Изобретение относится к области биотехнологии, в частности, к инженерной энзимологии и молекулярной биологии, и касается получения модифицированных гипертермофильных L-аспарагиназ I типа с улучшенными каталитическими характеристиками, получаемые путем экспрессии в клетках Escherichia coli кодирующих их последовательностей, содержащих точечные мутации, в составе рекомбинантной плазмидной ДНК. Настоящее изобретение описывает способ улучшения каталитических свойств гипертермофильных L-аспарагиназ I типа, которые могут найти широкое применение в биотехнологии, прежде всего, в высокотемпературных пищевых технологиях, а также в клинической практике при лечении чувствительных видов опухолей.
L-аспарагиназы (L-АСП) на протяжении нескольких десятилетий остаются одними из важнейших ферментов для биотехнологии [1]. L-АСП (L-аспарагин-амидогидролаза, КФ 3.5.1.1) катализирует гидролиз амидной группы боковой цепи L-аспарагина с образованием L-аспарагиновой кислоты и аммиака [2]. Это первый для клинической онкогематологии фермент со специфическим действием на опухолевые клетки. На протяжении более 40 лет он остается одним из основных препаратов комбинированной химиотерапии острого лимфобластного лейкоза [3,4]. Также имеются данные об эффективности фермента при лимфосаркоме, лимфогранулематозе, остром миелоидном лейкозе, ретикулосаркоме, солидных и ряде других видов опухолей [5-15].
Помимо биомедицинского применения в настоящее время проводятся исследования по введению L-АСП в технологии пищевых производств для снижения образования токсичных продуктов реакции Майяра, в частности, акриламида, потенциально канцерогенного и нейротоксического вещества [16,17]. Акриламид образуется в пищевых продуктах в процессе взаимодействия L-аспарагина с редуцирующими сахарами в реакции Майяра [18]. Образование акриламида протекает в условиях высокой температуры (свыше 120°C) и низкой влажности из сырья с высоким содержанием углеводов. Наибольшие его количества содержатся в пищевых продуктах, полученных в результате жарки, выпечки, запекания, обжаривания или приготовления на гриле [18]. Обработка L-АСП выступает одним из способов снижения уровня L-аспарагина, следовательно, акриламида в пищевых продуктах при сохранении их органолептических характеристик [16,19-22]. Однако для эффективного применения L-АСП в пищевых технологиях требуются варианты с повышенной активностью при высоких температурах и термостабильностью.
Еще одним направлением применения L-АСП является создание на ее основе биосенсоров для диагностических целей и мониторинга содержания уровня L-аспарагина в пищевых продуктах [23-28].
Таким образом, область применения L-АСП по изобретению достаточно широка. Принимая во внимание потенциал фермента в технологиях здравоохранения и здоровьесбережения, биоинженерия L-АСП с целью получения форм с улучшенными свойствами, представляет повышенный исследовательский интерес.
L-АСП, применяемые на данный момент в биотехнологии, выделены из мезофильных микроорганизмов и имеют ряд недостатков. Для использования в клинической практике при терапии чувствительных видов опухолей разрешены две L-АСП из мезофильных бактерий Esherichia coli и Erwinia chrysanthemi [29]. Тем не менее продолжительное применение этих препаратов зачастую сопровождается большим количеством побочных эффектов вплоть до развития анафилактического шока [30,31]. Нежелательные проявления связывают, в первую очередь, с сопутствующей L-глутаминазной активностью [10,12]. Считается также, что L-АСП мезофильного происхождения вызывают аллергическую реакцию у пациентов из-за мультисубъединичной структуры [32]. Существенным недостатком мезофильных L-АСП является низкая стабильность. Повышение стабильности применяемых в биомедицине L-АСП способствует улучшению их фармакокинетических характеристик - увеличению продолжительности циркуляции в крови, снижению кратности дозирования и вероятности развития аллергических проявлений [33,34].
В пищевых технологиях в настоящее время разрешены к применению L-АСП из грибов Aspergillus oryzae и Aspergillus niger с температурным оптимумом в диапазоне 35-50 °С [35,36]. В то же время, согласно экспериментальным данным, эффективному снижению акриламида в пищевых продуктах способствует обработка L-АСП при температуре свыше 80 °С [37]. Таким образом, для высокотемпературных пищевых технологий актуальным является поиск либо инженерия новых вариантов L-АСП с температурным оптимумом свыше 80 °С, высокой активностью и стабильностью в области повышенных температур.
Перспективными с биотехнологической точки зрения являются L-АСП термофильных и гипертермофильных микроорганизмов. Так, в исследованиях рекомбинантных L-АСП I типа гипертермофилов родов Pyrococcus и Thermococcus показано, что данные ферменты, активные в димерном состоянии, при температурном оптимуме свыше 85 °С обладают уникальными свойствами - высокой энзиматической активностью, стабильностью в широком диапазоне температур, рН, в присутствии солей и денатурирующих условиях, что расширяет возможности их применения в пищевой промышленности и диагностике, а также биотерапии [38-40]. Они имеют неоспоримые преимущества перед известными на данный момент препаратами при применении в высокотемпературных пищевых технологиях. В то же время повышенная стабильность этих L-АСП позволяет компенсировать потерю активности в неоптимальных, физиологических условиях и конкурировать с L-АСП мезофильных микроорганизмов в биомедицине [40,41]. Следует отметить, что в отличие от применяемых в клинической практике гомотетрамерных бактериальных L-АСП, ферменты гипертермофильного происхождения активны в димерном состоянии [1].
К настоящему времени биохимически охарактеризованы L-АСП I типа из гипертермофильных архей Pyrococcus и Thermococcus sp.: Pyrococcus furiosus [42], Pyrococcus yayanosii CH1 [38], Thermococcus kodakaraensis [40,43,44], Thermococcus zilligii [37], Thermococcus gammatolerans [45], Thermococcus sibiricus [46]. Данные нативные L-АСП превосходят по характеристикам улучшенные варианты мезофильных L-АСП с повышенной термостабильностью, разработанные для пищевых производств и описанные в патентах:
- мутантные формы L-АСП Aspergillus oryzae - их остаточная активность при инкубации в течение 20 минут при 66 °С не превышает 72 % [47];
- мутантные формы L-АСП Aspergillus oryzae и Aspergillus niger - термостабильные мутантные варианты характеризуются снижением активности в 2 и более раз при повышенной температуре 70 °С в сравнении с активностью при температурном оптимуме 50 °С [48];
- в качестве источника термостабильной L-АСП для биотехнологии в патенте KR101777904B1 описан штамм Bacillus methylotrophicus MKSY2013, однако выводы о термостабильности данной L-АСП в диапазоне температур 60-95 °С в течение 30 минут сделаны на основании оценки свойств не очищенного белка, а супернатанта [49].
Нативные L-АСП гипертермофильных архей при оптимуме 85-95 °С способны сохранять активность в интервале температур 70-90 °С до 32 часов [44]. Следует отметить, что активность этих L-АСП сильно варьирует. L-АСП Pyrococcus sp. проявляют более низкую энзиматическую активность в сравнении с L-АСП Thermococcus sp.: L-АСП Pyrococcus sp. - 550÷1483.8 МЕ/мг [38,42]; L-АСП Thermococcus sp. - 978.7÷7622 МЕ/мг [37,40,45] (активность для L-АСП мезофильных микроорганизмов - 50-450 МЕ/мг).
Активное изучение данной группы ферментов стимулировало работы по исследованию их структурно-функциональных свойств и получению мутантных вариантов с улучшенными характеристиками, свойства которых описаны в ряде патентов [50-52].
Для биотехнологического применения в патентах предлагается использовать мутантные варианты гипертермофильных L-АСП из архей Pyrococcus yayanosii CH1 (PyA) и Pyrococcus furiosus (PfA).
В группе патентов US-11001825-B2, US-2019185840-A1, WO-2019113965-A1, CN-108103049-A указано, что экспрессия в Bacillus subtilis 168 последовательности PfA, содержащей комбинацию мутаций S17G/A90S/R156S/K272A, позволяет получать ферментный препарат с L-аспарагиназной активностью 3108 МЕ/мг [50,51].
Для биомедицинского применения запатентованы последовательности L-АСП Pyrococcus furiosus, содержащие мутации K274E, T53Q и их комбинацию T53Q/K274E, - патент US9322008B2 [52]. При экспрессии в клетках E. coli данных мутантных форм, характеризующихся повышенной стабильностью и отсутствием сопутствующей глутаминазной активности, показана выраженная цитотоксическая активность.
