Область техники, к которой относится изобретение
Настоящее изобретение относится к микробиологической промышленности, в частности к способу получения L-аминокислот, более конкретно ароматических L-аминокислот, таких как L-фенилаланин, L-тирозин и L-триптофан ферментацией с использованием бактерии рода Escherichia, в которой аминокислотный белок-экспортер YddG модифицирован. Мутантный белок и способ его использования значительно улучшают продукцию L-фенилаланина и L-триптофана.
Уровень техники
Традиционно L-аминокислоты в промышленности получают способом ферментации с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано много способов повышения продукции L-аминокислот, например путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США №427,765). Эти способы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот, и/или снятии ингибирования ключевых ферментов продуцируемой L-аминокислотой по типу обратной связи (см., например, патент Японии, публикация №56-18596 (1981), WO 95/16042 или патенты США №5661012 и 6,040,160).
С другой стороны, повышенная экскреция L-аминокислот может способствовать увеличению продуктивности штамма, продуцирующего L-аминокислоту. Описан штамм бактерии рода Corynebacterium, обладающей повышенной экспрессией гена экскреции L-лизина (ген lysE) (WO 9723597A2). Кроме того, описаны гены, кодирующие белки, способные к секреции L-цистеина, L-цистина, N-ацетилсерина или производных триазолидина (патент США №5,972,633).
К настоящему времени описаны несколько генов из Escherichia coli (Е.coli), кодирующих, как предполагается, мембранные белки, которые увеличивают продукцию L-аминокислот. Дополнительная копия гена rhtB делает бактерию более устойчивой к L-гомосерину, что способствует увеличению продукции L-гомосерина, L-треонина, L-аланина, L-валина и L-изолейцина (Европейская патентная заявка ЕР994190 А2). Дополнительная копия гена rhtC делает бактерию более устойчивой к L-гомосерину и L-треонину, что способствует увеличению продукцию L-гомосерина, L-треонина и L-лейцина (Европейская патентная заявка ЕР 1013765 А1). Дополнительные копии генов yahN, yeaS, yfiK и yggA увеличивают продукцию L-глутаминовой кислоты, L-лизина, L-треонина, L-аланина, L-гистидина, L-пролина, L-аргинина, L-валина и L-изолейцина (Европейская патентная заявка ЕР 1016710 А2).
Было найдено, что ген rhtA расположен на хромосоме Е.coli вблизи оперона glnHPQ, который кодирует компоненты транспортной системы глутамина, и что ген rhtA идентичен открытой рамке считывания (OPC) ybiF, расположенной между генами рехВ и ompX. Элемент, экспрессирующий белок, кодируемый ОРС, был обозначен как ген rhtA (rht есть сокращение от resistance to homoserine and threonine - устойчивость к гомосерину и треонину).
Дополнительно было обнаружено, что амплификация гена rhtA также придает устойчивость к гомосерину и треонину. Мутация rhtA23 является заменой А на G в положении -1 относительно стартового кодона ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, Calif., Aug. 24-29, 1997, abstract No. 457). Известно, что состав нуклеотидов между последовательностью Шайна-Дальгарно (Shine-Dalgarno, SD) и стартовым кодоном, а в особенности последовательность непосредственно перед стартовым кодоном очень сильно влияет на транслируемость мРНК. Было обнаружено 20-кратное изменение уровня экспрессии гена в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold et al., Annu. Rev. Microbiol., 1981, 35:365-403; Hui et al., EMBO J., 1984, 3:623-629). Поэтому было высказано предположение, что мутация rhtA23 увеличивает экспрессию гена rhtA.
Ген rhtA кодирует белок, состоящий из 295 аминокислотных остатков и имеющий рассчитанный молекулярный вес 31.3 kDa. Анализ последовательности RhtA показал, что это весьма гидрофобный белок, содержащий 10 предсказанных трансмембранных сегментов. Поиск с помощью программы PSI-BLAST в нуклеотидной последовательности генома штамма Е.coli K-12, принадлежащего к роду Escherichia, выявил по меньшей мере 10 белков-паралогов RhtA (Blattner F.R. et al., Science, 1997, 277:1453-1474). Среди них оказались белки, кодируемые генами ydeD и yddG. Было показано, что ген ydeD вовлечен в процесс выброса из клетки веществ - метаболитов биосинтеза цистеина (Dassler Т. et al., Mol. Microbiol., 2000, 36:1101-1112; патент США №5,972,663).
Авторами настоящего изобретения белок YddG был идентифицирован как трансмембранный белок, образованный 10 спиралями (Airich L.G. et al., J. Mol. Microbiol. Biotechnol., 2010, 19:189-197) и способный в Е.coli экспортировать ароматические аминокислоты (Doroshenko V.G. et al., FEMS Microbiol. Lett., 2007, 275:312-318). Было обнаружено, что повышенная активность белка YddG положительно влияет на продукцию L-фенилаланина и L-триптофана мутантным штаммом Е.coli (патент РФ №2222596).
В настоящее время нет сообщений о влиянии мутантного белка YddG в соответствии с настоящим изобретением на продукцию ароматических L-аминокислот, в частности L-фенилаланина и L-триптофана с использованием бактерии, принадлежащей к роду Escherichia.
Раскрытие сущности изобретения
Целью настоящего изобретения является предоставление штамма-продуцента L-фенилаланина с повышенной продуктивностью и способа получения L-фенилаланина с использованием указанного штамма. Также целью настоящего изобретения является предоставление штамма-продуцента L-триптофана с повышенной продуктивностью и способа получения L-триптофана с использованием указанного штамма.
Цели настоящего изобретения были достигнуты благодаря обнаружению того факта, что мутантный ген yddG, который не вовлечен в биосинтетический путь целевой L-аминокислоты, сообщает микроорганизму повышенную продуктивность ароматических L-аминокислот, когда аминокислотный(е) остаток(ки) в положении(ях) 26 или 24 и 26 в аминокислотной последовательности аллеля гена yddG дикого типа заменен(ы) на другой(ие) L-аминокислотный(е) остаток(ки).
Целью настоящего изобретения является предоставление мутантного бактериального белка-транспортера аминокислоты, как определено в пунктах (А) или (В):
A) белка, включающего аминокислотную последовательность, представленную в SEQ ID NO:2; и
B) варианта белка, включающего аминокислотную последовательность, представленную в SEQ ID NO:2, но содержащую замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и имеющего активность бактериального белка-транспортера аминокислоты в соответствии с последовательностью, представленной в SEQ ID NO:2, отличающегося тем, что белок содержит замену, выбранную из группы, состоящей из:
(C) замены остатка L-изолейцина в положении №24 на остаток L-фенилаланина, или остаток L-триптофана, или остаток L-тирозина, или остаток L-метионина, или остаток L-лейцина;
(D) замены остатка глицина в положении №26 на остаток L-фенилаланина, или остаток L-изолейцина, или остаток L-лейцина, или остаток L-метионина, или остаток L-валина, или остаток L-триптофана, или остаток L-тирозина; или
(Е) их сочетание,
при этом положения указанных замен соответствуют положениям аминокислотных остатков в последовательности, представленной в SEQ ID NO:2.
