Изобретение относится к области органической химии и биотехнологии и может быть использовано для разработки биокатализаторов для биотехнологических процессов получения новых функциональных соединений. Описываемый фермент (трансаминаза из бактерии Haliscomenobacter hydrossis) состоит из стандартных аминокислот и содержит точечные аминокислотные замены относительно природной формы данного фермента, расширяющие его субстратную специфичность, что важно для разработки биокатализаторов стереоселективного аминирования органических соединений на основе природных ферментов-трансаминаз.
Введение
Трансаминирование - жизненно важный метаболический процесс, имеющийся у всех живых организмов и определяющий уровень и состав протеиногенных и непротеиногенных аминокислот в клетке (1). Ферментативное трансаминирование катализируется специализированными ферментами - пиридоксаль-5-фосфат (PLP) зависимыми трансаминазами (ТА), которые стереоселективно переносят аминогруппу от аминодонора (амина или аминокислоты) на кетосоединение (кетон, кетокислота или альдегид) с образованием новых амино- и кетопродуктов (1-3).
Биокатализаторы широко используются в настоящее время в промышленности для проведения химических превращений с высокой хемо-, регио- и стерео-селективностью (4). Ферментативное трансаминирование, в частности, является эффективным биотехнологическим методом получения оптически чистых аминов благодаря высокой энантиоселективности трансаминаз, применение которых обеспечивает получение оптически чистого продукта с энантиомерным избытком 99% и более (5-7). Успешным примером практического применения трансаминаз являются биотехнологический синтез антидиабетического препарата ситаглиптина (8,9) и препарата сакубитрила, улучшающего состояние сердечно-сосудистой системы (10). Стратегия асимметрического аминирования также была применена компаниями Merck и Pfizer в производственных циклах получения следующих фармацевтических препаратов: суворексанта (снотворный препарат) (11), гласдегиба (PF-04449913, препарат для лечения миелоидного лейкоза) (12), MK-7246 (препарат для лечения органов дыхания) (13), вернакалант (препарат для лечения аритмии сердца) (14). Во всех случаях в качестве биокатализаторов использовали модифицированные трансаминазы: (S)-селективная из Vibrio fluvialis или (R)-селективная из Arthrobacter sp. Инженерия ферментов включала классическую направленную эволюцию, компьютерный анализ и моделирование комплексов ферментов с субстратами, а также сайт-направленный мутагенез.
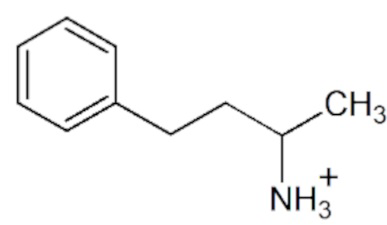
Аминосоединения находят свое применения в разных областях как добавки против коррозии металлов (15), компоненты косметических и парфюмерных средств (16), различных красителей (17) и др. Амины являются структурными блоками многих синтетических лекарственных средств (лиздексамфетамин (психостимулятор) (18), лабеталол (лекарство для лечения сердечных заболеваний) (19), репаглиндин (противодиабетический препарат) (20), ривастигмин (препарат, замедляющий развитие деменции) (21), долутегравир (препарат против ВИЧ) (22) и др.) и препаратов для сельского хозяйства (дифлуметорим, ципродинил (противогрибковые препараты) (23), продиамин, биалафос (гербициды) (24,25) и др.) (Рисунок 1). Для эффективного применения синтетических препаратов особенно важна хиральная чистота целевого энантиомера (26-28). Эффективное получение оптически чистых аминосоединений, таким образом, является одной из ключевых задач в области органического синтеза.
Ферментативное трансаминазы безусловно доказало свою эффективность в асимметрическом аминировании. Однако имеется ряд ограничений применения природных трансаминаз. Помимо нестабильности ферментов как таковых в промышленных условиях, ферменты трансаминазы отличаются узкой субстратной специфичностью, их активность ингибируется избытком субстрата или продукта, кроме того ферментативная реакция трансаминирования обратима (термодинамическая константа равновесия реакции трансаминирования - около единицы) (7,29-35). Для эффективного применения трансаминаз требуются разнообразные подходы к изменению их субстратной специфичности при сохранении стереоселективности.
Задачи заявляемого изобретения
Технической задачей настоящего изобретения является расширение субстратной специфичности природного фермента трансаминазы. Поставленная задача решается путем получения и биохимической характеристики генно-инженерного варианта трансаминазы из бактерии H. hydrossis с измененной субстратной специфичностью. Для этой цели была изменена нуклеотидная последовательность, кодирующая трансаминазу D-аминокислот из H. Hydrossis. Изменение привело к замене одного аминокислотного остатка в активном центре фермента и получению варианта трансаминазы с расширенной субстратной специфичностью, активной не только с аминокислотами и кетокислотами, но и с первичными (R)-аминами.
Технический результат изобретения заключается в повышении биотехнологического потенциала фермента путем расширения спектра его субстратов.
