Область техники
Настоящее изобретение относится к микробиологической промышленности, конкретно к способу продукции ароматической L-аминокислоты с использованием бактерии, принадлежащей к семейству Enterobacteriaceae, экспрессирующей дрожжевой ген ARO1, ранее введенный в бактерию.
Описание предшествующего уровня техники
Кластерный ген ARO1 был выделен путем комплементации в Saccharomyces cerevisiae после трансформации полной дрожжевой ДНК-библиотекой BamHI фрагментов рестрикции, встроенных в челночный вектор YEp13. Большинство трансформантов проявляли ожидаемое эписомальное наследование фенотипа ARO+; однако, 1 стабильный трансформант содержал фрагмент ARO1 и вектор YEp13, интегрированные в ARO1 локус. Вставка, содержащая ARO1, представляет собой BamHI фрагмент длиной 17.2 т.п.н., комплементирующий нонсенс- и миссенс- аллели aro1. Субклонирование фрагментов, образовавшихся в результате неполной рестрикции Sau3AI, привело к локализации сегмента ARO1 внутри области длиной 6.2 т.п.н. Автономно реплицирующаяся последовательность была на фрагменте 17.2 т.п.н. Дрожжевые мутанты по ARO1, трансформированные эписомным ARO1, экспрессировали ARO1 ферментативные активности в 5-12 раз выше нормального уровня и содержали повышенное количество белка ARO1. Дрожжевой фрагмент ARO1 также комплементировал aroA, aroB, aroD и aroE мутации Escherichia coli. Экспрессия ARO1 в S.cerevisiae и Е.coli не зависела от ориентации фрагмента в векторе (Larimer, F. et.al.; Molecular и Cellular Biology, 3 (9), 1609-14 (1983)).
Нуклеотидная последовательность гена S.cerevisiae ARO1, кодирующего мультифункциональный фермент биосинтеза ароматических аминокислот (arom multifunctional enzyme), была определена. Последовательность белка для пентафункционального полипептида имеет длину 1588 аминокислот, а ее вычисленная молекулярная масса - 174,555. Функциональные области внутри полипептидной цепи идентифицировали путем сравнения с 5 монофункциональными ферментами E.coli, активности которых соответствовали активностям мультифункционального фермента, Наблюдавшаяся гомология показала, что белок биосинтеза ароматических аминокислот представляет собой набор функциональных доменов, и согласовалась с гипотезой о том, что ген ARO1 эволюционировал по пути объединения Е.coli-подобных генов-предшественников (Duncan, К. et.al.; Biochemical Journal, 246(2), 375-86 (1987)).
Ген ARO1 S. cerevisiae кодирует мультифункциональный фермент биосинтеза ароматических аминокислот. Для получения доказательств того, что экспрессия клонированного гена ARO1 регулируется лимитированием по аминокислотам, использовали специфические ингибиторы биосинтеза аминокислот. Определили, что ARO1 регулируется хорошо охарактеризованным механизмом общего контроля S.cerevisiae. Это обеспечивает очень экономичные средства организации одновременного синтеза 5 ферментов пути шикимата для нужд клетки (Duncan, К. et. al.; FEBS Letters, 241 (1-2), 83-8 (1988)).
В настоящее время нет сообщений, описывающих продукцию ароматических L-аминокислот с использованием бактерии, принадлежащей к семейству Enterobacteriaceae, экспрессирующей введенный ранее ген ARO1.
Описание изобретения
Цели настоящего изобретения включают повышение продуктивности штаммов-продуцентов ароматических L-аминокислот и предоставление способа продукции ароматических L-аминокислот с использованием этих штаммов.
Эта цель была достигнута обнаружением того факта, что экспрессия гена ARO1 из дрожжей в бактерии, принадлежащей к семейству Enterobacteriaceae, увеличивает продукцию ароматической L-аминокислоты этой бактерией.
Целью настоящего изобретения является предоставление бактерии-продуцента ароматической L-аминокислоты, принадлежащей к семейству Enterobacteriaceae, при этом указанная бактерия модифицирована путем введения в нее дрожжевого гена ARO1.
Также целью настоящего изобретения является предоставление описанной выше бактерии, при этом указанная бактерия принадлежит к роду Escherichia.
Также целью настоящего изобретения является предоставление описанной выше бактерии, при этом указанная бактерия - Escherichia coli.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой указанный ген ARO1 является геном Saccharomyces cerevisiae.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой экспрессия указанного введенного гена усилена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой экспрессия указанного введенного гена усилена путем модификации последовательности, контролирующей экспрессию указанного гена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой экспрессия указанного введенного гена усилена путем увеличения числа копий указанного гена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Также целью настоящего изобретения является предоставление способа продукции ароматической L-аминокислоты,: включающего:
- выращивание описанной выше бактерии в питательной среде, и
- выделение указанной ароматической L-аминокислоты из культуральной жидкости.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Также целью настоящего изобретения является предоставление способа получения сложного эфира низших алкилов α-L-аспартил-L-фенилаланина, включающего выращивание описанной выше бактерии в питательной среде, вызывающее образование и накопление L-фенилаланина, при этом указанная бактерия обладает способностью к продукции L-фенилаланина, и синтез сложного эфира низших алкилов α-L-acnapran-L-фенилаланина из аспарагиновой кислоты или ее производных и полученного L-фенилаланина.
Также целью настоящего изобретения является предоставление описанного выше способа, дополнительно включающего этерифицирование L-фенилаланина с целью образования сложного эфира низшего алкила L-фенилаланина, конденсирование сложного эфира низшего алкила L-фенилаланина с производным аспарагиновой кислоты, при этом указанным производным является N-ацил-ангидрид L-аспарагиновой кислоты, выделение сложного эфира низших алкилов N-ацил-α-L-аспартил-L-фенилаланина из реакционной смеси и гидрогенизирование сложного эфира низших алкилов N-ацил-α-L-аспартил-L-фенилаланина для образования сложного эфира простых алкилов α-L-аспартил-L-фенилаланина.
Более детально настоящее изобретение описано ниже.
