Область техники изобретения
Настоящее изобретение представляет собой оральную вакцину, действующую против вируса человеческой папилломы (в дальнейшем в настоящем изобретении упоминаемого иногда просто как "HPV"). Уровень техники изобретения
HPV является малым делегированным безоболочечным ДНК-вирусом с икосаэдрической структурой. Геном вируса содержит открытые рамки считывания (ОРС), обозначаемые Е1-Е7 и L1 и L2, в которых "Е" означает ранние, a "L" означает поздние участки. Гены L1 и L2 кодируют капсидные белки вируса. Ранние гены (Е) связаны с такими функциями, как вирусная репликация и клеточная трансформация. Белок L1 является основным капсидным белком, имеющим молекулярный вес от 55 до 60 кДа, который определяют посредством электрофореза в полиакриламидном геле. Белок L2 представляет собой минорный капсидный белок, который имеет рассчитанный молекулярный вес также от 55 до 60 кДа и кажущийся молекулярный вес от 75 до 100 кДа.
В последнее время в развитых странах смертность от рака шейки матки уменьшилась, но, несмотря на это, во всем мире рак шейки матки является пятой из лидирующих причин смерти от злокачественных опухолей и второй наиболее распространенной злокачественной опухолью у женщин. Определенные типы HPV, передающиеся половым путем, являются наиболее важным фактором риска в отношении рака шейки матки. В последних сообщениях показано, что от 30 до 50% молодых женщин, имевших незадолго до анализа свое первое половое сношение, имели HPV-инфекцию шейки матки. Удивительно, что в основном HPV-инфекции шейки матки вызываются типами HPV с высоким риском, способными индуцировать рак. Считается, что типами высокого риска HPV являются HPV-типы 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 67 и 68, и возможно, также некоторые другие типы.
Этот тревожно высокий показатель распространенности HPV-инфекции среди молодых женщин предполагает, что образовательные и социальные программы здравоохранения, нацеленные на профилактику HPV-инфекции, не могут быть достаточно эффективными в борьбе с раком шейки матки. Особенно приоритетной является профилактика инфекции типа HPV высокого риска у женщин в развивающихся странах и у молодых женщин, не проходивших скринингового обследования по раку шейки матки. Текущий цитологический скрининг (скрининг на наличие рака шейки матки) и лечение рака после начала заболевания представляет собой экономически неэффективный выбор. Широкое применение в национальном масштабе профилактических вакцин против типов HPV с высоким риском может уменьшить частоту возникновения рака шейки матки. Предполагается, что частоту возникновения рака шейки матки наполовину уменьшает даже единственное применение вакцины HPV16.
В ходе разработок HPV-вакцин сообщалось, что высокий уровень продукции белка HPV11 L1 в клеточной системе насекомого приводил к сборке вирусоподобных частиц (в дальнейшем в настоящем изобретении иногда упоминаемых просто как "ВПЧ") (непатентный документ 1). Также сообщалось об успешном синтезе ВПЧ HPV16 в этой клеточной системе насекомого (непатентный документ 2).
Позже нам удалось достичь продукции ВПЧ, являющихся производными HPV6 и HPV16, в делящихся дрожжах Schizosaccharomyces pombe (в дальнейшем в настоящем изобретении иногда упоминаемых просто как "S. pombe") (непатентный документ 3). Хотя получаемый выход ВПЧ из делящихся дрожжей составляет меньше, чем их выход из клеточной системы насекомого, экспрессирующая система, использующая делящиеся дрожжи, обеспечивает преимущества в крупномасштабном производстве вирусоподобных частиц (в дальнейшем в настоящем изобретении иногда упоминаемых просто как "ВПЧ") и безопасности применения у человека.
Первые сообщения о том, что парентеральная вакцинация (путем инъекции) HPV16 ВПЧ обеспечивала 100% защиту против инфекции HPV16 у женщин, опубликованы Koustsky et al. (непатентный документ 4).
К сожалению, вакцина НРЧ16-ВПЧ для инъекций является дорогой, поскольку требует передовых технологий и специального оснащения для ее производства и хранения. Дополнительно, необходимые для эффективности повторные инъекционные вакцинации имеют ограниченную осуществимость и непригодны к использованию в развивающихся странах с ограниченным числом подготовленных специалистов среди медицинского персонала этих стран. Кроме того, сообщалось, что инъекционная вакцинация с ВПЧ недостаточно индуцирует секреторный IgA, играющий главную роль в защитных свойствах слизистых оболочек (непатентный документ 5).
Иммунизация лимфоидной ткани слизистых оболочек (ЛТСО), которая является иммунной тканью, расположенной в респираторном и пищеварительном тракте, способна защищать против вирусов типа HPV, вызывающих инфекции слизистого эпителия матки и влагалища. Исследователи Balmelli et al. успешно индуцировали антитела слизистых оболочек, которые нейтрализуют HPV16 во влагалище, посредством интраназального введения HPV16-ВПЧ (непатентный документ 6).
Вместе с тем интраназальная вакцинация, подобно вакцинации путем инъекции, также вызывает проблемы, поскольку для нее необходимо изготовление относительно больших количеств очищенных HPV-ВПЧ. Для индукции выраженного иммунитета слизистой оболочки влагалища проводили попытки стимуляции ассоциированной с кишечником лимфоидной ткани (GALT) оральными вакцинами вируса человеческой папилломы (иногда упоминаемыми в настоящем изобретении просто как "оральные HPV-вакцины"). Из растений табака и картофеля производили две группы оральных HPV-вакцин, которые экспрессируют гены L1 HPV11 (непатентный документ 7) и HPV16 (непатентный документ 8).
Очистка HPV-вирусоподобных частиц (HPV-ВПЧ) раскрыта в патентном документе 1. В патентных документах 2-4 и патентном документе 5, соответственно, раскрыто изготовление HPV-вакцины из экспрессирующих систем в бакуловирусах и в клетках насекомых. Также раскрыта вакцина из нуклеиновой кислоты для иммунотерапии HPV (патентный документ 6).
