Перекрестные ссылки на родственные заявки
Не применимо
Заявка в отношении поддержанного правительством исследования или разработки
Не применимо
Ссылка на "дополнение в виде микрофиша"
Не применимо
1. Область изобретения
Настоящее изобретение относится к улучшенному способу продуцирования нервных клеток в конечной стадии дифференцировки, таких как ГАМКергические нейроны, из полипотентных стволовых клеток, таких как мышиные эмбриональные стволовые клетки или эмбриональные стволовые клетки человека. Полученные согласно описанию настоящего изобретения ГАМКергические нейроны можно использовать в качестве отличного источника в клеточной заместительной терапии при нейродегенеративных нарушениях и нервных заболеваниях, таких, например, как инсульт, ишемия, болезнь Паркинсона, болезнь Альцгеймера, эпилепсия и болезнь Гентингтона.
2. Описание предшествующей области
Гамма-аминомасляная кислота (ГАМК) представляет собой основной тормозной нейромедиатор центральной нервной системы (ЦНС), и она широко распространена повсеместно в головном мозге и экспрессируется в межнейронных модулирующих локальных цепях. Синтезирующие ГАМК ГАМКергические нейроны представляют собой основные тормозные нейроны в ЦНС млекопитающих, и приблизительно 60-75% всех синапсов в ЦНС являются ГАМКергическими (Schwartz, R.D., 1988, Biochem. Pharmacol. 37: 3369-75). ГАМКергические нейроны локализованы в гиппокампе, мозжечке, коре головного мозга и гипоталамусе, и ГАМК связывается по меньшей мере с тремя рецепторами, включающими в себя ГАМК-А и ГАМК-Б. ГАМК-А-рецепторы опосредуют быстрые тормозные синаптические передачи, нейронную возбудимость и быстрые изменения настроения, такие как порог судорожной активности, тревога, паника и ответ на стрессы (например, ответ «нападение/бегство»). ГАМК-А-рецепторы также имеют сайты связывания бензодиазепинов, этанола, барбитуратов и нейростероидов. ГАМК-Б-рецепторы опосредуют медленные тормозные передачи и могут быть важны в памяти, настроении и боли.
Патогенез некоторых неврологических расстройств, включая некоторые формы эпилепсии, хроническую боль, тревогу и расстройства настроения, вероятно, связан со снижением ГАМКергической передачи. Например, позитрон-эмиссионное томографическое исследование (PET) показало, что пациенты с паническим расстройством имеют пониженный уровень связывания ГАМК-А-рецептора (Malizia et al., 1998, Arch. Gen. Psychiatry 55: 715-20). Кроме того, низкий уровень ГАМК в плазме может являться характеристикой подгруппы пациентов с расстройствами настроения (Brambilla et al., 2003, Mol. Psychiatry 8: 721-37). Показано, что при лечении этих нарушений эффективны определенные повышающие активность ГАМК лекарственные средства, такие как бензодиазепины, вальпроат и фенобарбитал.
Многие заболевания центральной нервной системы (ЦНС), такие как болезнь Паркинсона, болезнь Альцгеймера, рассеянный склероз, болезнь Гентингтона, боковой амиотрофический склероз, ишемия головного мозга и инсульт, характеризуются дегенерацией нейронов головного мозга и отделов спинного мозга. Дегенерирующие или поврежденные иным способом клетки или нейроны, по сути, не замещаются или не восстанавливаются организмом, что может приводить к долговременному и необратимому повреждению (During et al., 2001, Human Gene Ther. 12: 1589-1591). Инсульт или ишемия головного мозга могут происходить, когда кровяной сгусток закупоривает кровеносный сосуд или артерию, нарушая кровоток в области головного мозга, что вызывает гибель клеток головного мозга в ближайшей к месту закупорки области. При инфаркте гибель клеток головного мозга обычно настает в интервале от минут до нескольких часов после возникновения инсульта или ишемии. Гибель этих клеток может приводить к высвобождению химических соединений, вызывающих цепную реакцию, названную «ишемический каскад», подвергающую опасности клетки головного мозга в более обширной окружающей ткань головного мозга области с аномальным кровотоком. Без быстрого оказания медицинской помощи клетки головного мозга этой более обширной области также могут погибнуть, что может явиться причиной еще более серьезного и долговременного повреждения головного мозга. Если принять во внимание быстрый темп ишемического каскада, становится ясно, что «окно терапевтических возможностей» для активного лечения составляет лишь около шести часов. После этого временного промежутка восстановление тока крови и введение нейропротективных средств может оказаться недостаточным для помощи и может вызвать дальнейшее повреждение функций головного мозга (Padosch et al., 2001, Anaesthesist 50: 905-920; Nishino and Borlongan, 2000, Prog. Brain Res. 127: 461-476).
При возникновении инсульта нарушение тока крови в головном мозге оказывает вредное и потенциально смертельное воздействие на отдельные нейроны или группы нейронов. Недостаток кислорода и жизненно важных питательных веществ у большого количества нейронов в ограниченной области головного мозга в результате ишемии головного мозга может приводить к серьезной потере функциональных способностей пациента. Например, пациенты с инсультом могут испытывать потерю речи, памяти, когнитивных функций, ограниченную подвижность или даже паралич. Без адекватного кровоснабжения клетки головного мозга теряют свою способность вырабатывать энергию, в частности, аденозинтрифосфат (АТФ). Если достичь критических порогов такого недостатка энергии, клетки головного мозга повреждаются и погибают. Многие исследователи полагают, что вслед за недостатком энергии повреждение и гибель клеток головного мозга вызывают огромное количество механизмов и каждый из этих механизмов представляет собой потенциальный путь для терапевтического вмешательства.
Один из способов, посредством которого клетки головного мозга отвечают на недостаток энергии, связан с повышением концентрации внутриклеточного кальция. Такие концентрации могут приводить к опасным уровням посредством процесса, называемого эксцитотоксичность, при котором клетки головного мозга высвобождают избыточные количества глутамата, нейромедиатора, который приводит к деградации и деструкции живых клеток, локализованных в гиппокампе, коре головного мозга и таламической области головного мозга (Nishino and Borlongan, 2000, Prog. Brain Res. 127: 461-476). Кроме того, после инсульта в области гиппокампа головного мозга часто дегенерируют клетки, синтезирующие ГАМК (Nishino and Borlongan, 2000, Prog. Brain Res. 127: 461-476). На основе нейрогистопатологических и нейропсихологических исследований разработано несколько нейропротективных лекарственных средств для лечения неврологических расстройств, связанных с ишемией головного мозга или инсультом, таких как ГАМКергические агонисты, антагонисты кальция, антагонисты глутамата и антиоксиданты (Stutzmann et al., 2002, CNS Drug Rev. 8: 1-30; Rochelle et al., 2001, J. Neurochem. 77: 353-371; Blezer et al., 2002 Eur. J. Pharmacol. 444: 75-81). В настоящее время на разных стадиях разработки существуют сотни препаратов и соединений для предотвращения и экстренного инвазивного лечения инсульта (Rochelle et al., 2001, J. Neurochem. 77: 353-371). Планируется предоставить некоторые из этих препаратов на рассмотрение в FDA, и многие уже включены в последнюю фазу клинических испытаний. Обнаружено, что при лечении неврологических расстройств, связанных с ишемией головного мозга или инсультом, среди этих препаратов ГАМКергические препараты являются исключительно эффективными.
Однако, принимая во внимание многофакторную природу ишемического повреждения клеток головного мозга, специалисты по инсульту предполагают, что основанный на одном лекарственном средстве способ лечения не способен полностью защитить головной мозг во время инсульта и после него. Существует огромный интерес к разработке альтернативных способов лечения различных нейродегенеративных нарушений и нервных заболеваний, так как современные терапевтические средства не лечат адекватно связанное с ишемией головного мозга или инсультом повреждение. Только терапия на основе клеток может быть доступным средством для всестороннего лечения вызванного таким событием повреждения. Многие неврологические заболевания и состояния вызваны потерей нервных клеток в головном мозге и отделах спинного мозга. Широкий спектр таких неврологических заболеваний и состояний, включая - в качестве неограничивающих примеров - болезнь Паркинсона, болезнь Альцгеймера, болезнь Гентингтона и повреждение спинного мозга, могут быть излечимы посредством терапии на основе клеток. Например, пациентов с болезнью Паркинсона успешно лечили посредством трансплантации дофаминергических нейронов в головной мозг пораженных индивидов (Grisolia, 2002, Brain Res Bull 57: 823-826). Следовательно, когда у пациентов с ишемией головного мозга или инсультом поражены или повреждены продуцирующие ГАМК клетки, замещение этих поврежденных продуцирующих ГАМК клеток новыми и здоровыми продуцирующими ГАМК клетками было бы идеальной терапией при лечении ишемии головного мозга или инсульта.
Потребность в больших количествах нервных клеток, которые трудно выделить из эмбрионов или взрослых особей, является одной из основных проблем трансплантации клеток как терапевтического средства при нейродегенеративных нарушениях и нервных заболеваниях. Единственным решением этой дилеммы является доступность полипотентных стволовых клеток, которые можно применять для образования неограниченного количества типов клеток в конечной стадии дифференцировки. Полипотентные эмбриональные стволовые (ЭС) клетки представляют собой жизнеспособный альтернативный источник нервных клеток, которые можно применять для лечения различных нейродегенеративных нарушений и нервных заболеваний. ЭС клетки могут неограниченно пролиферировать в недифференцированном состоянии и являются полипотентными, что делает возможной их дифференцировку почти во все присутствующие в организме типы клеток. Так как ЭС клетки способны становиться почти всеми специализированными клетками организма, они могут замещать клетки широкого ряда тканей и органов, таких как сердце, поджелудочная железа, нервная ткань, мышца, хрящ и пр. ЭС клетки можно выделять из внутренней клеточной массы (ВКМ) бластоцисты, представляющей собой стадию эмбрионального развития, проходящую перед имплантацией. Человеческие ЭС клетки можно выделять из человеческой бластоцисты на ранней стадии эмбрионального развития, продолжающейся от 4 до 7 суток после оплодотворения. Выделенные из ВКМ ЭС клетки можно культивировать in vitro и в подходящих условиях для неограниченной пролиферации.
Линии ЭС клеток успешно созданы для ряда видов, включая мышь (Evans et al., 1981, Nature 292: 154-156), крысу (Iannaccone et al., 1994, Dev. Biol., 163: 288-292), свинью (Evans et al., 1990, Theriogenology 33: 125-128; Notarianni et al., 1990, J. Reprod. Fertil. Suppl. 41: 51-6), овцу и козу (Meinecke-Tillmann and Meinecke, 1996, J. Animal Breeding and Genetics 113: 413-426; Notarianni et al., 1991, J. Reprod. Fertil. Suppl. 43: 255-60), кролика (Giles et al., 1993, Mol. Reprod. Dev. 36: 130-138, Graves et al., 1993, Mol. Reprod. Dev. 36: 424-433), норку (Sukoyan et al., Mol. Reprod. Dev. 1992, 33: 418-431), хомяка (Doetschman et al., 1988, Dev. Biol. 127: 224-227), домашнюю птицу (Pain et al., 1996, Development 122(8): 2339-48), приматов (патент США №5843780), и человека (Thomson et al., 1998, Science 282: 1145-1147; Reubinoff et al., 2000, Nature Biotech. 18: 399-403). При введении иммунодефицитным мышам ЭС клетки человека, подобно другим ЭС клеткам млекопитающих, дифференцируются и формируют ткани всех трех зародышевых листков, доказывая свою полипотентность. Опубликованные сообщения показывают, что ЭС клетки человека поддерживали в культуре больше года, в течение которого они сохраняли полипотентность, способность к самообновлению и нормальный кариотип (Thomson et al., 1995, PNAS 92: 7844-7848).
Показано, что ЭС клетки дифференцировались в нейроны и глиальные клетки в моделях in vitro (Bain et al., 1995, Dev. Biol. 168: 342-357) и в моделях in vivo (Brustle, et al., 1999, Science 285: 754-56). Аналогично, после трансплантации стволовые клетки стадии бластулы могут дифференцироваться в дофаминергические и серотонинергические нейроны (Deacon et al., 1998, Exp. Neurol. 149: 28-41). Стволовые клетки человека или грызунов способны дифференцироваться в специфические типы нейронов при пересадке как в развивающуюся центральную нервную систему (Flax et al., 1998, Nat. Biotechnol. 16: 1033-39; Brustle et al., 1998, Nat. Biotechnol. 16: 1040-44; Reubinoff et al., 2001, Nat. Biotechnol. 19: 1034-40), так и в нейрогенные области ЦНС взрослой особи (Fricker et al., 1999, J. Neurosci. 19: 5990-6005; Shihabuddin et al., 2000, J. Neurosci. 20: 8727-35).
Сообщалось об одном способе получения продуцирующих ГАМК клеток из незрелых нервных клеток (Rubenstein et al., патент США №6602680, включенный в настоящее описание в качестве ссылки). Rubenstein et al. описали продукцию ГАМКергических клеток посредством повышения активности гена DLX в незрелых нервных клетках, например DLX1, DLX2 или DLX5. Повышение активности DLX является причиной дифференцировки незрелых нервных клеток в клетки с ГАМКергическим фенотипом. Также описаны способы получения продуцирующих ГАМК клеток из мышиных эмбриональных стволовых клеток (Hancock et al., 2000, Biochem. Biophys. Res. Commun. 271(2): 418-21, Westmoreland et al., 2001, Biochem. Biophys. Res. Commun. 284(3): 674-80; U.S. Publication №2003/0036195 Al, каждая конкретно включена в настоящее описание в качестве ссылки), но этими способами не получают высокого содержания ГАМКергических нейронов. Существует необходимость в дополнительных способах получения in vitro больших количеств ГАМКергических нейронов из полипотентных стволовых клеток, так как большие количества ГАМКергических нейронов необходимы для клеточной заместительной терапии.
Способы, посредством которых можно получать высокий выход ГАМКергических нейронов, имеют огромное клиническое значение для клеточной трансплантационной терапии, особенно для пациентов, страдающих от ишемии головного мозга или инсульта. В настоящее время доступные способы лечения являются крайне ограниченными в том, что касается лечения нейропатологии, связанной с ишемией головного мозга и инсультом, поэтому существует огромный интерес к развитию альтернативных способов лечения. Клеточная терапия требует больших количеств клеток или нейронов для лечения, которые не достижимы, если доступным источником клеток и нейронов является только эмбриональная ткань или ткань взрослой особи. Например, для изучения функционального восстановления двигательной функции необходимо пересадить в модельное животное с болезнью Паркинсона приблизительно 1 миллион клеток, продуцирующих дофаминергические нейроны (Grisolia, 2002, Brain Res. Bull. 57: 823-826). Получение такого большого количества клеток с применением эмбрионального материала вызывает множество этических проблем. Получение ГАМКергических нейронов из полипотентных стволовых клеток позволяет обеспечить потенциально неограниченную поставку ГАМКергических нейронов для применения в способах лечения на основе клеток. Однако, чтобы сделать данный источник осуществимым на практике, необходимы способы с высоким выходом ГАМКергических нейронов, особенно потому, что, вероятно, потребуются большие количества ГАМКергических нейронов для способов терапии с применением этих клеток.
Краткая сущность изобретения
Настоящее описание изобретения относится к улучшенным способам получения из полипотентных стволовых клеток, таких как эмбриональные стволовые (ЭС) клетки, нейрональных клеток-предшественников, а также глиальных клеток или нервных клеток в конечной стадии дифференцировки. В предпочтительном варианте осуществления полипотентные стволовые клетки, или ЭС клетки, представляют собой мышиные клетки. Полученные здесь клетки включают в себя в качестве неограничивающих примеров клетки с фенотипическими характеристиками нейрональных клеток-предшественников, нервных клеток, таких как ГАМКергические, дофаминергические, серотонинергические и глутаматергические нейроны, а также глиальные клетки, такие как олигодендроциты и астроциты. В настоящем описании изобретения показано, что полипотентные стволовые клетки, например мышиные ЭС клетки, могут дифференцироваться в клетки с высоким содержанием ГАМКергических нейронов (например, по меньшей мере 60%). Доля полученных способами согласно описанию ГАМКергических нейронов выше, чем в описанных ранее способах. ГАМКергические нейроны можно применять для поливалентных основанных на клетках способов лечения, например, в клеточной заместительной терапии или при лечении нейродегенеративных нарушений и нервных заболеваний, включая, например, инсульт, ишемию головного мозга, эпилепсию, болезнь Паркинсона, болезнь Гентингтона, болезнь Альцгеймера, хроническую боль, тревогу и другие расстройства настроения.
В настоящем описании представлена популяция дифференцированных клеток в культуре in vitro, полученных посредством дифференцировки полипотентных стволовых клеток, где по меньшей мере 60% дифференцированных нервных клеток представляют собой ГАМКергические нейроны, клетки, проявляющие фенотипы ГАМКергических нейронов, или клетки, продуцирующие гамма-аминомасляную кислоту (ГАМК). Предпочтительно, чтобы ГАМКергические нейроны экспрессировали GAD65, GAD67, рецептор ГАМК-А или рецептор ГАМК-Б, или их сочетание. В других вариантах осуществления по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% дифференцированных клеток представляют собой ГАМКергические нейроны или продуцируют ГАМК. Предпочтительно, чтобы ГАМКергические нейроны или продуцирующие ГАМК клетки были получены из ЭС клеток, более предпочтительно из человеческих или мышиных ЭС клеток. В других вариантах осуществления популяция дифференцированных клеток также содержит по меньшей мере приблизительно 15% дофаминергических нейронов, по меньшей мере приблизительно 10% глутаматергических нейронов, по меньшей мере приблизительно 10% глутаматергических нейронов, по меньшей мере приблизительно 5% серотонинергических нейронов, по меньшей мере приблизительно 5% астроцитов, или сочетание этих количеств.
Согласно изобретению также представлены способы получения дифференцированных клеточных популяций из полипотентных стволовых клеток, включающие в себя следующие этапы:
(a) наращивание культуры из полипотентных стволовых клеток;
(b) культивирование полипотентных стволовых клеток для отбора нестин-позитивных нейрональных клеток-предшественников;
(c) наращивание нестин-позитивных нейрональных клеток-предшественников; и
(d) дифференцировку нестин-позитивных клеток с образованием дифференцированной клеточной популяции посредством культивирования клеток в среде для дифференцировки, содержащей цитозин-β-d-арабинофуранозид (Ara-C).
