Настоящая заявка относится к рекомбинантному белку, несущему эпитопы вируса папилломы, встроенные в белок аденилатциклазу или его фрагмент.
Соответственно, данное изобретение относится к рекомбинантным белкам, в которых белок аденилатциклаза (CyaA) действует как белковый вектор, для формирования иммунного ответа на эпитопы, полученные из антигенов вируса папилломы (папилломавируса), в частности - из антигенов вируса папилломы человека.
Изобретение относится, в частности, к применению полученных таким образом белковых векторов для доставки эпитопов в эукариотические клетки, предпочтительно - в клетки млекопитающих, в частности - в клетки человека.
Данное изобретение также относится к полинуклеотидам, кодирующим рекомбинантный белок согласно данному изобретению, векторам, содержащим указанные полинуклеотиды, а также клетками-хозяевами, содержащим указанные полинуклеотиды или векторы.
Изобретение относится также к способам применения упоминавшегося выше рекомбинантного белка или полинуклеотидов для лечения или профилактики ВПЧ-инфекции (инфекции, вызванной вирусом папилломы человека) у хозяина, а также для лечения или профилактики злокачественных эффектов, возникающих при инфицировании вирусом папилломы человека, у хозяина, в частности у хозяина, представляющего собой млекопитающее. В конкретном способе реализации, данное изобретение обеспечивает средство, пригодное для создания соединений, подходящих для иммунотерапии, в частности - иммунотерапии, направленной против опухоль-специфических антигенов вирусов папилломы (папилломавирусов).
Некоторые из многочисленных типов вирусов папилломы человека (ВПЧ, HPV), определяемые как высокоопасные, связаны с развитием злокачественных заболеваний эпителиальных тканей при персистировании инфекции в организме хозяина: (1) Рак шейки матки, второе по распространенности в мире опухолевое гинекологическое заболевание, (1) ассоциирован (>99%), в основном, с выявлением ДНК вирусов ВПЧ16 и ВПЧ18 (2). Онкогенный потенциал этих вирусов относят на счет продуктов экспрессии ранних генов, т.е. генов E6 и E7, экспрессия которых определяется на протяжении всего цикла репликации вируса и которая необходима для начала и поддержания злокачественной трансформации. Однако высокая частота аногенитальных инфекций, вызываемых этими высокоопасными типами ВПЧ (2) контрастирует с низкой долей лиц, у которых в конце концов развиваются злокачественные заболевания, ассоциированные с ВПЧ, что позволяет сделать предположение о том, что инфекции, вызываемые высокоопасными типами ВПЧ сдерживаются посредством иммунного ответа. Данное предположение подтверждают такие наблюдения, как: спонтанное обратное развитие (регрессия) большинства предраковых поражений (1), инфильтрация регрессирующих генитальных бородавок CD4+ Т-лимфоцитами и макрофагами (1), а также повышенное число инфицированных лиц, выявленных среди пациентов с иммуносупрессией или иммунодефицитом (1). Кроме того, в крови пациентов, у которых были диагностированы злокачественные заболевания, ассоциированные с ВПЧ 16-, также как в крови здоровых лиц (9, 10), выявили ответы CD4+ и CD8+ Т-лимфоцитов против эпитопов Е6 и/или Е7 ВПЧ16. Эти факторы в совокупности создают предпосылки для разработки иммунотерапии, направленной на белки Е6 и/или Е7 вируса ВПЧ16.
Было разработано много стратегий вакцинирования для предотвращения роста опухолей из HPV16 -Е6 и -Е7-положительных линий клеток опухоли у мышей C57BL/6 путем выработки иммунных ответов на фрагмент эпитопа H-2Db HPV16-E749-57. В этих подходах к вакцинации использовали плазмидную ДНК, вирусные или бактериальные векторы, химерные вирусоподобные частицы, синтетические пептиды и рекомбинантные белки (11). К сожалению, в отношении клинической регрессии, эти подходы дали слабо удовлетворительные результаты (3). Поэтому все еще представляет интерес разработка новых средств для обеспечения взаимодействия эпитопов ВПЧ с иммунной системой с целью стимулирования клеточного (иммунного) ответа.
Таким образом, существует потребность в новых векторах, пригодных для доставки эпитопов антигенов ВПЧ к клеткам-мишеням в условиях, подходящих для возникновения гуморального и/или клеточного иммунного ответа на указанные антигены организме хозяина. Авторы настоящего изобртения обнаружили, что белок аденилатциклаза может представлять интерес для создания такого вектора. Были сделаны разнообразные наблюдения с использованием белка аденилатциклазы бактерии Bordetella pertussis. Эти наблюдения привели к заключению, что этот белок может быть подходящей основой для создания эффективного вектора.
Аденилатциклаза (CyaA) бактерии Bordetella pertussis обладает способностью доставлять свой каталитический домен в цитозоль эукариотических клеток (12). Следовательно, эпитопы CD4+ и CD8+ Т-клеток, встроенные в каталитический сайт аденилатциклазы процессируются и презентируются белками основного комплекса гистосовместимости классов I и II, соответственно, на поверхность антиген-презентирующих клеток (АПК) (13). Кроме того, недавно было показано, что аденилатциклаза специфическим образом связывается с интегрином αмβ2 (CD11b/CD18) (14, WO 02/22169) и, следовательно, нацеливает эти эпитопы Т-клеток на CD11b+ субпопуляцию отросчатых клеток (15). Иммунизация мышей рекомбинантной аденилатциклазой, несущей соответствующие эпитопы стимулирует мощный ответ со стороны цитотоксических Т-лимфоцитов (ЦТЛ), полный иммунитет к летальному инфицированию вирусом и эффективный профилактический и терапевтический противоопухолевый иммунитет (16, 17). Белок аденилатциклаза (CyaA) и его приготовление по технологии рекомбинантных ДНК описаны, например - в международных заявках WO 93/21324 и WO 02/22169. В WO 02/22169 описано, что фрагменты аденилатциклазы, содержащие остатки с 373 по 1706 содержат структуру, необходимую для взаимодействия с рецептором CD11b/CD18.
Позднее было более конкретно описано, что последовательность аминокислот, протяженностью от остатка 1166 до остатка аминокислоты 1281, содержит детерминанту взаимодействия с рецептором CD11b/CD18, а более точно - что последовательность аминокислот, от остатка 1208 до остатка 1243, ответственна за взаимодействие этого токсина с CD11b/CD18 (Евразийские патенты ЕР 03291486.3 и 45).
Авторы настоящего изобретения определили и исследовали условия создания рекомбинантного белка CyaA, несущего, т.е. содержащего, эпитопы антигенов ВПЧ, который может доставлять указанные эпитопы в клетки-мишени, в особенности - к антиген-презентирующим клеткам хозяина, в том числе хозяина, страдающего ВПЧ-инфекцией и ее злокачественными трансформациями.
Соответственно, данное изобретение относится, в частности, к рекомбинантному белку, содержащему один или несколько полипептидов, несущих один или несколько эпитопов одного или нескольких антигенов ВПЧ, причем указанные полипептиды встроены в один или разные подходящие сайты белка аденилатциклазы (CyaA) или его фрагмента, причем указанный фрагмент аденилатциклазы сохраняет свойство белка аденилатциклазы взаимодействовать с клетками-мишенями CyaA, такими как антиген-презентирующие клетки, в частности клетками, имеющими фенотип CD11b/CD18, такими как отросчатые клетки. В конкретном способе реализации указанный фрагмент также сохраняет свойство CyaA обеспечивать перемещение встроенного в нее эпитопа или полипептида, содержащего указанный эпитоп, в цитозоль клетки-мишени. Перемещение эпитопа или полипептида, содержащего указанный эпитоп, становится возможным, если фрагмент CyaA содержит домен, который обеспечивает перемещение каталитического домена этого белка.
Рекомбинантный белок согласно настоящему изобретению можно приготовить, с использованием технологии рекомбинантных ДНК. Его можно также получить путем синтеза, в частности, путем химического синтеза. Следовательно, термин «рекомбинантный белок» относится к химерной форме белка.
Способность рекомбинантного белка взаимодействовать с CD11b/CD18 клетками можно оценить, в частности, способами, раскрытыми в ЕР 03291486.3 и (45) или в WO 02/22169. Кроме того, способность рекомбинантного белка перемещать эпитоп или полипептид, содержащий указанный эпитоп, в цитозоль клетки-мишени можно оценить при помощи способа, описанного в WO 02/22169.
В конкретном способе реализации фрагмент аденилатциклазы может состоять из двух разных частей белка CyaA, которые не представляют собой неперерывный фрагмент природного CyaA. В качестве примера можно привести каталитический домен CyaA, т.е. 400 остатков аминокислот N-концевой части CyaA, и фрагмент, содержащий остатки аминокислот 1208-1243, необходимые для взаимодействия с CD11b/CD18 антиген-презентирующими клетками.
В приведенном выше определении выражение «полипептид» описывает любую последовательность аминокислот, включая последовательности аминокислот, которые содержат посттрансляционные модификации, в частности, последовательность аминокислот, содержащую по меньшей мере шесть остатков аминокислот, и включая последовательность аминокислот, содержащую, например, от 5 до 500 остатков, или от приблизительно 5 до приблизительно 100, или от приблизительно 5 до приблизительно 200, или от приблизительно 10 до приблизительно 50 остатков, или от приблизительно 30, или приблизительно 50 до приблизительно 200 остатков, от приблизительно 100 до приблизительно 210, или от приблизительно 100 до приблизительно 200 остатков, при условии, что указанная последовательность аминокислот содержит по меньшей мере один эпитоп, т.е. последовательность аминокислот, которая может вызвать иммунный ответ при ее доставке к клетке-мишени, предпочтительно, в организме хозяина, в частности хозяина, представляющего собой млекопитающее. Следовательно, полипептиды согласно этому определению могут представлять собой просто эпитопы и даже - единственный эпитоп, либо могут содержать несколько разных или идентичных эпитопов, либо могут включать также полноразмерные антигены патогенов, т.е. вируса папилломы человека. В рамках настоящего изобретения эпитопы охватывают последовательности аминокислот, которые участвуют в гуморальном иммунном ответе и/или клеточном иммунном ответе, в частности в Т-клеточном иммунном ответе. Соответственно, эпитопы в составе полипептидов рекомбинантных молекул согласно настоящему изобретению включают эпитопы, процессируемые антиген-презентирующими клетками в организме хозяина, в частности эпитопы, распознаваемые в комплексе с молекулами ГКГС I (главным комплексом гистосовместимости первого типа), такие как эпитопы, направленные на CD8+ Т-лимфоциты, или эпитопы, распознаваемые в комплексе с молекулами ГКГС II, такие как эпитопы, направленные на CD4+ Т-лимфоциты.
В конкретном способе реализации несущий эпитопы полипептид содержит несколько эпитопов, полученных из разных антигенов, в частности из антигенов одного типа разных штаммов ВПЧ или из антигенов нескольких типов разных штаммов ВПЧ. Следовательно, полипептид, полученный из антигенов ВПЧ, может быть мультивалентным, в частности, бивалентным или тривалентным, т.е. запускающим иммунный ответ против нескольких антигенов.
Согласно настоящему изобретению антигены ВПЧ, из которых можно создать полипептиды, несущие один или несколько эпитопов, предпочтительно получены из белков, являющихся основными участниками возникновения и/или поддержания злокачественных последствий ВПЧ-инфекции, и включают так называемые опухолевые антигены, т.е. антигены, ассоциированные с развитием опухоли, связанной с ВПЧ-инфекцией, которые могут вызывать иммунный ответ в организме хозяина и реагировать специфическим образом с антителами или Т-лимфоцитами в организме хозяина.
Несущие эпитопы полипептиды согласно настоящему изобретению можно получить из нативных или зрелых антигенов ВПЧ, в том числе путем использования целого антигена или путем выбора фрагментов, в частности антигенных фрагментов, в частности эпитопов указанных антигенов, вместо использования целого белка, или путем модификации указанного антигена или отдельных его антигенных частей или эпитопов, в частности, с целью улучшения их способности стимулировать или вызывать иммунный ответ в организме хозяина совместно с белком CyaA в составе рекомбинантной молекулы. Соответственно, эти полипептиды включают эпитопы, которые фланкированы (окружены) последовательностями антигена, из которого они получены, причем указанные последовательности в природе могут быть фланкирующими или нефланкирующими, а также включают эпитопы или последовательности аминокислот, которые были химически модифицированы с целью улучшения их иммунных свойств, что иллюстрирует разнообразных возможных модификаций таких эпитопов. Эти модификации могут быть полезны для улучшения эффективности получаемых полипептидов в комплексе с белком CyaA.
Некоторые конкретные модификации описаны ниже в качестве примеров, в том числе - модификации, включающие изменение заряда полипептидов, в частности, путем вставки дополнительных положительно заряженных остатков аминокислот.
Соответственно к полипептидам согласно настоящему изобретению относятся также полусинтетические и синтетические полипептиды.
Согласно конкретному способу реализации полипептиды, полученные из антигенов ВПЧ, содержат, каждый в отдельности или совместно, приблизительно от 5 до 500, или приблизительно от 5 до 100, или приблизительно от 5 до 200, например, приблизительно от 10 до 50 остатков аминокислот, или приблизительно от 30 или приблизительно 50 до 200 остатков аминокислот или от приблизительно 100 до приблизительно 210, или от приблизительно 100 до приблизительно 200 остатков аминокислот.
Полипептид(ы) специально выбирают таким образом, чтобы он(и) вызывал(и) антиген-специфический ответ в составе рекомбинантного белка согласно настоящему изобретению.
Рекомбинантный белок согласно настоящему изобретению можно специально сконструировать таким образом, чтобы он содержал полипептид или несколько полипептидов, состоящих из измененного (нарушенного) нативного антигена ВПЧ, причем указанное нарушение состоит в делеции одного или нескольких остатков аминокислот кислотной области указанного антигена ВПЧ и/или во добавлении (инсерции) по меньшей мере двух полипептидных фрагментов указанного антигена ВПЧ в по меньшей мере два подходящих (пермиссивных) сайта аденилатциклазы.
В частности, изменение (нарушение), подпадающее под это определение, осуществляют путем вставки по меньшей мере двух фрагментов нативного антигена ВПЧ в по меньшей мере два пермиссивных сайта аденилатциклазы. Причем взаимное положение таких фрагментов обращено относительно их естественного положения в нативном антигене, т.е. фрагмент, который в нативном антигене располагается ближе к N-концу при вставке в CyaA или ее фрагмент располагается ближе к С-концу и наоборот.
Согласно наблюдениям изменение взаимного положения N-концевого и С-концевого фрагментов может увеличить эффективность стимулирования сильного и продолжительного защитного иммунитета, в частности, при иммунотерапии рака, что было показано при использовании фрагментов Е7Δ (т.е. фрагментов антигена Е7).
Согласно данному изобретению аденилатциклазу (CyaA) применяют в форме полноразмерного белка или его фрагмента, как описано выше.
Преимуществом является то, что белок CyaA или его фрагмент представляют собой белок (или его фрагмент), который является продуктом экспрессии двух генов cyaA и cyaC в клетке, в частности, в рекомбинантной клетке. В самом деле, было показано, что для того, чтобы CyaA обладал свойствами, позволяющими ему проникать в клетки-мишени (способностью к инвазии), необходимо, чтобы он претерпел посттрансляционные модификации, которые возможны только при экспрессии обоих генов: cyaA и cyaC (WO 93/21324).
В одном из способов реализации настоящего изобретения фрагменты белка CyaA представляют собой фрагменты, содержащие по меньшей мере 30 остатков аминокислот, и которые могут содержать до приблизительно 1300, в частности, до приблизительно 500 остатков аминокислот, предпочтительно приблизительно от 50 до 150 остатков аминокислот, причем в одном из вариантов реализации фрагменты содержат остатки аминокислот с 1166 по 1281 белка CyaA, или остатки аминокислот с 1208 по 1243 белка CyaA, для взаимодействия с клетками-мишенями, несущими CD11b/CD18. Таким образом, конкретный фрагмент включает всю С-концевую нативного белка или ее часть, ответственную за связывание этого белка с мембраной клетки-мишени и/или рецептором CD11b/CD18, а также за последующую доставку эпитопа, содержащегося в полипептиде/ах (или эпитопов, содрежащихся в полипептиде/ах) в цитозоль. (12). Фрагмент белка CyaA согласно настоящему изобретению содержит остатки аминокислот с 372 по 1706 белка CyaA. Другой вариант фрагмента представляет собой фрагмент, соответствующий белку CyaA, из которого удалили остатки аминокислот с 225 по 234, получив, таким образом, фрагмент CyaA, содержащий остатки с 1 по 224 и с 235 по 1706.
В конкретном способе реализации настоящего изобретения белок аденилатциклаза представляет собой бактериальный белок. В предпочтительном способе реализации белок CyaA получают из одного из видов Bordetella.
Согласно настоящему изобретению одним из видов Bordetella, представляющих интерес, является Bordetella pertussis. Другие представляющие интерес штаммы Bordetella: Bordetella parapertussis или Bordetella bronchiseptica. Последовательности белка CyaA В. parapertussis описаны, в частности под номером NC 002928.3 (как последовательность из 1740 аминокислот) и в Parkhill J. et al (Nat. Genet. DOI, 10 (2003), a последовательности В. bronchiseptica - в Betsou F. et al (Gene 1995, August 30; 162(1): 165-6).
Bordetella pertussis является возбудителем коклюша. Среди нескольких других секретируемых им токсинов - хорошо известный коклюшный токсин и (токсин) аденилатциклазу (CyaA), который является ключевым фактором вирулентности этой бактерии и представляет собой один из антигенов, защищающих от инфекции, вызываемой В. pertussis.
Белок аденилатциклаза бактерии Bordetella pertussis представляет собой токсин, описанный как бифункциональный белок, состоящий из 1706 остатков, содержащий N-концевой каталитический домен из 400 остатков аминокислот и С-концевую часть из 1306 остатков, которая ответственна за связывание этого токсина с мембраной клетки-мищени и последующую доставку каталитического фрагмента в цитозоль клетки (12).
Белок CyaA синтезируют в форме неактивного протоксина, который превращается в активный токсин в ходе посттрансляционного пальмитоилирования двух внутренних остатков лизина (лизины 860 и 983). Для этой посттрансляционной модификации необходима экспрессия вспомагательного гена, а именно гена cyaC, который расположен вблизи гена cyaA в хромосоме В. pertussis.
Аденилатциклаза Bordetella pertussis описана в виде последовательности аминокислот и в виде последовательности нуклеотидов в Glaser, P. et al, 1988, Molecular Microbiology 2(1), 19-30. Соответственно, когда в описании настоящего изобретения упоминаются последовательности остатков аминокислот или последовательности нуклеотидов белка CyaA бактерии В. pertussis, их позиции даны в соответствии с последовательностями, раскрытыми в указанной публикации Glasser et al. 1988.
В рекомбинантном белке согласно настоящему изобретению полипептиды, несущие один или несколько эпитопов одного или нескольких антигенов ВПЧ встроены в один или несколько подходящих (пермиссивных) сайтов белка CyaA.
В настоящем изобретении "пермиссивный сайт" обозначает сайт или последовательность белка CyaA, в который можно встроить (вставить) полипептид таким образом, что это встраивание не отразится существенно на функциональных свойствах белка CyaA, в частности - на его способности взаимодействовать с клетками, и конкретно - на направленном взаимодействии (нацеливании) аденилатциклазы с антиген-презантирующими клетками, в том числе на специфическом связывании рецептора CD11b-CD18, и, предпочтительно, не затрагивает домены этого белка, задействованные в процессе перемещения эпитопа (эпитопов) в клетку-мишень.
Пермиссивные сайты аденилатциклазы Bordetella pertussis, обеспечивающие перемещение каталитического домена аденилатциклазы и, следовательно, перемещение эпитопов, встроенных в такие пермиссивные сайты включают, без ограничения, остатки 137-138 (Val-Ala), остатки 224-225 (Arg-Ala), остатки 228-229 (Glu-Ala), остатки 235-236 (Arg-Glu) и остатки 317-318 (Ser-Ala) ((44) Sebo et al., 1995). Следующие дополнительные пермиссивные сайты также входят в способы реализации настоящего изобретения: остатки 107-108 (Gly-His), остатки 132-133 (Met-Ala), остатки 232-233 (Gly-Leu), а также 335-336 (Gly-Gln) и 336-337. (43)
Для других видов Bordetella соответствующие пермиссивные сайты можно определить путем сравнения последовательностей и выявления соответствующих остатков.
Согласно другому способу реализации полипептид можно встраивать дополнительно, или в качестве альтернативы, на одном и/или другом конце белка CyaA или его фрагмента.
Конкретные фрагменты белков CyaA для применения в целях настоящего изобретения представляют собой фрагменты длиной до 1300 аминокислот или приблизительно от 30 до 500 остатков аминокислот, предпочтительно, приблизительно от 50 до 150 остатков аминокислот, в частности такие фрагменты, включающие остатки аминокислот с 1166 по 1281 нативного белка CyaA, предпочтительно, остатки с 1208 по 1243 нативного белка CyaA.
Следовательно, согласно настоящему изобретению «встраивание» (вставка) полипептида в белок CyaA с целью получить так называемый рекомбинантный белок, называемый также «гибридным белком», включает генетическое встраивание, в частности, при помощи доступной ДНК-технологии. В качестве альтернативы "встраивание" включает также встраивание негентическими способами, включая химическое встраивание, например, ковалентное присоединение к одному из концов аденилатциклазы или ее фрагмента, или нековалентное присоединение. Негенетическое встраивание может, в частности, предоставлять интерес в случае, когда полипептид, который нужно ввести, является синтетическим или полусинтетическим. Способы присоединения лекарственного средства к полипептиду хорошо известны в данной области и включают, например, присоединение дисульфидной связью при помощи N-пиридилсульфонил-активированного сульфгидрила.
В частности, возможно присоединять молекулы, в том числе молекулы, содержащие полипептиды согласно настоящему изобретению, к аденилатциклазе посредством химической связи или генетической вставки, с целью нацеливания in vivo на клетки-мишени аденилатциклазы, такие как антиген-презентирующие клетки, например клетки, несущие CD11b/CD18, и в частности, в цитозоль указанных клеток. В самом деле, было обнаружено, что при присоединении молекулы, соответствующей данному CD8+ Т-клеточному эпитопу, к каталитическому домену обезвреженной аденилатциклазы либо дисульфидной связью, либо при помощи генетической вставки, эта рекомбинантная молекула способна вызывать in vivo специфический ответ цитотоксических Т-лимфоцитов, демонстрируя таким образом, что указанный эпитоп CD8+ Т-лимфоцита перемещается в цитозоль клеток, экспрессирующих CD11b.
В конкретном способе реализации рекомбинантная аденилатциклаза, применяемая для изготовления вектора на основе белка, представляет собой аденилатциклазу или ее фрагмент, в частности, модифицированные путем встраивания остатков цистеина, содержащих одну или более молекул, включающих, в частности, полипептиды согласно настоящему изобретению, химически связанные посредством дисульфидной связи со встроенными генетическими методами остатками (остатком) цистеина, расположенными в пределах каталитического домена указанной аденилатциклазы.
В самом деле, множество молекул, в ососбенности - содержащих полипептиды согласно настоящему изобретению, можно химически присоединить к аденилциклазе посредством образования дисульфидной связи с различными остатками цистеина, расположенными в различных пермиссивных сайтах в пределах каталитического домена.
С намерением предложить рекомбинантный белок, подходящий для создания препаратов, обладающих способностью вызывать иммунный ответ, в особенности клеточный иммунный ответ, в организме хозяина, и в частности - для создания препаратов, способных вызывать клеточный иммунный ответ, направленный против злокачественных эффектов, наблюдаемых в организме хозяина, инфицированного ВПЧ, авторы настоящего изобретения предложили получать полипептиды, несущие эпитопы, из высоко онкогенных штаммов ВПЧ, в особенности из антигенов штаммов, выбранных среди HPV16, HPV18, HPV31, HPV33, HPV35, HPV45, HPV52 и HPV58.
Среди этих штаммов особый интерес представляют штаммы HPV18 и HPV16. В частности, HPV16 благодаря своей связи с развитием рака шейки матки у млекопитающих, в частности у людей, представляет особый интерес в качестве мишени для лечения животного - хозяина, инфицированного HPV.
Приняв за основу эти штаммы ВПЧ, изобретатели предлагают получать полипептиды, несущие эпитопы из антигенов, выбранные среди белков L1, L2, Е1, Е2, Е4 и Е5.
В качестве альтернативы или дополнительно изобретатели предлагают также получать такие полипептиды, несущие эпитопы из белков Е6 или Е7 ВПЧ.
В конкретном способе реализации данного изобретения для создания полипептидов, несущих эпитопы применяют белки Е6 или Е7 ВПЧ16, либо белки Е6 или Е7 из ВПЧ18.
Конкретный белок ВПЧ, который можно использовать для создания полипептида, полученного из антигенов ВПЧ, - белок Е7 ВПЧ, в частности, ВПЧ16 или ВПЧ18. Согласно одному из способов реализации данного изобретения при получении полипептида используют несколько белков Е7 различных штаммов ВПЧ, в частности HPV16 и HPV18. Например, полипептид представляет собой полноразмерный белок Е7 вируса ВПЧ16 и HPV18, либо один или несколько фрагментов каждого из белков Е7 вирусов ВПЧ16 и ВПЧ18, включая мультимеры, в особенности димеры указанных фрагментов.