Следующей целью настоящего изобретения является предоставление описанного выше мутантного белка, отличающегося тем, что остаток L-изолейцина в положении №24 замещен на остаток L-фенилаланина.
Следующей целью настоящего изобретения является предоставление описанного выше мутантного белка, отличающегося тем, что остаток глицина в положении №26 замещен на остаток L-фенилаланина.
Следующей целью настоящего изобретения является предоставление описанного выше мутантного белка, отличающегося тем, что остаток глицина в положении №26 замещен на остаток L-изолейцина.
Следующей целью настоящего изобретения является предоставление описанного выше мутантного белка, отличающегося тем, что мутантный белок происходит из бактерии, принадлежащей к роду Escherichia.
Следующей целью настоящего изобретения является предоставление описанного выше мутантного белка, отличающегося тем, что мутантный белок происходит из бактерии, принадлежащей к виду Escherichia coli.
Целью настоящего изобретения является предоставление ДНК, кодирующей описанный выше мутантный бактериальный белок-транспортер аминокислоты.
Целью настоящего изобретения является предоставление бактерии, продуцирующей ароматическую L-аминокислоту, принадлежащей к роду Escherichia и содержащей описанную выше ДНК.
Следующей целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что бактерия принадлежит к виду Escherichia coli.
Следующей целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что ароматическая L-аминокислота выбраны из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Следующей целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что ароматическая L-аминокислота есть L-фенилаланин.
Следующей целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что ароматическая L-аминокислота есть L-триптофан.
Целью настоящего изобретения является предоставление способа получения ароматической L-аминокислоты, включающего:
(F) выращивание описанной выше бактерии в питательной среде;
(G) накопление ароматической L-аминокислоты в питательной среде; и
(Н) выделение ароматической L-аминокислоты из культуральной жидкости.
Следующей целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Следующей целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что ароматическая L-аминокислота есть L-фенилаланин.
Следующей целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что ароматическая L-аминокислота есть L-триптофан.
Целью настоящего изобретения является предоставление способа получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина, включающего:
(I) выращивание описанной выше бактерии в питательной среде для получения и накопления L-фенилаланина в среде, при этом указанная бактерия способна продуцировать L-фенилаланин;
(J) этерификацию L-фенилаланина для образования низшего алкильного эфира L-фенилаланина; и
(K) получение низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из L-аспарагиновой кислоты или ее производного и низшего алкильного эфира L-фенилаланина.
Следующей целью настоящего изобретения является предоставление описанного выше способа, дополнительно включающего:
(L) конденсацию низшего алкильного эфира L-фенилаланина с производным L-аспарагиновой кислоты, при этом указанное производное есть ангидрид N-ацил-L-аспарагиновой кислоты;
(М) выделение низшего алкильного эфира N-(N-ацил-α-L-аспартил)-L-фенилаланина из реакционной смеси; и
(N) снятие защиты с низшего алкильного эфира N-(N-ацил-α-L-аспартил)-L-фенилаланина для получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина.
Целью настоящего изобретения является предоставление способа получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина, включающего:
(О) выращивание описанной выше бактерии в питательной среде для получения и накопления L-фенилаланина в среде, при этом указанная бактерия способна продуцировать L-фенилаланин;
(Р) этерификацию L-аспарагиновой кислоты для получения низшего α,β-диэфира L-аспарагиновой кислоты; и
(Q) получение низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из низшего алкильного α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина.
В настоящем изобретении аминокислота имеет L-конфигурацию, если не указано иное.
Настоящее изобретение более подробно будет описано ниже со ссылкой на последующие, но не ограничивающие настоящее изобретение Примеры.
Фигуры и их краткое описание.
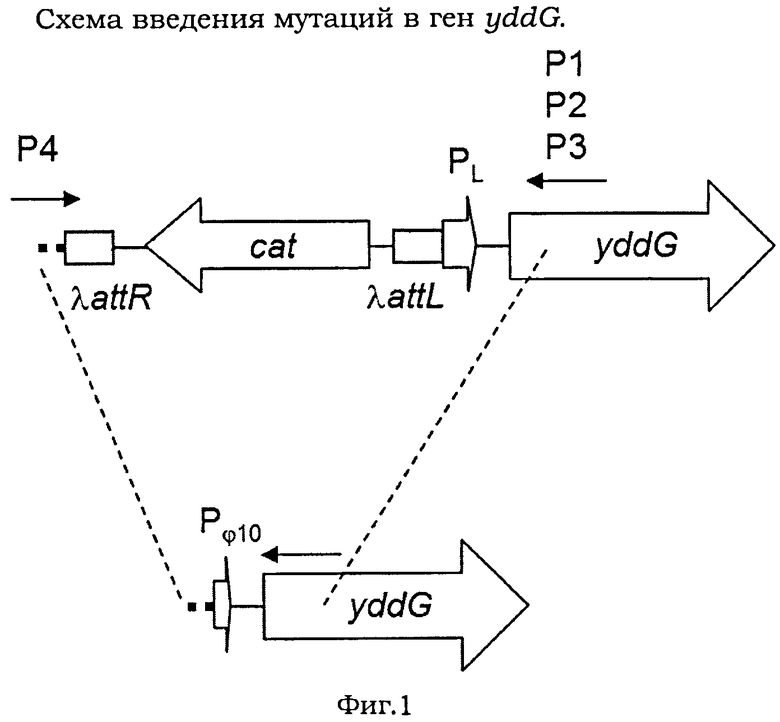
На Фигуре 1 изображена схема введения мутации в ген yddG.
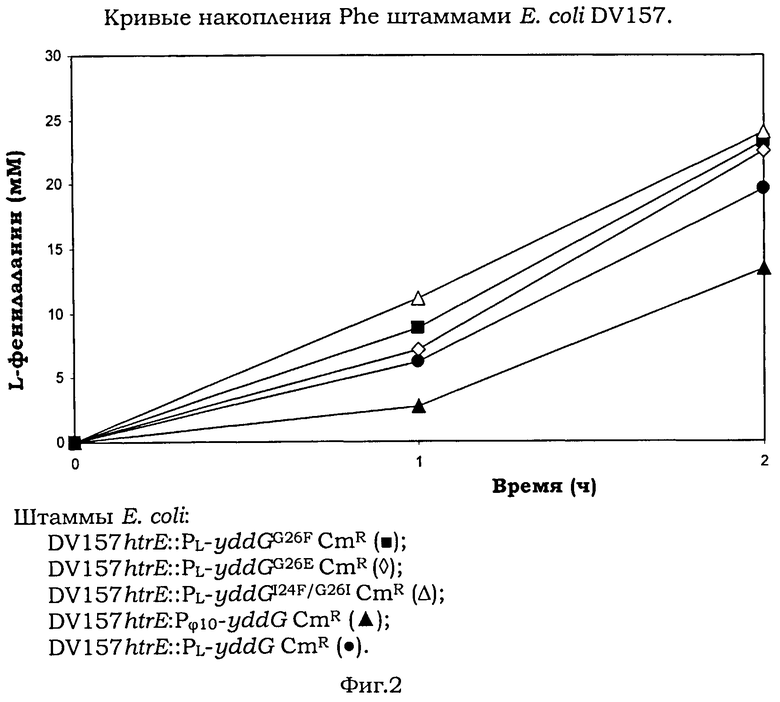
На Фигуре 2 изображены кривые накопления Phe штаммами Е.coli DV157.