В то же время в патентной литературе не представлено сведений по вопросам белковой инженерии L-АСП I типа из архей Thermococcus sp. Права патента WO2013027977A2, предусматривающего использование L-АСП Thermococcus kodakaraensis, ограничиваются способом получения данного фермента и разработкой композиции его содержащей для деградации L-аспарагина [53].
Таким образом, представлены единичные примеры изменения аминокислотных последовательностей L-АСП Pyrococcus yayanosii CH1 (PyA) и Pyrococcus furiosus (PfA), при этом ранее заявленные способы не направлены на инженерию гипертермофильных L-АСП Thermococcus sp.
Технической задачей заявляемого изобретения является создание модифицированных гипертермофильных L-АСП I типа, обладающих улучшенными каталитическими свойствами, для биотехнологического применения.
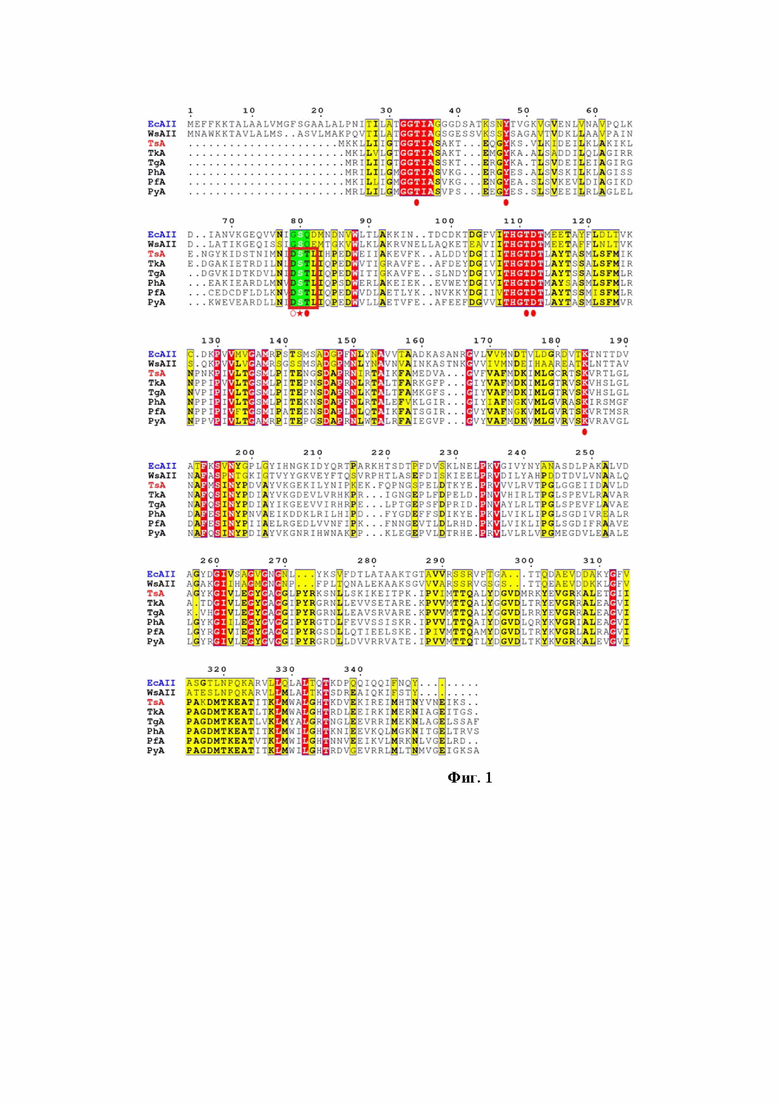
Поставленная задача решается путем изменения структуры L-АСП I типа в области активного центра двойной заменой аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL (приведен в однобуквенном коде) (Фиг. 1), примыкающих к субстрат-связывающему S, где в 1 положении произведена замена D/G, в третьем положении T/Q.
При сравнении последовательностей термофильных и мезофильных L-АСП выявлено, что в ряде ферментов мезофильного происхождения высококонсервативный субстрат-связывающий S58 окружен G57 и Q59 (нумерация по ферменту из E.coli EcAII). В гипертермофильных L-АСП I типа субстрат-связывающий S55 окружен остатками D54 и T56 (Фиг. 1). Здесь и далее нумерация аминокислотной последовательности гипертермофильных L-АСП I типа осуществляется по аминокислотной последовательности нативной формы фермента из T. sibiricus TsA (SEQ ID NO 1). Позиции аминокислотных остатков гипертермофильных L-АСП I типа, соответствующие аминокислотной последовательности TsA (SEQ ID NO 1), определяются согласно общепринятым методам выравнивания аминокислотных последовательностей L-АСП из разных источников [43,46].
Мутации D/G и T/Q в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, восстанавливающие триаду GSQ, характерную для ряда мезофильных гомологов, включая EcAII, способствуют повышению подвижности петли и улучшению ферментативных свойств гипертермофильных L-АСП I типа.
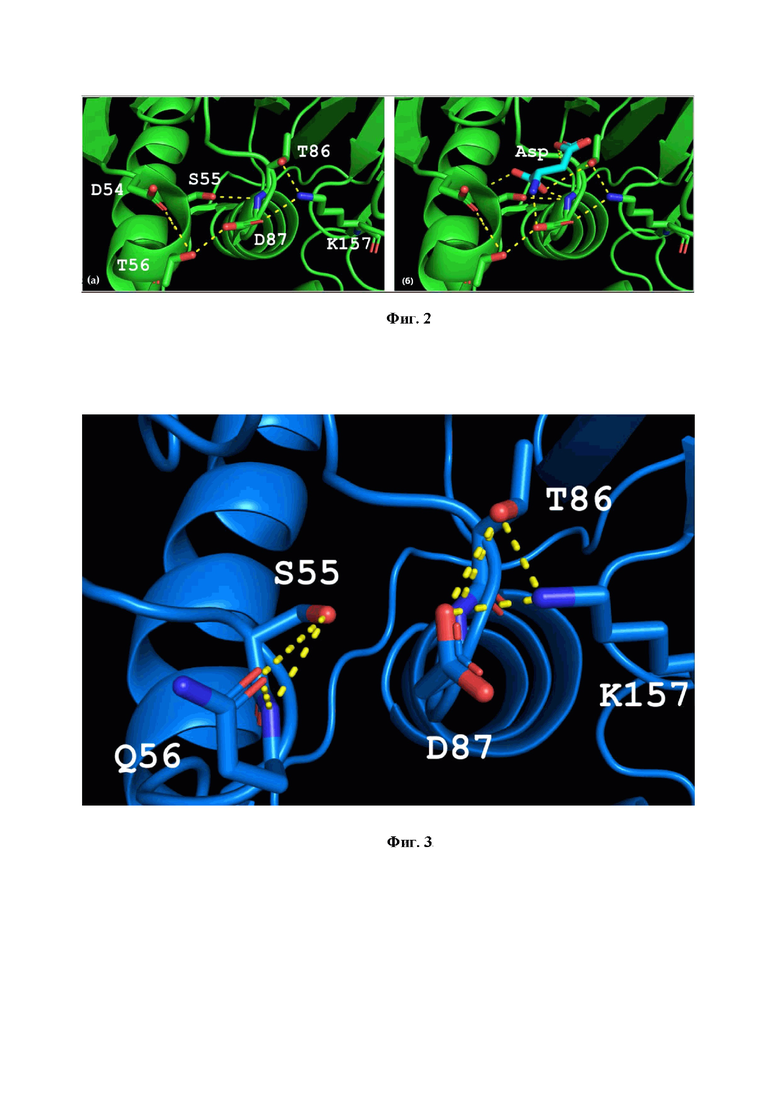
В гипертермофильных L-АСП остаток Т56 участвует в формировании дна активного центра с большим числом взаимодействий (Фиг. 2а). В основные взаимодействия этой части активного сайта вовлечены аминокислотные остатки D54, S55, T56, T86, D87 и K157. Они формируют сеть водородных связей, которая может ограничивать движение петли (аминокислотные остатки 85-88).
На модели комплекса TsA-субстрат, полученного путем включения аспартата из комплекса L-АСП EcAII (pdb 3eca) после выравнивания третичных структур данных ферментов, выявлено, что T86, D54 и K157 также участвуют во взаимодействии с субстратом (продуктом) (Фиг. 2б).