Также сообщалось о терапевтической системе доставки микроорганизма, в которой используется невакцинный активный компонент (патентный документ 7).
Патентный документ 1: JP-А-2003-520188
Патентный документ 2: JP-А-2001-519161
Патентный документ 3: JP-А-2002-516291
Патентный документ 4: JP-А-2002-510976
Патентный документ 5: JP-А-2004-269
Патентный документ 6: ТР-А-2004-121263
Патентный документ 7: JP-А-10-506791
Непатентный документ 1: Rose R.C., et al. J. Virol. J Virol. 1993; 67:1936-44.
Непатентный документ 2: Kirnbauer R., et al. J Virol. 1993; 67:6929-36.9.
Непатентный документ 3: Sasagawa T., et al. Virology, 1995; 206:126-35.
Непатентный документ 4: Koutsky L.А., et al. N. Engl. J Med. 2002; 347:1645-51.
Непатентный документ 5: Hagensee М.Е., et al. Virology 1995; 206:174-82.
Непатентный документ 6: Balmelli С., et al. J Virol. 1998; 72:8220-9.
Непатентный документ 7: Warzecha Н., et al. J Virol. 2003; 77:8702-11.
Непатентный документ 8: Biemelt S., et al. J Virol. 2003; 77:9211-20.
Раскрытие изобретения
Задачи изобретения
Задача согласно изобретению состоит в обеспечении оральной HPV-вакцины, которая является недорогой и доступной в больших количествах.
Средство решения задачи
Авторы выполнили настоящее изобретение, проводя исследования по изучению полезности рекомбинантных делящихся дрожжей, экспрессирующих HPV-белки в качестве вакцины, для решения вышеупомянутых задач.
В настоящем изобретении обеспечивается следующее:
1. Оральная вакцина вируса человеческой папилломы, полученная путем инкубации трансформанта авирулентных делящихся дрожжей-хозяев, в которой трансформант несет ген, кодирующий введенный в него антигенный белок вируса человеческой папилломы и накапливает в себе экспрессированный антигенный белок.
2. Вакцина по п.1, где авирулентными делящимися дрожжами-хозяевами являются Schizosaccharomyces pombe.
3. Вакцина по п.1 или 2, где вирус человеческой папилломы представляет собой вирус человеческой папилломы типа 16 (HPV16).
4. Вакцина по п.п.1, 2 или 3, где ген, кодирующий антигенный белок, представляет собой ген, кодирующий капсидный белок вируса человеческой папилломы.
5. Вакцина по п.4, где капсидный белок вируса человеческой папилломы представляет собой L1-белок.
6. Вакцина по п.4 или 5, где капсидный белок вируса человеческой папилломы представляет собой L1-белок вируса человеческой папилломы белкового типа 16 (HPV16-L1).
7. Вакцина по п.4, 5 или 6, где белок, накопленный в трансформанте, собран в вирусоподобные частицы.
8. Вакцина по любому из пп.1-7, представляющая собой лиофилизированный препарат трансформанта.
Эффекты, связанные с изобретением
Проводили наблюдения в случаях иммунизации оральной HPV-вакциной, в частности, оральной HPV16-вакциной согласно изобретению, и интраназальной иммунизации HPV-ВПЧ по индукции IgG в сыворотке и во влагалище и индукции IgA во влагалище. Все индуцированные антитела проявляли выраженную реакцию с HPV-ВПЧ-антигеном. Это указывает, что оральная HVP16-вакцина является эффективной в качестве профилактической вакцины против HPV16.
Краткие пояснения к чертежам
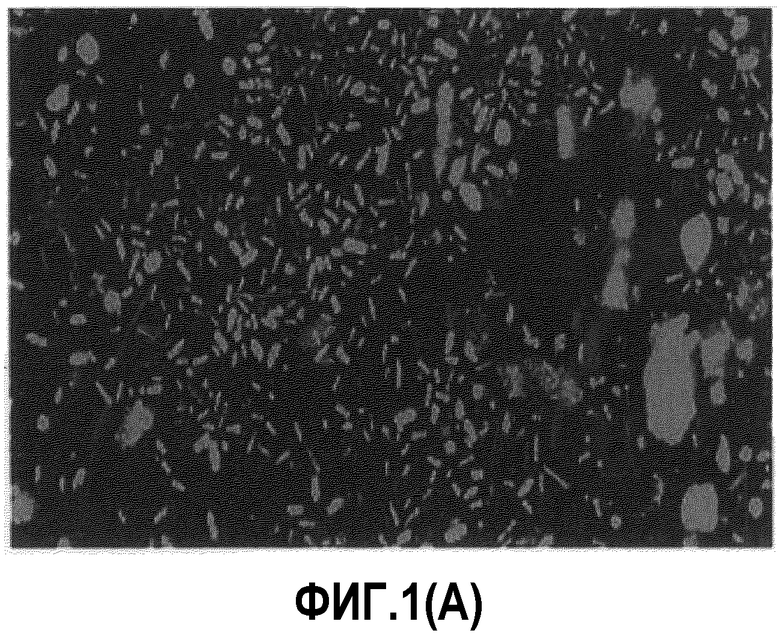
