Область техники, к которой относится изобретение
Настоящее изобретение касается новых идентифицированных полипептидов и применения таких полипептидов в терапевтических целях. Более конкретно полипептиды по настоящему изобретению применимы для стимуляции секреции инсулина бета-клетками поджелудочной железы в глюкозо-зависимом режиме, что тем самым обеспечивает возможный путь лечения тех субъектов, у которых имеется метаболическое расстройство, такое как диабет или нарушенная переносимость глюкозы, являющаяся преддиабетическим состоянием.
Предшествующий уровень техники
Диабет характеризуется нарушением метаболизма глюкозы, что, помимо прочего, проявляется в повышении содержания глюкозы в крови у больных-диабетиков. Лежащие в основе указанного дефекты позволяют классифицировать диабет на две основные группы: диабет 1-го типа или инсулин-зависимый сахарный диабет (IDDM), который возникает тогда, когда у больного наблюдается недостаток бета-клеток в поджелудочной железе, вырабатывающей инсулин, и диабет 2-го типа или инсулин-независимый сахарный диабет (NIDDM), который имеется у больных с нарушенной функцией бета-клеток и измененной активностью инсулина.
Больных с диабетом 1-го типа сейчас лечат инсулином, в то время как большинство больных с диабетом 2-го типа лечат с помощью средств, которые стимулируют функцию бета-клеток, или с помощью средств, которые усиливают восприимчивость тканей больных к инсулину. Со временем почти половина больных с диабетом 2-го типа утрачивают способность реагировать на указанные средства, после чего их переводят на инсулинотерапию. Лекарственными средствами, ныне используемыми в лечении диабета 2-го типа, являются:
Ингибиторы α-глюкозидазы (PRECOSE®, VOGLIBOSETM и MIGLITOL®). Ингибиторы α-глюкозидазы снижают перемещение глюкозы после принятия пищи за счет задержки всасывания глюкозы из кишечника. Такие лекарства безопасны и обеспечивают лечение больных с диабетом, выраженным в слабой и умеренной степени. Однако в научной литературе сообщается о побочных эффектах на желудочно-кишечный тракт.
Сенсибилизаторы инсулина. Сенсибилизаторы инсулина - это лекарственные средства, которые усиливают реакцию организма на инсулин. Тиозолидиндионы, такие как REZULINTM (троглитазон), активируют γ-рецептор PPAR и модулируют активность группы генов, которые пока точно не определены. Хотя указанные лекарства и являются эффективными, они связаны с гепатотоксичностыо. Из-за токсичного действия на печень REZULIN был изъят с фармацевтического рынка.
Средства, повышающие секрецию инсулина (сульфонилмочевины и другие средства, которые активны через АТФ-зависимый калиевый канал). СФМ обеспечивают стандартное лечение диабета 2-го типа, при котором имеется гликемия натощак от слабой до умеренной степени. Ограничения в применении СФМ связаны с возможной индукцией гипогликемии, увеличением веса и высокой частотой первичных и вторичных случаев безуспешного лечения. У 10-20% исходно получающих лечение больных не удается выявить существенный эффект лечения (первичные случаи безуспешного лечения). Вторичные случаи безуспешного лечения выявляют еще у 20-30% больных после 6 месяцев применения СФМ. Инсулинотерапия становится необходимой для 50% получавших СФМ по прошествии 5-7 лет лечения (A.J.Scheen et al., 1989, Diabetes Res. Clin. Pract., 6, 533-543).
GLUCOPHAGE™ (метформина гидрохлорид) является бигуанидом, который снижает содержание глюкозы в крови за счет подавления высвобождения глюкозы из печени и усиления периферического поглощения и утилизации глюкозы. Данное средство эффективно в снижении глюкозы в крови у больных от слабой до умеренной тяжести и не обладает побочными эффектами, связанными с увеличением веса или возможной индукции гипогликемии. Однако GLUCOPHAGE обладает рядом побочных действий, включая желудочно-кишечные расстройства и лактацидоз. GLUCOPHAGE противопоказан диабетикам старше 70 лет и больным с нарушениями функций почек или печени. Наконец, GLUCOPHAGE характеризуется той же частотой первичных и вторичных случаев безуспешного лечения, что и СМФ.
Инсулин назначают после того, как диета, упражнения и пероральные лекарственные средства не способны адекватно контролировать уровень глюкозы в крови. Такое лечение обладает недостатками, связанными с необходимостью введения инъекций, возможностью развития гипогликемии и набором избыточного веса.
С учетом имеющихся проблем в современных способах лечения терапия диабета 2-го типа нуждается в новых подходах. В частности, необходимы новые способы лечения для поддержания нормальной (глюкозо-зависимой) секреции инсулина. Такие новые лекарственные средства должны обладать следующими характеристиками: зависимостью стимуляции секреции инсулина от глюкозы, т.е. индукцией секреции инсулина только в присутствии повышенного содержания глюкозы в крови; низкой частотой первичных и вторичных случаев безуспешного лечения и сохранностью функционирования островковых клеток. Стратегия разработки новых способов лечения, описанных здесь, основывается на сигнальном механизме с участием циклического аденозинмонофосфата (цАМФ) и его влияния на секрецию инсулина.
Циклический АМФ является основным регулятором процесса секреции инсулина. Повышение количества указанной сигнальной молекулы стимулирует закрытие калиевых каналов с последующей активацией механизма протеинкиназы-А. Закрытие калиевых каналов обусловливает деполяризацию клеток и последующее открытие кальциевых каналов, что, в свою очередь, приводит к экзоцитозу инсулиновых гранул. В отсутствие глюкозы и при ее низких концентрациях отмечается слабая секреция инсулина или она вообще отсутствует (A.Weinhaus et al., 1998, Diabetes, 47, 1426-1435). Средства, повышающие секрецию, такие как пептид-активатор гипофизарной аденилатциклазы (РАСАР) и GLP-1, используют систему цАМФ для регуляции секреции инсулина в глюкозо-зависимом режиме (М.Komatsu et al., 1997, Diabetes, 46, 1928-1938). Средства, повышающие секрецию инсулина, механизм действия которых основан на повышении цАМФ, например, GLP-1 и РАСАР, также способны усиливать синтез инсулина в дополнение к секреции инсулина (G.Skoglund et al., 2000, Diabetes, 49, 1156-1164; P.Borboni et al., 1999, Endocrinology, 140, 5530-5537).
РАСАР является мощным стимулятором глюкозо-зависимой секреции инсулина бета-клетками поджелудочной железы. Были описаны три различных типа рецепторов РАСАР (R1, R2 и R3) (A.Harmar et al., 1998, Pharmacol. Reviews, 50, 265-270). РАСАР не проявляет избирательности по типу рецептора, обладая сопоставимыми активностью и сродством ко всем трем типам рецепторов. R1 в основном находится в ЦНС, в то время как R2 и R3 распределены более широко. R2 локализован в ЦНС, а также в печени, легких и кишечнике. R3 находится в ЦНС, поджелудочной железе, скелетной мускулатуре, сердце, почках, жировой ткани, семенниках и желудке. Недавние исследования подтвердили, что R3 ответствен за секрецию инсулина бета-клетками (N.Inagaki et al., 1994, Proc. Natl. Acad. Sci. USA, 91, 2679-2683). Инсулинотропное действие РАСАР опосредовано ГТФ-связывающим белком Gs. Накопление внутриклеточного цАМФ, в свою очередь, активирует неспецифичные катионные каналы в бета-клетках, повышая Са++, и стимулирует экзоцитоз инсулин-содержащих секреторных гранул.
РАСАР является новейшим представителем суперсемейства метаболических, нейроэндокринных и нейротрансмиттерных пептидных гормонов, действие которых опосредуется механизмом передачи сигнала с участием цАМФ (Arimura, 1992, Regul. Peptides, 37, 287-303). Биологически активные пептиды высвобождаются из биосинтетического предшественника в двух молекулярных формах - в виде 38-аминокислотного пептида (РАСАР-38) и/или 27-аминокислотного пептида (РАСАР-27) с амидированными С-концами (Arimura, цит. выше).
Наивысшие концентрации двух форм пептида найдены в головном мозге и семенниках (обзор Arimura, цит. выше). Более короткая форма пептида - РАСАР-27 - проявляет 68% структурного сходства с вазоактивным кишечным полипептидом (VIP). Однако распределение РАСАР и VIP в центральной нервной системе указывает на то, что указанные структурно родственные пептиды имеют различные нейротрансмиттерные функции (Koves et al., 1991, Neuroendocrinology, 54, 159-169).
Недавние исследования показали разнообразие биологических действий РАСАР-38 - от участия в репродуктивных процессах (McArdle, 1994, Endocrinology, 135, 815-817) до способности стимулировать секрецию инсулина (Yada et al., 1994, J.Biol. Chem., 269, 1290-1293).
Вазоактивный кишечный пептид (VIP) является пептидом из 28 аминокислот, который впервые был выделен из верхней части тонкого кишечника хряка (Said & Mutt, 1970, Science, 169, 1217-1218; патент США №3879371). Данный пептид относится к семейству структурно родственных небольших полипептидов, которое включает хелодермин, секретин, соматостатины и глюкагон. Биологические эффекты VIP опосредуются активацией локализованных на клеточных мембранах белков-рецепторов, которые вовлечены в сигнальную систему с участием внутриклеточного цАМФ. Указанные рецепторы исходно были известны как VIP-R1 и VIP-R2, однако впоследствии было установлено, что они являются теми же рецепторами, что и PACAP-R2 и PACAP-R3. VIP проявляет сопоставимые уровни активности и действия в отношении PACAP-R2 и PACAP-R3.
Для повышения стабильности VIP в легочной жидкости человека серия вариантов VIP (Bolin et al., 1995, Biopolymers, 37, 57-65) была сформирована для того, чтобы повысить склонность данного пептида к образованию спирали и снизить его протеолитическое разрушение. Замены проводили по положениям аминокислот 8, 12, 17 и 25-28, для которых была показана важность для связывания на рецепторе. Более того, последовательность "GGT" была присоединена в качестве метки к С-концу мутантных вариантов VIP с надеждой на более эффективное копирование спирали. Наконец, для дальнейшей стабилизации спирали был синтезирован ряд циклических вариантов (патент США №5677419). Хотя описанные усилия не были направлены на достижение рецепторной избирательности, в результате были получены два аналога (обозначаемые здесь как R3P0 и R3P4), которые обладали более чем 100-кратной избирательностью в отношении PACAP-R3 (Gourlet et al., 1997, Peptides, 18, 403-408; Xia et al., 1997, J.Pharmacol. Exp. Ther., 281, 629-633).
GLP-1 секретируется L-клеткой кишечника после принятия пищи и функционирует как гормон инкретин (т.е. он стимулирует зависимый от глюкозы выход инсулина из бета-клетки поджелудочной железы). Он является пептидом, состоящим из 37 аминокислот, который дифференцированно экспрессируется с гена глюкагона, что определяется типом ткани. Для GLP-1 были получены клинические данные, которые подтверждают полезный эффект повышения уровней цАМФ в бета-клетках. Инфузия GLP-1 пациентам с трудноизлечиваемым диабетом 2-го типа нормализовала у них уровни глюкозы в крови натощак (М.Gutniak et al., 1992, New England J.Med., 326, 1316-1322), а более продолжительная инфузия улучшала функцию бета-клеток до уровня, характерного для здоровых субъектов (J.Rachman et al., 1996, Diabetes, 45, 1524-1530). В недавнем сообщении было показано, что GLP-1 улучшает способность бета-клеток реагировать на глюкозу у субъектов с нарушенной переносимостью глюкозы (М.Byrne et al., 1998, Diabetes, 47, 1259-1265). Однако все указанные эффекты оказываются кратковременными из-за короткого времени полужизни пептида. Недавно фирма Novo Nordisk приостановила клинические испытания GLP-1. Как было сообщено, такая неудача обусловливалась очень коротким временем полужизни пептида в плазме, составляющим несколько минут.
EXENDIN-4TM. Фирма Amylin Pharmaceuticals проводит клинические испытания I фазы с препаратом EXENDIN-4 (AC2993) - 39-аминокислотным пептидом, который изначально был идентифицирован у ящерицы-ядозуба. Недавно испытания перешли во II фазу. Amylin сообщил о преклинических результатах, указывающих на 4-часовую эффективность и эффективность в животных моделях при подкожном, пероральном и интраназальном введении AC2993. Однако при дозах 0,2 и 0,3 мкг/мг часто возникали головные боли, постуральная гипотензия, понос и рвота.
Имеется необходимость в усовершенствованном пептиде, который бы обладал глюкозо-зависимой активностью РАСАР, GLP-1 или EXENDIN-4 по усилению секреции инсулина и также проявлял более слабые побочные действия.