Наилучший способ осуществления настоящего изобретения
1. Бактерия согласно настоящему изобретению
Бактерия, согласно настоящему изобретению, - это бактерия-продуцент ароматической L-аминокислоты, принадлежащая к семейству Enterobacteriaceae, модифицированная таким образом, что экспрессирует введенный ранее дрожжевой ген ARO1.
Согласно настоящему изобретению «бактерия-продуцент ароматической L-аминокислоты» означает бактерию, обладающую способностью к продукции и выделению ароматической L-аминокислоты в питательную среду, когда бактерия согласно настоящему изобретению выращивается в указанной питательной среде.
Используемый здесь термин «бактерия-продуцент ароматической L-аминокислоты» также означает бактерию, которая способна к продукции ароматической L-аминокислоты и вызывает накопление ароматической L-аминокислоты в ферментационной среде в больших количествах, по сравнению с природным или родительским штаммом Е.coli, таким как штамм Е.coli K-12, и, предпочтительно означает, что указанный микроорганизм способен накапливать в среде целевую L-аминокислоту в количестве не менее чем 0.5 г/л, более предпочтительно, не менее чем 1.0 г/л. Термин «ароматическая L-аминокислота» включает в себя L-фенилаланин, L-тирозин и L-триптофан.
Семейство Enterobacteriaceae включает в себя бактерии, принадлежащие к родам Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Morganella, Yersinia и т.д. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/htbinpost/Taxonomy/Browser/wwwtax.cgi?id=91347).
Предпочтительна бактерия, принадлежащая к роду Escherichia.
Термин "бактерия, принадлежащая к роду Escherichia" означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (Е.coli).
Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть включены в число бактерий согласно настоящему изобретению.
Фраза "бактерия модифицирована путем введения гена" означает, что бактерия модифицирована таким образом, что модифицированная бактерия содержит одну или несколько копий этого гена. В настоящем изобретении введение в бактерию фрагмента ДНК, содержащего ген ARO1, было осуществлено с использованием амплификации гена ARO1 S.cerevisiae методом ПЦР с последующим клонированном гена в векторе, способном функционировать в Е.coli, и трансформацией Е.coli вектором, содержащим ген ARO1.
