ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способам и композициям для лечения патологических состояний центральной нервной системы (ЦНС) посредством интраназального введения композиций, которые изменяют экспрессию и/или активность генов, вовлеченных в проявление вышеуказанных состояний, с помощью РНК-интерференции. Композиции согласно изобретению содержат короткие интерферирующие молекулы нуклеиновых кислот (siНК) и связанных с ними соединений. В предпочтительных вариантах осуществлений интраназальную доставку молекул siНК, нацеленных на гены тау, гентингтина, ацетилхолинэстеразы, а также на мутантные аллели указанных или иных генов ЦНС, используют для лекарственных средств, производимых для лечения заболеваний ЦНС, таких как слабоумие, болезни Альцгеймера, Гентингтона и/или Паркинсона, а также врожденных заболеваний, связанных с мутациями в других генах ЦНС.
УРОВЕНЬ ТЕХНИКИ
РНКi является инструментом для изменения экспрессии гена
РНК-интерференция относится к посттранскрипционному процессу последовательность-специфического сайленсинга, опосредованного двухцепочечными РНК (дцРНК). После открытия данного феномена у растений в начале 1990-х годов Andy Fire и Craig Mello показали, что дцРНК специфически и селективно подавляют экспрессию генов крайне эффективным способом у Caenorhabditis elegans (Fire et ah, 1998). Последовательность первой цепи (смысловая РНК) совпадала с таковой соответствующего участка матричной РНК-мишени (мРНК). Вторая нить (антисмысловая РНК) была комплементарна мРНК. Образовавшаяся дцРНК выключала ген на несколько порядков более эффективно, чем соответствующие одноцепочечные молекулы РНК (в частности, антисмысловая РНК).
Процесс РНКi начинается, когда фермент DICER встречается с дцРНК и расщепляет ее на фрагменты, называемые короткими интерфирирующими РНК, или siРНК. Этот белок принадлежит семейству нуклеаз РНКазы III. Комплекс белков включает в себя эти фрагменты РНК и использует эти коды для нахождения и разрушения любых РНК в клетке с совпадающей последовательностью, таких как мРНК-мишени (см. Bosher & Labouesse, 2000; и Akashi et al., 2001).
При попытках применить РНКi для нокдауна гена было выяснено, что клетки млекопитающих развили различные способы защиты против вирусных инфекций, которые могут препятствовать использованию данного подхода. Действительно, присутствие крайне малых уровней вирусных дцРНК усиливает ответ, опосредуемый интерфероном, что приводит к общему неспецифическому подавлению трансляции, которая, в свою очередь, запускает апоптоз (Williams, 1997, Gil & Esteban, 2000).
В 2000 году сообщалось, что дцРНК специфически ингибируют три гена в ооците мыши и раннем эмбрионе. Арест трансляции и, таким образом, PKR-ответ не наблюдался, так как эмбрионы продолжали развиваться (Wianny & Zernicka-Goetz, 2000). Исследования в Ribopharma AG (Kulmbach, Germany) показали, что функционирование РНКi в клетках млекопитающих с использованием коротких (20-24 пар оснований) дцРНК для выключения генов в человеческих клетках не приводит к инициации ответа с острой фазой. Подобные эксперименты были проведены другими исследовательскими группами и подтвердили эти результаты (Elbashir et al., 2001; Caplen et al., 2001). При проверке на разнообразных нормальных и опухолевых клеточных линиях человека и мыши было определено, что короткие шпилечные РНК (shРНК) могут заставить молчать гены так же эффективно, как и их siРНК-копии (Paddison et al., 2002). Недавно для другой группы коротких РНК (21-25 пар оснований) было показано, что они опосредуют даун-регуляцию экспрессии гена. Такие РНК, короткие временно регулирующие РНК (stРНК), регулируют экспрессию гена в процессе развития Caenorhabditis elegans (см. обзор Banerjee & Slack, 2002 и Grosshans & Slack, 2002).
Исследователи использовали РНКi в нескольких системах, включая Caenorhabditis elegans, дрозофилу, трипаносомы и других беспозвоночных. Несколько групп недавно представили результаты по специфическому подавлению биосинтеза белка в различных клеточных линиях млекопитающих (особенно в клетках HeLa), показав, что РНКi может быть широко используемым методом для генного сайленсинга in vitro. На основе этих результатов РНКi быстро стала общепризнанным инструментом для проверки (идентификация и определение) функций гена. В РНКi используются короткие олигонуклеотиды дцРНК, что обеспечит понимание функций генов, чья последовательность определена частично.
Недавно Krutzfeldt и коллеги показали, что особенным образом сконструированные соединения, названные «антагомирами», могут эффективно подавлять действие микроРНК (miРНК), не кодирующих кусочков РНК, которые регулируют экспрессию гена (Krutzfeldt et al., 2005).
Интраназальная доставка продуктов siНК
Аэрозольная доставка нуклеиновых кислот к легким с использованием вирусных векторов, полимеров, поверхностно-активных веществ или инертных наполнителей была описана для лечения заболеваний легких. Были предложены подходящие нуклеиновые кислоты для интраназальной доставки, включая дцДНК, дцРНК, оцДНК, оцРНК, короткие интерферирующие РНК, микроРНК и антисмысловые РНК (см. US2005/0265927 и WO2005/115358).
Предпочтительные вещества для доставки соединений, инициирующих РНКi, в легкие содержат катионные полимеры, модифицированные катионные полимеры, липиды и подходящие для введения поверхностно-активные вещества (см. US20050008617).
Доставка в ЦНС
Интраназальная доставка для лечения заболеваний ЦНС была только достигнута с помощью ингибиторов ацетилхолинэстеразы, таких как галантамин и различные соли и производные галантамина (для примера см. US2006003989, WO2004/002402, WO2005/102275), в то время как лечение нейродегенеративных заболеваний под действием незаряженных малых интерферирующих РНК в ЦНС было получено путем хирургической имплантации катетера (см. для примера WO2005/116212). В WO02/086105 описан способ доставки олигонуклеотидов в ЦНС посредством нейронных путей, берущих начало в носовой полости. Обсуждается использование антисмысловых олигонуклеотидов, но при этом не приведено ни одной ссылки на РНК-интерференцию. В данной публикации отсутствует также описание физиологической активности доставляемых олигонуклеотдов. При внутривенном введении siРНК было показано также, что они проходят через барьер кровь-сетчатка и изменяют экспрессию генов в глазе (WO03/087367, US2005/0222061). Изменение экспрессии определенных генов, участвующих в болезни Альцгеймера, таких как β-секретаза (BACE), амилоидный белок-предшественник (APP), PIN-1, пресенилин 1 (PS-1) и/или пресенилин 2 (PS-2), а также генов, вовлеченных в болезнь Гентингтона, таких как гентингтин или атаксин-1, было достигнуто с помощью siНК как в культуре клеток, так и in vivo посредством интратекального и интрацеребровентрикулярного введения, имплантации катетеров и помп, химическим или осмотическим открытием барьера кровь-мозг или путем прямой инъекции или перфузии в артериальную систему мозга (например, в полосатое тело, кору головного мозга, - см. для примера WO2005/003350, US2005/042646, and GB2415961).
Нацеливание на ген тау посредством РНКi
[Ген] Тау играет центральную роль в наследственных и приобретенных формах возрастных деменций, включая болезнь Альцгеймера (БА) (Hardy & Selkoe, 2002; Lee et al., 2001; Mullan et al., 1992; Poorkaj et al., 1998; Hutton et al., 1998). БА характеризуется двумя главными отличительными патологическими признаками: старческими бляшками, которые содержат бета-амилоид (AP), полученный после расщепления амилоидного белка-предшественника (APP), и нейрофибриллярные узлы, которые содержат нитчатый белок тау. Редкая наследственная форма БА раскрыла существенную роль продуцирования AP в патогенезе всех форм БА, как спорадической, так и наследственной (Hardy & Selkoe, 2002). Известно, что мутации в трех генах являются причиной наследственной БА - эти гены кодируют APP, пресенилин 1 и пресенилин 2, - обладают доминантным действием в усилении продукции нейротоксического бета-амилоида (Hardy & Selkoe, 2002).
Тау, главный компонент нейрофибриллярных узлов, также играет существенную роль в патогенезе БА (Lee et al., 2001). Мутации в тау вызывают подобные наследственные доминантные нейродегенеративные заболевания, лобно-височную деменцию с паркинсонизмом, связанным с 17 хромосомой (FTDP-17). В FTDP-17 мутации тау либо изменяет последовательность белка тау, либо ведут к ошибочному сплайсингу (Lee et al., 2001; Lewis et al., 2001; Oddo et al., 2003). Нарушения в экспрессии тау также вносят вклад в некоторые другие важные нейродегенеративные расстройства, включая прогрессирующий надъядерный паралич и кортикальную-базальную ганглионарную дегенерацию (Houlden et al., 2001). Таким образом, усилия по уменьшению экспрессии тау либо в целом, либо аллель-специфическим способом могут оказаться терапевтически полезными при FTDP-17, БА или других тау-связанных заболеваниях.
Аллель-специфический сайленсинг мутаций в тау и/или связанных однонуклеотидных полиморфизмах (SNP) посредством РНКi был уже достигнут на клеточных культурах (Miller et al. 2003, 2004). Кроме того, представляющие интерес siРНК были успешно доставлены на мышиной модели посредством инъекции в хвостовую вену (US2004/0241854).
Предшествующее описание приведено для пояснения специалистам из смежных областей понятий, относящихся к РНКi, а также к подходам для доставки в ЦНС. Пояснения представлены только для понимания последующего изобретения и не является признанием того, что любая из описанных публикаций относится к известному уровню техники по отношению к заявленному изобретению. В данной области есть большая потребность в удобных способах, посредством которых молекулы siНК могут быть доставлены в ЦНС, и в результате подобная доставка приводит к РНК-интерферирующей активности. Авторы изобретения разработали технологии для изменения экспрессии генов in vivo для лечения заболеваний ЦНС интраназальным путем посредством нацеливания молекул siРНК на ЦНС.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предоставляет способы и композиции для лечения патологий центральной нервной системы (ЦНС) путем интраназального применения соединений, которые вызывают РНК-интерференцию.
Композиции согласно изобретению содержат молекулы коротких интерферирующих нуклеиновых кислот (siНК) и связанных соединений, включая, но не ограничиваясь ими, короткие интерферирующие РНК (siРНК), двухцепочечные РНК (дцРНК), короткие шпилечные РНК (shРНК), микроРНК (miРНК), антагомиры и молекулы, способные опосредовать РНК-интерференцию.
Способы согласно изобретению включают в себя введение пациенту, в случае необходимости, эффективного количества одной или более siНК для лечения патологических состояний ЦНС. В предпочтительных вариантах осуществлений способы согласно изобретению включают в себя интраназальное применение терапевтических siРНК. Заслуживает большого внимания то, что композиции согласно изобретению могут быть использованы при получении лекарственного средства для лечения патологий ЦНС, включая деменцию, болезни Альцгеймера, Гентингтона и/или Паркинсона, а также врожденных заболеваний, связанных, помимо прочего, с мутациями генов ЦНС. Патологии и заболевания, которые могут быть подвержены лечению в соответствии со способами изобретения, предпочтительно включают те, которые действуют на гиппокамп, кору головного мозга и/или полосатое тело.
При одном варианте осуществления настоящее изобретение относится к siНК или к подобным химически синтезированным веществам, которые направленно препятствуют экспрессии мРНК с генов тау, гентингтина или ацетилхолинэстеразы, а также других мутантных аллелей указанных или других генов ЦНС, которые в конечном счете модулируют количество продуцируемого белка. В предпочтительном варианте осуществления композиции согласно настоящему изобретению применяют интраназально для специфического нацеливания на аномальный вариант представляющего интерес гена в ЦНС.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Изобретение будет описано только на примере, со ссылкой на соответствующие чертежи.
Фигура 1. Уровень экспрессии GFP после интраназального введения мыши 0,9% NaCl (контроль), 1 нмоль/мкл siРНК-GFP (мышь 1), 2 нмоль/мкл siРНК-GFP (мышь 2) и 2 нмоль/мкл siРНК-GFP + TransIT-TKO (мышь 3). Данные проведенного анализа коры головного мозга, гиппокампа, полосатого тела и луковицы ЦНС.
Фигура 2. siРНК уменьшает уровни белка GFP. siРНК, предназначенная против GFP, была интраназально введена трансгенным GFP-мышам. Животных умерщвляли в разное время, а ткани собирали и анализировали методом Вестерн-блотинга. Контрольный солевой раствор вводили мышам CI, CII и CIII в качестве контроля. Значения показывают уровни экспрессии белка GFP, нормализованные на белок GFP у контрольных мышей.
