Изобретение относится к области химии и медицины, а именно к новым соединениям, способным доставлять нуклеиновые кислоты в клетки млекопитающих.
Генная терапия предназначена для лечения наследственных, инфекционных и онкологических заболеваний и направлена либо на коррекцию наследственного заболевания, либо на придание клеткам новых функций. Успех генной терапии зависит от разработки эффективных способов доставки терапевтических генов в клетки-мишени, обеспечения их экспрессии и длительного функционирования в этих клетках [1, 2]. Вирусные векторы характеризуются высокой эффективностью доставки нуклеиновых кислот [1], однако, имунногенность, мутагенность, токсичность, а также проблемы масштабной наработки таких вирусных частиц стимулировали поиск альтернативных невирусных транспортных систем. В настоящее время в качестве невирусных векторов предложены катионные липиды, полимеры, дендримеры и пептиды [3-5]. Невирусные системы обладают рядом преимуществ, таких как низкая токсичность, неиммуногенность, а также возможность их легкого получения в ходе контролируемых процессов. Среди невирусных транспортных систем широкое применение находят катионные липиды и липосомы на их основе [3-12]. Липиды защищают нуклеиновые кислоты от деградации и способствуют их переносу через клеточную мембрану, инициируя эндоцитоз. Катионные липиды представляют собой положительно заряженные амфифильные соединения, которые состоят из трех структурных доменов: гидрофобного, гидрофильного и соединяющей их спейсерной группы. Гидрофобная часть чаще всего представлена остатком холестерина, диглицерида или длинноцепных углеводородных заместителей. В качестве гидрофильного домена используют монозарядные катионные группы, полученные кватернизацией алифатических или гетероциклических оснований, или остатки поликатионных природных или синтетических полиаминов. Гидрофобная и гидрофильная составляющие катионных липидов обычно разделены спейсером и соединяются карбамоильной, амидной, простой или сложноэфирной связью. Тип связывания определяет стабильность и токсичность катионного липида [3, 13].
Наиболее ближайшим к заявляемому соединению - прототипом, является катионный липид, rac-N-[2,3-ди(олеилокси)проп-1-ил]-N,N,N-триметиламмоний хлорид (DOTMA) [14]. В структуру данного липида входит остаток 1,2-ди-О-олеоил-rac-глицерина, который формирует гидрофобный домен, и триметиламмониевая группа, которая формирует катионный домен, который связывается с фосфатными группами нуклеиновых кислот.
Недостатками известного соединения являются высокая токсичность и низкая трансфицирующая активность при использовании в виде индивидуального соединения.
Технической задачей изобретения является получение нового эффективного синтетического соединения, обладающего высокой трансфицирующей активностью, низкой токсичностью и способного доставлять нуклеиновые кислоты в клетки млекопитающих.
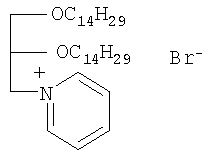
Поставленная техническая задача решается предлагаемым соединением, представляющим собой катионный липид на основе глицерина, структурная формула которого представлена на чертеже.
Заявляемое соединение получают способом, описанным в [15]. Исходным соединением в синтезе катионного липида rac-N-[2,3-ди(тетрадецилокси)проп-1-ил]пиридиний бромида служит 1-бром-1-дезокси-2,3-ди-O-тетрадецил-rac-глицерин, который получают обработкой 1,2-ди-О-тетрадецил-rac-глицерина избытком четырехбромистого углерода в присутствии трифенилфосфина. Нагревание 1-бром-1-дезокси-2,3-ди-O-тетрадецил-rac-глицерина с избытком пиридина в течение 43-45 часов при 90°С и последующая перекристаллизация из диэтилового эфира позволяют получить целевой продукт - rac-N-[2,3-ди(тетрадецилокси)проп-1-ил]пиридиний бромид.
