-рецептора. Как показано на фиг.2А, по сравнению с 224G11 [IgG1-Chim] наблюдалось значительное сокращение агонистического эффекта, связанного с изотипом hIgG1/kappa, как для конструкции на основе IgG2, так и для некоторых разработанных конструкций IgG1/kappa [MH, MUP9H и ТН7, фиг.2А]. Наиболее слабые и сопоставимые агонистические активности были получены для 224G11 [MH-IgG1], содержащего полную мышиную IgG1 шарнирную область, и для 224G11 [ТН7], содержащего наиболее человеческую разработанную IgG1 шарнирную область. Также было получено сопутствующее увеличение антагонистической эффективности [фиг.2 В]. Таким образом, и основанный на IgG2, и разработанный основанный на hIgG1/kappa ТН7 шарнирный мутант, связанный с мышиной 224G11 вариабельной областью, проявили функциональную активность, практически схожую с мышиным МКА 224G11. Тем не менее, сравнение агонистической/антагонистической активности 224G11 [MMCH-IgGI-Chim] и 224G11 [IgGI-Chim] показало, что увеличение антагонистической активности можно получить путем разработки антитела независимо от внутренних агонистических свойств такого антитела.
Вторую серию разработанных версий шарнирного домена тяжелой цепи конструировали и анализировали в анализе фосфорилирования c-Met-рецептора. Как показано на фиг.11А, аминокислотная замена в шарнирном домене тяжелой цепи, вводящая цистеиновый остаток, модифицирует агонистический эффект антител. Действительно, с одной стороны некоторые мутированные версии демонстрируют более слабый агонистический эффект, чем c224G11, например c224G11 [C2], c224G11 [С3], c224G11 [С5], c224G11 [6] или c224G11 [С7], в то время как другие демонстрируют увеличенный агонистический эффект, например c224G11 [C11], c224G11 [C12] и c224G11 [C14]. Более того, аминокислотные удаления в шарнирном домене тяжелой цепи, связанные или не связанные с аминокислотными заменами, также модифицируют агонистические свойства антител [фиг.11В]. Например, c224G11 [Δ1-3], c224G11 [Δ4-5-6], c224G11 [Δ5-6-7-8], c224G11 [С7Δ6], c224G11 [С6Δ9], c224G11 [С2Δ5-7], c224G11 [С5Δ2-6] или c224G11 [С9Δ2-7] демонстрируют более слабый агонистический эффект, чем c224G11, в то время как с224С11 [Δ8-11] демонстрирует более сильный агонистический эффект. Как и c224G11 [ТН7], все новые версии, демонстрирующие более слабый агонистический эффект, показывают сопутствующее увеличение антагонистической эффективности [фиг.12А и 12В], в то время как демонстрирующие более сильный агонистический эффект обладают более слабой антагонистической эффективностью.
В данной заявке применение квадратных скобок не является необходимым, и, в качестве примера, ссылку на [224G11] [IgG2chim] следует рассматривать как идентичную 224G11 IgG2chim. Таким же образом, чтобы показать, что антитело является мышиным, может быть добавлено выражение «murine» или буква m; чтобы показать, что антитело является химерным, может быть добавлено выражение «chim» или буква с; чтобы показать, что антитело является гуманизированным, может быть добавлено выражение «hum» или буква h. В качестве примера, химерное антитело 224G1IgG2 может рассматриваться как c224G11 IgG2, c224G11 [IgG2], с [224G11] IgG2, с [224G11] [IgG2], 224G11 IgG2chim, 224G11 [IgG2chim], [224G11] IgG2chim или [224G11] [IgG2chim].
Символ Δ означает удаление.
Пример 4: Анализ BRET
В первой серии экспериментов было проконтролировано, что нерелевантный мышиный IgG1, человеческий IgG1 и человеческий IgG2 не влияют на HGF-индуцированный BRET-сигнал в обеих моделях BRET (фиг.3). Эти МКА были использованы в дальнейшем в качестве контроля.
Затем было оценено влияние IgG1 химерных форм мышиного 224G11 МКА ([224G11]Chim), мышиного 11Е1 МКА ([11E1]Chim) и мышиного 227Н1 МКА ([227H1]Chim) на BRET модели c-Met-димеризации и c-Met-активации.
Хотя мышиное 224G11 МКА ингибирует 59% HGF-индуцированного BRET-сигнала на модели c-Met-димеризации, МКА [224G11]Chim ингибирует только 29% (фиг.4). [224G11]Chim антитело также было менее эффективным в ингибировании HGF-индуцированной c-Met-активации, так как антитела [224G11]Chim и m224G11 ингибировали 34,5% и 56,4% HGF-индуцированного BRET-сигнала (фиг.5). Более того, m224G11 в одиночку не влияло на c-Met-активацию, в то время как [224G11]Chim имел частичный агонистический эффект на c-Met-активацию, соответствующий 32,9% HGF-индуцированного сигнала. Этот частичный агонистический эффект [224G11]Chim также можно было видеть на BRET-модели c-Met-димеризации, так как [224G11]Chim в одиночку индуцировало увеличение BRET, соответствующее 46,6% HGF-индуцированного сигнала по сравнению с 21,3% для m224G11.
Агонистическую эффективность вторых серий разработанных версий шарнирного домена тяжелой цепи оценивали на BRET-модели c-Met активации (фиг.13А и 13В). В отличие от C224G11, который имеет частичное агонистическое влияние на c-Met-активацию, различные химерные формы антитела 224G11 с мутированным шарниром, включающие аминокислотную замену, аминокислотное удаление или и то, и другое, не показали значительного влияния на c-Met-активацию в одиночку для C224G11 [С2], C224G11 [С3], C224G11 [С5], C224G11 [С6], C224G11 [С7], c224G11 [Δ1-3], c224G11 [Δ4-5-6], c224G11 [Δ5-6-7-8], c224G11 [C7Δ6], C224G11 [С6Δ9], C224G11 [С2Δ5-7], c224G11 [C5Δ2-6] или c224G11 [C9Δ2-7] соответственно. Напротив, другие химерные формы с мутированным шарниром показали повышенное агонистическое влияние на c224G11 [Δ6], c224G11 [C11], c224G11 [C12] и c224G11 [C14].
Пример 5: Распознавание c-Met химерными и гуманизированными формами 224G11
Был проведен прямой ИФА, чтобы определить способность связывания различных химерных и гуманизированных форм по отношению к рекомбинантному c-Met. Кратко, рекомбинантный димерный c-Met от R&D Systems помещали в концентрации 1,25 мкг/мл в 96-луночные планшеты Immunol II. После инкубации в течение ночи при температуре 4°С лунки насыщали раствором 0,5% желатина/PBS. Планшеты инкубировали в течение 1 часа при температуре 37°С перед добавлением двукратных разведений тестируемых антител. Планшеты дополнительно инкубировали в течение часа перед добавлением HRP с козьим антимышиным IgG для выявления мышиного антитела и HRP с козьей античеловеческой легкой цепью каппа для распознавания химерного и гуманизированного антитела. Планшеты инкубировали в течение одного часа и добавляли пероксидазный субстрат ТМВ Uptima на 5 мин до нейтрализации с помощью 1М H2SO4. Результаты, представленные на фиг.6А и 6В, показывают, что все тестируемые формы были сопоставимы по распознаванию c-Met.
Пример 6: Влияние мышиных и химерных антител на HGF-индуцированную пролиферацию клеток NCI-H441 in vitro
Клетки NCl-H441 из АТСС стандартным образом культивировали в среде RPMI 1640 (Invitrogen Corporation, Шотландия, Великобритания), 10% FCS (Invitrogen Corporation), 1% L-глутамин (Invitrogen Corporation). Для анализа пролиферации клетки рассеивали за три дня до использования, чтобы они находились в конфлюэнтной фазе роста перед посевом. Клетки NCl-H441 помещали в 96-луночные платы для культивирования с плотностью 3,75×104 клеток/лунка в 200 мкл бессывороточной среды (RPMI 1640 плюс 1% L-глутамин). Через двадцать четыре часа после посева к NCl-H441 добавляли тестируемые антитела и инкубировали при 37°С в течение тридцати минут, прежде чем добавлять HGF в конечной концентрации 400 нг/мл (5 нМ) на 142 дополнительных часа. Диапазон тестируемых доз для каждого антитела составлял от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В этом эксперименте мышиное МКА IgG1 добавляли в качестве мышиного изотипического контроля, а протестированы были следующие антитела: m224G11, m11E1, m227H1 и их человеческие IgG1 химерные формы, соответственно идентифицированные как [224G11]Chim, [11E1]Chim и [227H1]Chim. Также были включены лунки, только засеянные клетками -/+HGF. Затем клетки подвергали импульсу 0,25 мкКи [3H]тимидина (Amersham Biosciences AB, Упсала, Швеция) в течение 7 часов и 30 минут. Значение [3H]тимидина, включенного в нерастворимую в трихлоруксусной кислоте ДНК, количественно оценивали с помощью жидкостного сцинтилляционного счетчика. Результаты представлены в виде нетрансформированных данных о cpm (импульсах в минуту) для лучшей оценки потенциальной внутренней агонистической активности, которая может возникнуть с анти-c-Met-MKA при добавлении их в одиночку к опухолевым клеткам.
