ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к способу определения гаплотипа нуклеиновой кислоты-мишени, имеющей по меньшей мере два гетерозиготных полиморфизма, и набору праймеров, используемому для такого определения.
УРОВЕНЬ ТЕХНИКИ
Гаплотип представляет собой цис-положение двух или больше полиморфизмов, расположенных на одной хромосоме. Полагают, что информация о гаплотипе очень важна, поскольку гаплотип связан с определенными заболеваниями или аномалиями и специфической чувствительностью к лекарственным средствам. До сих пор для анализа гаплотипа требовалось изучение генетической информации относительно полиморфизмов, сопровождающих несколько поколений одной семьи, и компьютеризованные алгоритмы оценки. В то же время достижения в методике полимеразной цепной реакции (ПЦР) обеспечили прямой анализ ДНК на молекулярном уровне, и было предложено применение результатов такого анализа для определения гаплотипа. Тем не менее, для определения гаплотипа с использованием ПЦР необходимо применение большого числа сочетаний праймеров, специфичных для аллельных генов, и многократное проведение ПЦР. При рассмотрении лишь 2 аллельных генов были предприняты попытки определить гаплотип путем расположения 2 пар праймеров на полиморфных сайтах и оценкой, пройдет ли какая-либо амплификация. Этот способ был предложен для определения гаплотипа с помощью единого лабораторного анализа (см. японскую выложенную заявку на патент № 2002-272482). Конкретно, прямой праймер, содержащий первый полиморфизм, модифицируют с помощью метки, которая дает различные сигналы в зависимости от типа аллеля на 5'-конце. Кроме того, в отношении обратного праймера, содержащего второй полиморфизм, к 5'-концу присоединяют клапанную последовательность (flap sequence), которая отсутствует в последовательности-мишени, таким образом, что праймер предназначен для получения ампликона, имеющего длину, которая зависит от аллеля. Используя эти праймеры, осуществляют анализ аллель-специфичной ПЦР в обоих направлениях. Ту пару праймеров, которая приведет к амплификации, можно узнать, определив флуоресцентную метку в ПЦР-ампликоне, и исходя из длины ампликона, и таким образом можно определить гаплотип.
В вышеописанном известном способе, тем не менее, существенно, что праймеры содержат сайты с полиморфными последовательностями. Это ограничивает свободу выбора последовательности оснований праймеров. В данном случае для проведения аллель-специфичной ПЦР Tm праймеров, используемых для проведения ПЦР, должны быть аналогичны. Тем не менее, из-за вышеописанных ограничений в выборе последовательностей оснований праймеров в некоторых случаях может быть затруднительно сделать Tm праймеров аналогичными. Другими словами, с помощью способа, описанного в японской выложенной патентной заявке № 2002-272482, высокоточное определение гаплотипа может быть ограничено случаями, когда полиморфизмы находятся в участках, для которых могут быть использованы праймеры с аналогичной Tm. Кроме того, в этом способе определения затруднительно смещать положения последовательностей как прямого, так и обратного праймеров. Таким образом, если прямой и обратный праймеры образуют димер праймеров, определение гаплотипа становится невозможным.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу определения гаплотипа с большей точностью независимо от расположения полиморфизмов. Конкретнее, изобретение относится к способу определения гаплотипа нуклеиновой кислоты-мишени, имеющей по меньшей мере два гетерозиготных полиморфизма, и набору праймеров, используемому для этой цели.
Способом по настоящему изобретению определяют гаплотип нуклеиновой кислоты-мишени, имеющей первый гетерозиготный полиморфный сайт на 3'-стороне и второй гетерозиготный полиморфный сайт на 5'-стороне, и способ включает:
(i) осуществление ПЦР нуклеиновой кислоты-мишени с использованием одного из прямых праймеров (а-1), т.е.
(а-1-1) прямой праймер, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой мутантный тип, и не удлиняет нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой дикий тип, и
(а-1-2) прямой праймер, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой дикий тип, и не удлиняет нуклеиновую кислоту-мишень, если этот первый полиморфизм представляет собой мутантный тип, и
(b-1) обратный праймер, который содержит последовательность оснований, комплементарную определенной последовательности оснований, которая не содержит второй полиморфизм, и располагается на 3'-стороне второго полиморфизма на комплементарной цепи нуклеиновой кислоты-мишени; и
(ii) гибридизацию продукта, полученного в результате (i), с первым зондом, который имеет последовательность оснований, идентичную определенному сегменту последовательности оснований нуклеиновой кислоты-мишени, который содержит нуклеотид дикого типа второго полиморфизма, и вторым зондом, который имеет последовательность оснований, идентичную определенному сегменту последовательности оснований нуклеиновой кислоты-мишени, который содержит мутантный нуклеотид второго полиморфизма, и определение сигналов образования гибрида с первым и вторым зондами.
Набор праймеров, используемый для определения гаплотипа по настоящему изобретению, представляет собой набор праймеров для определения гаплотипа нуклеиновой кислоты-мишени, имеющей первый полиморфный сайт на 3'-стороне и второй полиморфный сайт на 5'-стороне, и набор праймеров отличается наличием одного из прямых праймеров, т.е.
прямого праймера, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой мутантный тип, и не удлиняет нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой дикий тип, и
прямого праймера, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой дикий тип, и не удлиняет, если первый полиморфизм представляет собой мутантный тип, и
обратного праймера, который содержит последовательность оснований, комплементарную определенной последовательности оснований, которая не содержит второй полиморфизм, и располагается на 3'-стороне второго полиморфизма на комплементарной цепи нуклеиновой кислоты-мишени.