[Фиг.1А] Расщепление лиофилизированных дрожжей в желудке (Тестовый пример 1)
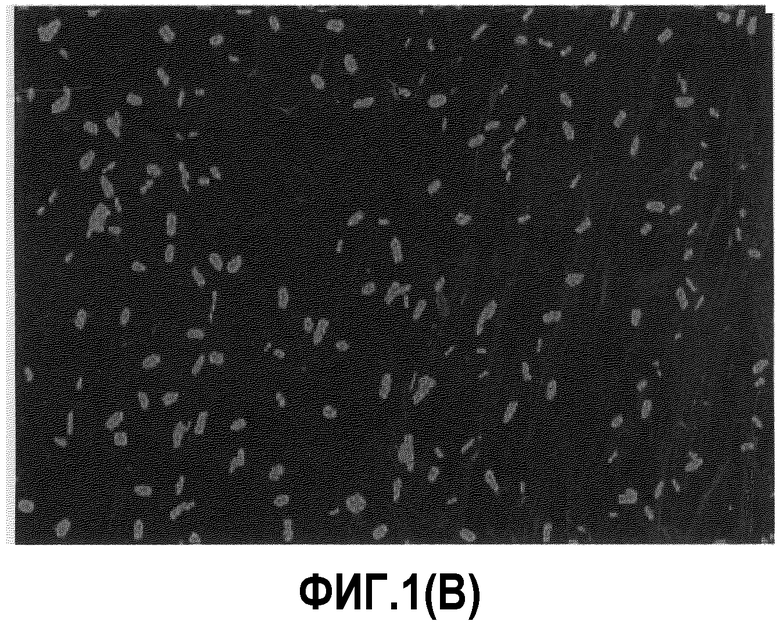
[Фиг.1B] Расщепление лиофилизированных дрожжей в брюшной полости (Тестовый пример 1)
[Фиг.1C] Расщепление лиофилизированных дрожжей в кишечнике (Тестовый пример 1)
[Фиг.2] Индукция антител путем иммунизации HPV16-вакцинами (Тестовый пример 2)
[Фиг.3] Индукция антител путем иммунизации HPV16-вакцинами, с последующей интраназальной бустер-иммунизацией HPV16-ВПЧ (Тестовый пример 3)
[Фиг.4] Изменение гуморальных иммунных ответов при иммунизации HPV16-вакциной, с последующей интраназальной бустер-иммунизацией HPV16-ВПЧ (Тестовый пример 5)
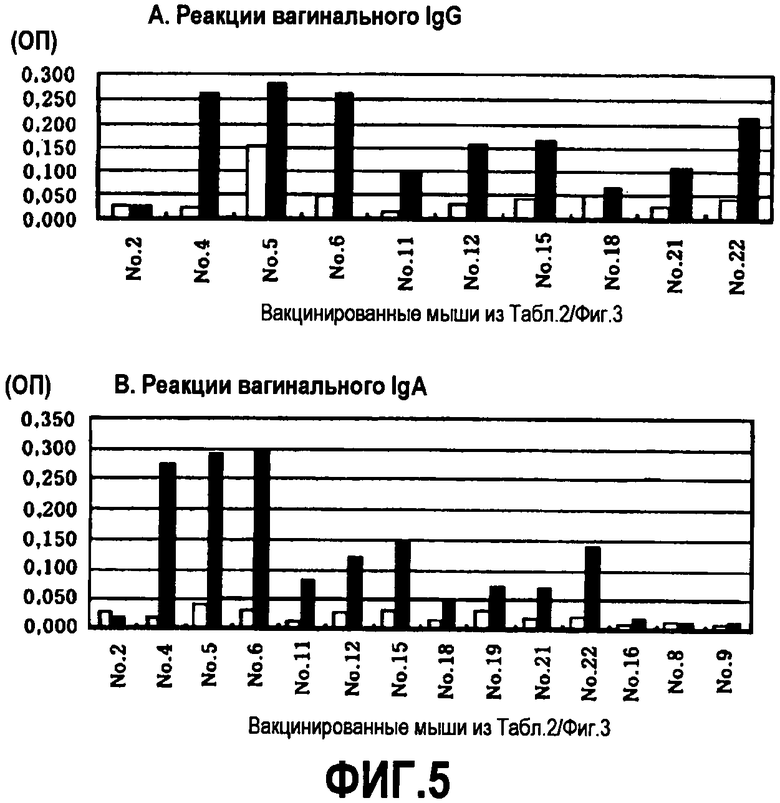
[Фиг.5] Изменения гуморальных иммунных ответов при иммунизации HPV16-вакцинами, с последующей интраназальной бустер-иммунизацией НРЧ16-ВПЧ (Тестовый пример 5)
Пояснения к обозначающим ссылкам
□ Реакция с денатурированным HPV16-L1-антигеном (фиг.4 и 5)
■ Реакция с HPV16-ВПЧ-антигеном (фиг.4 и 5)
Лучший вариант осуществления изобретения
Настоящее изобретение обеспечивает в качестве оральной HPV-вакцины инкубированный трансформант, несущий ген, кодирующий антигенный белок HPV, и экспрессирующий и накапливающий белок.
В настоящем изобретении хозяином, в который внедряют ген, кодирующий антигенный белок HPV для получения трансформанта, являются авирулентные делящиеся дрожжи, конкретно, делящиеся дрожжи S. pombe.
В настоящем изобретении "антигенный белок HPV" конкретно означает капсидный белок HPV, предпочтительно L1. В настоящем изобретении HPV предпочтительно является типом HPV высокого риска с опасностью возникновения рака шейки матки, конкретно, HPV16.
В настоящем изобретении можно без каких-либо ограничений использовать любой индуцибельный экспрессирующий вектор для S. pombe, в который вставлен чужеродный ген, такой как мультиклонирующие векторы, раскрытые в патентах JP-А-7-163373 и JP-А-11-192094, чтобы внедрить в хозяин S. pombe ген, кодирующий антигенный белок HPV. Например, можно использовать вектор, имеющий ген HPV16-L1, вставленный под контролем тиамин-репрессируемого промотора, для рекомбинантного синтеза вирусоподобных фрагментов (непатентный документ 3). Присутствие в вирусоподобных фрагментах белка L1 в 55 кДа можно подтвердить путем Вестерн-блоттинга или подобных анализов.
В настоящем изобретении "трансформант, в котором накапливается антигенный белок HPV", является трансформантом авирулентных делящихся дрожжей-хозяев, накапливающих антигенный белок HPV, и не имеет каких-либо конкретных ограничений. Говоря конкретно, трансформант (в дальнейшем в настоящем изобретении упоминаемый как "рекомбинант S. pombe") инкубируют при температуре от 23° до 37°С в течение 6-192 часов в известной культуральной среде, предпочтительно, в дрожжевой пептонно-декстрозной среде YPD при подходящих условиях для накапливания в нем экспрессируемого антигенного белка HPV. После инкубации культуральную среду, содержащую рекомбинантную S. Pombe, центрифугируют при подходящих условиях, например, при температуре от 0° до 50°С в течение от 1 до 60 минут при ускорении числа оборотов от 500 до 3000×g, для сбора преципитата перед пероральным введением.
