ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам и композициям для лечения глазных болезней; и в частности, но не только, изобретение относится к лечению глаукомы. В предпочтительных вариантах осуществления настоящее изобретение относится к применению технологии РНК-интерференции (РНКi) для подавления экспрессии генов образования водянистой влаги и генов оттока водянистой влаги. Также настоящее изобретение относится к способам и композициям для лечения глазных болезней.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
РНКi как инструмент подавления экспрессии гена
Для определения функции гена у млекопитающих обычно используют направленное воздействие на гены посредством гомологичной рекомбинации, но этот способ является дорогостоящим и отнимает много времени. С другой стороны, функции многих генов можно определить после ингибирования мРНК рибозимами или антисмысловыми технологиями. Несмотря на успешное применение этих технологий в некоторых ситуациях, существуют трудности для их универсального применения. Появление направленного "нокдауна" малыми интерферирующими РНК (siРНК) дало импульс революционным изменениям в генетике соматической клетки, давая возможность осуществлять недорогой и быстрый анализ функции гена у млекопитающих.
Последние 15 лет в молекулярной биологии постоянной темой является создание удобного и надежного способа экспрессии генного нокаута на уровне мРНК. В попытках создать клетки или организмы с потерей функции проводились исследования различных молекул, включая, например, антисмысловые последовательности, рибозимы и химерные олигонуклеотиды, но конструирование таких молекул было основано на методе проб и ошибок в зависимости от свойств гена-мишени. Кроме того, было трудно прогнозировать желаемые эффекты, и часто достигалась всего лишь слабая супрессия (Braasch&Corey, 2002).
Впервые феномен dsРНК (двунитевая РНК) был открыт у растений в начале 1990-ых годов, и позже, в 1998 году, исследователи Andy Fire и Craig Mello в работах с червем Caenorhabditis elegans впервые продемонстрировали, что dsРНК может чрезвычайно эффективным образом специфично и селективно ингибировать экспрессию гена (Fire et al., 1998). В эксперименте последовательность первой нити (так называемая смысловая РНК) совпадает с последовательностью соответствующей области матричной РНК-мишени (мРНК). Вторая нить (антисмысловая РНК) комплементарна этой мРНК. Получаемая dsРНК оказывается гораздо более эффективной (на несколько порядков), чем соответствующие однонитевые молекулы РНК (в частности, антисмысловая РНК). Исследователи Fire et al. в 1998 году назвали этот феномен интерференцией РНКi. Показано, что этот мощный механизм сайленсинга гена действует у нескольких видов большинства филогенетических типов.
РНКi начинается, когда фермент, называемый DICER, встречается с dsРНК и нарезает ее на части, называемые малыми интерферирующими РНК, или siРНК. Этот белок принадлежит семейству РНК-аза III нуклеаз. Комплекс белков собирает эти остатки РНК и использует их код в качестве проводника для поиска и уничтожения в клетке любых РНК с соответствующей последовательностью, таких как мРНК-мишень (для обзора см. Bosher & Labouesse, 2000).
Феномен РНКi (Akashi et al, 2001) в итоге можно суммировать следующим образом.
- Этап 1: процесс распознавания и сканирования dsРНК.
- Этап 2: расщепление dsРНК посредством РНК-азы III и продукция siРНК.
- Этап 3: связывание siРНК и связанных факторов в комплексах RISC.
- Этап 4: распознавание комплементарной мРНК-мишени.
- Этап 5: расщепление мРНК-мишени в центре области, комплементарной siРНК.
- Этап 6: распад мРНК-мишени и рециркуляции RISC-комплекса.
Вскоре в попытке использования феномена РНКi в качестве технологии генного нокдауна было выявлено, что в клетках млекопитающих разработаны различные защитные феномены против вирусных инфекций, которые могут препятствовать применению этого подхода. Действительно, присутствие вирусной dsРНК на чрезвычайно низком уровне запускает ответ интерферона, приводящий к общей неспецифической супрессии трансляции, которая в свою очередь запускает апоптоз (Williams, 1997, Gil & Esteban, 2000).
В 2000 году первая попытка работы с dsРНК привела к специфическому ингибированию в ооцитах и в ранних эмбрионах мышей 3 генов (MmGFP под контролем фактора элонгации 1a, E-кадгерин и c-mos). Не наблюдалось подавления трансляции, и таким образом, ответа РНК-зависимой протеинкиназы PKR, так как эмбрионы продолжали развитие (Wianny & Zernicka-Goetz, 2000). Через год в компании Ribopharma AG (Kulmbach, Германия) впервые были показаны функциональные возможности РНКi в клетках млекопитающих. Использование коротких (20-24 пар оснований) dsРНК, которые назвали SIRPLEX TМ, даже в клетках человека, специфически выключало гены без инициации стадии острого ответа. Сходные эксперименты, выполненные позже другими исследовательскими группами (Elbashir et al., 2001; Caplen et al., 2001), дополнительно подтвердили эти результаты.
Годом позже исследователи Paddison et al. (Paddison et al., 2002) для ингибирования функций специфических генов попробовали использовать уложенные в шпилечные структуры малые РНК. Стимулом для этой работы явились предыдущие исследования, показавшие, что у Caenorhabditis elegans некоторые гены посредством РНКi естественным образом регулируют другие гены путем кодирования РНК со шпилечной структурой. Тестированные в различных нормальных и опухолевых клеточных линиях человека и мыши короткие шпилечные РНК (shРНК) также эффективно, как их копии siРНК, способны к генному сайленсингу. Кроме того, in vivo shРНК обладают лучшей кинетикой реассоциации, чем дуплексные эквиваленты. Еще более важно, что указанные авторы создали трансгенные клеточные линии, сконструированные для синтеза shРНК, которые обладают длительным супрессивным эффектом во время деления клетки (Eurogentec). Недавно было показано, что другая группа малых РНК (также находящаяся в диапазое 21-25 нт) опосредует подавление экспрессии гена. Также РНК, известные как малые временные регулируемые РНК (stРНК), были описаны у Caenorhabditis elegans, где регулируют время экспрессии гена в ходе развития. Необходимо отметить, что stРНК и siРНК, несмотря на очевидные сходства, работают посредством различных механизмов действия (для обзора см. Banerjee&Slack, 2002). В отличие от siРНК, stРНК длиной в 22 нт подавляет экспрессию мРНК-мишени после инициации трансляции без нарушения целостности мРНК. В последних исследованиях указано, что две stРНК, впервые описанные у нематод, являются членами огромного семейства, с сотнями дополнительных микроРНК (miРНК), присутствующих у многоклеточных животных (Grosshans & Slack, 2002).
Ученые сначала использовали РНКi в нескольких системах, включающих в себя Caenorhabditis elegans, дрозофилу, трипаносомы и различные другие беспозвоночные. Более того, используя этот подход, недавно несколько групп исследователей представили специфическую супрессию биосинтеза белка в различных клеточных линиях млекопитающих, а именно в клетках HeLa, что указывает на то, что РНКi является широко применимым способом для сайленсинга гена in vitro. На основании этих результатов РНКi быстро стала широко признанным инструментом для подтверждения (идентификации и определения) функции гена. Интерференция РНК, использующая короткие олигонуклеотиды dsРНК, кроме того, позволит расшифровывать функцию генов, которые секвенированы только частично. Поэтому скоро РНКi станет обязательным в таких исследованиях, как:
- ингибирование экспрессии гена на посттранскрипционном уровне в эукариотических клетках. В этой связи РНКi является прямым инструментом быстрой оценки функции гена и выявления нулевых фенотипов;
- разработка технологии РНКi для применения у эмбрионов после имплантации;
- преимущественное экономическое значение интерференции РНК обусловлено ее применением в качестве терапии. В этой связи РНКi может приводить к созданию лекарственных препаратов на основе РНК для лечения заболеваний человека.
Глаукома
Глаукома является одной из ведущих причин слепоты. Приблизительно 15% случаев слепоты во всем мире является следствием глаукомы. Наиболее часто встречающимся типом глаукомы является первичная открытоугольная глаукома, уровень распространенности которой составляет 1/200 общего населения в возрасте старше 40 лет.
Глаукому определяют как процесс разрушения ткани глаза, вызванный постоянным повышением внутриглазного давления (ВГД) выше его нормальных физиологических границ.
Все более очевидно, что множество форм глаукомы имеют генетическую предрасположенность, и многочисленные современные исследования направлены на идентификацию хромосомных областей и генов, связанных с глаукомой. Вероятно, что этиология открытоугольной глаукомы (ОУГ) является мультифакторной, является результатом комбинации мутаций в более чем в одном гене и пока еще идентифицированных факторов внешней среды. В случае ювенильной ОУГ во взрослом возрасте было идентифицировано несколько локусов. Однако известен только один ген, а именно ген миоциллина/TIGR (трабекулярной сети индуцируемого глюкокортикоидного ответа) в локусе GLC1A на хромосоме 1q21-q31. В этнически различающихся популяциях во всем мире было идентифицировано более тридцати мутаций этого гена. Исследования показали, что этот ген отвечает примерно только за 5% всех случаев ОУГ (см. обзоры работ Wirtz&Samples, 2003, и Khaw et al., 2004a).
Патогенез
Большинство случаев глаукомы характеризуются повышенным ВГД, при этом степень повышения может различаться. В тех случаях, когда повышение давления является изначально невысоким (то есть открытоугольная глаукома, меланоцитарная глаукома), и в некоторых случаях вторичной глаукомы наблюдается медленное прогрессирование повреждения ретинальных ганглиозных клеток и зрительного нерва. При закрытоугольной глаукоме внезапное повышение ВГД часто приводит к слепоте, несомненно прежде всего в силу прекращения аксоплазматического потока на уровне lamina cribrosa.
При исследованиях людей принято считать, что в начале или в прогрессировании повреждения слепого пятна, происходящего при глаукоме, отчасти играет роль тканевая ишемия. Дегенерацией ретинальных ганглиозных клеток может быть некроз, но существует вероятность того, что возможно дегенерация является апоптозом, который вызван повышением ВГД, и соответствующие роли окиси азота и глутамата считаются существенными в ходе прогрессирования болезни (см. последние обзоры по этому предмету в работах Osborne et al, 2003).
Лечение
Притом что в комплекс факторов патогенеза глаукомы вовлечены несколько этиологических причин, абсолютным определением для выбора лечения является количество первичных и/или индуцированных нарушений давления, локализующихся в углу передней камеры.
Современные способы лечения включают в себя лекарственные препараты или хирургические вмешательства, направленные на снижение ВГД, хотя неизвестны патофизиологические механизмы глаукомы, по которым повышенное ВГД приводит к повреждениям нейронов.
Медикаментозное подавление повышенного ВГД может быть достигнуто, используя четыре типа лекарственных средств: средства, подавляющие образование водянистой влаги (среди которых ингибиторы карбоангидразы, бета-адреноблокирующие средства или агонисты альфа2-адренорецепторов), миотики (то есть парасимпатомиметики, представляющие собой холинэргические или антихолинэстеразные ингибиторы); средства, усиливающие увеосклеральный отток; и гиперосмотические средства (которые создают градиент осмотического давления через барьер крови/водянистой влаги внутри ресничного эпителия). При лечении глаукомы применяют все четыре типа средств, первые три типа обычно используют в качестве неотложного лечения и для продолжительного контроля, тогда как гиперосмотические средства неоценимы в качестве неотложного и предоперационного лечения. Появляющиеся в последнее время нейропротективные средства, которые представляют пятый тип лекарственных средств, являются важным возможным дополнением медикаментозной терапии. Действительно, наблюдение, что при глаукоме возрастают уровни NOS и глутамата и что они вовлечены в некроз или апоптоз ретинальных ганглиозных клеток, повышает возможность применения нейропротективных средств и даже нейрорегенерации. Таким образом, ингибиторы NOS, возбуждающие аминокислотные антагонисты, антагонисты глутаматных рецепторов, ингибиторы апоптоза и блокаторы кальциевых каналов, все они являются возможными лекарственными средствами в будущих разработках средств против глаукомы. Блокаторы кальциевых каналов могут уменьшать влияние повреждения капиллярного кровообращения в головке зрительного нерва, наряду с потенциальным повышением возможности оттока на уровне трабекулярных клеток.
Обзоры различных глазных болезней и способы их лечения приведены в ссылках, конкретно в работах Bunce (2005), Costagliola (1995, 2000), Cullinane (2002), Sakaguchi (2002), Shah (2000) и Wang (2005).
На сегодняшний день способы лечения не должны иметь известных и практических трудностей, связанных с оценкой способности оттока, точного мониторинга терапии и сложности хирургических технологий, объединяемых для запутывания прогноза. Важнейшим фактором всех глауком является дегенерация ретинальных ганглиозных клеток, таким образом, нейропротективое действие посредством эффективного снижения глазного давления является существенным требованием для любого применяемого средства (см. последние обзоры по предмету в работе Khaw et al., 2004b).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении описан способ лечения глазных болезней, характеризующихся изменением ВГД, у животных, включая человека. В частности, болезни глаз могут включать в себя глаукому, увеит и воспаление. Способ основан на подавлении экспрессии генов, вовлеченных в образование водянистой влаги или оттока водянистой влаги в глазу. Подавление можно осуществлять при помощи функциональных групп двунитевых нуклеиновых кислот, называемых siНК или малыми интерферирующими НК, которые направлены на интерференцию экспрессии различных возможных генов в мРНК. Предпочтительно siНК представляют собой siРНК, вместе с тем модифицированные нуклеиновые кислоты или сходные химически синтезированные объекты также включены в объем настоящего изобретения.
