Область техники, к которой относится изобретение
Изобретение относится к генерации in vitro нейрональных клеток-предшественников, или прогениторных клеток, или нейронов из полипотентных клеток, особенно эмбриональных стволовых (ЭС) клеток.
Предпосылки создания изобретения
Давно установлено, что полипотентные клетки, например, клетки эмбриональной карциномы (КЭК) и эмбриональные стволовые (ЭС) клетки могут быть дифференцированы в нейроны in vitro. В принципе, работа с ЭС клетками позволяет выделять клетки на определенных стадиях дифференциации и описывать нейрональные клетки-предшественники. ЭС клетки позволяют исследовать молекулярные и генетические пути развития in vitro, и, кроме того, являются мощным источником клеток для трансплантации в мозг для лечения неврологических заболеваний.
Однако осуществление этих и других применений затруднено из-за гетерогенности, беспорядочности и часто невоспроизводимости нейронального развития в культуре клеток. Клеточная гетерогенность является серьезной проблемой использования ЭС клеток для получения нервных клеток (см. ссылки 3 и 4 в списке литературы). Обычно культуры нейрональных клеток, сформировавшиеся из ЭС клеток, содержат множество разных нейрональных подтипов, а также клетки другой природы, включая глиальные клетки. Нехватка достаточно большого количества клеток с определенными и одинаковыми фенотипами является серьезной проблемой в нейробиологии. Ранее не было способа получения нейронов из ЭС клеток, в результате которого образовывалось бы стабильное количество нейронов или определенная популяция нейронов, поэтому нельзя было получать популяции гомогенных клеток в достаточных количествах для того, чтобы описать нейроны мозга с помощью биохимических подходов. Кроме того, взаимосвязи линий дифференцировки нейронов с их непосредственными предшественниками оставались неясными.
Для использования нейронов - производных ЭС клеток для трансплантации желательно получить определенные клетки-предшественницы, дающие определенное поколение клеток, а не смесь клеток, содержащую также те клетки, которые могут продолжать делиться и формировать опухоли (ссылки 3, 4). Гетерогенные клетки также могут вмешиваться в трофические и/или направляющие сигналы из ткани хозяина, которые стимулируют интеграцию имплантированной ткани в мозг. Функционально важен тип имплантированных клеток, например, допаминергические нейроны могут особенно быть востребованы для лечения такой болезни, как болезнь Паркинсона, поэтому для подобных медицинских целей желателен повышенный контроль над клетками-предшественниками и подтипами нейрональных клеток. Понижение гетерогенности клеток необходимо для снижения нежелательных побочных эффектов, риска развития опухолей, а также для повышения терапевтического действия за счет увеличения содержания доли клеток, нейрональная дифференциация которых идет в требуемом направлении.
В последнее время было получено важное достижение, заключающееся в использовании индуктивных сигналов и факторов транскрипции для значительного обогащения подтипов нейронов, в том числе допаминергических нейронов и двигательных нейронов. Таким образом, факторы транскрипции, например, Nurr1 (ссылка 5), или совместное культивирование стволовых клеток с клетками других типов (ссылка 6) значительно увеличивают генерацию допаминергических нейронов, хотя добавление несвойственных факторов, включая обработку звуком, усиливает генерацию двигательных нейронов, которые, что также было установлено, интегрируют в ткань хозяина после трансплантации (ссылка 7). Несмотря на эти достижения, все еще очень мало известно о генерации in vitro конкретных предшественников нервных клеток, которые могут повысить специфичность нейрональных фенотипов.
Имеется ряд протоколов, описывающих генерацию нейрональных и глиальных клеток (ссылки 14, 15, 25-31). Ранее на линии клеток эмбриональной карциномы Р19 было установлено, что обработка скоплений плюропотентных клеток ретиноевой кислотой индуцирует нейрональную дифференциацию (ссылка 32). Впоследствии было установлено, что обработка клеток зародышевых тел (ЗТ), являющихся производными ЭС клеток, РК также индуцирует экспрессию нейральных генов и репрессирует экспрессию мезодермальных генов (ссылка 33). ЗТ представляют трехмерные скопления клеток, возникающие за счет агрегации и пролиферации ЭС клеток. ЗТ могут быть получены культивированием ЭС клеток на основе, с которой они не могут срастись, обычно в чашках для культивирования бактерий (см., например, ссылку 41).
Один из способов генерации нейрональных клеток из ЭС клеток, представленный в работах Bain и др. (ссылка 14) и Li и др. (ссылка 10), включает следующие этапы:
культивирования ЭС клеток,
формирования ЗТ,
обработки ЗТ ретиноевой кислотой (РК),
диссоциации ЗТ и
высева и культивирования клеток диссоциированных ЗТ.
Обычно первоначальную культуру ЭС клеток получают на поддерживающем слое питающих клеток (инактивированных фибробластах) для поддержания ЭС клеток в форме колонии плюропотентных недифференцированных ЭС клеток. Предполагают, что фибробласты поддерживают ЭС клетки на недифференцированной стадии. Фактор подавления лейкоза (ФПЛ) может быть внесен в культуральную среду для подавления дифференциации. Однако было установлено, что даже в присутствии ФПЛ некоторые ЭС клетки проявляют тенденцию к дифференциации, и что при формировании ЗТ могут присутствовать клетки разных линий дифференцировки (ссылки 3, 34).
В способах, предложенных Bain и др. и Li и др. (ссылки 10, 14, 15), культивируемые ЭС клетки обрабатывались трипсином и/или растирались до получения небольших скоплений клеток и затем высевались на неадгезивную клеточную культуру для формирования ЗТ. Клетки культивировали на протяжении 4 суток без РК, а затем на протяжении 4 суток на среде с РК, после чего ЗТ диссоциировали и высевали в чашки с покрытием из ламинина. Высеянные клетки культивировали в средах, содержащих сыворотку.
Bain и др. сообщают, что с помощью описанного метода получали культуру, состоящую из популяции плоских клеток, плотно прикрепленных к покрытому ламинином субстрату, и популяции нейроноподобных клеток, преимущественно расположенных поверх плоских клеток. Отмечено, что примерно 38% клеток после двух суток культивирования обладали нейрональной морфологией. Эти культуры клеток имели смешанный состав из нейронов разных типов, главным образом ГАМКергических нейронов.
Некоторые методы основываются на применении селективных маркеров нейрональных клеток-предшественников, элиминируют, таким образом, во время процесса дифференциации клетки, отличные от нейрональных клеток-предшественников. Клетки-предшественники нервных клеток, полученные из ЭС клеток, преимущественно идентифицировались по экспрессии маркеров промежуточных филаментов, например, нестина (ссылка 9), или по факторам транскрипции, например, генам sox (ссылка 10). Li и др. применяли селекцию поколения дифференцирующих клеток для обогащения гетерогенной популяции клетками, экспрессирующими Sox2, за счет элиминации Sox2-отрицательных клеток (ссылка 10). Несмотря на то что селективные методы показали свою эффективность в обогащении популяции клеток нейрональными клетками-предшественниками, они неэффективны в тех случаях, когда отобранные клетки-предшественники нужно применить для генерации определенных нейрональных фенотипов. Из результатов Li и др. следует, что Sox-положительные клетки могут привести к возникновению клеток большинства типов, обнаруженных в центральной нервной системе (ЦНС), а не потомству определенной субпопуляции. Таким образом, хотя селекция Sox-положительных клеток может повысить долю нейрональных клеток-предшественников в популяции клеток, полученной из ЭС клеток, представляется маловероятным, что такая селекция может применяться для повышения содержания в популяции клеток конкретно одного подтипа нейрональных предшественников или нейронов.
В других известных методах РК не применяется. Например, в методах, использовавшихся Okabe и др. (ссылки 27, 43), РК не применяется, но включена промежуточная стадия культивирования сформированных ЗТ на адгезивном субстрате в специальной среде с последующей диссоциацией. Abe и др. (ссылка 30) также применяют промежуточную стадию, после которой выращенные ЗТ переносятся на субстрат, к которому они могут прикрепиться. Затем ЗТ культивируют в адгезивном состоянии, после чего подвергают диссоциации трипсином.
В некоторых способах, например, Abe и др. (ссылка 30) и Okabe и др. (ссылка 27), включено применение основного фактора роста фибробластов. Abe и др. затем применяют митотические ингибиторы, которые вызывают гибель нейронов и клеток, не относящихся к нейрональным (ссылка 30).
Описание изобретения
В настоящем изобретении описаны средства, с помощью которых дифференциация ЭС клеток в нервные клетки может быть оптимизирована с целью получения удивительных преимуществ, выраженных в генерации поколений определенных нервных клеток и в однородности популяций нервных клеток. Таким образом, настоящее изобретение описывает улучшенные способы индуцированного и/или активированного развития и/или дифференциации ЭС клеток в нейроны, или нейрональные клетки-предшественники, или прогениторные клетки для генерации нервных клеток из ЭС клеток in vitro.
В предпочтительных вариантах осуществления настоящего изобретения способы настоящего изобретения позволяют получать в значительной степени однородную популяцию нервных клеток, в которой нервные клетки практически все относятся к одной определенной линии дифференцировки нейрональных клеток, одному фенотипу, типу клеток и/или к одной стадии дифференциации.
В настоящем изобретении подробно описаны разработанные способы получения однородных нейрональных клеток-предшественников, которые были идентифицированы как радиальные глиальные клетки. В ходе последующего культивирования образованные из ЭС клеток радиальные глиальные клетки постепенно дифференцируют в пирамидальные нейроны. Предшественники и нейроны, полученные способами настоящего изобретения, были в основном однородными, причем процентное содержание нейрональных клеток одной линии дифференцировки было выше по сравнению с результатами применения способов, описанных в предшествующем уровне техники.
Таким образом, в более предпочтительных вариантах осуществления настоящего изобретения способы настоящего изобретения позволяют получать в значительной степени однородные популяции радиальных глиальных клеток и пирамидальных нейронов, которые являются одними из наиболее важных популяций нейрональных клеток коры головного мозга и которые было сложно получать из ЭС клеток способами, известными на предшествующем уровне техники.
Поскольку полученные согласно настоящему изобретению клетки-предшественники нервных клеток/прогениторные клетки обладают высоким уровнем однородности, эти клетки пригодны для дальнейшей дифференциации и/или созревания для образования нейрональных клеток определенной линии дифференциации. Предшественники нервных клеток/прогениторные клетки могут быть дифференцированы для получения пирамидальных нейронов, согласно описанному в настоящем изобретении способу, или могут быть изменены внешними или внутренними факторами для генерации популяций других нейрональных клеток.
Преимущество в количестве и однородности клеток, обеспечиваемое настоящим изобретением, отличает эти клетки от тех клеток, которые получают способами нейрогенеза и дифференциации нервных клеток предшествующего уровня техники, а также от ограниченного числа первичных нейронов, которые могут быть приготовлены из мозга мыши или крысы.
Биохимические исследования были ранее затруднены из-за ограниченного числа нервных клеток, которые могут быть получены известными методами предшествующего уровня техники. Настоящее изобретение облегчает исследование биохимических и генетических механизмов, вовлеченных в развитие нервных клеток, особенно в превращение предшественников нервных клеток в нейрональные клетки. ЭС клетки легко поддаются генетическому манипулированию, причем их можно получать в неограниченных количествах, и настоящее изобретение является идеально подходящим способом для получения большого числа нейронов определенных линий дифференциации для биохимического исследования.
Кроме того, поскольку эмбриональными стволовыми (ЭС) клетками можно легко генетически манипулировать или изолировать их из мышей, несущих значимые мутации, настоящее изобретение облегчает проведение сравнения нейронов дикого типа с мутантными нейронами и идентификацию механизмов, вызывающих потерю клеток определенного типа при нейродегенеративных заболеваниях. Хотя генетическая манипуляция ЭС клетками не является сложной, манипуляция с первичными нейронами представляет чрезвычайно трудную задачу, особенно устойчивая манипуляция. С помощью генетических манипуляций с ЭС клетками можно получить гомогенную модифицированную линию, в которой все дочерние клетки содержат одну и ту же мутацию, за 1-2 месяца, а получение линии мышей со стабильной мутацией может занять годы. Таким образом, описанные в настоящем изобретении способы получения клеток-предшественников и нейрональных клеток in vitro из ЭС клеток не требуют получения линий трансгенных мышей и, следовательно, позволяют исследовать мутантные нейроны на уровне, который ранее не был доступен и не применялся на практике.
Способы настоящего изобретения также предусматривают систему изучения клеток, а именно нейронов (например, элонгацию нейритов, гибель нейрональных клеток, нейрогенез и синаптогенез). Такие исследования необходимы в данной области, но возможность их осуществления была ограничена, поскольку нейроны нельзя было доступным способом получать в достаточном количестве. Настоящее изобретение позволят получать нейроны в больших количествах и со значительно большей гомогенностью по сравнению с ранее известными способами, что позволяет проводить нейрональные исследования.
Нейроны и/или нейрональные клетки-предшественники/прогениторные клетки, полученные по способу настоящего изобретения, также пригодны для медицинского применения, например, для имплантации в мозг для лечения нейродегенеративного заболевания или потери нервной ткани. Благодаря повышенной гомогенности нервных клеток требуемого подтипа, получаемых согласно настоящему изобретению, терапевтическая эффективность лечения повышается, а риск развития опухолей вследствие имплантации понижается.
Подробное описание изобретения
Настоящее изобретение относится к способам получения или генерации нервных клеток, например, нейронов и/или нейрональных клеток-предшественников /прогениторных клеток, активирующих или индуцирующих дифференциацию ЭС клеток в нейрональные клетки-предшественники или прогениторные клетки, и к способам активирования или индукции дифференциации или созревания клеток-предшественников или прогениторных клеток в нейроны.
Настоящее изобретение относится к усовершенствованному способу, позволяющему in vitro индуцировать и/или активировать развитие и/или дифференциацию эмбриональных стволовых (ЭС) клеток в нейрональные клетки-предшественники, или прогениторные клетки, или нейроны, и/или получению или генерации нервных клеток, причем указанный способ включает:
культивирование ЭС клеток,
формирование зародышевых тел (ЗТ),
обработку ЗТ ретиноевой кислотой (РК) и
диссоциацию ЗТ
в комбинации с одним или несколькими дополнительными свойствами или стадиями, описанными ниже.
В способах настоящего изобретения клетки диссоциированных ЗТ являются нейрональными клетками-предшественницами или нейрональными прогениторными клетками. Таким образом, диссоциация ЗТ может образовать культуру нейрональных клеток-предшественниц или прогениторных клеток.
Необязательно способ также включает высев в чашки клеток диссоциированных ЗТ, в результате чего получают культуру поверхностного роста нейрональных клеток-предшественниц или прогениторных клеток.
Способ настоящего изобретения может включать культивирование нейрональных клеток-предшественниц или прогениторных клеток для получения нейронов. Таким образом, в способы некоторых вариантов осуществления настоящего изобретения включены посевы в чашки и культивирование клеток диссоциированных ЗТ для получения нейронов.
Способы настоящего изобретения дополнительно включают одно или несколько свойств/этапов, описанных ниже. Любое свойство или стадия может использоваться отдельно или в комбинации с любым другим свойством или стадией, если в контексте не показано иначе.
