Область изобретения
Настоящее изобретение в целом относится к новому терапевтическому применению агонистов печеночного рецептора Х (LXR - от англ. liver X receptor). Более конкретно настоящее изобретение относится к применению агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения заболевания, обусловленного дегенерацией бета-клеток, такого как диабет, а также к способу повышения жизнеспособности ex vivo островковых клеток поджелудочной железы, при котором приводят в контакт указанные островковые клетки с агонистом LXR.
Предшествующий уровень техники
Диабет, как правило, классифицируют на две основных группы. При диабете типа 1 аутоиммунное разрушение бета-клеток внутри островков Лангерганса приводит к заметному нарушению в продуцировании инсулина. Напротив, диабет типа II характеризуется устойчивостью к инсулину в мышцах, жировой ткани и печени параллельно с относительным ухудшением продуцирования инсулина в бета-клетках. Множественные гены вносят вклад в склонность к обоим типам диабета, типу 1 и типу II, хотя во многих случаях их идентификация остается неизвестной.
Апоптоз представляет собой активный процесс саморазрушения клеток, который регулируется внешними и внутренними сигналами, имеющими место во время нормального развития. Документально хорошо подтверждено, что апоптоз играет ключевую роль в пластичности эндокринных бета-клеток поджелудочной железы. Возрастает количество данных, что у взрослых млекопитающих масса бета-клеток подвергается динамическим изменениям, чтобы адаптировать продуцирование инсулина к поддержанию эугликемии при особых состояниях, таких как беременность и ожирение. Регуляция массы бета-клеток зависит от тонкого равновесия между клеточной пролиферацией, ростом и клеточной гибелью (апоптозом). Нарушение этого баланса может привести к ухудшению гомеостаза глюкозы. Например, примечательно, что непереносимость глюкозы развивается с возрастом, когда скорость репликации бета-клетки снижена, и что у пациентов с инсулиннезависимым сахарным диабетом потеря массы бета-клеток составляет 40-60% по сравнению с субъектами, не являющимися диабетиками. Общепринято, что у субъектов, устойчивых к инсулину, нормогликемия поддерживается за счет компенсаторной гиперинсулинемии до тех пор, пока бета-клетки не становятся неспособными удовлетворять повышенную потребность в инсулине, и в этот момент возникает диабет типа II.
Сахарный диабет типа II, или инсулиннезависимый сахарный диабет (ИНСД), является полигенным заболеванием и составляет >90% случаев диабета. Это заболевание характеризуется устойчивостью к действию инсулина на всасывание глюкозы и нарушенным действием инсулина на ингибирование продуцирования глюкозы печенью.
Регуляция метаболизма глюкозы инсулином является ключевым механизмом, посредством которого поддерживается гомеостаз у животного. Инсулин стимулирует всасывание глюкозы из крови в ткани, в частности в мышечную и жировую. Это происходит за счет повышенного перемещения Glut4, чувствительного к инсулину транспортера глюкозы, от внутриклеточного везикулярного компартмента к плазматической мембране. Glut4 является важнейшим чувствительным к инсулину транспортером глюкозы в этих тканях. Инсулин связывается с его рецептором в плазматической мембране, генерируя серию сигналов, которые приводят к перемещению или движению везикул транспортера Glut4 к плазматической мембране.
Печеночные рецепторы Х (LXR) являются членами надсемейства ядерных рецепторов, которые индуцируют зависимую от лиганда транскрипционную активацию генов-мишеней. Они играют важную роль в метаболизме и гомеостазе холестерина. Известно, что у млекопитающих существует два белка LXR (альфа и бета). Экспрессия LXR альфа является высокой в органах, вовлеченных в гомеостаз липидов, таких как тонкий кишечник, бурая и белая жировая ткань, тогда как LXR бета является более распространенным и обильным в тканях нервного и эндокринного происхождения. Недавно обнаружено, что LXR альфа и бета экспрессируются в островках поджелудочной железы, а также в альфа-клетках и бета-клетках (Efanov et al, Diabetes, 53 (3), S75-78, 2004).
LXR альфа и LXR бета являются близкородственными и обладают 77% аминокислотной идентичностью как в их ДНК-, так и в связывающих лиганд доменах. LXR также являются консервативными у людей и других животных (например, у грызунов). Подобно другим ядерным рецепторам, LXR образуют гетеродимер с рецептором ретиноида Х (RXR) для функционирования. Известно, что LXR активируются некоторыми встречающимися в природе окисленными производными холестерина, включая 22(R)-гидроксихолестерин, 24(S)-гидроксихолестерин и 24,25(S)-эпоксихолестерин.
LXR альфа и бета являются регуляторами печеночных генов, вовлеченных в метаболизм холестерина и жирных кислот (HMGCoA синтазы/редуктазы, фарнезилдифосфатсинтазы, скваленсинтазы, SREBP1c, стеароил-СоА-десатуразы (SCD1 и 2), FAS), ингибируют экспрессию ферментов глюконеогенеза (РЕРСК, фруктозодифосфатазы-1, глюкозо-6-фосфатазы), индуцируют экспрессию трансмембранных транспортеров (АВСА1, Glut1 и Glut4), ингибируют экспрессию ферментов, вовлеченных в гликолиз (6-фосфофрукто-2-киназы) и индуцируют киназу пируватдегидрогеназы-4 (негативный регулятор гликолиза), снижают 11-бетагидроксистероиддегидрогеназу типа 1 (фермент, который повторно активирует неактивный кортизон в активный кортизол у людей). Лептин и UCP-1 идентифицированы как гены-мишени LXR (понижающая регуляция агонистами LXR). Кроме того, LXR имеют перекрывающиеся функции с PPAR (рецепторами активаторов пролиферации пероксисом) при негативной регуляции воспалительного ответа. LXR ингибируют продуцирование TNFα и IL-1β и экспрессию воспалительных медиаторов, таких как СОХ2, iNOS, IL-6. LXR могут играть ключевую роль в ответах на воспаление, и, поскольку показано, что они важны в метаболизме липидов, LXR могут быть также вовлечены в индуцированные ожирением воспалительные ответы (рассмотрено в Steffensen et al, Diabetes, 2004, 53(1), S36-42).
Показано, что у мышей db/db агонист LXR T0901317, описанный здесь ниже, снижает уровень глюкозы в плазме (не у нормальных мышей). Данное соединение ингибирует экспрессию РЕРСК (фосфоенолпируват-карбоксикиназы) с ограничением образования глюкозы печенью (Cao et al, J Biol Chem, 2003, 278, 1131-1136).
Культивирование островков поджелудочной железы или секретирующих инсулин клеток MIN6 с T0901317 вызывало повышение зависимой от глюкозы секреции инсулина и содержание инсулина в островках. Стимулирующий эффект данного соединения на секрецию инсулина наблюдали только после >72 ч культивирования островков с T0901317. В клетках MIN6 Tularik повышал белковую экспрессию липогенных ферментов, синтазы жирных кислот и ацетил-СоА карбоксилазы. Активация LXR также давала повышение уровня активности белков глюкокиназы и пируваткарбоксилазы (PC). LXR могут регулировать секрецию и биосинтез инсулина посредством регуляции метаболизма глюкозы и липидов в бета-клетках поджелудочной железы (Efanov et al, Diabetes, 53 (3), S75-78, 2004).
Панкреатический дуоденальный гомеобоксный ген-1 (Pdx-1) является главным регулятором как развития поджелудочной железы, так и дифференциации клеток-предшественников в фенотип бета-клеток. Кроме того, в дифференцированной бета-клетке Pdx1 является отвечающим на глюкозу регулятором экспрессии гена инсулина, и функция Pdx1 в ответ на глюкозу регулируется как его фосфорилированием, так и ядерной транслокацией. Во время более поздних стадий развития островков экспрессия Pdx-1 становится наиболее ограниченной зрелыми бета-клетками эндокринной поджелудочной железы. В поджелудочной железе взрослых субпопуляции клеток, продуцирующих соматостатин, и клеток, продуцирующих панкреатический полипептид, также экспрессируют Pdx-1, и только немногие клетки, продуцирующие глюкагон, экспрессируют его.
Дефект чувствительности к глюкозе бета-клеток поджелудочной железы наблюдали в нескольких животных моделях диабета типа II, и он коррелировал со сниженной экспрессией гена транспортера глюкозы типа 2 (Glut2). В модели трансгенной мыши показано, что экспрессия Glut2 антисмысловой РНК в бета-клетках поджелудочной железы связана с нарушенной индуцируемой глюкозой секрецией инсулина и развитием диабета. Экспрессия гена-транспортера глюкозы типа 2 (GLUT2) избирательно снижена в бета-клетках поджелудочной железы экспериментальных моделей диабета, и мышиный промотор GLUT2 регулируется PDX-1.
Сущность изобретения
Настоящее изобретение частично основано на неожиданном открытии, что активация LXR у крыс ZDF приводит к улучшению диабетического состояния при значительном снижении гликемии после обработки агонистом LXR. При оценке потенциальных механизмов, лежащих в основе противодиабетического действия агониста LXR, неожиданно обнаружено значительное снижение апоптоза в изолированных островках [Лангерганса] из крыс ZDF, обработанных известным агонистом LXR, названным "GW3965", связанное с увеличением числа островков;
кроме того, эти островки имели значительно более высокое содержание инсулина и лучшую чувствительность к глюкозе, чем островки из крыс ZDF, обработанных носителем. Эти открытия и дополнительная экспериментальная работа показали, что введение агониста LXR in vivo способно стимулировать секрецию инсулина, в то же время защищая островки поджелудочной железы от разрушения.
Кроме того, показано, что значительное повышение уровня мРНК PDX1 и Glut2, двух основных генов, вовлеченных в регенерацию островков, было получено в островках крыс ZDF, обработанных GW3965. Регенерация островков поджелудочной железы также подтверждена гистологическим анализом поджелудочной железы мышей NOD, обработанных GW3965.
Настоящее изобретение также основано на новом открытии, что агонист LXR не только защищает поджелудочную железу диабетиков от дальнейшего разрушения, в частности, посредством снижения апоптоза и инсулита в поджелудочной железе, но также усиливает регенеративные процессы поджелудочной железы и, таким образом, дает возможность восстановления функции поджелудочной железы, когда она подвергается риску. Эти эффекты являются дополнительными к снижению образования глюкозы печенью, описанному в печени (Cao et al, 2003, JBC, 278 (2), 1131-1136), и потенцируют противодиабетические свойства агонистов LXR.
Полезное свойство агонистов LXR увеличивать число островков и снижать апоптоз в островках дает возможность нового режима лечения: он состоит в лечении пациентов в течение ограниченного периода времени до тех пор, пока гликемия и/или HbA1c (гемоглобин А1 с) не снизится до уровня стабильного состояния (HbA1C ≤7% и/или гликемия ≤1,2-1,4 г/л); тогда лечение приостанавливают до тех пор, пока гликемия и/или HbA1c остается в приемлемом интервале (например, на один месяц). Если уровень гликемии и/или HbA1c повышается выше приемлемых значений, тогда лечение агонистом LXR будет назначено снова.
В первом аспекте настоящее изобретение, таким образом, направлено на применение агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения заболевания, обусловленного дегенерацией бета-клеток.
Настоящее изобретение также направлено на применение агониста LXR для изготовления лекарственного средства, полезного для регенерации бета-клеток и для повышения жизнеспособности бета-клеток.
Применение в соответствии с изобретением предусматривает лекарственное средство, которое можно применять в сочетании с известным противодиабетическим лекарственным средством.
Во втором аспекте настоящее изобретение направлено на способ повышения ex vivo жизнеспособности первичных островковых клеток поджелудочной железы, при котором приводят в контакт указанные островковые клетки с агонистом LXR.
В третьем аспекте в настоящем изобретении предложены способы лечения дегенерации бета-клеток, при которых субъекту вводят агонист LXR.
В первом воплощении в изобретении предложены способы предупреждения диабета типа 1 у субъекта, при которых субъекту вводят эффективное количество агониста LXR.
В другом воплощении в настоящем изобретении предложены способы лечения или облегчения диабета, при которых измеряют уровень циркулирующей глюкозы и/или HbA1c у субъекта и вводят этому субъекту ежесуточно эффективное количество агониста LXR до стабилизации гликемии и/или HbA1c (HbA1C ≤7% и/или гликемии ≤1,2-1,4 г/л, поддерживаемых в течение одного месяца по меньшей мере двумя дозами), затем приостанавливают лечение, как только гликемия и/или HbA1c стабилизируется, при этом ежесуточное лечение, возможно, возобновляют, если необходимо.
В другом воплощении в изобретении предложены способы повышения жизнеспособности трансплантированных донорских островковых клеток поджелудочной железы у реципиента трансплантата, при которых указанному реципиенту трансплантата вводят агонист LXR.
Дальнейшее понимание природы и преимуществ настоящего изобретения может быть реализовано со ссылкой на остальные разделы описания и формулы изобретения.