Результаты, описанные в фиг.7А, 7В и 7С, показывают, что, как и ожидалось, мышиные антитела не проявляли агонистического эффекта при добавлении их в одиночку к раковым клеткам при любых испытываемых дозах. В этом эксперименте не наблюдалось никакого существенного ингибирования HGF-индуцированной пролиферации в изотипическом контроле касательно значительных расхождений cpm, наблюдаемых для этого изотипического контроля. При добавлении в одиночку ни мышиный m224G11, ни m11E1 или m227H1 не проявляли агонистического эффекта по сравнению с изотипическим контролем mIgG1 MKA или только клетками. Дозозависимая антипролиферативная активность достигала 78%, 80% или 80%, соответственно для MKA m224G11, m11E1 или m227H1 (расчет % ингибирования: 100-[(cpm клеток+тестируемое MKA - среднее фоновое cpm mIgG1)×100/(среднее срт клеток+HGF - среднее cpm только клеток)]). Удивительно, что химерные формы этих трех MKA индуцировали значительный дозозависимый агонистический эффект при добавлении в одиночку со стимуляцией роста, близкой к той, которая наблюдалась с HGF для [11E1]chim и [227H1]chim соответственно. Для этих двух антител, демонстрирующих особенно высокую внутреннюю агонистическую активность, антагонистический эффект был значительно снижен до ингибирующего эффекта 53% и 21% по сравнению с 80%, наблюдаемым для обеих мышиных форм. Агонистический эффект, наблюдаемый для химерного [224G11]chim, также был дозозависим, но он был ниже, чем наблюдаемый для [11E1]chim и [227H1]chim. Тем не менее, этот агонистический эффект оказал влияние на ингибирование HGF-индуцированной пролиферации in vitro, которая спустилась с 78% для мышиного m224G11 до 50% для его химерной формы. Для определения того, совместима ли такая «низкая» собственная агонистическая активность in vitro с неизменным эффектом in vivo, m224G11 и [224G11]chim были получены для тестирования in vivo. Так как в предыдущих исследованиях доза 30 мкг/мышь продемонстрировала значительную активность in vivo, то такая доза была выбрана для оценки in vivo.
Пример 7: Сравнение in vivo мышиного и химерного MKA 224G11 на модели с ксенотрансплантатом NCl-H441
NCl-H441 получены из папиллярной аденокарциномы легких, экспрессируют высокий уровень c-Met и демонстрируют конститутивное фосфорилирование c-Met RTK.
Для оценки in vivo эффекта антител на модели с ксенотрансплантатом NCl-H441 атимичных мышей в возрасте от шести до восьми недель помещали в стерилизованные клетки с верхним фильтром, оставляли в стерильных условиях и проводили на них манипуляции в соответствии с французскими и европейскими принципами. Мышам вводили подкожно 9×106 клеток. Затем, через шесть дней после имплантации клеток, опухоли измеряли (примерно 100 мм3), животных делили на группы по 6 мышей с сопоставимыми размерами опухоли и вводили сначала нагрузочную дозу 60 мкг антитела/мышь, а затем два раза в неделю по 1 мг/доза каждого тестируемого антитела. У мышей наблюдали темпы роста ксенотрансплантата. Объем опухоли рассчитывали по формуле: π (Pi)/6×длина×ширина×высота. Результаты, описанные на фиг.8, показывают, что мышиное МКА, лишенное агонистической активности, in vivo ведет себя, как и ожидалось, как мощный антагонист даже при низких испытываемых дозах. В противоположность тому, что наблюдается с мышиным МКА, химерное МКА демонстрирует весьма транзиентную активность in vivo, и опухоль полностью пропадает при лечении на 20 день после инъекции клеток. Этот эксперимент ясно показывает, что увеличение агонистического эффекта in vitro, приведшее к снижению антагонистической активности, также было ответственным за значительные потери антагонистической активности in vivo.
Пример 8: Влияние мышиного МКА 224G11 и различных химерных и гуманизированных версий этого антитела на HGF-индуцированную пролиферацию клеток NCl-H441 in vitro
Клетки NCl-H441 из АТСС стандартным образом культивировали в среде RPMI 1640 (Invitrogen Corporation, Шотландия, Великобритания), 10% FCS (Invitrogen Corporation), 1% L-глутамин (Invitrogen Corporation). Для анализа пролиферации клетки рассеивали за три дня до использования, чтобы они находились в конфлюэнтной фазе роста перед посевом. Клетки NCl-H441 помещали в 96-луночные платы для культивирования с плотностью 3,75×104 клеток/лунка в 200 мкл бессывороточной среды (RPMI 1640 плюс 1% L-глутамин). Через двадцать четыре часа после посева к NCl-H441 добавляли тестируемые антитела и инкубировали при 37°С в течение тридцати минут, прежде чем добавлять HGF в конечной концентрации 400 нг/мл (5 нМ) на 142 дополнительных часа. Диапазон тестируемых доз для каждого антитела составлял от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В этом эксперименте мышиное МКА IgG1 добавляли в качестве мышиного изотипического контроля и в качестве агонистического отрицательного контроля. Были протестированы следующие антитела: i) m224G11, ii) его человеческие IgG1 химерные формы, соответственно идентифицированные как [224G11]Chim, [224G11] [МН Chim], [224G11] [MUP9H Chim], [224G11] [MMCH Chim], [224G11] [TH7 Chim], iii) его гуманизированные IgG1 формы, соответственно описанные как [224G11] [Hz1], [224G11] [Hz2], [224G11] [Hz3]. Также были включены лунки, только засеянные клетками -/+HGF. Целое антитело 5D5 из Genentech, коммерчески доступное из АТСС в виде гибридомной клеточной линии, было представлено как полный агонистический положительный контроль и затем названо как m5D5. Затем клетки подвергали импульсу 0,25 мкКи [3H]тимидина (Amersham Biosciences AB, Упсала, Швеция) в течение 7 часов и 30 минут. Значение [3H]тимидина, включенного в нерастворимую в трихлоруксусной кислоте ДНК, количественно оценивали с помощью жидкостного сцинтилляционного счетчика. Результаты представлены в виде нетрансформированных данных о cpm (импульсах в минуту) для лучшей оценки потенциальной внутренней агонистической активности, которая может возникнуть с анти-c-Met-MKA при добавлении их в одиночку к опухолевым клеткам.
Результаты, описанные в фиг.9А, показывают, что, как и ожидалось, ни изотипический контроль, ни m224G11 не проявляли агонистической активности по отношению к пролиферации NCl-H441. Изотипический контроль не влиял на HGF-индуцированную пролиферацию клеток, тогда как m224G11 демонстрировал 66%-ное ингибирование при добавлении в конечной концентрации 10 мкг/мл. m5D5, использованный в качестве агонистического контроля, показал, как и ожидалось, полный дозозависимый агонистический эффект при добавлении к клеткам в одиночку. Как уже отмечалось, МКА [224G11]chim демонстрировало значительный дозозависимый агонистический эффект, и наблюдалась сниженная ингибиторная активность этой химерной формы: 19% вместо 66% для мышиной формы. При добавлении в одиночку, три IgG1 гуманизированных МКА продемонстрировали дозозависимый агонистический эффект по сравнению с формой m224G11. [224G11] [Hz1], [224G11] [Hz2] и [224G11] [Hz3] имели сравнимую антагонистическую активность, примерно 46, 30 и 35%. Эти активности были значительно ниже, чем та, которая наблюдалась для m224G11. На фиг.9В были проверены различные IgG1 химерные формы. По сравнению с формой [224G11]chim, которая демонстрирует дозозависимый агонистический эффект при добавлении в одиночку к клеткам NCl-H441, формы [224G11] [МН chim], [224G11] [MUP9H chim], [224G11] [MMCH chim], [224G11] [TH7 chim] не проявляли значительного внутреннего агонистического эффекта. Их антагонистическая активность была выше, чем та, которая наблюдалась у МКА m224G11 (57%) с достижением ингибирования 79, 78, 84 и 93% соответственно [224G11] [MH Chim], [224G11] [MUP9H Chim], [224G11] [MMCH Chim] и [224G11] |TH7 Chim].
