Область применения
Изобретение относится к области биотехнологии и касается термостабильных ферментов с активностью алкогольдегидрогеназы. Предлагается новая алкогольдегидрогеназа из археи Thermococcus sibiricus и способ ее выделения.
Актуальность
Ферменты - биокатализаторы широко используются в различных отраслях промышленности, сельского хозяйства и медицины. Алкогольдегидрогеназы принадлежат к классу оксидоредуктаз, в клетке участвуют в детоксификации и метаболизме этанола и других спиртов, синтезе гормонов, модификации белков и полисахаридов. In vitro алкогольдегидрогеназы катализируют обратимое окисление спиртов или восстановление альдегидов и кетонов.
В последнее время алкогольдегидрогеназы (ЕС 1.1.1.1) привлекли внимание биотехнологов фармацевтической, агрохимической промышленности и производства парфюмерии. Алкогольдегидрогеназы способны превращать прохиральные кетоны в оптически активные вторичные спирты, которые необходимы далее в органическом синтезе лекарств, гормонов и других биологически активных веществ. В отличие от химического синтеза, применение биокатализаторов в стереоселективном синтезе позволяет получить 100%-ный чистый оптический изомер без примесей другого изомера с высоким выходом и из доступных исходных соединений. Примерами могут служить синтез ключевого интермедиата для лекарств против болезни Альцгеймера (S)-2-пентанола из 2-пентанона, синтез предшественника простагландинов (1S,2R-цис-2-карбоксиметил-3-циклопентен-1-ол лактона) (Machielsen et al., 2006; Patel, 2004; Hummel and Kula, 1989).
Однако для использования ферментов в индустриальном масштабе требуется преодолеть ряд трудностей: обратимость реакции, использование дорогостоящего кофактора и устойчивость к действию органических растворителей (это или исходные соединения, или растворители для субстратов) в условиях высокой температуры. Обратимость реакции успешно преодолевается избытком исходных соединений, для регенерации кофактора разработаны два подхода - двойная субстратная система (сбалансированное добавление субстрата обратной реакции) и двойная ферментативная система (введение в системы другого фермента, регенерирующего кофактор). В наибольшей степени нерешенной остается проблема устойчивости фермента к органическим растворителям и высокой температуре. Поэтому актуальной задачей, стоящей перед крупными исследовательскими центрами и ведущими биотехнологическими компаниями, является поиск новых алкогольдегидрогеназ, обладающих высокой термостабильностью и устойчивостью к органическим растворителям в сочетании с высокой удельной активностью и стереоселективностью.
Уровень техники
Абсолютное большинство применяемых в настоящее время в биотехнологической промышленности ферментов выделено из мезофильных организмов, поэтому они нестабильны при высоких температурах, чувствительны к действию органических растворителей и стадии иммобилизации на твердых носителях. Открытие и изучение термофильных бактерий и архей, развитие геномных технологий, определило новое направление поиска ферментов с требуемыми свойствами среди белков этих гиперустойчивых микроорганизмов. При общей повышенной устойчивости к температурам, рН среды, присутствию органических растворителей, ферменты из термо-, алкали- и ацидофильных микроорганизмов, как и их мезофильные аналоги, имеют широкую субстратную специфичность и используют NAD(H) и NADP(H) в качестве кофакторов. Компактность структуры, повышенная жесткость белковой глобулы позволяют проводить иммобилизацию без снижения активности фермента, а температуру реакции повысить до 80-90°С (Machielsen et al., 2006; Tripp et al., 1998).
Алкогольдегидрогеназы катализируют окисление спиртов до соответствующих альдегидов и кетонов, а также обратную реакцию восстановления. В качестве кофактора могут использоваться NAD, NADP, FAD или F420/Zn/PQQ. NADP - зависимые алкогольдегидрогеназы могут быть разделены на четыре семейства: короткоцепочечные (около 250 а.к.), среднецепочечные цинксодержащие (около 350 а.к.), длинноцепочечные железосодержащие (350-550 а.к.) ферменты, а также семейство, родственное альдо-кето редуктазам. В аминокислотных последовательностях ферментов первых трех групп присутствует мотив Россмана (Rossmann and Argos, 1981), обеспечивающий связывание NADP, у ферментов четвертой группы этот мотив отсутствует (Machielsen et al., 2006; Rondeau et al., 1992).
