Область техники
Изобретение относится к области экспериментальной биологии и медицины, и касается создания репортерных клеток, экспрессирующих мышиный рекомбинантный серотониновый рецептор типа 5-НТ4 и способных дозозависимо детектировать появление серотонина во внешней среде. В основе изобретения - репортерные моноклональные модифицированные клетки HEK-293/5-HT4R, гетерологически экспрессирующие мышиный рекомбинантный рецептор серотонина типа 5-НТ4, слитый с флуоресцентным белком GFP. Полученные клетки могут быть использованы для неинвазивного оптического мониторинга локализации 5-НТ4 рецептора, а так же, за счет обогащения флуоресцентным генетически кодируемым белком-сенсором внутриклеточного сАМР, дозозависимых внутриклеточных сАМР-сигналов, инициируемых серотонином и другими лигандами 5-НТ4 рецептора, в частности, для доклинического скрининга потенциальных лекарственных соединений и регистрации секреции серотонина различными типами клеток.
Предшествующий уровень техники
Серотонин - возможно наиболее известный и наиболее изученный нейромедиатор, модулирующий практически все поведенческие реакции человека, а также многочисленные физиологические процессы, включая регуляцию сердечно-сосудистой и дыхательной систем, перистальтику кишечника, контроль мочевого пузыря, агрегацию тромбоцитов, развитие тканей и т.д. Рецепторы серотонина распространены не только в центральной нервной системе, но и на периферии, они являются мишенями примерно для 40% одобренных к настоящему моменту терапевтических препаратов и остаются одной из самых прибыльных и многообещающих мишеней для разработки новых лекарств от нервных заболеваний, начиная от мигрени и заканчивая нервно-психическими расстройствами, такими как шизофрения и депрессия. Согласно современной классификации, существует семь типов 5-НТ рецепторов (5-НТ1-7), которые представляют собой семи-трансмембранные GPCR (G-белок связанные рецепторы), за исключением 5-НТ3, являющегося лиганд-управляемым катионным каналом [1, 2].
Серотониновые GPCR связаны со всеми тремя каноническими сигнальными путями через Gαi, Gαq/11 и Gαs белки, позволяя этому семейству рецепторов модулировать несколько биохимических сигнальных путей и приводя к самым разнообразным физиологическим последствиям. При скрининге потенциальных лекарственных препаратов широко используются клеточные линии, экспрессирующие какой-либо подтип серотонинового рецептора, позволяющие проводить наблюдения за клетками прижизненно в условиях физиологического эксперимента.
Известно изобретение, где описан химерный флуоресцентный белок psychLight, созданный на основе рецептора серотонина типа 5-НТ2А и флуоресцентного белка cpGFP, вставленного на место 3й трансмембранной петли в рецептор 5-НТ2А, что позволяет визуализовать конформационные изменения рецептора вследствие связывания с лигандом. Авторы предполагают, что изобретение будет полезно для скрининга лекарств из ряда нейролептиков и психоделиков, мишенью для которых является серотониновый 5-НТ2А рецептор [3]. Хотя предложенное изобретение вероятно позволит оценить величину галлюциногенного эффекта препаратов, авторами не описывается, как влияет конформационное изменение, внесенное в структуру 5-НТ2А рецептора, на последующее взаимодействие рецептора с G-белками и сигнальным фосфолипазным каскадом, им активируемым.
Известно изобретение химерного флуоресцентного белка GRAB на основе рецептора серотонина типа 5-НТ2С и флуоресцентного белка cpGFP, так же вставленного в 3ю внутриклеточную петлю белка рецептора и меняющего интенсивность флуоресценции при связывании лиганда. Авторы продемонстрировали работу фосфоинозитидного каскада, активируемого лигандами 5-НТ2С рецептора, методом кальциевого имиджинга при окрашивании клеток флуоресцентным химическим зондом Cal590. Помимо временной экспресии белка GRAB в клетках НЕК-293Т, авторы проводили исследования на срезах мозга мыши и на самцах дрозофиллы, что позволило визуализовать секрецию серотонина в этих объектах [4].
Известно изобретение, где GFP использовался для опосредованного мечения экспрессии 5-НТ1В рецептора серотонина. Для модификации первичной культуры нейронов и непосредственно животных авторы использовали генетическую конструкцию на базе вируса герпеса, в которой под двумя разными промоторами содержались нуклеотидные последовательности для независимой экспрессии GFP и 5-НТ1В [5]. Данный подход позволяет визуализовать модифицированные клетки, однако не позволяет оценить локализацию 5-НТ1В рецептора, так как рецептор должен встроится и функционировать в клеточной мембране, a GFP, являясь водорастворимым белком, полностью окрашивает цитоплазму клетки.
Известно изобретение линий клеток с экспрессией крысиного рецептора серотонина типа 5-НТ2А и флуоресцентного белка GFP, где слияние двух этих белков осуществлено в плазмиде pEGFP-N1. Авторы получили несколько клеточных линий НЕК-293, с разным уровнем экспрессии рецептора 5-НТ2А, слитого с GFP, и выбрали две наиболее яркие линии для дальнейших экспериментов по исследованию интернализации 5-НТ2А рецептора под воздействием разных факторов [6].
Все вышеперечисленные рецепторы - 5-НТ2А, 5-НТ2С и 5-НТ1В - связаны с фосфоинозитидным каскадом, тогда как 5-НТ4 рецептор, используемый в нашем изобретении связан с аденилатциклазным сигнальным каскадом.
Рецептор 5-НТ4 (5-HT4R) вызывает особый интерес, так как экспрессируется во многих тканях организма и его активация связана с обучением и памятью, депрессией, реакцией на стресс, контролем дыхания, питанием, транзитом пищи через желудочно-кишечный тракт и др. Так же предполагается, что антагонисты 5-HT4R обладают клиническим потенциалом в лечении мерцательной аритмии и сердечной недостаточности.
Известно изобретение, где предлагается способ идентификации химических соединений, специфически связывающихся с 5-HT4R млекопитающих. Авторы впервые выделили и клонировали нуклеотидную последовательность 5-НТ4 рецептора крысы и человека в плазмиды pBluescript-hS10, pcEXV-S10-87 и pcEXV-S10-95 (плазмиды, содержащие последовательности 5-HT4R, задепонированы в американской коллекции АТСС (№№75392, 75390, 75391) и в настоящий момент недоступны для поиска и приобретения). Авторы разработали антитела, специфичные к 5-HT4R, и антисмысловые нуклеотидные последовательности, уменьшающие уровень экспрессии 5-HT4R при введении в объект исследования. Для получения клеточных мембран, содержащих 5-HT4R, авторы осуществили временную экспрессию рецептора в клетках COS-7, для чего последовательность 5-HT4R была клонирована в плазмиду pcDNA1-Amp, которая не содержит гена маркера селекции для клеток млекопитающих и не содержит последовательности флуоресцентного белка, позволяющего неинвазивно оценить уровень экспрессии целевого белка в клетках [7, 8].
Известно изобретение, авторы которого описывают 2 сплайс-варианта серотонинового 5-HT4R человека (5-НТ4(с) и 5-HT4(d)). Основываясь на полученных нуклеотидных последовательностях, авторы описывают два полипептида (или любые их полипептидные фрагменты или биологически активные производные), имеющих фармакологическую активность серотонинового 5-HT4R. Для экспрессии рецептора в клетках млекопитающих, авторы клонировали 5-HT4R в плазмиду pRc/CMV, которая содержит ген маркера селекции для клеток млекопитающих, но не содержит последовательности флуоресцентного маркерного белка. При проведении исследований авторы использовали клетки COS-7, временно экспрессирующие 5-HT4R, и клетки СНО, стабильно экспрессирующие плазмиду pRc/CMV с рецептором 5-НТ4. Полученные клетки CHO+pRc/CMV+5-HT4R обладают устойчивостью к антибиотику генецитину (G418), что уменьшает возможности дальнейшей дополнительной модификации клеток, поскольку это наиболее распространенный маркер селекции. Неинвазивная визуализация локализации рецептора 5-НТ4 в клетках CHO+pRc/CMV+5-HT4R невозможна [9].
Известно изобретение, описывающее экспрессию 5-HT4R в разных тканях человека: мозге, желудочно-кишечном тракте, ткани сердца, печени, легких и т.д. Для скрининга экспрессии авторы брали последовательность 5-HT4R человека из базы данных под обозначением Y12505 SEQ ID NO: 1. Так же авторы описывают праймеры для выделения последовательности 5-HT4R и возможные способы получения плазмид, пригодных для гетерогенной экспрессии 5-HT4R в различных клетках, однако клеточной линии, экспрессирующей 5-HT4R получено не было [10].