В предпочтительном варианте осуществления полипотентные стволовые клетки представляют собой ЭС клетки, более предпочтительно человеческие или мышиные ЭС клетки.
В других вариантах осуществления описанные выше способы дополнительно включают в себя стадию культивирования полипотентных стволовых клеток со стадии (b) для формирования эмбриональных телец. Предпочтительно, чтобы эти эмбриональные тельца культивировали в условиях для отбора нестин-позитивных нейрональных клеток-предшественников, например, посредством культивирования полипотентных стволовых клеток или эмбриональных телец в бессывороточной среде предпочтительно в течение 6-10 суток. В предпочтительных вариантах осуществления бессывороточная среда представляет собой бессывороточную среду ITSFn определенного состава, предпочтительно включающую в себя один или несколько растворимых факторов, выбранных из группы, состоящей из инсулина, селенита натрия, основного фактора роста фибробластов, трансферрина и фибронектина. Предпочтительно, чтобы бессывороточная среда ITSFn определенного состава включала в себя инсулин, селенит натрия, трансферрин и фибронектин. В предпочтительных вариантах осуществления этими способами получают нейрональные клетки-предшественники, предпочтительно включающие в себя по меньшей мере приблизительно 60-75% нестин-позитивных клеток, более предпочтительно приблизительно 80-90% нестин-позитивных клеток и наиболее предпочтительно приблизительно 95-99% нестин-позитивных клеток.
В определенных вариантах осуществления указанные выше способы дополнительно включают в себя стадию наращивания нестин-позитивных нейрональных клеток-предшественников со стадии (c) в ростовой среде для клеток ЦНС предпочтительно в течение 6-10 суток. Предпочтительно, если ростовая среда для клеток ЦНС включает в себя один или несколько растворимых факторов, выбранных из группы, состоящей из добавки N2, добавки B27 и средства индукции нейронов. В предпочтительном варианте осуществления средство индукции нейронов представляет собой основной фактор роста фибробластов (bFGF). В еще одном предпочтительном варианте осуществления нестин-позитивные нейрональные клетки-предшественники вносят в культуральные чашки, предварительно покрытые поли-L-орнитином, поли-L-ламинином или их сочетанием. Нестин-позитивные нейрональные клетки-предшественники можно наращивать в культуре и последовательно пассировать в течение одного или нескольких удвоений популяций. Также эти клетки можно замораживать в жидком азоте.
Предпочтительно, чтобы нестин-позитивные нейрональные клетки-предшественники выращивали в среде для дифференцировки в течение 2 или более суток, как описано в стадии (d) указанных выше способов. Предпочтительно, чтобы среда для дифференцировки также включала в себя добавку N2, добавку B27 или обе эти добавки, но не основной фактор роста фибробластов (bFGF). В предпочтительных вариантах осуществления после этого нейрональные клетки-предшественники выращивают в среде для дифференцировки, содержащей цитозин-β-d-арабинофуранозид (Ara-C), дополнительно клетки культивируют в среде для дифференцировки без Ara-C предпочтительно в течение 8-16 суток, более предпочтительно в течение 12 суток.
Также в настоящем описании представлены способы образования ГАМКергических нейронов из нейрональных клеток-предшественников, включающие в себя обогащение нейрональных клеток-предшественников нестин-позитивными клетками и дифференцировку нестин-позитивных клеток с образованием ГАМКергических нейронов посредством культивирования клеток в присутствии Ara-C. Затем нестин-позитивные клетки культивируют в среде для дифференцировки с Ara-C, затем их предпочтительно дифференцируют в среде для дифференцировки без Ara-C. Предпочтительно, чтобы с применением этих способов по меньшей мере приблизительно 40-99% нестин-позитивных клеток дифференцировались в ГАМКергические нейроны.
В предпочтительных вариантах осуществления описанные выше способы применяют для образования дифференцированных популяций клеток, предпочтительно включающих в себя приблизительно 60-80% ГАМКергических нейронов, более предпочтительно приблизительно 75-90% ГАМКергических нейронов и наиболее предпочтительно 95-99% ГАМКергических нейронов. В других вариантах осуществления эти способы применяют для образования дифференцированных популяций клеток, предпочтительно включающих в себя приблизительно 15-30% дофаминергических нейронов, более предпочтительно приблизительно 20-40% дофаминергических нейронов и наиболее предпочтительно приблизительно 25-50% дофаминергических нейронов. В определенных вариантах осуществления эти способы применяют для образования дифференцированных популяций клеток, предпочтительно включающих в себя приблизительно 5-20% серотонинергических или глутаматергических нейронов, более предпочтительно приблизительно 10-25% серотонинергических или глутаматергических нейронов и наиболее предпочтительно приблизительно 15-30% серотонинергических или глутаматергических нейронов. В других вариантах осуществления эти способы применяют для образования дифференцированных популяций клеток, предпочтительно включающих в себя приблизительно 5-20% олигодендроцитов или астроцитов, более предпочтительно приблизительно 10-25% олигодендроцитов или астроцитов и наиболее предпочтительно приблизительно 15-30% олигодендроцитов или астроцитов.
Кроме того, настоящее изобретение представляет модель трансплантации in vitro для изучения эффективности, выживаемости и функционирования дифференцированных клеток, предпочтительно нейрональных или нервных клеток, в окружении, подобном хозяйскому, таком как окружение головного мозга. Например, описанные здесь ГАМКергические нейроны культивируют со зрелыми клетками головного мозга, предпочтительно нервными клетками, или клетками гиппокампа. Предпочтительно ГАМКергические нейроны переносят на клетки гиппокампа взрослой особи. Клетки культивируют вместе в течение по меньшей мере 3-20 суток, более предпочтительно 1 неделю, и определяют выживаемость нейрональных или нервных клеток. Предпочтительно, когда выживает по меньшей мере приблизительно 80% нейрональных или нервных клеток, более предпочтительно, когда выживает 90-99% клеток. Высокая степень выживаемости показывает, что клетки подходят для функционирования в окружении головного мозга взрослой особи, и их можно использовать для лечения нейродегенеративных нарушений или нервных заболеваний.
Также настоящее описание связано со способами лечения субъектов с нейродегенеративными нарушениями или нервными заболеваниями посредством введения субъекту нейрональных клеток-предшественников или дифференцированных нейрональных клеток, полученных из полипотентных стволовых клеток, например, как здесь описано, мышиных или человеческих ЭС клеток. Полученные здесь клетки можно применять также в клеточной заместительной терапии у человека. Например, популяцию дифференцированных нейрональных клеток можно получить следующим образом:
(a) наращивая культуру полипотентных стволовых клеток;
(b) культивируя полипотентные стволовые клетки для отбора нестин-позитивных нейрональных клеток-предшественников;
(c) наращивая нестин-позитивные нейрональные клетки-предшественники; и
(d) дифференцируя нестин-позитивные клетки для получения дифференцированной популяции клеток посредством культивирования клеток в среде для дифференцировки, включающей в себя цитозин-β-d-арабинофуранозид (Ara-C).
Согласно еще одному предпочтительному варианту осуществления дополнительно способствуют дифференцировке дифференцированных клеток со стадии (d) в среде для дифференцировки без Ara-C. В предпочтительных вариантах осуществления субъектом является пациент, более предпочтительно человек. Предпочтительно полученные из полипотентных стволовых клеток нейрональные клетки-предшественники или дифференцированные нейрональные клетки гистосовместимы с субъектом, например, если нейрональные клетки-предшественники или дифференцированные нейрональные клетки имеют, по существу, тот же геном, что и субъект.
В определенных вариантах осуществления ГАМКергические, дофаминергические, серотонинергические и глутаматергические нейроны, а также глиальные клетки, такие как олигодендроциты и астроциты, выделяют из дифференцированной нейрональной популяции клеток и вводят пациенту. В предпочтительном варианте осуществления субъекту вводят ГАМКергические нейроны. Эти клетки, а также популяции нейрональных клеток-предшественников или дифференцированных нейрональных клеток можно вводить субъекту для лечения ряда нейродегенеративных нарушений или нервных заболеваний, включая в качестве неограничивающих примеров инсульт, ишемию головного мозга, эпилепсию, болезнь Паркинсона, болезнь Гентингтона, болезнь Альцгеймера, повреждение спинного мозга, боковой амиотрофический склероз (ALS), эпилепсию и другие расстройства ЦНС, а также хроническую боль, тревожность и другие расстройства настроения. Этих субъектов также можно лечить посредством клеточной заместительной терапии. Предпочтительно клетки вводят посредством трансплантации, например посредством трансплантации желательных клеток в головной мозг субъекта.
Еще один вариант осуществления настоящего изобретения представляет собой способ лечения субъекта с нейродегенеративным расстройством или нервным заболеванием, включающий в себя следующие стадии:
(a) наращивание культуры полипотентных стволовых клеток;
(b) культивирование полипотентных стволовых клеток для отбора нестин-позитивных нейрональных клеток-предшественников;
(c) наращивание нестин-позитивных нейрональных клеток-предшественников;
(d) дифференцировку нестин-позитивных клеток для получения дифференцированной популяции клеток посредством культивирования клеток в среде для дифференцировки, включающей в себя цитозин-β-d-арабинофуранозид (Ara-C); и
(e) трансплантацию терапевтически эффективного количества дифференцированной нейрональной популяции клеток в центральную нервную систему пациента.
В предпочтительном варианте осуществления полипотентные стволовые клетки представляют собой мышиные или человеческие ЭС клетки. В других предпочтительных вариантах осуществления субъектом является пациент, более предпочтительно человек. Предпочтительно, когда дифференцированная популяция нейрональных клеток является гистосовместимой с пациентом. В еще одном варианте осуществления стадия (d) дополнительно включает в себя дифференцировку клеток в среде для дифференцировки в течение 2 или более суток и последующую дифференцировку клеток во второй, не содержащей цитозин-β-d-арабинофуранозид (Ara-C) среде для дифференцировки. В предпочтительных вариантах осуществления ГАМКергические нейроны выделяют из дифференцированной популяции нейрональных клеток и вводят субъекту, например, в головной мозг предпочтительно посредством трансплантации. В определенных вариантах осуществления нейродегенеративное расстройство или нервное заболевание выделяют из группы, состоящей из болезни Паркинсона, болезни Альцгеймера, болезни Гентингтона, деменции с тельцами Леви, рассеянного склероза, мозжечковой атаксии, прогрессирующего супрануклеарного паралича, повреждения спинного мозга, бокового амиотрофического склероза (ALS), эпилепсии, инсульта и ишемии.
В других вариантах осуществления популяцию нейрональных клеток-предшественников или дифференцированных нейрональных клеток, полученных из полипотентных стволовых клеток, как здесь описано, можно использовать для скрининга соединений, например небольших молекул и лекарственных средств, по их воздействию на популяцию клеток, особенно дифференцированных нервных или глиальных клеток, или активность этих клеток. Также соединения можно проверять на токсичность или модулирование нервных клеток. Например, соединение можно оценить посредством добавления соединения к популяции дифференцированных нервных клеток, таких как ГАМКергические нейроны, и сравнить выживаемость, морфологию, фенотип, функциональную активность или другие характеристики клеток с дифференцированными нервными клетками, культивируемыми в сходных условиях, но не подвергавшимися воздействию соединения. Соединения можно подвергать скринингу, например, чтобы определить, вызывают ли они изменения в синтезе, высвобождении или накоплении нейромедиатора клетками.
Еще один вариант осуществления настоящего описания связан со способом создания in vitro модели трансплантации нервных клеток, включающим в себя следующие стадии:
(a) выделение зрелых клеток гиппокампа;
(b) диссоциацию и культивирование клеток гиппокампа для образования культуры клеток гиппокампа; и
(c) культивирование нервных клеток на культуре клеток гиппокампа;
где оценивают выживаемость нервных клеток на культуре клеток гиппокампа. В других вариантах осуществления оценивают формирование синапсов между нервными клетками и культурой клеток гиппокампа. В одном варианте осуществления настоящего изобретения зрелые клетки гиппокампа выделяют из мыши. В определенных вариантах осуществления культивируемые на культуре клеток гиппокампа нервные клетки представляют собой, например, ГАМКергические, дофаминергические, серотонинергические или глутаматергические нейроны. В других вариантах осуществления на культуре клеток гиппокампа культивируют глиальные клетки, такие как олигодендроциты или астроциты. Культивируемые на культуре клеток гиппокампа нервные клетки или глиальные клетки можно получать с применением описанных здесь способов или же путем получения или выделения посредством других способов, хорошо известных специалистам в данной области. Предпочтительно, чтобы по меньшей мере приблизительно 50% нервных клеток, культивируемых на культуре клеток гиппокампа, выживали в течение по меньшей мере недели, более предпочтительно приблизительно 60%, 70%, 80%, 90% или 95%. В одном предпочтительном варианте осуществления после одной недели культивирования выживают более чем приблизительно 90% ГАМКергических нейронов.
Краткое описание некоторых чертежей
Следующие ниже чертежей составляют часть настоящего описания изобретения и включены в настоящее описание для дополнительной демонстрации конкретных аспектов настоящего изобретения, которое можно легче понять со ссылкой на один или несколько из этих чертежей в сочетании с детальным описанием представленных здесь конкретных вариантов осуществления.
На фиг.1 показано схематическое изображение получения ГАМКергических нейронов из мышиных ЭС клеток, а также пересадка in vitro ГАМКергических нейронов с полученными из зрелого мышиного головного мозга нейронами гиппокампа.
На фиг.2 показано схематическое изображение стадий дифференцировки мышиных ЭС клеток в нейрональные клетки в конечной стадии дифференцировки, включая (1) размножение недифференцированных клеток; (2) формирование эмбриональных телец; (3) отбор нестин-позитивных клеток; (4) размножение нестин-позитивных клеток; и (5) дифференцировку нейральных предшественников в окончательно дифференцированные нейрональные клетки.
На фиг.3 показана локализация следующих маркеров посредством иммунореактивности в нейрональной популяции, полученной из мышиных ЭС клеток: (a) GAD-65 и GAD-67; (b) GAT-1 и GAT-2; (c) глутамат и ГАМК; и (d) нестин и MAP-2. Иммунореактивность изучали на 4 различных стадиях: размножение нестин-позитивных клеток на 8, 12 и 16 сутки после дифференцировки.
На фиг.4 показана солокализация ГАМК и GAD-65/GAD-67 иммунореактивности в ГАМКергических нейронах, полученных из мышиных ЭС клеток с применением описанных здесь способов. Присутствие ГАМК иммунофлуоресценции в GAD-65 и GAD-67 позитивных клетках подтверждает, что эти ГАМКергические нейроны продуцируют ГАМК.
На фиг.5 показан профиль экспрессии генов ГАМКергических нейрон-специфических факторов в мышиных ЭС клетках и на разных стадиях во время дифференцировки in vitro этих клеток в ГАМКергические нейроны. Этими стадиями являются недифференцированные мышиные ЭС клетки (Un), эмбриональные тельца (EB), нестин-отбор (N), нестин-размножение (NE), 2 сутки дифференцировки (D-2); 4 сутки дифференцировки (D-4); 8 сутки дифференцировки (D-8); 12 сутки дифференцировки (D-12); и 16 сутки дифференцировки (D-16). Анализировали экспрессию следующих ГАМКергических нейрон-специфических генов: GAD1, GAD2, VIAAT и транскриптов эмбриональной GAD1. Экспрессию GAD1 наблюдали на всех стадиях, включая недифференцированные ЭС клетки. Экспрессию GAD2 наблюдали на стадиях отбора по нестину, размножения экспрессирующих нестин клеток и всех дифференцированных стадий, но экспрессию не обнаруживали на недифференцированной стадии и стадии эмбриональных телец. Экспрессию VIAAT наблюдали только на стадиях дифференцировки, особенно на 8 сутки и 12 сутки дифференцировки. Интересно, что VIAAT не экспрессировался после 16 суток дифференцировки. Экспрессию гена эмбриональной GAD1 наблюдали на всех стадиях, за исключением недифференцированной стадии. В качестве положительного контроля на всех стадиях определяли ген домашнего хозяйства β-актин.
Фиг.6 представляет собой сравнительный анализ нейрональных популяций, полученных из мышиных ЭС клеток. Суммарные количества клеток на различных стадиях получения считали с применением морфометрического анализа. Анализируемыми стадиями получения являлись размножение (a) нестин-позитивных клеток (NE); (b) 8 сутки дифференцировки; (c) 12 сутки дифференцировки; и (d) 16 сутки дифференцировки. Клетки считали по (1) общему количеству нейронов; (2)экспрессии глутамата; (3) экспрессии ГАМК; (4) экспрессии тирозингидроксилазы (TH); и (5) экспрессии HT.
На фиг.7 показаны определяемые посредством высокоэффективной жидкостной хроматографии с обращенной фазой (RP-HPLC) уровни внеклеточной ГАМК на следующих стадиях: размножения нестин-позитивных клеток (NE); на 8-е сутки дифференцировки (D-8); на 12-е сутки дифференцировки (D-12); и на 16-е сутки дифференцировки (D-16). ГАМК определяли только на дифференцированных стадиях, что подтверждает продукцию ГАМК нейронами, полученными из мышиных ЭС клеток с применением описанных здесь способов.
На фиг.8 показана иммунореактивность клеток, полученных из мышиных ЭС клеток, с применением описанных здесь способов после 12 суток дифференцировки с антителами к рецептору ГАМК-А. Приблизительно 80% клеток являются позитивными по иммунофлуоресценции с антителами к рецептору ГАМК-А.
На фиг.9 показана иммунореактивность клеток, полученных из мышиных ЭС клеток, с применением описанных здесь способов после 12 суток дифференцировки с антителами к рецептору ГАМК-Б. Приблизительно 25% клеток являются позитивными по иммунофлуоресценции с антителами к рецептору ГАМК-Б.