Методы усиления экспрессии гена включают увеличение числа копий гена. Введение гена в вектор, способный функционировать в бактерии семейства Enterobacteriaceae, позволяет увеличить число копий гена. Предпочтительно использование низкокопийных векторов. Примеры низкокопийных векторов включают, но не ограничиваются ими, pSC101, pMW118, pMW119 и т.п. Термин "низкокопийный вектор" используется для векторов, число копий которого - до 5 на клетку.
Усиление экспрессии гена также может быть достигнуто путем введения множества копий гена в хромосому бактерии, например, методом гомологичной рекомбинации. Mu интеграции, и т.п. Например, один акт µ-интеграции позволяет ввести в хромосому бактерии до 3 копий гена.
Увеличение числа копий гена ARO1 может также быть достигнуто путем введения множества копий гена ARO1 в хромосомную ДНК бактерии. Для того чтобы ввести множество копий гена в бактериальную хромосому, выполняют гомологичную рекомбинацию с использованием в качестве целевой последовательности такую, которая присутствует на хромосомной ДНК во множестве копий. Последовательности, присутствующие на хромосомной ДНК во множестве копий, включают, но не ограничиваются ими, повторяющуюся ДНК, инвертированные повторы на концах транспонируемых элементов. Также возможно включать ген ARO1 в транспозон с последующим переносом для введения множества копий гена в хромосомную ДНК.
Усиление экспрессии гена также может быть достигнуто путем подстановки ДНК настоящего изобретения под контроль сильного промотора. Например, известны как сильные промоторы Ptac промотор, lac промотор, trp промотор, trc промотор, PR или PL промоторы фага λ. Использование сильного промотора можно сочетать с увеличением копийности гена.
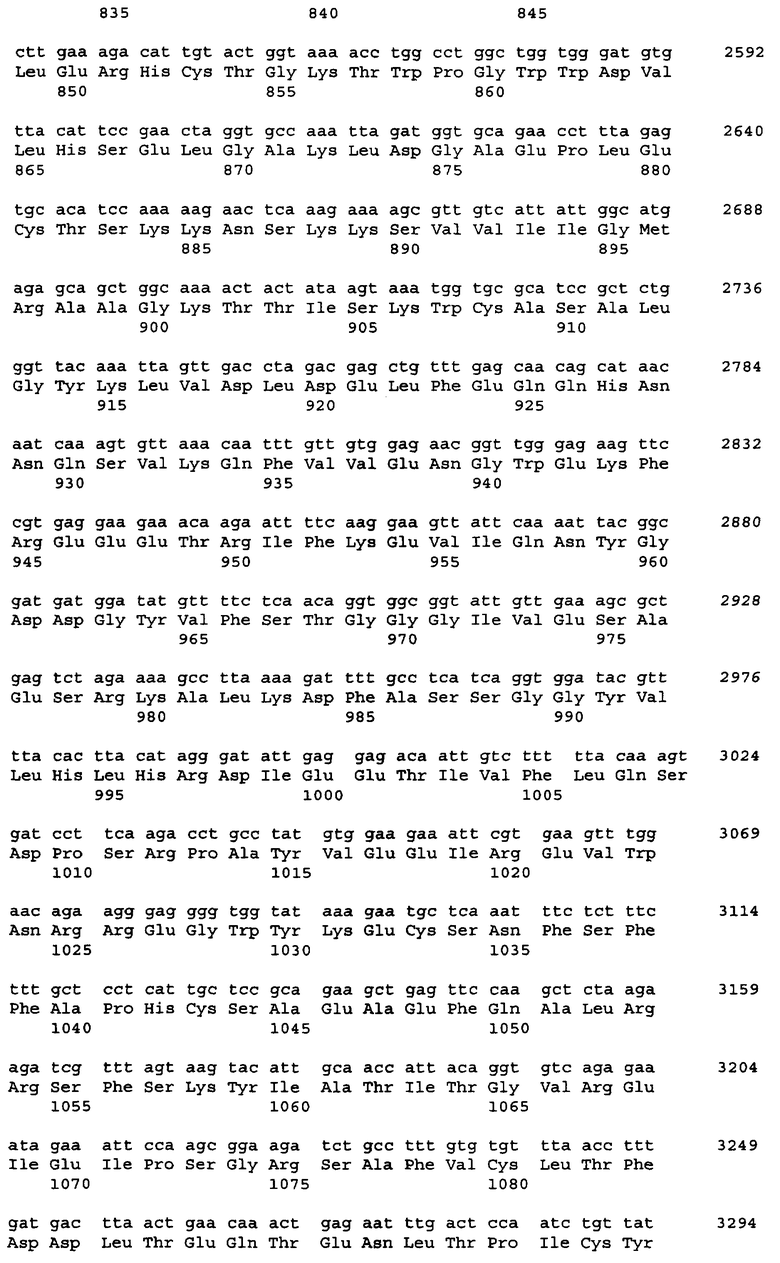
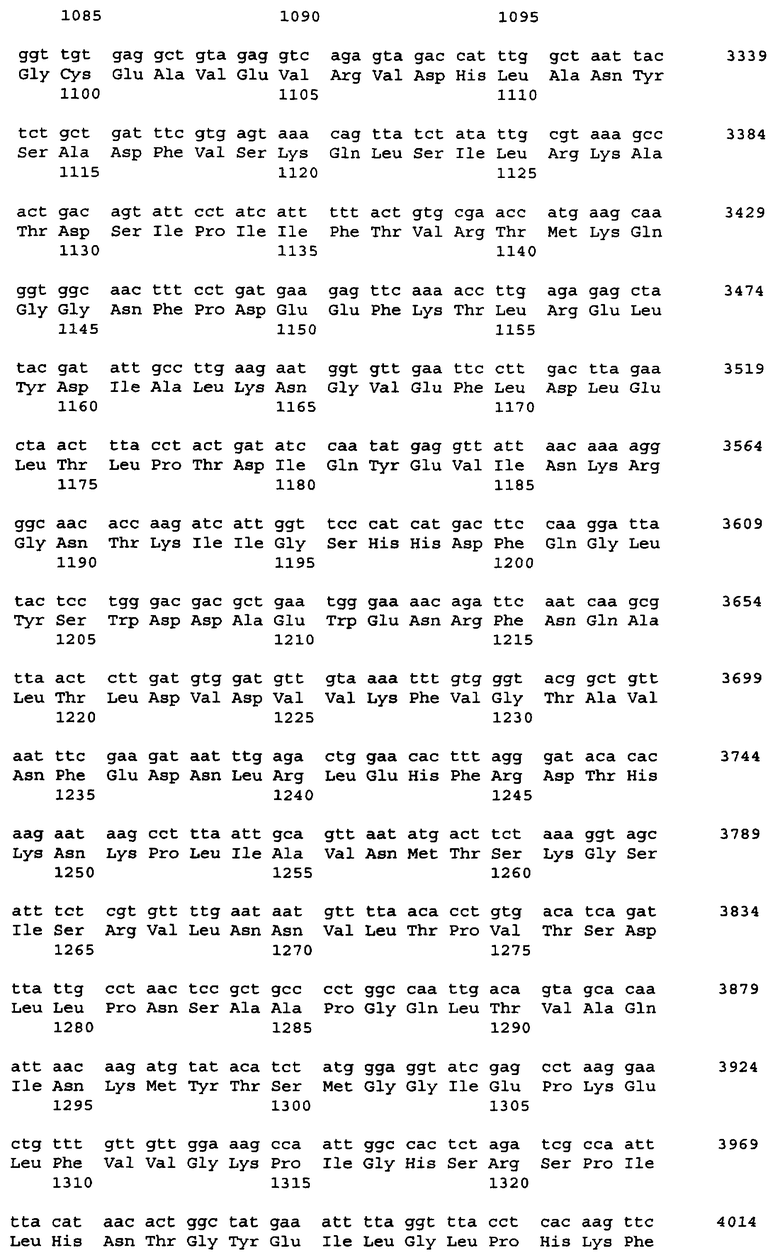
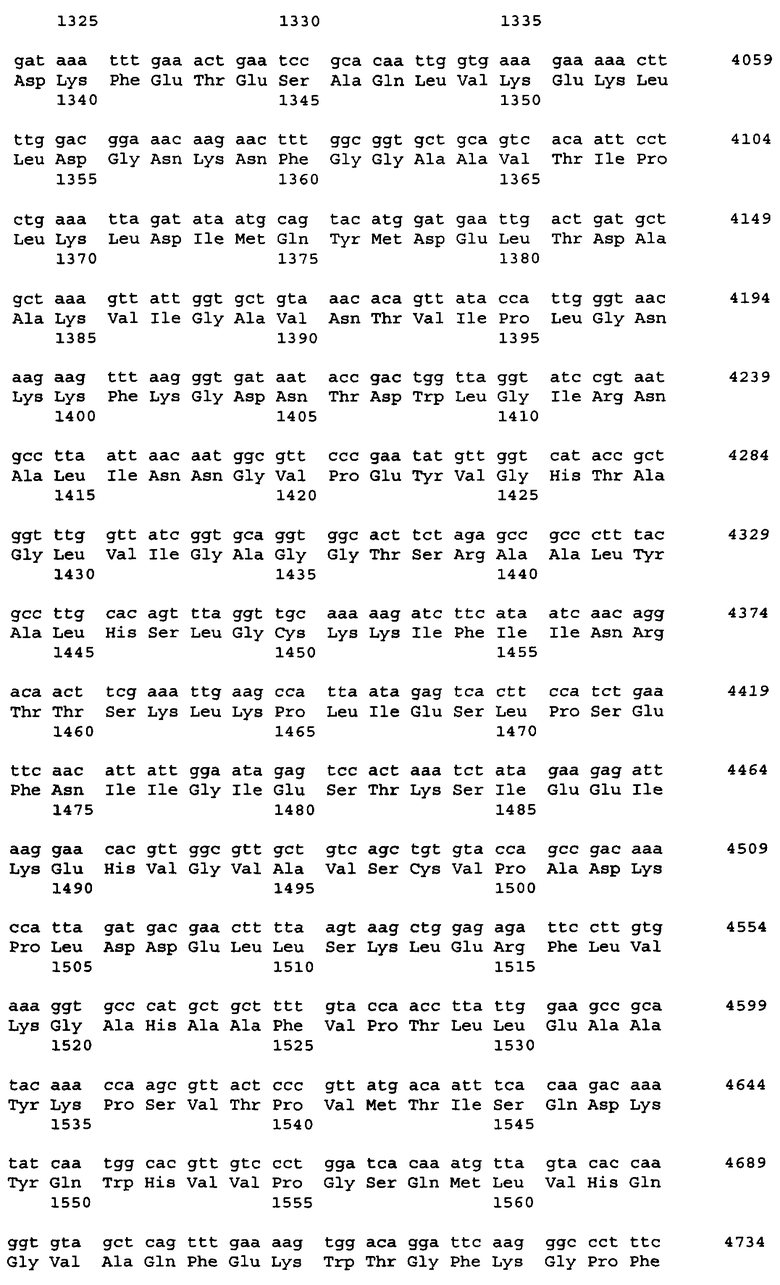
Примеры источника гена ARO1, кодирующего мультифункциональный фермент биосинтеза ароматических аминокислот, включают штаммы Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris и Cuida albicans. Нуклеотидная последовательность гена ARO1 S.cerevisiae DBY-746 и аминокислотная последовательность белка, кодируемого геном ARO1, показаны в Перечне последовательностей - SEQ ID NO:1 и SEQ ID NO:2, соответственно.