Представители всех четырех групп NADP - зависимых алкогольдегидрогеназ были найдены в геномах архей, но всего несколько ферментов были охаракетризованы биохимически. Большая их часть относится к среднецепочечным цинксодержащим ферментам, примерами могут являться алкогольдегидрогеназы из Sulfolobus solfataricus (Raia et al., 2001; Giordano et al., 2005), Aeropyrum pemix (Hirakawa et al., 2004), Picrophilus torridus (Hess et al., 2008), Sulfolobus tokodaii (Yanai et al., 2009), Thermoplasma acidophilum (Marino-Marmolejo et al., 2009). Также охарактеризовано несколько длинноцепочечных железосодержащих ферментов, в том числе алкгольдегидрогеназы из Thermococcus litoralis (Ma et al., 1994), Thermococcus strain AN1 (Li and Stevenson, 1997), Thermococcus hydrothermalis (Antoine et al., 1999) и одна альдо-кето редуктаза из археи Pyrococcus furiosus (Machielsen et al., 2006).
Короткоцепочечные алкогольдегидрогеназы имеют ряд преимуществ по сравнению с цинк- и железосодержащими ферментами: они не чувствительны к кислороду и присутствие акцепторов металлов не снижает их активности. Кроме того, учитывая зависимость устойчивости белка к органическим растворителям от его размера, наименьший размер белковой глобулы среди NAD(P)-зависимых алкогольдегидрогеназ дает предпочтение короткоцепочечным алкогольдегидрогеназам в средах с повышенным содержанием органических жидкостей. Компактная структура таких ферментов может также обуславливать их более высокую термостабильность, что было подтверждено в настоящем изобретении. Однако, до настоящего времени была охарактеризована только одна короткоцепочечная алкгольдегидрогеназа из термофильной археи, - AdhA из Pyrococcus furiosus (van der Oost et al., 2001).
Настоящее изобретение касается выделения и характеристики новой короткоцепочечной алкгольдегидрогеназы из гипертермофильной археи Thermococcus sibiricus (Miroshnichenko et al., 2001), выделенной из нефтяной скважины в Западной Сибири.
Раскрытие изобретения
Анализ определенной нами полной нуклеотидной последовательности генома археи Т. sibiricus выявил наличие в геноме этого организма как минимум трех генов, кодирующих короткоцепочечные алкгольдегидрогеназы. Один из этих генов, Tsib_0319, имеющий нуклеотидную последовательность SEQ ID NO1, кодирует белок, состоящий из 234 аминокислот, расчетная молекулярная масса которого составляет 26.2 кДа.
Методом культивирования рекомбинантного штамма Е.coli Rosetta-gami (DE3), трансформированного вектором рЕТ-15b, содержащим нуклеотидную последовательность SEQ ID NO 1, была выделена и очищена с помощью металло-хелатной хроматографии и гель фильтрации новая рекомбинантная термостабильная короткоцепочечная алкогольдегидрогеназа TsAdh319, содержащая аминокислотную последовательность SEQ ID NO2. Функциональная характеристика фермента показала, что рекомбинантная алкогольдегидрогеназа TsAdh319 использует NADP в качестве кофактора, обладает широкой субстратной специфичностью: окисляет алифатические, ароматические спирты и полиолы, при этом вторичные спирты окисляются стереоселективно. В обратной реакции TsAdh319 может восстанавливать альдегиды и кетоны. Фермент устойчив к высоким концентрациям солей (до 1М NaCl), не чувствителен к действию EDTA и присутствию ионов металлов. Окислительная активность TsAdh319 возрастает с ростом температуры от 40°С до 100С°, время полуинактивации при 100°С составляет один час. Таким образом, среди известных короткоцепочечных алкогольдегидрогеназ TsAdh319 имеет наиболее высокий температурный оптимум и является наиболее термостабильной.
Краткое описание чертежей.