Подробное описание изобретения
Настоящее описание представляет способы эффективного получения клеток нейрального происхождения, дифференцированных из полипотентных стволовых клеток. Полученные здесь клетки включают в себя в качестве неограничивающих примеров клетки с фенотипическими характеристиками нейрональных клеток-предшественников, ГАМКергических, дофаминергических, серотонинергических и глутаматергических нейронов, а также глиальные клетки, такие как олигодендроциты и астроциты. Полученные здесь клетки идентифицируют посредством фенотипических характеристик, морфологических характеристик и/или клеточных маркеров, хорошо известных специалистам в данной области, оценивающих такие клетки. Как употребляют здесь, термин «нейрональные клетки-предшественники» является равнозначным терминам нейральные или нейрональные клетки-родоначальники, а также нейральные или нейрональные клетки-предшественники и относится к клетке, образующей потомство, представляющее собой либо нейрональные клетки, такие как нейрональные предшественники, нервные клетки, или нейроны, или глиальные клетки, такие как глиальные предшественники, астроциты или олигодендроциты. Здесь описывают способы, включающие в себя культивирование клеток при сочетании растворимых факторов и окружающих условий, стимулирующих клетки к дифференцировке в клетки нейрального происхождения. Предпочтительно здесь описывают способы, применяемые для получения ГАМКергических нейронов из полипотентных стволовых клеток.
Эти предшественники и дифференцированные нервные клетки можно применять в ряде приложений, включающих в себя терапевтические и экспериментальные приложения, а также разработке и скрининге in vitro лекарственных средств, таких как скрининг соединения на токсичность для нервных клеток или способность модулировать функцию нейрональных клеток. Получение предшественников и дифференцированных нервных клеток, таких как ГАМКергические нейроны, а также других специализированных типов нейрональных клеток, из полипотентных стволовых клеток предоставляет потенциально неограниченный запас этих нейронов с огромной потенциальной выгодой для индивидов, страдающих истощающими нейродегенеративными нарушениями и нервными заболеваниями, включающими в себя в качестве неограничивающих примеров инсульт, ишемию, болезнь Гентингтона, эпилепсию, хроническую боль, тревогу и другие расстройства настроения. Описанные здесь предшественники и дифференцированные нервные клетки обычно представляют собой потомство популяции клеток, из которых они получены, и, следовательно, имеют тот же геном, что и родительская популяция, включающая в себя генетически измененную, трансформированную или трансфицированную родительскую популяцию.
Предпочтительный вариант осуществления настоящего описания относится к улучшению способов получения ГАМКергических нейронов из полипотентных стволовых клеток, предпочтительно эмбриональных стволовых (ЭС) клеток млекопитающих или эмбриональных зародышевых (ЭЗ) клеток млекопитающих. В особенно предпочтительных вариантах осуществления ЭС или ЭЗ клетки представляют собой мышиные или человеческие ЭС или ЭЗ клетки. Эти нейроны получают из полипотентных стволовых клеток посредством культивирования клеток в присутствии определенных растворимых факторов и окружающих условий.
Как применяют здесь, термин «ГАМКергические нейроны» относится к нейрональным клеткам, экспрессирующим, продуцирующим или секретирующим нейромедиатор ГАМК. Предпочтительно конечная дифференцировка включает в себя активацию и регуляцию генов, необходимых для синтеза ГАМК, а также формирование и высвобождение везикул. В других предпочтительных вариантах осуществления ГАМКергические нейроны экспрессируют рецепторы ГАМК-А и ГАМК-Б. ГАМК принадлежит к химическому семейству декарбоксилазы глутаминовой кислоты (GAD), и GAD представляет собой ключевой фермент в синтезе ГАМК. Виды млекопитающих экспрессируют две изоформы GAD, обозначенные GAD1 и GAD2, уровни экспрессии которых различаются в различных областях головного мозга. GAD1 и GAD2 также известны как GAD67 и GAD65 соответственно, указывающие на их относительную молекулярную массу в кДа. Так как GAD65 и GAD67 представляют собой ферменты, синтезирующие ГАМК, их можно применять в качестве маркеров для идентификации ГАМКергических нейронов. Кроме того, везикулярный транспортер ингибирующих аминокислот (VIAAT) необходим для синаптической упаковки ГАМК и также является маркером для идентификации ГАМКергических нейронов. Патогенез некоторых нейродегенеративных нарушений и нервных заболеваний обнаруживает потерю ГАМКергических нейронов или снижение ГАМКергической нейропередачи, включающих в себя некоторые формы инсульта, ишемии, болезни Гентингтона, эпилепсии, хронической боли, тревоги и других расстройств настроения.
Настоящее описание относится к улучшенным способам дифференцировки полипотентных стволовых клеток в нейрональные клетки-предшественники, а также в дифференцированную популяцию нервных клеток с фенотипическими, молекулярными и/или клеточными характеристиками, сходными с клетками нейронального происхождения. В предпочтительном варианте осуществления полипотентные стволовые клетки представляют собой мышиные или человеческие ЭС клетки, дифференцированные с применением конкретных условий культивирования в нервные клетки, предпочтительно ГАМКергические нейроны. Настоящее описание также относится к полученным посредством описанных способов клеткам или клеточным популяциям. В определенных вариантах осуществления описанные способы включают в себя следующие стадии:
1. Выделяют популяцию полипотентных стволовых клеток; предпочтительно полипотентные стволовые клетки представляют собой мышиные или человеческие ЭС клетки.
2. Наращивают полипотентные стволовые клетки для предоставления исходного материала.
3. Полипотентные стволовые клетки культивируют в суспензии для получения эмбриональных телец.
4. Эмбриональные тельца повторно наносят на субстрат и инкубируют в селективной для нейрональных клеток-предшественников бессывороточной среде.
5. Наращивают нейрональные клетки-предшественники в среде роста, включающей в себя растворимые факторы, относящиеся к нервной системе.
6. Нейрональные клетки-предшественники дифференцируются в зрелые нейроны в среде для дифференцировки, предпочтительно среде, включающей в себя сочетание растворимых факторов, относящихся к нервной системе, а также цистеин-β-d-арабинофуранозид (Ara-C).
Источники полипотентных стволовых клеток
Здесь описывают способы для дифференцировки клеток нейронального происхождения из полипотентных стволовых клеток, включающие в себя применение конкретных культуральных условий, направляющих дифференцировку необычно высокого относительного количества полипотентных стволовых клеток в конкретные нейрональные типы клеток. Полипотентные стволовые клетки получают из пред-эмбриональных, эмбриональных или фетальных тканей в любое время после оплодотворения, которые при соответствующих условиях способны дифференцироваться в несколько различных типов клеток, являющихся производными всех трех зародышевых листков (энтодермы, мезодермы и эктодермы). Также клетки нейронального происхождения можно получать из стволовых клеток, выделенных из фетальной или зрелой ткани, способной к дифференцировке или перепрограммированию в клетки нейронального происхождения. Полипотентные стволовые клетки включают в себя в качестве неограничивающих примеров ЭС и ЭЗ клетки млекопитающих, предпочтительно мышиные ЭС и ЭЗ клетки или ЭС и ЭЗ клетки приматов или человека. Предпочтительно недифференцированные полипотентные стволовые клетки способны бесконечно делиться и пролиферировать в культуре. Как применяют здесь, термин "дифференцировка" относится к процессу, в результате которого недифференцированные полипотентные стволовые клетки или клетки-предшественники приобретают более специализированное направление развития. Например, дифференцированная клетка имеет фенотип, характеризующий определенный тип клетки или ткань.
В предпочтительном варианте осуществления здесь применяют ЭС и ЭЗ линии клеток, полученные из внутренней клеточной массы бластоцисты. Эти бластоцисты можно выделять из полученных in vivo оплодотворенных преимплантационных эмбрионов или из оплодотворенных in vitro (IVF), например эмбрионов, оплодотворенных посредством обычного оплодотворения, внутриплазматической инъекции сперматозоида или пересадки ооплазмы. Человеческие бластоцисты получают от супружеских пар или доноров, добровольно передающих свои избыточные эмбрионы. Эти эмбрионы применяют для исследовательских целей после получения добровольно подписанного разрешения этих пар или доноров. Альтернативно, бластоцисты можно получать посредством переноса соматической клетки или ядра клетки в безъядерный ооцит человеческого происхождения или происходящий не от человека, который затем стимулируют для развития в стадию бластоцисты. Также можно применять замороженные бластоцисты, или происходить из эмбрионов, замороженных на ранней стадии и допущенных к продолжению развития до стадии бластоцисты эмбриона. Развитие бластоцисты и внутренней клеточной массы будет изменяться в соответствии с видами и хорошо известно специалистам в данной области.
Мышиные ЭС клетки можно получать in vitro из преимплантационных эмбрионов, таких как бластоцисты, с применением способов, хорошо известных специалистам в данной области, таких как стандартные иммунохирургические способы (Evans et al., Nature 292: 154-159, 1981; Martin, Proc. Natl. Acad. Sci. USA 78: 7634-7638, 1981, каждая включена в настоящее описание в качестве ссылки). Мышиные ЭЗ клетки можно получать из фетальных зародышевых клеток, снова применяя способы, хорошо известные специалистам в данной области (Matsui et al., Cell 70: 841-847, 1992, включенная в настоящее описание в качестве ссылки). Для поддержания мышиных ЭС клеток в недифференцированном состоянии предпочтительно клетки культивируют в присутствии фактора ингибирования лейкемии (LIF) на фидерных слоях из фибробластов (Williams et al., Nature 336: 684-687, 1988, включенная в настоящее описание в качестве ссылки).
ЭС клетки приматов или человека можно получать из бластоцисты с применением стандартных иммунохирургических способов, как сообщают в патентах США №№5843780 и 6200806, Thomson et al. (Science 282: 1145-1147, 1998) и Reubinoff et al. (Nature Biotech. 18: 399-403, 2000), каждая конкретно включена в настоящее описание в качестве ссылки. Хотя в описанных способах можно применять ЭС клетки, полученные любым рядом способов, известных специалисту в данной области, в предпочтительном варианте осуществления применяют ЭС клетки человека, полученные посредством уникального способа лазерной абляции (U.S. Serial №10/226711, конкретно включено в настоящее описание в качестве ссылки). Кратко, этим способом выделяют клетки из внутренней клеточной массы бластоцисты посредством лазерной абляции части прозрачной оболочки и трофэктодермы бластоцисты, формирующей отверстие или канал в бластоцисте, посредством которого можно отсасывать клетки внутренней клеточной массы. Затем эти клетки можно дополнительно культивировать для создания ЭС клеточных линий. Этот способ является предпочтительным, так как он позволяет выделять клетки внутренней клеточной массы без прохождения обычной трудоемкой иммунохирургической процедуры. Кроме того, полученные с применением этого способа ЭС клеточные линии можно выделять в отсутствие любых синтезированных животными антител и сыворотки, что минимизирует риск любой передачи микроорганизмов животных в ЭС клеточные линии. В еще одном варианте осуществления применяют человеческие ЭЗ клетки, выделенные из первичных половых клеток, присутствующих в человеческом фетальном материале (патент США №6090622 и Shamblott et al., 1998, Proc. Natl. Acad. Sci. USA. 95: 13726-13731, каждая конкретно включена в настоящее описание в качестве ссылки).
Предпочтительно ЭС клеточные линии можно поддерживать в культуре в недифференцированном состоянии в течение длительного периода времени, например более одного года, и сохранять нормальный эуплоидный кариотип. Человеческие ЭС клетки можно морфологически идентифицировать посредством высоких ядерно-цитоплазматических отношений, выделяющегося ядрышка и компактного колониеобразования, часто с различными краями клеток и колоний, часто более ровными, чем мышиные ЭС клетки. Также предпочтительно человеческие ЭС клетки иммунореактивны к маркерам человеческих полипотентных ЭС клеток, например SSEA-3, SSEA-4, GCTM-2 антигену и TRA 1-60, как описано у Thomson et al. (1998), Reubinoff et al. (2000), Buehr and Mclaren (1993), каждая конкретно включена в настоящее описание в качестве ссылки. Предпочтительно человеческие ЭС клетки также экспрессируют щелочную фосфатазу, а также OCT-4. В других вариантах осуществления ЭС клетки способны формировать эмбриональные тельца в неадгезивных культуральных условиях (патенты США №№5914268 и 6602711, каждый включен в настоящее описание в качестве ссылки). Эти эмбриональные тельца можно применять для получения дифференцированных производных энтодермального, мезодермального и эктодермального зародышевых листков, а также других желательных последовательностей клеточных поколений.
Полипотентные стволовые клетки, особенно ЭС и ЭЗ клетки, можно непрерывно размножать в условиях культивирования, поддерживающих клетки, по существу, в недифференцированном состоянии. ЭС клетки необходимо поддерживать в соответствующей клеточной плотности и многократно разделять и пересевать, так как частая замена культуральной среды предотвращает их дифференцировку. Что касается способов, относящихся к культуре клеток и культивированию ЭС клеток, исполнитель может обратиться к стандартным руководствам и обзорам, например: E. J. Robertson, "Teratocarcinomas and embryonic stem cells: A practical approach" ed., IRL Press Ltd. 1987; Hu and Aunins, 1997, Curr. Opin. Biotechnol. 8(2): 148-53; Kitano, 1991, Biotechnology 17: 73-106; Spier, 1991, Curr. Opin. Biotechnol. 2: 375-79; Birch and Arathoon, 1990, Bioprocess Technol. 10: 251-70; Xu et al., 2001, Nat. Biotechnol. 19(10): 971-4; and Lebkowski et al., 2001, Cancer J. 7 Suppl. 2: S83-93; каждый конкретно включен в настоящее описание в качестве ссылки.
Обычно ЭС клетки культивируют в среде для ЭС клеток на слое фидерных клеток. Слои фидерных клеток представляют собой клетки одного типа ткани, совместно культивируемые с ЭС клетками, и обеспечивают окружающую среду, в которой ЭС клетки могут расти без претерпевания существенной дифференцировки. Способы культивирования клеток на фидерных слоях хорошо известны специалистам в данной области (патенты США №№5843780 и 6200806, WO 99/20741, U.S. Serial №№09/530346 и 09/849022, WO 01/51616, каждый конкретно включен в качестве ссылки). Предпочтительно фидерный слой снижает, ингибирует или предотвращает дифференцировку ЭС клеток. Обычно фидерные слои представляют собой фидерный слой эмбриональных фибробластов человеческого или мышиного происхождения, например мышиные эмбриональные фибробласты, человеческие эмбриональные фибробласты, человеческие фибробласт-подобные клетки или мезенхимальные клетки, полученные из человеческих эмбриональных стволовых клеток или клеток STO.
Предпочтительно ЭС клетки культивируют в среде для ЭС клеток, снижающей, ингибирующей или предотвращающей дифференцировку ЭС клеток. Предпочтительно применяемая для культивирования ЭС клеток среда для ЭС клеток дополнена питательной сывороткой, например сывороткой или раствором на основе сыворотки, дополняющим питательные вещества, эффективные для поддержания роста и жизнеспособности ЭС клеток. Питательной сывороткой может быть сыворотка животного, такая как эмбриональная бычья сыворотка (FBS) или эмбриональная телячья сыворотка (FCS) (патенты США №№5453357 5670372 и 5690296, включенные в настоящее описание в качестве ссылки). Как применяют здесь, FBS можно применять вместо FCS, и наоборот. Также среда для ЭС клеток может быть бессывороточной (WO 98/30679, WO 01/66697, U.S. Serial №09/522030, каждый конкретно включен в настоящее описание в качестве ссылки). Примером пригодной среды для ЭС клеток с сывороткой для культивирования ЭС клеток является модифицированная Дульбекко среда Игла (DMEM) без пирувата натрия с высоким содержанием глюкозы (70-90%) (GIBCO), дополненная FBS или FCS (10-30%), β-меркаптоэтанолом (0,1 мМ), заменимыми аминокислотами (1%) и L-глутамином 2 мМ, 4 нг/мл основного фактора роста фибробластов (bFGF), 50 Ед/мл пенициллина и 50 мкг/мл стрептомицина. Также среда для ЭС клеток может включать в себя 1000 Ед/мл фактора ингибирования лейкемии (LIF). Пример пригодной бессывороточной среды для ЭС клеток для культивирования ЭС клеток представляет собой 80% нокаутирующую модифицированную Дульбекко среду Игла (DMEM) (GIBCO), 20% нокаутирующую SR (бессывороточный заменитель, GIBCO), β-меркаптоэтанола (0,1 мМ), заменимых аминокислот (1%) и L-глутамина 1 мМ.
Также ЭС клетки можно культивировать в культуральных условиях без фидерного слоя. Способы культивирования ЭС клеток в культуре без фидерного слоя хорошо известны специалистам в данной области (U.S. Publ. №2002/0022268, WO 03/020920, U.S. Serial №10/235094, каждый конкретно включен в настоящее описание в качестве ссылки). Предпочтительно в культуре без фидерного слоя ЭС клетки растят на пригодном культуральном субстрате, например внеклеточном матриксе, таком как MatrigelW (Becton Dickenson) или ламинин. Также предпочтительно для культур без фидера применяют кондиционированную среду для поддержания роста ЭС клеток. Кондиционированную среду получают посредством культивирования первичной популяции мышиных эмбриональных фибробластов или человеческих эмбриональных фибробластных клеток в среде в течение необходимого периода времени для продукции «кондиционированной» среды, которая поддержит культивирование ЭС клеток без значительной дифференцировки. Альтернативно, культуру без фидера можно комбинировать с внеклеточным матриксом с эффективной свежей средой, добавляемой в культуру без кондиционирования каким-либо другим типом клеток (U.S. Publ. №2003/0017589, конкретно включен в настоящее описание в качестве ссылки).