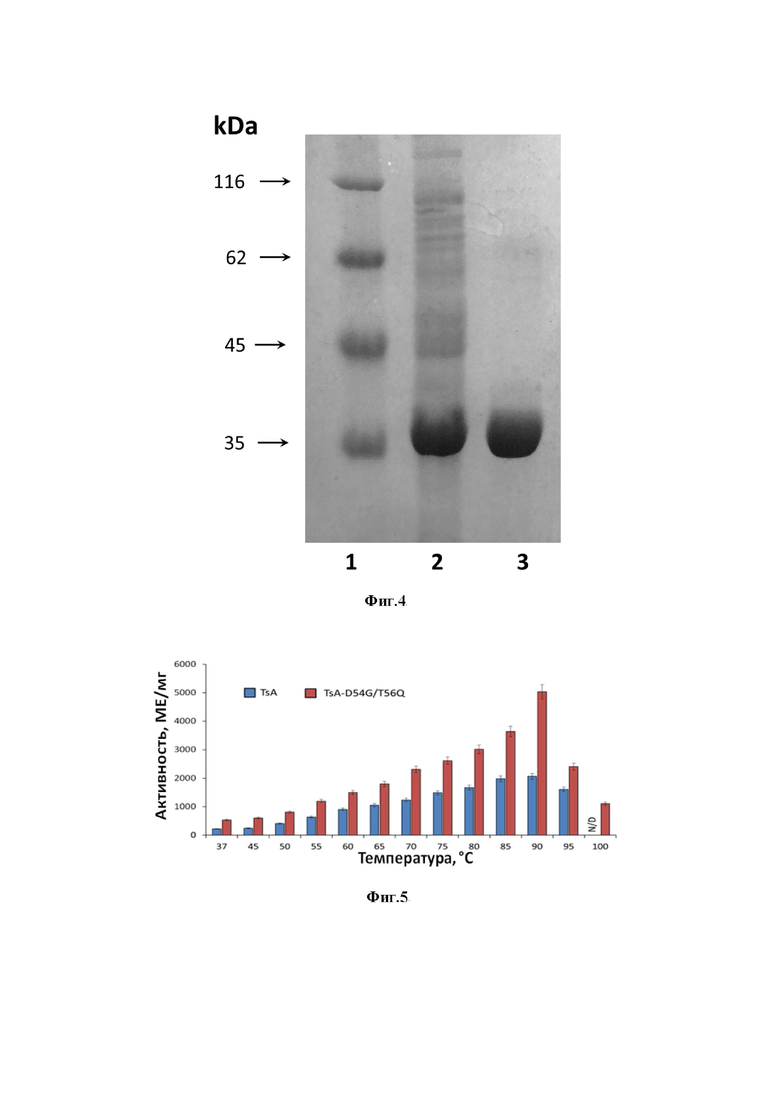
Замена короткого треонина на длинный глутамин в результате мутации T56Q способствует разрушению сети водородных связей (Фиг. 3), смещая амидную группу Q от D87 к D54. Разрушение этих водородных связей в жесткой структуре гипертермофильных L-АСП обуславливает повышение подвижности петли и способствует повышению катализа. В то же время, ввиду малых расстояний, появление длинного глутамина вместо короткого треонина связано с риском отдаления аминокислотных остатков глутамина и аспартата от субстрата, что может привести к снижению эффективности образования субстрат-ферментного комплекса. Для сохранения архитектуры активного центра и обеспечения высокой каталитической активности одновременно с мутацией T56Q необходима замена D54G (Фиг. 3).
Все вышесказанное относится в равной степени к гипертермофильным L-АСП I типа, имеющих общее c TsA строение активного центра в этой части.
Выводы подтверждаются ранее представленными экспериментальными данными [39,54]. Попытка одиночной замены T53Q в последовательности гипертермофильной L-АСП P. furiosus приводила к повышению аффинности PfA к субстрату при одновременном более чем 2-кратном снижении каталитической эффективности фермента [39]. Одиночная точечная замена D/G в этой области снижала L-аспарагиназную активность термо-АСП T. gammatolerans и P. yayanosii, ввиду нарушения удерживания субстрата каталитическим остаком S и соседним G после замены D [54].
В соответствии с вышесказанным данное изобретение направлено на получение рекомбинантных гипертермофильных L-АСП I типа с улучшенными каталитическими свойствами, которые отличаются от нативных последовательностей тем, что содержат комбинацию замен аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q.
Краткое описание чертежей, иллюстрирующих заявляемую группу изобретений
Заявляемая группа изобретений проиллюстрирована следующими фигурами.
Фиг. 1. Выравнивание аминокислотных последовательностей L-аспарагиназ различного происхождения:
- мезофильных L-аспарагиназ из Escherichia coli (EcAII, тип II-AAA23445.1, аминокислотная последовательность выделена синим цветом) и Wolinella_succinogenes (WsA, тип II-WP_011138590.1),
- гипертермофильных L-аспарагиназ I типа из Thermococcus sibiricus (TsA, WP_015849943, выделена красным цветом), Thermococcus kodakarensis (TkA, WP_011250607.1), Thermococcus gammatolerans (TgA, WP_015859055), Pyrococcus horikoshii (PhA, WP_010884185), Pyrococcus furiosus (PfA, WP_011013191), Pyrococcus yayanosii (PyA, WP_013906452).
Идентичные аминокислотные остатки выделены красным цветом, консервативные аминокислотные остатки выделены желтым цветом. Консервативные аминокислотные остатки, ключевые для каталитической активности L-аспарагиназ, отмечены красными кружками. Аминокислотный остаток субстрат-связывающего серина обозначен красной звездочкой, примыкающие к нему аминокислотные остатки, входящие в состав высококонсервативного для гипертермофильных L-аспарагиназ I типа мотива DSTL, выделенного красной рамкой, обозначены полым и заполненным красными кружками.
Фиг. 2. Структура дна активного центра гипертермофильных L-аспарагиназ I типа на примере модели L-аспарагиназы I типа из T. sibiricus:
(а) - сеть водородных связей (обозначены желтым) в активном центре фермента,
(б) - взаимодействие гипертермофильной L-аспарагиназы с аспартатом (продуктом).
Фиг. 3. Структура дна активного центра гипертермофильных L-аспарагиназ I типа на примере модели L-аспарагиназы I типа из T. sibiricus после замены аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q.
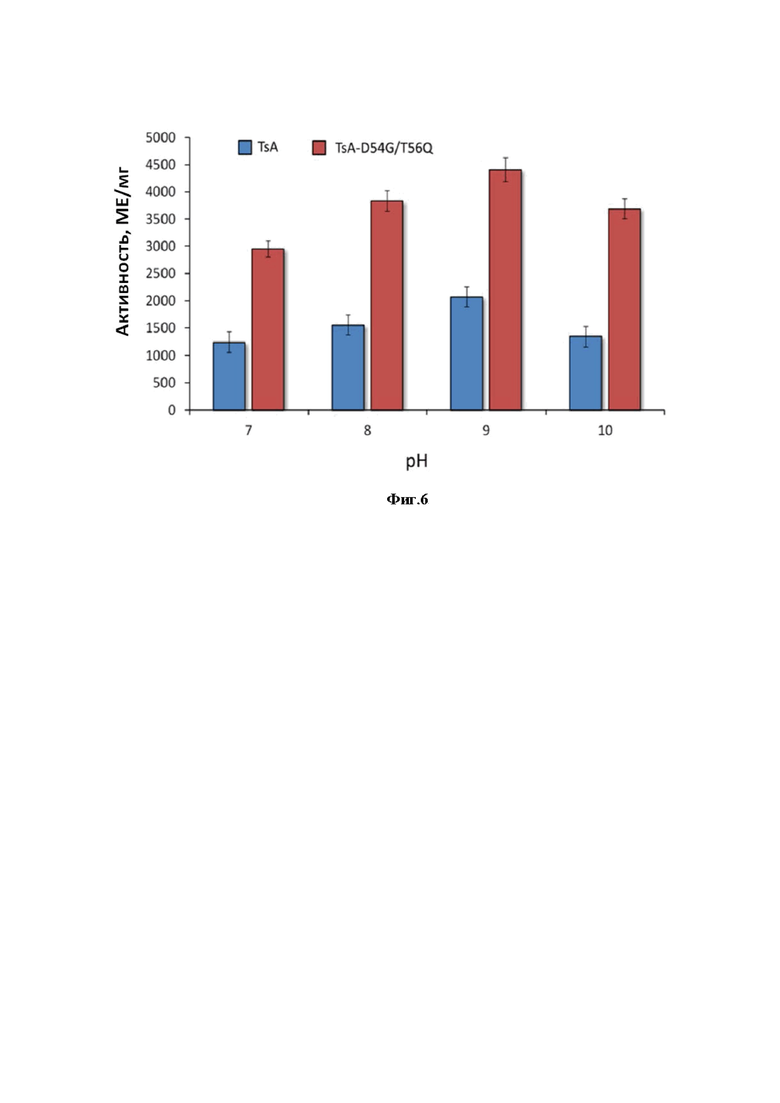
Фиг. 4. Экспрессия TsA_GQ в клетках E. coli и очистка белка.