Пример 9: Влияние in vitro различных IgG1 химерных и гуманизированных форм МКА 224G11
Клетки NCl-H441 из АТСС стандартным образом культивировали в среде RPMI 1640 (Invitrogen Corporation, Шотландия, Великобритания), 10% FCS (Invitrogen Corporation), 1% L-глутамин (Invitrogen Corporation). Для анализа пролиферации клетки рассеивали за три дня до использования, чтобы они находились в конфлюэнтной фазе роста перед посевом. Клетки NCl-H441 помещали в 96-луночные платы для культивирования с плотностью 3,75×104 клеток/лунка в 200 мкл бессывороточной среды (RPMI 1640 плюс 1% L-глутамин). Через двадцать четыре часа после посева к NCI-H441 добавляли тестируемые антитела и инкубировали при 37°С в течение тридцати минут, прежде чем добавлять HGF в конечной концентрации 400 нг/мл (5 нМ) на 142 дополнительных часа. Диапазон тестируемых доз для каждого антитела составлял от 10 до 0,0097 мкг/мл (конечная концентрация в каждой лунке). В этом эксперименте мышиное IgG1 МКА добавляли в качестве фонового отрицательного контроля агонистической активности, а протестированы были следующие антитела: i) m224G11, ii) его человеческие IgG1 химерные формы, соответственно идентифицированные как [224G11]Chim, [224G11] [ТН7 Chim], iii) его гуманизированные IgG1 формы, соответственно описанные как [224G11] [ТН7 Hz1], [224G11] [ТН7 Hz3]. Также были включены лунки, только засеянные клетками -/+HGF. Целое антитело 5D5 из Genentech, коммерчески доступное из АТСС в виде гибридомной клеточной линии, было представлено как полный агонистический положительный контроль и затем названо как m5D5. Затем клетки подвергали импульсу 0,25 мкКи [3H]тимидина (Amersham Biosciences AB, Упсала, Швеция) в течение 7 часов и 30 минут. Значение [3H]тимидина, включенного в нерастворимую в трихлоруксусной кислоте ДНК, количественно оценивали с помощью жидкостного сцинтилляционного счетчика. Результаты представлены в виде нетрансформированных данных о cpm (импульсах в минуту) для лучшей оценки потенциальной внутренней агонистической активности, которая может возникнуть с анти-c-Met-MKA при добавлении их в одиночку к опухолевым клеткам.
На фиг.10 показано, что МКА m224G11 демонстрирует обычный ингибиторный эффект (74% ингибирования). Химерная IgG1 форма [224G11]chim имела, как и ожидалось, дозозависимый внутренний агонистический эффект и сниженный антагонистический эффект по сравнению с мышиной формой: 33% против 74% ингибирования. [224G11] [ТН7 chim] имело слабую агонистическую активность в этом эксперименте. Кроме того, оно демонстрировало высокий ингибирующий эффект (81%), близкий к эффекту, отмеченному для мышиного МКА. Две гуманизированные формы не проявляли внутреннего агонистического эффекта и имели антагонистическую активность, близкую к наблюдаемой для мышиного МКА или [224G11] [ТН7 chim] с ингибированием 67 и 76% соответственно для [224G11] [ТН7 Hz1] и [224G11] [ТН7 Hz3].
Пример 10: Переключение изотипа для разработанной [ТН7] шарнирной области
Для того чтобы оценить модуляцию фармакологических свойств, индуцированную [ТН7] шарнирной последовательностью, соответствующей PKSCDCHCPPCP, в остовах иммуноглобулиновых изотипов кроме человеческого IgG1, мы генетически перенесли описанную выше последовательность [ТН7] в человеческие остовы IgG2 и IgG4. Модификацию шарнирной области проводили путем замены рестрикционного фрагмента {Nhe1I-Bcl1} на эквивалентную часть, несущую модификацию [ТН7], при этом фрагмент {Nhe1-Bcl1} синтезировали путем глобального синтеза гена (Genecust, LU). Этапы клонирования выполняли в соответствии с общепринятыми методиками молекулярной биологии, описанными в Laboratory manual (Sambrook and Russel, 2001), или в соответствии с инструкциями производителя. Каждую генетическую конструкцию полностью подтверждали путем секвенирования нуклеотидных последовательностей с помощью набора Big Dye terminator cycle sequencing kit (Applied Biosystems, US) и анализировали с помощью анализатора 3100 Genetic Analyzer (Applied Biosystems, US). Полученные последовательности были описаны как SEQ ID №78, 79, 80 и 81 для соответствующих аминокислотных и нуклеотидных последовательностей ТН7-разработанных человеческих изотипов IgG2 и IgG4 (только тяжелая цепь, легкая цепь была идентична c224G11/human Ckappa, используемой для всех других конструкций на основе IgG1). Эти новые конструкции были применены к химерному 224G11 анти-c-Met-MKA, описанному выше в примере 2.
Соответствующие разработанные антитела, c224G11 [IgG2TH7] и C224G11 [IgG4TH7], были получены, как описано выше, путем транзиентной экспрессии в адаптированных к суспензии клетках НЕК293 EBNA.
Пример 11: Оценка разработанных МКА c224G11 [IgG2TH7] и c224G11 [IgG4TH7] в фосфо-с-Met-специфическом анализе ИФА и BRET-анализе
ТН7-шарнир также был введен в IgG2 и IgG4 химерные МКА 224G11 и испытан в анализе фосфорилирования c-Met-рецептора. Как показано на фиг.14А и 14В, c224G11 [IgG2TH7] и c224G11 [IgG4TH7] индуцировали слабый агонистический эффект в одиночку, но значительно более слабый, чем МКА c224G11, и демонстрировали антагонистический эффект, сопоставимый с мышиной формой МКА 224G11 (m224G11). Этот результат был подтвержден на BRET-модели c-Met-активации (фиг.15), где c224G11 [IgG2TH7] и c224G11 [IgG4TH7] также показали себя как более слабые агонисты, чем МКА c224G11.
Таким образом, шарнирная мутация ТН7, введенная в МКА формата IgG2 или IgG4, дала функциональные антитела со свойствами, аналогичными c224G11 [ТН7].
Пример 12: Анализ адгезии клеток
Клетки рака предстательной железы РСЗ отделяли от чашек с помощью трипсина, промывали три раза бессывороточной средой F12k и ресуспендировали в этой же среде. Клетки (100000 клеток/лунка) высевали на 96-луночные планшеты, покрытые ламинином 1 в концентрации 1 мкг/мл. Следующие формы анти-CD151 МКА для тестирования были добавлены одновременно в конечной концентрации 10 мкг/мл: мышиное IgG1 МКА т214 В2, немодифицированная химерная форма IgG1 антитела, называемая с214 В2, и химерная форма антитела IgG1 с ТН7 модификацией, называемая СТН7-214 В2.
CD151 представляет собой мембранный белок, принадлежащий семейству тетраспанинов, и анти-С0151 МКА 214 В2, продуцируемое гибридомой 1-3919, зарегистрированной в CNCM 21 февраля 2008 года, и описан в опубликованной патентной заявке WO 2009/136070.
Мышиные и человеческие IgG1 антитела использовали в качестве изотипических контрольных антител. Окончательные условия были следующими:
100000 клеток/лунка и антитела в концентрации 10 мкг/мл. Через один час инкубации при 37°С планшеты стряхивали и промывали дважды бессывороточной средой F12k. Перед анализом 100 мкл бессывороточной среды F12k вводили в каждую лунку. Для оценки влияния антител на клеточную адгезию лунки фотографировали под фазово-контрастным микроскопом (фиг.16). Затем число прикрепленных клеток определяли с помощью анализа АТФ (фиг.17).