Культура ЭС клеток без использования питающих клеток
Предпочтительно настоящий способ включает культивирование ЭС клеток в отсутствие питающих клеток (обычно инактивированных фибробластов). Способы могут включать первоначальное культивирование ЭС клеток с питающими клетками с последующим культивированием без питающих клеток. Питающие клетки могут быть разбавлены и удалены повторным пассажем ЭС клеток. Предпочтительно, если проведен, по меньшей мере, один, более предпочтительно, по меньшей мере, два пересева без питающих клеток до формирования зародышевых тел. Таким образом, питающие клетки предпочтительно отсутствуют в культуре ЭС клеток, используемой для формирования ЗТ. «Пассаж» включает диссоциацию клеток и повторный высев ряда клеток. Например, пассаж может включать разъединение/диссоциацию клеток из культуры, выращенной в чашках (обычно с применением трипсина), диссоциацию агрегатов клеток, повторный высев ряда диссоциированных ЭС клеток (адгерентная культура) и культивирование ЭС клеток.
Подходящие культуральные клетки описаны в настоящем изобретении. Фактор подавления лейкоза (ФПЛ) необязательно может быть включен в состав культуральной среды для ЭС клеток.
Отбор и высев ЭС клеток для формирования ЗТ
Было установлено, что состояние пролиферации ЭС клеток влияет на их плюропотентность, и что плотность клеток, культивируемых таким способом, воздействует на их способность и тенденцию к дифференциации. Было установлено, что путем отбора и высева пролиферирующих ЭС клеток при той же густоте, что и в контроле, большее число нейрональных клеток-предшественников, относящихся к определенной линии дифференциации клеток, может быть получено, причем клетки являются в меньшей степени гетерогенными.
Способы настоящего изобретения предпочтительно включают отбор высоко пролиферативных и/или морфологически гомогенных ЭС клеток для формирования ЗТ. Предпочтительно способы включают высев измеренного/оцененного/определенного/установленного числа или плотности указанных ЭС клеток для формирования ЗТ. Предпочтительно настоящий способ включает отбор измеренного, оцененного, определенного или установленного числа ЭК клеток для высева для получения ЗТ.
Настоящий способ предпочтительно включает измерение, оценку, наблюдение или определение:
состояния пролиферации ЭС клеток (которое можно измерить или оценить путем определения времени удвоения клеток, увеличения числа клеток или другим приемлемым способом),
морфологии ЭС клеток и/или
числа или плотности ЭС клеток, высеянных для формирования ЗТ.
Таким образом, предпочтительно высевают клетки подсчитанной, определенной или заданной плотности. Подсчет, оценка или определение числа клеток могут быть проведены каким-либо из известных в данной области способов, например, подсчетом клеток в определенной заданной области с помощью микроскопии, или с помощью обычных счетчиков клеток, например, Casy®1 (фирма Schärfe System GmbH). Морфологию клеток изучают с помощью микроскопии.
Каждый из этих пунктов подробно обсуждается ниже.
Получение и отбор клеток с высокой пролиферативной активностью
В качестве клеток с высокой пролиферативной активностью могут применяться клетки, получаемые определенным способом культивирования, описанным в настоящем изобретении. Было установлено, что состояние пролиферации ЭС клеток может варьировать во время осуществления настоящего способа культивирования ЭС клеток.
Культивирование ЭС клеток или их пассаж предпочтительно приводят к получению клеток с высокой пролиферативной активностью. Предпочтительно пассаж повторяют примерно каждые 2 суток, и культура ЭС клеток предпочтительно включает, по меньшей мере, два пассажа на питающие клетки с последующими, по меньшей мере, двумя пассажами без питающих клеток. ЭС клетки следует снять с питающих клеток, когда они пребывают на стадии высокой пролиферативной активности, например, путем отделения в чашке диаметром 10 см ЭС клеток с питающих клеток и пересевом ЭС клеток на среду без питающих клеток (например, взять 1/4 объема клеточной суспензии и пересеять в новую среду, объем которой равен первоначальному), что должно опять привести к 60% слиянию культуры ЭС клеток уже на следующие сутки. Пассаж без использования питающих клеток может включать примерно 0,5×105 клеток/см2.
Предпочтительно культивирование ЭС клеток включает измерение, оценку или определение числа или плотности ЭС клеток, высеянных для получения клеточной культуры.
ЭС клетки с высокой пролиферативной активностью могут быть ЭС клетками, получаемыми путем культивирования или пересева ЭС клеток, по существу, следующим образом (в норме без питающих клеток):
высев ЭС клеток с плотностью примерно от 0,3×105 до 4×105 клеток/см2, например, примерно 0,5-2×105, предпочтительно примерно 1×105 клеток/см2, и
выделение/диссоциация ЭС клеток через 2 суток после высева, и, необязательно, повторный высев.
ЭС клетки могут быть выделены путем расщепления (диссоциации) через 2 суток после высева. В норме процедуру культивирования (пассаж) следует проводить, по меньшей мере, 2-3 раза до отбора клеток с высокой пролиферативной активностью для формирования ЗТ.
Например, около 2×106 клеток может быть высеяно в чашку для культивирования площадью 10 см2. Описанная выше методика позволяет в норме выделять от 10×106 до 35×106 клеток с площади 10 см2 через 2 суток, например, 10-20×106.
Стадия пролиферации может быть измерена в единицах времени удвоения ЭС клеток. Способы настоящего изобретения могут включать измерение времени удвоения ЭС клеток и выбор клеток с высокой пролиферативной активностью. Например, клетки с высокой пролиферативной активностью могут иметь время удвоения 8 ч или меньше, 16 ч или меньше, или 24 ч или меньше, обычно 8-24 ч.
Морфологические признаки
ЭС клетки, применяемые для формирования ЗТ, предпочтительно являются морфологически гомогенными, причем все или почти все ЭС клетки обладают одинаковыми или схожими морфологическими свойствами.
Предпочтительно способы настоящего изобретения включают отбор морфологически гомогенных ЭС клеток для формирования ЗТ и высев этих клеток для формирования ЗТ. Предпочтительно все или почти все (например, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 98% или, по меньшей мере, 99%) ЭС клетки, выбранные для формирования ЗТ, обладают одним или несколькими, а наиболее предпочтительно всеми, из следующих морфологических признаков (в культуре без питающих клеток): рост в виде плоского монослоя, клетки не находятся в прямом контакте с соседними клетками (но, тем не менее, плотно упакованы), крупные ядра, много ядрышек, клетки не растут поверх друг друга или в виде структур, напоминающих колонии. Предпочтительно клетки плотно упакованы, например, плотность клеток составляет примерно 20×106 клеток на чашку площадью 10 см2 (2×106 клеток/см2), предпочтительно примерно 10-30×106 клеток, например, 15-25×106 клеток на чашку площадью 10 см2. Настоящий способ может включать контроль за одним или несколькими предпочтительными морфологическими свойствами ЭС клеток и/или отбор клеток, обладающих одним или несколькими из этих признаков.
Морфология клеток также может применяться в качестве индикатора стадии пролиферации. Клетки с высокой пролиферативной активностью предпочтительно имеют один или несколько, предпочтительно все, из перечисленных выше морфологических признаков.
Предпочтительно все культивируемые ЭС клетки являются потомством одной ЭС клетки, например, на раннем этапе способа может проводиться отбор одной колонии ЭС клетки и культивирование ЭС клеток из этой колонии. За счет этого в культуре однородность и гомогенность ЭС клеток, в том числе морфологическая, могут быть повышены.
Плотность высева в чашки
Для формирования ЗТ соответствующие проявляющие высокую пролиферативную активность клетки и/или морфологически гомогенные клетки в норме следует высевать, используя примерно 0,5×106 и 5×106 клеток на 15 мл культуральной среды для формирования ЗТ, предпочтительно 2,5-2,5×106 клеток, например, 3×106 клеток в 15 мл среды. В норме следует высевать примерно 0,3-3,5×105 клеток·мл-1, предпочтительно 1,6-2,5×105 клеток·мл-1, наиболее предпочтительно 2×105 клеток·мл-1. Объем среды 15 мл является предпочтительным, хотя 10 мл или 10-15 мл в норме можно использовать для посева в чашки диаметром 10 см.
Плотность клеток, высеянных для формирования ЗТ, следует корректировать в зависимости от стадии пролиферации применяемых ЭС клеток. Следовательно, если культура ЭС клеток является более плотной, будет высеяно больше клеток, если культура менее плотная, будет высеяно меньше клеток. В настоящей работе было установлено, что наилучшие результаты достигаются при использовании наиболее быстро пролиферирующих ЭС клеток.
В качестве примера, ЭС клетки с гомогенной морфологией, имеющие время удвоения примерно 12-16 ч, могут быть выбраны и высеяны с плотностью примерно 0,5×105 клеток/см2.
Диссоциация клеток
В способах настоящего изобретения в основном диссоциация клеток предпочтительно заключается в разъединении клеток (ЭС клеток или ЗТ) для образования суспензии отдельных клеток, главным образом не оставляя агрегатов из более чем 2-3 клеток. Предпочтительно суспензия состоит из целых отдельных диссоциированных клеток (т.е. суспензия не содержит агрегатов клеток). Предпочтительно более 90%, 95%, 98% или 99% клеток в суспензии диссоциировано на отдельные клетки. Предпочтительно менее 5% клеток в суспензии образует агрегаты из 4 или более клеток.
Трипсин (например, 0,05%) и/или растирание могут применяться для диссоциации клеток, используя способы, подробно описанные в настоящем изобретении.
ЭС клетки следует хорошо диссоциировать перед высевом для образования ЗТ. Следовательно, предпочтительно способы настоящего изобретения включают диссоциацию ЭС клеток для формирования суспензии отдельных клеток, практически не содержащую агрегаты более чем из 2-3 клеток. Предпочтительно суспензия является полностью состоящей из отдельных диссоциированных клеток (т.е. суспензия не содержит агрегатов клеток). Предпочтительно более 90%, 95%, 98% или 99% клеток в суспензии диссоциировано на отдельные клетки. Предпочтительно менее 5% клеток в суспензии образует агрегаты из 4 или более клеток.
Способы настоящего изобретения могут включать определение или оценку уровня диссоциации ЭС клеток. Предпочтительно способы включают диссоциацию ЭС клеток и отбор суспензии диссоциированных клеток согласно настоящему изобретению. Микроскопирование или обычные счетчики клеток могут применяться для определения или оценки произошедшей диссоциации. Например, применение счетчика клеток Casy®1 позволяет выявить пики, соответствующие большему диаметру клеток в случае присутствия агрегатов.
Прямая диссоциация клеток ЗТ
Зародышевые тела (ЗТ) выращивают в суспензированной культуре, и затем клетки ЗТ подвергают диссоциации, в результате чего образуется суспензия клеток диссоциированных ЗТ. В норме ЗТ подвергают диссоциации через 8 суток, т.е. на 8-е сутки после высева клеток для образования ЗТ или через 4 суток после добавления РК. Диссоциация может быть выполнена раньше или позже этого срока, но обычно через 3-5 суток после добавления РК. Специалист в данной области может определить экспериментально оптимальное время для диссоциации.
Предпочтительно ЗТ не высевают на адгерентный субстрат перед диссоциацией, а вместо этого поддерживают в неадгерентной культуре до диссоциации клеток. Таким образом, ЗТ предпочтительно следует подвергать диссоциации до посева, а не высевать непосредственно.
Диссоциация ЗТ в норме включает инкубирование ЗТ с трипсином (в норме 0,05% или 0,01-0,5%). Предпочтительно способы настоящего изобретения включают фильтрацию суспензии диссоциированных ЗТ для удаления склеенных клеток, например, клетки могут фильтроваться через сито или фильтр, обычно нейлоновую ячеистую ткань или фильтр. В норме применяется сито или фильтр с ячейками 40 мкм. В вариантах осуществления настоящего изобретения размер пор предпочтительно составляет 20, 30 или 40 мкм, предпочтительно 100, 80, 60 или 50 мкм или менее.
Хранение клеток ЗТ
Способы настоящего изобретения могут включать хранение клеток диссоциированных ЗТ, например, путем замораживания клеток в жидком азоте. Например, хранение может включать центрифугирование клеток, ресуспендирование клеток после центрифугирования в среде для ЗТ+10% ДМСО и замораживание клеток в жидком азоте. Таким образом, в некоторых вариантах осуществления настоящего изобретения способ включает диссоциацию ЗТ и хранение клеток диссоциированных ЗТ. Таким образом, могут быть легко получены подходящие клетки-предшественники нервных клеток.
Замороженные исходные суспензии клеток могут размораживаться по мере необходимости, например, для высева и культивирования для получения нейронов. Ранее в данной области техники не было публикаций о возможности хранения таких клеток-предшественников для последующего использования. В норме клетки размораживают и немедленно после этого ресуспендируют в среде, обычно в 10 мл среды N2, центрифугируют (обычно в течение 5 мин при 1000 об/мин и комнатной температуре) и ресуспендируют (обычно в среде N2).
Плотность высева клеток диссоциированных ЗТ
При высевах клеток ЗТ было установлено, что плотность высеянных клеток ЗТ является важным показателем для выживания и дифференциации клеток. Слишком разреженно высеянные клетки приводят к снижению их выживаемости, а слишком плотные посевы неблагоприятно влияют на скорость дифференциации. Плотность высева также влияет на чистоту культуры, т.е. на соотношение нейрональных клеток и клеток иного типа. Предпочтительно следует высевать примерно от 0,5×105 до 2,5×105 клеток диссоциированных ЗТ на см2, например, примерно 1-2×105, наиболее предпочтительно примерно 1-1,5×105 клеток на см2.
Способы настоящего изобретения могут включать измерение, оценку или определение числа или плотности высеянных клеток ЗТ, используя способы, описанные в настоящем изобретении.
Замена культуральной среды
Было установлено, что выраженное значительное увеличение выживаемости клеток достигается, если культуральную среду заменяют примерно через 2 ч после высева клеток диссоциированных ЗТ. Эти открытия создали возможность получения длительно сохраняющихся нейрональных культур, которые ранее не были распространены в данной области.
В этом контексте замена культуральной среды означает обновление или замещение культуральной среды. Новая среда предпочтительно имеет тот же состав, что и среда, в которую клетки диссоциированных ЗТ были первоначально или предварительно высеяны, т.е. на среды того же типа, что и у применявшейся среды. Может применяться среда, близкая по составу, но предпочтителен состав такой же, что и у использовавшейся ранее среды. Например, в качестве среды может использоваться среда N2.
Таким образом, способы, описанные в настоящем изобретении, включают замену культуральной среды с последующей диссоциацией ЗТ и высевом клеток диссоциированных ЗТ в культуральную среду. Предпочтительно культуральная среда заменяется примерно через 1-6 ч после высева.
Культуральная среда может быть заменена в течение 6 ч после высева, предпочтительно в течение 5, 4, 3 или 2,5 ч после высева. Культуральная среда может быть заменена, по меньшей мере, примерно через 1, 1,5 или 2 ч после посева.
Наиболее предпочтительно культуральную среду заменяют примерно через 1-3 ч после посева, более предпочтительно примерно через 1,5-2,5 ч, и наиболее предпочтительно примерно через 2 ч.
Культура высеянных клеток диссоциированных ЗТ
Клетки диссоцированных ЗТ предпочтительно высевают на среду N2.
