Изобретение касается получения и использования нового фармацевтического продукта, нуклеиновой кислоты, которая при непосредственном введении в живую ткань позвоночного животного индуцирует иммунный ответ, который специфически узнает вирус папилломы.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Инфекции вируса папилломы (PV) имеют место у многих животных, в том числе у людей, овец, собак, кошек, кроликов, обезьян, змей и крупного рогатого скота. Вирусы папилломы инфицируют эпителиальные клетки, индуцируя обычно развитие доброкачественных эпителиальных или фиброэпителиальных опухолей в месте инфекции. Вирусы папилломы являются видоспецифическими инфекционными агентами; например, вирус папилломы человека обычно не инфицирует других животных.
Вирусы папилломы могут быть классифицированы на отдельные группы на основе хозяина, который они инфицируют. Человеческие вирусы папилломы (HPV) классифицируются далее на более чем 60 типов на основе гомологии последовательности ДНК (в качестве обзора см. Papilloma Viruses and Human Cancer, H. Pfister (ed.), CRC Press, Inc., 1990). По-видимому, инфекции вируса папилломы индуцируют тип-специфические иммуногенные реакции, в том смысле, что нейтрализующий иммунитет к инфекции для одного типа вирусов папилломы не создает иммунитета против другого типа вируса папилломы.
В случае людей, разные типы HPV вызывают различные заболевания. Типы HPV 1, 2, 3, 4, 7, 10 и 26-29 вызывают образование доброкачественных бородавок как у здоровых людей, так и у людей с ослабленным иммунитетом (иммунологически "скомпрометированных"). Типы HPV 5, 8, 9, 12, 14, 15, 17, 19-25, 36 и 45-50 вызывают плоские поражения у индивидуумов с ослабленным иммунитетом. Типы HPV 6, 11, 34, 39, 41-44 и 51-55 вызывают образование незлокачественных кондилом половых путей. Типы HPV 16 и 18 обусловливают эпителиальную дисплазию половых путей и ассоциируются с большинством in situ и инвазивных карцином шейки матки, влагалища, наружных женских половых органов и анального канала.
Иммунологические исследования в системах животных показали, что образование нейтрализующих антител к антигенам вируса папилломы предотвращает инфекцию гомологичным вирусом. Разработка эффективных вакцин человеческого вируса папилломы замедлялась неспособностью культивировать вирусы папилломы in vitro. Разработка эффективной вакцины HPV замедлялась, в частности, отсутствием подходящего животного-хозяина для прямого исследования HPV.
Нейтрализация вируса папилломы антителами является, по-видимому, тип-специфической и зависит от конформационных эпитопов на поверхности вируса.
Вирусы папилломы являются небольшими (50-60 нм), не имеющими оболочки, двадцатигранными (икосаэдральными) ДНК-вирусами, которые кодируют ранние и поздние гены. Открытые рамки считывания (ORF) геномов вируса обозначают Е1-Е7 и L1 и L2, где Е обозначает ранние и L обозначает поздние рамки считывания. L1 и L2 кодируют белки капсида вируса, Е1-Е3 и Е5-Е7 ассоциированы с функциями, такими как вирусная репликация и трансформация.
Белок L1 является основным капсидным белком и имеет мол. массу 55-60 К. Белок L2 является минорным капсидным белком, который имеет предсказанную мол. массу приблизительно 55 К и среднюю (кажущуюся) мол. массу 75-100 К, как определено электрофорезом в полиакриламидном геле. Электронно-микроскопические и иммунологические данные предполагают, что большая часть белка L2 является внутренней по отношению к белку L1. Белки L2 являются высококонсервативными среди различных вирусов папилломы, особенно 10 основных аминокислот при C-конце. ORF L1 является высококонсервативной среди различных вирусов папилломы.
Гены L1 и L2 использовали для образования рекомбинантных белков для потенциального использования в предотвращении и лечении инфекций вируса папилломы. Zhou et al. клонировали гены L1 и L2 HPV типа 16 в вектор-вирус коровьей оспы и инфицировали клетки млекопитающего CV-1 этим рекомбинантным вектором для получения вирус-подобных частиц (VLP). Эти исследования были интерпретированы как устанавливающие, что экспрессия как белка L1, так и белка L2 HPV типа 16 в эпителиальных клетках является необходимой и достаточной для сборки VLP. Экспрессия только белка L1 или только белка L2 или двойная инфекция клеток отдельными рекомбинантными векторными вирусами коровьей оспы, содержащими или L1, или L2 гены, не приводило к получению частиц.
Были получены рекомбинантные L1 и L2 бычьего вируса папилломы, происходящие из бактерий. Нейтрализующие сыворотки к рекомбинантным бактериальным белкам перекрестно реагировали с нативным вирусом при низких уровнях, предположительно благодаря различиям в конформациях нативных и происходящих из бактерий белков.
Рекомбинантные бакуловирусы, экспрессирующие ORF L1 или L2 HPV16, использовали для инфицирования клеток насекомых SF9 и получения рекомбинантных белков L1 и L2. Вестерн-блоттинги показали, что полученные из бакуловируса белки L1 и L2 реагировали с антителом к HPV16. Было также продемонстрировано образование белков L1 и L2 HPV16 рекомбинантными штаммами Saccharomyces cerevisiae.
Поскольку цитотоксические Т-лимфоциты (CTL) как в мышах, так и в человеке способны узнавать эпитопы, происходящие из консервативных внутренних вирусных белков, и считаются важными в иммунном ответе против вирусов, усилия были направлены на разработку CTL-вакцин, способных обеспечивать гетерологичную защиту против различных вирусных штаммов.
Известно, что CD8+ -CTL убивают инфицированные вирусами клетки, когда их Т-клеточные рецепторы узнают вирусные пептиды, ассоциированные с молекулами МНС (главного комплекса гистосовместимости) класса 1. Эти пептиды получают из эндогенно синтезированных вирусных белков, независимо от местоположения или функции белка внутри вируса. Таким образом, путем узнавания эпитопов из консервативных вирусных белков CTL могут обеспечивать перекрестную в отношении штаммов защиту. Пептиды, способные связываться с МНС класса 1 для узнавания цитотоксическими Т-лимфоцитами (CTL), происходят из белков, которые присутствуют в цитоплазме или эндоцитоплазматическом ретикулуме или проходят через него. Следовательно, как правило, экзогенные белки, вступающие в эндосомный путь процессинга (как в случае антигенов, предоставляемых молекулами МНС класса 11), являются неэффективными в генерировании CD8+-CTL-ответных реакций.
Усилия, предпринимаемые для получения CTL-реакций, включали в себя использование реплицирующихся векторов для продуцирования белкового антигена внутри клетки или фокусировались на введении пептидов в цитозоль. Оба эти подхода имеют ограничения, которые могут снижать их применимость в качестве вакцин. Ретровирусные векторы имеют ограничения в отношении размера и структуры полипептидов, которые могут экспрессироваться в виде слитых белков с сохранением способности рекомбинантного вируса к репликации, и эффективность векторов, таких как вирус коровьей оспы, для последующих иммунизаций может ослабляться иммунными ответными реакциями против самих векторов. Также вирусные векторы и модифицированные патогены имеют присущую им опасность, которая может препятствовать их использованию для человека. Кроме того, выбор пептидных эпитопов, которые должны предоставляться, зависит от структуры антигенов МНС индивидуума и, следовательно, пептидные вакцины могут иметь ограниченную эффективность благодаря разнообразию гаплотипов МНС в популяциях аутбридинга.