Краткое описание изобретения
Настоящее изобретение представляет новые полипептиды, которые функционируют in vivo в качестве агонистов рецептора PACAP-R3 (далее - R3) и эффективны в лечении заболеваний и состояний, которые могут быть улучшены с помощью средств, обладающих активностью агонистов R3. Предпочтительно полипептиды по настоящему изобретению являются селективными агонистами R3, обладающими большей эффективностью в отношении R3 по сравнению с R2 и R1. В качестве примера, но не с целью ограничения, указанные полипептиды стимулируют синтез инсулина и секрецию его бета-клетками поджелудочной железы в глюкозо-зависимом режиме с последующим снижением уровня глюкозы в плазме. Было показано, что указанные полипептиды-усилители секреции инсулина стимулируют секрецию инсулина островковыми клетками крысы и человека in vitro и in vivo. В отличие от РАСАР-27 указанные полипептиды-усилители секреции также снижают уровень глюкозы in vivo в большей степени, чем контрольные наполнители после провокацией глюкозой.
Полипептиды по настоящему изобретению предоставляют новый способ лечения больных, например, с метаболическими расстройствами, такими как те, которые вызваны сниженной секрецией эндогенного инсулина, в частности диабет 2-го типа, или больных с нарушенной переносимостью глюкозы, являющейся преддиабетическим состоянием, и со слабым изменением секреции инсулина.
В частности, в одном аспекте настоящего изобретения представлен полипептид, выбранный из группы, состоящей из SEQ ID NO 11-14, SEQ ID NO 18, SEQ ID NO 21-26, SEQ ID NO 32-36; SEQ ID NO 40-53; SEQ ID NO 57-61; SEQ ID NO 63-99; SEQ ID NO 102-119; SEQ ID NO 121-137; SEQ ID NO 139-177; SEQ ID NO 179, 180, SEQ ID NO 183-202, 322-341, а также их фрагменты, производные и варианты, которые проявляют по крайней мере одну биологическую функцию, которая по существу совпадает с таковой у полипептидов, перечисленных в SEQ ID NO (в целом обозначаются как "полипептиды по настоящему изобретению"), включая их функциональные эквиваленты. Предпочтительным вариантом настоящего изобретения является полипептид, выбранный из группы, состоящей из SEQ ID NO 12, 18, 21-26, 32-35, 41, 43-53, 63, 66, 70-92, 94-99, 102-104, 107, 109, 112-119, 121-137, 139, 140, 142-156, 156-174, 187, 322-341, а также их фрагменты, производные и варианты, которые проявляют по крайней мере одну биологическую функцию, которая по существу одинакова с таковой у полипептидов, перечисленных в SEQ ID NO. Более предпочтительным вариантом настоящего изобретения является полипептид, выбранный из группы, состоящей из SEQ ID NO 18, 24, 25, 32, 33, 43-50, 52, 53, 70-87, 92, 98, 99, 104, 107, 112-114, 129-131, 137, 140, 144, 147-151, 156-159 и 161-173, 323, 324, 326, 327, 335, 338, 341 и их фрагменты, производные и варианты, которые проявляют по крайней мере одну биологическую функцию, которая по существу одинакова с таковой у полипептидов, перечисленных в SEQ ID NO. Наиболее предпочтительным вариантом настоящего изобретения является полипептид, выбранный из группы, состоящей из SEQ ID NO 18, 32, 43, 45, 47, 50, 52, 71, 72, 83, 86 и 87, а также их фрагменты, производные и варианты, которые проявляют по крайней мере одну биологическую функцию, которая по существу одинакова с таковой у полипептидов, перечисленных в SEQ ID NO.
Другим вариантом воплощения настоящего изобретения являются полинуклеотид, который кодирует полипептиды по настоящему изобретению, а также соответствующие векторы и клетки-хозяева, необходимые для рекомбинантной экспрессии полипептидов по настоящему изобретению. Указанные полинуклеотидные последовательности включают те, которые идентифицированы как SEQ ID NO 204, 207-211, 214-230 и 232-321. Предпочтительные полинуклеотиды включают те, которые идентифицированы как SEQ ID NO 204, 207-209, 215, 217-230, 232-234, 237, 239, 242-268, 270-281 и 284-321. Более предпочтительные полинуклеотиды включают те, которые идентифицированы как SEQ ID NO 207, 218-224, 226-230, 234, 237, 242-244, 258-260, 266, 268, 272, 275-279, 284-287, 289-301 и 303, 304, 306, 307, 318 и 321. Наиболее предпочтительные полинуклеотиды включают те, которые идентифицированы как SEQ ID NO 217, 221 и 226.
Антитела и фрагменты антител, которые избирательно связываются с полипептидами, также представлены настоящим изобретением. Такие антитела могут использоваться для выявления полипептидов по настоящему изобретению и могут быть идентифицированы и сформированы с помощью процедур, хорошо известных в данной области техники, включая те способы, которые аналогичны описанному далее в примере 17.
Также изобретение представляет способ лечения диабета и/или других заболеваний или состояний, осуществляемого с использованием полипептидов по настоящему изобретению, предпочтительно на основе функции полипептидов по настоящему изобретению в качестве агонистов R3, у млекопитающего, включающий введение упомянутому млекопитающему терапевтически эффективного количества любого из полипептидов по настоящему изобретению или любого полипептида, активного в отношении R3, например, SEQ IN NO 5 и 9.
Также заявляются способы получения полипептидов по настоящему изобретению, как рекомбинантных, так и синтетических.
Краткое описание чертежей
На фигуре 1 приведены аминокислотные последовательности полипептидов SEQ ID NO 11-14, SEQ ID NO 18, SEQ ID NO 21-26, SEQ ID NO 32-36, SEQ ID NO 40-53, SEQ ID NO 57-61, SEQ ID NO 63-99, SEQ ID NO 102-119, SEQ ID NO 121-137, SEQ ID NO 139-177, SEQ ID NO 179, 180, SEQ ID NO 183-202 и SEQ ID NO 322-341, которые являются заявляемыми полипептидами.
На фигуре 2 показано сопоставление последовательностей мутантных вариантов VIP и нативных полипептидов VIP, PACAP38, GLP-1, EXENDIN-4 и примеров R3-иэбирательных полипептидов. Консервативные остатки выделены жирным шрифтом и затенены темно-серым цветом, в то время как консервативные замены затенены светло-серым цветом.
На фигуре 3 приведена рестрикционная карта типичной плазмиды, кодирующей GST-химерный пептид.
На фигурах 4А-4В показаны графики, отражающие влияние GLP-1 или R3P3 на секрецию инсулина островковыми клетками крысы in vitro.
На фигуре 5 приведен график, показывающий влияние пептида R3P3 на выведение глюкозы.
На фигуре 6 приведена диаграмма, показывающая влияние РАСАР и родственных полипептидов на содержание воды в кишечнике у мышей Balb/C.
На фигуре 7 показана диаграмма, указывающая на то, что доза 1 нмоль/кг R3P3, R3P12, R3P13 или GLP-1 усиливает выведение глюкозы у крысы после подкожного введения.
На фигуре 8 приведены полинуклеотидные последовательности SEQ ID NO 54-56 и 203-301, которые кодируют полипептиды по настоящему изобретению.
Первые 6 нуклеотидов представляют собой сайт распознавания рестриктазой BamHI, после которых расположены 12 нуклеотидов, кодирующих сайт распознавания "IEGR" фактора Ха. Последние 6 нуклеотидов представляют собой сайт распознавания рестриктазами Xhol или EcoRI, a 6 нуклеотидов перед ними кодируют два стоп-кодона. Нуклеотиды между сайтом фактора Ха и стоп-кодонами кодируют аминокислотную последовательность соответствующего полипептида. Нуклеотиды между двумя рестрикционными сайтами клонируют по соответствующим рестрикционным сайтам в векторе pGEX-6P-1 (Amersham Pharmacia Biotech). Номера SEQ ID NO приведены в скобках.
На фигуре 9 отображено влияние РАСАР-27, VIP и избирательных агонистов рецептора на частоту сердечных сокращений у собак, находящихся в сознании (см. пример 15).
На фигуре 10 показано выявление R3P66 методом ТИФА с помощью поликлональных антител, выработанных в кроликах, иммунизованных С-концевой последовательностью R3P66 (Ac-CRKQVAAKKYLQSIKNKRY-COOH).
Подробное описание предпочтительных вариантов
Настоящее изобретение представляет новые полипептиды и их фрагменты, производные и варианты, проявляющие по крайней мере одну биологическую функцию, которая по существу одинакова с таковой у полипептидов фигуры 1 (вместе "полипептиды по настоящему изобретению"). Полипептиды по настоящему изобретению функционируют in vivo в качестве агонистов R3 или, иными словами, используются для профилактики и/или лечения таких заболеваний или состояний, как диабеты, астма, артериальная гипертония, проблемы в репродуктивной сфере у мужчин, включая подвижность спермы у мужчин, сердечно-сосудистые заболевания, язвы и другие состояния, идентифицированные здесь, или функционируют иным путем в соответствии с описанным здесь ниже. Предпочтительно полипептиды по настоящему изобретению должны стимулировать секрецию инсулина бета-клетками поджелудочной железы в глюкозо-зависимом режиме.
Полипептиды по настоящему изобретению являются агонистами R3. Предпочтительно они являются избирательными - агонистами R3 по крайней мере с 10-кратной избирательностью в отношении R3 по сравнению с R2 и/или R1. Более предпочтительно они являются избирательными агонистами R3 по крайней мере с 100-кратной избирательностью в отношении R3 по сравнению с R2 и/или R1. Наиболее предпочтительно они стимулируют секрецию инсулина в плазму в глюкозо-зависимом режиме без индукции застойных явлений или повышения уровня глюкозы в плазме, что является противопоказаниями при лечении, например, диабета 2-го типа. Кроме того, предпочтительно, чтобы полипептиды по настоящему изобретению являлись избирательными агонистами рецептора R3, что тем самым обусловливает, например, повышение выхода инсулина в плазму, но не являлись избирательными в отношении других рецепторов, которые ответственны за такие неприятные или опасные побочные действия, как удерживание воды в желудочно-кишечном тракте, и/или за нежелательные сердечно-сосудистые проявления, такие как повышенная частота сердечных сокращений. В настоящее время установлено, что R3-опосредованная секреция инсулина не вызывает гипогликемию, а активация R2 обусловливает выход глюкозы в плазму, что противопоказано при лечении диабета 2-го типа, и удерживание воды в желудочно-кишечном тракте, и активация R1 приводит к сердечно-сосудистым проявлениям, таким как повышенная частота сердечных сокращений.
Полипептиды по настоящему изобретению представляют новый способ лечения больных со сниженной секрецией эндогенного инсулина или нарушенной переносимостью глюкозы, в частности, при диабете 2-го типа.
А. Обсуждение
РАСАР, VIP, GLP-1 и Exendin-4 являются полипептидами, способными стимулировать секрецию инсулина в глюкозо-зависимом режиме. Однако сам по себе этот факт не гарантирует снижения уровня глюкозы in vivo. Поскольку известно, что РАСАР связывается с рецепторами PACAP-R1, -R2 и -R3, а VIP, как известно, связывается с рецепторами PACAP-R2 и -R3, считалось, что они могут обладать сходными консервативными структурными свойствами. Ниже множественное сопоставление показывает соотношение первичных структур:

(где однобуквенные обозначения аминокислот можно найти у Zubay, 1988, "Biochemistry", 2d ed., MacMillan Publ., New York, p.33, и расшифрованы далее). Полипептиды по настоящему изобретению (фиг.1) наиболее близки к VIP по параметрам их первичной структуры за исключением SEQ ID NO 57-61, 66-69 и 176, 177, 179, 180, 183-202, которые наиболее близки к РАСАР.