Предпочтительные варианты осуществления настоящего изобретения относятся к местному применению siНК. Варианты осуществления настоящего изобретения также относятся к фармацевтическим композициям для использования при лечении глазных болезней. Настоящее изобретение можно использовать для препаратов местного лечения глаз, в генах-мишенях, вовлеченных в патогенез глаукомы, а также для применения химически синтезированных объектов для лечения заболеваний у животных (включающих в себя людей).
Кроме лечения глаукомы, способ по настоящему изобретению также может использоваться для лечения других заболеваний передней камеры глаза. В частности, способ можно применять при лечении заболеваний, характеризующихся нарушением образования или оттока водянистой влаги в глазу. Примеры возможных для лечения состояний включают в себя местные состояния, такие как инфекции или воспаления, и общие состояния, такие как увеит или проявление системных заболеваний. Дополнительно, некоторые варианты осуществления настоящего изобретения относятся к лечению диабетической ретинопатии.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Гены-мишени
В соответствии с настоящим изобретением авторы определили список генов-мишеней, уровни экспрессии которых могут влиять на ВГД. Эти гены могут входить в состав групп генов, задействованных в образовании водянистой влаги, или в группу генов, задействованных в оттоке водянистой влаги. Ниже представлен список таких генов-мишеней.
- Карбоангидразы II, IV и XII
- Адренергические рецепторы: бета1 и 2, и альфа 1А, 1B и 1D
- Ацетилхолинэстераза
- Циклооксигеназы 1 и 2
- АТФазы: альфа1, альфа2, альфа3, бета1, бета2
- Молекула адгезии лейкоцитов эндотелия (ELAM-1)
- Система ангиотензина: ангиотензин I, ангиотензин II превращающие ферменты (ACE I и ACE II), рецепторы ангиотензина II (ATR1 и ATR2) и ренин
- Кохлин
Конструирование siНК
Несмотря на то что механизмы для РНКi остаются неизвестными, этапы, необходимые для создания конкретных олигонуклеотидов dsРНК, очевидны. Показано, что для получения РНК-интерференции наиболее эффективно работают сдвоенные нити dsРНК длиной 21-26 нуклеотидов. Также важен выбор правильной гомологичной области в пределах гена. При рассмотрении создания dsРНК для РНКi также важны такие факторы, как расстояние от стартового кодона, содержание G/C и местоположение аденозиновых димеров. Однако одним из последствий этого является возможная необходимость тестирования нескольких различных последовательностей на наиболее эффективную РНКi, и это может стать дорогостоящим тестом.
В 1999 году авторы Tuschl et al. расшифровывали эффект сайленсинга siРНК, показывающий, что его эффективность зависит от длины дуплекса, длины липкого 3-конца и от последовательности в этих липких концах. На основании этих базовых работ Eurogentec рекомендует выбирать области мРНК-мишени и, следовательно, последовательность дуплекса siРНК, руководствуясь следующими принципами.
Поскольку РНКi основана на образовании сложных белковых взаимодействий, очевидно, что мРНК-мишень должна быть свободна от каких-либо чужеродных факторов связывания. В этой связи необходимо избегать 5′ и 3′ нетранслируемых областей (UTR) и областей, близких к стартовому кодону, поскольку они могут иметь больше сайтов связывания регуляторных белков. Следовательно, последовательность siРНК выбирают следующим образом.
- В мРНК последовательности выбирают область, расположенную на расстоянии от 50 до 100 нт в направлении 5′-3′ от стартового кодона AUG, или перед терминирующим кодоном.
- В этой области проводят поиск следующих последовательностей: AA(N19), CA(N19).
- Рассчитывают процент G/C для каждой идентифицированной последовательности. В идеале содержание G/C составляет 50%, но оно должно быть менее 70% и более 30%.
- Предпочтительно избегают последовательностей, содержащих следующие повторения: AAA, CCC, GGG, TTT, AAAA, CCCC, GGGG, TTTT.
- Также проводят прогнозирование доступности на основании вторичной структуры мРНК.
- Также осуществляют анализ BLAST (то есть поиск по базе данных NCBI EST) с нуклеотидной последовательностью, наилучшим образом отвечающей предыдущим критериям для гарантии того, что будет инактивирован только один ген.
Для максимальной интерпретации результата при использовании siРНК необходимо принять следующие меры предосторожности.
- Всегда проводить тестирование смысловых и антисмысловых одиночных нитей в отдельных экспериментах.
- Испытывать рандомизированный дуплекс siРНК. Он должен иметь такую же нуклеотидную композицию, как siРНК, но не иметь существенной гомологии последовательностей с любым другим геном (включая интересующий).
- Если возможно осуществлять нокдаун того же гена с двумя независимым дуплексами siРНК для контроля специфичности процесса сайленсинга.
На практике каждый из выбранных генов вводят в виде нуклеотидной последовательности в программе прогнозирования, которая рассматривает все вышеописанные варианты для конструирования оптимальных олигонуклеотидов. Эта программа просматривает любую нуклеотидную последовательность мРНК на наличие областей, восприимчивых в качестве мишеней для siРНК. Конечным результатом этого анализа является подсчет возможных показателей олигонуклеотидов siРНК. Самые высокие показатели используют для конструирования двунитевых олигонуклеотидов РНК (обычно длиной 21 п.о., хотя также возможны другие значения длины), которые обычно производят химическим синтезом.
Кроме siРНК также можно использовать модифицированные нуклеотиды. Авторы планируют тестирование нескольких химических модификаций, которые широко известны в данной области техники. Эти модификации направлены на повышение стабильности или доступности siНК. Примеры подходящих модификаций описаны в публикациях, упомянутых ниже, каждая из которых включена в настоящее изобретение в качестве ссылки.
Исследования показывают, что замена 3′-концевых нуклеотидных липких сегментов 21-мерного дуплекса siРНК, имеющего два нуклеотидных 3′-липких конца, на дезоксирибонуклеотиды не оказывает отрицательного эффекта на активность РНКi. Сообщалось, что замена до четырех нуклеотидов на каждом конце siРНК на дезоксирибонуклеотиды допустима, тогда как полная замена на дезоксирибонуклеотиды приводила к отсутствию активности РНКi (Elbashir, 2001). Дополнительно, исследователи Elbashir et al. также сообщают, что замена siРНК на 2′-O-метилнуклеотиды приводит к полной потери активности РНКi.
Можно использовать нуклеозиды с модифицированной аффинностью, как описано в патенте WO2005/044976. В этой публикации описаны олигонуклеотиды, содержащие модифицированные нуклеозиды, которые обладают повышенной или пониженной аффинностью к комплементарному им нуклеотиду в мРНК-мишени и/или в комплементарной нити siНК.
В патенте GB2406568 описаны альтернативные модифицированные олигонуклеотиды, химически модифицированные для получения повышенной устойчивости к расщеплению или для обеспечения повышенного поглощения. Примеры таких модификаций включают в себя фосфоротиоатные межнуклеотидные связи, 2′-O-метил-рибонуклеотиды, 2′-дезокси-фтор-рибонуклеотиды, 2′-дезоксирибонуклеотиды, "универсальные базовые" нуклеотиды, 5-C-метилнуклеотиды и включение инвертированного дезоксиабазического остатка.
В патенте WO2004/029212 описаны модифицированные олигонуклеотиды, обладающие повышенной устойчивостью siРНК или повышенной эффективностью направленного транспорта. Модификации включают в себя образование химических перекрестных связей между двумя комплементарными нитями siРНК и химическую модификацию 3′-конца нити siРНК. Предпочтительные модификации представляют собой внутренние модификации, например модификации сахаров, модификации нуклеиновых оснований и/или модификации остова. Описаны 2′-фтор-модифицированные рибонуклеотиды и 2′-дезоксирибонуклеотиды.
В патенте WO2005/040537 дополнительно рассмотрены модифицированные олигонуклеотиды, которые можно использовать в настоящем изобретении.
Наряду с использованием dsНК и модифицированной dsНК, в настоящем изобретении можно использовать короткую шпилечную НК (shНК); две нити молекулы siНК могут быть связаны сшивающей областью, которая может являться нуклеотидным линкером или ненуклеотидным линкером.
Кроме siНК, которая является абсолютно комплементарной области-мишени, можно использовать вырожденные последовательности siНК для направленного транспорта к гомологичным областям. В патенте WO2005/045037 описан дизайн молекул siNA для направленного транспорта к таким гомологичным последовательностям, например, путем включения в них неканонических пар оснований, например несоответствующих и/или неоднозначных пар оснований, которые могут обеспечивать дополнительные последовательности-мишени. В тех случаях, когда несоответствия идентифицированы, можно использовать неканонические пары оснований (например, несоответствующие и/или неоднозначные основания) для создания молекул siНК, которые направлены более чем на одну генную последовательность. В примере, не ограничивающем объем настоящего изобретения, используют неканонические пары оснований, такие как пары оснований UU и CC, для создания молекул siNA, которые способны направлять последовательности на различные мишени с долей гомологии последовательностей. Собственно, одно преимущество использования siНК настоящего изобретения состоит в том, что можно сконструировать единственную siНК для включения в нее последовательности нуклеиновой кислоты, которая комплементарна нуклеотидной последовательности, являющейся консервативной между гомологичными генами. В этом подходе можно использовать единственную siNA для ингибирования экспрессии более чем одного гена, вместо использования более чем одной молекулы siNA для направления к различным генам-мишеням.
Предпочтительные молекулы siNA по настоящему изобретению являются двунитевыми. Молекула siNA по настоящему изобретению может содержать тупые концы, то есть концы, которые не включают в себя какие-либо выступающие нуклеотиды. В одном из вариантов осуществления молекула siNA по настоящему изобретению может содержать один или несколько тупых концов. В предпочтительных вариантах осуществления молекулы siNA имеют 3′-липкий конец. Молекулы siNA по настоящему изобретению могут содержать дуплексные молекулы нуклеиновых кислот из n нуклеотидов (5≥n≥1) с 3′-липкими концами. Исследователем Elbashir (2001) показано, что дуплексы siРНК с 21 нуклеотидами наиболее активны при содержании 3′-концевых динуклеотидных липких концов. Для облегчения перехода от клинических исследований у животных к испытаниям на людях возможные олигонуклеотиды подвергают дополнительному фильтрованию на консервативные межвидовые последовательности. В предпочтительных вариантах осуществления настоящего изобретения используют консервативные олигонуклеотиды, что позволяет использовать единственную олигонуклеотидную последовательность и в животных моделях, и в клинических испытаниях у людей.