Было показано, что внутримышечное инокулирование полинуклеотидных конструкций, т.е. ДНК-плазмид, кодирующих белки, приводит в образованию in situ белка в мышечных клетках. Путем использования кДНК-плазмид, кодирующих вирусные белки, могут быть генерированы ответные реакции в виде образования антител, обеспечивающие гомологичную защиту против последующего заражения. Использование полинуклеотидных вакцин (PNV) для генерирования антител может приводить к увеличенной продолжительности гуморальных иммунных ответов, а также к обеспечению антигена, который может иметь правильные пост-трансляционные модификации и конформацию нативного белка (в сравнении с рекомбинантным белком). Вирусные белки, продуцируемые in vivo после иммунизации PNV, могут принимать их нативную конформацию, индуцируя тем самым образование нейтрализующих вирус антител. Генерирование CTL-ответов этим способом предоставляет преимущества перекрестной защиты относительно штаммов без применения живого потенциально патогенного вектора или ослабленного вируса.
Benvenisty et а1. сообщили, что осажденная CaCl2 ДНК, введенная в мышей внутрибрюшинно, внутривенно или внутримышечно, могла экспрессироваться. Более недавно сообщалось, что внутримышечная инъекция экспрессионных ДНК-векторов в мышей приводит к поглощению ДНК мышечными клетками и экспрессии белка, кодируемого этой ДНК (J. A. Wolff et al., 1990; G. Ascadi et al., 1991). Было показано, что инъецированные плазмиды сохраняются вне хромосом и не реплицируются. Затем сообщалось о стойкой экспрессии после внутримышечной инъекции в скелетную мышцу крыс, рыбы и приматов и в сердечную мышцу крысы. Способ применения нуклеиновых кислот в качестве иммуногенных агентов сообщался в WO 90/11092 (4 октября 1990 года), в котором "голые" полинуклеотиды использовали для вакцинации позвоночных.
Этот способ не ограничивается внутримышечной инъекцией. Например, введение золотых микроснарядов, покрытых ДНК, кодирующей бычий гормон роста (ВGH), в кожу мышей приводило к образованию антител против ВGН в этих мышах. Струйный инжектор использовали для трансфекции кожной, мышечной, жировой тканей и ткани молочной железы живых животных. Различные способы для введения нуклеиновых кислот обсуждались в обзоре Donelli, Ulmer and Liu (The Immunologist, 2:20, 1994).
Данное изобретение рассматривает множество способов введения нуклеиновых кислот в живую ткань для индукции экспрессии белков. Изобретение обеспечивает способы введения вирусных белков в путь процессинга антигена для генерирования вирус-специфических CTL и антител. Так, потребность в специфических терапевтических агентах, способных индуцировать желаемые профилактические иммунные ответы против вирусных патогенов, удовлетворяется этим изобретением для вируса папилломы. Таким образом, это изобретение обеспечивает конструкции ДНК, кодирующие вирусные белки человеческого вируса папилломы, которые индуцируют специфические CTL и антитела.
Защитная эффективность ДНК-вакцинации против последующего вирусного заражения демонстрируется иммунизацией нереплицирующейся плазмидной ДНК, кодирующей один или более из вышеупомянутых вирусных белков. Это является преимуществом, поскольку при этом не участвует инфекционный агент, не требуется сборки вирусных частиц и возможна селекция антигенных детерминант. Кроме того, поскольку последовательность некоторых из этих генных продуктов является консервативной среди различных типов вирусов папилломы, возможна защита против последующего заражения отличающимся типом вируса папилломы, который является гомологичным или гетерологичным по отношению к штамму, из которого получен этот клонированный ген.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩЕСТВА ИЗОБРЕТЕНИЯ
Конструкции ДНК, кодирующие генные продукты вируса папилломы, способные экспрессироваться при непосредственном введении в ткани животных, являются новыми профилактическими и терапевтическими фармацевтическими средствами, которые могут обеспечивать иммунную защиту против инфекции вирусом папилломы.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
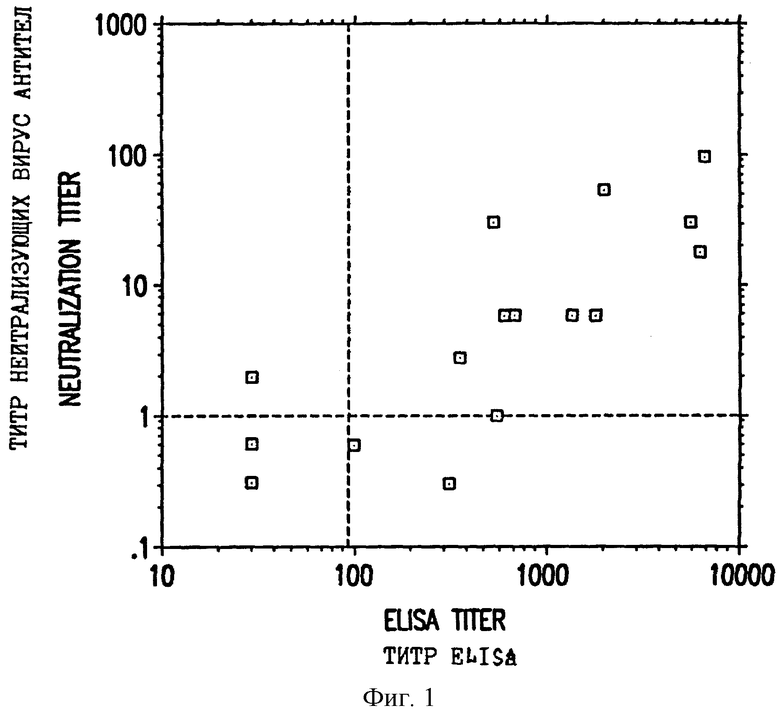
Фиг. 1 показывает ответную реакцию в виде нейтрализующих вирус антител, индуцируемую в кроликах, инъецированных ДНК L1 CRPV или смесью ДНК L1 и L2 (ось y), и соответствующие титры ELISA (твердофазного иммуноферментного анализа), индуцированные при этом.
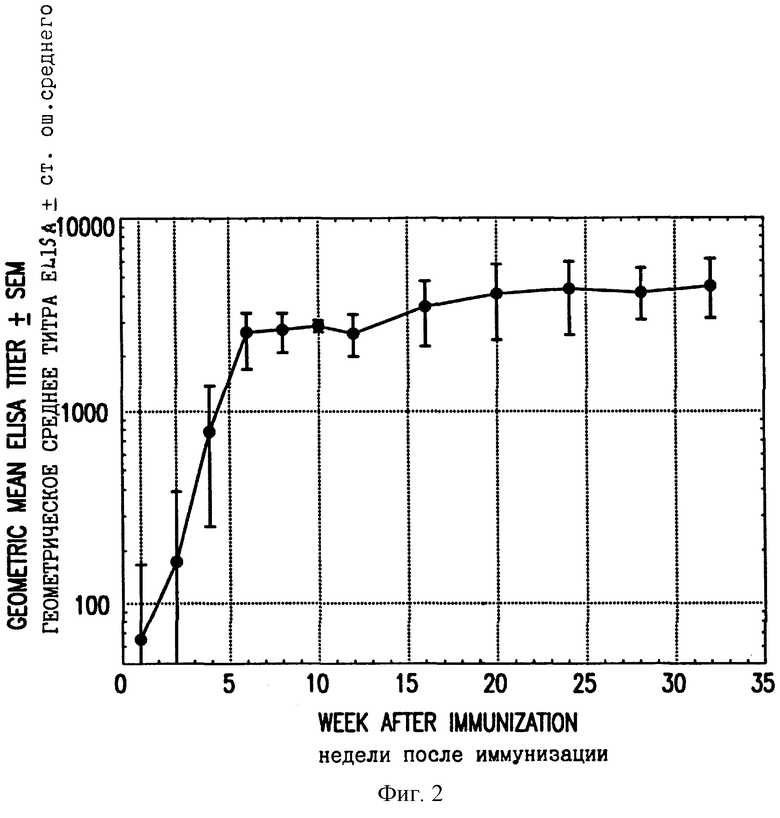
Фиг. 2. Образование антител в кроликах, инъецированных ДНК L1. Показаны титры ELISA против VLP L1 кроликов, получивших единственную иммунизацию произвольно выбранной дозой 1 мг ДНК L1. Кролики, инъецированные контрольной ДНК, не продуцировали детектируемых антител против VLP L1.