Авторы настоящего изобретения создали новый полипептид, являющийся агонистом рецептора R3, предпочтительно избирательным агонистом R3, и/или проявляющий избирательное глюкозо-зависимое действие по усилению секрецию инсулина, причем избирательная активация рецептора PACAP-R3 действительно приводит к глюкозо-зависимому механизму секреции инсулина бета-клетками поджелудочной железы с соответствующим снижением уровня глюкозы in vivo. В свете изложенного авторы изобретения вначале исследовали структуры РАСАР-27 и VIP с целью определения остатков, которые наиболее вероятно ответственны за рецепторную избирательность. Известно, что РАСАР и VIP не снижают уровень глюкозы in vivo, но, напротив, стимулируют высвобождение глюкозы из печени. Было показано, что активация R2 повышает уровни глюкозы в плазме in vivo. Ранее были получены разнообразные мутантные варианты и РАСАР, и VIP, что имело различные цели. Например, серийные делеции РАСАР-27 и РАСАР-38 с обоих концов подтвердили важность обоих концевых участков для связывания на рецепторе (Gourlet et al., 1995, Eur. J.Pharm., 287, 7-11; Gourlet et al., 1996, Regul. Peptides, 62, 125-130). Связывание на мембранах головного мозга крысы и аденилатциклазная активность химерных мутантных вариантов PACAP-27/VIP показали важность N-концевых остатков РАСАР для распознавания PACAP-R1 (Ando et al., 1996, Biomed. Pept. Proteins Nucleic Acids, 2, 41-46). Повышение основности Leul7-PACAP27 и Leu17-VIP за счет мутаций K15R, K20R и K21R и достройки С-конца последовательностью GKR привело к повышению длительности релаксантной активности в трахее морской свинки, что предположительно определяется защитой от связывания с гепарином (Kashimoto et al., 1996, Ann. NY Acad. Sci., 805, 505-510). Было показано (Gourlet et al., 1996, Biochim. Biophys. Acta, 1314, 267-273), что Q16R-мутантный вариант VIP и РАСАР проявляет более высокую аффинность по сравнению с соответствующими нативными полипептидами в отношении соответственно PACAP-R2 и R1. Высокоаффинный избирательный по R2 агонист был сформирован (Gourlet et al., 1997, Peptides, 18, 1539-1545) путем получения химерного замещенного пептида [К15,R16,L27]VIP(1-7)/GRF(8-27). Ацилирование по N-концу и замена на D-Phe2 у такого избирательного агониста привела к образованию мощного избирательного по R2 антагониста (Gourlet et al., 1997, Peptide, 18, 1555-1560). Мутантные варианты Y22L и Y22A VIP, но не Y22F, проявляют меньшую аффинность в отношении PACAP-R3, предполагая важность наличия ароматической группы в положении 22 для связывания на рецепторе R3, но не для связывания на рецепторе R2 (Gourlet, 1998, Eur. J.Biochem., 348, 95-99). Хелодермин и хелоспектин - VIP-подобные пептиды, выделенные из яда слюнных желез ящериц, - проявляют примерно 100-кратную избирательность в отношении рецептора PACAP-R3 (Gourlet, 1998, Ann. NY Acad. Sci., 865, 247-252). Фотоаффинное мечение РАСАР-27 путем замены F6 и Y22 на п-бензоил-L-фенилаланин (pBz) или К15, К20 и К21 на pBz2 предположило, что К15 и F22 ближе к PACAP-R1, чем F6, К20 и К21 (Сао et al., 1997, Eur. J.Biochem., 244, 400-406; Сао et al., 1998, Ann. NY Acad. Sci., 865, 82-91).
Авторы настоящего изобретения обнаружили ряд полипептидов, которые обусловливают стимуляцию секреции инсулина в глюкозо-зависимом режиме и приводят к снижению уровня глюкозы in vivo. Указанные полипептиды обладают некоторым сходством с VIP и РАСАР. В частности, множественное сопоставление показало следующее:

Однако в научной или патентной литературе нет информации, которая бы указывала на то, что выбранные модификации последовательностей VIP и РАСАР приводят к получению полипептида, обладающего способностью стимулировать секрецию инсулина в глюкозо-зависимом режиме и снижать концентрацию глюкозы в плазме.
Некоторые термины, используемые в данной заявке, будут определены теперь, а другие термины - по мере их введения. Однобуквенные обозначения конкретных аминокислот, соответствующие им аминокислоты и трехбуквенные аббревиатуры таковы: А - аланин (Ala); С - цистеин (Cys); D - аспарагиновая кислота (Asp); Е - глутаминовая кислота (Glu); F - фенилаланин (Phe); G - глицин (Gly); H - гистидин (His); I - изолейцин (Ile); К - лизин (Lys); L - лейцин (Leu); М - метионин (Met); N - аспарагин (Asn); P - пролин (Pro); Q - глутамин (Gln); R - аргинин (Arg); S - серин (Ser); Т - треонин (Thr); V - валин (Val); W - триптофан (Trp); Y - тирозин (Tyr).
Термин "полинуклеотид, кодирующий полипептид" охватывает полинуклеотид, который включает только кодирующую полипептид последовательность, а также полинуклеотид, который включает дополнительную кодирующую и/или некодирующую последовательность. Также настоящее изобретение касается полинуклеотидов, которые гибридизуют с описанными здесь выше последовательностями, если они характеризуются по крайней мере примерно 70%-ной, предпочтительно по крайней мере примерно 90%-ной и более предпочтительно по крайней мере примерно 95%-ной идентичностью последовательностей. Настоящее изобретение, в частности, касается, полинуклеотидов, кодирующих полипептиды, которые гибридизуют в жестких условиях с описанными здесь выше полинуклеотидами. По использованию в данном тексте термин "жесткие условия" обозначает "жесткие условия гибридизации". Предпочтительно гибридизация должна происходить только тогда, когда имеется по крайней мере примерно 90%-ная и предпочтительно примерно 95-97%-ная идентичность последовательностей. Полинуклеотиды, которые гибридиэуют с описанными здесь выше полинуклеотидами в предпочтительном варианте, кодируют полипептиды, которые сохраняют по существу такую же биологическую функцию или активность, что и зрелый полипептид, кодируемый молекулами кДНК.
Каждый из терминов "функциональный эквивалент" и "по существу одинаковая биологическая функция или активность" обозначает то, что уровень биологической активности составляет 30-100% или больше от той биологической активности, которую демонстрирует полипептид, с которым проводится сравнение в случае, когда биологическую активность каждого из полипептидов определяют в одной и той же процедуре. Например, полипептидом, который функционально эквивалентен полипептиду фигуры 1, будет тот полипептид, который в случае радиоиммунологического теста на сцинтилляционное приближение (RIA-SPA) цАМФ, описанного в частном примере 16, проявляет накопление цАМФ в линии клеток СНО, экспрессирующих рецептор PACAP/VIP-R2 (PACAP-R3) человека.
Полипептидом по настоящему изобретению, являющимся агонистом R3, является полипептид, который проявляет примерно 30-100% или больше максимальной активности агониста в отношении PACAP27-R3 при тестировании с помощью протокола примера 16. Предпочтительные полипептиды по настоящему изобретению, являющиеся избирательными агонистами R3 по сравнению с рецепторами PACAP-R2 и R1, - это такие полипептиды, которые проявляют отношение активности агониста R3 к активности в отношении R2 примерно 10:1 или выше, а более предпочтительно - примерно 100:1 или выше, и/или проявляют отношение активности агониста R3 по сравнению с активностью в отношении рецептора R1 примерно 10:1 или выше, а более предпочтительно - примерно 100:1 или выше тогда, когда полипептид тестируют в соответствии с протоколом примера 16 с использованием клеток, экспрессирующих соответствующие рецепторы.
"Жесткие условия гибридизации" указывают на инкубацию в течение ночи двух фрагментов полинуклеотидов, чтобы они гибридизовали при 42°С в растворе, содержащем 50% формамида, 5×SSC (750 мМ NaCl, 75 мМ цитрата натрия), 50 мМ фосфата натрия (рН 7,6), 5×раствор Денхардта, 10% декстрансульфата и 20 мкг/мл денатурированной, подвергнутой встряхиванию ДНК спермы лосося с последующей промывкой на фильтрах в 0,1×SSC при примерно 65°С.
Термины "фрагмент", "производное" и "вариант" при употреблении их в отношении полипептидов с фигуры 1 обозначают фрагменты, производные и варианты полипептидов, которые сохраняют по существу такую же биологическую функцию или активность таких полипептидов соответствии с тем, что дополнительно описано далее.
Аналог включает прополипептид, в составе которого имеется аминокислотная последовательность полипептида по настоящему изобретению. Активный полипептид по настоящему изобретению может быть отщеплен от дополнительных аминокислот, которые составляют полную полипептидную молекулу, с участием естественных процессов in vivo или с помощью процедур, хорошо известных в данной области техники, например, каталитического или химического расщепления. Например, 28-аминокислотный нативный пептид VIP в естественных условиях экспрессируется в виде существенно более длинного полипептида, который затем процессируется in vivo с высвобождением 28-аминокислотного активного зрелого пептида.
Фрагмент является частью полипептида, который по существу сохраняет такую же функциональную активность, которую выявляют в моделях in vivo, описанных далее в данном тексте.
Производное охватывает все модификации полипептида, которые по существу сохраняют функции, описанные здесь, и включает дополнительную структуру и сопутствующую ей функцию, например, ПЭГилированные полипептиды, которые характеризуются более высоким временем полужизни, химерные полипептиды, которые обладают свойствами специфичности в отношении какой-либо мишени, или дополнительной активностью, такой как токсичность в отношении конкретной мишени в соответствии с описанным здесь далее.
Полипептиды по настоящему изобретению могут быть рекомбинантными полипептидами, природными очищенными полипептидами или синтетическими полипептидами.
Фрагмент, производное или вариант полипептидов по настоящему изобретению может являться (i) таковым, в котором один или большее число аминокислотных остатков заменены на консервативные или неконсервативные аминокислотные остатки (предпочтительно на консервативный аминокислотный остаток), и такой замененный аминокислотный остаток может кодироваться или не кодироваться генетическим кодом; или (ii) таковым, в котором один или больше число аминокислотных остатков включают замещающую группу; или (iii) таковым, в котором зрелый полипептид химеризован с другим соединением так, что соединение увеличивает время полужизни полипептида (например, полиэтиленгликоль); или (iv) таковым, в котором дополнительные аминокислоты химеризованы со зрелым полипептидом так, что образуют лидерную или секреторную последовательность или последовательность, которая используется для очистки зрелого полипептида, или прополипептидную последовательность; или (v) таковым, в котором полипептидная последовательность химеризована с более крупным полипептидом, например, человеческий альбумин, антитело или фрагмент Fc, с целью продления действия. Такие фрагменты, производные или варианты и аналоги рассматриваются как попадающие в объем понятного специалистам в данной области техники исходя из изложенного в данном тексте.
Предпочтительно производные по настоящему изобретению должны включать консервативные аминокислотные замены (определены здесь далее), произведенные по одному или нескольким предсказанным, предпочтительно неключевым аминокислотным положениям. "Неключевой" аминокислотный остаток обозначает остаток, который может быть изменен в последовательности белка дикого типа без изменения биологической активности, в то время как "ключевой" аминокислотный остаток для биологической активности необходим. "Консервативной аминокислотной заменой" является замена, при которой аминокислотный остаток заменяется на аминокислотный остаток, имеющий сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, определены в данной области техники. Такие семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), β-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Неконсервативные замены не должны производиться по консервативным аминокислотным остаткам или по аминокислотным остаткам в пределах консервативного белкового домена, таким как остатки 19 и 27, поскольку указанные остатки являются ключевыми для активности белка, такой как активность в отношении R3 и/или избирательность в отношении R3. Фрагменты или биологически активные части охватывают полипептидные фрагменты, пригодные для использования в качестве лекарственного средства, для получения антител, в качестве исследовательского реагента и подобного. Фрагменты включают пептиды, имеющие аминокислотные последовательности, по существу сходные с или производные от аминокислотных последовательностей полипептида по настоящему изобретению и проявляющие по крайней мере одну активность такого полипептида, но которые включают меньшее число аминокислот по сравнению с описанными в данном тексте полнораэмерными полипептидами. Обычно биологически активная часть включает домен или мотив, имеющий по крайней мере одну активность полипептида. Биологически активная часть полипептида может являться пептидом, который, например, состоит из 5 или большего числа аминокислот. Такие биологически активные части могут быть получены синтетически или с помощью рекомбинантных методов и могут быть протестированы по одной или нескольким функциональным активностям полипептида по настоящему изобретению с помощью способов, описанных в данной заявке и/или известных в данной области техники.
Более того, предпочтительные производные по настоящему изобретению охватывают зрелые полипептиды, которые были химеризованы с другим соединением таким образом, что соединение увеличивает время полужизни полипептида и/или снижает потенциальный уровень иммуногенности полипептида (например, полиэтиленгликоль - "ПЭГ"). В случае "ПЭГилирования" химеризация полипептида на ПЭГ может быть осуществлена с помощью любого способа, известного специалистам в данной области техники. Например, ПЭГилирование можно осуществить сначала путем внесения мутантного цистеина в полипептид и затем путем сайт-специфичной дериватизации с ПЭГ-малеимидом. Цистеин может быть добавлен к С-концу пептидов (см., например, Tsutsumi et al., 2000, Proc. Natl. Acad. Sci. USA, 97(15), 8548-8553).
