Область применения
Изобретение относится к области молекулярной биологии и генетической инженерии и касается термостабильных ферментов с активностью ДНК-лигаз. Предлагается новая ДНК-лигаза из архей рода Thermococus и способ ее выделения.
Актуальность
Ферменты - биокатализаторы широко используются в различных отраслях промышленности, сельского хозяйства и медицины. Одним из наиболее быстрорастущих рынков является производство ферментов для молекулярно-биологических исследований в самых различных областях деятельности, от медицины до криминалистики. В качестве примера можно привести термофильные ДНК-полимеразы, используемые в полимеразной цепной реакции (ПЦР), внедрение которой в практику в 1990-х годах совершило по существу революцию в диагностике инфекционных заболеваний.
Наряду с ПЦР, известен ряд других методов детекции и амплификации специфических последовательностей ДНК. К ним относится метод лигазной цепной реакции (ligase chain reaction, LCR), эффективный для детекции ДНК-полиморфизмов, мутаций, генетических аномалий. Несмотря на то, что метод LCR известен уже давно и активно применяется в диагностической практике, например в США, в России он практически не используется, в значительной степени из-за отсутствия термостабильных ДНК-лигаз на Российском рынке. Другой областью использования термостабильных лигаз является их использование для «сборки» синтетических генов из олигонуклеотидов, проводимой при повышенной температуре для снижения вероятности образования неспецифических продуктов лигирования.
Возрастающая потребность в термостабильных ДНК-лигазах с различными характеристиками стимулирует интерес крупнейших исследовательских центров и ведущих биотехнологических компаний во всем мире к поиску и созданию новых ДНК-лигаз, обладающих улучшенными либо новыми свойствами - высокой термостабильностью, удельной активностью, устойчивостью к ингибиторам и т.д.
Уровень техники
ДНК-лигазы играют важную роль в репликации, рекомбинации и репарации ДНК [Lehman, 1974]. Они присутствуют у эукариот, бактерий и архей и могут быть разделены на два семейства в соответствии с используемыми высокоэнергетическими кофакторами: АТФ-зависимые ДНК-лигазы (КФ 6.5.1.1) и НАД - зависимые лигазы (КФ 6.5.1.2). ДНК-лигазы катализируют присоединение 5'-Р донорного конца к 3'-ОН акцепторному концу в результате трех последовательных реакций переноса [Lehman, 1974; Engler and Richardson, 1983]. Все известные в настоящее время эукариотические и архейные ДНК-лигазы, а также лигазы из бактериофагов и вирусов являются АТФ-зависимыми и содержат специфические мотивы в их аминокислотных последовательностях [Georlette et al., 2000].
Известны АТФ-зависимые ДНК-лигазы из гипертермофильных архей Thermococcus kodakaraensis и Thermococcus fumicolanus, которые могут использоваться в качестве кофактора как АТФ, так и НАДН [Nakatani et al., 2000; Rolland et al., 2004].
Известны рекомбинантные термостабильные белки, имеющие ДНК-лигазную активность. Белок выделен из Thermococcus sp. (штамм 9°N-7), является АТФ-зависимым и сохраняет активность при 100°C [WO 2007035439, опубл. 2007-03-29].
Настоящее изобретение касается выделения и характеристики новой ДНК-лигазы LigTh1519 из гипертермофильной археи, выделенной из глубоководных гидротерм Гуаймаса, а также разработки способа ее получения.
С помощью метода полимеразной цепной реакции был идентифицирован и затем секвенирован ген ДНК-лигазы из термофильных архей рода Thermococcus sp. (штамм 1519), имеющий нуклеотидную последовательность SEQ ID NO 1. Методом культивирования рекомбинантного штамма E.coli DLT1270, содержащего плазмиду pRARE, трансформированного вектором pQE30, содержащим нуклеотидную последовательность SEQ ID NO 1, была выделена и очищена методом металл-афинной хроматографии новая рекомбинантная термостабильная ДНК-лигаза, проявляющая лигазную активность в присутствии ионов магния, в широком диапазоне значений рН (7,0-10,7) и концентраций натрия хлорида (25-150 мМ), состоящая из 559 аминокислот, имеющая массу 63 кДа, определенную методом электрофореза в полиакриловом геле и содержащая аминокислотную последовательность SEQ ID NO 2, получаемую из рекомбинантного штамма E.coli.