Фигура 3. siРНК уменьшает уровни мРНК GFP. siРНК, предназначенная против мРНК GFP, была интраназально введена трансгенным GFP-мышам. Животных умерщвляли в разное время, и собирали мРНК из разных тканей. На фигуре 3 показана экспрессия мРНК GFP в полосатом теле и коре головного мозга, анализ которой осуществляли методом количественной ПЦР.
Фигура 4. siРНК уменьшает уровни транскриптов гена MAPT с различными мутациями. siРНК, сконструированные против различных мутаций, были проанализированы, SEQ ID NO:159 (мутация P301L) и SEQ ID NO:160 (мутация R406W). РНК получали из клеток, обработанных специфическими siРНК в течение 48 ч. Образцы анализировали методом количественной ПЦР, используя специфические праймеры для MAPT (описаны в тексте). Значения соответствуют среднему уровню экспрессии различных транскриптов, нормализованных на 18S, относительно трансфекции клеток пустым вектором в качестве контроля.
Фигура 5. siРНК уменьшает уровни транскриптов гена MAPT. РНК получали из клеток MDA-MB-435, обработанных различными siРНК в течение 24, 48 ч и 72 ч. Образцы анализировали методом ПЦР в режиме реального времени, используя специфические праймеры, описанные в тексте. Значения соответствуют среднему уровню экспрессии различных транскриптов, нормализованных на 18S, относительно трансфекции клеток пустым вектором в качестве контроля.
Фигура 6. siРНК SEQ ID NO:160, сконструированная против мутации R406W, уменьшает уровни белка MAPT in vivo. siРНК SEQ ID NO:160 интраназально вводили трансгенным мышам MAPT. Животных умерщвляли через семь дней после введения siРНК, а ткани гиппокампа собирали и анализировали методом Вестерн-блоттинга.
Фигура 7. Список мутаций и номера доступов последовательности гена MAPT.
Фигура 8. Список последовательностей участков MAPT, на которые нацелены siНК согласно изобретению, а также дуплексов siНК, нацеленных на данные участки.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам и композициям для лечения патологий центральной нервной системы (ЦНС) путем интраназального применения соединений, которые вызывают РНКi. Композиции согласно изобретению содержат молекулы коротких интерферирующих нуклеиновых кислот (siНК), которые модулируют экспрессию генов-мишеней, связанных с измененными состояниями ЦНС.
Способы согласно изобретению включают в себя введение пациенту, в случае необходимости, эффективного количества одной или более siНК согласно изобретению.
Конструирование siРНК
Ген «таргетируют» с помощью siНК согласно изобретению [или siНК согласно изобретению направляют на ген-мишень], при этом siНК селективно уменьшает или ингибирует экспрессию гена или аллеля гена, вовлеченного в патологическое состояние. Альтернативно siНК таргетирует ген, когда siНК гибридизуется с транскриптом при условии точного соответствия. siНК могут быть протестированы на способность к таргетингу гена либо in vitro, либо in vivo.
В 1999 г. Tuschl et al. раскрыл эффект сайленсинга с помощью siРНК, показав, что их эффективность есть функция от длины дуплекса, длины 3'-выступающих концов и последовательностей данных выступающих концов.
Правильный выбор гомологичной области внутри гена-мишени имеет большую значимость для точного сайленсинга. Короткие фрагменты последовательности гена-мишени (например, 19-40 нуклеотидов в длину) выбраны как последовательность siНК согласно изобретению. Альтернативно, вариабельный участок представляющего интерес аллеля выбирают в качестве мишеней для соединений siНК. При одном варианте осуществления siНК представляет собой siРНК. При данном варианте осуществления короткий фрагмент последовательности гена-мишени представляет собой фрагмент мРНК гена-мишени. В предпочтительных вариантах осуществлений критерии для выбора фрагмента последовательности из мРНК гена-мишени в качестве молекулы siРНК-кандидата включают в себя следующее: 1) последовательность мРНК из гена-мишени должна по меньшей мере на 50-100 нуклеотидов отстоять от 5'- или 3'-конца природной молекулы мРНК; 2) последовательность из мРНК гена-мишени должна иметь G/C-состав между 30% и 70%, более предпочтительно около 50%; 3) последовательность из мРНК гена-мишени не должна содержать повторяющихся последовательностей (например, AAA, CCC, GGG, TTT, AAAA, CCCC, GGGG, TTTT); 4) последовательность из мРНК гена-мишени должна быть доступна в мРНК; и 5) последовательность из мРНК гена-мишени должна быть уникальна для гена-мишени. Последовательность фрагмента из мРНК гена-мишени может соответствовать одному или более вышеуказанным установленным критериям. В предпочтительных вариантах осуществлений siРНК имеет G/C-состав менее 60% и/или в ней отсутствуют повторяющиеся последовательности.
Практически представляющий интерес ген вводят в виде нуклеотидной последовательности в программу для предсказания, в которой учитываются все переменные, описанные выше для конструирования оптимальных олигонуклеотидов. Данная программа сканирует любую нуклеотидную последовательность мРНК для выявления участков, пригодных для нацеливания siРНК. Результатом данного анализа является оценка возможного олигонуклеотида siРНК. Наиболее подходящие используют для конструирования олигонуклеотидов двухцепочечной РНК (обычно длиной в 21 п.н., хотя другие длины также возможны), которые обычно получают путем химического синтеза. Несколько химических модификаций, которые известны в данной области, направленные на увеличение стабильности или доступности олигонуклеотидов дцРНК, также возможны.
Олигонуклеотиды-кандидаты могут быть далее отобраны на основе межвидового консерватизма с тем, чтобы облегчить переход клинических исследований от животных к человеку.
Дополнительно к siНК, которые полностью комплементарны области-мишени, вырожденные последовательности siНК могут быть использованы также для нацеливания на гомологичные участки. В WO2005/045037 описано конструирование молекул siНК для нацеливания на такие гомологичные последовательности, например, путем введения неканонических пар оснований, для примера, мисматчей и/или болтающихся пар оснований, которые могут обеспечить дополнительные таргетируемые последовательности. В примерах, где определены мисматчи, неканонические пары оснований (например, мисматчи и/или вырожденные основания) могут быть использованы для синтеза молекул siНК, которые нацелены более чем на одну последовательность. В неограничивающем примере неканонические пары оснований, такие как UU- или CC-пары оснований, используют для образования молекул siНК, способных к нацеливанию на последовательности, у которых наблюдается гомология последовательностей. По существу, одним из преимуществ использования siНК согласно изобретению является то, что одна и та же siНК может содержать последовательность нуклеиновой кислоты, которая консервативна между гомологичными генами. В данном подходе одна siНК может быть использована для ингибирования экспрессии более чем одного гена, вместо того, чтобы использовать более чем одну молекулу siНК для нацеливания на разные гены.
Идентичность последовательности может быть подсчитана путем сравнения последовательностей и алгоритмов выравнивания, известных в данной области (см. Gribskov and Devereux, Sequence Analysis Primer, Stockton Press, 1991, и цитируемые здесь ссылки), и подсчета процента различий между нуклеотидными последовательностями, например, с помощью алгоритма Smith-Waterman, внедренного в программное обеспечение BESTFIT, используя значения по умолчанию (например, University of Wisconsin Genetic Computing Group). Предпочтительна более чем 90%, 95% или 99% идентичность последовательностей между siНК и областью гена-мишени. Альтернативно комплементарность между siНК и природной РНК может быть определена функционально путем гибридизации, а также по способности уменьшать или ингибировать экспрессию гена-мишени. Способность siНК воздействовать на экспрессию гена может быть определена опытным путем либо in vivo, либо in vitro.
В соответствии с изобретением предпочтительными молекулами siНК являются двухцепочечные. В одном из вариантов осуществления двухцепочечная молекула siНК содержит затупленные концы. В другом варианте осуществления двухцепочечная молекула siНК содержит выступающие нуклеотиды (например, 1-5 выступающих нуклеотидов, предпочтительно, 2 выступающих нуклеотида). В конкретном варианте осуществления выступающими нуклеотидами являются 3'-выступающие концы. В другом конкретном варианте осуществления выступающими нуклеотидами являются 5'-выступающие нуклеотиды. Любые типы нуклеотидов могут быть частью выступающего конца. В одном из вариантов осуществления выступающий нуклеотид или нуклеотиды являются рибонуклеиновыми кислотами. В другом варианте осуществления в качестве выступающего нуклеотида или нуклеотидов могут выступать дезоксирибонуклеиновые кислоты. В предпочтительном варианте осуществления в качестве выступающего нуклеотида или нуклеотидов могут выступать тимидиновые нуклеотиды. В другом варианте осуществления в качестве выступающего нуклеотида или нуклеотидов могут выступать модифицированные или неклассические нуклеотиды. Выступающие нуклеотид или нуклеотиды могут иметь неклассические внутринуклеотидные связи (например, отличные от фосфодиэфирных связей).
Синтез дуплексов siНК
siНК могут быть синтезированы любым из известных способов, известных в данной области. Предпочтительно РНК синтезируют химически, используя соответствующим образом защищенные фосфорамидаты рибонуклеозидов и серийный ДНК/РНК-синтезатор. Кроме того, siРНК могут быть получены от коммерческих поставщиков, включая, но не ограничиваясь ими, Proligo (Гамбург, Германия), Dharmacon Research (Лафейетт, CO, США), Glen Research (Sterling, VA, США), ChemGenes (Ашленд, MA, США), и Cruachem (Glasgow, UK), Qiagen (Германия) Ambion (США) и Invitrogen (Шотландия). Альтернативно, молекулы siНК в соответствии с изобретением могут быть экспрессированы в клетках в результате трансфекции клеток векторами, содержащими обратно комплементарную последовательность siНК под контролем промотора. После экспрессии siНК можно выделить из клеток, используя хорошо известные в данной области технологии.
Этап отжига необходим при работе с одноцепочечными молекулами РНК. Для отжига РНК 30 мкл 50 мкМ раствора каждого РНК-олигонуклеотида объединяют с 100 мМ ацетатом калия, 30 мМ HEPES-KOH с pH 7,4, с 2 мМ ацетатом магния. Растворы затем инкубируют в течение 1 минуты при 90°C, центрифугируют в течение 15 секунд и инкубируют в течение 1 часа при 37°С.
В вариантах осуществления, когда siРНК представляет собой короткую шпилечную РНК (shРНК), две нити молекулы siРНК могут быть связаны линкерным участком (например, нуклеотидным линкером или ненуклеотидным линкером).
Химические модификации siНК.
В соответствии с изобретением siНК могут включать в себя один или более модифицированных нуклеотидов и/или нефосфодиэфирные связи. Химические модификации, известные в данной области, могут увеличивать стабильность, доступность и/или клеточное поглощение siНК. Специалист может знать другие типы химических модификаций, которые могут быть введены в молекулы РНК (см. международные публикации WO03/070744 и WO2005/045037 для общего представления о типах модификаций).
В одном варианте осуществления модификации могут быть использованы для обеспечения повышенной стойкости к деградации или улучшенному поглощению. Примеры подобных модификаций включают фосфоротиоатные межнуклеотидные связи, 2'-O-метилрибонуклеотиды (в особенности на смысловой цепи двухцепочечной siРНК), 2'-дезокси-фторрибонуклеотиды, 2'- дезоксирибонуклеотиды, "универсальные основания" нуклеотидов, 5-C-метил нуклеотиды, а также введение обращенных остатков дезоксиоснований (см. в целом GB2406568).
В другом варианте осуществления модификации могут быть использованы для усиления стабильности siРНК или для увеличения эффективности нацеливания. Модификации включают в себя химическую сшивку между двумя комплементарными нитями siРНК, химическую модификацию 3'- или 5'-конца цепи siРНК, модификации сахара, модификации оснований нуклеотида и/или модификации боковых связей, 2'-фтор-модифицированные рибонуклеотиды и 2'-дезоксирибонуклеотиды (см. в целом международную публикацию WO2004/029212).
В другом варианте осуществления модификации могут быть использованы для увеличения или уменьшения аффинности для комплементарных нуклеотидов в мРНК-мишени и/или в комплементарной цепи siНК (см., в целом, международную публикацию WO2005/044976). Например, немодифицированный пиримидиновый нуклеотид может быть замещен на 2-тио-, 5-алкинил-, 5-метил- или 5-пропинилпиримидин. Кроме того, немодифицированный пурин может быть замещен на 7-деза-, 7-алкил- или 7-алкенилпурин.
В другом варианте осуществления, в котором siНК представляет собой двухцепочечную siРНК, 3'-концевые выступающие нуклеотиды замещены на дезоксирибонуклеотиды (см., в целом, Elbashir et al., 2001).