Как описано выше, с помощью настоящего изобретения можно одновременно и исчерпывающе определять гаплотип, если присутствует большое число гетерозиготных полиморфизмов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 представляет собой графическую схему определения гаплотипов двух аллельных генов по первому варианту осуществления настоящего изобретения;
Фиг.2 представляет собой диаграмму, которая иллюстрирует положение двух аллельных генов по первому варианту осуществления настоящего изобретения;
Фиг.3 представляет собой диаграмму, которая иллюстрирует протокол, используемый для проведения ПЦР-амплификации для получения специфических сегментов генома по первому варианту осуществления настоящего изобретения;
Фиг.4 представляет собой диаграмму, которая иллюстрирует протокол, используемый для проведения ПЦР-амплификации по первому варианту осуществления настоящего изобретения;
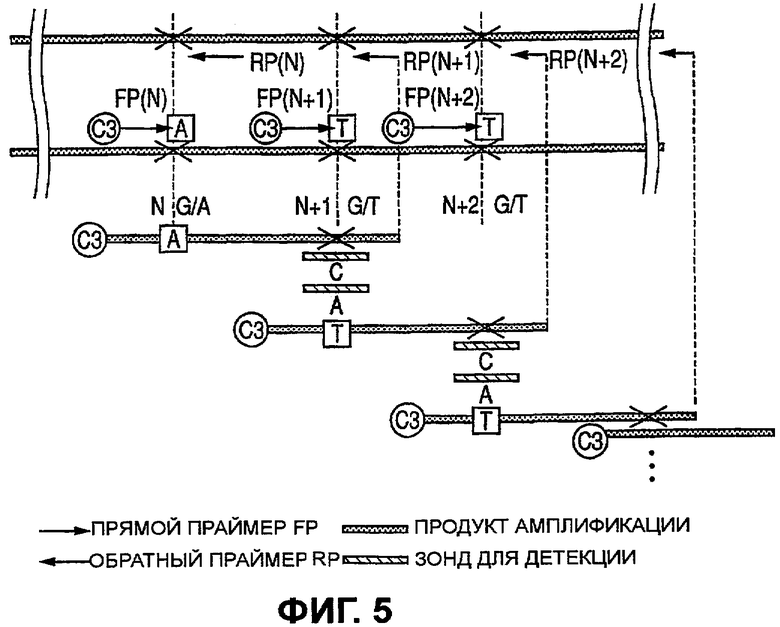
Фиг.5 представляет собой диаграмму, которая иллюстрирует положение двух или больше аллельных генов по второму варианту осуществления изобретения.
ЛУЧШИЕ СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В настоящем изобретении для определения гаплотипа с помощью микрочипа используют схему аллельной ген-специфической ПЦР-амплификации в отношении представляющих интерес полиморфизмов в геномной ДНК-мишени.
Сначала получают ампликон, который специфичен для одного из аллелей первого аллельного гена, а также содержит второй аллельный ген. Для идентификации аллеля второго аллельного гена этот ампликон затем гибридизуют с олигонуклеотидными зондами, каждый из которых специфичен к одному из аллелей второго аллельного гена. Эта методика может быть очень эффективным способом определения на молекулярном уровне, если гаплотип образуют первый и второй аллельные гены. Получают зонд к одной комплементарной цепи, который образует гибрид ДНК, если второй аллельный ген представляет собой мажорный аллель, и зонд к другой комплементарной цепи, который образует гибрид ДНК, если второй аллельный ген представляет собой минорный аллель, и фиксируют на твердой подложке, такой как микрочип. Гаплотип может быть определен выявлением с помощью метки зонда, с которым наблюдается гибридизация. Если существует большое число пар кандидатных аллельных генов, то N-ное количество гаплотипов может быть исчерпывающе проанализировано гибридизацией с Nx2 зондами, загруженными на твердую подложку. Если первый аллельный ген представляет собой предварительно выбранный ген в гетерозиготном состоянии, то для идентификации гаплотипов могут быть получены особые твердые подложки. Один из праймеров для проведения амплификации может быть сконструирован таким образом, что он станет специфичным для одного из аллелей первого аллельного гена, а другой праймер для проведения амплификации должен быть сконструирован так, чтобы включить в ампликон другие аллельные гены. Метка может представлять собой, например, флуорофор, введенный в праймер и сконструированный таким образом, чтобы можно было детектировать флуоресценцию в зонде, гибридизующемся с другим аллельным геном лишь в том случае, если аллель-специфическая амплификация прошла успешно.
Для получения ампликона, имеющего область, специфичную к одному из аллелей первого аллельного гена, по настоящему изобретению используют один из прямых праймеров, описанных ниже.
(а-1-1) Прямой праймер, который содержит последовательность оснований, комплементарную описанному выше первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой мутантный тип, и не удлиняет, если первый полиморфизм представляет собой дикий тип.
(а-1-2) Прямой праймер, который содержит последовательность оснований, комплементарную описанному выше первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой дикий тип, и не удлиняет, если первый полиморфизм представляет собой мутантный тип.
В этих прямых праймерах последовательность оснований праймера (и/или длина последовательности) может быть установлена стандартным способом на основе последовательности сегмента, содержащего известный первый полиморфизм, исходя из типирования нуклеиновой кислоты-мишени таким образом, чтобы этот сегмент был бы получен в ПЦР-ампликоне.
Может быть выбран полиморфизм либо мутантного типа, либо дикого типа.
Чтобы получить обратный праймер, в ампликоне конструируют сегмент, содержащий второй полиморфизм, для получения последовательности оснований, комплементарной определенной последовательности оснований, которая не содержит второй полиморфизм и располагается на 3'-стороне второго полиморфизма на цепи, комплементарной нуклеиновой кислоты-мишени. Нуклеотидная последовательность (длина последовательности) обратного праймера может быть установлена стандартным способом, используя за основу последовательность сегмента, содержащего известный второй полиморфизм, исходя из типирования нуклеиновой кислоты-мишени и таким образом, чтобы этот сегмент мог быть получен в ПЦР-ампликоне.
В отношении зондов, используемых для детекции сегмента, содержащего также второй полиморфизм, может быть выбрана последовательность, специфичная для этого сегмента, и последовательность оснований (и/или длина последовательности) зонда, который может определить выбранную последовательность, может быть установлена стандартным способом, поскольку сегмент, содержащий второй полиморфизм, уже известен из типирования нуклеиновой кислоты-мишени.
Ниже описываются варианты осуществления настоящего изобретения.
Первый вариант осуществления
Ниже описывается первый вариант осуществления настоящего изобретения со ссылкой на фиг.1. В этом варианте осуществления описывается методика определения типов аллелей множественных аллельных генов в геномной ДНК. Конкретно, далее последовательно описывается способ определения гаплотипа из 2 или больше чем 2 аллельных генов.