Мышиные 214В2 и химерные ТН7-214В2 антитела были способны модифицировать межклеточные взаимодействия (фиг.16) и увеличивать эквивалентно клеточную адгезию РСЗ (фиг.17), в то время как не наблюдалось эффекта с модифицированной химерной формой 214В2 (с214В2), которая была сопоставима с человеческим IgG1 контрольным изотипическим антителом.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ с-МЕТ | 2011 |
|
RU2607377C2 |
| НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2552161C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| АНТИТЕЛО ПРОТИВ cMet И ЕГО ПРИМЕНЕНИЕ ДЛЯ ОПРЕДЕЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2010 |
|
RU2582265C2 |
| КОМБИНАЦИЯ АНТАГОНИСТА С-МЕТ И АМИНОГЕТЕРОАРИЛА ДЛЯ ЛЕЧЕНИЯ РАКА | 2009 |
|
RU2526171C2 |
| АНТИ-МЕТ АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2765267C2 |
| АНТИТЕЛА К CXCR4 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ РАКА | 2009 |
|
RU2573897C2 |
| АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО VISTA И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2746994C2 |
| АНТИТЕЛО ПРОТИВ IGF-1R И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ АДРЕСУЮЩЕГО ПЕРЕНОСЧИКА ДЛЯ ЛЕЧЕНИЯ РАКА | 2015 |
|
RU2698977C2 |



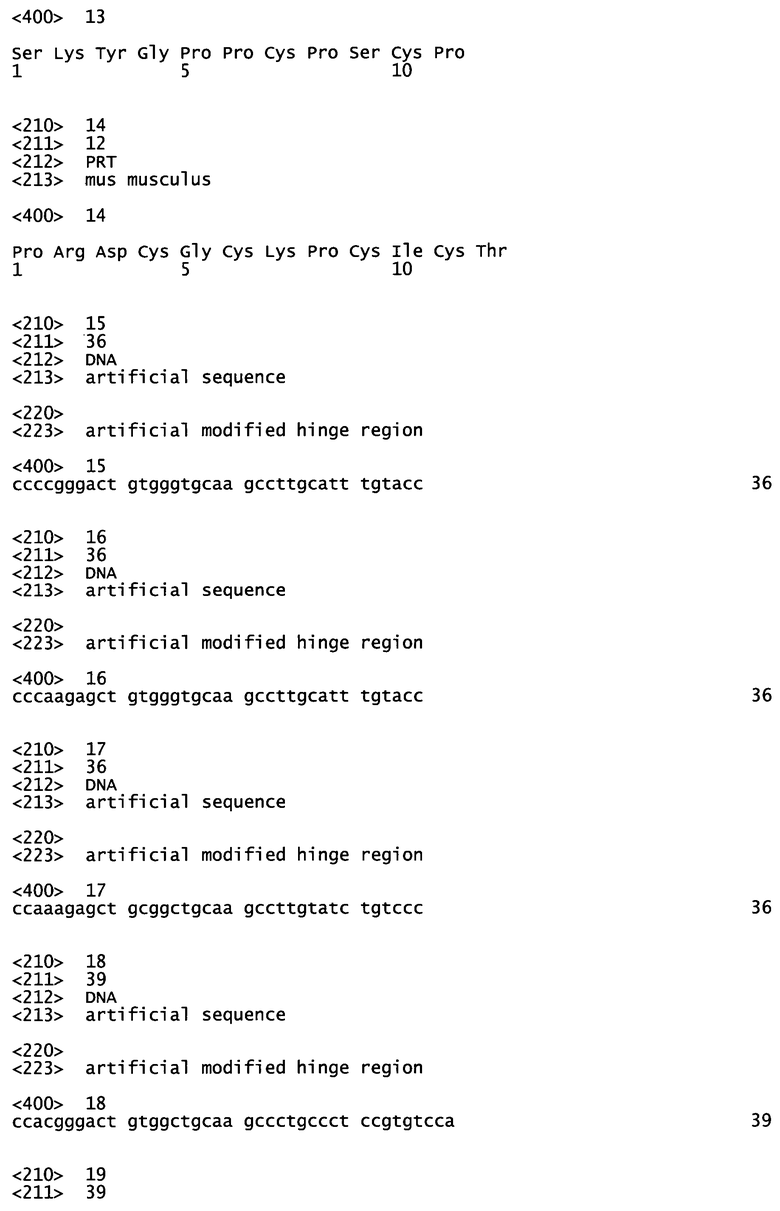

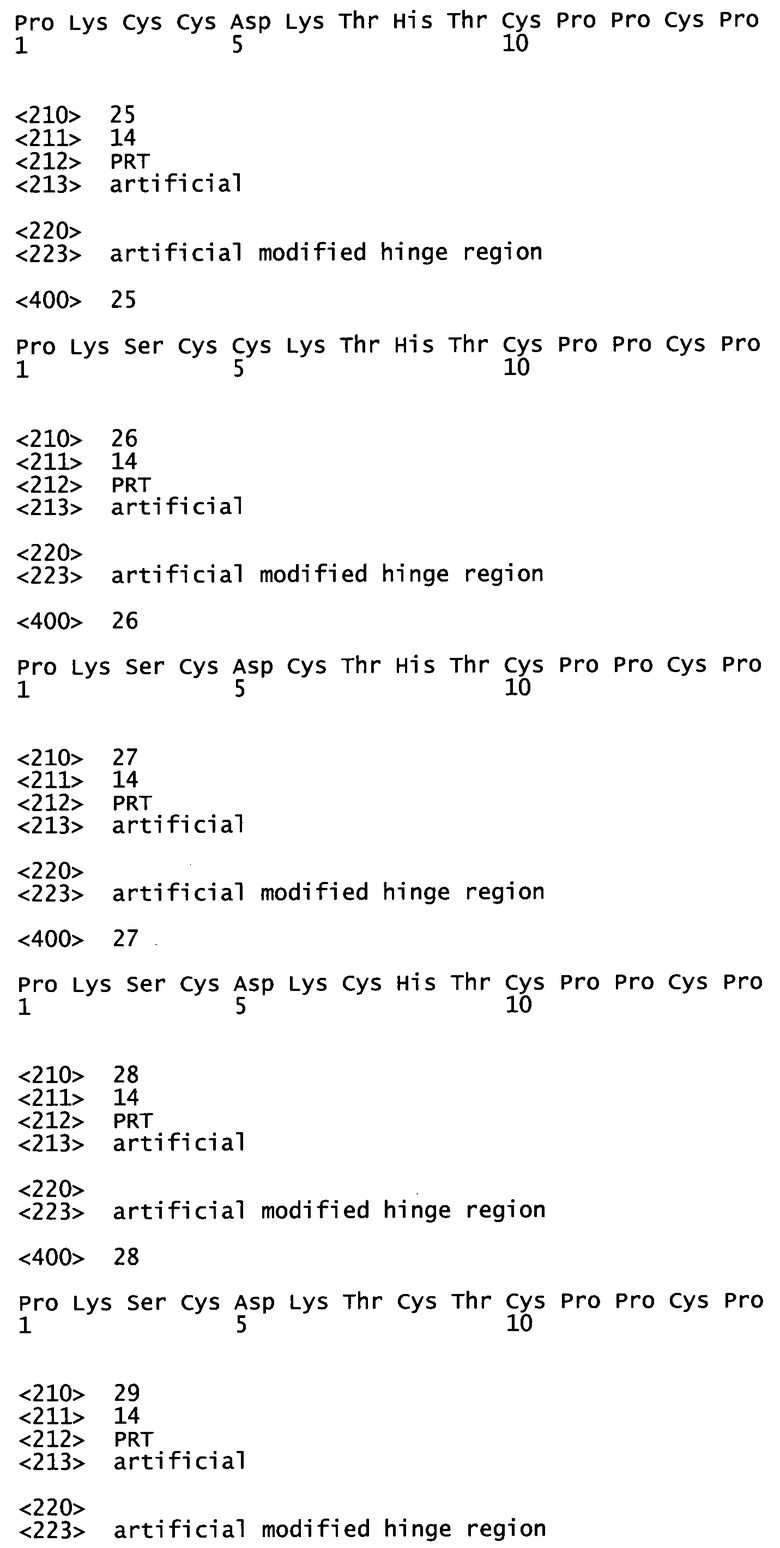

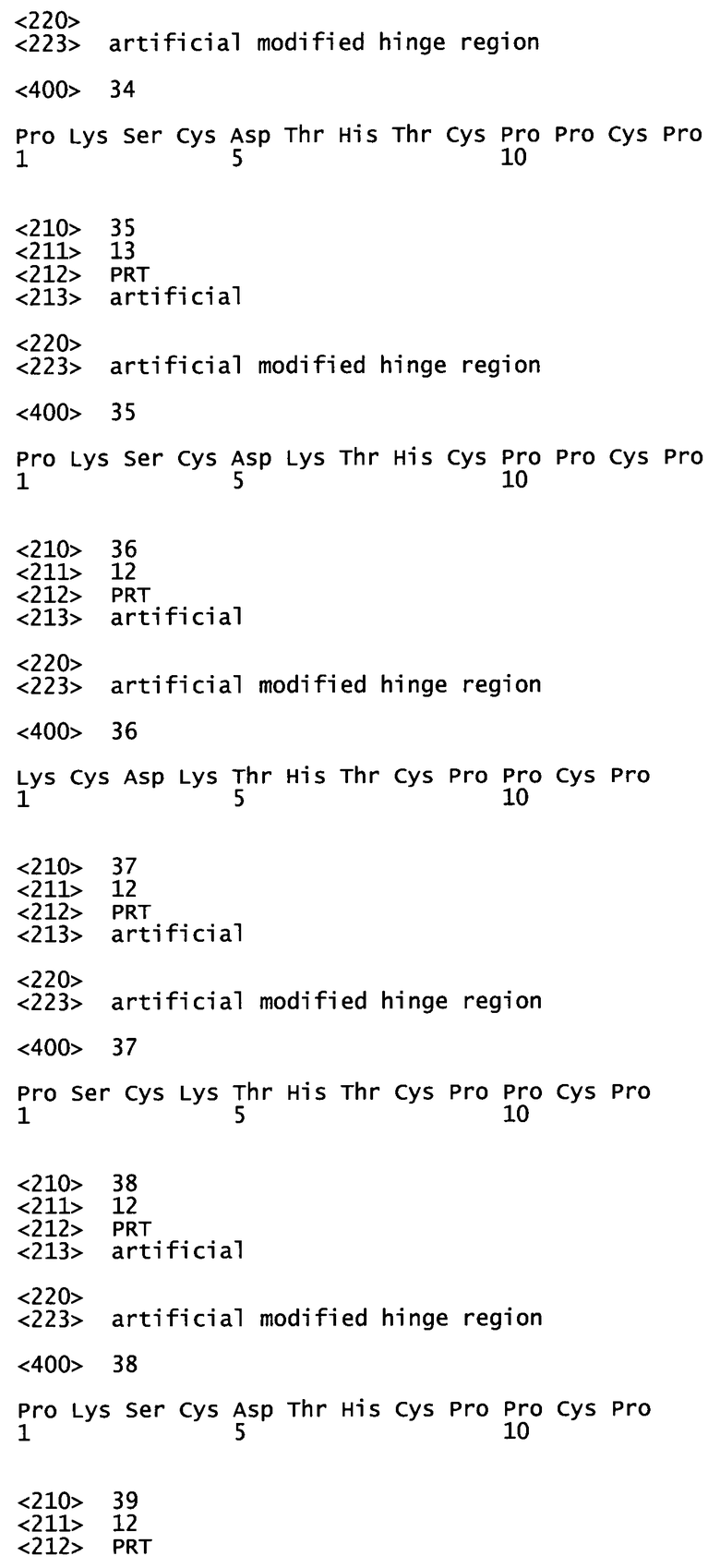