С другой стороны, действие промотора может быть усилено, например, путем введения мутаций в промотор с целью повышения уровня транскрипции гена, находящегося за этим промотором. Кроме того, известно, что замена нескольких нуклеотидов в промежутке между сайтом связывания (ribosome binding site - RBS) и стартовым кодоном, особенно в последовательности, находящейся непосредственно перед стартовым кодоном, значительно влияет на транслируемость мРНК. Например, обнаружена 20-кратная разница в уровне экспрессии в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold et al., Annu. Rev. Microbiol., 35, 365-403, 1981; Hui et al., EMBO J., 3, 623-629, 1984).
Поскольку у представителей различных родов и штаммов дрожжей возможны некоторые вариации в нуклеотидных последовательностях, понятие гена ARO1, который вводят в бактерию, не ограничивается геном, последовательность которого приведена в Перечне последовательностей под номером SEQ ID No:1, но также может включать и гены, гомологичные SEQ ID No:1, кодирующие варианты белка ARO1. Термин "вариант белка", используемый в настоящем изобретении, означает белок с изменениями в последовательности, будь то делеции, вставки, добавления или замены аминокислот. Число изменений в варианте белка зависит от положения или типа аминокислотного остатка в третичной структуре белка. Оно может быть от 1 до 30, предпочтительно от 1 до 15, более предпочтительно от 1 до 5 в SEQ ID No:2. Данные изменения в вариантах белка являются консервативными мутациями, при которых сохраняется функция белка. Другими словами, данные изменения могут иметь место в областях белка, некритичных для его трехмерной структуры. Это становится возможным благодаря тому, что некоторые аминокислоты обладают высокой гомологией друг к другу, и поэтому третичная структура при таких заменах не нарушается. Консервативная мутация - это мутация, при которой имеют место взаимные замены среди Phe, Trp, Туr, если сайт замены - ароматическая аминокислота; среди Leu, Ile, Val, если сайт замены - гидрофобная аминокислота; между Gln, Asn, если сайт замены - положительно заряженная аминокислота; среди Lys, Arg, His, если сайт замены - основная аминокислота; между Asp, Glu, если сайт замены - кислая аминокислота, и между Ser, Thr, если это аминокислота с гидроксильной группой. Консервативные замены являются типичными консервативными мутациями. Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ilе на Leu, Met, Val или Phe, замену Leu на Ilе, Met, Val или Phe, замену Lys на Asn, Glu, Gin, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp, и замену Val на Met, Ile или Leu. Описанные выше замены, делеции, вставки, добавления, перестановки и т.п. одного или нескольких аминокислотных остатков включают природные мутации (мутант или вариант) в зависимости от видовых различий или индивидуальных различий микроорганизмов, содержащих ген ARO1. Такой ген может быть получен модифицированием нуклеотидной последовательности, показанной в SEQ ID NO:1, с использованием, например, сайт-специфического мутагенеза, таким образом, что сайт-специфический аминокислотный остаток в кодируемом белке включает замены, делеции, вставки или добавления.
Кроме того, варианты белков, кодируемых генами ydiN и ydiB, могут иметь гомологию не менее 80%, предпочтительно не менее 90%, и наиболее предпочтительно не менее 95%, по отношению к полной аминокислотной последовательности, показанной в Перечне последовательностей под номером 2 (SEQ ID No:2).
Кроме того, ген ARO1 может быть вариантом, который гибридизуются в жестких условиях с нуклеотидной последовательностью, приведенной в Перечне последовательностей под номером 1 (SEQ ID No:1), или с зондом, который может быть синтезирован на основе указанной нуклеотидной последовательности. «Жесткие условия» включают такие условия, при которых специфические гибриды, например, гибриды с гомологией не менее 60%, предпочтительно не менее 70%, более предпочтительно не менее 80%, еще более предпочтительно не менее 90% и наиболее предпочтительно не менее 95%, образуются, а неспецифические гибриды, например, гибриды с меньшей гомологией, чем указано выше, - не образуются. Практическим примером жестких условий является однократная отмывка, предпочтительно двух- или трехкратная, при концентрации солей 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS, при 60°С. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для нейлоновой мембраны Hybond™ N+ (Amersham) при строгих условиях - 15 минут. Предпочтительна двух-, трехкратная отмывка. Длина зонда может быть выбрана в зависимости от условий гибридизации, обычно от 100 н.п. до 1 т.н.п.
Гомология между двумя аминокислотыми последовательностями может быть определена с использованием известных методов, например, компьютерной программы BLAST 2.0.