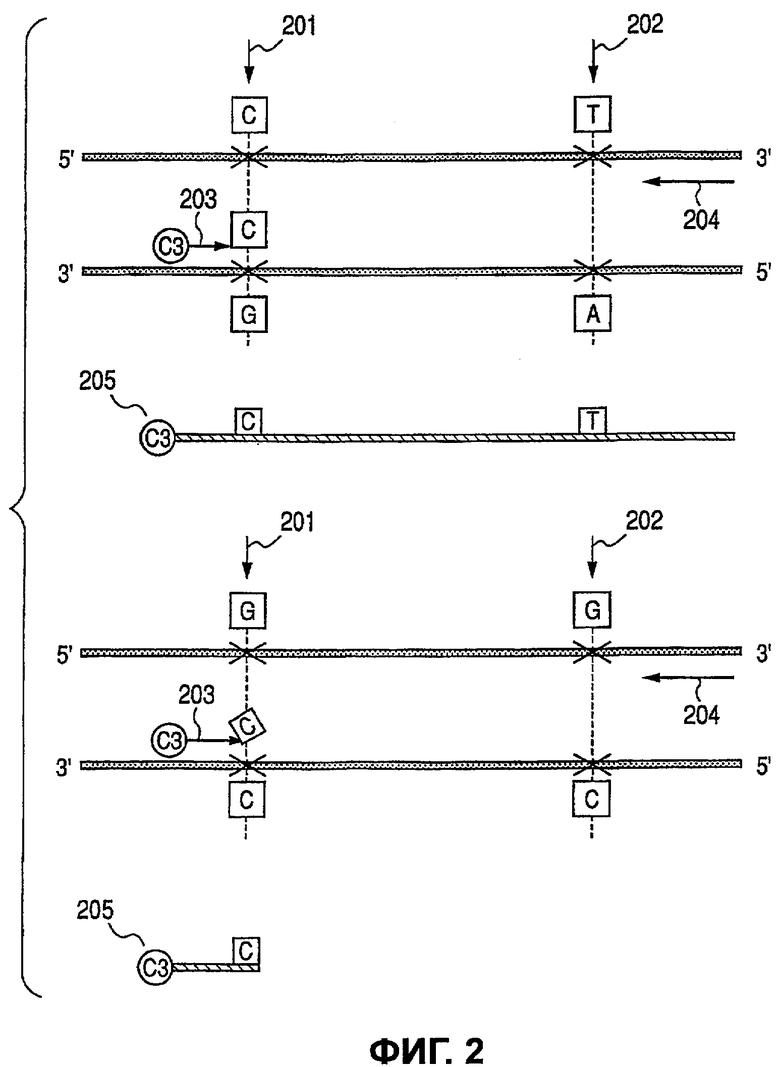
В этом варианте осуществления определяют гаплотипы двух гетерозиготных аллельных генов в геномной ДНК. Расстояние между двумя аллельными генами может находиться в интервале от около нескольких пар оснований до нескольких десятков пар оснований вплоть до нескольких сотен и даже нескольких тысяч пар оснований. Поэтому необходимо получить ампликон, имеющий оба аллельных гена. Два аллельных гена определяются в данном описании как первый аллельный ген и второй аллельный ген, считая от 5'-стороны. Мажорный аллель первого аллельного гена обозначается G, а минорный аллель обозначается С, и мажорный аллель второго аллельного гена обозначают G, а минорный аллель обозначают Т.
Для определения гаплотипа в этом варианте осуществления 3'-конец праймера подбирают в соответствии с локусом аллеля для избирательной амплификации либо мажорного, либо минорного аллеля первого аллельного гена. Например, первый праймер (прямой праймер) конструируют так, чтобы он был комплементарен минорному аллелю первого аллельного гена на его 3'-конце, чтобы удлинение происходило, если первый аллельный ген представляет собой минорный аллель, и удлинение не продолжалось бы, если бы это был мажорный аллель. С другой стороны, второй праймер (обратный праймер) противоположной цепи конструируют так, чтобы он затрагивал второй аллельный ген, используя в качестве контрольной точки позицию связывания первого праймера. Второй праймер не зависит от типа аллеля. При таком положении, если первый аллельный ген представляет собой минорный аллель, получают большое количество ампликона, что подтверждает тот факт, что первый аллельный ген представляет собой минорный аллель, и предпочтительно был получен ампликон, содержащий второй аллельный ген. Поэтому определив тип аллеля второго аллельного гена в ампликоне, возможно определить гаплотип первого и второго аллельных генов.
Способ определения гаплотипа по настоящему изобретению включает стадии 1) и 2), описанные ниже:
отбор одного из аллелей первого аллельного гена и получение ампликона, содержащего лишь второй аллельный ген;
гибридизация ампликона с комплементарными зондами, специфичными для аллелей второго аллельного гена, и, таким образом, идентификация аллеля второго аллельного гена.
Фиг.1 представляет собой графическую схему, иллюстрирующую описанные выше стадии. S-1 является стадией, подтверждающей, что как первый, так и второй аллельные гены находятся в гетерозиготном состоянии. В отношении способа типирования, используемого для каждого аллельного гена, ограничения не существует. Если по меньшей мере один (первый или второй) аллельный ген находится в гомозиготном состоянии, то становится известна комбинация гаплотипа двух аллельных генов и нет необходимости использовать настоящее изобретение.
В данном варианте осуществления нацеливаются лишь на комбинации, где гетерозиготность была подтверждена после каждого генотипирования. S-2 представляет собой стадию, в которой ампликон получают из генома-мишени путем сочетанного применения первого праймера, специфичного к минорному аллелю первого аллельного гена, и второго праймера противоположной цепи для включения второго аллельного гена. Первый праймер, специфичный к минорному аллелю первого аллельного гена, представляет собой праймер, в котором мутантный сайт располагается на его 3'-конце таким образом, что он комплементарен и праймер не приводит к удлинению, если мишенная геномная последовательность содержит мажорный аллель. В данном случае, поскольку первый аллельный ген находится в гетерозиготном состоянии, реакция удлинения продолжается лишь тогда, когда присутствует геном минорного аллеля и первый праймер отжигается. Другими словами, образуемый ампликон ограничивался бы ампликоном, содержащим минорный аллель первого аллельного гена. Второй праймер не зависит от генома-мишени, и будет продуцироваться ампликон, содержащий второй аллельный ген, соответствующий его конфигурации в геноме-мишени. В этом случае, когда первый аллельный ген представляет собой минорный аллель, образуется ампликон, содержащий соответствующий аллель второго аллельного гена. Представляет ли второй аллельный ген мажорный или минорный аллель, не имеет никакого значения, но гаплотип с первым аллельным геном может быть определен гибридизацией ампликона с зондами, специфичными к аллелям второго аллельного гена. Сущность настоящего изобретения не меняется, даже если первый праймер конструируют так, чтобы он был специфичен для мажорного аллеля, в отличие от описанного выше случая.