Через 2 суток среду предпочтительно заменяют подходящей средой для нейрональной дифференциации, например, «полноценной средой» (см. примеры). Выбор и состав среды может зависеть от того, какую линию дифференциации нейрональных клеток хотят получить. Например, основой полноценной среды, применяемой в настоящем изобретении, является среда Бревера, созданная для активации развития пирамидальных нейронов. Другие среды или факторы могут быть выбраны для поддержания разных линий дифференциации нейрональных клеток, например, Shh (сигнальный белок Sonic hedgehog) для получения холинергических двигательных нейронов.
Было установлено, что клетки-предшественники, полученные согласно настоящему изобретению, могут дифференцировать в ряд разных специфических линий дифференциации нейрональных клеток, включая двигательные нейроны, с последующей имплантацией в куриные эмбрионы.
В некоторых вариантах осуществления настоящего изобретения предпочтительно, чтобы культуральная среда не содержала ТЗ (трийодтиронин). Применяемая в настоящем изобретении культуральная среда основывается на среде Бревера, но ТЗ в состав среды не входит. Возможно, что ТЗ, обнаруженный в фетальной сыворотке теленка (ФСТ), может ингибировать нейрональную дифференциацию.
Предпочтительно среда Neurobasal не применяется. Среда Neurobasal + добавка В27 (оба продукта могут быть приобретены в фирме GIBCO) обычно использовалась в предшествующем уровне техники для нейрональной культуры. Однако установлено, что среда Neurobasal может активировать развитие глиальных клеток предпочтительнее, чем развитие нейрональных клеток. Следовательно, применение среды Neurobasal может привести к нежелательному присутствию глиальных клеток среди полученных нейрональных клеток. И напротив, установлено, что полноценная среда, применяемая в настоящем изобретении, подавляет развитие глиальных клеток и тем самым способствует нейрональному развитию.
Предпочтительно высеянные в чашки клетки (клетки диссоциированных ЗТ, нейрональные клетки-предшественники/прогениторные клетки) культивируются в отсутствие сыворотки. (Сыворотка может применяться для инактивирования трипсина после диссоциации клеток, но затем ее следует удалить, например, центрифугированием для осаждения клеток и почти полным удалением супернатанта).
Предпочтительно ростовые факторы (особенно EGF, FGF/bFGF и PDGF) не содержатся в культуральных средах и клетки-предшественники или прогениторные клетки не культивируют в присутствии этих или других ростовых факторов.
Способы могут включать культивирование нейронов, причем нейроны также предпочтительно не культивируют в присутствии сыворотки и предпочтительно не культивируют в присутствии ростовых факторов, особенно EGF, FGF/bFGF или PDGF.
Кроме того, способы настоящего изобретения не нуждаются в положительном или отрицательном отборе и предпочтительно не содержат эти стадии, например, генетический отбор по Sox-2, для обогащения нейральными клетками или нейронами, хотя при желании такие стадии отбора могут производиться. Способы настоящего изобретения позволяют получать в значительной степени гомогенные популяции нейральных клеток даже без стадии отбора. Предпочтительно способы настоящего изобретения не включают стадию выбраковки типов клеток, не относящихся к нервным или нейрональным (например, делящихся клеток). Предпочтительно способы настоящего изобретения не содержат стадии положительного отбора для обогащения нервными клетками или нейронами. Известные способы отбора включают генетический отбор по Sox-2, т.е. для уменьшения не содержащих Sox-2 клеток, и обработку клеток отрицательным селективным агентом для подавления и/или уничтожения клеток, не являющихся нервными или нейрональными, например, обработку клеток антимитогенным агентом, например, AraC или FRDU, для ингибирования и/или уничтожения делящихся клеток.
Эмбриональные стволовые клетки (ЭС клетки)
Эмбриональные стволовые клетки являются полипотентными стволовыми клетками, выделенными из внутренней клеточной массы бластоцистов млекопитающих. Эмбриональные стволовые клетки, используемые в настоящем изобретении, могут быть взяты у любого млекопитающего, человека или животного, например, морской свинки, крысы, мыши или другого грызуна, кошки, собаки, свиньи, овцы, козы, коровы, лошади или примата, например, обезьяны. Обычно используют ЭС клетки мыши.
В настоящем изобретении ЭС клетки в норме являются плюропотентными клетками, а не тотипотентными клетками, и не могут образовывать зародышевые клетки. ЭС клетки, используемые в примерах настоящего изобретения, являются плюропотентными. Необязательно могут использоваться тотипотентные ЭС клетки.
Ряд линий ЭС клеток, известных в данной области, может быть получен согласно настоящему изобретению (например, J1, Е14).
Могут применяться ЭС клетки, разработанные для того, чтобы избежать стадий отбора, например, отбора по Sox2.
В настоящем изобретении описано, что применяемые ЭС клетки могут быть целевыми клеточными линиями или генетически измененными линиями, содержащими интродуцированный ген, или мутированный ген, или сверхэкспрессируемый эндогенный ген.
Могут применяться линии ЭС клеток, включающих репортерный ген, оперативно связанный с промотором (например, промотором нейрон-специфической экспрессии). В настоящем изобретении описано применение линии клеток Tau-GFP. К свойствам локуса Tau относятся релевантные уровни экспрессии инсертированных молекул сДНК, высокая эффективность рекомбинации, экспрессия только в нейронах, а также то обстоятельство, что удаления Tau не обладают выраженным фенотипом. В настоящем изобретении применяется локус tau для инсерции исследуемой сДНК. Tau может быть легко заменен различными молекулами сДНК, или молекулы сДНК могут быть инсертированы в локус Tau (таким образом, что их экспрессия оперативно связана с промотором Tau) для того, чтобы быстро установить высокий уровень стабильной экспрессии именно в нейронах (ссылка 42).
Нервные клетки
В настоящем изобретении нервной клеткой называется клетка нервной системы, в том числе нервная стволовая клетка, нейрональная клетка-предшественник или прогениторная клетка, и нейрон (нейрональная клетка), если в контексте не указано иначе. Понятия «нейрон» и «нейрональная клетка» взаимозаменяемы.
Понятие «стволовая клетка» означает клетку любого типа, которая может самовозобновляться и, если она полипотентна или является нервной стволовой клеткой, может дать рост клеткам нервной системы всех типов, включая нейроны, астроциты и олигодендроциты. Стволовая клетка может экспрессировать один или несколько из следующих маркеров: Oct-4, Sox1-3, эмбриональных антигенов, специфичных для определенных стадий (SSEA-1, -3 и -4) (Tropepe и др. Neuron, 30, 2001, cc.65-78). Нервная стволовая клетка может экспрессировать один или несколько из следующих маркеров: нестин, нейротрофический рецептор р75, Notch 1, SSEA-1 (Capela и Temple Neuron 35, 2002, cc.865-875).
Понятие «нервная прогениторная клетка» означает дочернюю клетку или клетку последующих делений нервной стволовой клетки с более дифференцированным фенотипом и/или более пониженным потенциалом дифференциации по сравнению со стволовой клеткой. Понятие «клетка-предшественник» означает любую клетку, независимо от того, находится ли она в прямой связи с нейронами в ходе развития, но при определенных условиях внешней среды может быть индуцирована в трансдифференцированный, или заново дифференцированный, или приобретенный нейрональный фенотип.
Понятие «линия дифференциации клеток» означает потомство клеток одного определенного типа клеток. Понятие «сублиния дифференциации клеток» означает подтип определенной линии дифференциации клеток.
Выявление маркеров и идентификация типов клеток
С помощью способов настоящего изобретения предпочтительно образуется популяция клеток, в которой, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90% или, по меньшей мере, 95% нейрональных клеток-предшественников/прогениторных клеток, например, радиальных глиальных клеток, или нейронов, например, пирамидальных нейронов. Способы предпочтительно включают идентификацию, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 98% или, по меньшей мере, 99% клеток, являющихся нейрональными клетками-предшественниками/прогениторными клетками, например, радиальными глиальными клетками, или нейронами, например, пирамидальными нейронами. Способы получения культуры нейрональных клеток согласно настоящему изобретению предпочтительно позволяют получать популяцию клеток, содержащую менее 5% астроцитов, например, менее 4%, 3%, 2% или 1%.
Способы настоящего изобретения, описанные выше, предпочтительно являются способами, достигающими эти пропорции. Настоящее изобретение предусматривает способы достижения, получения или генерации этих пропорций клеток, используя одну или несколько из числа описанных выше стадий и свойств.
Способы настоящего изобретения могут включать идентификацию клеток диссоциированных ЗТ в качестве нейрональных клеток-предшественников или (последующая высеянная культура) в качестве нейронов. Способ может включать определение, наблюдение или подтверждение, что, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или не менее 95% клеток, и идентификацию, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или не менее 99% клеток являются нейрональными клетками-предшественниками/прогениторными клетками, например, радиальными глиальными клетками, или нейронами, например, пирамидальными нейронами. Обычно менее 5% клеток, полученных в культуре нейрональных клеток способами, описанными в настоящем изобретении, являются астроцитами, например, менее 4%, 3%, 2% или 1%.
Линии дифференциации клеток и/или клеточных типов могут быть определены по морфологическим признакам, например, методом микроскопирования. Настоящий способ может включать изучение морфологии нейрональных клеток-предшественников/прогениторных клеток или нейрональных клеток, по меньшей мере, в описанных пропорциях образовавшихся клеток. Нейрональные клетки-предшественники/прогениторные клетки могут быть вытянутой и/или биполярной веретенообразной формы. Линия дифференциации нейрональных клеток может быть определена по морфологическим признакам нейрональных клеток, например, пирамидальные нейроны имеют треугольную форму и разветвленные отростки нейриты, а холинергические нейроны имеют биполярную форму.
Полученные согласно способам настоящего изобретения клетки могут идентифицироваться другим способом или дополнительно идентифицироваться по выявлению маркеров, обычно маркеров поверхности клетки, определяемых с помощью антител. Способ может включать определение наличия одного или нескольких маркеров, чье присутствие показывает, что клетка относится к определенной линии или сублинии клеток, или к определенному типу или подтипу клеток. Специалисту известны маркеры, которые могут быть идентифицированы и применимы в качестве показателя принадлежности к линии дифференциации или типу клеток.
Например, способ может включать выявление маркера Рах6 на поверхности клеток и идентификацию клеток, являющихся нейрональными предшественниками, например, радиальными глиальными клетками. К другим выявляемым маркерам относятся нестин, RC2 и BLBP, которые присутствуют на поверхности радиальных глиальных клеток, и р75, GluR1, синаптофизин, Trks (например, TrkA, TrkB, TrkC) и АРР, которые присутствуют на поверхности определенных нейрональных клеток.
Настоящий способ может включать определение высокого процентного содержания клеток, экспрессирующих маркеры нейрональных клеток-предшественников, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% клеток, и идентификацию, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% нейрональных клеток-предшественников.
Настоящий способ может включать выявление высокого процента клеток, экспрессирующих маркеры нейрональных клеток, например, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90% или, по меньшей мере, 95% клеток, и идентификацию, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 98% или, по меньшей мере, 99% клеток в виде нейронов, предпочтительно нейронов определенных линий дифференциации клеток, например, пирамидальных нейронов или допаминергических нейронов.
Таким образом, способ позволяет получить почти полностью гомогенные популяции нейрональных клеток-предшественников или нейронов. По меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 98% или, по меньшей мере, 99% клеток может быть одного и того же типа/линии дифференциации или подтипа/линии дифференциации, например, нейрональных клеток-предшественников одного и того же типа, например, радиальных глиальных клеток, или нейронов одной и той же линии дифференциации, например, пирамидальных нейронов.
Приводимые в настоящем описании примеры показывают, в какие сроки происходит экспрессия различных маркеров и какие морфологические стадии развития в какие сроки наступают. Способы настоящего изобретения могут включать выявление маркеров и/или наблюдение за определенной морфологией в определенное время после диссоциации ЗТ (как отмечено в примерах), например, нейрональная морфология наблюдается менее чем через двое суток после диссоциации ЗТ и/или проявляется экспрессия рецепторов Trk примерно через 7 суток. Например, в настоящем изобретении показано, что примерно 99% клеток, полученных описанным способом, были радиальными глиальными клетками, что было установлено выявлением экспрессии RC2+ у 99% клеток диссоциированных ЗТ. В настоящем изобретении также показано, что, по меньшей мере, 80% нейронов могут стабильно получаться способами настоящего изобретения, что подтверждается измерением экспрессии или подсчетом vGLUT1 и GFP примерно через 7 суток после диссоциации ЗТ.
Процентное содержание может быть подсчитано в виде процента жизнеспособных клеток или процента клеток, экспрессирующих ядерный маркер, например, после обработкой красителем DAPI или Hoechst.
Формирование ЗТ и их обработка РК
В способах настоящего изобретения ЗТ культивируют и формируют в культуральной среде. Во время формирования ЗТ и культивирования культуральную среду обычно заменяют каждые двое суток.
В норме в способах настоящего изобретения ЗТ культивируют в присутствии РК в течение одних или более суток, обычно двух, трех или предпочтительно четырех суток, или вплоть до пяти, шести, семи или восьми суток. Изначально ЗТ могут культивироваться в отсутствие РК на протяжении одних суток или более, обычно от двух до шести суток, обычно в течение двух, трех или предпочтительно четырех суток, или вплоть до пяти или шести суток до обработки РК. Методика 4 суток -/4 суток + использовалась Bain и др. и Li и др.
Специалист может выбрать подходящую концентрацию РК. Например, концентрация может быть, по меньшей мере, 0,25 мкмолей, по меньшей мере, 0,5 мкмолей, или, по меньшей мере, 1 мкмоль. Концентрация может быть, например, 10 мкмолей или меньше, 7,5 мкмолей или меньше, или 5 мкмолей или меньше. Предпочтительно концентрация составляет от 0,5 до 5 мкмолей включительно. Например, концентрация может быть 1 мкмоль или 5 мкмолей.
Изучение нервных клеток
Другими объектами настоящего изобретения являются способы изучения клеток, выполняемые с нейрональными клетками-предшественниками, или прогениторными клетками, или нейрональными клетками, которые в норме формируются in vitro (эти нейроны не являются первичными), и предпочтительно клетки, образуемые по способу настоящего изобретения. К способам изучения может относиться описанный в настоящем изобретении способ получения нейрональных клеток-предшественников, или прогениторных клеток, или нейронов. Способы настоящего изобретения могут представлять способ настоящего изобретения для нейрональной дифференциации (образования нейрональных клеток-предшественников, или прогениторных клеток, или нейронов), а также дополнительно включать стадии способа изучения клеток, описанные в настоящем изобретении.
Таким образом, указанные выше способы нейральной дифференциации могут применяться в контексте исследований.
Кроме того, поскольку с помощью настоящего изобретения впервые были получены в значительной степени гомогенные культуры/популяции нейрональных клеток-предшественников, или прогениторных, или нейрональных клеток, настоящее изобретение также предусматривает способы изучения, выполняемые с высокой степени гомогенными культурами/популяциями нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток, которые могут быть получены (или не могут быть получены) способами дифференциации нервных клеток настоящего изобретения, но в норме их получают способами in vitro.
Способы изучения настоящего изобретения могут включать обнаружение, подсчет, наблюдение или определение одного или нескольких характеристик нейрональных клеток-предшественников, или прогениторных клеток, или нейронов («нейрональные характеристики»), например, роста нейритов или их элонгацию/дегенерацию, нейрональную форму, гибель нейрональных клеток, нейрогенез, нейрональную дифференциацию, электрическую активность клеток, синаптогенез и/или маркеров нейрональных клеток.