Приведены результаты электрофоретического анализа белковых препаратов:
1 - маркер молекулярного веса белков,
2 - TsA_GQ в бесклеточном гомогенате в концентрации 20 мкг,
3 - образец очищенного TsA_GQ в концентрации 10 мкг.
Фиг. 5. Зависимость абсолютной активности TsA и TsA_GQ от температуры.
ND - недетектируемое значение.
Фиг. 6. Зависимость абсолютной активности TsA и TsA_GQ от рН.
При осуществлении изобретения, помимо методов, подробно раскрытых в нижеследующих примерах, использовались хорошо известные методики, описанные в руководствах по молекулярной биологии и генетической инженерии [55].
Изобретение проиллюстрировано на примерах, которые не ограничивают объем изобретения, определяемый формулой изобретения. Все сходные замены и модификации, очевидные для специалистов в данной области, находятся в рамках идеи, объема и концепции данного изобретения, определяемые прилагаемой формулой изобретения.
Осуществление изобретения раскрыто на примере гипертермофильного фермента I типа из T. sibiricus TsA, характеризующегося высокой активностью - более 2000 МЕ/мг - при температуре 90 °С и повышенным сродством к субстрату Km=3 мМ в сравнении с другими термо-аспарагиназами - Km=5÷12 мМ [46,56].
Пример 1. Получение модифицированной формы TsA и рекомбинантного штамма-продуцента.
Модифицированный вариант гипертермофильной L-АСП I типа TsA получали путем конструирования мутантного гена tsA_mod_gq (SEQ ID NO: 2) методом сайт-направленного мутагенеза. Для этого в качестве матрицы использовали вариант гена tsA_mod с нуклеотидной последовательностью SEQ ID NO: 3, ранее оптимизированный для экспрессии архебактериальной L-АСП в клетках E. coli и клонированный в вектор pET-28a(+) [46].
Сконструированную плазмиду pET-28a(+)_TsA_GQ после подтверждения секвенированием результатов сайт-направленного мутагенеза трансформировали в компетентные клетки штамма E. coli BL21 (DE3). Штамм-продуцент модифицированной формы TsA (TsA_GQ, SEQ ID NO: 4.), содержащей 4-членный мотив GSQL, был обозначен как E. coli BL21(DE3)/pMЕ_GQ.
Пример 2. Экспрессия, выделение и очистка модифицированной гипертермофильной L-аспарагиназы
Для экспрессии модифицированной гипертермофильной L-АСП TsA_GQ рекомбинантный штамм E. coli BL21(DE3)/pMЕ_GQ культивировали в среде LB с добавлением канамицина 0.05 мг/мл при 37°C. Лактозу в качестве индуктора добавляли в среду при OD600 = 1.9 до конечной концентрации 0.2 %. Клеточную биомассу собирали через 17-20 ч после индукции центрифугированием при 4000 g, 15 мин.
Все стадии выделения и очистки TsA_GQ проводили при 4°С. 8 г охлажденной биомассы ресуспендировали в 100 мл буфера А (20 мМ Na-фосфатный буфер рН 7.2, 1 мМ глицин, 1 мМ ЭДTA), далее проводили ультразвуковую обработку суспензии клеток [56] и удаление дебриса центрифугированием (35000 g, 30 мин). Очистка фермента из клеток штамма-продуцента E. coli BL21(DE3)/pMЕ_GQ включала стадию ионообменной хроматографии на колонках с SP-Sepharose. Фермент элюировали линейным градиентом концентрации NaCl (0-1.0 М). Фракции, содержавшие TsA_GQ (зона элюции - 0.46-0.70 М NaCl), концентрировали в ячейке Amicon («Millipore», США). Препараты хранили при -20°C.
Концентрацию белка определяли методом Sedmak [57], используя в качестве стандарта бычий сывороточный альбумин. Чистоту белка и его молекулярную массу определяли при помощи ДСН-ПААГ-электрофореза (Фиг. 4).
Экспрессия tsA_mod_gq (SEQ ID NO: 2) в составе вектора pET-28a(+)_TsA_GQ в клетках рекомбинантного штамма E. coli BL21(DE3)/pMЕ_GQ обеспечивала высокий выход активного целевого белка. В ходе выделения и очистки TsA_GQ из клеток штамма-продуцента методом ионообменной хроматографии был получен белок со степенью очистки 2.7 (таблица 1, Фиг. 4).
Таблица 1 - Очистка рекомбинантной модифицированной гипертермофильной L-АСП TsA_GQ
Пример 3. Сравнительное исследование энзиматической активности, субстратной специфичности и кинетических характеристик нативной и модифицированной форм TsA
При определении ферментативной активности различных форм L-АСП T. sibiricus за 1 единицу активности принимали количество фермента, катализирующее высвобождение 1 мкМ аммиака в минуту при экспериментальных условиях.
Количество образовавшегося аммиака определяли методом прямой несслеризации [58,59].
Реакции для TsA_wt (SEQ ID NO: 1) проводили в 0.05 М трис-HCl буфере, рН 9.0, для модифицированной формы TsA_GQ (SEQ ID NO: 4) в 0.01 М буфере глицин-NaOH, рН 9.3 при 90°C.
L-глутаминазную активность определяли аналогичным образом, используя в качестве субстрата L-глутамин.
Удельную активность выражали в ME/мг белка.
Определение кинетических параметров TsA_GQ проводили в 0.01 М буфере глицин-NaOH, рН 9.3, содержащем 20-250 мкМ L-аспарагина, при 90 °C. Обработку полученных данных и расчет KМ и Vmax проводили с использованием метода Лайнуивера-Берка [56].
В сравнительном исследовании выявлено более чем 2-кратное повышение каталитической активности для модифицированной формы TsA_GQ относительно фермента дикого типа. Активность TsA_GQ составила 5037.7 МЕ/мг, TsA - 2066.1 МЕ/мг. При этом также отмечалось небольшое снижение относительной L-глутаминазной активности: TsA - 7 %, TsA_GQ - 5 %.
Кинетические константы при использовании в качестве субстрата L-аспарагина для TsA_GQ составили: Vmax 8170 мкМ/мин, Kм 6.0 мМ. В аналогичных экспериментальных условиях для TsA были получены следующие значения кинетических констант: Vmax 4020 мкМ/мин, Kм 3.0 мМ.
Таким образом, комбинация 2 точечных мутаций D54G/T56Q в области активного центра гипертермофильной L-АСП TsA или иначе замена аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q, привела к росту активности модифицированной формы на 240 % относительно нативной формы при одновременном повышении значений кинетических параметров Kм и Vmax.
Пример 4. Исследование влияния температуры и рН на ферментативную активность нативной и модифицированной форм TsA.
Специфическую активность очищенной нативной и модифицированной форм TsA определяли при различной температуре и рН методом прямой несслеризации. Активность ферментов измеряли в диапазоне температур 45÷100 °C с шагом 5°С: TsA_GQ в 0.01 М буфере глицин-NaOH (рН 9.3), TsA в 0.05 М трис-HCl буфере (рН 9.0) Дополнительно активность была измерена при физиологической температуре 37 °C.
Влияние рН на активность нативной и модифицированной форм TsA изучали при температуре 90°С в различных буферных системах: 0.05 М Na-ацетатной (pH 4.0-6.0), 0.05 М Na-фосфатной (рН 6.0-7.0), 0.05 М трис-HCl (рН 7.0-9.0), 0.05 М глицин-NaOH (рН 9.0-10.0).
В исследовании влияния температуры на активность фермента установлено, что введение мутаций D54G/T56Q в высококонсервативный мотив DSTL не приводило к изменению температурного оптимума фермента (Фиг. 5). Обе формы TsA проявляли максимальную активность при температуре 90 °C. При этом при 100 °C относительная активность модифицированной формы TsA_GQ составила 22 %, в то же время у нативной формы TsA при данном значении температуры активность отсутствовала (Фиг. 5).
Между исследуемыми формами TsA выявлены различия в температурном профиле активности. Более узкий температурный оптимум для TsA_GQ в сравнении с нативной формой свидетельствует о его большей чувствительности к температурным изменениям. Во всем тестируемом температурном диапазоне активность изучаемых форм TsA превышала 1000 МЕ/мг. При этом, несмотря на более высокую скорость снижения активности вне температурного оптимума, по абсолютным значениям активности TsA_GQ превосходил TsA (Фиг. 5). При температуре оптимума 90 °C рост каталитической активности модифицированной формы TsA_GQ составил 240 % (Фиг. 5).