Для определения второго аллельного гена также необходимо использовать метку. Мечеными могут быть либо первый, либо второй праймер. Альтернативно, во время реакции удлинения в ампликон встраивают меченый нуклеотид. Метка может представлять собой флуорофор, хемилюминесцентное вещество или радиоизотоп. В данном варианте осуществления используют метку, представляющую собой флуоресцентный краситель Су3. S-3 представляет собой стадию, когда ампликон оставляют гибридизоваться с твердой подложкой, на которой были фиксированы зонды, специфичные к второму аллельному гену. Получают зонды, каждый из которых специфичен к мажорному или минорному аллелю. Эти зонды могут быть зафиксированы на твердой подложке или использованы в положении, которое вызвало бы FRET в ходе реакции в жидкой фазе. В данном варианте осуществления второй аллельный ген определяют, используя твердую подложку. Иначе говоря, если флуоресценция наблюдается лишь в определенном участке на твердой подложке, где располагается зонд, специфичный для минорного аллеля второго аллельного гена, это означает, что первый и второй аллельные гены образуют минор/минорный гаплотип. В то же время это также подтверждает существование мажор/мажорного гаплотипа. С другой стороны, если флуоресценция наблюдается лишь в определенном участке на твердой подложке, где располагается зонд, специфичный для мажорного аллеля второго аллельного гена, это означает, что первый и второй аллельные гены образуют минор/мажорный гаплотип, что одновременно подтверждает существование мажор/минорного гаплотипа. Более того, если флуоресценция наблюдается в обоих сайтах на твердой подложке, где располагаются два зонда, один из которых специфичен для мажорного аллеля, а другой - для минорного аллеля второго аллельного гена, это должно быть интерпретировано как тот факт, что первый и второй аллельные гены имеют слабую ассоциацию в гаплотипе. Таким образом, S-4 представляет собой стадию, на которой определяют аллели второго аллельного гена и гаплотип первого и второго аллельных генов.
Фиг.2 представляет собой диаграмму, иллюстрирующую взаимное расположение описанных выше первого аллельного гена 201 и второго аллельного гена 202 в геноме-мишени. На диаграмме также показаны первый праймер 203 и второй праймер 204 и их ампликон 205. Как описано ранее, 3'-конец первого праймера 203 соответствует мутантному сайту первого аллельного гена 201, а второй праймер 204 находится ниже области, которая содержит второй аллельный ген 202. Исходя из первого праймера 203 получение продукта реакции удлинения, который содержит один из аллелей первого аллельного гена 201 (верхняя часть Фиг.2), больше, чем получение продукта реакции удлинения, содержащего другой аллель (нижняя часть Фиг.2). Иначе говоря, с помощью ПЦР амплифицируется лишь та часть генома-мишени, которая содержит единственный из двух аллелей первого аллельного гена 201. Поэтому второй аллельный ген 202 также будет иметь аллель, соответствующий лишь той же части генома-мишени. Кроме того, зонды конструируют так, чтобы они содержали аллели второго аллельного гена 202, показанные на диаграмме.
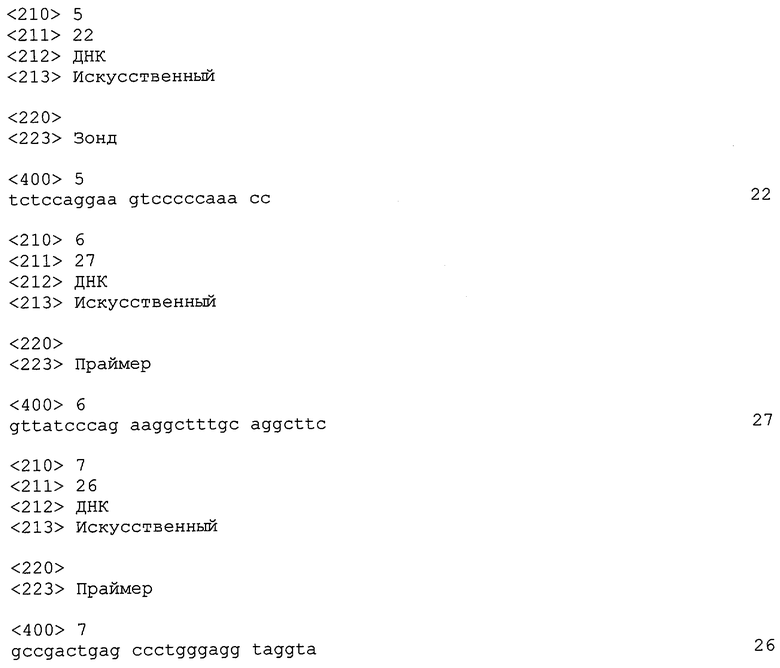
В таблице 1 представлены последовательности оснований пары амплификационных праймеров, их ампликона и зондов для второго аллельного гена, все они относятся к этому варианту осуществления. Начиная с 5'-стороны жирный шрифт соответствует аллельному локусу (мутантный нуклеотид С) первого аллельного гена и аллельному локусу (мутантный нуклеотид Т) второго аллельного гена в этом порядке. Для прямого праймера (FP) был выбран мутантный нуклеотид С.
CATGGATGGT
GGGTGAAACCACAGGCTGGACCAGAAGCCAGGCTGAGAA
GGGGAAGCAG
GTTTGGGGGACTTCCTGGAGAAGGGCATTTATACATGGCAT
GAAGGACTG
GATTTTCCAAAGGCCAAGGAAGAGTAG-3'
Второй вариант осуществления
Ниже описывается второй вариант осуществления изобретения со ссылкой на Фиг.5. Полиморфный локус каждого аллельного гена помечают с помощью «×». Здесь иллюстрируются лишь полиморфизмы в локусах N, N+1 и N+2. Другие опускаются, что не предполагает каких-либо ограничений в количестве полиморфизмов. В данном случае меченым с помощью Су3 является прямой праймер, но существенные признаки изобретения, конечно, не изменились бы, даже если бы им оказался обратный праймер или сама удлиняющаяся нуклеотидная цепь, которую пометили.