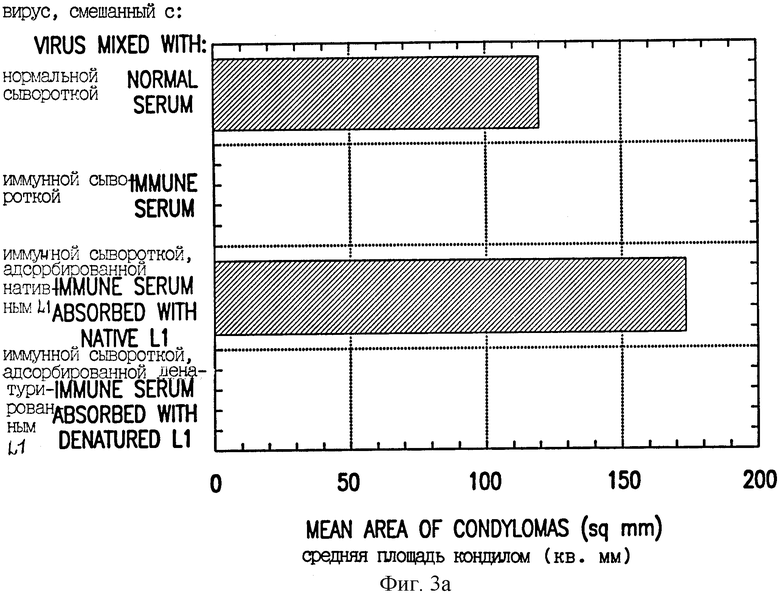
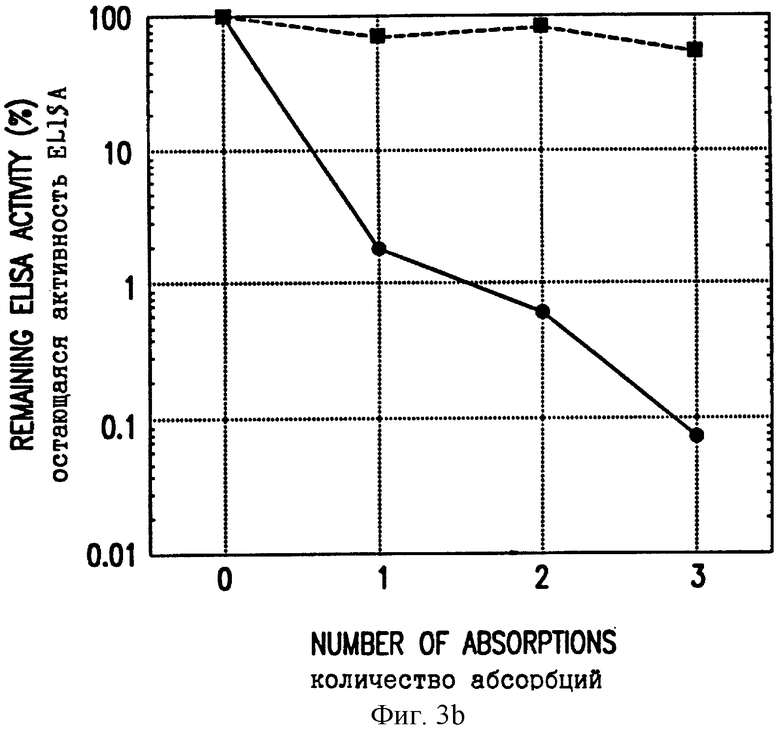
Фиг. 3. Влияние адсорбции VLP L1 на антисыворотку, полученную иммунизацией ДНК L1. A, Нормальную сыворотку и иммунную сыворотку, адсорбированную нативными или денатурированными VPL, как в (15), тестировали на нейтрализующую вирус активность. Показаны средние площади кондилом на 3 подвергнутых заражению местах, измеренные через 7 недель после заражения. B, Иммунную сыворотку из кролика, которого инъецировали ДНК L1, последовательно адсорбировали три раза нативными (кружки) или денатурированными (квадраты) VLP L1, экспрессируемыми в рекомбинантном штамме дрожжей (Saccharomyces cerevisiae). После каждой последовательной адсорбции аликвоты сыворотки анализировали на активность антител против полученных из бакуловируса VLP L1 при помощи ELISA. Титр ELISA адсорбированного материала наносили на график в виде процента от исходного титра ELISA неадсорбированной сыворотки.
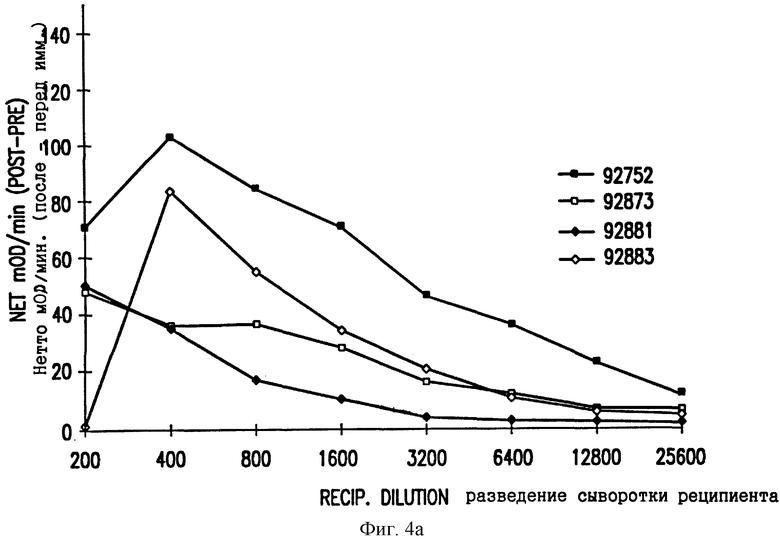
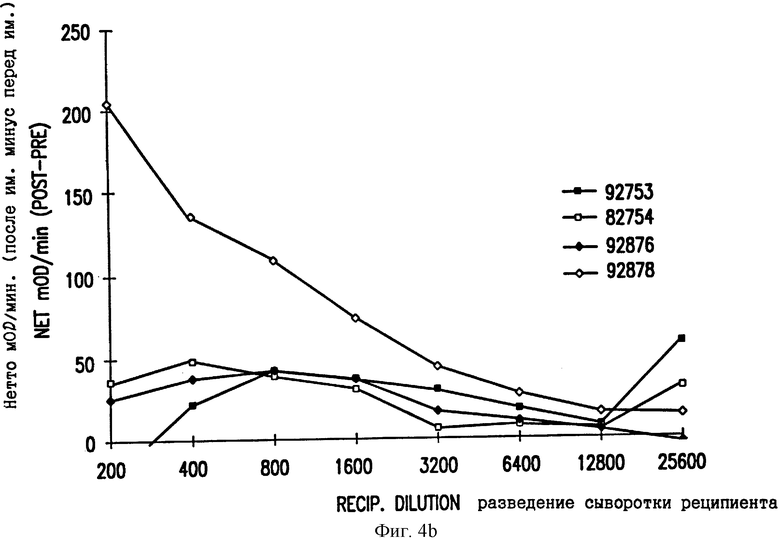
Фиг. 4. Ответные реакции ELISA в тестах на антитела против E2 CRPV (A) и против E7 CRPV (B). Нетто-скорость реакции (скорость после дозы 4 минус скорость перед иммунизацией при том же самом разведении) в OD/мин. показана для каждого отдельного кролика.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Конструкции ДНК, кодирующие генные продукты вируса папилломы, способные экспрессироваться при непосредственном введении в ткани животных, являются новыми профилактическими и терапевтическими фармацевтическими средствами, которые могут обеспечивать иммунную защиту против инфекции вирусом папилломы.
