Изобретение относится к области клеточной биологии, молекулярной биологии и биотехнологии, конкретно, к получению белкового препарата модифицированного варианта цитокина TRAIL DR5-B - Манселана, который может быть использован в качестве терапевтического средства для лечения злокачественных опухолей.
Природный цитокин человека TRAIL (TNF-related apoptosis inducing ligand) рассматривается как потенциальное средство для терапии опухолевых заболеваний, поскольку он способен селективно индуцировать апоптоз в опухолевых клетках, не затрагивая нормальные клетки. Однако клинические испытания показали его низкую эффективность в связи с тем, что многие типы опухолей резистентны к TRAIL. Один из механизмов резистентности связан с наличием пяти рецепторов TRAIL, из которых лишь два, DR4 и DR5 (рецепторы смерти), проводят сигнал апоптоза, a DcR1, DcR2 и OPG являются рецепторами - «ловушками», которые не проводят сигнал апоптоза и ингибируют TRAIL опосредованный гибель опухолевых клеток. Ранее был получен уникальный рецептор-специфичный вариант TRAIL DR5-B с заменами аминокислотных остатков Y189N/R191K/Q193R/H264R/I266L/D269H в белке TRAIL, который селективно связывается лишь с одним из пяти рецепторов TRAIL, рецептором смерти DR5 (Gasparian ME, Chernyak BV, Dolgikh DA, Yagolovich AV et al. Apoptosis, 2009, 14:778-787). Исследования на линиях опухолевых клеток показали, что мутантный вариант DR5-B значительно эффективнее индуцирует апоптоз по сравнению с TRAIL дикого типа. В связи с этим рецептор-специфичный цитокин DR5-В является перспективным средством для лечения различных типов опухолей. Ранее авторами данного патента была описана методика получения рецептор-селективных мутантных вариантов TRAIL DR5-B, путем экспрессии в штамме Е. coli BL21(DE3) в виде гибридного белка с тиоредоксином и последующей очистки после расщепления слитного белка энтеропептидазой (Гаспарян М.Э., Яголович А.В., Долгих Д.А., Кирпичников М.П. Способ получения мутантного белка TRAIL человека. Роспатент №2405038 от 27.11.2010). Этот способ позволяет получить препаративные количества белка, однако он не подходит для использования препарата в клинических исследованиях, и, потенциально, для терапии опухолей по ряду причин. Несмотря на то, что энтеропептидаза расщепляет белок по специфическому сайту, расположенному между несущим белком тиоредоксином и целевым белком DR5-B, при расщеплении образуются следовые количества низкомолекулярных пептидов - продуктов неспецифического расщепления, способных оказывать побочные эффекты при введении в организм. Процесс очистки целевого белка включает дополнительный этап удаления энтеропептидазы методом аффинной хроматографии, при этом полностью избавится от следов фермента энтеропептидазы невозможно. Следовые количества энтеропептидазы приводят к постепенному накоплению низкомолекулярных белковых фрагментов неспецифического расщепления и снижению содержания активного белка при длительном хранении препарата. Кроме того, использование коммерческого препарата энтеропептидазы значительно повышает стоимость итогового продукта в связи с высокой стоимостью фермента, поскольку к использованию фермента в ходе очистки лекарственных препаратов предъявляются высокие требования к качеству: во-первых, фермент должен быть высокоочищенным и высокоактивным, во-вторых, в целевом белковом препарате должны отсутствовать даже следовые количества неспецифически расщепленных пептидов. Кроме того, трудозатратные и дорогостоящие этапы получение белка путем гибридной экспрессии с доменом тиоредоксина в качестве «несущего» белка затрудняют задачу масштабирования производства препарата для клинических испытаний. Еще один недостаток существующей методики заключается в том, что при экспрессии рекомбинантных белков в бактериальных клетках в препаратах превышен допустимый уровень бактериальных эндотоксинов, которые вызывают иммунный ответ в организме животных и способны привести к эндотоксиновому шоку. Разработанная ранее методика очистки рекомбинантного цитокина DR5-B включала только стадии метал-аффинной хроматографии, что не позволяет полностью избавиться от бактериальных эндотоксинов, следовых количеств энтеропептидазы и низкомолекулярных пептидных фрагментов неспецифического расщепления в препарате.
Известен другой способ цитоплазматической экспрессии цитокина TRAIL в виде слитного белка с мальтоз связывающим белком (МВР), где слитный белок расщепляют протеиназой вируса гравировки табака с последующей очисткой целевого белка (Do ВН, Nguyen МТ, Song JA, Park S, Yoo J, Jang J et al. Soluble prokaryotic expression and purification of bioactive Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand// J Microbiol Biotechnol. 2017, 27:2156-2164). Выход белка по данным этой работы составил 36 мл с 1 литра культуры клеток.