Для данного варианта осуществления авторам изобретения представляется целесообразным описать общую методику определения гаплотипа большого числа аллельных генов. В данной работе тестируют количество гетерозиготных полиморфизмов N. Как можно видеть на Фиг.5, определяют FP (N) (прямой праймер для N-ого полиморфизма) и RP (N) (обратный праймер для противоположного направления). Для N+1 далее также авторы изобретения могут аналогичным образом определить прямые праймеры FP (N+1), FP (N+2), … и обратные праймеры RP (N+1), RP (N+2), …. Для получения ПЦР-ампликона, содержащего N-й полиморфизм и N+1-й полиморфизм, конструируют прямой праймер FP (N) и обратный праймер RP (N+1). FP (N) конструируют таким образом, чтобы на его 3'-конце иметь N-й полиморфизм с одним из двух выбранных аллелей. RP (N+1) конструируют на противоположной стороне N-го полиморфизма таким образом, чтобы ПЦР-ампликон содержал бы N+1-й полиморфизм. Пара праймеров FP (N) и RP (N+1) представляет собой пару, которая создала бы ПЦР-ампликон, содержащий N-й полиморфизм и N+1-й полиморфизм, если N-й полиморфизм соответствует лишь одному из двух аллелей. Продукты реакции затем получают исходя из количества ПЦР-амплификаций m, для N=1, 2, 3, …, m, и гибридизуют с зондами, которые могут определять N+1-й аллельный ген для N-го аллельного гена, выбранного с помощью FP (N). В описанном здесь случае зонды, созданные для соответствия 2×m аллелям, фиксируют в желаемых позициях на микрочипе.
Ампликоны, полученные в m ПЦР-амплификациях, смешивают и гибридизуют с зондами на микрочипе согласно протоколу, описанному для следующего примера 1. Поступая таким образом, можно исчерпывающе определить с помощью единственного мечения комбинации гаплотипа большого числа аллельных генов. Более того, методика данного изобретения, т.е. идентификация аллеля путем обнаружения, какой из двух зондов с последовательностями, соответствующими двум аллелям одного аллельного гена, подвергается гибридизации, повышает точность анализа гаплотипа.
В данной работе авторы изобретения ограничились обсуждением гетерозиготных аллельных генов. Для определения могут быть получены два набора праймеров, каждый из которых специфичен для одного из двух аллелей первого аллельного гена, и гибридизация проведена раздельно.
Примеры
Пример 1
Ниже описывается пример по первому варианту осуществления.
I Получение образца и выделение специфической области генома
Используемую матричную ДНК получали амплификацией области размером лишь 5 т.п.н. гена метаболического фермента CYP2D6 геномной ДНК, выделенной из линии В-клеток, полученных от японца, и разведением ампликона особо чистой водой до концентрации 8 нг/мкл. Эту процедуру осуществляли для устранения областей повторяющихся последовательностей, таких как CYP2D7 и CYP2D8, которые представляют собой псевдогены, и для получения чистой области CYP2D6. Конкретно, как правило, использовали широко применяемые праймеры, перечисленные в таблице 2.
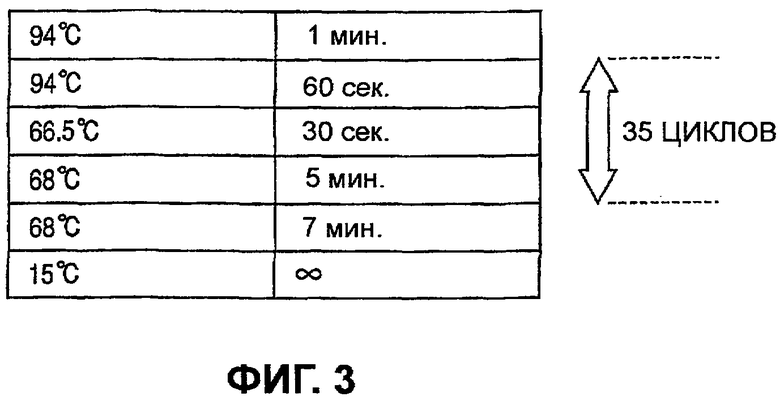
Раствор для проведения ПЦР получали, как показано в таблице 3, и цикл ПЦР осуществляли, как описано на Фиг.3, и в результате получали ампликон (размером 5079 нп) области гена CYP2D6.
Во время проведения реакции 3-стадийный цикл, включающий денатурацию, отжиг и удлинение, повторяли 35 раз и после охлаждения ампликон очищали с помощью процесса очистки.
ПЦР-ампликон очищали на колонке для очистки (набор для очистки продуктов ПЦР Qiagen QIAquick) и объем раствора, содержащего ПЦР-ампликон, доводили до 50 мкл. Часть раствора, содержащего очищенный ПЦР-ампликон, полученного таким образом, отбирали и подвергали электрофорезу стандартным способом и исходя из размера продукта подтверждали, что был синтезирован желаемый ПЦР-продукт. Продукт дополнительно разбавляли чистой водой до концентрации 8 нг/мкл.
II ПЦР-амплификация
Создание праймеров и очистка ПЦР-ампликона
В таблице 4 перечислены реагенты, использованные в реакции амплификации, и их соотношение в смеси. Здесь также перечисляются использованные концентрации и количества буфера для проведения ПЦР, ДНК-полимеразы, нуклеотидов, праймеров и матричной ДНК. В данной работе использовали праймер, меченный Су3.
dNTP
(dATP, dGTP, dCTP, dTTP)
Ampli Taq Gold (TAKARA)
Матричная ДНК
2 мМ
каждого
8 нг/мкл
2,5
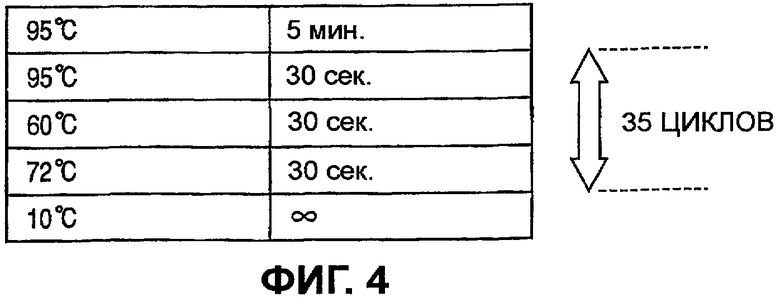
На Фиг.4 иллюстрируется протокол, использованный для реакции амплификации. В ходе реакции 3-стадийный цикл, включающий денатурацию, отжиг и удлинение, повторялся 35 раз, и затем после охлаждения ампликон очищали посредством процесса очистки.