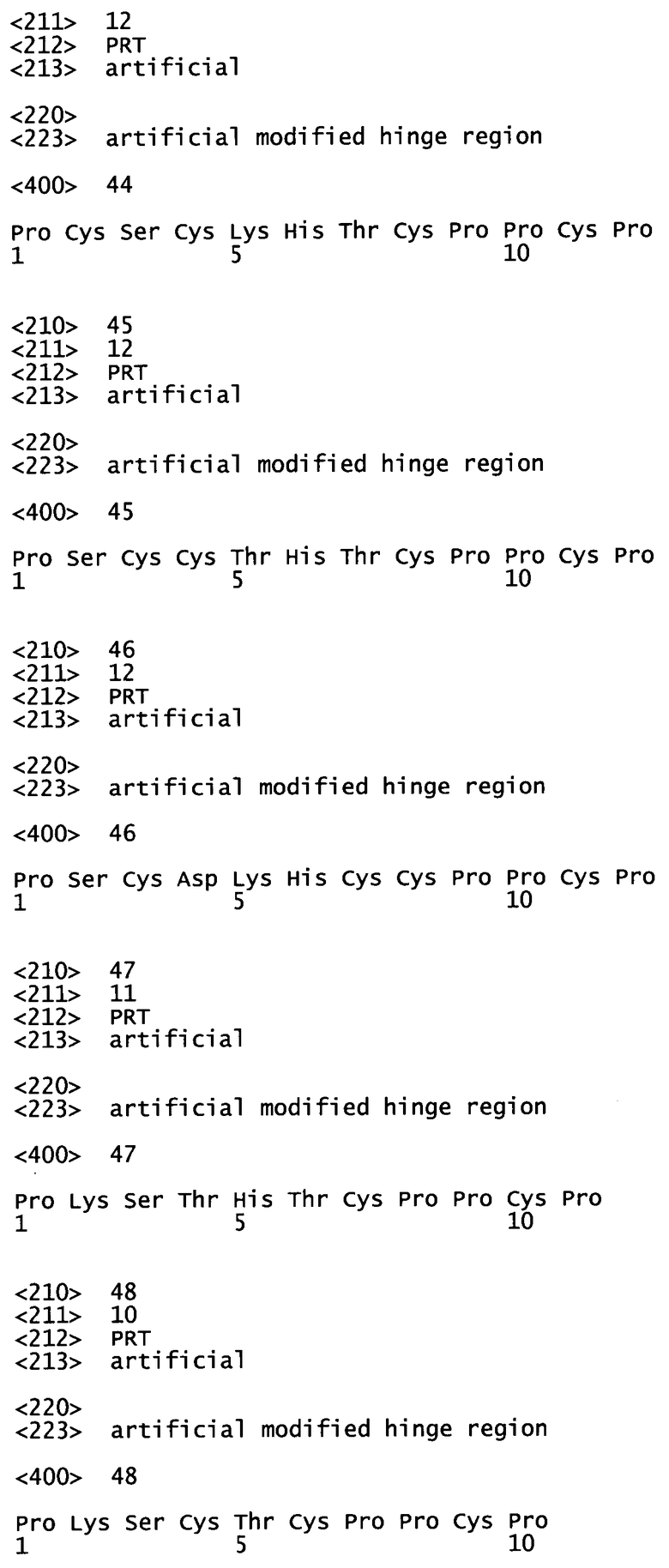

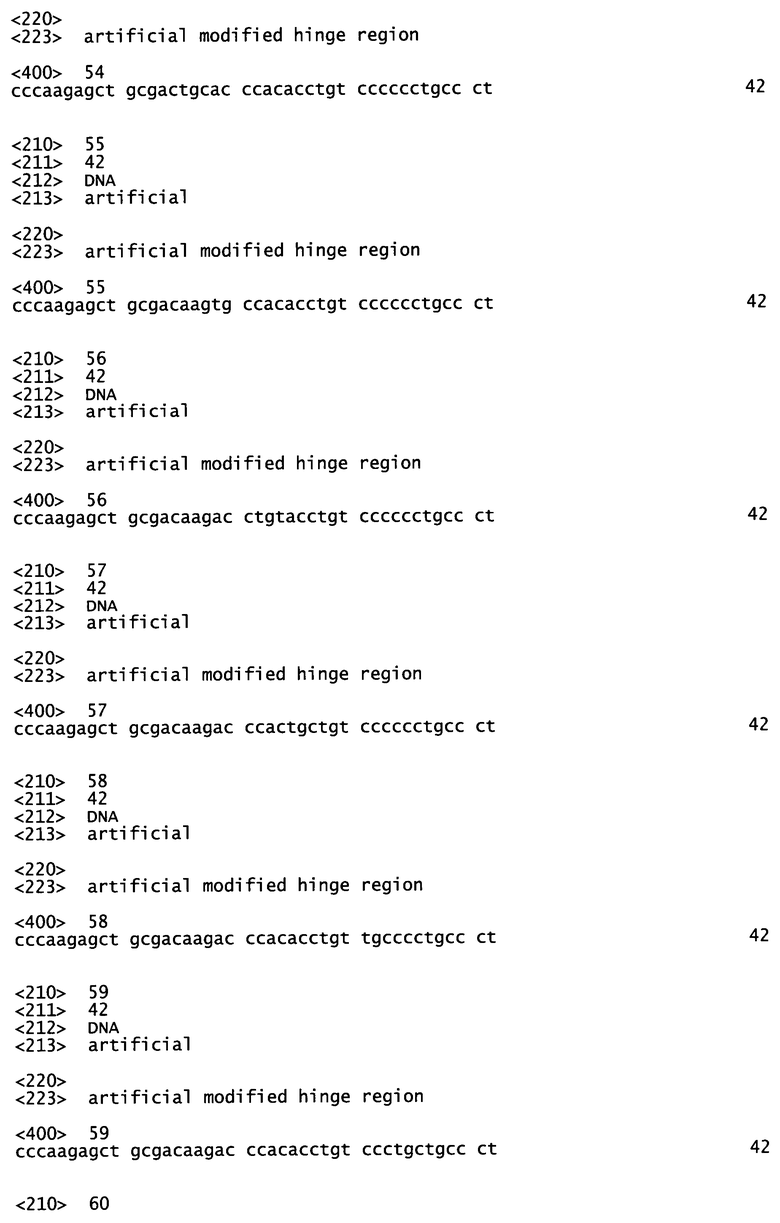









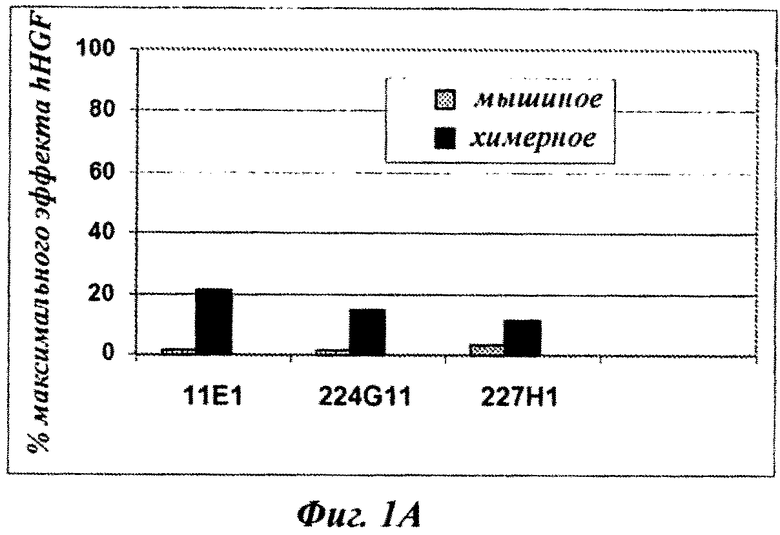
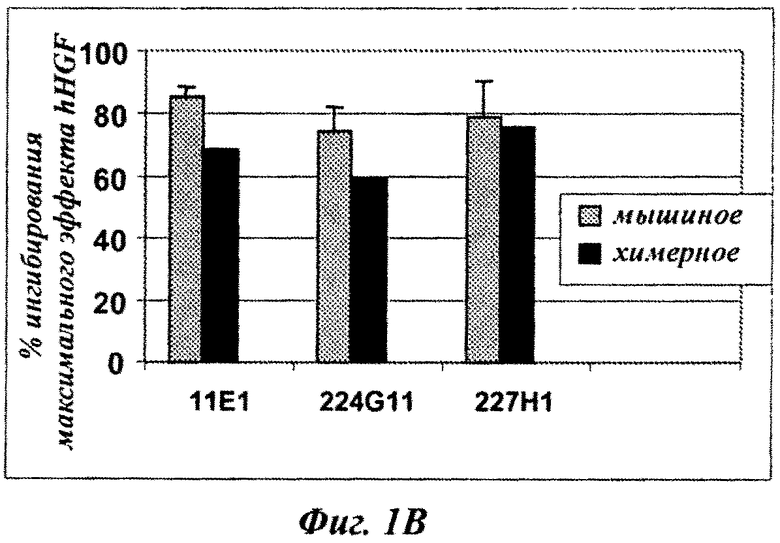

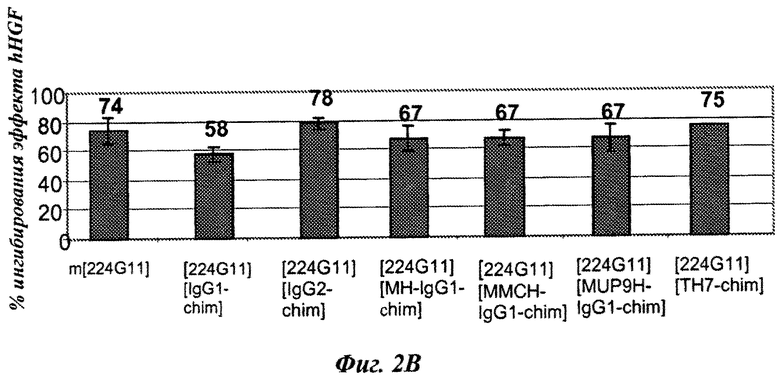

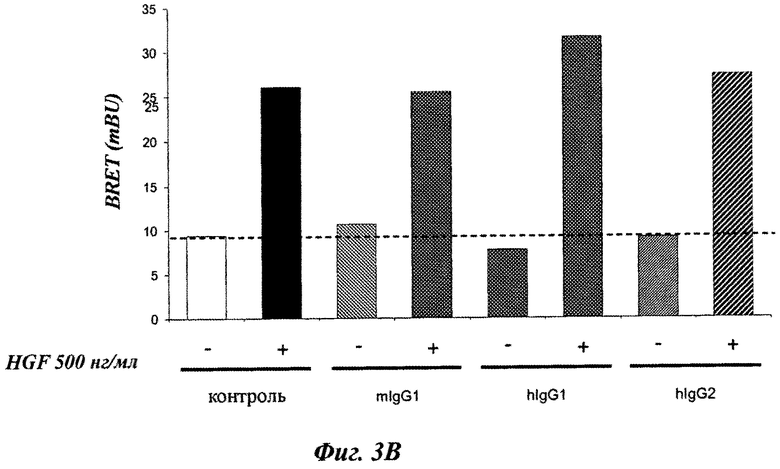
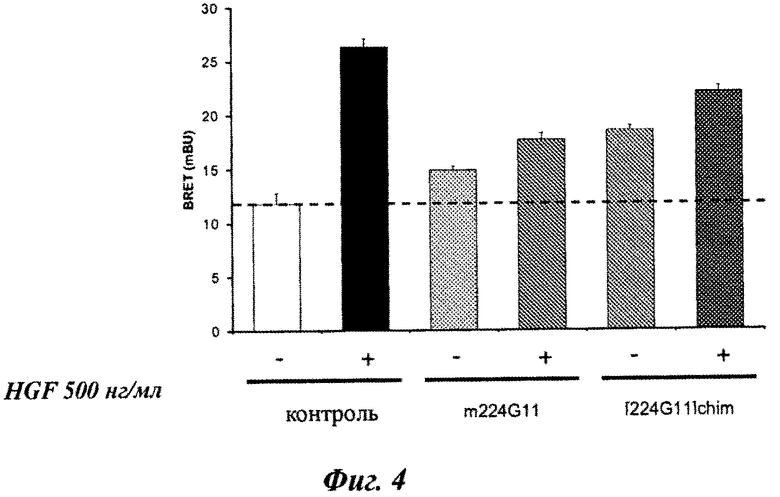

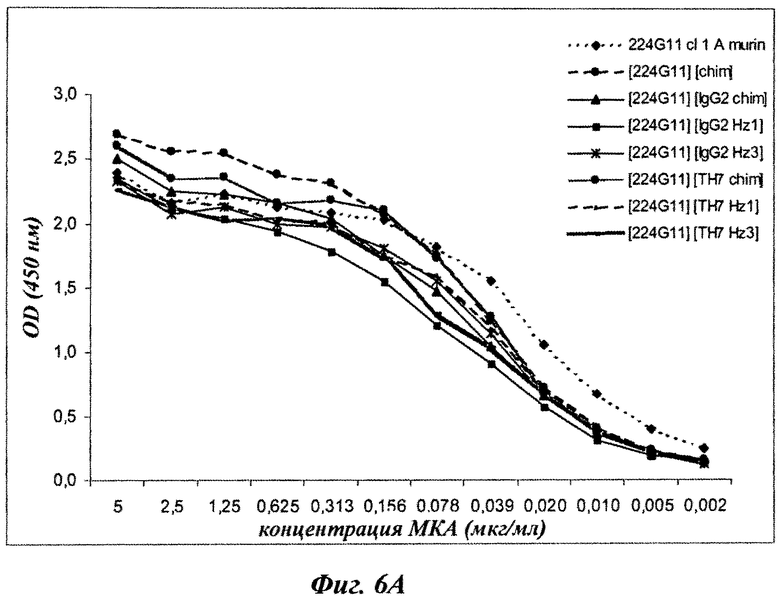


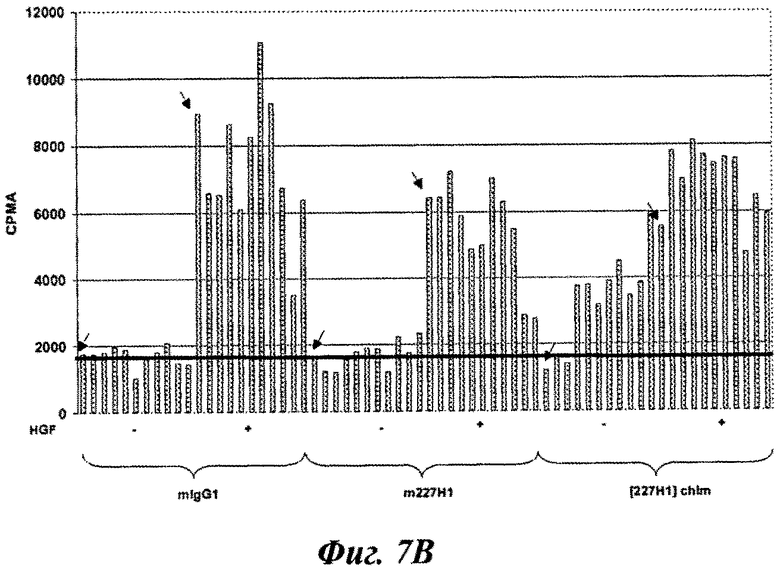

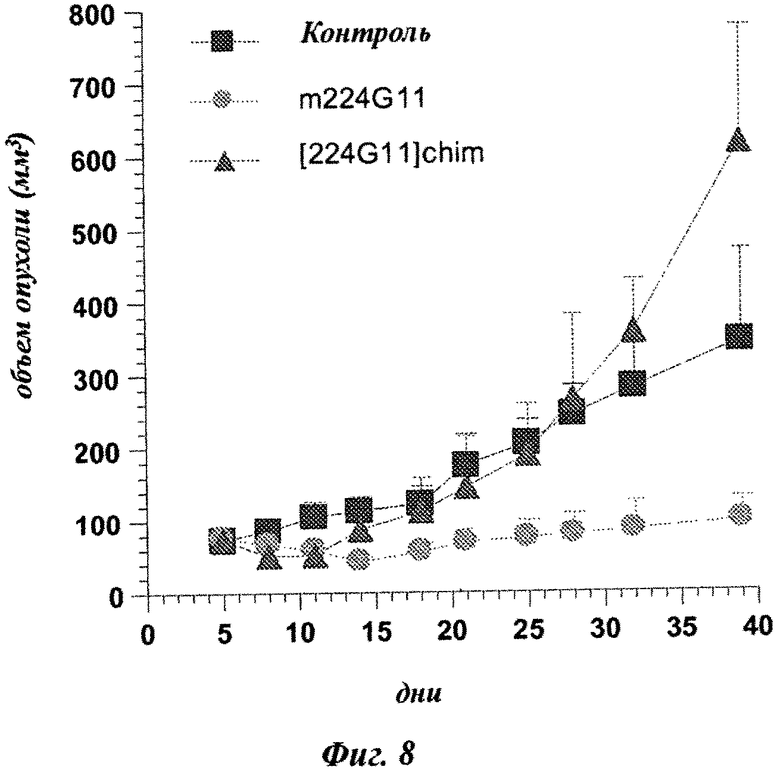


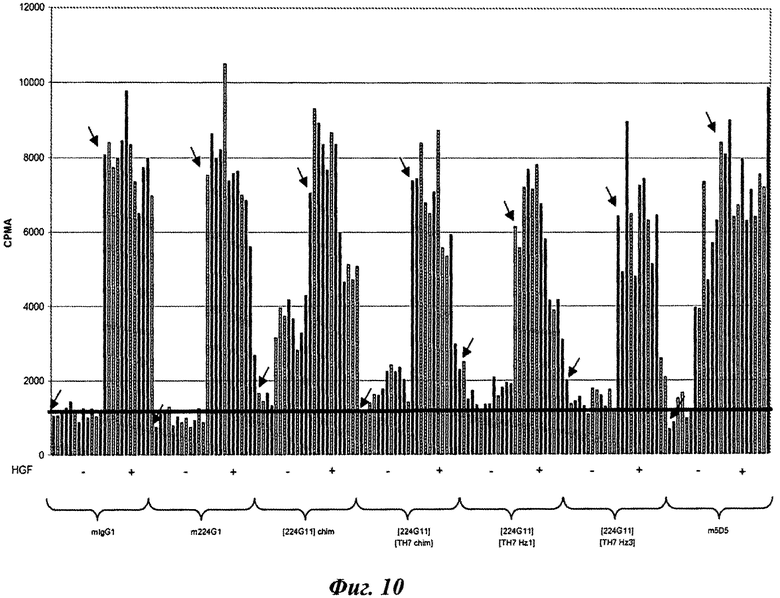
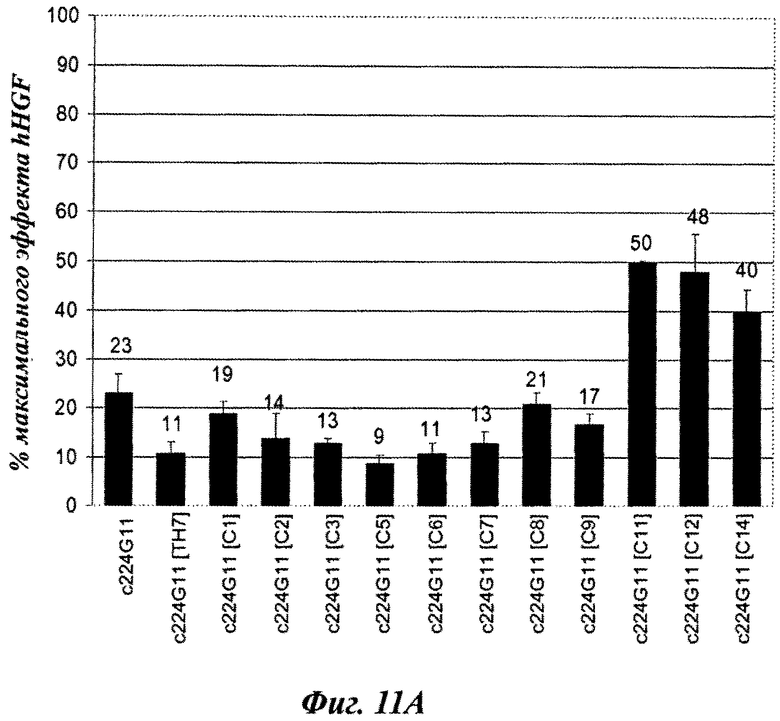
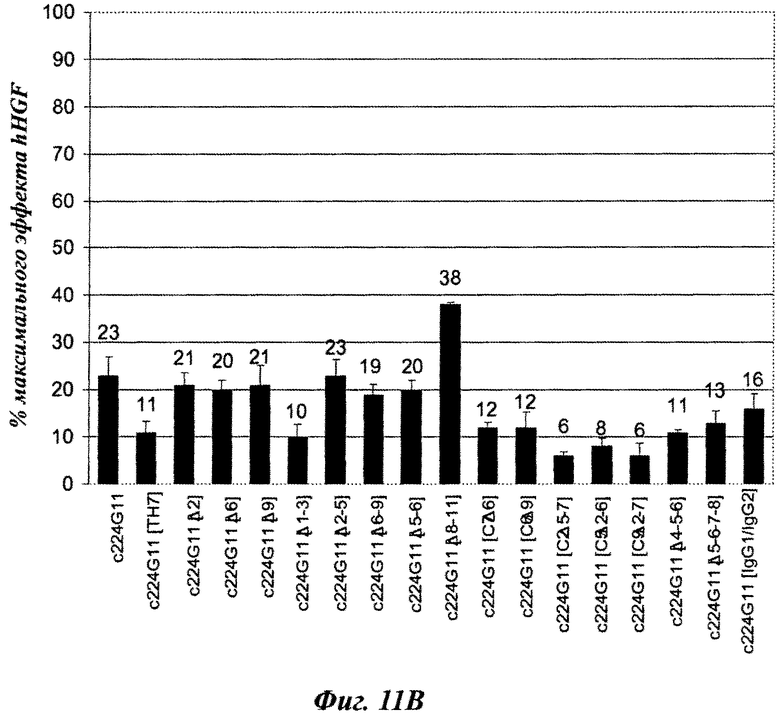
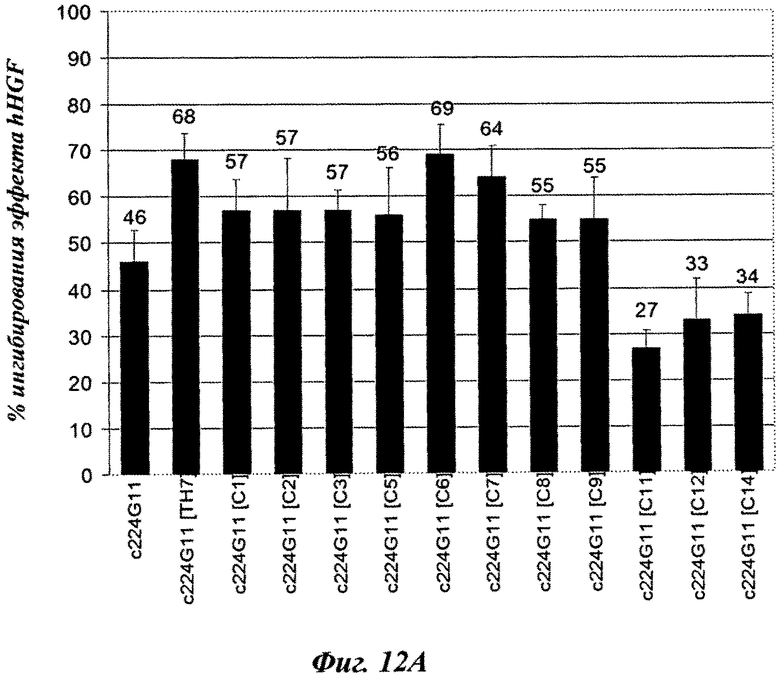
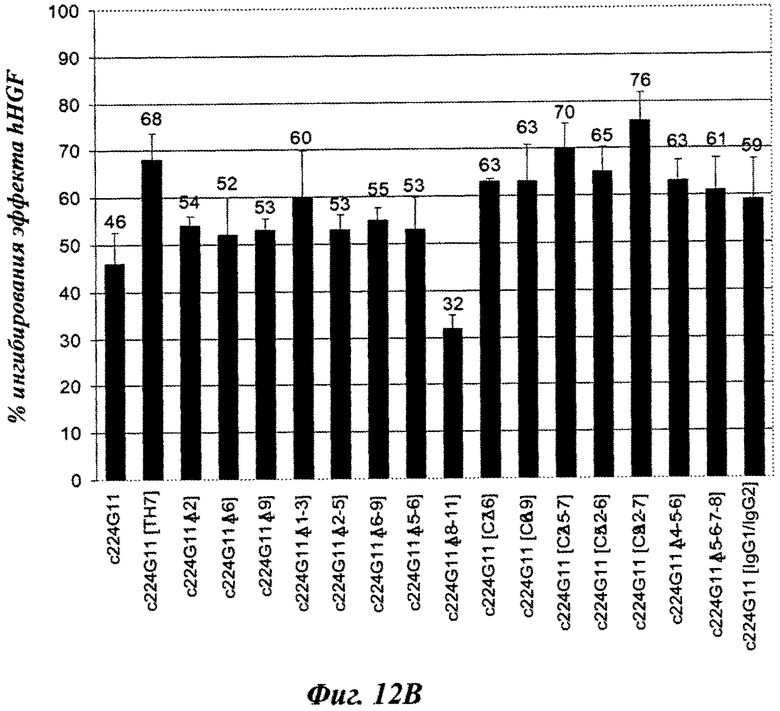

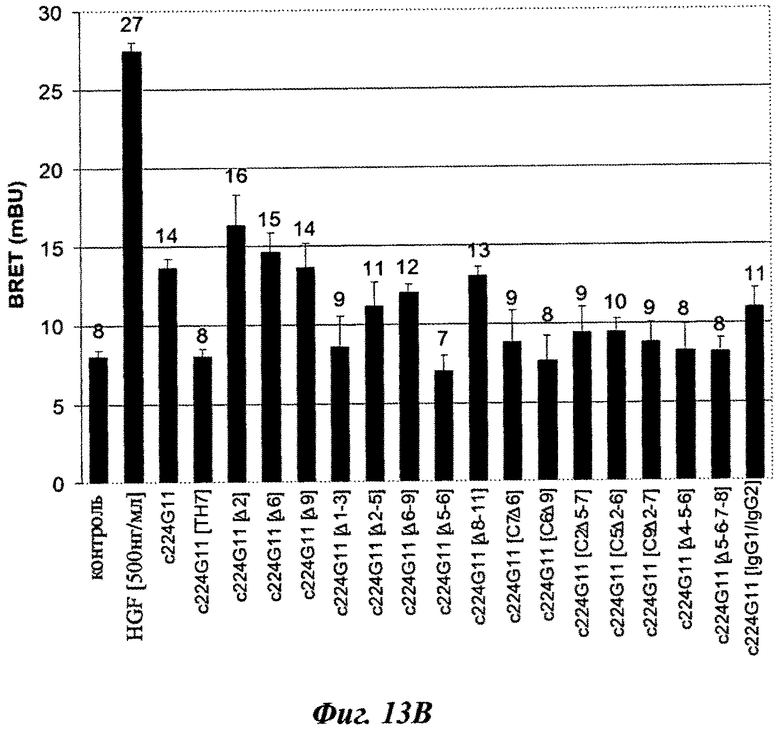
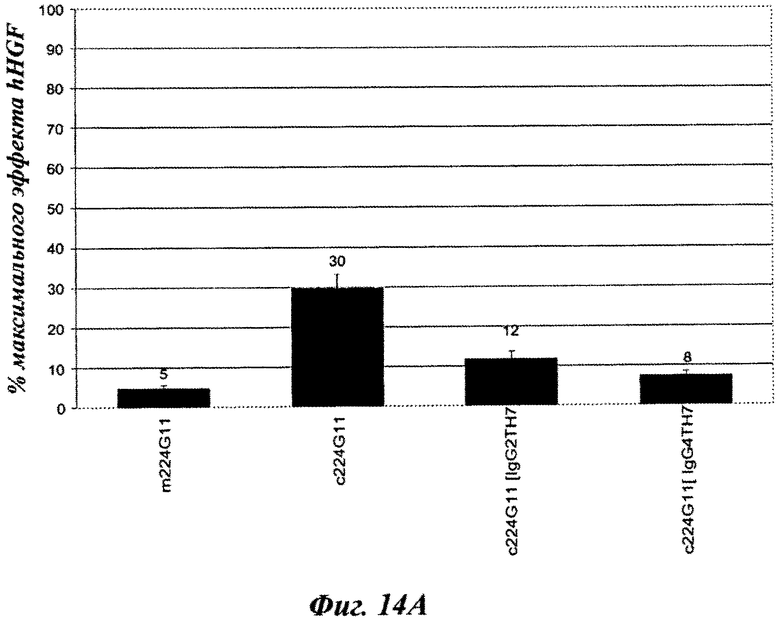
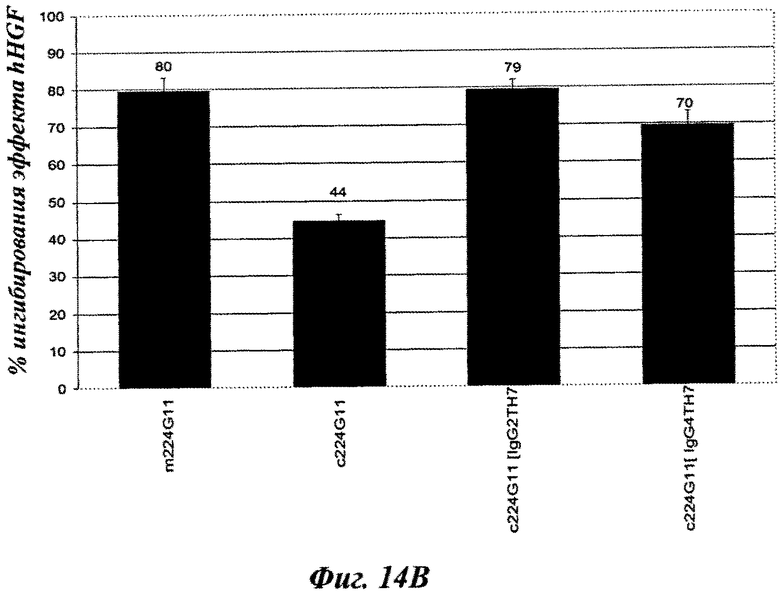


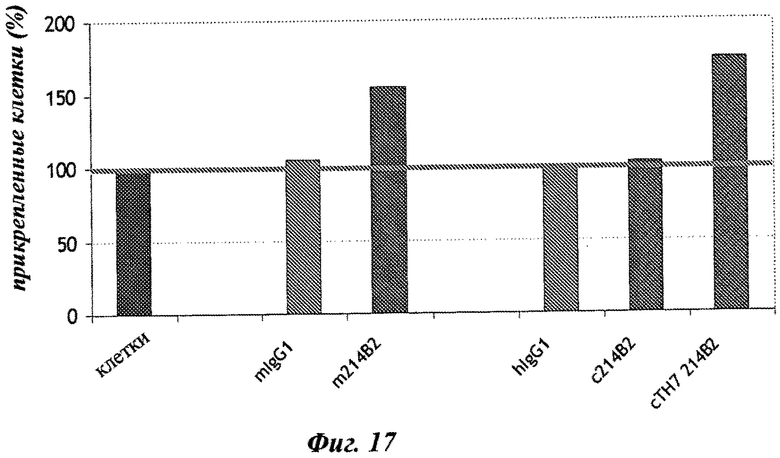
Настоящее изобретение относится к области биотехнологии. Предложен способ повышения антагонистической активности моноклонального антитела, содержащего константную область IgG1 человека и направленного против специфической молекулы-мишени, включающий модификацию аминокислотной последовательности шарнирной области. Кроме того, рассмотрен способ скрининга антагонистического моноклонального антитела, основанный на использовании способа повышения антагонистической активности по изобретению, моноклональное