Варианты полипептида по настоящему изобретению охватывают полипептиды, характеризующиеся аминокислотной последовательностью, по существу сходной с аминокислотной последовательностью номеров SEQ ID NO с фигуры 1 или ее домена. Термин "по существу сходный" обозначает первую аминокислотную последовательность, которая включает достаточное или минимальное число идентичных или эквивалентных аминокислотных остатков по сравнению со второй аминокислотной последовательностью таким образом, что первая и вторая аминокислотные последовательности имеют общий структурный домен и/или общую функциональную активность. Например, аминокислотные последовательности, включающие общий структурный домен, который идентичен по крайней мере на 45%, предпочтительно примерно на 75-98%, определяются здесь как по существу сходные. Предпочтительно варианты должны быть по существу сходными с аминокислотной последовательностью предпочтительных полипептидов по настоящему изобретению. Варианты включают варианты полипептидов, кодируемые полинуклеотидом, который гибридизует с полинуклеотидом по настоящему изобретению или его комплементом при жестких условиях. Такие варианты в целом сохраняют функциональную активность полипептидов по настоящему изобретению. Библиотеки фрагментов полинуклеотидов могут быть использованы для создания гетерогенной выборки фрагментов для скрининга и последующего отбора. Например, библиотека фрагментов может быть сформирована путем обработки двухцепочечного ПЦР-фрагмента полинуклеотида нуклеазой в условиях, при которых разрыв происходит в одной молекуле только один раз, с последующими денатурацией двухцепочечной ДНК, ренатурацией ДНК с образованием двухцепочечной ДНК, которая может включать смысловые/антисмысловые пары различных разорванных продуктов, удалением одноцепочечных фрагментов от вновь образованных дуплексов путем обработки нуклеазой S1 и лигированием полученной в результате библиотеки фрагментов в состав экспрессирующего вектора. С помощью такого способа можно сформировать экспрессионную библиотеку, которая кодирует N-концевые и внутренние фрагменты различного размера из состава полипептида по настоящему изобретению.
Варианты охватывают полипептиды, которые отличаются по аминокислотной последовательности вследствие мутагенеза. Варианты, функционирующие в качестве агонистов R3, могут быть идентифицированы путем скрининга на активность агониста R3 комплексных библиотек мутеинов, например, укороченных мутантных вариантов полипептидов по настоящему изобретению.
В одном варианте гетерогенную библиотеку аналогов формируют путем комбинаторного мутагенеза на уровне нуклеиновой кислоты и она кодируется гетерогенной генной библиотекой. Гетерогенная библиотека вариантов может быть получена, например, каталитически путем лигирования смеси синтетических олигонуклеотидов в генные последовательности так, что вырожденный набор возможных вариантов аминокислотных последовательностей экспрессируется в виде отдельных полипептидов или, как альтернатива, в виде группы более крупных химерных белков (например, для представления на фаговой поверхности), включающих в себя набор последовательностей. Имеется ряд методов, которые можно использовать для формирования библиотек потенциальных вариантов на материале вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной генной последовательности можно осуществить на автоматическом ДНК-синтезаторе с последующим лигированием синтетического гена в состав подходящего экспрессирующего вектора. Использование вырожденного набора генов обеспечивает наличие в одной смеси всех последовательностей, кодирующих желательный набор потенциальных вариантных последовательностей. Способы синтеза вырожденных олигонуклеотидов известны в данной области техники (см., например, Narang, 1983, Tetrahedron, 39, 3; Itakura et al., 1984, Annu. Rev. Biochem., 53, 323; Itakura et al., 1984, Science, 198, 1056; Ike et al., 1983, Nucl. Acid Res., 11, 477).
В данной области техники известен ряд методов скрининга генных продуктов из состава комбинаторных библиотек, сформированных с помощью точковых мутаций или укорочения, и скрининга библиотек кДНК на генные продукты, обладающие выбранным свойством. Такие методы могут быть приспособлены для оперативного скрининга генных библиотек, сформированных с помощью комбинаторного мутагенеза полипептидов-агонистов R. Наиболее широко применяемые методы, которые пригодны для высокопроизводительного скрининга больших генных библиотек, обычно включают клонирование генной библиотеки в компетентные по репликации экспрессирующие векторы, трансформацию подходящих клеток полученными библиотечными векторами и экспрессию перекомбинированных генов в условиях, при которых выявление желательной активности обеспечивает выделение вектора, кодирующего ген, продукт которого был выявлен. Рекурсивный комплексный мутагенез (REM) математический метод, который повышает частоту функциональных мутаций в библиотеках, может быть использован в сочетании с тест-скринингом с целью идентификации желательных вариантов.
Настоящее изобретение также предусматривает химерные или гибридные полипептиды. Примерами являются те полипептиды по настоящему изобретению, которые описаны в SEQ ID NO 18 и 172, являющиеся химерами последовательности-мишени поджелудочной железы "SWCEPGWCR" (D.Rajotte et al., 1998, J.Clin. Invest., 102, 430-437) с SEQ ID NO 8 и 32 соответственно. Последовательность-мишень сформирована так, чтобы локализовать доставку полипептида в поджелудочную железу и свести к минимуму возможные побочные эффекты. Полипептиды по настоящему изобретению могут быть составлены аминокислотами, соединенными друг с другом пептидными связями или модифицированными пептидными связями, например, являться изостерными пептидами, и могут включать иные аминокислоты, нежели те 20 аминокислот, которые генетически детерминированы. Полипептиды могут быть модифицированы либо в результате естественных процессов, таких как посттрансляционный процессинг, либо с помощью методов химической модификации, хорошо известных в данной области техники. Такие модификации подробно описаны в учебниках и специальных монографиях, а также в обширной научной литературе. Модификации могут затрагивать любую часть полипептида, включая пептидный скелет, боковые цепи аминокислот и N- или С-концевые участки. Должно быть понятно, что одинаковый тип модификаций может присутствовать в одинаковой или разной степени в разных сайтах данного полипептида. Также данный полипептид может включать много типов модификаций. Полипептиды могут быть разветвленными, например, в результате убихитинизации, и они могут быть циклическими с разветвлениями или без них. Циклические, разветвленные и разветвленные циклические полипептиды могут образовываться в результате естественных посттрансляционных процессов или могут быть получены методами синтеза. Модификации включают ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гема, ковалентное присоединение нуклеотида или нуклеотидного производного, ковалентное присоединение липида или липидного производного, ковалентное присоединение фосфотидилинозитола, перекрестное сшивание, циклизацию, образование дисульфидных связей, деметилирование, образование ковалентных перекрестных сшивок, образование цистеина, образование пироглутамата, формилирование, γ-карбоксилирование, гликозилирование, образование GPI-якоря, гидроксилирование, иодирование, метилирование, миристоилирование, оксидирование, ПЭГидирование, протеолитическое расщепление, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, опосредованное транспортными РНК добавление аминокислот к белкам, например, аргинилирование, и убихитинизацию (см., например, "Proteins, Structure and Molecular Properties", 2d ed., Т.Е.Creighton, W.H.Freeman and Co., New York, 1993; "Posttranslational Covalent Modification of Proteins", B.C.Johnson ed., Academic Press, New York, 1983, pp. 1-12; Seifter et al., 1990, Meth. Enzymol., 182, 626-646; Rattan et al., 1992, Ann. NY Acad. Sci., 663, 48-62).
Полипептиды по настоящему изобретению охватывают полипептиды фигуры 1, соответствующие SEQ ID NO 11-14, SEQ ID NO 18, SEQ ID NO 21-26, SEQ ID NO 32-36, SEQ ID NO 40-53, SEQ ID NO 57-61, SEQ ID NO 63-99, SEQ ID NO 102-119, SEQ ID NO 121-137, SEQ ID NO 139-177, SEQ ID NO 179, 180, SEQ ID NO 183-202, 322-341, а также последовательности, характеризующиеся "несущественной" изменчивостью их последовательностей. "Несущественная изменчивость" должна охватывать любой вариант последовательности, замену или делецию, которые по существу поддерживают по крайней мере одну биологическую функцию полипептидов по настоящему изобретению, предпочтительно активность агониста R3, более предпочтительно активность избирательного агониста R3 и наиболее предпочтительно демонстрируемую здесь активность по секреции инсулина. Такие функциональные эквиваленты предпочтительно могут охватывать полипептиды, которые характеризуются по крайней мере примерно 90%-ной идентичностью полипептидам фигуры 1, а более предпочтительно по крайней мере 95%-ной идентичностью полипептидам фигуры 1 и еще более предпочтительно по крайней мере 97%-ной идентичностью полипептидам фигуры 1, а также включают части таких полипептидов, обладающие по существу одинаковой биологической активностью. Однако любой полипептид, обладающий несущественной изменчивостью аминокислотной последовательности полипептидов фигуры 1, который проявляет функциональную эквивалентность в соответствии с описанным далее в данном тексте, включается в описание настоящего изобретения.
Как известно в данной области техники, "сходство" между двумя полипептидами определяют путем сравнения аминокислотной последовательности и ее консервативных аминокислотных замен одного полипептида с последовательностью второго полипептида. Такие консервативные замены включают то, что описано выше и у Dayhoff, 1978, "The Atlas of Protein Sequence and Structure", 5, и Argos, 1989, EMBO J., 8, 779-785. Например, консервативные замены представляют аминокислоты, относящиеся к одной из следующих групп:
- Ala, Pro, Gly, Gln, Asn, Ser, Thr;
- Cys, Ser, Tyr, Thr;
- Val, Ile, Leu, Met, Ala, Phe;
- Lys, Arg, His;
- Phe, Tyr, Trp, His и
- Asp, Glu.
Также настоящее изобретение касается векторов, которые включают полинуклеотиды по настоящему изобретению, клеток-хозяев, которые были генетически преобразованы векторами по настоящему изобретению, и получения полипептидов по настоящему изобретению рекомбинантными методами. Клетки-хозяева могут быть генетически преобразованы (трансдуцированы или трансформированы, или трансфицированы) с помощью векторов по настоящему изобретению, которые могут быть, например, клонирующим вектором или экспрессирующим вектором. Вектор может иметь форму, например, плазмиды, вирусной частицы, фага и т.п. Преобразованные клетки-хозяева можно прокультивировать в стандартных питательных средах, модифицированных в соответствии с активирующими промоторами или способом отбора трансформантов. Условия культивирования, такие как температура, рН и подобное, соответствуют тем, которые ранее использовались для клеток-хозяев, отобранных для экспрессии, и будут ясны специалистам в данной области техники. Полинуклеотид по настоящему изобретению может быть использован для получения полипептида с помощью рекомбинантных методов. Таким образом, например, полинуклеотидная последовательность может быть включена в состав любой из различных экспрессионных систем, в частности, векторов или плазмид для экспрессии полипептида. Такие векторы включают хромосомные, нехромосомные и синтетические последовательности ДНК, например, производные SV40; бактериальные плазмиды; фаговую ДНК; дрожжевые плазмиды; векторы, производные от сочетания плазмид и фаговой ДНК, вирусную ДНК, например, вирусов коровьей оспы, аденовируса, поксвируса птиц и вируса ложного бешенства. Однако любой другой вектор или плазмида могут быть использованы, лишь бы они были способными к репликации и жизнеспособными в клетке-хозяине.
Подходящая последовательность ДНК может быть встроена в состав вектора с помощью различных процедур. В целом, последовательность ДНК встраивают по подходящему рестрикционному сайту с помощью методов, известных в данной области техники. Такие методы и другое рассматриваются как известные специалистам в данной области техники. Последовательность ДНК в экспрессирующем векторе функционально присоединена к подходящим последовательностям-регуляторам экспрессии (промотор), обеспечивающим синтез мРНК. В качестве характерных примеров таких промоторов можно упомянуть: промотор LTR или SV40, промотор lac или trp E.coli, промотор PL λ-фага и другие промоторы, для которых известна регуляция экспрессии генов в прокариотических или эукариотических клетках или их вирусах. Также экспрессирующий вектор включает инициирующий трансляцию сайт связывания на рибосомах и терминатор транскрипции. Также вектор может включать последовательности, подходящие для амплифицирующей экспрессии. Кроме того, экспрессирующие векторы предпочтительно включают ген, детерминирующий фенотипический признак для отбора трансформированных клеток-хозяев, такой как дегидрофолатредуктаза, или резистентность к неомицину для культур эукариотических клеток, или такой как резистентность к тетрациклину или ампициллину у E.coli. Вектор, включающий подходящую последовательность ДНК, как это было описано здесь выше, равно как подходящий промотор или регуляторную последовательность, может быть использован для трансформации подходящего хозяина, чтобы хозяин мог экспрессировать белок. В качестве характерных примеров подходящих хозяев можно упомянуть: бактериальные клетки, такие как E.coli, Salmonella typhimurium, Streptomyces; грибковые клетки, такие как дрожжи; клетки насекомых, такие как Drosophila S2 и Spodoptera Sf9; клетки животных, такие как СНО, COS или меланома Бойеса; аденовирусы; растительные клетки и т.п. Выбор подходящего хозяина, как можно считать, будет ясен специалисту в данной области техники с учетом изложенного в данном тексте.