Известен также способ цитоплазматической экспрессии цитокина TRAIL с новым тагом Fh8-ΔI-CM, где Fh8 является полипептидом 8-кДа, полученный из низкомолекулярного антигена печеночной двуустка (Fasciola hepatica), который улучшает растворимость белка, a ΔI-CM это кальмодулин-связывающий полипептид способный расщепляться при низкиз рН раствора. (Zhang М, Wang Z, Chi L, Sun J, Shen Y. Enhanced production of soluble tumor necrosis factor-related apoptosis-inducing ligand in Escherichia coli using a novel self-cleavable tag system Fh8-ΔI-CM. Protein Expr Purif. 2018, 148:16-23). Выход белка в данной работе составил 77,5 мг белка с 1 литра культуры клеток с чистотой на 89%.
Препараты, полученные путем гибридной экспрессии, позволяют получить растворимый белок, однако для очистки целевого белка требуется дополнительный этап расщепления слитного белка с последующей очистки целевого белка. Это увеличивает трудоемкость очистки, а также влияет на себестоимость препарата, так как стоимость протеиназ достаточно высока. Кроме того, в ходе расщепления слитных белков часто происходит неспецифическое расщепление целевого белка в скрытых неспецифических сайтах, что приводит к образованию низкомолекулярных пептидных примесей от которых очень трудно избавиться. Поэтому препараты, полученные путем гибридной экспрессии, часто удовлетворяют требованиям, предъявляемым к качеству белка для биологических исследований (исследование структуры и функций белка, связывания с рецепторами, биологической активности на линиях раковых клеток и пр.), но не подходит для клинических исследований, а также для его применения в качестве лекарственного препарата.
Известны способы получение TRAIL в системах прямой экспрессии белка в штаммах Е. coli, где белок экспрессируется в нерастворимой форме в составе телец включения (Kang Н, Sun AY, Shen YL, Wei DZ. Refolding and structural characteristic of TRAIL/Apo2L inclusion bodies from different specific growth rates of recombinant Escherichia coli. Biotechnol Prog. 2007; 23:286-292), (Zhihua L, Huanzong Lei, Peng C. Expression, purification, and in vitro refolding of soluble tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Protein Expression and Purification, 2007; 51:276-282). При экспрессии белка в таких системах возникает необходимость проведения процедуры ренатурации, что сильно увеличивает на трудоемкость и стоимость работы и влияет на окончательные выходы препарата.
Изобретение решает задачу получения высокоочищенного рекомбинантного модифицированного белка DR5-B - Манселана - с высоким выходом без примесей и эндотоксинов с качеством, удовлетворяющим требованиям к лекарственным препаратам. Задача решается за счет модификации белка DR5-B аминокислотным остатком метионина на N-конце и разработки принципиально нового способа получения модифицированного белка DR5-B Манселана с высокой противоопухолевой активностью, включающего создание генетической конструкции (плазмидного вектора) для прямой экспрессии белка Манселана с последующей трансформацией высокопроизводительного относительно нового штамма Е. coli SHuffle В, экспрессию и очистку целевого белка с помощью методов аффинной и ионообменной хроматографии, а также удаление эндотоксинов из очищенного препарата для терапии злокачественных опухолей различного происхождения.
Плазмидный вектор pET-32a/DR5-B ранее был получен для гибридной экспрессии белка Trx/DR5-B с доменом тиоредоксина в качестве «несущего» белка (Gasparian ME, Chernyak BV, Dolgikh DA и др. Generation of new TRAIL mutants DR5-A and DR5-B with improved selectivity to death receptor 5. 2009, Apoptosis, 14:778-787). Плазмидный вектор для прямой экспрессии модифицированного белка DR5-B - Манселана - получают, вырезая ген тиоредоксина Trx из вектора pET-32a/DR5-B. Для этого предварительно вводят сайт рестрикции NdeI между генами тиоредоксина и DR5-В методом сайт-специфического мутагенеза. Плазмиду pET32a/DR5-B с введенным сайтом рестрикции NdeI обрабатывают рестриктазами XhoI и NdeI, получившиеся фрагменты ДНК разделяют в 1% агарозном геле, после чего фрагмент ДНК линеаризованного плазмидного вектора и фрагмент гена DR5-B элюируют из геля и лигируют с помощью Т4 ДНК лигазы. Таким образом получают новый плазмидный вектор pET-32a/manselan для прямой экспрессии целевого белка Манселана. В результате введения сайта рестрикции NdeI (С ATATG) непосредственно перед геном TRAIL DR5-B появляется кодон метионина ATG для инициации экспрессии модифицированного цитокина Манселана.