ПЦР-ампликон очищали на колонке для очистки (набор для очистки продуктов ПЦР Qiagen QIAquick). После очистки объем раствора, содержащего ПЦР-ампликон, доводили до 50 мкл. Часть раствора, содержащего очищенный ПЦР-ампликон, полученного таким образом, отбирали и подвергали электрофорезу стандартным способом и исходя из размера продукта подтверждали, что был синтезирован желаемый ПЦР-продукт. Последовательности использованных праймеров и ампликона представлены ранее в таблице 1.
III Получение микрочипа
(1) Конструирование зондов
Для описанного выше ПЦР-продукта конструировали два зонда. Как и при конструировании праймеров зонды конструировали с особым вниманием, чтобы каждый зонд мог специфически распознавать последовательность оснований аллелей второго аллельного гена. В этой конструкции зондов цепь ДНК, которая удлиняется начиная от прямого праймера FP, представляет собой цепь, которая образует с зондом гибрид. Последовательность оснований каждого зонда тщательно конструировали, регулируя длину нуклеотидных пар и т.д., принимая во внимание устойчивость образуемого гибрида. Последовательности двух созданных зондов представлены в таблице 1.
(2) Синтез зондов и получение микрочипа
Синтез зондов и получение микрочипа осуществляли, используя способы получения микрочипов ДНК, описанные Canon Inc. (японская выложенная патентная заявка № 11-187900). Вкратце, для подготовки подложки кварцевое стекло обрабатывали агентом, связывающим силан, и с ним связывали EMCS для введения на поверхность малеимидных групп. В отношении синтеза зондов, синтезировали зонды, в которые на 5'-конце встраивали тиольную группу, и очищали их с помощью HPLC. Для получения ДНК-микрочипа каждый зонд помещали на стеклянную подложку, используя модифицированную версию струйного принтера (патентованное название BJF-850, продукция фирмы Canon Inc.). Размер стеклянной подложки составлял 25×75×1 мм (W × L × T).
IV Гибридизация
Гибридизацию на микрочипе осуществляли, используя ДНК-микрочип, полученный в пункте III, и ПЦР-ампликон, полученный в пункте II в качестве образца нуклеиновой кислоты.
(1) Блокирование ДНК-микрочипа
BSA (бычий сывороточный альбумин фракции V, продукция фирмы Sigma Co.) растворяли в 100 мМ NaCl/10 мМ фосфатном буфере до концентрации 1 мас.%. ДНК-микрочип, полученный в пункте III, погружали в этот раствор на 2 часа при комнатной температуре, чтобы добиться блокировки поверхности стеклянной подложки. После блокировки ее отмывали с помощью 2-кратного раствора SSC (NaCl в концентрации 300 мМ и цитрат натрия (тринатриевого цитрата дигидрат, т.е. C6H5Na3·2H2O) в концентрации 30 мМ, рН 7,0), содержащего 0,1 мас.%. SDS (додецилсульфат натрия). Затем ее промывали чистой водой. После этого ДНК-микрочип высушивали в устройстве для центробежной сушки.
(2) Получение гибридизационного раствора
Гибридизационный раствор получали, используя 2 мкл раствора, содержащего ПЦР-ампликон, для достижения конечной концентрации, приведенной ниже. Гибридизационный раствор имел следующий состав:
6-кратный SSPE/10% формамид/раствор, содержащий ПЦР-ампликон
(6-кратный SSPE: NaCl в концентрации 900 мМ, NaH2PO4·H2O в концентрации 60 мМ и EDTA в концентрации 6 мМ, рН 7,4).
(3) Гибридизация
Высушенный ДНК-микрочип помещали в прибор для гибридизации (гибридизационную камеру, продукция фирмы Genomic Solutions Inc.) и реакцию гибридизации осуществляли, используя гибридизационный раствор описанного выше состава исходя из процедур и условий, приведенных ниже.
Использованные условия и схема гибридизации были следующими.
Гибридизационный раствор нагревали до 65°С, оставляя при этой температуре в течение 3 минут, и поддерживали при 92°С в течение 2 минут и затем при 50°С в течение 4 часов. После этого его отмывали при 40°С с помощью 2-кратного SSC, содержащего 0,1% SDS. При необходимости его дополнительно отмывали при 25°С с помощью 2-кратного SSC и промывали чистой водой, следуя принятой инструкции, и окончательно высушивали в устройстве для центробежной сушки.
(4) Измерение флуоресценции
По окончании реакции гибридизации измеряли флуоресценцию, исходящую от гибрида, на высушенном с помощью центробежной сушки ДНК-микрочипе, используя сканер для измерения флуоресценции ДНК-микрочипа (Genepix 4000B, продукция фирмы Axon). Результаты измерения, полученные для каждого зонда, приводятся ниже в таблице 5.
При расчете яркости в качестве фонового значения использовали интенсивность флуоресценции на участках ДНК-микрочипа, не содержащих пятна ДНК-зонда, и в качестве измеренной интенсивности флуоресценции принимали наблюдаемую интенсивность флуоресценции каждого пятна минус фоновое значение. Измерения проводили дважды, и в данном документе приводятся средние величины.
Исходя из результатов гаплотип первого аллельного гена и второго аллельного гена определяли как С/Т (минор/минор). Кроме того, поскольку оба аллельных гена находились в гетерозиготном состоянии, диплотип определяли как G/G (мажор/мажор). В данной работе авторы изобретения ограничились гетерозиготными аллельными генами. Также могут быть получены два набора праймеров, каждый из которых специфичен к одному из двух аллелей первого аллельного гена, и гибридизация может быть осуществлена раздельно.