Данные белки ВПЧ, а также последовательности аминокислот и нуклеотидов этих белков раскрыты в Seedorf, К. et al (Human papillomavirus type 16 DNA sequence (Последовательность ДНК вируса папилломы человека 16 типа). Virology, 145: 181-185, 1985) для ВПЧ16, Cole S.T., Danos O. (Nucleotide sequence and comparative analysis of the human papillomavirus type 18 genome. Phylogeny of papillomaviruses and repeated structure of the E6 and E7 gene products (Последовательность нуклеотидов и сравнительный анализ генома вируса папилломы человека 18 типа. Филогенетика папилломавирусов и повторяющаяся структура продуктов генов E6 и E7). J. Mol. Biol. 193: 599-606 (1987)) или в Fernando GJ. et al (T-helper epitopes of the E7 transforming protein of cervical cancer associated human papillomavirus type 18 (HPV18) (Эпитопы Т-хелперов трансформирующего белка ассоциированного с раком шейки матки папилломавируса человека 18 типа (ВПЧ18) рака шейки матки) Virus Res. 1995 Апрель. 36(1): 1-13).
Белки E6 и E7 представляют собой онкобелки, экспрессируемые, в частности, вирусами ВПЧ16 или ВПЧ18 на протяжении всего цикла репликации вируса. Было показано, что они необходимы для начала и поддержания злокачественной трансформации клеток хозяина, следующей за инфицированием штаммом ВПЧ. Следовательно, оба эти опухоль-специфических антигена рассматриваются как возможные мишени для адаптивной иммунотерапии, опосредованной цитотоксическими Т-лимфоцитами.
Согласно конкретному способу реализации данного изобретения рекомбинантный белок содержит множество полипептидов, каждый из которых несет один или несколько эпитопов одного или нескольких антигенов ВПЧ.
Например, такие множественные полипептиды можно получить из белков E6 и E7 одного штамма HPV, в частности HPV16 или HPV18. Согласно другому примеру, такие множественные полипептиды могут содержать эпитопы, полученные из белков E6 или E7, из обоих вирусов HPV16 и HPV18.
Множественные полипептиды могут также состоять из разных эпитоп-определяющих фрагментов одного белка, например, белка E7 или E6, которые встроены в разные пермиссивные сайты представляющего интерес белка CyaA.
Другой конкретный рекомбинантный белок, соответствующий приведенным выше определениям, представляет собой рекомбинантный белок CyaA, в котором множественные полипептиды, несущие эпитопы, включают (содержат) фрагмент, содержащий остатки с 1 по 29 или фрагмент, состоящий из остатков с 1 по 29, либо фрагмент, содержащий остатки с 42 по 98, или фрагмент, состоящий из остатков с 42 по 98, белка Е7 вируса ВПЧ16, либо множественные полипептиды, состоящие из, или содержащие оба фрагмента, встроенные в разные пермиссивные сайты белка CyaA.
Другой рекомбинантный белок согласно данному изобретению представляет собой белок, указанные множественные полипептиды в котором включают фрагмент, который имеет последовательность аминокислот RAHYNIVTF (E749-57) и/или GQAEPDRAHYNIVTFCCKCDSTLRLCVQSTHVDIR (Е743-77).
Согласно наблюдениям число остатков аминокислот в полипептидах, встраиваемых в пермиссивные сайты белка CyaA, таково, что оно позволяет встраивать в белок CyaA или его фрагмент полипептиды, состоящие из полноразмерных антигенов, в частности полноразмерных белков Е6 или Е7 вируса папилломы человека.
Согласно конкретному способу реализации данного изобретения полипептид, всоставе рекомбинантной аденилатциклазы, представляет собой белок Е7, конкретно - белок Е7 вируса ВПЧ16, встроенный между кодонами 224 и 235 CyaA или между кодонами 319 и 320 CyaA.
В другом способе реализации рекомбинантный белок согласно настоящему изобретению содержит множественные полипептиды, некоторые из которых представляют собой полипептиды, несущие эпитоп или несколько эпитопов одного или нескольких ВПЧ, а другие полипептиды несут эпитопы других патогенов.
В другом частном способе реализации рекомбинантный белок согласно настоящему изобретению дополнительно содержит один или несколько эпитопов, полученных из разных патогенных агентов. Особый интерес может представлять объединение эпитопов, полученных из Chlamydia или из ретровируса ВИЧ или вируса папилломы человека, вируса гепатита В, вируса гепатита С, аденовируса Эпштейна-Барр, вируса герпеса, вируса Т-клеточной лимфомы человека 1 (HTLV.1) и цитомегаловируса с эпитопами, полученными из вируса папилломы человека.
Согласно другому частному способу реализации данного изобретения нативная последовательность аминокислот полипептидов, несущих эпитопы, была модифицирована, например, таким образом, чтобы уменьшить число отрицательно заряженных остатков аминокислот в последовательности. Такой модификации можно достичь путем удаления некоторых отрицательно заряженных остатков аминокислот или путем добавления нескольких положительно зараженных остатков аминокислот, например, в виде фланкирующих остатков эпитопов. Полипептиды, содержащие вследствие такой модификации меньшее число отрицательно заряженных остатков, могут обладать преимуществом при перемещении каталитического домена белка CyaA в цитозоль клеток-мишеней.
Полипептиды, несущие эпитопы, можно также сконструировать таким образом, чтобы при встраивании в аденилатциклазу они находились в «развернутом» (не сложенном) состоянии, что повышает эффективность внедрения рекомбинантного белка CyaA в клетки-мишени. Такого развертывания полипетидов, сворачиваение которых является следствием их аминокислотного состава, можно достичь, например, путем удаления или замещения остатков цистеина, что позволяет предотвратить образование дисульфидных связей, которые могут быть задействованы в свертывании полипептидов. В некоторых случаях можно предотвратить сворачивание полипептидов, готовя их в присутствии восстанавливающих агентов, что позволяет избежать повторного свертывания in vivo.
В отдельном способе реализации полипептиды, несущие эпитопы, могут представлять собой скрытые (криптические) эпитопы.
В отдельном аспекте данного изобретения, изобретатели фактически установили, что химерные белковые конструкции, полученные из рекомбинантных белков, которые содержат (i) аденилатциклазу (CyaA) или ее фрагмент согласно раскрытым выше определениям и (ii) полипептид, несущий один или несколько антигенных фрагментов одного или нескольких антигенов, обеспечивают иммуногенность скрытых эпитопов указанных антигенов как результат их презентации в составе рекомбинантной конструкции. В частности, указанные химерные конструкции, содержащие CyaA или ее фрагмент, охарактеризованные в настоящем изобретении, и полипептиды, полученные из антигенов, представляющих интерес для специфических криптических эпитопов антигена, которые могут стать иммуногенными, в частности, вызвать Т-клеточный ответ, в особенности, цитотоксических Т-лимфоцитов, в организме хозяина.
Таким образом, данное изобретение относится также к рекомбинантному белку, содержащему один или несколько полипептидов, несущих один или несколько эпитопов одного или нескольких антигенов, причем указанный полипептид (полипептиды) встроены в один и тот же или в разные пермиссивные сайты аденилатциклазы (белка CyaA) или его фрагмента, где указанный фрагмент аденилатциклазы сохраняет свойство указанного белка аденилатциклазы направленно взаимодействовать с антиген-презентирующими клетками, где по меньшей мере один из указанных эпитопов представляет собой субдоминантный криптический Т-клеточный эпитоп и где указанный рекомбинантный белок способен вызвать антиген-специфический ответ против указанного полипептида (полипептидов).
В частности, скрытые эпитопы происходят из антигена ВПЧ, в том числе из антигенов ВПЧ16 или ВПЧ18, в частности, входят в состав антигена Е7.
Определенный таким образом рекомбинантный белок, в частности, может содержать пептид, полученный из белка Е7 вируса ВПЧ18, т.е. пептид, который имеет последовательность аминокислот IDGVNHQHL.
Согласно одному из способов реализации настоящего изобретения криптический эпитоп может быть модифицирован, например, он может содержать замены в первых двух позициях и может иметь, например, аминокислотную последовательность: ASGVNHQHL.
Изобретение, в частности, относится к пептидам IDGVNHQHL.
Изобретение также относится к пептидам, которые содержат замены в этой последовательности, в частности в позициях 1 и/или 2, в частности к пептидам, которые имеют последовательность ASGVNHQHL.
Изобретение также охватывает варианты указанных пептидов, при условии, что они сохраняют иммуногенные свойства, в частности, если они способны вызывать Т-клеточный ответ, в частности ответ цитотоксических Т-лимфоцитов.
Для приготовления рекомбинантного белка согласно настоящему изобретению предпочтительно инактивировать ферментативную активность белка CyaA, т.е. его способность превращать АТФ в цАМФ. Такая инактивации может быть результатом генетической модификации. Например, генетический инактивации можно достичь, при встраивании дипептида в участок последовательности аденилатциклазы, который является частью каталитического сайта (например, между остатками 188 и 189). Некоторые варианты таких инактивированных белков CyaA приведены в последующих примерах.
Рекомбинантный белок согласно настоящему изобретению предпочтительно способен вызывать опосредуемый клетками иммунный ответ (клеточный иммунный ответ). Такой ответ включает активацию цитотоксических Т-лимфоцитов и Т-хелперов (Th-клеток), в частности ответ Th1, включая ответ CD4+ Т лимфоцитов и/или CD8+ Т-лимфоцитов.
В частности, было показано, что способность рекомбинантного белка вызывать клеточный иммунный ответ достаточна для предотвращения роста опухоли in vivo, и даже для того, чтобы обеспечить регрессию опухоли у животного.
Изобретение также относится к полинуклеотиду, который кодирует описанный выше рекомбинантный белок.
Полинуклеотид согласно настоящему изобретению можно встроить в вектор экспрессии, чтобы получить рекомбинантный вектор, подходящий для экспрессии рекомбинантного белка согласно данному изобретению. Такие векторы экспрессии включают плазмиды, космиды, фагмиды и вирусные векторы.
Рекомбинантный вектор может представлять собой вектор, подходящий для экспрессии в прокариотических клетках, в частности в бактериях, или может представлять собой вектор экспрессии, подходящий для экспрессии в эукариотических клетках, в частности в клетках млекопитающих, и предпочтительно - в клетках человека.
Изобретение относится, в частности, к векторам, состоящим из плазмид, кодирующих рекомбинантный белок согласно настоящему изобретению, таким как:
pTRACE5-HPV16E7full(полноразмерный) (также обозначаемый CyaAE5-HPV16E7FULL), помещенный в Национальную коллекцию культур микроорганизмов CNCM (Париж, Франция) 18 Марта 2004 г. под номером CNCM I-3191; pTRACE5-HPV16E7Δ30-42, (также обозначаемый CyaAE5-HPV16E7Δ30-42), помещенный в Национальную коллекцию культур микроорганизмов CNCM (Париж, Франция) 18 Марта 2004 г. под номером CNCM I-3190, или конструкция pTRACE5-HPV16E749-57.
Изобретение также включает клетку-хозяина, в частности - прокариотические клетки или эукариотические клетки, например клетки млекопитающих, включая клетки человека, трансформированные полинуклеотидом или вектором согласно настоящему изобретению.
Данное изобретение относится, в частности, к клеткам, депонированным в Национальной коллекции культур микроорганизмов института Пастера (CNCM) под номером №CNCM I-3190 и номером № CNCM I-3191.
Изобретение относится также к иммуногенной композиции, которая содержит в качестве активного компонента описанный выше рекомбинантный белок или описанный выше вектор экспрессии. Указанный активный компонент иммуногенной композиции можно приготовить в комплексе с физиологически приемлемой средой, приемлемым наполнителем, носителем или разбавителем, или их комбинацией, подходящими для введения в организм хозяина.
Иммуногенную композицию предпочтительно разрабатывают таким образом, чтобы она стимулировала клеточный иммунный ответ, в частности иммунный ответ, опосредуемый Т-лимфоцитами, в организме хозяина-млекопитающего. Предпочтительно, она способна стимулировать опосредуемый клетками цитолитический иммунный ответ цитотоксических Т-лимфоцитов, в частности CD8+.
Другая иммуногенная композиция согласно настоящему изобретению представляет собой композицию, способную вызывать гуморальный клеточный ответ.
Для того чтобы повысить способность иммунной композиции согласно настоящему изобретению стимулировать иммунный ответ, можно комбинировать активный компонент с адъювантом и/или поверхностно-активным веществом и/или иммуномодулирующими веществами (такими как цитокины и хемокины).
Адъюванты включают, например, липосомы, масляные фазы, такие как адъюванты типа адъюванта Фрейнда, применяемые обычно в форме эмульсии с водной фазой, или содержат нерастворимые в воде неорганические соли, такие как гидроксид алюминия, сульфат цинка, фосфат кальция или хлорид кальция.
Иммуногенную композицию согласно настоящему изобретению предпочтительно применяют для стимулирования иммунного ответа в организме хозяина, инициируя и/или поддерживая (усиливая) указанный ответ, в частности, в целях иммунотерапии. В частности, иммуногенная композиция согласно настоящему изобретению может представлять интерес для предотвращения начала или продолжения злокачественной трансформации, причиной которой является инфекция организма хозяина ВПЧ, либо для лечения пациента, страдающего злокачественной трансформацией, причиной которой является инфекция организма хозяина ВПЧ, в частности инфекция ВПЧ-16 или ВПЧ-18.
Такая иммунотерапевтическая композиция может представлять особенный интерес для лечения неконтролируемой пролиферации клеток в организме хозяина, приводящей к онкологическим заболеваниям, в частности для иммунотерапии рака, конкретно для иммунотерапии рака шейки матки, ассоциированного с ВПЧ-инфекцией. Таким образом, она обеспечивает средства для создания терапевтических вакцин, подходящих, в частности, для лечения злокачественных заболеваний, вызванных онковирусными инфекциями, включая опухолевые заболевания.
В описании настоящего изобретения выражение "лечение" или "терапевтическое лечение" охватывает действия соединений, раскрытых в настоящей заявке, которые в результате оказывают благотворный эффект на пациента, проходящего лечение. Указанные эффекты можно наблюдать либо на клеточном уровне, либо на клиническом уровне, они включают улучшение состояния пациента, либо ремиссию, либо возвращение к здоровому состоянию, происходящие в результате лечения. В случае когда злокачественное заболевание, лечение которого проводят, представляет собой неконтролируемую пролиферацию клеток или персистирование опухоли, благотворный эффект может включать стабилизацию или, предпочтительно, предотвращение, остановку или обращение неконтролируемой пролиферации клеток, либо регрессию опухоли.
Описанная выше композиция, предназначенная для лечения злокачественного заболевания, может предпочтительно содержать дозу активного компонента, составляющую приблизительно от 1 до 1000 мкг рекомбинантного белка, предпочтительно приблизительно от 10 до 500 мкг рекомбинантного белка. В случае, когда композиция содержит в качестве активного компонента рекомбинантный белок согласно настоящему изобретению, доза может содержать приблизительно от 0.05 до 10 мкг рекомбинантного белка, предпочтительно, приблизительно от 0.1 до 1 мкг белка.
В зависимости от заболевания, которое подлежит лечению, данную композицию можно вводить местно на уровне пораженного участка, один или несколько раз, например, с регулярными интервалами продолжительностью несколько дней, например продолжительностью от 5 до 10 дней. Возможно также системное введение композиции.
Изобретение также относится к составу вакцины, в частности к композиции, составленной для введения млекопитающему, предпочтительно, человеку, содержащей рекомбинантный белок, соответствующий данному выше определению, или описанный выше полинуклеотид, или вектор, содержащий такой полинуклеотиотид, предпочтительно для введения человеку, и, если требуется, фармацевтически приемлемый разбавитель для стимулирования иммунного ответа, включая клеточный иммунный ответ, и/или гуморальный ответ.
Изобретение относится также к лекарственному препарату, который содержит рекомбинантный белок, или полинуклеотид, или вектор согласно настоящему изобретению и фармацевтически приемлемый разбавитель, для предотвращения или лечения ВПЧ-инфекций.
Согласно другому способу реализации лекарственный препарат содержит рекомбинантный белок, полинуклеотид или вектор согласно настоящему изобретению с фармацевтически приемлемым разбавителем, для предотвращения или лечения начала или продолжения злокачественной трансформации, причиной которой является ВПЧ-инфекция хозяина.
Лекарственный препарат, содержащий рекомбинантный белок, полинуклеотид или вектор и фармацевтически приемлемый разбавитель, для иммунотерапии рака. Изобретение также относится к применению в организме пациента рекомбинантных белков, содержащих бактериальный белок, в частности бактериальный токсин (предпочтительно, в обезвреженной форме) или его фрагмент, подходящих для применения в качестве вектора с целью получения (стимуляции) иммунного ответа, т.е. гуморального и/или клеточного иммунного ответа, в организме хозяина, причем указанный (бактериальный) белок или фрагмент белка модифицированы путем введения в них одного или нескольких эпитопов одного или нескольких антигенов одного или нескольких онковирусов, с целью лечения инфекции, вызванной одним из онковирусов. Такой рекомбинантный белок предназначен, в частности, для лечения злокачественного действия, в частности, опухолей, вызываемых такими онковирусами.
Примерами бактериальных белков, подходящих для применения в качестве векторов, несущих эпитопы антигенов онковирусов являются OmpA из бактерии Klebsiella или следующие токсины: токсин Шига (Shiga), включая его β-субединицу (Haicher N. et al J. Immunol. 2000, 165: 3301-8) токсин сибирской язвы (Goletz TJ et al, PNAS USA 1997, 94: 12059-64), дифтерийный токсин (Stenmark H. et al, J. Cell. Biol. 1991, 113: 1028-32) или Экзотоксин А псевдомонады (Donnelly JJ. et al, PNAS USA 1993, 90: 9530-4). Онковирусы, из антигенов которых можно получить эпитопы для встраивания в бактериальный белок, включают вирус папилломы человека, вирус гепатита В, вирус гепатита С, аденовирус Эпштейна-Барр, вирус герпеса, вирус Т-клеточной лейкимии человека 1 и цитомегаловирус.
Описание, приведенное выше для рекомбинантного белка CyaA или его применения в качестве активного компонента, можно адаптировать для других бактериальных белков или антигенов онковирусов.
Изобретение относится также к набору для диагностики инфекции ВПЧ или для иммунологического мониторинга этой инфекции, который включает рекомбинантный белок, полинуклеотид или вектор экспрессии согласно данному изобретению.
Изобретение также относится к применению описанного выше рекомбинантного белка, полинуклеотида или вектора согласно настоящему изобретению для лечения или предотвращения ВПЧ-инфекции у пациента.
Изобретение также относится к применению вышеописанных рекомбинантного белка, полинуклеотида или вектора согласно настоящему изобретению для иммунотерапии, направленной против начала или продолжения (применению для предотвращения или лечения) злокачественной трансформации, вызванной ВПЧ-инфекцией у пациента.
Изобретение также относится к способу in vitro диагностики или иммунологического мониторинга ВПЧ-инфекции, включающему:
- приведение в контакт Т-лимфоцитов, полученных из организма млекопитающего, в частности человека, с рекомбинантным белком согласно настоящему изобретению,
- детектирование изменения активации Т-лимфоцитов.
В частном способе реализации, рекомбинантный белок можно применять для предотвращения ВПЧ-инфекции или для лечения животного, страдающего от ВПЧ-инфекции, включая животных, у которых в результате такой инфекции развивается опухоль.
Изобретение также относится к способу скринига (обнаружения) неизвестных или субдоминантных скрытых (криптических) эпитопов Т-клеток в полипептиде, который находится в составе химерного белка: CyaA-полипептид, где CyaA представляет собой описанные выше аденилатциклазу или ее фрагмент, который включает:
- введение указанных химерных белков в организм животного-хозяина,
- определение Т-клеточного ответа, в частности ответа цитотоксических Т-лимфоцитов у указанного хозяина.
Изобретение относится, в частности, к способу скрининга (обнаружения) неизвестных или субдоминантных скрытых (криптических) эпитопов Т-клеток (в частности, эпитопов для CD8+ Т-клеток) в полипептиде (полипетидах) антигена (антигенов) ВПЧ, содержащихся в составе рекомбинантных белков, описанных в настоящем изобретении, который включает:
- введение указанных химерных белков в организм животного-хозяина (не человеку),
- определение Т-клеточного ответа у указанного животного, в частности ответа цитотоксических Т-лимфоцитов
Другие свойства, отличающие данное изобретение, раскрыты в приведенных ниже примерах и на фигурах.
Описание фигур
Фигура 1. Конструирование и очистка рекомбинантных аденилатциклаз, несущих белок Е7 вируса ВПЧ16 (HPV16-E7).
(А) Схематическая карта pTRACE5, на которой указаны имеющие в данном случае значение сайты рестрикции и всроенные последовательности. (В) Схематическое изображение аденилатциклазы, показывающее сайт встраивания (вставки) дипептида LQ для ингибирования ее ферментативной активности. Также показаны позиции вставки белка HPV16-E7. Подчеркнут рестриктированный по Н-2b эпитоп белка HPV16-E7. (С) Анализ рекомбинантных аденилатциклаз с HPV16-E7 в ПАГ с додецилсульфатом натрия. Пять микрограмм очищенных белков разделяли на полиакриламидном геле с 4-15% додецилсульфата натрия и окрашивали Кумасси синим. Полоса 1: CyaA дикого типа; полоса 2: CyaA-Е749-57; полоса 3: CyaA-E7Full(полноразмерный); полоса 4: CyaA-Е7Δ30-42. (D) Анализ рекомбинантных аденилатциклаз с HPV16-E7 методом Вестерн-блот. После очистки в полиакриламидном геле с додецилсульфатом натрия очищенные белки переносят в электрическом поле на мембрану из нитроцеллюлозы, которую затем зондируют моноклональным анти-НРУ16-Е7 антителом. Полоса 1, 2: CyaA дикого типа (2 и 0.4 мкг соответственно); полосы 3, 4 и 5: CyaA-Е749-57, CyaA-E7Full(полнор) и CyaA-Е7Δ30-42, соответственно, 0.4 мкг каждого белка.
Фигура 2. Стимуляция ответов Т-лимфоцитов рекомбинантными аденилатциклазами, несущими белок Е7 вируса ВПЧ16 (HPV16-E7 CyaA).
(А) Мышей C57BL/6 (а, b, с), ТАР1-/- (d), ГКГС класс II-/- (е) и CD40-/- (f) иммунизировали в день 0 путем внутривенного введения 50 мкг CyaA-Е749-57 (а), CyaA-E7Full; (b), или CyaA-Е7Δ30-42 (с, d, е и f). Через семь дней животных умерщвляли, а спленоциты повторно стимулировали in vitro в течение 5 дней 1 мкг/мл пептида HPV16-E743-77 в присутствии облученных сингенных спленоцитов и использовали в качестве эффектора против клеток-мишеней ТС-1 (сплошные квадраты) или EL4 (пустые квадраты). Взятые у мышей спленоциты обрабатывали CyaAE5-cysOVA, несущей не имеющий отношения к настоящему изобретению эпитоп OVA257-264, повторно стимулировали in vitro в течение 5 дней 1 мкг/мл пептида HPV16-E743-77 в присутствии облученных сингенных спленоцитов. Этот эксперимент также проиллюстрирован (а, сплошные треугольники). Лизис мишеней оценивали по высвобождению 51Cr. Данные представляют медианной значение процентных долей значений специфического лизиса (n = число мышей, указанное на каждой диаграмме) а также межквартильные размахи. (В) Детектирование HPV16-Е7-специфических ИФН-γ-продуцирующих клеток после иммунизации рекомбинантными аденилатциклазами, несущими HPV16-E7. Мышей C57BL/6 иммунизировали, как в А, конструктами CyaAE5-cysOVA (кружки), CyaA-Е749-57 (квадраты), CyaA-E7Full (треугольники), или CyaA-Е7Δ30-42 (ромбы). Чрез семь дней клетки селезенки, выделенные у иммунизированных мышей, культивировали in vitro в течение 36 часов без стимуляции (т.е. без пептида, пустые символы) или с 1 мкг/мл пептида Е749-57 (сплошные символы) в присутствии сингенных облученных спленоцитов. Данные выражены в числе пятнообразующих клеток на селезенку, представлены результаты отдельных мышей из трех независимых экспериментов для каждой группы. Горизонтальные полосы представляют медианное значение ответов каждой группы.
Фигура 3. Рекомбинантные аденилатциклазы, несущие HPV16-E7, стимулируют HPV16 Е7-специфический ответ хелперов Th1.
(А) Мышей C57BL/6 либо оставляли без лечения (квадратики), либо подвергали первичному воздействию антигена путем внутривенного введения 50 мкг CyaAE5-cysOVA (кружки), CyaA-Е749-57 (треугольники), CyaA-E7Full (перевернутые треугольники) или CyaA-Е7Δ30-42 (ромбы). Спустя семь дней клетки селезенки стимулировали in vitro 10 микрограммами/мл белка с полигистидиновой меткой His-Tag-HPV16-E7 и через 72 часа исследовали надосадочную жидкость на содержание ИФН-γ. Представлены результаты для отдельных мышей из 4 независимых экспериментов. Результаты выражены в форме концентрации высвобожденного ИФН-γ в надосадочной жидкости (из двух лунок дупликата). Из этих значений вычитали фоновые значения, измеренные в спленоцитах, которые не подвергали повторной стимуляции. Вкладка: in vitro стимуляция 1 мкг/мл пептида Е743-77. Горизонтальные линии представляют медиану ответа в каждой группе. (В) То же, что на (А), за тем исключением, что надосадочные жидкости исследовали на содержание ИЛ-5. Представлены результаты для отдельных мышей из 2 независисых экспериментов. Результаты выражены в форме концентрации высвобожденного ИЛ-5 в надосадочной жидкости (в двух лунках дупликата).
Фигура 4. Терапевтическая вакцинация рекомбинантными аденилатциклазами, несущими HPV16-E7, ликвидирует развитые опухоли.