Наилучший способ осуществления изобретения
1. Бактерия
Бактерия, используемая в настоящем изобретении, есть бактерия-продуцент L-аминокислоты, принадлежащая к роду Escherichia, отличающаяся тем, что продукция L-аминокислоты указанной бактерией повышена за счет модификации в бактерии аминокислотной последовательности трансмембранного белка согласно настоящему изобретению.
Используемый в настоящем изобретении термин «бактерия-продуцент L-аминокислоты» может означать бактерию, в том числе и более предпочтительно подвижную бактерию, которая способна продуцировать и вызывать накопление L-аминокислоты в культуральной жидкости, когда указанная бактерия выращивается (культивируется) в питательной среде. Бактерия может обладать способностью к продукции аминокислоты изначально в соответствии со своими природными характеристиками или может быть модифицирована таким образом, чтобы она получила способность продуцировать аминокислоту.
Термин "ароматическая L-аминокислота" включает L-фенилаланин, L-тирозин и L-триптофан.
Термин "продуцировать и накапливать L-аминокислоту в среде" может означать способность продуцировать, экскретировать или секретировать и вызывать накопление L-аминокислоты в среде.
Предпочтительным примером бактерии согласно изобретению является бактерия, продуцирующая L-фенилаланин и принадлежащая к роду Escherichia, которая имеет трансмембранный белок согласно настоящему изобретению, аминокислотная последовательность которого модифицирована. Другим предпочтительным примером бактерии согласно изобретению является бактерия, продуцирующая L-триптофан и принадлежащая к роду Escherichia, которая имеет трансмембранный белок согласно настоящему изобретению, аминокислотная последовательность которого модифицирована. Также бактерией согласно изобретению может быть бактерия, продуцирующая L-тирозин и принадлежащая к роду Escherichia, которая имеет трансмембранный белок согласно настоящему изобретению, аминокислотная последовательность которого модифицирована.
Бактерия согласно изобретению может содержать ДНК, имеющую ген yddG, модифицированный с целью введения мутации в положение №24, или 26, или оба положения, соответствующее(ие) положению(ям) аминокислотного(ых) остатка(ов) в аминокислотной последовательности белка YddG дикого типа, кодируемого немодифицированным геном yddG, представленным в SEQ ID NO:1. Примером замены аминокислотного остатка в положении №24, которым является остаток L-изолейцина, как следует из аминокислотной последовательности белка YddG дикого типа, представленной в SEQ ID NO:2, может быть замена на остаток L-триптофана, или остаток L-тирозина, или остаток L-метионина, или остаток L-лейцина. Примером замены аминокислотного остатка в положении №26, которым является отстаток глицина, как следует из аминокислотной последовательности белка YddG дикого типа, представленной в SEQ ID NO:2, может быть замена на остаток L-фенилаланина, или остаток L-изолейцина, или остаток L-лейцина, остаток L-метионина, или остаток L-валина, или остаток L-триптофана, или остаток L-тирозина. Примеры замен в положении №24, или 26, или в обоих положениях в аминокислотной последовательности, кодируемой геном yddG дикого типа, не ограничиваются указанными выше заменами. Это возможно ввиду того, что аминокислоты, как протеиногенные, так и непротеиногенные или другие, могут быть разделены на группы в соответствии со свойствами их боковых групп при С2-атоме, такими как пространственная геометрия, полярность (заряд), гидрофобность или гидрофильность и так далее при условии, что, в целом, третичная структура аминокислотной последовательности или белка не затронута.
Бактерия согласно изобретению может иметь мутантный ген yddG, введенный в хромосому бактерии вместо аллеля гена yddG дикого типа или в дополнение к аллелю гена yddG дикого типа. В том случае когда два или более мутантных гена yddG содержатся в бактерии согласно изобретению, эти гены могут находиться вместе на одной плазмиде или отдельно на разных плазмидах. Также допустимо, чтобы один ген находился на хромосоме, а другой ген был расположен на плазмиде. Мутантный ген yddG может быть введен в бактерию таким способом, что уровень экспрессии мутантного гена yddG повышен по сравнению с немодифицированной бактерией.
Усиление экспрессии гена yddG может быть достигнуто путем подстановки фрагмента ДНК, содержащего мутантный ген yddG, под контроль сильного промотора. Примерами сильных промоторов являются lac промотор, trp промотор, trc промотор, tac промотор, PR или PL промоторы фага λ. В данном изобретении могут быть использованы сильные промоторы, обеспечивающие высокий уровень экспрессии гена в бактерии, принадлежащей к семейству Enterobacteriaceae. С другой стороны, действие промотора может быть усилено, например, введением мутации в промоторный участок гена или генов оперона на бактериальной хромосоме, что приводит к увеличению уровня транскрипции гена, находящегося под промотором. Кроме того, известно, что замена нескольких нуклеотидов в области между сайтом связывания рибосомы (ribosome binding site, RBS) и стартовым кодоном, особенно в последовательности непосредственно перед стартовым кодоном, существенно влияет на трансляционную способность мРНК. Например, обнаружен 20-кратный разброс в уровне экспрессии гена в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold L. et al., Annu. Rev. Microbiol., 1981, 35:365-403; Hui A. et al., EMBO J., 1984, 3:623-629). Использование сильного промотора можно сочетать с увеличением числа копий гена.
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в данной области. Эти методы описаны, например, в Sambrook J., Fritsch E.F. and Maniatis Т., "Molecular Cloning: A Laboratory Manual", 2nd ed., Cold Spring Harbor Laboratory Press (1989). Методы молекулярного клонирования и экспрессии гетерологичных генов описаны в Bernard R. Glick, Jack J. Pasternak and Cheryl L. Patten, "Molecular Biotechnology: principles and applications of recombinant DNA", 4th ed., Washington, D.C: ASM Press (2009); Evans Jr., T.C. and Xu M.-Q., "Heterologous gene expression in E.coli", 1st ed., Humana Press (2011). Методы экспрессии рекомбинантных генов описаны в Balbas P. and Lorence A., "Recombinant gene expression: reviews and protocols", 2nd ed., Totowa N.J., Humana Press (2004).
Уровень экспрессии гена можно определить измерением количества транскрибируемой геном мРНК, используя для этого различные известные методы, включая Нозерн-блоттинг, количественную ОТ-ПЦР (RT-PCR) и т.п. Количество белков, кодируемых геном, можно определить известными методами, включая электрофорез в СДС-ПААГ (SDS-PAGE) с последующим иммуноблоттингом (Вестерн-блоттинг) (Western blotting analysis) и подобными.