В некоторых вариантах осуществления настоящего изобретения способы изучения настоящего изобретения могут включать описанный способ дифференциации нервных клеток для получения нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток, причем указанный способ также включает культивирование ЭС клеток и/или ЗТ в изучаемом состоянии, а также выявление, подсчет, наблюдение или определение одной или нескольких нейрональных характеристик нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток.
Согласно другим вариантам осуществления настоящего изобретения, предусмотренные в них способы изучения могут включать культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в исследуемом состоянии, а также выявление, подсчет, наблюдение или определение одной или нескольких нейрональных характеристик клеток. Необязательно клетки могут быть получены и/или выращены в соответствии со способами дифференциации нервных клеток, описанными в настоящем изобретении.
Способы изучения могут необязательно включать сопоставление нейрональных свойств в исследуемом состоянии («тест-культуры») с нейрональными свойствами клеток, культивируемых во втором состоянии («контрольной культуры»), необязательно с установленными данными о протекании процесса культивирования клеток второй, контрольной, культуры. Способы могут включать культивирование клеток во втором состоянии.
Таким образом, способы изучения могут включать способ дифференциации нервных клеток, описанный в настоящем изобретении, для получения нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток, включающий культивирование ЭС клеток или ЗТ в первых и во вторых условиях, а также сопоставления одного или нескольких свойств нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток, культивируемых в первых условиях, с аналогичными нейрональными свойствами или свойствами нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток, культивируемых во вторых условиях, соответственно.
Культивирование в тест-культуре, т.е. в первом варианте культивирования, может включать обработку клеток тестируемым соединением или культивирование клеток в присутствии исследуемого соединения, которое может быть добавлено к культуральной среде или включено в ее состав. Во втором варианте культивирования, т.е. при культивировании в контроле, клетки могут выращиваться в отсутствие исследуемого соединения, или не вступают в контакт с исследуемым соединением, или не подвергаются его воздействию.
Исследуемым соединением может быть любая молекула, например, из библиотеки исследуемых соединений. В некоторых вариантах осуществления настоящего изобретения исследуемое соединение является молекулой двунитевой РНК (днРНК) и первый вариант культивирования, т.е. с исследуемым соединением, заключается в обработке ЭС клеток или клеток ЗТ молекулами двунитевых РНК и, в результате такой обработки, в ингибировании гена в клетках по механизму РНК-интерференции (РНКи).
Было установлено, что днРНК для подавления экспрессии гена даже более эффективны, чем отдельно взятые нити смысловой или антисмысловой РНК (Fire А. и др., Nature, 391, 1998). Подавление экспрессии с помощью днРНК является геноспецифичным и его часто называют РНК-интерференцией (РНКи) (см. также Fire в Trends Genet., 15, 1999, cc.358-363, Sharp в Genes Dev., 15, 2001, cc.485-490, Hammond и др. в Nature Rev. Genes, 2, 2001, cc.1110-1119, Tuschl в Chem. Biochem. 2, 2001, cc.239-245).
РНК-интерференция является двухступенчатым процессом. Сначала молекулы днРНК расщепляются внутри клетки с формированием коротких молекул интерферирующих РНК (киРНК), длиной примерно 21-23 нуклеотида с 5'-концевым фосфатом и коротким 3'-выступающим концом (примерно из 2 нуклеотидов). Молекулы киРНК нацеливаются конкретно на соответствующую последовательность иРНК для ее разрушения (Zamore P.D. в Nature Structural Biology, 8, 2001, cc.746-750).
РНКи может быть эффективно индуцирована с помощью химически синтезированных дуплексов киРНК той же структуры с 3'-выступающими концами (Zamore в Cell, 101, 2000, cc.25-33). Установлено, что синтетические дуплексы киРНК специфически подавляют экспрессию эндогенных и чужеродных генов в широком диапазоне линий клеток млекопитающих (Elbashir и др. в Nature, 411, 2001, cc.494-498). Могут применяться дуплексы киРНК, содержащие 20-25 пар оснований, более предпочтительно 21-23 пары оснований, ингибируемой последовательности.
Другим способом киРНК могут быть получены с помощью вектора, in vitro (для выделения и применения) или in vivo.
В других вариантах осуществления настоящего изобретения исследуемым соединением может быть нуклеиновая кислота (ДНК, кДНК или РНК), необязательно кодирующая ген, например, кДНК. Таким образом, исследуемое соединение может быть вектором, кодирующим ген, причем экспозиция клеток нуклеиновой кислотой или вектором приводит к экспрессии гена в клетках. В одном из вариантов осуществления настоящего изобретения вектор может включать последовательность нуклеиновой кислоты, которая согласно настоящему изобретению может быть и смысловой, и антисмысловой ориентации, при этом в случае экспрессии РНК смысловые и антисмысловые фрагменты ассоциируют с формированием двунитевой РНК. Такой РНК, например, может быть длинная двунитевая РНК (например, состоящая более чем из 23 нуклеотидов), которая в результате процессинга в клетке может привести к образованию молекул киРНК для РНКи (см., например, Myers в Nature Biotechnology 21, 2003, cc.324-328).
В другом варианте осуществления настоящего изобретения исследуемое соединение может быть антителом.
Способы изучения также могут идентифицировать соединения или состояния, которые повышают или понижают интересуемое свойство.
Обычно сопоставляют с нейрональными клетками, например, через неделю после высева клеток диссоциированных ЗТ.
Исследуемая и контрольная культуры обычно являются двумя разными культурами, выращиваемыми в одинаковых условиях по всем другим параметрам. Если культивируют вместе с исследуемым соединением, особенно с нуклеиновой кислотой, это культивирование, являющееся культивированием первого типа, может заключаться в обработке клеток (обычно ЭС клеток или клеток диссоцированных ЗТ) исследуемым соединением и затем в культивировании клеток.
Нейрональные свойства (например, рост или удлинение нейритов) могут быть выявлены путем индукции или допущения экспрессии нейрон-специфического репортерного гена, а также обнаружения или количественной оценки экспрессии репортерного гена. Репортерный ген может кодировать флуоресцирующий белок, например, зеленый флуоресцентный белок (GFP). Репортерный ген может быть нацелен или оперативно связан с нейрон-специфическим локусом или промотором, например, tau локусом или промотором для нейрон-специфической экспрессии. Была описана экспрессия нейрон-специфического репортерного гена с локуса tau (Tucker и др., ссылка 42). Экспрессия репортерного гена включается сразу после дифференциации клетки в нейрон, причем только в нейрон, а не в клетки-предшественники или клетки других типов нервной системы. В способах настоящего изобретения, в том числе в исследованиях нейрональных клеток, может использоваться линия клеток (ЭС клеток), содержащих репортерный ген, обладающий нейрон-специфической экспрессией, причем репортерный ген оперативно связан с промотором или локусом, экспрессируемым только в нейронах (например, линия Tau-GFP, описанная в настоящем изобретении).
В настоящем изобретении предусмотрены способы исследования, предназначенные для идентификации агента, который ингибирует или редуцирует рост нейрональной характеристики, вызванный условием, о котором известно, что оно увеличивает эту характеристику или ассоциировано с ростом этой характеристики (например, в некоторых вариантах осуществления настоящего изобретения, согласно которым условие заключается в культивировании с амилоидным бета-пептидом), т.е. идентификации агента, который редуцирует или ингибирует эффекты, ассоциированные с таким условием.
Такое исследование может включать:
культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в присутствии исследуемого агента при условии, о котором известно, что оно повышает или связано с повышением нейронального свойства,
культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в отсутствие исследуемого агента при условии, о котором известно, что оно повышает нейрональное свойство,
измерение или определение уровней нейронального свойства и
сравнение уровней нейронального свойства в присутствии исследуемого агента с уровнями нейронального свойства в отсутствие исследуемого агента,
при этом пониженный уровень нейронального свойства в присутствии исследуемого агента по сравнению с уровнем в отсутствие исследуемого агента показывает, что этот агент ингибирует или редуцирует усиление проявления нейронального свойства, вызванного или ассоциированного с данным условием проведения исследования.
Состояние, о котором известно, что оно повышает нейрональную характеристику или связано с ее повышением, может быть состоянием, выявленным способом исследования настоящего изобретения в качестве состояния, которое увеличивает нейрональную характеристику.
Например, если изучаемое соединение является нуклеиновой кислотой (например, днРНК), культивирование при соблюдении условия, о котором известно, что оно повышает нейрональную характеристику, может включать экспозицию клеток нуклеиновой кислотой и последующее культивирование клеток.
Культивирование с исследуемым агентом и культивирование при соблюдении определенного условия может быть выполнено одновременно или культивирование с исследуемым агентом может быть проведено до культивирования при соблюдении определенного условия, или культивирование при соблюдении определенного условия может быть выполнено до культивирования с исследуемым агентом. Специалист может выбрать подходящую очередность, причем в некоторых вариантах осуществления настоящего изобретения одна очередность может быть предпочтительнее другой. Например, предпочтительно клетки подвергаются воздействию нуклеиновой кислоты и затем их культивируют в присутствии исследуемого агента.
Элонгация или дегенерация нейритов
Способы настоящего изобретения могут включать количественную оценку роста нейритов, удлинения нейритов или дегенерации нейритов. Количественная оценка может включать определение уровней экспрессии нейрит-специфического белка, при этом повышенный уровень экспрессии свидетельствует о повышенном уровне роста нейритов, и/или элонгации нейритов, и/или пониженном уровне дегенерации нейритов, а пониженный уровень экспрессии свидетельствует о пониженном уровне роста нейритов, и/или элонгации нейритов, и/или повышенном уровне дегенерации нейритов. Количественная оценка может включать индукцию или допущение экспрессии нейрон-специфического репортерного гена и измерение уровней экспрессии репортерного гена, тем самым оценивается количественно рост нейритов, элонгация нейритов или дегенерация нейритов. Например, если репортерный ген кодирует флуоресцирующий белок, например, GFP, измерение уровней экспрессии заключается в измерении флуоресценции. Способы настоящего изобретения могут включать количественную оценку роста нейритов, элонгации нейритов или дегенерации нейритов путем обработки нейронов антителом к маркеру нейрита (например, к тубулину, нейрофиламенту, синаптофизину), определяя или подсчитывая связывание антитела с маркером и, таким образом, определяя или подсчитывая формирование выростов или элонгацию нейритов.
Для обработки нейронов антителом могут быть использованы клеточные экстракты, полученные после лизиса клеток (например, при Вестерн-блоттинге). Согласно другому варианту, целые нейроны могут контактировать с антителом. Способы исследования могут включать культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток при соблюдении первого или второго условия, соответственно, и сравнение уровней роста нейритов, элонгации или дегенерации нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток при соблюдении первого условия с уровнем роста нейритов, элонгацией или дегенерацией нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток при соблюдении второго условия, соответственно. Например, если уровни роста нейритов, элонгации или дегенерации выше (например, по данным повышенного/пониженного уровня экспрессии нейрит-специфического белка, см. выше) в клетках, культивируемых при соблюдении первого условия, по сравнению с клетками, культивируемыми при соблюдении второго условия, следует вывод о том, что при соблюдении первого условия (по сравнению со вторым) увеличивается рост, элонгация или дегенерация нейритов, соответственно.
В предпочтительном варианте осуществления настоящего изобретения культивирование при соблюдении первого условия заключается в культивировании клеток в присутствии исследуемого соединения, предпочтительно являющегося амилоидным β пептидом (Аβ) (который образуется из предшественника амилоидного белка, АРР).
Настоящее изобретение предусматривает способы изучения, позволяющие идентифицировать агент, который ингибирует или редуцирует рост дегенерации нейритов, возникающий под воздействием состояния, о котором известно, что оно увеличивает дегенерацию нейритов (например, при культивировании в присутствии амилоидного бета-пептида), т.е. идентифицировать агент, который редуцирует или ингибирует эффекты, связанные с таким состоянием. Такое изучение может включать:
культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в присутствии исследуемого агента при соблюдении условия, о котором известно, что оно повышает дегенерацию нейритов,
культивирование нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в отсутствие исследуемого агента при соблюдении условия, о котором известно, что оно повышает дегенерацию нейритов,
подсчет или определение уровней дегенерации нейритов в присутствии исследуемого агента, или в его отсутствие, и
сравнение уровней дегенерации нейритов в присутствии исследуемого агента с уровнями дегенерации нейритов в отсутствие исследуемого агента,
при этом пониженный уровень дегенерации нейритов в присутствии исследуемого агента по сравнению с уровнем дегенерации нейритов в отсутствие исследуемого агента показывает, что этот агент ингибирует или редуцирует усиление дегенерации нейритов, вызванной или ассоциированной с данным условием проведения исследования.
Согласно указанному выше, сравнение уровней дегенерации нейритов может включать сравнение уровней экспрессии нейрит-специфического белка, при этом повышенный уровень экспрессии (пониженный уровень деградации) в присутствии исследуемого агента по сравнению с уровнем экспрессии в отсутствие исследуемого агента показывает, что исследуемый агент ингибирует или редуцирует усиление дегенерации нейритов, вызванной данным условием проведения исследования.
Условие проведения исследования может заключаться в присутствии соединения, которое может быть соединением, выявленным с помощью способа исследования настоящего изобретения благодаря присущей ему способности усиливать дегенерацию нейритов, а именно Аβ-пептидом.
Гибель нейрональных клеток
Существует потребность исследований гибели нейрональных клеток в данной области и такие исследования описаны в настоящем изобретении.
Исследования гибели нейрональных клеток могут использоваться для определения или оценки чувствительности популяции нейронов или нейрональных клеток к заданному условию, например, к присутствию одного или нескольких соединений, например, для идентификации условия (например, соединения), которое увеличивает или снижает гибель нейрональных клеток.
Например, исследование согласно настоящему изобретению может включать:
культивирование нейронов при соблюдении первого условия («изучаемая культура»),
культивирование нейронов при соблюдении второго условия («контрольная культура»),
количественная оценка или определение гибели нейрональных клеток при соблюдении первого и второго условий и
сравнение уровней гибели нейрональных клеток при соблюдении первого условия с уровнями гибели нейрональных клеток при соблюдении второго условия,
при этом повышенный уровень гибели нейрональных клеток при соблюдении первого условия по сравнению с уровнем гибели нейрональных клеток при соблюдении второго условия свидетельствует, что при соблюдении первого условия гибель клеток увеличивается, и/или
пониженный уровень гибели нейрональных клеток при соблюдении первого условия по сравнению с уровнем гибели нейрональных клеток при соблюдении второго условия свидетельствует, что при соблюдении первого условия гибель клеток уменьшается.
При изучении гибели нейрональных клеток, особенно при идентификации состояния, которое снижает гибель нейрональных клеток, нейроны предпочтительно генетически предрасположены к апоптозу. Например, нейроны могут экспрессировать рецептор нейротрофина р75 и/или экспрессировать апоптический белок (например, каспазу), оперативно связанный с нейрон-специфическим промотором (например, локусом Tau). Таким образом, ЭС клетки, применяемые в настоящем изобретении для получения нейронов для изучения гибели нейрональных клеток, могут экспрессировать апоптический белок (например, каспазу), оперативно связанный с нейрон-специфическим промотором (например, локусом Tau).