Настоящее изобретение также охватывает рекомбинантные конструкции, включающие одну или большее число последовательностей, в широком смысле описанных в данном тексте. Конструкции включают вектор, такой как плазмида или вирусный вектор, в состав которого последовательность по настоящему изобретению была встроена в прямой или обратной ориентации. В предпочтительном аспекте данного варианта воплощения конструкция дополнительно включает регуляторные последовательности, включая, например, промотор, функционально присоединенный к последовательности. Большое число подходящих векторов и промоторов известно специалистам в данной области техники и имеется в продаже. Следующие векторы приведены в качестве примера. Бактериальные: pQE70, pQE60, pQE-9 (Qiagen), pBS, phagescript, psiX174, pBluescript SK, pBsKS, pNHSa, pNH16a, pNH18a, pNH46a (Stratagene); pTRC99A, pKK223-3, pKK233-3, pDR540, PRIT5 (Pharmacia). Эукариотические: pWLneo, pSV2cat, pOG44, pXTl, pSG (Stratagene), pSVK3, pBPV, pMSG, PSVL (Pharmacia). Однако может быть использована любая другая плазмида или вектор, лишь бы они были способными реплицироваться и выживать в хозяине. Промоторные участки могут быть выбраны из состава любого желательного гена с использованием векторов CAT (хлорамфениколтрансфераза) или других векторов, имеющих селективные маркеры. Двумя подходящими векторами являются рКК232-8 и рСМ7. Конкретно упоминаемыми бактериальными промоторами являются lacI, lacZ, Т3, Т7, gpt, PR, λ-фага и trp. Эукариотические промоторы включают предранний промотор CMV, промотор тимидинкиназы вируса простого герпеса, ранний и поздний промоторы SV40, LTR ретровирусов и промотор металлотионеина-1 мыши. Выбор подходящих вектора и промотора хорошо понятен специалистам в данной области техники.
Также настоящее изобретение касается клеток-хозяев, несущих описанные выше конструкции. Клетка-хозяин может являться клеткой высших эукариот, такой как клетка млекопитающего, или клеткой низших эукариот, таких как дрожжевая клетка, или же клетка-хозяин может являться прокариотической клеткой, такой как бактериальная клетка. Внесение конструкции внутрь клетки-хозяина можно осуществить путем трансфекции с фосфатом кальция, трансфекции с помощью DEAE-декстрана или электропорации (L.Davis, M.Dibner, I.Battey, 1986, "Basic Methods in Molecular Biology"). Конструкции в клетках-хозяевах могут быть использованы стандартным образом для выработки генного продукта, кодируемого рекомбинантной последовательностью. Как альтернатива, полипептиды по настоящему изобретению могут быть получены синтетически с использованием стандартных пептидных синтезаторов.
Зрелые белки могут быть экспрессированы в клетках млекопитающих, дрожжей, бактерий или иных клетках под контролем соответствующих промоторов. Бесклеточные трансляционные системы также можно использовать для выработки таких белков с использованием РНК, производных от ДНК-конструкций по настоящему изобретению. Подходящие клонирующие и экспрессирующие векторы для использования с прокариотическими и эукариотическими организмами-хозяевами описаны у Sambrook et al., 1989, "Molecular Cloning: A Laboratory Manual", 2d edition. Cold Spring Harbor, NY: содержание руководства включено здесь для сведения в виде библиографической ссылки.
Транскрипция ДНК, кодирующей полипептиды по настоящему изобретению, у высших эукариот усиливается в результате встраивания в вектор энхансерной последовательности. Энхансеры - это цис-действующие элементы ДНК длиной обычно примерно 10-300 пар нуклеотидов (п.н.), которые действуют на промотор, усиливая транскрипцию с него. Примерами являются энхансер вируса SV40 в области позднего гена со стороны сайта начала репликации (нуклеотиды 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомавируса со стороны позднего гена от сайта начала репликации и аденовирусные энхансеры. В целом, рекомбинантные эрепрессирующие векторы должны включать сайты начала репликации и селективные маркеры при трансформации клетки-хозяина, например, ген резистентности к ампициллину E.coli и ген TRP1 S.cerevisiae, а также промотор, производный от интенсивно экспрессируемого гена, с целью обеспечения транскрипции нижерасположенной структурной последовательности. Такие промоторы могут происходить из оперонов, кодирующих, помимо прочего, ферменты гликолиза, такие как 3-фосфоглицераткиназа (PGK), α-фактор, кислую фосфатазу или белки теплового шока. Гетерологичную структурную последовательность собирают в соответствующей фазе с последовательностями трансляции, инициации и терминации, включая предпочтительно лидерную последовательность, способную обеспечивать секрецию транслированного белка в периплазматическое пространство или внеклеточную среду. Необязательно гетерологичная последовательность может кодировать химерный белок, включающий N-концевой идентификационный пептид, придающий желательные свойства, например, стабилизацию или упрощение очистки экспрессированного рекомбинантного продукта.
Применимые для использования в бактериях экспрессирующие векторы конструируют путем встраивания структурной последовательности ДНК, кодирующей желательный белок, вместе с подходящими трансляционными, инициирующими и терминирующими сигналами в функциональной фазе считывания с активным промотором. Вектор должен включать один или несколько фенотипических селективных маркеров и сайт начала репликации, чтобы обеспечить поддержание вектора и, если это желательно, обеспечить амплификацию в организме хозяина. Подходящими прокариотическими хозяевами для трансформации являются E.coli, Bacillus subtilis. Salmonella typhimurium и различные виды, входящие в роды Pseudomonas, Streptomyces и Staphylococcus, хотя также могут быть выбраны и другие хозяева. Применимые экспрессирующие векторы для использования в бактериях могут включать селективный маркер и бактериальный сайт начала репликации, происходящие из состава имеющихся в продаже плазмид, включающие генетические элементы хорошо известного клонирующего вектора pBR322 (ATCC 37017). Такими коммерчески доступными векторами являются, например, рКК223-3 (Pharmacia Fine Chemicals, Uppsala, Швеция) и GEM1 (Promega Biotec, Madison, WI, США). Указанные фрагменты "скелета" pBR322 объединяют с подходящим промотором и структурной последовательностью, которую предстоит экспрессировать.
После трансформации подходящего штамма клеток-хозяев и культивирования штамма-хозяина до подходящей плотности клеток выбранный промотор дерепрессируют соответствующим способом (например, сменой температуры или химической индукцией), и клетки культивируют в течение дополнительного периода времени. Обычно клетки собирают центрифугированием, разрушают физическими или химическими способами и полученный неочищенный экстракт оставляют для дальнейшей очистки. Микробные клетки, использованные для экспрессии белков, могут быть разрушены с помощью любого стандартного способа, включая циклическое замораживание-оттаивание, облучение ультразвуком, механическое разрушение или использование агентов, лизирующих клетки.
Культуральные системы различных клеток млекопитающих также могут быть использованы для экспрессии рекомбинантного белка. Примерами экспрессионных систем млекопитающих являются клеточные линии COS-7 фибробластов почек зеленой мартышки, описанные у Gluzman, 1981, Cell, 23, 175, и другие клеточные линии, способные экспрессировать совместимый вектор, например, клеточные линии С127, 3Т3, СНО, HeLa, и ВНК. Экспрессирующие векторы для клеток млекопитающих должны включать сайт начала репликации, подходящий промотор и энхансер, а также по необходимости сайты связывания на рибосомах, сайт полиаденилирования, донорные и акцепторные сайты сплайсинга, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности. Последовательности ДНК, производные от генома вируса SV40, например, сайт начала репликации, ранний промотор, энхансер, сайты сплайсинга и сайты полиданилирования SV40, могут быть использованы для обеспечения необходимых нетранскрибируемых генетических элементов.
Полипептиды по настоящему изобретению могут быть выделены и очищены из культур рекомбинантных клеток с помощью методов, используемых здесь, включая осаждение сульфатом аммония или этанолом, кислотную экстракцию, анион- или катион-обменную хроматографию, фосфоцеллюлозную хроматографию, гидрофобную хроматографию взаимодействия, аффинную хроматографию, хроматографию на гидроксиапатите и лектиновую хроматографию. Если необходимо, может быть применен этап белкового рефолдинга для завершения пространственной конфигурации зрелого белка. Наконец, в качестве окончательных стадий очистки может быть применена высокоразрешающая жидкостная хроматография (ВЭЖХ).
Полипептиды по настоящему изобретению могут являться продуктом процедур химического синтеза или быть выработанными с помощью рекомбинантных технологий в прокариотическом или эукариотическом хозяине (например, культивируемыми клетками бактерий, дрожжей, высших растений, насекомых и млекопитающих). В зависимости от используемого организма-хозяина в процедуре рекомбинантной выработки полипептиды по настоящему изобретению могут быть гликозилированы углеводами млекопитающих или других эукариот или могут не быть гликозилированы. Полипептиды по настоящему изобретению также могут включать исходный остаток метионина. Выделенный или очищенный полипептид по настоящему изобретению или его биологически активная часть по существу свободны от другого клеточного материала или от культуральной среды в случае выработки его с помощью рекомбинантных технологий, или по существу свободны от химических предшественников или других химических веществ в случае химического синтеза. Предпочтительно выделенный полипептид по настоящему изобретению по существу свободен от клеточного материала и содержит менее чем примерно 30% (по сухой массе) неполипептидного или загрязняющего материала. Когда полипептид по настоящему изобретению или его биологически активную часть получают рекомбинантным путем, предпочтительно культуральная среда составляет менее чем примерно 30% объема полипептидного препарата. Когда настоящее изобретение осуществляется с помощью химического синтеза, препараты предпочтительно содержат менее чем примерно 30% по сухой массе химических предшественников или не связанных с изобретением химических веществ.
Полипептиды по настоящему изобретению могут быть эффективно выделены в соответствии с описанным в конкретных примерах далее. Препарат очищенного полипептида является чистым по крайней мере примерно на 70%; предпочтительно препарат является чистым на 85-99%. Чистоту препаратов можно оценить с помощью любого из способов, известных в данной области техники, такого как электрофорез в ДСН-полиакрил-амидном геле или масс-спектроскопия с жидкостной хроматографией.
Полинуклеотидные последовательности, кодирующие полипептид по настоящему изобретению, могут быть синтезированы целиком или частично с использованием химических методов, хорошо известных в данной области техники (см., например, Caruthers et al., 1980, Nucl. Acids Res. Symp. Ser., 215-223; Horn et al., 1980, Nucl. Acids Res. Symp. Ser., 225-232). Полинуклеотид, который кодирует полипептид, может быть затем клонирован в состав экспрессирующего вектора для экспрессии полипептида.
Как должно быть понятно специалисту в данной области техники, преимущество может иметь получение нуклеотидных последовательностей, кодирующих полипептида, включающих не встречающиеся в естественных условиях кодоны. Например, кодоны, предпочтительные для конкретного прокариотического или эукариотического хозяина, могут быть подобраны с целью увеличения интенсивности экспрессии полипептида или получения РНК-транскрипта, обладающего желательными свойствами, например, временем полужизни, которое больше по сравнению с таковым у транскрипта, полученного с последовательности, встречающейся в естественных условиях.
Нуклеотидные последовательности, описанные в данном тексте, могут быть сконструированы с применением способов, известных в данной области техники, для изменения кодирующих полипептид последовательностей с различными целями, включая, но тем самым не исчерпываясь, изменения, которые модифицируют образование, процессинг и/или экспрессию полипептида или мРНК-продукта. Для преобразования нуклеотидных последовательностей может быть использовано "перемешивание" ДНК с помощью случайной фрагментации и повторной ПЦР-сборки генных фрагментов и синтетических олигонуклеотидов. Например, сайт-специфичный (направленный) мутагенез может быть применен для внесения новых рестрикционных сайтов, изменения параметров гликозилирования, изменения предпочтения кодонов, внесения сплайсинговых вариантов, внесения мутаций и т.д.