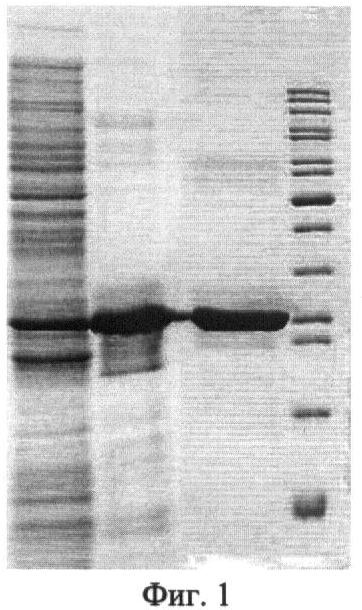
Фиг.1. Экспрессия TsAdh319 в E.coli и очистка белка.
Приведены результаты электрофоретического анализа белковых препаратов:
1 - суммарный белковый препарат, выделенный из клеток штамма Rosetta-gami (DE3), содержащего плазмиду pET_Tsib0319, через 15 часов после индукции синтеза TsAdh319 внесением в среду 1 мМ изопропил-бета-D-тиогалактозида (ИПТГ).
2 - препарат TsAdh319 после стадии очистки на аффинной колонке Hi-Trap chelating HP column.
3 - препарат TsAdh319 после конечной стадии очистки методом гель фильтрации на колонке Superdex 200.
4 - маркер молекулярной массы (PageRuler Unstained Protein Ladder, Fermentas).
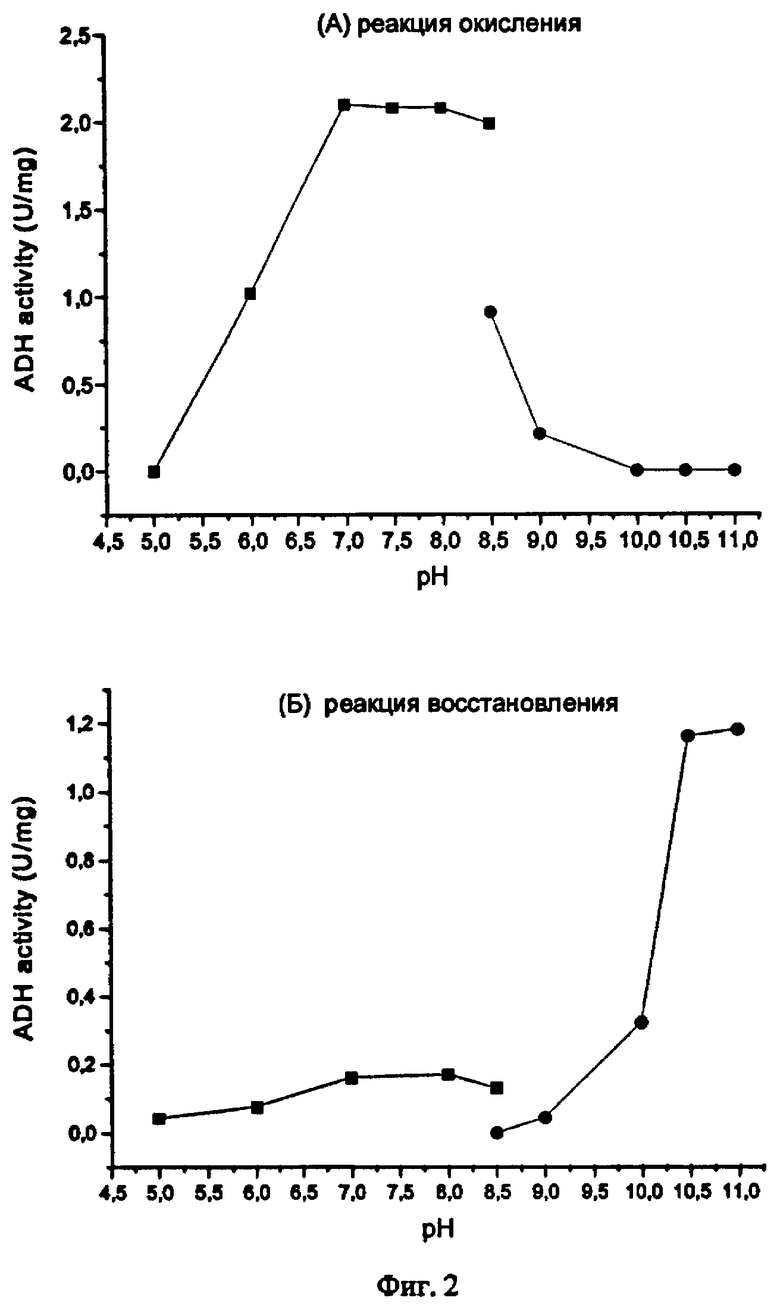
Фиг.2. Влияние рН на активность алкогольдегидрогеназы TsAdh319 в реакциях окисления 2-пропанола (А) и восстановления метилглиоксаля (Б).