Получение нейрональных клеток-предшественников
Выделенные полипотентные стволовые клетки можно наращивать, а затем содержать их в условиях культивирования, что вызывает их дифференцировку в нейрональные клетки-предшественники. Клетки культивируют в соответствии с описанными здесь протоколами дифференцировки для продвижения полипотентных стволовых клеток только по пути нейрональной дифференцировки. Полипотентные стволовые клетки культивируют на пригодном субстрате в питательной среде для дифференцировки, содержащей средства для дифференцировки, такие как растворимые факторы и ростовые факторы. Пригодные субстраты включают в себя в качестве неограничивающих примеров покрытые положительными зарядами твердые поверхности, например поли-L-лизин или полиорнитин, субстраты, покрытые компонентами внеклеточного матрикса, например фибронектин, ламинин, PDGF, EGF, коллаген V, человеческую амниотическую мембрану или Matrigel®, или их сочетание. Предпочтительными питательными средами для дифференцировки являются те, которые поддерживают пролиферацию, дифференцировку и выживаемость желательных типов нервных клеток и могут включать в себя один или несколько пригодных средств для дифференцировки. Как применяют здесь, термин «ростовой фактор» относится к белкам, связывающимся с рецепторами на поверхности клеток с основным результатом активации клеточной пролиферации и дифференцировки. Пригодные растворимые факторы включают в себя в качестве неограничивающих примеров нейротрофины, митогены, факторы стволовых клеток, ростовые факторы, факторы для дифференцировки (например, суперсемейства TGF-β), агонисты суперсемейства TGF-β, нейротрофические факторы, антиоксиданты, нейромедиаторы и факторы выживаемости. Многие растворимые факторы являются достаточно универсальными, стимулируя клеточное деление в ряде различных типов клеток, тогда как другие являются специфичными для особых типов клеток.
Пригодные средства для дифференцировки, которые особенно поддерживают дифференцировку нейрональных типов клеток, включают в себя в качестве неограниченных примеров прогестерон, путресцин, ламинин, инсулин, селенит натрия, трансферрин, нейртурин, sonic hedgehog (SHH), ноггин, фоллистатин, эпидермальный фактор роста (EGF), любой тип ростовых факторов фибробластов, цитозин-β-d-арабинофуранозид (Ara-C), фактор роста и дифференцировки 5 (GDF-5), члены семейства нейротрофинов (фактор роста нервов (NGF), нейротрофин 3 (NT-3), нейротрофин 4 (NT-4), выделенный из головного мозга нейротрофический фактор (BDNF)), трансформирующий фактор роста-α (TGF-α), трансформирующий фактор роста бета-3 (TGF β3), выделенный из тромбоцитов фактор роста (PDGF-AA), инсулиноподобный фактор роста (IGF-1), костные морфогенетические белки (BMP-2, BMP-4), выделенный из глиальных клеток нейротрофический фактор (GDNF), midkine, аскорбиновую кислоту, дибутирил цАМФ, дофамин и лиганды рецепторов, связанные с gpl30 (например, LIF, CNTF, SCF, IL-11 и IL-6). Как применяют здесь, термин «ростовой фактор фибробластов», или «FGF», относится к любому пригодному фактору роста фибробластов, выделенному из любого организма, экспрессирующего такие факторы и их функциональные фрагменты. Специалистам в данной области известно множество FGF, включающих в себя, но не ограниченных FGF-1 (кислым фактором роста фибробластов), FGF-2 (основным фактором роста фибробластов), FGF-3 (int-2), FGF-4 (hst/K- FGF), FGF-5, FGF-6, FGF-7, FGF-8 и FGF-9. Также питательные среды для дифференцировки могут содержать добавки, помогающие поддержать культуры нервных клеток, например добавки N2 и B27 (Gibco). Предпочтительно ни в каком из описанных здесь способов не применяют такие средства для дифференцировки, как ретиноевая кислота, 13-цис-ретиноевая кислота и транс-ретиноевая кислота.
Первая стадия дифференцировки полипотентных стволовых клеток включает в себя индукцию клеток к формированию эмбриональных телец. Эмбриональные тельца помещают прямо на пригодный субстрат с добавлением или без добавления компонента внеклеточного матрикса, такого как фибронектин или ламинин, и культивируют в пригодной питательной среде для дифференцировки, адаптированной для стимуляции дифференцировки в нейрональные клетки-предшественники, такие как нестин-позитивные нейрональные клетки-предшественники. Нестин представляет собой клеточный маркер, характеризующий нейрональные клетки-предшественники. В еще одном варианте осуществления сначала полипотентные стволовые клетки агрегируют в гетерогенную клеточную популяцию, формируя эмбриональные тельца, например, посредством культивирования полипотентных стволовых клеток в суспензии. Эти клетки можно культивировать в питательной среде с добавлением или без добавления сыворотки, а также с одним или несколькими перечисленными выше средствами для дифференцировки для стимулирования дифференцировки клеток в эмбриональные тельца. Предпочтительно полипотентные стволовые клетки культивируют в среде для ЭС клеток без LIF.
Как применяют здесь, термин «эмбриональные тельца» относится к агрегации дифференцированных клеток, полученных, когда полипотентные стволовые клетки растят в суспензионной культуре, или наращивают в монослойных культурах. Также эмбриональные тельца могут иметь в массе клеток недифференцированные клетки. Предпочтительно эта масса клеток окружена примитивной энтодермой. Обычно эмбриональные тельца содержат клетки, выделенные из всех трех зародышевых листков, эктодермы, мезодермы и энтодермы. В зрелых человеческих эмбриональных тельцах возможно различить клетки, несущие маркеры разных типов клеток, таких как нейрональные клетки, гематопоэтические клетки, клетки печени и клетки сердечной мышцы. Некоторые клетки в зрелых эмбриональных тельцах могут вести себя подобно функционально дифференцированным клеткам. Например, активные клетки сердечной мышцы могут заставлять пульсировать эмбриональное тельце. Предпочтительно дифференцировку полипотентных стволовых клеток контролируют таким образом, чтобы можно было получить конкретные типы клеток для терапевтических целей.
Эмбриональные тельца культивируют до тех пор, пока они не достигнут необходимого размера или желательной дифференцировки, например, через 3-10 суток культивирования, предпочтительно 4-8 суток, и затем наносят на субстрат. Предпочтительно субстрат покрыт компонентами внеклеточного матрикса, включающими в себя, но не ограниченными, поли-L-лизином, поли-L-орнитином, ламинином, коллагеном, фибронектином, Matrigel® или их сочетанием. Предпочтительно эмбриональные тельца наносят непосредственно на субстрат без диспергирования клеток. Затем эмбриональные тельца культивируют в условиях, стимулирующих дальнейшую дифференцировку нанесенных клеток в нейрональные или клетки-предшественники ЦНС. Например, эмбриональные тельца можно культивировать в описанной бессывороточной среде, селективной для нестин-позитивных клеток, такой как среда ITSFn. Альтернативно, наросшие полипотентные стволовые клетки можно наносить непосредственно на субстрат и культивировать в описанной бессывороточной среде для отбора нестин-позитивных клеток. Нестин представляет собой белок промежуточных филаментов, экспрессированный в нейроэпителии.
Предпочтительно описанная бессывороточная среда, применяемая для размножения эмбриональных телец, представляет собой DMEM:F12, дополненную одним или несколькими факторами роста, выбранными из группы, состоящей из прогестерона, путресцина, ламинина, инсулина, селенита натрия, трансферрина, фибронектина, FGF, SHH, EGF и BDNF. Более предпочтительно описанная бессывороточная среда представляет собой среду ITSFn, дополненную питательными веществами инсулином, селенитом натрия, трансферрином и фибронектином. Как правило, клетки растят в этих условиях в течение периода 5-16 суток, более предпочтительно в течение 7 суток. В предпочтительных вариантах осуществления отбор с описанной выше бессывороточной средой обогащает популяцию жизнеспособных нестин-позитивных клеток приблизительно до 40-70%, более предпочтительно приблизительно до 80%-90% и наиболее предпочтительно приблизительно до 95%-99%.
Затем полученные нейрональные клетки-предшественники или клетки-предшественники ЦНС наращивают в среде для роста клеток ЦНС. Как применяют здесь, термины «наращивание» или «размножение» относят к процессу, посредством которого количество клеток увеличивается благодаря клеточному росту и делению. Термин «пролиферировать» можно применять взаимозаменяемо с «наращивание» или «размножение». Предпочтительно среда для роста клеток ЦНС включает в себя минимальную необходимую среду, такую как DMEM:F12, и дополнена добавками, помогающими поддерживать культуры нейрональных клеток, например добавки N2 и B27. Также предпочтительно среда для роста клеток ЦНС включает в себя одно или несколько индуцирующих нейроны средств для стимуляции пролиферации клеток-предшественников ЦНС и повышения эффективности образования ГАМКергических нейронов, например, основной фактор роста фибробластов (bFGF), а также другие факторы, контролирующие развитие ГАМКергических нейронов при эмбриогенезе in vivo. Предпочтительно нейрональные клетки-предшественники или клетки-предшественники ЦНС растят в среде для роста клеток ЦНС в течение 4-10 суток. Кроме того, предпочтительно клетки наносят на поверхность, предотвращающую адгезию нейрональных клеток-предшественников или клеток-предшественников ЦНС, такую как поверхности, покрытые поли-L-лизином, поли-L-орнитином, ламинином, коллагеном, фибронектином, Matrigel® или их сочетанием.
По одному варианту осуществления эмбриональные тельца получают из мышиных ЭС клеток посредством культивирования клеток на бактериологической чашке в отсутствие фидерных клеток в соответствующей среде. Предпочтительно мышиные ЭС клетки сначала диссоциируют, например, подвергая воздействию трипсина, с последующим соскабливанием и разбивкой клеток на небольшие кластеры. Затем эти кластеры в соответствующей плотности наносят на бактериологические чашки, предпочтительно имеющие неадгезивную поверхность, что предотвращает прикрепление клеток, стимулируя таким образом дифференцировку и образование эмбриональных телец. Клетки культивируют в соответствующей среде, например в среде для ЭС клеток, предпочтительно включающей в себя DMEM с высоким содержанием глюкозы или нокаут DMEM, дополненной 10-20% FCS, FBS или нокаутирующим заменителем сыворотки, а также другими добавками, такими как β-меркаптоэтанол, L-глутамин (2 мМ) и антибиотики. Среду меняют, по меньшей мере, каждые сутки, и эмбриональные тельца растят предпочтительно в течение 4-8 суток.
По еще одному варианту осуществления эмбриональные тельца получают из человеческих ЭС клеток посредством культивирования клеток на бактериологической чашке в отсутствие фидерных клеток в соответствующей среде. Предпочтительно человеческие ЭС клетки диссоциируют в кластеры и затем вносят в неадгезивные чашки, чтобы способствовать развитию эмбриональных телец. Предпочтительно соответствующая среда включает в себя DMEM с высоким содержанием глюкозы и дополнена 10-20% FCS. В среду также могут быть добавлены другие добавки, такие как 0,1 мМ 2-меркаптоэтанол, 2 мМ L-глутамина, 50 Ед/мл пенициллина и 50 мкг/мл стрептомицина.
После выделения эмбриональных телец их повторно вносят в культуральный планшет, покрытый от 0,1% до 0,2% желатина в бессывороточной среде для выделения нейрональных клеток-предшественников и клеток-предшественников ЦНС, предпочтительно нестин-позитивных клеток. Предпочтительно бессывороточная среда представляет собой базальную среду, такую как DMEM:F12, дополненную факторами роста. Среда ITSFn является средой для отбора нестин-позитивных клеток, включающая в себя базальную среду DMEM:F12 (1:1) или среду IMDM, дополненную факторами роста: инсулином, селенитом натрия, трансферрином и фибронектином.
Предпочтительно эти нейрональные клетки-предшественники или клетки-предшественники ЦНС, предпочтительно нестин-позитивные клетки, затем культивируют в среде для роста клеток ЦНС, содержащей индуцирующие нейроны факторы роста, выбираемые для нейрональных предшественников, предпочтительно ГАМКергических нейрональных предшественников. Один пример среды для роста клеток ЦНС представляет собой среду DMEM:F12, дополненную N2, B27 и bFGF. Нейрональные клетки-предшественники или клетки-предшественники ЦНС также повторно вносят в еще одну культуральную чашку, предварительно покрытую внеклеточными матриксами, например поли-L-лизином, поли-L-орнитином, ламинином, коллагеном или их сочетанием. Несмотря на отсутствие желания быть связанными конкретным механизмом, полагают, что эти различные присутствующие в среде для роста клеток ЦНС факторы способствуют суммарному повышению доли нейрональных клеток и в дальнейшем индуцируют дифференцировку фенотипа ГАМКергического нейрона. В предпочтительных вариантах осуществления нестин-позитивные клетки-предшественники растят в среде для роста клеток ЦНС в течение 5-8 суток.
Дифференцировка ГАМКергических нейронов
Полученные по описанным здесь способам нейрональные клетки-предшественники затем могут дифференцироваться в большие количества зрелых нейронов, например ГАМКергические нейроны, а также дофаминергические, серотонинергические и глутаматергические нейроны. Также нейрональные клетки-предшественники затем могут дифференцироваться в глиальные клетки, такие как олигодендроциты или астроциты. Окончательной дифференцировки нейрональных клеток-предшественников или клеток-предшественников ЦНС достигают посредством культивирования клеток в соответствии с описанными здесь протоколами дифференцировки.
Предпочтительно нестин-позитивные клетки-предшественники или клетки-предшественники ЦНС растят 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, или 20 суток в среде для дифференцировки, способствующей дифференцировке нейрональных клеток-предшественников или клеток-предшественников ЦНС в окончательно дифференцированные нервные клетки, или зрелые нейроны. Кроме того, можно облегчить дифференцировку посредством применения некоторых или всех факторов роста, таких как β-FGF, стимулирующих дифференцировку, пролиферацию нейрональных клеток-предшественников или клеток-предшественников ЦНС или и то, и другое. Например, выросшие нейрональные клетки-предшественники или клетки-предшественники ЦНС можно дифференцировать посредством культивирования клеток в среде для дифференцировки, включающей в себя среду DMEM:F12 или среду Neurobasal A, дополненную FCS, N2, B27 или их сочетанием с добавлением или без β-FGF. Также для усиления дифференцировки или выхода ГАМКергических нейронов среда для дифференцировки может содержать множество дополнительных факторов, например Ara-C. По одному предпочтительному варианту осуществления нейрональные клетки-предшественники или клетки-предшественники ЦНС культивируют в содержащей Ara-C среде для дифференцировки в течение одних или нескольких суток и затем культивируют в среде для дифференцировки без Ara-C в течение 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 суток. По еще одному предпочтительному варианту осуществления нейрональные клетки-предшественники или клетки-предшественники ЦНС культивируют в среде для дифференцировки, включающей в себя среду DMEM:F12, дополненную N2 и B27, в течение одних суток; затем культивируют в среде для дифференцировки, содержащей среду DMEM:F12, дополненную 5% FBS, B27 и Ara-C, в течение двух суток; и в заключение культивируют в той же среде для дифференцировки без Ara-C в течение 2-16 суток, предпочтительно 12 суток.
Предпочтительно большое количество нейрональных клеток-предшественников дифференцируются в ГАМКергические нейроны, например, по меньшей мере приблизительно 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% клеток. По одному предпочтительному варианту осуществления полипотентные стволовые клетки, выращенные и дифференцированные в соответствии с описанными здесь способами, приводят к образованию большого количества (по меньшей мере приблизительно 60%) ГАМКергических нейронов. Кроме того, затем ГАМКергические нейроны можно очистить из популяции дифференцированных нервных клеток посредством хорошо известных специалистам в данной области способов, таких как иммунное мечение и флуоресцентная сортировка, например твердофазная адсорбция, FACS, MACS и т.п. Другие полученные здесь дифференцированные нервные клетки, например дофаминергические, серотонинергические и глутаматергические нейроны, а также олигодендроциты и астроциты, также можно выделять с применением сходных способов.
Применения для нейрональных клеток-предшественников и дифференцированных нервных клеток
Описанные здесь нейрональные клетки-предшественники и дифференцированные нервные клетки (например, ГАМКергические, дофаминергические, серотонинергические и глутаматергические, а также олигодендроциты и астроциты) можно применять для различных приложений, таких как терапевтические применения, а также для определения и скрининга in vitro и in vivo различных соединений, таких как низкомолекулярные лекарственные средства по их эффектам на эти клетки. Также эти клетки можно применять для получения экспрессионных библиотек кДНК для анализа профилей экспрессии этих клеток, а также для получения моноклональных и поликлональных антител, специфичных для маркеров для конкретных применяемых клеток, с применением хорошо известных специалистам в данной области способов. Также эти клетки можно применять терапевтически для оказания помощи индивидам, страдающим истощающими нейродегенеративными нарушениями и нервными заболеваниями.
По настоящему описанию представлено применение описанных здесь нейрональных клеток-предшественников и дифференцированных нервных клеток для лечения или предотвращения различных нейродегенеративных нарушений и нервных заболеваний, при которых повреждаются или погибают нейроны или глиальные клетки центральной нервной системы (ЦНС) или спинного мозга. Нуждающихся в такой терапии субъектов будут лечить посредством терапевтически эффективного количества таких клеток для восстановления функций ЦНС или периферической нервной системы (ПНС). Как применяют здесь, «терапевтически эффективное количество» клеток представляет собой необходимое количество для угнетения или улучшения у субъекта физиологических эффектов, вызванных гибелью, повреждением или дегенерацией нервных клеток, таких как зрелые нейроны (например, ГАМКергические, дофаминергические и серотонинергические нейроны), астроциты и олигодендроциты. Например, эти клетки можно было бы терапевтически применять посредством их трансплантации непосредственно в паренхимные или интратекальные участки ЦНС, в зависимости от заболевания или условий лечения.
Эти клетки можно применять для лечения острого или хронического повреждения нервной системы, а также истощающих нейродегенеративных нарушений и нервных заболеваний, включающих в себя расстройства или заболевания нервной системы, включая ЦНС и ПНС. Нейродегенеративные нарушения и нервные заболевания включают в себя в качестве неограничивающих примеров болезнь Паркинсона, болезнь Альцгеймера, болезнь Гентингтона, деменцию с тельцами Леви, рассеянный склероз, мозжечковую атаксию, прогрессирующий супрануклеарный паралич, повреждение спинного мозга, боковой амиотрофический склероз (ALS), эпилепсию, инсульт, ишемию, повреждение или травму нервной системы, нейротоксическое повреждение и т.п. Также определенные неврологические расстройства можно лечить полученными из полипотентных стволовых клеток дифференцированными нервными клетками, например, связанные с узнаванием и психологией нарушения, включающие в себя, в качестве неограничивающих примеров, тревожные расстройства, расстройства настроения, пристрастия, обсессивно-компульсивные расстройства (OCD), изменения личности, синдром нарушения внимания (ADD), синдром дефицита внимания с гиперактивностью (ADHD) и шизофрению.