Собранная путем центрифугирования рекомбинантная S. pombe называется дрожжевым дебрисом и может использоваться в качестве "трансформанта, накапливающего в себе антигенный белок HPV" согласно изобретению. При необходимости дрожжевой дебрис можно подходящим способом обрабатывать, например, путем лиофилизации, и "трансформант, накапливающий в себе антигенный белок HPV" согласно изобретению, охватывает такой обработанный дебрис. Лиофилизацию можно осуществлять в обычных условиях, например, при максимальной пригодной температуре 20°С в течение ночи, хотя конкретных ограничений во время лиофилизации S. pombe не существует. Оральная HPV-вакцина согласно изобретению охватывает трансформант, накапливающий полученный таким образом антигенный белок HPV.
Пероральная иммунизация имеет некоторые преимущества по сравнению с профилактической иммунизацией через другие пути введения. Например, оральные вакцины легко вводятся и воспринимаются реципиентами вакцины. Дополнительно, оральные вакцины могут содержать активные компоненты в более низких концентрациях, чем вакцины для инъекций, и поэтому стоимость их производства может быть ниже.
Рецептуру оральной HPV-вакцины согласно изобретению можно составлять в виде препаратов по меньшей мере с одним приемлемым с медицинской точки зрения дополнительным компонентом, таким как носитель, разбавитель, адъювант и/или буфер. Кроме того, оральную HPV-вакцину согласно изобретению можно использовать в виде смеси с продуктами питания или подобным образом. Можно использовать известные адъюванты, такие как адъювант LT (R192) слизистой оболочки, полученный из Е. coli.
Оральную HPV-вакцину согласно изобретению вводят с дозой дрожжей (в пересчете на сырую массу), выбранной в диапазоне от 10 до 500 мг/кг, предпочтительно от 20 до 200 мг/кг, или с дозой белка HPV16-L1, выбранной в диапазоне от 0,05 до 5 мг/кг, предпочтительно от 0,1 до 2 мг/кг. Оральную HPV-вакцину можно вводить однократно или повторно.
Предполагается, что оральная HPV-вакцина согласно изобретению является полезной в качестве профилактической вакцины против HPV16. Оральную HPV-вакцину согласно изобретению можно вводить совместно с обычными HPV-вакцинами для инъекций и/или с интраназальными HPV-вакцинами при обычных для них режимах вакцинации, например, в виде "сопутствующей вакцины" для использования в комбинации с известными вакцинами. Также ее можно применять в качестве "бустер-вакцины" для поддержки титров индуцированных антител.
ПРИМЕРЫ
В этом разделе настоящее изобретение будет описано со ссылкой на примеры и сравнительные примеры. Вместе с тем, примеры представляют собой простые варианты осуществления согласно изобретению, которые помогают воспроизводить настоящее изобретение, и никаким образом не ограничивают или не сужают объем согласно изобретению.
ПРИМЕР 1
Конструирование рекомбинантного штамма S. Pombe, экспрессирующего белок HPV16-L1
Рекомбинантный штамм S. pombe получали согласно непатентному документу 3. Для повышения экспрессии гена L1 ген HPV16-L1 (В27; дикий тип HPV16) вставляли в новый вектор, pTL2M (патент JP-А-7-163373). Рекомбинантный штамм S. pombe инкубирововали в 2 л среды YPD, чтобы дать возможность экспрессировать белок HPV16-L1. Методом Вестерн-блоттинга подтверждали высокий уровень экспрессии белка L1 в 55 кДа рекомбинантной S. pombe (pTL2-HPV16-L1). Методом электронной микроскопии подтверждали, что экспрессированный белок собран в вирусоподобные частицы.
Общее количество белков, экспрессируемых дрожжами, составляло около 10% от сырого веса дрожжей и около 50% от веса лиофилизированных дрожжей, и количество белка L1 составляло от 5 до 10% экспрессированных белков.
ПРИМЕР 2
Очистка инактивированных лиофилизированных дрожжей для изготовления вакцины
Рекомбинантный штамм S. pombe инкубировали в 2 л среды YPD и собирали путем центрифугирования при 4°С в течение 10 минут при 2000×g. Дрожжевой дебрис промывали фосфатно-буферным раствором (ФБР) и ресуспендировали в ФБР в количестве 150 мг/мл в пересчете на сырой вес. Суспензию S. pombe лиофилизировали в течение ночи при максимальной пригодной температуре от 13 до 20°С. Лиофилизированныые рекомбинантные S. pombe герметично закрывали в воздухонепроницаемые пластмассовые пробирки и сохраняли до применения при 4°С. Получаемую оральную вакцину для упрощения называют "дрожжи HPV16-L1".
Перед применением в виде оральной вакцины лиофилизированные дрожжи HPV16-L1 ресуспендировали более чем в десяти объемах 70% раствора этанола в воде и инкубировали при 4°С в течение 30 минут для инактивирования дрожжей, затем отделяли путем фильтрации и высушивали. Раздельные аликвотные количества лиофилизированных штаммов S. pombe обрабатывали этанолом, как перед пероральным введением. Затем "свежие живые дрожжевые клетки" обрабатывали этанолом, как перед пероральным введением.
ПРИМЕР 3
Вакцины для исследования расщепления дрожжей в пищеварительном тракте мышей
Рекомбинантный штамм S. pombe, экспрессирующий красный флуоресцентный белок (RFP), получали таким же способом, что и в примере 1. Рекомбинантый штамм S. pombe инкубировали, как описано выше, для экспрессии двух указанных белков. После инкубации штамм дрожжей центрифугировали при 4°С в течение 10 минут при 2000×g, и собирали дрожжевой дебрис.
ПРИМЕР 4
Адъювант
В качестве слизистого адъюванта использовали термо-лабильный токсин Escherichia coli LT (R192G), обеспечиваемый согласно публикации Curr Top Microbiol. Immunol. 1999; 236:216-36. Порошкообразный адъювант суспендировали в ФБР и сохраняли при температуре -30°С.