Для синтеза заявляемого соединения использовали растворители и реагенты отечественного производства. ТСХ осуществляли на силикагеле (Silicagel 60 F254, Merck). Идентификацию соединений проводили с помощью реактива Драгендорфа или раствора Ce(SO4)2 - фосформолибденовой кислоты с последующим прогреванием до 100°С. Колоночную хроматографию проводили на силикагеле Kieselgel 60 (40-63 мкм, Merck). Спектры 1Н-ЯМР (δ, м.д. КССВ, Гц) регистрировали на импульсном фурье-спектрометре «Bruker AMD-400» с рабочей частотой 400 МГц в CDCl3 или смеси CDCl3-CD3OD, внутренний стандарт - Me4Si. Масс-спектры получали на время-пролетном масс-спектрометре LC/MSD Agilent 1100 Series ионизацией с помощью электрораспыления (ESI).
Для изучения способности предлагаемого соединения доставлять нуклеиновые кислоты в клетки млекопитающих использовали 25-звенный олигодезоксирибонуклеотид 5'-dTACAGTGGAATTGTATGCCTATTA-3', модифицированный флуоресцеинизотиоцианатом (FITC) по 3'-концу, плазмидную ДНК (pEGFP-C2, «Clontech» (Германия)) и 21-звенную двухцепочечную РНК (siPHK, ИХБФМ СО РАН) (последовательность смысловой цепи 5'-GCGCCGAGGUGAAGUUCGATT-3', антисмысловой цепи - 5'-UCGAACUUCACCUCGGCGCGG-3'). FITC-ON был синтезирован фосфитамидным методом [16] и очищен ионообменной и обращенно-фазовой хроматографией. Чистоту олигонуклеотида проверяли с помощью электрофореза в 15% ПААГ в денатурирующих условиях, визуализацию олигонуклеотида в геле проводили с помощью окраски Stains-All.
Эффективность проникновения нуклеиновых кислот с использованием заявляемого соединения в клетки млекопитающих in vitro была исследована в экспериментах по трансфекции клеток FITC-меченым олигонуклеотидом, плазмидной ДНК, кодирующей зеленый флуоресцирующий белок (EGFP), и короткой интерферирующей РНК, направленной против матричной РНК гена, кодирующего EGFP.
Сопоставительный анализ заявляемого соединения с известными и широко используемыми коммерческими агентами трансфекции, такими как Lipofectamine® 2000 и Oligofectamine, показал, что предлагаемое соединение обладает следующими преимуществами.
1. Заявляемое соединение обладает способностью доставлять в клетки млекопитающих нуклеиновые кислоты, как короткие, так и протяженные, что позволяет рассматривать его как перспективный агент для трансфекции.
2. Заявляемое соединение доставляет нуклеиновые кислоты в клетки в индивидуальном состоянии в виде раствора в этиловом спирте и не требует использования дополнительных вспомогательных липидов.
3. Заявляемое соединение малотоксично для клеток млекопитающих.
4. Заявляемое соединение не требует сложной процедуры приготовления, свойственной липосомальным композициям.
Кроме того, заявляемое соединение стабильно при хранении, как в сухом виде, так и в виде концентрированного спиртового раствора.