Изучение гибели нейрональных клеток может применяться для идентификации агента, который ингибирует или редуцирует повышение гибели нейрональных клеток, получаемое при соблюдении определенного условия, о котором известно, что оно усиливает гибель нейрональных клеток, т.е. агента, который редуцирует или ингибирует влияние такого условия. Такое исследование может включать:
культивирование нейронов в присутствии исследуемого агента при соблюдении условия, о котором известно, что оно увеличивает гибель нейрональных клеток,
культивирование нейронов в отсутствие исследуемого агента при соблюдении условия, о котором известно, что оно увеличивает гибель нейрональных клеток,
количественная оценка или определение уровней гибели нейрональных клеток в присутствии исследуемого агента, или в его отсутствие, и
сравнение уровней гибели нейрональных клеток в присутствии исследуемого агента с уровнями гибели нейрональных клеток в отсутствие исследуемого агента,
при этом пониженный уровень гибели нейрональных клеток в присутствии исследуемого агента по сравнению с уровнем гибели нейрональных клеток в отсутствие исследуемого агента показывает, что данный агент ингибирует или редуцирует усиление гибели нейрональных клеток, вызванное этим условием.
Гибель клеток может быть определена способами, известными в данной области техники, например, путем определения механизмов индукции апоптоза в нейронах. К признакам гибели клеток, которые могут быть определены, относятся индукция апоптических белков (например, каспаз, особенно каспазы-3, см. ссылку 43), окраска иодидом пропидия и/или фрагментация ДНК, и/или разрушение нуклеосом (выявляемое, например, связыванием антитела с ДНК /или гистоном, см. ссылку 44),
Нейрогенез и нейрональная дифференциация
Способы настоящего изобретения могут включать исследования нейрогенеза или нейрональной дифференциации, причем получение или генерация нейронов или дифференциация ЭС клеток, и/или нейрональных клеток-предшественников, или прогениторных клеток выявляют и/или оценивают количественно. Способ может включать выявление и/или количественную оценку одного или нескольких нейрон-специфических маркеров. Способы настоящего изобретения могут включать мониторинг уровней нейрогенеза для одного или нескольких определенных нейрональных подтипов или линий дифференциации клеток, или уровни нейронов в целом, в зависимости от выбранных маркеров. Поколения нейронов определенных линий дифференциации клеток могут изучаться путем выявления и/или количественного определения маркеров, специфичных для линий дифференциации клеток. Способы могут включать обработку клеток антителом к клеточному маркеру и определение связывания, причем наличие маркера (и связывание антитела, соответственно) показывает, что клетки относятся к определенному типу, подтипу, линии дифференциации клеток или суб-линии дифференциации клеток. Способы могут включать определение или подсчет уровней связывания антитела и, таким образом, определение или подсчет уровней дифференциации, стадии дифференциации клеток и/или процента клеток определенного типа, подтипа, линии дифференциации клеток или суб-линии дифференциации клеток, или определенной стадии дифференциации. Более подробно выявление маркеров идентификации типов клеток представлены в настоящем изобретении, а подходящие маркеры известны специалистам в данной области.
Способы изучения нейрональной дифференциации согласно настоящему изобретению пригодны для определения маркеров, которые могут использоваться для идентификации ЭС клеток и/или нервных клеток на определенных стадиях дифференциации, или для идентификации типа или подтипа клеток и, таким образом, для установления стадии дифференциации клетки, или типа клетки, или подтипа. Например, способы изучения могут включать индукцию или допуск дифференциации ЭС клеток для получения нейрональных клеток-предшественников, или прогениторных клеток, и/или культивирования нейрональных клеток-предшественников или прогениторных клеток для получения нейронов (предпочтительно используя способы нервной дифференциации, описанные в настоящем изобретении), сравнения уровней экспрессии белков в клетках на одной стадии дифференциации с уровнями экспрессии белков в клетках на второй стадии дифференциации, и идентификации белков, уровень экспрессии которых отличается в клетках на первой и второй стадиях дифференциации. Различие в уровнях экспрессии показывает, что белок может использоваться в качестве маркера для идентификации стадии дифференциации, типа или подтипа клетки и/или для различия клеток на первой и второй стадиях дифференциации. Уровни экспрессии могут быть сопоставлены с помощью подходящего способа, который может выбрать специалист в данной области. Предпочтительно сопоставляется экспрессия белков на поверхности клетки, например, путем изучения взаимодействия клеток или клеточных экстрактов с библиотекой экспрессируемых на поверхности клеток антител и определения связывания. Например, способ может включать сопоставление экспрессии белков в нейрональных клетках-предшественниках/прогениторных клетках (например, радиальных глиальных клетках) с экспрессией белков в ЭС клетках. Различие в уровнях экспрессии может быть, по меньшей мере, в 1,2 раза, по меньшей мере, в 1,5 раза, по меньшей мере, в 1,6 раза, по меньшей мере, в 1,8 раза, по меньшей мере, в 2 раза, по меньшей мере, в 3 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз или более. Экспрессия может быть обнаружена в клетках на первой стадии дифференциации и совсем не обнаруживается в клетках на второй стадии дифференциации.
Электрическая активность
Уровни электрической активности в нейронах, например, электрической активности, свидетельствующей об открытии специфических каналов (например, ионных каналов), могут наблюдаться, выявляться, определяться или подсчитываться.
Способы исследования могут применяться для идентификации соединения, способного модулировать электрическую активность нейронов. Способы могут включать культивирование нейронов при соблюдении первого условия, культивирование нейронов при соблюдении второго условия и сравнение электрической активности нейронов, культивируемых при соблюдении первого условия, с электрической активностью нейронов, культивируемых при соблюдении второго условия, соответственно. Различие в электрической активности показывает, что условие культивирования модулирует электрическую активность.
Синаптогенез
Способы изучения могут включать выявление или количественную оценку синаптогенеза в нейрональных клетках. Выявление или количественная оценка могут включать измерение электрофизиологической активности клеток и/или выявление или измерение экспрессии одного или нескольких маркеров синаптогенеза, например, синаптофизина.
Сопоставление генетически различных нейронов
В настоящем изобретении описаны способы сравнения стандарта (обычно клетки дикого типа) нейрональных клеток-предшественников, или прогениторных клеток, или нейронов с мутантами нейрональных клеток-предшественников, или прогениторных клеток, или нейронов, нейронов с другими генотипами. Способ настоящего изобретения может включать описанный выше способ получения нервных клеток.
Таким образом, настоящее изобретение описывает способ, включающий:
получение первой и второй культур нейрональных клеток, или нейрональных клеток-предшественников, или прогениторных клеток, которые в первой культуре имеют генотип, отличающийся от генотипа клеток во второй культуре, и
сравнение нейрональных клеток-предшественников, или прогениторных клеток, или нейронов в первой культуре с нейрональными клетками-предшественниками, или прогениторными клетками, или нейронами во второй культуре.
Могут сравниваться нейрональные клетки-предшественники, или прогениторные клетки, или нейрональные клетки, содержащие или не содержащие мутацию в гене интереса. Мутация может быть, например, делецией во всех или в части генов, делецией во всех или в части генных промоторов и/или энхансеров, или заменой одного или нескольких нуклеотидов в кодирующей области промотора или энхансера. В норме мутация приводит к изменению (понижению или повышению) уровня экспрессии гена или экспрессии мутированного белка (например, к укорочению или содержанию одной или более делеций в аминокислотной последовательности). В другом варианте нейрональные клетки-предшественники, или прогениторные клетки, или нейрональные клетки в первой культуре могут обладать интродуцированным геном (например, инсертированным геном или инсертированной кДНК) или сверхэкспрессией эндогенного гена, но нейрональные клетки-предшественники, или прогениторные клетки, или нейрональные клетки во второй культуре ими не обладают.
ЭС клетки могут быть генетически изменены, мутации могут быть индуцированы в ЭС клетки или ЭС клетки могут быть выделены из животного, клетки которого несут мутацию, например, ЭС клетки мыши, несущие мутацию в интересующем гене, для получения нервных клеток, например, нейронов или нейрональных прогениторных клеток/клеток-предшественников, обладающих или не обладающих интересующей мутацией, соответственно. Таким образом, с помощью настоящего изобретения могут быть получены мутантные нервные клетки и нервные клетки дикого типа, например, нейроны или нейрональные прогениторные клетки/клетки-предшественники. Сравнение нервных клеток, полученных из ЭС клеток разных типов (из которых одни несут интересующую мутацию, а другие нет), может, например, быть выполнено для идентификации механизма, отвечающего за потерю типа нервных клеток при нейродегенеративном заболевании, или влияющего на эту потерю, и для идентификации соответствующих целей, связанных с фенотипами заболеваний.
В некоторых вариантах осуществления настоящего изобретения способ может включать получение нейрональных клеток-предшественников/прогениторных клеток или нейронов из первой и второй культур ЭС клеток, соответственно, причем ЭС клетки в первой и второй культурах имеют разные генотипы. Необязательно нейрональные клетки-предшественники/прогениторные клетки или нейроны могут быть получены из ЭС клеток способами, описанными в настоящем изобретении. ЭС клетки в первой культуре могут содержать мутацию интересующего гена, а ЭС клетки во второй культуре не содержат этой мутации (например, клетки дикого типа). В другом варианте ЭС клетки в первой культуре могут содержать интродуцированный ген или сверхэкспрессировать эндогенный ген, а ЭС клетки во второй культуре - нет.
В другом варианте для применения генетически различных ЭС клеток генетическим манипуляциям могут подвергаться ЭС клетки. Способы могут включать трансфекцию первой культуры клеток диссоциированных ЗТ, или нейрональных клеток-предшественников, или прогениторных клеток конструкцией нуклеиновой кислоты и, в результате, изменение генотипа клеток в первой культуре по сравнению с клетками во второй культуре. Например, конструкция нуклеиновой кислоты может кодировать эндогенный ген, или кодировать интересующий ген, содержащий мутацию. Такие способы настоящего изобретения в норме допускают экспрессию (обычно транзиторную экспрессию, длящуюся примерно 2, 3 или 4 суток) с конструкции нуклеиновой кислоты. Настоящий способ может включать культивирование клеток для получения нейрональных клеток. Клетки в первой культуре следует сравнить с нейрональными клетками-предшественниками, прогениторными клетками или нейрональными клетками во второй культуре, причем клетки во второй культуре не содержат конструкцию нуклеиновой кислоты, интродуцированный ген и/или мутацию.
Сравнение нейрональных клеток-предшественников, или прогениторных клеток, или нейронов может заключаться в сравнении (обычно в определении или подсчете) одной или нескольких характеристик, например, роста нейритов, или элонгации нейритов, нейрональной формы, гибели нейрональных клеток, нейрогенеза, нейрональной дифференциации, электрической активности, синаптогенеза и/или маркеров нейрональных клеток. В других вариантах осуществления настоящего изобретения сравнение может заключаться в считывании интересующего гена, например, интродуцированного, или мутированного, или сверхэкспрессируемого гена, или в проявлениях этого гена. Природа считывания зависит от гена и может быть определена специалистом для данного гена. Таким образом, механизмы передачи нейронального сигнала могут быть выяснены, заблокированы и/или подвергнуты воздействию.
Сравнение нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток может включать сравнение одной или нескольких характеристик клеток при определенном изучаемом условии, причем могут применяться способы сравнения генетически различных нейрональных клеток-предшественников, или прогениторных клеток, или нейрональных клеток в контексте способов исследования, описанных в настоящем изобретении. Таким образом, в предпочтительных вариантах осуществления настоящего изобретения и первая, и вторая культура клеток культивируется при исследуемом условии, а нейрональные характеристики клеток сравниваются. Другие способы и вариации способов исследования клеток описаны выше. Например, культивирование при исследуемом условии может заключаться в культивировании присутствия Аβ-пептида.
Антитела
В контексте настоящего изобретения понятие «антитело» или «антитела» означает любое вещество, способное специфически связываться, или вещества, имеющие связывающий домен с требуемой специфичностью. Таким образом, данное понятие относится к фрагментам антител, производным, функциональным эквивалентам и гомологам антител, включая любые полипептиды, содержащие иммуноглобулин-связывающий домен, природный или синтетический. К этому понятию также относятся химерные молекулы, включающие иммуноглобулин-связывающий домен, или эквивалент, связанный с другим полипептидом. Клонирование и экспрессия химерных антител описаны в заявках ЕР-А-0120694 и ЕР-А-0125023.
Было показано, что фрагменты целого антитела могут выполнять функцию связывающих антигенов. Примерами связывающих фрагментов являются (i) фрагмент Fab, состоящий из доменов VL, VH, CL и СН1; (ii) фрагмент Fd, состоящий из доменов VH и СН1; (iii) фрагмент Fv, состоящий из доменов V1 и VH одного антитела; (iv) фрагмент dAb (Ward, E.S. и др., Nature 341, 1989, cc.544-546), состоящий из домена VH; (v) выделенные области CDR; (vi) F(ab')2 фрагменты, бивалентный фрагмент, включающий два связанных фрагмента Fab, и (vii) одноцепочечные молекулы Fv (scFv), в которых домен VH и домен VL связаны пептидным линкером, позволяющим двум доменам ассоциировать с формированием антиген-связывающего сайта (Bird и др., Science, 242, 1988, cc.423-426; Huston и др., PNAS USA, 85, 1988, cc.5879-5883; (viii) биспецифические одноцепочечные димеры Fv (PCT/US92/09965) и (ix) «димеры», поливалентные или полиспецифические фрагменты, сконструированные слиянием генов (WO94/13804; Р Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, cc.6444-6448).
Димеры являются мультимерами полипептидов, и каждый полипептид, включающий первый домен, содержащий область связывания легкой цепи иммуноглобулина, и второй домен, включающий область связывания тяжелой цепи иммуноглобулина, причем два домена связаны (например, пептидным линкером), но не могут ассоциировать друг с другом для образования антиген-связывающего сайта: антиген-связывающие сайты формируются ассоциацией первого домена полипептида в составе мультимера со вторым доменом другого полипептида в составе мультимера (WO 94/13804).
Антитела могут быть модифицированы рядом способов, например, на них может быть нанесена метка, например, флуоресцентным красителем, позволяя количественно оценивать связанные антитела путем измерения уровней флуоресценции.
Различные дополнительные объекты и варианты осуществления настоящего изобретения, описанные в настоящем изобретении, понятны специалистам в данной области.
Все документы, упоминаемые в настоящем описании, включены в него в виде ссылок на их сущность.
Определенные объекты и варианты осуществления настоящего изобретения ниже иллюстрируются примером и ссылкой на чертеж, на котором показаны проценты Рах-6-положительных клеток в определенное время после высева клеток диссоциированных ЗТ. Сначала Рах-6 экспрессируется большинством клеток, но затем экспрессия быстро исчезает. Результаты представляют среднее значение по 4 экспериментам, выполненным с двумя разными линиями ЭС клеток. Они выражены в виде процента ± среднеквадратичное отклонение от числа DAPI-положительных ядер, принятых за 100%.
Примеры
Культура ЭС клеток
Способ, приводящий к генерации нейронов из ЭС клеток, включает следующие стадии, суммируемые следующим образом:
1. Клетки, выращенные на питающем слое, растут в виде колоний, хотя после депривации питающего слоя они растут в виде плоского монослоя клеток.