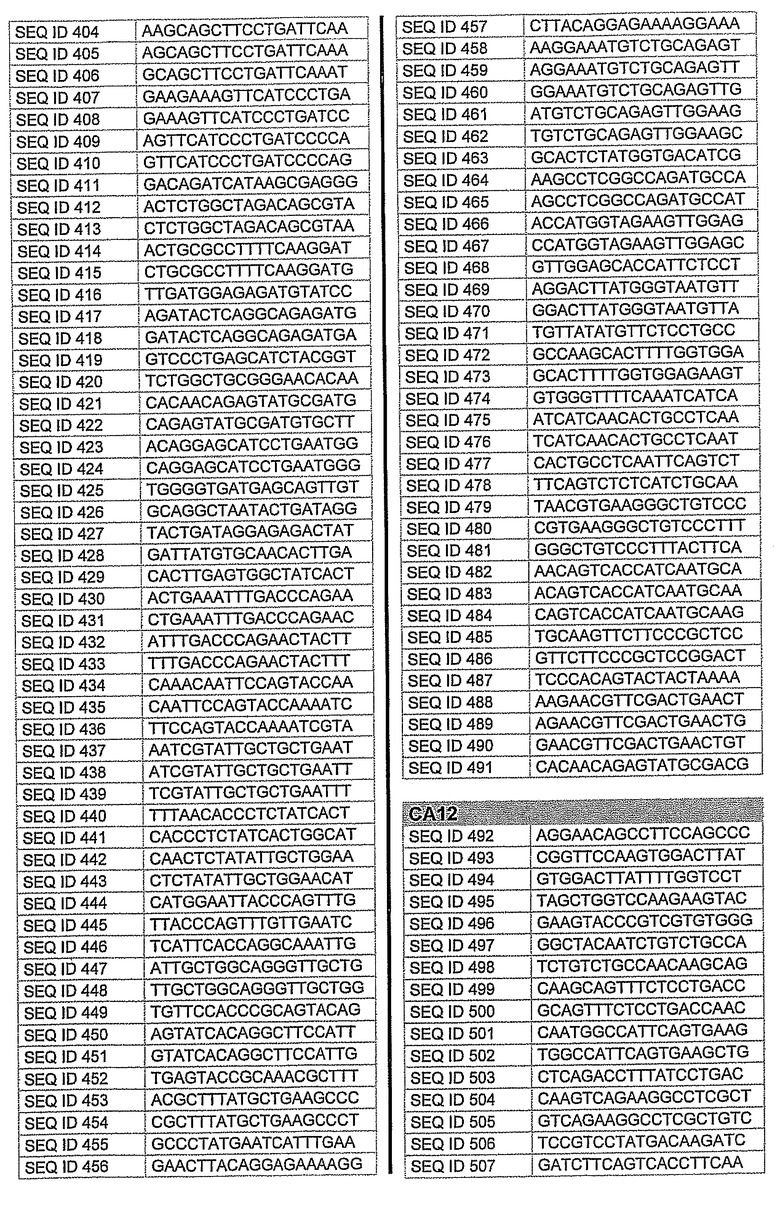
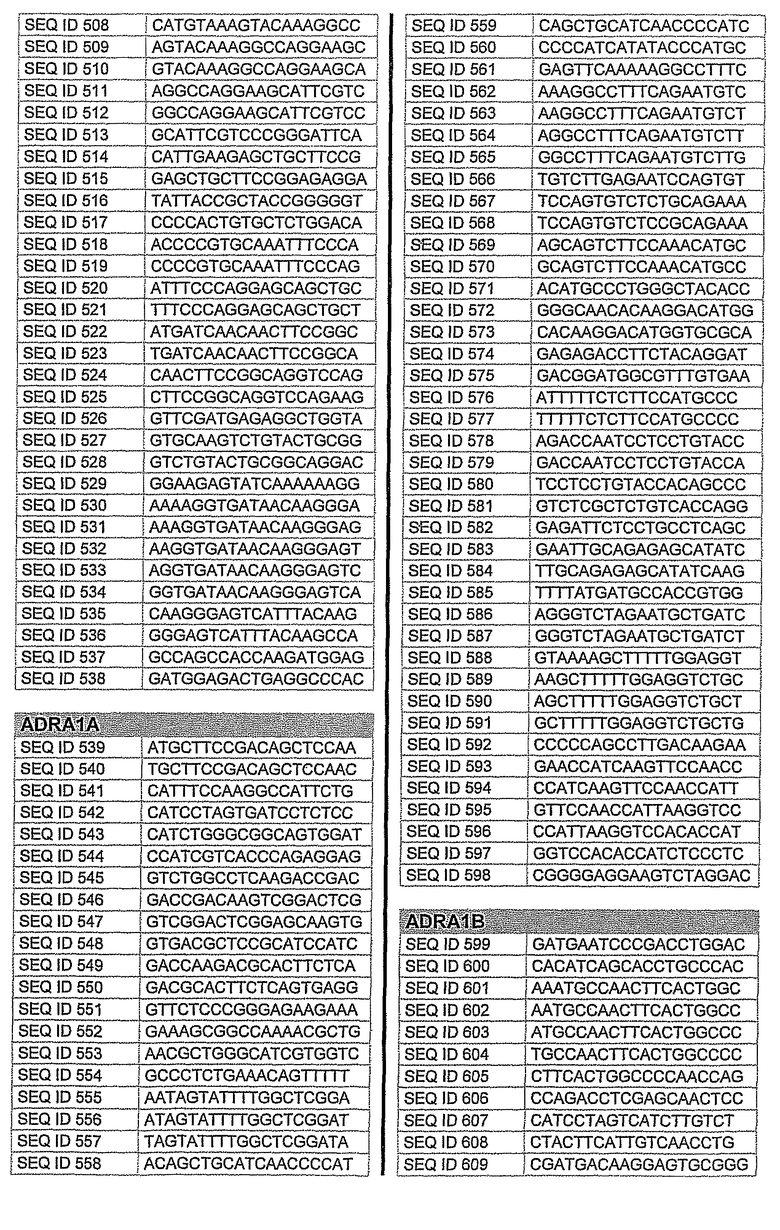
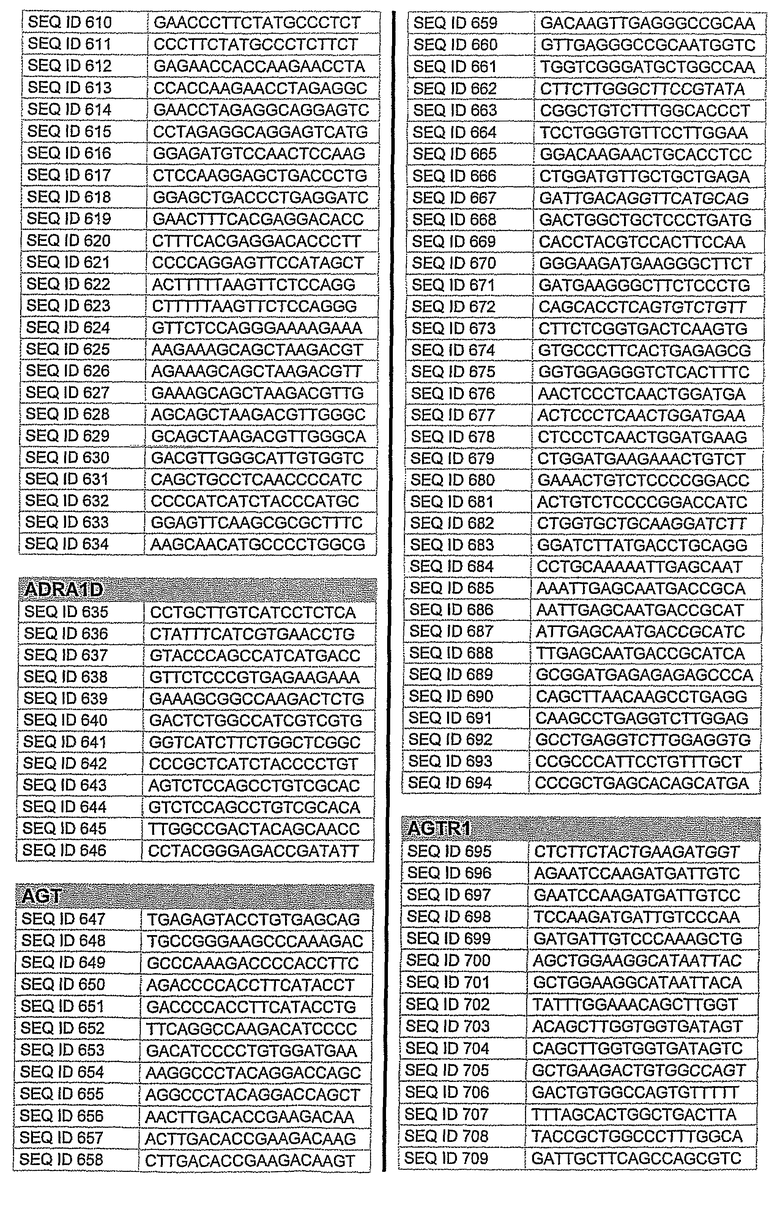
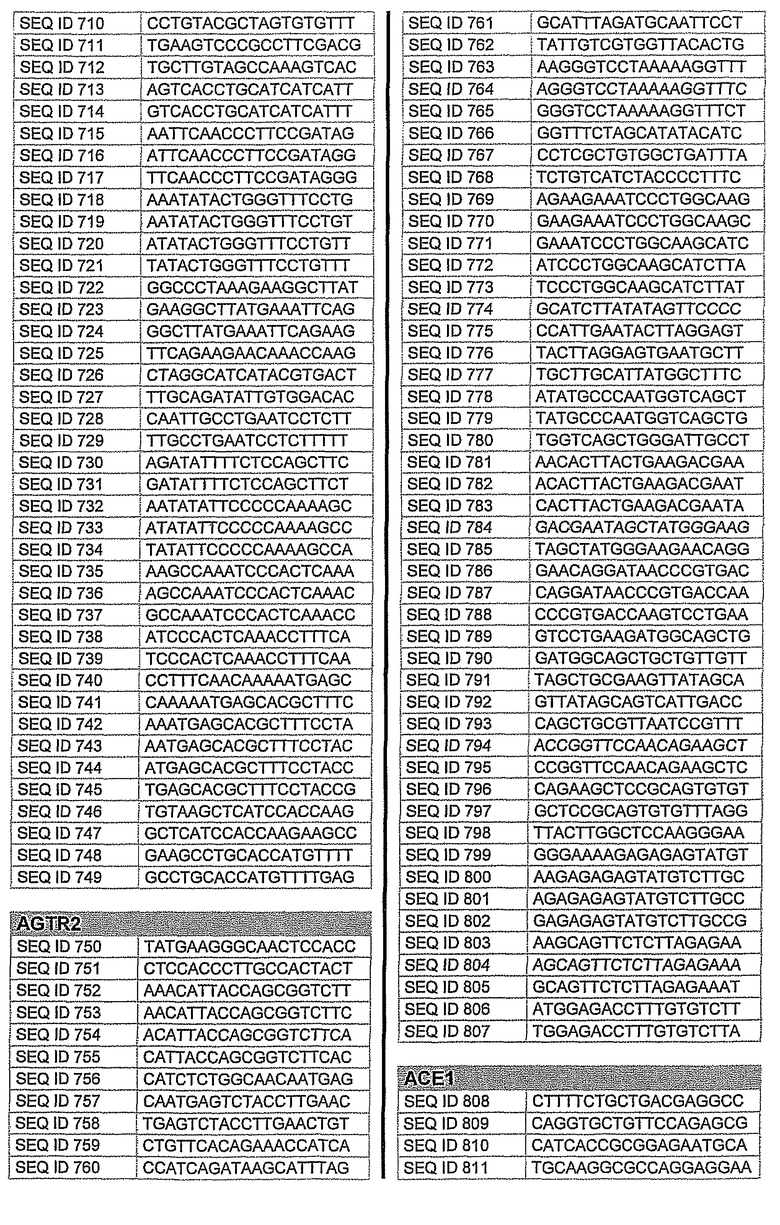
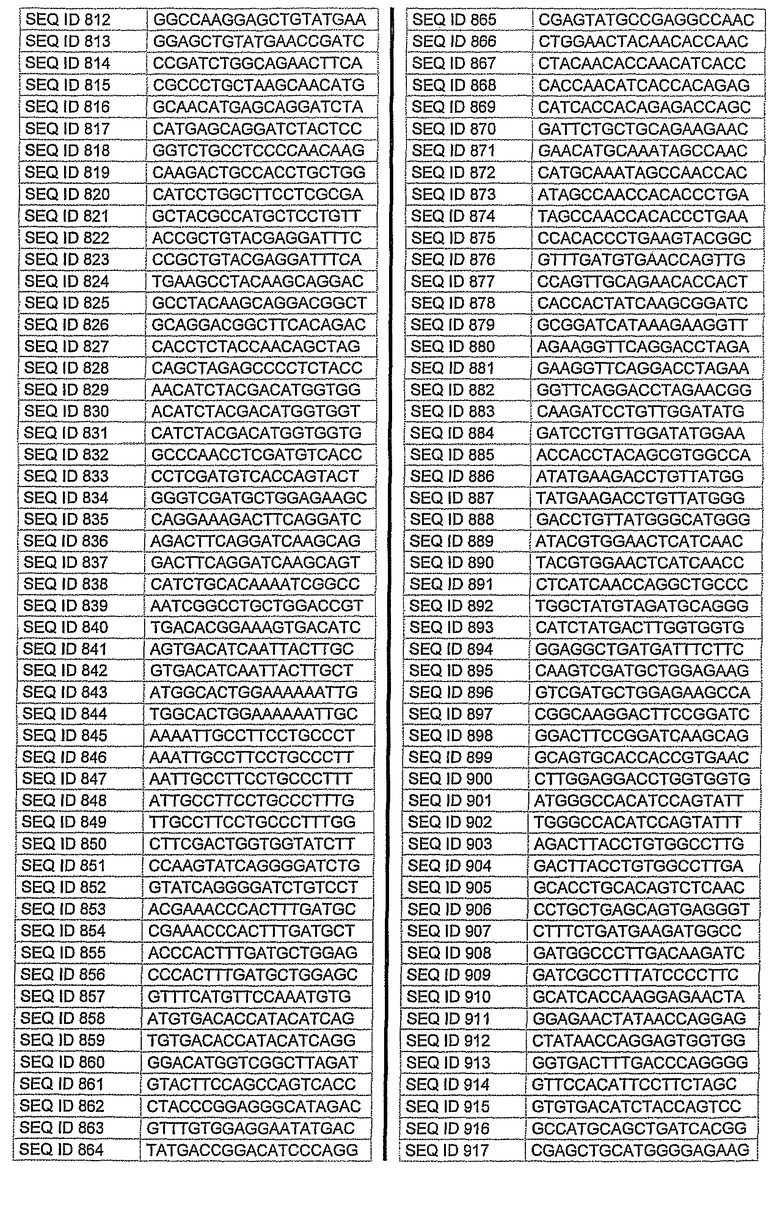
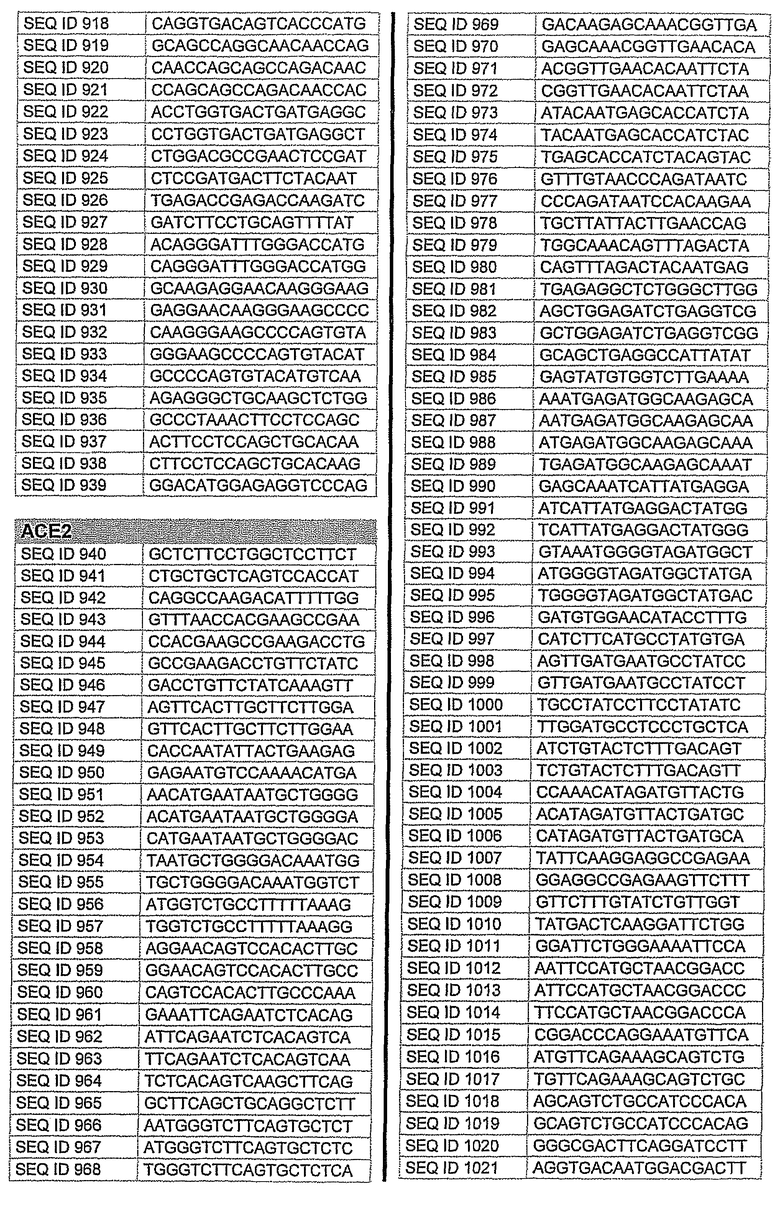
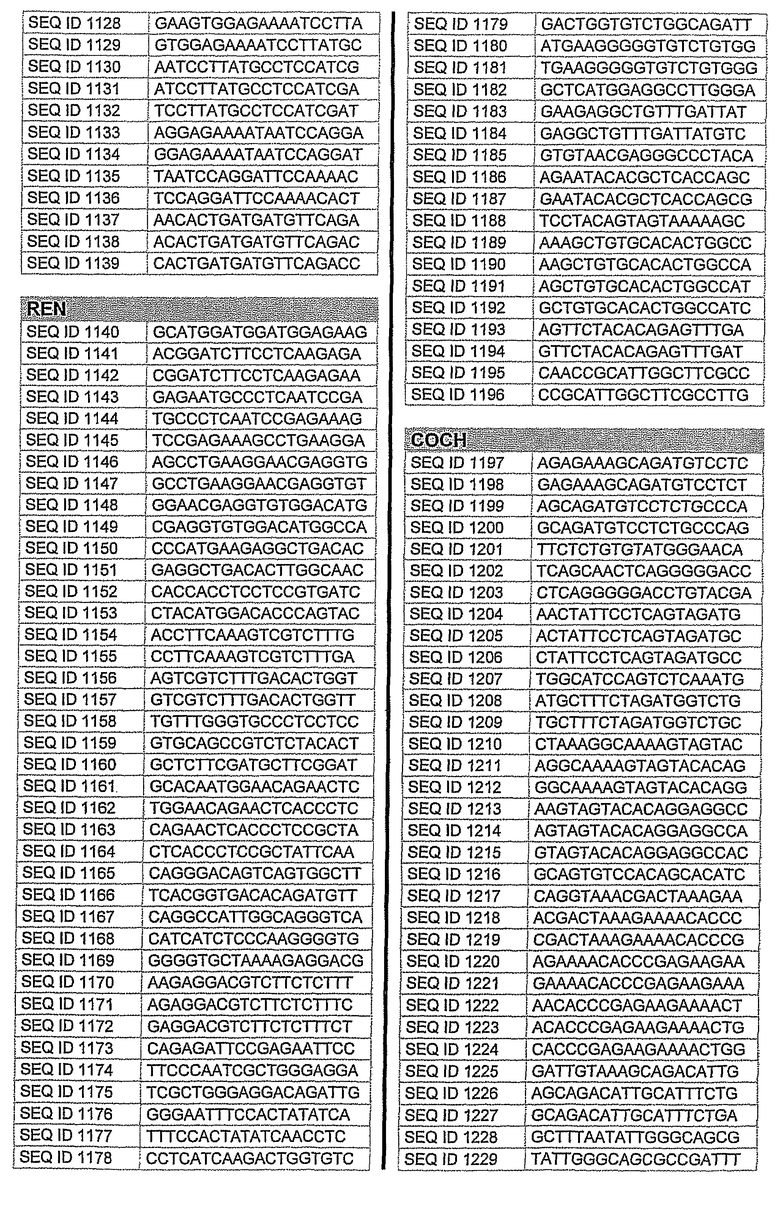
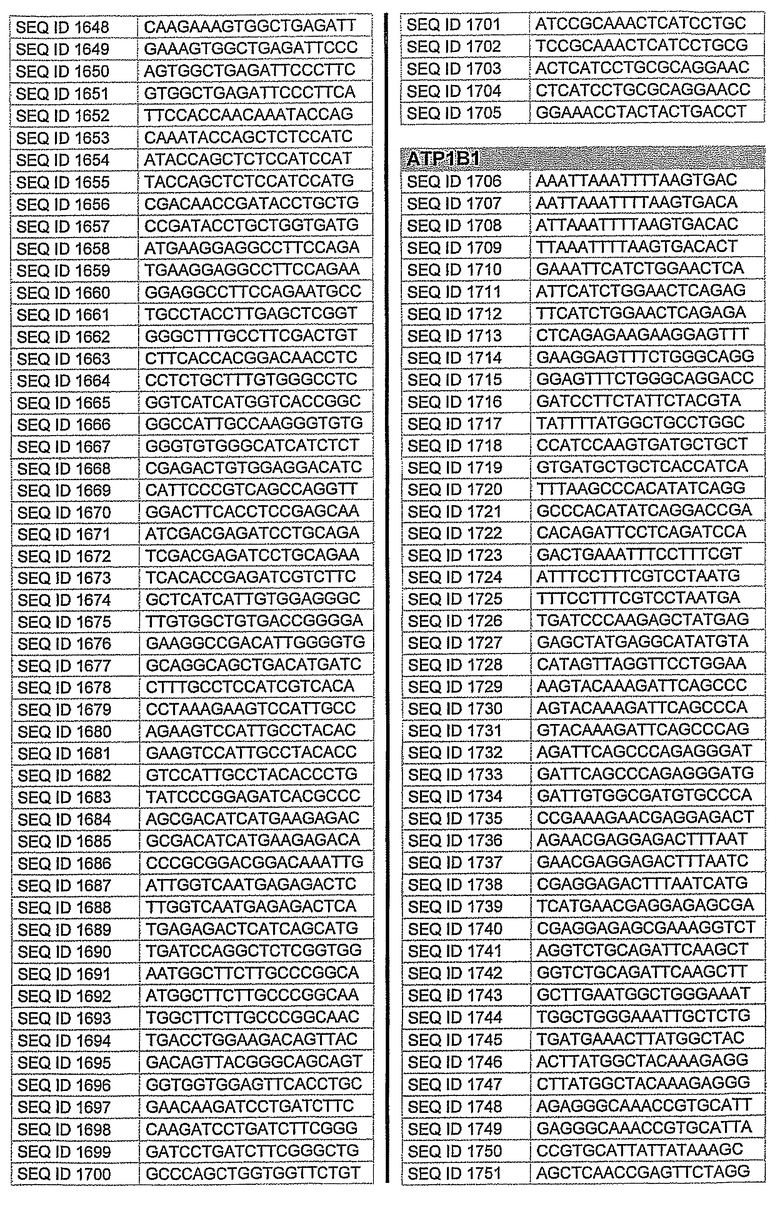
На фиг.1 показаны инвентарные номера GenBank, соответствующие выбранным генам-мишеням человека. В некоторых из этих генов альтернативный сплайсинг производит семейство транскриптов, которые отличаются составом экзонов. Настоящее изобретение позволяет индивидуально направлять каждую из форм транскриптов.