Как альтернатива, полипептиды по настоящему изобретению могут быть получены с применением химических методов для синтеза соответствующей аминокислотной последовательности, например, путем прямого пептидного синтеза с использованием твердофазных методологий (см., например, Merrifield, 1963, J.Amer. Chem. Soc., 85, 2149-2154; Roberge et al., 1995, Science, 269, 202-204). Синтез полипептидов может быть осуществлен с использованием ручных методик или автоматически. Автоматический синтез можно осуществить, например, с использованием пептидного синтезатора модели 431А от Applied Biosystems (Perkin-Elmer). Необязательно фрагменты полипептида могут быть синтезированы по отдельности и объединены с использованием химических методов с получением полнораэмерной молекулы.
Новосинтезированный полипептид после этого может быть в существенной степени очищен с помощью высокоразрешающей жидкостной хроматографии (см., например, Creighton, 1983, "Proteins: Structures and Molecular Principles", W.H.Freeman&Co., New York, NY). Состав синтетического полипептида по настоящему изобретению может быть подтвержден с помощью аминокислотного анализа или секвенирования, например, с помощью процедуры деградации по Эдману (см. Creighton, цит. выше). Кроме того, любая часть аминокислотной последовательности полипептида может быть изменена в ходе прямого синтеза и/или объединена с помощью химических методов с последовательностями других белков с получением вариантного полипептида или химерного полипептида.
Полипептиды по настоящему изобретению вследствие способности стимулировать секрецию инсулина островковыми клетками поджелудочной железы in vitro и снижать уровень глюкозы в крови in vivo могут быть использованы для лечения диабета 2-го типа (инсулин-независимого сахарного диабета). Также полипептиды могут быть использованы для профилактики у субъектов с нарушенной переносимостью глюкозы для предотвращения развития диабета 2-го типа. Кроме того, полипептиды по настоящему изобретению могут быть использованы для лечения астмы (Bolin et al., 1995, Biopolymer, 37, 57-66; патент США №5677419) (показано, что полипептид R3PO активен по релаксации гладкой мускулатуры трахеи морской свинки); индукции пониженного артериального давления (VIP индуцирует гипотонию, тахикардию и прилив крови к лицу у пациентов-астматиков (А.Н.Morice & P.S.Sever, 1986, Peptides, 7, 279-280; A.Morice et al., 1983, The Lancet II, 1225-1227), проблем мужской репродуктивной функции (Y.Slow et al., 1999, "Effects of vasoactive intestinal peptide on human sperm motility". Arch. Androl., 43 (1), 67-71); в качестве противоапоптозного и нейропротекторного средства (D.E. Brenneman et al., 1998, "VIP neurotrophism in the central nervous system" multiple effectors and identification of a femtomolar-acting neuroprotective peptide", Ann. NY Acad. Sci., 865, 207-212); в кардиозащите при приступах ишемии (R.Kalfin et al., 1994, "Protective role of intracoronary vasoactive intestinal peptide in ischemic and reperfused myocardium", J.Pharmacol. Exp. Ther., 268 (2), 952-958; D.K.Das et al., 1998, "Coordinated role of vasoactive intestinal peptide and nitric oxide in cardioprotection", Ann. NY Acad. Sci., 865, 297-308); и наконец в качестве противоязвенного средства (Tuncel et al., 1998, "The protective effect of vasoactive intestinal peptide (VIP) on stress-induced gastric ulceration in rats", Ann. NY Acad. Sci., 865, 309-322).
Полипептиды по настоящему изобретению могут быть использованы в сочетании с подходящим фармацевтическим носителем для составления фармацевтической композиции для парентерального введения. Такие композиции содержат терапевтически эффективное количество полипептида и фармацевтически приемлемый носитель или наполнитель. Таким носителем является, тем самым не ограничиваясь, физиологический раствор, забуференный физиологический раствор, декстроза, вода, глицерин, этанол и их сочетания. Форма препарата должна удовлетворять пути введения. Также изобретение представляет фармацевтическую упаковку или набор, содержащие одну или большее число емкостей, заполненных одним или несколькими компонентами фармацевтических композиций по настоящему изобретению. В связи с указанными емкостями может находиться маркировка в форме, предусмотренной федеральным агентством, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, так чтобы такая маркировка отражала одобрение агентством по производству, применению или продаже для введения человеку. Кроме того, полипептиды по настоящему изобретению могут быть использованы в сочетании с другими терапевтическими средствами.
Фармацевтические композиции могут быть введены стандартным образом, например пероральным, топическим, внутривенным, внутрибрюшинным, внутримышечным, подкожным, интраназальным или внутрикожным путями. Фармацевтические композиции вводят в количестве, которое эффективно по лечению и/или профилактике конкретного показания. В целом, их вводят в количестве по крайней мере примерно 350 нг (0,1 нмоль) на 1 кг массы тела, а в большинстве случаев их нужно вводить в количестве, не превышающем примерно 35 мкг (10 нмоль) на 1 кг массы тела в день. В большинстве случаев дозировка составляет от примерно 0,1 мкг/кг до примерно 100 мг/кг массы тела ежедневно, принимая во внимание пути введения, симптомы и др. Указанные параметры не учитывают биологическую доступность пептида in vivo: в этом случае может быть использовано большее или меньшее количество для достижения желательной эффективной дозы. Специалист в данной области техники сможет определить с помощью экспериментальных дозировок или других стандартных подходов общее количество, которое нужно использовать для достижения эффективной дозы.
Полипептид по настоящему изобретению также может быть использован в соответствии с настоящим изобретением путем экспрессии такого полипептида in vivo, что часто обозначается как "генотерапия". Таким образом, например, клетки могут быть сконструированы с использованием полинуклеотидов (ДНК или РНК), кодирующих полипептид ех vivo, а затем сконструированные клетки вводят больному, которому проводят лечение полипептидом. Такие способы хорошо известны в данной области техники. Например, клетки могут быть сконструированы с помощью процедур, известных в данной области техники, путем использования ретровирусной частицы, включающей РНК, которая кодирует полипептид по настоящему изобретению.
Местная доставка усилителей секреции инсулина с использованием генотерапии может доставить терапевтическое средство в сайт-мишень, а именно в поджелудочную железу. Например, специфичный для поджелудочной железы промотор использовали для создания модельной мышиной опухоли бета-клеток поджелудочной железы (D.Hanahan, 1985, "Heritable formation of pancreatic beta-cell tumors in transgenic mice expressing recombinant insulin/simian virus 40 oncogenes", Nature, 315 (6015), 115-122).
Предусматриваются генотерапевтические методологии как in vitro, так и in vivo. Известен ряд методов переноса потенциальных терапевтических генов в конкретные клеточные популяции: см., например, Mulligan, 1993, "The basic science of gene therapy", Science, 260, 926-931. Указанные методы включают:
1) прямой перенос гена - см., например, Wolff et al., 1990, "Direct gene transfer into, mouse muscle in vivo". Science, 247, 1465-1468;
2) перенос ДНК с помощью липосом: см., например, Caplen et al., 1995, "Liposome-mediated CFTR gene transfer to the nasal epithelium of patients with cystic fibrosis", Nature Med., 3, 39-46; Crystal, 1995, "The gene as a drug", Nature Med., 1, 15-17; Gao& Huang, 1991, "A novel cationic liposome reagent for efficient transfection of mammalian cells", Biochem. Biophys. Res. Commun., 179, 280-285;
3) перенос ДНК с помощью ретровирусов: см., например, Кау et al., 1993, "In vivo gene therapy of hemophilia B: sustained partial correction in factor IX-deficient dogs", Science, 262, 117-119; Anderson, 1992, "Human Gene Therapy", Science, 256, 808-813;
4) перенос ДНК с использованием ДНК-вирусов. Такими ДНК-вирусами являются аденовирусы (предпочтительно векторы на основе Ad-2 или Ad-5), вирусы герпеса (предпочтительно векторы на основе вируса простого герпеса) и парвовирусы (предпочтительно векторы на основе "дефектных" или неавтономных парвовирусов, более предпочтительно векторы на основе адено-ассоциированных вирусов, наиболее предпочтительно вектором на основе AAV-2): см., например. Ali et al., 1994, "The use of DNA viruses as vectors for gene therapy". Gene Therapy, 1, 367-384; патент США №4797368, включенный здесь для сведения в виде библиографической ссылки, и патент США №5139941, включенный здесь для сведения в виде библиографической ссылки.
Выбор конкретной векторной системы для переноса интересующего гена диктуется рядом факторов. Одним из важных факторов является природа клеточной популяции-мишени. Хотя ретровирусные векторы были детально исследованы и нашли свое место в большом числе генотерапевтических применений, такие векторы в целом не пригодны для инфицирования неделящихся клеток. Кроме того, ретровирусы обладают онкогенным потенциалом. Однако недавние разработки в области лентивирусных векторов могут преодолевать некоторые из указанных ограничений: см. Naldini et al., 1996, "In vivo gene delivery and stable transduction of non-dividing cells by a lentiviral vector". Science, 272, 263-267.
Ретровирусами, от которых могут быть получены плазмидные ретровирусные векторы, упоминавшиеся выше, являются, тем самым не ограничиваясь, вирус лейкоза мыши Молони, вирус некроза селезенки, ретровирусы, такие как вирус саркомы Рауса, вирус саркомы Харвея, вирус лейкоза птиц, вирус лейкоза гиббоновых обезьян, вирус иммунодефицита человека, аденовирус, вирус миелопролиферативной саркомы и вирус опухоли молочной железы. В одном варианте воплощения ретровирусный плазмидный вектор происходит от вируса лейкоза мыши Молони.
Преимущество аденовирусов связано с тем, что они характеризуются широким кругом хозяев, могут инфицировать покоящиеся или окончательно дифференцировавшиеся клетки, такие как нейроны или гепатоциты, и, по-видимому, не являются онкогенными: см., например. Ali et al., цит. выше, стр.367. Считается, что аденовирусы не встраиваются в геном клетки-хозяина. Вследствие их внехромосомного существования риск инсерционного мутагенеза в значительной степени снижен: Ali et al., цит. выше, стр.373.
Адено-ассоциированные вирусы (AAV) имеют сходные преимущества, что и векторы на основе аденовирусов. Однако AAV характеризуются сайт-специфичным встраиванием в хромосому 19 человека (Ali et al., цит. выше, стр.377).
В предпочтительном варианте воплощения ДНК, кодирующая полипептид-усилитель секреции инсулина по настоящему изобретению, используется в генотерапии таких заболеваний, как диабет.
В соответствии с данным вариантом генотерапия с использованием ДНК, кодирующей полипептид-усилитель секреции инсулина или мутантные варианты по настоящему изобретению, применяют в отношении нуждающегося в этом больного одновременно с постановкой диагноза или сразу после этого.
Для специалиста в данной области техники должно быть понятно, что любой подходящий генотерапевтический вектор, кодирующий полипептид-усилитель секреции инсулина, ДНК или ДНК-фрагмент, производное или вариант полипептида-усилителя секреции инсулина, может быть использован в соответствии с данным вариантом воплощения. Известны методы конструирования таких векторов: см., например, W.F.Anderson, 1998, "Human gene therapy". Nature, 392, 25-30; I.M.Verma & N.Somia, 1998, "Gene therapy - promises, problems, and prospects". Nature, 389, 239-242. Внесение вектора, включающего ДНК полипептида-усилителя секреции инсулина, в сайт-мишень может быть осуществлено с помощью известных методов.
Вектор включает один или более промоторов. Подходящими для использования промоторами являются, тем самым не ограничиваясь, ретровирусный LTR; промотор SV40 и промотор цитомегаловируса (CMV) человека, описанные у Miller et al., 1989, Biotechniques, 7 (9), 980-990, или любой иной промотор (например, клеточные промоторы, такие как эукариотические клеточные промоторы, включая, но тем самым не исчерпываясь, промоторы генов гистонов, полимеразы-III и β-актина). Другими вирусными промоторами, которые можно использовать, являются, тем самым не ограничиваясь, аденовирусные промоторы, промоторы тимидинкиназы (ТК) и промоторы парвовируса В19. Выбор подходящего промотора будет ясен специалистам в данной области техники исходя из описанного в данном тексте.
Последовательность нуклеиновой кислоты, кодирующая полипептид по настоящему изобретению, находится под контролем подходящего промотора. Подходящими промоторами, которые можно использовать, являются, тем самым не ограничиваясь, аденовирусные промоторы, такие как промотор главного позднего гена аденовируса, или гетерологичные промоторы, такие как цитомегаловирусный (CMV) промотор, промотор респираторно-синцитиального вируса (RSV), индуцибельные промоторы, такие как промотор ММТ, промотор металлотионеина, промоторы теплового шока, промотор альбумина, промотор ApoAl, промоторы генов глобина человека, промоторы вирусных тимидинкиназ, такие как промотор тимидинкиназы вируса простого герпеса, ретровирусные LTR (включая модифицированные ретровирусные LTR, описанные здесь выше), промотор гена β-актина и промоторы генов гормона роста человека. Также промотор может являться нативным промотором, под контролем которого находится ген, кодирующий полипептид.