Это изобретение обеспечивает полинуклеотиды, которые при прямом введении в позвоночное животное, такое как американский кролик и человек, индуцируют экспрессию кодируемых пептидов в тканях этого животного. Если пептид является пептидом, который не встречается в этом животном, за исключением случая его инъекции, например, в случае инъекции белков, ассоциированных с вирусом папилломы (PV), иммунная система этого животного активируется для запуска защитной реакции. Поскольку эти экзогенные белки продуцируются клетками хозяина-животного, они процессируются и предоставляются главным комплексом гистосовместимости (МНС). Это узнавание аналогично узнаванию, которое имеет место при истинной инфекции родственным организмом. Результатом, как показано здесь, является индукция иммунных ответов, которые могут защищать против вирулентной инфекции.
В применении здесь полинуклеотид представляет собой нуклеиновую кислоту, которая содержит основные регуляторные элементы, так что при введении в живую клетку позвоночного он может направлять клеточные механизмы на продуцирование продуктов трансляции, кодируемых генами, содержащимися в этом полинуклеотиде. Имеется много вариантов данного изобретения, которые могут использовать специалисты в данной области. Например, можно использовать различные транскрипционные промоторы, терминаторы, векторы-носители или специфические последовательности генов.
Это изобретение обеспечивает нуклеиновые кислоты, которые при введении в ткани животного in vivo индуцируют экспрессию генного продукта вируса папилломы. Так, например, инъекция конструкций ДНК этого изобретения в мышцу кролика индуцирует экспрессию генных продуктов и вызывает образование нейтрализующих вирус антител. При последующем заражении вирусом папилломы американского кролика (CRPV) дозами, которые вызывают поражения на всех контрольных кроликах, животные, инъецированные этой полинуклеотидной вакциной, обнаруживают значительно уменьшенные поражения. Таким образом, это изобретение описывает вакцину, применимую в человеке, для предотвращения инфекций вируса папилломы.
Конструкции ДНК, кодирующие белки вируса папилломы, вызывают защитные иммунные ответы в животных. Как будет описано более подробно ниже, иммунные ответы в животных включали в себя нейтрализующие вирус антитела и защиту от вирусного заражения в кроликах гомологичными типами вируса папилломы.
В одном варианте продукт вакцины будет состоять из отдельных ДНК-плазмид, кодирующих, например, белки L1, L2, E2, E4, либо по отдельности, либо в комбинации.
Ожидаемые преимущества над другими вакцинами включают в себя (но не ограничиваются ими) увеличенный диапазон защиты благодаря CTL-ответам, увеличенный диапазон антител и увеличенную продолжительность защиты.
В одном варианте изобретения последовательность ДНК L1 или L2 или L1+L2 из HPV типа 6а, 6b, 11, 16 или 1, полученную из клинических изолятов, клонируют в экспрессионный вектор. Вектор содержит промотор для транскрипции РНК-полимеразой и терминатор транскрипции в конце кодирующей HPV последовательности. Примерами промотора являются промоторы CMV (но не только). Примеры терминаторов транскрипции включают в себя (но не ограничены ими) ВGН. Кроме того, для облегчения приготовления фармацевтического средства маркер устойчивости к антибиотику, экспрессируемый в Е. coli, также предпочтительно включают в экспрессионный вектор. Можно использовать гены устойчивости к неомицину или любой другой приемлемый маркер устойчивости к антибиотику. Далее, для содействия продуцированию с высокими уровнями фармацевтического агента посредством ферментации в прокариотических организмах, выгодно, чтобы вектор содержал начало репликации и имел высокую копийность. Многие коммерчески доступные прокариотические клонирующие векторы обеспечивают эти преимущества. Желательно удалять несущественные последовательности ДНК.
Таким образом, это изобретение обеспечивает экспрессионные векторы, кодирующие белок PV в качестве иммуногена. Изобретение предоставляет средство для индукции перекрестного в отношении типа защитного иммунитета без необходимости в самореплицирующихся агентах. Кроме того, иммунизация при помощи ДНК предоставляет ряд других преимуществ. Во-первых, этот подход к вакцинации должен быть применимым к опухолям, а также к инфекционным агентам, так как CD8+-CTL-реакция важна для иммунологического вмешательства в обоих патофизиологических процессах. Поэтому индуцирование иммунного ответа против белка, критического для процесса перерождения, может быть эффективным средством противораковой защиты или иммунотерапии. Во-вторых, образование антител против экспрессируемых белков после инъекции ДНК, кодирующей вирусный белок, предполагает, что эта технология обеспечивает легкое и эффективное средство для получения индуцирующих антитела вакцин.
Легкость получения и очистки конструкций ДНК выгодно отличается от традиционной очистки белков, что облегчает получение комбинаторных вакцин. Так, можно приготовить, смешать и совместно вводить множественные конструкции, например конструкции, кодирующие белки L1 и L2 одного или более типов HPV. Наконец, поскольку экспрессия белка может поддерживаться в течение периода времени после инъекции ДНК, стойкость памяти B- и Т-клеток может быть усилена, что порождает тем самым длительный гуморальный и клеточно-опосредованный иммунитет.
Ограничения предложенных HPV-вакцин придают особое значение необходимости разработки более эффективных средств для предотвращения инфекции и ослабления заболевания. Генерирование улучшенной CTL-реакции против консервативного белка может обеспечить значительный долгосрочный перекрестный иммунитет.
Мы продемонстрировали экспрессию белка из PNV-конструкций в кроликах путем детектирования иммунного ответа хозяина, направленного против антигенов CRPV. Результаты этих экспериментов на животных указывают, что прямая инъекция ДНК может обеспечить способ для защиты людей против инфекции HPV и заболевания.
Диапазон доз сравнивается по иммуногенности для оптимизации концентраций для использования. Прогнозируется, что дозы 10, 50, 100 и 200 мкг ДНК являются эффективными для человека.
Эффективность для человека показана на добровольцах, которые получают ДНК-вакцину HPV. Состав, доза и режим введения этой вакцины основаны на предшествующих исследованиях. Клиническая эффективность показана в виде скорости инфекции, показателей заболевания и продолжительности болезни. Эти клинические результаты сравниваются с лабораторной оценкой иммунного ответа хозяина и обнаружением вируса, для определения суррогатных маркеров, которые коррелируют с защитой.
Молекулярная биология для получения и очистки конструкций ДНК позволяет получение фармацевтических ДНК-агентов этого изобретения. Хотя стандартные способы молекулярной биологии являются достаточными для получения продуктов изобретения, специфические конструкции, описанные здесь, обеспечивают новые терапевтические средства, которые могут обеспечивать перекрестную защиту.
Количество экспрессируемой ДНК, которое должно быть введено реципиенту вакцины, будет зависеть от силы транскрипционных и трансляционных промоторов, используемых в конструкции ДНК, и от иммуногенности экспрессируемого генного продукта. Обычно иммунологически или профилактически эффективную дозу приблизительно 1 мкг - 1 мг и предпочтительно приблизительно 10 мкг - 300 мкг вводят непосредственно в мышечную ткань. Подкожная инъекция, внутрикожное введение, вдавливание через кожу и другие способы введения, такие как внутрибрюшинная, внутривенная доставка или доставка ингаляцией, также обсуждаются. Также обсуждается, что должны обеспечиваться ревакцинации (бустер-иммунизация).
Полинуклеотид может быть "обнаженным", т.е. не связанным с какими-либо белками, адъювантами или другими агентами, которые влияют на иммунную систему реципиента. В этом случае желательно, чтобы полинуклеотид находился в физиологически приемлемом растворе, таком как (но не только) стерильный солевой или стерильный забуференный солевой раствор. Альтернативно, полинуклеотид может быть ассоциирован с липосомами, такими как лецитиновые липосомы или другие известные в этой области липосомы, в виде ДНК-липосомной смеси, или ДНК может быть ассоциирована с адъювантом, известным в этой области знаний, для усиления иммунных ответов, таким как белок или другой носитель. Выгодно использоваться могут агенты, которые способствуют клеточному поглощению ДНК, такие как (но не только) ионы кальция, вирусные белки и другие облегчающие трансфекцию агенты. Эти агенты обычно называют облегчающими трансфекцию агентами и фармацевтически приемлемыми носителями.