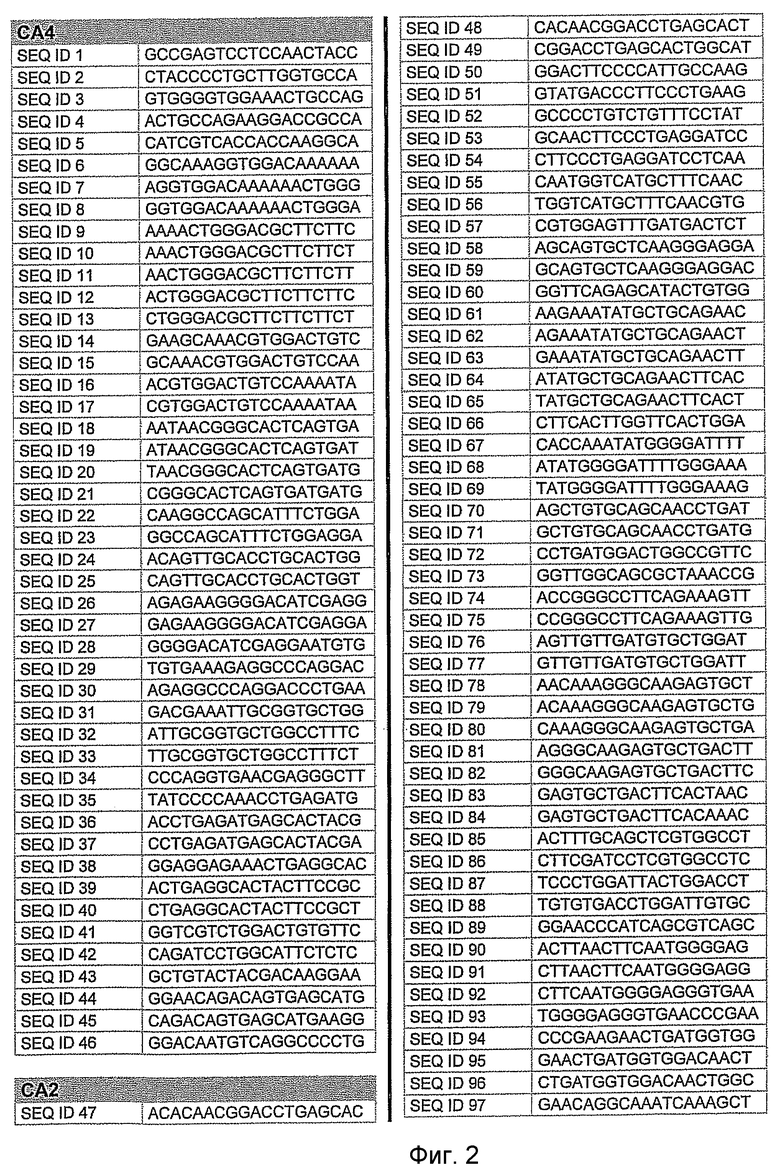
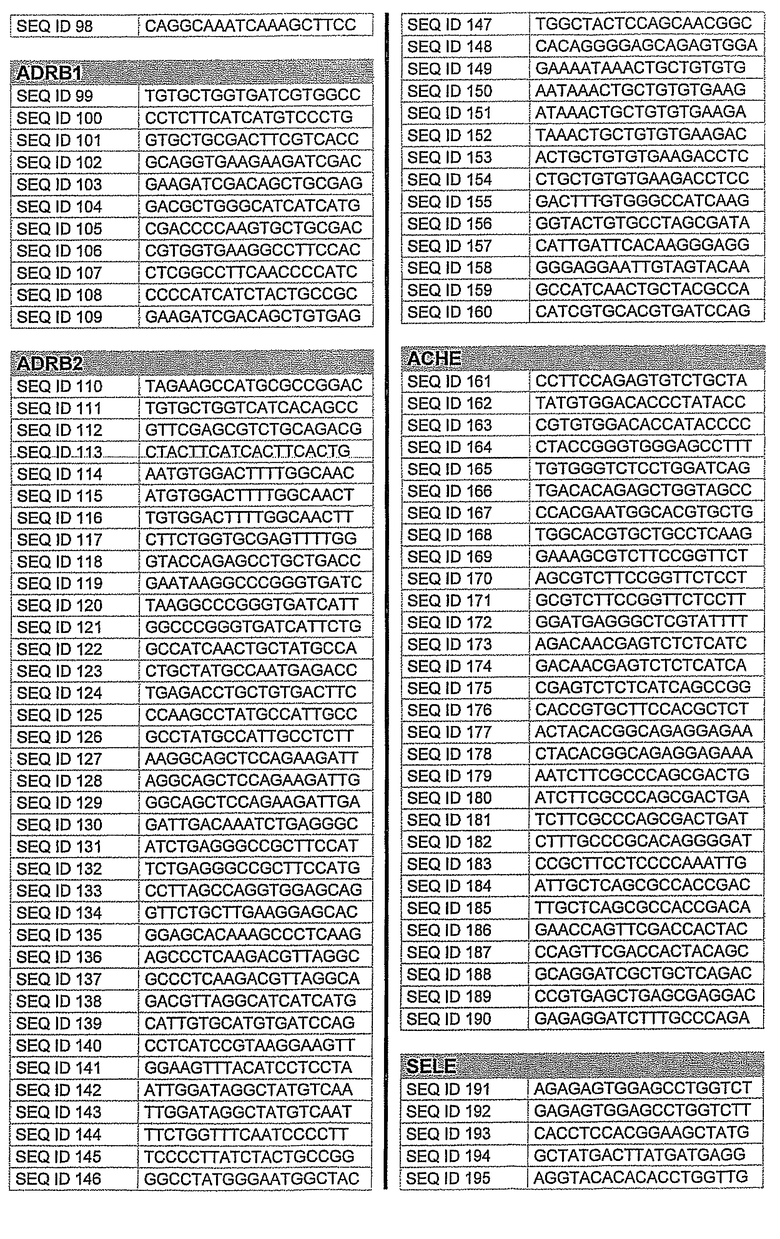
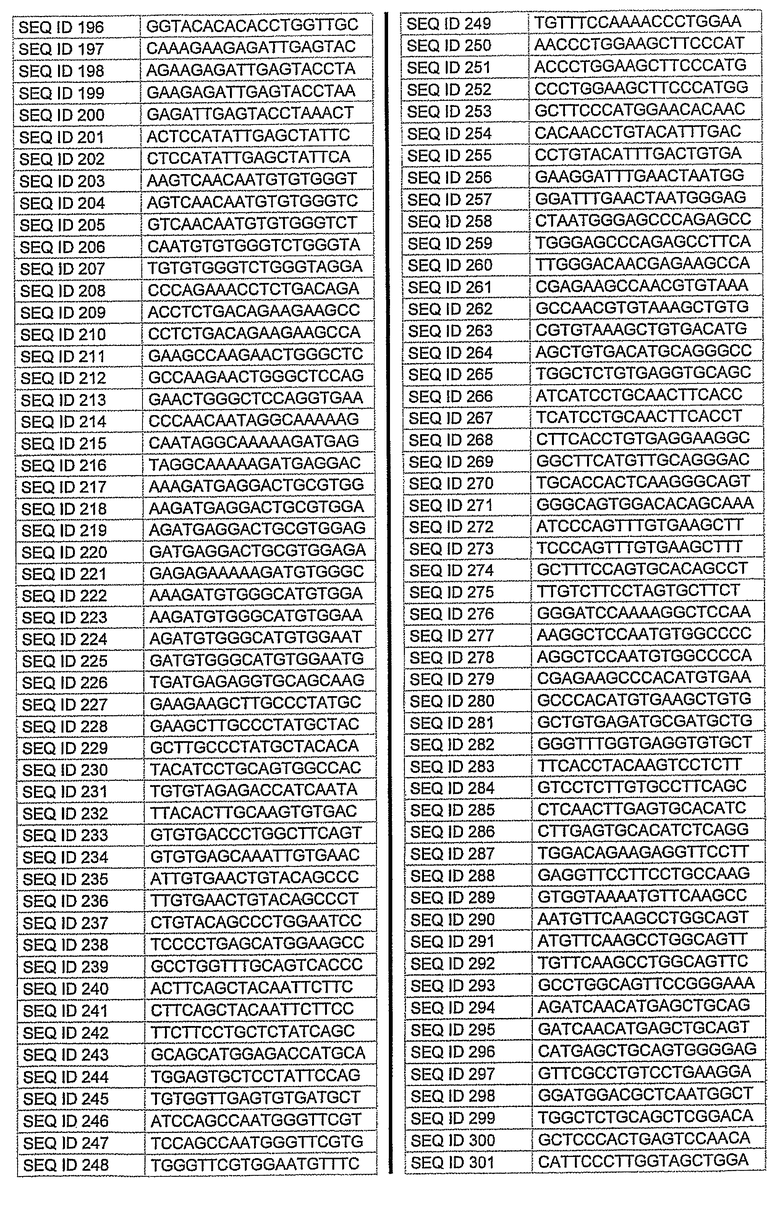
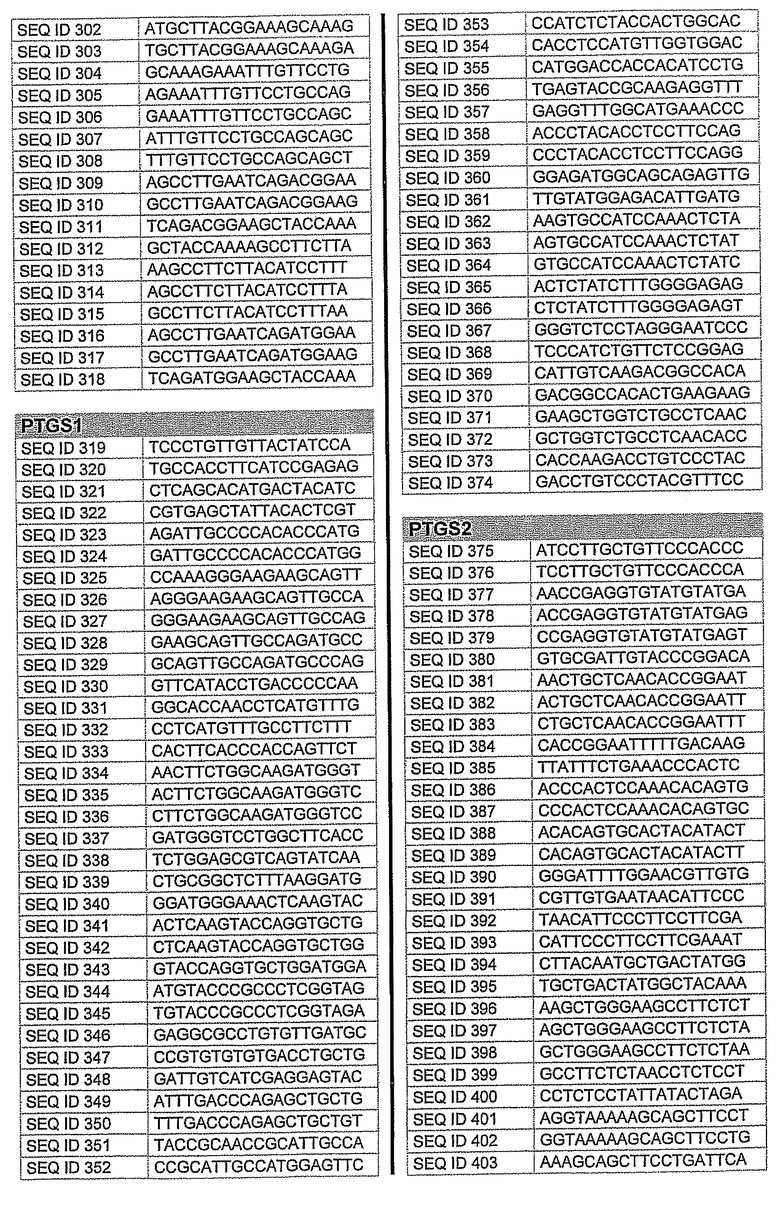
На фиг.2 показаны выбранные олигонуклеотидные последовательности, против которых направляют РНКi. Показанные последовательности представляют собой последовательности ДНК, являющиеся мишенями направляемой siНК. Поэтому в настоящем изобретении будут использованы дуплексы НК с последовательностями, комплементарными обозначенным последовательностям ДНК.
Последовательности, показанные на фиг.2, не ограничивают объем настоящего изобретения. На самом деле, не нужно, чтобы ДНК-мишени обязательно предшествовали AA или СА. Дополнительно, ДНК-мишень может состоять из последовательностей, представленных на фиг.2, фланкированных любой смежной последовательностью.
Исследования in vitro и на животных
Получение дуплексов siРНК
Предпочтительно РНК синтезируют химически, используя соответствующим образом защищенные рибонуклеозидные фосфорамидиты и общепринятый синтезатор ДНК/РНК. Замена одной или обеих нитей дуплекса siРНК на 2′-дезокси или 2′-O-метил-олигорибонуклеотиды ликвидирует сайленсинг в экстракте мухи (Elbashir et al., 2001). Вместе с тем, по-видимому, в клетках млекопитающих возможна замена смысловой siРНК на 2′-O-метил-олигорибонуклеотид (Ge et al., 2003).
Наиболее удобно siРНК приобретают от поставщиков промышленного олигосинтеза РНК, которые продают продукты синтеза РНК различного качества и стоимости. В целом получение РНК размером 21 нт не имеет больших трудностей для синтеза, и их легко получить с подходящим для РНКi качеством.
Поставщиками реактивов для синтеза РНК являются компании Prоligo (Hamburg, Германия), Dharmacon Research Lafayette, (CO, США), Glen Research (Sterling, VA, США), ChemGenes (Ashland, MA, США) и Cruachem (Glasgow, Великобритания), Qiagen (Германия), Ambion (США) и Invitrogen (Шотландия). Предыдущие компании, специализирующиеся на синтезе РНК, обладали правом лицензирования siРНК в целях признания направленного транспорта. В частности, поставщиками siРНК являются компании Ambion, Dharmacon и Invitrogen, которые предлагают традиционное специализированное обслуживание химического синтеза для siРНК и обеспечивают получение siРНК очисткой высокоэффективной жидкостной хроматографией ВЭЖХ и поставляют в сухой форме вместе с водой без РНК-азы. На веб-сайтах вышеупомянутых поставщиков можно найти основной Интернет-ресурс для методологий РНКi и siРНК, наряду с выходом на дополнительные продукты и услуги siРНК.
При работе с однонитевыми молекулами РНК необходим этап ренатурации. Крайне важно, что все этапы обработки проводят в стерильных условиях при отсутствии РНК-азы. Для ренатурации РНК вначале необходимо количественное определение олиго- с помощью УФ-поглощения при 260 нанометрах (нм). В этом случае для ренатурирования используют следующий протокол, основанный на исследованиях Elbashir et al. (2001).
- Раздельно берут аликвоты олиго-РНК и растворяют каждую до концентрации 50 мкМ.
- Объединяют 30 мкл раствора каждой олиго-РНК и 15 мкл 5× ренатурирующего буфера. Конечная буферная концентрация составляет: 100 мМ ацетата калия, 30 мМ HEPES-KOH с уровнем pH 7,4, 1 мМ ацетата магния. Конечный объем составляет 75 мкл.
- Раствор инкубируют в течение 1 минуты при 90°C, центрифугируют пробирку в течение 15 секунд, оставляют на 1 час при 37°C и затем используют при температуре окружающей среды. Раствор можно хранить замороженным при -20°C и замораживать-оттаивать до 5 раз. Конечная концентрация дуплекса siРНК обычно составляет 20 мкМ.
В качестве альтернативы у поставщиков можно приобретать уже ренатурированные dsРНК. Также можно использовать химически модифицированные нуклеиновые кислоты. Например, краткий обзор возможных для использования типов модификации приводится в патенте WO03/070744, содержание которого включено в настоящее изобретение в качестве ссылки. Отдельное внимание следует уделить страницам 11-21 указанной публикации. Выше описаны другие возможные модификации. Специалисту в данной области техники известны другие типы химической модификации, которые можно включать в молекулы РНК.
Система in vitro
Для проверки специфичности интерференции siРНК использовали различные клеточные культуры, которые экспрессируют гены-мишени. Для этих экспериментов использовали следующие клетки: непигментированные клетки цилиарного эпителия NPE кролика, человеческие клетки цилиарного эпителия OMDC и человеческие клетки эмбриональной почки HEK293. Клетки инкубировали с соответствующими дуплексами siРНК и проводили анализ подавления экспрессии гена-мишени. Для связывания siРНК с нокдауном со специфическими фенотипами в культивируемых клетках необходимо проявление снижения белка-мишени или по меньшей мере проявление снижения мРНК-мишени.