При изучении зависимости активности от рН среды в различных буферных системах в диапазоне значений рН 4.0÷10.0 установлено, что ферменты проявляют схожий характер рН-зависимости. TsA проявляет максимальную активность при рН 9.0, рН оптимум TsA_GQ составляет рН 9.3. Обе формы сохраняют высокую активность в интервале рН 7.0÷10.0 независимо от буферной системы (Фиг. 6). При этом модифицированная форма TsA_GQ в диапазоне рН 7.0-10.0 превосходит по абсолютной активности фермент дикого типа ~ в 2 раза (Фиг. 6).
Пример 5. Изучение химической стабильности, эффектов ионов металлов для нативной и модифицированной форм TsА
Устойчивость ферментов к химической денатурации изучали в присутствии мочевины 0-8 М при инкубации в течение 1 ч. Инкубацию проводили в 0.05 М трис-HCl буфере (рН 9.0) - для TsА, в 0.01 М буфере глицин-NaOH (рН 9.3) - для TsA_GQ. За 100% принимали активность фермента при 90°С в отсутствие мочевины. Измеренную активность фермента сопоставляли с таковой без добавления мочевины в данных экспериментальных условиях.
Установлено, что обе формы фермента проявляют высокую устойчивость к химической денатурации. Мочевина до концентрации 6 М не приводила к значимому снижению активности TsA. Модифицированная форма TsA_GQ проявляла еще большую устойчивость к химической денатурации, сохраняя 54.6% от первоначальной активности при инкубации в присутствии 8 М мочевины.
Активность нативной и модифицированной форм TsA определяли при добавлении ионов металлов (Ni2+, Cu2+, Mg2+, Zn2+, Ca2+, Fe3+), а также ЭДТА. Все вещества (NiCl2, CuSO4, MgCl2, ZnCl2, CaCl2, FeCl3 и ЭДТА) добавляли в концентрации 10.0 мМ. Ферментативную активность определяли при температуре 90°С и оптимуме рН при добавлении L-аспарагина и соответствующих ионов металлов либо ЭДТА в трех повторностях. Полученные значения выражали в % от активности фермента без добавления катионов металлов, которую принимали за 100%.
Согласно экспериментальным данным, добавление ионов металлов Ni2+, Cu2+, Mg2+, Zn2+, Ca2+, а также ЭДТА не оказывало значимого ингибирующего эффекта на активность обеих форм фермента. В присутствии ионов Fe3+ наблюдалось снижение специфической активности TsA_GQ на 82.5%, TsA - на 85.0%.
Таким образом, экспериментально подтверждено, что замены в 1 и 3 положении 4-членного высококонсервативного мотива DSTL в аминокислотной последовательности TsA, соответствующие мутациям D54G и T56Q, повышают каталитическую активность фермента на 240 % при 90 °С. При этом гипертермофильные L-АСП I типа, представителем которых является TsA, имеют общее c TsA строение активного центра в этой части. Комбинация замен аминокислотных остатков в 1 и 3 положении 4-членного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q, в аминокислотных последовательностях гипертермофильных L-АСП I типа способствует повышению гибкости активного центра, облегчая конформационные изменения при связывании с субстратом.
Каждая из мутаций в отдельности в высококонсервативном мотиве DSTL, D/G либо T/Q, приводит к нарушению оптимального связывания и удерживания субстрата, снижая каталитическую эффективность/активность гипертермофильных L-АСП I типа [39,54]. При этом комбинация замен D/G и T/Q в 1 и 3 положении мотива DSTL, заявляемая в данном изобретении, обеспечивает необходимую стабилизацию субстрата в процессе каталитического превращения, а также повышает гибкость структуры вокруг активного центра, способствуя связыванию, реориентации субстрата и улучшая каталитические свойства этих ферментов.
Для модифицированного фермента TsA_GQ, полученного указанным способом, активность в физиологических условиях при 37°C, а также при температурном оптимуме 90°C возросла в сравнении с нативным TsA в 2 раза, что делает модифицированную форму более перспективной как для применения в высокотемпературных пищевых технологиях, так и в биомедицине.
Список использованной информации
1. Dumina, M.V.; Eldarov, M.A.; Zdanov, D.D.; Sokolov, N.N. L-Asparaginases of Extremophilic Microorganisms in Biomedicine. Biochem. Suppl. Ser. B Biomed. Chem. 2020, 14, doi:10.1134/S1990750820040046.
2. Oettgen HF; Old LJ; Boyse EA Inhibition of Leukemias in Man by L-Asparaginase. Cancer Res. 1967, 27, 2619-2631.
3. Solomon, B.; Parihar, N.; Ayodele, L.; Hughes, M. Global Incidence and Prevalence of Acute Lymphoblastic Leukemia: A 10-Year Forecast. J Blood Disord Transfus 2017, 8, 24, doi:10.4172/2155-9864-C1-028.
4. Brumano, L.P.; da Silva, F.V.S.; Costa-Silva, T.A.; Apolinario, A.C.; Santos, J.H.P.M.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C. de O.; Benyahia, B.; Junior, A.P. Development of L-Asparaginase Biobetters: Current Research Status and Review of the Desirable Quality Profiles. Front. Bioeng. Biotechnol. 2018, 6, 1-22, doi:10.3389/fbioe.2018.00212.
5. Avramis, V.I.; Tiwari, P.N. Asparaginase (Native ASNase or Pegylated ASNase) in the Treatment of Acute Lymphoblastic Leukemia. Int. J. Nanomedicine 2006, 1, 241-254.
6. Verma, N.; Kumar, K.; Kaur, G.; Anand, S. L-Asparaginase: A Promising Chemotherapeutic Agent. Crit. Rev. Biotechnol. 2007, 27, 45-62, doi:10.1080/07388550601173926.
7. Abakumova, O.Y.; Podobed, O. V.; Karalkin, P.A.; Kondakova, L.I.; Sokolov, N.N. Antitumor Activity of L-Asparaginase from Erwinia Carotovora against Different Human and Animal Leukemic and Solid Tumor Cell Lines. Biochem. Suppl. Ser. B Biomed. Chem. 2012, doi:10.1134/s1990750812040026.
8. Kumar, K.; Kaur, J.; Walia, S.; Pathak, T.; Aggarwal, D. L-Asparaginase: An Effective Agent in the Treatment of Acute Lymphoblastic Leukemia. Leuk. Lymphoma 2014, 55, 256-262, doi:10.3109/10428194.2013.803224.
9. Абакумова О. Ю.; Подобед О. В.; Каралкин П.А.; Кондакова Л.И.; Соколов Н.Н. Противоопухолевая Активность L-Аспарагиназы Erwinia Carotovora Для Клеток Различных Лейкемий Человека и Солидных Опухолей Человека и Животных. Биомедицинская химия 2013, 59, 498-513.
10. Соколов Н.Н.; Эльдаров М.А.; Покровская М.В.; Александрова С.С.; Абакумова О.Ю.; Подобед О.В.; Мелик-Нубаров Н.С.; Кудряшова Е.В.; Гришин Д.В.; Арчаков А.И. Бактериальные Рекомбинантные L-Аспарагиназы: Свойства, Строение и Антипролиферативная Активность. Биомедицинская химия 2015, 61, 312-324.
11. Avramis, V.I. Is Glutamine Depletion Needed in ALL Disease? Blood 2014, 123, 3532-3533, doi:10.1182/blood-2014-04-565523.
12. Matsumoto, Y.; Nomura, K.; Kanda-Akano, Y.; Fujita, Y.; Nakao, M.; Ueda, K.; Horiike, S.; Yokota, S.; Kusuzaki, K.; Kitoh, T.; et al. Successful Treatment with Erwinia L-Asparaginase for Recurrent Natural Killer/T Cell Lymphoma. Leuk. Lymphoma 2003, 44, 879-882, doi:10.1080/1042819031000067873.
13. Müller, H.J.; Boos, J. Use of L-Asparaginase in Childhood ALL. Crit. Rev. Oncol. Hematol. 1998, 28, 97-113., doi:10.1016/S1040-8428(98)00015-8.
14. Hill, J.M.; Roberts, J.; Loeb, E.; Khan, A.; Maclellan, A.; Hill, R.W. L-Asparaginase Therapy for Leukemia and Other Malignant Neoplasms: Remission in Human Leukemia. JAMA J. Am. Med. Assoc. 1967, 202, doi:10.1001/jama.1967.03130220070012.