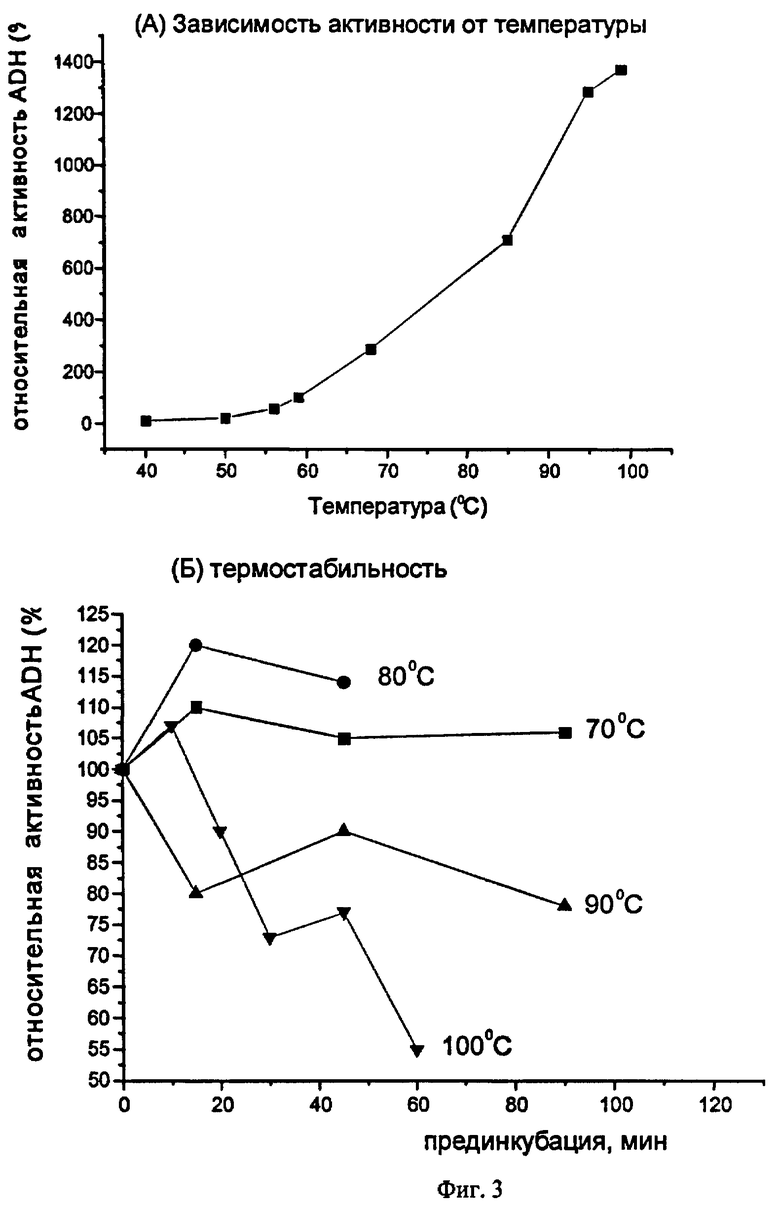
Фиг.3. Влияние температуры на активность алкогольдегидрогеназы TsAdh319 (А) и характеристика ее термостабильности (Б).
Осуществление изобретения
Пример 1. Идентификация гена короткоцепочечной алкогольдегидрогеназы в геноме термофильной археи Т. sibiricus.