Количественный анализ уровней мРНК гена-мишени можно осуществить количественной полимеразной цепной реакцией в реальном времени (RT/PCR). Дополнительно, можно определить уровни белка различными путями, хорошо известными в данной области техники, такими как анализом Вестерн-блоттинг со специфическими антителами к различным мишеням, что позволяет проводить прямой контроль снижения белка-мишени.
Трансфекция дуплексов siРНК
Ниже приведены несколько примеров способов, известных в данной области техники. Можно осуществлять единственную трансфекцию дуплекса siРНК, используя катионный липид, такой как реагент трансфекции RNAiFect Transfection Reagent (Qiagen) и реагент Lipofectamine 2000 (Invitrogen), и проводить анализ сайленсинга через 24, 48 и 72 часа после трансфекции.
Обычный протокол трансфекции можно осуществлять следующим образом. В одной лунке 6-луночного планшета проводили трансфекцию с использованием siРНК в конечной концентрации 100 нМ. Следуя протоколу РНКiFect, за один день перед трансфекцией засевали от 2 до 4×105 клеток на лунку в 3 мл подходящей среды для выращивания, содержащей DMEM, 10% сыворотки, антибиотики и глутамин, и инкубировали клетки при нормальных условиях роста (37°C и 5% CO2). В день трансфекции конфлуэнтность клеток должна составлять от 30 до 50%. Растворяли 15 мкл 20 мкМ дуплекса siРНК (соответствующих конечной концентрации в 100 нМ) в 85 мкл буферного раствора ЕС-R, получая конечный объем в 100 мкл, и перемешивали встряхиванием. Для образования комплекса добавляли 19 мкл реагента трансфекции РНКiFect к растворенной siРНК и перемешивали с помощью пипетки или встряхиванием. После инкубирования образцов в течение 10-15 минут при комнатной температуре для возможности образования комплексов трансфекции, по капле добавляли комплексы на клетки с 2,9 мл свежей среды для выращивания с низким содержанием антибиотиков. После вращения планшета для обеспечения однородного распределения комплексов трансфекции клетки инкубировали при нормальных условиях их роста. Через день удаляли комплексы и добавляли свежую и полную среду для выращивания. Для контроля сайленсинга гена клетки собирали через 24, 48 и 72 часа после трансфекции. Протокол реагента липофектамина 2000 является весьма сходным. За один день перед трансфекцией засевали от 2 до 4×105 клеток на лунку в 3 мл подходящей среды для выращивания, содержащей DMEM, 10% сыворотки, антибиотики и глутамин, и инкубировали клетки при нормальных условиях роста (37°C и 5% CO2). В день трансфекции конфлуэнтность клеток должна составлять от 30 до 50%. Растворяли 12,5 мкл 20 мкМ дуплекса siРНК (соответствующих конечной концентрации в 100 нМ) в 250 мкл DMEM для получения конечного объема 262,5 мкл, и перемешивали. Также 6 мкл липофектамина 2000 растворяли в 250 мкл DMEM и смешивали. Для образования комплексов через 5 минут инкубирования при комнатной температуре растворенный олигомер и растворенный липофектамин соединяли в течение 20-минутного инкубирования при комнатной температуре. После этого по капле добавляли комплексы на клетки с 2 мл свежей среды для выращивания с низким содержанием антибиотиков и аккуратно смешивали, покачивая планшет назад и вперед, что гарантировало однородное распределение комплексов трансфекции. Клетки инкубировали при нормальных условиях их роста и через день удаляли комплексы и добавляли свежую и полную среду для выращивания. Для контроля сайленсинга генов клетки собирали через 24, 48 и 72 часа после трансфекции.
Эффективность трансфекции может зависеть от типа клетки, но также и от количества пассирований и конфлуэнтности клеток. Также решающими являются время и способ формирования siРНК-липосомных комплексов (например, переворачивание по сравнению с встряхиванием). Низкая эффективность трансфекции является наиболее частой причиной неудачного сайленсинга. Успешная трансфекция представляет собой нетривиальную задачу и нуждается в тщательном исследовании на каждую новую используемую клеточную линию. Эффективность трансфекции можно тестировать трансформирующими репортерными генами, например cmv-стимулируемой EGFP-экспрессирующей плазмидой (например, от компании Clontech) или B-GaI-экспрессирующей плазмидой, и затем на следующий день оценивать посредством фазово-контрастной и/или флюоресцентной микроскопии.
Исследование дуплексов siРНК
В зависимости от относительного количества и времени жизни (или метаболизма) белка-мишени, фенотип с нокдауном может стать заметным спустя от 1 до 3 дней или даже позже. В случаях, когда не наблюдается какого-либо фенотипа, истощение белка можно выявлять иммунофлуоресценцией или Вестерн-блоттингом.
Экстрагированные из клеток общие фракции РНК после трансфекций предварительно обрабатывали ДНК-азой I и использовали для обратного считывания с применением рандомизированного праймера. ПЦР амплифицировали со специфической праймерной парой, охватывающей по меньшей мере одну точку соединения экзон-экзона для контроля амплификации пред-мРНК. В качестве контроля также необходимо проводить RT/PCR мРНК-немишени. Эффективное истощение мРНК еще неопределяемого снижения белка-мишени может указывать на то, что в клетке возможно существует большой запас устойчивого белка. Альтернативно, можно использовать амплификацию ПЦР в реальном времени для более точного тестирования уменьшения или исчезновения мРНК. Количественный анализ исходного количества матрицы наиболее специфически, точно и воспроизводимо осуществляют путем ПЦР в реальном времени с обратной транскриптазой (РВ). ПЦР в реальном времени мониторирует флюоресценцию, испускаемую во время реакции, как индикатор продукции ампликона в течение каждого цикла ПЦР. Этот сигнал возрастает прямо пропорционально общему продукту ПЦР в реакции. Регистрируя число флюоресцентных эмиссий в каждом цикле, можно контролировать ПЦР реакцию во время экспоненциальной фазы, когда первое значительное повышение количества продукта ПЦР коррелирует с исходным количеством матрицы-мишени.
Чтобы подтвердить свойство интерференции по-разному экспрессирующихся генов, идентифицированных в клеточных культурах, проводили количественный кRT/PCR согласно протоколу изготовителя. Для количественного RT/PCR (кRT/PCR) использовали около 250 нг общей РНК для обратной транскрипции, с последующей ПЦР-амплификацией со специфическими праймерами для каждого гена в реакционной смеси, содержащей Master SYBR Green I. Основные условия ПЦР содержали начальный этап в 30 минут при 91°C, с последующими 40 циклами в 5 с при 95°C, 10 с при 62°C и 15 с при 72°C. Использовали специфические праймерные последовательности, соответствующие каждому гену-мишени. В качестве контроля для стандартизации данных применяли количественный анализ мРНК b-актина. Сравнения относительной экспрессии гена работают лучше всего, когда, среди проб, экспрессия гена выбранного эндогенного/внутреннего контроля выражена сильно и остается постоянной, по отношению к общей РНК. Используя инвариантный эндогенный контроль в качестве активного эталона, можно стандартизировать количественный анализ мРНК-мишени на различия в количестве общей РНК, добавляемой к каждой реакции.
Фармацевтические композиции
Настоящее изобретение может предусматривать одновременное введение одного или более видов молекул siНК. Эти виды можно выбирать для направления к одному или более генам-мишеням.
Молекулы siНК по настоящему изобретению и их композиции можно вводить непосредственно в глаз или применять местно (например, локально), что, в общем, известно специалистам в данной области техники. Например, молекула siNA может содержать средство-носитель для доставки, включающее в себя липосомы, для введения их субъекту. В фармацевтически приемлемых композициях могут присутствовать носители и разбавители и их соли. Молекулы нуклеиновой кислоты можно вводить в клетки различными известными специалистам в данной области техники способами, включающими в себя инкапсулирование в липосомы, ионофорез, или внедрением в другие носители, такие как биодеградируемые полимеры, гидрогели, циклодекстрины сополимера молочной и гликолевой кислоты (PLGA) и микросферы PLGA, биодеградируемые нанокапсулы и биоадгезивные микросферы, или посредством белковоподобных векторов, но способы не ограничены вышеперечисленным. В другом варианте осуществления молекулы нуклеиновых кислот по настоящему изобретению можно также создавать или образовывать комплексы с полиэтиленимином и его производными, такими как производные полиэтиленимин-полиэтиленгликоль-N-ацетилгалактозамина (PEI-PEG-GAL) или полиэтиленимин-полиэтиленгликоль-три-N-ацетилгалактозамина (PEI-PEG-triGAL).
Молекулу siNA по настоящему изобретению можно образовывать в комплексе со средствами разрушения мембраны и/или катионным липидом или хелперной липидной молекулой.
Системы доставки, которые можно использовать по настоящему изобретению, включают в себя, например, водные и неводные гели, кремы, гетерогенные эмульсии, микроэмульсии, липосомы, мази, водные и неводные растворы, лосьоны, аэрозоли, углеводородные основы и порошки и могут содержать наполнители, такие как солюбилизаторы, усилители проникновения (например, жирные кислоты, сложные эфиры жирных кислот, жирные спирты и аминокислоты), и гидрофильные полимеры (например, поликарбофил и поливинилпирролидон). В одном из вариантов осуществления фармацевтически приемлемый носитель представляет собой липосому или трансдермальный усилитель.
Фармацевтическая композиция по настоящему изобретению находится в форме, подходящей для введения, например для системного введения или местного применения, в клетку или у индивидуума, включая, например, человека. Подходящие формы отчасти зависят от применения или от пути введения, например перорального, трансдермального или инъекционного пути. Другие факторы известны в данной области техники и включают в себя, например, токсичность и формы, которые создают препятствия для проявления композицией своего эффекта. Настоящее изобретение также включает в себя композиции, подготовленные для хранения или для введения, которые включают в себя фармацевтически эффективное количество желательных соединений в фармацевтически приемлемом носителе или разбавителе. В области фармации известны приемлемые носители или разбавители для терапевтического использования. Например, можно использовать консерванты, стабилизаторы, красители и ароматизаторы. Они включают в себя бензоат натрия, сорбиновую кислоту и сложные эфиры п-гидроксибензойной кислоты. Дополнительно, можно использовать антиоксиданты и суспендирующие средства.
Фармацевтически эффективная доза представляет собой дозу, необходимую для профилактики, предупреждения возникновения или для лечения болезни (в некоторой степени для частичного снятия симптома, предпочтительно всех симптомов). Фармацевтически эффективная доза зависит от типа заболевания, от применяемой композиции, пути введения, вида млекопитающего, лечение которого осуществляется, от физикальных характеристик рассматриваемого конкретного млекопитающего, от сопутствующего лечения и от других факторов, которые известны специалистам в области медицины.
Обычно количество вводимых активных компонентов находится в диапазоне от 0,1 мг/кг до 100 мг/кг массы тела в день.
Композиции по настоящему изобретению можно вводить в композициях лекарственных форм, содержащих общепринятые нетоксичные фармацевтически приемлемые носители, адьюванты и/или носители. Композиции могут находиться в форме, подходящей для перорального применения, например в виде таблеток, пастилок, пластинок, водных или масляных суспензий, диспергируемых порошков или в виде гранул, эмульсии, твердых или мягких капсул, сиропов или эликсиров. Композиции, предназначенные для перорального использования, можно изготовлять согласно любому способу для изготовления фармацевтических композиций, известному в данной области техники, и такие композиции могут содержать одно или более подслащивающих веществ, ароматизирующих веществ, окрашивающих веществ или консервирующих веществ, для обеспечения фармацевтически изящных препаратов с привлекательным вкусом. Таблетки содержат активный компонент в смеси с нетоксичными фармацевтически приемлемыми наполнителями, которые подходят для изготовления таблеток.
Такими наполнителями могут быть, например, инертные разбавители, такие как карбонат кальция, карбонат натрия, лактоза, фосфат кальция или фосфат натрия; гранулирующие и дезинтегрирующие вещества, например кукурузный крахмал или альгиновая кислота; связующие вещества, например крахмал, желатин или камедь; и смазывающие вещества, например стеарат магния, стеариновая кислота или тальк. Таблетки могут быть непокрытыми, или их можно покрывать известными технологиями. В некоторых случаях такие покрытия можно изготовлять известными технологиями для задержки дезинтеграции и абсорбции в желудочно-кишечном тракте и, таким образом, обеспечивать пролонгированное действие в течение более длительного времени. Например, можно использовать материал для замедления времени, такой как глицерил моностеарат или глицерил дистеарат.
Композиции для перорального применения также могут быть представлены в виде твердых желатиновых капсул, в которых активный компонент смешивают с инертным твердым наполнителем, например с карбонатом кальция, фосфатом кальция или с каолином, или в виде мягких желатиновых капсул, в которых активный компонент смешивают с водной или масляной средой, например с арахисовым маслом, жидким вазелином или с оливковым маслом.