В одном варианте осуществления изобретение характеризуется двухцепочечной молекулой короткой интерферирующей нуклеиновой кислоты (siНК), которая снижает экспрессию гена-мишени, предпочтительно, экспрессию гена в ЦНС, более предпочтительно, гена MAPT, где сборка молекулы siНК происходит из двух отдельных олигонуклеотидных фрагментов, где один фрагмент содержит смысловой участок, а второй фрагмент содержит антисмысловой участок молекулы siНК. В другом варианте осуществления приблизительно 19 нуклеотидов каждого фрагмента молекулы siНК представляют собой пары оснований, комплементарных нуклеотидам другого фрагмента молекулы siНК, и где два 3'-концевых нуклеотида каждого фрагмента молекулы siНК не представляют собой пары оснований нуклеотидов другого фрагмента молекулы siНК (то есть молекула siНК содержит выступающие концы по меньшей мере с 2 нуклеотидами на каждой цепи). В одном варианте осуществления каждый из двух 3'-концевых нуклеотидов каждого фрагмента молекулы siНК представляет собой 2'-дезоксипиримидиновый нуклеотид, такой как 2'-дезокситимидин. В другом варианте осуществления все нуклеотиды в количестве 21 нуклеотида каждого фрагмента молекулы siНК являются парами оснований, комплементарных нуклеотидам другого фрагмента молекулы siНК. В другом варианте осуществления приблизительно 19 нуклеотидов антисмысловой области представляют собой парные основания для нуклеотидной последовательности или части РНК, кодируемой геном-мишенью. В другом варианте осуществления около 21 нуклеотида антисмысловой области спарены с нуклеотидной последовательностью или частью РНК, кодируемой геном-мишенью. В любом из вышеописанных вариантов осуществления 5'-конец фрагмента, включающий в себя указанный антисмысловой участок, может дополнительно содержать фосфатную группу.
В одном варианте осуществления изобретение характеризуется молекулой siНК, в которой либо одна, либо обе - и смысловая, и антисмысловая - цепи содержат одну или более, например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, предпочтительно, от 1 до 5, фосфоротиоатных межнуклеотидных связей, и/или одну или более (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более), предпочтительно, от 1 до 5, 2'-дезокси, 2'-O-метил, 2'-дезокси-2'-фтор, и/или приблизительно один или более (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более), предпочтительно, от 1 до 5 нуклеотидов, модифицированных универсальных оснований и дополнительно концевую молекулу кэпа на 3'-конце, 5'-конце или на обоих, 3'- и 5'-концах одной или обеих смысловой или антисмыловой цепей. В другом варианте осуществления один или более, например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, предпочтительно, от 1 до 5 пиримидиновых нуклеотидов смысловой и/или антисмыловой цепи siНК являются химическими модифицированными 2'-дезокси-, 2'-O-метил-, 2'-дезокси-2'-фтор-нуклеотидами, дополнительно одним или более, например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, или более, предпочтительно, от 1 до 5, фосфоротиоатных межнуклеотидных связей, и/или концевых молекул кэпа на 3'-конце, на 5'-конце или на обоих, 3'- и 5'-концах, которые присутствуют на одной или на разных цепях.
В одном варианте осуществления для изобретения характерно то, что химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК) содержит приблизительно от 1 и приблизительно до 5 или более (в частности, приблизительно от 1, 2, 3, 4, 5 или более) фосфоротиоатных межнуклеотидных связей в каждой цепи молекулы siНК.
В другом варианте осуществления для изобретения характерно то, что молекула siНК содержит 2'-5'-межнуклеотидные связи. 2'-5' межнуклеотидная(ые) связь(и) может находиться на 3'-конце, на 5'-конце или на обоих, 3'- и 5'-концах, одной или обеих нитей последовательности siНК. Кроме того, 2'-5'-межнуклеотидная(ые) связь(и) может находиться в различных других положениях внутри одной или обеих цепей последовательности siНК, например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, включая каждую межнуклеотидную связь пиримидинового нуклеотида на одной или обеих цепях последовательности молекулы siНК; в различных положениях внутри одной или обеих цепей последовательности siНК может содержаться 2'-5'-межнуклеотидная(ые) связь(и), например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, может содержаться 2'-5'-межнуклеотидная(ые) связь(и).
В одном варианте осуществления молекула siНК согласно изобретению может содержать один или более (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более) блокирующих нуклеиновую кислоту (LNA) нуклеотидов, например, на 5'-конце, на 3'-конце, на обоих, 5'- и 3'-концах, или их комбинациях, молекулы siНК.
В другом варианте осуществления молекула siРНК согласно изобретению содержит один или более (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более) ациклических нуклеотидов, например, на 5'-конце, на 3'-конце, на обоих, 5'- и 3'-концах, или их комбинациях, молекулы siНК.
В одном варианте осуществления для изобретения характерно то, что согласно изобретению химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК), в которой любые (например, один или более, или все) пиримидиновые нуклеотиды, присутствующие либо на одном, либо на обоих, смысловом или антисмысловом, участках, являются 2'-дезокси-2'-фторпиримидиновыми нуклеотидами, и в которой любые (например, один или более, или все) пуриновые нуклеотиды, присутствующие либо на одном, либо на обоих, смысловом или антисмысловом, участках, являются 2'-дезоксипуриновыми нуклеотидами. Кроме того, любые нуклеотиды, содержащие 3'-концевой выступающий нуклеотид, которые присутствует на указанном смысловом или антисмысловом участке, представляет собой 2'-дезоксинуклеотиды.
В одном варианте осуществления для изобретения характерно то, что согласно изобретению химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК), в которой любые (например, один или более, или все) пиримидиновые нуклеотиды, присутствующие либо на одном, либо на обоих, смысловом или антисмысловом, участках, являются 2'-дезокси-2'-фторпиримидиновыми нуклеотидами, и в которой любые (например, один или более, или все) пуриновые нуклеотиды, присутствующие либо на одном, либо на обоих, смысловом или антисмысловом, участках, являются 2'-O-метилпуриновыми нуклеотидами. Кроме того, любые нуклеотиды, содержащие 3'-концевой выступающий нуклеотид, которые присутствует на указанном смысловом или антисмысловом участке, представляет собой 2'-дезоксинуклеотиды.
В одном варианте осуществления для изобретения характерно то, что химически синтезированная двухцепочечная молекула РНК, которая направляет расщепление РНК-мишени, предпочтительно, РНК, которая экспрессируется в ЦНС, более предпочтительно, РНК MAPT, посредством РНК интерференции, в которой каждая нить указанной молекулы РНК имеет длину приблизительно от 21 до 23 нуклеотидов; одна нить молекулы РНК содержит нуклеотидную последовательность, обладающую достаточной комплементарностью к РНК-мишени, с тем, чтобы направить расщепление РНК-мишени посредством РНК интерференции; и в которой по меньшей мере одна нить молекулы РНК содержит один или более описанных здесь химически модифицированных нуклеотидов, таких как дезоксинуклеотиды, 2'-O-метил-нуклеотиды, 2'-дезокси-2'-фтор-нуклеотиды, 2'-O-метоксиэтил-нуклеотиды и т.д.
В одном варианте осуществления для изобретения характерно то, что лекарственный препарат содержит молекулу siНК согласно изобретению.
В одном варианте осуществления для изобретения характерно то, что активный ингредиент содержит молекулу siНК согласно изобретению.
В одном варианте осуществления для изобретения характерно то, что двухцепочечная молекула короткой интерферирующей нуклеиновой кислоты (siНК) осуществляет понижающую регуляцию экспрессии гена-мишени, предпочтительно, экспрессию гена в ЦНС, более предпочтительно, гена MAPT, и в котором молекула siНК содержит одну или более химических модификаций, и каждая нить двухцепочечной siНК имеет длину приблизительно от 18 до 28 или более (например, приблизительно 18, 19, 20, 21, 22, 23, 24, 25, 26, 27 или 28 или более) нуклеотидов.
В одном варианте осуществления для изобретения характерно то, что двухцепочечная молекула короткой интерферирующей нуклеиновой кислоты (siНК), которая снижает экспрессию гена-мишени, предпочтительно, экспрессию гена в ЦНС, более предпочтительно, гена MAPT, в которой одна из нитей двухцепочечной молекулы siНК представляет собой антисмысловую нить, которая содержит нуклеотидную последовательность, комплементарную нуклеотидной последовательности РНК-мишени или части ее, а другая нить представляет собой смысловую нить, которая содержит нуклеотидную последовательность, комплементарную нуклеотидной последовательности антисмысловой нити и в которой большинство пиримидиновых нуклеотидов, присутствующих в двухцепочечной молекуле siНК, содержит модификацию сахарного остатка. Предпочтительно, чтобы РНК-мишень или часть ее кодировала белок или часть его. Кроме того, 5'- конец антисмысловой цепи включает в себя фосфатную группу. Нуклеотидная последовательность антисмысловой цепи или ее части может быть комплементарна нуклеотидной последовательности нетранслируемой области РНК-мишени или ее части.
В одном варианте осуществления каждая нить молекулы siНК содержит приблизительно от 18 до 29 или более (например, приблизительно 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28 или 29 или более) нуклеотидов, и каждая нить содержит по меньшей мере приблизительно 18 нуклеотидов, которые комплементарны нуклеотидам другой цепи. В одном варианте осуществления сборка молекулы siНК происходит из двух отдельных олигонуклеотидных фрагментов, где один фрагмент содержит нуклеотидную последовательность антисмысловой нити молекулы siНК, а второй фрагмент содержит нуклеотидную последовательность смыслового участка молекулы siНК. В одном варианте осуществления смысловая цепь соединена с антисмысловой цепью через линкерную молекулу, такую как полинуклеотидный линкер или ненуклеотидный линкер. В дальнейшем варианте осуществления пиримидиновые нуклеотиды, присутствующие в смысловой нити, представляют собой 2'-дезокси-2'-фтор-пиримидиновые нуклеотиды, а пуриновые нуклеотиды, присутствующие в смысловой области, представляют собой 2'-дезокси-пуриновые нуклеотиды. В другом варианте осуществления пиримидиновые нуклеотиды, присутствующие в смысловой нити, представляют собой 2'-дезокси-2'-фтор-пиримидиновые нуклеотиды, а пуриновые нуклеотиды, присутствующие в смысловой области, представляют собой 2'-О-метилпуриновые нуклеотиды. В еще одном варианте осуществления пиримидиновые нуклеотиды, присутствующие в антисмысловой нити, представляют собой 2'-дезокси-2'-фтор-пиримидиновые нуклеотиды, а любые пуриновые нуклеотиды, присутствующие в антисмысловой нити, представляют собой 2'-дезокси-пуриновые нуклеотиды. В другом варианте осуществления антисмысловая нить содержит один или более 2'-дезокси-2'-фтор-пиримидиновый нуклеотид и один или более 2'-О-метил-пуриновый нуклеотид. В другом варианте осуществления пиримидиновый нуклеотид, присутствующий в антисмысловой цепи, представляет собой 2'-дезокси-2'-фторпиримидиновый нуклеотид, и любые пуриновые нуклеотиды, присутствующие в антисмысловой цепи представляют собой 2'-О-метил-пуриновые нуклеотиды. В дальнейшем варианте осуществления смысловая нить содержит 3'-конец и 5'-конец, в которых концевой остаток кэпа (например, инвертированный дезокси-остаток без основания или остаток инвертированного дезоксинуклеотида, такого как инвертированный тимидин) находится на 5'-конце, на 3'-конце или на обоих, 3'- и 5'-концах, смысловой цепи. В другом варианте осуществления антисмысловая нить содержит фосфоротиоатную межнуклеотидную связь на 3'-конце антисмысловой нити. В другом варианте осуществления антисмысловая нить содержит глицериловую модификацию на 3'-конце. В другом варианте осуществления 5'-конец антисмысловой цепи дополнительно содержит фосфатную группу.
В любом из вышеуказанных вариантов осуществления двухцепочечная молекула короткой интерферирующей нуклеиновой кислоты (siНК), которая снижает экспрессию гена-мишени, предпочтительно, экспрессию гена в ЦНС, более предпочтительно, гена MAPT, где большинство пиримидиновых нуклеотидов, присутствующих в двухцепочечной молекуле siНК, содержит модификацию сахарного остатка, и каждая из двух нитей siНК может содержать приблизительно 21 нуклеотид. В одном варианте осуществления приблизительно 21 нуклеотид каждой цепи молекулы siНК спарен основаниями с комплементарными нуклеотидами другой цепи молекулы siНК. В другом варианте осуществления приблизительно 19 нуклеотидов каждой цепи молекулы siНК спарены основаниями с комплементарными нуклеотидами другой цепи молекулы siНК, в которой по меньшей мере два 3'-концевых нуклеотида каждой цепи молекулы siНК не спарены основаниями с нуклеотидами другой цепи молекулы siНК. В другом варианте осуществления каждый из двух 3'-концевых нуклеотидов каждого фрагмента молекулы siНК представляет собой 2'-дезокси-пиримидин, такой как 2'-дезокси-тимидин. В одном варианте осуществления каждая цепь молекулы siНК спарена основаниями с комплементарными нуклеотидами другой цепи молекулы siНК. В одном варианте осуществления приблизительно 19 нуклеотидов антисмысловой цепи спарены основаниями с нуклеотидной последовательностью РНК-мишени или частью ее. В одном варианте осуществления приблизительно 21 нуклеотид антисмысловой цепи спарены основаниями с нуклеотидной последовательностью РНК-мишени или частью ее.