Изобретатели настоящего изобретения предположили, что неэффективная экспрессия клонированного дрожжевого гена в бактерии объясняется присутствием значительного количества в кодирующей части очень редких для E.coli кодонов. Эти кодоны присутствуют в генах E.coli в исключительно редких случаях, кроме того, предполагается важная регуляторная роль этих кодонов (или, возможно, соответствующих тРНК) в E.coli. В S.cerevisiae, напротив, эти типы кодонов встречаются наиболее часто. Изобретатели настоящего изобретения предложили несколько объяснений, почему присутствие редких кодонов может значительно снижать уровень экспрессии ARO1 в E.coli: возможный сдвиг рамки считывания в положении, где два редких кодона следуют друг за другом; возможная нестабильность мРНК из-за неэффективной элонгации трансляции; неправильное сворачивание белка из-за низкой (вместо высокой) скорости трансляции. Представляется целесообразным заменить такие кодоны другими, присутствующими в E.coli более часто. Другой путь решения этой проблемы - трансформация бактериальной клетки плазмидой, содержащий ген, кодирующий тРНК, способную узнавать эти редкие кодоны. Конкретный пример такой плазмиды - плазмида pDY(Arg4-tRNA) (Gurskiy Ia. G., et al, Mol Biol (Mosk), 26 (5): 1080-7 (1992)), содержащая ген argU, кодирующий Arg4-tRNA, способный узнавать кодоны AGG и AGA.
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия - продуцент ароматической L-аминокислоты
В качестве бактерии согласно настоящему изобретению, модифицированной путем введения дрожжевого гена ARO1, может быть использована бактерия, способная к продукции ароматической L-аминокислоты.
L-триптофан, L-фенилаланин и L-тирозин являются ароматическими аминокислотами и имеют общий путь биосинтеза. Примеры генов, кодирующих ферменты биосинтеза этих ароматических аминокислот, включают гены, кодирующие дезоксиарабиногептулозонатфосфатсинтазу (aroG), 3-дегидрохиннатсинтазу (aroB), шикиматдегидрогеназу, шикиматкиназу (aroL), 5-енолпирувилшикимат-3-фосфатсинтазу (aroA) и хоризматсинтазу (aroC) (EP763127). Следовательно, способность к продукции ароматических аминокислот может быть усилена увеличением числа копий генов, кодирующих эти ферменты, на плазмиде или хромосоме. Известно, что эти гены могут контролироваться регулятором транскрипции (tyrR), поэтому активность ферментов биосинтеза ароматических аминокислот может также быть увеличена путем делеции гена tyrR (EP 763127).
Для увеличения продукции ароматических аминокислот в бактерии может быть ослаблен биосинтез других аминокислот, иных чем целевая ароматическая амнокислота. Например, когда целевая аминокислота - L-триптофан, может быть ослаблен биосинтез L-фенилаланина и/или L-тирозина (US 4371614). Кроме того, 3-дезокси-D-арабиногептулозонат-7-фосфатсинтетаза (DS), кодируемая генами aroF или aroG, подвержена ингибированию по типу обратной связи ароматическими аминокислотами. Следовательно, бактерия может быть модифицирована таким образом, чтобы содержать мутантную DS, не подверженную ингибированию по типу обратной связи. Такая мутантная DS может быть получена, например, заменой в aroF L-аспарагиновой кислоты в положении 147 или L-серина в положении 181 на другую аминокислоту. В случае aroG мутантная DS может быть получена, например, заменой L-аспарагиновой кислоты в положении 146, L-метионина в положении 147, L-пролина в положении 150 или L-аланина в положении 202 на другую аминокислоту или заменой L-метионина в положении 157 и L-аланина в положении 219 на другую аминокислоту. Бактерия-продуцент ароматической L-аминокислоты может быть получена введением в бактерию мутантного гена, кодирующего такую матантную DS (EP 0488424).
Бактерия-продуцент L-фенилаланина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-фенилаланина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм AJ 12739 (tyrA::Tn10, tyrR) (ВКМП В-8197); штамм HW1089 (АТСС-55371), содержащий ген pheA34 (патент США 5354672); мутантный штамм MWEC101-b (KR 8903681); штаммы NRRL B-12141, NRRL В-12145, NRRL В-12146 и NRRL В-12147 (патент США 4407952) и подобные им. Также в качестве родительских штаммов могут быть использованы бактерии, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм E.coli K-12[W3110(tyrA)/pPHAB] (FERM ВР-3566), штамм E.coli К-12[W3110(tyrA)/pPHAD] (FERM ВР-12659), штамм E.coli K-12[W3110(tyrA)/pPHATerm] (FERM BP-12662) и штамм E.coli K-12[W3110(tyrA)/pBR-aroG4, pACMAB], названный как AJ12604 (FERM BP-3579) (Европейский патент ЕР 488424 В1). Кроме того, также могут быть использованы бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia с повышенной активностью белков, кодируемых геном yedA или геном yddG (патентные заявки США 2003/0148473 А1 и 2003/0157667 А1).
Бактерия-продуцент L-триптофана
Примеры родительских штаммов, которые могут использоваться для получения бактерии-продуцента L-триптофана согласно настоящему изобретению, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к роду Escherichia, такие как Е.coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), лишенные активности триптофанил-тРНК синтетазы, кодируемой мутантным геном trpS (патент США 5756345); Е.coli SV164 (pGH5), содержащий аллель serA, кодирующий фосфоглицератдегидрогеназу, не ингибируемую серином по типу обратной связи, и аллель trpE, кодирующий антранилатсинтазу, не ингибируемую триптофаном по типу обратной связи (патент США 6180373); Е. coli AGX17 (pGX44) (NRRL В-12263) и AGX6(pGX50)aroP (NRRL B-12264), в которых отсутствует активность триптофаназы (патент США 4371614); штамм Е.coli AGX17/pGX50, pACKG4-pps, в котором усилена способность к синтезу фосфоенолпирувата (заявка РСТ WO 9708333, патент США 6319696), и подобные им. Также могут быть использованы бактерии - продуценты L-триптофана, принадлежащие к роду Escherichia, в которых увеличена активность белка, кодируемого геном yedA или геном yddG (заявки на патент США 2003/0148473 А1 и 2003/0157667 А1).