Задачи решены путем
Для модификации субстратной специфичности трансаминазы D-аминокислот из H. hydrossis был проведен точечный мутагенез природного фермента путем замены в нуклеотидной последовательности кодона, кодирующего аминокислотный остаток R90, на кодон, кодирующий остаток изолейцина (SEQ ID: 1).
Рекомбинантная форма нового варианта фермента была наработана путем гетерологической экспрессии в клетках E. coli с последующей очисткой целевого фермента до гомогенного состояния. Наличие замененного аминокислотного остатка I90 в активном центре фермента подтверждено методом масс-спектрометрии, а нативность структуры нового белка - методом рентгеноструктурного анализа. Новый вариант фермента был охарактеризован путем измерения его активности в реакциях с субстратами D-аминокислотами и первичными (R)-аминами (см. далее).
Создание варианта R90I трансаминазы D-аминокислот из H. hydrossis (HalhyTA)
Замену R90I в аминокислотную последовательности HalhyTA вводили методом сайт-направленного мутагенеза с помощью ПЦР с одним праймером (Таблица 1) на амплификаторе T100 Thermal Cycler (Bio-Rad Laboratories, США) по методике, описанной в работах (36,37). Реакционная смесь объемом 25 мкл содержала 5-10 нг/мкл ДНК-матрицы (плазмидная ДНК, содержащая ген фермента), 0,8 мкМ праймера, несущего нуклеотидную замену, а также ДНК-полимеразу, смесь дезоксинуклеотидов и буфер, взятые из специального набора Encyclo Plus PCR kit (Евроген, Россия) в концентрациях, рекомендуемых производителем. ПЦР проводили в следующем режиме: 1-я стадия - 95°С, 30 с; 2-я стадия - 55°С, 30 с; 3-я стадия - 72°С - 6,5 мин, всего 18 циклов. В конце смесь выдерживали еще 7 минут при 72°С.
Таблица 1. Олигонуклеотидные последовательности праймеров для введение замены R90I в последовательность HalhyTA, а также для отбора клонов, несущих соответствующую замену.
Для поиска колоний клеток E. coli, несущих плазмиду с нужной мутацией, проводил проверочный ПЦР с двумя праймерами на колониях. Первым праймером (Таблица 1 Check-праймер) была олигонуклеотидная последовательность длиной 18-21 нуклеотидов, комплементарная участку гена с введенной нуклеотидной заменой. При этом на 3’-конце праймера были 1-3 нуклеотида, комплементарные нуклеотидной замене и не комплементарные соответствующему участку в гене WT. Если нуклеотидная замена находилась ближе к 5’-концу гена, то Check-праймер конструировали в прямом направлении, если ближе к 3’-концу, то в обратном. Вторым праймером использовали стандартный Т7-праймер в прямом или обратном направлении, в зависимости от направления Check-праймера. Таким образом, в присутствие плазмидной ДНК, несущей нуклеотидную замену, в ходе ПЦР накапливался фрагмент ДНК определенной длины. Реакционная смесь объемом 25 мкл содержала два праймера в концентрациях по 0,8 мкМ, а также Taq ДНК-полимеразу, буфер и смесь дезоксинуклеотидов, взятых из набора TaqDNA Polymerase dNTP mix (Евроген, Россия) в концентрациях, рекомендуемых производителем. В пробирки с приготовленной ПЦР-смесью инокулировали одиночные колонии с чашки Петри и проводили ПЦР в следующем режиме: 1-я стадия - 95°С, 20 с; 2-я стадия - 49°С, 20 с; 3-я стадия - 72°С - 50 с, всего 25 циклов. В конце смесь выдерживали еще 5 минут при 72°С. В качестве отрицательного контроля использовали плазмидную ДНК-матрицу, несущую ген WT, в качестве положительного контроля - реакционной смесь после ПЦР-мутагенеза. По окончанию проверочного ПЦР пробы анализировали с помощью ДНК-электрофореза.
После мутагенезного ПЦР в реакционную смесь добавляли ДНКазу Dpn (0,1 Ед/мкл) и инкубировали ее при 37°С в течение 2 часов, далее этой смесью трансформировали клетки E. coli Mach1, которые после растили на чашках Петри. Выросшие одиночные колонии проверяли на наличие плазмид, несущих нуклеотидную последовательность варианта R90I HalhyTA, методом проверочного ПЦР с колоний. Смеси проверочного ПЦР анализировали с помощью ДНК-электрофореза, отбирали по 2 колонии, несущих целевую нуклеотидную замену, выделяли из них плазмидную ДНК и проверяли ее последовательность методом секвенирования ДНК. После клетки E. coli Rosetta (DE3) pLysS трансформировали плазмидной ДНК pET-21d-HisTEV, несущей целевой ген с заменой.