Подробное описание изобретения
В одном аспекте настоящее изобретение направлено на применение агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения заболевания, обусловленного дегенерацией бета-клеток.
Применение согласно настоящему изобретению находит приложение при лечении заболеваний, обусловленных, в частности, потерей функции бета-клеток или дисфункции бета-клеток и/или гибели бета-клеток, таких как, но не ограниченных ими, диабет, гипергликемия и ожирение. Настоящее изобретение также полезно для предупреждения или модулирования развития таких заболеваний или расстройств у субъекта, подозреваемого на склонность к таким заболеваниям или расстройствам, либо если эта склонность известна.
Настоящее изобретение, таким образом, относится к применению агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения диабета типа I и типа II, и предпочтительно диабета типа I.
Как объяснено выше, у пациентов с диабетом типа II могут проявляться в большей или меньшей степени признаки диабета типа I, в основном, связанные с дефицитом продуцирования инсулина. Соответственно, в одном воплощении настоящее изобретение направлено на применение агониста LXR для изготовления лекарственного средства, полезного для предупреждения, остановки или замедления прогрессирования диабета типа II в направлении последующего диабетического состояния, характеризующегося дефицитом продуцирования инсулина.
Поскольку введение in vivo агониста LXR увеличивает число островков и снижает апоптоз в островках, применение агониста LXR для изготовления лекарственного средства, полезного для регенерации бета-клеток и для повышения жизнеспособности бета-клеток, представляет собой два дополнительных воплощения настоящего изобретения.
В отношении применения для повышения жизнеспособности клеток поджелудочной железы должно быть понятно, что жизнеспособные клетки определяют как способные к пролиферации, дифференциации, росту и развитию. Жизнеспособность можно измерить любыми способами, известными в данной области техники, и, например, путем использования окрашивания трипановым синим.
В другом воплощении настоящее изобретение направлено на применение агониста LXR для изготовления лекарственного средства, полезного для повышения жизнеспособности трансплантированных донорских островковых клеток поджелудочной железы у реципиента трансплантата. Островковые клетки поджелудочной железы представляют собой первичные островковые клетки.
Для данного конкретного применения лекарственное средство можно вводить местно в трансплантированную область или предпочтительно вводить системно.
В одном воплощении лекарственное средство вводят реципиенту трансплантата до и/или после трансплантации донорских островковых клеток поджелудочной железы. В другом воплощении лекарственное средство вводят реципиенту трансплантата одновременно с трансплантацией донорских клеток поджелудочной железы.
Любые агонисты LXR, известные и описанные в данной области техники, можно применять для всех применений и способов, как описано здесь в соответствии с настоящим изобретением. Предпочтительно агонист LXR представляет собой агонист LXR альфа, и, в частности, агонист LXR выбран из группы, состоящей из GW3965 и Т0901317.
Кроме того, согласно предпочтительному воплощению, лекарственное средство, которое следует применять для всех применений и способов настоящего изобретения, находится в форме, пригодной для перорального введения.
При всех применениях и способах, описанных здесь, лекарственное средство предпочтительно вводят ежесуточно в течение по меньшей мере 30 суток. Его можно также вводить в течение по меньшей мере 60 суток, 90 суток или дольше.
Кроме того, лекарственное средство согласно настоящему изобретению можно применять в сочетании с известным противодиабетическим лекарственным средством, в частности, выбранным из группы, состоящей из метформина, пиоглитазона, розиглитазона, глимепирида, глипизида, глибурида/метформина, глибурида, миглитола, глипизида + метформина, репаглинида, акарбозы, троглитазона, натеглинида и агониста GLP-1 (глюкагоноподобного пептида-1).
Во втором аспекте настоящее изобретение направлено на способ повышения жизнеспособности первичных островковых клеток поджелудочной железы ex vivo, при котором приводят в контакт указанные островковые клетки поджелудочной железы с агонистом LXR.
Предпочтительно агонист LXR представляет собой агонист LXR альфа, и, в частности, агонист LXR, который следует применять, выбран из группы, состоящей из GW3965 и Т0901317.
В третьем аспекте в настоящем изобретении предложены способы лечения дегенерации бета-клеток. Эти способы включают введение субъекту агониста LXR. При таких способах применяемый агонист LXR может представлять собой агонист LXR альфа, и, в частности, агонист LXR, который следует применять, выбран из группы, состоящей из GW3965 и Т0901317.
В способах согласно изобретению субъект предпочтительно страдает диабетом типа I или типа II. Возможно, агонист LXR вводят субъекту одновременно с известным противодиабетическим лекарственным средством или с любым другим лекарственным средством, снижающим гликемию и/или HbA1c. В частности, известное противодиабетическое лекарственное средство выбрано из группы, состоящей из метформина, пиоглитазона, розиглитазона, глимепирида, глипизида, глибурида/метформина, глибурида, миглитола, глипизида+метформина, репаглинида, акарбозы, троглитазона, натеглинида и агониста GLP-1.
В одном воплощении в изобретении предложены способы предупреждения диабета типа I у субъекта. Эти способы включают введение субъекту эффективного количества агониста LXR. Подобным образом, агонист LXR, применяемый в указанных способах предупреждения диабета типа I, может представлять собой агонист LXR альфа, и, в частности, агонист LXR, который следует применять, выбран из группы, состоящей из GW3965 и Т0901317.
Возможно, агонист LXR вводят субъекту одновременно с известным противодиабетическим лекарственным средством или с любым другим лекарственным средством, снижающим гликемию и/или HbA1c. В частности, известное противодиабетическое лекарственное средство выбрано из группы, состоящей из метформина, пиоглитазона, розиглитазона, глимепирида, глипизида, глибурида/метформина, глибурида, миглитола, глипизида+метформина, репаглинида, акарбозы, троглитазона, натеглинида и агониста GLP-1.
В другом аспекте в настоящем изобретении предложены способы лечения или облегчения диабета, при которых измеряют уровень циркулирующей глюкозы и/или HbA1c у субъекта и вводят этому субъекту ежесуточно эффективное количество агониста LXR до стабилизации гликемии и/или HbA1c (HbA1C ≤7% и/или гликемия ≤1,2-1,4 г/л, поддерживаемая в течение одного месяца по меньшей мере двумя дозами), затем приостанавливают лечение, как только гликемия и/или HbA1c стабилизируется, при этом ежесуточное лечение возможно возобновляют при необходимости.
Ежесуточное лечение может быть возобновлено, если гликемия и/или HbA1c более не стабилизированы или потеряна чувствительность к инсулину.
Способы измерения уровней циркулирующей глюкозы и/или HbA1c представляют собой способы, хорошо известные на предшествующем уровне техники, и можно использовать любой вид способов.
Подобным образом агонист LXR, применяемый в указанных способах лечения или облегчения диабета, может представлять собой агонист LXR альфа, и, в частности, агонист LXR, который следует применять, выбран из группы, состоящей из GW3965 и Т0901317. Возможно, агонист LXR вводят субъекту одновременно с известным противодиабетическим лекарственным средством или с любым другим лекарственным средством, снижающим гликемию и/или HbA1c. В частности, известное противодиабетическое лекарственное средство выбрано из группы, состоящей из метформина, пиоглитазона, розиглитазона, глимепирида, глипизида, глибурида/метформина, глибурида, миглитола, глипизида+метформина, репаглинида, акарбозы, троглитазона, натеглинида и агониста GLP-1.
В указанных способах агонист LXR предпочтительно вводят субъекту по меньшей мере ежесуточно в течение по меньшей мере 30 суток.
В другом воплощении настоящее изобретение направлено на способ повышения жизнеспособности трансплантированных донорских островковых клеток поджелудочной железы реципиенту трансплантата, при котором указанному реципиенту трансплантата вводят агонист LXR.
В последующих разделах приведено руководство по получению и применению лекарственного средства в соответствии с изобретением и по осуществлению способов по изобретению.
Определения
Если не указано иное, все технические и научные термины, используемые здесь, имеют такое же значение, как обычно их понимает обычный специалист в области техники, к которой принадлежит данное определение. Приведенные ниже ссылки дают специалисту в данной области техники общее определение многих терминов, используемых в данном изобретении: Singleton et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY (2d ed. 1994); THE CAMBRIDGE DICTIONARY OF SCIENCE AND TECHNOLOGY (Walker ed., 1988); и Hale & Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY (1991). Кроме того, приведенные ниже определения приведены, чтобы помочь читателю в осуществлении изобретения.
Термин "LXR" (печеночный рецептор X) или "рецептор LXR" включает все подтипы данного рецептора. Конкретно LXR включает LXR альфа и LXR бета. Ряд названий, таких как LXRU, LXRa, LXR, RLD-1, NR1H3, использовали для обозначения LXR альфа. Он охватывает любой полипептид, кодируемый геном с существенной идентичностью последовательности с номером по каталогу GenBank U22662. Подобным образом, LXR бета включает любой полипептид, кодируемый геном, который называют как LXRb, LXRP, LXR бета, NER, NER1, UR, OR-1, RIP 15, NR1H2, или геном с существенной идентичностью последовательности с номером по каталогу GenBank U07132.
Термин "лиганд" относится к агонисту или частичному агонисту LXR. Лиганд может быть избирательным к LXR альфа или LXR бета, либо он может обладать смешанным связывающим сродством к обоим рецепторам LXR альфа и LXR бета. Хотя лиганд может либо агонизировать, либо антагонизировать функцию рецептора, если не указано иное, лиганд LXR, используемый здесь, прежде всего, относится к агонисту LXR, который активирует рецептор LXR.
Термин "модулировать" в отношении рецептора LXR относится к активации рецептора LXR и его биологических активностей, связанных с биохимическим путем LXR (например, регуляции транскрипции гена-мишени). Модулирование рецептора LXR может представлять собой повышающую регуляцию (то есть агонизм, активацию или стимуляцию) или понижающую регуляцию (то есть антагонизм, ингибирование или подавление). Механизм действия модулятора LXR может быть прямым, например, посредством связывания с рецептором LXR в качестве лиганда. Модулирование также может быть косвенным, например, посредством связывания с другой молекулой и/или модификацией этой другой молекулы, которая в противном случае связывается с рецептором LXR и активирует его. Таким образом, модулирование LXR включает изменение в биологических активностях лиганда агониста LXR (то есть его активности при связывании и/или активации рецептора LXR) или изменение в клеточном уровне лиганда.
Как используют здесь, выражение "скрининг на агонисты LXR" относится к использованию соответствующей системы анализа для идентификации новых агонистов LXR из тестируемых агентов. Анализ может представлять собой анализ in vitro или in vivo, пригодный для идентификации, может ли тестируемый агент стимулировать или активировать одну или более чем одну из биологических функций рецептора LXR. Примеры подходящих биологических анализов включают, но не ограничены ими, анализы на исследование связывания тестируемых агентов с полипептидом LXR (например, фрагментом LXR, содержащий его домен связывания с лигандом), анализы на основе транскрипции, анализы креатинкиназы, анализы, основанные на дифференциации преадипоцитов, анализы, основанные на регуляции захвата глюкозы в адипоцитах, и иммунологические анализы.
"Субъект" предпочтительно представляет собой млекопитающее, более предпочтительно человека.
Термин "дегенерация бета-клеток" предназначен для обозначения потери функции бета-клеток, дисфункции бета-клеток и гибели бета-клеток, такой как некроз или апоптоз бета-клеток.
Термин "лечение заболевания" определяют как помощь и уход за пациентом с целью борьбы с заболеванием, состоянием или расстройством и включают в него введение агониста LXR для предупреждения появления симптомов или осложнений, либо для облегчения симптомов или осложнений, либо для устранения заболевания, состояния или расстройства. Лечение включает модулирование, ингибирование, снижение, уменьшение или остановку дегенерации бета-клеток, такой как некроз или апоптоз бета-клеток, в частности, программируемой гибели бета-клеток, известной как апоптоз бета-клеток, а также предупреждение дегенерации бета-клеток, такой как некроз или апоптоз бета-клеток, в частности, предупреждение апоптоза бета-клеток. Лечение включает также дегенерацию бета-клеток.
Термин "для лечения", как используют здесь, следует понимать как охватывающий прямое применение соединения для лечения или косвенное применение указанного соединения при лечении указанного заболевания.
Термин "дефицит продуцирования инсулина" предназначен для обозначения проявления значимой гипергликемии и нормо- или гипоинсулинемии. Способы измерения уровней гликемии или инсулина и оценки гипергликемии и нормо- или гипоинсулинемии хорошо известны в данной области техники.
Термин "в сочетании с" следует относить либо к лекарственному средству, в котором два активных вещества (агонист LXR и противодиабетическое лекарственное средство) являются существенными составными частями одной и той же композиции, либо к двум отдельным лекарственным средствам, которые можно вводить одновременно или последовательно.
Термины "стабилизация гликемии и/или HbA1c" или "как только гликемия и/или HbA1c стабилизируется", как используют здесь, предназначены для обозначения значения HbA1C ≤7% и/или значения гликемии ≤1,2-1,4 г/л, поддерживаемые в течение одного месяца по меньшей мере двумя дозами.