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-триптофана, согласно настоящему изобретению также включают в себя штаммы, в которых увеличена активность одного или нескольких ферментов, выбранных из группы, состоящей из антранилатсинтазы, фосфоглицератдегидрогеназы и триптофансинтазы. И антранилатсинтаза, и фосфоглицератдегидрогеназа подвержены ингибированию L-триптофаном и L-серином по типу обратной связи, так что в эти ферменты могут быть введены мутации, снижающие чувствительность к ингибированию по типу обратной связи. Конкретные примеры штаммов с такой мутацией включают E.coli SV164, антранилатсинтаза которой не чувствительна к ингибированию по типу обратной связи, и штамм-трансформант, полученный введением в Е.coli SV164 плазмиды pGH5 (заявка РСТ WO 94/08031), которая содержит мутантный ген serA, кодирующий фосфоглицератдегидрогеназу, которая не чувствительна к ингибированию по типу обратной связи.
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-триптофана, согласно настоящему изобретению также включают в себя штаммы, в которые введен триптофановый оперон, содержащий ген, кодирующий антранилатсинтазу, которая не чувствительна к ингибированию по типу обратной связи (заявка Японии 57-71397 А, заявка Японии 62-244382 А, патент США 4371614). Кроме того, способность к продукции L-триптофана может быть придана путем усиления экспрессии гена (из триптофанового оперона), кодирующего триптофансинтазу (trpBA). Триптофансинтаза состоит из двух субъединиц α и β, которые кодируются trpA и trpB соответственно. Кроме того, способность к продукции L-триптофана может быть увеличена усилением экспрессии оперона изоцитратлиазы-малатсинтазы (заявка РСТ WO 2005/103275).
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-триптофана, согласно настоящему изобретению также включают в себя штаммы, содержащие мутации, приводящие к конститутивной высокоэффективной транскрипции генов Trp-оперона, снижающие ингибирование по типу обратной связи ферментов, вовлеченных в путь биосинтеза Trp, и блокирующие способность клеток к деградации триптофана. Конкретные примеры штаммов, содержащих такие мутации, включают штамм E.coli 202 (WO 2005/103275), содержащий мутации ΔtrpR, trpLE8Fbr, Δtna и штамм E.coli 202ΔiclR, дополнительно содержащий делецию гена iclR, кодирующего репрессор транскрипции IclR.
2. Способ согласно настоящему изобретению.
Способом согласно настоящему изобретению является способ получения ароматической L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления ароматической L-аминокислоты в питательной среде и выделения ароматической L-аминокислоты из культуральной жидкости.
Согласно настоящему изобретению выращивание, выделение и очистка ароматической L-аминокислоты из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Выбранная для выращивания питательная среда может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от характера ассимиляции выбранного микроорганизма могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно, выращивание в течение от 1 до 5 дней приводит к накоплению целевой ароматической L-аминокислоты в культуральной среде.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем ароматическая L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и/или кристаллизации.
Фенилаланин, образующийся способом настоящего изобретения, может использоваться, например, для получения сложного эфира низших алкилов α-L-аспартил-L-фенилаланина (также называемого "аспартам"). А именно, способ настоящего изобретения включает способ получения сложного эфира низших алкилов α-L-аспартил-L-фенилаланина с использованием L-фенилаланина в качестве сырья. Способ включает синтез сложного эфира низших алкилов α-L-аспартил-L-фенилаланина из L-фенилаланина, получаемого описанным выше способом настоящего изобретения, и аспарагиновой кислоты или ее производных. В качестве сложных эфиров низших алкилов могут быть упомянуты метиловый эфир, этиловый эфир, пропиловый эфир и т.п.
В способе настоящего изобретения процесс синтеза сложного эфира низших алкилов α-L-аспартил-L-фенилаланина из L-фенилаланина и аспарагиновой кислоты или ее производных никак особо не ограничивается и может использоваться любой традиционный способ при условии, что для синтеза сложного эфира низших алкилов α-L-аспартил-L-фенилаланина могут использоваться L-фенилаланин или его производные. Конкретно, сложный эфир низших алкилов α-L-аспартил-L-фенилаланин может быть получен, например, следующим способом (US Pat. No 3786039). L-фенилаланин этерифицируется с целью получения сложного эфира низшего алкила L-фенилаланина. Сложный эфир низшего алкила L-фенилаланина взаимодействует с производным L-аспарагиновой кислоты, у которого аминогруппа и β-карбоксильная группа защищены, а α-карбоксильная группа свободна для этерификации. К производным относятся N-ацилангидрид L-аспарагиновой кислоты, такой как N-формил-, N-карбобензокси- или N-p-метоксикарбобензокси- ангидрид L-аспарагиновой кислоты. При реакции конденсации образуется смесь N-ацил-α-L-аспартил-L-фенилаланина и N-ацил-β-L-аспартил-L-фенилаланина. Если реакция осуществляется в присутствии органической кислоты, константа диссоциации которой при 37°С 10-4 или менее, отношение α-формы к β-форме в смеси увеличивается (Japanese Patent Laid-Open Publication No. 51-113841). Затем N-ацил-α-L-аспартил-L-фенилаланин выделяется из смеси и гидрогенизируется с целью получения α-L-аспартил-L-фенилаланина.
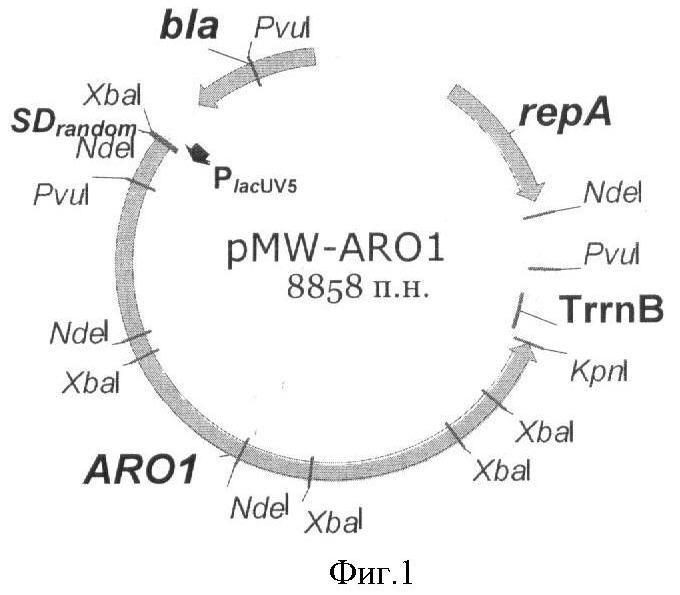
Краткое описание фигур.