2. ЭС клетки в неадгерентных бактериологических чашках формируют клеточные агрегаты (зародышевые тела, ЗТ), которые растут в суспензии.
3. Через 4 суток, во время которых формируются ЗТ, добавляют РК на последующие 4 суток.
5. ЗТ подвергают диссоциации в общей сложности через 8 суток и высевают в чашки с покрытием из ПДЛ/ламинина в среду с N2.
6. Среду N2 заменяют через 2 ч и еще раз через 12-24 ч. На этой стадии большинство клеток-предшественников имеет веретеновидную форму. Среду для нейрональной дифференциации добавляют через 30-48 ч.
Эта методика была разработана с использованием ЭС клеток, которые экспрессируют GFP с локуса tau (ссылка 13). Экспрессия GFP с эндогенного промотора позволяет визуализировать нейроны и их отростки в УФ свете, что было использовано для максимизации генерации флуоресцирующих клеток.
После оттаивания ЭС клетки сначала культивируют на питающих клетках на протяжении 2-3 пассажей и затем постепенно убирают питающие клетки. Определенные количества (3×106) клеток затем используют для формирования агрегатов (зародышевых тел, ЗТ), инкубируемых в неадгезивных бактериологических чашках (чашка диаметром 10 см, 15 мл среды) в течение 8 суток. Ретиноевую кислоту (РК, 5 мкмолей) вносят через 4 суток и оставляют на последующие 4 суток. Важным этапом является отбор ЭС клеток без питающих клеток, обладающих гомогенной плоской морфологией и высокой пролиферативной активностью (см. материалы и методы).
Через 8 суток ЗТ подвергают диссоциации с помощью свежеприготовленной суспензии трипсина и высевают на субстрат, содержащий поли-D-лизин (ПДЛ) и ламинин. Установлено, что плотность высева клеток (1,5×105 клеток/см2) является критической, поскольку при меньших плотностях клетки склонны к быстрой гибели. Диссоциированные клетки высевают на среду без сыворотки, которую заменяют через 2 ч после высева для удаления фрагментов клеток и мертвых клеток. Через сутки (примерно через 24 ч) среду заменяют еще раз. Через 48 ч среду заменяют средой без сыворотки, обогащенной добавками (ссылка 12). Помимо ЭС клеток, экспрессирующих GFP с обеих tau аллелей, также применяют более 7 других линий ЭС клеток с результатами, которые не отличаются от результатов, описанных в настоящем исследовании. К ним относятся ЭС клетки дикого типа J1 и Е14, а также J1 с GFP в одной или обеих tau аллелях. Также были выделены 4 разные линии ЭС клеток из бластоцистов смешанной BL6/SV129 генетической среды и использованы в протоколе дифференциации со схожими результатами.
Нейрональные клетки-предшественницы
ЭС клетки дифференцируют в гомогенную популяцию радиальных глиальных клеток.
Клетки, дифференцированные из ЗТ, принимают четкую удлиненную веретеновидную форму, напоминающую форму радиальных глиальных клеток (см. ссылку 16). Фазово-контрастная микроскопия иллюстрирует биполярную веретеновидную форму через 2 ч процесса дифференциации.
Эти клетки идентифицируют в качестве нейрональных клеток-предшественников окрашиванием антителом к промежуточному филаментному белку нестину (ссылка 9). Через 2 ч после высева установлено, что в подавляющем большинстве клетки являются нестин-положительными, что следует из сравнительного количественного анализа общего числа высеянных клеток, после окраски ядер (см. таблицу).
Затем применяют RC2 - маркер, который экспрессируют все радиальные глиальные клетки, и практически все клетки являются позитивными (см. таблицу). Окрашивание антителами к белку, связывающемуся с липидами мозга (BLBP), - антигеном, который также экспрессируется радиальными глиальными клетками в развивающейся ЦНС (ссылка 18), дополнительно подтверждает идентичность клеток, свежеполученных в результате диссоциации из ЗТ (см. таблицу). Гомеодоменный фактор транскрипции Рах-6 экспрессируют все кортикальные радиальные клетки (ссылка 19), и установлено, что почти все клетки в ЗТ экспрессируют его перед диссоциацией, следовательно, на этой стадии они уже являются клетками-предшественниками.
Подсчет через 2 ч после высева показывает, что в подавляющем большинстве клетки все еще позитивны для Рах-6 (см. чертеж), и что его экспрессия быстро снижается за последующие сутки до полного исчезновения через 7 суток (см. чертеж).
Нестин, RC2, BLBP, Рах-6 анализируют методом иммуноцитохимии через 2 ч после высева диссоциированных ЗТ. Процент (± среднеквадратичное отклонение) позитивных клеток определяют от общего числа клеток, окрашенных ядерным маркером DAPI.
Нейрональная дифференциация
Клетки с признаками нейрональной морфологии начинают появляться не позднее, чем через 2 суток после диссоциации ЗТ. Все дифференцирующиеся клетки экспрессируют GFP, что означает их принадлежность к нейронам. Этот вывод подтверждают эксперименты по окрашиванию, в которых используют антитело, распознающее нейрон-специфическую форму тубулина.
Через 4 суток примерно 85% клеток являются GFP-позитивными и тубулин-позитивными. С помощью фазово-контрастной и флуоресцентной микроскопии было показано возникновение замечательно гомогенных нейрональных клеточных тел. Со временем в культуре они все больше и больше принимают пирамидальную форму, отмечаемую для клеток, выделенных из гиппокампа грызунов (ссылка 20). После окраски антителами к синаптофизину наблюдают процессы выстраивания многочисленных кластеров GFP-позитивных клеток, что свидетельствует о возможности возникновения синаптического контакта в этих культурах.
Для того чтобы установить, используют ли эти нейроны глутамат в качестве нейротрансмиттера, клетки окрашивают с антителом к везикулярному транспортеру глутамина (vGlut1) - мембранному белку, экспрессируемому большинством пирамидальных нейронов в коре головного мозга и в гиппокампе (ссылка 21). Через 7 суток 93±4,7% клеток культуры окрашивают антителами к vGlut1. Результаты, полученные с применением антител vGlut1, подтверждают идентификацию нейронов в качестве пирамидальных клеток. К концу первой недели после диссоциации ЗТ менее 0,1% клеток окрашивается позитивно в отношении Is1-1, трозингидроксилазы и холинацетилтрансферазы. Через 3 недели менее 5% клеток являются ГАМК-положительными.
Для идентификации белков, экспрессируемых во время перехода от радиальных глиальных клеток к нейронам, проводят Вестерн-блоттинг, используя дифференцированные in vitro нейроны, полученные в разные интервалы времени.
Хотя АМРА-рецептор субъединицы GluR1, например, синаптофизин, не выявляют в лизатах радиальных глиальных клеток, после нескольких суток роста их четко обнаруживают в культуре. Уровни белков GluR1 и синаптофизина увеличиваются после начала дифференциации нейронов.
Поскольку пирамидальные нейроны экспрессируют высокие уровни рецепторов Trk и в коре головного мозга, и в гиппокампе (ссылка 22), также анализируют их экспрессию, используя антисыворотку к внутриклеточному домену этих нейротрофиновых рецепторов. Хотя рецепторы Trk с трудом выявляются на 5 сутки, их уровень резко возрастает в последующие сутки. Значительную экспрессию рецепторов Trk наблюдают примерно через 7 суток in vitro, после чего она резко возрастает. И, напротив, установили, что уровни нейротрофического рецептора р75 падают во время нейронального созревания, что во многом сходно с процессом in vivo (ссылка 23).
В заключение исследуют экспрессию белка - амилоидного предшественника (АРР). Установили, что этот мембранный белок экспрессируют радиальные глиальные клетки (ссылка 24), а также ряд клеток, включая нейроны на более поздней стадии развития. Установлено, что, в отличие от других исследованных мембранных белков, АРР четко выявляют в лизатах радиальных глиальных клеток. Уровни экспрессии позднее возрастают, по-видимому, вследствие созревания нейронов, которое включает выраженный рост нейрональных процессов.
Дифференциация in vivo имплантированных клеток-предшественниц
Потенциал развития нейрональных клеток-предшественников согласно настоящему изобретению исследуют путем имплантации этих клеток в куриные эмбрионы, в которых они могут дифференцировать в различные специфические линии дифференциации нейрональных клеток, в том числе в двигательные нейроны.
Электрофизиология
Электрофизиологические эксперименты показывают, что нейроны формируют синапсы, демонстрируют потенциалы действия и являются весьма однородными по электрофизиологическим характеристикам. Эти нейроны преимущественно глутаматергические (что показано блокированием синаптической передачи NBQX, vGAT окрашиванием) с некоторым ГАМКергическим входом (блокируемым бикукуллином, иначе культура не может выжить). С помощью электрофизиологии четко установлено, что при данных условиях отсутствуют другие типы нейрональных клеток.
Для описания электрофизиологических свойств полученных из ЭК клеток нейронов проводят распознавание пэтч-клампов на поверхности отдельных клеток, находящихся в культуре на протяжении от 10 до 22 суток. Все исследованные клетки (n=22) показывают спонтанные или индуцированные деполяризацией потенциалы действия, и во всех случаях они могут быть блокированы применением тетродотоксина. Кроме того, электрофизиологические характеристики исследованных клеток показывают, что они в высшей степени гомогенны по функциональным свойствам, схожим с ранее описанными свойствами пирамидальных нейронов. Могут наблюдаться спонтанные синаптические передачи (ССП), которые могут быть полностью блокированы добавлением NBQX/AP-5 и бикукуллина, или только одного NBQX/AP-5. Эти результаты показывают, что полученные из ЭС клеток нейроны формируют функциональные синапсы, которые потребляют глутамат в качестве нейротрансмиттера. Поскольку настоящие эксперименты также выявляют присутствие функциональных ГАМК-синапсов в длительно поддерживаемых культурах, подсчитывают количество ГАМК-нейронов через 3 недели. Одновременно с утратой окрашивания для нейротрансмиттеров, не связанных с глутаматом и ГАМК-системой, не обнаруживают какой-либо синаптической активности, которая не была бы связана с глутаматом или ГАМК.
Обсуждение
Применение мышиных ЭС клеток позволило установить условия, которые приводят к генерации, по существу, чистых популяций нейрональных клеток-предшественников, которые называют радиальными глиальными клетками. Затем эти клетки продолжают развиваться в гомогенные популяции нейронов со свойствами пирамидальных клеток.
Было установлено, что если высокопролиферативные недифференцированные стволовые клетки отбирают для трансформации ЗТ, обработка РК преобразует целые популяции клеток в нейрональные клетки-предшественники определенного типа. Отбор недифференцированных ЭС клеток является важным, поскольку было установлено, что даже в присутствии LIF некоторые ЭС клетки проявляют тенденцию к дифференциации, и что в ходе формирования ЗТ часто наблюдают клетки разных линий дифференциации (см. обзоры, ссылки 3, 34).
Для отбора ЭС клеток с высокой пролиферативной активностью после удаления питающих клеток проводят мониторинг скорости деления путем подсчета клеток с применением фазово-контрастной микроскопии, а также определения достигнутой степени слияния клеток до начала формирования ЗТ с определенным числом клеток.
Ранее сообщалось о присутствии радиальных глиальных клеток в недиссоциированных ЗТ, установленном с применением либо ЭС клеток, либо клеток эмбриональной карциномы Р19 (ссылка 35). Если ЗТ высевают на полилизиновый субстрат, можно наблюдать радиальную миграцию удлиненных клеток по направлению от ЗТ с последующей трансформацией в астроциты (ссылка 35). Идентификацию клеток осуществляют путем диссоциации ЗТ, основываясь на морфологии клеток и их количестве после окрашивания антителами к RC2, BLBP и Рах-6. Ранее было установлено, что этот набор маркеров экспрессируют радиальные глиальные клетки в коре головного мозга (ссылки 11, 16). Известно, что не все радиальные глиальные клетки экспрессируют Рах-6. В частности клетки, локализованные в ганглионарном бугорке, не экспрессируют Рах-6 и не являются нейрогенными (ссылка 11). Ранее было сделано интересное наблюдение, заключающееся в том, что добавление РК к ЗТ приводит к индукции Wnt сигнального антагониста sFRP2 (ссылка 36). Возможно, что подавление Wnt сигнального антагониста молекулами, присутствующими при развитии переднего мозга, индуцирует у клеток фенотип радиальных глиальных клеток. Пространственная и временная экспрессия структуры sFRP1 сочетается с этим наблюдением (ссылка 37).
В условиях настоящего изобретения in vitro ключевым моментом является добавление РК (ссылка 38). Если после 4 суток обработки РК практически все клетки в ЗТ экспрессируют Рах-6, то в отсутствие РК Рах-6-позитивные клетки обнаружить невозможно, и после диссоциации необработанных ЗТ нейроны обнаружены не были. Хотя представляется маловероятным, что РК имеет физиологическое значение, заключающееся в индукции Рах-6 в развивающейся коре головного мозга, это может происходить в других частях развивающейся ЦНС. В самом деле, хотя Рах-6 имеет ограниченный характер экспрессии в ЦНС, включая кору головного мозга, также происходит экспрессия Рах-6 в большей части брюшной части нервной трубки во время развития, и полученные результаты означают, что сегмент-производная РК имеет физиологическое значение при развитии определенной структуры брюшной части нервной трубки (ссылки 39, 40). В случае ЗТ, обработанных РК, Renoncourt и др. (ссылка 28) и Wichterle и др. (ссылка 7) также наблюдали, что некоторые клетки в ЗТ являются Рах-6-позитивными после обработки РК. Однако Рах-7-позитивные клетки также наблюдались при схожем воздействии (ссылка 7), что подтверждает гетерогенность состава клеток после обработки ЗТ ретиноевой кислотой (РК).
На поликатионном субстрате, покрытом ламинином, радиальные глиальные клетки быстро теряют типичную веретеновидную форму. При использовании линий EGP-ES установлено, что число флуоресцирующих клеток быстро возрастает и что по размеру и форме нейроны выглядят в высокой степени однородными. К 4-м суткам после диссоциации практически все клетки уже имеют нейрональные характеристики. В последующий период практически все нейроны принимают пирамидальную форму и являются позитивными на наличие везикулярного транспортера глутамина. Все клетки окрашивают в разные сроки независимо от их идентичности. Напротив, менее 0,1% клеток очевидно позитивны, что следует из окраски через 1 неделю в культуре с антителами к Is1-1, тирозингидроксилазе или холинтрансферазе. Через три недели менее 5% клеток являются vGAT-положительными. Отсутствие окрашивания ГАМК и Is1-1 исключает ряд интернейронов и нейронов большой протяженности, включая, в частности, двигательные нейроны, многие из которых также являются производными Рах-6-позитивных клеток in vivo. Предположительно, индуктивные сигналы, например, sonic hedgehog, должны присутствовать для движения потомства Рах-6-позитивных радиальных глиальных клеток по определенному пути дифференциации (ссылка 7). Глутаматергический фенотип нейронов в настоящем изобретении соответствует их идентичности в качестве кортикальных пирамидальных нейронов, что следует из их формы. Наиболее важно, что этот признак согласуется с наблюдением, что все эти нейроны происходят от радиальных глиальных клеток. Действительно, ранее Malatesta и др. (ссылка 11) установили, что потомство кортикальных радиальных глиальных клеток является пирамидальными нейронами, расположенными во всех слоях коры и в гиппокампе. Таким образом, предусмотренные в настоящем изобретении условия культивирования могут быть описаны в качестве «разрешительных (пермиссивных)», позволяющих внутренней программе дифференциации глиальных клеток развертываться in vitro. В соответствии с этим используемая в настоящем изобретении среда первоначально создавалась для поддержания выживаемости и дифференциации пирамидальных нейронов, выделенных из гиппокампа эмбрионов грызунов (ссылка 12). Свойством этой среды также является способность предупреждать или репрессировать размножение таких клеток, как астроциты. Следует ожидать, что эти клетки присутствуют в культурах, полученных согласно настоящему изобретению, поскольку они также относятся к потомству радиальных глиальных клеток.