Сравнительный пример1
Очистка HPV16-ВПЧ для интраназальной иммунизации
Белок HPV16-ВПЧ очищали от рекомбинантных S. pombe, pTL2-HPV16-L1, полученных в примере 1, путем ультрацентрифугирования в градиенте хлорида цезия (Giga-Ната Y, et. Biotechnology (N Y) 1994; 12:400-4).
Клетки центрифугировали при температуре 4°С при 2000×g в течение 10 минут, ресуспендировали в 50 мМ фосфатно-калиевом буфере [50 мМ КН2Р04 (уровень рН 6,5), содержащем 20 мМ этилендиаминтетрауксусную кислоту (ЭДТА), и охлаждали на льду. Суспензию снова центрифугировали и дрожжевой дебрис ресуспендировали в 10 мл буфера KKC [20 мМ КРО4 (уровень рН 6,5), 800 мМ KCl, 0,1 мМ CaCl2, 1,5 мМ MgCl2, содержащем 5 мг/мл Novozyme] и инкубировали при 32°С в течение 30 минут для расщепления оболочки дрожжевой клетки, и разрушали клетки ультразвуком в течение 1 минуты при 60 W. Клеточный экстракт получали путем центрифугирования при 4°С в течение 10 минут при 7000×g. Осадок ресуспендировали в 10 мл буфера VLP (10 мМ HEPES, 10 мМ KCl (уровень pH 7,0)), содержащего 0,5% сурфактанта (NP-40), и клетки аккуратно разрушали ультразвуком, как указано выше.
Супернатант собирали и наслаивали на 40% сахарозный ВПЧ буфер и центрифугировали при 4°С в течение 2 часов при 27000×g в роторе Beckman SW28 (Beckman Coulter). Дебрис ресуспендировали в ВПЧ-буфере и центрифугировали при 4°С в течение 20 часов при 27000×g в уравновешенном CsCl ВПЧ-буфере в SW-роторе. Подходящие фракции разводили до плотности 1,29 г/мл и центрифугировали при 4°С в течение 2,5 часов при 27000×g.
Дебрис ресуспендировали в ВПЧ-буфере и хранили при температуре -30°С.
Сборку белка HPV16-ВПЧ подтверждали твердофазным иммуноферментным анализом (ELISA) с использованием двух анти-HPV16-моноклональных антител, Camvir-5 и Camvir-6 (предоставленных Margaret Stanley, Cambridg University), которые распознают конформационно-зависимые детерминанты.
Белок HPV16-ВПЧ использовали для интраназальной иммунизации. Интраназальную вакцину для упрощения называют "HPV16-ВПЧ".
Тестовый пример 1
Расщепление дрожжей в пищеварительном тракте мышей
Четырем мышам BALB/c в возрасте 9 недель после 12 часов голодания скармливали с часовыми интервалами 6 раз по 20 мг (в пересчете на сырой вес) свежих живых или лиофилизированных дрожжей, полученных в примере 3, экспрессирующих флуоресцентный белок (RFP). После последнего кормления мышей препарировали и исследовали на переваривание дрожжей. Пищеварительные тракты мышей рассекали, и на предметных стеклах получали мазок-отпечаток. На каждом слайде наблюдали флуоресценцию дрожжевых клеток под микроскопом Axiovert S-100 (Carl Zeiss, Германия) и фотографировали камерой Fuji 3CCD (Fuji Film).
Поскольку свежие живые дрожжевые клетки не расщеплялись, они выявлялись в большинстве участков пищеварительного тракта и выделялись со стулом.
Напротив, лиофилизированные дрожжевые клетки не разрушались в желудке (фиг.1A) или двенадцатиперстной кишке (фиг.1В), но от подвздошной кишки до толстой кишки количество клеток уменьшалось и было очень немногочисленным в прямой кишке (фиг.1С). Предполагается, что лиофилизированные дрожжевые клетки расщепляются в кишечнике, где расположена лимфоидная ткань, ассоциированная с кишечником.
Тестовый пример 2
Продукция антител, индуцированная оральной вакциной "Дрожжи HPV16-L1"
Самкам мышей BALB/c в возрасте 9 недель после 12 часов голодания скармливали свежие живые дрожжи. Мышей разделяли на 6 групп, как показано в таблице 1, вакцинировали с 4-недельными интервалами и исследовали на предмет продукции антител. Группа 1 являлась отрицательным контролем, и группа 2 служила положительным контролем. Дозы обозначены в пересчете на сырой вес.
После вакцинации у мышей забирали сывороточные и вагинальные образцы. Образцы сыворотки получали из крови, отобранной из хвоста мыши, и вагинальные образцы получали смывом из влагалища 100 мкл ФБР, используя микропипетку. Чтобы избежать влияния эстрального цикла мыши на продукцию антител, вагинальные образцы отбирали дважды с 5-дневным интервалом, и для анализа использовали два этих смешанных образца. Образцы отбирали за несколько дней до первой пероральной иммунизации и через четыре недели после каждой иммунизации. Чтобы избежать повторного размораживания, все образцы разделяли на аликвоты и хранили до применения при -30°С.