Известно изобретение, в котором для оценки эффективности препаратов, мишенью которых является 5-HT4R, использовались клетки СНО-К1, с постоянной оверэкспрессией 5-НТ4(b) - человеческого сплайс-варианта рецептора. Авторы использовали слияние 5-HT4R на N- конце с FLAG-тагом, клетки использовались для анализа продукции сАМР под влиянием исследуемых препаратов методом иммуногистохимии, проведение исследований осуществлялось на фиксированных (дополнительно обработанных неживых) клетках [11].
Известно использование эндогенных рецепторов серотонина 5-НТ4, слитых с разными метками (НА, FLAG, RhoTag), для изучения влияния димеризации рецептора 5-HT4R на активацию G-белка в клетках COS-7, временно экспрессирующих рецепторы 5-НТ4 [12].
Известно изучение разницы в интернализации двух изоформ 5-HT4R после активации серотонином и экзогенным лигандом и участие в этом процессе внутриклеточных молекул на клетках НЕК-293 с постоянной экспрессией серотониновых рецепторов. Авторы работы использовали мечение рецепторов специфичными тагами, узнаваемыми коммерческими антителами, используемыми в методе ELISA и при получении микрофотографий на конфокальном микроскопе. Хотя в работе показана зависимость процента интернализованного рецептора от времени, измерения выполнялись в отдельных временных точках на разных препаратах фиксированных клеток, мониторинг в реальном времени не проводился, так же для визуализации экспрессии рецептора клетки дополнительно обрабатывались [13].
Известно исследование взаимодействия С-конца 5-HT4R с внутриклеточными сигнальными белками, где авторы использовали клетки НЕК-293, временно экспрессирующие RhoTag-5-HT4R и нексин HA-SNX27, участвующий в интернализации 5-HT4R. При помощи иммуноцитохимии, требующей фиксации и обработки клеток, было показано взаимодействие и колокализация этих белков [14].
Существенным недостатком ряда перечисленных изобретений является использование для визуализации рецептора меток-тагов, специфичных для определенных коммерческих вторичных антител. Этот подход достаточно информативен, но используя его можно визуализовать локализацию белка только постфактум на обработанных неживых клетках. Так же отметим, что при использовании иммуноферментного/радиоиммунного анализа, измерения проводятся в одной временной точке каждый раз на новом препарате клеток, что не позволяет вести наблюдения в реальном времени и затрудняет осуществление внутреннего контроля. Описанные выше примеры не дают возможности мониторинга изменений концентрации внутриклеточных молекул на коротких промежутках времени, что особенно важно при исследовании влияния агонистов рецепторов. Так же данные, полученные на клеточном лизате, могут не отражать важных характеристик физиологического ответа живых клеток.
Использование экспрессионной плазмиды, обеспечивающей слияние рецептора с флуоресцентным белком, позволяет осуществить мониторинг локализации рецептора (провести физиологические эксперименты) в реальном времени на живых клетках.
Наиболее близким по сути к предлагаемому изобретению является работа, в которой осуществлена прижизненная визуализация 5-HT4R, где использовался эндогенный рецептор серотонина, клонированный в плазмиду pcDNA 3.1 и слитый на С-конце с флуоресцентным белком (eGFP, mCherry или mCerulean), временно экспрессируемый в клетках нейробластомы N1E-115, в которой была визуализована ретракция мембраны при участии G-белка G13 и малой GTPaзы семейства Rho - RhoA. [15]. Изобретение можно считать прототипом, однако авторы использовали другую клеточную линию. Мы предлагаем использовать клетки НЕК-293, доступные к приобретению в России, которые наиболее часто используются в качестве исходных клеток для получения репортерных линий.
Другим существенным недостатком прототипа (как и ряда других работ [7-9, 12, 14]) является использование временной трансфекции без получения стабильной клеточной линии. Следует отметить, что сама процедура липофекции отрицательно сказывается на физиологии клеток, что может оказывать влияние на достоверность полученных результатов. Так же требуется перед каждым экспериментом повторять процедуру трансфекции клеток, для чего требуется большое количество исходных клеток, реактивов для осуществления трансфекции, плазмиды, кодирующей белок интереса. После осуществления трансфекции (временная экспрессия) только часть клеток популяции экспрессирует целевой белок (5-30% в зависимости от условий трансфекции), уровень экспрессии значительно варьируется от клетки к клетке, так же клетки без селекции могут экспрессировать целевой белок ограниченное время после введения фрагмента его ДНК в клетку, поскольку защитные системы элиминируют введенные чужеродные ДНК-последовательности (спустя 7 и более дней после трансфекции не наблюдалось экспрессии флуоресцентных целевых белков в клеточной популяции, прошедшей трансфекцию). Это приводит к тому, что при каждом эксперименте объект исследований будет несколько различным, что может привести к получению противоречивых результатов исследований.
Получение клеточной линии, стабильно экспрессирующей белок интереса, что осуществлено в некоторых изобретениях [9, 11, 13], более удобно в эксперименте по сравнению с временной экспрессией, однако так же имеет свои недостатки - поскольку получаемая линия поликлональна (имеет в составе множество клеток-родителей), только около половины клеток полученной популяции несут целевые белки, уровень экспрессии от клетки к клетке различается, их содержание падает с течением времени (при постоянном культивировании клеток более 1 месяца в популяции может остаться менее 10% позитивных клеток) за счет случайных мутаций.
В настоящее время не описано постоянных клеточных линий, позволяющих осуществлять прижизненный мониторинг локализации 5-HT4R на мембране и его возможное перераспределение под действием агонистов. Ни одно из существующих технических решений не позволяет проводить серию экспериментов по прижизненной регистрации локализации мышиного серотонинового рецептора типа 5-НТ4 и его перераспределения под воздействием лигандов на мембране клеток.
Задачей изобретения является получение репортерной моноклональной линии клеток, стабильно и на высоком уровне экспрессирующей мышиный рекомбинантный рецептор серотонина типа 5-НТ4, слитый с флуоресцентным белком.
Поставленная задача решается за счет получения клеточной линии НЕК-293/5-HT4R, все клетки которой будут характеризоваться стабильной оверэкспрессией мышиного рецептора серотонина типа 5-НТ4, слитого на С-конце с зеленым флуоресцентным белком GFP, на основе линии клеток почек эмбриона человека НЕК-293.
Техническим результатом является стабильная оверэкспрессия мышиного рекомбинантного рецептора серотонина типа 5-НТ4, слитого на С-конце с зеленым флуоресцентным белком GFP (химерный белок AcGFPl/5-HT4R) в полученной репортерной моноклональной линии HEK-293/5-HT4R. Экспрессия белка AcGFPl/5-HT4R сохраняется в клетках полученной линии как минимум в течение 3-х месяцев непрерывного культивирования.
Приобретенные (экспрессия химерного белка AcGFP1/5-HT4R и гена устойчивости к селективному антибиотику гигромицину) и сохраненные (адгезия, эпителиоподобная морфология с наличием дендрито-подобных отростков у клеток, жизнеспособность и время удвоения популяции) свойства делают полученную репортерную моноклональную линию клеток HEK-293/5-HT4R пригодной для исследования прижизненной динамики локализации 5-HT4R под действием различных лигандов. С помощью дополнительной модификации путем встраивания в клетки флуоресцентного белка-сенсора сАМР для визуализации работы аденилатциклазного сигнального каскада, линия HEK-293/5-HT4R может использоваться для проведения доклинического тестирования потенциальных лекарственных соединений, а так же для детекции секреции серотонина клетками различных типов [16]. Заданные и приобретенные свойства репортерной моноклональной линии клеток НЕК-293/5-HT4R делают ее уникальным объектом для проведения неинвазивного оптического мониторинга в соответствующей области исследований.
Изобретение иллюстрируют графические материалы:
Фиг. 1 Схематичная карта плазмидного вектора pAcGFP1-Hyg-N1, где
pUC ori - область начала репликации (ориджин) плазмиды;
Р CMV IE - конститутивный промотор цитомегаловируса, регулирующий экспрессию целевого белка (5-HT4R);
MCS - кассета множественных сайтов для встраивания последовательности целевого белка, содержащая сайт HindIII - непосредственно сайт встраивания 5-HT4R;
AcGFP1 - последовательность гена флуоресцентного зеленого белка GFP;
SV40 poly А - сигнал полиаденилирования;
SV40 ori Р SV40 е - регуляторный элемент для экспрессии гена устойчивости к гигромицину;
Hyg - ген устойчивости к антибиотику гигромицину для селекции стабильных линий клеток млекопитающих;
Р - бактериальный промотор гена устойчивости к ампициллину;
Amp - Ampicillin resistance gene - ген устойчивости к ампициллину для наработки плазмиды в E.coli.