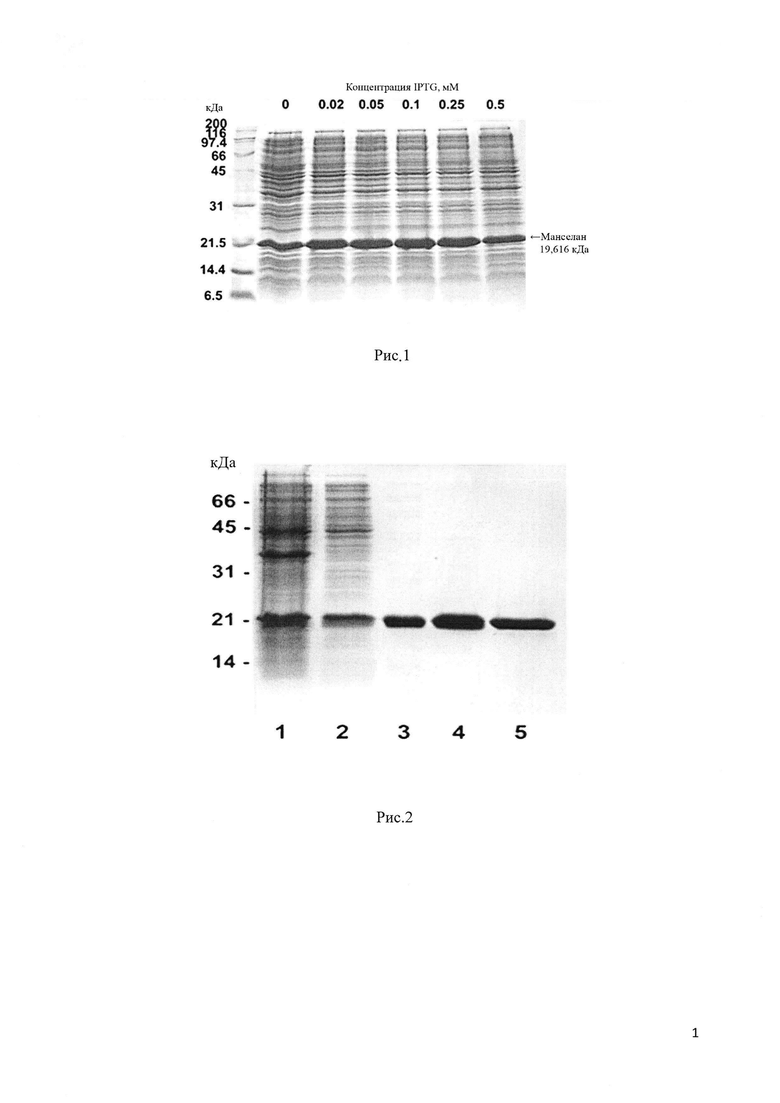
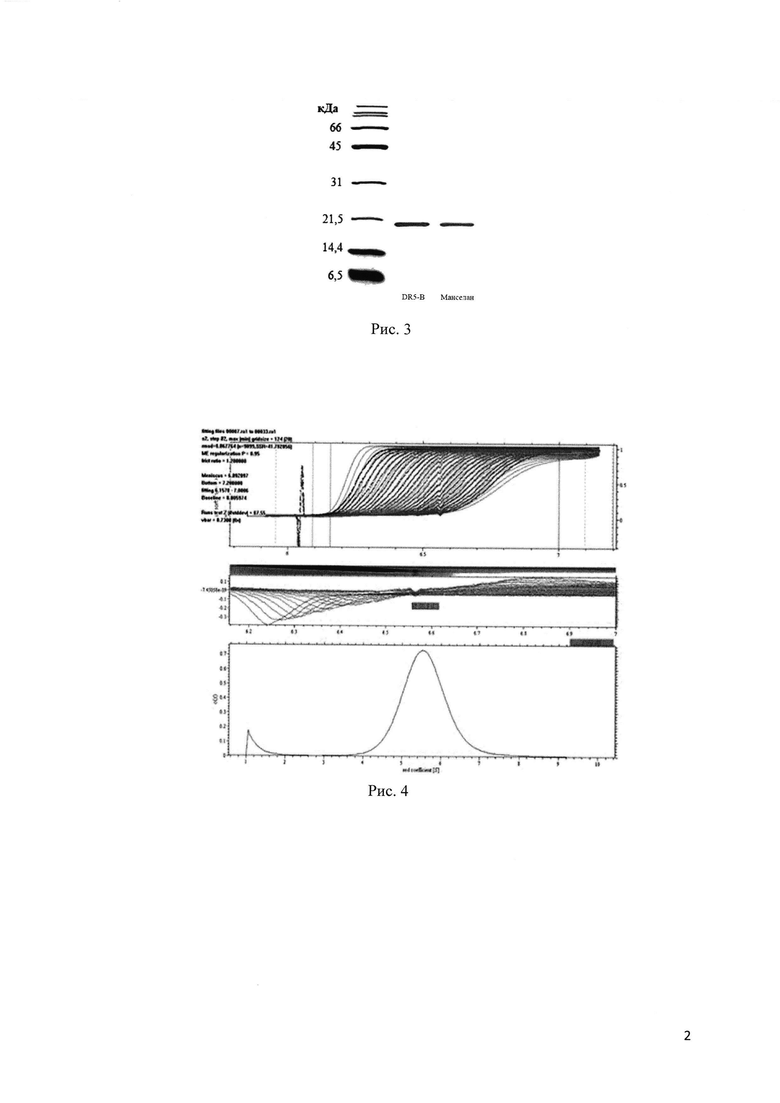
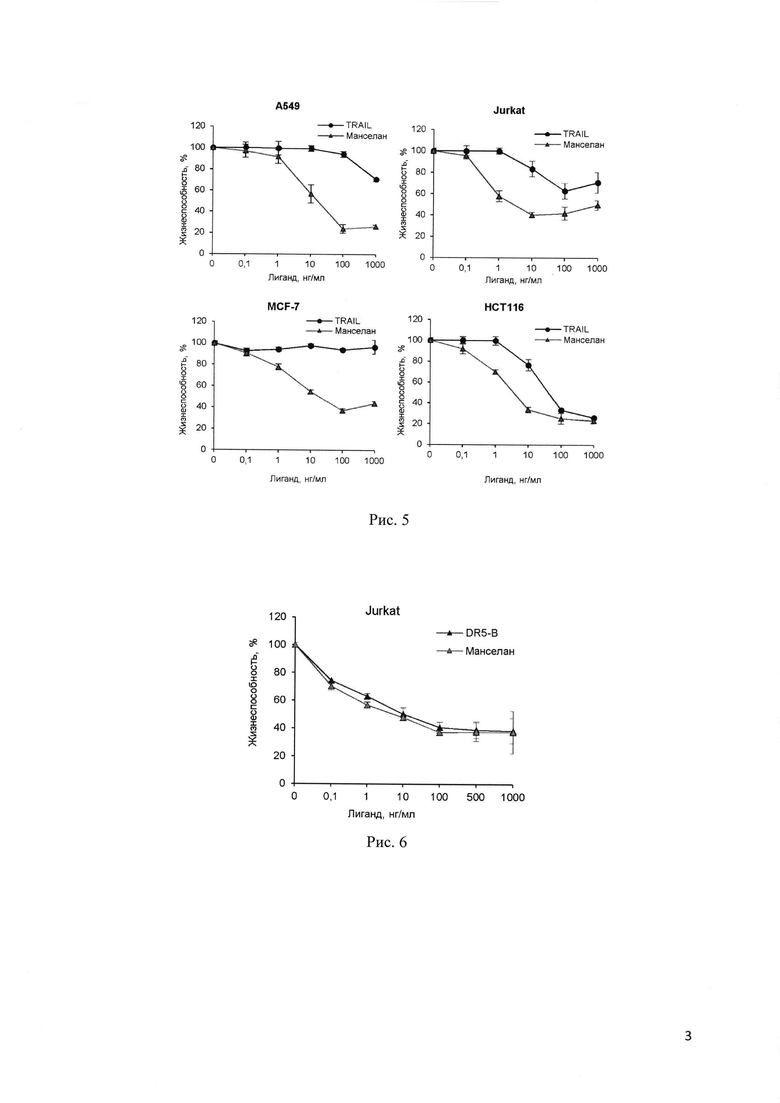
Для эффективной экспрессии растворимого белка Манселана, плазмидный вектор рЕТ-32а/ manselan трансформируют в высокопроизводительный штамм Е. coli SHuffle В. Это относительно новый штамм Е. coli, позволяющий получить высокий уровень цитоплазматической экспрессии активных белков с правильно образованными дисульфидными связями благодаря сниженной активности редуктаз gor и trxB наряду с повышенной экспрессией изомеразы дисульфидных связей DsbC (Lobstein et al. Microbial Cell Factories 2012, 11:56). Экспрессию белка индуцируют 0.25 mM IPTG (изопропил_β-D-1-тиогалактопиранозид) (Рис. 1). На первом этапе целевой белок Манселан очищают из растворимой фракции клеточного лизата с помощью металл-аффинной хроматографии на сорбенте Ni-NTA агарозе. Благодаря наличию нескольких гистидиновых остатков в аминокислотной последовательности белка Манселан аффинно связывается с сорбентом Ni-NTA агарозой даже в отсутствие полигистидинового тага. Данный этап позволяет получить белок, очищенный примерно на 85% (Рис. 2). Для получения высокоочищенного белка Манселан проводят второй этап очистки с помощью катион-обменной хроматографии на сорбенте SP Sepharose в О - 500 тМ линейном градиенте NaCl на установке  (GE Healthcare, Sweden). Для удаления остаточных примесей, дополнительно очищают белок на сорбенте Ni-NTA (Рис. 2,3). Описываемый в изобретении способ позволяет получить высокоочищенный белок Манселан в количестве более 0.2 г из 1 литра клеточной культуры (Таблица 1).
(GE Healthcare, Sweden). Для удаления остаточных примесей, дополнительно очищают белок на сорбенте Ni-NTA (Рис. 2,3). Описываемый в изобретении способ позволяет получить высокоочищенный белок Манселан в количестве более 0.2 г из 1 литра клеточной культуры (Таблица 1).
Гомогенность и тримеризацию полученного препарата Манселан исследуют с помощью аналитического ультрацентрифугирования (Рис. 4). Полученный препарат является высоко гомогенным и состоит на 98% из тримерных молекул.