(Эксп. А) В День 0 мышам C57BL/6 трансплантировали 5×104 опухолевых клеток ТС-1. В день 10 мышам давали одну внутривенную инъекцию CyaA-Е749-57 (С), CyaA-E7Full (D) или CyaA-Е7Δ30-42 (Е). Мышей без обработки (лечения) (А) либо мышей, которым делали инъекцию CyaAE5-cysOVA, (В) брали в качестве контроля. Мышей умерщвляли, когда размер опухоли достигал 1000 мм3 либо когда этого требовало санитарное состояние животных (некроз опухоли, быстрая потеря веса >20%), чтобы таким образом предотвратить ненужные страдания. Двух мышей, получавших CyaA-E7Full, у которых позже развились прогрессирующие опухоли (*) забили для дальнейшего исследования (см. Фиг.6).
(Эксп. В) Постановка эксперимента - такая же, как в (Эксп. А). Терапевтическую вакцинацию проводили путем инъекций в кожу ушей в дни +10 и +17. Вводили 10 мкг CyaAE5-CysOVA (а, сплошные линии) или 10 мкг CyaA-Е7Δ30-42 (b). Каждая кривая изображает рост опухоли у отдельного животного. Также представлены результаты для двух необработанных животных (а, пунктирные линии). Справа вверху каждого квадранта (а, b) указано число забитых животных по отношению к общему числу животных. Показаны кривые выживаемости этих мышей (с): необработанные (пустые треугольники), получавшие инъекции симулятора CyaAE5-CysOVA (заполненные треугольники), получившие CyaA-Е7Δ30-42 (кружки).
Фигура 5. Терапевтическая вакцинация рекомбинантными аденилатциклазами, несущими HPV16-E7, приводит к продлению сроков выживаемости. Терапевтическую вакцинацию проводили как описано для Фиг.4. После инъекции опухолевых клеток ТС-1 мышей (5-10 на группу) иммунизировали рекомбинантными аденилатциклазами, несущими HPV16-E7. Как показано на диаграммах, иммунизацию проводили в дни +1, +5 или +10. Мышей оставляли без обработки (сплошные квадраты, сплошная линия), делали симулирующие инъекции CyaAE5-cysOVA (пустые квадраты, пунктирная линия) или вводили им CyaA-Е749-57 (пустые треугольники), CyaA-E7Full (пустые кружки) или CyaA-Е7Δ30-42 (пустые ромбы). Видно, что в День +5 терапевтического эксперимента кривые выживаемости животных, получавших CyaA-E7Full и CyaA-Е7Δ30-42, полностью накладываются друг на друга. В каждом случае выживаемость животных, получивших рекомбинантные аденилатциклазы с HPV16-E7, значительно повышена по сравнению с мышами, не получавшими никакого лечения или получавшими симулирующее лечение (р<0.05).
Фигура 6. Клетки опухоли ТС-1, взятые из растущих непродолжительное время опухолей и культивируемые вне организма, утрачивают экспрессию молекулы H-2Db.
В экспериментах по отторжению опухоли у некоторых животных, которых вакцинировали конструктом CyaA-E7Full, на поздних стадиях эксперимента появились опухоли (Фиг.4, *). Двух животных забили, чтобы культивировать опухоли вне организма. Клетки этих опухолей, ТС-1 А1 и А2, а также нативные клетки ТС-1 анализировали методом FACS® (сортировки клеток с возбужденной флуоресценцией), чтобы определить уровень экспрессии молекулы H-2Db (жирная линия). Указаны медианы интенсивности флуоресценции (MedFi). Показаны результаты, полученные с изотипическим контролем (серый заштрихованный).
Фигура 7. Персистирование HPV16-E749-57 - специфических CD8* Т-лимфоцитов у мышей, получавших рекомбинантную CyaAs с HPV16-E7.
Умерщвляли мышей C57BL/6, иммунизированных CyaA-Е749-57 (А), CyaA-E7Full (В) или CyaA-Е7Δ30-42 (С) и выживших после трансплантации ТС-1 в серии терапевтических экспериментов. Спленоциты повторно стимулировали in vitro (в течение 5 дней 1 мкг/мл пептида HPV16-E743-77 в присутствии облученных сингенных спленоцитов. Лизис клеток-мишеней (ТС-1, обычные квадраты; EL4, пустые квадраты) оценивали по высвобождению 51Cr. Данные представляют медианные значения процентной доли лизиса (n=6 для каждой группы), а также интерквартильный размах. Число животных, у которых развился ответ, определяли по специфическому лизису ≥20% при максимальном отношении эффектор/мишень. Это число указано в правой верхней части квадрантов.
Фигура 8. Продолжительная защита против роста опухоли ТС-1, стимулируемая рекомбинантными аденилатциклазами, несущими HPV16-E7. Мышам C57BL/6, выжившим после трансплантации ТС-1 в серии терапевтических экспериментов, в день 100 повторно трансплантировали под кожу 5×104 клеток ТС-1. В качестве контролей брали не получавших лечения мышей соответствующего возраста (А). Показан рост опухолей у мышей, первоначально иммунизированных CyaA-Е749-57, CyaA-E7Full, и CyaA-Е7Δ30-42 (В, С и D соответственно). Мышей умерщвляли, когда размер опухоли достигал 1000 мм3, или когда этого требовало санитарное состояние животных. (Е) Кривые выживаемости животных: не получавшие лечения, пустые квадраты; или иммунизированные Cya-Е749-57, пустые треугольники; CyaA-E7Full, пустые кружки или CyaA-Е7Δ30-42, (пустые ромбы) и животные, которым повторно трансплантировали клетки ТС-1 (день трансплантации взят за 0). Во всех случаях выживаемость мышей, получавших рекомбинантные аденилатциклазы, несущие HPV16-E7, значительно повышена по сравнению с мышами, получавшими препарат - симулятор (р<0.05).
Фигура 9. Сравнение терапевтической активности CyaA-Е7Δ30-42 с терапевтической активностью HPV16E743-77 (фрагмента белка Е7 вируса ВПЧ16) в адъюванте CpG ODN 1826 (цитозин гуанин динуклеотид, содержащий олигодезоксинуклеотиды).
В День 0 мышам C57BL/6 подкожно трансплантировали 5х104 клеток опухоли ТС-1. В дни +10 и +17 мышам делали по одной инъекции 10 мкг HPV16-E743-77 (n=5, треугольники), 1 мкг CpG-ODN 1826 (n=5, квадратики), 10 мкг HPV16-E743-77 + 1 мкг CpG-ODN 1826 (n=5, ромбы), 10 мкг CyaA-CysOVA (n=3, перевернутые треугольники) или 10 мкг CyaA-Е7Δ30-42 (n=7, кружки). Мышей умерщвляли, когда размер опухоли превышал 1000 мм3, либо когда этого требовало санитарное состояние животных.
Фигура 10. Анализ влияния предшествующего иммунитета к аденилатциклазе на способность CyaA-Е7Δ30-42 стимулировать отторжение опухоли ТС-1.
(А) Мышей C57BL/6 либо оставляли без обработки либо иммунизировали в день -90 или -30 (две инъекции в день с 7-дневными интервалами, 10 мкг CyaAE5). В День -1 у животных брали кровь. Сыворотку крови каждого животного исследовали отдельно методом твердофазного иммуноферментного анализа (ELISA) на присутствие анти-CyaAE5 Иммуноглобулинов G. Результаты выражены в индивидуальных титрах антител, рассчитанных методом линейной регрессии по графику разведение - А492 (оптическая плотность на длине волны 492 нм). Горизонтальные линии представляют медианное значение ответа в каждой группе. (В) В день 0 необработанным (а, b), иммунизированным в день -30 CyaAE5 (с, d) и иммунизированным в день -90 CyaAE5 (е, f) животным трансплантировали под кожу 5×104 клеток опухоли TC-1, а в дни +10 и +17 делали им по одной инъекции 10 мкг CyaA-cysOVA (а, с, е) или 10 мкг CyaA-Е7Δ30-42 (b, d, f). Вклейки (b, d, f) представляют собой увеличенные изображения за период с Дня 0 по День 35, показывающие, что у всех животных к моменту вакцинации развились пальпируемые опухоли. Каждая кривая изображает рост опухоли у отдельного животного. Мышей умерщвляли, когда размер опухоли превышал 1000 мм3 либо когда этого требовало санитарное состояние животных. Справа вверху каждого квадранта указано число забитых животных по отношению к общему числу животных, включенных в эксперимент.
Фигура 11. Стимулирование ответов цитотоксических Т-лимфоцитов у Мышей HHD рекомбинантной аденилатциклазой CyaA-HPV16E7Δ30-42.
Белки, которыми «нагружены» клетки EL4-HHD, указаны над каждым графиком. В расположенных ниже таблицах в столбце под заголовком Vacc (вакцина) указан тип аденилатциклазы, которую вводили путем инъекции (HPV16E7 или HPV18E7), а в столбце под заголовком Stim (стимуляция) указан пептид, который использовали для повторной стимуляции in vitro. Можно увидеть, что после иммунизации аденилатциклазой CyaA-HPV16E7Δ30-42 нам удалось стимулировать цитотоксические Т-лимфоциты, специфичные только к пептиду #253 (правая картинка). Эта активность цитотоксических Т-лимфоцитов была специфичческой, поскольку повторно стимулированные in vitro пептидом #255 спленоциты не проявляли цитотоксичности в отношении клеток EL4-HHD, покрытых пептидом #253 (правая картинка). Отсутствие ЦТЛ-специфических ответов по отношению к двум другим рестриктированным по HLA-A2 пептидам может быть следствием различных явлений: (i) иммунодоминантность пептида #253, (ii) отсутствие переработки пептидов #255 и #258 протеосомами клеток EL4-HHD, (iii) плохая растворимость пептида #258, для которого пришлось использовать ацетонитрил (50%), который, возможно, токсичен для клеток. Очень интересно, что при одновременном введении (путем инъекции) CyaA-HPV18E7Δ32-42 не оказывала отрицательного влияния на способность CyaA-HPV16E7Δ30-42 стимулировать цитотоксические Т-лимфоциты (правый рисунок).
Фигура 12. Стимулирование ответов цитотоксических Т-лимфоцитов аденилатциклазой CyaA-HPV18E7Δ32-42.
Белки, которыми «нагружены» клетки EL4-HHD, указаны над каждым графиком. В расположенных ниже таблицах в столбце под заголовком Vacc (вакцина) указан тип аденилатциклазы, которую вводили путем инъекции (HPV16E7 или HPV18E7), а в столбце под заголовком Stim (стимуляция) указан пептид, который использовали для повторной стимуляции in vitro. Можно увидеть, что после иммунизации аденилатциклазой CyaA-HPV18E7Δ32-42 нам удалось стимулировать цитотоксические Т-лимфоциты, специфические только к пептиду #251 (левый рисунок). Эта активность цитотоксических Т-лимфоцитов была специфической, поскольку повторно стимулированные in vitro пептидом #257 спленоциты не проявляли цитотоксичности в отношении клеток EL4-HHD, покрытых пептидом #251 (левый рисунок). Что касается пептидов HPV16E7, рестриктированных по HLA-A2, то отсутствие ЦТЛ-специфических ответов по отношению к пептиду HPV18E7 #257, рестриктированному по HLA-A2, может быть следствием различных явлений: (i) иммунодоминантности пептида #251, (ii) отсутствие процессинга пептида #257 в протеосомах клеток EL4-HHD, (iii) плохая растворимость пептида #257, для которого пришлось использовать ацетонитрил (50%), который, возможно, токсичен для клеток. Очень интересно, что при одновременном введении (путем инъекции) CyaA-HPV16E7Δ30-42 не оказывала отрицательного влияния на способность CyaA-HPV18E7Δ32-42 стимулировать цитотоксические Т-лимфоциты (левый рисунок).
Фигура 13. Конструирование pTRACE5-E7HPV18
Фигура 14. Конструирование pTRACE5-E7ΔHPV16+18
Раскрыты 3 этапа приготовления плазмиды.
Фигура 15: Карта кодирования полинуклеотида:
CyaA-HPV18E7Full(полноразмерный)
CyaA-HPV18E7Δ32-42
CyaA-HPV16+18E7Full(полноразмерный)
CyaA-HPV16+18Е7Δ
Фигура 16: Стимуляция цитотоксических Т-лимфоцитов рекомбинантными аденилатциклазами, несущими HPV18E7 у мышей C57BL/6.
Фигура 17: Induction of CTL by recombinant CyaAs carrying HPV18E7 in C57BL/6 mice.
Депонированный материал (I-3190 и I-3191) содержится в штамме BLR бактерии E.coli, которую можно выращивать в жидкой среде Luria Broth (LB), сеять в LB со 100 мкг/мл ампициллина и инкубировать при 30°С в воздухе со встряхиванием (175 об./мин) и освещением. Возможна консервация в течение ночи в среде LB с 7-10% ДМСО.
ПРИМЕРЫ
Пример 1
В этом примере мы сконструировали рекомбинунтную аденилатциклазу, содержащую либо полноразмерную последовательность белка Е7 вируса HPV16, либо более мелкие фрагменты этого полипептида (в частности, пептид, содержащий остатки 49-57 белка Е7, который соответствуют эпитопу, растриктированному по H-2Db, и остатки 43-98 плюс 1-29 белка HPV16-E7). Мы показали, что при введении путем инъекции мышам C57BL/6, эти рекомбинантные аденилатциклазы с белком Е7 вируса ВПЧ16 способны стимулировать специфический отфет цитотоксических Т-лимфоцитов и хелперов Th1, характеризующиеся секрецией ИФН-γ. Кроме того, при терапевтическом исследовании эти конструкции продемонстрировали способность обеспечивать 100% зациты против подкожного трансплантата клеток ТС-1. Эти исследования впервые продемонстрировали in vivo противоопухолевую терапавтическую активность аденилатциклазы против опухольспецифических антигенов.
Материалы и Методы
Линии мышей и клеток. Беспатогенных самок мышей линии C57BL/6 в возрасте 6-10 недель получали из CER Janvier (Ле Гене Сент-Иль, Франция) или Charles River (Л'Арбрель, Франция). Также в этом исследовании использовали мышей ТАР1-/- (18), ГКСГ класс II-/- (19) и CD40-/- (20) выведенных на фоновом фенотипе C57BL/6. Животных содержали в виварии Института Пастера в беспатогенных условиях, со свободным доступом к пище и воде. Эксперименты с участием животных проводили согласно установленным рекомендациям по уходу за животными.
Клетки ТС-1, экспрессирующие белки Е6 и Е7 вируса ВПЧ16 (21), и клетки тимомы мыши EL4 (17) получали из АТСС (американская коллекция типовых культур). Клетки поддерживали в среде RPMI 1640 с Glutamax, дополненной 10% термоинактивированной сыворотки крупного рогатого скота, 100 U/мл пенициллина, 100 мкг/мл стрептомицина, 0.4 мг/мл генетицина (только для клеток ТС-1) и 5·10-5 М 2-меркаптоэтанола (Gibco BRL, Сержи-Понтуаз, Франция).
Пептиды. Синтетические пептиды Е749-57 (RAHYNIVTF, однобуквенный код для аминокислот), соответствующий рестриктированному по H2-Db эпитопу белка Е7 вируса ВПЧ16 (22), и Е743-77 (GQAEPDRAHYNIVTFCCKCDSTLRLCVQSTHVDIR) соответствующий эпитопу Е749-57 для цитотоксических Т-лимфоцитов с его естественными фланкирующими последовательностями и эпитоп для Т-хелперов (жирный) (23) приобретали в Neosystem (Страсбург, Франция). CpG ODN 1826 приобретали в PROLIGO (Париж, Франция).
Конструирование и очистка рекомбинантной аденилатциклазы В. pertussis, несущей эпитопы белка Е7 вируса папилломы человека типа 16 (HPV16-E7).
Рекомбинантную аденилатциклазу, применяемую в этом пункте, экспрессировали в Е.coli, используя производные плазмид pTRACE5 (Фиг.1А), которые кодируют CyaA (24) (25), не обладающую ферментативной активностью. Плазмида pTRACE5 представляет собой вектор экспрессии для экспрессии не обладающих ферментативной активностью и, следовательно, цитотоксических вариантов CyaA бактерии В. pertussis. Она также экспрессирует белок CyaC бактерии В. pertussis, который необходим для посттрансляционного ацилирования СуаА. Эту плазмиду получают из описанной ранее плазмиды pTRACG (Gmira et al., 2001, Res. Mic. 152:889). Ее получили путем вставки гексануклеотида CTGCAG в сайт EcoRV, расположенный в 5'-части последовательности ДНК cyaA. Результатом является внутрирамочная вставка дипептида Leu-Gln между Asp188 и IIe189 белка CyaA в пределах основной части каталитического сайта (Guermonprez et al. 2000, Meth. Enzymol. 326:527).
Плазмида pTRACE5 содержит ориджин репликации CoIE1 и сайт устойчивости к ампициллину. В этой плазмиде гены CyaC и модифицированной CyaA расположены в одной транскрипционной единице под контролем промотора Pr фага λ. Плазмида pTRCAG кодирует также термочувствительный репрессор cl857 фага λ, который сильно подавляет транскрипцию с промотора Pr фага λ при температуре ниже 32°С.
Для всех манипуляций с ДНК, которые проводили, следуя стандартным протоколам (Maniatis et al.)., использовали Штамм Е. coli XL1-Blue (Stratagene, Ла Хойа, Калифорния, США).
CyaA-Е749-57 содержит последовательность из 9 аминокислот (RAHYNIVTF), встроенную между кодонами 224 и 235 аденилатциклазы. Плазмиду для экспрессии CyaA-Е749-57 конструировали следующим образом. Два синтетических олигонуклеотида (MWG, Куртабеф, France), BTP1 (5'-СТА GCC GTG ССС АТТ АСА АТА TTG ТАА ССТ TTG GТА С-3' кодирующая нить) и ВТР2 (5'-САА AGG ТТА САА TAT TGT ААТ GGG САС GG-3' некодирующая нить) отжигали и лигировали в pTRACE5, расщепленную ферментами Nhel и Kpnl. CyaA-E7Full содержит полную последовательность белка HPV16-E7, т.е. 98 аминокислот, встроенных в той же позиции 224 аденилатциклазы, не обладающей ферментативной активностью. Последовательность ДНК, кодирующую белок Е7 амплифицировали из ДНК вируса ВПЧ16 (Seedorf К et al, см. выше) с использованием специфических праймеров ВТР3, (5'-GGG CGC TAG CAT GCA TGG AGA TAC ACC TAC-3') и ВТР4 (5'-GGG CGG TAC CTG GTT TCT GAG AAC AGA TGG G-3'). Полученный в результате ПЦР продукт расщепляли ферментами Nhel и Kpnl и лигировали в pTRACE5, расщепленную ферментами Nhel и Kpnl. Сайт Sspl, присутствующий в олигонуклеотиде после отжига, а также в полной последовательности ВПЧ16-Е7, позволял быстро идентифицировать мутанты, несущие вставку. CyaA-Е7Δ30-42 содержит первые 29 остатков аминокислот белка HPV16-E7, встроенные между кодонами 319 и 320 аденилатциклазы, а также остатки 43-98 белка HPV16-E7, введенные между кодонами 224 и 235 аденилатциклазы. Плазмиду для экспрессии CyaA-Е7Δ30-42 конструировали в два этапа. Первый фрагмент ДНК, кодирующий (остатки аминокислот с 1 по 29) белок HPV16-E7 амплифицировали методом ПЦР, используя в качестве целевой ДНК синтетический ген HPV16-E7 (оптимизированный для продуцирования в E.coli, созданный в GTP Technology, Лабеж, Франция), и праймеры ВТР5 (5'-GGG САС CGG ТАА ACG TAT GCA CGG CGA TAC ТСС G-3') и ВТР6 (5'-CGT GAG CAT CTG GCT TTC ACT AGT ACG TTT GTT CAG CTG CTC GTA GCA-3'). Второй фрагмент ДНК, кодирующий кодоны 320-372 аденилатциклазы амплифицировали методом ПЦР, используя pTRACE5 в качестве целевой DNA и праймеры ВТР7 (5'-GGG САС TAG TGA AAG ССА GAT GCT САС GCG CGG G-3') и ВТР8 (5'-AGT АСА ТСС GGC GAG ААС-3'). Эти два фрагмента ДНК (которые частично перекрываются) очищали и комбинировали с праймерами ВТР5 и ВТР8 в третьей ПЦР, целью которой была амплификация фрагмента ДНК длиной 294 пар оснований. Эти фрагменты расщепляли ферментами Agel и BstBl и вставляли между соответствующими сайтами плазмиды pTRACE5, получая в результате плазмиду. pTRACE5-E71-29. Затем по методу ПЦР амплифицировали фрагмент ДНК, кодирующий с 43 по 98 остатки аминокислот белка HPV16-E7, используя синтетический ген HPV16-E7 в качестве целевой ДНК и праймеры ВТР9 (5'-GGG CGC TAG CGG ТСА AGC AGA ACC GGA С-3') и ВТР10 (5'-GGG CGG TAC CAG GTT TTT GAG AGC ААА TCG GAC ААА САА ТСС ССА GAG TAC ССА ТС-3'). Полученные в результате ПЦР и очищенные фрагменты расщепляли ферментами Nhel и Kpnl и лигировали в плазмиду pTRACE5-E71-29, расщепленную теми же ферментами рестрикции.
Рекомбинантную аденилатциклазу полностью получали в штамме BLR Escherichia coli (Novagen, Мэдисон, Висконсин, США), описанным ранее способом (26). Рекомбинантные белки очищали от включений до состояния, близкого к гомогенному (Фиг 1В), посредством двухэтапной процедуры, которая включает хроматографию на ДЭАЭ-Сефарозе и фенил-Сефарозе, проводимые описанным ранее способом (26). В случае хроматографии на фанил-Сефарозе в процедуру включали этап дополнительной промывки 60% изопропанолом в 20 мМ Hepes-Na, pH 7.5, с целью устранения большей части загрязняющих липолисахаридов. Содержание липополисахаридов определяли при помощи набора QCL-1000 (Biowhittaker, Уокесвилль, Мэрилэнд, США). Очищенные рекомбинантные белки исследовали в ДСН-геле. Концентрацию белков определяли методом спектрофотометрии по поглощению на длине волны 280 нм, используя в расчетах коэффициент молярной экстинкции, равный 142,000 М-1, см-1.
Конструирование и очистка рекомбинантного белка HPV16-E7. Оптимизированную для экспрессии в E.coli кДНК, кодирующую последовательность ДНК белка HPV16-E7 (можно заказать в GTP technology), субклонировали в векторе pIVEX2.4b (Roche Molecular Biochemicals, Мейлдан, Франция) между сайтами рестрикции Ncol и Xhol. Полученную в результате плазмиду трансформировали в E.coli, штамм BL21λDE3 (Novagen). Экпрессия белка Е7 вируса ВПЧ16 с полигистидиновой меткой (His-Tag-HPV16-E7) происходила после стимуляции 0.5 миллимолями изопропил-β-D-тиогалактопиранозида (Euromedex, Souffelweyersheim, Франция) и очищали на Никель-тринитирилацетат-агарозе (Qiagen, Hilden, Германия), следуя инструкциям производителя. Для удаления загрязняющих липополисахаридов использовали промывки изопропанолом, описанные в (27).
Иммуноблотинг. Белки разделяли в полиакриламидном геле с додецилсульфатом натрия и переносили в электрическом поле на нитроцеллюлозную мембрану (0.45 мкм, BioRad, Марн-ля-Кокетт, Франция), которую зондировали моноклональным анти- HPV16 Е7 антителом мыши (Zymed, Сан-Франциско, Калифорния, США) или поликлональной сывороткой против BLR Е.coli, приготовленной в мышах C57BL/6. Иммунный комплексы детектировали при помощи анти-мышиных иммуноглобулинов козы, конъюгированных с щелочной фосфатазой (Chemicon, Темекула, Калифорния, США) и проявляли 5-бром-4-хлор-3-индолилфосфат/нитросиний тетеразолий (BCIP/NBT) (Sigma, Сент-Луис, Миссури, США).
Иммунизация мышей и эксперименты по отторжению опухоли. Животных иммунизировали, делая им одну внутривенную инъекцию 50 микрограмм или две внутрикожные инъекции (по 10 мкг каждая) контрольного состава или рекомбинантных аденилатциклаз с белком Е7 вируса HPV16, разбавленных фосфатным буферным раствором (Gibco BRL). Внутрикожные инъекции делали в кожу ушей (47). Для проведения анализа in vivo, у умерщвленных животных (CO2) брали селезенку. Эту процедуру проводили через 7 дней после инъекций, а для анализа продолжительных ответов - через 3 месяца после инъекции. В экспериментах по отторжению опухоли мыши получали 5×104 клеток ТС-1 подкожно, а затем рекомбинантные аденилатциклазы с белком Е7 вируса ВПЧ16 (HPV16-E7-CyaA) в течение 1, 5 или 10 дней после инокуляции опухоли. За ростом опухоли ТС-1 следили, измеряя ее размер циркулем и рассчитывая объем по формуле V=(L×w2)/2 (V: объем, L: длина, w: ширина) (48).