ДНК, используемая для модификации описанной в изобретении бактерии, может кодировать трансмембранный белок, обладающий активностью экскретировать L-аминокислоту. Более точно, указанная ДНК представлена мутантным геном yddG, являющимся вариантом нуклеотидной последовательности аллеля гена yddG дикого типа, представленного в SEQ ID NO:1.
Ген yddG дикого типа кодирует экспортер ароматических аминокислот YddG (KEGG, Kyoto Encyclopedia of Genes and Genomes, Киотская энциклопедия генов и геномов, № b1473). Ген yddG (инвентарный номер в GenBank NC_000913.2; нуклеотиды, комплементарные нуклеотидам в положении с 1544312 по 1545193; Gene ID: 945942) расположен между геном fdnG на противоположной цепи и геном yddL на той же цепи хромосомы E.coli K-12. Нуклеотидная последовательность гена yddG и аминокислотная последовательность белка YddG, кодируемого геном yddG, представлены в SEQ ID NO:1 и SEQ ID NO:2 соответственно.
Ввиду того что могут быть некоторые различия в последовательностях ДНК между родами и штаммами бактерий рода Escherichia, модифицируемый ген yddG не ограничивается нуклеотидной последовательностью гена, приведенного в SEQ ID NO:1, и может включать гены, гомологичные нуклеотидной последовательности SEQ ID NO:1, кодирующие варианты дикого или мутантного белка YddG. Замена аминокислотного остатка/ов в положении №24, или 26, или в обоих положениях в белке YddG дикого типа, представленного в SEQ ID NO:2, может быть достигнуто путем модификации нуклеотидной последовательности гена yddG дикого типа, представленного в SEQ ID NO:1, или варианта этой нуклеотидной последовательности.
Ввиду того, что авторы настоящего изобретения методом компьютерного анализа (in silico} предположили, что белок YddG дикого типа может содержать сайт расщепления между G26 и V27 согласно SEQ ID NO:2 для получения предполагаемого сигнального пептида, очевидно, что другая(ие) замена(ы), которая(ые) отлична(ы) от тех, что находятся в позиции 124 и G26 согласно SEQ ID NO:2, может(гут) быть использована(ы) в соответствии с настоящим изобретением. Такая(ие) замена(ы) может(гут) быть локализована(ы) в пределах 3-х аминокислотных остатков или в пределах 5-и аминокислотных остатков в направлении С- и/или N-конца белка YddG относительно предполагаемого сайта расщепления, как приведено выше.
Термин "вариант белка" может означать белок, содержащий одно или несколько изменений в последовательности при сравнении с последовательностью дикого типа, представленной в SEQ ID NO:2, или его мутантной последовательностью, соответствующей мутантному белку YddG согласно изобретению, которыми могут быть замены, делеции (удаления), вставки и/или добавления аминокислотных остатков при условии, что активность такого белка такая же, как и у белка YddG дикого и мутантного типа. Количество изменений в варианте белка зависит от вида или положения аминокислотного остатка в третичной структуре белка. Оно может быть, не ограничиваясь этим, от 1 до 30, предпочтительно от 1 до 15 и еще более предпочтительно от 1 до 5 в SEQ ID NO:2 или его мутантной последовательности, соответствующей мутантному белку YddG, как здесь описано. В варианте белка подобные изменения могут быть в областях, не критичных для функции белка, так как некоторые аминокислоты в большей степени гомологичны другим, и поэтому третичная структура или активность белка не затрагивается ввиду этих изменений. Следовательно, варианты белков, кодируемые геном yddG, могут иметь идентичность не менее чем 98% или не менее чем 99% по отношению к полной аминокислотной последовательности, представленной в SEQ ID NO:2, при условии что функциональность белков YddG дикого и мутантного типа сохранена.
Замены, делеции, вставки и/или добавления одного или нескольких аминокислотных остатков могут быть консервативными мутациями при условии, что активность и/или функция варианта белка сохранена или подобна активности белка YddG дикого или мутантного типа. Примером консервативной мутации может быть консервативная замена. Консервативными заменами могут быть взаимные замены между Phe, Trp и Tyr, если сайт замещения содержит ароматическую аминокислоту; между Leu, Ile и Val, если сайт замещения содержит гидрофобную аминокислоту; между Gln и Asn, если сайт замещения содержит полярную аминокислоту; между Lys, Arg и His, если сайт замещения содержит основную аминокислоту; между Asp и Glu, если сайт замещения содержит кислую аминокислоту; между Ser и Thr, если сайт замещения содержит аминокислоту, имеющую гидроксильную группу. Специфическими примерами консервативных замен могут быть замены Ser или Thr на Ala; замена Gln, His или Lys на Arg; замена Glu, Gln, Lys, His или Asp на Asn; замена Asn, Glu или Gln на Asp; замена Ser или Ala на Cys; замена Asn, Glu, Lys, His, Asp или Arg на Gln; замена Gly, Asn, Gln, Lys или Asp на Glu; замена Pro на Gly; замена Asn, Lys, Gln, Arg или Tyr на His; замена Leu, Met, Val или Phe на Ile; замена Ile, Met, Val или Phe на Leu; замена Asn, Glu, Gln, His или Arg на Lys; замена Ile, Leu, Val или Phe на Met; замена Trp, Tyr, Met, Ile или Leu на Phe; замена Thr или Ala на Ser; замена Ser или Ala на Thr; замена Phe или Tyr на Trp; замена His, Phe или Trp на Tyr; и замена Met, Ile или Leu на Val.
Гомология или идентичность между белками или ДНК может быть определена с использованием нескольких известных подходов, например компьютерных алгоритмов BLAST и FASTA и метода ClustalW. Алгоритм BLAST (Basic Local Alignment Search Tool, www.ncbi.nlm.nih.gov/BLAST/), позволяющий проводить иерархический поиск, заложен в программах blastp, blastn, blastx, megablast, tblastn и tblastx; эти программы присваивают уровень значимости найденным объектам, используя статистические методы, описанные в Samuel K. and Altschul S.F. ("Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes" Proc. Natl. Acad. Sci. USA, 1990, 87:2264-2268; "Applications and statistics for multiple high-scoring segments in molecular sequences". Proc. Natl. Acad. Sci. USA, 1993, 90:5873-5877). Алгоритм BLAST вычисляет три параметра: число аминокислотных остатков, идентичность и сходство. Поисковый алгоритм FASTA описан в Pearson W.R. ("Rapid and sensitive sequence comparison with FASTP and FASTA", Methods Enzymol., 1990, 183:63-98). Метод ClustalW описан в Thompson J.D. et al. ("CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice". Nucleic Acids Res., 1994, 22:4673-4680).