Настоящее изобретение не ограничивается вышеуказанными вариантами осуществления, и большое число изменений и модификаций может быть осуществлено в рамках сущности и объема настоящего изобретения. Поэтому для информирования научного мира об объеме настоящего изобретения приводится следующая формула изобретения.
По данной заявке испрашивается приоритет японской патентной заявки № 2006-325951, поданной 1 декабря 2006, которая приводится в данном документе в качестве ссылки в полном объеме.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ АНАЛИЗА ГЕНЕТИЧЕСКОГО ПОЛИМОРФИЗМА, ОПРЕДЕЛЯЮЩЕГО ПРЕДРАСПОЛОЖЕННОСТЬ К ОНКОЛОГИЧЕСКИМ ЗАБОЛЕВАНИЯМ И ИНДИВИДУАЛЬНУЮ ЧУВСТВИТЕЛЬНОСТЬ К ФАРМАЦЕВТИЧЕСКИМ ПРЕПАРАТАМ С ИСПОЛЬЗОВАНИЕМ ОЛИГОНУКЛЕОТИДНОГО БИОЛОГИЧЕСКОГО МИКРОЧИПА (БИОЧИПА) | 2005 |
|
RU2303634C2 |
| ОБНАРУЖЕНИЕ МУТАЦИЙ И ПЛОИДНОСТИ В ХРОМОСОМНЫХ СЕГМЕНТАХ | 2015 |
|
RU2717641C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ВЫСОКОМУЛЬТИПЛЕКСНОЙ ПЦР | 2012 |
|
RU2650790C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ПОПУЛЯЦИОННЫХ НАРУШЕНИЙ БИОТРАНСФОРМАЦИИ ЧУЖЕРОДНЫХ ВЕЩЕСТВ, ОБУСЛОВЛЕННЫХ ВОЗДЕЙСТВИЕМ ТЕХНОГЕННЫХ ХИМИЧЕСКИХ ФАКТОРОВ СРЕДЫ ОБИТАНИЯ | 2012 |
|
RU2497120C1 |
| СПОСОБЫ ВЫЯВЛЕНИЯ И МОНИТОРИНГА РАКА ПУТЕМ ПЕРСОНАЛИЗИРОВАННОГО ВЫЯВЛЕНИЯ ЦИРКУЛИРУЮЩЕЙ ОПУХОЛЕВОЙ ДНК | 2019 |
|
RU2811503C2 |
| Способ идентификации генетических полиморфизмов, влияющих на метаболизм противоопухолевых препаратов, с использованием биологических микрочипов | 2018 |
|
RU2697096C1 |
| Способ анализа полиморфных маркеров в генах VKORC1, CYP4F2, CYP2C9, CYP2C19, ABCB1, ITGB3 для определения индивидуальной чувствительности к противосвертывающим препаратам | 2018 |
|
RU2689400C1 |
| МУЛЬТИАЛЛЕЛЬНОЕ ГЕНОТИПИРОВАНИЕ ОДНОНУКЛЕОТИДНЫХ ПОЛИМОРФИЗМОВ И ИНДЕЛ-МУТАЦИЙ | 2016 |
|
RU2706203C1 |
| ГЕНЕТИЧЕСКИЕ МАРКЕРЫ ДЛЯ ПРОГНОЗИРОВАНИЯ ОТВЕТА НА ТЕРАПИЮ ПОДНИМАЮЩИМИ УРОВЕНЬ HDL ИЛИ ИМИТИРУЮЩИМИ HDL АГЕНТАМИ | 2015 |
|
RU2703192C2 |
| ГЕНЕТИЧЕСКИЕ МАРКЕРЫ ДЛЯ ПРОГНОЗИРОВАНИЯ ВОСПРИИМЧИВОСТИ К ТЕРАПИИ | 2014 |
|
RU2809215C2 |

Изобретение относится к области молекулярной биологии и может быть использовано в медицинской генетике. Предложен способ и набор праймеров для определения гаплотипа ДНК-мишени, содержащей два гетерозиготных полиморфных сайта. Способ по изобретению предусматривает а) получение образца геномной ДНК и амплификацию области, на 3'-стороне которой располагается первый полиморфный сайт, а на 5'-стороне - второй полиморфный сайт; б) проведение ПНР указанной специфической области с использованием одного из двух аллель-специфичных в отношении первого полиморфного сайта праймеров и обратного праймера, который располагается так, чтобы включить в ампликон второй полиморфный сайт; в) гибридизацию полученного ПЦР-продукта с двумя зондами, один из которых специфичен к последовательности мутантного типа, а другой - к последовательности дикого типа в области второго полиморфизма, и г) оценку гаплотипа, где первый полиморфизм определяется видом удлиняемого прямого праймера, а второй полиморфизм - видом зонда, гибридизовавшегося с ПНР-продуктом. Применение изобретения обеспечивает относительно простую и при этом высокоточную диагностическую систему. 2 н. и 2 з.п. ф-лы, 5 ил., 5 табл.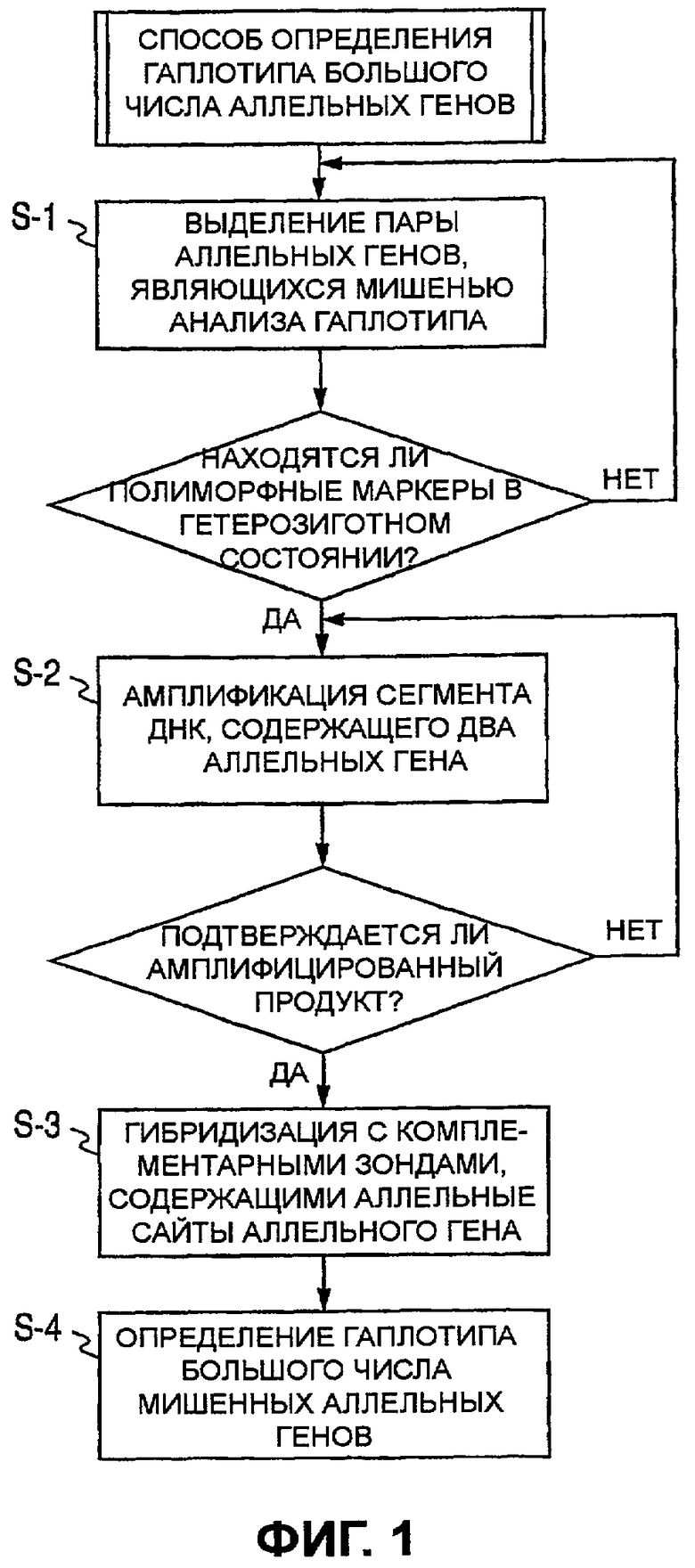
1. Способ определения гаплотипа геномной ДНК-мишени, имеющей два гетерозиготных полиморфных сайта, который отличается тем, что включает:
(i) получение образца геномной ДНК и выделение специфической области генома путем амплификации области, в которой первый гетерозиготный полиморфный сайт располагается на 3'-стороне, а второй гетерозиготный полиморфный сайт располагается на 5'-стороне области;