Определяют уровень эндотоксинов в препарате Манселан, полученном описываемым в изобретении методом. Уровень эндотоксинов в препарате TRAIL DR5-В, полученным путем гибридной экспрессии (Гаспарян М.Э., Яголович А.В., Долгих Д.А., Кирпичников М.П. Способ получения мутантного белка TRAIL человека. Роспатент №2405038 от 27.11.2010), составляет 250 ед./мг. Уровень эндотоксинов в препарате Манселан, полученном описываемым методом прямой экспрессии, на порядок ниже (25 ед./мг) благодаря использованию стадии катион-обменной хроматографии. Исследуют влияние низких концентраций (0.05-0.2%) детергента Тритон Х-100 в ходе очистки на содержание эндотоксинов в препарате. Оптимизируя отмывку сорбента детергентом Triton Х-100 на разных этапах очистки, уровень эндотоксинов снижают с 25 ед./мг до 8 ед./мг. Такое содержание эндотоксинов позволяет применять препарат в биологических и доклинических исследованиях, однако превышает предельно допустимую дозу для использования в клинике (не более 5 ед./кг/час). Поэтому препарат Манселан дополнительно очищают на аффинном сорбенте Pierce High Capacity Endotoxin Removal Resin (Thermo Scientific) с ковалентно-связанным модифицированным ε-поли-L-лизином для удаления эндотоксинов (этот сорбент имеет преимущество перед обычно используемыми сорбентами на основе полимиксина Б благодаря отсутствию токсичности). Итоговое содержание эндотоксинов в очищенном препарате Манселан составляет не более 0.5 ед./мг (Таблица 2).
Для корректной оценки цитотоксической активности белка Манселан аналогичным способом получают высокоочищенный рекомбинантный белок TRAIL дикого типа (114-281), модифицированный аминокислотным остатком метионина на N-конце, используя в качестве основы плазмидный вектор pET-32a/TRAIL, ранее полученный для гибридной экспрессии белка Trx/TRAIL с доменом тиоредоксина в качестве «несущего» белка.
Противоопухолевую активность полученных препаратов Манселана и TRAIL исследуют на линиях опухолевых клеток (Рис. 5). Препараты Манселан и TRAIL, полученные описываемым в изобретении методом прямой экспрессии в штаммах-продуцентах Е. coli SHuffle B/pE-T32a/manselan и Е. coli SHuffle B/pET-32a/trail, индуцируют апоптоз в клетках колоректальной карциномы НСТ-116, аденокарциномы легких А549, Т-клеточной лейкемии Jurkat и карциномы молочной железы MCF-7. Показывают, что Манселан эффективнее вызывает гибель в клеточных линиях по сравнению с TRAIL (Рис. 5). Активность препарата Манселан, полученного путем прямой экспрессии, сравнивают с препаратом DR5-B, полученным ранее описанным методом гибридной экспрессии, в линии Т-клеточной лейкемии Jurkat. Препараты, полученные обоими способами, индуцируют апоптоз с одинаковой эффективностью (Рис. 6).
Техническим результатом заявленного изобретения является способ получения модифицированного противоопухолевого белка Манселана на основе мутантного варианта цитокина TRAIL DR5-B, который заключается в повышении выхода целевого белка и степени очистки от низкомолекулярных белковых примесей и бактериальных эндотоксинов, а также в сокращенном количестве стадий очистки и снижении стоимости получения препарата. Описываемый в изобретении способ позволяет использовать препарат Манселан не только в качестве инструмента для молекулярно-биологических исследований, но и в качестве лекарственного средства при терапии онкологических заболеваний.
Результаты изобретения отражаются в рисунках.
Рис. 1. Экспрессия белка Манселан в штамме E.coli SHuffle В при индукции с индуктором Т7-ого промотора IPTG в различных концентрациях. Пробы, содержащие по 20 мкл ночной культуры, центрифугируют при 5000 × g в течении 5 мин., растворяют в буфере для проб и анализируют на 15% ДСН-ПААГ.
Рис. 2. Прямая экспрессия в штамме E.coli SHuffle В и очистка белка Манселан. Образцы, содержащие по 15 мкг белка, анализируют в 15% трис-глициновом ДСН-полиакриламидном геле после окрашивания краской Кумасси бриллиантовым синий R-250. Линия 1: клеточный лизат, Линия 2: фракция растворимых цитоплазматических белков. Линия 3: фракция белка Манселан после очистки на сорбенте Ni-NTA агарозы. Линия 4: Белок после очистки с помощью ионообменной хроматографии на SP сефарозе. Линия 5: фракция белка после дополнительной очистки на сорбенте Ni-NTA агарозы.
Рис. 3. Анализ чистоты препаратов Манселана и TRAIL. Образцы, содержащие по 0.25 мкг очищенного белка, анализируют в 16.5% трис-глициновом ДСН-полиакриламидном геле после окрашивания нитратом серебра. Линии 1: TRAIL дикого типа. Линия 2: Манселан.
Рис. 4. Анализ аналитического ультрацентрифугирования препарата Манселан. Препарат центрифугируют при 60000 rpm в стандартных 0.4 мл ячейках. Профили белка зарегистрированы после 1, 10, 20, 45, 54, 60, 74 и 86 мин при поглощении 280 нм. Посчитанный коэффициент седиментации s20, w = 5.49±0.03 и средняя молекулярная масса Mw = 59511 кДа свидетельствуют о содержании исключительно глобулярных тримерных молекул в препарате.
Рис. 5. Цитотоксичность рекомбинантных белковых препаратов TRAIL и Манселана на различных опухолевых клеточных линиях. Клетки карциномы прямой кишки НСТ116, аденокарциномы молочной железы MCF-7, карциномы легких А549 и Т-лимфобластной лейкемии Jurkat инкубировали с указанными концентрациями TRAIL или Манселана в течении 24 часов и жизнеспособность определяли с помощью МТТ теста.