Препаративная экспрессия гена варианта R90I HalhyTA
Гены, кодирующие аминокислотную последовательность варианта R90I HalhyTA экспрессировали в клетках E. coli Rosetta (DE3) pLysS. Для препаративной экспрессии сначала получали 30 мл ночной культуры клеток, которую затем разбавляли в 100 раз (до 3 л) в среде LB в присутствие 20 мкг/мл ампициллина и 30 мкг/мл хлорамфеникола и разливали по 250 мл в 12 колб объемом 1 л. Клетки растили на качалке (180 об/мин) при 37°С до значения оптической плотности при 600 нм 0,6-0,8, затем охлаждали до 25°С, индуцировали экспрессию 0,2 мМ ИПТГ и инкубировали клетки около 18 часов на качалке (180 об/мин) при 25°С. После клетки центрифугировали, удаляли супернатант и для хранения замораживали при -70°С.
Выделение и очистка рекомбинантной формы варианта R90I HalhyTA
Составы буферов, используемых на стадиях выделения и очистки рекомбинантных форм ферментов, приведены в Таблице 2. Размороженные клетки ресуспендировали в буфере 1, охлаждали во льду, и обрабатывали ультразвуком в течение 10-15 минут на приборе Vibra-Cell VCX130 (Sonic and materials, США) в следующем режиме: импульс - 5 с, покой - 10 с, амплитуда - 65%. Затем добавляли 0,005 мг/мл ДНКазы (Sigma, США) и откручивали разрушенные клетки в течение 45 минут при 18,500 об/мин и 4°С. Далее супернатант фильтровали через 0,45 мкм фильтр (Millipore, США) и наносили на 5 мл колонку HisTrap HP (Cytiva, США), уравновешенную буфером 2 (здесь и далее скорость потока на колонке HisTrap HP была 1 мл/мин). Адсорбировавшийся на колонке фермент с (His)6-тагом затем промывали буфером 3 и элюировали линейным градиентом от буфера 3 к буферу 4, пропустив 10 объемов колонки буфера 4. В случае трансаминаз после элюции белок инкубировали с 1 мМ PLP в течение 1 часа при 25°С, далее ферменты концентрировали на концентраторах с мембраной, размер пор которой составлял 30 кДа, (Millipore, США), переводили в буфер 5, разбавляли глицерином в два раза и хранили при -20°С. Вариант с (His)6-тагом использовали для проведения функциональной характеристики варианта.
Таблица 2. Составы буферов, используемых на стадиях выделения и очистки рекомбинантной формы варианта.
Методы определения активности и pH и температурного оптимума реакций, катализируемых вариантом R90I HalhyTA.
Активность варианта R90I HalhyTA определяли несколькими альтернативными методами:
ЛДГ метод:
Скорость реакций трансаминирования между D-аланином и α-кетоглутаратом определяли по второй ферментативной реакции с NAD-зависимыми лактатдегидрогеназой (ЛДГ метод) из мышц кролика (Roche, Швейцария) и рекомбинантной (R)-2-гидроксиглутаратдегидрогеназой (ГГДГ метод) из A. fermentas (см. 2.4.3. и 2.4.4.), соответственно. Скорость реакций регистрировали спектрофотометрически при 340 нм по убыли кофактора NADH (ε(NADH) = 6220 М-1 см-1) в ходе восстановления продукта реакции трансаминирования пирувата в случае ЛДГ метода или α-кетоглутарата в случае ГГДГ метода на планшетном спектрофотометре SPECTROstar Omega в 96-лучночных УФ-планшетах (Greiner Bio-One, Германия). Анализ активности проводили в присутствии 5-500 мМ D-аланина и 0,5-20 мМ α-кетоглутарата или 1-30 мМ D-глутамата и 10-500 мМ пирувата, 0,01-1 мкМ WT AmicoTA или варианта, 60 мкМ PLP, 330 мкМ NADH и 4 U/мл ЛДГ или ГГДГ, соответственно. Реакцию запускали добавлением D-аминокислоты после прогрева реакционной смеси в течение 10 минут. Стандартный анализ активности AmicoTA и вариантов проводили в 50 мМ CHES буфере, pH 9,0, при 60°С с 50 мМ D-аланином и 5 мМ α-кетоглутаратом. Ферментативную активность определяли по тангенсу угла наклона начального линейного участка кинетической кривой. Одна единица ферментативной активности (U) соответствует количеству фермента, который катализирует превращение 1 мкмоля субстрата в продукт за минуту.