АГОНИСТЫ LXR
Существуют многие агонисты LXR, которые пригодны для осуществления способов по настоящему изобретению. Они могут представлять собой любые известные агенты, которые активируют рецептор LXR, например GW3965 (см. Примеры ниже) или другие имеющиеся в продаже соединения, такие как F3метилАА (от Merck; см. Menke et al., Endocrinology 143: 2548-58, 2002) и Т0901317 (Tularik, Calif.). Они могут также представлять собой новые агонисты LXR, подлежащие скринингу в соответствии с настоящим изобретением. Как подробно описано ниже, агонисты LXR, пригодные для настоящего изобретения, могут представлять собой полипептиды, пептиды, малые молекулы или другие агенты. Агонисты LXR могут представлять собой агонисты для LXR человека, а также других животных.
Большое число агонистов LXR описано в данной области техники. Примеры низкомолекулярных агонистов LXR включают хорошо известные оксистерины и родственные соединения (Janowski et al., Nature 383: 728-31, 1996); Т0901317 и Т0314407 (Schultz et al., Genes Dev 14: 2831-8, 2000); 24(S)-гидроксихолестерин и 22(R)-гидроксихолестерин (Janowski et al., Nature 383: 728-731, 1996); а также 24,25-эпоксихолестерин (патент США №6316503). Примерные полипептидные агонисты LXR также описаны в данной области техники, например, в WO 02/077229. Дополнительные агонисты LXR описаны в данной области техники, например в патенте США №6316503; Collins et al., J Med Chem. 45: 1963-6, 2002; Joseph et al., Proc Natl Acad Sci USA 99: 7604-9, 2002; Menke et al., Endocrinology 143: 2548-58, 2002; Schultz et al., Genes Dev. 14: 2831-8, 2000 и Schmidt et al., Mol Cell Endocrinol. 155:51-60, 1999.
Многие агонисты LXR эффективны при активации как LXR альфа, так и LXR бета (например, GW3965, как описано в Collins et al., J Med Chem. 45: 1963-6, 2002). Некоторые агонисты LXR активируют LXR альфа и LXR бета в различных условиях. Например, 6-альфа-гидроксилированные желчные кислоты являются агонистами LXR альфа, но также активируют LXR бета при более высоких концентрациях (Song et al., Steroids 65: 423-7, 2000). Некоторые агонисты LXR действуют исключительно на LXR альфа, тогда как некоторые другие активируют только LXR бета. Например, в результате введения атома кислорода на стерольное В-кольцо оксистерина образуется лиганд с избирательностью к LXR альфа-подтипу (Janowski et al., Proc Natl Acad Sci USA 96: 266-71, 1999). С использованием зависимых от лиганда транскрипционных анализов было обнаружено, что 5-тетрадецилокси-2-фуранкарбоновая кислота (TOFA) и гидроксихолестерин трансактивируют химерные рецепторы, состоящие из ДНК-связывающего домена рецептора глюкокортикоидов и связывающих лиганд областей рецепторов LXR бета, PPAR альфа и PPAR бета (Schmidt et al., Mol Cell Endocrinol. 155:51-60, 1999).
Агонисты LXR могут быть также получены из производных известных полипептидных агонистов рецептора LXR. Они могут быть получены с помощью ряда методик, известных в данной области техники. Например, можно синтезировать специфичные олигопептиды (например, 10-25 аминокислотных остатков), охватывающие известный полипептидный агонист LXR (например, химическим или рекомбинантным путем) и тестировать на их способность к активации рецептора LXR. Фрагменты агониста LXR можно синтезировать, используя стандартные методики, такие как описаны в Bodansky, M. Principles of Peptide Synthesis, Springer Verlag, Berlin (1993) и Grant, G. A. (ed.). Synthetic Peptides: A User's Guide, W.H. Freeman and Company, New York (1992). Автоматические синтезаторы пептидов имеются в продаже, например, от Advanced ChemTech Model 396; Milligen/Biosearch 9600. Альтернативно такие агонисты LXR могут быть получены путем ферментативного расщепления нативных или продуцированных рекомбинантным путем полипептидных агонистов LXR, используя протеазу, например трипсин, термолизин, химотрипсин или пепсин. Компьютерный анализ (с использованием имеющегося в продаже программного обеспечения, например MacVector, Omega, PCGene, Molecular Simulation, Inc.) можно использовать для идентификации сайтов протеолитического расщепления.
Полипептидные или пептидные агонисты для применения в способах по настоящему изобретению являются предпочтительно изолированными и по существу свободными от клеточного материала или других загрязняющих белков из клетки или тканевого окружения, из которых выделяют агонисты LXR, или по существу свободными от химических предшественников или других химических веществ при химическом синтезе. Протеолитические или синтетические полипептидные агонисты или их фрагменты могут содержать столько аминокислотных остатков, сколько необходимо для активации рецепторной активности LXR, и могут содержать по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45, 50 или более аминокислотных остатков в длину.
Кроме известных соединений и полипептидов, которые активируют рецептор LXR, агонисты LXR могут быть также получены путем скрининга тестируемых агентов (например, библиотек соединений) для идентификации новых агонистов LXR, которые связываются с рецепторными активностями LXR и/или активируют их. Для скрининга на такие новые агонисты LXR можно использовать человеческий LXR или LXR других животных в системе точного анализа. Полинуклеотидные и аминокислотные последовательности рецепторов LXR известны и описаны в данной области техники. Их структуры и функциональные организации, включая их домены, связывающие лиганд, также охарактеризованы. См., например, Apfel et al., Mol Cell Biol 14: 7025-7035, 1994; Willy et al., Genes Dev 9: 1033-1045, 1995; Song et al., Proc Natl Acad Sci USA 91: 10809-10813, 1994; Shinar et al., Gene 147: 273-276, 1994; Teboul et al., Proc Natl Acad Sci USA 92: 2096-2100, 1995; и Seol et al., Mol Endocrinol 9:72-85, 1995.
Агонисты могут активировать либо LXR, либо LXR альфа. Кроме того, вместо полноразмерной молекулы LXR в некоторых из скрининговых анализов можно использовать полипептид LXR, который содержит фрагмент молекулы LXR. Например, два функциональных домена рецептора LXR, N-концевой ДНК-связывающий домен (DBD) и С-концевой домен, связывающий лиганд (LBD), опосредуют функцию транскрипционной активации ядерных рецепторов. Полипептид LXR, содержащий любой из этих доменов, можно использовать в скрининге на новые агонисты LXR. Ряд аналитических систем можно использовать для скрининга тестируемых агентов на агонисты рецептора LXR. Как подробно описано ниже, тестируемые агенты можно подвергать скринингу на прямое связывание с полипептидом LXR или его фрагментом (например, его доменом, связывающим лиганд). Альтернативно или дополнительно потенциальные агонисты LXR можно исследовать на способность к активации биохимического пути рецептора LXR или к стимуляции других биологических активностей рецептора LXR. в скрининге можно использовать либо систему анализа in vitro, либо систему анализа на клеточной основе.
Избирательность потенциальных агонистов LXR к различным рецепторам (например, LXR альфа, LXR бета, RXR или PPAR) можно тестировать, используя способы, хорошо известные в данной области техники, например конкурентные анализы приближенности сцинтилляции радиоактивного лиганда LXR описаны, например, в WO 01/41704, и конкурентные анализы связывания PPAR описаны, например, в Berger et al., J Biol Chem 274: 6718-6725,1999).
Тестируемые агенты, которые можно подвергать скринингу на новые агонисты LXR, включают полипептиды, бета-вращающие миметики, полисахариды, фосфолипиды, гормоны, простагландины, стероиды, ароматические соединения, гетероциклические соединения, бензодиазепины, олигомерные N-замещенные глицины, олигокарбаматы, полипептиды, сахариды, жирные кислоты, стероиды, пурины, пиримидины, производные, структурные аналоги или их комбинации. Некоторые тестируемые агенты представляют собой синтетические молекулы, а другие природные молекулы.
Тестируемые агенты получают из широкого ряда источников, включая библиотеки синтетических или природных соединений. Комбинаторные библиотеки можно получать для многих типов соединения, которые можно синтезировать ступенчато. Большие комбинаторные библиотеки соединений можно конструировать способом кодируемых синтетических библиотек (ESL), описанным в WO 95/12608, WO 93/06121, WO 94/08051, WO 95/35503 и WO 95/30642. Пептидные библиотеки можно также создавать способами фагового дисплея (см., например, Devlin, WO 91/18980). Библиотеки природных соединений в форме бактериальных, грибных, растительных и животных экстрактов могут быть получены из коммерческих источников или собраны в поле. Известные фармакологические агенты могут быть предметом направленных или случайных химических модификаций, таких как ацилирование, алкилирование, эстерификация, амидирование, с получением структурных аналогов.
Комбинаторные библиотеки пептидов или других соединений могут быть полностью рандомизированными без предпочтений последовательностей или констант в любом положении. Альтернативно библиотека может быть смещенной, то есть некоторые положения в пределах последовательности либо сохраняют постоянными, либо выбирают из ограниченного числа возможностей. Например, в некоторых случаях нуклеотиды или аминокислотные остатки рандомизированы в пределах определенного класса, например, гидрофобных аминокислот, гидрофильных остатков, стерически смещенных (либо малых, либо больших) остатков, в направлении образования цистеинов, для поперечной сшивки, пролинов для SH-3 доменов, серинов, треонинов, тирозинов или гистидинов для сайтов фосфорилирования, либо пуринов.
Тестируемые агенты могут представлять собой встречающиеся в природе белки или их фрагменты. Такие тестируемые агенты могут быть получены из природного источника, например клеточного или тканевого лизата. Библиотеки полипептидных агентов могут быть также получены, например, из библиотеки кДНК, имеющейся в продаже или созданной рутинными способами. Тестируемые агенты могут также представлять собой пептиды, например пептиды из аминокислот в количестве от примерно 5 до примерно 30, где предпочтительно от примерно 5 до примерно 20 аминокислот, и особенно предпочтительно от примерно 7 до примерно 15. Пептиды могут представлять собой продукты ферментативного расщепления встречающихся в природе белков, случайных пептидов или "смещенных" случайных пептидов. При некоторых способах тестируемые агенты представляют собой полипептиды или белки.
Тестируемые агенты могут также представлять собой нуклеиновые кислоты. Нуклеиново-кислотные тестируемые агенты могут представлять собой встречающиеся в природе нуклеиновые кислоты, случайные нуклеиновые кислоты или "смещенные" случайные нуклеиновые кислоты. Например, продукты ферментативного гидролиза прокариотических или эукариотических геномов можно использовать подобным образом, как описано выше для белков.
При некоторых предпочтительных способах тестируемые агенты представляют собой малые органические молекулы (например, молекулы с молекулярной массой не более чем примерно 1000). Предпочтительно анализы с высоким выходом адаптированы и используются для скрининга на такие малые молекулы. При некоторых способах комбинаторные библиотеки низкомолекулярных тестируемых агентов, как описано выше, можно легко использовать для скрининга на низкомолекулярные модуляторы рецептора LXR. Ряд анализов доступен для такого скрининга, например, как описано в Schultz (1998) Bioorg Med Chem Lett 8: 2409-2414; Weller (1997) Mol Divers. 3: 61-70; Fernandes (1998) Curr Opin Chem Biol 2: 597-603; и Sittampalam (1997) Curr Opin Chem Biol 1: 384-91.
Потенциальные агонисты LXR можно также идентифицировать на основании рационального дизайна. Например, Janowski et al. (Proc Natl Acad Sci USA 96: 266-71, 1999) раскрыты структурные требования к лигандам для LXR альфа и LXR бета. Было показано, что специфичное для положения моноокисление боковой цепи стерина оксистерина является необходимым для связывания LXR с высоким сродством и его активации. Усиленное связывание и активация могут быть также достигнуты посредством использования 24-оксо-лигандов, которые действуют как акцепторы водородной связи в боковой цепи. Кроме того, результатом введения атома кислорода на стериновое В-кольцо является лиганд с избирательностью к LXR альфа-подтипу.
Библиотеки тестируемых агентов, подлежащие скринингу заявленными способами, можно также создать на основании структурных исследований рецепторов LXR, их фрагментов или аналогов. Такие структурные исследования дают возможность идентификации тестируемых агентов, которые более вероятно связываются с рецептором LXR. Трехмерную структуру рецептора LXR можно исследовать рядом путей, например кристаллической структуры и молекулярного моделирования. Способы исследования белковых структур с использованием рентгенокристаллографии хорошо известны в литературе. См. Physical Biochemistry, Van Holde, K.E. (Prentice-Hall, N.J. 1971), pp.221-239, и Physical Chemistry with Applications to the Life Sciences, D. Eisenberg & D.C. Crothers (Benjamin Cummings, Menio Park 1979). Способы молекулярного моделирования описаны в литературе, например в патенте США №5612894, озаглавленном "System and method for molecular modeling utilizing a sensitivity factor", и в патенте США №5583973, озаглавленном "Molecular modeling method and system". Кроме того, белковые структуры можно также определять с помощью дифракции нейтронов и ядерного магнитного резонанса (ЯМР). См., например, Physical Chemistry, 4th Ed. Moore, W.J. (Prentice-Hall, N.J. 1972), и NMR of Proteins and Nucleic Acids, K. Wuthrich (Wiley-lnterscience, New York 1986).