Рис. 6. Сравнительный анализ цитотоксичной активности препаратов DR5-B и Манселан. Клетки Т-лимфобластной лейкемии Jurkat инкубируют с указанными концентрациями DR5-B или Манселана в течении 24 часов и жизнеспособность определяли с помощью МТТ теста.
Изобретение иллюстрируется примерами.
Пример 1. Создание генетической конструкции для прямой экспрессии модифицированного белка DR5-B - Манселана.
На первом этапе в последовательности слитного белка Trx/DR5-B (SEQ ID NO 1) в составе плазмидного вектора pET32a/DR5-B (ранее полученного авторами Gasparian ME, Chernyak BV, Dolgikh DA, Yagolovich AV, et al. Apoptosis, 2009, 14:778-787) вводят 3 точечные мутации (G469C, C471T, A473T) методом сайт-специфического мутагенеза, основанного на полимеразной цепной реакции. Реакционная смесь для полимеразной цепной реакции содержит буфер с MgSO4 для ДНК-полимеразы Pfu (Fermentas, Литва), ДНК-полимеразу Pfu, смесь из 4-х дезоксинуклеотидтрифосфатов, плазмидную ДНК pET-32a/DR5-B, прямой (SEQ ID NO 2) и обратный (SEQ ID NO 3) олигонуклеотидные праймеры. Поверх реакционной смеси наслаивают минеральное масло. Проводят 30 циклов амплификации при следующих условиях: 10 секунд при 95°С, 30 секунд при 95°С, 30 секунд при 55°С, 14.5 минут при 72°С.Синтез ДНК проверяют с помощью электрофореза в 1% агарозном геле. Для расщепления исходной метилированной плазмидной ДНК реакционную смесь обрабатывают рестриктазой Dpn I в течение 1 часа при 37°С. Для наработки мутантной ДНК проводят трансформацию бактериального штамма Escherichia coli XL-1 Blue реакционной смесью ПЦР. Выращивают клетки в среде LB с ампициллином (100 мкг/мл) при 37°С и перемешивании 250 об/мин в течение 18 часов, после чего выделяют плазмидную ДНК из клеточной биомассы. Наличие мутаций подтверждают определением нуклеотидной последовательности мутантной ДНК. Таким образом вводят сайт рестрикции NdeI перед началом гена DR5-B.
На втором этапе обрабатывают плазмиду pET32a/DR5-B с дополнительным сайтом рестрикции NdeI рестриктазами XhoI и NdeI. В результате образуются 4 фрагмента: ДНК линеаризованного плазмидного вектора (5445 п.н.), фрагмент гена DR5-B (504 п.н.), а также фрагмент с геном тиоредоксина (345 п.н.) и фрагмент тага (129 п.н.) (так как в нем находится еще один сайт рестрикции NdeI). Фрагменты разделяют в 1% агарозном геле, после чего фрагменты ДНК линеаризованного плазмидного вектора и гена DR5-B элюируют из геля с помощью набора QuiaQuick Gel Extraction kit (Quiagen) и лигируют с помощью Т4 ДНК лигазы при 10°С в течение 16 часов. Реакционную смесь трансформируют в штамм Escherichia coli XL-1 Blue и выделяют плазмидную ДНК. Правильную вставку гена в плазмиду подтверждают определением нуклеотидной последовательности встроенного гена. Результатом проделанной работы является получение плазмидного вектора pET32a/manselan, который не содержит Trx-, His- и S-тагов и служит для прямой экспрессии модифицированного белка DR5-B - Манселана - благодаря внесенному аминокислотному остатку метионина на N-конце аминокислотной последовательности белка (SEQ ID NO 4).
Для корректной оценки цитотоксической активности белка Манселан аналогичную процедуру проводят с плазмидным вектором pET-32a/TRAIL, экспрессирующим цитокин TRAIL дикого типа. Результатом проделанной работы является плазмидный вектор pET-32a/trail для прямой экспрессии белка TRAIL дикого типа, который не содержит Trx-, His- и S-тагов и служит для прямой экспрессии белка TRAIL без тагов благодаря внесенному аминокислотному остатку метионина на N-конце (SEQ ID NO 5).
Пример 2. Экспрессия и очистка рекомбинантного белка Манселана.
Экспрессия модифицированного белка Манселана проводится в бактериальном штамме Е. coli SHuffle В. Клетки трансформируют плазмидным вектором рЕТ-32a/manselan и получают экспрессирующий штамм продуцент Е. coli SHuffle В/рЕТ-32a/manselan. Трансформированные клетки выращивают в жидкой питательной среде LB с ампициллином (100 мкг/мл) при 37°С и интенсивном перемешивании (250 об/мин) в течение 17 часов. После этого клетки инокулируют в жидкую среду ТВ с ампициллином (100 мкг/мл) разбавляя 1:100, растят при 37°С до оптической плотности ОД600=0,6 и индуцируют экспрессию целевого белка добавлением индуктора Т7-ого промотора IPTG (изопропил_β-D-1-тиогалактопиранозид). Культуру клеток выращивают при 28°С в течение 20 часов. Уровень экспрессии целевого белка анализируют в 15% полиакриламидном геле в присутствии додецилсульфата натрия (Рис. 1). После этого клетки осаждают центрифугированием при 5000 × g при 4°С в течение 10 минут, ресуспендируют в буфере, содержащем 300 mM NaCl, 50 мМ Tris-HCl (рН 8.0) и разрушают под давлением 2000 psi на установке «French press» (Spectronic Instruments, Inc., США). Разрушенную клеточную массу осаждают центрифугированием при 25000 об/мин в течение 25 мин.