В одном варианте осуществления для изобретения характерно то, что композиция, содержащая молекулу siНК согласно изобретению, представлена в фармацевтически приемлемом носителе или растворителе.
В неограничивающем примере введение химически модифицированных нуклеотидов в молекулы нуклеиновых кислот обеспечивает мощный инструмент для преодоления потенциальных ограничений стабильности in vivo и биодоступности, присущих нативным молекулам РНК, которые доставляются экзогенно. Например, использование химически модифицированных молекул нуклеиновых кислот может дать возможность предоставить желаемый терапевтический эффект при использовании пониженных доз конкретной молекулы нуклеиновой кислоты, поскольку химически модифицированные молекулы нуклеиновый кислоты проявляют тенденцию к более высоким временам полужизни в сыворотке крови. Более того, определенные химические модификации могут улучшить биодоступность молекул нуклеиновых кислот при направлении в конкретные клетки или ткани, и/или улучшить клеточный захват молекулы нуклеиновой кислоты. Следовательно, даже если активность молекулы химически модифицированной нуклеиновой кислоты уменьшена по сравнению с таковой природной молекулы нуклеиновой кислоты, например, когда сравнивают с молекулой, являющейся полностью-РНК нуклеиновой кислотой, то общая активность модифицированной молекулы нуклеиновой кислоты может быть больше чем для таковой природной молекулы вследствие повышенной стабильности и/или доставки молекулы. В отличие от природных немодифицированных siНК, химически модифицированные siНК могут также минимизировать возможность активации интерферона у человека.
В любом варианте осуществления описанных здесь молекул siНК антисмысловая область молекулы siНК согласно изобретению может содержать фосфоротиоатную межнуклеотидную связь на 3'-конце указанной антисмысловой области. В любом варианте осуществления описанных здесь молекул siНК их антисмысловая область может содержать приблизительно от одной до пяти фосфоротиоатных межнуклеотидных связей на 5'-конце указанной антисмысловой области. В любом варианте осуществления описанных здесь молекул siНК их выступающий 3'-концевой нуклеотид согласно изобретению может содержать рибонуклеотиды или дезоксирибонуклеотиды, которые химически модифицированы по сахару нуклеиновой кислоты, основанию или боковой связи. В любом варианте осуществления описанных здесь молекул siНК выступающий 3'-концевой нуклеотид может содержать один или более универсальных оснований рибонуклеотидов. В любом варианте осуществления описанных здесь молекул siНК выступающий 3'-концевой нуклеотид может содержать один или более ациклических нуклеотидов.
В других вариантах осуществления молекулы siНК имеют тупые концы.
В одном варианте осуществления изобретение включает молекулы siНК в 40 или менее нуклеотидов, содержащие любую нуклеотидную последовательность с SEQ ID NO:1-160 на фигуре 8. В определенном варианте осуществления siНК имеют длину 21-30 нуклеотидов и содержат любую из SEQ ID NO:161-318 на фигуре 8.
В одном варианте осуществления изобретение характеризуется способом модуляции экспрессии гена-мишени, предпочтительно экспрессии гена в ЦНС, более предпочтительно, гена MAPT, у субъектов или в организмах, содержащих (a) синтезированную молекулу siНК согласно изобретению, которая может быть химически модифицирована, в которой одна из цепей siНК содержит последовательность, комплементарную РНК гена-мишени; (b) введение молекулы siНК субъекту или в организм в условиях, пригодных для модуляции экспрессии гена-мишени у субъектов или в организмах. Уровень белка-мишени или РНК-мишени можно определить, используя различные методы, хорошо известные в данной области.
В другом варианте осуществления изобретение характеризуется способом модуляции экспрессии более чем одного гена-мишени, предпочтительно, генов, экспрессирующихся в ЦНС, более предпочтительно, включая по меньшей мере один ген MAPT, у субъектов или в организмах, содержащих (a) синтезированные молекулы siНК согласно изобретению, которые могут быть химически модифицированы, в которых одна из цепей siНК содержит последовательность, комплементарную РНК генов-мишеней; (b) введение молекул siНК субъекту или в организм в условиях, пригодных для модуляции экспрессии генов-мишеней у субъектов или в организмах. Уровень белка-мишени или РНК-мишени может быть определен известным в данной области способом.
В одном варианте осуществления изобретение характеризуется способом модуляции экспрессии гена-мишени, предпочтительно, экспрессии гена в ЦНС, более предпочтительно, гена MAPT, внутри клетки, содержащей (a) синтезированную молекулу siНК согласно изобретению, которая может быть химически модифицирована, где siНК содержит одноцепочечную последовательность, обладающую комплементарностью к РНК гена-мишени; (b) введение молекулы siНК в клетку в условиях, пригодных к модуляции экспрессии гена-мишени в клетке.
В другом варианте осуществления изобретение характеризуется способом модуляции экспрессии более чем одного гена-мишени, предпочтительно, экспрессирующихся генов в ЦНС, более предпочтительно, включая по меньшей мере один ген MAPT, внутри клетки, содержащей (a) синтезированные молекулы siНК согласно изобретению, которые могут быть химически модифицированы, где siНК содержит одноцепочечную последовательность, обладающую комплементарностью к РНК гена-мишени; (b) контактирование клетки in vitro или in vivo с молекулой siНК в клетке в условиях, пригодных для модуляции экспрессии генов-мишеней в клетке.
Испытание in vitro дуплексов siРНК.
Чтобы проверить специфичность интерференции siРНК, можно провести предварительное тестирование на культуре клеток, экспрессирующих ген-мишень.
По существу, клетки инкубируют с соответствующими дуплексами siРНК, после чего исследуют уровни экспрессии гена. Чтобы привязать нокдаун с участием siРНК к специфическому фенотипу в культивируемых клетках, необходимо показать уменьшение уровня белка-мишени или по меньшей мере уменьшение мРНК-мишени. Уровни мРНК гена-мишени могут быть подсчитаны методом количественной ПЦР в режиме реального времени (ОТ-ПЦР). Также и уровни белка могут быть определены различными способами, известными в данной области, такими как Вестерн-блот-анализ со специфическими антителами к различным мишеням, позволяющий напрямую контролировать уменьшение уровня белка-мишени.
siРНК вводят в клетки посредством техники трансфекции, хорошо известной в данной области. Отдельная трансфекция дуплексом siРНК может быть выполнена, например, с использованием катионного липида, такого как реагент Lipofectamine 2000 (Invitrogen), после чего следует анализ эффективности сайленсинга через 24, 48 и 72 часа после трансфекции.
Эффективность трансфекции может зависеть от типа клеток, а также от номера пассажа и степени конфлуэнтности клеток. Время и способ образования комплексов siРНК-липосома (например, переворачивание вместо встряхивания) также является критичным. Низкие эффективности трансфекции являются наиболее распространенной причиной неудачного сайленсинга. Хорошая трансфекция является нетривиальным вопросом и требует тщательного рассмотрения для каждой новой используемой линии клеток. Эффективность трансфекции может быть определена путем трансфекции репортерным геном, например EGFP-экспрессирующей плазмидой под CMV-промотором (например, от Clontech), или B-Gal экспрессирующей плазмиды, а затем оценена с помощью фазово-контрастной и/или флуоресцентной микроскопии на следующий день.
В зависимости от обилия и времени жизни (или оборота) белка-мишени фенотип после нокдауна может проявиться от 1 и до 3 дней или даже позже. В тех случаях, когда фенотип не проявляется, уменьшение белка можно наблюдать с помощью иммунофлуоресценции или Вестерн-блотинга.
Фармацевтические композиции и способы применения.
Настоящее изобретение может включать в себя применение одного или более видов молекул siНК одновременно. Эти виды могут быть выбраны для нацеливания на один или более генов-мишеней.
В одном варианте осуществления один тип siНК применяют в терапевтических способах согласно изобретению. В другом варианте осуществления siНК согласно изобретению применяют в комбинации с другой siНК согласно изобретению и/или с одним или более других, не содержащих siНК, терапевтических соединений, пригодных для лечения, предотвращения или управления болезненным состоянием ЦНС. Понятие «в комбинации с» не ограничивается применением терапевтических соединений в точно то же самое время, но предпочтительно означает, что siНК согласно изобретению и другие соединения вводят пациентам последовательно и в течение такого промежутка времени, чтобы польза от введения комбинации была выше, чем польза от их введения иным способом. Например, каждое терапевтическое средство можно вводить одновременно или последовательно в любом порядке в разное время; однако, если их не вводят одновременно, то интервал между их введениями должен быть достаточно малым, таким, чтобы обеспечить желаемый терапевтический эффект. Каждое терапевтическое средство может быть введено отдельно в любой подходящей форме и в любом подходящем курсе.
siНК согласно изобретению могут входить в состав фармацевтических композиций любым из обычных технологических приемов, известных в данной области (см., например, Alfonso, G. et al., 1995, в: The Science and Practice of Pharmacy, Mack Publishing, Easton PA, 19th ed.). Композиции, содержащие одну или более siНК, для использования в способах согласно изобретению, могут находиться в многочисленных формах и могут зависеть от различных факторов, специфичных для каждого пациента (например, типа и тяжести заболевания, типа применяемой siНК, возраста, массы тела, реакции и истории болезни пациента в прошлом), количества и типа siНК в композиции, от формы композиции (например, в жидкой, полужидкой или твердой форме) и/или терапевтического режима (например, от того, применяется ли терапевтическое вещество в виде длительной инфузии, однократного болюса, один раз в день, несколько раз в день или один раз каждые несколько дней).
Молекулы siНК согласно изобретению, а также составы или композиции на их основе могут быть введены непосредственно или местно, как общеизвестно в данной области. Например, молекулы siНК могут содержать средство для доставки, включая липосомы, для применения субъектом. Носители, растворители и их соли могут присутствовать в фармацевтически приемлемых композициях. Молекулы нуклеиновых кислот могут быть доставлены в клетки различными способами, известными специалистам в данной области, включая, но не ограничиваясь ими, заключение в липосомы путем ионтофореза или путем введения в другие средства для доставки, такие как биодеградируемые полимеры, гидрогели, циклодекстриновая поли(лактик-со-гликолевая) кислота (PLGA), PLCA микросферы, биодеградируемые нанокапсулы, биоадгезивные нанокапсулы или белковые векторы. В другом варианте осуществления молекулы нуклеиновых кислот согласно изобретению могут также входить в состав или образовывать комплекс с полиэтиленимином или его производными, такими как полиэтиленимин-полиэтиленгликоль-N-ацетилгалактозаминовым (PEI-PEG-GAL) или полиэтиленимин-полиэтиленгликоль-три-N-ацетилгалактозаминовым (PEI-PEG-triGAL) производными.
Молекулы siНК согласно изобретению могут образовывать комплекс с мембраноразрушаемыми веществами и/или катионным липидом или вспомогательной липидной молекулой.
Системы доставки, которые могут применяться в соответствии с изобретением, включают, например, водные и неводные гели, крема, многочисленные эмульсии, микроэмульсии, липосомы, мази, водные и неводные растворы, лосьоны, аэрозоли, углеводородные основания и порошки и могут содержать инертные носители, такие как солюбилизаторы, усилители проницаемости (например, жирные кислоты, эфиры жирных кислот, жирные спирты и аминокислоты) и гидрофильные полимеры (например, поликарбофильные и поливинилпирролидон). В одном варианте осуществления фармацевтически приемлемым носителем является липосома или трансдермальный усилитель.
Фармацевтическая композиция согласно изобретению представлена в форме, подходящей для применения, например, общего или локального, внутрь клетки или субъекта, включая, например, человека. Выбор подходящих форм отчасти зависит от применения или пути введения, например орального, трансдермального или путем инъекции. Другие факторы известны в данной области, включая такие факторы, как токсичность, и формы, которые предохраняют композицию и состав от производимого ими эффекта.