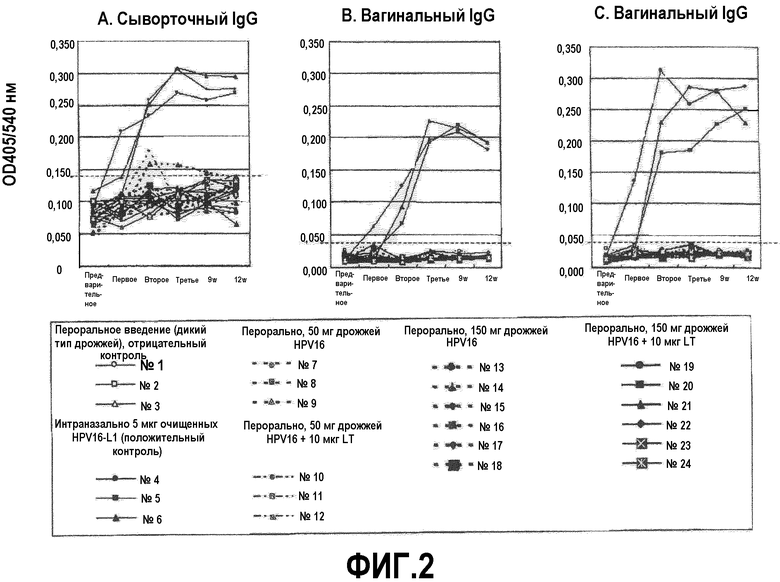
После трех иммунизаций оценивали уровни HPV16-специфических (IgG и IgA) антител методом ELISA (фиг.2).
1) ELISA
В качестве HPV16-ВПЧ-антигена для ELISA использовали белок HPV16-ВПЧ, очищенный от клеток насекомого, согласно Rose (J Virol. 1933; 67:1936-44, J Gen. Virol. 1994; 75:2445-9).
Для поверхностных антигенов, 100 нг и 300 нг HPV16-ВПЧ- антигена инкубировали в ФБР на планшетах ELISA (Nunc Immunoplate Maxisorp; Nalgene Nunc International) при 4°С в течение ночи.
Покрытые планшеты однократно промывали буфером ФБРТ (ФБР, 0,1% Tween-20) и инкубировали в течение 1 часа с блокирующим буфером (3% альбумина, 0,5% фетальной телячьей сыворотки ФТС в ФБРТ) при комнатной температуре (КТ; от 20° до 24°С). Все последующие промывки проводили буфером с ФБРТ.
В реакции антитела 1 мкл образцов сыворотки или 20 мкл вагинальных образцов смешивали с реакционным буфером (1,5% бычьего альбумина, 0,25% ФТС в ФБРТ), высевали в планшеты ELISA и инкубировали при комнатной температуре в течение 3 часов.
После трех промывок в планшеты добавляли биотинилированные антимышиные IgA- или IgG-антитела (разведенные реакционным буфером в отношении 1:1500 для IgA и 1:1000 для IgG) и инкубировали при комнатной температуре в течение 1 часа.
После трех промывок в планшеты добавляли 100 мкл конъюгата стрептавидина с хреном (DAKO, Германия), разведенного ФБРТ в соотношении 1:5000 и инкубировали в течение 30 минут.
После трех промывок в планшеты добавляли 100 мкл 50 мМ цитратного буфера (уровень рН 5,0; 0,0075% перекись водорода), содержащего одну таблетку АВТ [2,2'-азино-би-(3-этилбензеотиазолин-6-сульфоновой кислоты) (Sigma). После 1 часа проявления цвета при комнатной температуре измеряли значения оптической плотности (ОП) двухволновым способом (405/540 нм) посредством автоматизированного спектрофотометра для считывания планшетов (Iems Reader MS; Labsystems; Labsystems). Конечные значения ОП рассчитывали путем вычитания значения ОП при 540 нм из значения ОП при 405 нм.
2) Оценка
В 10 образцах сывороток и вагинальных образцов неиммунизированных мышей для титров IgA и IgG в сыворотке устанавливали точки среза в средних значениях ОП плюс допустимые отклонения в 2,5 раза. Анализ ELISA выполняли два или более раз для каждого образца, и для оценки использовали средние значения ОП. В каждый эксперимент включали два набора положительных и отрицательных контрольных образцов для поправки на межтестовую вариабельность.
У всех мышей положительного контроля наблюдали высокие значения ОП для сывороточного IgG, вагинального IgG и вагинального IgA, вместе с тем не выявлено реакций сывороточного IgA.
Наконец, у двух из этих 18 мышей (11%) (номер 7 в группе 3 и номер 14 в группе 5) выявлены транзиторные слабые реакции сывороточного IgG после второй иммунизации (фиг.2А). Напротив, никаких реакций вагинальных IgG или IgA не наблюдалось ни у одной перорально вакцинированной мыши (фиг.2В и 2С).
Тестовый пример 3
Повышение иммунных реакций путем интраназального введения очищенных HPV16-ВПЧ
Через 12 недель после конечной иммунизации интраназально в качестве бустера вводили субоптимальную дозу (1 мкг) HPV16-ВПЧ всем мышам, включая мышей положительного контроля, отрицательного контроля и мышей, перорально иммунизированных дрожжами HPV16-L1.
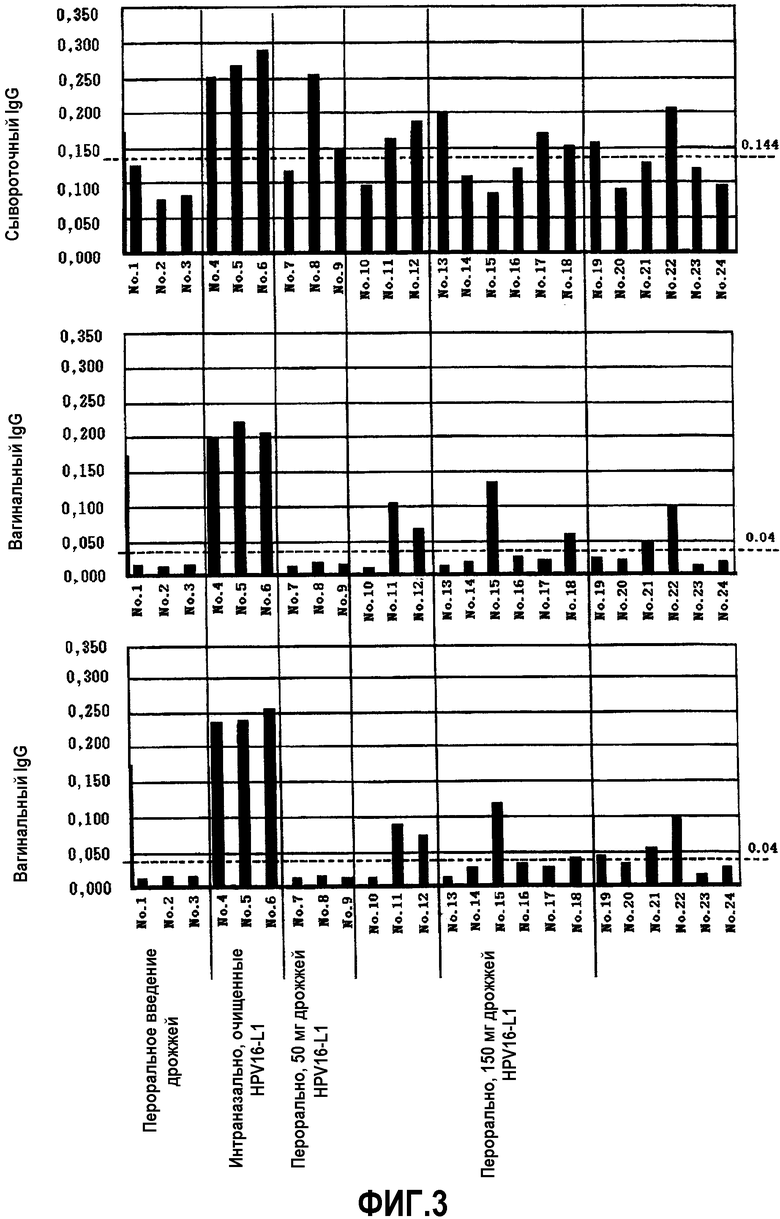
Результаты показаны в таблице 2 и на фиг.3.