Термофильная архея T. sibiricus (Miroshnichenko et al., 2001) была выделена из водного горизонта высокотемпературной нефтяной скважины в Западной Сибири. Нами определена полная нуклеотидная последовательность генома этого микроорганизма, в результате чего были идентифицированы белок-кодирующие гены (Mardanov et al., 2009). Сравнение аминокислотных последовательностей предсказанных белков Т. sibiricus с представленными в GenBank аминокислотными последовательностями выявило наличие в геноме Т. sibiricus как минимум трех генов короткоцепочечных алкогольдегидрогеназ. Аминокислотная последовательность продукта одного из этих генов, Tsib_0319, имеет 85% идентичности с последовательностью алкогольдегидрогеназы AdhA из Р. furiosus (van der Oost et al., 2001).
Как и большинство известных короткоцепочечных алкгогольдегидрогеназ, TSIB_0319 включает два функциональных домена, N-концевой NADP - связывающий домен и С-концевой субстрат-связывающий домен. Как и AdhA из Р. furiosus, N-концевой домен TSIB_0319 содержит консервативный мотив (Gly8-X-X-X-Glyl2-X-Glyl4), характерный для сайтов связывания NAD(P) (Kallberg and Persson, 2006). В С-концевом домене TSIB_0319 и AdhA присутствуют входящие в активный центр короткоцепочечных алкогольдегидрогеназ каталитические остатки Asp59, Tyr150 и Lys154 (нумерация по AdhA).
Пример 2. Получение штамма Е.coli - продуцента алкогольдегидрогеназы, выделение и очистка фермента.
Праймеры TSIB0319-XhoI (GTTCTCGAGATGAAGGTTGCTGTGATAACAGGG) и TSIB0319-BamHI (GCTGGATCCTCAGTATTCTGGTCTCTGGTAGACGG) были использованы для ПЦР-амплификации полноразмерного гена Tsib_0319, в качестве матрицы использовали геномную ДНК Т. sibiricus. Полученный фрагмент обрабатывали рестриктазами XhoI и BamHI и клонировали по сайтам XhoI и BamHI в экспрессионном векторе рЕТ-15b, позволяющем экспрессировать целевой белок, слитый с 6 аминокислотными остатками гистидина на N-конце. В результате был получен экспрессионный вектор pET_Tsib0319, обеспечивающий продукцию рекомбинантаого фермента, TsAdh319, содержащего N-концевой (His)6 - таг, полилинкер и последовательность TSIB_0319.
Для продукции рекомбинантной алкогольдегидрогеназы TsAdh319 экспрессионным вектором pET_Tsib0319 трансформировали штамм Е. coli Rosetta-gami (DE3). Указанный штамм, содержащий pET_Tsib0319, выращивали в среде LB с добавлением ампициллина (100 мг/л) и хлорамфеникола (20 мг/л) на шейкере при 37°С до середины логарифмической фазы роста (OD600 ~0.5), затем индуцировали синтез рекомбинантного белка внося изопропил-β-D-тиогалактопиранозид (ИПТГ) до 1 мМ, и продолжали выращивать культуру в течение 15 ч при 37°С.
Клетки штамма-продуцента собирали центрифугированием, ресуспендировали в буфере, содержащем 50 mM Tris (рН 7.5), 200 mM NaCl, 20 mM имидазола, 10% (v/v) глицерина, 10 mM β-меркаптоэтанол, 0.1% (v/v) Triton X-100, ImM PMSF, и лизировали, обрабатывая ультразвуком. Клеточный экстракт центрифугировали в течение 25 мин при 10000 g, супернатант наносили на аффинную колонку Hi-Trap chelating HP (1ml), уравновешенную буфером, содержащим 50 mM Tris (рН 7.5), 500 mM NaCl, 20 mM имидазола с добавлением 0.1% (v/v) Triton X-100. Рекомбинантный белок TsAdh319, содержащий (His)6-таг, элюировали линейным градиентом (от 20 до 500 mM) имидазола в том же буфере без добавления Triton Х-100. Активную фракцию далее концентрировали в центрифужных концентраторах (Millipore) и дочищали гель фильтрацией на колонке Superdex 200 10/300 GL (Amersham), уравновешенной буфером, содержащим 50 mM Tris (рН 7.5) и 200 mM NaCl. Чистый активный препарат TsAdh319 хранили при 4°С в том же буфере.