In vitro анализ цитотоксичности. Взятые у иммунизированных мышей спленоциты in vitro стимулировали пептидами Е749-57 или Е743-77 в концентрации 1 мкг/мл в присутствии облученных сингенных нативных клеток селезенки в полной среде (RPMI 1640 с Glutamax, дополненная 10% термоинактивированной сывороткой крупного рогатого скота FCS, 100 U/мл пеницилиина, 100 мкг/мл стрептомицина и 5·10-5 М 2-меркаптоэтанола) в течение 5 дней. Цитотоксичную активность этих эффекторных клеток определяли в пятичасовом анализе высвобождения 51Cr на клетках ТС-1. Введение радиоактивной метки осуществляли следующим образом: культивируемые в атмосфере, содержащей 7.5% CO2, при температуре 37°С клетки ТС-1 в экспоненциальной фазе роста быстро трипсинизировали (Trypsin-EDTA, GibcoBRL) и инкубировали в течение одного часа при температуре 37°С с 100 мкКи 51Cr. Использовали различные отношения эффектор: мишень, все тесты проводили в дупликатах. Измеряли радиоактивность, высвобождаемую в надосадочной жидкости из каждой ячейки. Процентную долю специфического лизиса рассчитывали как 100 × (определенное экспериментально высвобождение - спонтанное высвобождение) / (максимальное высвобождение - высвобождение). Максимальное высвобождение определяли в клетках-мишенях, к которым добавляли 10% Triton X-405, а спонтанное высвобождение определяли в клетках-мишенях, которые инкубировали в полной среде без добавок. Считали, что у мыши развился ответ, если при самом высоком отношении эффектор:мишень наблюдали по меньшей мере 20% специфический лизис. Результаты выражены как медиана ± межквартильный разброс числа мышей, у которых развился ответ на группу.
Исследование отдельной клетки, продуцирующей ИФН-γ методом иммуноферментный спот-анализа для секретирующих клеток. Фильтрационные планшеты Multiscreen (96 лунок; Millipore, Мольшен, Франиция) покрывали 4 мкг на мл антитела крысы против гамма-интерферона мыши (ИФН-γ) (клон R4-6A2; PharMingen, Сан-Диего, Калифорния, США). Процедуру проводили при комнатной температуре в течение ночи. Затем планшеты промывали и блокировали полной средой. В лунки добавляли серийные двукратные разведения клеток селезенки иммунизированных мышей и 5×105 сингенных фидерных клеток, подвергнутых гамма-облучению (2,500 рад). Клетки инкубировали в течение 36 часов с пептидом Е749-57 в концентрации 1 мкг/мл или без него. После тщательных промывок планшеты проявляли путем инкубации с 5 мкг на мл биотинилированных антител крысы против ИФН-γ мыши (клон XMG 1.2; PharMingen) за которой следовала инкубация с конъюгатом страптавидин-щелочная фосфатаза (PharMingen). Наконец, пятна проявляли, используя в качестве субстрата 5-бром-4-хлор-3-индолил фосфат (BCIP) и нитротетразол синий (NBT). Число продуцирующих ИФН-γ клеток определяли, подсчитывая число пятнообразующих клеток в каждой лунке (Bioreader, Карбен, Германия). Результаты выражали в общем числе пятнобразующих клеток на селезенку (17).
Твердофазный иммуноферментный анализ (ELISA). У мышей, иммунизированных путем внутрикожного введения пустого вектора CyaAE5, через 30 или 90 дней брали кровь и тестировали сыворотку крови отдельных мышей на ответы антител методом твердофазного иммуноферментного анализа. Микротитрационные планшеты (Nunc, Роскилд, Дания) в течение ночипокрывали пустым вектором CyaAE5 (3 мкг/мл) в ФБР. После промывок в смеси ФБР-tween 20 (0.1%), в лунки добавляли разбавленную сыворотку и инкубировали в течение 1 часа при 37°С. После промывок в смеси ФБР-tween 20 планшеты инкубировали с конъюгатом антитела козы против IgG мыши с пероксидазой (Sigma) в течение 1 часа при 37°С. Планшеты проявляли при помощи о-фенилнедиамина и перекиси водорода (Sigma). Останавливали реакцию серной кислотой и анализировали планшеты на длине волны 492 нм в считывающем устройстве для ELISA (Dynatech, Марн-ля-Кокетт, Франция). Результаты выражены в титрах антител, рассчитанных методом линейной регрессии по графику разведение против А492(оптическая плотность на длине волны 492 нм). Титры рассчитывали как log10 самого высокого разведения, которое дает двойную плотность объединенной контрольной сыворотки, разведенной 1/100.
Продукция цитокинов. Клетки селезенки иммунизированных мышей стимулировали in vitro 10 мк/мл белка HisTag-HPV16-E7 (HPV16-E7 с полигистидиновой меткой) или 1 мкг/мл пептида Е743-77 в полной среде в течение 72 часов. Продукцию ИФН-γ и ИЛ-5 определяли в надосадочных жидкостях культур методом твердофазного иммуноферментного анализа (ELISA) типа «Сэндвич», как было описано ранее (28). Все тесты стандартизировали по соответствующим рекомбинантным цитокинам мыши (Pharmingen).
Анализ методом сортировки клеток с активацией флуоресценцией (FACS®). Для анализа уровня экспрессии молекулы Н-2Db главного комплекса гистосовместимости класса I методом поточной цитометрии с использованием специфического моноклонального антитела, конъюгированного с флуоресцина изотиоцианатом (FITC) (клон КН95, Pharmingen, Ле Пон-де-Кле, Франция), клетки ТС-1 обрабатывали, как описано других местах данной заявки (29).
Статистический анализ. Учитывая небольшой размер различных выборок, применяли непараметрические статистические критерии (30). Анализ проводили при помощи программного обеспечения StatXact 4 (Cytel corporation, Кембридж, Массачусетс, США). Кривые выживаемости строили при помощи программного продукта Prism (GraphPad Software Inc., Калифорния, США) и сравнивали при помощи встроенного логарифмического рангового критерия. Данные считали существенно различными при р<0.05.
Результаты
Конструирование и описание рекомбинантной аденилатциклазы, несущей эпитопы белка Е7 вируса ВПЧ16 (HPV16-E7). Чтобы изучить способность аденилатциклазы стимулировать HPV16-E7 - специфические ответы Т-климфоцитов, мы сконструировали 3 различные рекомбинантные молекулы. CyaA-Е749-57 содержит последовательность полипептида длиной 9 аминокислот (RAHYNIVTF), соответствующую описанному ранее рестриктированному по Н-2Db эпитопу, распознаваемому ЦТЛ (22), которые встраивали между кодонами 224 и 235 аденилатциклазы, не обладающей ферментативной активностью (следовательно, нетоксичной). CyaA-E7Full(полноразмерный) содержит полную последовательность (98 аминокислот) белка HPV16-E7, встроенного в той же позиции 224 аденилатциклазы, не обладающей ферментативной активностью. CyaA-Е7Δ30-42 содержит первые 29 аминокислот белка HPV16-E7, встроенные между кодонами 319 и 320 CyaA, не обладающей ферментативной активностью, а также остатки с 43 по 98 белка HPV16-E7, встроенные между кодонами 224 и 235. Чтобы сделать возможным проведение анализов in vitro и in vivo assays, получали содержащие CyaA конструкты и очищали их до состояния, близкого к гомогенному (Фиг.1В) В протокол очистки включали процедуру удаления липополисахаридов (26), что позволяло получать рекомбинантные белки, содержащие менее 100 единиц эндотоксина на 50 мкг. Присутствие белка Е7 в конструктах CyaA-E7Full и CyaA-Е7Δ30-42 подтвердили методом Вестерн-блот с использованием специфического моноклональнго антитела (Zymed) (Фиг.1C). CyaA-Е749-57, содержащий только эпитоп, рестриктированный по H-2Db, наоборот, не распознавался анти-НРV16-Е7 антителом. Общие биохимические свойства трех рекомбинантных аденилатциклаз не изменились, эти молекулы проявляли гемолитическую активность, сходную с гемолитической активностью аденилатциклазы дикого типа (17).
Иммунизация рекомбинантными аденилатциклазами, несущими белок Е7 вируса ВПЧ16, стимулирует Е7-специфические ответы цитотоксических Т-лимфоцитов. Чтобы исследовать способность CyaA стимулировать ответы цитотоксических Т-лимфоцитов против эпитопов белка Е7 вируса ВПЧ16, мышей C57BL/6 однократно иммунизировали, вводя им внутривенно 50 мкг различных рекомбинантных аденилатциклаз, несущих HPV16-E7. Собирали спленоциты и стимулировали их in vitro 1 мкг/мл пептида Е743-77. Через 5 дней в анализе, основанном на высвобождении 51Cr, определяли их способность лизировать клетки ТС-1. Как показано на фиг.2А, однократная иммунизация мышей C57BL/6 путем внутривенного введения рекомбинантных аденилатциклаз, несущих белок HPV16-E7, стимулирует сильный и специфический ответ цитотоксических Т-лимфоцитов на ТС-1. Иммунизация аденилатциклазой, содержащей полноразмерный белок HPV16-E7 (Фиг.2А, b) или его делегированную форму, CyaA-Е7Δ30-42 (Фиг.2А, с) вызывала более высокие значения максимальной активности ЦТЛ, по сравнению с активностью, стимулируемой CyaA-Е749-57, которая содержит лишь минимальный ограниченный по H-2Db эпитоп (Фиг.2А, а). Однако мы не можем продемонстрировать значимое различие, исходя из наших статистических данных. Сходные результаты были получены при использовании пептида Е749-57 для повторной стимуляции in vitro (данные не показаны). Спленоциты мышей, вакцинированных рекомбинантной аденилатциклазой, несущей не имеющий отношения к данному изобретению эпитоп (OVA257-264), которые повторно стимулировали in vitro пептидом Е743-77 в концентрации 1 мкг/мл давали лишь слабый неспецифический лизис ТС-1 клеток (Фиг.2А, а). Ранее было показано, что доставка эпитопа OVA для клеток CD8+(SIINFEKL) к молекуле ГКГС класса I аденилатциклазой CyaA in vivo зависит от функции ТАР1 (15). Мы исследовали, будет ли то же самое наблюдаться в случае использования CyaA-Е7Δ30-42. Как показано на фиг.2А, d, спленоциты вакцинированных путем внутривенной инъекции мышей ТАР1-/-, после стимуляции in vitro были не способны лизировать клетки ТС-1. Мы также исследовали необходимость участия CD4+ Т-лимфоцитов в in vivo стимуляции ответа цитотоксических Т-лимфоцитов рекомбинантной аденилатциклазой, содержащей HPV16E7. Мы наблюдали, что внутривенная вакцинация мышей ГКГМ класса II-/- конструктом CyaA-Е7Δ30-42 вызывала высокие уровни специфических ответов ЦТЛ на клетки ТС-1 (Фиг.24А, е), что согласуется с более ранними наблюдениями (15). В этой модели мы, напротив, наблюдали некоторую зависимость от передачи сигнала через CD40, а также низкий уровень ответа ЦТЛ на клетки ТС-1 в мышах CD40-/- (Фиг.2А, f).
Чтобы оценить ex vivo встречаемость HPV16-Е7-специфических спленоцитов у мышей, иммунизированных рекомбинантными аденилатциклазами, методом ферментно-опосредованного иммуноспота (ELISPOT) подсчитывали число клеток, продуцирующих ИФН-γ в ответ на стимуляцию in vivo пептидом HPV16-Е749-57. На фиг.2В показано, что существует лишь незначительная разница в числе спленоцитов, продуцирующих ИФН-γ, в частности спленоцитов, продуцирующих ИФН-γ, полученных из мышей, иммунизированных CyaE5-CysOVA, по сравнению с мышами, иммунизированными CyaA-Е749-57. Напротив, число полученных спленоцитов, продуцирующих ИФН-γ, у мышей, вакцинированных рекомбинантными аденилатциклазами с HPV16-E7, содержащими либо полноразмерный белок HPV16-E7, либо его делегированную форму, было намного выше (р<0.05). Наблюдаемые ответы были эпитот-специфичными, поскольку очень лишь небольшое число клеток селезенки продуцировало ИФН-γ без стимуляции пептидом (Фиг.2В). Эти результаты показывают, что аденилатциклаза обладает способностью доставлять in vivo иммунодоминантный рестриктированный по H-2Db CD8+ T-лимфоцитов эпитоп белка HPV16-E7 в цитозоль иммунокомпетентных клеток, где он перерабатывается и презентируется по пути ГКГС класса I, что приводит к напряженным ответам цитотоксических Т-лимфоцитов. Мы подтверждаем, что в аденилатциклазу можно вводить крупные фрагменты полипептидов, поскольку аденилатциклазы, несущие полноразмерный белок HPV16-E7, либо его делетированную и инвертированную форму, также способны стимулировать напряженные ответы токсических Т-лимфоцитов, что согласуется с более ранними наблюдениями (25, 31). Наши данные также демонстрируют, что эти последние молекулы стимулируют значительно более высокую встречаемость HPV16-Е749-57-специфических ответов у мышей.
Иммунизация рекомбинантными аденилатциклазами, несущими белок Е7 вируса ВПЧ16 стимулирует Е7-специфические ответы Т-хелперов Th1.
Ответы Th1 играют важную роль в обеспечении защиты против внутриклеточных патогенов и развития опухолей (32, 33). Соответственно, мы описали ответы этого типа Т-лимфоцитов, стимулированные рекомбинантными аденилатциклазами, несущими HPV16-E7. Мышей C57BL/6 однократно иммунизировали путем внутривенного введения 50 микрограмм трех разных рекомбинантных аденилатциклаз, несущих HPV16-E7, и определяли синтез цитокинов после стимуляции in vitro клеток селезенки 10 мкг/мл очищенного белка HPV16-E7 с полигистидиновой меткой. Как показано на Фиг.3, иммунизация аденилатциклазами, несущими полноразмерный белок HPV16-E7 или его делетированную форму, давала Th1-подобный профиль, характеризующийся продукцией высоких уровней ИФН-γ и отсутствием детектируемых уровней ИЛ-5. Этот ответ был специфическим, поскольку уровни ИФН-γ, полученный после иммунизации CyaA-E7Full или CyaA-Е7Δ30-42 были существенно выше уровней, полученных у мышей в результате иммунизации симулятором CyaAE5-CysOVA (p<0.05). Однако в случае со спленоцитами, иммунизированными CyaA-Е749-57, это было не так. Сходные результаты были достигнуты, когда повторную стимуляцию проводили 1 мкг/мл пептида Е743-77 (Фиг.3А, вкладка).
Взятые вместе, эти результаты указывают на то, что в использованных нами условиях, CD4+ Т-лимфоциты играют важную роль в секреции ИФН-γ, поскольку уровни, полученные при использовании аденилатциклаз, несущих белок HPV16-E7, которые содержат рестриктированные по Н-2b класса II Т-клеточные эпитопы, намного превышают уровни, полученные при использовании CyaA-Е749-57, которая содержит только эпитоп, рестриктированный по Н-2b класса I.
Иммунизация рекомбинантными аденилатциклазами, несущими белок Е7 вируса ВПЧ16 (HPV16-E7), стимулируют обратное развитие развитых опухолей, экспрессирующих HPV16. Учитывая получение устойчивых иммунологических ответов, мы затем оценивали in vivo терапевтическую активность аденилатциклаз, несущих HPV16-E7, в предклинических моделях, представляющих собой линию опухолевых клеток Н-2b, экспрессирующих белки Е6 и Е7 вируса ВПЧ16 (клетки ТС-1), при введении мышам C57BL/6 путем подкожной инъекции. Ранее на этой модели было показано, что отторжение опухоли опосредуют исключительно Е749-57-специфические CD8+ Т-лимфоциты (21, 22, 34, 35). Соответственно, мышам C57BL/6 вводили 5×104 клеток ТС-1 путем подкожной инъекции в правый бок, а через 1, 5 или 10 дней мышам вводили 50 мкг CyaAE5-HPV16-E749-57, -E7full или -Е7Δ30-42 путем внутривенной инъекции. Фиг.4 изображает рост опухоли у мышей, получивших терапевтическое лечение через 10 дней после трансплантации опухоли. Примечательно, что в этих условиях у 100% мышей к моменту вакцинации развивались пальпируемые опухоли. Чтобы предотвратить ненужные страдания, мышей умерщвляли, когда размер опухоли достигал 1000 мм3. У всех животных, не получавших лечения, а также у животных, получавших препарат-симулятор CyaAE5-cysOVA развились опухоли такого же размера (>1000 mm3) в пределах максимум 49 дней. С этим разительно контрастирует тот факт, что у большинства животных, получавших рекомбинантные аденилатциклазы, несущие HPV16-E7, на протяжении всего эксперимента опухоли не развивались (Фиг.4, С, D, Е). На Фиг.5 показан график выживаемости животных, которым трансплантировали клетки ТС-1 и которых подвергли терапии по трем различным протоколам, им вводили рекомбинантные аденилатциклазы в день 1, день 5 или день 10 после трансплантации ТС-1. Медианные значения продолжительности выживания животных, не получавших никакого лечения и получавших симулятор лежали в пределах от 31 до 40 дней. И наоборот, выживаемость мышей, вакцинированных аденилатциклазами, несущими антигены HPV16-E7, была значительно выше выживаемости контрольных животных (р<0.05). Можно выявить разницу в защитной активности между различными конструктами, хотя из-за небольшого объема выборок эта разница не может быть признана статистически значимой. Если заметную разницу в степенях обратного развития опухолей, вызываемого CyaA-Е749-57 и -E7Full, выявить не удалось, то CyaA-Е7Δ30-42 явно превосходила их с точки зрения стимуляции обратного развития и подавления роста опухолей, поскольку во всех трех терапевтических схемах степень защиты превышала 90%.
Оказалось, что у некоторых животных, вакцинированных CyaA-Е749-57 и CyaA-E7Full, опухоли развились позже в ходе эксперимента (Фиг.4 (*) и не показанные данные). Учитывая, что этот феномен может отражать механизмы ускользания опухоли от иммунологического контроля, мы культивировали растущие опухоли, взятые у этих животных, вне организма и анализировали эти линии клеток, названные ТС-1 А1 и ТС-1 А2 методом сортировки клеток с возбужденной флуоресценцией (FACS) на предмет экспрессии H-2Db. Как показано на Фиг.6, по сравнению с их нативными аналогами, культивируемые вне организма клетки ТС-1 А1 и А2, взятые из поздних растущих опухолей, утратили экспрессию H-2Db, что, по всей вероятности, привело к тому, что они не распознаются Е749-57-специфическими CD8+ Т-лимфоцитами. Поскольку клетки ТС-1 росли in vivo в среде без селектирующего давления антибиотиков, мы также проверили экспрессию HPV16-E7 в клетках ТС-1 А1 и А2 методом Вестерн-блот. Нам не удалось обнаружить никакой разницы в экспрессии этого белка между клетками ТС-1 А1 и А2 (данные не показаны).
Вместе взятые, эти результаты демонстрируют аденилатциклазного вектора в качестве терапевтической вакцины, подходящей для стимуляции обратного развития опухолей, экспрессирующих ВПЧ16, в доклинической модели.
Мы также исследовали другой путь инъекционного введения, представляющий интерес с терапевтической точки зрения. Для этого мы вводили 10 мкг CyaA-Е7Δ30-42 путем внутрикожной инъекции дважды с 7-дневным интервалом, начиная с 10-го дня после трансплантации. Интересно, что в то время как у всех мышей, которым не вводили никаких препаратов или вводили препарат-симулятор, развивались опухоли, мы наблюдали обратное развитие опухоли у всех животных, которым вводили CyaA-Е7Δ30-42 (Фиг.4В, а, b). Результатом этой терапевтической иммунизации была 100% выживаемость мышей, получавших CyaA-Е7Δ30-42, на 90-й день, в то время как медианные значения времени выживаемости для мышей, не получавших никаких препаратов и получавших симулятор, составляли 30 и 32 дней соответственно (Фиг.4, с).
Продолжительное персистирование HPV16-Е749-57-специфических CD8+ Т-лимфоцитов, стимулированое иммунизацией аденилатциклазой CyaA.
Чтобы оценить устойчивость иммунного ответа, стимулируемого рекомбинантными аденилатциклазами, несущими HPV16-E7, мышей, выживших после терапевтических экспериментов, через 3 месяца умерщвляли и подвергали взятые у них стимуляции in vitro 1 (одним) мкг/мл пептида Е743-77 в течение 5 дней. Затем определяли их способность лизировать клетки ТС-1 по высвобождению 51Cr. Как показано на Фиг.7, специфические ответы цитотоксических Т-лимфоцитов на пептид HPV16-E743-77 имели место даже у спленоцитов животных, иммунизированных за три месяца до проведения анализа.
Исходя из максимальной процентной доли специфического лизиса при отношении эффектор: мишень, равном 30:1, можно сделать вывод, что иммунологический ответ более устойчив у животных, получавших CyaA-Е7Δ30-42, хотя исходя из этих данных нельзя продемонстрировать статистическую значимость этого результата. Чтобы оценить физиологическую значимость такой продолжительной иммуногенности оставшимся животным в день 100 повторно вводили 5×104 клеток ТС-1 подкожно. В таких условиях у всех подобранных по возрасту контрольных животных развились опухоли. Медианное значение периода выживаемости составило 37.5 дней (Фиг.8). У мышей, которых 3 три месяца до этого иммунизировали рекомбинантными аденилатциклазами, CyaA, несущими HPV16-E7, напротив, наблюдали очень сильную защиту против развития опухолей. Согласно наблюдениям (см. выше) животные, вакцинированные CyaAE5-HPV16-Е7Δ30-42, показали высокий уровень защиты. Однако в отличие от результатов, полученных в первой серии терапевтических экспериментов, в данном случае имеет место убедительная разница в уровне защиты между животными, получавшими CyaA-Е749-57, и теми, которые получали CyaA-E7Full, в пользу последних, хотя небольшой объем выборок не позволил однозначно продемонстрировать статистическую значимость. Эти наблюдения позволяют предположить, что в этой модели помощь Т-клеток, обеспечиваемая CyaA, несущей полный белок HPV16-E7, важна для эффективных продолжительных ответов против клеток ТС-1.
Взятые вместе, наши результаты демонстрируют, что одной внутривенной инъекции 50 мкг рекомбинантных аденилатциклаз, несущих хелперный эпитоп белка HPV16-E7 (DRAHYNIVTF), достаточно для стимуляции продолжительного ответа Е749-57-специфических CD8+ Т-лимфоцитов, которые способны обеспечить защиту против двух введений опухолевых клеток ТС-1 в течение периода времени продолжительностью по меньшей мере 6 месяцев.
Терапевтическая эффективность CyaA-Е7Δ30-42 выгодно отличается от терапевтической активности пептида, вводимого CpG-ODN 1826. Для того чтобы лучше оценить эффективность аденилатциклазы в качестве системы доставки антигенов, мы сравнили терапевтическую эффективность CyaA-Е7Δ30-42 с терапевтической эффективностью пептида HPV16-E743-77, дополненного CpG-ODN 1826 (37). С этой целью мышей иммунизировали, вводя им путем подкожной инъекции 5×104 клеток ТС-1, а затем в дни 10 и 17 мыши получали терапевтические вещества: внутрикожным путем им вводили 10 мкг CyaA-Е7Δ30-42, или 10 мкг пептида HPV16-E743-77, который вводили с 1 мкг CpG-ODN 1826. Степени выживаемости в этих двух группах были приблизительно одинаковы (Фиг.9). Хотя результаты, полученные при применении CyaA-Е7Δ30-42 были несколько лучше, однако статистически они не отличались от результатов, полученных при применении пептида HPV16-E743-77, смешанного с CpG-ODN 1826. Следует отметить, что эти результаты получили, когда количество используемого пептида HPV16-E743-77 было в 50 раз больше количества CyaA-Е7Δ30-42 (в молярном выражении). При использовании пептида HPV16-E743-77 без добавок, он не оказывал влияния на рост опухоли ТС-1.
Предшествующий иммунитет к вектору CyaA слабо влияет на терапевтическую эффективность CyaA-Е7Δ30-42. В клинических условиях может возникнуть необходимость давать пациенту с повреждениями множественные поддерживающие иммунизации для получения эффективного клеточного иммунного ответа. Следовательно, необходимо продемонстрировать, что предшествующий иммунитет к вектору CyaA не ослабляет его способность запускать реакцию отторжения опухли. Для этого мы иммунизировали мышей, вводя им дважды с семидневным интервалом путем внутрикожных инъекций 10 мкг пустого вектора CyaAE5 за 90 или 30 дней до введения путем подкожной инъекции 5×104 клеток ТС-1. Терапевтическое введение CyaA-Е7Δ30-42 (две внутрикожные инъекции с 7-дневным интервалом, по 10 мкг CyaA-Е7Δ30-42) проводили в день 10. Анализ ответов антител показал, что мыши, иммунизированные пустым вектором CyaA во время инъекции клеток ТС-1 были иммунными (Фиг.10А). Затем мы сравнили способность введенной CyaA-Е7Δ30-42 стимулировать отторжение опухоли в подобранных по возрасту животных и животных с иммунитетом к аденилатциклазе (CyaA). Вне зависимости от наличия иммунитета к CyaA, у большинства животных, которым вводили CyaA-Е7Δ30-42, на протяжении эксперимента опухоли не развивались (Фиг.10В). Только у одного животного в группе, которую иммунизировали на 30 день, и у двух животных в группе, которую иммунизировали на 90 день, развились опухоли (Фиг.10В b, d, f). Напротив, у 100% животных, которым вводили симулятор, развились опухоли и их забили (Фиг.10В, а, с, е). Мы не наблюдали никакой корреляции между уровнем титра анти-CyaA антител и развитием опухоли ТС-1 (данные не показаны). Кроме того, кривые выживаемости мышей, получавших CyaA-Е7Δ30-42 (Фиг.10В, b, d, f), статистически не отличались (р=0.324).
Следовательно, эти данные указывают на то, что предшествующий иммунитет к CyaA оказывает лишь очень ограниченное влияние на способность этого вектора стимулировать впоследствии эффективные ответы против конкретного чужеродного антигена.