Существует несколько преимуществ иммунизации геном, а не продуктом гена. Одним преимуществом является относительная простота, с которой нативный или почти нативный антиген может предоставляться иммунной системе. Другим преимуществом иммунизации полинуклеотидом является потенциальная возможность вхождения этого иммуногена в путь МНС класса 1 и индуцирования CTL-реакции. Поскольку иммунизация полинуклеотидом может вызывать как гуморальный, так и клеточно-опосредованный ответы, следующим преимуществом может быть то, что она обеспечивает относительно простой способ обследования вирусных генов и вирусных типов на вакцинный потенциал. Иммунизация путем инъекции полинуклеотидов делает также возможной сборку многокомпонентных вакцин смешиванием отдельных компонентов.
В применении здесь термином ген называют сегмент нуклеиновой кислоты, кодирующий отдельный полипептид. Термины фармацевтическое средство и вакцина используются взаимозаменяемо для обозначения композиций, применимых для индуцирования иммунных ответов. Термины конструкция и плазмида используются взаимозаменяемо. Термин вектор используется для обозначения ДНК, в которую могут быть клонированы гены для использования в соответствии со способом этого изобретения.
В соответствии с этим, одним из вариантов этого изобретения является способ использования генов PV для индукции иммунных ответов in vivo в позвоночном, таком как млекопитающее, в том числе человек, предусматривающий:
а) выделение по меньшей мере одного гена PV,
б) соединение этого гена с регуляторными последовательностями таким образом, что ген оперативно связывается с регуляторными последовательностями, которые, при введении в живую ткань, направляют инициацию транскрипции и последующую трансляцию этого гена,
в) введение этого гена в живую ткань, и
г) необязательно, повторную иммунизацию дополнительным геном PV.
Другим вариантом этого изобретения может быть способ защиты против гетерологичных типов PV. Это выполняют введением иммунологически эффективного количества нуклеиновой кислоты, кодирующей консервативный эпитоп PV.
В другом варианте этого изобретения полинуклеотидная вакцина кодирует другой белок PV, такой как L1 или L2 или E1-E7 или их комбинации.
В другом варианте этого изобретения конструкция ДНК кодирует белки HPV типов 6a, 6b, 11, 16 или 18, причем эта конструкция ДНК способна экспрессироваться при введении в ткани животных in vivo и индуцировать иммунный ответ против экспрессируемого продукта, кодируемого геном HPV. Комбинации, содержащие такие конструкции с полинуклеотидами, кодирующими другие антигены, не относящиеся к HPV, также обсуждаются этим изобретением.
Примеры конструкций ДНК с геном HPV включают в себя: VIJ-L1, VIJ-L2, VIJ-E1, VIJ-E2, VIJ- E3, VIJ-E4, VIJ-E5, VIJ-E6, VIJ-E7, VIJ-Eli^E4, VIJ-E1^ E4-L1, VIJ-E2-C.
В специфических вариантах этого изобретения конструкция ДНК кодирует белок L1 CRPV, причем эта конструкция ДНК способна экспрессироваться при введении в ткани животного in vivo и индуцировать иммунный ответ против экспрессируемого продукта, кодируемого геном CRPV. Комбинации, содержащие такие конструкции с полинуклеотидами, кодирующими другие антигены, не относящиеся к CRPV, рассматриваются данным изобретением.
Примеры конструкций ДНК, кодирующих ген CRPV, включают в себя: VIJ-L1, VIJ-L2, VIJ-E1, VIJ-E2, VIJ-E3, VIJ-E4, VIJ-E5, VIJ-E6, VIJ-E7, VIJ-Eli^E4, VIJ-E1^E4-L1, VIJ-E2-C.
Фармацевтически применимые композиции, содержащие эту ДНК, могут быть приготовлены в соответствии с известными способами, например, смешиванием с фармацевтически приемлемым носителем. Примеры таких носителей и способов приготовления можно найти в Remington's Pharmaceutical Sciences. Для образования фармацевтически приемлемой композиции, пригодной для эффективного введения, такие композиции должны содержать эффективное количество ДНК HPV.
Терапевтические или диагностические композиции этого изобретения вводят индивидууму в количествах, достаточных для лечения или диагностики инфекций PV. Эффективное количество может изменяться в зависимости от таких факторов, как состояние индивидуума, вес, пол и возраст. Другие факторы включают в себя способ введения. Обычно эти композиции вводят в дозах от приблизительно 1 микрограмма до приблизительно 1 миллиграмма.
Фармацевтические композиции могут вводиться индивидууму различными путями, такими как подкожный, топический, пероральный и внутримышечный.
Вакцины этого изобретения содержат ДНК HPV, кодирующие рекомбинантные белки HPV, содержащие антигенные детерминанты, которые вызывают образование нейтрализующих антител в человеке-хозяине. Такие вакцины также достаточно безопасны для введения без риска клинической инфекции; не имеют токсичных побочных эффектов; могут вводиться эффективным способом; и совместимы с вакцинными носителями.
Эти вакцины могут вводиться множеством способов, например перорально, парентерально, подкожно или внутримышечно. Вводимая доза может варьировать в зависимости от состояния, пола, веса и возраста индивидуума; способа введения; и типа PV вакцины. Вакцину можно использовать в виде таких лекарственных форм, как капсулы, суспензии, эликсиры или жидкие растворы. Вакцина может быть приготовлена с иммунологически приемлемым носителем.
Вакцины вводят в профилактически или терапевтически эффективных количествах, т. е. в количествах, достаточных для генерирования иммунологически защитной реакции. Эффективное количество может изменяться в соответствии с типом PV. Вакцину можно вводить в виде единственной дозы или в виде множественных доз.
Способы данного изобретения позволяют готовить моновалентные и поливалентные вакцины для предотвращения инфекций PV. При помощи этих способов могут быть приготовлены либо моновалентные, либо поливалентные вакцины PV. Например, моновалентная вакцина HPV типа 16 может быть приготовлена с использованием ДНК, кодирующей белок L1 или белок L2 HPV 16 или белки L1+L2. Альтернативно, поливалентная вакцина HPV может быть приготовлена смешиванием ДНК, кодирующей белки L1 или L2 HPV или белки L1+L2 из различных типов HPV.
Эта ДНК может быть использована для генерирования антител. Термин "антитело" используют здесь для обозначения как поликлональных, так и моноклональных антител, а также их фрагментов, таких как Fv, Fab и F(ab)2, которые способны связывать антиген или гаптен.
ДНК PV и антитела данного изобретения могут быть использованы для серотипирования инфекции HPV или CRPV и для скрининга HPV. ДНК HPV и CRPV пригодны для приготовления наборов (китов) для детектирования и серотипирования HPV и CRPV. Такой набор мог бы содержать упакованный отдельно носитель, пригодный для плотного удерживания по меньшей мере одного контейнера. Кроме того, этот носитель мог бы содержать реагенты, такие как ДНК HPV или антитела против HPV, пригодные для детектирования множества типов HPV. Носитель может содержать также средства для детектирования, такие как меченый антиген или субстраты для ферментов или т.п.
Следующие далее примеры даны для дальнейшего раскрытия данного изобретения, однако без ограничения этого изобретения только этими примерами.
ПРИМЕР 1
ВЕКТОРЫ ДЛЯ ПОЛУЧЕНИЯ ВАКЦИНЫ
A) V1: Этот экспрессионный (экспрессирующий) вектор конструировали из pCMVIE-AKI-DHFR [Y. Whang et al., J. Virol. 61, 1796 (1987)]. Гены АКI и DHFR удаляли разрезанием этого вектора рестриктазой EcoR 1 и самолигированием. Этот вектор не содержит интрона A в промоторе CMV, так что его добавляли в виде ПЦР-фрагмента, в котором был делетирован внутренний сайт Sac 1 [при 1855 согласно нумерации, данной в B.S. Chapman et al., Nuc. Acids Res. 19, 3979 (1991)] . Матрицей, использованной для ПЦР-реакций, была плазмида pCMVintA-Lux, полученная лигированием фрагмента Hind III и Nhe 1 из pCMV6al20 [см. B. S. Chapman et al. , ibid.], включающего в себя энхансер/промотор hCMV-IE1 и интрон A, в сайты Hind 111 и Xba 1 pBL3 для образования pCMVintBL. Фрагмент гена люциферазы из 1881 п.н. (Hind 111 - Sma 1, заполненный фрагментом Кленова) из RSV-Lux [J.R. de Wet et al., Mol. Cell Biol. 7, 725, 1987] , клонировали в сайт Sal 1 pCMVIntBL, который был заполнен фрагментом Кленова и обработан фосфатазой.