Один вариант осуществления настоящего описания относится к способам лечения или предотвращения нейродегенеративных нарушений или нервных заболеваний, характеризуемых дегенерацией или деструкцией ГАМКергических нейронов, посредством введения терапевтически эффективного количества полученных из полипотентных стволовых клеток ГАМКергических нейронов, предпочтительно мышиных или человеческих полипотентных стволовых клеток. Предпочтительно страдающего нейродегенеративным нарушением или нервным заболеванием пациента (человека) лечат пересадкой пациенту терапевтически эффективного количества нейрональных клеток-предшественников и дифференцированных нервных клеток настоящего описания. Если пациент страдает ишемией головного мозга или инсультом, предпочтительно введение терапевтически эффективного количества ГАМКергических нейронов приведет к уменьшению количества или тяжести симптомов, связанных с церебральной ишемией или инсультом, таких как потеря памяти, когнитивные расстройства или двигательные расстройства.
Терапевтически эффективное количество применяемых клеток будет зависеть от нужд субъекта, возраста субъекта, физиологического состояния и жизнеспособности, желательного терапевтического эффекта, размера предназначенного для лечения участка ткани, участка имплантации, степени патологии (например, уровня нейрональной дегенерации), выбранного пути доставки и стратегии лечения. Например, лечение поражающего больший участок головного мозга нарушения может потребовать большего количества клеток для достижения терапевтического эффекта по сравнению с меньшим участком поражения. Также клетки можно вводить в более чем один участок данной пораженной ткани с несколькими небольшими трансплантатами с низкими дозами клеток. Перед введением клетки по настоящему описанию можно полностью диссоциировать для создания клеточной суспензии одиночных клеток или перед введением почти полностью диссоциировать для создания небольших агрегатов клеток. Клетки можно вводить способом, позволяющим им трансплантироваться или мигрировать в предназначенный участок ткани и восстанавливать или регенерировать функционально неполноценный участок. Предпочтительно клетки применяют в аутологичной терапии, минимизируя или исключая проблемы иммунного отторжения после трансплантации, например, гистосовместимость с намеченным реципиентом. Альтернативно, клетки применяют для аллогенной терапии.
Подходящий разброс клеток, вводимых для достижения терапевтического эффекта, может составлять приблизительно от 100 до приблизительно 1000000 нейронов, предпочтительно приблизительно от 500 до приблизительно 500000 нейронов или приблизительно от 1000 нейронов до приблизительно 100000 нейронов. Количество вводимых клеток будет напрямую зависеть от числа клеток, выживающих при терапевтическом введении. Терапевтические концентрации вводимых субъекту нервных клеток могут колебаться приблизительно от 10, 100, 500, 1000, 5000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 150000, 200000, 250000, 300000, 350000, 400000, 450000 до приблизительно 500000 клеток в микролитре фармацевтически допустимого носителя. Разбросы концентраций клеток в носителе включают в себя, например, 100-50000 клеток/мкл, 1000-10000 клеток/мкл, 5000-25000 клеток/мкл, 15000-45000 клеток/мкл, 20000-50000 клеток/мкл, 55000-200000 клеток/мкл, 100000-40000 клеток/мкл, 150000-50000 клеток/мкл и т.д. Также количество перенесенных в участок трансплантации клеток окажет влияние на терапевтическую эффективность.
Для терапевтических применений часто предпочтительно, чтобы популяции предшественников или дифференцированных нервных клеток были в значительной степени очищенными от любых недифференцированных полипотентных стволовых клеток. Одна стратегия для выделения полипотентных стволовых клеток из терапевтического препарата представляет собой трансфекцию клеток вектором, имеющим ген, предпочтительно экспрессированный в недифференцированных клетках, экспрессия которого позволяет отбор от полипотентных стволовых клеток. Пригодные промоторы, предпочтительно экспрессированные в недифференцированных клетках, представляют собой промотор теломеразной обратной транскриптазы (TERT) и промотор OCT-4. Экспрессирующийся в векторе ген может являться, например, литическим для клетки, таким как токсин, или напротив, его можно выбрать противостоящим применению внешнего средства.
Способность образовывать ГАМКергические нейроны, а также дофаминергические и серотонинергические нейроны из полипотентных стволовых клеток, как описывают здесь, представляет собой огромную клиническую значимость для терапевтического предотвращения или лечения множества нейродегенеративных нарушений и нервных заболеваний. Например, ГАМКергические нейроны можно применять для лечения или предотвращения нейродегенеративных нарушений и нервных заболеваний, характеризуемых отклонениями в быстрых тормозных синаптических передачах, возбудимости нейронов и быстрых изменениях настроения, таких как порог судорожной активности, тревога, паника и ответ на стресс (т.е. ответ «нападение или бегство»), а также отклонения памяти, настроения или болевых функций. Например, ГАМКергические нейроны можно применять для лечения или предотвращения болезни Паркинсона, болезни Альцгеймера, эпилепсии, деменции с тельцами Леви, рассеянного склероза, мозжечковой атаксии, прогрессирующего супрануклеарного паралича, повреждения спинного мозга, бокового амиотрофического склероза (ALS) и болезни Гентингтона, а также инсульта, ишемии, церебральной ишемии, повреждения или травмы нервной системы, нейротоксического повреждения и т.п. Также ГАМКергические нейроны можно применять для лечения или предотвращения определенных неврологических нарушений, включающих в себя в качестве неограничивающих примеров связанные с узнаванием и психологией нарушения, такие как тревожные расстройства, расстройства настроения, привыкание, обсессивно-компульсивные расстройства (OCD), изменения личности, синдром нарушения внимания (ADD), синдром дефицита внимания с гиперактивностью (ADHD) и шизофрению.
Болезнь Паркинсона представляет собой двигательное расстройство, вызываемое прогрессирующей деградацией продуцирующих дофамин клеток в черном веществе среднего мозга. Выявлено, что терапия на основе клеток трансплантируемых дофаминергических нейронов в черное вещество пациентов с болезнью Паркинсона терапевтически эффективна, но симптоматическая помощь является недостаточной (Lindvall, O., 1997, Neuroreport. 8(14): iii-x). Следовательно, трансплантации дофаминергических нейронов может быть недостаточно для лечения болезни Паркинсона, и недавние исследования свидетельствуют, что существует еще один участок головного мозга, а именно субталамическое ядро (STN), вовлеченное в нейропатологию болезни Паркинсона (Bergman et al., 1998, Trends Neurosci. 21: 32-38; Luo et al., 2002, Science 298: 425-29). Субталамическая область головного мозга содержит глутаматергические и ГАМКергические нейроны (Nishino et al., 1988, Japn. J. Pharmacol. 48: 331-339). У пациентов с болезнью Паркинсона локализованные в STN нейроны дегенерируют вместе с нейронами черного вещества (During et al., 2001, Hum. Gene Ther. 12(12): 1589-91). Следовательно, глутаматергические и ГАМКергические нейроны также могут дегенерировать у пациентов с болезнью Паркинсона (Luo et al., 2002, Science 298(11): 425-429).
Недавние исследования свидетельствуют, что введение критичного для биосинтеза ГАМК фермента GAD в STN в модели болезни Паркинсона снижает моторные нарушения, связанные с болезнью Паркинсона (Luo et al., 2002, Science 298: 425-29). Следовательно, ГАМКергические нейроны также могут быть полезными при лечении болезни Паркинсона, например, посредством введения или трансплантации ГАМКергических нейронов в STN. Можно вводить или пересаживать только ГАМКергические нейроны или в ассоциации, или в сочетании с введением или трансплантацией дофаминергических нейронов в черное вещество. Также ГАМКергические нейроны можно применять для лечения болезни Паркинсона в сочетании с препаратами, полученными из растений, экстрактами на основе растений или синтетических источников с активностью против болезни Паркинсона, антинейродегенеративной или нейропротективной активностями.
Болезнь Альцгеймера включает в себя недостаточность преимущественно холинергических клеток в базальном ядре головного мозга. Хотя введение клеток или трансплантация холинергических клеток представляют собой эффективную терапию болезни Альцгеймера, этого может быть недостаточно для излечения заболевания, так как у пациентов с болезнью Альцгеймера в головном мозге, особенно в гиппокампе, также гибнут другие типы нервных клеток. Например, пациент с болезнью Альцгеймера страдает очень сильной потерей функции памяти, что отчасти соответствует гибели нейронов гиппокампа, большинство которых являются ГАМКергическими нейронами (Seidl et al., 2001, Arch. Pharmacol. 363: 139-145). Кроме того, обнаружено значительное снижение содержания ГАМК в височной коре, затылочной коре и мозжечке пациентов с болезнью Альцгеймера (Seidl et al., 2001). По одному варианту осуществления для лечения пациентов с болезнью Альцгеймера в область коры гиппокампа головного мозга вводят или пересаживают полученные из полипотентных стволовых клеток ГАМКергические нейроны, как описано здесь, отдельно или в сочетании с другими нейронами, такими как холинергические нейроны или дофаминергические нейроны.
Существует безусловная необходимость улучшения свойства клеток выдерживать различные нейродегенеративные нарушения и нервные заболевания, включающие в себя в качестве неограничивающих примеров болезнь Паркинсона и болезнь Альцгеймера, а также церебральную ишемию и инсульт. На дегенерацию и гибель нейронов оказывают влияние различные факторы. По предпочтительному варианту осуществления индуцирующие деградацию и гибель нейронов факторы, например внеклеточный кальций, избыточное высвобождение глутамата или высвобождение кислородных радикалов, блокируют до введения или пересадки в головной мозг пациента описанных здесь нейрональных клеток-предшественников или дифференцированных нервных клеток. Посредством блокировки этих факторов или противодействия им в участке введения или пересадки клеток процедуру может перенести большая доля клеток.
По другим вариантам осуществления настоящее изобретение относится к совместному введению одного или нескольких факторов выживания нейронов с нейрональными клетками-предшественниками или дифференцированными нервными клетками по настоящему изобретению для лечения нейродегенеративного нарушения или нервного заболевания. Фактор(ы) выживания нейронов можно вводить до введения желательных клеток, в сцеплении с ним, в сочетании с ним или после него. Как применяют здесь, «фактор выживания нейронов» представляет собой любое вещество, вызывающее выживание контактирующих с фактором нейронов (in vitro или in vivo) в течение большего периода времени, по сравнению с периодом, который мог бы быть без присутствия фактора. Факторы выживания нейронов, которые можно применять по настоящему терапевтическому варианту осуществления, включают в себя в качестве неограничивающих примеров агонисты ГАМК (например, бензодиазепины, вальпроат и фенобарбитал), антагонисты кальция, антагонисты глутамата, антиоксиданты, тканевой активатор плазминогена (t-PA), глиальный нейротрофический фактор (GDNF), фактор роста нервов (NGF), цилиарный нейротрофический фактор (CNTF), выделенный из головного мозга нейротрофический фактор (BDNF), нейротрофин-3 (NT-3), нейротрофин-4 (NT-4), FGF, IL-1β, TNFα, инсулиноподобный фактор роста (IGF-1, IGF-2), трансформирующий фактор роста бета (TGF-β, TGF-βl), полученные из растений препараты, экстракты на основе растений или синтетические источники с активностью против болезни Паркинсона, инсульта, церебральной ишемии, антинейродегенеративной или нейропротективной активностями.
По одному предпочтительному варианту осуществления факторы выживания нейронов совместно вводят с нейрональными клетками, предпочтительно ГАМКергическими нейронами, для лечения страдающих церебральной ишемией или инсультом субъектов. В настоящее время агонисты ГАМК (например, бензодиазепины, вальпроат и фенобарбитал), антагонисты кальция, антагонисты глутамата, антиоксиданты и другие нейропротективные средства или препараты представляют собой применяемые для лечения церебральной ишемии или инсульта химические средства. В то время как эти препараты помогают ослабить у пациентов симптомы, связанные с неврологическими нарушениями инсульта или ишемии, они не способны излечить эти нарушения. Принимая во внимание их критическую роль у пациентов с инсультом или ишемией, эти средства или препараты можно применять для лечения связанных с инсультом или ишемией неврологических нарушений и заболеваний в сочетании с полученными из полипотентных стволовых клеток ГАМКергическими нейронами, до терапевтического введения ГАМКергических нейронов, во время или после него.
По еще одному предпочтительному варианту осуществления для лечения субъектов, страдающих церебральной ишемией или инсультом до терапевтического введения нейрональных клеток, предпочтительно ГАМКергических нейронов, во время или после него совместно вводят тканевой активатор плазминогена (t-PA). Рекомбинантные формы t-PA применяют у пациентов для удаления кровяных сгустков или связанной с церебральной ишемией или инсультом закупорки, и они представляют собой только одобренную FDA терапию церебральной ишемии или инсульта. Прогноз для страдающего инсультом или церебральной ишемией пациента улучшается при наискорейшем снабжении кровью поврежденного участка(ов), предпочтительно гиппокампа, коры и областей таламуса головного мозга, особенно до введения нейрональных клеток любого типа в поврежденный участок(и) головного мозга. Этого можно достичь посредством введения пациенту t-PA, который улучшает выживание введенных нейронов в окружение хозяина посредством разблокирования кровеносных сосудов, таким образом предоставляя нейронам необходимый кислород и питательные вещества.
Как применяют здесь, термины «для лечения», «лечение» или «терапия» относятся к терапевтическому лечению и профилактическим или превентивным мерам. Терапевтическое лечение включает в себя в качестве неограничивающих примеров ослабление или устранение симптомов определенного заболевания или нарушения, или замедление или устранение прогрессии, или излечение существующего заболевания или нарушения. Следовательно, субъекты с необходимостью лечения включают в себя субъектов с уже поставленным диагнозом нейродегенеративного нарушения или нервного заболевания, а также субъектов, у которых нейродегенеративное нарушение или нервную болезнь необходимо предотвратить. Способы по настоящему описанию можно применять для любых нуждающихся в лечении млекопитающих, включающих в себя в качестве неограничивающих примеров человека, приматов и домашних животных, животных, выращенных на ферме, комнатных животных или спортивных животных, таких как собаки, лошади, кошки, овцы, свиньи, крупный рогатый скот, крысы, мыши и т.д. «Нарушение» представляет собой любое состояние, которому помогает лечение нейрональными клетками-предшественниками, дифференцированными нервными клетками или любым полученным по способам настоящего описания типом клеток. Примеры нарушений и заболеваний, которым помогает лечение клетками по настоящему описанию, особенно ГАМКергическими нейронами, представляют собой болезнь Паркинсона, болезнь Альцгеймера, болезнь Гентингтона, деменцию с тельцами Леви, рассеянный склероз, мозжечковую атаксию, прогрессирующий супрануклеарный паралич, повреждение спинного мозга, боковой амиотрофический склероз (ALS), эпилепсию, инсульт, ишемию и т.п., а также связанные с узнаванием и психологией нарушения, включающие в себя в качестве неограничивающих примеров тревожные расстройства, обсессивно-компульсивные расстройства (OCD), изменения личности, синдром нарушения внимания (ADD), синдром дефицита внимания с гиперактивностью (ADHD) и шизофрению.
Способы по настоящему изобретению преимущественно можно выполнять посредством прямого введения в пораженный участок нейрональных клеток-предшественников или дифференцированных нервных клеток по настоящему описанию. Специалистам в данной области хорошо известны способы трансплантации нейронов и культивирования клеток, например, патент США №5514552; Yurek and Sladek, 1990, Annu. Rev. Neurosci. 13: 415-440; Rosenthal, 1998, Neuron 20: 169-172; Vescovi et al., 1999, J. Neurotrauma 16(8): 689-93; Vescovi et al., 1999, Exp. Neuro. 156(1): 71-83; Brustle et al., 1999, Science 285: 754-56; каждый конкретно включен в настоящее описание в качестве ссылки. Клетки можно доставлять или отдельно, или в сочетании с другими факторами, например фактором выживания нейронов, и можно доставлять параллельно с фармацевтически приемлемым носителем. В идеале такой носитель повысит свойства стабильности и доставки клеток.
Также по настоящему описанию представлены фармацевтические композиции, содержащие клетки, которые можно вводить с применением пригодного носителя, такого как липосомы, микрочастицы или микрокапсулы. Клетки по настоящему описанию также можно доставлять в форме фармацевтической композиции, содержащей изотонический наполнитель и полученной в достаточно стерильных для введения человеку условиях. Основные принципы медицинских препаратов клеточных композиций находятся в Cell Therapy: Stem Cell Transplantation, Gene Therapy, and Cellular Immunotherapy, G. Morstyn & W. Sheridan eds, Cambrigge University Press, 1996, and Hematopoietic Stem Cell Therapy, E. Ball, J. Lister & P. Law, Churchill Livingstone, 2000, конкретно включенные в настоящее описание в качестве ссылки. Кроме того, желательным может являться локальное введение в нуждающийся в лечении участок фармацевтической композиции, содержащей фактор выживания нейронов, чего можно достичь, например, посредством локальной инфузии при хирургической операции, инъекции, катетерного механизма или имплантационного устройства, где таким имплантатом может быть пористый, непористый или гелеобразный материал, включающий в себя мембраны, такие как силиконовые мембраны или волокна.
По настоящему описанию субъекту можно вводить нейрональные клетки-предшественники и дифференцированные нервные клетки в виде гомогенной в значительной степени, почти гомогенной или гетерогенной популяции клеток. Гомогенная в значительной степени популяция клеток содержит более чем 75% одного типа клеток, таких как ГАМКергические нейроны, более предпочтительно более чем 90% и наиболее предпочтительно более чем 95%-99%. Гетерогенная популяция клеток состоит из двух или нескольких типов клеток, смешанных в одну клеточную популяцию, например ГАМКергические нейроны, дофаминергические нейроны, серотонинергические нейроны, Шванновские клетки, олигодендроциты, астроциты и глиальные клетки. Также посредством способов, хорошо известных специалистам в данной области, клетки можно генетически изменять для экспрессии или высвобождения трофических факторов, факторов роста, факторов выживания нейронов или других терапевтических соединений в поврежденной области головного мозга, центральной нервной системы, периферической нервной системы или других тканях. Применение сочетаний промотора и типа клеток для экспрессии белка хорошо известны специалистам в области молекулярной биологии, например, см. Sambrook, et al. 1989, Molecular Cloning: A Laboratory Manual 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, конкретно включенные в настоящее описание в качестве ссылки.