Водные суспензии содержат активные материалы в смеси с наполнителями, подходящими для изготовления водных суспензий. Такими наполнителями являются суспендирующие вещества, например натрия карбоксиметилцеллюлоза, метилцеллюлоза, гидроксипропил-метилцеллюлоза, альгинат натрия, поливинилпирролидон, трагакантовая камедь и гуммиарабик; диспергирующие или увлажняющие вещества могут представлять собой фосфатид природного происхождения, например лецитин, или продукты конденсации окиси алкилена с жирными кислотами, например с полиоксиэтилена стеаратом, или продукты конденсации окиси этилена с длинноцепочечными алифатическими спиртами, например с гептадекаэтиленоксиэтанолом, или продукты конденсации окиси этилена с неполными эфирами, полученным из жирных кислот и гексита, из таких как моноолеат сорбита полиоксиэтилена, или продукты конденсации окиси этилена с неполными эфирами, полученными из жирных кислот и ангидридов гексита, например моноолеата сорбита полиэтилена. Водные суспензии могут также содержать один или более консервантов, например этил, или n-пропил п-гидроксибензоат, одно или более окрашивающих веществ, одно или более ароматизирующих веществ и одно или более подслащивающих веществ, таких как сахароза или сахарин.
Можно создавать композиции масляных суспензий путем суспендирования активных компонентов в растительном масле, например в арахисовом масле, оливковом масле, сезамовом масле или в кокосовом масле, или в минеральном масле, таком как жидкий вазелин. Масляные суспензии могут содержать загустители, например пчелиный воск, твердый вазелин или цетиловый спирт. Можно добавлять подслащивающие и ароматизирующие вещества для обеспечения препаратов с желаемым вкусом. Эти композиции можно сохранять путем добавления антиоксиданта, такого как аскорбиновая кислота.
Диспергируемые порошки и гранулы, подходящие для приготовления водной суспензии путем добавления воды, обеспечивают активный компонент в смеси вместе с диспергирующим или увлажняющим веществом, суспендирующим веществом и одним или более консервантом. В качестве примеров подходящих диспергирующих или увлажняющих веществ или суспендирующих веществ приведены упомянутые выше вещества. Также могут присутствовать дополнительные наполнители, например подслащивающие, ароматизирующие и окрашивающие вещества.
Фармацевтические композиции по настоящему изобретению также могут быть в виде эмульсий масло в воде. Масляная фаза может представлять собой растительное масло или минеральное масло или их смеси. Подходящими эмульгирующими веществами могут быть камеди природного происхождения, например гуммиарабик или трагакантовая камедь, фосфатиды природного происхождения, например соевые бобы, лецитин, и сложные эфиры или неполные эфиры, полученные из жирных кислот и гексита, ангидриды, например моноолеата сорбит, и продукты конденсации упомянутых неполных эфиров с окисью этилена, например моноолеат сорбита полиоксиэтилена. Эмульсии могут также содержать подслащивающие и ароматизирующие вещества.
Можно создавать композиции сиропов и эликсиров с подслащивающими веществами, например с глицерином, пропиленгликолем, сорбитом, глюкозой или сахарозой. Такие композиции могут также содержать смягчающее средство, консервирующие, и ароматизирующие, и окрашивающие вещества. Фармацевтические композиции могут находиться в виде стерильной водной или маслянистой суспензии для инъекций.
Композиции этой суспензии можно создавать согласно известной технике, используя вышеупомянутые подходящие диспергирующие или увлажняющие вещества и суспендирующие вещества.
Стерильные препараты для инъекций могут также представлять собой стерильный раствор или суспензию для инъекций в нетоксичном приемлемом разбавителе или растворителе для парентерального введения, например, в виде раствора в 1,3-бутандиоле. Среди приемлемых носителей и растворителей, возможных для применения, находится вода, раствор Рингера и изотонический раствор хлорида натрия. Дополнительно, в качестве растворяющей или суспендирующей среды общепринято используют стерильные жирные масла. Для этих целей можно использовать любое мягкое жирное масло, включающее в себя синтетические моно- или диглицериды. Дополнительно, в приготовлении препаратов для инъекций находят применение жирные кислоты, такие как олеиновые кислоты.
Молекулы нуклеиновых кислот по настоящему изобретению также можно вводить в виде суппозиториев, например, для ректального введения лекарственного препарата. Эти композиции можно изготовлять, смешивая лекарственный препарат с подходящим нераздражающим наполнителем, который является твердым при обычной температуре, но жидким при ректальной температуре, и поэтому в прямой кишке он будет таять для высвобождения лекарственного препарата. Такие материалы включают в себя масло какао и полиэтиленгликоли.
Молекулы нуклеиновых кислот по настоящему изобретению также можно вводить парэнтерально в стерильной среде. Лекарственный препарат, в зависимости от носителя и применяемой концентрации, может быть или суспендирован или растворен в носителе. Предпочтительно, что адьюванты, такие как местноанестезирующие средства, консерванты и буферные вещества, можно растворять в носителе.
Подразумевается, что конкретный уровень дозы для любого конкретного субъекта зависит от различных факторов, включающих в себя активность конкретного используемого соединения, возраст, вес тела, общее состояние здоровья, пол, диету, время введения, путь введения и скорость выведения, комбинации препаратов и тяжесть конкретного заболевания, лечение которого проводят.
Также для введения животным нечеловеческого происхождения композиции можно добавлять в корма или в питьевую воду. Может быть удобным создание рецептуры композиции кормов и питьевой воды таким образом, чтобы животное принимало терапевтически соответствующее количество композиции вместе с кормлением. Также может быть удобным представлять композиции в виде премикса для добавления к кормам или к питьевой воде.
Для повышения общего терапевтического эффекта молекулы нуклеиновых кислот по настоящему изобретению также можно вводить субъекту в комбинации с другими терапевтическими соединениями. Использование сложных композиций для лечения симптомов может увеличивать полезные эффекты наряду со снижением наличия побочных эффектов.
Альтернативно, некоторые молекулы siНК по настоящему изобретению могут экспрессироваться внутри клеток их эукариотических промоторов. Можно доставлять и сохранять в клетках-мишенях рекомбинантные векторы, способные к экспрессии молекул siNA. Альтернативно, можно использовать векторы, которые обеспечивают транзиторную экспрессию молекул нуклеиновой кислоты. Такие векторы можно вводить повторно по мере необходимости. После экспрессирования молекула siНК взаимодействует с мРНК-мишенью и вызывает ответ РНКi. Доставка векторов, экспрессирующих молекулы siНК, может быть системной, например, путем внутривенного или внутримышечного введения, путем введения в клетки-мишени эксплантата, взятого у субъекта, с последующим повторным введением субъекту, или любыми другими средствами, которые дают возможность вводить их в желательную клетку-мишень.
Исследования на животных
Новозеландский кролик является золотым стандартом в экспериментальных платформах, разработанных для изучения ВГД. С ним легко обращаться, и у него большие глаза, сходные по размерам с человеческими. Кроме того, современное оборудование для измерения ВГД не подходит для применения у животных с маленькими глазами, такими как у мышей или крыс. Наконец, у кроликов значение ВГД (около 23 мм рт. ст.), которое можно снизить до 40% от его значения, используя серийные лекарственные препараты для местного гипотензивного лечения. Таким образом, несмотря на возможность создания модели глаукомы у кролика (например, хирургическим путем, блокируя эписклеральные вены или создавая искусственную окклюзию трабекулярной сети), были использованы нормотензивные кролики, пока не появилось легкое и воспроизводимое измерение фармакологического снижения ВГД.
Экспериментальный протокол
Использовали нормотензивных новозеландских белых кроликов (самцов весом 2-3 кг). Животных содержали в отдельных клетках со свободным доступом к корму и воде. Они находились при 12-часовых искусственных циклах свет/тьма, чтобы избежать неконтролируемых циркадных колебаний ВГД. Обращение с животными и лечение выполняли в соответствии с Директивой Совета Европейского Союза 86/609/EEC и заявлением Ассоциации офтальмологических исследований и зрения по использованию животных в офтальмологических исследованиях и изучении зрения.
Обычно лекарственные препараты вводили закапыванием небольшого объема (обычно 40 мкл) на роговичную поверхность. Контрлатеральные глаза обрабатывали единственным носителем, и их можно было использовать в качестве контроля в каждом эксперименте, что не является феноменом взаимовлияния на другой глаз. Было необходимо исключить многократные эксперименты на одном и том же животном.
Проводили измерения ВГД, используя контактный тонометр (TonoPen XL, Mentor, Norwell, Massachusetts). Тонометр TonoPen является очень удобным в силу его надежности и небольшого размера. Измерения этим инструментом выполняли, мягко прикладывая датчик тонометра на поверхность роговицы. Было показано, что это устройство является тонометром выбора для измерения внутриглазного давления у кроликов в пределах диапазона от 3 до 30 мм рт. ст. (Abrams et al., 1996). Все измерения попадали в пределы этого интервала: среднее исходное значение внутриглазного давления составляло 17,0±0,39 мм рт. ст. (n=100). Поскольку ВГД меняется от ночного значения до дневного, все эксперименты выполняли в одно и то же время, чтобы получить более стабильное ВГД и дать возможность объективного сравнения с лечением носителем. Чтобы избежать стресса у животных, проводили местную анестезию кроликов (оксибупрокаин/тетракаин, 0,4%/1%, в физиологическом растворе (объем:объем 1/4)). Раствор (10 мкл) наносили на роговицу перед каждым измерением внутриглазного давления. siРНК или физиологический раствор в объемах 40 мкл местно наносили на роговицу.
Стандартный протокол для применения siРНК у кролика был следующим. Дозы siРНК в физиологическом растворе (0,9% вес/объем) до конечного объема 40 мкл наносили на один глаз каждый день в течение четырех дней подряд. Контрлатеральный глаз принимали в качестве контроля, и в него закапывали 40 мкл стерильного физиологического раствора (0,9% в/о), в те же интервалы времени. Проводили измерение ВГД перед каждым применением и через 2 ч, 4 ч и 6 ч после закапывания в течение 10 дней. Максимальные ответы наблюдали между вторым и третьим днем. Для сравнения эффекта siРНК с другими гипотензивными соединениями проводили анализы ксалатана (0,005% латанапрост) и трастопа (2% дорзоламид) и измеряли ВГД при тех же условиях.
Результаты
Пример 1 анализов in vitro
Для определения ингибирования с использованием технологии РНКi различных мишеней, вовлеченных в глаукому, первым этапом было проведение экспериментов в клеточных культурах. Для каждой мишени было разработано несколько siРНК, используя специальное программное обеспечение согласно ранее описанным правилам. Для тестирования выбирали мишени с наилучшими характеристиками. siРНК применяли в клеточных культурах, таких как NPE, OMDC и HEK293. Проводили анализ эффекта siРНК на ген-мишень с помощью ПЦР в режиме реального времени и путем полуколичественной ПЦР согласно стандартным протоколам. Стандартизировали уровни транскрипта гена-мишени, используя актин в качестве гена домашнего хозяйства. Нижеприведенная таблица 1 показывает репрезентативные результаты экспериментов ПЦР в режиме реального времени для некоторых из ранее описанных генов-мишеней. Данные представляют средние значения процента интерференции siРНК по каждому экспрессируемому гену после стандартизации с контрольными клетками и их допустимые отклонения. По сравнению с контрольными клетками, уровень различных транскриптов и в точке времени 24 часа и в точке 48 часа после лечения siРНК был значительно снижен. В таблицу включены некоторые из различных протестированных siРНК и их различная эффективность в интерференции гена-мишени. siРНК, используемые в таблице, относятся к перечисленным человеческим siРНК-мишеням, приведенным на фигуре 2 следующим образом.