Фиг. 2 Конфокальные изображения клеток HEK-293/5-HT4R, где
А - фотография клеток в проходящем свете, Б - флуоресценция клеток, индуцированная экспрессией мышиного рекомбинантного рецептора серотонина типа 5-НТ4, слитого на С-конце с зеленым флуоресцентным белком GFP (химерный белок AcGFP1/5-HT4R) (длина волны возбуждения флуоресценции = 496 нм, длина волны эмиссии флуоресценции = 500 - 560 нм); г - те же клетки в проходящем свете. Масштаб изображений указан в правом нижнем углу.
Фиг. 3 Конфокальные изображения клеток в составе разных моноклональных линий HEK-293/5-HT4R в проходящем свете с наложенной флуоресценцией белка AcGFP1/5-HT4R, где:
А - изображение клеток «плохого» клона, который был отбракован; Б - изображение клеток «хорошего» клона, клетки которого были протестированы в физиологических экспериментах и криоконсервированы. Длина волны возбуждения флуоресценции=496 нм, длина волны эмиссии флуоресценции = 500 - 560 нм, масштаб изображений указан в правом нижнем углу.
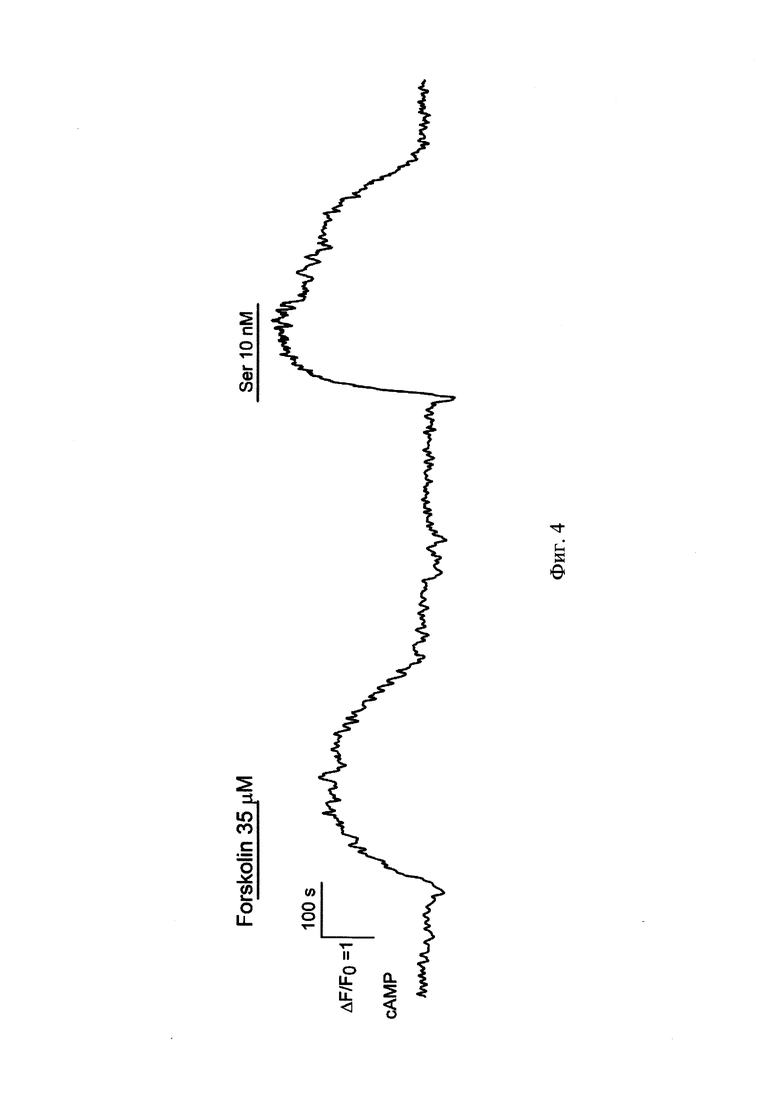
Фиг. 4 Репрезентативная регистрация флуоресценции Pink Flamindo (λex = 572 ± 17.5 λem =634 ± 34 нм) одиночной клетки HEK-293/5-HT4R при стимуляции активатором аденилатциклазы Forskolin (35 мкМ), активатором 5-HT4R серотонином (10 нМ). Моменты и продолжительность аппликаций веществ обозначены горизонтальными линиями выше экспериментальной кривой.
Фиг. 5 Репрезентативная регистрация флуоресценции Pink Flamindo (λex = 572 ± 17.5 нм, λem = 634 ± 34 нм) одиночной клетки HEK-293/5-HT4R при стимуляции активатором 5-HT4R серотонином в разных концентрациях (0,3 - 100 нМ), демонстрирующая дозозависимость клеточных ответов. Моменты, продолжительность аппликаций обозначены горизонтальными линиями выше экспериментальной кривой, над линиями указана концентрация серотонина.
Описание изобретения
Поскольку большинство животных моделей различных заболеваний создано на мышах для удобства содержания, воспроизводства и проведения исследований, представляется актуальным получение клеточной модели с подтипом именно мышиного серотонинового рецептора 5-НТ4, что позволит проводить эксперименты и интерпретировать результаты, сопоставляя их с данными, полученными на животных (мышиных) моделях. В изобретении использована последовательность белка мышиного рецептора серотонина 5-НТ4, соответствующая аминокислотной последовательности NP_032339 мышиного белка 5-НТ4 из базы данных NCBI [https://www.ncbi.nlm.nih.gov/protein/NP_032339.2], соответствующая нуклеотидам 207-1376 последовательности NM_008313.4 из базы данных NCBI [https://www.ncbi.nlm.nih.gov/nuccore/NM_008313.4] и имеющая нуклеотидную последовательность SEQ ID No. 3.
Для экспрессии белка мышиного рекомбинантного серотонинового рецептора типа 5-НТ4 использовалась коммерческая плазмида (вектор) для экспрессии белков в клетках млекопитающих pAcGFP1-Hyg-N1 (Takara Bio) (Фиг. 1), где по сайту HindIII в рамку слияния с геном белка GFP была вставлена последовательность SEQ ID No. 3, соответствующая участку гена мышиного рецептора серотонина 5-НТ4, что позволяет экспрессировать в клетках химерный белок, состоящий из функциональных фрагментов рецептора и флуоресцентного белка GFP. Полученная плазмида pAcGFP1-Hyg-N1/5-HT4R содержит ген устойчивости к ампициллину для наработки плазмиды в E.coli, и ген устойчивости к селективному антибиотику гигромицину для возможности осуществления селективного отбора в клетках млекопитающих [https://www.takarabio.com/documents/Vector%20Documents/PT3837-5.pdf], что позволит при необходимости осуществить дополнительную модификацию клеток плазмидами с генами устойчивости к другим антибиотикам, в частности наиболее распространенному G418 (генецитин).
Флуоресцентный белок GFP, экспрессия которого обусловлена использованием плазмиды pAcGFP1-Hyg-N1, обладает спектральными характеристиками (λex = 498 нм, λem = 522 нм), которые в дальнейшем при необходимости позволят одновременно использовать: 1) белок-сенсор Pink Flamindo (λex = 567 нм, λem = 590 нм) для мониторинга уровня с AMP; 2) флуоресцентный химический Са2+ зонд - Fura-2 (λex = 340 нм, λех =380 нм, λem1 = 520 нм); 3) белок-сенсор GEM-GECO1 (λex = 390 нм, λem1 = 455 нм, λem2 = 511 нм) для мониторинга уровня внутриклеточного Са2+, и т.д. [16-18]. Спектры возбуждения перечисленных белков-сенсоров и красителей не пересекаются, что позволяет осуществлять прижизненный мониторинг локализации рецептора в мембране и его возможную интернализацию, и проводить после дополнительной модификации клеток одновременный и независимый мониторинг работы внутриклеточных сигнальных каскадов, через наблюдение за динамикой внутриклеточных сАМР и Са2+.
Исходным материалом для получения репортерной моноклональной линии клеток HEK-293/5-HT4R является линия НЕК-293 - клетки почки эмбриона человека женского пола, трансформированные ДНК аденовируса типа 5 (Ad 5), которая часто используется как модельная система для встраивания различных мембранных и цитоплазматических белков, и которая доступна для приобретения в России (Российская коллекция клеточных культур (РККК) при Институте Цитологии РАН (Санкт-Петербург, Россия). Исходная линия культивируется в среде DMEM с высоким содержанием глюкозы (4.5 г/л) с добавлением 10% эмбриональной бычьей сыворотки (HyClone), 100 мг/мл гентамицина (Sigma), 2 мМ глутамина (Sigma) (ростовая среда) во влажной атмосфере с 5% содержанием СО2 в воздухе при 37°С как адгезионная культура. Для пересева используется смесь трипсин 0.25%: версен 0.02% (1:2), кратность рассева составляет 1:2 - 1:6, оптимальная плотность 3.0-5.0×104 кл/см2, жизнеспособность 85%.