Для достижения экспрессии трофических факторов, факторов роста, факторов выживания нейронов или других терапевтических соединений в нейрональных клетках-предшественниках и дифференцированных нервных клетках по настоящему описанию пригодные регуляторные элементы можно получать из разных источников, и их легко может выделять специалист в данной области. Примеры регуляторных элементов включают в себя транскрипционный промотор, энхансер, связывающую РНК-полимеразу последовательность, а также связывающую рибосомы последовательность, включающую в себя сигнал инициации трансляции. В рекомбинантную молекулу можно вводить другие дополнительные генетические элементы, такие как селективные маркеры. Рекомбинантную молекулу можно вводить в полипотентные стволовые клетки или в нейрональные клетки-предшественники или дифференцированные нервные клетки, полученные из полипотентных стволовых клеток, с применением in vitro доставляющих носителей или способов in vivo. Примеры способов доставки включают в себя ретровирусные векторы, аденовирусные векторы, векторы вирусной ДНК, липосомы, физические способы, такие как микроинъекции и трансфекция посредством электропорации или преципитации фосфатом кальция, или другие известные в данной области способы переноса для создания рекомбинантных клеток. Генетически измененные клетки можно инкапсулировать в микросферы и имплантировать в больную или пораженную ткань или вблизи от нее. Применяемые протоколы хорошо известны специалистам в данной области, и их можно найти, например, у Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, New York, N. Y., 1997, включенные в настоящее описание в качестве ссылки.
Предпочтительно лечение трансплантацией клеток по настоящему описанию также включает в себя несколько средств хранения и консервации нейрональных клеток-предшественников и дифференцированных нервных клеток для применения при пересадке тканей, например длительное хранение посредством криоконсервации или кратковременное хранение в консервирующей среде. Замороженную эмбриональную ткань среднего мозга успешно хранили до 70 суток и пересаживали в качестве гомотрансплантата грызунам (Collier et al., Progress in Brain Research, Vol.78, New York, Elsevier (1988), pp.631-36, конкретно включенная в настоящее описание в качестве ссылки) и приматам (Collier et al., 1987, Brain Res. 436: 363-66, конкретно включенная в настоящее описание в качестве ссылки). Также показано, что после криоконсервации можно успешно культивировать эмбриональные клетки среднего мозга. Ткань среднего мозга также можно кратковременно (2-5 суток) хранить в консервирующей среде при 4°C и впоследствии трансплантировать с количеством выживающих трансплантатов, сходным с таковым для свежей ткани (Sauer et al., 1989, Restor. Neurol. Neurosci. (Suppl.: 3.sup.rd Int. Symp.Neural Tranplan.): 56, конкретно включенная в настоящее описание в качестве ссылки). Для хранения и консервации нейрональных клеток-предшественников и дифференцированных нервных клеток по настоящему описанию можно применять подобные способы, и такие способы хорошо известны специалистам в данной области.
Еще одним применением для описанных здесь нейрональных клеток-предшественников и дифференцированных нервных клеток является скрининг факторов, таких как фармацевтические соединения, растворы, небольшие молекулы, пептиды или полинуклеотиды, а также факторы внешней среды, такие как условия культивирования или оказывающие влияние на фенотип или характеристики этих клеток манипуляции. Например, биологически активные молекулы, представленные в растениях, экстрактах на основе растений или у животных, человека или из синтетических источников, можно скринировать и оценивать с применением этих клеток. Кроме того, эти клетки можно применять для определения возможных факторов роста или факторов дифференцировки. Например, подходящее фармацевтическое соединение можно добавлять к нейрональным клеткам-предшественникам или зрелым нейронам отдельно или в сочетании с другими препаратами, и можно определять и оценивать любые изменения в морфологии, фенотипе или функциональной активности клеток. По еще одному варианту осуществления ГАМКергические нейроны применяют для скрининга воздействующих на рецепторы факторов (например, агонисты или антагонисты) ГАМКергических нейронов в ЦНС, или ПНС, или конкретных тканях и органах. Также ГАМКергические нейроны можно применять для скрининга агонистов и/или антагонистов нейропептидов, нейротрансмиттеров, нейрогормонов или ГАМК. Также ГАМКергические нейроны можно применять для тестирования нейротоксичности биологически активных молекул.
Кроме того, затем описанные здесь нейрональные клетки-предшественники и дифференцированные нервные клетки можно модифицировать на любой стадии дифференцировки. Например, эти клетки можно генетически модифицировать для получения одной или многочисленных временных или стабильных генетических модификаций. Генетические изменения этих клеток могут быть желательными по многим причинам, таким как представление модифицированных клеток для генной терапии или замещение тканей для пересадки или имплантации. Клетки по настоящему описанию можно генетически модифицировать посредством введения экспрессирующих селективный маркер векторов под контролем специфичного для нейронов промотора, хорошо известного специалистам в данной области. Также эти клетки можно модифицировать на любой стадии для экспрессии определенных маркеров или генов, что затем можно применять для очищения полученных из полипотентных стволовых клеток дифференцированных клеток или, альтернативно, для индукции дифференцировки в конкретные линии клеток. Эти клетки можно модифицировать для снижения или предотвращения иммунного отторжения после трансплантации, т.е. гистосовместимости с определенным реципиентом.
Для повышения репликативной способности полученных с применением настоящего изобретения клеток эти клетки можно теломеризовать посредством генетического изменения их пригодным вектором так, чтобы они экспрессировали теломеразный каталитический компонент (TERT). Применяемую последовательность TERT можно получить из человека или мыши (WO 98/14592 и WO 99/27113, конкретно включенные в настоящее описание в качестве ссылки), а также других видов млекопитающих. Альтернативно, можно повысить транскрипцию эндогенного гена TERT. Применяемые способы генетического модифицирования клеток хорошо известны специалистам в данной области. В этих способах применяют различные молекулярно-биологические технологии, многие из которых полностью описаны у Sambrook, et al. 1989, Molecular Cloning: A Laboratory Manual 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, конкретно включенная в настоящее описание в качестве ссылки.
Трансплантация in vitro клеток головного мозга взрослой особи
Человеческий головной мозг имеет невысокую способность к самовосстановлению, и разработка способов лечения вызванных заболеванием или травмой повреждений головного мозга остается огромной клинической проблемой. Хотя существовал большой интерес к возможности лечения повреждения головного мозга посредством трансплантации клеток, пока еще этот подход находится на ранней экспериментальной стадии. Одним большим препятствием для трансплантации нервных клеток является выживаемость этих клеток, так как эксперименты показывают, что большинство пересаженных клеток после трансплантации дегенерируют. Например, обычный коэффициент выживаемости дофаминергических клеток, пересаженных экспериментальным животным или субъектам с болезнью Паркинсона, ограничен приблизительно 10% (Brundin et al., 1987, Ann. N.Y. Acad. Sci. 495: 473-96; Nakao et al., 1995, Nat. Med. 1(3): 226-31). Хотя причина(ы) такого низкого коэффициента выживаемости неизвестна(ы), это может являться результатом нейротоксических воздействий клеток головного мозга реципиента или недостаточного снабжения пересаженных клеток питательными веществами и кислородом. Очевидно, что факторы, вызывающие гибель нейронов, следует изучать дополнительно для того, чтобы можно было минимизировать гибель нейронов после трансплантации.
Одно направление идентификации и изучения этих факторов представляет собой создание подобного головному мозгу окружения и затем тестирование эффективности или выживаемости нейрональных или нервных клеток, полученных из полипотентных стволовых клеток или выделенных из других источников. По одному варианту осуществления настоящего описания создают такое подобное головному мозгу окружение с применением технологии трансплантации in vitro выделенных нейрональных или нервных клеток вместе со зрелыми нервными клетками, предпочтительно выделенными из зрелого гиппокампа клетками. Как применяют здесь, термин «трансплантация in vitro» относится к культивированию двух различных типов клеток вместе в одинаковом культуральном окружении. Трансплантацию in vitro можно применять для определения совместимости двух типов клеток в сходном окружении и изучения и прогнозирования факторов, воздействующих на выживаемость и функциональность пересаженных клеток. Эту систему можно применять для определения выживаемости и функциональности выделенных нейрональных или нервных клеток, особенно полученных из полипотентных клеток. Это определение, в свою очередь, помогает поочередно определить возможность лечения этими клетками нейродегенеративных нарушений или нервных заболеваний, включающих в себя в качестве неограничивающих примеров инсульт, ишемию, болезнь Паркинсона, болезнь Альцгеймера, эпилепсию и болезнь Гентингтона. По предпочтительному варианту осуществления этот способ применяют для определения выживаемости или эффективности полученных, как здесь описано, из полипотентных стволовых клеток ГАМКергических нейронов в подобном хозяину окружении. По предпочтительным вариантам осуществления выживаемость нейрональных или нервных клеток, предпочтительно ГАМКергических нейронов, полученных из полипотентных стволовых клеток, составляет по меньшей мере 90% в окружении клеток зрелого гиппокампа после трансплантации in vitro. В других вариантах осуществления в этом окружении выживают по меньшей мере приблизительно 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 95% или 99% ГАМКергических нейронов.
Новую описанную здесь модель трансплантации in vitro также можно применять для изучения следующих параметров: (1) формирование синапса между пересаженными клетками и клетками хозяина и у них; (2) факторы, вовлеченные в гибель нейрональных клеток или нейронов после трансплантации; и (3) коэффициент гибели нейрональных клеток или нейронов. Модель трансплантации in vitro помогает не только предоставить подробную информацию, необходимую для улучшения выживаемости и функциональности клеток после трансплантации, но также помогает нейробиологам и нейрохирургам определить стратегию до трансплантации нейронов пациенту.
Следующие ниже примеры включены для демонстрации предпочтительных вариантов осуществления изобретения. Специалистам в данной области ясно, что описанные ниже в примерах способы представляют найденные авторами способы для правильного исполнения изобретения на практике, и таким образом их можно рассматривать для составления предпочтительных способов его применения. Кроме того, в свете настоящего описания специалистам в данной области ясно, что можно сделать много изменений в конкретных описанных вариантах осуществления и все еще достичь подобного или сходного результата без отклонения от сущности и цели изобретения.
Пример 1
В следующих примерах продемонстрировано получение in vitro функциональных ГАМКергических нейронов из мышиных эмбриональных стволовых клеток. На фиг.1 проиллюстрировано получение ГАМКергических нейронов из мышиных ЭС клеток, тогда как на фиг.2 проиллюстрированы различные стадии дифференцировки мышиных ЭС клеток в окончательно дифференцированные нейроны.
1) Культивирование и наращивание мышиных эмбриональных стволовых клеток:
В настоящей серии экспериментов применяют мышиные ЭС клетки, выделенные из внутренней клеточной массы мышиной бластоцисты с применением способов, хорошо известных специалистам в данной области. Мышиные ЭС клетки представляют собой клетки происхождения J-1 (получены из National Institute of Dental and Craniofacial Research, National Institute of Health, Bathesda, Maryland, USA) и были на 14 пассаже. Клетки держали на обработанных митомицином-C фидерных клетках мышиных эмбриональных фибробластов, митотически инактивированных в ЭС клеточной среде. Мышиные ЭС клетки культивируют в ЭС клеточной среде для увеличения количества недифференцированных клеток. Как правило, ЭС клетки можно наращивать без потери полипотентности по меньшей мере приблизительно 1000-кратно. Обычно применяемая среда для ЭС клеток включает в себя источник углерода и буфер для поддержания желательного pH. ЭС клеточная среда, состоящая из модифицированной Дульбекко среды Игла (DMEM) или нокаут DMEM (Gibco), дополнена 10-20% пригодной для ЭС клеток эмбриональной бычьей сывороткой (FBS) (Hyclone) или нокаутирующим заменителем сыворотки (Gibco), 1% MEM раствора заменимых аминокислот, 2 мМ L-глутамина и 0,1 мМ β-меркаптоэтанола. Также среда для ЭС клеток дополнена фактором ингибирования лейкемии (LIF) в концентрации 1000 Ед/мл (ESGRO, Chemicon International Inc.) для ингибирования дифференцировки мышиных ЭС клеток.
Мышиные ЭС клетки наращивают посредством культивирования и регулярного пересаживания ЭС клеток в ЭС клеточной среде для ингибирования дифференцировки снова с применением способов, хорошо известных специалистам в данной области. ЭС клетки культивируют на тканевых культуральных планшетах, обработанных от 0,1% до 0,2% желатином в фосфатно-солевом буфере (PBS) при 37°C в течение по меньшей мере 1-2 часов. ЭС клетки растят на инактивированных митомицином-C мышиных фидерных клетках в концентрации 1×105/мл. ЭС клетки инкубируют в течение 4 суток при температуре приблизительно между 35°C и 40°C, предпочтительно приблизительно при 37°C, приблизительно между 1% и 10% CO2 в атмосфере, более предпочтительно при 5% CO2 в атмосфере. Среду для ЭС клеток меняют каждые сутки или через сутки, в зависимости от роста ЭС клеток в культуре.
2) Образование эмбриональных телец
После того как недифференцированные мышиные ЭС клетки пролиферируют и размножаются, их культивируют для формирования эмбриональных телец. Сначала ЭС клетки диссоциируют посредством короткой экспозиции с 0,05% трипсин-ЭДТА, с последующим удалением с поверхности и разбивкой клеток в небольшие кластеры. Затем эти кластеры наносят на 60 мм бактериологические чашки с плотностью приблизительно 4×105 клеток/мл в соответствующей среде в отсутствие фидерных клеток. Применяемые бактериологические чашки имеют предотвращающую прикрепление неадгезивную поверхность, стимулируя таким образом дифференцировку ЭС клеток и формирование эмбриональных телец. Эти клетки культивируют в виде суспензионной культуры в среде для ЭС клеток. Применяемая для культивирования этих клеток среда для ЭС клеток представляет собой DMEM с высоким содержанием глюкозы, или нокаут DMEM, дополненную 10-20% FBS или нокаутирующим заменителем сыворотки, а также другими добавками, такими как β-меркаптоэтанол (0,1 мМ), L-глутамин (2 мМ) и антибиотики. В среду для ЭС клеток не добавляют bFGF или LIF. На следующие сутки клеточную суспензию переносят в новый культуральный планшет, оставляя на предыдущем планшете любые прикрепленные к нему клетки. Среду ЭС клеток меняют через сутки посредством центрифугирования клеток в старой среде, ресуспендируя клетки в свежей среде и возвращая клетки в неадгезивный культуральный планшет. Эмбриональные тельца растят в течение 4-8 суток. В конце 4-8 суток эмбриональные тельца собирают и осаждают на низкой скорости (1000 об/мин, 5 минут) и ресуспендируют в среде для ЭС клеток. Затем приблизительно 30-40 эмбриональных телец переносят в непокрытый планшет для тканевых культур и инкубируют в течение 24 часов.
3) Отбор и наращивание нестин-позитивных нейрональных клеток-предшественников
Через 24 часа нестин-позитивные клетки (нейрональные клетки-предшественники) отбирают посредством замены среды ЭС клеток на определенную бессывороточную среду ITSFn (селекция по нестину). Среда ITSFn состоит из среды DMEM:F12 (Gibco), дополненной ростовыми факторами, инсулином (5-25 мкг/мл) (Sigma), селенитом натрия (1-5 нМ) (Sigma), трансферрином (1-10 мкг/мл) (Gibco) и фибронектином (1-5 мкг/мл). Как правило, клетки инкубируют в среде ITSFn в течение приблизительно 6-10 суток, более предпочтительно в течение приблизительно 7 суток, с дополнением среды ITSFn через сутки. Как правило, клетки растят приблизительно при 35°C и 40°C, предпочтительно приблизительно при 37°C, в атмосфере приблизительно от 1% до 10% CO2, более предпочтительно в атмосфере приблизительно от 2% до 6% CO2. После завершения отбора предпочтительно в течение 7 суток нейрональные клетки-предшественники характеризуют по экспрессии нестина с применением экспериментального иммунофлуоресцентного способа, показывающего, что приблизительно 90% клеток позитивны по экспрессии нестина. Затем нестин-позитивные клетки наращивают, как описано ниже.
Нестин-позитивные клетки диссоциируют с применением 0,05% трипсин-ЭДТА и наносят на покрытый поли-L-орнитином/ламинином планшет, содержащий среду для роста клеток ЦНС. Среда для роста клеток ЦНС содержит DMEM:F12 с добавлением N2 (10 мкг/мл) и B27 (20 мкг/мл). Также добавляют в среду для роста клеток ЦНС фактор роста нервов, β-FGF (10-20 нг/мл), как известно, увеличивающие образование нейронов. Для образования большого количества нейрональных клеток нейрональные клетки-предшественники растят в течение 6 суток. Клетки растят в среде для роста клеток ЦНС в течение 6 суток с дополнением среды каждые двое суток.
4) Дифференцировка нейрональных клеток-предшественников
Наросшие нейрональные клетки-предшественники дифференцируют посредством культивирования клеток в среде для дифференцировки, содержащей среду Neurobasal A (Gibco), FCS (10-20%) (HYCLONE) и добавку B27 (2-10%) (Gibco), но не β-FGF. Кроме того, среда для дифференцировки содержит факторы для увеличения выхода ГАМКергических нейронов, предпочтительно Ara-C (Sigma Chemical Co, USA). Культивирование нейрональных клеток-предшественников в содержащей Ara-C (20 мкг/мл) среде для дифференцировки повышает долю ГАМКергических нейронов в полученной из мышиных ЭС клеток нейрональной популяции. После 3 суток культивирования в содержащей Ara-C среде для дифференцировки клетки растят в той же среде без Ara-C в течение 8, 12 или 16 суток. При культивировании нейрональных клеток-предшественников в содержащей Ara-C среде для дифференцировки в течение 3 суток и последующем росте в среде для дифференцировки без Ara-C в течение 16 суток по меньшей мере приблизительно 60% нейрональных клеток-предшественников дифференцируются в ГАМКергические нейроны (фиг.6).