В некоторых скрининговых анализах определяют связывание тестируемого агента с LXR или полипептидом LXR, содержащим его домен, связывающий лиганд. Связывание тестируемых агентов (например, полипептидов) с полипептидом LXR можно анализировать с помощью ряда способов, включая, например, меченые анализы связывания белок-белок in vitro, анализы сдвига электрофоретической подвижности, иммунологические анализы на связывание белка, функциональные анализы (анализы фосфорилирования и т.д.) и тому подобное. См., например, патенты США №№4366241; 4376110; 4517288; и 4837168; а также Bevan et al., Trends in Biotechnology 13: 115-122, 1995; Ecker et al., Bio/Technology 13: 351-360, 1995; и Hodgson, Bio/Technology 10: 973-980, 1992. Тестируемый агент можно идентифицировать путем обнаружения прямого связывания с полипептидом LXR, например, совместной иммунопреципитации с полипептидом LXR антителом, направленным на полипептид LXR. Тестируемый агент можно также идентифицировать путем обнаружения сигнала, который указывает на то, что этот агент связывается с полипептидом LXR, например гашения флуоресценции.
Конкурентные анализы обеспечивают подходящий формат для идентификации тестируемых агентов (например, пептидов или низкомолекулярных соединений), которые специфично связываются с полипептидом LXR. В таких форматах тестируемые агенты подвергают скринингу в конкуренции с соединением, уже известным как связывающее полипептид LXR. Известное связывающее соединение может представлять собой синтетическое соединение. Оно также может представлять собой антитело, которое специфично распознает полипептид LXR, например, моноклональное антитело, направленное против полипептида LXR. Если тестируемый агент ингибирует связывание соединения, известного как связывающее полипептид LXR, тогда тестируемый агент также связывает полипептид LXR.
Известны различные типы анализов конкурентного связывания, например: твердофазный прямой или косвенный радиоиммунологический анализ (РИА), твердофазный прямой или косвенный ферментативный иммунологический анализ (EIA), конкурентный сэндвич-анализ (см. Stahli et al., Methods in Enzymology 9: 242-253 (1983)); твердофазный прямой EIA биотин-авидин (см. Kirkland et al., J. Immunol. 137: 3614-3619 (1986)); твердофазный прямой меченый анализ, твердофазный прямой меченый сэндвич-анализ (см. Hariow and Lane, "Antibodies, A Laboratory Manual," Cold Spring Harbor Press (1988)); твердофазный прямой меченый РИА с использованием метки 125I (см. Morel et al., Mol. Immunol. 25(1): 7-15 (1988)); твердофазный прямой EIA биотин-авидин (Cheung et al., Virology 176: 546-552 (1990)); и прямой меченый РИА (Moldenhauer et al., Scand. J. Immunol. 32: 77-82 (1990)). Типично в такой анализ вовлечено использование очищенного полипептида, связанного с твердой поверхностью или клетками, несущими любой из них, немеченого тестируемого агента и меченого сравнительного соединения. Конкурентное ингибирование измеряют путем определения количества метки, связанной с твердой поверхностью или клетками в присутствии тестируемого агента. Обычно тестируемый агент присутствует в избытке. Модулирующие агенты, идентифицированные конкурентным анализом, включают агенты, связывающиеся с тем же эпитопом, что и сравнительное соединение, и агенты, связывающиеся с прилежащим эпитопом, существенно близким к эпитопу, связываемому сравнительным соединением, чтобы произошло стерическое затруднение. Обычно, когда конкурирующий агент присутствует в избытке, он будет ингибировать специфичное связывание сравнительного соединения с общим полипептидом-мишенью по меньшей мере на 50 или 75%.
Скрининговые анализы можно проводить либо в нерастворимом, либо в растворимом формате. Одним из примеров нерастворимых анализов является иммобилизация полипептида LXR или его фрагментов на твердофазном матриксе. Затем этот твердофазный матрикс приводят в контакт с тестируемыми агентами на период, достаточный для возможности связывания тестируемых агентов. После отмывки какого-либо не связавшегося материала от твердофазного матрикса присутствие агента, связанного с твердой фазой, дает возможность идентификации этого агента. Эти способы могут дополнительно включать стадию элюции связанного агента из твердофазного матрикса, выделяя посредством этого агент. Альтернативно, кроме иммобилизации полипептида LXR, тестируемые агенты связывают с твердым матриксом, а затем добавляют молекулу полипептида LXR.
Анализы в растворе включают некоторые из способов скрининга комбинаторных библиотек, описанных выше. В форматах анализа в растворе ни тестируемые агенты, ни полипептид LXR не связан с твердым носителем. Связывание полипептида LXR или его фрагмента с тестируемым агентом можно определить, например, по изменениям флуоресценции либо полипептида LXR, либо тестируемых агентов, либо того и другого. Флуоресценция может быть собственной или приданной путем мечения любого компонента флуорофором.
В некоторых анализах связывания либо полипептид LXR, либо тестируемый агент, либо третья молекула (например, антитело против полипептида LXR) могут быть представлены в виде меченых группировок, то есть ковалентно связанных или сшитых с детектируемой меткой или группой, либо поперечно сшиваемой группой, чтобы способствовать идентификации, обнаружению и количественному определению полипептида в данной ситуации. Эти детектируемые группы могут содержать обнаружимую полипептидную группу, например анализируемый фермент или эпитоп антитела. Альтернативно детектируемая группа может быть выбрана из ряда других детектируемых групп или меток, таких как радиоактивные метки (например, 125I, 32P, 35S), либо хемилюминесцентная или флуоресцентная группа. Подобным образом, детектируемая группа может представлять собой субстрат, кофактор, ингибитор или аффинный лиганд.
Связывание тестируемого агента с LXR можно также тестировать косвенно при анализе на клеточной основе. Например, ДНК-связывающий домен не рецепторного фактора транскрипции GAL4 можно слить с доменом, связывающим лиганд, LXR (например, LXR альфа). Полученную в результате конструкцию вводят в клетку-хозяина (например, клетки 293) вместе с конструкцией-репортером (например, UAS-содержащая конструкция люциферазного репортера). Затем трансфецированные клетки обрабатывают библиотеками тестируемых агентов и измеряют активность полипептида-репортера (например, активность люциферазы). Эффекты индивидуальных тестируемых агентов на активность полипептида-репортера оценивают относительно контроля (то есть, когда тестируемое соединение не присутствует).
Бесклеточный анализ чувствительности к лиганду (USA, cell-free ligand sensing assay) можно также использовать для идентификации новых агонистов LXR. Его можно проводить, как описано в данной области техники, например, Collins et al., J Med Chem. 45: 1963-6, 2002; и Spencer et al., J. Med. Chem. 44: 886-97, 2001. В этом анализе измеряют зависимый от лиганда рекрутмент пептида от соактиватора 1 стероидного рецептора (SRC1) к ядерному рецептору. При данном анализе (LISA) можно исследовать структурные требования к активации рецептора LXR тестируемыми агентами.
Кроме того, или в дополнение к обнаружению прямого связывания тестируемого агента с полипептидом LXR, потенциальные агонисты LXR для применения в способах по настоящему изобретению можно также исследовать на способность к активации других биологических активностей или клеточных активностей рецептора LXR. Тестируемые агенты, которые активируют рецептор LXR, можно идентифицировать путем мониторинга их эффектов на ряд клеточных активностей LXR. Клеточные активности LXR включают любую активность, опосредованную активированным рецептором LXR (например, транскрипционную регуляцию гена-мишени). Например, LXR трансактивирует экспрессию ряда генов-мишеней (например, АВСА1), ингибирует дифференциацию фибробластов в адипоциты, модулирует продуцирование ферментов, специфичных для мышц, например, креатинкиназы, модулирует захват глюкозы клетками и стимулирует пролиферацию клеток-миобластов. Степень, с которой тестируемый агент активирует рецептор LXR, можно идентифицировать путем тестирования на способность агента к усилению таких активностей LXR.
Таким образом, новый агонист LXR можно идентифицировать путем идентификации тестируемого агента, который усиливает экспрессию гена-мишени LXR (например, АВСА1, ABCQ1, SREBP1 или гена холестерин-7-гидроксилазы). Способы идентификации тестируемых агентов, которые индуцируют экспрессию гена-мишени LXR (например, повышение уровней мРНК АВСА1), раскрыты в данной области техники, например, Menke et al., Endocrinology 143: 2548-58, 2002; Sparrow et al., J. Biol. Chem. 277: 10021-7, 2002; и Murthy et al., J Lipid Res. 43:1054-64, 2002.
Кроме мониторинга экспрессии гена-мишени LXR, агонисты LXR можно также идентифицировать путем исследования других клеточных активностей, стимулируемых биохимическим путем LXR. Например, агонисты LXR модулируют уровень белка и, следовательно, активность специфичного для мышц фермента, креатинкиназы. Следовательно, агонисты LXR можно подвергать скринингу путем исследования тестируемых агентов на способность к модулированию активности креатинкиназы, например, как описано в Somjen et al., J Steroid Biochem Mol Biol 62: 401-8, 1997. Этот анализ можно проводить в клеточной линии, например, линии клеток-миобластов скелетных мышц мыши или клеточной линии первичных клеток-миобластов цыпленка. Эффекты тестируемых соединений на активность креатинкиназы в культивируемых клетках можно измерить в клеточных лизатах, используя имеющийся в продаже набор (поставляемый Sigma, St Louis, Mo.).
Модулирование других клеточных биологических активностей рецептора LXR можно также обнаружить, используя способы, хорошо известные и обычно практикуемые в данной области техники. Например, тестируемый агент можно анализировать на его активности при повышении оттока холестерина из клеток, таких как макрофаги (Menke et al., Endocrinology 143: 2548-58, 2002; и Sparrow et al., J. Biol. Chem. 277: 10021-7, 2002). Другие анализы включают зависимые от лиганда транскрипционные анализы (Schmidt et al., Mol Cell Endocrinol 155: 51-60, 1999), способы измерения способности агонистов LXR к вмешательству в процесс дифференциации преадипоцитов (фибробластов) в адипоциты (Plaas et al., Biosci Rep 1: 207-16, 1981; Hiragun et al., J Cell Physiol 134: 124-30, 1988; и Liao et al., J Biol Chem 270: 12123-32, 1995) или способности к стимуляции пролиферации клеток-миобластов (Konishi et al., Biochemistry 28: 8872-7, 1989; и Austin et al., J Neurol Sci 101: 193-7, 1991). В качестве контроля все эти анализы могут включать измерения до и после добавления тестируемого агента в систему анализа.
ТЕРАПЕВТИЧЕСКИЕ ПРИМЕНЕНИЯ
В настоящем изобретении предложены способы лечения дегенерации бета-клеток у субъекта, при которых указанному субъекту вводят агонист LXR.
Эти способы также находят применение при лечении заболевания, характеризующегося недостаточностью бета-клеток и, следовательно, дисфункцией инсулина (например, устойчивостью, отсутствием активности или дефицитом) и/или недостаточным транспортом глюкозы в клетки. Такие заболевания включают, но не ограничены ими, диабет, гипергликемию и ожирение. Лечение дегенерации бета-клеток также полезно для предупреждения или модулирования развития таких заболеваний или расстройств у субъекта, подозреваемого на склонность к таким заболеваниям или расстройствам, либо если эта склонность известна.
Более конкретно, поскольку теперь неожиданно обнаружено, что возможно избежать дегенерации бета-клеток, изобретение также направлено на способ предупреждения диабета типа I у субъекта, при котором субъекту вводят эффективное количество агониста LXR.
Агонисты LXR, которые следует использовать в данных применениях, могут представлять собой любой из известных агонистов LXR, которые описаны в данной области техники. Альтернативно способы терапии включают скрининг тестируемых агентов для идентификации новых агонистов LXR, как описано выше, и введение таких новых агонистов для лечения дегенерации бета-клеток или для лечения отмеченных выше заболеваний у субъекта.
Авторы настоящего изобретения наблюдали, что остановка дегенерации бета-клеток может быть достигнута после применения агониста LXR в течение очень короткого периода времени, например 21 сутки. Однако когда целью является усиление чувствительности к инсулину или облегчение симптомов диабета у субъекта, необходим более длительный период лечения. Для таких применений агонист LXR типично вводят субъекту в течение продолжительного периода времени, например по меньшей мере 30 суток, 60 суток, 90 суток или дольше.