Праймеры, использованные для получения интрона A:
5'-праймер, 5'-CTATATAAGCAGAGCTCGTTTAG-3'; (SEQ ID N 1)
3'-праймер, 5'-GTAGCAAAGATCTAAGGACGGTGACTGCAG-3'; (SEQ ID N 2)
Праймеры, использованные для удаления сайта Sac 1:
Смысловой праймер, SEQ ID N 3:
5'-GTATGTGTCTGAAAATGAGCGTGGAGATTGGGCTCGCAC-3'; и
Антисмысловой праймер, SEQ ID N 4:
5'-GTGCGAGCCCAATCTCCACGCTCATTTTCAGACACA TAC-3'.
Этот фрагмент ПЦР разрезали Sac 1 и Bgl 11 и встраивали в вектор, который был разрезан теми же самыми рестриктазами.
B) ЭКСПРЕССИОННЫЙ ВЕКТОР VIJ
Нашей целью при создании VIJ было удаление промотора и элементов терминации транскрипции из нашего вектора VI, для того чтобы поместить их в более определенный контекст, создать более компактный вектор и улучшить выходы очистки плазмиды.
VIJ получен из векторов VI и pUC19, коммерчески доступной плазмиды. VI расщепляли рестриктазами Sspl и EcoR1, получая два фрагмента ДНК. Меньший из этих фрагментов, содержащий промотор CMVintA и элементы терминации транскрипции бычьего гормона роста (ВGН), которые управляют экспрессией гетерологичных генов, очищали при помощи электрофореза из агарозного геля. Концы этого фрагмента ДНК затем "затупляли" при помощи фермента ДНК-полимеразы T4 для облегчения его лигирования с другим ДНК-фрагментом с "затупленными" концами.
Для обеспечения "каркаса" экспрессионного вектора была выбрана плазмида pUC19. Известно, что она продуцирует высокие выходы плазмиды, хорошо охарактеризована относительно ее последовательности и функции и имеет минимальный размер. Мы удалили весь lac оперон из этого вектора, который не был обязательным для наших целей и мог быть вредным для выходов плазмиды и экспрессии гетерологичных генов, частичным расщеплением рестриктазой Hael 1. Оставшуюся плазмиду очищали электрофорезом на агарозном геле, концы ее затупляли ДНК-полимеразой T4, обрабатывали ее щелочной фосфатазой кишечника теленка и лигировали с описанным выше элементом CMVint A/BGH. Получали плазмиды, обнаруживающие любую из двух возможных ориентаций элементов промотора в каркасе pUC. Одна из этих плазмид дала гораздо более высокие выходы ДНК в Е. coli и была названа VIJ. Эта структура вектора была подтверждена анализом последовательности участков соединения, и затем было показано, что она дает сравнимую или более высокую экспрессию гетерологичных генов в сравнении с VI.
C) ЭКСПРЕССИОННЫЙ ВЕКТОР VIJneo
Было необходимо удалить ген ampr, используемый для отбора на устойчивость к антибиотику бактерий, несущих VIJ, поскольку ампициллин нельзя использовать в широкомасштабных ферментерах для производства человеческих клинических продуктов. Ген ampr удаляли из каркаса pUC расщеплением рестриктазами Ssp 1 и Eam 11051. Оставшуюся плазмиду очищали электрофорезом в агарозном геле, затупляли на концах ДНК-полимеразой T4 и затем обрабатывали щелочной фосфатазой кишечника теленка. Коммерчески доступный ген kanr, полученный из транспозона 903 и содержащийся внутри плазмиды pUC4K, вырезали при помощи рестриктазы Pst1, очищали электрофорезом в агарозном геле и затупляли ДНК-полимеразой T4. Этот фрагмент лигировали с каркасом VIJ и получали плазмиды с геном kanr в любой ориентации, которые были названы VIJ N 1 и VIJ N 3. Каждая из этих плазмид была подтверждена рестрикционным анализом, секвенированием ДНК в участках соединения и было показано, что она продуцирует сходные количества плазмиды в сравнении с VIJ. Экспрессия продуктов гетерологичных генов также была сравнима с экспрессией VIJ для этих векторов VIJneo. Мы произвольно выбрали VIJneo N 3, называемую далее VIJneo, которая содержит ген kanr в той же ориентации, что и ген ampr в VIJ, в качестве экспрессионной конструкции.
D) ЭКСПРЕССИОННЫЙ ВЕКТОР VIJns
Сайт Sfi 1 добавляли к VIJneo для облегчения исследований интеграции. Коммерчески доступный Sfi 1-линкер (New England BioLabs) добавляли при сайте Kpn 1 внутри последовательности BGH этого вектора. VIJneo линеаризовали при помощи Kpn 1, очищали электрофорезом в агарозном геле, затупляли ДНК-полимеразой T4 и лигировали с затупленным Sfi 1-линкером. Клональные изоляты отбирали посредством рестрикционного картирования и подтверждали секвенированием сквозь линкер. Этот новый вектор был назван VIJns. Экспрессия гетерологичных генов в VIJns (с Sfi 1) была сравнима с экспрессией тех же генов в VIJneo (с Kpn 1).
ПРИМЕР 2
Получение конструкций ДНК, кодирующих белки вируса папилломы американского кролика
Источником ДНК CRPV для всех клонированных генов является CRPV-pLA11. Это весь геном CRPV, клонированный в pBR322 при сайте Sal 1 (Nasseri, М., Meyers, C. and Wettstein, F.O. (1989) Genetic analysis of CRPV pathogenesis: The L1 open reading frame is dispensable for cellular transformation but is required for papilloma formation, Virology 170, 321-325).
1. VIJns: Кодирующую L1 последовательность получали при помощи ПЦР с использованием ДНК CRPV-pLA11 в качестве матрицы. Были сконструированы праймеры ПЦР, содержащие сайты Bam H1 для расщепления после того, как ПЦР-фрагмент очищали на геле.
Праймеры, использованные для получения кодирующего района L1:
Смысловой праймер:
5'-GGNACAGGATCCACCATGGCAGTGTGGCTGTCTACGCAG-3' (SEQ ID N 7) Bam H1
Антисмысловой праймер:
5'-CCACATGGATCCTTAAGTACGTCTCTTGCGTTTAGATG-3'
(SEQ ID N 8) Bam H1
Этот ПЦР-фрагмент очищали электрофорезом в агарозном геле, разрезали Bam H1 и лигировали с VIJns, разрезанной Bgl 11.
2. VIJns-L2: Кодирующий район L2 получали при помощи ПЦР. Вектор CRPV-pLA11 имеет ген L2, прерываемый сайтом Sal 1, используемым при встраивании CRPV в pBR322. Поэтому матрицу для ПЦР получали разрезанием CRPV-pLA11 рестриктазой Sal 1 и лигированием ДНК CRPV в кольцевую форму при сайте Sal 1. Эту лигированную ДНК CRPV использовали в качестве матрицы для ПЦР. Конструировали праймеры ПЦР, содержащие сайты Bam H1 для расщепления после очистки ПЦР - фрагмента на геле.