Ретровирусный плазмидный вектор используют для трансдукции упаковочных клеточных линий с целью получения линий клеток-продуцентов. Примерами упаковочных клеток, которые можно трансфицировать, являются, тем самым не ограничиваясь, клеточные линии РЕ501, РА317, ψ-2, ψ-AM, РА12, Т19-14Х, VT-19-17-H2, ψCRE, ψCRIP, GP+E-86, GP+envAml2 и DAN, описанные у Miller, 1990, Human Gene Therapy, 1, 5-14, что в полном объеме включено здесь для сведения в виде библиографической ссылки. Трансдуцировать упаковочные клетки вектором можно с помощью любого метода, известного в данной области техники. Такими методами являются, тем самым не ограничиваясь, электропорация, использование липосом и осаждение фосфатом кальция. В альтернативном варианте ретровирусный плазмидный вектор может быть упакован в липосому или соединен с липидом и после этого введен реципиенту. Линия клеток-продуцентов формирует инфекционные ретровирусные векторные частицы, которые несут последовательность(ти) нуклеиновой кислоты, кодирующую(ие) полипептиды. Такие ретровирусные векторные частицы затем можно использовать для трансдукции эукариотических клеток как in vitro, так и in vivo. Трандуцированные эукариотические клетки будут экспрессировать последовательность(ти) нуклеиновой кислоты, кодирующую(ие) полипептид. Эукариотическими клетками, которые можно трансдуцировать, являются, тем самым не исчерпываясь, эмбриональные стволовые клетки, клетки эмбриональной карциномы, а также кроветворные стволовые клетки, гепатоциты, фибробласты, миобласты, кератиноциты, клетки эндотелия и клетки эпителия бронхов. Отдельным генотерапевтическим подходом является так называемая "транскариотическая терапия", при которой клетки больного обрабатывают ex vivo с целью индукции собственных "покоящихся" хромосомных генов к выработке интересующего белка после обратного внесения больному. Транскариотическая терапия подразумевает, что у больного имеется нормальный хромосомный набор, необходимый для активации. Транскариотическая терапия включает внесение промотора или других экзогенных регуляторных последовательностей, способных активировать гены-мишени, в состав хромосомной ДНК клеток больного ех vivo, культивирование и отбор клеток, активно вырабатывающих белок, и затем обратное внесение активированных клеток в организм больного, предполагая, что после этого они полностью "приживутся". После этого "активированные по гену" клетки вырабатывают интересующий белок в течение некоторого достаточного количества времени, возможно, на протяжении всей жизни больного. Патенты США №№5641670 и 5733761 подробно описывают данный принцип: они в полном объеме включены здесь для сведения в виде библиографических ссылок.
С целью лучшего понимания настоящего изобретения приведены нижеследующие примеры. Данные примеры даны исключительно в иллюстративных целях и не призваны в чем-либо ограничить объем настоящего изобретения. Все упоминающиеся в данном тексте публикации включены в полном объеме для сведения в виде библиографических ссылок.
Пример 1. Методика выделения островков Лангерганса у крыс
Крыс линии Sprague-Dawley (275-320 г) использовали в качестве доноров островков Лангерганса. Вкратце, поджелудочные железы наполняли 10 мл холодной реконституированной Liberase RI (Boehringer Mannheim), собирали и инкубировали с дополнительными 5 мл ферментного раствора на водяной бане в течение 30 минут. Тканевые суспензии дважды промывали холодным буфером Хенкса с 10% ПТС (Gibco), ресуспендировали в 8 мл 25% фиколла (Sigma) и затем наслаивали фиколл 23%, 20% и 11% по 5 мл каждого. После центрифугирования островки Лангерганса в слое 20% отбирали, дважды промывали холодным буфером Хенкса с 10% ПТС и ресуспендировали в культуральной среде RPMI-1640 с 10% ПТС (Sigma).
Пример 2. Методика тестирования индуцированного пептидом повышения уровней инсулина у крыс
Крысы линии Wistar голодали в течение ночи (17 часов), и затем их обездвиживали фенобарбиталом (0,1 мл на 100 г массы тела). Глюкозу (0,4 г/кг, растворенную в физиологическом растворе с 1% альбумина человека) с пептидом (растворенным в физиологическом растворе с 1% альбумина человека) или без него инъецировали внутривенно в хвостовую вену.
Спустя 1 минуту после инъекции у крыс брали глазную пробу крови, и 50-100 мкл плазмы тестировали на уровень инсулина с помощью набора Linco RIA kit (Linco Research Inc., St. Charles, МО).
Пример 3. Методика определения влияния пептидов на внутрибрюшинную переносимость глюкозы у крыс
Крысы линии Wistar голодали в течение ночи, и затем их обездвиживали фенобарбиталом. У крыс брали глазные пробы крови (время 0), и пептид (в 1% человеческого альбумина) инъецировали в хвостовую вену. Через пять минут 1 г/кг глюкозы (в физиологическом растворе) инъецировали внутрибрюшинно, и у крыс брали глазные пробы крови через 15, 30 и 60 минут. Уровни плазматической глюкозы определяли с использованием автоанализатора Technicon Axon (Bayer Diagnostics Division of Bayer Corp., Tarrytown, NY), работающего в соответствии с программой SM4-2143F90 "Glucose".
Пример 4. Методика определения влияния пептидов на удерживание воды в кишечнике у крыс
Самцы крыс голодали в течение 24 часов, а поилки с водой у них отбирали за 2-3 часа до начала эксперимента. Пептид или физиологический раствор инъецировали крысам подкожно без анестезии. Через 10 минут после введения дозы крыс умерщвляли углекислотой и тонкий кишечник вырезали и взвешивали (1). Кишечник открывали разрезом, воду из просвета промахивали фильтровальной бумагой, и кишечник вновь взвешивали (2). Количество воды в кишечнике (в граммах) определяли как разницу масс: (1) минус (2).
Пример 5. Методология пептидного синтеза
Следующая базовая процедура была использована для синтеза некоторых полипептидов по настоящему изобретению. Пептидный синтез осуществляли с применением FMOC/трет-бутильного метода ("Peptide Synthesis Protocol", 1994, Vol. 35, Michael W.Pennington & Ben M.Dunn) в условиях непрерывного потока с использованием ПЭГ-полистиреновых смол Rapp-Polymere (Rapp-Polymere, Tubingen, Германия). По завершении синтеза пептиды отщепляли от смолы и снимали защиту с использованием смеси ТФК/ДТТ/вода/триизопропилсилан (88:5:5:2). Пептиды осаждали из отщепляющей смеси с использованием холодного диэтилового эфира. Преципитат трижды промывали холодным эфиром и затем растворяли в 5%-ной уксусной кислоте с последующей лиофилизацией. Пептиды проверяли с помощью хроматографии с обращенной фазой на колонке YMC-Pack ODS-AQ (YMC Inc., Wilmington, NC) с системой Waters ALLIANCE® (Waters Corp., Milford, MA) с использованием воды/ацетонитрила с 3% ТФК в градиенте от 0% до 100% ацетонитрила, а также с помощью масс-спектрометрии MALDI на масс-спектрометре VOYAGER DETM MALDI (модель 5-2386-00: PerSeptive BioSystems, Framingham, MA). Пептидный образец добавляли к матричному буферу (50:50 дистиллированной воды и ацетонитрила с 3% ТФК) в соотношении 1:1. Те пептиды, которые не удовлетворяли критериям чистоты >95%, очищали с помощью хроматографии с обращенной фазой в ВЭЖХ-системе Waters Delta Prep 4000 (Waters Corp., Milford, MA).
Пример 6. Клонирование пептидов
Попытка рекомбинантной экспрессии белка VIP была предпринята ранее с получением неоднозначных результатов. Мотивы Leu17,Gly29-VIP или Leu17,Gly29Lys30Arg31-VIP были проэкспрессированы (Simoncsits et al., 1988, Eur. J.Biochem., 178, 343-350) в виде С-концевой химеры с N-концевой частью гена β-галактозидазы E.coli в клетках E.coli. Удаление метионина из 17-го положения удаляет сайт расщепления бромцианом. Добавление с С-конца Gly или Gly-Lys-Arg было осуществлено с целью достижения возможного С-концевого амидирования in vivo действием ПАМазы млекопитающих. После расщепления химерных белков бромцианом по метионину, внесенному с N-концов мутантных вариантов VIP, высвобожденные мутантные VI Р очищали, и было показано, что они проявляют активности, сходные с таковыми у нативного VIP, хотя эти активности были измеримыми только тогда, когда пептиды имели концентрации насыщения. VIP был проэксперссирован (Raingeaud et al., 1996, Biochimie, 78, 14-25) в виде полимерных С-концевых химер с глутатион-3-трансферазой (GST) в клетках E.coli. Свободные мономерные или полимерные пептиды VIP высвобождали путем последовательного расщепления фактором Ха и гидроксиламином. Необходимость двухстадийного расщепления приводила к неэффективности и получению смеси продуктов. Полученные таким путем полимерные или мономерные пептиды VIP были менее активными по сравнению с нативным VIP. Улучшенный вариант конструкции с использованием сайта расщепления только для фактора Ха привел к получению мутантного VIP с 7-аминокислотным С-концевым удлинением и был также менее активен, чем нативный VIP (Ottavi et al., 1998, Biochimie, 80, 289-293). По экспрессии РАСАР опубликованных данных к настоящему времени нет. Для разработки эффективного способа экспрессии РАСАР, VIP и их мутантных вариантов генетические коды названных пептидов были клонированы как С-концевые к гену GST с единственным сайтом расщепления фактором Ха, разделяющим мономерный пептид и GST. Ген, кодирующий сайт распознавания фактором Ха, химеризованный на последовательность ДНК, кодирующей пептид, который должен быть получен, был синтезирован путем гибридизации двух перекрывающихся одноцепочечных ДНК-фрагментов (70-90-нуклеотидных), включающих рестрикционные сайты для BamHI или XhoI непосредственно с 5'-стороны от последовательности ДНК того гена, который предстоит клонировать, с последующим синтезом ДНК в противоположных цепях с использованием большого фрагмента ДНК-полимеразы I (Life Technologies Inc., Gaithersburg, MD). Последовательность ДНК, выбранная для каждого из генов, основывалась на обратной трансляции сконструированной аминокислотной последовательности каждого пептида. В некоторых случаях ген, кодирующий пептид, получают методом ПЦР-мутагенеза (V.Picard et al., 1994, Nucleic Acids Res., 22, 2587-2591; J.Sambrook, E.F.Fritsch & T.Maniatis, 1989, "Molecular Cloning: A Laboratory Manual", 2d ed., Cold Spring Harbor Laboratory Press, New York) уже имеющегося гена с помощью способа, описанного выше. Двухцепочечный продукт затем разрезают рестриктазами BamHI и XhoI и лигируют в состав вектора pGEX-6P-1 (Amersham Pharmacia Biotech), который был предварительно также расщеплен рестриктазами BamHI и XhoI. Последовательности ДНК клонированных пептидных генов приведены на фигуре 8.
Например, в случае клонирования последовательностей ДНК SEQ ID NO 54, 55 и 56 в состав pGEX-6P-1 следующие полипептидные последовательности были экспрессированы в виде химер с глутатион-3-трансферазой (GST):
РАСАР38: IEGRHSDGIFTDSYSRYRKQMAVKKYLAAVLGKRYQRVKNK (SEQ ID NO:2)
VIP: IEGRHSDAVFTDNYTRLRKQMAVKKYLNSILN (SEQ ID NO:1)
R3P3: IEGRHSDAVFTENYTKLRKQLAAKKYLNDLKKGGT (SEQ 10 NO:8)
Рестрикционная карта типичной плазмиды, кодирующей GST-пептидную химеру, показана на фигуре 3.