На Фигуре 1 изображена схема плазмиды pMW-aro1, содержащей ген ARO1.
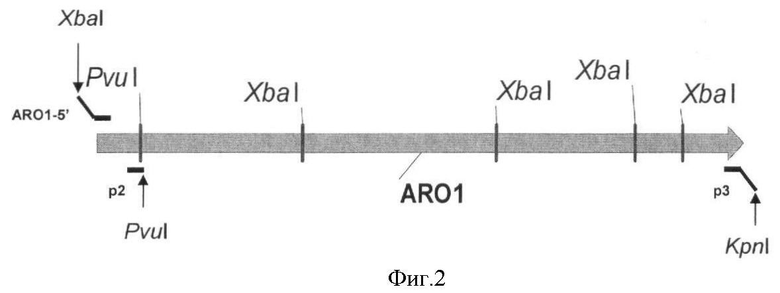
На Фигуре 2 изображено положение праймеров ARO1-5', р2 и p3 на гене ARO1.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Продукция L-триптофана штаммом Е.coli AJ12739-ΔydiN.
Для оценки влияния экспрессии гена ARO1 на продукцию L-триптофана штамм Е.coli 202 трансформировали различными вариантами плазмиды pMW-arol (конструирование плазмиды pMW-arol описано в Справочном примере 1): pMW-aro1-3 (содержащей ACAAGG SD-последовательность) и pMW-aro1-6 (содержащей AATAGG SD- последовательность). Таким образом были сконструированы штаммы 202[pMW-aro1-3] и 202[pMW-aro1-6], соответственно. Эти штаммы затем трансформировали плазмидой pDY(Arg4-t RNA).
Штаммы Е.coli, 202, 202 [pMW119], 202 [pMW-aro1-3], 202 [pMW-aro1-6], 202 [pMW119, pDY(Arg4-t RNA)], 202 [pMW-aro1-3, pDY(Arg4-t RNA)] и 202 [pMW-aro1-6, pDY(Arg4-t RNA)] выращивали при 32°С в течение 18 часов в питательном бульоне. 0.3 мл полученных культур вносили в 3 мл ферментационной среды с добавлением ИПТГ (0.1 мМ) в пробирки размером 20×200 мм и культуры выращивали при 32°С в течение 72 часов на роторной качалке (250 об/мин). По окончании ферментации количество накопленного в среде триптофана определяли с помощью тонкослойной хроматографии (TLC). Для этой цели использовали TLC-пластинки размером 10×15 см, покрытые 0.11 мм-слоем силикагеля Сорбфил без флуоресцентного индикатора (Акционерное Общество Сорбполимер, Краснодар, Россия). Пластинки Сорбфил экспонировали в подвижной фазе следующего состава: пропан-2-ол: этилацетат: 25% водного аммиака: вода = 40:40:7:16 (v/v). Раствор (2%) нингидрина в ацетоне использовали для визуализации.
Результаты по крайней мере восьми независимых пробирочных ферментации приведены в Таблице 1. Как следует из Таблицы 1, 202 [pMW-aro1-3] накапливал больше L-триптофана, чем 202 и 202 [pMW119]; 202 [pMW-aro1-3, pDY (Arg4-t RNA)] и 202 [pMW-aro1-6, pDY (Arg4-t RNA)] накапливали больше L-триптофана, чем 202, 202 [pMW119], 202 [pMW-aro1-3], 202[pMW-aro1-6], 202 [pMWl 19, pDYtArg4-t RNA)].
Компоненты использованной ферментационной среды представлены в Таблице 2; группы компонентов А, В, С, D, Е, F и Н стерилизуют отдельно, как и показано в Таблице 2, чтобы избежать нежелательных взаимодействий во время стерилизации.
Пример 2. Продукция L-фенилаланина штаммом Е.coli, содержащим ген ARO1.
Для оценки влияния экспрессии гена ARO1 на продукцию L-фенилаланина штамм-продуцент L-фенилаланина Е.coli AJ12739 может быть трансформирован различными вариантами плазмиды pMW-arol. Полученный штамм может быть затем трансформирован плазмидой pDY(Arg4-t RNA).
Штаммы Е.coli, AJ 12739[pMW-aro1, pDY (Arg4-t RNA)], AJ 12739[pMW-arol] и AJ 12739 могут быть выращены при 37°С в течение 18 часов в питательном бульоне, 0.3 мл полученных культур может быть внесено в 3 мл ферментационной среды в пробирки размером 20×200 мм и культуры могут быть выращены при 37°С в течение 48 часов на роторной качалке. По окончании ферментации количество накопленного в среде фенилаланина может быть определено с помощью тонкослойной хроматографии (TLC) как описано в Примере 1.
Может быть использована ферментационная среда следующего состава (г/л):
Глюкозу и сульфат магния стерилизуют отдельно. СаСО3 стерилизуют сухим жаром при 180°С в течение 2 часов. рН доводят до 7.0.
Справочный пример 1. Конструирование плазмид pMW119-ARO1-3 и pMW119-ARO1-6.
Поскольку ген ARO1 содержит четыре сайта рестрикции XbaI, плазмиды pMW119-ARO1 сконструировали путем лигирования следующих трех фрагментов ДНК:
1) фрагмент PvuI-KpnI (4790 п.н.) гена ARO1 получили методом ПЦР с использованием праймеров ARO1-5' (SEQ ID NO:3, где R=A/G, N - любой нуклеотид) и р3 (SEQ ID NO:4) и геномной ДНК S. cerevisiae DBY-746 (этот штамм можно получить в The Yeast Genetic Stock Center, Berkeley, Calif. 94720 или в American Type Culture Collection, Rockville, Md. 20852) в качестве матрицы. Праймер ARO1-5' содержит сайт узнавания для эндонуклеазы XbaI, вырожденную SD-последовательность, и сайт узнавания для эндонуклеазы NdeI, содержащий ATG-кодон гена ARO1. Праймер р3 содержит сайт узнавания для эндонуклеазы KpnI. Полученный продукт ПЦР последовательно обработали эндонуклеазами PvuI и KpnI.