Обсуждение
Предшествующие исследования продемонстрировали, что аденилатциклаза из бактерии Bordetella pertussis представляет собой мощный инструмент для доставки in vivo, CD4+ и CD8+ Т-клеточных эпитопов на путь презентации с главным комплексом гистосовместимости класса I и II отросчатых клеток. В экспериментальных моделях с использованием мышей эту систему применяли для запуска эффективных ответов Т-хелперов Th1 и цитотоксических Т-лимфоцитов, обеспечивающих противоопухолевую и противовирусную защиту (36). Для оценки возможности применения аденилатциклаз у людей для лечения ассоциированных с ВПЧ16 злокачественными заболеваниями шейки матки, мы продолжили исследования, чтобы продемонстрировать, что этот вектор эффективно доставляет эпитопы из белка Е7 вируса ВПЧ16.
Мы сконструировали разнообразные рекомбинантные аденилатциклазы, несущие эпитопы, содержащие полноразмерный белок из вируса ВПЧ16 или более мелкие фрагменты этого полипептида, включая, в частности, рестриктированный по H-2Db минимальный ЦТЛ-эпитоп, соответствующий остаткам 49-57. Мы показали, что эти разные рекомбинантные белки при введении путем инъекции мышам C57BL/6 способны инициировать специфические и напряженные ответы цитотоксических Т-лимфорцитов. Наши данные подтвердили, что доставка ЦТЛ-эпитопов аденилатциклазой требует полностью функционального пути презентации класса I, поскольку нам не удалось инициировать ЦТЛ после введения путем инъекции CyaA-Е7Δ30-42 мышам TAP1-/- (15). Инициирование ЦТЛ, опосредуемое аденилатциклазой, не зависело от присутствия CD4+ Т-лимфоцитов, на что указывает эффективность ответов цитотоксических Т-лимфоцитов, полученных у мышей с генотипом ГКГС Класс II-/-, что согласуется с более ранними результатами (15). Это свойство аденилатциклазы как вектора для вакцины очень важно в случае вакцинации пациентов с иммуносупрессией или иммунодефицитом, у которых изменено число CD4+ Т-лимфоцитов. Однако у CD40-/- мышей получали слабые ответы цитототоксических Т-лимфоцитов, что указывает на то, что инициация цитотоксических Т-лимфоцитов частично зависит от передачи сигнала по пути CD40. Эти наблюдения позволяют предположить, что рекомбинантные аденилатциклазы, содержащие HPV16-E7, непосредственно вызывают появление рестриктированных по ГКГС класса I цитотоксических Т-лимфоцитов путем прямой стимуляции иммунных антиген-презентирующих клеток. Тем не менее, для получения оптимальной инициации ответов ЦТЛ необходимо взаимодействие CD40 с его лигандом (CD40L).
Мы сравнили иммуногенность минимального рестриктированного по H-2Db-ЦТЛ-эпитопа белка Е7 вируса ВПЧ17 (HPV16-E7) с иммуногенностью полноразмерного белка Е7 или его части Δ30-42 Е7, которые содержат помимо, возможно, других описанный хелперный эпитоп DRAHYNIVTF (37). Инициация ЦТЛ, а равно и встречаемость HPV16-Е7-специфических спленоцитов, стимулируемых CyaA-E7Full и CyaAE-Е7Δ30-42, превосходили эти же показатели для CyaA-Е749-57, несущей лишь эпитоп для ЦТЛ Е49-57. Эти наблюдения указывают на то, что одновременная доставка эпитопов для ЦТЛ и Т-хелперов аденилатциклазой дает более устойчивый ответ цитотоксических Т-лимфоцитов. Это согласуется с опубликованными ранее данными, полученными на других моделях на доклинической (37) и клинической стадиях (38). Доставку эпитопов для Т-хелперов белка Е7 вируса ВПЧ16 рекомбинантными аденилатциклазами также подтвердил анализ цитокинов, продуцируемых HPV16-Е7-специфическими спленоцитами, подвергнутыми повторной стимуляции in vitro рекомбинантными белком HPV16-E7 с полигистидиновой меткой или пептидом Е743-77. В самом деле, мы наблюдали специфический синтез ИФН-γ только у мышей, вакцинированных рекомбинатными аденилатциклазами с HPV16-E7, содержащими эпитоп для Т-хелперов. Типичный профиль Т-хелперов Th1, характеризующийся высоким уровнем ИФН-γ и отсутствием секреции ИЛ-5, который мы наблюдали после однократной иммунизации путем внутривенной инъекции рекомбинантными аденилатциклазами CyaA-E7Full и CyaA-Е7Δ30-42 подчеркивает возможные преимущества применения этого вектора в иммунотерапии.
Это исследовали в модели отторжения опухоли, основанной на введенных подкожно туморогенных клетках ТС-1 (21). В соответствии с нашими данными, демонстрирующими иммуногенность рекомбинантных аденилатциклаз, мы наблюдали, что эти рекомбинантные белки способны стимулировать обратное развитие привитых опухолей ТС-1. CyaA-Е7Δ30-42 превосходила CyaA-Е749-57 и CyaAE5-HPV16-E7Full по выживаемости в течение периода продолжительностью 90 дней. Поскольку аденилатциклазы, несущие полноразмерный белок Е7 и Δ30-42 Е7 давали сравнимые результаты по способности инициировать цитотоксические Т-лимфоциты, встречаемости HPV16-Е749-57-специфичных спленоцитов и продуцированию ИФН-γ, мы ожидали, что CyaA-E7Full превзойдет CyaA-Е749-57 по выживаемости. По всей вероятности, большее число исследуемых животных помогло бы исключить это явное несоответствие. Однако следует обсудить два аспекта, связанных с биохимией CyaA. Во-первых, было показано, что присутствие отрицательно заряженной аминокислоты в области 224-235 аденилатциклазы CyaA ингибирует перемещение каталитического домена CyaA в цитозоль эукариотических клеток (39). В этом отношении кислый белок Е7 (pKi=4.17), возможно, препятствовал бы эффективному перемещению N-концевого домена аденилатциклазы CyaA в цизоль отросчатых клеток, CyaA-Е7Δ30-42 была специально сконструирована таким образом, чтобы удалить участок отрицательно заряженных аминокислот, расположенный между остатками 30-42 (DSSEEEDEIDGPA), чтобы облегчить доставку аденилатциклазы в отросчатые клетки (и кроме того, с каждой стороны введенного N-концевого домена Е7 вируса ВПЧ16 ввели две положительно зараженные аминокислоты (KR)). Во-вторых, Gmira et al. (25) продемонстрировали, что разворачивание гетерологичного белка, встроенного в CyaA, необходимо для проникновения рекомбинантного белка в клетки-мишени. Встраивание двух разных фрагментов полипептида Е7 в два разных пермиссивных сайта CyaA-Е7Δ30-42 может предотвратить повторное свертывание Е7 и, следовательно, должно облегчить его перемещение в клетки-мишени.
В ходе экспериментов по отторжению опухолей у некоторых мышей на поздних стадиях начался рост ВПЧ16-положительных опухолей. Это происходило уже после отторжения привитых ранее опухолей ТС-1. Анализ методом сортировки клеток с возбужденной флуоресценцией (FACS®) показал, что клетки этих опухолей не экспрессируют молекулы H-2Db. Это наблюдение заставляет рассмотреть возможность проведения поддерживающей иммунизации путем введения гомологичной рекомбинантной аденилатциклазы с целью добиться более выраженного уничтожения клеток опухоли и предотвратить рецидив опухоли по механизму ускользания от иммунитета. Учитывая данные, полученные другими группами, работающими в данной области (37, 40, 41), может иметь смысл поддерживать вакцинацию у мышей аденилатциклазой, увеличив общее количество вводимой рекомбинантной аденилатциклазы, несущей Е7 вируса ВПЧ16, до 100 мкг, т.е. до 0,56 нМоль. Эксперименты, целью которых является проверка этого последнего наблюдения, а также другие эксперименты по исследованию различных путей введения аденилатциклаз проводятся в настоящее время.
После повторного введения клеток ТС-1 выжившие мыши, иммунизированные рекомбинантными аденилатциклазами, несущими HPV16-E7, были выборочно защищены. Это коррелировало с присутствием HPV16-Е749-57-специфических CD8+ Т-лимфоцитов среди спленоцитов этих животных. Это наблюдение подтвердило тот факт, что причиной поздних рецидивов, наблюдаемых на Фиг.4, были механизмы ускользания опухоли от иммунологического контроля, а не затухание иммунитета к белку Е7. Лучшие показатели выживаемости мышей, иммунизированных рекомбинантными аденилатциклазами, содержащими эпитопы для Т-хелперов, показали, что обеспечение помощи Т-лимфоцитов против того же антигена также приводит к эффективному вторичному иммунному ответу HPV16-E749-57 CD8+ Т-лимфоцитов. В связи с этим было выдвинуто предположение, что CD4+ Т-лимфоциты, возможно, оставляют посредством CD40L уникальную молекулярную подпись на эффекторных CD8+ Т-лимфоцитах, наделяя их своей способностью к улучшенному функционированию (42).
В модели, утвержденной для исследования новых иммунотерапевтических средств для лечения новообразований, ассоциированных с ВПЧ (21), мы продемонстрировали, что аденилатциклаза является эффективным вектором для стимуляции обратного развития развитых опухолей, равно как и для обеспечения защиты против введения туморогенных клеток в течение продолжительного периода времени. В отличие от других подходов, разрабатываемых в настоящее время (11), иммунотерапия, основанная на применении аденилатциклазы, исключает необходимость выбора эпитопов, рестриктированных по HLA, поскольку в аденилатциклазу можно вводить полные белки. Этот подход также позволяет избежать применения вирусных векторов и потенциально онкогенных последовательностей ДНК вируса папилломы человека. Кроме того, мы получили наилучшие результаты, применяя аденилатциклазу, которая содержит два фрагмента белка Е7 вируса папилломы человека, встроенные в два разных пермиссивных сайта аденилатциклазы.
Тот факт, что эта последняя конструкция включает все эпитопы белка HPV16-Е7 для HLA класса I и класса II, описанные (8, 46) подтверждает выбор CyaA-Е7Δ30-42 для применения в качестве вакцины.
Исходя из представленных здесь данных мы планируем исследовать эффективность аденилатциклазы, содержащей Е7 вируса ВПЧ16, в клинических испытаниях, ориентированных на дисплазии шейки матки анальной области, ассоциированные с ВПЧ-инфекцией.
Пример 2
Целью было создание бивалентной терапевтической вакцины, направленной на белки Е7 обоих вирусов ВПЧ16 и ВПЧ18, для лечения ВПЧ16- и ВПЧ18-ассоциированных заболеваний у человека. Для этого спроектировали, сконструировали, продуцировали и очистили кандидатные вакцины, названные CyaAE5-HPV16E7Δ30-42 и CyaAE5-HPV18E7Δ32-42. Мыши HHD - это мыши, у которых инактивированы два гена: H-2D-/- β2m-/-, экспрессирующие трансген HHD, содержащий домены α1 (Н) и α2 (Н) комплекса HLA*0201, связанные с α3 трансмембранными цитоплазматическими доменами H-2Db (D), причем домен α1 связан с микроглобулином β2 человека. Таким образом, единственная молекула ГКГС 1 класса, которую экспрессируют мыши HHD, - это модифицированная молекула HLA*0201 (Pascolo et Lemonnier).
Целью этого эксперимента было продемонстрировать:
1 - что рекомбинантная аденилатциклаза способна доставлять рестриктированные по HLA-A2 эпитопы белков Е7 вирусов ВПЧ16 и ВПЧ18 Е7.
2 - не имеет место феномен иммунодоминантности одной рекомбинантной аденилатциклазы, несущей белок ВПЧ, над другой.
Для достижения этой цели мышей HHD вакцинировали, вводя им путем внутривенной инъекции 50 мкг CyaAE5-HPV16E7Δ30-42 (3 мыши), CyaAE5-HPV18E7Δ32-42 (3 мыши), либо CyaAE5-HPV16E7Δ30-42 + CyaAE5-HPV18E7Δ32-42 путем одной инъекции в заглазничную область (50 мкг в 200 мкг) (5 мышей). Через 7 дней объединенные спленоциты повторно стимулировали in vitro рестриктированными по HLA-A2 эпитопами белков Е7 из ВПЧ16 или ВПЧ18. Эти эпитопы описаны в литературе, либо их определяли при помощи программного обеспечения SYFPEITHI.
Активность цитотоксических Т-лимфоцитов анализировали через 5 дней по высвобождению 51Cr. Клетки-мишени представляли собой клетки HHD-EL4, нагруженные различными имеющими отношение к настоящему изобретению пептидами, либо без них.
Результаты показывают, что вакцинация CyaAE5-HPV16E7Δ30-42 или CyaAE5-HPV18E7Δ32-42 стимулировала специфический лизис клеток EL4-HHD, нагруженных пептидом HPV16E7 или HPV18E7, после повторной стимуляции аутологичным пептидом in vitro.
Эти результаты также демонстрируют, совместное введение (путем инъекции) CyaAE5-HPV16E7Δ30-42 и CyaAE5-HPV18E7Δ32-42 не ослабляет иммуногенность ни одной из рекомбинантных аденилатциклаз, несущих фрагменты вируса паппилломы человека, поскольку наблюдали сходные ответы на соответствующие пептиды.
Этот результат демонстрирует, что CyaAE5-HPV16E7Δ30-42 и CyaAE5-HPV18E7Δ32-42 способны стимулировать in vivo ответ цитотоксических лимфоцитов на рестриктированные по HLA-A2 человека эпитопы соответствующего белка Е7.
- Мыши
Мышей HHD не имеющих специфических патогенов выращивали в Институте Пастера. Для этого эксперимента использовали одиннадцать самцов в возрасте 6-10 недель
- Реагенты и биологический материал
- Реагенты и буферы
Среда RPMI 1640 - glutamax (invitrogen GIBCO, обр: 6187010)
Этиловый спирт 70° (Prolabo, обр: МС311631)
Пенициллин-стрептомицин (invitrogen GIBCO, обр: 15140122,)
Фетальная сыворотка крупного рогатого скота (PERBIO, обр: СН30160.03)
β меркаптоэтанол (BIO-RAD, обр: 161-0710)
Пиролизованная вода
Трипановый синий (SIGMA, Т-8154)
51Cr
Сцинциллятор Trilux Scintillant (Wallac)
- Пептиды
Для стимуляции спленоцитов in vitro перед проведением анализа высвобождения 51Cr использовали пять синтетических пептидов (Neosystem, Страсбург, Франция):
- E711-20 (YMLDLQPETT, однобуквенный код для аминокислот, #253) соответствующий рестриктированному по HLA-A2 эпитопу белка Е7 вируса ВПЧ16 (1),
- Е782-90 (LLMGTLGIV, #258), соответствующий рестриктированному по HLA-A2 эпитопу белка Е7 вируса ВПЧ16 (1),
- Е786-93 (TLGIVCPI, #255), соответствующий рестриктированному по эпитопу HLA-А2 эпитопу белка Е7 вируса ВПЧ16 (1),
- Е77-15 (TLQDIVLHL, #251), соответствующий рестриктированному по HLA-А2 эпитопу белка Е7 вируса ВПЧ18, предсказанному программным обеспечением SYFPEITHI,
- Е786-94 (FQQLFLNTL, #257), соответствующий рестриктированному по HLA-A2 эпитопу белка Е7 вируса ВПЧ18 (2).
Пептиды разбавляли до концентрации 1 мг/мл в стерильной апирогенной воде (#253, 255, 251) или в апирогенной воде с 0.1 М NaHCO3 и ацетонитрилом (50/50) (#257 и 258).
- Рекомбинантные аденилциклазы (CyaA)
В этом эксперименте исследовали две аденилатциклазы:
- CyaA-HPV16E7Δ30-42 (рекомбинантная аденилатциклаза несущая фрагмент Δ30-42 белка Е7 вируса папилломы человека 16 типа)
- CyaA-HPV18E7Δ32-42 (рекомбинантная аденилатциклаза несущая фрагмент Δ32-42 белка Е7 вируса папилломы человека 18 типа)
Эти аденилатциклазы экспрессировали в штамме BLR E.coli, используя производные плазмиды pTRACE5, кодирующие аденилатциклазу, не обладающую ферментативной активностью (CyaAE5). Конструирование плазмид, получение и очистку всех рекомбинантных белков проводили, как описано выше в Примере 1 для случая конструирования и очистки рекомбинантной аденилатциклазы В. pertussis, несущей эпитопы белка Е7 вируса ВПЧ16, но с эпитопами для данного эксперимента.
Перед введением мышам путем внутривенной инъекции исходные растворы (хранили при - 20°С), содержащие, соответственно 1.22 мг/мл и 1.33 мг/мл аденилатциклазы в 8 М мочевине размораживали и разбавляли до концентрации 250 мкг/мл фосфатным буферным раствором (Gibco BRL). Конечная концентрация мочевины лежала, таким образом, между 1.6 и 1.5 М соответственно.
- Клеточные линии
В качестве мишеней в анализе цитотоксических Т-лимфоцитов по высвобождению 51Cr использовали клетки EL4-HHD.
Эти клетки поддерживали в полной среде: RPMI 1640 с Glutamax, дополненной 10% термоинактивированной фетальной сыворотки крупного рогатого скота, 100 U/мл пенициллина, 100 мкг/мл стрептомицина и 5·10-5 М β2-меркаптоэтанола (Gibco BRL, Сержи-Понтуаз, Франция).
Методы
Иммунизация мышей
Животные Vigil иммунизировали путем одной внутривенной инъекции (в заглазничную область) (50 мкг в 200 мкг) CyaAE5-HPV16E7Δ30-42 и/или CyaAE5-HPV18E7Δ32-42. Для инъекций использовали шприцы для инсулина объемом 0.3 мл (Terumo).
Планы экспериментов
В Таблице 1 описаны кандидатные вакцины и лечение, назначенное каждой группе мышей.
- Анализ ЦТЛ
Для оценки in vitro клеточного иммунного ответа, стимулируемого иммунизацией, у умерщвленных через 7 дней после иммунизации животных (CO2) удаляли селезенку. Перед проведением анализа ЦТЛ объединяли спленоциты каждой группы.
- Стимуляция цитотоксических Т-лимфоцитов (Приготовьте 2 флакона 2 Т25 на селезенку).
- Отбор и измельчение селезенки в среде RPMI 1640-Glutamax 1% антибиотиков.
- Декантировали.
- Центрифугировали на 1200 об/мин в течение 10 минут.
- Ресуспендировали клеток в 2 мл RMPI 1640-Glutamax 1% антибиотиков - 10% фетальной сыворотки - 5·10-5М β меркаптоэтанола (полная среда: ПС).
- В каждый флакон Т25 добавляли:
10 мл ПС.
5·107 эффекторных клеток.
Соответствующий пептид в конечной концентрации 10 мкг/мл
Инкубация в течение 5 дней, не передвигая флаконы.
- Анализ цитотоксичности.
1/ Клетки-мишени:
- За день до проведения анализа клетки разбавляли 1/3 или 1/2, чтобы собрать их в фазе экспоненциального роста.
- Перенесли в 15 мл пробирки.
- Центрифугировали со скоростью 1200 об/мин в течение 10 минут.
- Ресуспендировали в 1 мл RPM11640 - 1% антибиотиков и
- Нумеровали.
- Приготовили 2 пробирки: с пептидом и без пептида.
- Ресуспендировали в 150 мкл:
пептида (50 мкмоль) в одной из двух пробирок.
100 мкКи 51Cr (50 мкл) на 3·106 клеток.
Добавили среды до объема 150 мкл.
- Инкубация в течение 1 часа в водяной бане при 37°С, при аккуратном встряхивании каждые 15 минут.
2/ Эффекторные клетки
- Удалили приблизительно 5 мл надосадочной жидкости при помощи пипетки Пастера.
- Тщательно ресуспендировали клетки пипетированием.
- Центрифугировали со скоростью 1200 об/мин в течение 10 минут.
- Ресуспендировали в 1 мл ПС.
- Довели до концентрации 107 клеток/ мл.
- Рзбавили таким образом, чтобы получить следующие отношения эффектор: мишень: 200/1, 100/1, 50/1, 25/1, 12/1, 6/1.
- Разлили по 100 мкл в лунки микротитрационного планшета с круглым дном.
- Инкубировали при 37°С в атмосфере с 7.5% CO2 до окончания приготовления клеток-мишеней.
3/ Анализ:
- Промыли клетки-мишени 10 миллилитрами RPMI 1640 - 1% антибиотиков
- Снова промыли 10 миллилитрами ПС.
- Ресуспендировали в 2 мл ПС.
- Пронумеровали.
- Довели до концентрации 105 клеток/ мл.
- Распределили по лункам планшета (100 мкл/лунка).
- Приготовили 6 лунок для спонтанного высвобождения (Довели объем до 100 мкл полной средой).
- Приготовили 6 лунок для максимального высвобождения (Добавили 100 мкл 20% Triton X-405).
- Incubate at 37°С 7.5% CO2 for 4 to 5 hours.
4/ Подсчет:
- Центрифугировали от 5 до 10 минут при 2000 об/мин.
- Поместили по 100 мкл сцинциллятора Trilux в лунки 96-луночного микротитрационного планшета.
- Отобрали 50 мкл надосадочной жидкости и перенесли в гибкий планшет.
- Закрыли планшет полимерной пленкой.
- На следующий день измерили сцинциляцию на детекторе wallac.
- HPV16E7 - Специфические ответы цитотоксических Т-лимфоцитов
Ответы цитотоксических Т-лимфоцитов, стимулированные CyaA-HPV16E7Δ30-42 показаны на Фиг.11.
- HPV18E7 - Специфические ответы цитотоксических Т-лимфоцитов
Ответы цитотоксических Т-лимфоцитов, стимулированные CyaA-HPV18E7Δ32-42 показаны на Фиг.12.
- ЗАКЛЮЧЕНИЕ
В этой модели с химерными животными HDD и CyaA-HPV16E7Δ30-42 и CyaA-HPV18E7Δ32-42 способны стимулировать специфические ответы цитотоксических Т-лимфоцитов против соответствующих рестриктированных по HLA-A2 пептидов.
Одновременное введение путем инъекции CyaA-HPV16E7Δ30-42 и CyaA-HPV18E7Δ32-42 не сказывается на способности каждой аденилатциклазы стимулировать цитотоксические Т-лимфоциты против соответствующих специфических рестриктированных по HLA-A2 пептидов. Это указывает на то, что не имеет место иммунодоминантность одной конструкции по отношению к другой. Это - ключевое наблюдение с точки зрения стратегии создания бивалентной вакцины.
Пример 3
Конструирование и иммунологическая оценка рекомбинантных аденилатциклаз, несущих белок Е7 вируса папилломы человека ВПЧ18 и белки Е7 вирусов ВПЧ16 и ВПЧ18
Карциномы аногенитального тракта составляют приблизительно 12% всех случаев рака у женщин, что делает карциному шейки матки вторым по распространенности в мире гинекологическим раковым заболеванием. Важное наблюдение, что ВПЧ-инфекция может вызывать карциному шейки матки впоследствии подтвердили эпидемиологические исследования. Самые распространенные типы ВПЧ, ассоциированные с карциномой шейки матки - это ВПЧ16 и ВПЧ18 (Распространенность 55% и 12% соответственно) {Clifford, 2003}. Чтобы охватить большую популяцию было решено создать (сконструировать) бивалентную терапевтическую вакцину, несущую Е7 вируса HPV16 и Е7 вируса HPV18. Исследовали две возможные стратегии; (i) первая состояла в том, чтобы смешивать эквимолярные количества двух рекомбинантные аденилатциклаз, одна из которых несет Е7 вируса ВПЧ16, а другая - Е7 вируса ВПЧ18; (ii) вторая стратегия состояла в том, чтобы создать рекомбинантную аденилатциклазу, несущую белки Е7 обоих вирусов ВПЧ16 и ВПЧ18. В настоящем отчете описано создание описано конструирование рекомбинантных аденилатциклаз, несущих Е7 вируса ВПЧ16 или вируса ВПЧ18, либо Е7 обоих вирусов. Иммуногенность таких конструктов оценивали в анализе цитотоксических Т-лимфоцитов.
- Идентификация эпитопов
В процессе поиска Н-2b-эпитопов последовательность аминокислот белка Е7 вируса ВПЧ18 обработали в SYFPEITHI (www.syfpeithi.de). Это программное изобретение использует алгоритм предсказания эпитопов, способных связываться с определенной молекулой главного комплекса гистосовместимости. Программа выдала возможный эпитоп, рестриктированный по молекуле H-2Db, с баллом 25. Обычно мы рассматриваем эпитопы с баллом выше 22. Затем предполагаемый пептид, IDGVNHQHL, синтезировали в компании Neosystem.
- Конструирование рекомбинантных аденилатциклаз
Схемы, изображающие стратегию, применяемую для конструирования плазмид для получения CyaA-HPV18E7, CyaA-HPV18E7Δ30-42, CyaA-HPV16+18E7, CyaA-HPV16+18ΔE7, показаны на фигурах 13 и 14.