Раскрытие изобретения
Лигаза LigTh1519 выделена из археи Thermococcus sp. штамм 1519 и характеризуется тем, что состоит из 559 аминокислот и имеет массу 63 кДа, определенную методом электрофореза в полиакриловом геле. LigTh1519 имеет гомологию по аминокислотным последовательностям с другими АТФ-зависимыми ДНК-лигазами археи, в том числе с лигазой Т. kodakaraensis (91%), Т. fumicolans (89%), Pyrococcus abyssi (82%). Анализ аминокислотной последовательности LigTh1519 выявил три характерных домена АТФ-зависимых ДНК-лигаз. Первый домен, N-концевой район ДНК-лигазы (DNA_Ligase_A_N), встречается в большинстве, но не во всех АТФ-зависимых ДНК-лигазах. Этот регион необходим для связывания фермента с ДНК и участвует в катализе реакции. Второй домен - DNA_Ligase_A_M, встречается в ДНК-лигазах, точные функциональные свойства его не известны. Третий, C-концевой домен (DNA_Ligase_A_C), составляет основную часть каталитического центра АТФ-зависимых ДНК-лигаз. Более 50% активности сохраняется после инкубации ферменты в течении 30 минут при температуре 80°C. Отличие от близких по структуре лигаз в том, что новый фермент активен в широком диапазоне значений рН (7,0-10,7) и концентраций натрия хлорида (25-150 мМ).
Одной из основных областей практического применения термостабильных ДНК-лигаз является амплификация ДНК в лигазной цепной реакции (ЛЦР). Проведение ЛЦР требует высокой термостабильности фермента, и в этом плане LigTh1519 не дает особо значимых преимуществ. Однако новый фермент неожиданно обнаруживает ряд полезных свойств: он обеспечивает дотирование лишь фрагментов ДНК, имеющих протяженные (12-нт в случае BstEII-фрагментов фага 1), но не короткие (4-нт концы HindIII фрагментов фага 1) липкие концы, и не способен лигировать фрагменты ДНК с «тупыми» концами. Поэтому одним из перспективных применений нового фермента может быть «сборка» синтетических генов из олигонуклеотидов. Проведение реакции лигирования при повышенной температуре (60-70°C) должно повысить эффективность «сборки» фрагмента ДНК из олигонуклеотидов за счет снижения вероятности образования неспецифических продуктов лигирования. Такие продукты могут образовываться при использовании обычной мезофильной ДНК лигазы (например, ДНК лигазы фага Т4) в результате отжига случайным образом слипшихся коротких комплиментарных участков олигонуклеотидов, использованных для синтеза гена. Термостабильность LigTh1519 и ее способность лигировать лишь протяженные липкие концы должны снизить вероятность образования неспецифических продуктов при «сборке» генов с ее применением.
Краткое описание чертежей
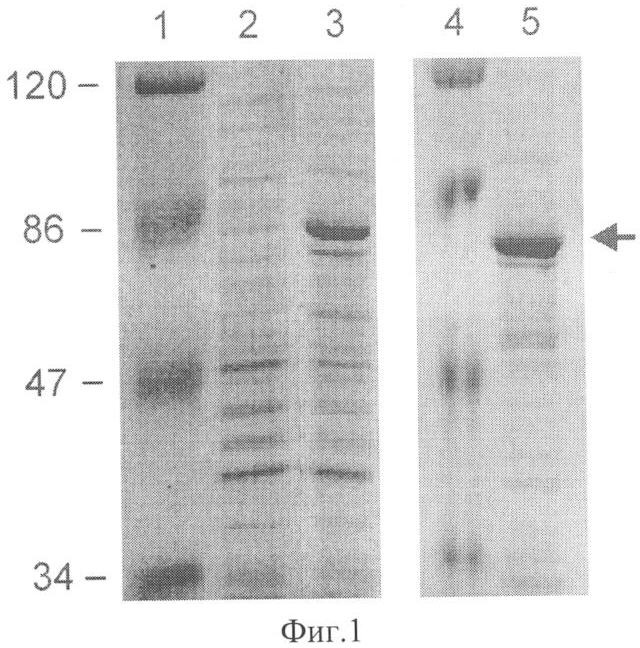
Фиг 1. Экспрессия LigTh1519 в Е.coli и очистка белка.