Кроме того, гены yddG дикого и мутантного типа могут быть вариантами нуклеотидных последовательностей. Термин «вариант нуклеотидной последовательности» может означать нуклеотидную последовательность, кодирующую «вариант белка». Термин «вариант нуклеотидной последовательности» также может означать нуклеотидную последовательность, гибридизующуюся в жестких условиях с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности, приведенной в Перечне последовательностей под номером SEQ ID NO:1, или ее мутантной последовательностью, кодирующей мутантный белок YddG как описано выше, или с зондом, который может быть синтезирован на основе указанных нуклеотидных последовательностей, при условии что он кодирует функциональный белок YddG дикого или мутантного типа до инактивации. Под «жесткими условиями» понимаются такие условия, в которых образуется специфический гибрид, например гибрид, имеющий гомологию не менее чем 97%, или не менее чем 98%, или не менее чем 99%, и в которых не образуется неспецифический гибрид, например гибрид, имеющий гомологию меньшую, чем указанно выше. Практическим примером жестких условий является однократная отмывка, или двух- или трехкратная, при концентрации солей 1×SSC, 0.1% SDS или 0.1×SSC, 0.1% SDS, при 60°С или 65°С. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для положительно заряженной нейлоновой мембраны Hybond™-N+ (GE Healthcare) при жестких условиях составляет 15 минут. Промывка может быть произведена двух- или трехкратно. В качестве зонда может быть использована часть последовательности, комплементарная последовательности, приведенной в SEQ ID NO:1. Подобный зонд может быть получен с помощью ПЦР с использованием олигонуклеотидных праймеров, приготовленных на основе последовательности, приведенной в SEQ ID NO:1, и фрагмента ДНК, содержащего нуклеотидную последовательность в качестве матрицы. Рекомендуемая длина зонда может быть подобрана в зависимости от условий гибридизации и составляет >50 п.н., обычно около 100-1000 п.н. Например, при использовании в качестве зонда фрагмента ДНК длиной около 300 п.н. условия отмывки могут быть следующие: 2×SSC, 0.1% SDS при 50°С, или при 60°С, или при 65°С.
Ген, кодирующий белок YddG из бактерии рода Escherichia, известен (см. описание выше), поэтому этот ген и гены, кодирующие варианты указанного белка, могут быть получены с использованием метода ПЦР (полимеразная цепная реакция; White T.J. et al., Trends Genet, 1989, 5:185-189) с использованием праймеров, синтезированных на основе нуклеотидной последовательности гена yddG. Ген, кодирующий белок YddG и варианты указанного белка из других микроорганизмов, могут быть получены аналогичным способом.
Бактерия-продуцент ароматической L-аминокислоты
В качестве бактерии согласно изобретению, модифицированной для продукции мутантного белка YddG, может быть использована бактерия, способная продуцировать ароматическую L-аминокислоту.
L-триптофан, L-фенилаланин и L-тирозин являются ароматическими аминокислотами и имеют общий путь биосинтеза. Примеры генов, кодирующих ферменты биосинтеза этих ароматических аминокислот, включают гены, кодирующие дезоксиарабиногептулозонатфосфатсинтазу (aroG), 3-дегидрохиннатсинтазу (aroB), шикиматдегидрогеназу, шикиматкиназу (aroL), 5-енолпирувилшикимат-3-фосфатсинтазу (aroA) и хоризматсинтазу (aroC) (Европейская патентная заявка ЕР763127). Следовательно, способность к продукции ароматических аминокислот может быть усилена увеличением числа копий генов, кодирующих эти ферменты, на плазмиде или хромосоме. Известно, что эти гены могут контролироваться регулятором транскрипции (tyrR), поэтому активность ферментов биосинтеза ароматических аминокислот также может быть увеличена путем делеции гена tyrR (Европейская патентная заявка ЕР763127).
Для увеличения продукции ароматических аминокислот в бактерии может быть ослаблен биосинтез других аминокислот, иных чем целевая ароматическая аминокислота. Например, когда целевая аминокислота-L-триптофан, может быть ослаблен биосинтез L-фенилаланина и/или L-тирозина (патент США №4371614). Кроме того, 3-дезокси-D-арабиногептулозонат-7-фосфатсинтетаза (DS), кодируемая генами aroF или aroG, подвержена ингибированию по типу обратной связи ароматическими аминокислотами. Следовательно, бактерия может быть модифицирована таким образом, чтобы содержать мутантную DS, не подверженную ингибированию по типу обратной связи. Такая мутантная DS может быть получена, например, заменой в aroF L-аспарагиновой кислоты в положении 147 или L-серина в положении 181 на другую аминокислоту. В случае aroG мутантная DS может быть получена, например, заменой L-аспарагиновой кислоты в положении 146, L-метионина в положении 147, L-пролина в положении 150 или L-аланина в положении 202 на другую аминокислоту или заменой L-метионина в положении 157 и L-аланина в положении 219 на другую аминокислоту. Бактерия-продуцент ароматической L-аминокислоты может быть получена введением в бактерию мутантного гена, кодирующего такую мутантную DS (Европейская патентная заявка ЕР0488424).
Бактерия-продуцент L-фенилаланина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-фенилаланина, включают, но не ограничиваются ими, штаммы, принадлежащие к роду Escherichia, такие как штамм AJ12739 (tyrA::Tn10, tyrR) (ВКМП В-8197), штамм HW1089 (АТСС-55371), содержащий ген pheA34 (патент США № 5354672), мутантный штамм MWEC101-b (KR8903681), штаммы NRRL B-12141, NRRL В-12145, NRRL В-12146 и NRRL B-12147 (патент США №4407952) и подобные им. В качестве родительских штаммов также могут быть использованы бактерии, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм Е.coli K-12[W3110(tyrA)/pPHAB] (PERM ВР-3566), штамм Е.coli K-12[W3110(tyrA)/pPHAD] (FERM ВР-12659), штамм Е.coli K-12[W3110(tyrA)/pPHATerm] (PERM ВР-12662) и штамм Е.coli K-12[W3110(tyrA)/pBR-aroG4, рАСМАВ], названный какАJ12604 (PERM ВР-3579) (Европейский патент ЕР488424 В1). Кроме того, также могут быть использованы бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia, с повышенной активностью белков, кодируемых геном yedA или геном yddG (патентные заявки РФ №2001131570 и 2001131571). Также могут быть использованы бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia, который устойчивы к L-фенилаланину и/или аналогу аминокислоты, такому как р-фторфенилаланин, 5-фтор-DL-триптофан, или подобные им (патент США №7,666,655).
Бактерия-продуцент L-триптофана
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-триптофана, включают, но не ограничиваются ими, штаммы, принадлежащие к роду Escherichia, такие как штаммы Е.coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), лишенные активности триптофанил-тРНК-синтетазы, кодируемой мутантным геном trpS (патент США №5756345); штамм Е.coli SV164 (pGH5), содержащий аллель serA, кодирующий фосфоглицератдегидрогеназу, не ингибируемую серином по типу обратной связи, и аллель trpE, кодирующий антранилатсинтазу, не ингибируемую триптофаном по типу обратной связи (патент США №6180373); штаммы Е.coli AGX17 (pGX44) (NRRL В-12263) и AGX6(pGX50)aroP (NRRL B-12264), в которых отсутствует активность триптофаназы (патент США №4371614); штамм Е.coli AGX17/pGX50,pACKG4-pps, в котором усилена способность к синтезу фосфоенолпирувата (заявка РСТ WO9708333, патент США №6319696); и подобные им. Также могут быть использованы бактерии-продуценты L-триптофана, принадлежащие к роду Escherichia, в которых увеличена активность белка, кодируемого геном yedA или геном yddG (заявки на патент США №2003/0148473 А1 и 2003/0157667 А1).