Индуцированные титры сывороточного IgG и вагинального IgA у мышей
Через четыре недели у всех трех мышей положительного контроля (100%) и у 9 мышей, вакцинированных перорально, наблюдали положительные реакции на сывороточный IgG (фиг.3А). Что касается вагинального IgG, у этих трех мышей положительного контроля (100%) и у 6 мышей, вакцинированных перорально (33%), реакции были положительными, тогда как ни у одной мыши отрицательного контроля не было положительной реакции (фиг.3В). Относительно вагинальных IgA, эти три мыши положительного контроля (100%) и 7 мышей, вакцинированных перорально (39%), имели положительную реакцию, и положительной реакции не наблюдалось ни у одной мыши из отрицательного контроля (фиг.3С).
Не выявлено индукции каких-либо антител в отрицательном контроле даже при интраназальной бустер-иммунизации с субоптимальным количеством HPV16-ВПЧ после пероральной вакцинации дрожжами, тогда как у мышей, вакцинированных перорально дрожжами HPV16-L1, выявлены положительные иммунные реакции, что предполагает, что HPV16-специфические иммунные реакции могут быть премированы дрожжами HPV16-L1.
Тестовый пример 4
Эффекты пероральных доз дрожжей HPV16-L1 и адъюванта на иммунные реакции
Не было различий в антитело-позитивной интенсивности у мышей, вакцинированных HPV16-L1, при низких дозах дрожжей HPV16-L1 (50 мг) и при высоких дозах дрожжей HPV16-L1 (100 мг).
Некоторым мышам вводили слизистый адъювант LT (R192G) с дрожжами HPV16-L1 для повышения реакции антител. Адъювант не вызывал серьезных побочных эффектов у мышей. Адъювант не вызывал каких-либо отличий в реакциях сывороточных IgG среди групп, вакцинированных перорально дрожжами HPV16-L1, тогда как в группах адъюванта антитело-позитивная интенсивность вагинальных IgG была в два раза выше, и у вагинальных IgA была в 2,5 раза выше, чем в неадъювантных группах, хотя статистически существенных отличий не выявлено. Значения ОП для вагинальных IgA были слегка выше в адъювантной группе мышей, вакцинированных дрожжами HPV16-L1, чем в неадъювантной группе мышей, вакцинированных дрожжами HPV16-L1 (Р=0,085; тест Mann-Whitney), что предполагает наличие небольшого эффекта адъюванта.
Пример 5
Характеристика антител, активированных в результате пероральной и интраназальной вакцинации
Реактивность антител анализировали способом ELISA с использованием НРУ16-ВПЧ и денатурированных HPV16-L1 поверхностных белков.
HPV16-ВПЧ кипятили в бикарбонатном буфере (100 нг для IgG и 300 нг для IgA) в течение 10 минут и использовали в качестве денатурированного HPV16-L1 белкового антигена для ELISA. Сравнивали реактивность к ВПЧ-антигену или к денатурированному HPV16-L1-антигену, отражающуюся в ОП. Сыворотку и вагинальные смывы подвергали серийному разведению и измеряли титры антител.
Предыдущие исследования показали, что антитела, распознающие конформационно-зависимые детерминанты на HPV-ВПЧ, нейтрализуют вирус. В настоящем изобретении было изучено то, проявляют ли индуцированные антитела более выраженную реакцию на HPV16-ВПЧ-антиген, чем на денатурированный HPV16-L1-антиген.
Антитела, индуцированные при первой интраназальной иммунизации у двух интраназально вакцинированных HPV16-L1-мышей (№4 и №6), проявляли более выраженную реакцию на HPV16-L1-антиген. Вместе с тем, более выраженную реакцию к сывороточному IgG проявляли антитела, индуцированные HPV16-ВПЧ-антигеном при второй иммунизации или позже (фиг.4А). Это указывает на сероконверсию от неспецифического типа реакции к HPV16-специфическому типу реакции после второй иммунизации (фиг.4А)
Антитела, индуцированные HPV16-ВПЧ-антигеном после интраназальной бустер-иммунизации у мышей, вакцинированных перорально HPV16-L1-дрожжами, проявляли более выраженную реакцию.
Напротив, две мыши, вакцинированные перорально HPV16-L1-дрожжами, проявляли транзиторную позитивную реакцию, но в итоге негативную реакцию после бустер-иммунизации. Антитела в одном случае (№ 7) из двух проявляли более выраженную реакцию с денатурированным HPV16-L1-антигеном, тогда как антитела в другом случае (№ 14) показывали одинаковую реакцию с обоими типами HPV16-L1-антигена в течение всего периода эксперимента (фиг.4В).