Приведены результаты ДСН-ПААГ анализа белковых препаратов:
1 и 4 - маркеры молекулярной массы в кД,
2 - суммарный белковый препарат, выделенный из клеток штамма DLT1270/pRARE, содержащего плазмиду pQE30-LigTh1519, до индукции синтеза LigTh1519,
3 - то же, что на дорожке 2, но через 3 часа после индукции синтеза LigTh1519 внесением в среду 1 мМ Изопропил-бета-В-тиогалактозид (ИПТГ),
5 - белковый препарат.
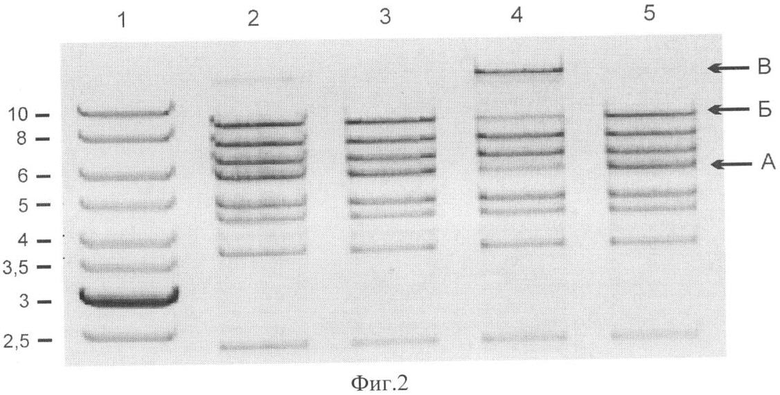
Фиг.2. Анализ способности LigTh1519 лидировать фрагменты, образованные при обработке ДНК фага λ эндонуклеазой BstEII.
Приведены результаты анализа препаратов ДНК с помощью гель-электрофореза в агарозном геле.
1 - маркер молекулярной массы ДНК (длины фрагментов указаны в т.п.н.),
2 - ДНК фага λ после обработки рестриктазой BstEII,
3 - то же, что на дорожке 2, но перед нанесением на электрофорез препарат ДНК был прогрет в течение 10 мин при 70°C,
4 - препарат BstEII-фрагментов ДНК фага λ после инкубации в течение 1 ч при 45°C с 20 н. лигазы LigThl519 в реакционном буфере (40 мМ трис-HCl рН 7,0, 2 мМ MgCl2), содержащем 1 мМ АТФ. Перед нанесением на электрофорез препарат ДНК был прогрет в течение 10 мин при 70°C,
5 - то же, что на дорожке 4, но без добавления АТФ в буфер для лигирования.
Справа стрелками показаны концевые BstEII-фрагменты А (5678 п.н.) и Б (8453 п.н), а также продукт их лигирования, В (14131 п.н.).
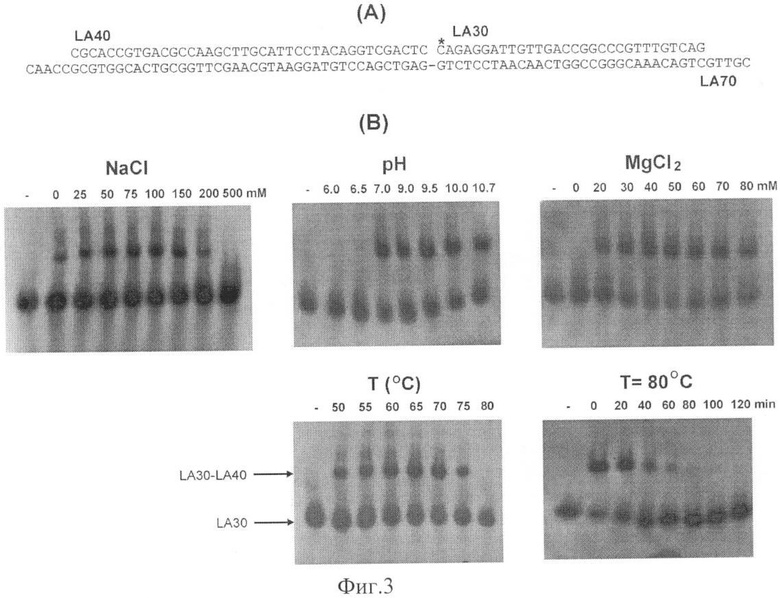
Фиг.3. Ферментативная характеристика ДНК лигазы LigTh1519.