Изобретение также относится к новому способу с улучшенным соблюдением пациентом режима и схемы лечения. Хорошо известно, что субъект с диабетом должен принимать большое количество лекарств. Настоящее изобретение дает возможность уменьшить число принимаемых лекарств и даже дает возможность приостановить лечение, как только значение гликемии и/или HbA1c стабилизируется или восстановить чувствительность к инсулину.
Таким образом, настоящее изобретение также направлено на способ лечения или облегчения диабета, при котором измеряют уровень циркулирующей глюкозы и/или HbA1c у субъекта и вводят этому субъекту ежесуточно эффективное количество агониста LXR до стабилизации гликемии и/или HbA1c, затем приостанавливают лечение, как только гликемия и/или HbA1c стабилизируется.
Агонист LXR предпочтительно вводят субъекту ежесуточно в течение по меньшей мере 30 суток. Его можно также вводить в течение по меньшей мере 60 суток, 90 суток или дольше.
Ежесуточное лечение может быть возобновлено, если гликемия и/или HbA1c более не стабилизированы или потеряна чувствительность к инсулину.
Способы измерения уровней циркулирующей глюкозы и/или HbA1c представляют собой способы, хорошо известные на предшествующем уровне техники, и можно использовать любой вид способов.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ (ИЛИ ЛЕКАРСТВЕННОЕ СРЕДСТВО)
Агонисты LXR по настоящему изобретению можно непосредственно вводить в стерильных условиях субъекту, подлежащему лечению. Модуляторы можно вводить отдельно или в виде активного ингредиента фармацевтической композиции. Терапевтическую композицию по настоящему изобретению можно комбинировать или применять в сочетании с другими терапевтическими агентами. Например, субъекта можно лечить агонистом LXR параллельно с другими общепринятыми противодиабетическими лекарственными средствами. Примеры таких известных противодиабетических лекарственных средств включают Actos (пиоглитазон, Takeda, Eli Lilly), Avandia (розиглитазон, Smithkline Beacham), Amaryl (глимепирид, Aventis), глипизид сульфонилмочевину (Generic) или Glucotrol (Ptizer), Glucophage (метформин, Bristol Meyers Squibb), Glucovance (глибурид/метформин, Bristol Meyers Squibb), Glucotrol XL (глипизид пролонгированного высвобождения, Pfizer), Глибурид (Micronase; Upjohn, Glynase; Upjohn, Diabeta; Aventis), Glyset (миглитол, Pharmacia & Upjohn), Metaglip (глипизид+метформин; таблетка фиксированной комбинации), Prandin (репаглинид, NOVO), Precose (акарбоза, Bayer), Rezulin (троглитазон, Parke Davis) и Starlix (натеглинид, Novartis).
Субъекта можно также лечить агонистом LXR параллельно с аналогами и производными GLP-1. Такие аналоги и производные GLP-1, которые можно применять согласно настоящему изобретению, включают те, на которые ссылаются в WO 99/43705 (Novo Nordisk A/S), WO 99/43706 (Novo Nordisk A/S), WO 99/43707 (Novo Nordisk A/S), WO 98/08871 (Novo Nordisk A/S), WO 99/43708 (Novo Nordisk A/S), WO 99/43341 (Novo Nordisk A/S), WO 87/06941 (The General Hospital Corporation), WO 90/11296 (The General Hospital Corporation), WO 91/11457 (Buckley et al.), WO 98/43658 (Eli Lilly & Co.).
Фармацевтические композиции по настоящему изобретению типично содержат по меньшей мере один активный ингредиент вместе с одним или более чем одним его приемлемым носителем. Фармацевтически приемлемые носители усиливают или стабилизируют композицию или облегчают изготовление препарата композиции. Фармацевтически приемлемые носители отчасти определяются конкретной композицией, подлежащей введению (например, нуклеиновая кислота, белок или модуляторные соединения), а также конкретным способом, применяемым для введения композиции. Они также должны быть как фармацевтически, так и физиологически приемлемыми в смысле совместимости с другими ингредиентами и безвредности для пациента. Этот носитель может принимать широкое разнообразие форм в зависимости от формы препарата, желательной для введения, например пероральной, подъязычной, ректальной, назальной или парентеральной.
Существует широкое разнообразие пригодных фармацевтически приемлемых носителей для практики настоящего изобретения (см., например, Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000). Без ограничения они включают среди прочего сироп, воду, изотонический солевой раствор, 5% декстрозу в воде или буферном растворе ацетата натрия или аммония, масла, глицерин, спирты, корригирующие агенты, консерванты, красящие агенты, крахмалы, сахара, разбавители, гранулирующие агенты, смазывающие агенты и связующие агенты. Можно также образовывать комплекс агониста LXR с белками-носителями, такими как овальбумин или сывороточный альбумин, перед его введением с целью усиления стабильности.
Фармацевтические композиции можно готовить в различных формах, таких как гранулы, таблетки, пилюли, суппозитории, капсулы, суспензии, мази, лосьоны и тому подобное. Концентрация терапевтически активного соединения в препарате может варьировать от примерно 0,1-100% мас./мас. Терапевтические препараты готовят любыми способами, хорошо известными в области фармацевтики. См., например, Oilman et al., eds., Goodman and Oilman's: The Pharmacological Bases of Therapeutics, 8th ed., Pergamon Press, 1990; Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000; Avis et al., eds., Pharmaceutical Dosage Forms: Parenteral Medications, published by Marcel Dekker, Inc., N.Y., 1993; и Lieberman et al., eds., Pharmaceutical Dosage Forms: Disperse Systems, published by Marcel Dekker, Inc., N.Y., 1990.
Препарат фармацевтической композиции по изобретению готовят таким образом, чтобы он был совместим с предназначенным путем введения. Примеры пригодных путей введения включают парентеральное, например внутривенное, внутрикожное, подкожное, пероральное (например, ингаляцию), чрескожное (местное), введение в слизистую оболочку и ректальное введение. Для парентерального введения препараты агонистов LXR по настоящему изобретению можно готовить разнообразными путями. Водные растворы модуляторов можно инкапсулировать в полимерных гранулах, наночастицах или других инъекционных препаратах депо, известных специалистам в данной области техники. Кроме того, соединения по настоящему изобретению можно также вводить инкапсулированными в липосомах. Композиции, в зависимости от их растворимости, могут находиться как в водном слое, так и в липидном слое, либо в том, что обычно называют липосомной суспензией. Гидрофобный слой, обычно, но не исключительно, содержит фосфолипиды, такие как лецитин и сфингомиелин, стероиды, такие как холестерин, больше или меньше ионных сурфактантов, таких как диацетилфосфат, стеариламин или фосфатидная кислота, и/или другие вещества гидрофобной природы.
В композиции могут быть добавлены другие активные фармацевтические ингредиенты, где желательно. Возможные антибактериальные, антисептические агенты и антиоксиданты могут также присутствовать в композициях, где они будут выполнять свои обычные функции. При некоторых применениях агонисты LXR готовят с носителями, которые будут защищать соединение от быстрого удаления из организма, такие как препарат регулируемого высвобождения, включая имплантаты и микроинкапсулированные системы доставки. Материал замедленного высвобождения, такой как глицерилмоностеарат или глицерилдистеарат, отдельно или с воском, можно включать в композиции. Можно также использовать биоразрушаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Способы изготовления таких препаратов будут очевидны специалистам в данной области техники. Эти материалы могут быть также получены из коммерческих источников, таких как Alza Corporation и Nova Pharmaceuticals, Inc.
Фармацевтические композиции могут быть включены в контейнер, упаковку или диспенсер вместе с инструкциями по применению.
ДОЗИРОВКИ
Субъектов, страдающих диабетом или родственными расстройствами, типично лечат фармацевтическими композициями по настоящему изобретению в течение продолжительного периода времени (например, по меньшей мере 30 суток, 60 суток, 90 суток или дольше). Фармацевтические композиции содержат фармацевтически эффективное количество или профилактически эффективное количество агониста LXR. Под термином "терапевтически эффективное количество" следует понимать то количество лекарственного средства или фармацевтического агента, которое будет вызывать биологический или медицинский ответ ткани, системы, животного или человека, который наблюдается исследователем, ветеринаром, врачом или другим клиницистом. Под термином "профилактически эффективное количество" следует понимать то количество фармацевтического лекарственного средства, которое будет предупреждать или уменьшать риск возникновения биологического или медицинского процесса, который стремятся предотвратить в ткани, системе, у животного или человека.
Пригодную терапевтическую дозу можно определить любым из хорошо известных способов, таким как клинические исследования на видах млекопитающих для определения максимальной переносимой дозы и на нормальных субъектах-людях для определения безопасной дозы. В частности, доза лиганда LXR, которую получает субъект, может быть выбрана таким образом, чтобы достичь регенерации бета-клеток; дозу, которую получает субъект, можно также титровать по времени с целью стабилизации гликемии и/или HbA1c или получения чувствительности к инсулину. Токсичность и терапевтическую эффективность агонистов LXR можно определить с помощью стандартных фармацевтических методик на клеточных культурах или подопытных животных, например, для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной в 50% популяции). Отношение дозы между токсическим и терапевтическим эффектами представляет собой терапевтический индекс, и его можно выразить в виде отношения LD50/ED50. Агонисты LXR, которые проявляют высокие терапевтические индексы, являются предпочтительными. Хотя агонисты LXR, которые проявляют токсические побочные эффекты, можно применять, следует позаботиться о разработке системы доставки, которая направляет такие агонисты LXR к месту пораженной ткани с целью минимизации потенциального вреда непораженным клеткам, посредством чего уменьшаются побочные эффекты.
Данные, полученные на основании анализов на клеточных культурах и исследований на животных, можно использовать при подборе диапазона дозировок для применения людьми. Дозировка таких соединений предпочтительно находится в интервале концентраций в кровообращении, который включает ED50 при небольшой токсичности или ее отсутствии. Дозировка может варьироваться в пределах этого интервала в зависимости от применяемой лекарственной формы и используемого пути введения. Для любого агониста LXR, применяемого в способе по изобретению, терапевтически эффективную дозу можно оценить первоначально на основании анализов на клеточных культурах. Дозу можно подобрать на животных моделях для достижения интервала концентрации в плазме крови, который включает IC50 (то есть концентрацию тестируемых агонистов LXR, при которой достигается половина максимального ингибирования симптомов), определенную на клеточной культуре. Такую информацию можно использовать для более точного определения полезных доз у людей. Уровни в плазме можно измерить, например, с помощью высокоэффективной жидкостной хроматографии.
В целом, за исключением некоторых обстоятельств, когда могут требоваться более высокие дозировки, предпочтительная дозировка агониста LXR обычно находится в интервале от примерно 0,001 до примерно 1000 мг, более обычно от примерно 0,01 до примерно 500 мг в сутки. Предпочтительная дозировка и режим введения агониста LXR может варьироваться для различных субъектов в зависимости от факторов, которые могут индивидуально наблюдаться лечащим врачом, таких как состояние или состояния, подлежащие лечению, выбор композиции для введения, включающей конкретный агонист LXR, возраст, масса и ответ индивидуального субъекта, тяжесть симптомов у субъекта и выбранный путь введения. В качестве общего правила количество вводимого агониста LXR составляет самую маленькую дозировку, которая эффективно и надежно предупреждает или минимизирует состояния субъектов. Таким образом, приведенные выше интервалы дозировки предназначены для обеспечения общего руководства и подтверждения изложенных здесь положений, но не предназначены для ограничения объема изобретения.
При некоторых применениях первый агонист LXR применяют в комбинации со вторым агонистом LXR или известным противодиабетическим лекарственным средством с целью достижения терапевтических эффектов, которые не могут быть достигнуты при применении одного агониста LXR индивидуально.
ОБРАБОТКА EX VIVO
Следующим признаком изобретения является способ ингибирования потери бета-клеток в островковой ткани поджелудочной железы путем приведения в контакт островковой ткани поджелудочной железы с агонистом LXR. Под уменьшением потери подразумевают, что в ткани поджелудочной железы находится на 10%, 20%, 30%, 40% или более бета-клеток в присутствии агониста LXR по сравнению с отсутствием агониста LXR.
Признаком изобретения также является способ повышения жизнеспособности или пролиферации островковых клеток поджелудочной железы путем приведения в контакт клетки с агонистом LXR. Кроме того, жизнеспособность островковых клеток поджелудочной железы повышают путем введения реципиенту трансплантата агониста LXR. Островковые клетки поджелудочной железы представляют собой первичные клетки островков. Альтернативно клетки представляют собой трансплантированные донорские клетки поджелудочной железы. Под жизнеспособностью подразумевают, что клетка исключает витальный краситель, такой как трипан. Жизнеспособные клетки также способны к пролиферации, дифференциации, росту и развитию. Жизнеспособность измеряют способами, известными в данной области техники, такими как окрашивание трипановым синим. Клетки приводят в контакт in vivo, in vitro или ex vivo. Агонист LXR вводят локально в область трансплантации. Альтернативно агонист LXR вводят системно. Агонист LXR вводят реципиенту трансплантата до или после трансплантации донорских островковых клеток поджелудочной железы. Возможно, агонист LXR вводят реципиенту трансплантата одновременно с трансплантацией донорских островковых клеток поджелудочной железы.