- Анализ цитотоксических Т-лимфоцитов
Целью этого эксперимента было продемонстрировать in vivo стимуляцию цитотоксических лимфоцитов рекомбинантной аденилатциклазой, несущей либо фрагменты белка Е7 вируса ВПЧ18, либо этот белок целиком (HPV18E7), а также аденилатциклазой, несущей белки Е7 обоих вирусов HPV16 и HPV18. Эпитопами, направленными на H-2Db, являются HPV18E741-49 (IDGVNHQHL), предсказанный специальным программным обеспечением, и HPV16E749-57. Мышей C57BL/6 вакцинировали, вводя им путем внутривенной инъекции 50 мкг CyaA-CysOVA, CyaA-HPV18E7, CyaA-НРУ18Е7Δ32-42, CyaA-HPV16+18E7 или CyaA-HPV16+18ΔE7 (2 мыши в каждой группе). Через 7 объединенные спленоциты повторно стимулировали in vitro пептидами HPV18E741-49 (10 мкг/мл), OVA257-264 (1 мкг/мл) или HPV16E743-77 (1 мкг/мл). Активность цитотоксических Т-лимфоцитов оценивали спустя 5 дней по высвобождению 51Cr. Клетки-мишени представляли собой клетки EL4, нагруженные пептидом HPV18E741-49 (8 нмоль), или без него, либо клетки ТС-1. Подписи на фигурах указывают: вакцинация/повторная стимуляция. Пример: OVA/OVA относится к мышам, которых вакцинировали рекомбинантной аденилатциклазой CyaAE5-CysOVA, а затем повторно стимулировали пептидом OVA.
Эти результаты (фиг. 16) показывают, что в отличие от CyaA-HPV18E7Full, CyaA-HPV18E7Δ32-42 способна in vivo доставлять предсказанный компьютером рестриктированный по CD8+ H-2Db Т-клеток эпитоп белка HPV18-E7 в цитозоль иммунокомпетентных клеток для обработки и презентации на путь главного комплекса гистосовместимости класса I и, таким образом, вызывать напряженные ответы цитотоксических Т-лимфоцитов. Странно, что CyaA-HPV18E7Δ32-42 может инициировать ответ цитотоксических Т-лимфоцитов против HPV18E741-49, поскольку в этой рекомбинантной аденилатциклазе отсутствует первые две аминокислоты пептида HPV18E741-49. Однако в сайте встраивания фрагмента 43-105 белка HPV18E7, присутствует один аланин и один серин, что превращает предполагаемый пептид в ASGVNHQHL вместо IDGVNHQHL. При обработке этого пептида программа syfpeithi выдала балл 29 (у нативного пептида - 25). Таким образом, похоже, что, заменив две первые аминокислоты этого пептида в ходе клонирования, мы перевели его из скрытой формы в иммуногенную. Впервые эпитоп белка HPV18E7 описан применительно к H-2Db.
Несмотря на высокий уровень неспецифического фона, эти результаты также показали, что CyaA-HPV16+18E7Full и CyaA-HPV16+18Е7Δ способны in vivo доставлять рестриктированный по H-2Db Т-лимфоцитов эпитоп белка HPV16-E7 в цитозоль иммунокомпетентных клеток для обработки и презентации по пути ГКГС класса I, что вызывает напряженные ответы цитотоксических Т-лимфоцитов. CyaA-HPV16+18Е7Δ демонстрирует более высокую способность к этому по сравнению с CyaA-HPV16+18E7Full. Эти данные также впервые показали, что аденилатциклаза, несущая крупные фрагменты полипептидов (до 203 аминокислот), остается иммуногенной.
Следующий эксперимент провели для того, чтобы подтвердить описанное на фиг.16. Двух оставшихся мышей в каждой группе обрабатывали отдельно от первых. Эксперимент организован аналогичным образом. Выводы идентичны приведенным на фиг.16 за тем исключением, что вероятно, причиной высокого уровня неспецифического лизиса клеток EL4 цитотоксическими лимфоцитами, взятыми у мышей, вакцинированных CyaAE5-HPV18E7Δ32-42, является избыток пептида HPV18E741-49, остающегося во флаконах.
Также в этом случае имеет место лизис клеток ТС-1 спленоцитами, взятыми у мышей, вакцинированных CyaA-HPV16+18ΔE7, но не CyaA-HPV16+18E7. В этом эксперименте максимум неспецифического фона приходится на 40%. Тем не менее, вывод, который можно сделать из фигуры 17, подтверждает выводы фигуры 16, а именно:
- CyaA-HPV18E7Δ32-42 способна in vivo доставлять предсказанный компьютером рестриктированный по H-2Db CD8+ Т-лимфоцитов эпитоп белка HPV18-E7 в цитозоль иммунокомпетентных клеток для обработки и презентации по пути ГКГС класса I, что вызывает напряженные ответы цитотоксических Т-лимфоцитов. Впервые эпитоп белка HPV18E7 описан применительно к H-2Db.
Рекомбинантная аденилатциклаза, несущая крупные фрагменты полипептидов (до 203 аминокислот), остается иммуногенной.
- Заключение
Было решено создать бивалентную терапевтическую вакцину, несущую Е7 вируса ВПЧ16 и вируса ВПЧ18. Были рассмотрены две возможные стратегии; (i) первая состояла в том, чтобы смешивать эквимолярные количества двух рекомбинантные аденилатциклаз, одна из которых несет Е7 вируса ВПЧ16, а другая - Е7 вируса ВПЧ18: (ii) вторая стратегия состояла в том, чтобы создать рекомбинантную аденилатциклазу, несущую белки Е7 обоих вирусов ВПЧ16 и ВПЧ18. Приведенные в настоящей заявке результаты описывают конструирование рекомбинантных аденилатциклаз, несущих Е7 вируса ВПЧ16 или вируса ВПЧ18, либо Е7 обоих вирусов. Иммуногенность таких конструктов оценивали в анализе цитотоксических Т-лимфоцитов.
Главное заключение этого исследования: обе стратегии являются приемлемыми, поскольку рекомбинантная аденилатциклаза, несущая более мелкие фрагменты белка Е7 вируса ВПЧ18 эффективна, т.к. она впервые демаскировала скрытый (криптический) рестиктированный по H-2Db эпитоп в последовательности белка Е7 вируса. Кроме того, весьма интересно, что рекомбинантные аденилатциклазы, несущие белки Е7 (или более мелкие фрагменты) обоих вирусов ВПЧ16 и ВПЧ18 оставались иммуногенными, т.к. они проявляли способность инициировать ответ цитотоксических Т-лимфоцитов против рестриктированных по H-2Db эпитопов белка Е7, присутствующих в норме в клетках ТС-1.
Ссылки
1. Zur Hausen, H. Papillomaviruses and cancer: from basic studies to clinical application (Папилломавирусы человека: от базовых исследований к клиническому применению). Nat Rev Cancer, 2: 342-350, 2002.
2. Clifford, G.M., Smith, J.S., Plummer, M., Munoz, N., and Franceschi, S. Human papillomavirus types in invasive cervical cancer worldwide: a meta-analysis (Типы вирусов папилломы человека при инвазивном раке шейки матки (по всему миру): мета-анализ). Br J Cancer, 88: 63-73, 2003.
3. Frazer, I.H. Prevention of cervical cancer through papillomavirus vaccination (Предотвращение рака шейки матки посредством вакцинации папилломавирусом). Nat Rev Immunol, 4: 46-55, 2004.
4. de Gruijl, T.D., Bontkes, H.J., Walboomers, J.M., Stukart, M.J., Doekhie, F.S., Remmink, A.J., Helmerhorst, T.J., Verheijen, R.H., Duggan-Keen, M.F., Stern, P.L., Meijer, C.J., and Scheper, R.J. Differential T helper cell responses to human papillomavirus type 16 E7 related to viral clearance or persistence in patients with cervical neoplasia: a longitudinal study (Дифференциальные ответы Т-хелперов на E7 Вируса Папилломы Человека типа 16, связанные с клиренсом или персистеированием вируса у пациентов с цервикальными новообразованиями: продолжительное исследование). Cancer Res, 58; 1700-1706, 1998.
5. Evans, E.M., Man, S., Evans, A.S., and Borysiewicz, L.K. Infiltration of cervical cancer tissue with human papillomavirus-specific cytotoxic T-lymphocytes (Инфильтрация ткани опухоли шейки матки ПВЧ-специфическими цитотоксическими Т-лимфоцитами). Cancer Res, 57: 2943-2950, 1997.
6. Nimako, M., Fiander, A.N., Wilkinson, G.W., Borysiewicz, L.K., and Man, S. Human papillomavirus-specific cytotoxic T lymphocytes in patients with cervical intraepithelial neoplasia grade III (ПВЧ-специфические цитотоксические лимфоциты у пациентов с цервикальной интраэпителиальной неоплазией III степени). Cancer Res, 57: 4855-4861, 1997.
7. Ressing, M.E., van Driel, W.J., Celis, E., Sette, A., Brandt, M.P., Hartman, M., Anholts, J.D., Schreuder, G.M., ter Harmsel, W.В., Fleuren, G.J., Trimbos, B.J., Kast, W.M., and Melief, C.J. Occasional memory cytotoxic T-cell responses of patients with human papillomavirus type 16-positive cervical lesions against a human leukocyte antigen-A *0201-restricted E7-encoded epitope (Атипичный ответ цитотоксических Т-лимфоцитов памяти у пациентов с ПВЧ16-положительными поражениями шейки матки против рестриктированного по антигену-А *0201 Е7-кодируемого эпитопа лейкоцитов человека). Cancer Res, 56: 582-588, 1996.
8. van der Burg, S.H., Ressing, M.E., Kwappenberg, K.M., de Jong, A., Straathof, K., de Jong, J., Geluk, A., van Meijgaarden, K.E., Franken, K.L., Ottenhoff, T.H., Fleuren, G.J., Kenter, G., Melief, C.J., and Offringa, R. Natural T-helper immunity against human papillomavirus type 16 (HPV16) E7-derived peptide epitopes in patients with HPV16-positive cervical lesions: identification of 3 human leukocyte antigen class ll-restricted epitopes (Иммунитет естественных Т-хелперов против эпитопов, полученных из пептида Е7 вируса папилломы человека 16 (ВПЧ16): идентификация трех эпитопов, рестриктированных по антигенов лейкоцитов человека класса II). Int J Cancer, 91: 612-618, 2001.
9. Schreurs, M.W., Scholten, К.В., Kueter, E.W., Ruizendaal, J.J., Meijer, C.J., and Hooijberg, E. In vitro generation and life span extension of human papillomavirus type 16-specific, healthy donor-derived CTL clones (Наработка и продление клеточного цикла клонов ВПЧ16-специфических цитотоксических Т-лимфоцитов, полученных от здоровых доноров, in vitro). J Immunol, 171: 2912-2921, 2003.
10. Welters, M.J., de Jong, A., van den Eeden, S.J., van der Hulst, J.M., Kwappenberg, K.M., Hassane, S., Franken, K.L., Drijfhout, J.W., Fleuren, G.J., Kenter, G., Melief, C.J., Offringa, R., and van der Burg, S.H. Frequent display of human papillomavirus type 16 Е6-specific memory T-Helper cells in the healthy population as witness of previous viral encounter (Многократное обнаружение специфичных к Е7 вируса папилломы человека 16 Т-хелперных клеток памяти в здоровой популяции как свидетельство предшествующих случаев заражения вирусами). Cancer Res, 63: 636-641, 2003.
11. Roden, R. and Wu, Т.С. Preventative and therapeutic vaccines for cervical cancer (Профилактические и терапевтические вакцины рака шейки матки). Expert Rev Vaccines, 2: 495-516, 2003.
12. Ladant, D. and Ullmann, A. Bordetella pertussis adenylate cyclase: a toxin with multiple talents (Тоскин аденилатциклаза Bordetella pertussis: тосин с многочисленными талантами). Trends Microbiol, 7: 172-176, 1999.
13. Morón, G., Dadaglio, G., and Leclerc, С. New tools for antigen delivery to the MHC class I pathway (Новые средства доставки антигенов по пути ГКГС клесса I). Trends in Immunology, 25: 92-97, 2004.
14. Guermonprez, P., Khelef, N., Blouin, E., Rieu, P., Ricciardi-Castagnoli, P., Guiso, N., Ladant, D., and Leclerc, С. The adenylate cyclase toxin of Bordetella pertussis binds to target cells via the alpha(M)beta(2) integrin (CD11b/CD18) (токсина аденилатциклаза бектерии Bordetella pertussis связывается с клетками-мишенями через альфа(М)бета(2) интегрин (CD11b/CD18)). J Exp Med, 193: 1035-1044, 2001.
15. Guermonprez, P., Fayolle, С., Rojas, M.J., Rescigno, M., Ladant, D., and Leclerc, С. In vivo receptor-mediated delivery of a recombinant invasive bacterial toxoid to CD11c + CD8 alpha - CD11bhigh dendritic cells (Рецептор-опосредованная доставка рекомбинантного инвазивного обезвреженного бектериального токсина к CD11c + CD8альфа - CD11bвыс. отросчатым клеткам In vivo). Eur J Immunol, 32: 3071-3081, 2002.
16. Fayolle, С., Ladant, D., Karimova, G., Ullmann, A., and Leclerc, С.Therapy of murine tumors with recombinant Bordetella pertussis adenylate cyclase carrying a cytotoxic Т cell epitope (Лечение мышиных опухолей рекомбинантной аденилатциклазой бектерии Bordetella pertussis, несущей эпитоп цитотоксических Т-лимфоцитов). J Immunol, 162: 4157-4162, 1999.
17. Fayolle, С., Osickova, A., Osicka, R., Henry, Т., Rojas, M. J., Saron, M.F., Sebo, P., and Leclerc, С.Delivery of multiple epitopes by recombinant detoxified adenylate cyclase of Bordetella pertussis induces protective antiviral immunity (Доставка множественных эпитопов обезвреженной аденилатциклазой бектерии Bordetella pertussis стимулирует зафитный противовирусный иммунитет). J Virol, 75: 7330-7338, 2001.
18. Van Kaer, L., Ashton-Rickardt, P.G., Ploegh, H.L., and Tonegawa, S. TAP1 mutant mice are deficient in antigen presentation, surface class I molecules, and CD4-8 + Т cells (У мышей, мутантных по TAP1 отсутствует презентация антигенов, поверхностные молекулы класса I и CD4-8 + Т-лимфоциты). Cell, 71: 1205-1214, 1992.
19. Madsen, L., Labrecque, N., Engberg, J., Dierich, A., Svejgaard, A., Benoist, C., Mathis, D., and Fugger, L. Mice lacking all conventional MHC class II genes (Мыши, у которых отсутствуют все обычные гены ГКГС класса II). Proc Natl Acad Sci USA, 96: 10338-10343, 1999.
20. Kawabe, Т., Naka, Т., Yoshida, К., Tanaka, Т., Fujiwara, H., Suematsu, S., Yoshida, N., Kishimoto, Т., and Kikutani, H. The immune responses in CD40-deficient mice: impaired immunoglobulin class switching and germinal center formation (Иммунные ответы у CD40-дефицитных мышей: нарушенное переключение классов иммуноглобулинов и формирование зародышевого центра). Immunity, 1: 167-178, 1994.
21. Lin, К.Y., Guarnieri, F.G., Staveley-O'Carroll, K.F., Levitsky, H.I., August, J.Т., Pardoll, D.M., and Wu, T.C. Treatment of established tumors with a novel vaccine that enhances major histocompatibility class II presentation of tumor antigen (Лечение развитых опухолей новой вакциной, усиливает презентацию опухолевых антигенов рецепторами ГКГС класса II). Cancer Res, 56: 21-26, 1996.
22. Feltkamp, M.C., Smits, H.L., Vierboom, M.P., Minnaar, R.P., de Jongh, B.M., Drijfhout, J.W., ter Schegget, J., Melief, C.J., and Kast, W.M. Vaccination with cytotoxic T lymphocyte epitope-containing peptide protects against a tumor induced by human papillomavirus type 16-transformed cells (Вакцинация пептидном, содержащим эпитопы цитотоксических Т-лимфоцитов защищает от опухолей, индуцируемых клетками, трансформированными вирусами папилломы человека типа 16). Eur J Immunol, 23: 2242-2249, 1993.
23. Tindle, R.W., Fernando, G.J., Sterling, J.C., and Frazer, I.H. A "public" T-helper epitope of the E7 transforming protein of human papillomavirus 16 provides cognate help for several E7 B-cell epitopes from cervical cancer-associated human papillomavirus genotypes ("Общий" эпитоп Т-хелперов трансформирующего белка E7 вируса папилломы человека обеспечивает когнатную помощь для нескольких эпитопов Е7 для В-клеток ассоциированных с раком шейки матки вирусов папилломы человека). Proc Natl Acad Sci USA, 88: 5887-5891, 1991.
24. Dadaglio, G., Morel, S., Bauche, С., Moukrim, Z., Lemonnier, F.A., Van Den Eynde, B.J., Ladant, D., and Leclerc, C. Recombinant adenylate cyclase toxin of Bordetella pertussis induces cytotoxic Т lymphocyte responses against HLA*0201 - restricted melanoma epitopes (Рекомбинантная аденилатциклаза Bordetella pertussis стимулирует ответ цитотоксических Т-лимфоцитов против рестриктированных по HLA*0201 эпитопов меланомы). Int Immunol, 15: 1423-1430, 2003.
25. Gmira, S., Karimova, G., and Ladant, D. Characterization of recombinant Bordetella pertussis adenylate cyclase toxins carrying passenger proteins (Описания рекомбинантных токсинов Bordetella pertussis аденилатциклазы, несущей белки-пассажиры). Res Microbiol, 152: 889-900, 2001.
26. Guermonprez, P., Fayolle, C., Karimova, G., Ullmann, A., Leclerc, C., and Ladant, D. Bordetella pertussis adenylate cyclase toxin: a vehicle to deliver CD8-positive T-cell epitopes into antigen-presenting cells (Токсин аденилатциклаза бактерии Bordetella pertussis: транспортировщик для доставки эпитопов СD8-положительных Т-клеток в антиген-презентирующие клетки). Methods Enzymol, 326: 527-542, 2000.
27. Franken, К.L., Hiemstra, H.S., van Meijgaarden, K.E., Subronto, Y., den Hartigh, J., Ottenhoff, T.H., and Drijfhout, J.W. Purification of his-tagged proteins by immobilized chelate affinity chromatography: the benefits from the use of organic solvent (Очистка меченых гистидином белков при помощи тфердофазной хелат-аффинной хроматографии: преимущества использования органического растворителя). Protein Expr Purif, 18: 95-99, 2000.
28. Dadaglio, G., Moukrim, Z., Lo-Man, R., Sheshko, V., Sebo, P., and Leclerc, C. Induction of a polarized Th1 response by insertion of multiple copies of a viral T-cell epitope into adenylate cyclase of Bordetella pertussis (Стимуляция направленного ответа Th1-хелперов путем введения множественных копий вирусного Т-клеточного эжпитопа в аденилатциклазу Bordetella pertussis). Infect Immun, 68: 3867-3872, 2000.
29. Michallet, M.С., Preville, X., Flacher, M., Fournel, S., Genestier, L., and Revillard, J.P. Functional antibodies to leukocyte adhesion molecules in antithymocyte globulins (Функциональные антитела к адгезионным молекулам лейкоцитов в глобулинах антитимоцитов). Transplantation, 75: 657-662, 2003.
30. Siegel, S. and Castellan, N.J. Non parametric statistics for the behavioral sciences (Непараметрическая статистика в бихевиоризме). New York: McGraw-Hill, 1988.
31. Sebo, P., Moukrim, Z., Kalhous, M., Schaft, N., Dadaglio, G., Sheshko, V., Fayolle, C., and Leclerc, C. In vivo induction of CTL responses by recombinant adenylate cyclase of Bordetella pertussis carrying multiple copies of a viral CD8(+) T-cell epitope (Стимуляция ответа цитотоксических Т-лимфоцитов рекомбинантной аденилатциклазой Bordetella pertussis, несущей множественные копии вирусного CD8(+) Т-клеточного эпитопа). FEMS Immunol Med Microbiol, 26: 167-173, 1999.
32. Matsui, S., Ahlers, J.D., Vortmeyer, A.O., Terabe, M., Tsukui, Т., Carbone, D.P., Liotta, L.A., and Berzofsky, J.A. A model for CD8 + CTL tumor immunosurveillance and regulation of tumor escape by CD4 Т cells through an effect on quality of CTL (Модель иммунного надзора опухолей CD8+ цитотоксическими Т-лимфоцитами и регуляции ускользания опухолей CD4 Т-клетками посредством воздействия на свойства цитотоксических Т-лимфоцитов). J Immunol, 163: 184-193, 1999.
33. Romagnani, S. Th1 and Th2 in human diseases (Ролье хелперов Th1 и Th2 в заболеваниях человека). Clin Immunol Immunopathol, 80: 225-235, 1996.
34. Greenstone, H.L., Nieland, J.D., de Visser, K.E., De Bruijn, M.L., Kirnbauer, R., Roden, R.В., Lowy, D.R., Kast, W.M., and Schiller, J.T. Chimeric papillomavirus virus-like particles elicit antitumor immunity against the E7 oncoprotein in an HPV16 tumor model (Химерные ПВЧ-подобные частицы вызывают противоопухолевый иммунитет против онкобелка E7 в модели опухоли, вызванной ВПЧ16). Proc Natl Acad Sci USA, 95: 1800-1805, 1998.
35. De Bruijn, M.L., Schuurhuis, D.H., Vierboom, M.P., Vermeulen, H., de Cock, K.A., Ooms, M.E., Ressing, M.E., Toebes, M., Franken, K.L., Drijfhout, J.W., Ottenhoff, T.H., Offringa, R., and Melief, C.J. Immunization with human papillomavirus type 16 (HPV16) oncoprotein-loaded dendritic cells as well as protein in adjuvant induces MHC class l-restricted protection to HPV16-induced tumor cells (Иммунизация отросчатыми клетками, "нагруженными" вирусам папилломы человека (HPV16), а также белком в адъюванте стимулирует рестриктированную по ГКГС класс I защиту от ВПЧ16-стримулированных опухолевых клеток). Cancer Res, 58: 724-731, 1998.
36. El Azami El Idrissi, M., Ladant, D., and Leclerc, C. The adenylate cyclase of Bordetella pertussis: a vector to target antigen presenting cells (Аденилатциклаза бактерии Bordetella pertussis: вектор для нацеливания на антиген-презентирующие клетки). Toxicon, 40: 1661-1665, 2002.
37. Zwaveling, S., Ferreira Mota, S.C., Nouta, J., Johnson, M., Lipford, G.В., Offringa, R., van der Burg, S.H., and Melief, C.J. Established human papillomavirus type 16-expressing tumors are effectively eradicated following vaccination with long peptides. J Immunol, 169: 350-358, 2002.
38. Knutson, К.L., Schiffman, К., and Disis, M.L. Immunization with a HER-2/neu helper peptide vaccine generates HER-2/neu CD8 T-cell immunity in cancer patients. J Clin Invest, 107: 477-484, 2001.
39. Karimova, G., Fayolle, C., Gmira, S., Ullmann, A., Leclerc, C., and Ladant, D. Charge-dependent translocation of Bordetella pertussis adenylate cyclase toxin into eukaryotic cells: implication for the in vivo delivery of CD8(+) Т cell epitopes into antigen-presenting cells. Proc Natl Acad Sci USA, 95: 12532-12537, 1998.
40. van der Burg, S.H., Kwappenberg, K.M., O'Neill, Т., Brandt, R.M., Melief, C.J., Hickling, J.K., and Offringa, R. Pre-clinical safety and efficacy of TA-CIN, a recombinant HPV16 L2E6E7 fusion protein vaccine, in homologous and heterologous prime-boost regimens. Vaccine, 19: 3652-3660, 2001.
41. Chu, N.R., Wu, H.В., Wu, Т., Boux, L.J., Siegel, M.I., and Mizzen, L.A. Immunotherapy of a human papillomavirus (HPV) type 16 E7-expressing tumour by administration of fusion protein comprising Mycobacterium bovis bacille Calmette-Guerin (BCG) hsp65 and HPV16 E7. Clin Exp Immunol, 121: 216-225, 2000.
42. Bourgeois, C., Rocha, В., and Tanchot, C. A role for CD40 expression on CD8+ Т cells in the generation of CD8+ Т cell memory. Science, 297: 2060-2063, 2002.
43. Glaser, P. et al, 1988, The calmodulin-sensitive adenylate cyclase of Bordetella pertussis: cloning and expression in E.coli Molecular Microbiology 2(1), 19-30.
44. Sebo P. et al, 1995, Cell-invasive activity of epitope-tagged adenylate cyclase of Bordetella pertussis allows in vitro presentation of a foreign epitope to CD8+ cytotoxic Т cells. Infection and Immunity, p.3851-3857.
45. EI-Azami-EI-Idrissi M. et al, 2003 October 3, Interaction of Bordetella pertussis adenylate cyclase with CD11b/CD18: Role of toxin acylation and identification of the main integrin interaction domain. J. Biol. Chem. 278 (40): 38514-21.
46. Kast, W.M. et al, 1994, Role of HLA-A motifs in identification of potential CTL epitopes in human papillomavirus type 16 E6 and E7 proteins, J. Immunol, 152: 3904-3912.
47. Courret N, Lang T, Milon G, Antoine JC. Intradermal inoculations of low doses of Leishmania major and Leishmania amazonensis metacyclic promastigotes induce different immunoparasitic processes and status of protection in BALB/c mice. Int J Parasitol 2003; 33: 1373-83.
48. Chu NR, Wu HB, Wu T, Boux LJ, Siegel MI, Mizzen LA. Immunotherapy of a human papillomavirus (HPV) type 16 E7-expressing tumour by administration of fusion protein comprising Mycobacterium bovis bacille Calmette-Guerin (BCG) hsp65 and HPV16 E7. Clin Exp Immunol 2000; 121: 216-25.