На первом этапе очистки растворимую клеточную фракцию наносят на хроматографическую колонку с сорбентом Ni-NTA агарозой (Qiagen) предварительно уравновешенную в буфере для разрушения клеток. Колонку промывают буфером, содержащим 300 мМ NaCl, 50 мМ NaH2PO4 20 mM имидазола, 0.1% Triton Х-100. Целевой белок Манселан элюируют в буфере, содержащем 300 мМ NaCl, 50 мМ NaH2PO4, 250 мМ имидазола (рН 8.0). Очищенный препарат диализуют против буфера, содержащего 45 мМ NaCl, 50 мМ Tris-HCl, 5 мМ β-МЕ (β-меркаптоэтанол), 0.02% NaN3 (рН 7.5) при комнатной температуре в течение 18 часов. На втором этапе белок очищают с помощью катион-обменной хроматографии на сорбенте SP Sepharose (GE Healthcare) в 0-500 mM линейном градиенте NaCl на установке (GE Healthcare, Sweden). Перед нанесением белка на колонку к раствору белка добавляют 0.05% Triton Х-100. На третьем этапе белок дополнительно очищают на сорбенте Ni-NTA агароза. Целевой белок Манселан элюируют с помощью буфера, содержащего 300 мМ NaCl, 50 мМ NaH2PO4, 250 мМ имидазола (рН 8.0). Очищенный белок диализуют против буфера, содержащего 150 мМ NaCl (рН 7.5) в течении 18-20 часов при 4°°С. Полученный высокоочищенный препарат Манселан стерилизуют фильтрацией через стерильный шприцевой фильтр (Millipore, США) с порами 0,2 мкм с мембраной поливинил-дифторид (PVDF) и хранят при 4°С. Пробы после каждой стадии очистки анализируют в полиакриламидном геле при окрашивании краской Кумасси бриллиантовой синий R250 (Рис. 2) или нитратом серебра (Рис. 3). Аналогичную процедуру проводят для цитокина TRAIL дикого типа.
Выходы белковых препаратов Манселан и TRAIL приведены в Таблице 1.

Гомогенность и тримеризацию полученного препарата Манселан исследуют с помощью аналитического ультрацентрифугирования. Показывают, что очищенный препарат Манселан является высоко гомогенным и состоит на 98% из тримерных молекул (Рис. 4).
Пример 3. Очистка препарата Манселан от эндотоксинов.
После аффинной и ионообменной хроматографии препараты Манселан и TRAIL дополнительно очищают на аффинном сорбенте Pierce High Capacity Endotoxin Removal Resin (Thermo Scientific). Каждые 50 мл образца инкубируют с 10 мл сорбента в течение 2 часов при комнатной температуре при медленном помешивании. Затем смесь белков с сорбентом наносят на колонку и собирают белковую фракцию без сорбента.
Для количественной оценки содержания эндотоксинов используют набор ToxinSensor™ Chromogenic LAL Endotoxin Assay Kit (USA). Содержание эндотоксинов в препарате оценивают спектрофотометрически по поглощению при 545 нм. Серийные разведения белков Манселан и TRAIL проводят в воде для ЛАЛ-теста. В качестве отрицательного контроля используют воду для ЛАЛ-теста, в качестве положительного контроля используют стандартные количества эндотоксинов, входящих в набор реактивов. Содержание эндотоксинов в препарате определяют графически по калибровочной кривой (Табл. 2).


Пример 4. Исследование биологической активности препарата на линиях опухолевых клеток.