15. Duval, M.; Suciu, S.; Ferster, A.; Rialland, X.; Nelken, B.; Lutz, P.; Benoit, Y.; Robert, A.; Manel, A.M.; Vilmer, E.; et al. Comparison of Escherichia Coli-Asparaginase with Erwinia-Asparaginase in the Treatment of Childhood Lymphoid Malignancies: Results of a Randomized European Organisation for Research and Treatment of Cancer - Children’s Leukemia Group Phase 3 Trial. Blood 2002, doi:10.1182/blood.V99.8.2734.
16. Baskar G; Subanjalin Joy S; Aiswarya R Optimization of Enzymatic Pretreatment and Frying Conditions for Acrylamide Mitigation in Fried Tapioca Chips. Int. J. Mod. Sci. Technol. 2016, 6, 224-229.
17. Aiswarya, R.; Baskar, G. Enzymatic Mitigation of Acrylamide in Fried Potato Chips Using Asparaginase from Aspergillus Terreus. Int. J. Food Sci. Technol. 2018, doi:10.1111/ijfs.13608.
18. Baskar, G.; Aiswarya, R. Overview on Mitigation of Acrylamide in Starchy Fried and Baked Foods. J. Sci. Food Agric. 2018.
19. Gurunathan, B.; Sahadevan, R. Production of L-Asparaginase from Natural Substrates by <em>Aspergillus Terreus</Em> MTCC 1782: Optimization of Carbon Source and Operating Conditions. Int. J. Chem. React. Eng. 2011, doi:10.2202/1542-6580.2479.
20. Abboudi, M.; Al-Bachir, M.; Koudsi, Y.; Jouhara, H. Combined Effects of Gamma Irradiation and Blanching Process on Acrylamide Content in Fried Potato Strips. Int. J. Food Prop. 2016, doi:10.1080/10942912.2014.968790.
21. Acrylamide Reduction in Potato Crisps Using: Asparaginase from Candida Utilis, Commercial Asparaginase, Salt Immersion, and PH Treatment. Int. J. Eng. 2016, doi:10.5829/idosi.ije.2016.29.07a.01.
22. Enhancement of L-Asparaginase Production by Candida Utilis in a 13 L Fermenter and Its Purification. Int. J. Eng. 2015, doi:10.5829/idosi.ije.2015.28.08b.04.
23. Verma, N.; Bansal, M.; Kumar, S. Whole Cell Based Miniaturized Fiber Optic Biosensor to Monitor L-Asparagine. Adv Appl Sci Res 2012.
24. Verma, N.; Kumar, K.; Kaur, G.; Anand, S. E. Coli K-12 Asparaginase-Based Asparagine Biosensor for Leukemia. Artif. Cells, Blood Substitutes, Biotechnol. 2007, doi:10.1080/10731190701460358.
25. Hurst, P.L.; Boulton, G.; Lill, R.E. Towards a Freshness Test for Asparagus: Spear Tip Asparagine Content Is Strongly Related to Post Harvest Accumulated Heat Units. Food Chem. 1998, doi:10.1016/S0308-8146(97)00031-9.
26. Izco, J.M.; Torre, P.; Barcina, Y. Ripening of Ossau-Iraty Cheese: Determination of Free Amino Acids by RP-HPLC and of Total Free Amino Acids by the TNBS Method. Food Control 2000, doi:10.1016/S0956-7135(99)00031-6.
27. Sharma, S.K.; Sehgal, N.; Kumar, A. Biomolecules for Development of Biosensors and Their Applications. Curr. Appl. Phys. 2003.
28. Kumar, K.; Kataria, M.; Verma, N. Plant Asparaginase-Based Asparagine Biosensor for Leukemia. Artif. Cells, Nanomedicine Biotechnol. 2013, doi:10.3109/10731199.2012.716062.
29. Shrivastava, A.; Khan, A.A.; Khurshid, M.; Kalam, M.A.; Jain, S.K.; Singhal, P.K. Recent Developments in L-Asparaginase Discovery and Its Potential as Anticancer Agent. Crit. Rev. Oncol. Hematol. 2016, 100.
30. El-Naggar, N.E.A.; El-Ewasy, S.M.; El-Shweihy, N.M. Microbial L-Asparaginase as a Potential Therapeutic Agent for the Treatment of Acute Lymphoblastic Leukemia: The Pros and Cons. Int. J. Pharmacol. 2014, doi:10.3923/ijp.2014.182.199.
31. Lopes, A.M.; Oliveira-Nascimento, L. de; Ribeiro, A.; Tairum, C.A.; Breyer, C.A.; Oliveira, M.A. de; Monteiro, G.; Souza-Motta, C.M. de; Magalhães, P. de O.; Avendaño, J.G.F.; et al. Therapeutic L-Asparaginase: Upstream, Downstream and Beyond. Crit. Rev. Biotechnol. 2017, 37.
32. Goodsell DS The Molecular Perspective: L-Asparaginase. Oncologist 2005, 10, 238-239.
33. Nunes, J.C.F.; Cristóvão, R.O.; Freire, M.G.; Santos-Ebinuma, V.C.; Faria, J.L.; Silva, C.G.; Tavares, A.P.M. Recent Strategies and Applications for L-Asparaginase Confinement. Molecules 2020, 25.
34. Varshosaz, J.; Anvari, N. Enhanced Stability of L-Asparaginase by Its Bioconjugation to Poly(Styrene-Co-Maleic Acid) and Ecoflex Nanoparticles. IET Nanobiotechnology 2018, 12, doi:10.1049/iet-nbt.2017.0156.
35. Dias, F.F.G.; Santos Aguilar, J.G. dos; Sato, H.H. L-Asparaginase from Aspergillus Spp.: Production Based on Kinetics, Thermal Stability and Biochemical Characterization. 3 Biotech 2019, 9, doi:10.1007/s13205-019-1814-5.
36. Dias, F.F.G.; Ruiz, A.L.T.G.; Torre, A. Della; Sato, H.H. Purification, Characterization and Antiproliferative Activity of L-Asparaginase from Aspergillus Oryzae CCT 3940 with No Glutaminase Activity. Asian Pac. J. Trop. Biomed. 2016, 6, doi:10.1016/j.apjtb.2016.07.007.
37. Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Reduction of Acrylamide Level through Blanching with Treatment by an Extremely Thermostable L-Asparaginase during French Fries Processing. Extremophiles 2015, doi:10.1007/s00792-015-0763-0.
38. Li, X.; Zhang, X.; Xu, S.; Zhang, H.; Xu, M.; Yang, T.; Wang, L.; Qian, H.; Zhang, H.; Fang, H.; et al. Simultaneous Cell Disruption and Semi-Quantitative Activity Assays for High-Throughput Screening of Thermostable L-Asparaginases. Sci. Rep. 2018, doi:10.1038/s41598-018-26241-7.
39. Bansal, S.; Srivastava, A.; Mukherjee, G.; Pandey, R.; Verma, A.K.; Mishra, P.; Kundu, B. Hyperthermophilic Asparaginase Mutants with Enhanced Substrate Affinity and Antineoplastic Activity: Structural Insights on Their Mechanism of Action. FASEB J. 2012, doi:10.1096/fj.11-191254.
40. Chohan, S.M.; Rashid, N. TK1656, a Thermostable l-Asparaginase from Thermococcus Kodakaraensis, Exhibiting Highest Ever Reported Enzyme Activity. J. Biosci. Bioeng. 2013, doi:10.1016/j.jbiosc.2013.04.005.
41. Hatanaka, T.; Usuki, H.; Arima, J.; Uesugi, Y.; Yamamoto, Y.; Kumagai, Y.; Yamasato, A.; Mukaihara, T. Extracellular Production and Characterization of Two Streptomyces L-Asparaginases. Appl. Biochem. Biotechnol. 2011, doi:10.1007/s12010-010-9087-9.
42. Bansal, S.; Gnaneswari, D.; Mishra, P.; Kundu, B. Structural Stability and Functional Analysis of L-Asparaginase from Pyrococcus Furiosus. Biochem. 2010, doi:10.1134/s0006297910030144.
43. Guo; Coker; Wood; Cooper; Chohan; Rashidc; Akhtar Structure and Function of the Thermostable L-Asparaginase from Thermococcus Kodakarensis. Acta Cryst. ( 2017, 889-895.