В изобретение также включены способы ингибирования клеточной гибели путем приведения в контакт клетки с агонистом LXR. Клетки приводят в контакт in vivo, in vitro или ex vivo. Клетка представляет собой клетку поджелудочной железы, такую как островковая клетка поджелудочной железы. Клеточная гибель представляет собой клеточную гибель, индуцированную оксидантным стрессом, или апоптотическую клеточную гибель.
Краткое описание графических материалов
На фиг.1 показан эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на глюкозу плазмы и после окончания обработки.
На фиг.2 показан эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на стимулируемую глюкозой секрецию инсулина в изолированных островках поджелудочной железы.
На фиг.3 показан эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на содержание инсулина в изолированных островках поджелудочной железы.
На фиг.4 показан эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на число собранных островков поджелудочной железы.
На фиг.5 показан эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 4 недель, на апоптоз в изолированных островках поджелудочной железы и срезах поджелудочной железы.
На фиг.6 показан эффект GW3965 на экспрессию мРНК PDX1 и GLUT2.
На фиг.7 показан эффект GW3965 (50 мг/кг) на регенерацию бета-клеток у мышей NOD.
На фиг.8 показан эффект GW3965 (50 мг/кг) на развитие инсулита у мышей NOD, где на пластине А представлены срезы поджелудочной железы контрольных мышей NOD, обработанных одним носителем, а на пластине В проиллюстрированы срезы поджелудочной железы обработанных GW3965 мышей NOD.
Примеры
Приведенные ниже примеры предложены для иллюстрации, но не для ограничения настоящего изобретения.
ЖИВОТНЫЕ
Самцы крыс Zucker Diabetic Fatty fa/fa (ZDF) представляют собой модель диабета типа II. Эти крысы устойчивы к инсулину, но имеют нормогликемию от рождения, и у них развивается диабет в возрасте примерно от недели 7 до недели 10. Во время переходного периода животные проходят через состояние нарушенной толерантности к глюкозе. Хотя эти животные имеют гиперинсулинемию перед возникновением диабета и во время ранних стадий диабета, впоследствии они утрачивают стимулированную глюкозой секрецию инсулина и, наконец, приобретают почти полную инсулинопению.
Самцов крыс ZDF fa/fa и ZDF fa+ получали от Genetic Models (Indianapolis, IN) в возрасте 6 недель и использовали в экспериментах в возрасте 12 недель после 3 недель пероральной обработки носителем и GW3965 (10 мг/кг). Соединения давали один раз в сутки (в 17 ч) через пероральный зонд. В момент использования крысы ZDF fa/fa весили 310-350 г, тогда как крысы ZDF fa+ такого же возраста весили 245-285 г. Животные ZDF fa+ и ZDF fa/fa имели свободный доступ к диабетогенному корму (Purina 5008). Животных держали по 5 на клетку в комнате с регулируемой температурой и влажностью при световом цикле 12/12 ч. Образцы крови брали через 16 ч после введения соединения из хвостовой вены и измеряли на клиническом химическом анализаторе Konelab 300 (Labsystem).
Мыши NOD (non-obese diabetes, диабет без ожирения) представляют собой экспериментальную модель спонтанного диабета, напоминающего диабет типа I (инсулинозависимый) человека, который является результатом прогрессирующей инвазии островков и разрушения бета-клеток аутоиммунными Т-клетками. Эта модель спонтанного диабета дает уникальную возможность исследования аутоиммунных Т-клеток, вовлеченных в процесс разрушения бета-клеток, и установки превентивных стратегий до клинического проявления заболевания.
ПОЛУЧЕНИЕ ПРЕПАРАТОВ ОСТРОВКОВ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ КРЫС
Препараты островков поджелудочной железы крыс получали путем ферментативного гидролиза коллагеназой и очистки в градиенте плотности. Фермент коллагеназу типа XI (Sigma Chemical, St. Louis, МО) использовали для ферментативного гидролиза поджелудочной железы. После голодания в течение ночи животных подвергали глубокой анестезии и умерщвляли. Затем проток поджелудочной железы канюлировали и медленно инъецировали 20 мл переваривающего раствора при 37°С (коллагеназа 30 мг/мл, растворенная в 20 мл сбалансированного солевого раствора Хэнкса [HBSS]) (Sigma Chemical, St. Louis, МО) для расправления ткани. После расправления поджелудочную железу помещали в 20 мл стеклянный стакан, содержащий 5 мл переваривающего раствора при 37°С, который устанавливали в водяную баню с качалкой при 37°С на 8 минут. После центрифугирования при 633 g в течение 1 мин при 4°С надосадочную жидкость удаляли и островки промывали три раза HBSS путем центрифугирования при 633 g в течение 1 мин при 4°С. Для процедуры очистки островки ресуспендировали в 4 мл 80% Histopaque 1,077 (Sigma) и 4 мл HBSS. После центрифугирования при 1943 g в течение 20 мин при 4°С островки выделяли на границе раздела между слоями Histopaque и HBSS. Островки ресуспендировали в среде RPMI 1640, содержащей 11 мМ глюкозу, 10% ФСТ (фетальную сыворотку теленка), пенициллин (100 ед/мл), стрептомицин (100 мкг/мл) и L-глутамин (2 мМ), и культивировали в течение ночи при 37°С в СО2 инкубаторе.
ОЦЕНКА АПОПТОЗА ПУТЕМ ИЗМЕРЕНИЯ ФРАГМЕНТОВ ДНК
Апоптоз островковых клеток оценивали с помощью твердофазного иммуноферментного анализа обнаружения клеточной гибели (ELISA) с набором от Roche в соответствии с методиками, рекомендованными изготовителем и применимыми к эксперименту с островками Лангерганса крыс. После инкубации в течение ночи в среде RPMI 1640, содержащей 11 мМ глюкозу, 10% ФСТ, пенициллин (100 ед/мл), стрептомицин (100 мкг/мл) и L-глутамин (2 мМ), группы из 10 островков инкубировали в течение 30 мин с лизирующим буфером при комнатной температуре, а затем центрифугировали при 200 g в течение 10 мин при 4°С для апоптоза. Аликвоты надосадочной жидкости (20 мкл) помещали в лунки микротитрационного планшета, покрытые стрептавидином. Затем добавляли суммарно 80 мкл смеси, содержащей антитело анти-гистон-биотин и антитело анти-ДНК-пероксидаза, и давали возможность инкубации в течение 120 мин при 37°С. Затем препараты отмывали и добавляли 100 мкл раствора, содержащего ABTS (2,2'-азино-ди[3-этилбензтиазалинсульфонат]) (субстрат для пероксидазы). По окончании 5-минутной инкубации поглощение образцов считывали спектрофотометрическим путем при 405 нм. Результаты выражали в виде процента от внутреннего стандарта (нуклеосомных частиц).
ОЦЕНКА АПОПТОЗА С ПОМОЩЬЮ ОКРАШИВАНИЯ TUNEL
Апоптоз островковых клеток определяли путем окрашивания опосредованным трансферазой концевым мечением dUTP (TUNEL. transferase-mediated dUTP nick-end labeling). Клеточную гибель идентифицировали по 3'-концевому мечению in situ фрагментированной ДНК биотинилированным дезоксиуридинтрифосфатом. Каждый кусочек поджелудочной железы резали в длину на срезы толщиной 5 мкм, которые собирали на стекла, покрытые желатином. Четыре среза через регулярные интервалы собирали как из кусочков головки, так и хвоста. После ферментативного гидролиза протеиназой К (2 мкг/мл; Sigma) срезы последовательно промывали в буфере TdT (0,5 моль/л какодилата [рН 6,8], 1 ммоль/л хлорида кобальта, 0,15 моль/л NaCl), а затем инкубировали при 37°С с TdT (25U; Boehringer-Mannheim, Mannheim, Germany) и биотинилированным dUTP (1 нмоль/мкл). После отмывки в буфере ТВ (300 ммоль/л NaCl, 30 ммоль/л цитрата натрия) для остановки реакции включение биотинилированного dUTP обнаруживали модифицированным способом комплекса авидин-биотин. Наносили комплекс стрептавидин-биотин, конъюгированный с пероксидазой хрена (Vector Laboratories). Затем срезы проявляли, используя DAB в качестве хромогена, и слегка окрашивали контрастно гематоксилином и укрепляли на dePex (BDH Laboratory, Poole, England).
СТИМУЛИРУЕМАЯ ГЛЮКОЗОЙ СЕКРЕЦИЯ ИНСУЛИНА И СОДЕРЖАНИЕ ИНСУЛИНА В ОСТРОВКАХ
После 30 мин периода предварительной инкубации при 37°С в течение 30 мин в бикарбонате Кребса-Рингера (КРВ)-растворе HEPES (129 мМ NaCl, 4,8 мМ KCl, 1,2 мМ MgSO4, 1,2 мМ КН2РО4, 2,5 мМ CaCl2, 5 мМ NaHCO3 и 10 мМ HEPES при рН 7,4), содержащем 0,5% БСА (бычьего сывороточного альбумина) и без глюкозы партии из 3 островков сравнимого размера помещали в лунки микротитрационного планшета, содержащие 200 мкл раствора KRB-HEPES, содержащего 0,5% БСА и 3,3 мМ глюкозы, при 37°С в течение 1 часа при качании и атмосфере 5% СО2 и 95% О2. В конце этой провокации среду полностью удаляли и заменяли KRB-HEPES, содержащим 16,7 ммоль/л глюкозы. После дополнительной 1-часовой инкубации среду удаляли и замораживали. Содержание инсулина измеряли в тех же островках после экстракции кислотой-спиртом в течение ночи (этанол 75%+HCl 0,15 н.). Концентрации инсулина в различных средах определяли с помощью теста ЭЛАЙЗА на инсулин крысы (Insulin Rat Elite plus, MERCODIA).
КОЛИЧЕСТВЕННАЯ ОБРАТНАЯ ТРАНСКРИПЦИЯ-ПЦР В РЕАЛЬНОМ ВРЕМЕНИ
Экспрессию генов панкреатического дуоденального гена 1 гомеобокса, Pdx1 (Genbank: NM_022852) и усиленного транспортера глюкозы, члена 2, Glut2 (Genbank: NM_012879) измеряли с помощью количественной обратной транскрипции (ОТ)-ПЦР в реальном времени.
Препараты суммарной РНК из свежевыделенных островков поджелудочной железы крыс получали, используя набор Rneasy 96 Kit в соответствии с протоколом изготовителя (Qiagen, France), обрабатывали ДНКазой I для удаления загрязняющей ДНК (Stratagene, France) и определяли количественно, используя наборы iScript cDNA Synthesis и IQ SYBR Green Supermix (Biorad, France). Количественную ОТ-ПЦР в реальном времени проводили, используя прибор и программное обеспечение iCycler Detection System (Bio-Rad). Поскольку предсказанный размер амплифицированных кДНК и значения Tm для праймеров были сходными, использовали единые условия амплификации (то есть отжиг при 60°С). Мониторинг специфичности амплифицированных продуктов проводили путем построения кривых отжига в конце каждой амплификации. Уровни экспрессии количественно определяли (единицы суммарной РНК) путем построения стандартной кривой из серий по шесть точек. Образцы РНК нормализовали по сравнению с определенными уровнями 18S рРНК.
Последовательности праймеров
PDX1 (прямой): CCACAGCCCTCCAGCATCG
PDX1 (обратный): CAGACCCGCTCACCCTCAG
Glut2 (прямой): ACCAGCACATACGACACCAGAC
Glut2 (обратный): GACACAGACAGAGACCAGAGCATAG
18S (прямой): CGTCTGCCCTATCAACTTTCG
18S (обратный): GATGTGGTAGCCGTTTCTCAG
АНАЛИЗ ДАННЫХ
Данные выражают в виде среднего ±СОС (стандартная ошибка среднего). Сравнения между различными группами были получены с помощью t-критерия Стьюдента. Различия при р<0,05 между экспериментальными группами считали статистически значимыми. Для сравнений группу ZDF fa/fa сравнивают с группой ZDF fa+, а группу ZDF fa/fa GW3965 сравнивают с группой ZDF fa/fa.
СРЕЗЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ И ГИСТОЛОГИЧЕСКИЙ АНАЛИЗ
Не голодавших мышей умерщвляли через двое суток после последнего введения соединений. Поджелудочные железы вырезали и обрабатывали для общепринятых гистологических исследований после фиксации в спиртовом растворе Буэна. Срезы толщиной 5 мкм окрашивали гематоксилином-эозином, как описано ранее Thivolet C.H. et al., 1991, Diabetologia, 34, 314-319.
Пример 1
Эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на глюкозу плазмы и после окончания обработки
Графики изменений глюкозы в плазме показаны на фиг.1. В начале исследования гликемия была сходной у крыс ZDF, обработанных носителем и GW3965.