Поиск по источникам научно-технической и патентной литературы показал, что соединение rac-N-[2,3-ди(тетрадецилокси)пропил]пиридиний бромид, обладающее способностью доставлять нуклеиновые кислоты в клетки млекопитающих, в известных источниках не описано.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение rac-N-[2,3-ди(тетрадецилокси)проп-1-ил]пиридний бромида
К охлажденному до 0°С раствору 1,2-ди-О-тетрадецил-rac-глицерина (96.5 мг, 0.199 ммоль) и трифенилфосфина (109.5 мг, 0.417 ммоль) в безводном дихлорметане (10 мл) добавили по каплям раствор тетрабромуглерода (139.6 мг, 0.420 ммоль) в безводном дихлорметане. Через 17 часов к реакционной смеси добавили метанол (2 мл) и перемешивали 10 мин, растворитель удаляли в вакууме, остаток хроматографировали на колонке с силикагелем, выделяя 1-бром-1-дезокси-2,3-ди-O-тетрадецил-rac-глицерин (108.0 мг, 99%). Смесь 1-бром-1-дезокси-2,3-ди-O-тетрадецил-rac-глицерина (108 мг, 0.19 ммоль) и пиридина (1 мл) выдерживали 44 часа при 90°С. Пиридин удаляли в вакууме, продукт перекристаллизовывали из безводного диэтилового эфира, получая rac-N-[2,3-ди(тетрадецилокси)проп-1-ил]пиридиний бромид (82 мг, 66%). Масс-спектр, m/z (Iотн(%)): 546.7 [М-Br]+ (100%). Спектр ЯМР 1Н: (δ, м.д., J/Гц): 0.81 (т, 6Н, J=7.0, 2 (CH2)11СН 3), 1.24 (уш. с, 44Н, 2(СН 2)11СН3), 1.27-1.50 (м, 4Н, 2 OCH2CH 2), 3.19 и 3.52 (оба дт по 1Н, J=6.6 и 9.3, OCH 2CH2), 3.38 (т, 2Н, J=6.7, OCH 2CH2), 3.58 (дд, J=4.0 и 10.9, ОСНН а), 3.70 (дд, J=3.1 и 10.9, OCHH b), 3.94-4.00 (м, 1Н, CHOC14H29), 4.72 (дц, 1Н, J=8.3 и 13.2) и 5.53 (дд, 1Н, J=2.9 и 13.2, CH2N+), 7.97 (м, 2Н), 8.43 (м, 1Н) и 9.41 (м, 2 Н, C5H5N+).
Пример 2. Исследование эффективности трансфекции в клетки FITC-меченого олигонуклеотида с использованием заявляемого катионного липида
Исследование проникновения FITC-меченого олигонуклеотида в клетки НЕК 293 и ВНК проводили с помощью проточной цитофлуориметрии. Эффективность трансфекции оценивали по количеству трансфицированных клеток от общего количества клеток в образце. Клетки высаживали в 24-луночные планшеты (2·105 клеток на лунку для клеток НЕК 293 и 1·105 клеток на лунку для клеток ВНК) и культивировали 24 ч при 37°С в атмосфере, содержащей 5% CO2. Перед проведением трансфекции для экспериментов в присутствии или в отсутствие сыворотки среду в лунках заменяли на 200 мкл среды DMEM, содержащей 10% сыворотки, или DMEM, не содержащей сыворотку, соответственно. Раствор катионного липида (10 мкМ) в среде OptiMEM смешивали с раствором FITC-меченого олигонуклеотида (5 мкМ) в этой же среде, полученные комплексы добавляли к клеткам и инкубировали в течение 4 ч. По окончании инкубации клетки промывали фосфатно-солевым буфером (PBS) (300 мкл), добавляли 50 мкл раствора трипсина и инкубировали 1-2 мин (37°С, 5% CO2). По окончании инкубации в лунки добавляли 200 мкл DMEM с 10% сыворотки, клетки суспендировали и переносили в пробирки. Полученную клеточную суспензию центрифугировали при 1000-1200 об/мин, 4°С, отбирали среду и промывали 1 мл PBS. Затем клетки фиксировали 500 мкл 2% раствора формальдегида в PBS. Для исследования трансфицирующей активности катионного липида использовали клеточный сортер BD FACSAria (Becton Dickinson), измеряющий флуоресценцию клеток, содержащих FITC-меченый олигонуклеотид, определяемый при длине волны возбуждения 488 нм (в приборе используется когерентный сапфировый лазер (20 мВ)).
Результаты проникновения в клетки FITC-меченого олигонуклеотида приведены в таблице 1.
Из приведенных данных видно, что в присутствии заявляемого катионного липида FITC-меченый олигонуклеотид эффективно (количество трансфицированных клеток превышает 75%) проникает в клетки в отсутствие сыворотки в ростовой среде. Наличие в ростовой среде 10% сыворотки уменьшает эффективность трансфекции, однако количество трансфицированных клеток при этом составляет примерно 50% или выше.