Настоящее изобретение включает в себя также композиции, готовые для хранения или применения, которые содержат фармацевтически эффективное количество желаемых соединений в фармацевтически подходящем носителе или растворителе. Подходящие для терапевтического использования носители или растворители хорошо известны в фармацевтической области. Например, могут быть использованы консерваторы, стабилизаторы, красители и ароматизаторы. Они включают в себя бензоат натрия, сорбиновую кислоту и эфиры п-гидроксибензойной кислоты. Кроме того, могут быть использованы антиоксиданты и суспендирующие вещества.
Фармацевтической эффективной дозой является та доза, которая необходима для предотвращения, подавления возникновения или лечения (облегчения в некоторой степени признака, предпочтительно, всех признаков) болезненного состояния. Фармацевтическая эффективная доза зависит от типа заболевания, применяемой композиции, способа применения, типа млекопитающих, которых подвергают лечению, физических характеристик конкретных млекопитающих при рассмотрении, параллельного лечения и других факторов, известных специалистам в области медицины.
Композиции согласно изобретению можно вводить в виде дозированных композиций, содержащих общепринятые нетоксичные допустимые носители, вспомогательные средства и/или наполнители. Композиции для орального применения также могут быть представлены в виде твердых желатиновых капсул, в которых активный ингредиент смешан с инертным твердым растворителем, например карбонатом кальция, фосфатом кальция или каолином, или мягких желатиновых капсул, где активный ингредиент смешан с водой или масляным связующим, например арахисовым маслом, жидким парафином или оливковым маслом.
Водные суспензии содержат активные вещества в смеси с наполнителями, применяемыми для производства водных суспензий. Такие наполнители содержат суспендирующие вещества, например натрий-карбоксиметилцеллюлозу, метилцеллюлозу, гидропропилметилцеллюлозу, альгинат натрия, поливинилпирролидон, трагакантовая камедь и гуммиарабик; диспергирующими и увлажняющими веществами могут быть природные фосфатиды, например лецитин или продукты конденсации алкиленоксида с жирными кислотами, например полиоксиэтиленстеарат, или продукты конденсации окиси этилена с алифатическими спиртами с длинной цепью, например гептадекаэтиленоксицетанол, или продукты конденсации окиси этилена с неполными эфирами, полученными из жирных кислот и гексита, такие как полиоксиэтиленсорбитол моноолеат, или продукты конденсации окиси этилена с неполными эфирами, полученными из жирных кислот и ангидридов гексита, например полиэтиленсорбитан моноолеат. Водная суспензия может также содержать один или более консервантов, например этил- или н-пропил-п-гидроксибензоат, один или более красителей, один или более ароматизаторов и один или более подсластителей, таких как сахароза или сахарин.
Масляные суспензии могут быть составлены путем суспендирования активных ингредиентов в растительном масле, например арахисовом масле, оливковом масле, кунжутном масле или кокосовом масле, или в минеральном масле, таком как жидкий парафин. Масляные суспензии могут содержать загустители, например пчелиный воск, твердый парафин или цетиловый спирт. Эти композиции можно хранить с добавлением антиоксидантов, таких как аскорбиновая кислота.
Диспергируемые порошки и гранулы, подходящие для получения водных суспензий путем добавления воды, обеспечивают активный ингредиент в смеси с диспергирующим или увлажняющим веществом, суспендирующим веществом и одним или более консервантами. Подходящие диспергирующие, увлажняющие или суспендирующие вещества указаны выше. Дополнительные наполнители, например подсластители, ароматизаторы и красители, также могут присутствовать.
Фармацевтические композиции согласно изобретению могут также находиться в форме эмульсии масло-в-воде. В качестве масляной фазы может выступать растительное масло или минеральное масло, или их смесь. В качестве подходящих эмульгирующих веществ могут выступать встречающиеся в природе камеди, например гуммиарабик или трагакантовая камедь, встречающиеся в природе фосфатиды, например, из бобов сои, лецитин, эфиры или неполные эфиры, полученные из жирных кислот и гексита, ангидридов, например, сорбитан моноолеат и продукты конденсации указанных неполных эфиров с окисью этилена, например полиоксиэтиленсорбитан моноолеат. Эмульсии могут также содержать подсластители и ароматизаторы.
Такая суспензия может быть составлена известными в данной области способами с использованием подходящих диспергаторов или увлажнителей и суспендирующих веществ, упомянутых выше.
В качестве стерильного лекарственного средства для инъекций может выступать также раствор или суспензия для инъекций в нетоксичном парентерально приемлемом разбавителе или растворителе, например раствор в 1,3-бутандиоле. К приемлемым наполнителям и растворителям, которые могут быть использованы, относятся вода, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, легкие нелетучие масла традиционно используются в качестве растворителей или суспендирующей среды. Для этой цели может быть использовано любое легкое нелетучее масло, включая синтетические моно- и диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, находят применение при получении средств для инъекций.
Молекулы нуклеиновых кислот согласно настоящему изобретению также могут быть введены субъекту в комбинации с другими терапевтическими соединениями для увеличения общего терапевтического эффекта. Использование многочисленных соединений для лечения проявлений может увеличить благотворные эффекты, в то же время уменьшая побочные эффекты.
Альтернативно определенные молекулы siНК могут быть экспрессированы внутри клеток с эукариотических промоторов. Рекомбинантные векторы, способные экспрессировать молекулы siНК, могут быть доставлены в клетки-мишени и могут в них находиться постоянно. Альтернативно могут быть использованы векторы для обеспечения временной экспрессии молекул нуклеиновых кислот. Такие векторы, в случае необходимости, могут быть введены многократно. При экспрессировании молекула siНК взаимодействует с мРНК-мишенью и вызывает РНКi ответ. Доставка экспрессирующих векторов для молекул siНК может быть системной, как, например, при внутривенном или внутримышечном введении, она может осуществляться путем введения клеток-мишеней, эксплантированных из субъекта, с последующим повторным введением в организм субъекта, или любыми другими способами введения в желаемую клетку-мишень.
Интраназальное применение siНК.
Исследования интраназальной доставки siНК были проведены на мышах GFP C57BL/6-TG (ACTB-EGFP). Данную линию трансгенных мышей приобретали у "The Jackson Laboratory". Были использованы трансгенные мыши, так как гомозиготные мыши по этому трансгену погибают в течение 2 недель после рождения. У мышей трансгенной линии с кДНК «усиленного» GFP (EGFP) под контролем куриного промотора бета-актина и цитомегаловирусного энхансера во всех тканях, за исключением эритроцитов и шерсти, наблюдается зеленый цвет флуоресценции при облучении возбуждающим светом. Эта линия была образована на основе мышей линии C57BL/6. кДНК для этой линии, кодирующую усиленный зеленый флуоресцирующий белок (EGFP), присоединяли к промотору куриного бета-глобина и цитомегаловирусному энхансеру. В конструкцию был также введен бычий глобиновый сигнал полиаденилирования. Сайты EcoRI были включены в праймеры для ПЦР, которые были использованы для вставки амплифицированной кДНК EGFP в экспрессионный вектор pCAGGS, содержащий куриный бета-актиновый промотор и цитомегаловирусный энхансер, бета-актиновый интрон и бычий глобиновый сигнал полиаденилирования. Целая вставка с промотором и кодирующей последовательностью была вырезана с помощью Bam-HI и SalI и очищена в геле.
siРНК, использованная для понижающей регуляции экспрессии мРНК EGFP, была нацелена на следующую последовательность в мРНК EGFP (SEQ ID NO:319): 5'-GGC UAC GUC CAG GAG CGC ACC-3'. Смысловая цепь дуплекса siРНК была 5'-P GGC UAC GUC CAG CGC AСС-3', а антисмысловая цепь была 5'-P U GCG CUC CUG GAC GUA GCC UU-3' (SEQ ID NO:320). Дуплекс siРНК, использованный в описанных ниже экспериментах, содержал 2 3'-выступающих тимидиновых нуклеотида.
Протокол эксперимента
Для экспериментов по интраназальной доставки были использованы мыши-самцы C57BL/6-TG (ACTB-EGFP) (в возрасте 8 недель). Мышей, которым интраназально вводили siРНК, разведенную в 0,9% NaCl, сравнивали с контрольными мышами, которым вводили носитель (NaCl 0,9%). Животных анестезировали изофлураном, и по 20 мкл каждого раствора добавляли по каплям в каждую ноздрю.
Разные дозы siРНК против мРНК EGFP (-/+ трансфекция липидом) вводили интраназально в конечном объеме 20 мкл. Контрольных животных обрабатывали одним только носителем. Во всех случаях животных умерщвляли в диапазоне нескольких дней после применения лекарства с целью отработки оптимального времени для интерференции.
Для анализа ткани мышей умерщвляли с помощью CO2, мозг быстро препарировали на ледяной подложке. Одну половину использовали для Вестерн-блот-анализа, вторую в иммуногистохимическом анализе.
Образцы тканей собирали из различных участков мозга и анализированы методами Вестерн-блотинга и ПЦР в режиме реального времени. Экспрессию GFP при различных условиях лечения измеряли с помощью программы Adobe Photoshop. Ингибирование уровней получали после нормирования по отношению к гену бета-актина, который конститутивно экспрессируется в различных тканях.
Экспериментальные условия были размещены как указаны в Таблице 1 (условия были проанализированы в дупликатах или трипликатах). Мышей, интраназально обработанных одной дозой в 530 мкг (40 наномоль) голой siРНК для GFP, были обозначены как мыши 1, 2 и 3 и умерщвлены на 3 и 5 дни после введения siРНК. Другая экспериментальная группа (пронумерованная как 4, 5, 6, 7, 8 и 9) состояла из животных, обработанных двумя дозами по 265 мкг (20 наномолей) стабилизированной siРНК и умерщвленных на 3, 5 и 8 дни (таблица 1).
Образцы тканей были собраны двумя методами: в белковом буфере среды для лизиса и в РНКlater (Ambion). Впоследствии маленькие кусочки тканей были включены в OCT для анализа иммунофлуоресцентного сигнала белка GFP в ткани. Все образцы хранили при -80°C до применения.
Схематическое распределение экспериментальных условий для интраназальной доставки siРНК. Дозы siРНК приведены в таблице
Экстракты для Вестерн-блот-анализа получали путем гомогенизации областей мозга в ледяном экстракционном буфере, состоящем из 20 мМ HEPES, pH 7,4, 100 мМ NaCl, 20 мМ NaF, 1% Triton X-100, 1 мМ ортованадата натрия, 5 мМ EDTA, 1 мкM окадаевой кислоты и ингибиторов протеаз (2 мМ PMSF, 10 мкг/мл апротинин, 10 мкг/мл лейпептин и 10 мкг/мл пепстатин). Образцы гомогенизировали и центрифугировали при 15000 X g в течение 20 мин при 4°C. Белок, содержащийся в супернатанте, определяли по Бредфорду. Тридцать микрограммов общего белка разделяли методом электрофореза в полиакриламидном геле с 10% додецилсульфатом натрия и переносили на нитроцеллюлозные мембраны. В качестве первичных антител использовали антитела против трансгена EGFP (1/1000) (Sigma) и против бета-актина (1/2500) (Sigma). Мембраны инкубировали с антителами при 4°C в течение ночи в 5% обезжиренном сухом молоке. Вторичные козьи антитела против мышиных антител (1/1000; Invitrogen, San Diego, CA) и реагент для детекции ECL (Amersham Biosciences, Arlington Heights, IL) были использованы для иммунодетекции. Уровень белка рассчитывали с помощью денситометрии, значения GFP были нормализованы по отношению к актину для корректировки любого отклонения в количествах нанесенного белка.
Мозг для иммуногистохимии фиксировали в 4% параформальдегиде в буфере Соренсена в течение ночи и криозащищены в растворе 30% сахарозы. Мозг нарезали на тридцатимикрометровые сагиттальные срезы на замораживающем микротоме (Leica, Nussloch, Germany) и собирали в криозащищающий раствор, состоящий из 30% этиленгликоля, 26% глицерина и фосфатного буфера, pH 7,2. Затем срезы мозга анализировали с помощью флуоресцентной микроскопии.
Ткани собирали в RNAlater и хранили при -80°C. RNAlater удаляли перед экстракцией РНК вследствие его плотности. РНК выделяли с помощью реагента Trizol (Invitrogen) в соответствии с протоколом производителя. Обработку ДНКазой проводили перед измерением экспрессии GFP методом количественной ПЦР.
Применение siРНК связано с тем, чтобы определить, имеет ли место доставка siРНК в мозг. Так как целью было определение понижающей регуляции транскрипта гена GFP, то уровни флуоресценции измеряли после применения siРНК. Никаких вторичных эффектов у животных в процессе проведения экспериментальных методов обнаружено не было.