На сутки 6 уровни глюкозы в плазме в группе, обработанной носителем, повысились выше 400 мг/мл. Обработка GW3965 в значительной степени предотвращала этот подъем и поддерживала гликемию на низком уровне вплоть до окончания обработки на сутки 21.
От суток 21 до конца эксперимента животные не получали никакой обработки. На сутки 44 гликемия все еще была значимо снижена у крыс ZDF, которые прежде были обработаны GW3965. Это указывает на то, что долгосрочная регуляция гликемии, которая следует за ограниченным периодом обработки GW3965, вероятно, являлась результатом восстановления функции и массы бета-клеток в соответствии с данными ex vivo. Данный пример подтверждает предположение о новом способе лечения, основанном на введении агониста LXR в течение ограниченных периодов времени пациентам с диабетом.
Результаты показывают, что, хотя эффект на триглицериды снижается после окончания обработки, снижение глюкозы в крови поддерживалось в течение более чем одного месяца. Агонистические лиганды LXR можно, таким образом, вводить либо в виде ударной дозы в течение одних или нескольких суток и последующего повторного введения ударной дозы на еженедельной основе или дольше в зависимости от ударной дозы, либо чередовать через каждые следующие сутки, либо чередовать через каждую следующую неделю (или несколько суток).
Пример 2
Стимулируемая глюкозой секреция инсулина в островках поджелудочной железы, выделенных из крыс ZDF. обработанных перорально в течение 3 недель GW3965 (10 мг/кг).
Результаты представлены на фиг.2. В присутствии 3,3 ммоль/л и 16,7 ммоль/л глюкозы секреция инсулина островками крыс ZDF по сравнению с крысами fa+ была снижена на 82,6 и 69,6%, соответственно. В островках крыс ZDF, обработанных GW3965, по сравнению с обработанными носителем секреция инсулина в ответ на 3,3 ммоль/л и 16,7 ммоль/л глюкозы значимо повышалась на 133,8% и 74,5%, соответственно. Таким образом, показано, что стимулируемая глюкозой секреция инсулина в островках поджелудочной железы крыс ZDF улучшается посредством агониста LXR.
Пример 3
Эффект GW3965 (10 мг/кг), вводимого перорально крысам ZDF в течение 3 недель, на содержание инсулина изолированных островков поджелудочной железы
Результаты представлены на фиг.3. Содержание инсулина было сильно снижено в островках крыс ZDF по сравнению с островками крыс fa+ (4,78±0,86 против 27,74±6,85 нг/островок). Содержание инсулина островков ZDF, обработанных GW3965 (10 мг/кг), значимо повышалось по сравнению с островками ZDF, обработанных носителем (13,05±2,49 против 4,78±0,86 нг/островок).
Данный пример демонстрирует, что содержание инсулина островков поджелудочной железы повышается агонистом LXR.
Пример 4
Эффект GW3965 (10 мг/кг). вводимого перорально крысам ZDF в течение 3 недель, на число собранных островков Лангерганса поджелудочной железы
Результаты представлены на фиг.4. Число островков поджелудочной железы, собранных у крыс ZDF, было ниже, чем собранных у крыс fa+ (51±13 и 120±28 островков/крыса, соответственно). У крыс ZDF, обработанных GW3965, было повышенное число островков поджелудочной железы по сравнению с крысами, обработанными носителем (80±16 и 51±13 островков/крыса, соответственно).
Таким образом, число островков Лангерганса поджелудочной железы повышается после обработки in vivo агонистом LXR.
Пример 5
Эффект GW3965 ПО мг/кг). вводимого перорально крысам ZDF в течение 4 недель, на апоптоз в изолированных островках поджелудочной железы и срезах поджелудочной железы
В данном примере исследовали скорость апоптоза островков поджелудочной железы. Результаты представлены на фиг.5 (см. панель А). Число фрагментов ДНК, как измерено с помощью специфичного анализа ELISA после культивирования в течение ночи в среде RPMI, было значимо повышено в островках крыс ZDF по сравнению с островками контрольных крыс fa+ (43,1±9,2% против 19,9±6,2%). Апоптоз в островках поджелудочной железы, выделенных из крыс ZDF, обработанных GW39065, по сравнению с крысами, обработанными носителем, был снижен (34,0±7,5% и 43,1±9,2%, соответственно).
Апоптоз также оценивали, используя способ окрашивания TUNEL, на срезах поджелудочной железы 5 животных из 8-недельных крыс ZDF, обработанных GW3965 при 10 мг/кг в течение 4 недель. Результаты представлены на фиг.5 (см. панель В) и показывают, что апоптоз в срезах поджелудочной железы крыс ZDF, обработанных GW3965, был значимо снижен по сравнению с крысами, обработанными носителем (1,41±0,96% и 4,55±1,29% апоптоза. соответственно (p<0,05).
Пероральная обработка in vivo агонистом LXR позволяет снизить апоптоз соответствующих изолированных островков поджелудочной железы. Конкретно, поскольку бета-клетки представляют 85-95% островка поджелудочной железы, эти результаты показывают, что введение агониста LXR значимо снижает апоптоз бета-клеток.
Пример 6
Эффект GW3965 на экспрессию мРНК PDX1 и GLUT2 (Фиг.6)
В данном примере определяли экспрессию мРНК PDX1 и Glut2. Как описано ранее, PDX1 и Glut2 являются маркерами регенерации бета-клеток. Экспрессия мРНК PDX1 и Glut2 была значительно снижена в островках из крыс ZDF против контрольных крыс fa+ (0,17 против 0,38±0,21 и 0,27 против 0,40±0,22, соответственно). Экспрессия мРНК PDX1 и Glut2 была повышена в островках из крыс ZDF, обработанных GW3965, по сравнению с обработанными носителем крысами ZDF. Значения составляли 0,35±0,11 для PDX1 и 0,55±0,23 для Glut2.
Данный пример демонстрирует, что бета-островковые клетки поджелудочной железы претерпевают регенерацию после обработки агонистом LXR.
Пример 7
Эффект GW3965 (50 мг/кг) на регенерацию бета-клеток у мышей NOD (Фиг.7)
Мышей NOD в возрасте 4 недель обработали в течение 4 недель (5 суток/7) носителем или GW3965 при 50 мг/кг. Гистологические исследования поджелудочной железы провели, как описано выше.
Как четко показано путем образования новых островков, указанных в кружках на фиг.7, бета-островковые клетки поджелудочной железы претерпевают регенерацию после обработки агонистом LXR. Кроме того, регенерация бета-клеток происходит при морфологической структуре островков Лангерганса поджелудочной железы, эквивалентной структуре нормальных островков Лангерганса поджелудочной железы.
Пример 8
Эффект GW3965 (50 мг/кг) на развитие инсулита у мышей NOD (Фиг.8)
Развитие инсулита анализировали в поджелудочной железе. Обычно описывают пять состояний инсулита: состояние 0 характеризуется нормальными островками без инфильтрации моноцитов (Т лимфоцитов), состояние 1 соответствует периинсулиту, состояние 2 с инсулитом ниже 50%, состояние 3 с инсулитом выше 50% и состояние 4 с апоптозом. Состояние 4 не наблюдалось у мышей в возрасте 8 недель, это состояние появлялось около 9-10 недель.
Мыши NOD в возрасте 4 недель были обработаны в течение 4 недель (5 суток/7) носителем или GW3965 при 50 мг/кг. Гистологические исследования поджелудочной железы провели, как описано выше.
Как показано на панели А, на срезах поджелудочной железы контрольных мышей NOD, обработанных только носителем, проявляется инсулит, тогда как поджелудочная железа обработанных GW3965 мышей NOD оказывается защищенной против инфильтрации лимфоцитов, как проиллюстрировано на панели В. Действительно, 94% нормальных островков наблюдали в поджелудочной железе обработанных GW3965 мышей NOD, тогда как у контрольных мышей NOD показано только 32% нормальных островков (данные не показаны).
Данный пример демонстрирует, что введение in vivo агониста LXR GW3965 способно ингибировать инсулит, развившийся у мышей NOD.
Данные примеры показывают, что агонист LXR, GW3965, повышает стимулируемую глюкозой секрецию инсулина, содержание инсулина и число островков, снижает апоптоз бета-клеток и стимулирует регенерацию бета-клеток у крыс ZDF с тяжелым диабетом. Это было связано с улучшением диабетического состояния и значимым снижением гликемии и HbA1c после 21 суток обработки.
При определении возможных механизмов, лежащих в основе противодиабетических действий агонистов LXR, авторами изобретения сделано открытие, что обработка GW3965 привела к значимому снижению апоптоза бета-клеток и значимому повышению содержания инсулина. Кроме того, авторами изобретения обнаружено восстановление уровня мРНК PDX1 и Glut2 до контрольных значений, полученных у крыс fa+.
Через тридцать суток после окончания обработки GW3965 авторы изобретения продемонстрировали, что соединение обладает долгосрочным защитным и превентивным эффектом на развитие диабета типа II, поскольку глюкоза в плазме все еще поддерживалась на низком уровне.
Экспериментальные результаты, подобные продемонстрированным выше в примерах, получены путем использования других агонистов LXR, таких как Т0901317 (данные не представлены).
Понятно, что примеры и варианты осуществления изобретения, описанные здесь, предназначены только для иллюстративных целей, и что различные модификации или изменения в их свете будут предложены специалистам в данной области техники и должны быть включены в сущность и объем данной заявки и объем прилагаемой формулы изобретения. Хотя любые способы и материалы, подобные или эквивалентные описанным здесь, можно использовать в практике или тестировании настоящего изобретения, описаны предпочтительные способы и материалы.