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-триптофана согласно настоящему изобретению, также включают в себя штаммы, в которых увеличена активность одного или нескольких ферментов, выбранных из группы, состоящей из антранилатсинтазы, фосфоглицератдегидрогеназы и триптофансинтазы. И антранилатсинтаза, и фосфоглицератдегидрогеназа подвержены ингибированию L-триптофаном и L-серином по типу обратной связи, так что в эти ферменты могут быть введены мутации, снижающие чувствительность к ингибированию по типу обратной связи. Специфические примеры штаммов с такой мутацией включают Е.coli SV164, антранилатсинтаза которой не чувствительна к ингибированию по типу обратной связи, и штамм-трансформант, полученный введением в Е.coli SV164 плазмиды pGH5 (заявка РСТ WO94/08031), которая содержит мутантный ген serA, кодирующий фосфоглицератдегидрогеназу, не чувствительную к ингибированию по типу обратной связи.
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-триптофана, также включают в себя штаммы, в которые введен триптофановый оперон, содержащий ген, кодирующий антранилатсинтазу, не чувствительную к ингибированию по типу обратной связи (патентные заявки Японии, публикации №57-71397 (1982) и 62-244382 (1987), патент США №4371614). Кроме того, способность к продукции L-триптофана может быть придана путем усиления экспрессии гена из триптофанового оперона, кодирующего триптофансинтазу (trpBA). Триптофансинтаза состоит из двух субъединиц α и β, которые кодируются генами trpA и trpB соответственно. Кроме того, способность к продукции L-триптофана может быть увеличена усилением экспрессии оперона изоцитратлиазы-малатсинтазы (заявка РСТ WO2005/103275).
Бактерия согласно изобретению может быть получена введением вышеупомянутой ДНК в бактерию, изначально имеющую способность продуцировать ароматическую L-аминокислоту. С другой стороны, бактерия согласно изобретению может быть получена сообщением способности продуцировать ароматическую L-аминокислоту бактерии, уже имеющей упомянутую ДНК.
2. Способ получения L-аминокислот
Способ получения L-аминокислот, конкретно ароматических L-аминокислот, таких как L-фенилаланин и L-триптофан, включает стадии выращивания описанной выше бактерии в питательной среде для продукции и накопления L-аминокислоты в питательной среде и выделения L-аминокислоты из культуральной жидкости.
В настоящем изобретении выращивание, выделение и очистка L-аминокислоты из культуральной или подобной ей жидкости могут быть осуществлены способами, подобными традиционным способам ферментации, в которых аминокислота продуцируется с использованием микроорганизма. Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизма. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза, лактоза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза, рибоза и гидролизаты крахмала; спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая кислота, фумаровая кислота, лимонная кислота, яблочная кислота и янтарная кислота; и подобные им. В зависимости от типа ассимиляции используемого микроорганизма может также использоваться спирт, включая этанол и глицерин. В качестве источника азота могут использоваться различные неорганические аммонийные соли, такие как сульфат аммония, хлорид аммония и фосфат аммония; органический азот, такой амины, пептон, разрушенные микроорганизмы и гидролизаты соевых бобов; аммиачный газ; водный раствор аммиака; и подобные им. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. При необходимости витамины, такой как витамин В1, и некоторые другие питательные вещества могут быть добавлены в среду.
Выращивание осуществляют предпочтительно в аэробных условиях, таких как встряхивание или перемешивание питательной среды с аэрацией, при температуре в интервале 20-42°С, предпочтительно 35-40°С. Кислотность (рН) питательной среды поддерживают в интервале 5-9, предпочтительно 6,8-7,0. рН можно регулировать с использованием аммиака, карбоната кальция, различных кислот и оснований, различных солей и буферов. Обычно выращивание в течение от 1 до 3 дней приводит к накоплению целевой аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки и клеточный дебрис, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевая L-аминокислота может быть выделена и очищена с использованием любой комбинации традиционных методов, таких как ионообменная хроматография, концентрация и кристаллизация.
Фенилаланин, полученный способом, описанным в настоящем изобретении, может быть использован, например, для получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина (также называемого "аспартам"). То есть способ настоящего изобретения включает способ получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина с использованием L-фенилаланина в качестве исходного материала. Способ включает этерификацию L-фенилаланина, полученного представленным выше способом согласно изобретению, для получения низшего алкильного эфира L-фенилаланина и синтез низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из полученного низшего алкильного эфира L-фенилаланина и L-аспарагиновой кислоты. Если для взаимодействия с низшим алкильным эфиром L-фенилаланина используется производное L-аспарагиновой кислоты, то может потребоваться стадия снятия защиты для получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина. В качестве низшего алкильного эфира можно привести метиловый эфир, этиловый эфир и пропиловый эфир или подобные им.
В способе согласно изобретению процесс синтеза низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из низшего алкильного эфира L-фенилаланина и L-аспарагиновой кислоты или ее производного не ограничивается приведенным частным случаем, и любой обычный способ может быть использован при условии, что L-фенилаланин или его производное может быть использовано для синтеза низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина. Более конкретно, например, низший алкильный эфир N-(α-L-аспартил)-L-фенилаланина может быть получен способом, описанным в патенте США №3786039. L-Фенилаланин этерифицируют для получения низшего алкильного эфира L-фенилаланина. Алкиловый эфир L-фенилаланина взаимодействует с производным L-аспарагиновой кислоты, аминогруппа и β-карбоксильная группа которой защищены, а α-карбоксильная группа этерифицирована или трансформирована в галогенкарбонильную группу или подобную ей с целью активации. Примеры производного включают N-ацил-L-аспарагиновый ангидрид, такой как N-формил-, N-карбобензокси- или N-n-метоксикарбобензокси-L-аспарагиновый ангидрид. В результате реакции конденсации образуется смесь N-(N-ацил-α-L-аспартил)-L-фенилаланина и N-(N-ацил-β-L-аспартил)-L-фенилаланина. Если реакция конденсации проходит в присутствии органической кислоты, у которой константа диссоциации при 37°С составляет 10-4 или меньше, отношение α-формы к β-форме в смеси увеличивается (патентная заявка Японии, публикация №51-113841 (1976)). Затем N-(N-ацил-α-L-аспартил)-L-фенилаланин выделяют из смеси с последующим гидрированием с образованием N-(α-L-аспартил)-L-фенилаланина. Гидрирование, или снятие защиты с аминогруппы остатка α-L-аспартила, может осуществить в условиях кислотного гидролиза.
Низший алкильный эфир N-(α-L-аспартил)-L-фенилаланина может быть получен путем выращивания бактерии согласно настоящему изобретению, способной к продукции L-фенилаланина, в питательной среде для получения и накопления в среде L-фенилаланина; этерификации L-аспарагиновой кислоты для получения низшего алкила α,β-диэфира L-аспарагиновой кислоты; и образования низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из низшего алкильного α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина (Европейская патентная заявка ЕР 1587941 А1).