Пример 3. Трансфекция клеток НЕК 293 плазмидной ДНК с использованием заявляемого катионного липида
Исследование проникновения плазмидной ДНК в клетки НЕК. 293 проводили с помощью проточной цитофлуориметрии. Эффективность трансфекции оценивали по количеству клеток, содержащих зеленый флуоресцентный белок (EGFP) от общего количества клеток в образце. Клетки высаживали в 24-луночные планшеты (2·105 клеток на лунку для клеток НЕК 293) и культивировали в течение суток при 37°С в атмосфере, содержащей 5% CO2. Перед проведением трансфекции для экспериментов в присутствии или в отсутствие сыворотки среду в лунках заменяли на 200 мкл среды DMEM, содержащей 10% сыворотки, или DMEM, не содержащей сыворотку, соответственно. Раствор катионного липида (20 мкМ) в среде OptiMEM смешивали с раствором плазмиды pEGFP-C2 (0.3 мкг/мкл) в этой же среде, полученные комплексы добавляли к клеткам и инкубировали в течение 24 ч. Через 4 часа после начала трансфекции в лунках со средой без сыворотки заменяли среду на DMEM с 10% сывороткой. По окончании инкубации клетки обрабатывали, как описано в примере 1. В таблице 2 представлены результаты трансфекции клеток плазмидной ДНК pEGFP-C2 с использованием заявляемого катионного липида.
Из приведенных данных видно, что плазмидная ДНК в присутствии заявляемого катионного липида проникает в клетки в заметном количестве в отсутствие сыворотки в ростовой среде.
Пример 4. Трансфекция клеток ВНК IR-780 короткой интерферирующей РНК с использованием заявляемого катионного липида
Исследование проникновения siPHK, направленной на подавление синтеза зеленого флуоресцирующего белка (EGFP), проводили на клетках линии ВНК IR-780, стабильно экспрессирующих этот белок. В качестве мишени была выбрана мРНК, кодирующая белок EGFP, таким образом по уменьшению флуоресценции клеток можно сделать выводы об эффективности доставки короткой интерферирующей РНК в цитоплазму. Клетки высаживали в 24-луночные планшеты (0.2·105 клеток на лунку для клеток ВНК IR-780) и культивировали в течение суток при 37°С в атмосфере, содержащей 5% CO2. Перед проведением трансфекции для экспериментов в присутствии или в отсутствие сыворотки среду в лунках заменяли на 200 мкл среды DMEM, содержащей 10% сыворотки, или DMEM, не содержащей сыворотку, соответственно. Раствор катионного липида (10 мкМ) в среде OptiMEM смешивали с раствором короткой интерферирующей РНК (50 нМ) в этой же среде, полученные комплексы добавляли к клеткам и инкубировали в течение 72 ч. Через 4 часа после начала трансфекции в лунках со средой без сыворотки заменяли среду на DMEM с 10% сывороткой. По окончании инкубации клетки обрабатывали, как описано в примере 1. В таблице 3 представлены результаты доставки короткой интерферирующей РНК заявляемым катионным липидом.
Из приведенных данных видно, что заявляемый катионный липид в количестве, малотоксичном для клеток, способствует эффективному проникновению короткой интерферирующей РНК в клетки в отсутствие сыворотки в ростовой среде.