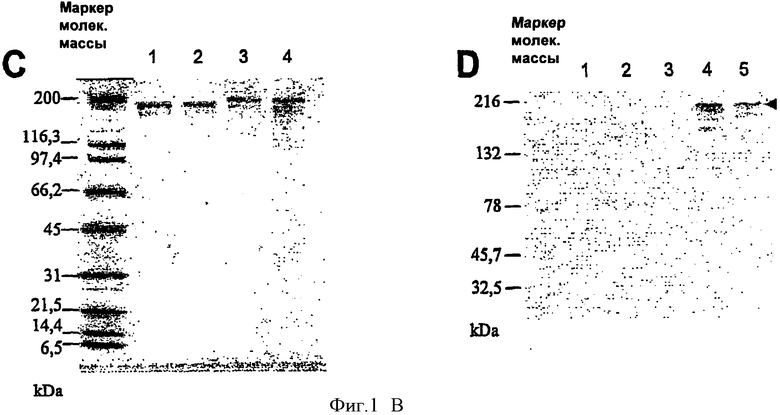
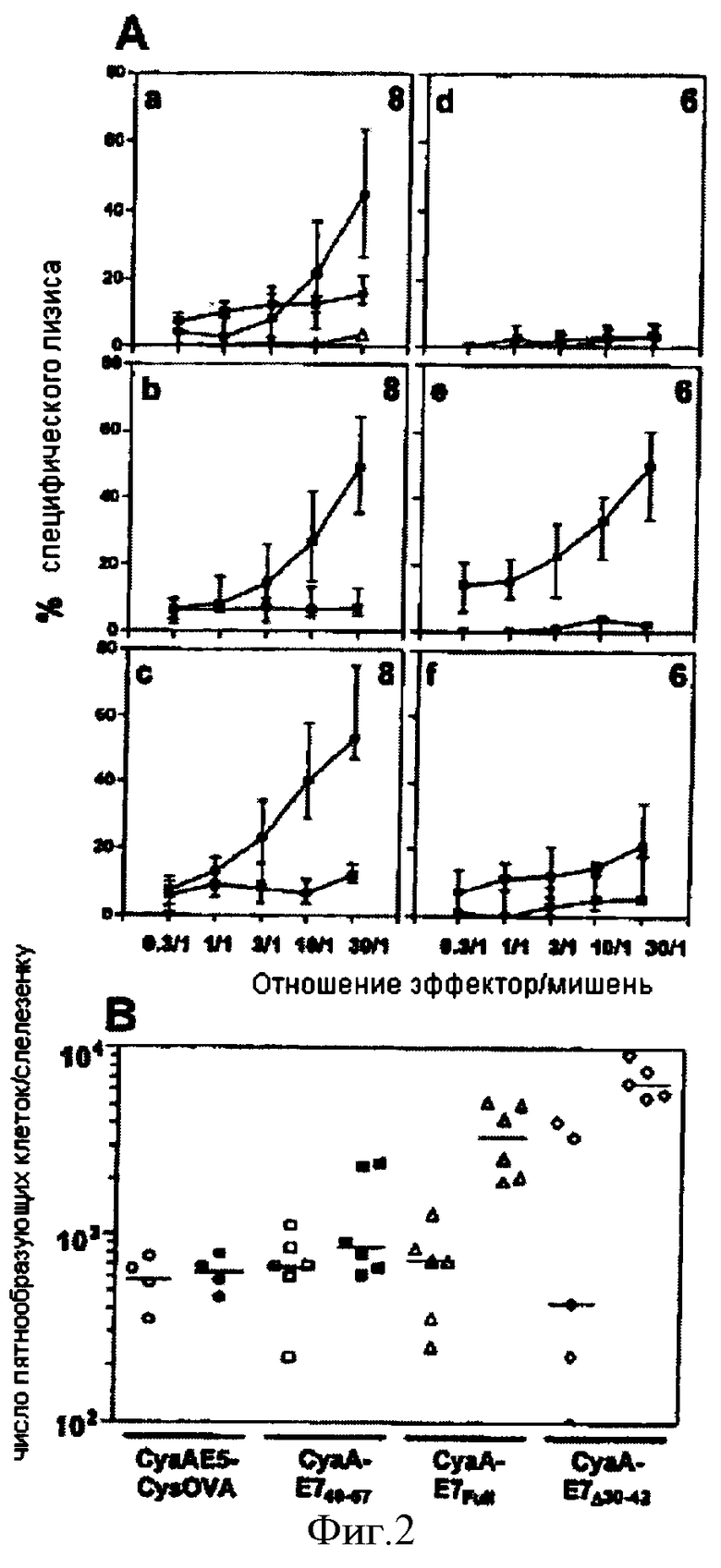
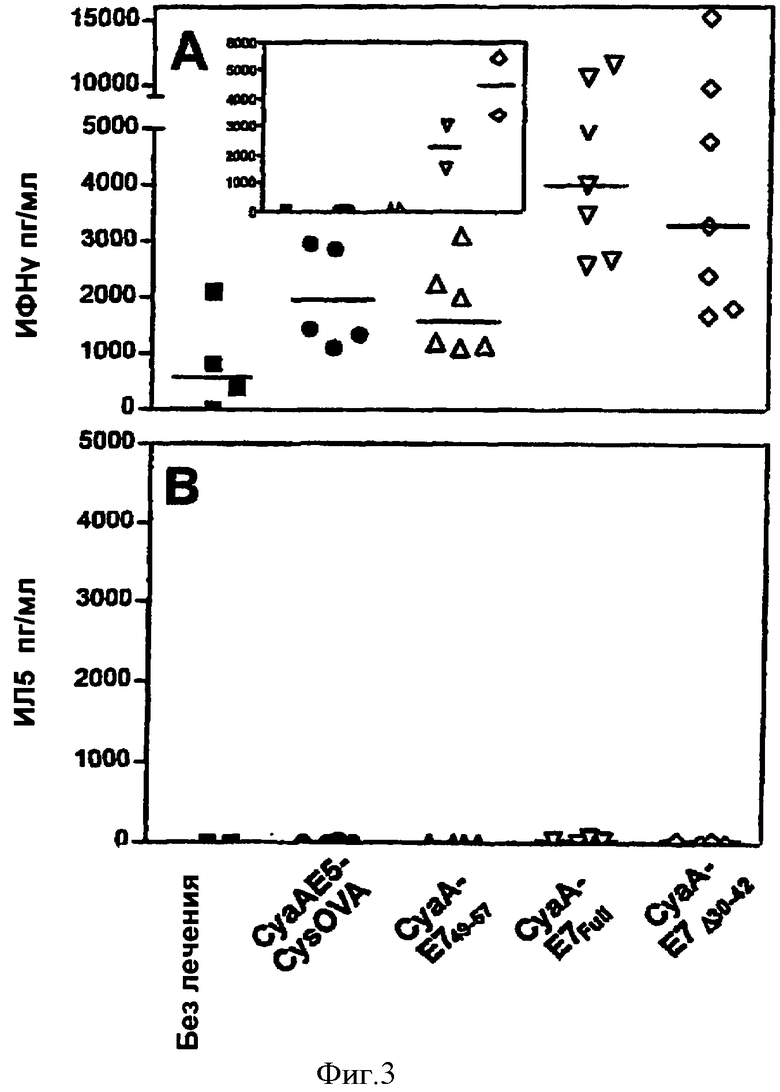
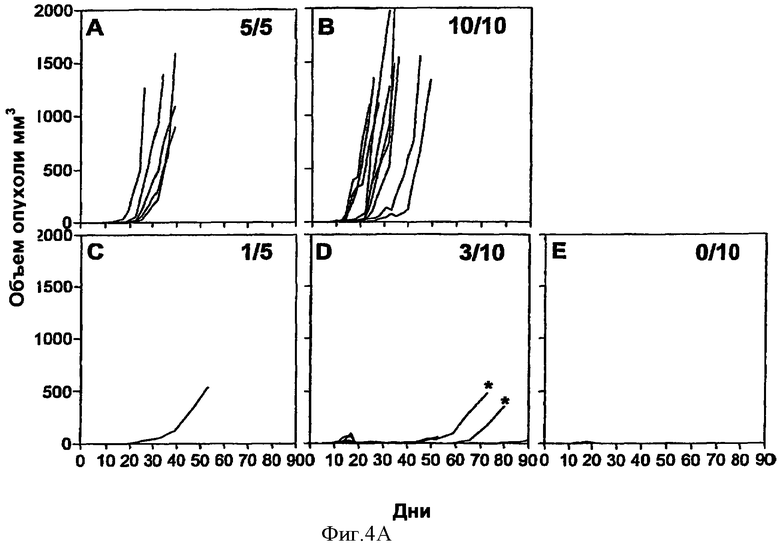
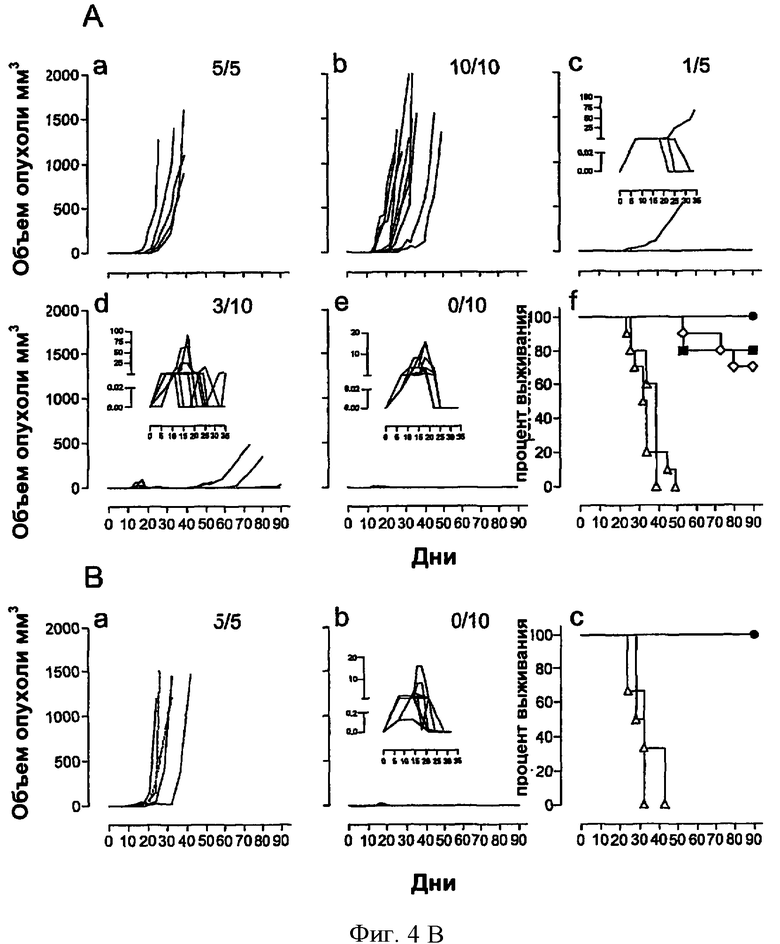
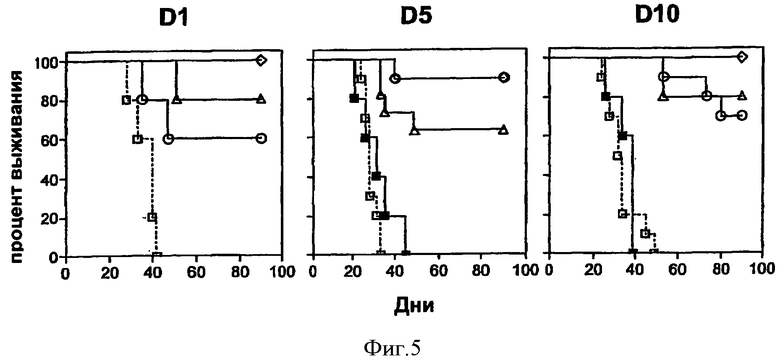
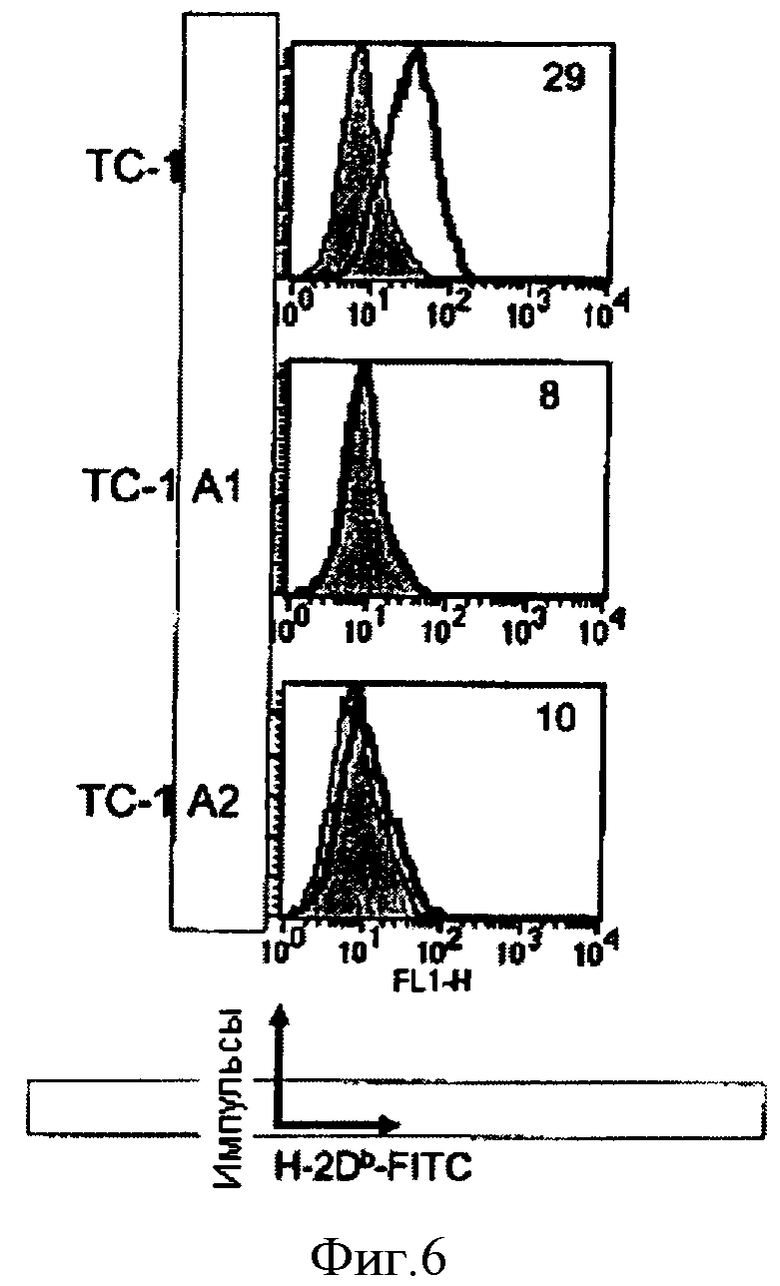
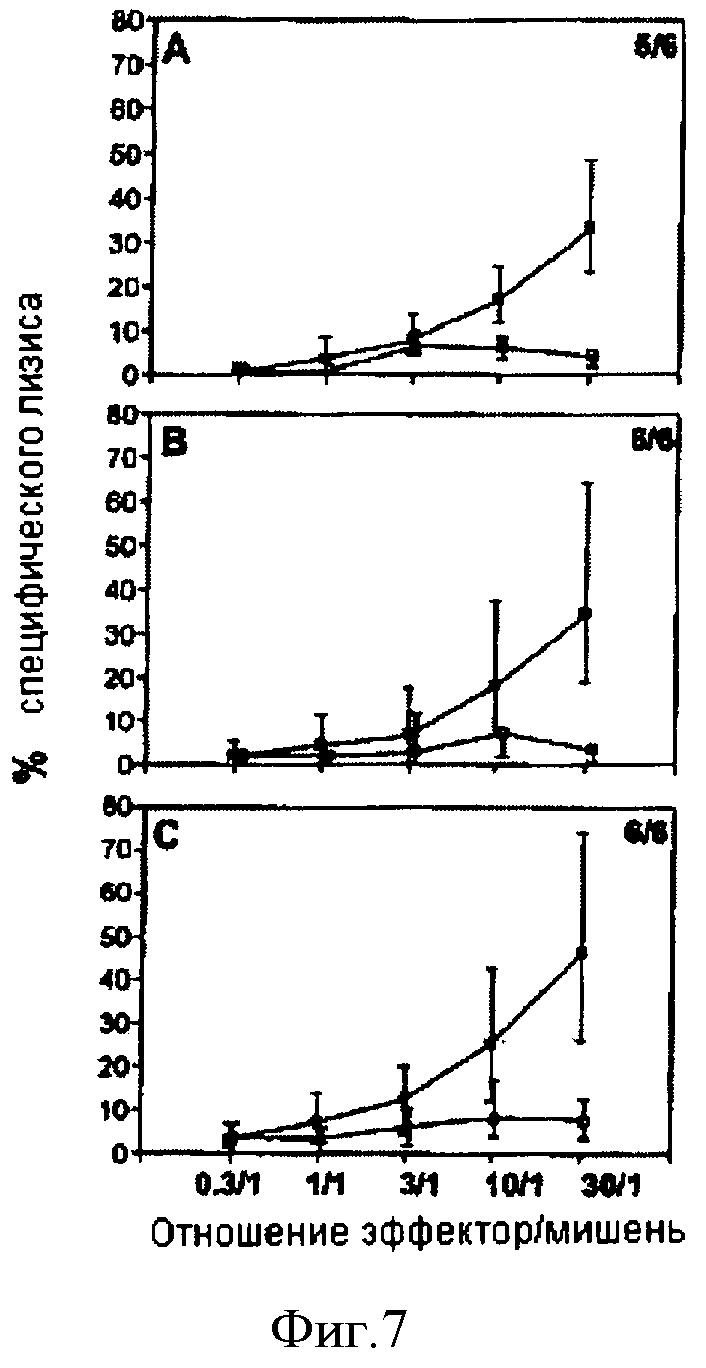
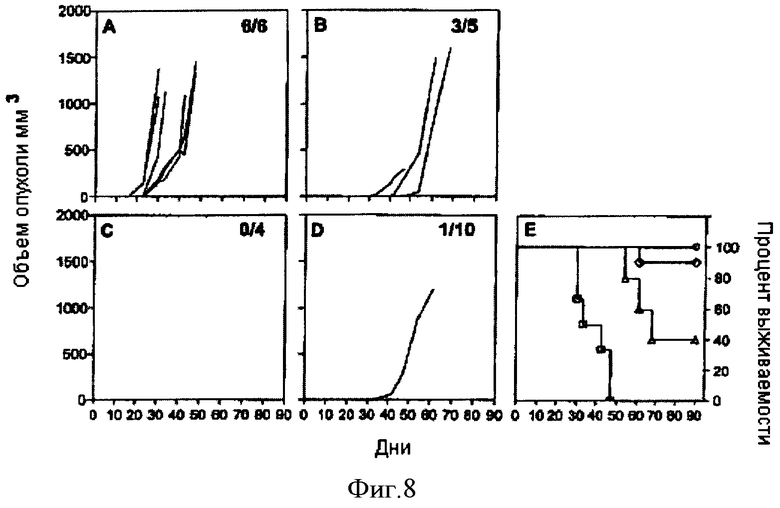

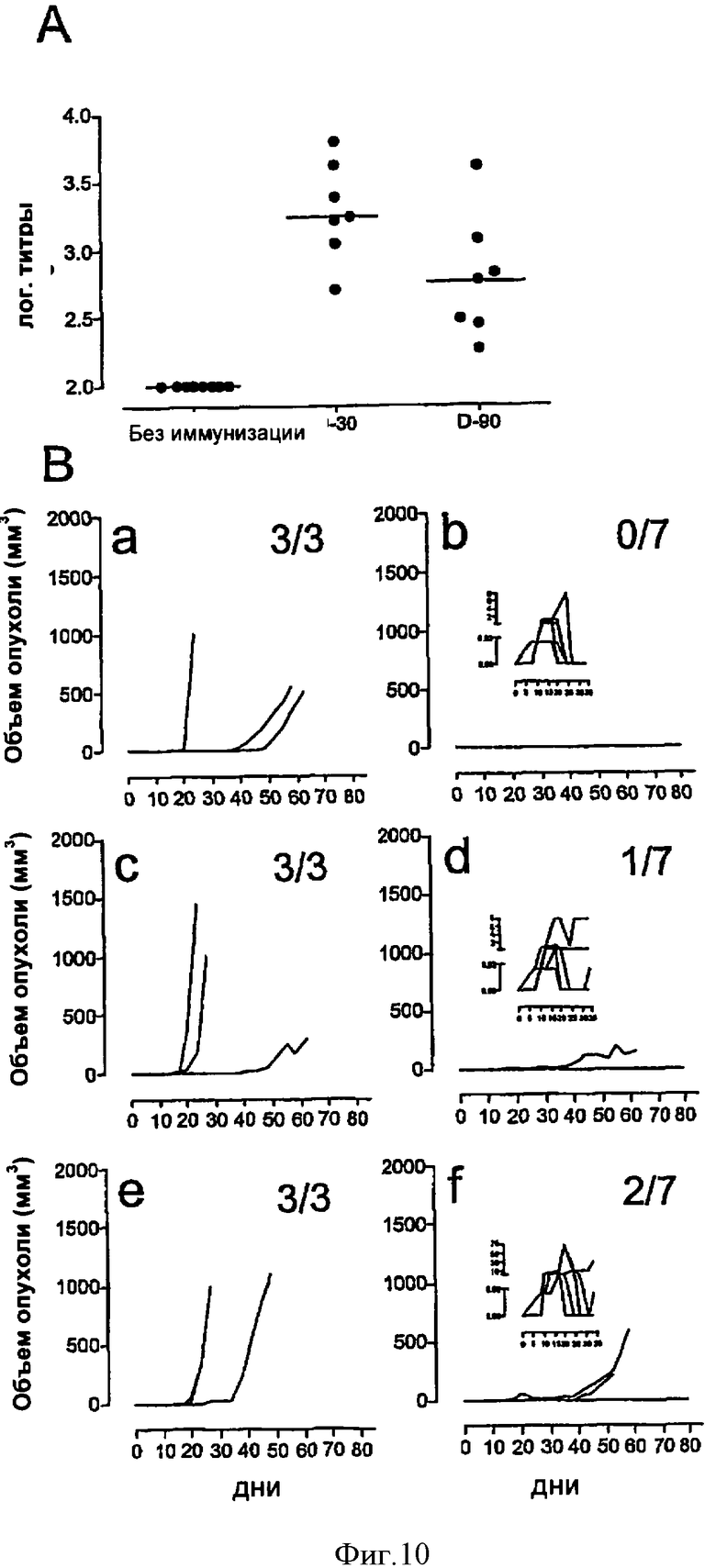
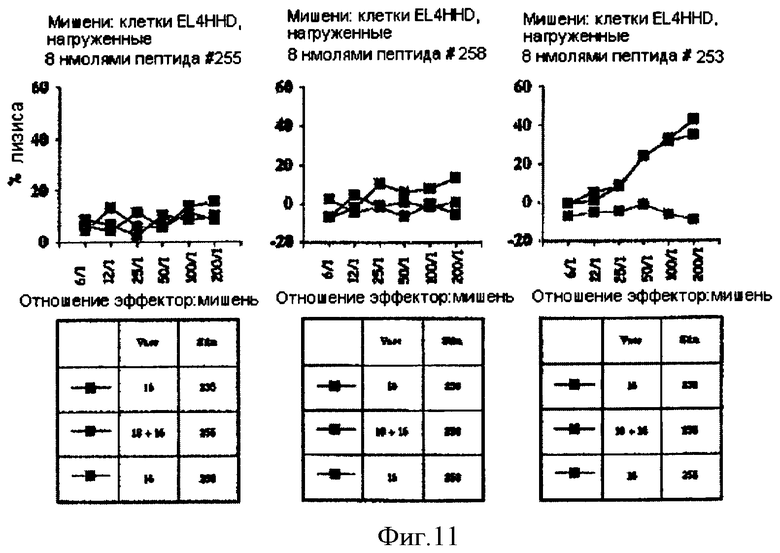
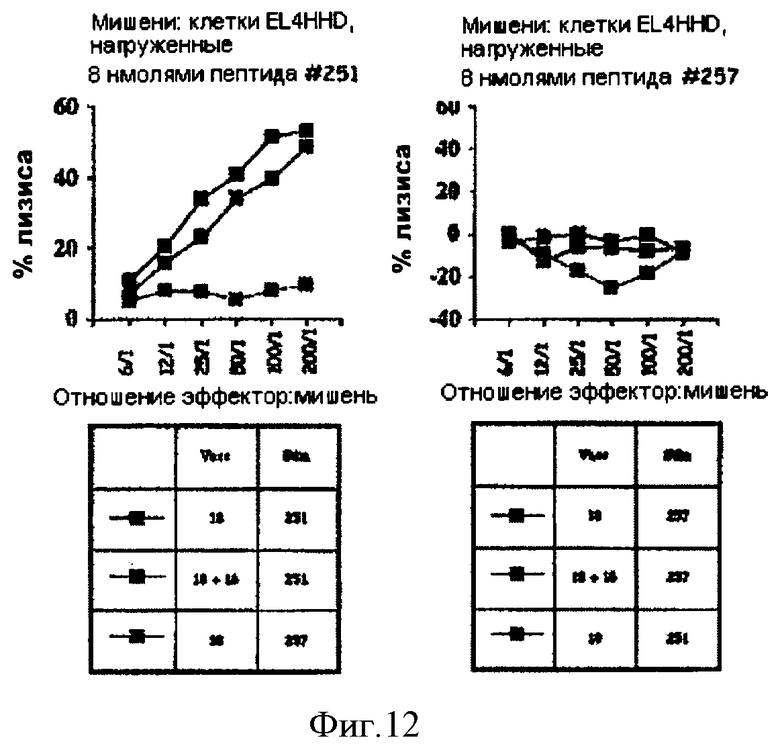

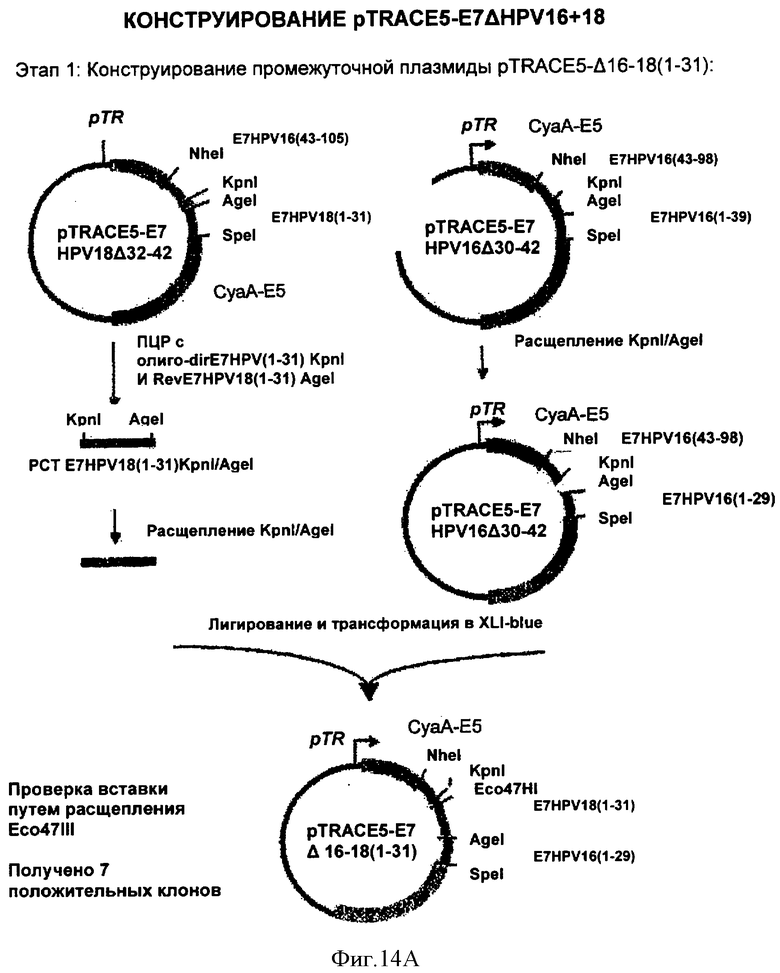
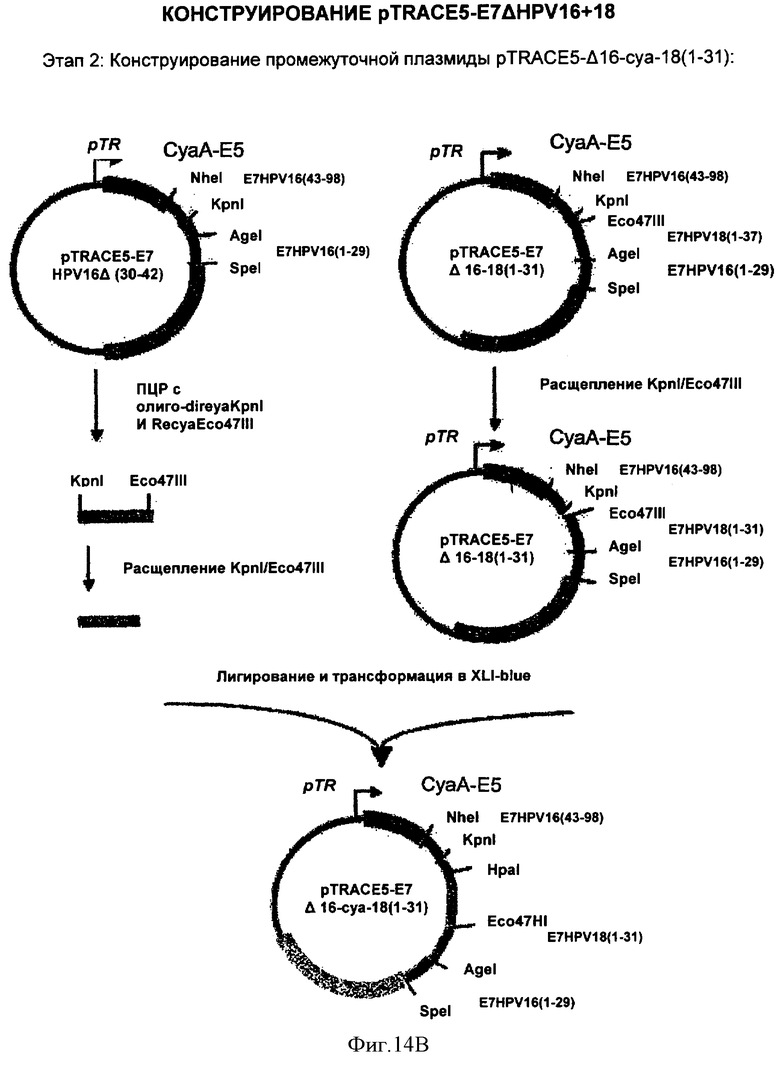
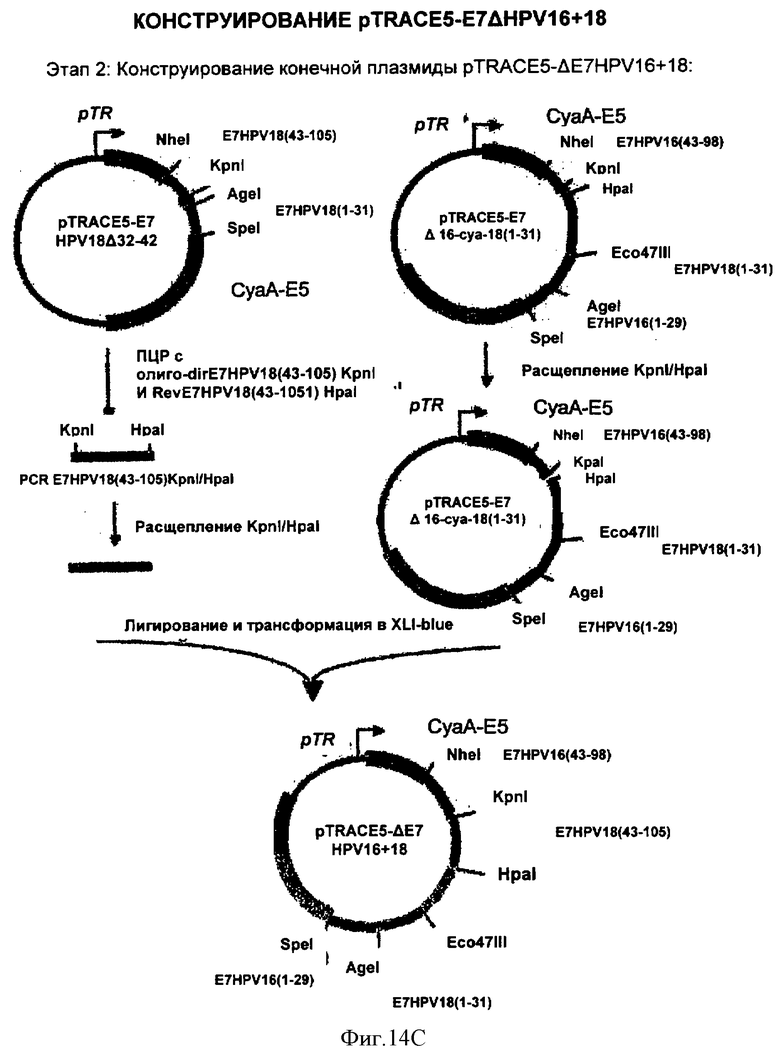

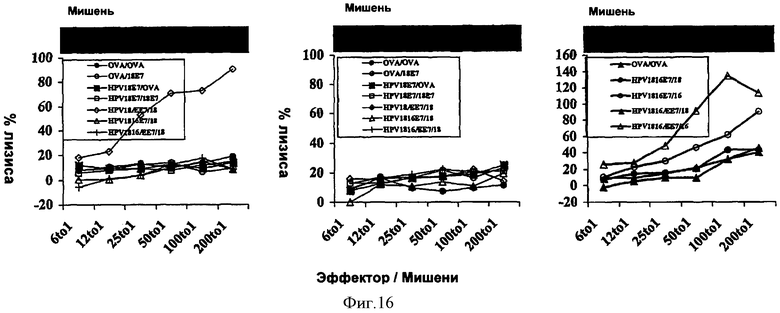
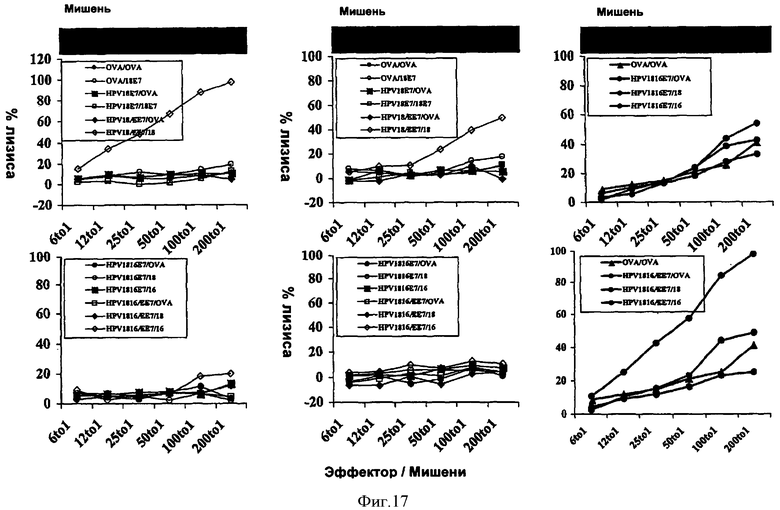
Изобретение относится к области биотехнологии, вирусологии и медицины. Описан рекомбинантный белок, содержащий один или несколько полипептидов, несущих один или несколько эпитопов одного или нескольких антигенов вируса папилломы человека ВПЧ. При этом указанные полипептиды встроены в один или в различные пермиссивные сайты белка аденилатциклазы (CyaA) или его фрагмента. Указанный фрагмент CyAa сохраняет свойство указанного белка аденилатциклазы направленно взаимодействовать с антиген-презентирующими клетками. Также описан полинуклеотид, кодирующий такой белок, и их использование в системах экспрессии для получения иммуногенных композиций и лекарственных средств. Изобретение может быть использовано в медицине. 17 н. и 43 з.п. ф-лы, 21 ил., 1 табл.
1. Рекомбинантный белок для лечения инфекции, вызванной вирусом папилломы человека (ВПЧ), или для профилактики или лечения злокачественных эффектов, возникающих при инфицировании ВПЧ, содержащий один или несколько полипептидов, несущих один или несколько эпитопов одного или нескольких антигенов ВПЧ, причем указанные полипептиды встроены в один или разные пермиссивные сайты белка аденилатциклазы CyaA одного из видов Bordetella или его фрагмента, и при этом указанные фрагменты аденилатциклазы сохраняют свойство направленно взаимодействовать с антиген-презентирующими клетками.
2. Рекомбинантный белок по п.1, отличающийся тем, что фрагмент белка CyaA сохраняет свойство белка CyaA направленно взаимодействовать с CD11b/CD18 антиген-презентирующими клетками.
3. Рекомбинантный белок по п.1 или 2, отличающийся тем, что фрагмент белка CyaA сохраняет свойство CyaA обеспечивать транслокацию указанных одного или нескольких эпитопов указанных одного или нескольких полипептидов или транслокацию указанных одного или нескольких полипептидов в цитозоль клеток-мишеней.
4. Рекомбинантный белок по п.3, содержащий полипептид, несущий один или несколько эпитопов одного или нескольких антигенов ВПЧ, встроенные в один или несколько пермиссивных сайтов фрагмента белка CyaA, отличающийся тем, что указанный фрагмент содержит приблизительно от 30 до 1300 остатков аминокислот и включает остатки аминокислот 1208-1243 белка CyaA или остатки аминокислот 1166-1281 белка CyaA.
5. Рекомбинантный белок по п.1, отличающийся тем, что полипептид содержит и/или несколько полипептидов вместе содержат приблизительно от 5 до 500, или приблизительно от 5 до 200, или приблизительно от 10 до 50 остатков аминокислот, или приблизительно от 30 или 50 до 200 остатков аминокислот.
6. Рекомбинантный белок по п.5, способный вызывать антиген-специфический ответ против указанных одного или нескольких полипептидов.
7. Рекомбинантный белок по любому из пп.1, 2 или 4-6, отличающийся тем, что по меньшей мере один из полипептидов, несущих один или несколько эпитопов, получен из онкогенного ВПЧ.
8. Рекомбинантный белок по п.7, отличающийся тем, что полипептид, несущий один или несколько эпитопов, получен из ВПЧ16, ВПЧ18, ВПЧ31, ВПЧ33, ВПЧ35, ВПЧ45, ВПЧ52 или ВПЧ58.
9. Рекомбинантный белок по п.8, отличающийся тем, что полипептид, несущий один или несколько эпитопов, получен из антигена ВПЧ, выбранного из белков L1, L2, Е1, Е2, Е4 и Е5.
10. Рекомбинантный белок по п.8, отличающийся тем, что полипептид, несущий один или несколько эпитопов, получен из белков Е6 или Е7 ВПЧ16 и/или ВПЧ18.
11. Рекомбинантный белок по п.10, который содержит множественные полипептиды, несущие один или несколько эпитопов одного или нескольких антигенов ВПЧ.
12. Рекомбинантный белок по п.8 или 11, отличающийся тем, что полипептид, несущий один или несколько эпитопов, получен из белка Е7 вируса ВПЧ16.
13. Рекомбинантный белок по п.8 или 11, отличающийся тем, что полипептид, несущий один или несколько эпитопов, получен из белка Е7 вируса ВПЧ18.
14. Рекомбинантный белок по п.8 или 11, отличающийся тем, что полипептид представляет собой полноразмерный белок Е6 или Е7.
15. Рекомбинантный белок по п.14, содержащий множество полипептидов, встроенных в различные пермиссивные сайты последовательности CyaA или ее фрагмента.
16. Рекомбинантный белок по п.15, отличающийся тем, что указанные множественные полипептиды включают фрагмент, содержащий остатки с 1 по 29, или фрагмент, содержащий остатки с 42 по 98 белка Е7 вируса ВПЧ16, либо оба фрагмента, причем указанные полипептиды встроены в различные пермиссивные сайты белка CyaA.
17. Рекомбинантный белок по п.12, отличающийся тем, что полипептид содержит фрагмент, имеющий последовательность аминокислот RAHYNIVTF (E749-57) и/или GQAEPDRAHYNIVTFCCKCDSTLRLCVQSTHVDIR (E743-77).
18. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что полипептид или полипептиды состоят из измененного нативного антигена ВПЧ, где указанное изменение состоит в удалении одного или нескольких остатков аминокислот в кислотной области указанного антигена ВПЧ и/или во встраивании по меньшей мере двух полипептидных фрагментов указанного антигена ВПЧ в по меньшей мере два пермиссивных сайта аденилатциклазы.
19. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что полипептиды, несущие один или несколько эпитопов одного или нескольких антигенов ВПЧ, модифицируют с целью улучшения иммуногенности получаемого в результате рекомбинантного белка.
20. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что белок CyaA получен из Bordetella pertussis.
21. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что ферментативная активность белка CyaA инактивирована.
22. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что ферментативная активность белка CyaA инактивирована генетическими способами.
23. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что ферментативная активность белка CyaA инактивирована посредством встраивания дипептида в сайт аминокислотной последовательности CyaA, который участвует в циклазной активности.
24. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что способен вызывать клеточный иммунный ответ в организме хозяина-млекопитающего.
25. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что способен вызывать гуморальный иммунный ответ в организме хозяина-млекопитающего.
26. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8-11, 15 или 16, отличающийся тем, что содержит несколько полипептидов, несущих один или несколько эпитопов разных антигенов ВПЧ, встроенных в один и тот же или в различные пермиссивные сайты белка CyaA или его фрагмента.
27. Рекомбинантный белок по п.26, отличающийся тем, что полипептиды представляют собой полноразмерные полипептиды Е7, полученные из соответственно ВПЧ16 и ВПЧ18, причем указанные полипептиды встроены в различные пермиссивные сайты белка CyaA.
28. Рекомбинантный белок по любому из пп.1, 2, 4-6, 8 или 9, отличающийся тем, что указанный белок кодирует вставка, содержащаяся в плазмиде, выбиранной из pТRАСЕ5-НРV16Е7FULL (CNCM I-3191) и рТКАСЕ5-НРV16Е7Δ30-42 (CNCM I-3190).
29. Рекомбинантный белок для лечения инфекции, вызванной ВПЧ, или для профилактики или лечения злокачественных эффектов, возникающих при инфицировании ВПЧ, содержащий один или несколько полипептидов, несущих один или несколько эпитопов одного или нескольких антигенов, причем указанные один или несколько полипептидов встроены в один или в различные пермиссивные сайты белка аденилатциклазы CyaA одного из видов Bordetella или его фрагмента, причем указанный фрагмент CyaA сохраняет свойство указанного белка аденилатциклазы направленно взаимодействовать с антиген-презентирующими клетками, отличающийся тем, что по меньшей мере один из указанных эпитопов представляет собой субдоминантный криптический Т-клеточный эпитоп, причем указанный рекомбинантный белок способен вызывать антиген-специфический ответ против указанных одного или нескольких полипептидов.