Линии клеток колоректальной карциномы НСТ116 и карциномы легких А549 карциномы культивируют в жидкой питательной среде DMEM, содержащей 10% (по объему) бычьей эмбриональной сыворотки. Линию клеток аденокарциномы молочной железы MCF7 культивируют в жидкой питательной среде ЕМЕМ, содержащей 10 мкг/мл рекомбинантного человеческого инсулина (Sigma) и 10% бычьей эмбриональной сыворотки. Линию Т-клеточной лейкемии Jurkat культивируют в жидкой питательной среде RPMI 1640, содержащей 5% бычьей эмбриональной сыворотки. Все линии клеток культивируют при 37°С и 5% СО2. Разведения белков TRAIL, DR5-B и Манселан производят в культуральной среде. Для определения количества живых клеток клетки рассаживают на 96-луночные планшеты (1×105 клеток/лунку) и инкубируют с серийными разведениями препаратов TRAIL, DR5-B и Манселан в течение 24 часов при 37°С и 5% СО2. После этого добавляют реагент МТТ (3-[4,5-диметилтиазол-2]-2,5-дифенил тетразолия бромид) в окончательной концентрации 0.5 мг/мл и инкубируют в течении 3 часов при 37°С. Планшеты центрифугируют при 3000 об/мин в течение 5 мин, удаляют надосадок и в каждую лунку добавляют ДМСО для растворения кристаллов формазана. Количество живых клеток определяют путем измерения поглощения раствора формазана в ДМСО при 540 нм на установке BioRad 680 (USA). Показывают, что Манселан эффективнее вызывает гибель всех тестированных клеточных линиях по сравнению с TRAIL (Рис. 5). Активность препарата Манселан, полученного описываемым методом прямой экспрессии, сравнивают с препаратом DR5-B, полученным ранее описанным методом гибридной экспрессии в линии Т-клеточной лейкемии Jurkat. Показывают, что препараты, полученные обоими способами, индуцируют апоптоз с одинаковой эффективностью (Рис. 6).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОГО БЕЛКА TRAIL ЧЕЛОВЕКА | 2009 |
|
RU2405038C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET-32a, КОДИРУЮЩАЯ ГЕН ЛИГАНД-СВЯЗЫВАЮЩЕГО ДОМЕНА РЕЦЕПТОРА II ТИПА ТРАНСФОРМИРУЮЩЕГО ФАКТОРА РОСТА-β ЧЕЛОВЕКА (TβRII), ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ СЛИТНОГО БЕЛКА ТИОРЕДОКСИН/TβRII И СПОСОБ РЕНАТУРАЦИИ И ОЧИСТКИ ЦЕЛЕВОГО БЕЛКА TβRII | 2009 |
|
RU2413769C1 |
| СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ | 2011 |
|
RU2499052C2 |
| Многофункциональные гибридные рекомбинантные белковые препараты для терапии опухолевых заболеваний | 2022 |
|
RU2801367C1 |
| Белковая конструкция на основе рецептор-специфичного мутантного варианта противоопухолевого цитокина TRAIL в виде гибридного белка с пептидом, специфичным к интегрину αvβ3, для терапии солидных опухолей | 2022 |
|
RU2802488C1 |
| Комбинированные наночастицы на основе амфифильных сополимеров N-винилпирролидона для совместной таргетной доставки лекарственных препаратов | 2023 |
|
RU2830606C2 |
| Способ очистки рекомбинантного белка, содержащего в своем составе последовательности миелопептидов | 2016 |
|
RU2630302C1 |
| ПЛАЗМИДНЫЙ ВЕКТОР pE-Trx-Aur, ШТАММ ESCHERICHIA COLI ДЛЯ ЭКСПРЕССИИ АНТИМИКРОБНОГО ПЕПТИДА АУРЕЛИНА И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ПЕПТИДА | 2009 |
|
RU2412999C1 |
| Способ получения рекомбинантного бруцеллезного белка rLptD | 2023 |
|
RU2815099C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTRX-TEVRS-РТН, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СПОСОБНЫЙ К ПРОТЕОЛИТИЧЕСКОМУ РАСЩЕПЛЕНИЮ С ОБРАЗОВАНИЕМ ФРАГМЕНТА ЭНДОГЕННОГО ЧЕЛОВЕЧЕСКОГО ПАРАТИРЕОИДНОГО ГОРМОНА (1-34), ШТАММ ESCHERICHIA COLI BL21(DE3)/PTRX-TEVRS-РТН - ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО PTH (1-34) | 2019 |
|
RU2700452C1 |




Изобретение относится к области биотехнологии, конкретно к рекомбинантному получению терапевтических белков, и может быть использовано для получения рекомбинантного противоопухолевого белка DR5-B в Е. coli. Способ предусматривает трансформацию клеток штамма Е. coli SHuffle В полученной рекомбинантной плазмидной ДНК рЕТ-32а без Trx-, His- и S-тагов, кодирующей DR5-B с последующей экспрессией, выделением и очисткой целевого белка. Изобретение позволяет получить модифицированный противоопухолевый белок Манселан на основе мутантного варианта цитокина TRAIL DR5-B с высоким выходом и степенью очистки от низкомолекулярных белковых примесей и бактериальных эндотоксинов. 2 табл., 6 ил., 4 пр.
Способ получения рекомбинантного противоопухолевого белка DR5-B, модифицированного аминокислотным остатком метионина на N-конце с SEQ ID NO 4, ген которого находится в составе плазмидной ДНК рЕТ-32а без Trx-, His- и S-тагов с последующей трансформацией штамма Е. coli SHuffle В полученной рекомбинантной плазмидной ДНК, прямой экспрессией, выделением и очисткой целевого белка.
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОГО БЕЛКА TRAIL ЧЕЛОВЕКА | 2009 |
|
RU2405038C1 |
| СПОСОБ ИНДУКЦИИ ГИБЕЛИ ОПУХОЛЕВЫХ КЛЕТОК | 2015 |
|
RU2620165C2 |
| GASPARIAN M.E., et al., Generation of new TRAIL mutants DR5-A and DR5-B with improved selectivity to death receptor 5, 2009, Apoptosis, v.14, n.6, p.778-787 | |||
| US 9403892 B2, 02.08.2016. | |||