Пример 5. Влияние заявляемого катионного липида на жизнеспособность клеток ВНК, ВНК IR-780, HeLa, HEK 293
Клетки линий ВНК (эмбриональные клетки почки сирийского хомячка), ВНК IR-780 (модифицированные эмбриональные клетки почки сирийского хомячка), HeLa (клетки карциномы шейки матки человека), HEK 293 (эмбриональные клетки почки человека) культивировали в среде DMEM, содержащей 10%-ную эмбриональную телячью сыворотку в атмосфере 5%-ного CO2 при 37°С. Жизнеспособность клеток после инкубации с катионным липидом определяли с помощью МТТ теста, который основан на способности живых клеток превращать соединения на основе тетразола (МТТ) в ярко окрашенные кристаллы формазана, что позволяет спектрофотометрически оценивать количество живых клеток в препарате. Для этого клетки высаживали в 96-луночные планшеты (0.1·105 клеток на лунку для клеток HEK 293 и 0.03·105 клеток на лунку для клеток HeLa, BHK, BHK IR-780). Через 24 ч в лунках меняли среду и к клеткам добавляли раствор соединения в среде DMEM до конечной концентрации в лунке от 1 до 80 мкМ. Клетки инкубировали в присутствии катионного липида еще в течение 24 часов в тех же условиях. По окончании инкубации без смены среды к клеткам добавляли раствор МТТ (5 мг/мл) в фосфатно-солевом буфере до концентрации 0.5 мг/мл и инкубировали в течение 3 ч в тех же условиях. Среду удаляли, к клеткам добавляли по 100 мкл диметилсульфоксида, в котором происходит растворение образовавшихся в клетках кристаллов формазана, и измеряли оптическую плотность на многоканальном спектрофотометре на длинах волн 570 и 630 нм, где А570 - поглощение формазана, а А630 - фон клеток.
Из экспериментальных данных вычисляли значение IC50, концентрацию соединения, при которой наблюдается гибель 50% клеток. Значения IC50 катионного липида для клеток приведены в таблице 4.
Из приведенных данных видно, что обработка клеток заявляемым катионным липидом вызывает их эффективную гибель только при концентрациях соединений выше 10 мкМ, что свидетельствует о низкой токсичности катионного липида.
Таким образом, предлагаемое соединение, представляющее собой катионный липид на основе глицерина со структурной формулой, представленной на чертеже, обладает способностью эффективно доставлять нуклеиновые кислоты в клетки млекопитающих, что позволяет использовать его в качестве агента для доставки генетического материала в клетки.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Lundstrom К. // Trends Biotechnol. 2003. V.21. P.117-122.
2. Rubanyi G.M. // Handbook of pharmaceutical biotechnology / Ed. Gad S.C., John Wiley & Sons, 2007. P. 943-970.
3. Martin В., Sainlos M., Aissaoui A., Oudrhiri N., Hauchecorne M., Vigneron J.-P., Lehn J.-M., Lehn P. II Curr. Pharm. Design. 2005. V.11. P.375-394.
4. Mintzer M.A., Simanek E.E. // Chem. Rev. 2009. V.109, P.259-302.
5. Dufes С., Uchegbu I.F., Schatzlein A.G. // Adv. Drug Deliv. Rev. 2005. V.57. P.2177-2202.
6. Wasungu L., Hoekstra D. // J. Controlled Release. 2006. V.116. P.255-264.
7. Morille M, Passirani С, Vonarbourg A., Clavreul A., Benoit J.-P. // Biomaterials. 2008. V.29. P.3477-3496.
8. KirbyA.J., Camilleri P., Engberts J.B.F.N., Fetters M.C., Nolte R.J.M., Soderman O., Bergsma M., BellP.C., Fielden M.L., Rodriguez C.L.G, Guedat P., Kremer A., McGregor C., Perrin C., Ronsin G., van Eijk M.C.P. // Angew. Chem. Int. Ed. 2003. V.42. P.1448-1457.
9. Kostarelos К., Miller A.D. // Chem. Soc. Rev. 2005. V.34. P.970-994.
10. Bhattacharya S., Bajaj A. // Curr. Opin. Chem. Biol. 2005. V.9. P.647-655.
11. Маслов М.А., Сычева Е.В., Морозова Н.Г., Серебренникова Г.А. // Изв. АН. Сер. хим. 2000. С.385-400.
12. Zhanga S., Xua Y., Wanga B., Qiaob W., Liub D., LiZ. // J. Controlled Release. 2004. V.100. P.165-180.
13. Lv H., Zhang S., Wang В., Cui S., Yan J. // J. Controlled Release. 2006. V.114. P.100-109.
14. синтез ДОТМА.
15. Маслов М. А., Сычева Е.В., Морозова Н.Г., Серебренникова Г.А. // Изв. АН. Сер. хим. 1999. С.1381-1384.