3А - синтетический субстрат, использованный для определения эффективности лигирования однонитевого разрыва (Положение γ32Р - радиоактивной метки, введенной на 5'-конец олигонукдлеотида LA30, указано звездочкой),
3Б - влияние NaCl, величины рН, MgCl2, температуры проведения реакции и термостабильность фермента при 80°C.
Осуществление изобретения
Пример 1. Молекулярная идентификация штамма 1519.
Штамм 1519 был выделен из проб с гидротермального поля Гуаймас (1986 г.) и поддерживался в коллекции Лаборатории гипертермофильных микробных сообществ ИНМИ РАН. Штамм 1519 является облигатным анаэробом, органотрофом, использующим пептон или крахмал в качестве субстратов роста при обязательном присутствии в среде элементной серы, которую восстанавливает в сероводород. Оптимальная температура роста 85°C, оптимальный рН среды 6.2-6.4. Культивирование проводили в 15 мл пробирках Хангейта на среде следующего состава (г л-1): NH4Cl - 0.33, KCl - 0.33, KH2PO4 - 0.33, CaCl2·2H2O - 0.33, MgCl2·6H2O - 0.33, NaCl - 25, дрожжевой экстракт - 0.2, пептон - 2.0, элементная сера - 10.0; NaHCO3 - 0.5, Na2S·9H2O - 0.5, раствор микроэлементов [Balch et al., 1979] - 1 мл·л-1, витамины [Wolin et al., 1963] - 1 мл·л-1. Анаэробно приготовленная среда (рН 6.4) разливалась под током очищенной от кислорода смеси N2/CO2 (80:20) порциями по 10 мл и автоклавировалась при избыточном давлении (0.5 атм), температура инкубации 85°C.
Геномную ДНК штамма 1519 выделяли стандартными методами [Ausubel et al., 1997], последовательность гена 16S рРНК определяли, как описано в работе [Sokolova et al., 2002]. Филогенетическое положение штамма 1519 определяли путем построения дендрограммы, основанной на последовательностях генов 16S рРНК этого штамма и сравниваемых микроорганизмов.
Сравнение последовательности гена 16S рРНК штамма 1519 (1366 нт.) с представленными в GenBank сиквенсами показало, что штамм 1519 относится к отделу Euryarchaeota, классу Thermococci, порядку Thermococcales, семейству Thermococcaceae и роду Thermococcus, где его наиболее близкородственным организмом является Thermococcus kodakaraensis.
Пример 2. Выделение гена ДНК-лигазы Thermococcus sp 1519.
Анализ представленных в базе данных GenBank нуклеотидных последовательностей генов ДНК-лигаз архей выявил несколько консервативных участков, два из которых были использованы для создания пары специфических праймеров на гены ДНК-лигазы: Lig2F (AGGAGGAAGTACAACATCGA) и Lig5R (TTGAACCTCTCCTGGAGC).
С помощью этих праймеров на суммарной ДНК Thermococcus sp1519 был амплифицирован фрагмент длиной около 600 п.н. Была использована следующая программа проведения ПЦР: 94°C - 1 мин, 50°C - 140 сек и 72°C - 1,5 мин, всего 40 циклов. Нуклеотидная последовательность полученного фрагмента имела высокую гомологию с последовательностями генов АТФ-зависимых ДНК-лигаз архей. Для клонирования полноразмерного гена ДНК-лигазы LigTh1519 был использован метод «инверсного» ПЦР. Для этого были использованы две пары направленных в противоположные стороны праймеров: Lig7F (TGAGGAGAGCGAGAAGATAA) и Lig6R (CTATGAGGCTCTCGCCGTCA) для выделения 5' - конца гена, а также Lig8R (TATCTTCCCTCAACGCCACG) и Lig9F (ACACGATAGAGAGGATTGCC) для определения последовательности 3' - конца гена. Эти пары праймеров расположены вблизи концов просеквенированного 600-нт фрагмента. ДНК Thermococcus sp 1519 обрабатывали ферментами SacI и BstEII, полученные фрагменты замыкали в кольцо, обрабатывая Т4 ДНК-лигазой и использовали в качестве матрицы для ПЦР с парами праймеров Lig7F-Lig6R и Lig9F-Lig8R соответственно. Полученные в результате этих ПЦР фрагменты ДНК были просеквенированы, в результате объединения полученных последовательностей была определена полная нуклеотидная последовательность гена ДНК-лигазы Thermococcus sp 1519, представленная в SEQ ID NO 1.