Кинетические параметры полных реакций трансаминирования: Vmax (максимальная скорость), 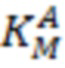 , (константа Михаэлиса субстрата А) и
, (константа Михаэлиса субстрата А) и 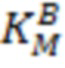 (константа Михаэлиса субстрата В) определяли из аппроксимации зависимости наблюдаемой скорости реакции (V) от концентрации субстрата при фиксированной концентрации косубстрата уравнением (3):
(константа Михаэлиса субстрата В) определяли из аппроксимации зависимости наблюдаемой скорости реакции (V) от концентрации субстрата при фиксированной концентрации косубстрата уравнением (3):
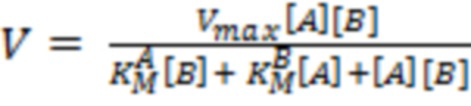 , (3)
, (3)
Ацетофеноновый метод:
Активность варианта в реакции с (R)-1-фенилэтиламином (R-PEA) измеряли спектрофотометрически по скорости изменения поглощения при длине волны 245 нм, соответствующей поглощению продукта деаминирования R-PEA - ацетофенона. Реакционная смесь сдержала: 50 мМ CHES буфера, pH 9.0, 50 мМ NaCl, 30 мкМ PLP, 1-10 мМ R-PEA и 5 мМ a-кетоглутарата. Смесь инкубировали в течение 3 мин при температуре 25°C. Реакцию запускали добавлением 0,1 мг/мл фермента. Удельную активность рассчитывали из начального линейного участка зависимости накопления ацетофенона от времени, используя молярный коэффициент экстинкции ацетофенона, 11600 М-1 см-1. Один U активности определяли как количество фермента, необходимого для образования 1 мкмоль ацетофенона в минуту.
Метод полуреакций трансаминирования:
Скорость полуреакции трансаминирования варианта R90I HalhyTA с D-аминокислотами и (R)-аминами регистрировали спектрофотометрически по убыли оптической плотности в области 407-415 нм. Полуреакции анализировали с 35 мкМ PLP-формы варианта и 0,5-500 мМ субстрата. Так как субстрат находится в значительном избытке по отношению к ферменту, каждую полуреакцию рассматривали как реакцию первого порядка и константу скорости полуреакции kobs определяли из аппроксимации зависимости поглощения А407-415 от времени уравнением (1):
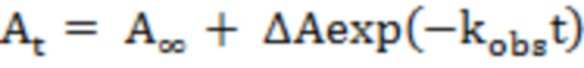 , (1)
, (1)
где 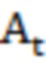 - поглощение во время t,
- поглощение во время t, 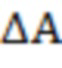 - изменение поглощения за период t = 0 и t = ∞, A∞ - конечное поглощение и kobs - наблюдаемая константа скорости первого порядка.
- изменение поглощения за период t = 0 и t = ∞, A∞ - конечное поглощение и kobs - наблюдаемая константа скорости первого порядка.
Кинетические параметры полуреакций трансаминирования: 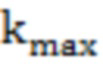 (максимальная константа скорости),
(максимальная константа скорости), 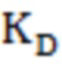 (константа диссоциации фермент-субстратного комплекса),
(константа диссоциации фермент-субстратного комплекса),  (константа специфичности) и
(константа специфичности) и 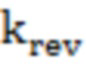 (константа скорости обратной реакции) определяли из аппроксимации зависимости наблюдаемой константы скорости полуреакции (kobs) от концентрации субстратов уравнением (2):
(константа скорости обратной реакции) определяли из аппроксимации зависимости наблюдаемой константы скорости полуреакции (kobs) от концентрации субстратов уравнением (2):
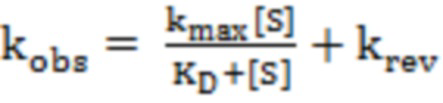 , (2)
, (2)
Определение рН- и температурных профилей:
Влияние рН и температуры на активность варианта анализировали в полной реакции трансаминирования между 50 мМ D-аланином и 5 мМ α-кетоглутаратом ЛДГ методом или между 5 мМ (R)-PEA и 5 мМ α-кетоглутарата. рН-оптимум реакций определяли при 25°С в следующих буферах: 50 мМ Na-ацетат, рН 4,0-5,6, 25 мМ Трис-HCl и 25 мМ К-фосфат, рН 5,8-9,0, 50 мМ CHES, pH 9,0-10,0. Температурную зависимость определяли в диапазоне 30-70°С в 50 мМ K-фосфатном буфере, рН 8,0 или 50 мМ Na-ацетатном буфере, рН 5,2. Для измерения ферментативной активности при 65 и 70°С из реакционной смеси отбирали аликвоты через определенные промежутки времени и замораживали их. Затем определяли концентрацию образовавшегося пирувата в аликвоте с помощью ЛДГ при 25°С в 100 мМ K-фосфатном буфере, рН 7,0, используя линейный калибровочный график - зависимость активности ЛДГ от концентрации пирувата.
Анализ функциональных свойств варианта R90I HalhyTA
Показана активность варианта в полных реакциях трансаминирования с аминокислотой D-аланином и амином (R)-фенилэтиламином (R-PEA) (Таблица 3).
Таблица 3. Кинетические параметры полных реакций трансаминирования D-аланин + α-кетоглутарат в 50 мM K-фосфатном буфере, pH 8.0, и (R)-PEA + α-кетоглутарат в 50 мM CHES буфере, pH 9.0, катализируемых вариантом R90I при 25°C.
Активность варианта с разными аминами определена методом полуреакций (Таблица 4).