16. D.Proudnikov, A.Mirzabekov // Nucleic Acids Res. 1996. V.24. P.4535-4542.
| название | год | авторы | номер документа |
|---|---|---|---|
| УГЛЕВОДСОДЕРЖАЩИЕ КАТИОННЫЕ АМФИФИЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ ДОСТАВЛЯТЬ НУКЛЕИНОВЫЕ КИСЛОТЫ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2009 |
|
RU2394834C1 |
| КОМПОЗИЦИЯ ДЛЯ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2010 |
|
RU2423147C1 |
| Незаряженный липид, композиция на его основе с поликатионным амфифилом и нейтральным фосфолипидом и способ ее получения для доставки нуклеиновых кислот in vitro | 2020 |
|
RU2747559C1 |
| Полиэтиленгликоль-содержащий липид, композиция на его основе с катионным амфифилом и нейтральным фосфолипидом и способ ее получения для доставки нуклеиновых кислот in vivo | 2017 |
|
RU2683572C1 |
| ДИСУЛЬФИДНЫЙ ПОЛИКАТИОННЫЙ АМФИФИЛ, КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ С НЕЙТРАЛЬНЫМ ФОСФОЛИПИДОМ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2016 |
|
RU2610271C1 |
| ФРАГМЕНТЫ ДВУЦЕПОЧЕЧНОЙ РНК, ОБЛАДАЮЩИЕ АНТИПРОЛИФЕРАТИВНОЙ И ИНТЕРФЕРОН-ИНДУЦИРУЮЩЕЙ АКТИВНОСТЯМИ | 2008 |
|
RU2391405C1 |
| БИОДЕГРАДИРУЕМЫЕ КАТИОННЫЕ ПОЛИМЕРЫ | 2005 |
|
RU2440380C2 |
| РЕПОРТЕРНАЯ МОНОКЛОНАЛЬНАЯ КЛЕТОЧНАЯ ЛИНИЯ НЕК-293/5-HT4R, ЭКСПРЕССИРУЮЩАЯ МЫШИНЫЙ РЕКОМБИНАНТНЫЙ СЕРОТОНИНОВЫЙ РЕЦЕПТОР ТИПА 5HT4 | 2022 |
|
RU2817234C2 |
| Способ получения системы доставки фрагментов нуклеиновых кислот в клетки млекопитающих | 2016 |
|
RU2646113C1 |
| ПРОИЗВОДНЫЕ АЛКИЛАММОНИЙНОГО ГЕМИНАЛЬНОГО ПОВЕРХНОСТНО-АКТИВНОГО ВЕЩЕСТВА, ОБЕСПЕЧИВАЮЩИЕ ЭФФЕКТИВНУЮ ДОСТАВКУ ДНК В КЛЕТКИ | 2012 |
|
RU2513726C2 |

Изобретение относится к новому химическому соединению, а именно к rac-N-[2,3-ди(тетрадецилокси)проп-1-ил]пиридиний бромиду, обладающему способностью доставлять нуклеиновые кислоты в клетки млекопитающих:
 Технический результат: описанное соединение, обладает низкой токсичностью и способно в виде спиртового раствора доставлять нуклеиновые кислоты в клетки млекопитающих. 4 табл., 1 ил.
Технический результат: описанное соединение, обладает низкой токсичностью и способно в виде спиртового раствора доставлять нуклеиновые кислоты в клетки млекопитающих. 4 табл., 1 ил.
rac-N-[2,3-Ди(тетрадецилокси)проп-1-ил]пиридиний бромид со структурной формулой
в качестве агента для доставки нуклеиновых кислот в клетки млекопитающих.
| Способ получения композиционных материалов | 1971 |
|
SU451763A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| ПРОИЗВОДНЫЕ ГЛИЦЕРИНА, СРЕДСТВО ДЛЯ ДОСТАВКИ ФИЗИОЛОГИЧЕСКИ АКТИВНОГО ВЕЩЕСТВА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1994 |
|
RU2123492C1 |