Клеточная линия HEK-293/5-HT4R получена путем трансфекции клеток исходной линии клеток человека НЕК-293 плазмидой pAcGFP1-Hyg-N1/5-HT4R, содержащей участок гена мышиного серотонинового рецептора 5-НТ4 и последующей многоэтапной селекцией (отбор клеток на антибиотике, отбор на клеточном сортере, визуальный отбор, физиологическое тестирование).
Клетки полученной репортерной моноклональной линии HEK-293/5-HT4R культивируются прикрепленными к пластику до достижения монослоя в среде DMEM с высоким содержанием глюкозы (4.5 г/л) с добавлением 10% эмбриональной бычьей сыворотки (HyClone), 100 мг/мл гентамицина (Sigma), 2 мМ глутамина (Sigma), 200 мкг/мл селективного антибиотика гигромицина во влажной атмосфере с 5% содержанием СО2 в воздухе при 37°С как адгезионная культура. Для пересева используется смесь трипсин 0.25%: версен 0.02% (1:2), кратность рассева составляет 1:2 - 1:4, оптимальная плотность 3.0-5.0×104 кл/см2, время удвоения около 30 часов. Для криоконсервации при температуре -196°С используется смесь из 90% эмбриональной бычьей сыворотки и 10% диметилсульфоксида при концентрации клеток 1-2×106 в 1 мл. Жизнеспособность после криоконсервации составила 75-85% по окраске трипановым синим, после разморозки клетки восстанавливают исходные ростовые и морфологические свойства в течение 2-3 пассажей. Контроль полученной клеточной линии на посторонние контаминанты осуществлялся культивированием в среде без антибиотиков с последующей оценкой с помощью световой микроскопиии и окраской по Хехсту - бактерии, грибы и микоплазма не обнаружены.
Полученная клеточная линия HEK-293/5-HT4R характеризуется экспрессией серотонинового рецептора мыши типа 5-НТ4, слитого с GFP (Фиг. 2, 3), и фактора устойчивости к селективному антибиотику гигромицину, а так же следующими свойствами:
- средней степенью адгезивности;
- эпителиоподобной морфологией;
- клетки равномерно распределяются по дну культуральной посуды при пересеве, прикрепляются к пластику в течение 24-30 часов, распластываются и формируют длинные и дендритоподобные отростки, которые укорачиваются в процессе культивирования, по мере роста и деления образуют сеть на поверхности и приобретают полигональную форму с небольшими отростками, со временем образуют плотный монослой, полностью покрывающий дно культуральной посуды;
- вариативностью формы и размеров (диапазон 10-15 мкм), ядра клеток крупные с неправильной овальной формой, цитоплазма слабозерниста;
- время удвоения роста клеток составляет около 30 часов;
- жизнеспособность клеток составляет 75-85%;
- культивирование клеток проводят в стандартных условиях (подробно описано выше) в присутствии селективного антибиотика гигромицина.
В клеточной популяции полученной репортерной моноклональной клеточной линии HEK-293/5-HT4R все клетки стабильно и на высоком уровне экспрессируют мышиный рекомбинантный рецептор серотонина типа 5-НТ4, сохраняющий физиологическую активность, что позволяет использовать полученную линию для проведения физиологических экспериментов с высокой эффективностью за счет однородности клеточной популяции (Фиг. 3Б). Высокий стабильный уровень экспрессии 5-HT4R в клетках полученной линии сохранялся длительное время - не детектировалось заметного уменьшения экспрессии целевого белка спустя 3 месяца постоянного культивирования клеточного клона. Репортерная моноклональная клеточная линия HEK-293/5-HT4R, экспрессирующая мышиный рекомбинантный рецептор серотонина типа 5НТ4, слитый на С-конце с флуоресцентным белком GFP, может использоваться в in vivo экспериментах по изучению локализации рецептора, в частности, его интернализации во время и после воздействия некоторых препаратов. Полученная линия предполагает как возможность прижизненной визуализации, так и, при необходимости, визуализацию рецептора можно осуществить на обработанных (зафиксированных тем или иным образом) клетках, причем это не требует дополнительных манипуляций (окрашивания антителами), поскольку при обработке происходит фиксация белка рецептора 5-НТ4, слитого с флуоресцентным белком GFP.
Получение репортерной моноклональной линии HEK-293/5-HT4R.
Получение линии клеток HEK-293/5-HT4R, стабильно и на высоком уровне экспрессирующих мышиный серотониновый рецептор типа 5-НТ4, соответствующий аминокислотной последовательности NP_032339 мышиного белка 5-HT4R из базы данных NCBI [https://www.ncbi.nlm.nih.gov/protein/NP_032339.2], слитый с зеленым флуоресцентным белком GFP выполнялось в несколько последовательных этапов (раундов селекции/отбора):
A) На первом этапе получали экспрессионный вектор pAcGFP1-Hyg-N1/5-HT4R, содержащий последовательность участка гена серотонинового рецептора типа 5-НТ4, соответствующий SEQ ID No. 3, для чего последовательно осуществляли:
1) Клонирование полноразмерной кДНК, кодирующей серотониновый рецептор 5-НТ4 из ткани мозга мыши;
2) Амплификацию полноразмерной кДНК рецептора 5-НТ4 мыши;
3) Очистку полученного ПЦР-фрагмента при помощи набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen);
4) Клонирование очищенного ПЦР-фрагмента в вектор pAcGFP1-Hyg-N1 (Takara Bio). Идентичность последовательности 5-HT4R в составе рекомбинантного плазмидного вектора pAcGFP1-Hyg-N1/5-HT4R была подтверждена секвенированием по Сенгеру.
Б) На втором этапе осуществляли трансфекцию клеток НЕК-293 экспрессионным плазмидным вектором pAcGFP1-Hyg-N1/5-HT4R для временной экспрессии белка 5-HT4R.
B) На третьем этапе при помощи многоэтапной селекции (отбора) получали репортерную моноклональную линию клеток HEK-293/5-HT4R, все клетки которой стабильно и на высоком уровне экспрессируют химерный белок AcGFP1/5-HT4R (слитый с флуоресцентным белком GFP и сцепленный с аденилатциклазным каскадом серотониновый рецептор мыши типа 5-НТ4, сохраняющий функциональную активность), процедура получения включала:
1) химическую селекцию путем культивирования трансфицированных на этапе Б) клеток в присутствии антибиотика гигромицина;
2) механический отбор клеток HEK-293/5-HT4R, обладающих наибольшей интенсивностью флуоресценции GFP (λex = 488 нм, λem = 525 ± 20, где: λех - длина волны возбуждения флуоресценции, λem - длина волны эмиссии флуоресценции) с помощью клеточного сортера;
3) визуальный отбор (селекцию) моноклональных линий клеток HEK-293/5-HT4R с помощью флуоресцентного микроскопа по интенсивности свечения GFP и процентному отношению флуоресцирующих клеток в популяции (Фиг. 2, 3) и последующее наращивание отобранных моноклональных линий клеток HEK-293/5-HT4R в количестве, достаточном для проведения физиологических экспериментов и криоконсервации (до 10-20×106 клеток каждого клона);
4) физиологическое тестирование клеток моноклональных линий HEK-293/5-HT4R с помощью метода микрофотометрии по чувствительности клеток к серотонину (пример 7, Фиг. 4, 5) для оценки функциональности рецептора 5-НТ4.
Г) На четвертом этапе проводили криоконсервацию отобранных клонов и их размещение в криохранилище.
Таким образом изобретение состоит в создании репортерной моноклональной линии клеток HEK-293/5-HT4R, полученной на основе клеток линии НЕК-293, экспрессирующей на клеточной мембране мышиный рекомбинантный рецептор серотонина типа 5-НТ4, слитый с флуоресцентным белком GFP. Экспрессия обеспечивается плазмидным вектором pAcGFP1-Hyg-N1/5-HT4R, содержащим участок гена 5-НТ4 рецептора, имеющий нуклеотидную последовательность SEQ ID No. 3, соответствующую номерам нуклеотидов от 207 по 1376 последовательности NM_008313.4 базы данных NCBI. Полученная моноклональная клеточная линия сохраняет уровень экспрессии рецептора серотонина 5-НТ4 в период не менее 3-х месяцев, что обеспечивает многоэтапный отбор: селекция антибиотиком гигромицином, отбор на клеточном сортере, визуальный отбор; депонирована в Государственной коллекции патогенных микроорганизмов и клеточных культур (ГКПМ-Оболенск) при ФБУН ГНЦ ПМБ под регистрационным номером С-4 (свидетельство о депонировании №448).