Примеры
Настоящее изобретение более подробно будет описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1.
Конструирование штаммов Е.coli DV157, имеющих мутантный ген yddG
Штаммы Е.coli DV157htrE::Pφ10-yddG и DV157htrE::PL-yddG, продуцирующие L-фенилаланин, были описаны ранее (Doroshenko V.G. et al., FEMS Microbiol. Lett., 2007, 275:312-318). Оба штамма содержат дополнительную копию гена yddG, интегрированного в ген htrE. В штамме Е.coli DV157htrE::Pφ10-yddG копия yddG не активна благодаря промотору Pφ10 фага Т7, расположенному перед геном.
Мутации вводили в хромосому штамма Е.coli DV157htrE::Pφ10-yddG с использованием модифицированного метода λRed-зависимой интеграции (Doroshenko V.G. et al., FEMS Microbiol. Lett, 2007, 275:312-318; Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645). Полимеразную цепную реакцию (ПЦР) проводили с использованием прямого праймера P1 (SEQ ID NO:3) для введения мутации G26F, или праймера Р2 (SEQ ID NO:4) для введения мутации G26E, или праймера Р3 (SEQ ID NO:5) для введения мутации I24F/G26I и обратного праймера Р4 (SEQ ID NO:6), и хромосомной ДНК в качестве матрицы. Температурный профиль был следующим: начальная денатурация ДНК в течение 5 мин при 94°С; затем 30 циклов: денатурация в течение 30 сек при 94°С, отжиг в течение 30 сек при 50°С, элонгация в течение 1 мин при 72°С и финальная элонгация в течение 7 мин при 72°С. Праймеры подбирали таким образом, чтобы при ПЦР неактивный промотор Pφ10 заместить на активный промотор PL, маркированный геном cat, кодирующим маркер устойчивости к хлорамфениколу (CmR). Структура фрагмента htrE::attR-cat-attL-PL-yddG и схема интеграции фрагмента показаны на Фигуре 1. Мутации в штаммах Е.coli DV157htrE::PL-yddGG26FCmR, DV157htrE::PL-yddGG26ECmR и DV157htrE::PL-yddGI24F/G26ICmR подтверждали анализом последовательности методом секвенирования.
Пример 2.
Продукция L-фенилаланина модифицированными штаммами Е.coli DV157
Модифицированные штаммы Е.coli DV157htrE::PL-yddGG26FCmR, DV157htrE::PL-yddGG26ECmR, DV157htrE::PL-yddGI24F/G26ICmR и контрольные штаммы DV157htrE::PL-yddGCmR, DV157htrE::Pφ10-yddGCmR, каждый, выращивали отдельно при 37°С и энергичном качании (250 об/мин) до OD600нм 2 (логарифмическая фаза роста) в среде М9 (Sambrook, J. and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press) с добавлением Tyr (125 mg/L). Клетки отмывали в 0.9% (w/v) NaCl и концентрировали дважды в свежей культуральной среде М9 (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Laboratory Press, Cold Spring Harbor) в общем объеме 10 мл до получения плотности клеток ОD600нм 4 (~2×109 CFU/mL, CPU означает колониеформирующая единица). Клетки выращивали в пробирках 20×200 мм при 37°С в течение 2 ч на роторной качалке при 250 об/мин, как описано в Doroshenko V.G. et al., FEMS Microbiol. Lett., 2007, 275:312-318 (краткосрочное выращивание).
Аликвоты (1.5 мл) ростовой среды отбирали после 0, 1 и 2 часов от начала выращивания, клетки осаждали при 10000 об/мин и супернатант анализировали с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ).
Условия ВЭЖХ:
Колонка: Eclipse XDB-C18, 4.6×75 мм, 3.5 мкм (Agilent Technologies).
Температура: 23°С.
Скорость потока: 1 мл/мин.
Объем инъекции: 10 мкл.
Детекция: УФ 220 нм.
Буферы:
А: 25 мМ NaH2PO4, pH 2.5 (регулируется H3PO4),
В: H2O,
С: 70% водный CH3CN.
Профиль градиента:
От 0 мин до 12 мин: А-100%,
От 12 мин до 32 мин: А-85.7% и С-14.3%,
От 32 мин до 34 мин: А-42.8% и С-57.2%,
От 34 мин 36 мин: С-100%,
От 36 мин до 42 мин: В-100%,
От 42 мин до 44 мин: А-100%,
От 44 мин до 66 мин: А-100%.
Средние результаты трех независимых пробирочных ферментации представлены на Фигуре 2 и в Таблице 1. Как можно видеть из Фигуры 2 и Таблицы 1, модифицированные штаммы Е.coli DV157htrE::PL-yddGG26FCmR, DV157htrE::PL-yddGG26ECmR и DV157htrE::PL-yddGI24F/G26ICmR накапливали большее количество L-фенилаланина при сравнении с контрольными штаммами.
Пример 3.
Конструирование штаммов Е.coli DV1060, имеющих мутантный ген yddG
Модификации PL-yddGG26F, PL-yddGI24F/G26I, PL-yddGG26E и PL-yddG, маркированные CmR-маркером, были, каждая, трансформирована с использованием P1-трансдукции в штамм Е.coli DV1060, продуцирующий L-триптофан (Doroshenko V.G. et al., FEMS Microbiol. Lett., 2007, 275:312-318; Miller J.H. (1972) Experiments in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor). Штаммы, описанные в Примере 1, использовались в качестве штаммов-доноров. Так были получены штаммы Е.coli DV1060htrE::PL-yddGG26FCmR, DV1060htrE::PL-yddGI24F/G26ICmR, DV1060htrE::PL-yddGG26ECmR и DV1060htrE::PL-yddGCmR. Конструирование штамма DV1060htrE::Pφ10-yddG описано в Doroshenko V.G. et al., FEMS Microbiol. Lett., 2007, 275:312-318.
Пример 4.
Продукция L-триптофана модифицированными штаммами Е.coli DV1060
Модифицированные штаммы Е.coli DV1060htrE::PL-yddGG26FCmR, DV1060htrE::PL-yddGI24F/G26ICmR, DV1060htrE::PL-yddGG26ECmR и контрольный штамм DV1060htrE::PL-yddGCmR, DV1060htrE::Pφ10-yddG, каждый, выращивали отдельно при 37°С и энергичном качании (250 об/мин) до OD600нм 2 (логарифмическая фаза роста) в среде М9 (см. Пример 2), содержащей добавки Phe (125 мг/л) и Tyr (125 мг/л). Клетки отмывали в 0.9% (w/v) NaCl и дважды концентрировали в среде М9 (см. Пример 2) в общем объеме 10 мл до получения плотности клеток OD600нм 4 (~2×109 CFU/мл, CFU означает колониеформирующая единица). Клетки выращивали в пробирках 20×200 мм при 37°С в течение 3 ч на роторной качалке при 250 об/мин, как описано в Doroshenko V.G. et al., FEMS Microbiol. Lett, 2007, 275:312-318 (краткосрочное выращивание).