Таблица 4. Наблюдаемая скорость полуреакции с аминами варианта R90I: условия 10 mM амина в 50 мM CHES буфере, pH 9.0, при 25°C.
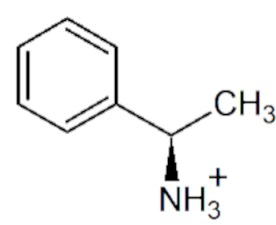
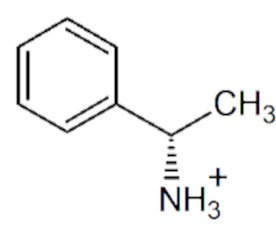
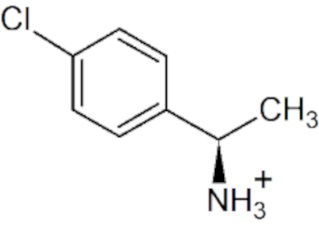
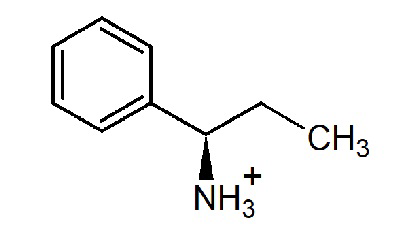
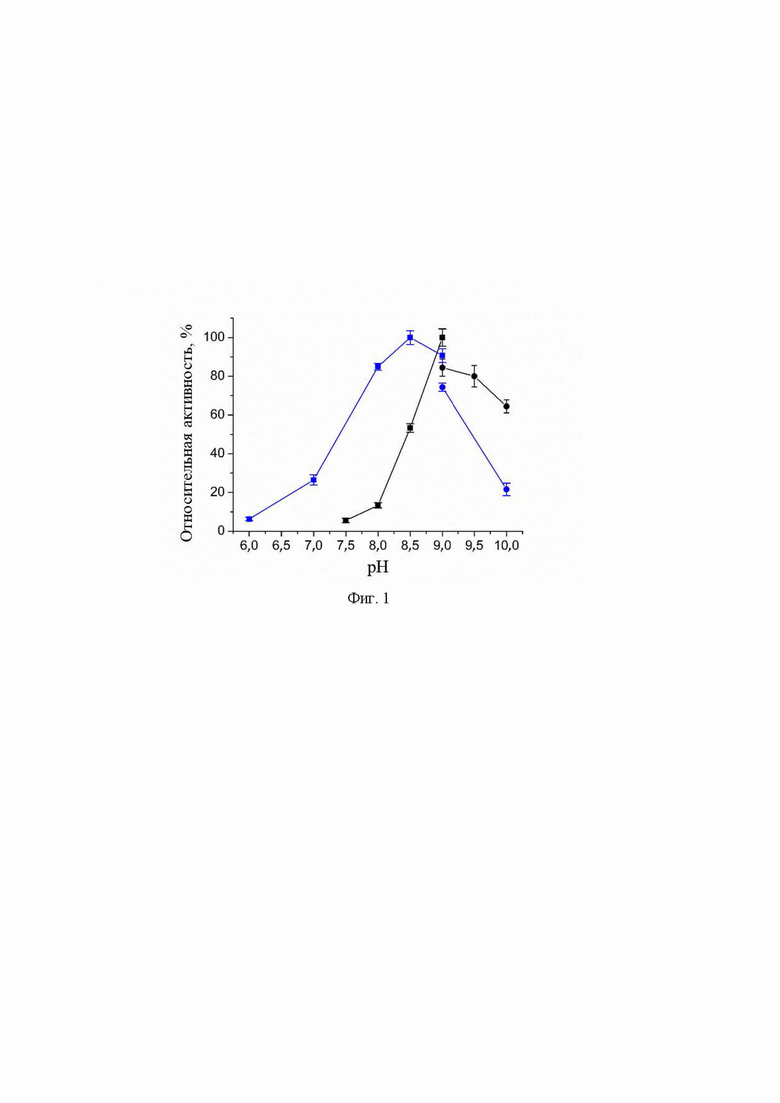
pH- и температурная зависимости активности варианта R90I в полных реакциях трансаминирования.
pH- и температурная зависимости активности варианта R90I определяли в полных реакциях трансаминирования с субстратами D-аланин и R-PEA. Анализ рН оптимума приведен на Фиг. 1. Анализ температурного оптимума реакций показал, что температура 25°С оптимальна для обеих реакций.
На Фиг.1 представлены рН -зависимости активности варианта R90I в полных реакциях трансаминирования D-аланин + α-кетоглутарат (синий); (R)-PEA + α-кетоглутарат (черный). 100% активности соответствует 1,7 ± 0,1 U/мг (синий), 0,050 ± 0,002 U/мг (черный) при 25°C. Синие/черные квадраты: смешанный буфер 25 мМ Трис-HCl и 25 мМ K-фосфат, pH 6,0-9,0. Синие/черные круги: 50 мМ CHES буфер, pH 9,0 - 10,0. Реакционная смесь содержала 5 мМ D-аланин, 2 мМ α-кетоглутарат, 30 мкМ PLP, 330 мкM NADH, 1 U/мл LDH и 0,01 мг/мл R90I вариант или 5 мМ (R)-PEA, 2 мМ α-кетоглутарат, 30 мкМ PLP и 0,1 мг/мл R90I вариант.