Изобретение иллюстрируют следующие примеры:
Пример 1. Получение экспрессионного вектора pAcGFP1-Hyg-N1/5-HT4R
Для экспрессии мышиного серотонинового рецептора типа 5-HT4R плазмидный вектор pAcGFP1-Hyg-N1/5-HT4R должен содержать вставку участка гена, кодирующего последовательность мышиного рецептора серотонина 5-НТ4 без стоп-кодона и имеющего нуклеотидную последовательность SEQ ID No. 3, соответствующую аминокислотной последовательности NP_032339 мышиного белка 5-HT4R из базы данных NCBI для чего последовательно осуществляли:
1) Клонирование полноразмерной кДНК, кодирующей серотониновый рецептор 5-НТ4 из ткани мозга мыши.
Для этого из образца мозга мыши выделяли тотальную РНК с помощью набора реагентов RNeasy Mini Kit (Qiagen). РНК обрабатывали ДНказой I (BioLabs) при следующем температурном режиме: 10 мин - 37°С, 10 мин - 75°С. Реакцию обратной транскрипции проводили при помощи коммерческого набора реактивов Superscript IV Reverse Transcriptase (Invitrogen) по протоколу производителя: 20 мкл реакционной смеси содержали реакционный буфер, смесь дезоксирибонуклеотидов, 5 мМ DTT, 2,5 мкМ и рандомные праймеры, 150 нг РНК. Температурный режим проведения реакции: 50°С - 10 мин, 80°С- 10 мин.
2) Амплификацию полноразмерной кДНК рецептора 5-НТ4 мыши проводили с использованием ДНК-полимеразы Phusion Hot Start II DNA Polymerase (ThermoFisher), одноцепочечной кДНК в качестве матрицы, полученной на первом этапе, и ген-специфичных праймеров TATAAGCTTCCTGTAATGGACAAACTTGATG (SEQ ID No. 1) и TATAAGCTTAGTATCACTGGGCTGAGC (SEQ ID No. 2), подобранных на основании последовательности NM_008313.4, представленной в базе данных NCBI [https://www.ncbi.nlm.nih.gov/nuccore/NM_008313.4]. В состав обоих ген-специфичных праймеров входит сайт рестрикции HindIII для вставки в плазмидный вектор pAcGFP1-Hyg-N1 (выделено курсивом и подчеркнуто). Прямой (forward) специфичный праймер также содержит последовательность Козак (выделена жирным шрифтом). Размер ожидаемого продукта реакции - 1169 п.н. Температурный режим программы амплификации: денатурация при 98°С в течение 30 с, затем 30 циклов, включающих денатурацию при 98°С в течение 10 секунд, отжиг праймеров при температуре 69°С в течение 15 секунд и элонгацию при 72°С в течение 30 с. Размер полученного ПЦР-фрагмента при проведении электрофореза совпал с ожидаемым.
3) Очистку полученного ПЦР-фрагмента при помощи набора для экстракции из геля QIAquick Gel Extraction Kit (Qiagen).
4) Клонирование очищенного ПЦР-фрагмента в вектор pAcGFP1-Hyg-N1 (Фиг. 1) (Takara Bio), осуществляли по сайту HindIII в рамку слияния с белком GFP. Последовательность 5-HT4R не содержит стоп-кодон, поэтому транскрипция рецептора происходит вместе с последовательностью флуоресцентного белка GFP, в результате чего в клетках млекопитающих в дальнейшем экспрессируется химерный рекомбинантный белок AcGFP1/5-HT4R. Встройка в экспрессионную плазмиду происходит таким образом, что GFP располагается в С-концевой области белка рецептора и не влияет на конформацию рецепторного белка и его чувствительность к лигандам. В дальнейшем зеленая флуоресценция GFP позволяет визуализировать экспрессию и локализацию рекомбинантного белка 5-HT4R в трансфицированных клетках и не мешает проведению физиологических экспериментов (Фиг. 3).
5) Идентичность последовательности 5-HT4R в составе рекомбинантного плазмидного вектора pAcGFP1-Hyg-N1/5-HT4R была подтверждена секвенированием по Сенгеру.
Полученная экспрессионная плазмида pAcGFP1-Hyg-N1/5-HT4R включает:
1) фрагмент ДНК, содержащий последовательность Козак длиной 6 п.о.; последовательность ДНК химерного белка AcGFP1/5-HT4R (кодирует мышиный рецептор серотонина 5-НТ4 размером 1169 п.о. без стоп-кодона (имеющий нуклеотидную последовательность SEQ ID No. 3) слитый с зеленым флуоресцентным белком AcGFP1);
2) регуляторные элементы, обеспечивающие экспрессию химерного белка AcGFP1/5-HT4R в клетках млекопитающих: промотор-энхансер цитомегаловируса (CMV) и сигнал полиаденилирования SV40;
3) ген устойчивости к антибиотику гигромицину (Hyg) для селекции стабильных клеточных линий млекопитающих, промотор и сигнал полиаденилирования вируса SV40 для его экспрессии;
4) промотер pUC, обеспечивающий конститутивную репликацию плазмиды в клетках E. coli.;
5) ген устойчивости к антибиотику ампициллину (Amp) и бактериальный промотор гена устойчивости к ампициллину, позволяющие проводить препаративную наработку плазмиды в E. coli.
Пример 2. Культивирование и описание материнской линии клеток НЕК-293
Клетки линии НЕК-293 (Российская коллекция клеточных культур позвоночных) - гипотриплоидная линия клеток почки эмбриона человека, трансформированных ДНК аденовируса типа 5 [19]. Растут прикрепленными к пластику в монослое, имеют эпителиоподобную структуру. Условия культивирования: среда ЕМЕМ + NEAA 1% или DMEM (ИНЦ РАН) с высоким содержанием глюкозы (4.5 г/л) с добавлением 10% эмбриональной бычьей сыворотки или инактивированной лошадиной, 100 мг/мл гентамицина (Sigma), 2 мМ глутамина (Sigma) (ростовая среда) во влажной атмосфере с 5% содержанием СО2 в воздухе при 37°С. Процедура пересева включает снятие клеток, используя смесь трипсин 0.25%: версен 0.02% (1:2 - 1:3), кратность рассева составляет 1:2 - 1:3, оптимальная плотность 3.0-5.0×104 кл/см2, клетки прикрепляются к субстрату в течение суток - нескольких дней. Криоконсервация осуществляется в сыворотке или ростовой среде с добавлением 10% DMSO, плотность клеток в криоконсервирующем растворе - 2-5×106/мл. Жизнеспособность после криоконсервации по окраске трипановым синим на нулевом пассаже составляет 90%. Контаминации бактериями, грибами и микоплазмой не обнаружено. Видовая идентичность контролировалась кариологическим анализом; кариология: 2n = 46, модальное число хромосом 72, количество маркеров - 12 (дифференциальная окраска), количество полиплоидов - 2.4%. ДНК профиль по STR: Amelogenin: X, X; CSF1PO: 11, 12; D13S317: 12,12; D16S539: 9,13; D5S818: 8, 9; D7S820: 11,12; THO1: 7, 9.3; ТРОХ: 11, 11; vWA: 16, 19. Линия НЕК-293 характеризуется чувствительностью к аденовирусам человека и астровирусам; присутствием и экспрессией трансформирующих генов Ad 5. Клетки линии НЕК-293 туморогенны в иммунодефицитных мышах линии nude. Основная область применения: биотехнология (титрование аденовирусов человека), вирусология, трансформация. Линия клеток НЕК-293 (РКККП) соответствует линиям в других коллекциях под обозначениями: №300192*CLS, CRL-1573*ATCC, №85120602*ЕСАСС.
Пример 3. Трансфекция клеток НЕК-293 экспрессионным плазмидным вектором pAcGFP1-Hyg-N1/5-HT4R для временной экспрессии белка 5-HT4R
Накануне трансфекции клетки НЕК-293 рассевали в ростовой среде из расчета 5-10×105 клеток в 1 лунку 12-лучночного планшета. К 800 мкл ростовой среды в лунку добавляли трансфекционную смесь, которая содержала 200 мкл среды OptiMEM (Gibco), 2 мкл Р3000 Reagent (Invitrogen), 2 мкл Lipofectamine 3000 (Invitrogen), 1 мкг плазмидного вектора pAcGFPl-Hyg-N1/5-HT4R. Через 24 ч оценивали эффективность трансфекции непосредственно в планшете на инвертированном флуоресцентном микроскопе по интенсивности флуоресценции GFP. Далее клетки использовали для получения стабильной репортерной моноклональной клеточной линии HEK-293/5-HT4R.