С помощью GFAP-антител наблюдают развитие нескольких разветвленных астроцитов в культуре. Однако их число является слишком маленьким (в диапазоне 1-2% от общего числа клеток через 3 недели).
Относительная однородность полученных нейрональных культур стимулировала исследовать экспрессию мембранных белков, о которых известно, что они экспрессируются в определенные сроки развития. В соответствии с полученными in vivo результатами (ссылка 23) установили, что экспрессия р75 тесно коррелирует с появлением нейронов в этих культурах и позднее регуляция понижается. Напротив, хотя экспрессию рецептора Trk не обнаруживают на ранние сроки, она резко возрастает через несколько суток, подтверждая, что развитие нейронов происходит синхронно. Высокие уровни экспрессии рецептора Trk являются характеристикой пирамидальных нейронов in vivo (ссылка 22). Эксперименты с применением метода ПЦР подтверждают, что и TrkB, и TrkC воздействуют на сигнал, полученный с использованием pan-Trk антител, хотя через несколько первых суток in vitro обнаруживают только экспрессию TrkA. В отличие от рецепторов р75 и Trk, APP четко обнаруживают уже через 2 ч после диссоциации ЗТ и их уровни повышаются во время курса нейрональной дифференциации. Это находится в соответствии с результатами иммуногистохимических экспериментов, показывающих, что APP специфически метит радиальные глиальные клетки в развивающейся коре головного мозга грызунов (ссылка 24).
Материалы и методы
Материалы
Ингредиенты для среды для культуры клеток были получены фирмы Gibco, LIF получен от фирмы Chemicon, ПДЛ и исходные растворы для среды N2 и полноценной среды - от фирмы Sigma. Порошковая фракция БСА V производства фирмы Gibco. Ламинин выделен из клеток саркомы Engelbreth-Holm-Swann (фирма Roche). PK приобретена в фирме Sigma, причем результаты не отличались при использовании разных доз.
Антитела
Первичные антитела для иммуноцитохимии являются мышиным моноклональным антителом антинестином (rat401, IgG1; 1:10; фирма DSHB - Банк гибридом для научных исследований,), мышиным моноклональным антителом RC2 (IgM; 1:4; фирма DSHB), кроличьим поликлональным антителом анти-BLBP (1:2000; любезно предоставлено N. Heintz для М. Goetz, Рокфеллеровский Университет, Нью-Йорк), мышиным моноклональным антителом анти-Рах6 (IgG1; 1:100; фирма DSHB), мышиным моноклональным антителом анти-βтубулином III (IgG2b; 1:100; фирма Sigma) и кроличьим поликлональным антителом анти-vGlut1 (1:5000; фирма SYSY). Подкласс-специфические Cy2- или Cy-3-связанные антисыворотки применяют в качестве вторичных антител. Для Вестерн-блоттинга применяют мышиное моноклональное антитело анти- синаптофизин (IgG1; 1:1000; фирма Sigma), кроличье поликлональное антитело анти-GluR1 (1:1000; фирма Upstate), кроличье поликлональное антитело анти-Trk (С-14, sc-11; 1:1000; фирма Santa Cruz), кроличье поликлональное антитело анти-АРР (1:3000; любезно предоставлено Р. Paganetti, фирма Novartis, Базель), кроличье поликлональное антитело анти-р75 (1:2000; фирма Promega).
Среды
Среда для ЭС клеток (500 мл):
(инактивируют нагреванием при 55°С 30 мин)
Среда для ЗТ (500 мл):
Среда N2:
П/С обозначает антибиотики, например, пенициллин/стрептомицин. Антибиотики необязательно могут быть исключены из среды и замещены равным объемом DMEM.
Исходные растворы для среды N2:
БСА фирмы Gibco, номер в каталоге А-9418, порошковая фракция V 100 г
Аликвоты 10 мг/мл хранят при -20°С
Конечная концентрация 50 мкг/мл
Инсулин фирмы Sigma, номер в каталоге I-6634 100 мг
Исходный раствор 5 мг/мл в воде (подкисляют каплей концентрированной HCl до величины рН 2 для растворения инсулина)
Хранят при -80°С
Трансферрин фирмы Sigma, номер в каталоге I-1147, апотрансферрин человека 100 мг
Исходный раствор 2 мг/мл в воде
Хранят при -80°С
Прогестерон фирмы Sigma, номер в каталоге Р-8783 5 г
Исходный раствор 2 мМ в этаноле хранят при -80°С.
Рабочий раствор 20 мкМ получают разведением исходного раствора в воде и хранят при -80°С.
Путресцин фирмы Sigma, номер в каталоге Р-5780
Исходный водный раствор 100 мкМ хранят при -80°С.
Селенит натрия фирмы Sigma, номер в каталоге S-5261 25 г
Исходный водный раствор концентрацией 300 мкМ сохраняют при 4°С.
Полноценная среда:
Водные растворы:
L-аланин (фирмы Sigma, номер в каталоге А-7627) [исходный раствор 2 мг/мл] 2 мкг/мл
Биотин (фирмы Sigma, номер в каталоге В-4501) [исходный раствор 0,1 мг/мл] 0,1 мкг/мл
L-карнитин (фирмы Sigma, номер в каталоге С-0283) [2 мг/мл] 2 мкг/мл
Этаноламин (фирмы Sigma, номер в каталоге Е-9508) [1 мг/мл) 1 мкг/мл
D+ - галактоза (фирмы Sigma, номер в каталоге G-0625) [15 мг/мл] 15 мкг/мл
L-пролин (фирмы Sigma, номер в каталоге Р-0380) [7,76 мг/мл] 7,76 мкг/мл
Путресцин (фирмы Sigma, номер в каталоге Р-7505) [16,1 мг/мл] 16,1 мкг/мл
Пируват натрия (фирмы Sigma, номер в каталоге Р-5280) [25 мг/мл] 25 мкг/мл
Селенит натрия (фирмы Sigma, номер в каталоге S-1382) [0,016 мг/мл] 0,016 мкг/мл
Витамин В12 (фирмы Sigma, номер в каталоге V-2876) [0,34 мг/мл] 0,34 мкг/мл
Сульфат цинка (фирмы Sigma, номер в каталоге Z-4750) [0,194 мг/мл] 0,194 мкг/мл
Каталаза (фирмы Sigma, номер в каталоге С-40) [16 мг/мл] 16 мкг/мл
Глютатион (фирмы Sigma, номер в каталоге G-6013) [1 мг/мл] 1 мкг/мл
SOD (фирмы Sigma, номер в каталоге S-2515) [2,5 мг/мл] 2,5 мкг/мл
Этанольные растворы:
Линолевая кислота (фирмы Sigma, номер в каталоге L-1376) [100 мг/мл] 1 мкг/мл
Линоленовая кислота (фирмы Sigma, номер в каталоге L-2376) [100 мг/мл] 1 мкг/мл
Прогестерон (фирмы Sigma, номер в каталоге Р-8783) [0,63 мг/мл] 6,3 нг/мл
Ретинол (транс-форма) Retinol (фирмы Sigma, номер в каталоге R-7632) [10 мг/мл] 100 нг/мл
Ретинилацетат (фирмы Sigma, номер в каталоге R-7882) [10 мг/мл] 100 нг/мл
Токоферол (фирмы Sigma, номер в каталоге Т-3251) [100 мг/мл] 1 мкг/мл
Токоферолацетат (фирмы Sigma, номер в каталоге Т-3001) [100 мг/мл] 1 мкг/мл
Растворяют:
в 400 мл DMEM и добавляют указанные выше растворы.
Культура ЭС клеток
Первоначально ЭС клетки культивируют на питающих клетках, представляющих инактивированные митомицином фибробласты эмбрионов мышей, по меньшей мере, на протяжении двух пассажей после оттаивания. Последующие пассажи ЭС клеток культивируют без питающих клеток, и дифференциация может начаться либо немедленно через, по меньшей мере, два пассажа без питающих клеток, или из биомассы замороженных ЭС клеток без питающих клеток. Биомассу клеток, используемую для дифференциации, пассируют, по меньшей мере, дважды до начала процедуры. После культивирования ЭС клеток на питающих клетках весьма важным является первый пассаж на среду без питающих клеток. Успешная дифференциация зависит от плотности ЭС клеток, применяемых для первого пассажа. ЭС клетки должны занять, по меньшей мере, одну треть чашки через 1 сутки после расщепления. Основу среды для ЭС клеток составляет среда DMEM, содержащая 15% ФСТ (фетальная сыворотка теленка специально исследована для культуры ЭС клеток, впоследствии претерпевающих нейрональную дифференциацию), LIF (1000 ЕД/мл), не являющиеся незаменимыми аминокислоты и β-меркаптоэтанол. В чашки для культур клеток всегда наносят раствор 2% желатина, по меньшей мере, на 10 мин. Установлено, что температура инкубации является важным фактором и при температуре выше 37°С успешной дифференциации не происходит. Все среды предварительно подогревают до 37°С.
ЭС клетки расщепляют каждые 2 суток и с определенной плотностью, составляющей 1,5×106-4×106 клеток, высевают в чашки для культур клеток диаметром 10 см (фирмы Corning). Через 2 суток 10-25×106 клеток может быть выделено, причем их высокая пролиферативная активность является необходимым условием для успешного проведения эксперимента. Клетки находятся в фазе быстрого роста и формируют плоский монослой.
Расщепление клеток проводят промыванием в двукратном объеме ФСФ и инкубированием клеток в тонком слое раствора трипсина (однократно в растворе трипсина фирмы Gibco - 0,05% в 0,02% EDTA) при 37°С, 7% CO2 в течение 3 мин, чашки встряхивают вручную, клетки отделяют и ресуспендируют в новой порции среды для ЭС клеток путем засасывания и выдувания из пипетки (инактивация трипсином). Затем центрифугируют 5 мин при 1000 об/мин при комнатной температуре. Осадок опять ресуспендируют в новой порции среды для ЭС клеток с помощью пипетки, несколько раз засасывая и выдувая суспензию. Клетки следует диссоциировать для получения культуры отдельных клеток, хотя могут присутствовать агрегаты из 2-3 клеток, более крупных скоплений клеток не должно быть. Необходимое количество клеток заново высевают в чашки, поверхность которых покрыта желатином.
Для депривации ЭС клеток от питающих клеток их можно культивировать после оттаивания примерно дважды на питающих клетках и затем, по меньшей мере, 2 раза пассировать без питающих клеток таким образом, чтобы удалить фибробласты. ЭС клетки, таким образом, меняют форму с колониеподобной на плоскую.
Оттаивание ЭС клеток включает быстрое оттаивание исходной пробирки с ЭС клетками в количестве 3×106, ресуспендирование клеток в 10 мл среды для ЭС клеток и центрифугирование в течение 5 мин при 1000 об/мин при комнатной температуре. Осажденные клетки ресуспендируют еще раз в среде для ЭС клеток количество клеток высевают в чашку для культуры клеток диаметром 6 см. Замораживание ЭС клеток проводят путем ресуспендирования клеток, расщепившихся после обработки трипсином и центрифугирования в среде для ЭС клеток + 10% ДМСО.
Протокол нейрональной дифференциации
Для формирования ЗТ 3×106 ЭС клеток помещают в неадгерентные бактериологические чашки (фирма Greiner) в 15 мл среды ЭС (среда ЭС без ФПЛ и только с 10% ФСТ) и инкубируют в течение 8 суток.
Среду заменяют каждые двое суток, для чего удаляют всю культуру клеток с бактериологической чашки (в пробирку объемом 50 мл фирмы Falcon) и дают ЗТ осесть (примерно за 3-5 мин). Затем надосадочную жидкость тщательно отсасывают и к ЗТ заново добавляют 15 мл среды для ЗТ. ЗТ тщательно ресуспендируют в среде с помощью пипетки с достаточно широким кончиком для того, чтобы избежать разрушения или диссоциации ЗТ (например, с помощью 10 мл пластиковой пипетки).
РК (фирмы Sigma) в количестве 5 мкмолей добавляют через 4 суток непосредственно в чашку и распределяют аккуратным покачиванием чашки. РК не следует оставлять длительное время на свету, поскольку она разрушается под воздействием света. ЗТ затем подвергают диссоциации и клетки высевают в чашки с покрытием ПДЛ/ламинин следующим образом.
В чашки для культуры клеток вносят раствор 10 мкг/мл ПДЛ в боратном буфере (150 мМ рН 8,4) и помещают на ночь в инкубатор (37°С, 7% СО2). Полиорнитин также используют в концентрации 100 мкг/мл со схожим результатом. После трехкратного промывания чашек ФСБ (при использовании полиорнитина промывают H2O) ламинин (примерно 0,5 мкг/см2) добавляют непосредственно в раствор ФСБ и чашки возвращают в инкубатор, по меньшей мере, на 2 ч.
Через 8 суток формирования полученные ЗТ отмывают 2 раза ФСБ и обрабатывают трипсином путем инкубирования ЗТ в течение 3 мин на водяной бане при 37°С в 0,05% растворе трипсина в 0,04% EDTA/ФСБ (свежеприготовленный раствор с порошком трипсина, ТРСК-обработанный, фирмы Sigma). Во время инкубирования пробирку фирмы Falcon два раза вручную осторожно встряхивают, после чего дезинтеграция клеток ЗТ становится заметной. Диссоциированные ЗТ осторожно, но тщательно, ресуспендируют в 10 мл среды ЗТ, содержащей сыворотку для инактивации трипсина. Диссоциация может быть выполнена примерно путем пятикратного засасывания в пипетку и выдувания из пипетки. Наилучшим образом диссоциацию проводят пастеровской пипеткой малого объема с оттянутым в пламени кончиком (примерно 1,5 мл), а затем пластиковой пипеткой объемом 5 мл. После измельчения центрифугируют 5 мин при 1000 об/мин при комнатной температуре. Надосадочную жидкость полностью удаляют, осадок ресуспендируют в среде N2 и суспензию клеток фильтруют через нейлоновый фильтр с размером ячеек 40 мкм (фирма Falcon).
Ламинин удаляют с чашек с покрытием и немедленно добавляют суспензию клеток, не допуская подсушивания чашек. Диссоциированные клетки высевают с плотностью 1,5×105 клеток/см2. Среду N2 заменяют через 2 ч и еще раз через 1 сутки. Через 2 суток среду заменяют обогащенной не содержащей сыворотку средой, описанной Brewer и Cotman (ссылка 12), но в модифицированном варианте - без содержания глутамата, HEPES, кортикостерона, липоевой кислоты и Т3.
Нейрональную дифференциацию завершают и нейрональные культуры могут поддерживать в течение нескольких недель.