Аликвоты (1.5 мл) ростовой среды отбирали после 0, 1 и 2 часов от начала выращивания, клетки осаждали при 10000 об/мин и супернатант анализировали с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ), как описано в Примере 2.
Средние результаты трех независимых пробирочных ферментации представлены в Таблице 2. Как можно видеть из Таблицы 2, модифицированные штаммы Е.coli DV1060htrE::PL-yddGG26FCmR, DV1060htrE::PL-yddGG26FCmR и DV1060htrE::PL-yddGI24F/G26ICmR накапливали большее количество L-триптофана по сравнению с контрольными штаммами.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ЭКСПРЕССИРУЮЩЕЙ ГЕН ARO1 ИЗ ДРОЖЖЕЙ | 2008 |
|
RU2408723C2 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| ДНК, КОДИРУЮЩАЯ ДИПЕПТИД-СИНТЕЗИРУЮЩИЙ ФЕРМЕНТ (ВАРИАНТЫ), БАКТЕРИЯ РОДА Escherichia И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДОВ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2560980C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПРОДУКЦИИ АРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Methylophilus | 2005 |
|
RU2333950C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНА pckA | 2003 |
|
RU2268300C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОПЕРОН relBE | 2005 |
|
RU2313574C2 |



Изобретение относится к биотехнологии и представляет собой мутантный бактериальный белок-экспортер аминокислоты YddG. Изобретение относится также к бактерии рода Escherichia, продуцирующей ароматическую аминокислоту, при этом бактерия содержит ДНК, которая кодирует мутантный белок-экспортер аминокислоты YddG. Также представляется способ получения ароматических L-аминокислот, таких как L-фенилаланин и L-триптофан, а также способ получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина с использованием указанной бактерии. Изобретение позволяет получать ароматические L-аминокислоты, а также низший алкильный эфир N-(α-L-аспартил)-L-фенилаланина с высокой степенью эффективности. 6 н. и 13 з.п. ф-лы, 2 ил., 2 табл., 4 пр.
1. Мутантный бактериальный белок-экспортер аминокислоты, как определено в пунктах (А) или (В):
A) белок, включающий аминокислотную последовательность, представленную в SEQ ID NO:2; и
B) вариант белка, включающий аминокислотную последовательность, представленную в SEQ ID NO:2, но содержащую замену, делецию, вставку или добавление одного или нескольких аминокислотных остатков, и имеющий активность бактериального белка-транспортера аминокислоты в соответствии с последовательностью, представленной в SEQ ID NO:2,
отличающийся тем, что белок содержит замену, выбранную из группы, состоящей из:
(C) замены остатка L-изолейцина в положении №24 на остаток L-фенилаланина, или остаток L-триптофана, или остаток L-тирозина, или остаток L-метионина, или остаток L-лейцина;
(D) замены остатка глицина в положении №26 на остаток L-фенилаланина, или остаток L-изолейцина, или остаток L-лейцина, или остаток L-метионина, или остаток L-валина, или остаток L-триптофана, или остаток L-тирозина; или
(Е) их сочетание,
при этом положения указанных замен соответствуют положениям аминокислотных остатков в последовательности, представленной в SEQ ID NO:2.
2. Мутантный белок по п.1, отличающийся тем, что указанный остаток L-изолейцина в положении №24 замещен на остаток L-фенилаланина.
3. Мутантный белок по п.1, отличающийся тем, что указанный остаток глицина в положении №26 замещен на остаток L-фенилаланина.
4. Мутантный белок по п.1, отличающийся тем, что указанный остаток глицина в положении №26 замещен на остаток L-изолейцина.
5. Мутантный белок по п.1, отличающийся тем, что указанный мутантный белок происходит из бактерии, принадлежащей к роду Escherichia.
6. Мутантный белок по п.1, отличающийся тем, что указанный мутантный белок происходит из бактерии, принадлежащей к виду Escherichia coli.
7. ДНК, кодирующая мутантный бактериальный белок-экспортер аминокислоты по любому из пп.1-6.
8. Бактерия-продуцент ароматической L-аминокислоты, принадлежащая к роду Escherichia и содержащая ДНК по п.7.
9. Бактерия по п.8, отличающаяся тем, что указанная бактерия принадлежит к виду Escherichia coli.
10. Бактерия по п.8, отличающаяся тем, что указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
11. Бактерия по п.10, отличающаяся тем, что указанная ароматическая L-аминокислота есть L-фенилаланин.
12. Бактерия по п.10, отличающаяся тем, что указанная ароматическая L-аминокислота есть L-триптофан.
13. Способ получения ароматической L-аминокислоты, включающий:
(F) выращивание бактерии по любому из пп.8-12 в питательной среде;
(G) накопление указанной ароматической L-аминокислоты в питательной среде; и
(Н) выделение указанной ароматической L-аминокислоты из культуральной жидкости.
14. Способ по п.13, отличающийся тем, что указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
15. Способ по п.14, отличающийся тем, что указанная ароматическая L-аминокислота есть L-фенилаланин.
16. Способ по п.14, отличающийся тем, что указанная ароматическая L-аминокислота есть L-триптофан.
17. Способ получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина, включающий:
(I) выращивание бактерии по любому из пп.1-11 в питательной среде для получения и накопления L-фенилаланина в среде, при этом указанная бактерия способна продуцировать L-фенилаланин;
(J) этерификацию L-фенилаланина для образования низшего алкильного эфира L-фенилаланина; и
(K) получение низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из L-аспарагиновой кислоты или ее производного и низшего алкильного эфира L-фенилаланина.
18. Способ по п.17, включающий:
(L) конденсацию низшего алкильного эфира L-фенилаланина с производным L-аспарагиновой кислоты, при этом указанное производное есть ангидрид N-ацил-L-аспарагиновой кислоты;
(М) выделение низшего алкильного эфира N-(N-ацил-α-L-аспартил)-L-фенилаланина из реакционной смеси; и
(N) снятие защиты с низшего алкильного эфира N-(N-ацил-α-L-аспартил)-L-фенилаланина для получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина.
19. Способ получения низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина, включающий:
(О) выращивание бактерии по любому из пп.1-11 в питательной среде для получения и накопления L-фенилаланина в среде, при этом указанная бактерия способна продуцировать L-фенилаланин;
(Р) этерификацию L-аспарагиновой кислоты для получения низшего α,β-диэфира L-аспарагиновой кислоты; и
(Q) получение низшего алкильного эфира N-(α-L-аспартил)-L-фенилаланина из низшего алкильного α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина.
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2002 |
|
RU2222596C1 |
| DOROSHENKO V | |||
| et.al., YddG from Escherichia coli promotes export of aromatic amino acids, FEMS Microbiol Lett | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ЭКСПРЕССИРУЮЩЕЙ ГЕН ARO1 ИЗ ДРОЖЖЕЙ | 2008 |
|
RU2408723C2 |
| US 20030157667, 21.08.2003 | |||