44. Hong, S.J.; Lee, Y.H.; Khan, A.R.; Ullah, I.; Lee, C.; Park, C.K.; Shin, J.H. Cloning, Expression, and Characterization of Thermophilic L-Asparaginase from Thermococcus Kodakarensis KOD1. J. Basic Microbiol. 2014, doi:10.1002/jobm.201300741.
45. Zuo, S.; Xue, D.; Zhang, T.; Jiang, B.; Mu, W. Biochemical Characterization of an Extremely Thermostable L-Asparaginase from Thermococcus Gammatolerans EJ3. J. Mol. Catal. B Enzym. 2014, doi:10.1016/j.molcatb.2014.08.021.
46. Dumina, M.; Zhgun, A.; Pokrovskaya, M.; Aleksandrova, S.; Zhdanov, D.; Sokolov, N.; El’darov, M. A Novel L-Asparaginase from Hyperthermophilic Archaeon Thermococcus Sibiricus: Heterologous Expression and Characterization for Biotechnology Application. Int. J. Mol. Sci. 2021, 22, 9894, doi:10.3390/ijms22189894.
47. Tomoko Matsui; Esben Peter Friis; Akihiko Yamagishi Thermostable Asparaginases 2008.
48. Tomoko Matsui; Aki Tomiki; Allan Svendsen; Hanne Vang Hendriksen; Mary Ann Stringer; Keiichi Ayabe Thermostable Asparaginase Variants and Polynucleotides Encoding Same.
49. Novel Bacillus Methylotrophicus MKSY2013 Producing Enhanced Thermostable Asparaginase and Its Producing Method 2014.
50. Zhiming, R.; Xu, L.; Xian, Z.; Shuqin, X.; Jingyi, H.; Meijuan, X.; Taowei, Y. Thermophilic L-Asparaginase Mutant and Screening and Fermentation Methods Thereof 2019.
51. National Library of Medicine Thermophilic L-Asparaginase. Https://Pubchem.Ncbi.Nlm.Nih.Gov/#query=thermophilic%20l-Asparaginase&tab=patent.
52. Bishwajit, K.; Saurabh, B.; Prashant, M. Mutants of L-Asparaginase 2011.
53. SHIN, J.-H.; HONG, S.J.; LEE, Y.H. Composition for Breaking down L-Asparagine Comprising l-Asparaginase, and Production Method for l-Asparaginase 2012.
54. Li, X.; Zhang, X.; Xu, S.; Xu, M.; Yang, T.; Wang, L.; Zhang, H.; Fang, H.; Osire, T.; Rao, Z. Insight into the Thermostability of Thermophilic L-Asparaginase and Non-Thermophilic L-Asparaginase II through Bioinformatics and Structural Analysis. Appl. Microbiol. Biotechnol. 2019, doi:10.1007/s00253-019-09967-w.
55. Vennison. J Laboratory Manual for Genetic Engineering.; 2010;
56. Dumina, M.; Zhgun, A. Thermo-L-Asparaginases: From the Role in the Viability of Thermophiles and Hyperthermophiles at High Temperatures to a Molecular Understanding of Their Thermoactivity and Thermostability. Int. J. Mol. Sci. 2023, 24, 2674, doi:10.3390/ijms24032674.
57. Mardanov, A. V.; Ravin, N. V.; Svetlitchnyi, V.A.; Beletsky, A. V.; Miroshnichenko, M.L.; Bonch-Osmolovskaya, E.A.; Skryabin, K.G. Metabolic Versatility and Indigenous Origin of the Archaeon Thermococcus Sibiricus, Isolated from a Siberian Oil Reservoir, as Revealed by Genome Analysis. Appl. Environ. Microbiol. 2009, 75, doi:10.1128/AEM.00718-09.
58. Mardanov,A.V., Ravin,N.V., Svetlitchnyi,V.A., Beletsky,A.V., Miroshnichenko,M.L., Bonch-Osmolovskaya,E.A. and Skryabin, K.G. Thermococcus Sibiricus MM 739, Complete Genome Available online: https://www.ncbi.nlm.nih.gov/nuccore/NC_012883.1.
59. Swain, A.L.; Jaskolski, M.; Housset, D.; Rao, J.K.; Wlodawer, A. Crystal Structure of Escherichia Coli L-Asparaginase, an Enzyme Used in Cancer Therapy. Proc. Natl. Acad. Sci. 2006, doi:10.1073/pnas.90.4.1474.
60. Yao, M.; Yasutake, Y.; Morita, H.; Tanaka, I. Structure of the Type I L-Asparaginase from the Hyperthermophilic Archaeon Pyrococcus Horikoshii at 2.16 Å Resolution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2005, doi:10.1107/S0907444904032950.
61. Miroshnichenko, M.L.; Hippe, H.; Stackebrandt, E.; Kostrikina, N.A.; Chernyh, N.A.; Jeanthon, C.; Nazina, T.N.; Belyaev, S.S.; Bonch-Osmolovskaya, E.A. Isolation and Characterization of Thermococcus Sibiricus Sp. Nov. from a Western Siberia High-Temperature Oil Reservoir. Extremophiles 2001, 5, doi:10.1007/s007920100175.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="СПОСОБ УЛУЧШЕНИЯ
КАТАЛИТИЧЕСКИХ СВОЙСТВ ГИПЕРТЕРМОФИЛЬНЫХ L-АСПАРАГИНАЗ I ТИПА,
L-АСПАРАГИНАЗА I ТИПА ИЗ ГИПЕРТЕРМОФИЛЬНОЙ АРХЕИ THERMOCOCCUS
SIBIRICUS, ПОЛУЧЕННАЯ ТАКИМ СПОСОБОМ2.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2023-12-11">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023130911/20(068635)</ApplicationNumberText>
<FilingDate>2023-11-27</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>HH</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023130911/20(068635)</ApplicationNumberText>
<FilingDate>2023-11-27</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ФИЦ Биотехнологии
РАН</ApplicantName>
<ApplicantNameLatin>Research Center of Biotechnology
RAS</ApplicantNameLatin>
<InventorName languageCode="ru">Мария Владимировна
Думина</InventorName>
<InventorNameLatin>Maria V. Dumina</InventorNameLatin>
<InventionTitle languageCode="ru">СПОСОБ УЛУЧШЕНИЯ КАТАЛИТИЧЕСКИХ
СВОЙСТВ ГИПЕРТЕРМОФИЛЬНЫХ L-АСПАРАГИНАЗ I ТИПА, L-АСПАРАГИНАЗА I ТИПА
ИЗ ГИПЕРТЕРМОФИЛЬНОЙ АРХЕИ THERMOCOCCUS SIBIRICUS, ПОЛУЧЕННАЯ ТАКИМ
СПОСОБОМ</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>331</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..331</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Thermococcus
sibiricus</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MKKLLIIGTGGTIASAKTEQGYKSVLKIDEILKLAKIKLENGYKIDSTN
IMNIDSTLIHPEDWEIIAKEVFKALDDYDGIIITHGTDTLAYTASMLSFMIKNPNKPIVLTGSMLPITEN
GSDAPRNIRTAIKFAMEDVAGVFVAFMDKIMLGCRTSKVRTLGLNAFMSINYPDVAYVKGEKILYNIPKE
KFQPNGSPELDTKYEPRVVVLRVTPGLGGEIIDAVLDAGYKGIVLEGYGAGGLPYRKSNLLSKIKEITPK
IPVIMTTQALYDGVDMRKYEVGRKALETGIIPAKDMTKEATITKLMWALGHTKDVEKIREIMHTNYVNEI
KS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>996</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..996</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q11">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgaagaagttactgattattggcactggcggcacaatcgcttcggcaa
agacggagcagggttataagagcgtgctgaagatcgacgagattctcaagcttgcaaagatcaagcttga
gaacggctacaagattgactccacgaacatcatgaatattggttcgcagttgattcatccagaggattgg
gagatcatcgcgaaggaagtctttaaagcactcgacgactacgacggtatcattattactcacgggaccg
ataccttagcgtatactgcatctatgttgtcttttatgattaagaatccgaataagcctatcgtgttgac
cgggagcatgctcccgattaccgagaacggctcagacgctccgcgtaatattcgtaccgctatcaagttc
gccatggaagacgttgccggcgtgttcgtggcgtttatggacaagattatgttgggctgccgtacgagca