Для получения кодирующего района L2 использовали следующие праймеры:
Смысловой праймер:
5'-GGTACAGGATCCACCATGGTTGCACGGTCACGAAAACGC-3'
(SEQ ID N 9) Bam H1
Антисмысловой праймер:
5'-CCACATGGATCCTTATTCTGCGTAGACAGCCACACT-3' (SEQ ID N 10) Bam H1
3. VIJns-E2: Кодирующий район E2 получали при помощи ПЦР с использованием ДНК CRPV-pLA1 1 в качестве матрицы. Конструировали праймеры ПЦР, содержащие сайты Bgl 11 для расщепления после очистки ПЦР-фрагмента на геле.
Для получения кодирующего района E2 использовали следующие праймеры:
Смысловой праймер:
5'-GGTACAAGAT CTACCATGGA GGCTCTCAGCCAGCGCTTA-3'
(SEQ ID N 11) Bgl 11
Антисмысловой праймер:
5'-CCACATAGAT CTCTAAAGCC CATAAAAATTCCCTAAAAACAC-
Bgl 11 -AC-3'(SEQ ID N 12)
4. VIJns-E4: Кодирующий район E4 получали при помощи ПЦР с использованием в качестве матрицы ДНК CRPV-pLA11. Конструировали праймеры ПЦР, содержащие сайты Bgl 11 для расщепления после очистки ПЦР-фрагмента на геле.
Для получения кодирующего района E4 использовали следующие праймеры:
Смысловой праймер:
5'-GGTACAAGAT CTACCATGAG CCATGGACATTGCAGGATAC-3'
(SEQ ID N 13) Bgl 11
Антисмысловой праймер:
5'-CCACATAGAT CTTTATAAGC TCGCGAAGCCGTCTATTCC-3'
(SEQ ID N 14) Bgl 11
5. VIJns-E7: Кодирующий район E7 получали при помощи ПЦР с использованием CRPV-pLA11 в качестве матрицы в одном случае и очищенную ДНК из штамма CRPV Крайдера в другом случае. Одни и те же праймеры ПЦР использовали для обеих матриц. Конструировали праймеры ПЦР, содержащие сайты Bgl 11 для расщепления после очистки ПЦР-фрагмента на геле при помощи электрофореза.
Для получения кодирующего района E7 использовали следующие праймеры:
Смысловой праймер:
5'-GGTACAAGAT CTACCATGAT AGGCAGAACTCCTAAGCTTA-3'
Bgl 11
Антисмысловой праймер:
5'-CCACATAGAT CTTCAGTTAC AACACTCCGGGCACAC-3'
6. pGEX-2T-E2: Кодирующий район E2 получали при помощи ПЦР, как описано для VIJns-E2. Этот фрагмент клонировали в pGEX-2Т в сайт Bam H1 для получения слияния внутри рамки с глутатион-S-трансферазой (GST). Эту конструкцию использовали для получения белка в Е. coli.
7. pGEX-2T-E4: Кодирующий район E4 получали при помощи ПЦР, как описано для VIJns-E4. Этот фрагмент клонировали в pGEX-2Т в сайт Bam H1 для получения слияния внутри рамки с глутатион-S-трансферазой (GST). Эту конструкцию использовали для получения белка в Е. coli.
8. pGEX-2T-E7: Кодирующий район E7 получали при помощи ПЦР, как описано для VIJns-E7. Этот фрагмент клонировали в pGEX-2Т в сайт Bam H1 для получения слияния внутри рамки с глутатион-S-трансферазой (GST). Эту конструкцию использовали для получения белка в Е. coli.
ПРИМЕР 3
Очистка плазмиды из Е. coli
Конструкции VIJ выращивали в течение ночи до насыщения. Клетки собирали и лизировали при помощи модификации способа со щелочным ДСН (Sambrook, J., Fritsch, E. F. , and Maniatis, Т., Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor, N. Y., ed. 2 (1989). Модификация состояла в увеличении объемов в три раза для клеточного лизиса и экстракции ДНК. ДНК очищали двойным бэндингом на градиентах CsCl-EtBr. Этидийбромид удаляли экстракцией 1-бутанолом. Полученную ДНК экстрагировали смесью фенол/хлороформ и осаждали этанолом. ДНК ресуспендировали в ТЕ (10 мМ Трис, 1 мМ ЭДТА), pH 8, для трансфекций и в 0,9% NaCl для инъекции в мышей. Концентрацию и чистоту каждого препарата ДНК определяли по OD260/280. Отношения 260/280 были более или равны 1,8.
ПРИМЕР 4
Получение специфических для CRPV антител in vivo
У пяти кроликов на группу брали кровь и затем их инъецировали 1,2 мл солевого раствора, содержащего 1 мг VIJns-L1, VIJns-L2, VIJns-L1 в смеси с VIJns-L2 (в целом 2 мг) или один VIJns (контрольный вектор, не кодирующий белок). Инокулят делили поровну между 6 внутримышечными местами на обеих задних лапах, обеих передних лапах и нижней части спины. Через три недели после первоначальной инъекции ДНК у кроликов брали кровь и проводили вторую инъекцию той же ДНК таким же образом. Через 4 недели после второй инъекции у животных опять брали кровь.
Сыворотки тестировали на вирус-нейтрализующие антитела путем смешивания 10-кратных серийных разведения иммунной сыворотки с разведением 1:3 исходного вируса CRPV (штамма Крейдера). Разведения готовили в модифицированной по способу Дульбекко среде Игла (ДМЕМ), дополненной 1% бычьим сывороточным альбумином (БСА). Исходный вирус CRPV (приобретенный у Dr.J.Kreider, Hershey, PA) получали из кожных фрагментов, полученных из диких американских кроликов, которые были инфицированы CRPV, и имплантированных под почечными капсулами бестимусных мышей. Полученные кондиломы гомогенизировали и осветляли центрифугированием, получая исходный вирусный препарат. Смеси иммунной сыворотки и исходного вируса инкубировали на льду по меньшей мере в течение 60 минут и затем 50 мкл каждой смеси наносили на площадь 1 кв. см выбритой скарифицированной кожи на спинках трех Новозеландских белых кроликов. Этих животных наблюдали спустя 7 недель для обнаружения присутствия бородавок и измеряли передне-задние и латеральные размеры (в мм) эллипсоидных бородавок. Титры в конечной точке определяли из частоты бородавок при разных разведениях интерполяцией по Reed-Muench. Титры нейтрализующих антител кроликов, инъецированных ДНК L1 или обеими ДНК L1 и L2, нанесены на ось y фиг. 1.
Сыворотки из кроликов, которые были инъецированы ДНК L1, тестировали также на антитела при помощи твердофазного иммуноферментного анализа (ELISA). Планшеты ELISA из полистирола покрывали в течение ночи при 4oC 1 мг/лунку полуочищенного рекомбинантного полученного из дрожжей белка L1 CRPV. Рекомбинантный L1 получали в S. cerevisiae и очищали, как описано Kirnbauer et al. (Proc. Nat. Acad. Sci. USA 89 : 12180-4, 1992), с небольшими модификациями, добавляли разведенные сыворотки и инкубировали в течение 1 часа при комнатной температуре (с качанием на орбитальном шейкере). Затем планшеты промывали и добавляли меченый пероксидазой хрена козлиный антикроличий IgG (Fc-специфический). После одного часа инкубирования с качанием планшеты промывали и добавляли субстрат. Планшеты прочитывали при 450 нм при помощи планшет-ридера ELISA (Molecular Devices Corp.), и полученные величины корректировали на фон вычитанием скорости реакции сыворотки, взятой перед иммунизацией, из скорости реакции сыворотки после иммунизации при одинаковом разведении. Титры определяли интерполяцией полученной кривой корректированных скоростей реакции в зависимости от величины скорости 10 мOD/мин. Титры ELISA кроликов, инъецированных ДНК L1 или ДНК L1+L2, наносили на ось x фиг. 1. Фиг. 1 показывает, что 12 из 13 сывороток, которые были положительными в тесте на нейтрализующую активность (log титра более или равен 1, т.е. положительный с неразведенной сывороткой), были также положительными в отношении антител в ELISA (титр ELISA более или равен 100). Четыре из 4 сывороток, которые были негативными в отношении нейтрализующих антител, имели титры ELISA, равные или меньшие, чем 350. Все сыворотки из кроликов, получивших либо только ДНК L2, либо контрольный вектор VIJ, имели титры ELISA менее 100. Взятые вместе эти данные показывают, что антитела, специфические для CRPV и способные нейтрализовать этот вирус, были получены после инъекции ДНК L2. Титры ELISA в кроликах сохранялись неснижающимися в течение по меньшей мере 32 недель после иммунизации (фиг. 2).