Гомогенность препарата TsAdh319 и соответствие молекулярного веса рекомбинантного белка расчетному значению 28.7 кД была подтверждена при помощи SDS/PAGE (фиг.1).
Пример 3. Субстратная специфичность и кинетические характеристики TsAdh319.
Короткоцепочечные алкогольдегидрогеназы относятся к семейству NADP-зависимых оксидоредуктаз, которые в прямой реакции окисляют спирты до альдегидов и кетонов, а в обратной - восстанавливают альдегиды и кетоны до спиртов.
Для характеристики субстратной специфичности фермента TsAdh319 в прямой реакции окисления использовали ряд первичных, вторичных, ароматических спиртов, полиолов и сахаров (Табл. 1). Реакции проводили при температуре 60°С в 50 мМ глицин/NaOH буфере рН 10.5 с добавлением 0.3 мМ NADP и субстрата в концентрации 250 мМ (моно- и дисахариды - в концентрации 50 мМ). Реакцию инициировали добавлением 15-35 мкг фермента в 2 мл термостатированной реакционной смеси. Скорость реакции контролировали по восстановлению NADP на длине волны 340 нм на спектрофотометре Helios Alfa (Thermosystems), снабженном термостатируемым кюветным отделением. За единицу активности принимали количество фермента, необходимое для окисления (или восстановление в обратной реакции) 1 мкмоль NADPH (или NADP) в минуту. В приведенных выше условиях для прямой реакции линейность соблюдалась в интервале концентрации фермента 7-40 мкг/мл в реакционной смеси, что позволило далее анализировать скорости как прямой, так и обратной реакции по схеме Михаэлиса-Ментен. Было установлено, что фермент имеет широкую субстратную специфичность. В реакции окисления наибольшая активность наблюдалась с 2-пропанолом (Vmax, 1.1 ед/мг) и бензиловым спиртом. TsAdh319 проявляла также высокую активность в реакциях окисления D-арабинозы и D-ксилозы при концентрациях субстрата в 5 раз ниже, чем в стандартной реакции с 2-пропанолом (Табл. 1). Окисления 2-пропанола в присутствии NAD не наблюдалось, что подтвердило специфичность алкогольдегидрогеназы к NADP.
Алкогольдегидрогеза TsAdh319 обладает стереоселективностью, о чем свидетельствует вдвое более высокая скорость ферментативного окислении (S)-(+)-2-бутанола по сравнению с (RS)-(±)-2-бутанолом. Еще более высокая стереоселективность была показана в реакции окисления моносахаридов (10-кратная разница в активности относительно D-и L-арабинозы, Табл. 1).
Реакции восстановления проводили при температуре 60°С в 100 мМ натрий-фосфатном буфере рН 7.0 с добавлением 0.3 мМ NADPH и субстрата в концентрации 250 мМ (моносахаридов - в концентрации 50 мМ). Реакцию инициировали добавлением фермента (20-40 мкг) в 2 мл предварительно термостатированной реакционной смеси. Скорость реакции контролировали по окислению NADPH на длине волны 340 нм. Субстратную специфичность TsAdh319 определяли по восстановлению альдегидов, кетонов, моносахаридов и др. (Табл. 2). Фермент проявлял максимальную активность в реакции восстановления диметилглиоксаля (5.6 ед/мг) и метилглиоксаля (2.2 ед/мг). В отличие от прямой реакции окисления, TsAdh319 не проявляла активность в реакции восстановления моносахаридов (Табл. 2). В таблице 3 представлены кинетические характеристики TsAdh319, определенные для реакций с наиболее эффективно используемыми субстратами.
Пример 4. Влияние рН, концентрации солей, ионов металлов и хелатирующих агентов на активность алкогольдегидрогеназы TsAdh319.