Пример 4. Получение репортерной моноклональной клеточной линии НЕК-293/5-HT4R
Для получения стабильной репортерной моноклональной клеточной линии НЕК-293/5-HT4R, экспрессирующей мышиный серотониновый рецептор типа 5-НТ4, слитый с флуоресцентным белком GFP, использовали клетки, временно экспрессирующие 5-HT4R (химерный белок AcGFP1/5-HT4R), спустя 24 часа после трансфекции. Клетки в лунке 12 луночного планшета промывали раствором Версена (Sigma) и обрабатывали 0.125% раствором трипсина (Sigma), затем рассевали в соотношении 1:10 в чашку Петри 60 мм.
Первый раунд селекции: для осуществления химической селекции в чашку Петри к клеткам спустя 24 часа после пересева (48 часов после трансфекции) непосредственно в ростовую среду добавляли 1000 мкг (200 мкг/мл) селективного антибиотика гигромицина (Hygromycin В, InvivoGen), так как использованная плазмида несет ген устойчивости к нему. В присутствии селективного антибиотика клетки культивировали в течение 2-3 недель, каждые 2-3 дня среда менялась на свежую с добавлением 200 мкг/мл гигромицина и последующее наращивание клеток, стабильно экспрессирующих 5-HT4R до количества, достаточного для проведения отбора с помощью клеточного сортера (5-10×106 клеток). За это время большая часть клеток гибла, погибшие клетки всплывали и удалялись при смене среды, выживали только клетки, несущие плазмиду pAcGFP1-Hyg-N1/5-HT4R и способные экспрессировать ген устойчивости к гигромицину. Выжившие клетки сохраняли способность к адгезии и образовывали на дне культуральной посуды неравномерные колонии, когда большая часть колоний достигла размеров 2-6 мм, клетки были пересеяны в новую культуральную чашку Петри, где образовали монослой. В результате химической селекции была получена поликлональная клеточная линия НЕК-293/5-HT4R, стабильно экспрессирующая 5-HT4R, слитый с GFP (химерный белок AcGFP1/5-HT4R) примерно в 50% клеточной популяции, то есть при визуальной оценке уровня экспрессии целевого белка клетки в популяции заметно варьировали - от видимого отсутствия до достоверно визуально детектируемого уровня флуоресценции разной интенсивности.
Второй раунд селекции: полученную популяцию клеток использовали для получения клеточных моноклонов с высоким уровнем экспрессии белка 5-HT4R с помощью механического отбора клеточным сортером FACSAria SORP (BD Biosciences) по интенсивности флуоресценции GFP. Накануне сортировки 3-4×106 клеток переносили в 25 см2 культуральный флакон в 5 мл ростовой среды. Непосредственно перед сортировкой клетки однократно обрабатывали раствором Версена и затем инкубировали в 0.5 мл 0.125% раствора трипсина до полного открепления клеток от пластика, ферментативную реакцию останавливали добавлением 1 мл ростовой среды и переносили клетки в коническую микропробирку объемом 1,5 мл. Затем клетки осаждали центрифугированием при 50 g и ресуспендировали в 1 мл среды для сортировки клеток на основе фосфатного буфера (рН 7.4) и 1% эмбриональной бычьей сыворотки. Для получения суспензии одиночных клеток их пропускали через клеточное сито с размером ячейки 40 мкм (Corning cell strainer, Sigma). Полученную суспензию клеток анализировали с помощью сортера клеток FACSAria SORP (BD Biosciences). Флуоресценцию клеток возбуждали при длине волны 488 нм, эмиссию регистрировали в области 523 ± 23 нм. Для получения моноклонов отбирали клетки, обладающие наибольшей интенсивностью флуоресценции, и с помощью программы клеточного сортера помещали их по одной в каждую лунку 96-луночного планшета (всего было отобрано 3 планшета), содержащую 150 мкл ростовой среды с повышенным до 15% содержанием сыворотки. Планшеты с клетками помещали в СО2-инкубатор, на 7 день культивирования в лунки добавляли по 50 мкл ростовой среды, содержащей 10% сыворотки. На 14 день проводили частичную смену среды в лунках, для этого из каждой лунки отбирали 100 мкл старой среды и затем добавляли такой же объем новой ростовой среды, содержащей 200 мкг/мл гигромицина. На 17 день проводили первичный визуальный контроль наличия в лунках живых моноклонов и оценку интенсивности их флуоресценции с использованием инвертированного флуоресцентного микроскопа Axiovert 135 (Zeiss), флуоресценцию клеток возбуждали при 495 ± 5 нм, эмиссию регистрировали в области 545 ± 25 нм. Всего в трех 96-луночночных планшетах в 83 лунках наблюдались выжившие клетки, из которых: в 19-ти лунках клетки не обладали визуально детектируемой флуоресценцией; в 17 лунках наблюдался более чем один клон - в лунке присутствовали или несколько точек роста или две популяции клеток - со свечением и без - такие клетки не исследовались далее; в 30 лунках клетки в популяции клона сильно различались по интенсивности свечения или обладали низкой его интенсивностью; в 5 лунках клеточные клоны прекратили деление и не достигли размера, достаточного для перенесения в культуральную посуду большего размера.
Третий раунд селекции: в результате для дальнейшего наращивания (культивирование подробно описано в примере 5) было отобрано 12 клонов по наиболее яркой и однородной флуоресценции клеток в составе клона (первичный визуальный отбор), эти клоны были пересеяны на 24-луночный планшет. По мере нарастания клеточные клоны пересаживали в культуральную посуду большей площади, последовательно в 12- и 6 луночный. На этом этапе часть клеток каждого клона оценивали визуально на конфокальном микроскопе (вторичный визуальный отбор) для визуализации локализации рецептора и оценки однородности клеточной популяции по содержанию клеток с флуоресценцией GFP. По результатам визуального тестирования (Фиг. 2, 3) было отобрано 6 моноклональных клеточных линий, 100% клеток которых обладало флуоресценцией GFP, локализованной в мембране клеток. Поскольку GFP слит с рецептором 5-HT4R (химерный белок AcGFP1/5-HT4R), это подтверждает и локализацию белка рецептора.
Данный пример, в частности, демонстрирует возможность использования полученной клеточной линии для наблюдения локализации мышиного рекомбинантного рецептора серотонина типа 5-НТ4 и ее изменения под воздействием лигандов, лекарственных соединений и физиологических агентов.
Четвертый раунд селекции: оценка функциональных свойств 5-HT4R, экспрессируемого клетками HEK-293/5-HT4R по чувствительности клеток к серотонину с помощью дополнительной модификации клеток плазмидой Pink Flamindo и физиологических экспериментов методом микрофотометрии с использованием регистрации флуоресцентного сигнала от генетически кодируемого белка-сенсора сАМР Pink Flamindo (пример 7). В результате для криоконсервации (пример 6) было отобрано 4 моноклональных клеточных линии, клетки которых характеризовались наибольшей амплитудой флуоресцентного сигнала, полученного от Pink Flamindo в ответ на аппликацию 10 нМ серотонина.
Пример 5. Культивирование клеточных клонов HEK-293/5-HT4R
Все полученные репортерные моноклональные клеточные линии HEK-293/5-HT4R сохранили морфо-функциональные характеристики материнской линии НЕК-293. Полученные линии клеток обладают адгезионными характеристиками, сопоставимыми с материнской линией НЕК-293, и растут прикрепленными к пластику в монослое, сохранили эпителиоподобную структуру (клетки имеют отростки и некоторую зернистость цитоплазмы). Жизнеспособность клеточных линий составляет 75-85% по тестированию окрашиванием трипановым синим, время удвоения клеточной популяции около 30 часов.
Все клетки линии экспрессируют слитый с флуоресцентным белком GFP целевой рецепторный белок 5-HT4R (химерный белок AcGFP1/5-HT4R), сохраняющий функциональность, и ген устойчивости к селективному антибиотику гигромицину.
Условия культивирования линии HEK293/5-HT4R: среда DMEM (Gibco) с высоким содержанием глюкозы (4.5 г/л) с добавлением 10% эмбриональной бычьей сыворотки (HyClone), 100 мг/мл гентамицина (Sigma), 200 мкг/мл селективного антибиотика гигромицина (Hygromycin В, InvivoGen), 2 мМ глутамина (Sigma) (ростовая среда) во влажной атмосфере с 5% содержанием CO2 в воздухе при 37°С. Для пересева последовательно удаляли старую ростовую среду, аккуратно отмывали клетки раствором Версена и обрабатывали 0.125% трипсином, кратность рассева составляла 1:2 - 1:4. Оптимальная плотность клеток при культивировании 1-3×105 кл/мл, пересев рекомендован 2-3 раза в неделю. После хранения в жидком азоте выживаемость составляет 75-85%, клетки восстанавливают физиологические и морфологические свойства в течение 2-3 пассажей.