Ссылки:
1. Braunstein AE. Amino Group Transfer. In: P.D. B, editor. The Enzymes [Internet]. 3d ed. London: Acad. Press; 1973. p. 379-481. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1874604708601225.
2. Eliot AC, Kirsch JF. Pyridoxal Phosphate Enzymes: Mechanistic, Structural, and Evolutionary Considerations. Annu Rev Biochem [Internet]. 2004 Jun;73(1):383-415. Available from: http://www.annualreviews.org/doi/10.1146/annurev.biochem.73.011303.074021.
3. Toney MD. Reaction specificity in pyridoxal phosphate enzymes. Arch Biochem Biophys [Internet]. 2005 Jan;433(1):279-87. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0003986104005338.
4. Eds. J. Polaina and A.P. MacCabe, editor. Industrial Enzymes: Structure, Function and Applications.
5. Brundiek H, Höhne M. Transaminases - A Biosynthetic Route for Chiral Amines. In: Applied Biocatalysis: From Fundamental Science to Industrial Applications [Internet]. Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA; 2016. p. 199-218. Available from: http://doi.wiley.com/10.1002/9783527677122.ch10.
6. Steffen-Munsberg F, Vickers C, Kohls H, Land H, Mallin H, Nobili A, et al. Bioinformatic analysis of a PLP-dependent enzyme superfamily suitable for biocatalytic applications. Biotechnol Adv [Internet]. 2015 Sep;33(5):566-604. Available from: http://dx.doi.org/10.1016/j.biotechadv.2014.12.012.
7. Slabu I, Galman JL, Lloyd RC, Turner NJ. Discovery, Engineering, and Synthetic Application of Transaminase Biocatalysts. ACS Catal [Internet]. 2017 Dec 7;7(12):8263-84. Available from: http://pubs.acs.org/doi/10.1021/acscatal.7b02686.
8. Desai AA. Sitagliptin Manufacture: A Compelling Tale of Green Chemistry, Process Intensification, and Industrial Asymmetric Catalysis. Angew Chemie Int Ed. 2011 Feb;50(9):1974-6.
9. Savile CK, Janey JM, Mundorff EC, Moore JC, Tam S, Jarvis WR, et al. Biocatalytic Asymmetric Synthesis of Chiral Amines from Ketones Applied to Sitagliptin Manufacture. Science (80- ) [Internet]. 2010 Jul 16;329(5989):305-9. Available from: http://www.sciencemag.org/cgi/doi/10.1126/science.1188934.
10. Gu X, Zhao J, Chen L, Li Y, Yu B, Tian X, et al. Application of transition-metal catalysis, biocatalysis, and flow chemistry as state-of-the-art technologies in the synthesis of LCZ696. J Org Chem [Internet]. 2020 Jun 5;85(11):6844-53. Available from: https://pubs.acs.org/doi/10.1021/acs.joc.0c00473.
11. Mangion IK, Sherry BD, Yin J, Fleitz FJ. Enantioselective synthesis of a dual orexin receptor antagonist. Org Lett. 2012;14(13):3458-61.
12. Peng Z, Wong JW, Hansen EC, Puchlopek-Dermenci ALA, Clarke HJ. Development of a Concise, Asymmetric Synthesis of a Smoothened Receptor (SMO) Inhibitor: Enzymatic Transamination of a 4-Piperidinone with Dynamic Kinetic Resolution. Org Lett [Internet]. 2014 Feb 7;16(3):860-3. Available from: https://pubs.acs.org/doi/10.1021/ol403630g.
13. Molinaro C, Bulger PG, Lee EE, Kosjek B, Lau S, Gauvreau D, et al. CRTH2 Antogonist MK-7246: Synthetic Evolution from Discovery through Development. J Org Chem. 2012;77:2299-309.
14. Limanto J, Ashley ER, Yin J, Beutner GL, Grau BT, Kassim AM, et al. A highly efficient asymmetric synthesis of vernakalant. Org Lett. 2014;16(10):2716-9.
15. Obot IB, Solomon MM, Umoren SA, Suleiman R, Elanany M, Alanazi NM, et al. Progress in the development of sour corrosion inhibitors: Past, present, and future perspectives. J Ind Eng Chem. 2019;79:1-18.
16. Barel AO, Paye M, Maibach HI. Handbook of Cosmetic Science and Technology. New York: Informa Healthcare USA, Inc; 2009. 1-887 p.
17. Landim Neves MI, Silva EK, Meireles MAA. Natural blue food colorants: Consumer acceptance, current alternatives, trends, challenges, and future strategies. Trends Food Sci Technol. 2021;112(September 2020):163-73.