Для определения рН-оптимума реакций окисления и восстановления (соответственно, с 2-пропанолом и метилглиоксалем) использовали фосфатный (100 mM, диапазон рН 5.0-8.5) или глициновый (50 mM, диапазон рН 8.5-11.0) буферы, реакцию проводили в стандартных условиях при 60°С.Было установлено (фиг.2), что для реакции окисления спиртов требуется щелочной рН (более 10, оптимум при рН 11). Реакция восстановления имеет относительно узкий диапазон рН вблизи нейтрального (оптимум при рН 6.5-8.5). Влияние солей на активность фермента было охарактеризовано в стандартных условиях реакциях, в присутствии 200-600 мМ NaCl или KCl. Было установлено, что активность фермента не только не ингибируется солями, но что повышение концентрации NaCl до 600 мМ примерно вдвое увеличивает скорости окисления 2-пропанола (Табл. 4) и восстановления метилглиоксаля, меньший эффект наблюдался при добавлении KCl.
Для изучения возможного влияния ионов металлов на активность TsAdh319 фермент прединкубировали в буфере, содержащем 10 мМ соответствующей соли металла, а затем определяли активность фермента в стандартной реакции при 60°С в присутствии 1 мМ соли металла. Было установлено (Табл. 4), что двухвалентные катионы Mg+2, Zn+2, Fe+2 Ni+2 и Со+2 не оказывают существенного ингибирующего или активирующего эффекта на активность TsAdh319 в реакции окисления. Для оценки влияния хелатирующих агентов на активность TsAdh319 в реакцию вносили EDTA в концентрации 1 или 5 мМ. Было установлено, что EDTA не оказывает существенного влияния на активность фермента.
Пример 5. Температурный оптимум и термостабильность алкогольдегидрогеназы TsAdh319.
Зависимость ферментативной активности TsAdh319 от температуры определяли в реакции окисления 2-пропанола. Реакцию проводили в 50 мМ глицин/NaOH буфере рН 10.5, 0.3 мМ NADP и 250 мМ изопропанола при разных температурах, изменение оптической плотности на длине волны 340 нм регистрировали в течение 2 мин. Реакцию инициировали добавлением 20 мкг фермента в предварительно термостатированную реакционную смесь. Было установлено (Фиг.3а), что активность TsAdh319 в реакции окисления изопропанола непрерывно возрастает при увеличении температуры от 40°С (0.09 ед/мг) до 100°С (7.3 ед/мг). Таким образом, температурный оптимум фермента превышает 100°С.
Для характеристики термостабильности алкогольдегидрогеназы TsAdh319 раствор фермента (0.4 мг/мл) в буфере, содержащем 50 mM Tris (рН 7.5) и 200 mM NaCl, инкубировали различное время при температурах 70, 80, 90 или 100°С. После инкубации активность TsAdh319 определяли в стандартной реакции окисления 2-пропанола при 60°С (фиг.3б). Было показано, что фермент обладает высокой термостабильностью: инкубация при 70°С или 80°С не только не снижает, но даже увеличивает активность фермента. Время полуинактивации TsAdh319 при температуре 90°С составляет 2 часа, а при температуре 100°С - 1 час. Таким образом, среди известных короткоцепочечных алкогольдегидрогеназ TsAdh319 имеет наиболее высокий температурный оптимум и является наиболее термостабильной.