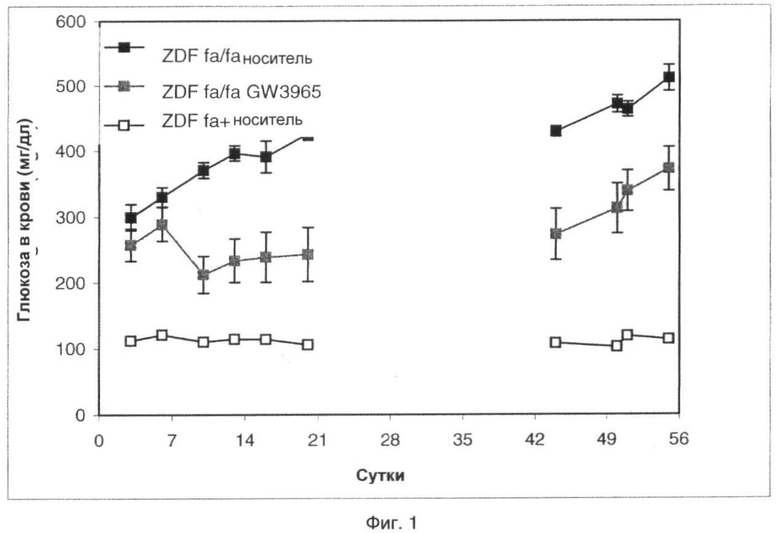
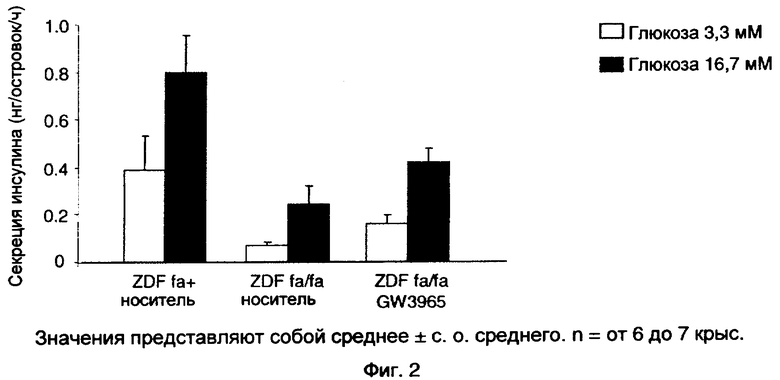
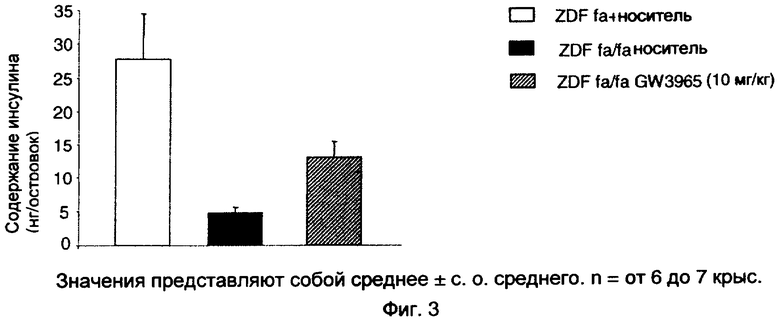
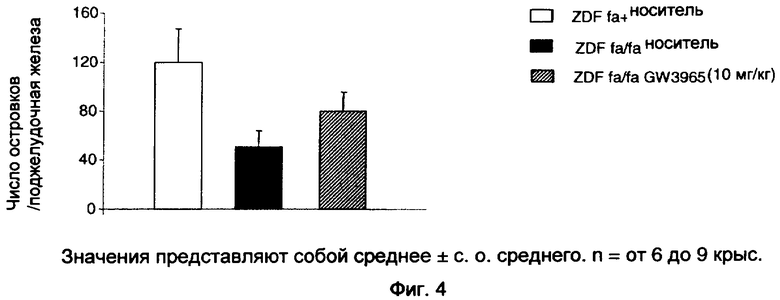
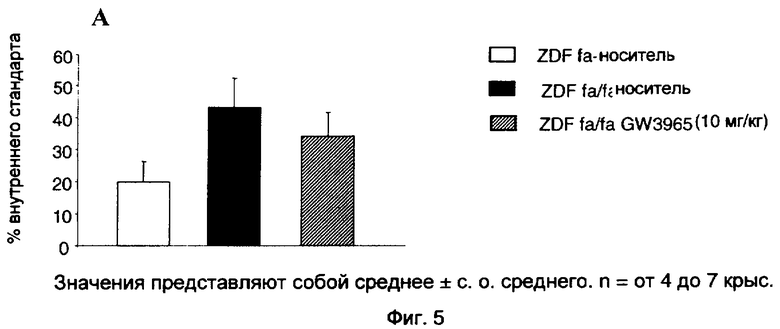
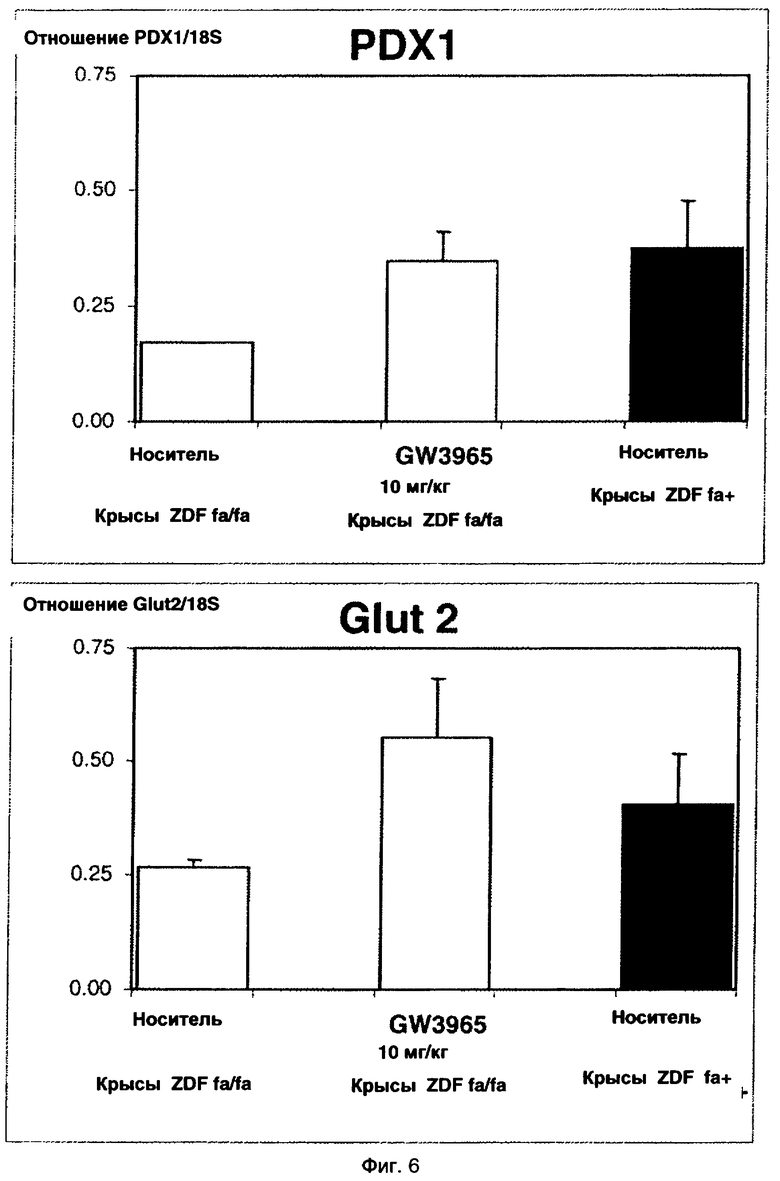
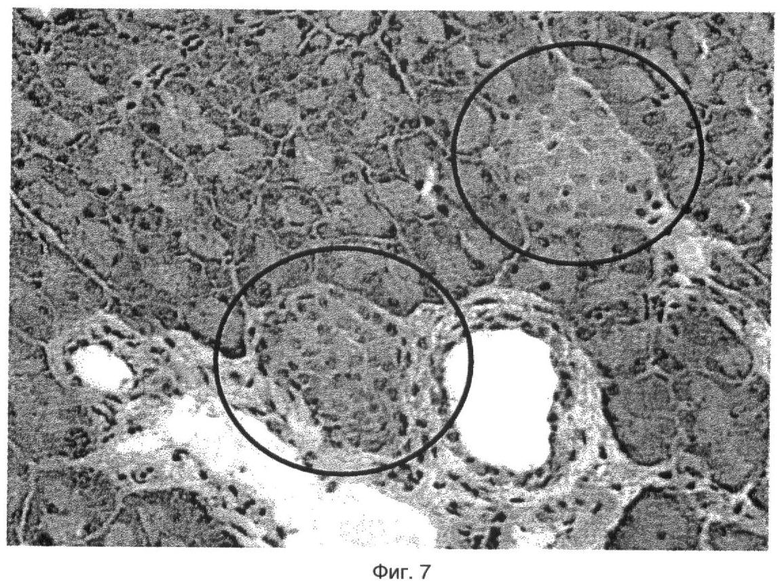
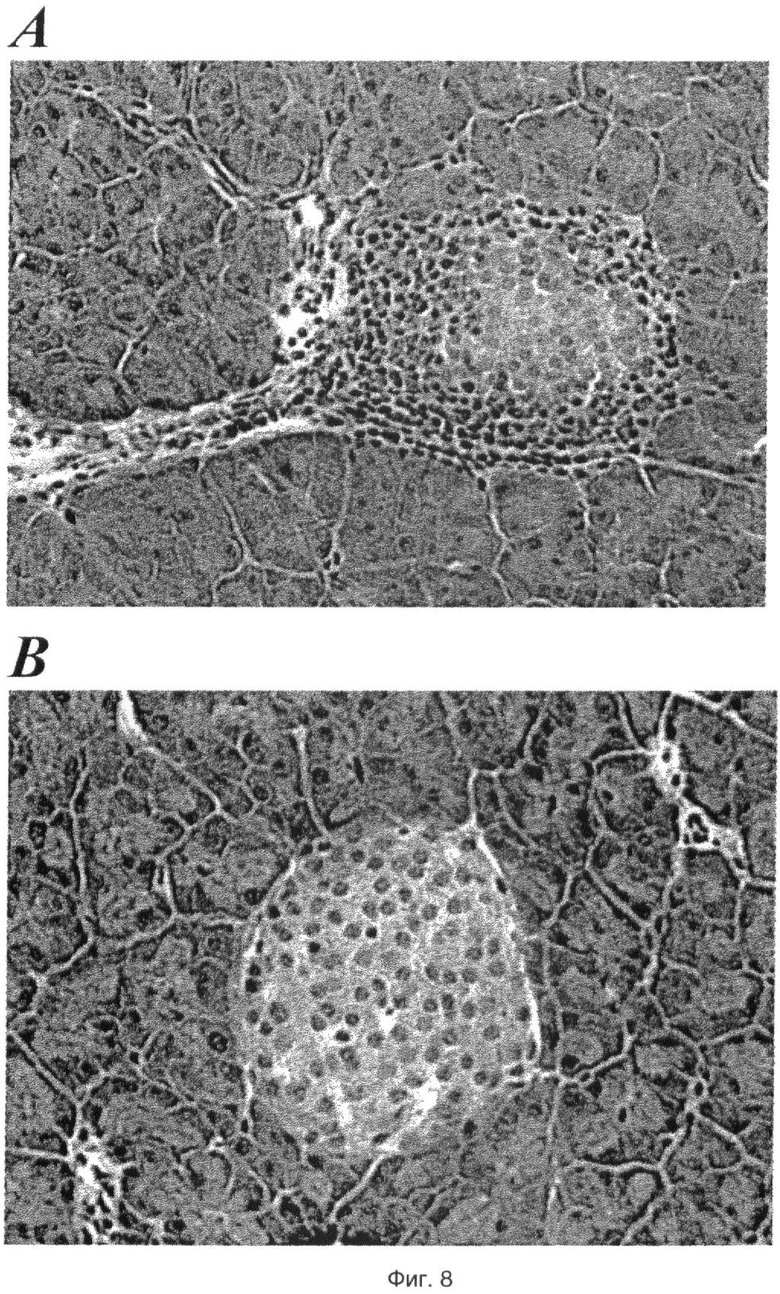
Изобретение относится к лекарственным средствам и касается применения агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения диабета типа I и способа повышения жизнеспособности первичных островковых клеток поджелудочной железы ex vivo, при котором приводят в контакт указанные островковые клетки поджелудочной железы с агонистом LXR. Изобретение направлено на расширение арсенала средств для лечения диабета типа I. 2 н. и 11 з.п. ф-лы, 8 ил.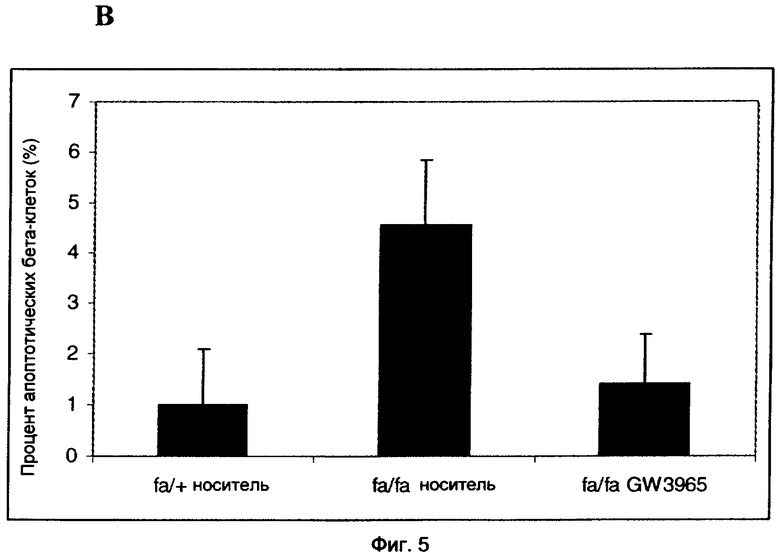
1. Применение агониста LXR для изготовления лекарственного средства, полезного для лечения и/или предупреждения диабета типа I.
2. Применение по п.1, где указанное лекарственное средство применяют для регенерации бета-клеток.
3. Применение по п.1, где указанное лекарственное средство применяют для повышения жизнеспособности бета-клеток.
4. Применение по п.3, где указанное лекарственное средство применяют для повышения жизнеспособности трансплантированных донорских островковых клеток поджелудочной железы у реципиента трансплантата.
5. Применение по п.1, где агонист LXR представляет собой агонист LXR альфа.
6. Применение по п.1, где агонист LXR выбран из группы, состоящей из GW3965 и T0901317.
7. Применение по п.1, где указанное лекарственное средство находится в форме, пригодной для перорального введения.
8. Применение по п.1, где указанное лекарственное средство применяют ежесуточно в течение по меньшей мере 30 суток.
9. Применение по п.1, где указанное лекарственное средство применяют в сочетании с известным противодиабетическим лекарственным средством.
10. Применение по п.1, где указанное известное противодиабетическое лекарственное средство выбрано из группы, состоящей из метформина, пиоглитазона, розиглитазона, глимепирида, глипизида, глибурида/метформина, глибурида, миглитола, глипизида + метформина, репаглинида, акарбозы, троглитазона, натеглинида и агониста GLP-1.
11. Способ повышения жизнеспособности первичных островковых клеток поджелудочной железы ex vivo, при котором приводят в контакт указанные островковые клетки поджелудочной железы с агонистом LXR.
12. Способ по п.11, где агонист LXR представляет собой агонист LXR альфа.
13. Способ по п.11, где агонист LXR выбран из группы, состоящей из GW3965 и T0901317.
| EFANOV A.M | |||
| et al | |||
| Liver X receptor activation stimulates insulin secretion via modulation of glucose and lipid metabolism in pancreatic beta-cells | |||
| Diabetes | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| CAO G | |||
| et al | |||
| Antidiabetic action of a liver x receptor agonist mediated by inhibition of hepatic gluconeogenesis | |||
| The Journal of Biological Chemistr., 2003, | |||