18. Blick SKA, Keating GM. Lisdexamfetamine. Pediatr-Drugs. 2007;9(2):129-35.
19. Abdullah A, Yusof MKM. Lebetalol: a brief current review. Pharmacophore. 2019;10(6):50-6.
20. Scott LJ. Repaglinide A Review of Its Use in Type 2 Diabetes Mellitus. Drugs. 2012;72(2):249-72.
21. Nguyen K, Hoffman H, Chakkamparambil B. Evaluation of rivastigmine in Alzheimer ’ s disease. Neurodegener Dis Manag. 2021;11:35-48.
22. Paton NI, Musaazi J, Sc M, Kityo C, Ph D, Walimbwa S, et al. Dolutegravir or Darunavir in Combination with Zidovudine or Tenofovir to Treat HIV. N Engl J Med. 2021;385:330-41.
23. Yan Z, Liu A, Ou Y, Li J, Yi H, Zhang N, et al. Design, synthesis and fungicidal activity evaluation of novel pyrimidinamine derivatives containing phenyl-thiazole/oxazole moiety. Bioorganic Med Chem. 2019;27(15):3218-28.
24. Fermanian TW, E. HJ. Fall Application of Prodianline for Spring Crabgrass. Weed Technol. 1994;8:612-6.
25. Thompson CJ, Seto H. Bialaphos. In: Genetics and Biochemistry of Antibiotic Production. New York: Elsevier Inc.; 1995. p. 197-222.
26. Adams JP, Brown MJB, Diaz-Rodriguez A, Lloyd RC, Roiban GD. Biocatalysis: A Pharma Perspective. Adv Synth Catal. 2019 Jun;361(11):2421-32.
27. Winkler CK, Schrittwieser JH, Kroutil W. Power of Biocatalysis for Organic Synthesis. ACS Cent Sci [Internet]. 2021 Jan 27;7(1):55-71. Available from: https://pubs.acs.org/doi/10.1021/acscentsci.0c01496.
28. Wu S, Snajdrova R, Moore JC, Baldenius K, Bornscheuer UT. Biocatalysis: enzymatic synthesis for industrial applications. Angew Chemie Int Ed [Internet]. 2021 Jan 4;60(1):88-119. Available from: https://onlinelibrary.wiley.com/doi/10.1002/anie.202006648.
29. Guo F, Berglund P. Transaminase biocatalysis: optimization and application. Green Chem [Internet]. 2017;19(2):333-60. Available from: http://dx.doi.org/10.1039/c6gc02328b.
30. Höhne M, Schätzle S, Jochens H, Robins K, Bornscheuer UT. Rational assignment of key motifs for function guides in silico enzyme identification. Nat Chem Biol [Internet]. 2010 Nov 26;6(11):807-13. Available from: http://dx.doi.org/10.1038/nchembio.447.
31. Mutti FG, Fuchs CS, Pressnitz D, Sattler JH, Kroutil W. Stereoselectivity of Four ( R ) -Selective Transaminases for the Asymmetric Amination of Ketones. Adva. 2011;353:3227-33.
32. Yu H. Biochemical and Structural Characterization of an ( R ) -Selective Transaminase in the Asymmetric Synthesis of Chiral Hydroxy Amines. Adv Synth Catal. 2021;363:4582-9.
33. Bezsudnova EY, Popov VO, Boyko KM. Structural insight into the substrate specificity of PLP fold type IV transaminases. Appl Microbiol Biotechnol [Internet]. 2020 Mar 27;104(6):2343-57. Available from: http://link.springer.com/10.1007/s00253-020-10369-6.
34. Kelly SA, Pohle S, Wharry S, Mix S, Allen CCR, Moody TS, et al. Application of ω-Transaminases in the Pharmaceutical Industry. Chemical Reviews. 2018.
35. Patil MD, Grogan G, Bommarius A, Yun H. Recent advances in ω-transaminase-mediated biocatalysis for the enantioselective synthesis of chiral amines. Catalysts [Internet]. 2018 Jun 21;8(7):254. Available from: http://www.mdpi.com/2073-4344/8/7/254.
36. Makarova O, Kamberov E, Margolis B. Generation of deletion and point mutations with one primer in a single cloning step. Biotechniques [Internet]. 2000 Nov;29(5):970-2. Available from: https://www.future-science.com/doi/10.2144/00295bm08.