AC2: siРНК1: кроличья последовательность, гомологичная человеческой последовательности SEQ. ID. 73
siРНК2: кроличья последовательность, идентичная человеческой SEQ. ID. 54
siРНК3: кроличья последовательность, идентичная человеческой SEQ. ID. 66
PTGS1 siРНКl: кроличья последовательность, гомологичная человеческой SEQ. ID. 353
siРНК2: кроличья последовательность, гомологичная человеческой SEQ. ID. 369
PTGS2 siРНКl: кроличья последовательность, идентичная человеческой SEQ. ID. 426
siРНК2: кроличья последовательность, гомологичная человеческой SEQ. ID. 421
siРНК3: кроличья последовательность, гомологичная человеческой SEQ. ID. 477
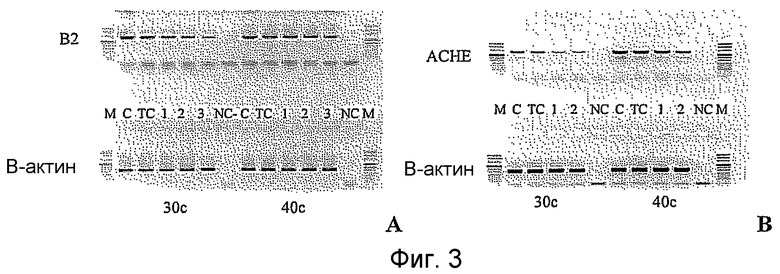
Фиг.3 показывает некоторые репрезентативные полуколичественные гели для некоторых из описанных выше мишеней. Уменьшение экспрессии гена для каждого гена-мишени зависит от эффективности сайленсинга siРНК. Для каждой мишени животной модели вводили наиболее эффективную siРНК, полученную в исследованиях in vitro. РНК приготовляли из клеток, обработанных различными siРНК. Пробы анализировали полуколичественной ПЦР с использованием специфических праймеров. Фигура показывает репрезентативный полуколичественный гель для экспрессии бета-адреноэргического рецептора 2 (A) и также для экспрессии ацетилхолинэстеразы (B). М: MW маркер; C: контрольные клетки; TC: контроль трансфекции; 1: siРНК1; 2: siРНК2; 3: siРНК3; NC: отрицательный контроль. Уровни экспрессии для каждой мишени зависят от эффективности сайленсинга siРНК. Показанные на фигуре siРНКs относятся к человеческим мишеням, приведенным на фигуре 2 следующим образом:
Столбец А (бета-адреноэргический рецептор 2)
1: кроличья последовательность, гомологичная человеческой SEQ. ID. № 122
2: кроличья последовательность, идентичная человеческой SEQ. ID. № 125
3: кроличья последовательность, гомологичная человеческой SEQ. ID. № 139
Столбец B (ацетилхолинэстераза)
1: кроличья последовательность, гомологичная человеческой SEQ. ID. № 162
2: кроличья последовательность, гомологичная человеческой SEQ. ID. № 177
Пример 2 анализов in vitro
Перед терапевтическим применением siРНК получали подтверждения в исследованиях in vivo для определения надлежащей доставки siРНК.
Выбранные в исследованиях in vitro siРНК применяли у животной модели согласно ранее описанному протоколу. Чтобы избежать эффекта колебаний ВГД по причине циркадных циклов, осуществляли все введения в одно и то же время. Для определения эффекта siРНК измерение значений внутриглазного давления (ВГД) проводили ранее упомянутым образом.
Поскольку глаукомная патология проявляется в повышении внутриглазного давления, задача состояла в том, чтобы достичь снижения его уровня после применения siРНК.
Большинство результатов для различных мишеней показывали значительное снижение уровней ВГД по сравнению с контролем, а также с серийными лекарственными средствами (латанопростом и дорзоламидом) и у животных, которым вводили единственный носитель (отрицательный контроль), не проявлялось какого-либо значительного изменения исходного уровня ВГД. Данные суммированы в таблице 2, где представлены стандартизированные средние значения максимального процента снижения ВГД после лечения siРНК и их допустимые отклонения. Снижение ВГД было статистически значимым для всех обработанных мишеней. Эти результаты указывают, что siРНК и серийные лекарственные средства действуют сходным образом, снижая уровни ВГД примерно на 20%, хотя siРНК проявляют более постоянный эффект. У животных не наблюдали каких-либо вторичных эффектов в ходе экспериментальных протоколов. Используемые в этих экспериментах siРНК относятся к человеческим мишеням, приведенным на фигуре 2 следующим образом:
AC2: кроличья последовательность, гомологичная человеческой SEQ. ID. № 73
AC4: кроличья последовательность, идентичная человеческой SEQ. ID. № 5
AC12: кроличья последовательность, идентичная человеческой SEQ. ID. № 522
ADRB1: кроличья последовательность, идентичная человеческой SEQ. ID. № 105
ADRB2: последовательность кролика, гомологичная человеческой SEQ. ID. № 139
ADRA1A: кроличья последовательность, гомологичная человеческой SEQ. ID. № 546
ADRA1B: кроличья последовательность, гомологичная человеческой SEQ. ID. № 619
ACHE: кроличья последовательность, гомологичная человеческой SEQ. ID. № 189
PTGS1: кроличья последовательность, гомологичная человеческой SEQ. ID. № 322
PTGS2: кроличья последовательность, идентичная человеческой SEQ. ID. № 426
SELE: кроличья последовательность, гомологичная человеческой SEQ. ID. № 262
ACE1: кроличья последовательность, гомологичная человеческой SEQ. ID. № 866
AGTR1: кроличья последовательность, гомологичная человеческой SEQ. ID. № 705
AGTR2: кроличья последовательность, идентичная человеческой SEQ. ID. № 774
ATP1A1: кроличья последовательность, идентичная человеческой SEQ. ID. № 1399
ATP1B2: кроличья последовательность, идентичная человеческой SEQ. ID. № 1820
СПИСОК ССЫЛОК



| название | год | авторы | номер документа |
|---|---|---|---|
| ОПОСРЕДОВАННОЕ РНКi ИНГИБИРОВАНИЕ Rho-КИНАЗЫ ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ НАРУШЕНИЙ | 2006 |
|
RU2432165C2 |
| ЛЕЧЕНИЕ НАРУШЕНИЙ ЦНС | 2007 |
|
RU2426544C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ КИШЕЧНИКА | 2006 |
|
RU2418594C2 |
| iPHK-ОПОСРЕДОВАННОЕ ИНГИБИРОВАНИЕ СВЯЗАННЫХ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ АЛЬФА СОСТОЯНИЙ | 2007 |
|
RU2469090C2 |
| МОДУЛИРОВАНИЕ ЭКСПРЕССИИ 11БЕТА-ГИДРОКСИСТЕРОИДНОЙ ДЕГИДРОГЕНАЗЫ 1 ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2006 |
|
RU2420582C2 |
| ЛЕЧЕНИЕ СИРТУИН 1 (SIRT1)-СВЯЗАННЫХ ЗАБОЛЕВАНИЙ ПУТЕМ ИНГИБИРОВАНИЯ НАТУРАЛЬНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА | 2009 |
|
RU2551234C2 |
| НОВЫЕ МИНИМАЛЬНЫЕ UTR-ПОСЛЕДОВАТЕЛЬНОСТИ | 2017 |
|
RU2759737C2 |
| КиРНК И ИХ ИСПОЛЬЗОВАНИЕ В СПОСОБАХ И КОМПОЗИЦИЯХ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ГЛАЗНЫХ ЗАБОЛЕВАНИЙ | 2012 |
|
RU2653766C2 |
| ЛЕЧЕНИЕ СВЯЗАННЫХ С ЭРИТРОПОЭТИНОМ (ЕРО) ЗАБОЛЕВАНИЙ ПУТЕМ ИНГИБИРОВАНИЯ ПРИРОДНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА К ЕРО | 2009 |
|
RU2620970C2 |
| СПОСОБЫ ГЕНЕТИЧЕСКОГО КОНТРОЛЯ ПОРАЖЕНИЯ РАСТЕНИЙ НАСЕКОМЫМИ И ПРИМЕНЯЕМЫЕ ДЛЯ ЭТОГО КОМПОЗИЦИИ | 2006 |
|
RU2478710C2 |







Изобретение относится к области биологии и медицины. Предложено применение технологии РНК-интерференции при лечении глазных болезней, предпочтительно глаукомы и увеита. Лекарственное средство составляют для местного нанесения на поверхность роговицы. В качестве мишеней предложено использовать карбоангидразу II, IV и XII, адренергические рецепторы: бета 1 и 2 и альфа 1А, 1В и 1D, ацетилхолинэстеразу, циклооксигеназы 1 и 2, АТФ-азы: альфа-1, альфа-2, альфа-3, бета-1, бета-2, молекулу адгезии лейкоцитов эндотелия (ELAM-1), ангиотензин I превращающие ферменты (АСЕ I и АСЕ II), рецепторы ангиотензина I (ATR1 и ATR2) и ренин; кохлин. 4 н. и 32 з.п. ф-лы, 2 табл., 3 ил.
1. Применение siHK для получения лекарственного средства для лечения глазных болезней с повышенным внутриглазным давлением (IO), где лекарственное средство составляют для местного нанесения на поверхность роговицы, и оно подавляет в глазу экспрессию гена-мишени, выбранного из группы, содержащей карбоангидразу II, IV и XII, адренергические рецепторы: бета 1 и 2, и альфа 1А, 1В и 1D, ацетилхолинэстеразу, циклооксигеназы 1 и 2, АТФ-азы: альфа-1, альфа-2, альфа-3, бета-1, бета-2, молекулу адгезии лейкоцитов эндотелия (ELAM-1), ангиотензин I превращающие ферменты (АСЕ I и АСЕ II), рецепторы ангиотензина II (ATR1 и ATR2) и ренин; кохлин.
2. Применение по п.1, где глазная болезнь выбрана из группы, состоящей из глаукомы, инфекции, воспаления, увеита и проявления системных заболеваний.
3. Применение по п.1, где глазной болезнью является глаукома.
4. Применение по п.1, где глазной болезнью является диабетическая ретинопатия.
5. Применение по пп.1-4, где siHK представляет собой siPHK.
6. Применение по п.5, где siPHK представляет собой dsPHK.
7. Применение по п.5, где siPHK представляет собой shPHK.
8. Применение по пп.1-4, где siHK содержит модифицированный олигонуклеотид.
9. Применение по любому из пп.1-4, где используют множество видов siHK.
10. Применение по п.9, где на указанное множество видов направлены одинаковые виды мРНК.
11. Применение по п.9, где на указанное множество направлены различные виды мРНК.
12. Применение по любому из пп.1-4, где siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1-646 или SEQ ID NO: 695-1829.
13. Применение по п.12, где ген-мишень представляет собой карбоангидразу IV, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1-46.
14. Применение по п.12, где ген-мишень является карбоангидразой II, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 47-98.
15. Применение по п.12, где ген-мишень представляет собой бета-адреноергический рецептор 1, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 99-109.
16. Применение по п.12, где ген-мишень представляет собой бета-адреноергический рецептор 2, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 110-160.
17. Применение по п.12, где ген-мишень является ацетилхолинэстеразой, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 161-190.
18. Применение по п.12, где ген-мишень представляет собой ELAM-1 (селектин Е), и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 191-318.
19. Применение по п.12, где ген-мишень представляет собой простагландинэндопероксидсинтетазу 1, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 319-374.
20. Применение по п.12, где ген-мишень представляет собой простагландинэндопероксидсинтетазу 2, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 375-491.
21. Применение по п.12, где ген-мишень является карбоангидразой XII, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 492-538.
22. Применение по п.12, где ген-мишень является альфа-адренергическим рецептором 1А, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 539-598.
23. Применение по п.12, где ген-мишень является альфа-адреноергическим рецептором 1В, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 599-634.
24. Применение по п.12, где ген-мишень является альфа-адреноергическим рецептором 1D, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 635-646.
25. Применение по п.12, где ген-мишень является ангиотензин II рецептором 1 типа, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 695-749.
26. Применение по п.12, где ген-мишень является ангиотензин II рецептором 2 типа, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO:750-807.
27. Применение по п.12, где ген-мишень является ренином, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1140-1196.
28. Применение по п.12, где ген-мишень является кохлином, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1197-1307.
29. Применение по п.12, где ген-мишень является АТФ-азой альфа 1, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1308-1500.
30. Применение по п.12, где ген-мишень является АТФ-азой альфа 2, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1501-1606.
31. Применение по п.12, где ген-мишень является АТФ-азой альфа 3, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1607-1705.
32. Применение по п.12, где ген-мишень представляет собой АТФ-азу бета 1, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1706-1780.
33. Применение по п.12, где ген-мишень представляет собой АТФ-азу бета 2, и siHK направлена на последовательность, выбранную из последовательностей SEQ ID NO: 1780-1829.
34. Выделенная молекула siHK для применения в лечении глазных болезней у пациента, характеризующихся изменением внутриглазного давления (IO), siHK, являющаяся комплементарной нуклеотидной последовательностью, выбранной из последовательностей SEQ ID NO: 1-646 или SEQ ID NO: 695-1829.
35. Применение выделенной молекулы siHK с последовательностью, которая комплементарна нуклеотидной последовательности, выбранной из последовательностей SEQ ID NO: 1-646 или SEQ ID NO: 695-1829, для изготовления лекарственного препарата для лечения глазных болезней.
36. Фармацевтическая композиция для лечения глазных болезней, содержащая siHK с последовательностью, которая комплементарна нуклеотидной последовательности, выбранной из последовательностей SEQ ID NO: 1-646 или SEQ ID NO: 695-1829.
| WO 2004042024, 21.05.2004 | |||
| WO 03087367, 23.10.2003 | |||
| WO 2005076998, 25.08.2005 | |||
| WO 2005079815, 01.09.2005 | |||
| HERKEL U ET AL | |||
| Update on topical carbonic anhydrase inhibitors | |||
| CURRENT OPINION IN OPHTHALMOLOGY | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Шланговое соединение | 0 |
|
SU88A1 |
| WOODWARD D F ET AL | |||
| The inflow and outflow of anti-glaucoma drugs TRENDS IN | |||