2) фрагмент XbaI-PvuI (338 п.н.) гена ARO1 получили методом ПЦР с использованием праймеров ARO1-5' (SEQ ID NO:3) и р2 (SEQ ID NO:5) и геномной ДНК S. cerevisiae DBY-746 в качестве матрицы. Полученный продукт ПЦР последовательно обработали эндонуклеазами XbaI и PvuI.
3) большой XbaI-KpnI фрагмент плазмиды pMW119-PlaclacI получили путем обработки pMW119-PlaclacI эндонуклеазами XbaI и KpnI. Вектор pMW119-PlaclacI сконструировали таким же образом, как и вектор pMW118-PlaclacI (Skorokhodova, A.Yu. et al., Biotekhnologiya (Rus), 5, 3-21 (2004)), за исключением того, что содержит полилинкер из плазмиды pMW119 вместо pMW118.
Получили библиотеку плазмид pMW119-ARO1 с рандомизированной SD-последовательностью гена ARO1. Среди этих плазмид выделили плазмиды, способные комплементировать мутации aroA-, aroB-, aroD- и aroE- в соответствующих штаммах Е. coli. Получены и секвенированы только две плазмиды с требуемыми свойствами.
Плазмида pMW-ARO1-3 содержит ACAAGG SD-последовательность, a pMW-ARO1-6 содержит AATAGG SD-последовательность.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.
Промышленная применимость
Согласно настоящему изобретению в бактерии, принадлежащей к семейству Enterobacteriaceae, может быть усилена продукция ароматических L-аминокислот.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| МУТАНТНЫЙ БЕЛОК, КОДИРУЕМЫЙ ГЕНОМ yddG, И СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ РОДА Escherichia | 2012 |
|
RU2530171C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНА pckA | 2003 |
|
RU2268300C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН hipA | 2005 |
|
RU2320718C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНОВ УТИЛИЗАЦИИ КСИЛОЗЫ | 2005 |
|
RU2283346C1 |
| 6-ФОСФОГЛЮКОНОЛАКТОНАЗА ИЗ ESCHERICHIA COLI, ФРАГМЕНТ ДНК, БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2005 |
|
RU2288268C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА ИЛИ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН fdrA | 2006 |
|
RU2337957C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН pnp, - ПРОДУЦЕНТ L-ТРЕОНИНА, И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2005 |
|
RU2330883C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН leuO | 2006 |
|
RU2312894C1 |












Изобретение относится к биотехнологии и представляет собой способ получения ароматической L-аминокислоты. Способ включает выращивание бактерии семейства Enterobacteriaceae, которая модифицирована путем введения дрожжевого гена ARO1, и выделение аминокислоты. Изобретение относится также к способу получения сложного эфира низших алкилов α-L-аспартил-L-фенилаланина, включающего выращивание указанной бактерии, накопление L-фенилаланина и синтез сложного эфира низших алкилов α-L-аспартил-L-фенилаланина из аспарагиновой кислоты или ее производных и полученного L-фенилаланина. Изобретение позволяет получать ароматические аминокислоты и сложные эфиры с высокой степенью эффективности. 3 н. и 9 з.п. ф-лы, 2 ил., 2 табл.
1. Бактерия семейства Enterobacteriaceae - продуцент ароматической L-аминокислоты, отличающаяся тем, что указанная бактерия модифицирована путем введения дрожжевого гена ARO1.
2. Бактерия по п.1, отличающаяся тем, что указанная бактерия принадлежит к роду Escherichia.
3. Бактерия по п.1, отличающаяся тем, что указанная бактерия - Escherichia coli.
4. Бактерия по п.1, отличающаяся тем, что указанный ген ARO1 (SEQ ID NO:1) является геном из Saccharomyces cerevisiae.
5. Бактерия по п.1, отличающаяся тем, что экспрессия указанного введенного гена усилена.
6. Бактерия по п.5, отличающаяся тем, что экспрессия указанного введенного гена усилена путем модификации последовательности, контролирующей экспрессию указанного гена.
7. Бактерия по п.5, отличающаяся тем, что экспрессия указанного введенного гена усилена путем увеличения числа копий указанного гена.
8. Бактерия по любому из пп.1-7, отличающаяся тем, что указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
9. Способ получения ароматической L-аминокислоты, включающий
выращивание бактерии по любому из пп.1-8 в питательной среде; и
выделение указанной ароматической L-аминокислоты из культуральной жидкости.
10. Способ по п.9, отличающийся тем, что указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
11. Способ получения сложного эфира низших алкилов α-L-аспартил-L-фенилаланина, включающий
выращивание бактерии по любому из пп.1-8 в питательной среде, приводящее к продукции и накоплению L-фенилаланина, при этом указанная бактерия обладает способностью к продукции L-фенилаланина;
синтез сложного эфира низших алкилов α-L-аспартил-L-фенилаланина из аспарагиновой кислоты или ее производных и полученного L-фенилаланина.
12. Способ по п.11, дополнительно включающий этерификацию L-фенилаланина с образованием сложного эфира низшего алкила L-фенилаланина, конденсирование сложного эфира низшего алкила L-фенилаланина с производным аспарагиновой кислоты, при этом указанным производным является N-ацил-ангидрид L-аспарагиновой кислоты, выделение сложного эфира низших алкилов N-ацил-α-L-аспартил-L-фенилаланина из реакционной смеси и гидрогенизирование сложного эфира низших алкилов N-ацил-α-L-аспартил-L-фенилаланина с образованием сложного эфира низших алкилов α-L-аспартил-L-фенилаланина.
| DUNCAN K et | |||
| al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| An example of the co-ordinate regulation of five enzymes on a single biosynthetic pathway | |||
| FEBS Lett | |||
| Механическая топочная решетка с наклонными частью подвижными, частью неподвижными колосниковыми элементами | 1917 |
|
SU1988A1 |
| US 6346405 B1, 12.02.2002 | |||
| RU 2004117766 A, 20.11.2005 | |||
| RU 2005126704 A, 27.01.2006. | |||