30. Рекомбинантный белок по п.29, отличающийся тем, что криптический эпитоп представляет собой эпитоп ВПЧ, в частности эпитоп белка Е7 ВПЧ.
31. Рекомбинантный белок по п.30, отличающийся тем, что криптический эпитоп представляет собой эпитоп белка Е7 вируса ВПЧ18, в частности эпитоп, имеющий последовательность IDGVNHQHL.
32. Полинуклеотид, который кодирует рекомбинантный белок для лечения инфекции, вызванной вирусом папилломы человека (ВПЧ), или для профилактики или лечения злокачественных эффектов, возникающих при инфицировании ВПЧ, по любому из пп.1-31.
33. Полинуклеотид по п.32, отличающийся тем, что указанный полинуклеотид содержится в векторе pТRАСЕ5-HPV16E7FULL (CNCM I-3191) или pТRАСЕ5-НРV16Е7Δ30-42 (CNCM I-3190).
34. Рекомбинантный вектор экспрессии, который содержит нуклеотид по п.32.
35. Рекомбинантный вектор экспрессии по п.34, отличающийся тем, что указанный вектор подходит для экспрессии в бактерии.
36. Рекомбинантный вектор экспрессии по п.34, отличающийся тем, что указанный вектор подходит для экспрессии в эукариотических клетках, в частности в клетках млекопитающих.
37. Рекомбинантный вектор экспрессии по п.34, отличающийся тем, что указанный вектор выбран из pТRАСЕ5-НРV16Е7FULL (CNCM I-3191) и pТRАСЕ5-НРV16Е7Δ30-42 (CNCM I-3190).
38. Рекомбинантная клетка-хозяин, содержащая полинуклеотид по п.32 или 33 или рекомбинантный вектор экспрессии по пп.34-37.
39. Рекомбинантная клетка-хозяин по п.38, отличающаяся тем, что указанная клетка-хозяин представляет собой клетку E.coli, депонированную в Национальной коллекции культур микроорганизмов CNCM под номером I-3190 или I-3191.
40. Иммуногенная композиция для лечения или профилактики ВПЧ-инфекции или для лечения или профилактики злокачественных эффектов, возникающих при инфицировании ВПЧ, которая содержит рекомбинантный белок по любому из пп.1 - п.31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37, совместно с физиологически приемлемыми средой, наполнителем, носителем и/или разбавителем.
41. Иммуногенная композиция по п.40, отличающаяся тем, что стимулирует клеточный иммунный ответ в организме хозяина - млекопитающего.
42. Иммуногенная композиция по п.41, отличающаяся тем, что стимулирует клеточный цитолитический иммунный ответ.
43. Иммуногенная композиция по любому из пп.40-42, отличающаяся тем, что иммунный ответ включает гуморальный иммунный ответ.
44. Иммуногенная композиция по п.43, отличающаяся тем, что содержит дополнительно адъювант, и/или поверхностно-активное вещество, и/или иммуномодулирующие вещества.
45. Иммуногенная композиция по любому из пп.40-44, отличающаяся тем, что указанная композиция предназначена для иммунотерапии рака.
46. Иммуногенная композиция по любому из пп.40-44, отличающаяся тем, что указанная композиция предназначена для профилактики или лечения начала или продолжения злокачественной трансформации, причиной которой является ВПЧ-инфекция организма-хозяина.
47. Способ лечения и/или профилактики ВПЧ-инфекции у пациента, отличающийся тем, что указанный способ включает введение эффективного количества рекомбинантного белка по любому из пп.1-31, или полинуклеотида по п.32 или 33, или вектора по любому из пп.34-37.
48. Способ по п.47, отличающийся тем, что указанный рекомбинантный белок, полинуклеотид или вектор предназначены для иммунотерапии, направленной против начала или продолжения злокачественной трансформации, причиной которой является ВПЧ-инфекция организма-хозяина.
49. Вакцина против ВПЧ-инфекции и/или ассоциированных с ними злокачественных новообразований, которая содержит рекомбинантный белок по любому из пп.1-31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37, пригодный для формирования клеточного и/или гуморального ответа организма-хозяина.
50. Лекарственный препарат, содержащий рекомбинантный белок по любому из пп.1-31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37, для профилактики или лечения ВПЧ-инфекций.
51. Лекарственный препарат, содержащий рекомбинантный белок по любому из пп.1-31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37, для профилактики или лечения начала или продолжения злокачественной трансформации, вызванной ВПЧ-инфекцией организма-хозяина.
52. Лекарственный препарат, содержащий рекомбинантный белок по любому из пп.1-31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37, для иммунотерапии рака, ассоциированного с ВПЧ.
53. Набор для диагностики или иммунологического мониторинга ВПЧ-инфекций, включающий рекомбинантный белок по любому из пп.1-31, или полинуклеотид по п.32 или 33, или вектор по любому из пп.34-37.
54. Способ in vitro диагностики или иммунологического мониторинга ВПЧ-инфекций, включающий:
приведение в контакт Т-лимфоцитов, полученных из организма млекопитающего, в частности человека, рекомбинантным белком по любому из пп.1-31, детектирование изменения активации Т-лимфоцитов.
55. Способ скрининга неизвестных субдоминантных криптических Т-клеточных эпитопов, содержащихся в полипептиде, который находится в составе химерного белка: CyaA-полипептид, где CyaA представляет собой аденилатциклазу или ее фрагмент, описанные в пп. 1-31, а полипептид получен из антигенов ВПЧ, в частности из белков Е7 ВПЧ16 и/или ВПЧ18, причем указанный способ включает: введение указанного химерного белка в организм животного - хозяина, определение Т-клеточного ответа, в частности ответа цитотоксических Т-лимфоцитов у указанного хозяина.
56. Бивалентная терапевтическая вакцина против ВПЧ-инфекций и/или ассоциированных с ними злокачественных новообразований, включающая:
а) рекомбинантный белок по п.1, в котором указанный антиген ВПЧ представляет собой белок Е7 ВПЧ16; и
б) рекомбинантный белок по п.1, в котором указанный антиген ВПЧ представляет собой белок Е7 ВПЧ18.
57. Бивалентная терапевтическая вакцина по п.56, отличающаяся тем что:
- рекомбинантный белок а) содержит первые 29 остатков аминокислот Е7 ВПЧ16, встроенных между аминокислотами 319 и 320 CyaA, а также остатки с 43 по 98 Е7 ВПЧ16, встроенные между аминокислотами 224 и 235 CyaA; и
- рекомбинантный белок б) содержит первые 31 остатка аминокислот Е7 ВПЧ18, встроенные между аминокислотами 319 и 320 CyaA, а также остатки с 43 по 105 Е7 ВПЧ18, встроенные между аминокислотами 224 и 235 CyaA;
и при этом CyaA лишена ферментативной активности в результате вставки дипептида LQ между остатками аминокислот Asp 188 и Ilе 189.
58. Бивалентная терапевтическая вакцина против ВПЧ-инфекций и/или ассоциированных с ними злокачественных новообразований, содержащая рекомбинантный белок по п.1, причем указанный рекомбинантный белок содержит множество полипептидов, каждый из которых содержит эпитопы, которые представляют собой один или несколько фрагментов белка Е7 ВПЧ16 и белка Е7 ВПЧ18.
59. Бивалентная терапевтическая вакцина против ВПЧ-инфекций и/или ассоциированных с ними злокачественных новообразований, содержащая рекомбинантный белок по п.1, причем указанные полипептиды, встроенные в различные пермиссивные сайты, включают эпитопы, которые представляют собой один или несколько фрагментов белка Е7 ВПЧ16 и белка Е7 ВПЧ18.
60. Бивалентная терапевтическая вакцина по п.59, отличающаяся тем, что указанный рекомбинантный белок содержит:
(а) остатки с 43 по 98 Е7 ВПЧ16 и следующие за ними остатки с 43 по 105 Е7 ВПЧ18, встроенные между аминокислотами 224 и 235 CyaA;
(б) первые 31 остатков аминокислот Е7 ВПЧ 18 и следующие за ними первые 29 остатков аминокислот Е7 ВПЧ 16, встроенные между аминокислотами 319 и 320 CyaA,
и при этом CyaA лишена ферментативной активности в результате вставки дипептида LQ между остатками аминокислот Asp 188 и Ile 189.
| US 6458368, 01.10.2002 | |||
| US 6251406, 26.06.2001. |