Пример 3. Получение штамма Е.coli - продуцента ДНК-лигазы Thermococcus sp 1519, выделение и очистка фермента.
Праймеры 1519-F2 (GGATCCAAGTATTCCGAGTTGGCTG) и праймер 1519R (TAGATCTTACCTCTTTGCCTTGAACTTCTC) были использованы для ПЦР-амплификации полноразмерного гена LigTh1519, в качестве матрицы использовали геномную ДНК Thermococcus sp1519. Полученный фрагмент обрабатывали рестриктазами BamHI и BglII и клонировали в сайте BamHI в экспрессионном векторе pQE30 (Qiagen), позволяющем экспрессировать целевой белок, слитый с 6 аминокислотными остатками гистидина на N-конце. В результате был получен рекомбинантный экспрессионный вектор pQE30-LigTh1519.
Штамм Е.coli DLT1270, являющийся производным штамма DH10B [Grant et al., 1990], содержит ген репрессора лактозного оперона lacI, интегрированный в хромосому. Штамм DLT1270/pRARE, содержащий плазмиду pRARE-2 (Novagen), кодирующую «редкие» для Е.coli т-РНК, был использован для экспрессии ДНК-лигазы LigTh1519. Для этого pQE30-LigTh1519 вводили в штамм Е.coli DLT1270/pRARE.
Указанный штамм, содержащий рекомбинантную плазмиду pQE30-LigTh1519, выращивали на шейкере при 37°C до достижения Д600 ~0.5, затем индуцировали синтез рекомбинантного белка, внося изопропил-β-D-тиогалактопиранозид (ИПТГ) до 1 мМ, и продолжали выращивать культуру в течение 18 ч при 37°C. Выделение и очистку лигазы LigTh1519 осуществляли методом металл-афинной хромотографии с помощью набора Spin-NTA KIT (Qiagen, Германия). В результате был получен гомогенный препарат белка LigTh1519 (фиг1).
Пример 4. Определение активности ДНК-лигазы.
Для определения лигазной активности in vitro использовали два метода. Во-первых, метод, описанный в работе [Muerhoffet al., 2004], который основан на дотировании BstEII - фрагментов ДНК фага λ. Концевые фрагменты длиной 8453 п.н. и 5678 п.н., содержащие когезивные концы длиной 12 нуклеотидов, образуют стабильный дуплекс при температуре 45°C. Анализ лигазной активности проводили в буфере, содержащем 40 мМ трис-HCl (рН 7.0), 1 мМ АТФ, 100 мМ NaCl и 10 мМ MgCl2. 1 мкг ДНК фага X, обработанной BstEII, инкубировали с 20 нг белка LigTh1519 в 40 мкл указанного буфера, реакцию проводили при 45°C в течение 1 ч. Если тестируемый фермент обладает лигазной активностью, то из двух концевых BstEII-фрагментов длиной 8453 нт и 5678 нт, образуется фрагмент длиной 14131 нт. Реакцию лигирования останавливали добавлением 50 мМ ЭДТА и прогревали в течение 10 мин при 70°C для денатурации нелигированных концов. Пробы анализировали с помощью электрофореза в 0.7%-ном агарозном геле (фиг.2).
Представленные на фиг.2 результаты показывают, что рекомбинантный белок LigTh1519 действительно проявлял активность ДНК-лигазы, активность фермента LigTh1519 зависела от АТФ. Так в реакции, не содержащей АТФ (фиг.2, дор. 5), фрагмент длиной 14131 нт не образовывался. Таким образом, LigThl519 является АТФ-зависимой ДНК-лигазой. Отметим, что лигирование остальных (не концевых) рестрикционных фрагментов не наблюдалось ни в описанном эксперименте с BstEII, ни в случае обработки ДНК фага λ рестриктазой HindIII, образующей липкие концы длиной 4 нт.