aagtgcgcactttggggcttaacgccttcatgtctatcaactatccggacgtcgcctacgtgaagggcga
gaaaattctgtataacattccaaaggagaagtttcagccgaacggctctccagaactggacaccaaatac
gagccgcgcgtggtagtactgcgtgtaactccgggactgggtggggaaattattgacgctgtgctggacg
cgggttacaagggtattgttttggaaggttacggcgccggcgggctgccgtaccgtaaaagcaacctgct
ctctaagatcaaagagatcactccgaagattcccgttatcatgacgacccaggcgctgtacgacggagtg
gatatgcgtaagtacgaggtgggccgcaaagctcttgagaccggtatcattccggcgaaggatatgacca
aagaggctaccattacgaaactgatgtgggcgctggggcataccaaggatgtggagaaaattcgtgagat
tatgcacaccaattatgtgaacgagattaagagttga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>996</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..996</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q12">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgaagaagttactgattattggcactggcggcacaatcgcttcggcaa
agacggagcagggttataagagcgtgctgaagatcgacgagattctcaagcttgcaaagatcaagcttga
gaacggctacaagattgactccacgaacatcatgaatattgattcgaccttgattcatccagaggattgg
gagatcatcgcgaaggaagtctttaaagcactcgacgactacgacggtatcattattactcacgggaccg
ataccttagcgtatactgcatctatgttgtcttttatgattaagaatccgaataagcctatcgtgttgac
cgggagcatgctcccgattaccgagaacggctcagacgctccgcgtaatattcgtaccgctatcaagttc
gccatggaagacgttgccggcgtgttcgtggcgtttatggacaagattatgttgggctgccgtacgagca
aagtgcgcactttggggcttaacgccttcatgtctatcaactatccggacgtcgcctacgtgaagggcga
gaaaattctgtataacattccaaaggagaagtttcagccgaacggctctccagaactggacaccaaatac
gagccgcgcgtggtagtactgcgtgtaactccgggactgggtggggaaattattgacgctgtgctggacg
cgggttacaagggtattgttttggaaggttacggcgccggcgggctgccgtaccgtaaaagcaacctgct
ctctaagatcaaagagatcactccgaagattcccgttatcatgacgacccaggcgctgtacgacggagtg
gatatgcgtaagtacgaggtgggccgcaaagctcttgagaccggtatcattccggcgaaggatatgacca
aagaggctaccattacgaaactgatgtgggcgctggggcataccaaggatgtggagaaaattcgtgagat
tatgcacaccaattatgtgaacgagattaagagttga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>331</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..331</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Thermococcus
sibiricus</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MKKLLIIGTGGTIASAKTEQGYKSVLKIDEILKLAKIKLENGYKIDSTN
IMNIGSQLIHPEDWEIIAKEVFKALDDYDGIIITHGTDTLAYTASMLSFMIKNPNKPIVLTGSMLPITEN
GSDAPRNIRTAIKFAMEDVAGVFVAFMDKIMLGCRTSKVRTLGLNAFMSINYPDVAYVKGEKILYNIPKE
KFQPNGSPELDTKYEPRVVVLRVTPGLGGEIIDAVLDAGYKGIVLEGYGAGGLPYRKSNLLSKIKEITPK
IPVIMTTQALYDGVDMRKYEVGRKALETGIIPAKDMTKEATITKLMWALGHTKDVEKIREIMHTNYVNEI
KS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ГЕН tsA, КОДИРУЮЩИЙ ТЕРМОСТАБИЛЬНУЮ L-АСПАРАГИНАЗУ ИЗ АРХЕИ THERMOCOCCUS SIBIRICUS, РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ L-АСПАРАГИНАЗЫ И СПОСОБ ОЧИСТКИ ЭТОГО ФЕРМЕНТА | 2022 |
|
RU2794225C1 |
| ШТАММ-ПРОДУЦЕНТ РЕКОМБИНАНТНОЙ ТЕРМОСТАБИЛЬНОЙ L-АСПАРАГИНАЗЫ ИЗ MELIORIBACTER ROSEUS | 2022 |
|
RU2831613C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pHisTevTSIB0821, ТРАНСФОРМИРОВАННЫЙ ЕЮ ШТАММ Escherichia coli Rosetta(DE3)/pHisTevTSIB0821 И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ПРОЛИДАЗЫ TSIB_0821 | 2012 |
|
RU2509154C1 |
| МУТАНТНАЯ РЕКОМБИНАНТНАЯ L-АСПАРАГИНАЗА Wolinella succinogenes (ВАРИАНТЫ) | 2014 |
|
RU2545722C1 |
| СПОСОБ ВЫЯВЛЕНИЯ ШТАММОВ БАКТЕРИЙ-ПРОДУЦЕНТОВ L-АСПАРАГИНАЗЫ | 2008 |
|
RU2398876C2 |
| ТЕРМОСТАБИЛЬНАЯ АЛКОГОЛЬДЕГИДРОГЕНАЗА ИЗ АРХЕИ THERMOCOCCUS SIBIRICUS | 2009 |
|
RU2413766C1 |
| АСПАРАГИНАЗА, ПОЛУЧЕННАЯ ИЗ БАЗИДОМИЦЕТОВ | 2011 |
|
RU2560597C2 |
| ГИБРИДНЫЙ БЕЛОК НА ОСНОВЕ L-АСПАРАГИНАЗЫ Wolinella succinogenes, ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОГО БЕЛКА, ОБЛАДАЮЩЕГО ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2562166C1 |
| НОВАЯ ГИДРОГЕНАЗА SEQ ID NO:5, ОЧИЩЕННАЯ ИЗ THERMOCOCCUS ONNURIENUS NA1 С ПОМОЩЬЮ МОНОOKCИДА УГЛЕРОДА, КОДИРУЮЩИЕ ЕЕ ГЕНЫ, И СПОСОБЫ ПОЛУЧЕНИЯ ВОДОРОДА С ИСПОЛЬЗОВАНИЕМ МИКРООРГАНИЗМА, ИМЕЮЩЕГО УКАЗАННЫЕ ГЕНЫ | 2009 |
|
RU2460789C2 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОЙ БЕТА-ГАЛАКТОЗИДАЗЫ AsBgl 1390 ИЗ АРХЕИ SACCHAROVORANS В КАЧЕСТВЕ БЕТА-ГЛЮКОЗИДАЗЫ, БЕТА-КСИЛОЗИДАЗЫ И БЕТА-МАННОЗИДАЗЫ | 2012 |
|
RU2523581C2 |
Изобретение относится к области инженерной энзимологии, молекулярной биологии и биотехнологии. Предложен способ получения модифицированной гипертермофильной L-аспарагиназы I типа из Thermococcus sibiricus, имеющей SEQ ID NO: 4, в которой содержатся мутации D54G и T56Q. Способ включает изменение структуры исходной L-аспарагиназы I типа из Thermococcus sibiricus с SEQ ID NO: 1 путем замены аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q. Также предложена модифицированная гипертермофильная L-аспарагиназа I типа из Thermococcus sibiricus, полученная указанным способом и имеющая SEQ ID NO: 4. Изобретение обеспечивает получение гипертермофильной L-аспарагиназы I типа из Thermococcus sibiricus с улучшенными каталитическими свойствами. 2 н.п. ф-лы, 6 ил., 1 табл., 5 пр.
1. Способ получения модифицированной гипертермофильной L-аспарагиназы I типа из Thermococcus sibiricus, имеющей SEQ ID NO: 4, в которой содержатся мутации D54G и T56Q, включающий изменение структуры исходной L-аспарагиназы I типа из Thermococcus sibiricus с SEQ ID NO: 1 путем замены аминокислотных остатков в 1 и 3 положении 4-членного высококонсервативного мотива DSTL, где в 1 положении произведена замена D/G, в третьем положении T/Q.
2. Модифицированная гипертермофильная L-аспарагиназа I типа, полученная по п. 1, характеризующаяся тем, что источником исходного фермента является Thermococcus sibiricus, соответствующая аминокислотной последовательности SEQ ID NO: 4, в которой выбранные замены в 1 и 3 положении 4-членного высококонсервативного мотива DSTL соответствуют мутациям D54G и T56Q.
| МОДИФИЦИРОВАННЫЙ ГЕН tsA, КОДИРУЮЩИЙ ТЕРМОСТАБИЛЬНУЮ L-АСПАРАГИНАЗУ ИЗ АРХЕИ THERMOCOCCUS SIBIRICUS, РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ L-АСПАРАГИНАЗЫ И СПОСОБ ОЧИСТКИ ЭТОГО ФЕРМЕНТА | 2022 |
|
RU2794225C1 |
| CHOHAN S.M, RASHID N | |||
| Регулирующее приспособление для ветряных двигателей со складными перьями или лопастями | 1924 |
|
SU1656A1 |
| J Biosci Bioeng | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| ZUO S | |||
| et al | |||
| Biochemical characterization of an extremely thermostable l-asparaginase from | |||