антитело, полученное способом по изобретению, и кодирующая его нуклеиновая кислота. Данное изобретение позволяет модулировать антигенсвязывающую активность путем изменения только шарнирного участка константной области антитела. 4 н. и 9 з.п. ф-лы, 17 ил., 3 табл., 12 пр.
1. Способ повышения антагонистической активности моноклонального антитела, содержащего константную область IgG1 человека и направленного против специфической молекулы-мишени, при этом указанное антитело способно ингибировать одну или более биологическую активность указанной молекулы-мишени, где указанный способ включает этап реконфигурации шарнирной области, заключающийся в модификации аминокислотной последовательности указанной шарнирной области путем удаления, добавления или замены по меньшей мере одной аминокислоты, и где указанная реконфигурированная шарнирная область, полученная модификацией ее аминокислотной последовательности, обладает последовательностью, выбранной из группы, состоящей из последовательностей SEQ ID № 2 - SEQ ID № 6, SEQ ID № 22 - SEQ ID № 26, SEQ ID № 28, SEQ ID № 29, SEQ ID № 33 - SEQ ID № 39 и SEQ ID № 42 - SEQ ID № 49.
2. Способ по п.1, где указанное моноклональное антитело является двухвалентным антителом.
3. Способ по п.1, где указанное моноклональное антитело является химерным антителом.