Результаты анализа вагинальных образцов от мышей, вакцинированных перорально дрожжами HPV16-L1, показали, что IgG-антитела вырабатывались у 6 мышей (фиг.5А). У всех этих мышей активированные антитела проявляли более выраженную реакцию с HPV16-ВПЧ-антигеном.
Оральная вакцина с дрожжами HPV16-L1 вызывает HPV16-L1-специфические вагинальные IgA- и IgG-антитела, а также сывороточный IgG. Несмотря на то, что отдельно пероральное введение лиофилизированных дрожжей HPV16-L1 не вызывало индукции анти-HPV16-L1-антител, при последующей интраназальной бустер-иммунизации субоптимальными количествами HPV16-ВПЧ наблюдалось активирование сывороточного IgG, вагинального IgG и вагинального IgA, соответственно, у 50%, у 33% и у 39% мышей. У мышей отрицательного контроля не происходило выработки анти-HPV16-L1-антител даже после такой же интраназальной бустер-иммунизации. Интраназальное введение HPV16-ВПЧ повышало продукцию антител, вероятно, посредством усиления активности примированных В-клеток памяти, распознающих HPV16. Все индуцированные антитела проявляли более выраженную реакцию с HPV16-ВПЧ-антигеном, чем с денатурированным HPV16-L1-антигеном, что предполагает, что эти антитела распознают конформационно-зависимые HPV16-L1 детерминанты и являются нейтрализующими.
Промышленная применимость
Как описано выше, оральная вакцина HPV16-L1-дрожжей согласно изобретению индуцирует нейтрализующие антитела и является эффективной в качестве вакцины. В этой связи обеспечивается вакцина для отдельного применения, введение которой является менее болезненным для пациентов, чем интраназальные вакцины и инъекционные вакцины. Оральная вакцина не требует очистки и поэтому доступна в широком масштабе при низкой стоимости. Вакцину можно применять различными способами, прежде всего, для индукции анти-HPV-антител у неиммунизированных людей, а также при бустер-иммунизации для поддержки титров индуцированных антител.
Полное раскрытие Японской патентной заявки № 2004-263580, зарегистрированной 10 сентября 2004 года, включающей в себя описание, формулу изобретения, чертежи и реферат, включено в настоящее описание в виде ссылки во всей ее полноте.
| название | год | авторы | номер документа |
|---|---|---|---|
| СТАБИЛЬНЫЙ ВЕКТОР КОНСТИТУТИВНО ВЫСОКОЙ ЭКСПРЕССИИ ДЛЯ ПОЛУЧЕНИЯ ВАКЦИНЫ ПРОТИВ ВПЧ И ТРАНСФОРМИРОВАННЫЕ ЭТИМ ВЕКТОРОМ РЕКОМБИНАНТНЫЕ МОЛОЧНОКИСЛЫЕ БАКТЕРИИ | 2010 |
|
RU2492240C2 |
| МУКОЗАЛЬНАЯ ВАКЦИНА ДЛЯ ИММУНОТЕРАПИИ ЗАБОЛЕВАНИЙ, ОБУСЛОВЛЕННЫХ ВИРУСАМИ ПАПИЛЛОМЫ ЧЕЛОВЕКА, И СПОСОБ ЛЕЧЕНИЯ С ЕЕ ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2008 |
|
RU2377305C1 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2014 |
|
RU2546241C1 |
| ПОЛИНУКЛЕОТИДНАЯ ВАКЦИНА ДЛЯ ВИРУСА ПАПИЛЛОМЫ | 1995 |
|
RU2173170C2 |
| ОЧИЩЕННЫЕ БЕЛКИ ВИРУСА ПАПИЛЛОМЫ | 1995 |
|
RU2161651C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ДЛЯ ВВЕДЕНИЯ ЧЕРЕЗ СЛИЗИСТУЮ И СПОСОБ ПРЕДОТВРАЩЕНИЯ ГРИППА | 2004 |
|
RU2390351C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2011 |
|
RU2445357C1 |
| РЕКОМБИНАНТНЫЙ БЕЛОК, НЕСУЩИЙ ЭПИТОПЫ ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА, ВСТРОЕННЫЕ В БЕЛОК АДЕНИЛАТЦИКЛАЗУ ИЛИ ЕГО ФРАГМЕНТ, ЕГО ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2441022C2 |


Изобретение относится к области медицины и касается иммуногенной композиции. Сущность изобретения заключается в разработке иммуногенной композиции, полученной путем культивирования трансформанта авирулентных делящихся дрожжей-хозяев Schizosaccharomyces pombe, в которой трансформант несет ген, кодирующий введенный в него антигенный белок, представляющий собой капсидный белок L1 вируса человеческой папилломы типа 16 (HPV-16) и накапливает в себе экспрессированный антигенный белок для орального применения. Преимущество изобретения заключается в разработке иммуногенной композиции для орального применения. 2 з.п. ф-лы, 2 табл., 7 ил.
1. Иммуногенная композиция, полученная путем культивирования трансформанта авирулентных делящихся дрожжей-хозяев Schizosaccharomyces pombe, в которой трансформант несет ген, кодирующий введенный в него антигенный белок, представляющий собой капсидный белок L1 вируса человеческой папилломы типа 16 (HPV-16) и накапливает в себе экспрессированный антигенный белок, для орального применения.
2. Композиция по п.1, где белок, накопленный в трансформанте, собран в вирусоподобные частицы.
3. Композиция по п.1, где композиция представляет собой лиофилизированный препарат трансформанта.
| WARZECHA H., et al., Oral Immunogenicity of human papillomavirus-like particles expressed in potato, J | |||
| of Virology, 2003, v.77, №16, pp.8702-8711 | |||
| SASAGAWA Т., et al., Synthesis and assembly of virus-like particles papillomaviruses type 6 and type 16 in fission yeast Schizosacharomyces pombe | |||
| BIEMELT S | |||
| et al., Production of Human |