Второй метод, описанный в работе [Nakatani et al., 2000], позволял количественно определять лигазную активность по «зашиванию» однонитевого разрыва в двунитевом фрагменте ДНК. В качестве субстрата использовали дуплекс, образованный в результате отжига трех синтетических олигонуклеотидов: LA-30 (CAGAGGATTGTTGACCGGCCC GTTTGTCAG), LA-40 (CGCACCGTGACGCCA AGCTTGCATTCCTACAGGTCGACTC) и LA-80 (CGTTGCTGACAAACGGGCCGGTCA ACAATCCTCTGGAGTCGACCTGTAGGAATGCAAGCTTGGCGTCACGGTGCGCCAAC).
В олигонуклеотид LA-30 с помощью Т4 полинуклеотид киназы вводили радиоактивную метку на 5'-конец. Лигазную реакцию проводили в указанных выше условиях (состав буфера и температуру реакции меняли для определения оптимальных условий реакции), по окончании реакции денатурировали ДНК прогреванием в течение 15 мин при 94°C. Продукты реакции анализировали с помощью электрофореза в денатурирующем полиакриламидном геле и радиоавтографии. В результате «зашивания» однонитевого разрыва ДНК-лигазой радиоактивно меченный праймер LA30 соединялся с LA40, что приводило к образованию олигонуклеотида длиной 70 нуклеотидов (нт), детектируемого при помощи электрофореза с последующей радиоавтографией.
Пример 5. Влияние температуры, рН и концентрации солей на активность ДНК лигазы LigTh1519.
Полуколичественное определение лигазной активности LigTh1519 проводили на полученном в результате отжига трех олигонуклеотидов фрагменте ДНК, содержащем однонитевой разрыв в одной из цепей (фиг.3А). Эффективность лигирования определяли по способности фермента «зашивать» этот разрыв и образованию продукта лигирования, - олигонуклеотида длиной 70 нт [Nakatani et al., 2000]. Определяли зависимость активности LigTh1519 от рН буфера, концентраций NaCl и MgCl2, а также температуры.
Представленные на фиг.3 результаты показывают, что LigTh1519 активен в широком диапазоне значений рН (7,0-10,7) и концентраций NaCl (25-150 мМ). В этом отношении LigTh1519 отличался от имеющей с ней высокую гомологию ДНК-лигазы Т. fumicolans [Rolland et al., 2004], проявляющей максимальную активность в узком диапазоне значений рН (6,5-7,0). Авторы установили, что для лигазной активности LigTh1519 необходимо наличие в среде двухвалентного катиона (Mg+), причем фермент активен в широком диапазоне концентраций MgCl2 (5-60 мМ), в отличие от лигазы Т. fumicolans, активность которой максимальна при 2 мМ MgCl2 и резко снижалась при концентрациях выше 5 мМ [5]. ДНК лигаза LigTh1519 проявляла активность в широком диапазоне температур. Хотя оптимальная температура реакции составляла 65-70°C (рис 3), фермент активен при температурах от 50 до 75°C и выше.
Термостабильность LigTh1519 оценивали, инкубируя фермент в реакционном буфере (без АТФ) при 80°C или 94°C, затем активность фермента определяли в стандартной реакции (40 мМ Трис-HCl buffer (рН 7.0), 60 мМ MgCl2, 100 мМ NaCl, 1 мМ АТФ) при 70°C. Авторы установили, что фермент терял половину своей активности в течение 40 мин при 80°C (фиг.3) и 5 мин при 94°C (данные не представлены). Таким образом, уровень термостабильности LigTh1519 ниже, чем у ДНК лигазы Т. fumicolanus (время полужизни 3 ч при 80°C и 15 мин при 90°C) и Pyrococcus furiosus [Патент США 5506137], но выше, чем у лигазы Sulfolobus shibatae [Lai et al., 2002].
СПИСОК ЛИТЕРАТУРЫ
1. Ausubel P.M., Brent R., Kingston R.E., Moore D.D., Seidman G., Smith J.A., Struhl K. Current Protocols in Molecular Biology. New York: John Wiley and Sons, 1997. P.112-118.