5) Характеристика дифференцированных нейронов
Анализ общей популяции полученных с применением указанных выше способов нейрональных клеток демонстрирует, что более 90% дифференцированных клеток представляют собой нейрональные клетки. Полученные по настоящему описанию дифференцированные типы нейрональных клеток оценивают посредством общей морфологии клеток, а также идентифицированных посредством иммунофлуоресценции фенотипов. Иммунофлуоресцентный анализ проводят на стадии роста нейрональных предшественников, а также после 4, 8, 12 и 16 суток дифференцировки, как описывают выше в протоколе. Сначала выделенные клетки растят на стеклах с 2-луночными камерами, отмывают в PBS и фиксируют в течение 10 минут 4% параформальдегидом при комнатной температуре. Затем клетки пермеабилизируют 0,2% Тритоном X-100 в PBS, содержащим 1% нормальной козьей сыворотки, и затем блокируют 1% бычьего сывороточного альбумина (BSA)/PBS в течение 1 часа при комнатной температуре.
Затем клетки инкубируют в течение ночи при 4°C с первыми антителами (разведение антител выполняют в 1% BSA). В иммунофлуоресцентном исследовании применяли следующие антитела: моноклональные ГАМК, 1:200 (Chemicon Inc. USA); поликлональные ГАМК, 1:500 (Chemicon Inc. USA); GAD65, 1:500 (Chemicon Inc. USA); GAD67, 1:500 (Chemicon Inc. USA); GAT-1, 1:500 (Chemicon Inc. USA), GAT-2, 1:500 (Chemicon Inc. USA); глутамат, 1:500 (Chemicon Inc. USA); нестин, 1:50 (Chemicon Inc. USA); олигодендроциты, 1:500 (Chemicon Inc. USA); серотонин, 1:500 (Chemicon Inc. USA); тирозингидроксилаза, 1:800 (Chemicon Inc. USA); MAP-2, 1:500 (Chemicon Inc. USA) и GFAP, 1:500 (Chemicon Inc. USA). После ночной инкубации с первыми антителами клетки отмывают в PBS и инкубируют с меченными FITC вторыми антителами в течение 1 часа в темноте. После этого клетки отмывают три раза в PBS и покрывают заключающей средой. Лунки стекол смотрят во флуоресцентном микроскопе для оценки иммунопозитивных клеток на различных стадиях дифференцировки.
Иммунофлуоресцентный анализ дифференцированных культур показал, что приблизительно 90% дифференцированных нейрональных клеток являются иммунореактивными к нейрон-специфическим маркерам, таким как GAD-65, GAD-67, GAT-1, GAT-2, глутамат, ГАМК, нестин и MAP-2 (фиг.3). Приблизительно 60% всех нейрональных клеток в культуре окрашиваются по ГАМК, маркеру ГАМКергических нейронов, тогда как приблизительно 15% нейрональных клеток окрашиваются по тирозингидроксилазе (TH), маркеру дофаминергических нейронов. Кроме того, приблизительно 10% нейрональных клеток окрашиваются по глутамату, тогда как 5% нейрональных клеток окрашиваются по серотонину, маркеру серотонинергических нейронов. Наконец, приблизительно 5-10% клеток окрашиваются как олигодендроциты. Также культура клеток включает в себя глиальные клетки, обычно приблизительно от 5% до 10% популяции клеток. Анализ продемонстрировал, что по меньшей мере некоторые из дифференцированных нейронов в культуре клеток являются синаптически активными. Как правило, зрелые нейроны идентифицируют посредством присутствия длинных аксональных отростков с множеством тонких шиповидных структур. Присутствие ассоциированного с миелином белка-2 (MAP-2) в аксональных отростках также служит признаком созревания нейронов.
Также дифференцированные клетки анализируют посредством двойного мечения клеток первыми антителами для определения совместной локализации экспрессии GAD65 и GAD67 (фиг.4). GAD65 и GAD67 представляют собой экспрессированные в ГАМКергических нейронах два гена, необходимые для синтеза ГАМК. Популяцию дифференцированных нейрональных клеток исследуют посредством применения способа двойного иммунного мечения с применением антител к GAD65/GAD67 и ГАМК, как описано выше. Результаты двойного иммунного мечения предполагают, что экспрессирующие GAD65 и GAD67 белки нейрональные клетки также экспрессируют нейромедиаторы ГАМК (фиг.4). Эти результаты подтверждают, что полученные с применением описанных способов ГАМКергические нейроны представляют собой продуцирующие ГАМК клетки. Экспрессирующие GAD65 и GAD67 белки нейрональные клетки также продемонстрировали экспрессию ГАМК на раннем развитии, так как в ГАМКергических нейронах в присутствии ферментов GAD65 и GAD67 преобразуется глутамат.
Также анализируют профили генной экспрессии полученных на разных стадиях дифференцировки клеток. Клетки собирают для анализа посредством обратной транскрипции-полимеразной цепной реакции (ОТ-ПЦР) на каждой следующей стадии описанного способа: (1) Недифференцированные клетки; (2) Эмбриональные тельца (EBS); (3) Наращивание нестин-позитивных клеток; (4) 2 суток дифференцировки; (5) 4 суток дифференцировки; (6) 8 суток дифференцировки; (7) 12 суток дифференцировки; и (8) 16 суток дифференцировки. После сбора клеток их осаждают и из осадков клеток выделяют общую клеточную РНК с применением коммерческого набора RNeasy Qiagen. Выделенную РНК хранят при -20°C. Общую клеточную РНК обрабатывают ДНКазой RQ без РНКаз (Promega Corp., Madison, WI) для удаления всех остатков ДНК. кДНК синтезируют из выделенной РНК с применением обратной транскриптазы Moloney Leukemia virus superscript II, следуя инструкциям производителя. Для проведения реакций обратной транскрипции (ОТ) применяют случайные гексамерные праймеры (GIBCO/BRL).
Для определения того, какие гены экспрессируются в полученных клетках, синтезированную посредством этой реакции обратной транскрипции кДНК применяли для амплификации ПЦР с различными наборами специфических праймеров. Для идентификации экспрессируемых в ГАМКергических нейронах мРНК, конкретно генов глутамат-декарбоксилазы (GAD1 и GAD2), альтернативно сплайсированной эмбриональной РНК GAD1, и транскрипта везикулярного транспортера ингибирующих аминокислот (VIAAT) конструировали праймеры. GAD1 во время развития подвергается альтернативному сплайсингу, и предпочтительно во время эмбрионального развития в нейрональных стволовых клетках/клетках-предшественниках экспрессируется эмбриональный транскрипт. В качестве положительного контроля также проверяют экспрессию повсеместно экспрессируемого гена β-актина. Эти реакции ПЦР проводили с применением только 10% общего количества после реакции образования первой цепи (синтезированной посредством ОТ кДНК) в качестве матрицы и платиновой Taq-полимеразы в стандартных условиях ПЦР, хорошо известных специалистам в данной области. Обычные применяемые параметры циклов для амплификации продуктов ДНК представляют собой следующее:
1. денатурация матрицы при 94°C в течение 30 секунд;
2. отжиг праймеров при 55-65°C в течение 1 минуты, в зависимости от применяемых праймеров; и
3. инкубация реакции при 72°C в течение 1 минуты и повторение стадий 1-3 (циклов) от 25 до 40 раз.
После реакции ПЦР продукты разделяют в агарозном геле с применением электрофореза вместе с маркером размера ДНК. Экспрессию GAD1, GAD2, эмбриональной GAD1, VIAAT и β-актина анализируют посредством ОТ-ПЦР с применением праймеров, как описано в таблице:
Антисмысловой: GCG CAG TTT GCT CCC CGT TCT T (SEQ ID NO:2)
Антисмысловой: GAA TCA CAC TGT CTG TTC CAA TCC CTA A (SEQ ID NO:4)
Антисмысловой: TCC CAT CAC CTT TAT TTG ACC ATC C (SEQ ID NO:6)
Антисмысловой: GCA CCA CCT CCC CGT CTT CGT TCT CCT C (SEQ ID NO:8)
Антисмысловой: GTA ACA ATG CCA CCA TGT TCA AT (SEQ ID NO: 10)
Указанным выше анализом ОТ-ПЦР продемонстрировано, что многие полученные из ЭС клеток нейрональные клетки спонтанно дифференцируются в экспрессирующие маркеры ГАМКергического фенотипа нейроны. Обнаружена разница в экспрессии GAD1 и GAD2 на всех различных стадиях анализа культур клеток. Ген GAD1 экспрессирован почти на всех стадиях (от стадии 1 до стадии 5), включая в себя стадию недифференцированных клеток (фиг.5). Это сопоставимо с недавним сообщением об экспрессии GAD1 в ЭС клетках (Bain et al., 1993, Brain Res. Mol. Brain Res. 17: 23-30). С другой стороны, как показано на фиг.5, GAD2 экспрессирован только на стадии дифференцированных клеток. Этот результат предполагает, что полученные с применением описанных способов ГАМКергические нейроны имеют неодинаковую фенотипическую экспрессию генов GAD1 и GAD2. Сплайсированный вариант эмбриональной GAD1 экспрессирован на всех стадиях, за исключением стадии недифференцированных клеток, тогда как ген нейромедиаторов (VIAAT), умеющий транспортировать ГАМК, экспрессирован только на стадиях дифференцированных клеток (фиг.5). В качестве положительного контроля применяют экспрессию гена домашнего хозяйства β-актина (фиг.5).
Для анализа экспрессии ГАМК полученными с применением описанных способов ГАМКергическими нейронами также применяют высокоэффективную жидкостную хроматографию с обращенной фазой (RP-HPLC). Посредством прямого измерения внутриклеточных уровней ГАМК с применением RP-HPLC оценивают функциональную способность продукции ГАМК полученными из ЭС клеток ГАМКергическими нейронами, так как продукция ГАМК является отличительной характеристикой ГАМКергических нейронов. Детектируемую в каждом образце концентрацию ГАМК определяли посредством сравнения со стандартным раствором ГАМК, внесенным в колонку непосредственно перед каждым экспериментом и после него.
Сначала клетки собирали на разных стадиях описанного способа: наращивания нестин-позитивных клеток и дифференцированных клеток, выделенных после 8, 12 и 16 суток дифференцировки. Супернатанты культур собирали на разных стадиях, немедленно обрабатывали 7,5% ортофосфорной кислотой и метабисульфитом (0,22 мг/мл), чтобы стабилизировать нейромедиатор для измерения, и хранили при -80°C до проведения анализа посредством RP-HPLC. Внутриклеточные уровни ГАМК определяли с применением способа изократической HPLC, основанного на электрохимической детекции ГАМК образования с OPA/т-бутантиолом (Kehr and Ungerstedt, 1988, J. Neurochem. 51(4): 1308-10; Osborne et al., 1991, J. Neurosci. Method 34: 99-106, обе конкретно включены в настоящее описание в качестве ссылки). Данные HPLC нормализовали по стандартам ГАМК с варьирующими отношениями потока и чувствительностей (фиг.7).
Так как ГАМК не определяли на стадиях отбора по нестину и наращивания нестин-позитивных клеток, измеряемые уровни ГАМК на 8 сутки дифференцировки составили 20,95 пг/мл, тогда как на 12 и 16 сутки дифференцировки 26,18 пг/мл и 18,60 пг/мл соответственно. Этот анализ продемонстрировал, что самые высокие уровни ГАМК, полученные, как указано выше, ГАМКергические нейроны продуцируют на 12 сутки дифференцировки. Общее высвобождение ГАМК из культивируемых клеток составляло 21,51 пг/мл.
Так как самые высокие уровни ГАМК ГАМКергические нейроны продуцируют на 12 сутки дифференцировки, на этой же стадии дифференцировки клетки оценивают по присутствию рецепторов ГАМК-А и ГАМК-Б. Распределение функциональных рецепторов нейромедиаторов, например ГАМК-А- и ГАМК-Б-рецепторов, на поверхности нейронов, например ГАМКергических нейронов, четко соответствуют синаптической передаче и обработке сигнала (Eder et al., 2001, Eur. J. Neurosci. 13: 1065-69). Известно, что рецепторы ГАМК-А и ГАМК-Б критичны для нормального функционирования этих нейронов. Для определения экспрессии полученными, как здесь описано, из мышиных ЭС клеток ГАМКергическими нейронами ГАМКергических рецепторов получали иммунное окрашивание клеток с применением антител к рецептору ГАМК-А, 1:250 (Chemicon, USA) и рецептору ГАМК-Б, 1:250 (Chemicon, USA) с применением описанного ранее иммунофлуоресцентного протокола.
Иммунофлуоресцентный анализ культур на 12 сутки дифференцировки продемонстрировал, что рецепторы ГАМК-А и ГАМК-Б локализованы в дифференцированных нейронах. Интересно, что приблизительно 80% дифференцированных клеток экспрессируют рецептор ГАМК-А (фиг.8), тогда как только приблизительно 25% дифференцированных клеток экспрессируют рецептор ГАМК-Б (фиг.9). Так как экспрессия рецепторов ГАМК-А и ГАМК-Б четко соответствует синаптической передаче и обработке сигнала, выделяемые ГАМКергические нейроны, экспрессирующие оба рецептора, могут быть критичными для установления клеточной функциональности после трансплантации.
Пример 2
Заявители настоящего описания применяют следующий ниже пример в модели трансплантации in vitro для изучения эффективности, выживаемости и функциональности полученных из мышиных стволовых клеток ГАМКергических нейронов. На фиг.1 проиллюстрирована модель трансплантации in vitro.
Выживаемость и функциональность полученных из мышиных ЭС клеток ГАМКергических нейронов изучают посредством пересадки in vitro ГАМКергических нейронов к мышиными клетками гиппокампа головного мозга взрослой особи. Клетки гиппокампа выделяют посредством первичного выделения клеток из мышиного гиппокампа головного мозга взрослой особи, диссоциируют эти клетки 0,05% трипсин-ЭДТА и культивируют эти клетки в течение одной недели в среде Neurobasal-A, дополненной B27 (20 мкг/мл) и N2 (10 мкг/мл). Клетки гиппокампа головного мозга взрослой особи растят на 100 мм тканевых культуральных планшетах, покрытых поли-L-орнитином и ламинином или желатином. Через одну неделю полученные из мышиных ЭС клеток ГАМКергические нейроны и собранные после 12 суток дифференцировки наносят на клетки гиппокампа головного мозга взрослой особи и оба типа клеток культивируют вместе в течение одной недели. Приблизительно 90% ГАМКергических нейронов выживали после одной недели трансплантации in vitro с клетками гиппокампа головного мозга взрослой особи. Этот результат предполагает, что полученные из полипотентных стволовых клеток нейроны функциональны в окружении головного мозга взрослой особи и создают синаптические связи с клетками головного мозга взрослой особи, что также предполагает, что эти клетки можно терапевтически применять для лечения множества нейродегенеративных нарушений или нервных заболеваний.