4. Способ по п.1, где указанное моноклональное антитело является гуманизированным антителом.
5. Способ по п.1, где указанное моноклональное антитело является человеческим антителом.
6. Способ по п.1, где указанное моноклональное антитело представляет собой IgG1.
7. Способ по п.1, где указанная молекула-мишень является трансмембранным рецептором.
8. Способ по п.7, где указанный трансмембранный рецептор выбран из группы, состоящей из тирозинкиназных рецепторов, тетраспанина и GPCR (рецепторов, связанных с G белком).
9. Способ скрининга антагонистического моноклонального антитела, направленного против специфической молекулы-мишени, при этом указанное антитело способно ингибировать одну или более биологическую активность указанной молекулы-мишени, где указанный способ включает этапы:
(а) выбора исходного антитела с исходным уровнем ингибирования указанной одной или более биологической активности указанной молекулы-мишени,
(б) изменения аминокислотной последовательности шарнирной области указанного исходного антитела с помощью способа по п.1,
(в) оценки модифицированного антитела этапа (б) в отношении его способности ингибировать указанную одну или более биологическую активность указанной молекулы-мишени, и
(г) выбор, как положительный результат, антитела этапа (в) с уровнем ингибирования одной или более биологической активности указанной молекулы-мишени более высоким, чем исходный уровень указанного ингибирования.