37. Mikhailova AG, Rakitina T V., Timofeev VI, Karlinsky DM, Korzhenevskiy DA, Agapova YК, et al. Activity modulation of the oligopeptidase B from Serratia proteamaculans by site-directed mutagenesis of amino acid residues surrounding catalytic triad histidine. Biochimie [Internet]. 2017 Aug;139:125-36. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0300908416303236.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Фермент
трансаминаза D-аминокислот из Haliscomenobacter hydrossis с
расширенной субстратной специфичностью.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2023-12-01">
<ApplicationIdentification>
<IPOfficeCode></IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate>2023-12-01</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>HH</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1</ApplicationNumberText>
<FilingDate>2023-12-01</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ФИЦ Биотехнологии
РАН</ApplicantName>
<ApplicantNameLatin>Research Center of Biotechnology RAS
</ApplicantNameLatin>
<InventorName languageCode="ru">Екатерина Юрьевна
Безсуднова</InventorName>
<InventorNameLatin>Ekaterina Y. Bezsudnova</InventorNameLatin>
<InventionTitle languageCode="ru">Фермент трансаминаза D-аминокислот
из Haliscomenobacter hydrossis с расширенной субстратной
специфичностью</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>843</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..843</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q3">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Haliscomenobacter
hydrossis</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgattaaatattacaacatcaacggtcagcaggttccggtagaaaacg
ctactctgcacgtttctgacctgagcatcctgcgcggctacggtatcttcgactactttctcgctcgtga
aggccacccgctgttcctcgatgattatttaaaccgtttttaccgtagcgcggcagaactgtatctggaa
atcccgttcgacaaagctgaacttcgccgtcagatctatgctctgctgcaggcgaatgaggttcgtgaag
cgggtatcattctggtgctgacgggtggctactctcctgacggttataccccggttaacccgaacctgct
gatcatgatgtacgatctgccggcttccgcttgggaattcagcgctcagggcattaaaatcattacccac
ccgtttcagcgtgaactgccggaagtcaaaaccatcaactactccactggtatccgcatgctgaagacga
tcaaagaacgtggtgctacagacctgatctacgtggaccagggtgaatggattcgtgaaagcgctcgttc
taacttcttcctggtgatgccggacaacaccattgttaccgcggatgaaaagatcctctgggggatcacg
cgccgtcaagtcatcgatgcagcacgtgaagccggctatgccgttgaggaacgtcgcattcacatcactg
aactggaccaggctcgtgaagcattctttactagcaccattaaaggtgtgatggcaatcggccagatcga
cgatcgtgttttcggtgatggcactatcggtaaagtgacccaagaactgcaggatctcttcgttggcaag
gttaaagcatacctggaaacctgc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к области органической химии и биотехнологии. Предложен фермент трансаминаза D-аминокислот из Haliscomenobacter hydrossis, кодируемый нуклеотидной последовательностью SEQ ID:1, применяемый для реакции трансаминирования первичных R-аминов. Изобретение позволяет получить трансаминазу D-аминокислот из Haliscomenobacter hydrossis с расширенной субстратной специфичностью, что важно для разработки биокатализаторов стереоселективного аминирования органических соединений на основе природных ферментов-трансаминаз. 1 ил., 4 табл.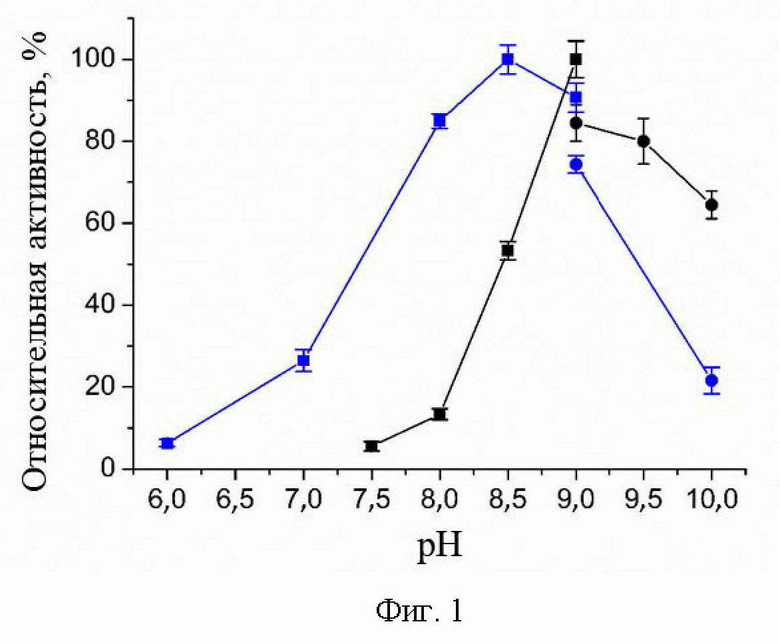
Фермент трансаминаза D-аминокислот из Haliscomenobacter hydrossis, кодируемый нуклеотидной последовательностью SEQ ID:1, применяемый для реакции трансаминирования первичных R-аминов.
| BAKUNOVA A.K | |||
| et al | |||
| The Uncommon Active Site of D-Amino Acid Transaminase from Haliscomenobacter hydrossis: Biochemical and Structural Insights into the New Enzyme | |||
| Molecules | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| NCBI Reference Sequence: WP_013764869.1, 22.05.2021 | |||
| Aminotransferase class IV [Haliscomenobacter hydrossis] | |||