2. Balch W.E., Fox G.E., Magnum G.E., Woes G.E., Wolf R.S. // Microbiol. Rev. 1979. V.43. N 2. P.260-296.
3. Engler M.J., Richardson C.C. //J. Biol. Chem. 1983. V.258. N 18 P.11197-11205.
4. Georlette D., Jonsson Z.O., Van Petegem F., Chessa J., Van Beeumen J., Hubscher U., Gerday C. // Eur. J. Biochem. 2000. V.267. N 12. P.3502-3512.
5. Grant S.G., Jessee J., Bloom F.R., Hanahan D. // Proc. Natl. Acad. Sci. USA. 1990. V.87. N 12. P.4645-4649.
6. Lai X., Shao H., Hao F., Huang L. // Extremophiles. 2002. V.6. N 6. P.469-477.
7. Lehman I.R. // Science. 1974. V.186. N 4166. P.790-797.
8. Muerhoff A.S., Dawson G.J., Desai S.M. // J. Virol. Methods. 2004. V.119. N 2. P.171-6.
9. Nakatani M., Ezaki S., Atomi H., Imanaka T. // J. Bacteriol. 2000. V.182. N22. P.6424-6433.
10. Rolland J.L., Gueguen Y., Persillon С., Masson J.M., Dietrich J. // FEMS Microbiol. Lett. 2004. V.236. N 2. P.267-73.
11. Sokolova T.G., Kostrikina N.A., Chernyh N.A., Tourova T.P, Kolganova T.V., Bonch-Osmolovskaya E.A. // Int. J. Syst. Evolut. Microbiol. 2002. V.59. N 6. P.1961-1967.
12. Wiedmann M., Wilson W. J., Czajka J., Luo J., Barany F., Batt C.A. // PCR Methods Appl. 1994. V.3. N4. P.51-64.
13. Wolin E.A., Wolin M.J., Wolf R.S. // J. Biol. Chem. 1963. V.238, P.2882-2888.

Изобретение относится к биотехнологии и генетической инженерии и представляет собой рекомбинантную термостабильную ДНК-лигазу Thermococcus sp 1519, проявляющую лигазную активность в присутствии ионов магния, в широком диапазоне значений рН (7,0-10,7) и концентраций натрия хлорида (25-150 мМ). Такая лигаза состоит из 559 аминокислот, имеет массу 63 кДа, определенную методом электрофореза в полиакриловом геле, и содержит аминокислотную последовательность SEQ ID NO:2, получаемую из рекомбинантного штамма E.coli. Изобретение относится также к ДНК, кодирующей упомянутую лигазу и способу получения лигазы. Изобретение позволяет получить новую ДНК-лигазу с улучшенными свойствами. 3 н.п. ф-лы, 3 ил.
1. Рекомбинантная термостабильная ДНК-лигаза Thermococcus sp 1519, проявляющая лигазную активность в присутствии ионов магния, в широком диапазоне значений рН (7,0-10,7) и концентраций натрия хлорида (25-150 мМ), состоящая из 559 аминокислот, имеющая массу 63 кДа, определенную методом электрофореза в полиакриловом геле и содержащая аминокислотную последовательность SEQ ID NO:2, получаемая из рекомбинантного штамма Е.соli.
2. Выделенная нуклеотидная последовательность ДНК, кодирующая термостабильную ДНК-лигазу по п.1, включающая нуклеотидную последовательность SEQ ID NO:1.
3. Способ получения термостабильной ДНК-лигазы по п.1, включающий культивирование рекомбинантного штамма Е.соli DLT1270, содержащего плазмиду pRARE, трансформированного вектором pQE30, содержащим нуклеотидную последовательность SEQ ID NO:1, кодирующую ДНК-лигазу Thermococcus sp.1 519, выделение и очистку ДНК-лигазы методом металлаффинной хроматографии.
| MASARU NAKATANI et.al | |||
| A DNA Ligase from a Hyperthermophilic Archaeon with Unique Cofactor Specificity | |||
| J | |||
| Bacteriol | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| SEO M.S | |||
| et.al | |||
| Cloning and expression of a DNA ligase from the hyperthermophilic archaeon Staphylothermus marinus and properties of the enzyme | |||
| J | |||
| Biotechnol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Epub. | |||