Пример 6. Криоконсервация клеток HEK-293/5-HT4R
Для криоконсервации клетки 3х отобранных в результате 4х раундов селекции клеточных клонов наращивались до объема 20-25×106 клеток, а 1 клон - до объема 60×106 клеток (приготовление большего количества криоампул для параллельного депонирования). Предпочтительная плотность клеточной культуры при заморозке - 80%. Клетки промывали Версеном, обрабатывали 0,125% трипсином, ресуспендировали в присутствии ростовой среды и центрифугировали при 1,2 g в течении 2 минут. Супернатант удаляли, полученный осадок ресуспендировали в сыворотке с добавлением 10% DMSO, плотность клеток в среде для криоконсервации составляла 2-5×106/мл, затем клетки размещались в несколько криоампул, по 2-3×106 клеток в 1 ампуле для криоконсервации. Далее криопробирки с клетками помещали в криобокс и в холодильник с температурой -80°С, спустя 1-3 дня криоампулы переносили в жидкий азот.
Пример 7. Применение репортерных моноклональных линий НЕК-293/5-HT4R для физиологических методов исследования внутриклеточной сигнализации, запускаемой лигандами, активирующими/ингибирующими 5-HT4R.
Клеточная линия HEK-293/5-HT4R может использоваться для прижизненных физиологических методов исследования внутриклеточной сигнализации, запускаемой лигандами, активирующими/ингибирующими 5-HT4R. Известно, что 5-НТ4 рецептор серотонина сопряжен с аденилатциклазным сигнальным каскадом, то есть активация этих рецепторов приводит к изменению концентраций внутриклеточного сАМР. Таким образом, стимуляция клеток HEK-293/5-HT4R серотонином опосредует активацию аденилатциклазы и приводит к повышению концентрации внутриклеточного сАМР, как и в случае воздействия на клетки форсколином (специфический активатор аденилатциклазы) непосредственно [16-18]. Осуществить неинвазивный оптической мониторинг уровня внутриклеточного сАМР на клетках НЕК-293/5-HT4R можно посредством встраивания в клетки полученной линии генетически кодируемого флуоресцентного белка сенсора цитоплазматического сАМР - Pink Flamindo [17].
Процедура подготовки клеток для физиологических экспериментов методом микрофотометрии с использованием регистрации флуоресцентного сигнала от генетически кодируемого белка-сенсора с AMP Pink Flamindo включает:
1) Встраивание в клетки HEK-293/5-HT4R плазмидного вектора Pink Flamindo (#102356, Addgene). Для этого 5-10×105 клеток отсаживали на 1 лунку 12 луночного планшета и через сутки осуществляли липофекцию плазмидного вектора Pink Flamindo, для чего к клеткам в 800 мкл ростовой среды добавляют трансфекционную смесь, содержащую 200 мкл среды OptiMEM, 2 мкл Р3000 Reagent, 2 мкл Lipofectamine 3000, 1 мкг плазмидного вектора Pink Flamindo.
2) Проведение физиологических экспериментов осуществляется на клетках НЕК-293/5-HT4R с временной экспрессией белка-сенсора Pink Flamindo спустя 24-48-72 часа после трансфекции. Оценка активности аденилатциклазного сигнального каскада методом микрофотометрии осуществляется по характеристикам флуоресцентного сигнала, полученного от Pink Flamindo (λex = 572 ± 17.5 нм, λem = 634 ± 34 нм) в ответ на стимуляцию активатором аденилатциклазы Forskolin (10-50 мкМ) и лигандом 5-НТ4 рецептора серотонином (0,1-10 нМ). Клетки снимали 0,125% трипсином и ресуспендировали в 1 мл ростовой среды, затем 50 мкл клеточной суспензии помещали в одноразовые фотометрические камеры, где клетки прикреплялись ко дну с помощью адгезивного материала Cell Так (Corning), и выдерживали при комнатной температуре 30 мин. Внеклеточный раствор содержал (мМ): NaCl - 110, KCl - 5.5, CaCl2 - 2, MgCl2 -0.8, HEPES - 10, глюкозу - 10. Фотометрические эксперименты проводили с использованием инвертированного флуоресцентного микроскопа Axiovert 135 (Zeiss), оборудованного объективом Plan NeoFluar 20×/0.75 и цифровой ECCD камерой LucaR (Andor Technology). Флуоресценцию клеток возбуждали при 572 ± 17.5 нм, эмиссию регистрировали в области 634 ± 34 нм. Количественный фотометрический анализ изображений осуществляли с использованием программы NIS Elements (Nikon). В ходе физиологических экспериментов клетки, обладающие флуоресценцией Pink Flamindo, демонстрировали обратимое увеличение ее интенсивности в ответ на стимуляцию Forskolin (10-50 мкМ) и серотонином (1-100 нМ), что соответствует существующим представлениям о работе 5-НТ4 рецептора (Фиг. 4, 5).
Данный пример демонстрирует возможность использования полученной клеточной линии не только для исследования изменения локализации белка рецептора, но и для исследования внутриклеточного аденилатциклазного сигнального пути, активированного лигандами (потенциальными лекарственными соединениям) мышиного рецептора серотонина типа 5-НТ4.
Экспериментальная процедура, описанная в данном примере, использовалась для подтверждения физиологической функциональности серотонинового рецептора мыши типа 5-HT4R, встроенного в клетки репортерной линии HEK-293/5-HT4R. Для этого от каждого клеточного клона HEK-293/5-HT4R отсевали в 12-ти луночный планшет небольшую часть клеток (5-10×105) и в эти клетки встраивался плазмидный вектор Pink Flamindo с помощью набора для липофекции Lipofectamine 3000. Спустя 24-72 часа после трансфекции клетки HEK-293/5-HT4R+PinkFlamindo тестировались физиологически как описано выше.
Промышленная применимость
Описываемая в настоящем изобретении репортерная моноклональная клеточная линия HEK-293/5-HT4R позволяет проводить прижизненную визуализации локализации рецептора в клетках. Преимуществом применения клеток HEK-293/5-HT4R, получаемых описываемым способом, является возможность проведения неинвазивного оптического мониторинга локализации рецептора в мембране и его интернализации под воздействием лигандов - потенциальных лекарственных препаратов, мишенью которых является 5-НТ4 рецептор.
Источники информации:
1. Barnes N.M. et al. International Union of Basic and Clinical Pharmacology. CX. Classification of Receptors for 5-hydroxytryptamine; Pharmacology and Function. Pharmacol Rev. 2021. 73(1): 310-520.
2. McCorvy JD, Roth BL. Structure and Function of Serotonin G protein Coupled Receptors. Pharmacol Ther. 2015. 150:129-142.
3. Dong C. et al. Psychedelic-inspired drug discovery using an engineered biosensor. Cell. 2021. 184(10):2779-2792.e18.
4. Wan J. et al. A genetically encoded sensor for measuring serotonin dynamics. Nat Neurosci. 2021. 24(5):746-752.
5. Riegert C. et al. Increased expression of 5-HT(1B) receptors by Herpes simplex virus gene transfer in septal neurons: New in vitro and in vivo models to study 5-HT(1B) receptor function. Brain Res Bull. 2008. 76(4): 439-53.
6. Bhattacharyya S. et al. Internalization and recycling of 5-HT2A receptors activated by serotonin and protein kinase C-mediated mechanisms. Proc Natl Acad Sci USA. 2002. 99(22): 14470-5.
7. Gerald C. et al. Uses of the 5-HT4 receptor. Патент США US 6,331,401 B1, опубл. 1992-12-24/2001-12-18.
8. Gerald C. et al. DNA encoding 5-HT4 serotonin receptors and uses thereof. Патент США US5766879 (А), опубл. 1998-06-16.
9. Fischmeister R. et al. Splice variants for human 5-HT4 serotonin receptor and their applications, in particular for screening. Патент США US 6,506,580 B1, опубл. 1997-11-28 / 2003-01-14.
10. Golz S. et al. Diagnostics and therapeutics for diseases associated with 5-hydroxytryptamine (serotonin) receptor 4 (5-HT4). Патент США US 2007/0065801 A1, опубл. 2007-03-22/2003-01-14. Международный номер WO 2004/072651 A2, WO/2004/104577.
11. Konen J.R. et al. Prokinetic actions of luminally acting 5-HT4 receptor agonists. Neurogastroenterol Motil. 2021. 33(4):e14026.
12. Pellissier L.P. et al. G protein activation by serotonin type 4 receptor dimers:
evidence that turning on two protomers is more efficient. J Biol Chem. 2011. 286(12):9985-97.