Результаты
Модель доставки in vivo в центральную нервную систему
Пример 1
siРНК использовали для определения подходящих условий для интраназальной доставки siНК в центральную нервную систему (ЦНС). Мышей, обработанных 20 мкл NaCl (0,9%) (контроль) или 20 мкл siРНК в концентрации 1 нмоль/мкл (мышь 1), 2 нмоль/мкл (мышь 2) или 2 нмоль/мкл + липидная трансфекция TransIT-TKO (мышь 3), умерщвляли через 48 ч поле обработки. Никаких вторичных эффектов в процессе проведения эксперимента у животных не наблюдали.
Образцы из различных областей ЦНС (коры головного мозга, гиппокампа, полосатого тела или луковицы), а также из различных тканей (трахеи, легких, носового эпителия, пищевода) экстрагировали и анализировали методом Вестерн-блотинга с антителами, которые специфически распознают GFP, и иммунофлуоресценции, как описано выше. В качестве контроля использовали антитела против бета-актина.
Результаты по уровням ингибирования экспрессии GFP в различных областях ЦНС после нормализации на бета-актин представлены на фигуре 1. Как можно видеть, самый высокий ингибирующий эффект был получен в дозе 2 нмоль/мкл без липидной трансфекции. Кроме того, ингибирование наблюдалось в различных тканях ЦНС, включая кору головного мозга, гиппокамп, полосатое тело и луковицу. Эти результаты были также подтверждены ПЦР в режиме реального времени.
Пример 2
Различные концентрации и времена интраназального введения siРНК использовали на моделях GFP мышей. Мышей, обработанных 20 мкл NaCl (0,9%) (контроль) или 20 мкл siРНК в концентрации 40 нмоль или 20 нмоль, умерщвляли на 3, 5 и 8 дни после обработки. Никаких вторичных эффектов у животных в процессе эксперимента не наблюдали.
Образцы из различных областей ЦНС (коры головного мозга, гиппокампа, полосатого тела, мозжечка, ствола мозга или луковицы) экстрагировали и затем анализировали методами Вестерн-блотинга и иммунофлуоресценции с антителами, которые специфически узнают GFP. В качестве контроля использовали антитела против бета-актина.
Результаты по уровням подавления экспрессии GFP в различных областях ЦНС после нормализации на бета-актин представлены на фигуре 2. Как можно видеть, ингибирующий эффект зависит от области мозга. Максимальный сайленсинг GFP наблюдали в коре головного мозга, гиппокампе, полосатом теле и луковице. Результаты экспериментов Вестерн-блотинга были подтверждены методом количественной ПЦР (фигура 3). На фигуре 3 приведены результаты по понижающей регуляции уровней мРНК GFP в коре головного мозга и полосатом теле, отчетливое уменьшение этих уровней наблюдали у мышей в условиях 8 и 9.
Пример 3. Испытание дуплексов MAPT siРНК in vitro.
Для проверки специфичности siРНК интерференцию MAPT (белок тау, ассоциированный с микротрубочками) анализировали на клеточных культурах, экспрессирующих MAPT. Клетки, использованные для данных экспериментов, представляли собой человеческие клетки MDA-MB-435. Уровни мРНК MAPT анализировали после инкубации с соответствующими дуплексами siРНК. Чтобы “привязать” нокдаун с участием siРНК к специфическому фенотипу в культивируемых клетках, необходимо продемонстрировать уменьшение уровня белка-мишени или по меньшей мере уменьшение уровня мРНК-мишени.
Трансфекция дуплексами siРНК в клеточных культурах
Несколько примеров технологий трансфекции siРНК хорошо известны в данной области. Трансфекция дуплексами siРНК состоит из одиночной трансфекции дуплексом siРНК при использовании катионного липида, такого как Lipofectamine 2000 Reagent (Invitrogen), с последующим анализом сайленсинга через 24, 48 и 72 часа после трансфекции.
Обычно трансфекцию выполняют по следующей методике: одну лунку 6-луночного планшета с человеческими клетками MDA-MB-435 трансфицируют siРНК в конечной концентрации 100 нМ. Следуя инструкциям производителя реагента Lipofectamine 2000, за день до трансфекции высевали до 2-4·105 клеток на лунку в 3 мл соответствующей ростовой среды, содержащей DMEM, 10% сыворотку, антибиотики и глутамин, и инкубировали клетки при нормальных для роста условиях (37°C и 5% CO2). В день трансфекции степень конфлуэнтности клеток составляла 30-50%. 12,5 мкл 20 микромолярного дуплекса siРНК (соответствует конечной концентрации 100 нМ) или 25 мкл 20 мкM дуплекса siРНК (соответствует конечной концентрации 200 нМ) разводили в 250 мкл DMEM и смешивали. Также 6 мкл Lipofectamine 2000 разводили в 250 мкл DMEM и смешивали. После 5 минут инкубации при комнатной температуре разведенный олигомер (дуплекс siРНК) и разведенный Lipofectamine объединяли для образования комплекса во время 20 минутной инкубации при комнатной температуре. Впоследствии по каплям к клеткам с 2 мл свежей среды с низким содержанием антибиотика добавляли комплексы и аккуратно перемешивали, качая планшет назад и вперед, чтобы обеспечить равномерное распределение комплексов для трансфекции. Клетки инкубировали в нормальных для роста условиях, и через день комплексы удаляли и добавляли свежую и полную ростовую среду. Для наблюдения за сайленсингом гена клетки собирали через 24, 48 и 72 ч после трансфекции.
Эффективность трансфекции может зависеть от типа клеток, а также от номера пассажа и степени конфлуэнтности клеток. Время и способ образования комплексов siРНК-липосома (например, переворачивание вместо встряхивания) также являются критическими. Низкие эффективности трансфекции являются наиболее распространенной причиной неудачного сайленсинга. Хорошая трансфекция является нетривиальным вопросом и требует тщательного рассмотрения для каждой новой используемой линии клеток. Эффективность трансфекции может быть определена путем трансфекции репортерным геном, например, EGFP-экспрессирующей плазмиды под CMV-промотором (например, от Clontech), или B-Gal экспрессирующей плазмиды, а затем оценена с помощью фазово-контрастной и/или флуоресцентной микроскопии на следующий день.
В зависимости от обилия и времени жизни (или оборота) белка-мишени, фенотип после нокдауна может проявиться от 1 до 3 дней или даже позже. В тех случаях, когда фенотип не проявляется, уменьшение белка можно обнаружить методом иммунофлуоресценции или Вестерн-блотинга.
После трансфекций суммарные фракции РНК экстрагировали из клеток, обрабатывали ДНКазой I и использовали для обратной транскрипции с помощью случайных праймеров. ПЦР-амплификацию производили со специфической парой праймеров, покрывающих по меньшей мере одно экзон-экзонное соединение, с тем, чтобы контролировать амплификацию пре-мРНК. ОТ/ПЦР не являющейся мишенью мРНК также использовали в качестве контроля. Эффективное уменьшение мРНК все же может не обнаруживаться на белке-мишени, что показывает, что большой запас стабильного белка может существовать в клетке. Альтернативно может быть использован ПЦР в режиме реального времени для более точного тестирования уменьшения или исчезновения мРНК. При методе количественной ПЦР наблюдают появление флуоресценции во время реакции, что служит индикатором продукции ампликонов во время каждого цикла ПЦР. Этот сигнал увеличивается в прямой пропорциональности к количеству ПЦР продукту в реакции. Регистрируя количество испускаемой флуоресценции на каждом цикле, возможно наблюдать реакцию ПЦР во время экспоненциальной фазы, в которой первое значительное увеличение в количестве ПЦР продукта коррелирует с начальным количеством таргетируемой матрицы.
Для подтверждения степени интерференции дифференциально экспрессирующегося гена MAPT в клеточных культурах был проведен qОТ-ПЦР в соответствии с протоколом производителя (Applied Biosystems). Условия реакции были установлены для Applied Biosystems 7300, и была проведена реакция ОТ-ПЦР в один шаг. Объем реакции в 25 мкл, включающий 2X SyBr green, MultiscribeTM обратную транскриптазу 6,25U, ингибитор РНКазы, 50 нМ прямого и обратного праймеров, который смешивали со 100 нг РНК-матрицы. Были сконструированы специфические праймеры для MAPT, и ген 18S анализировали в качестве гена домашнего хозяйства. Прямой праймер имел последовательность AAGAGCCGCCTGCAGACA (SEQ ID NO 321), в то время как обратный праймер имел последовательность GAGCCGATCTTGGACTTGACA (SEQ ID NO 322).
Обратную транскрипцию на начальной стадии проводили в условиях 30' при 48°C. Термические параметры циклического процесса следующие: 95°C в течение 10 мин, 40 циклов при 95°C в течение 15 с и 60°C в течение 1 мин. Кривые диссоциации также анализировали для проверки специфичности амплификации. Значения порогового цикла (Ct) для каждого образца сравнивали с контрольным 24 ч образцом для определения процента понижающей регуляции каждого гена после siРНК-трансфекции.
Для определения специфичности амплифицированного продукта ПЦР проводили анализ кривых плавления продукта. Результирующие кривые плавления позволяют отличить димеры-праймеры и специфичный продукт ПЦР.
In vitro анализы siРНК MAPT
Для того чтобы определить подавление MAPT в качестве мишени, используя технологию РНКi, первый этап экспериментов выполняли на клеточных культурах MDA-MB-435. Эти анализы выполняли в два этапа. Во-первых, конструировали siРНК против мутаций в MAPT, описанных на фигуре 7, и осуществляли трансфекцию. Впоследствии конструировали также и siРНК против MAPT дикого типа, и анализировали понижающую регуляцию после трансфекции. На фигуре 4 показаны репрезентативные результаты экспериментов по количественной ПЦР для некоторых мутаций MAPT, предварительно описанных на фигуре 7. На фигуре 8 показаны последовательности-мишени в MAPT (SEQ ID NO:1-160), против которых были сконструированы siНК. Дуплексы siНК представлены как SEQ ID NO:161-318.
siРНК конструировали против мутаций MAPT P301L (область-мишень представлена в SEQ ID NO:159) и MAPT R406W (область-мишень представлена в SEQ ID NO:160) для снижения уровней анализируемой мРНК MAPT. Значения, приведенные на фигуре 4, представляют собой средний процент siРНК интерференции в отношении экспрессии гена, уровень которой нормализован по контрольным клеткам, а также их стандартные отклонения. По сравнению с контрольными клетками уровень транскрипта МАРТ через 48 ч уменьшался на 20% после обработки siРНК SEQ ID NO:159 (специфичной в отношении MAPT P301L). Однако уменьшение MAPT при трансфекции siРНК SEQ ID NO:160 (специфичной в отношении MAPT R406W) достигало 40% от уровней в контроле.
Во втором цикле экспериментов были сконструированы различные siРНК, направленные на MAPT дикого типа. Вследствие того, что MAPT имеет разные изоформы, было проведено выравнивание последовательности, и были сконструированы siРНК к общей последовательности. Номера доступа ссылочных последовательностей изоформ MAPT перечислены как NM_005910, NM_016834, NM_016835 и NM_016841. Двумя разными siРНК к MAPT дикого типа, соответствующими SEQ ID NO:128 и SEQ ID NO:139, трансфицировали клетки MDA-MB-435. Ниже на фигуре 5 показаны репрезентативные результаты количественного ПЦР-анализа. Значения представляют собой средний процент siРНК интерференции в отношении экспрессии гена, нормализованной по контрольным клеткам, а также их стандартные отклонения. По сравнению с контрольными клетками уровень MAPT транскрипта через 24, 48 или 72 ч значительно уменьшался после специфической обработки siРНК. siРНК, соответствующая SEQ ID NO:128, уменьшала уровень MAPT-транскрипт на 70%, при этом данное уменьшение поддерживалось в течение 48 ч и 72 ч. siРНК SEQ ID NO:129 также вызывала очень хороший уровень понижающей регуляции MAPT, приблизительно до 60% по сравнению с трансфекцией клеток пустым вектором.
Пример 4. Тестирование дуплексов siРНК МАРТ in vivo.
Для доказательства концепции интраназальной доставки на патологической модели осуществляли понижающую регуляцию MAPT, вовлеченного в лобно-височную деменцию, на подходящей трансгенной мышиной модели МАРТ.