10. Моноклональное антитело, полученное способом по п.1, с увеличенной антагонистической активностью, характеризующееся тем, что оно включает аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID SEQ ID № 2 - SEQ ID № 6, SEQ ID № 22 - SEQ ID № 26, SEQ ID № 28, SEQ ID № 29, SEQ ID № 33 - SEQ ID № 39 и SEQ ID № 42 - SEQ ID № 49.
11. Моноклональное антитело по п.10, где указанное антитело является человеческим антителом.
12. Моноклональное антитело по п.10, где указанное антитело представляет собой антитело IgG1.
13. Выделенная нуклеиновая кислота, кодирующая моноклональное антитело по п.10, где указанная нуклеиновая кислота содержит нуклеиновую последовательность, выбранную из группы, состоящей из SEQ ID № 16 - SEQ ID № 20, SEQ ID № 50 - SEQ ID № 54, SEQ ID № 56, SEQ ID № 57, SEQ ID № 61 - SEQ ID № 67 и SEQ ID № 70 - SEQ ID № 77.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| CAVACINI L.A | |||
| et al., "Influence of Heavy Chain Constant Regions on Antigen Binding and HIV-1 Neutralization by a Human Monoclonal Antibody", The Journal of Immunology, 1995, 155: 3638-3644 | |||
| ENDO S., ARATA Y., "Proton Nuclear Magnetic Resonance Study of Human Immunoglobulins G1 and Their Proteolytic Fragments: | |||