Все из описанных и заявленных здесь композиций и способов можно разрабатывать и выполнять без лишнего проведения экспериментов в свете настоящего описания. Специалистам в данной области очевидно, что можно применять вариации к композициям и/или способам, и в стадиях или в последовательности стадий описанных здесь способов без потери концепции, сущности и объема изобретения, так как композиции и способы этого изобретения описаны в терминах предпочтительных вариантов осуществления. Более конкретно, очевидно, что определенные химически или физиологически родственные средства можно заменять описанными здесь средствами, пока можно достигать тех же самых или сходных результатов. Полагают, что специалистам в данной области очевидно, что все подобные сходные замены и модификации не меняют сущности, объема и концепции изобретения, определенных посредством прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНО ДИФФЕРЕНЦИРОВАННЫХ ДОФАМИНЕРГИЧЕСКИХ НЕЙРОНОВ ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2004 |
|
RU2345133C2 |
| СПОСОБ ДИФФЕРЕНЦИАЦИИ НЕЙРОНАЛЬНЫХ КЛЕТОК ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ (ЭС) КЛЕТОК | 2005 |
|
RU2407790C2 |
| ТРАНСПЛАНТАЦИЯ НЕРВНЫХ КЛЕТОК ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ СОСТОЯНИЙ | 2005 |
|
RU2434636C2 |
| ПРИМЕНЕНИЕ СТВОЛОВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, И ИХ КОМПОЗИЦИЙ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ (ВАРИАНТЫ) | 2017 |
|
RU2742828C2 |
| СТВОЛОВЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ МЕЗЕНХИМАЛЬНЫЕ И НЕЙРОНАЛЬНЫЕ МАРКЕРЫ, ИХ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2017 |
|
RU2741839C2 |
| СПОСОБ ТКАНЕВОЙ ИНЖЕНЕРИИ СПИННОГО МОЗГА ПОСЛЕ ЕГО АНАТОМИЧЕСКОГО РАЗРЫВА | 2012 |
|
RU2489176C1 |
| НОВОЕ ПРИМЕНЕНИЕ | 2004 |
|
RU2416426C2 |
| ПРИМЕНЕНИЕ ПРОГРАММЫ ПРОТИВОПОЛОЖНОЙ ДИФФЕРЕНЦИРОВКИ КЛЕТОК (ППДК) ДЛЯ ЛЕЧЕНИЯ ДЕГЕНЕРИРОВАВШИХ ОРГАНОВ В ПАТОЛОГИЧЕСКОМ СОСТОЯНИИ | 2008 |
|
RU2528084C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОПУЛЯЦИИ СТРОМАЛЬНЫХ КЛЕТОК ЖИРОВОЙ ТКАНИ, ИНДУЦИРОВАННЫХ К НЕЙРАЛЬНОЙ ДИФФЕРЕНЦИРОВКЕ | 2008 |
|
RU2396345C1 |
| СПОСОБЫ, ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИЗДЕЛИЯ ДЛЯ ВВЕДЕНИЯ ТЕРАПЕВТИЧЕСКИХ КЛЕТОК В ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ ЖИВОТНОГО | 2008 |
|
RU2468818C2 |
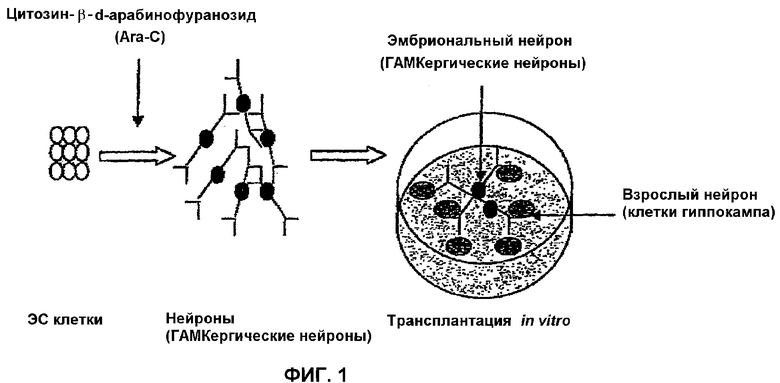
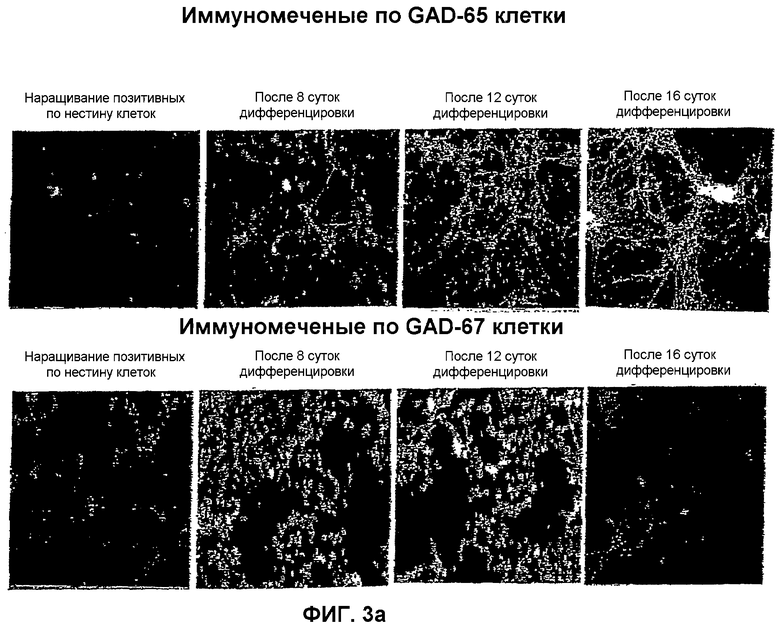
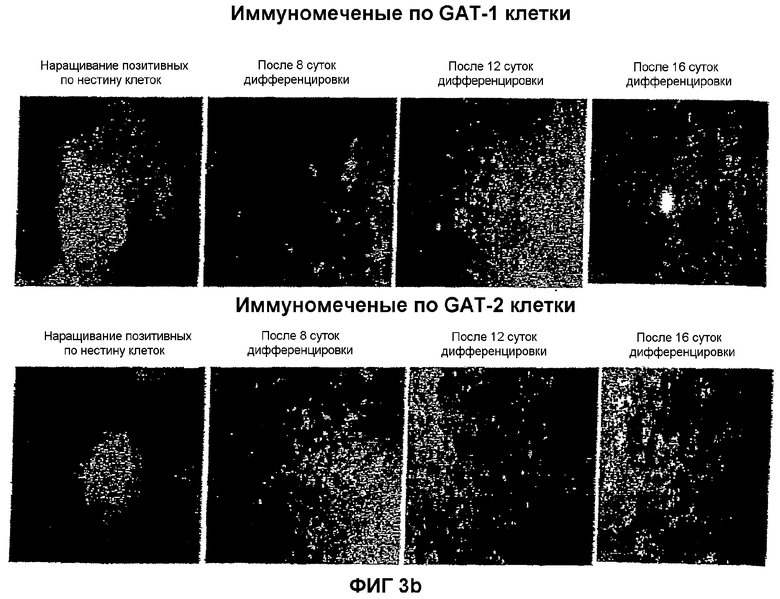
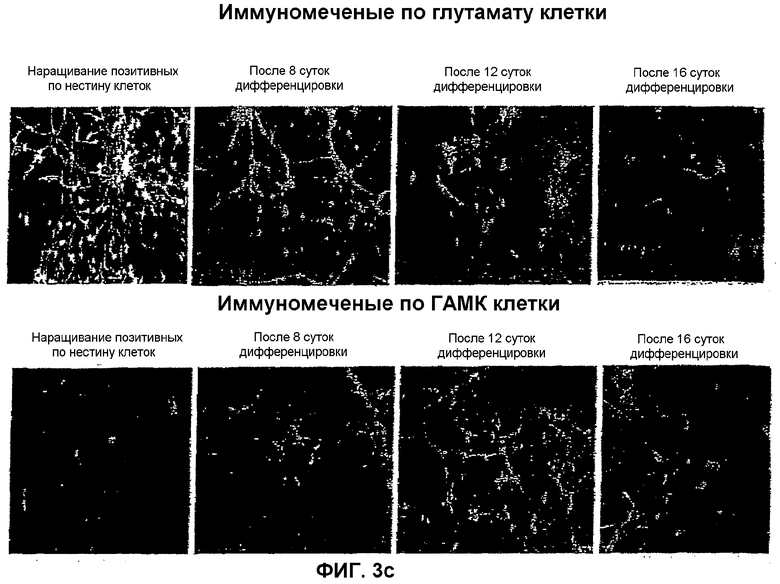
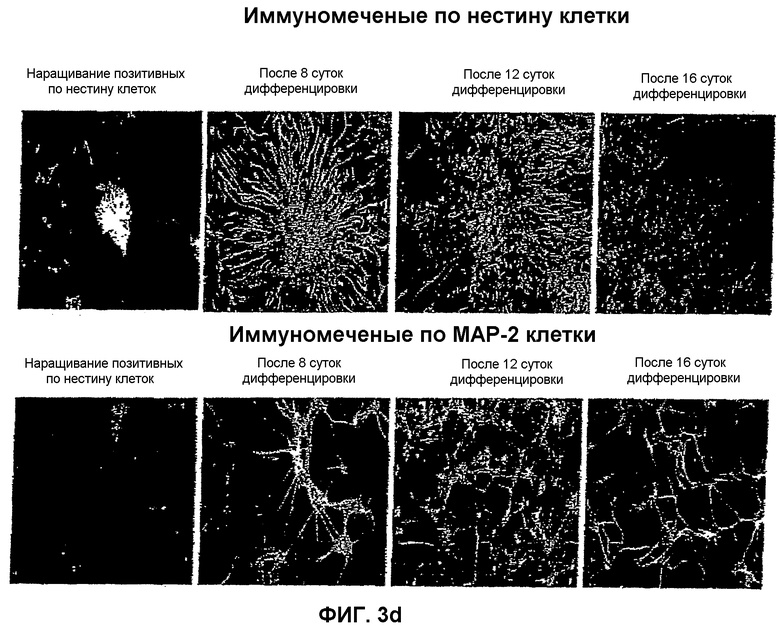
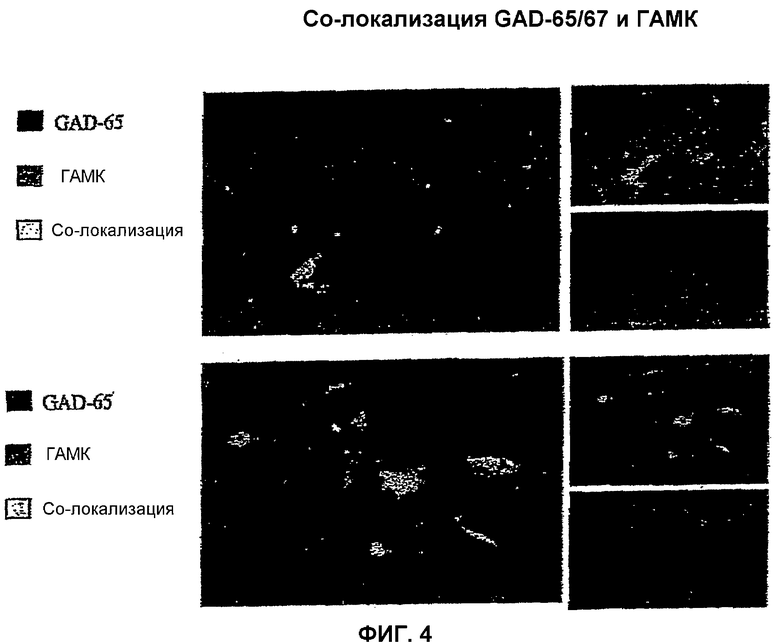
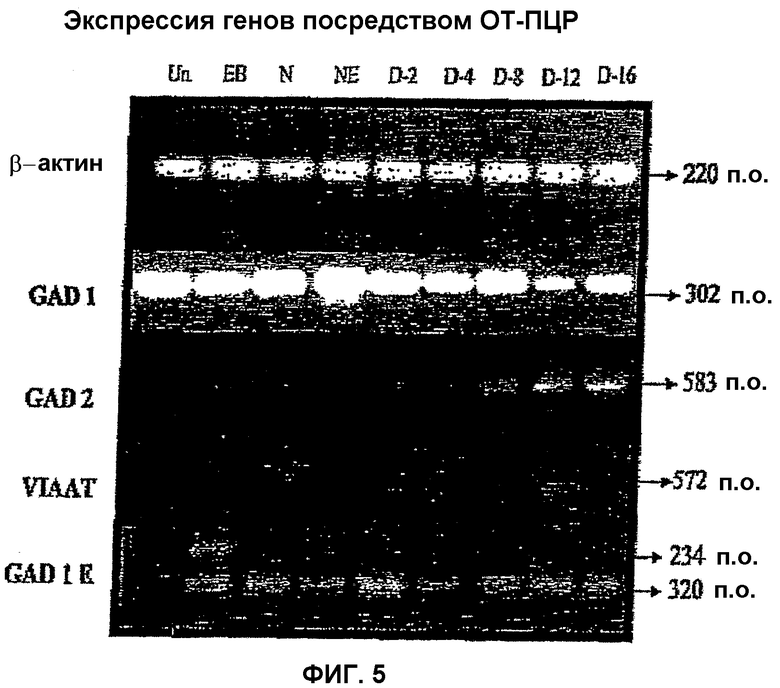




Изобретение относится к области клеточной трансплантологии. Предложена обогащенная популяция клеток, где ГАМКергические нейроны получены в результате дифференцировки популяции полипотентных стволовых клеток, обогащенной нестин-положительными клетками. Высокий выход ГАМКергических нейронов обусловлен селекцией нестин-положительных клеток, повышением содержания (обогащения) нестин-положительных клеток, а затем дальнейшей дифференцировкой в присутствии цитозин-β-d-арабинофуранозида (Ara-С). Изобретение может быть использовано в качестве источника для клеточной заместительной терапии при нейродегенеративных нарушениях и нервных заболеваниях, таких как инсульт, ишемия, эпилепсия и болезнь Гентингтона. 4 н. и 33 з.п. ф-лы, 12 ил., 1 табл.
1. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, в культуре in vitro для использования в клеточной терапии, полученных в результате дифференцировки полипотентных стволовых клеток млекопитающего, за исключением человека, где по меньшей мере 60% дифференцированных клеток представляют собой ГАМКергические нейроны.
2. Клеточная популяция по п.1, где полипотентные стволовые клетки млекопитающего, за исключением человека, представляют собой мышиные эмбриональные стволовые клетки.
3. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, где ГАМКергические нейроны экспрессируют GAD65.
4. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, где ГАМКергические нейроны экспрессируют GAD67.
5. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, где ГАМКергические нейроны экспрессируют рецептор ГАМК-А.
6. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, где ГАМКергические нейроны экспрессируют рецептор ГАМК-Б.
7. Обогащенная популяция ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, где ГАМКергические нейроны экспрессируют рецепторы ГАМК-А и ГАМК-Б.
8. Способ получения обогащенной популяции ГАМКергических нейронов млекопитающего, за исключением человека, по п.1 из полипотентных стволовых клеток млекопитающего, за исключением человека, предусматривающий следующие стадии:
(a) наращивание культуры полипотентных стволовых клеток млекопитающего, за исключением человека;
(b) культивирование полипотентных стволовых клеток для образования эмбриоидных телец и отбора нестин-положительных нейрональных клеток-предшественников в бессывороточной среде в течение 6-10 сут, где бессывороточная среда содержит один или более растворимых факторов, выбранных из группы, состоящей из инсулина, селенита натрия, трансферрина и фибронектина;
(c) наращивание нестин-положительных нейрональных клеток-предшественников; и необязательно дополнительное наращивание нестин-положительных нейрональных клеток-предшественников со стадии (с) в среде для роста клеток ЦНС, где нестин-положительные нейрональные клетки-предшественники наносят на предварительно покрытую поли-L-орнитином или ламинином культуральную чашку, где среда для роста клеток ЦНС содержит один или более растворимых факторов, выбранных из группы, состоящей из добавки N2, добавки В27 и необязательно содержит индуцирующее образование нейронов средство;
(d) дифференцировку нестин-положительных клеток-предшественников с получением популяции дифференцированных клеток в среде для дифференцировки, которая содержит цитозин-β-d-арабинофуранозид (Ara-С), причем популяция дифференцированных клеток содержит по меньшей мере 60% ГАМКергических нейронов; и необязательно предусматривающий стадию
(е) дополнительной дифференцировки популяции дифференцированных клеток со стадии (d) путем культивирования клеток в течение 8-16 сут во второй среде для дифференцировки клеток, которая не содержит цитозин-β-d-арабинофуранозид (Ara-С).
9. Способ по п.8, где популяция дифференцированных клеток содержит наибольшее возможное количество 15% дофаминергических нейронов.
10. Способ по п.8, где популяция дифференцированных клеток содержит не более чем 10% глутаматергических нейронов.
11. Способ по п.8, где популяция дифференцированных клеток содержит не более чем 5% серотонинергических нейронов.
12. Способ по п.8, где популяция дифференцированных клеток содержит наибольшее возможное количество 5% олигодендроцитов.
13. Способ по п.8, где популяция дифференцированных клеток содержит не более чем 5% астроцитов.
14. Способ по п.8, где нестин-положительные нейрональные клетки-предшественники отбирают путем культивирования стволовых клеток в бессывороточной среде.
15. Способ по п.14, где бессывороточная среда представляет собой определенную бессывороточную среду ITSFn.
16. Способ по п.15, где клетки растят в определенной бессывороточной среде ITSFn в течение 6-10 сут.
17. Способ по п.14, где бессывороточная среда содержит один или несколько растворимых факторов, выбранных из группы, состоящей из инсулина, селенита натрия, трансферрина и фибронектина.
18. Способ по п.8, где эмбриоидные тельца культивируют для отбора нестин-положительных нейрональных клеток предшественников.
19. Способ по п.18, где нейрональные клетки-предшественники содержат по меньшей мере 90% нестин-положительных клеток.
20. Способ по п.8, где нестин-положительные нейрональные клетки-предшественники отбирают путем культивирования эмбриоидных телец в бессывороточной среде.
21. Способ по п.20, где бессывороточная среда представляет собой определенную бессывороточную среду ITSFn.
22. Способ по п.20, где бессывороточная среда содержит один или несколько растворимых факторов, выбранных из группы, состоящей из инсулина, селенита натрия, основного фактора роста фибробластов, трансферрина и фибронектина.
23. Способ по п.8, дополнительно предусматривающий наращивание нестин-положительных нейрональных клеток-предшественников со стадии (с) в среде для роста клеток ЦНС.
24. Способ по п.23, где нестин-положительные нейрональные клетки-предшественники наносят на предварительно покрытую поли-L-орнитином или ламинином культуральную чашку.
25. Способ по п.23, где среда для роста клеток ЦНС содержит один или несколько растворимых факторов, выбранных из группы, состоящей из добавки N2, добавки В27, и индуцирующее нейроны средство.
26. Способ по п.25, где индуцирующее нейроны средство представляет собой основной фактор роста фибробластов (bFGF).
27. Способ по п.23, где клетки растят в среде для роста клеток ЦНС в течение 6-10 сут.
28. Способ по п.8, где среда для дифференцировки содержит добавку N2, добавку В27 или обе эти добавки, но не содержит основного фактора роста фибробластов (bFGF).
29. Способ по п.8, где клетки растят в среде для дифференцировки клеток в течение 2 или более суток.
30. Способ по п.8, дополнительно предусматривающий стадию (е), где дополнительной дифференцировке популяции дифференцированных клеток со стадии (d) способствуют путем культивирования клеток во второй среде для дифференцировки клеток, не содержащей цитозин-β-d-арабинофуранозид (Ara-С).
31. Способ по п.30, где популяцию дифференцированных клеток растят во второй среде для дифференцировки клеток в течение 8-16 сут.
32. Способ получения обогащенной популяции нейронов млекопитающего, за исключением человека, по п.1 из нейрональных клеток-предшественников млекопитающего, за исключением человека, предусматривающий обогащение нейрональных клеток-предшественников нестин-положительными клетками и дифференцировку нестин-положительных клеток, с получением ГАМКергических нейронов путем культивирования клеток в присутствии цитозин-β-d-арабинофуранозида (Ara-С).
33. Способ получения модели трансплантации in vitro обогащенной популяции ГАМКергических нейронов млекопитающего, за исключением человека, по п.1, предусматривающий следующие стадии:
(a) выделение клеток гиппокампа взрослой особи;
(b) диссоциацию и культивирование клеток гиппокампа с получением культуры клеток гиппокампа и
(c) культивирование нервных клеток на культуре клеток гиппокампа,
при этом производят оценку выживаемости нервных клеток на культуре клеток гиппокампа.
34. Способ по п.33, где клетки гиппокампа взрослой особи выделяют из мыши.
35. Способ по п.33, где нервные клетки представляют собой ГАМКергические нейроны.
36. Способ по п.35, где более чем 90% ГАМКергических нейронов выживают после одной недели культивирования.
37. Способ по п.33, дополнительно предусматривающий оценку формирования синапса между нервными клетками и культурой клеток гиппокампа.
Приоритет по пунктам:
03.09.2003 - пп.8-37;
31.03.2004 - пп.1-7.
| JP 2002291469, 08.10.2002 | |||
| WO 03000852, 03.01.2003 | |||
| SOSUNOV A.A., CHELYSHEV IUA «Neural stem cells in the brain» | |||
| - Usp | |||
| Fiziol Nauk | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| TAKESHIMA et al., «Mesencephalic type 1 astrocytes rescue dopaminergic neurons from death by deprivation | |||
| - J | |||
| Neurosci | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| KAZANTSEV A | |||
| et al., | |||