Иммуноцитохимия
Покровные стекла приготавливают промывкой в воде и погружением в 65% азотную кислоту в течение 1-2 суток. Впоследствии их флотируют в H2O в течение нескольких часов, промывают этанолом, высушивают на воздухе и стерилизуют УФ светом. Клетки фиксируют 4% параформальдегидом (ПФА) в течение 10 мин, промывают ФСБ и блокируют в течение 1 ч в блокирующем буфере (0,03% карагенан, 10% NGS, 0,3% тритон Х-100). Препарат монтируют в AquaPoly/Mount (фирма Polysciences).
Вестерн-блоттинг
Диссоциированные ЗТ высевают согласно указанному выше способу, и образцы для Вестерн-блоттинга собирают в определенные установленные сроки. Перед отбором клеток из чашек их дважды отмывают ледяным фосфатно-солевым буфером (ФСБ). Экстракты целых клеток приготавливают в 750 мкл литического буфера и помещают в чашку диаметром 6 см (50 мМ Tris pH 7,4, 150 мМ NaCl, 10% глицерин, 1% тритон Х-100), в которую добавлена смесь ингибиторов протеаз (фирма Roche). После центрифугирования в течение 30 мин при 4200 об/мин в центрифуге фирмы Eppendorf удаляют супернатант, а содержание белка определяют с помощью метода DC Protein Assay (фирма BioRad). Образцы кипятят в буфере Laemmli и по 5 мкг помещают в полиакриламидные гели. Пятна блокируют 5% раствором молока, инкубируют в течение ночи с первым антителом и 2 ч со вторым антителом. Определение проводят с помощью прибора ECL Plus (фирма Amersham).
Источники информации
1. Evans, М. J., Kaufman, М. Н. Nature 292, 1981, cc.154-156.
2. Martin, G. R. Proc. Natl. Acad.Sci. USA 78, 1981, cc.7634-7638.
3. Stavridis, М. Р. & Smith, А. G. Biochem. Soc. Trans. 31, 2003, cc.45-49.
4. Rathjen, J. & Rathjen, P. D. Curr. Opin. Genet. Dev. 11, 2001, cc.587-594.
5. Chung, S. и др., Eur. J. Neurosci. 16, 2002, cc.1829-1838.
6. Kawasaki, Н. и др., Neuron. 28, 2000, cc.31-40.
7. Wichterle, Н. и др., Cell 110, 2002, cc.385-397.
8. Kondo, М. и др., Annu. Rev. Immunol. 21, 2003, cc.759-806.
9. Lendahl, U., Zimmerman, L.B., McKay, R.D. Cell 60, 1990, cc.585-595.
10. Li, М. и др., Curr. Biol. 8, 1998, cc.971-974.
11. Malatesta, Р. и др., Neuron 37, 2003, cc.751-764.
12. Brewer, G. J., Cotman, С.W. Brain Res. 494, 1989, cc.65-74.
13. Tucker, К. L., Meyer, М., Barde, Y.A. Nat. Neurosci. 4, 2001, cc.29-37.
14. Bain, G., Kitchens, D., Yao, М., Huettner, J. E., Gottlieb, D. I. Dev Biol. 168, 1995, cc.342-357.
15. Li, М. в кн.: «Methods in Molecular Biology: Lineage selection for generation and amplification of neural precursor cells. Embryonic Stem Cells: Methods and Protocols», изд-во Humana Press Inc., 2002, Тотова, США, 185, cc.205-215.
16. Hartfuss, E., Galli, R., Heins, N., Gotz, М. Dev. Biol. 229, 2001, cc.15-30.
17. Misson, J. и др., Brain Res. Dev. Brain Res. 44, 1988, cc.95-108.
18. Feng, L., Hatten, M.E. & Heintz, N. Neuron 12, 1994, cc.895-908.
19. Gotz, M., Stoykova, A. & Gruss, P. Neuron 21, 1998, cc.1031-1044.
20. Banker, G. A. & Cowan, W. M. Brain Res. 126, 1977, cc.397-442.
21. Fremeau, R.T. и др., Jr Neuron 31, 2001, cc.247-260.
22. Klein, R. и др., Development 109, 1990, cc.845-850.
23. Bothwell, M. Annu. Rev. Neurosci. 18, 1995, cc.223-53.
24. Trapp, В. D. & Hauer, P. E. J. Neurosci. Res. 37, 1994, cc.538-550.
25. Fraichard, А. и др., J. Cell Sci. 108, 1995, cc.3181-3188.
26. Strübing, С. и др., Mech. Dev. 53, 1995, cc.275-287.
27. Okabe, S., Forsberg-Nilsson, К., Spiro, А. С., Segal, M., McKay, R. D. Mech. Dev. 59. 1996, cc.89-102.
28. Renoncourt, Y. и др., Mech. Dev. 79, 1998, cc.185-197.
29. Rathjen, J. и др., Development 129, 2002, cc.2649-2661.
30. Abe, Y. и др., J. Neurosci. 23, 2003, cc.8513-8525.
31. Barberi, Т. и др., Nat. Biotechnol. 21, 2003, cc.1200-1207.
32. Jones-Villeneuve, E. и др., J. Cell Biol. 94, 1982, cc.253-262.
33. Bain, G., и др., Biochem. Biophys. Res. Commun. 223, 1996, cc.691-694.
34. Murray, P., Edgar, D. Proc. Roy. Soc. Meeting Review (в печати).
35. Liour, S. S., Yu, R. K. Glia 42, 2003, cc.109-117.
36. Aubert, J., и др., Nat. Biotechnol. 20, 2002, cc.1240-1245.
37. Kim, А. и др., Mech. Dev. 103, 2001, cc.167-172.
38. Gajovic, S., и др., Differentiation 62, 1997, cc.187-192.
39. Diez del Corral, R. и др., Neuron 40, 2003, cc.65-79.
40. Novitch, В. и др., Neuron 40, 2003, cc.81-95.
41. Robertson, E. J. в кн.: «Teratocarcinoma and Embryonic Stem Cells: a Practical Approach» под ред. Robertson, E. J., изд-во IRL Press, 1987, Вашингтон, cc.71-112.
42. Tucker и др., Nat Neurosci 4, 2001, cc.29-37.
43. Okabe в кн.: «Current Protocols in Neuroscience», изд-во John Wiley & Sons, 1997, раздел 3.6, cc.1-13.
44. Weil и др., Cell Science 111, 1998, cc.2707-2715.
45. Leist и др., J. Immunol. 153, 1994, cc.1778-1788.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНО ДИФФЕРЕНЦИРОВАННЫХ ДОФАМИНЕРГИЧЕСКИХ НЕЙРОНОВ ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2004 |
|
RU2345133C2 |
| ПОЛУЧЕНИЕ ГАМКергических НЕЙРОНОВ in vitro ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ НЕВРОЛОГИЧЕСКИХ РАССТРОЙСТВ | 2004 |
|
RU2380410C2 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях гипоксии, и способ ее получения | 2019 |
|
RU2732600C1 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА В ДОФАМИНЭРГИЧЕСКИЕ НЕЙРОНЫ | 2023 |
|
RU2823290C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из глиальных прогениторных клеток в условиях теплового шока, и способ её получения | 2019 |
|
RU2732599C1 |
| Ноотропная композиция на основе полипептидных комплексов, выделенных из нейрональных прогениторных клеток в условиях теплового шока, и способ ее получения | 2019 |
|
RU2752906C2 |
| ТРАНСПЛАНТАЦИЯ НЕРВНЫХ КЛЕТОК ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ СОСТОЯНИЙ | 2005 |
|
RU2434636C2 |
| МАТРИЦА ДЛЯ КЛЕТОЧНОЙ ТРАНСПЛАНТОЛОГИИ | 2011 |
|
RU2521194C2 |
| СПОСОБ ПОЛУЧЕНИЯ АДГЕЗИВНОЙ КУЛЬТУРЫ НЕЙТРАЛЬНЫХ СТВОЛОВЫХ/ПРОГЕНИТОРНЫХ КЛЕТОК ОБОНЯТЕЛЬНОЙ ВЫСТИЛКИ НОСА МЛЕКОПИТАЮЩИХ ДЛЯ ЛЕЧЕНИЯ ТРАВМ СПИННОГО МОЗГА | 2020 |
|
RU2749156C1 |
| ПРИМЕНЕНИЕ СТВОЛОВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, И ИХ КОМПОЗИЦИЙ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ (ВАРИАНТЫ) | 2017 |
|
RU2742828C2 |
Изобретение относится к области биотехнологии, в частности к способу индуцированной дифференциации эмбриональных стволовых (ЭС) клеток в нейрональные клетки-предшественники. Данный способ включает культивирование ЭС клеток путем высева ЭС клеток с плотностью примерно 0,5×105 - 2×105 клеток на см2 и диссоциацию ЭС клеток через 2 суток после высева. После чего производят формирование клеточных агрегатов (КА), которое включает отбор клеток с высокой пролиферативной активностью со временем удвоения от 0 до 24 часов и высев этих клеток с плотностью примерно 0,5×105 - 5×105 клеток в мл для формирования КА. Далее проводят обработку клеточных агрегатов ретиноевой кислотой (РК). Затем проводят диссоциацию КА для получения культуры нейрональных клеток-предшественников. Предложенное изобретение позволяет получать в значительной степени однородную популяцию нервных клеток, в которой нервные клетки практически все относятся к одной определенной линии дифференцировки нейрональных клеток, одному фенотипу, типу клеток и/или к одной стадии дифференцировки. 25 з.п. ф-лы, 1 ил., 1 табл.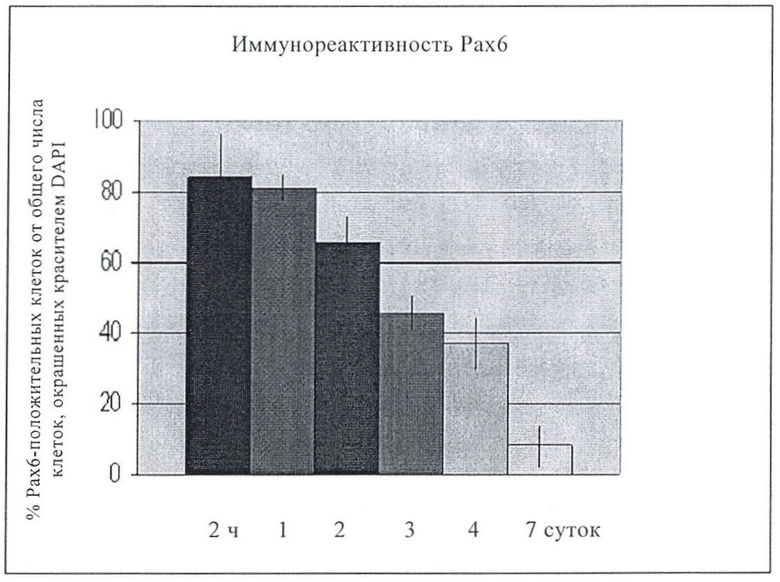
1. Способ индуцированной дифференциации эмбриональных стволовых (ЭС) клеток в нейрональные клетки-предшественники, включающий
культивирование ЭС клеток,
формирование клеточных агрегатов (КА),
обработку клеточных агрегатов (КА) ретиноевой кислотой (РК) и
диссоциацию КА для получения культуры нейрональных клеток-предшественников,
в котором культивирование ЭС клеток включает пассирование путем высева ЭС клеток с плотностью примерно 0,5×105 - 2×105 клеток на см2 и диссоциацию ЭС клеток через 2 суток после высева, а формирование КА включает отбор ЭС клеток с высокой пролиферативной активностью со временем удвоения от 0 до 24 ч и высев этих клеток с плотностью примерно 0,5×105 - 5×105 клеток в мл для формирования КА.
2. Способ по п.1, в котором формирование КА включает высев ЭС клеток с плотностью примерно 2,5×105 - 3,5×105 клеток в мл.
3. Способ по п.1 или 2, в котором КА поддерживают в неадгерентной культуре до диссоциации клеточных агрегатов.
4. Способ по п.1 или 2, включающий изучение морфологии ЭС клеток и отбор морфологически гомогенных ЭС клеток для формирования КА.
5. Способ по п.4, включающий отбор ЭС клеток, обладающих одним или несколькими из следующих морфологических свойств: ростом в виде плоского монослоя, отсутствием непосредственного контакта между соседним клетками, крупными ядрами, большим количеством ядрышек, отсутствием роста клеток поверх друг друга или в виде форм, напоминающих колонии.
6. Способ по п.1 или 2, включающий определение стадии пролиферации ЭС клеток и отбор клеток с высокой пролиферативной активностью для формирования КА.
7. Способ по п.1 или 2, в котором культивирование ЭС клеток включает пассирование ЭС клеток в отсутствии питающих клеток.
8. Способ по п.7, в котором пассирование повторяют по меньшей мере дважды в отсутствии питающих клеток.
9. Способ по п.1 или 2, включающий диссоциацию ЭС клеток для получения суспензии отдельных клеток, в которой примерно менее 5% клеток образуют агрегаты из 4 или более клеток, и высев клеток для формирования КА.
10. Способ по п.1 или 2, в котором диссоциация КА заключается в диссоциации клеток КА трипсином для формирования суспензии клеток диссоциированных КА с последующим фильтрованием суспензии для удаления скоплений клеток.
11. Способ по п.10, в котором клетки диссоциированных КА фильтруют через ячейки размером примерно 40 мкм.
12. Способ по п.1 или 2, предусматривающий дополнительный этап хранения клеток диссоцированных КА путем замораживания клеток.
13. Способ по п.1 или 2, дополнительно включающий высев и культивирование диссоциированных клеток КА для получения нейронов и необязательно культивирование нейронов.
14. Способ по п.13, в котором клетки диссоциированных КА высевают с плотностью примерно 0,5×105 - 2,5×105 клеток на см2.
15. Способ по п.14, в котором клетки диссоциированных КА высевают с плотностью примерно 1×105 - 1,5×105 клеток на см2.
16. Способ по п.13, включающий замену культуральной среды клеток диссоциированных КА примерно через 1-6 ч после высева клеток диссоциированных КА.
17. Способ по п.16, включающий замену культуральной среды примерно через 1-3 ч после высева.
18. Способ по п.13, в котором клетки диссоциированных КА или нейроны не культивируют в присутствии сыворотки.
19. Способ по п.13, в котором нейрональные клетки-предшественники или нейрональные клетки не культивируют в присутствии ростовых факторов.
20. Способ по п.19, в котором нейрональные клетки-предшественники или нейрональные клетки не культивируют на среде Neurobasal.
21. Способ по п.1 или 2, в котором отсутствует стадия выбраковки клеток.
22. Способ по п.1 или 2, в котором ЭС клетки не являются ЭС клетками человека.
23. Способ по п.1 или 2, включающий идентификацию по меньшей мере 80% диссоциированных клеток КА в качестве нейрональных клеток-предшественников.
24. Способ по п.1 или 2, включающий идентификацию по меньшей мере 99% клеток диссоциированных КА в качестве нейрональных клеток-предшественников.
25. Способ по п.13, включающий идентификацию по меньшей мере 80% клеток в качестве нейронов.
26. Способ по п.25, включающий идентификацию по меньшей мере 90% клеток в качестве нейронов.
| BAIN G | |||
| et al, Embryonic stem cells express neuronal properties in vitro, Developmental Biology, Academic press, New York, 1995, v.168, n.2, p.342-357 | |||
| US 6432711, 13.08.02. |