Описание линии мышей
Для образования трансгенных мышей MAPT была использована плазмида pSGT42 (Montejo de Garcini et al., 1994), которая кодирует человеческий 4-повтор изоформы тау с двумя N-концевыми экзонами, в качестве матрицы для введения FTDP-17-мутаций G272V и P301L, разделенных с помощью процедуры Quikchange (Stratagene). Тройная мутантная тау-кДНК была впоследствии собрана путем лигирования рестрикционных фрагментов SacII/AseI (содержащего мутацию G272V) и AseI/HindIII (содержащего мутацию P301L) в плазмиду pSGTR406W (Perez, M., Lim, F., Arrasate, M., and Avila, J. (2000). The FTDP-17-linked mutation R406W abolishes the interaction of phosphorylated tau with microtubules. J. Neurochem. 74: 2583-2589), предварительно расщепленную SacII/HindIII, с получением плазмиды pSGTVLW. Мутантная открытая рамка считывания тау была вырезана из pSGTVLW в виде фрагмента BamHI/BglII и лигирована в BamHI-сайт PBKCMV (Stratagene) в прямой ориентации по отношению к CMV-промотору для получения pBKVLW. Фрагмент SalI/XhoI pBKVLW был затем лигирован в XhoI-сайт pTSC21k (Luthi et al., 1997) в прямой ориентации по отношению к thyl-промотору. Секвенированием участка SacII-HindIII подтвердили, что окончательная плазмида pTTVLW кодирует три специфические аминокислотные замены. Последовательности вектора удаляли обработкой NotI, и очищали большой фрагмент, который был впоследствии введен путем пронуклеарной инъекции в одноклеточные CBA эмбрионы 3C57BL/6. Мышей-основателей идентифицировали с помощью ПЦР и скрещивали с мышами дикого типа C57BL/6. Все анализируемые трансгенные мыши являлись гетерозиготами. Мышей размещали по четыре на клетку, предоставляя им пищу и воду в неограниченном количестве, при поддержании управляемой температуры среды и 12/12-часовых циклов света и темноты, с включением света в 07:00 ч. ПЦР-скрининг осуществляли на ДНК из хвоста, используя олигонуклеотиды TT1, 5'-CTCTGCCCTCTGTTCTCTGG-3' (SEQ ID NO:323, в экзоне 2 мышиного гена thyl), TT2, 5'-CCTGTCCCCCAACCCGTACG-3' (SEQ ID NO:324, на 5'-конец человеческой кДНК тау); и THY, 5'-CGCTGATGGCTGGGTTCATG-3' (SEQ ID NO:325, в интрон 2 мышиного гена thyl). Использовали TTl и TT2 для амплификации продукта в 470 п.н., специфичного для трансгена, а не для эндогенной мышиной ДНК, в то время как для внутреннего контроля использовали TT1 и THY для амплификации продукта в 450 п.н., специфичного для мышиной геномной ДНК, но не для трансгена. Трансгены преимущественно экспрессировались в гиппокампе и лобно-височной коре головного мозга.
Протокол эксперимента
Были использованы разные концентрации и времена интраназального применения siРНК на трансгенных мышиных моделях МАРТ. Мышей обрабатывали 20 мкл NaCl (0,9%) (контроль) или 20 мкл siРНК в концентрации 20 нмоль.
Экспериментальные условия соответствовали тому, как описано в таблице 2 (условия анализировали в трех параллельных пробах). Мышей обрабатывали интраназально одной или двумя дозами siРНК против MAPT (SEQ ID NO:160) и умерщвляли в разное время после введения siРНК.
Схематическое распределение экспериментальных условий для интраназальной доставки siРНК. Дозы siРНК показаны в таблице
Образцы из различных областей ЦНС (коры головного мозга, гиппокампа, полосатого тела, мозжечка, ствола мозга или луковицы) экстрагировали и затем анализировали методами Вестерн-блотинга, иммунофлуоресценции и количественной ПЦР. Были использованы антитела, которые специфически распознают мутантный MAPT человека. В качестве контроля использовали антитела против бета-актина. Экспрессию MAPT при различных условиях обработки измеряли с помощью программы Adobe Photoshop. Уровни ингибирования были получены после нормализации по отношению к гену бета-актина, который конститутивно экспрессируется в различных тканях.
Результаты по подавлению уровней экспрессии MAPT в различных областях ЦНС после нормализации на бета-актин представлены на фигуре 6. Как можно видеть, ингибирующий эффект в отношении экспрессии MAPT наблюдался в гиппокампе, в котором трансгенный белок экспрессировался на высоких уровнях. Животные, у которых понижающая регуляция экспрессии гена была наибольшей, соответствовали животным, умерщвленным через 7 дней после инстилляции siРНК. Результаты Вестерн-блот-экспериментов были подтверждены методом количественной ПЦР.