ПРИМЕР 5
Защита кроликов при заражении вирулентным CRPV
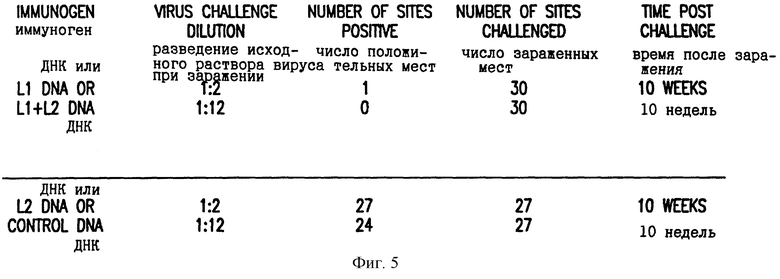
Пять кроликов на группу инъецировали внутримышечно 1 мг VIJns-L1, VIJnsL2, VIJns-L1, смешанными с VIJ-L2 (2 мг в целом), или только VIJns (контрольным вектором, не кодирующим белок), как описано выше. Через три недели после первоначальной инъекции ДНК кролики получали вторую инъекцию той же самой ДНК. Через 4 недели после второй инъекции кроликов заражали CRPV. Заражение CRPV проводили нанесением 50 мкл двух разведений исходного вируса (разведенного 1: 2 или 1:12 при помощи ДМЕМ +1% БСА) в тройных повторностях на места размером 1 кв. см выбритой скарифицированной кожи на спинке каждого кролика. Сыворотки, взятые при времени заражения, из животных, инъецированных ДНК L1 или ДНК L1+L2, содержали антитела к L1 согласно ELISA и имели вирус-нейтрализующую активность, как описано выше. Животные наблюдались в отношении образования бородавок при 3, 6 и 10 неделях после заражения. Среди кроликов, которые не получали ДНК L1, 51 из 54 мест, зараженных CRPV, развили бородавки, тогда как на животных, которые получали ДНК L1, только 2 из 60 мест развили бородавки. Одна из двух бородавок, которые наблюдались на кролике, иммунизированном ДНК L1, регрессировала в пределах 3 недель после их появления. Таблица на фиг.5 показывает распределение бородавок на кроликах после заражения CRPV. Профилактическая иммунизация ДНК L1 защищала кроликов от развития бородавок при инфицировании вирулентным CRPV.
ПРИМЕР 6
Конформационная специфичность антител, индуцируемых ДНК L1
Для демонстрации того, что защитные нейтрализующие антитела узнают конформационные эпитопы на VLP, проводили эксперименты по адсорбции. Адсорбция иммунной сыворотки VLP L1 (15) удаляла все нейтрализующие антитела и активность ELISA (фиг. 3A, B). Нанесение на скарифицированную кожу CRPV, смешанного с предиммунной кроличьей сывороткой, приводило к образованию кондилом на всех зараженных местах, тогда как при смешивании CRPV с иммунной сывороткой и таком же нанесении не были обнаружены кондиломы, вследствие активности нейтрализующих антител. При смешивании CRPV с иммунной сывороткой, которую адсорбировали белком VLP L1, из которой должны были удаляться нейтрализующие антитела, все (3/3) места были положительными в отношении кондилом. В противоположность этому, при адсорбции иммунной сыворотки денатурированным, не образующим частиц белком L1 (денатурированным восстановлением и алкилированием в 8 М мочевине) сыворотка все еще была способна нейтрализовать CRPV (фиг. 3A) и сохраняла свою активность в ELISA (фиг. 3B). Таким образом, вирус-нейтрализующие антитела, индуцируемые иммунизацией при помощи ДНК L1, могли удаляться VLP L1 в нативной конформации, но не денатурированным L1. Анализ ELISA, по-видимому, детектирует первичные конформационно-специфические антитела, реагирующие с интактными VLP L1, так как устранение активности в ELISA адсорбцией соответствовало удалению нейтрализующих антител.
ПРИМЕР 7
Гуморальные ответы (образование антител), индуцируемые ДНК E4 и E7
Группы из 4 Новозеландских белых кроликов инъецировали внутримышечно 1 мг ДНК VIJ-E2 или VIJ-E7 на одну иммунизацию. Проводили 4 иммунизации при 0, 4, 8 и 20 неделях и затем извлекали кровь при 22 неделях. Антитела использовали в качестве суррогатных маркеров экспрессии кодируемых белков. Сывороточные антитела анализировали с применением планшетов ELISA (NUNC Maxisorp), покрытых 1 мкг/лунку GST-E2 или слитого белка GST-E7, очищенного из Е. coli, которая была трансформирована экспрессирующим вектором pGEX, кодирующим E2 или E7 CRPV, и индуцирована IPTG. Анализ ELISA проводили, как описано в Примере 3. Нетто-скорости реакции (после иммунизации дозой 4 минус результат перед иммунизацией) в мOD/мин показаны на фиг. 4. Нетто-скорости более 10 мOD/мин считаются положительными; образцы с высокими титрами антител могут иметь низкие нетто-скорости реакции при самом низком разведении из-за пересыщения системы детектирования. Таким образом, белки E2 и E7, кодируемые их ДНК, экспрессируются и узнаются иммунной системой реципиента.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОЧИЩЕННЫЕ БЕЛКИ ВИРУСА ПАПИЛЛОМЫ | 1995 |
|
RU2161651C2 |
| ПАПИЛЛОМАВИРУСНЫЕ ВАКЦИНЫ | 1995 |
|
RU2206608C2 |
| ЭКСПРЕССИРУЮЩИЙ ВЕКТОР (ВАРИАНТЫ) | 1995 |
|
RU2177999C2 |
| КОНСТРУКЦИЯ ДНК (ВАРИАНТЫ), ДНК-ВЕКТОР, ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГРИППА, СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА, ВАКЦИНА И СПОСОБ ВАКЦИНАЦИИ | 1994 |
|
RU2193065C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ HPV 58 L1 В ДРОЖЖАХ | 2004 |
|
RU2370538C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ L1 HPV45 В ДРОЖЖАХ | 2004 |
|
RU2360001C2 |
| МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ HPV31 L1, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ВИРУСОПОДОБНАЯ ЧАСТИЦА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБЫ С ИХ ИСПОЛЬЗОВАНИЕМ | 2004 |
|
RU2356943C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ HPV 52 L1 В ДРОЖЖАХ | 2005 |
|
RU2373219C2 |
| ПОЛИНУКЛЕОТИДНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА | 1995 |
|
RU2186109C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, ПРИГОДНАЯ ПРИ ИНФЕКЦИЯХ HPV И ВИРУСОМ ГЕПАТИТА В, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2009 |
|
RU2509570C2 |
Изобретение относится к биотехнологии и иммунологии и может быть использовано для получения вакцины против вируса папилломы (НРV). Вакцина содержит полинуклеотид, содержащий ген человеческого вируса папилломы, выбранный из группы, состоящей из HPV 6а, HPV 6b, HPV 11, HPV 16 и HPV 18, и экспрессирующий белок L1 или L1+L2, промотор CMV, терминатор транскрипции из гена бычьего гормона роста, маркерный ген устойчивости к неомицину и физиологически приемлемый раствор. Изобретение позволяет обеспечивать иммунную защиту против инфекции вирусом папилломы. 2 с. и 2 з.п.ф-лы, 5 ил.
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |
| Наклонный подъемник | 1976 |
|
SU586076A1 |