(ii) осуществление ПНР специфической области, полученной в результате (i), с использованием одного из прямых праймеров (а-1), перечисленных ниже:
(а-1-1) прямой праймер, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой мутантный тип, и не удлиняет нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой дикий тип, и
(а-1-2) прямой праймер, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм представляет собой дикий тип, и не удлиняет нуклеиновую кислоту-мишень, если этот первый полиморфизм представляет собой мутантный тип, и
(b-1) обратного праймера, который содержит последовательность оснований, комплементарную определенной последовательности оснований, которая не содержит второй полиморфизм и располагается на 3'-стороне второго полиморфизма на комплементарной цепи нуклеиновой кислоты-мишени,
где один из используемых праймеров является меченным;
(iii) гибридизацию продукта, полученного в результате (ii) с первым зондом, который имеет последовательность оснований, идентичную определенному сегменту последовательности оснований нуклеиновой кислоты-мишени, которая содержит нуклеотид(ы) дикого типа второго полиморфизма, и вторым зондом, который имеет последовательность оснований, идентичную определенному сегменту последовательности оснований нуклеиновой кислоты-мишени, которая содержит мутантный(ые) нуклеотид(ы) второго полиморфизма, и определение сигналов образования гибрида с первым и вторым зондами; и
(iv) определение гаплотипа геномной ДНК, где использованный прямой праймер соответствует первому полиморфизму, а зонд, с которым происходит гибридизация в (iii), соответствует второму полиморфизму.
2. Способ определения гаплотипа по п.1, в котором гибридизация (iii) дополнительно включает гибридизацию носителя зонда, на котором первый зонд и второй зонд фиксируются с продуктом, полученным в результате (ii).
3. Способ определения гаплотипа по п.1 или 2, в котором прямой праймер (а-1-1) имеет на своем 3'-конце или вблизи него нуклеотид(ы), комплементарный(ые) первому полиморфизму, имеющему нуклеотид(ы) мутантного типа, и
прямой праймер (а-1-2) имеет на своем 3'-конце или вблизи него нуклеотид(ы), комплементарный(ые) первому полиморфизму, имеющему нуклеотид(ы) дикого типа.
4. Набор праймеров для определения гаплотипа геномной ДНК-мишени, имеющей два гетерозиготных полиморфных сайта, способом по пп.1-3, где набор праймеров отличается содержанием одного из прямых праймеров, т.е. прямого праймера, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой мутантный тип, и не удлиняет нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой дикий тип, и прямого праймера, который содержит последовательность оснований, комплементарную первому полиморфному сайту, и может удлинять нуклеиновую кислоту-мишень, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой дикий тип, и не удлиняет, если первый полиморфизм нуклеиновой кислоты-мишени представляет собой мутантный тип; и обратного праймера, который содержит последовательность оснований, комплементарную определенной последовательности оснований, которая не содержит второй полиморфизм и располагается на 3'-стороне второго полиморфизма на комплементарной цепи нуклеиновой кислоты-мишени.
| VERDUYN W | |||
| ET AL, Tissue antigens, 1996, 48(2), 80-86 | |||
| US 20050260607 A1, 24.11.2005 | |||
| ГЕНЕТИЧЕСКИЙ ТЕСТ ДЛЯ ИДЕНТИФИКАЦИИ НОСИТЕЛЕЙ РЕЦЕССИВНОГО ГЕНА КОМПЛЕКСНЫХ ВЕРТЕБРАЛЬНЫХ МАЛЬФОРМАЦИЙ У КРУПНОГО РОГАТОГО СКОТА | 2001 |
|
RU2276690C2 |