ССЫЛКИ



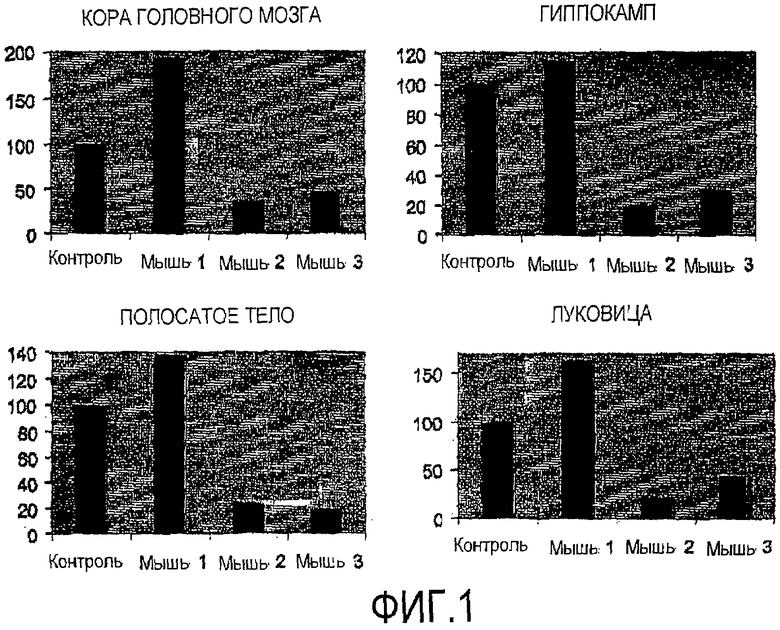
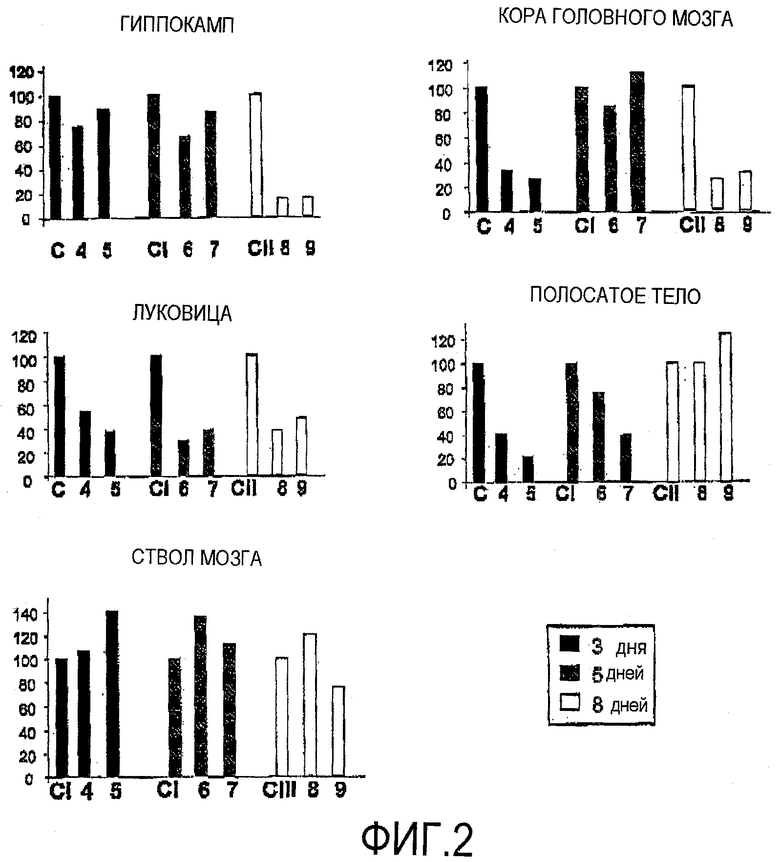

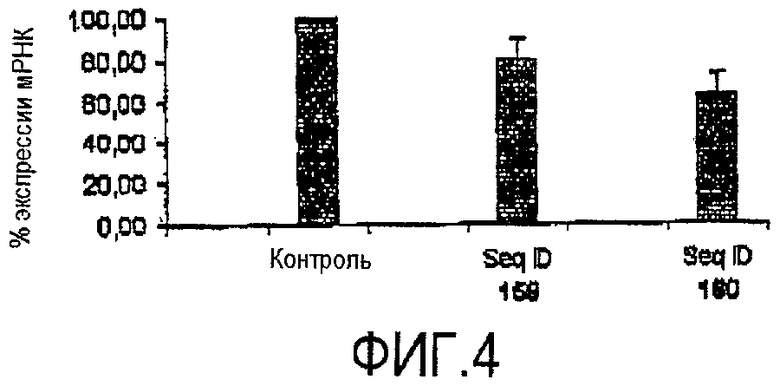
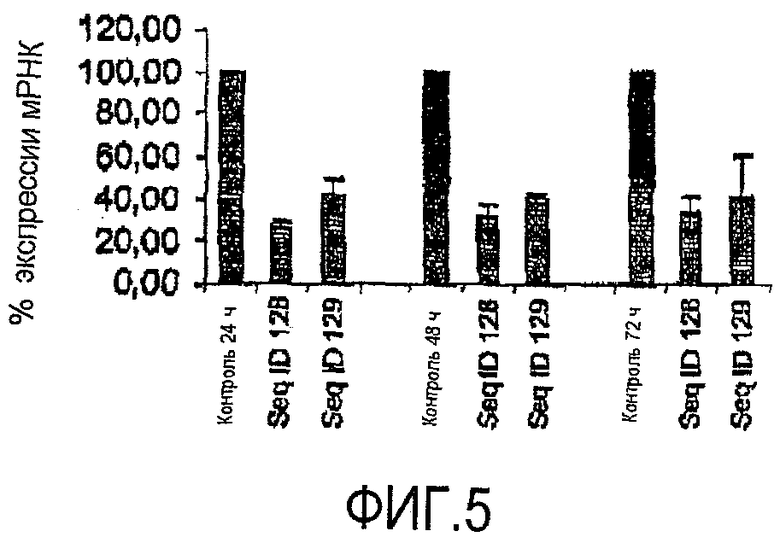
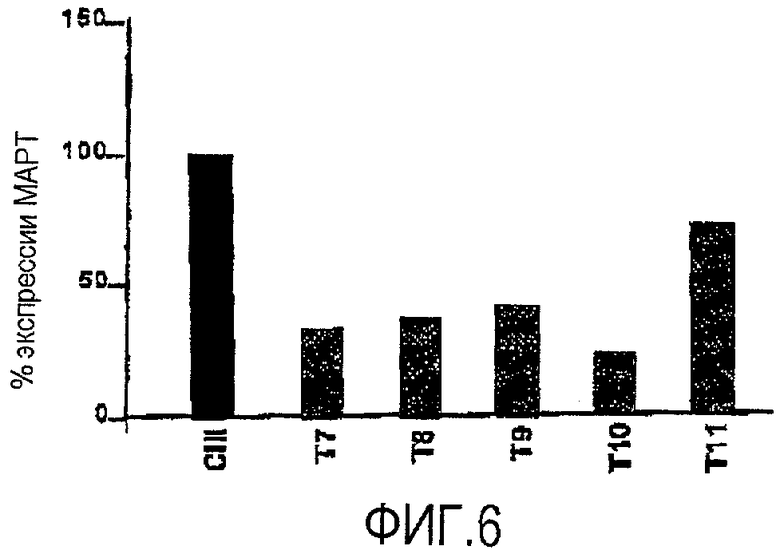
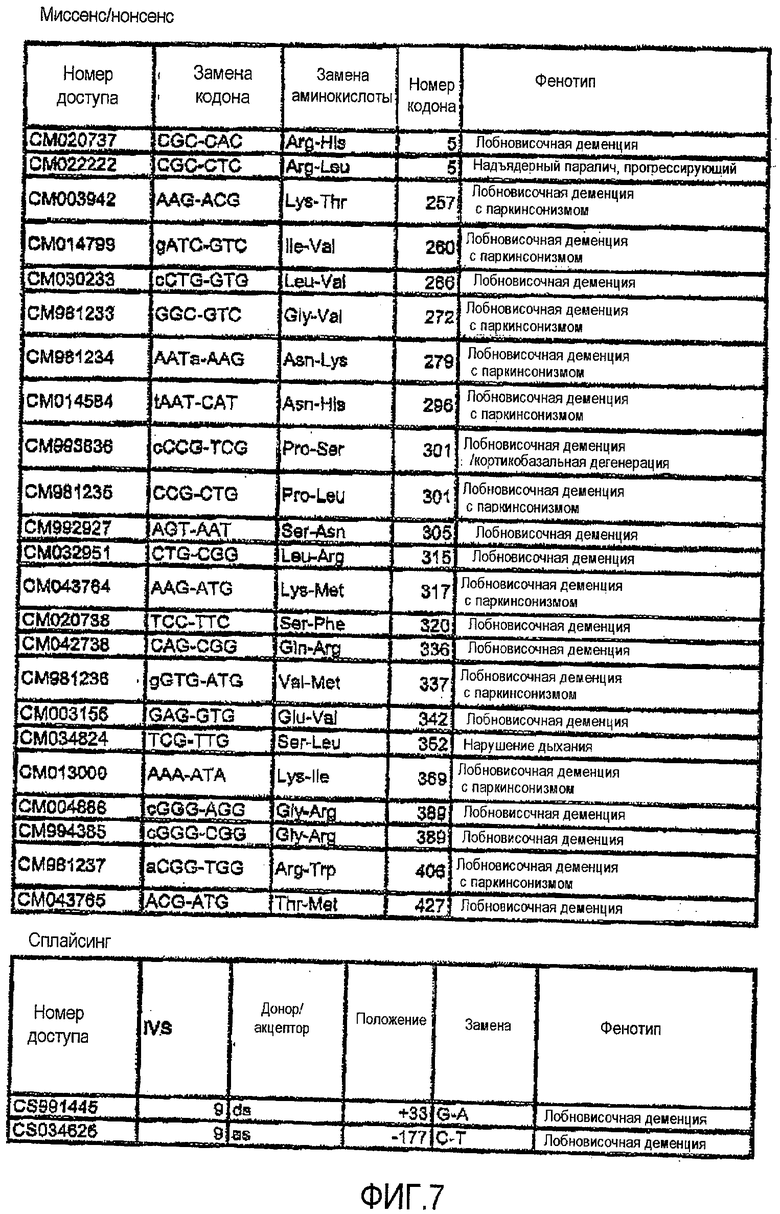
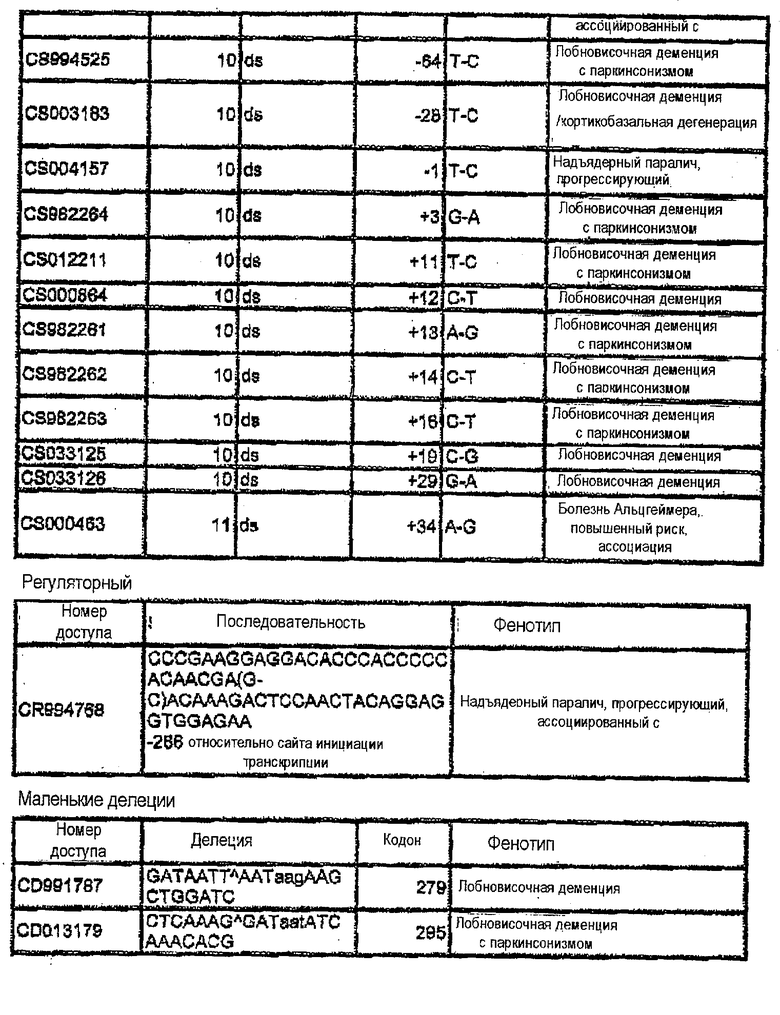


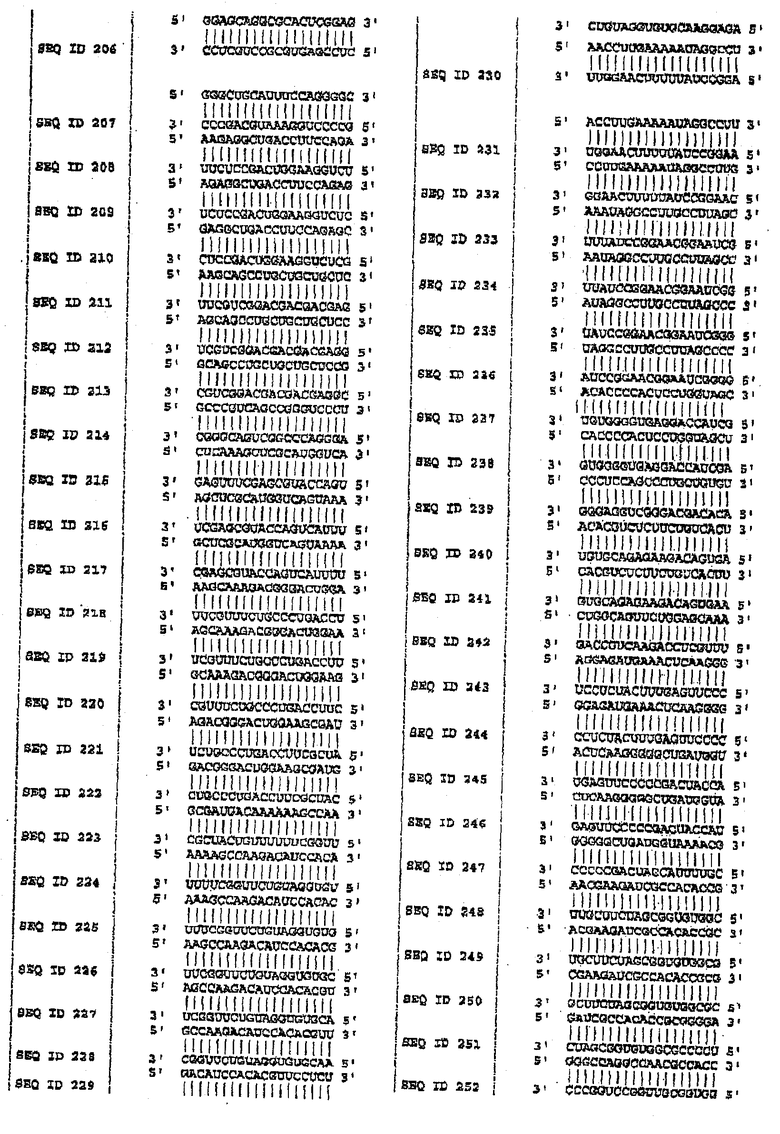

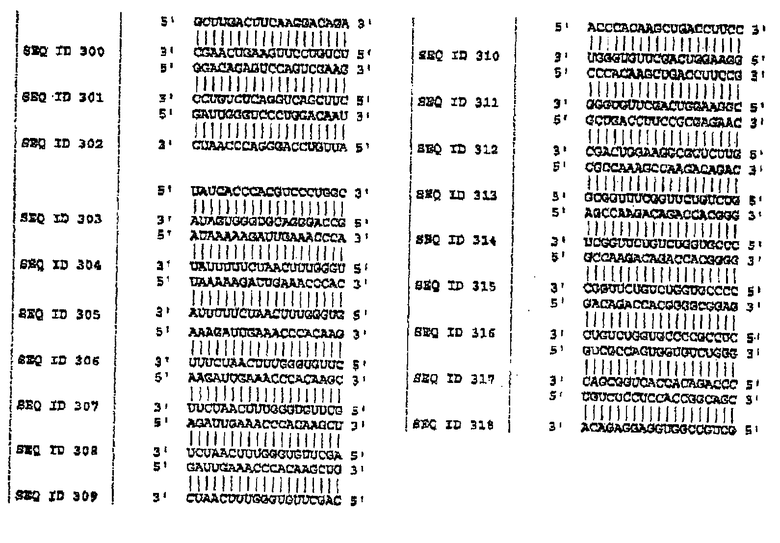
Изобретение относится к области медицины. Настоящее изобретение относится к способу и композиции для лечения нарушения центральной нервной системы (ЦНС) путем интраназального применения соединений, которые вызывают РНК-интерференцию. Композиция согласно изобретению содержит молекулы коротких интерферирующих нуклеиновых кислот (siPHK). При этом нарушение выбрано из деменции, болезни Альцгеймера, болезни Гентингтона и/или болезни Паркинсона, а также врожденных заболеваний, ассоциированных с мутациями в генах ЦНС. Способ включает в себя введение пациенту в случае необходимости эффективного количества одной или более siPHK согласно изобретению. Изобретение обеспечивает подавление экспрессии генов-мишеней, связанных с измененными состояниями ЦНС. 3 н. и 17 з.п. ф-лы, 8 ил., 2 табл.
1. Применение siPHK, которая вызывает РНК-интерференцию в гене, экспрессируемом в центральной нервной системе (ЦНС), для получения лекарственного средства для лечения нарушения ЦНС, где лекарственное средство составлено в форме для интраназального введения.
2. Применение по п.1, согласно которому нарушение выбрано из деменции, болезни Альцгеймера, болезни Гентингтона и/или болезни Паркинсона, а также врожденных заболеваний, ассоциированных с мутациями в генах ЦНС.
3. Применение по п.2, согласно которому ген-мишень выбран из генов тау, гентингтина, ацетилхолинэстеразы или мутантного аллеля этих генов.
4. Применение по п.3, согласно которому экспрессию гена-мишени модулируют в клетке ЦНС.
5. Применение по любому из пп.1-4, согласно которому соединение содержит модифицированный олигонуклеотид.
6. Применение по п.1, согласно которому siPHK имеет в длину 40 или менее пар оснований.
7. Применение по п.5, согласно которому siPHK имеет 3'-выступающие концы.
8. Применение по п.7, согласно которому 3'-выступающие концы являются тимидиновыми динуклеотидами.
9. Применение по любому из пп.1-4 и 6-8, согласно которому используют множество видов siPHK.
10. Применение по п.5, согласно которому используют множество видов siPHK.
11. Применение по п.9, согласно которому указанное множество видов siPHK нацелено на один и тот же вид мРНК или на разные виды мРНК.
12. Применение по п.10, согласно которому указанное множество видов siPHK нацелено на один и тот же вид мРНК или на разные виды мРНК.
13. Применение по любому из пп.1-4, 6-8, 11 и 12, согласно которому геном-мишенью является ген тау, а siPHK нацелена на нуклеотидную последовательность, выбранную из SEQ ID NO:1-160.
14. Применение по п.5, согласно которому геном-мишенью является ген тау, а siPHK нацелена на нуклеотидную последовательность, выбранную из SEQ ID NO:1-160.
15. Применение по п.6, согласно которому геном-мишенью является ген тау, а siPHK нацелена на нуклеотидную последовательность, выбранную из SEQ ID NO:1-160.
16. Применение по п.9, согласно которому геном-мишенью является ген тау, а siPHK нацелена на нуклеотидную последовательность, выбранную из SEQ ID NO:1-160.
17. Применение по п.13, согласно которому siPHK представляет собой последовательность, выбранную из SEQ ID NO:161-318.
18. Фармацевтическая композиция, содержащая одну или множество siPHK, нацеленных на экспрессию гена в ЦНС, составленная в форме для интраназального введения.
19. Композиция по п.18, где указанный ген специфически экспрессируется в ЦНС.
20. Применение siPHK, которая вызывает РНК-интерференцию в гене, экспрессируемом в центральной нервной системе (ЦНС), для получения лекарственного средства для лечения нарушения ЦНС, при этом siPHK вводится пациенту интраназально.
| WO 2005004794 А2, 20.01.2005 | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Аравин А.А и др | |||
| Роль двухцепочечной РНК в подавлении экспрессии генов эукариот | |||
| - Молекулярная Биология, 2002, том 36, №2, с.240-251. | |||