13. Mnie-Filali O. et al. Serotonin 4 receptor (5-HT4R) internalization is isoform-specific: Effects of 5-HT and RS67333 on isoforms A and B. Cellular Signalling. 2010. 22(3):501-509.
14. Joubert L. et al. New sorting nexin (SNX27) and NHERF specifically interact with the 5-HT4(a) receptor splice variant: roles in receptor targeting. J Cell Sci. 2004. 117(22):5367-5379.
15. Schill Y. et al. Serotonin 5-HT4 receptor boosts functional maturation of dendritic spines via RhoA-dependent control of F-actin. Commun Biol. 2020. 14; 3(1): 76.
16. Cherkashin A.P. et al. Taste Cells of the Type III Employ CASR to Maintain Steady Serotonin Exocytosis at Variable Ca2+ in the Extracellular Medium. Cells. 2022. 11(8): 1369.
17. Harada K. Red fluorescent protein-based cAMP indicator applicable to optogenetics and in vivo imaging. Scientific Reports. 2017. 7(1), 7351.
18. Кочкина E.H., Рогачевская O.A., Котова П.Д. Анализ клеточных ответов на норадреналин с использованием синхронного мониторинга Са2+ и сАМР в клетках НЕК 293. Биологические мембраны. 2021. 38(6), с 474-477.
19. Атлас хромосом постоянных линий человека и животных, С.Е. Мамаева, 2002. М. Научный мир.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="5-НТ4R,
ЭКСПРЕССИРУЮЩАЯ МЫШИНЫЙ РЕКОМБИНАНТНЫЙ СЕРОТОНИНОВЫЙ РЕЦЕПТОР ТИПА
5НТ4.xml" softwareName="WIPO Sequence" softwareVersion="2.2.0"
productionDate="2022-11-23">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022125909/20(056519)</ApplicationNumberText>
<FilingDate>2022-10-04</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>-</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022125909/20(056519)</ApplicationNumberText>
<FilingDate>2022-10-04</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки «Федеральный исследовательский центр
Пущинский научный центр биологических исследований Российской
академии наук</ApplicantName>
<ApplicantNameLatin>Federal Research Center Pushchino Scientific
Center for Biological Research of the Russian Academy of
Sciences</ApplicantNameLatin>
<InventionTitle languageCode="ru">РЕПОРТЕРНАЯ МОНОКЛОНАЛЬНАЯ
КЛЕТОЧНАЯ ЛИНИЯ НЕК-293/5-НТ4R, ЭКСПРЕССИРУЮЩАЯ МЫШИНЫЙ
РЕКОМБИНАНТНЫЙ СЕРОТОНИНОВЫЙ РЕЦЕПТОР ТИПА 5НТ4.</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>31</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..31</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tataagcttcctgtaatggacaaacttgatg</INSDSeq_sequence
>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>27</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..27</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tataagcttagtatcactgggctgagc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>1170</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1170</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q3">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgtaatggacaaacttgatgctaatgtgagttccaacgagggtttca
ggtccgtggagaaggtcgtgctgctcacgttccttgcagtggttatcctgatggccatcttgggcaacct
gctggtgatggtggctgtgtgcagggacaggcagctcaggaaaataaaaaccaactatttcattgtgtct
ctcgcctttgctgacctgctggtttcggtgctggtgatgccctttggtgccattgagctggtccaagaca
tctgggcttatggggagatgttctgcctggtccggacctctctggatgtcctacttaccacagcatcgat
ctttcacctgtgctgtatttccctggacaggtattacgccatctgctgccagcctttggtttataggaac
aagatgacccctctacgcatcgcattaatgttgggaggctgctgggtccttcccatgtttatatcttttc
tccccataatgcaaggctggaacaacatcggcatagttgatgtgatagagaaaaggaaattcagccacaa
ctctaactccacgtggtgtgtcttcatggtcaacaagccctatgctatcacctgctctgtggtggccttc
tacatcccgtttctcctcatggtgctggcctattaccgaatctatgtcactgctaaggagcatgcccagc
agatacagatgttacaacgggcaggagccacctctgaaagcaggccccagccagctgaccagcacagcac
acatcgcatgaggacagagaccaaggcagccaagactttatgtgtcatcatgggctgcttctgtttctgc
tgggcccccttctttgtcaccaatattgtggaccctttcatagactacactgtccccgagcaggtgtgga
ctgctttcctctggcttggctatatcaattcggggttgaacccttttctctatgccttcttgaataagtc
tttcagacgtgccttcctcatcatcctctgctgtgatgatgagcgctacaaaagaccccccattctgggc
cagactgtcccctgttcaaccacaaccattaatggatccacccatgtactaagggatacagtggaatgtg
gtggccagtgggagagtcggtgtcacctcacagcaacttctcctttggtggctgctcagcccagtgatac
t</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЛУОРЕСЦЕНТНЫЕ РЕПОРТЕРНЫЕ СИСТЕМЫ ДЛЯ ОЦЕНКИ ЭПИТЕЛИАЛЬНОГО И/ИЛИ МЕЗЕНХИМАЛЬНОГО СОСТОЯНИЯ КЛЕТКИ | 2017 |
|
RU2705251C2 |
| ПРОМОТОР ДЛЯ ТКАНЕСПЕЦИФИЧЕСКОЙ ЭКСПРЕССИИ ГЕНОВ В ГЕРМИНАЛЬНЫХ ТКАНЯХ МЛЕКОПИТАЮЩИХ | 2010 |
|
RU2459870C2 |
| КОМПОЗИЦИЯ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ ТРАНСГЕНА В ЭУКАРИОТИЧЕСКИХ КЛЕТКАХ И СПОСОБ УВЕЛИЧЕНИЯ ПРОДУКЦИИ ЦЕЛЕВОГО БЕЛКА, КОДИРУЕМОГО ТРАНСГЕНОМ | 2013 |
|
RU2546249C2 |
| ПЛАЗМИДА ДЛЯ ВЫЯВЛЕНИЯ МЕЗЕНХИМАЛЬНОГО СОСТОЯНИЯ КЛЕТКИ | 2019 |
|
RU2715643C1 |
| ПЛАЗМИДА ДЛЯ ВЫЯВЛЕНИЯ ЭПИТЕЛИАЛЬНОГО СОСТОЯНИЯ КЛЕТКИ ЧЕЛОВЕКА | 2019 |
|
RU2716054C1 |
| ЛЕНТИВИРУСНАЯ РЕПОРТЕРНАЯ ВЕКТОРНАЯ СИСТЕМА ДЛЯ ИЗМЕРЕНИЯ АКТИВНОСТИ КЛЮЧЕВЫХ ТРАНСКРИПЦИОННЫХ ФАКТОРОВ, ОТРАЖАЮЩИХ ФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ СИГНАЛЬНЫХ ПУТЕЙ КЛЕТКИ | 2009 |
|
RU2420584C1 |
| ИДЕНТИФИКАЦИЯ МОЛЕКУЛ, МОДУЛИРУЮЩИХ БЕЛОК-БЕЛКОВОЕ ВЗАИМОДЕЙСТВИЕ | 2008 |
|
RU2476891C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2015 |
|
RU2699715C2 |
| ПРИМЕНЕНИЕ ВИТАМИНОВ И ГЕНОВ И БЕЛКОВ МЕТАБОЛИЗМА ВИТАМИНОВ ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2016 |
|
RU2736705C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 2014 |
|
RU2711322C1 |




Изобретение относится к биотехнологии. Предложена репортерная моноклональная линия клеток HEK-293/5-HT4R, экспрессирующая мышиный рекомбинантный рецептор серотонина типа 5-НТ4, слитый на С-конце с зеленым флуоресцентным белком GFP, и депонированная в Государственной коллекции патогенных микроорганизмов и клеточных культур (ГКПМ-Оболенск) при ФБУН ГНЦ ПМБ под регистрационным номером С-4. Изобретение обеспечивает стабильную оверэкспрессию мышиного рекомбинантного рецептора серотонина типа 5-НТ4, слитого на С-конце с зеленым флуоресцентным белком GFP. 5 ил., 7 пр.
Репортерная моноклональная линия клеток HEK-293/5-HT4R, экспрессирующая мышиный рекомбинантный рецептор серотонина типа 5-НТ4, слитый на С-конце с зеленым флуоресцентным белком GFP, и депонированная в Государственной коллекции патогенных микроорганизмов и клеточных культур (ГКПМ-Оболенск) при ФБУН ГНЦ ПМБ под регистрационным номером С-4.
| CHERKASHIN A.P | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Cells | |||
| Способ получения продуктов конденсации фенолов с формальдегидом | 1924 |
|
SU2022A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9030112/ Дата обращения 06.06.2023 | |||
| MNIE-FILALI O | |||
| et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