СПИСОК ЛИТЕРАТУРЫ



| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРМОСТАБИЛЬНАЯ ДНК-ЛИГАЗА ИЗ АРХЕИ РОДА THERMOCOCCUS, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, КОДИРУЮЩАЯ ЭТУ ДНК-ЛИГАЗУ | 2009 |
|
RU2405823C2 |
| МОДИФИЦИРОВАННЫЙ ГЕН tsA, КОДИРУЮЩИЙ ТЕРМОСТАБИЛЬНУЮ L-АСПАРАГИНАЗУ ИЗ АРХЕИ THERMOCOCCUS SIBIRICUS, РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ L-АСПАРАГИНАЗЫ И СПОСОБ ОЧИСТКИ ЭТОГО ФЕРМЕНТА | 2022 |
|
RU2794225C1 |
| СПОСОБ УЛУЧШЕНИЯ КАТАЛИТИЧЕСКИХ СВОЙСТВ ГИПЕРТЕРМОФИЛЬНЫХ L-АСПАРАГИНАЗ I ТИПА, L-АСПАРАГИНАЗА I ТИПА ИЗ ГИПЕРТЕРМОФИЛЬНОЙ АРХЕИ THERMOCOCCUS SIBIRICUS, ПОЛУЧЕННАЯ ТАКИМ СПОСОБОМ | 2023 |
|
RU2836049C1 |
| ТЕРМОСТАБИЛЬНАЯ ДНК-ЛИГАЗА ИЗ АРХЕИ РОДА ACIDILOBUS | 2009 |
|
RU2413767C1 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОЙ БЕТА-ГАЛАКТОЗИДАЗЫ AsBgl 1390 ИЗ АРХЕИ SACCHAROVORANS В КАЧЕСТВЕ БЕТА-ГЛЮКОЗИДАЗЫ, БЕТА-КСИЛОЗИДАЗЫ И БЕТА-МАННОЗИДАЗЫ | 2012 |
|
RU2523581C2 |
| Рекомбинантный штамм бактерий Escherichia coli Rosetta 2(DE3)/pET28c-TstP36H-Sso7d - продуцент химерной термостабильной ДНК-полимеразы TstP36H-Sso7d из Thermococcus stetteri | 2024 |
|
RU2840236C1 |
| Фермент трансаминаза D-аминокислот из Haliscomenobacter hydrossis с расширенной субстратной специфичностью | 2023 |
|
RU2824015C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pHisTevTSIB0821, ТРАНСФОРМИРОВАННЫЙ ЕЮ ШТАММ Escherichia coli Rosetta(DE3)/pHisTevTSIB0821 И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ПРОЛИДАЗЫ TSIB_0821 | 2012 |
|
RU2509154C1 |
| Способ прогнозирования формирования развернутой астматической триады у больных полипозным риносинуситом после полипотомии | 2018 |
|
RU2679414C1 |
| СПОСОБ ПРОГНОЗА ЧУВСТВИТЕЛЬНОСТИ К ИНТЕРФЕРОНУ У БОЛЬНЫХ ПОЧЕЧНО-КЛЕТОЧНЫМ РАКОМ | 2010 |
|
RU2437101C1 |
Изобретение относится к биотехнологии и представляет собой рекомбинантную термостабильную алкогольдегидрогеназу TsAdh319 из археи Thermococcus sibiricus, проявляющую активность в реакциях окисления спиртов и сахаров до соответствующих альдегидов и кетонов и в обратных реакциях восстановления альдегидов и кетонов при температурах до 100°С. Последовательность алкогольдегидрогеназы состоит из 234 аминокислот и представлена в SEQ ID NO:2. Изобретение относится также к ДНК, кодирующей термостабильную алкогольдегидрогеназу, включающее нуклеотидную последовательность SEQ ID NO:1. Термостабильная алкогольдегидрогеназа TsAdh319 может быть использована в биотехнологической промышленности для органического синтеза биологически активных веществ. 2 н.п. ф-лы, 3 ил., 4 табл.
1. Рекомбинантная термостабильная алкогольдегидрогеназа TsAdh319 из археи Thermococcus sibiricus, проявляющая активность в реакциях окисления спиртов и сахаров до соответствующих альдегидов и кетонов и в обратных реакциях восстановления альдегидов и кетонов при температурах до 100°С, состоящая из 234 аминокислот и содержащая аминокислотную последовательность SEQ ID NO:2, полученная из рекомбинантного штамма Escherichia coli.
2. Выделенная нуклеотидная последовательность ДНК, кодирующая термостабильную алкогольдегидрогеназу по п.1, включающая нуклеотидную последовательность SEQ ID NO:1.
| VAN DER OOST J et.al | |||
| Genetic and biochemical characterization of a short-chain alcohol dehydrogenase from the hyperthermophilic archaeon Pyrococcus furiosus | |||
| Eur J Biochem | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| ANTOINE E et.al | |||
| Cloning and over-expression in Escherichia coli of the gene encoding NADPH group III alcohol dehydrogenase from Thermococcus | |||