Пример 7. Экспрессия и очистка рекомбинантного пептида
Клетки BL21 (Stratagene), трансформированные плазмидами, кодирующими GST-пептидную химеру, культивировали при 37°С до достижения OD600=0,6-1,0 и индуцировали действием 1 мМ IPTG (Life Technologies) в течение 2 часов при 37°С. 2 л клеток центрифугировали при 7700g в течение 15 минут, взвешивали и хранили при -20°С в течение по крайней мере 3 часов. Сгусток замороженных клеток ресуспендировали в 100 мл охлажденного на льду ФСБ с 250 мкл смеси протеазных ингибиторов (Sigma Chemical: каталожный №Р-8465) в расчете на 1 г клеток, обрабатывали ультразвуком трижды по 1 минуте с 15-секундными интервалами. Клеточный дебрис осаждали центрифугированием при 10000g в течение 20 минут. Надосадочную фракцию смешивали с 2 мл 50%-ной глутатион-сефарозной смолы 4В (Pharmacia) в шейкере в течение ночи при 4°С. Смолы центрифугировали при 1500g в течение 15 минут, упаковывали в пустые хроматографические колонки Poly-Prep (Bio-Rad), промывали 30 мл ФСБ и затем 10 мл буфера для фактора Ха (1 мм CaCl2, 100 мМ NaCl и 50 мМ Трис-HCl, рН 8,0). Пептиды отщепляли от колонки действием 60 единиц фактора Ха (Pharmacia) в 1 мл буфера для фактора Ха в течение ночи при 4°С и пропускали через ВЭЖХ-колонку С18 (Beckman System Gold) с использованием 2-мл петли и скорости потока 2 мл/мин со следующей программой: 10 минут для буфера А (0,1% ТФК в воде), 30 минут для градиента к буферу В (0,1% ТФК в ацетонитриле), 10 минут для буфера А, 10 минут для градиента и 10 минут для буфера А. Пиковые фракции (каждая по 1 мл) собирали и анализировали электрофоретически в ДСН-геле с 10-20% трицина. Фракции, содержащие пептиды таблицы 1, объединяли и высушивали. Типичные выходы составили по несколько сот микрограмм свободных пептидов на 1 литр культуры E.coli. Рекомбинантные пептиды, как было показано, обладают теми же активностями, что и их синтетические версии.
Таблица 1 включает некоторые отобранные полипептиды, полученные в соответствии с обсуждавшейся выше методикой пептидного синтеза (пример 5) или рекомбинантным путем, что описано в примере 7. Пептиды, полученные рекомбинантным путем, отмечены строчной буквой "r" впереди от обозначения пептида. Пептиды, полученные и рекомбинантным, и синтетическим путями, обозначены звездочкой после обозначения пептида. Пептиды R3PO и R3P4 были описаны у Bolin et al., 1995, Biopolymers, 37, 57-66 и в патенте США №5677419.



Пример 8. Секреция инсулина островками Лангерганса крыс
Пептид R3P3 (0,1-100 нМ) стимулирует секрецию инсулина выделенными островками Лангерганса крыс в глюкозо-зависимом режиме. В проведенных исследованиях сравнивают влияние пептида R3P3 и GLP-1 на панкреатические клетки.
Островки Лангерганса крыс были выделены и обработаны GLP-1 или R3P3 при 3 или 8 мМ глюкозы в культуральной среде в соответствии с методикой выделения островков Лангерганса крыс, описанной в примере 1. Как показано на фигурах 4А-4В, пептид R3P3 значительно усиливает секрецию инсулина островками Лангерганса в зависимости от концентрации, и такой эффект сходен с выявленным для GLP-1 - известного усилителя секреции инсулина.
Пример 9. Инсулиновая и глюкозная реакция in vivo
Как показано далее в таблицах 2 и 3, полипептиды, которые активируют рецептор R3, также усиливает индуцируемое глюкозой повышение уровней плазматического инсулина по сравнению с действием одной глюкозы. Такое повышение инсулина обусловливает согласованное снижение уровня плазматической глюкозы.
В соответствии с методикой примера 2, описанной выше, голодавших в течение ночи крыс линии Wistar обездвиживали фенобарбиталом, внутривенно инъецировали глюкозу с пептидом или без него и брали глазную пробу крови через 1 минуту. В каждой группе было по 12 крыс. График на фигуре 5 показывает, что полипептид R3P3 обусловливает усиление выведения глюкозы, что сопровождается интенсификацией секреции инсулина. Пептид или наполнитель вводили внутривенно с последующей глюкозной загрузкой внутрибрюшинно в соответствии с описанным в примере 3. В течение некоторого времени после этого определяли уровень плазматической глюкозы. Как видно из графика, R3P3 существенно ускоряет выведение глюкозы из крови.
Пример 10. Диарейные побочные эффекты
Как описано выше в методике примера 4, голодавшим крысам подкожно вводили исследуемый пептид (5 нмоль/кг или 22-24 нмоль/кг). Через пять минут после инъекции 0,3 мл воды давали перорально. Через 5 минут после водной дозы животных умерщвляли и определяли содержание воды в тонком кишечнике. Как показано на фигуре 6, инъекции VIP при двух дозах приводили к существенному увеличению содержания воды в просвете тонкого кишечника по сравнению с контролем (наполнитель: физиологический раствор). При наивысшей дозе пептиды R3 обусловливали только очень небольшое повышение (т.е. грубо около 10%) по сравнению с VIP. При дозе 5 нмоль/кг пептиды не вызывали каких-либо изменений в содержании воды в тонком кишечнике. Степень удерживания воды использовали в качестве показателя активации R2 in vivo.
Пример 11. Влияние пептидов на внутрибрюшинную переносимость глюкозы у крыс
Крысы линии Wistar голодали в течение ночи, и затем их обездвиживали фенобарбиталом. У крыс брали глазные пробы крови (время 0) и подкожно вводили пептид (в 1% человеческого альбумина). Через пять минут внутрибрюшинно вводили 1 г/кг глюкозы (в физиологическом растворе) и у крыс брали глазные пробы крови через 30 минут. Уровни плазматической глюкозы определяли на автоанализаторе Axon, а результаты показаны на фигуре 7.
Из фигуры 7 видно, что уровень глюкозы в плазме возрастает до 160 мг/дл по сравнению с нормой (100 мг/дл) у крыс, которым вводили только наполнитель, через 30 минут после теста IPGTT (тест на внутрибрюшинную переносимость глюкозы). У крыс, которым вводили пептиды R3P3, R3P12 и R3P13, такое повышение плазматической глюкозы было существенно снижено, что указывает на эффект выработки инсулина. При подкожном введении 1 нмоль/кг эффект по снижению глюкозы у каждого пептида был сходным с эффектом, наблюдаемым при эквивалентной дозе GLP-1.
Пример 12. Полипептиды, секретирующие инсулин в глюкозо-зависимом режиме
В таблице 2 приведен список пептидов, которые стимулируют секрецию инсулина in vivo в тесте IPGTT или in vitro в тесте с островками Лангерганса крыс. Как видно из этих данных, пептиды усиливают опосредованную глюкозой секрецию инсулина и in vivo, и in vitro.
Плазматический инсулин. Данные выражены в процентах от уровня плазматического инсулина через 1 минуту после IVGTT (0,4 г/кг глюкозы) вместе либо с физиологическим раствором, либо 0,1 нмоль/кг Р51, Р55, Р60, Р66, либо 1 нмоль/кг других пептидов у крыс линии Wistar. Кровь брали из глаза, и уровень инсулина измеряли с помощью набора для анализа крысиного инсулина Rat Insulin RIA kit (Linco Research Inc., St. Charles, МО).
Плазматическая глюкоза: Данные выражены в процентах от области плазматической глюкозы под кривой после IPGTT (1 г/кг глюкозы) варианта с наполнителем после введения дозы 1 нмоль/кг пептида. Как было сообщено (К.Filipsson et al., 1997, J.Clin. Endocrinol. & Metabolism, 82, 3093-3098), PACAP-27 индуцирует секрецию инсулина, но не влияет на уровни плазматической глюкозы. Авторы настоящего изобретения впервые показали, что это обусловливается взаимным влиянием рецепторов R2 и R3 на тенденцию к выведению глюкозы. Избирательный по R2 агонист [K15,R16,L27]VIP(1-7)/GRF(8-27) (P.Gourlet et al., 1997, Peptides, 18, 1539-1545), обозначенный как R2P1, повышает уровень плазматической глюкозы на 14%, в то время как избирательные по R3 агонисты снижают уровень плазматической глюкозы примерно на 20%. Следовательно, избирательность в отношении R3 является желательным свойством лекарственных средств, которые должны быть использованы для достижения снижения уровней глюкозы в крови с целью лечения диабета 2-го типа.
Секреция инсулина островками Лангерганса. Островки Лангерганса были выделены у крыс линии Sprague-Dawley в соответствии с описанным в примере 1 и обработаны либо наполнителем, либо конкретным пептидом (10 нМ) в течение 2 часов. Концентрацию инсулина в культуральной среде определяли с помощью набора реактивов Linco Rat Insulin RIA kit. Концентрация глюкозы в среде составляла 8 мМ. Данные выражены в процентах (инсулина) от варианта с присутствием только 8 мМ глюкозы. При 3 мМ глюкозы не было выявлено индуцируемого полипептидом повышения концентрации инсулина: следовательно, активность по секреции инсулина у указанных полипептидов является глюкозо-зависимой.
Пример 13. Фармацевтическая композиция: внутривенный препарат
Стерильный препарат для инъекции приготавливают из 4 мг полипептида SEQ ID NO 72 и 1 литра стерильного физиологического раствора с использованием производственного процесса, хорошо известного в данной области техники.
Пример 14. Фармацевтическая композиция: внутривенный препарат
Стерильный препарат для инъекции приготавливают из 400 мг полипептида SEQ ID NO 174 и 1 литра стерильного физиологического раствора с использованием производственного процесса, хорошо известного в данной области техники.
Пример 15. Влияние рецепторно-избирательных пептидных агонистов РАСАР-27, VIP и РАСАР на частоту сердечных сокращений у собак в сознании
Методика:
Гончих собак помещали в повязку так, чтобы приучить их находиться на ногах в течение 3 часов. Манжету кардиографа располагали вокруг хвоста собаки.
Физиологический раствор инъецировали в головную вену и частоту сердечных сокращений контролировали каждые 2 минуты для установления нормального значения. Спустя 10 минут инъецировали пептид и контролировали частоту сердечных сокращений каждые 2 минуты в течение еще 20 минут. Если в течение указанного времени частота сокращений была нормальной, давали более высокую дозу и контролировали частоту сокращений в течение 20 минут. Область под кривой (ОПК) в первые 10 минут индуцированного полипептидом изменения частоты сердечных сокращений как процент превышения ОПК наполнителя соотносили на графике с концентрацией полипептида. РАСАР-27 и являющийся избирательным по R1 агонистом максидилан (О.Moro, 1997, J.Biol. Chem., 272, 966-970) проявляют сходную активность по повышению частоты сердечных сокращений. VIP, который активирует и R2, и R3, но не R1, избирательный по R2 агонист R2P1 и избирательные по R3 агонисты R3P0, R3P3, R3P19, R3P36, R3P51 и R3P53 по крайней мере в 10 раз менее активны по сравнению с РАСАР-27 или максидиланом. Таким образом, сердечно-сосудистое действие РАСАР-27 в основном может определяться активацией PACAP-R1. Все пептиды являются абсолютными агонистами в отношении соответствующих рецепторов при сходном уровне аффинности. R3P0 является Roche-аналогом RO 25-1553, для которого было показано проявление избирательности PACAP-R3 по крайней мере в 100 раз более высокой, чем по отношению к R1 и R2 (Gourlet et al., 1997, Peptides, 18, 403-408).
Пример 16. SPA-тест циклического АМФ
Клетки СНО, экспрессирующие PACAP-R3, высевали на 96-луночные планшеты (Costar) при плотности 8×104 клеток на лунку и культивировали при 37°С в течение 24 часов в среде α-МЕМ с нуклеозидами, глутамином (Gibco BRL), 10% ПТС, 100 мкг/мл пенициллина/стрептомицина, 0,3 мг/мл глутамина, 1 мМ HEPES, 0,5 мг/мл генетицина (Gibco BRL). Среду удаляли и планшеты промывали ФСБ. Клетки инкубировали с пептидом в HEPES-ФСБ-БСА с 0,4 мг/мл соевого трипсинового ингибитора, 0,5 мг/мл бацитрацина, 100 мкМ IBMX в течение 15 минут при 37°С. Циклический АМФ в клеточных экстрактах оценивали количественно с использованием тест-системы для прямого SPA-скрининга цАМФ (Amersham Pharmacia Biotech Inc., Piscataway, NJ). Пептиды, показанные в таблице 3, были протестированы на активность цАМФ.
В таблице 3 показаны результаты SPA-анализа цАМФ in vitro в клетках СНО, трансфецированных PACAP-R2 или PACAP-R3. ЕС50 определяют как концентрацию полипептида, при которой достигается 50% максимальной активности РАСАР-27. "NA" обозначает отсутствие выявляемой активности. "Избирательность по R3" определяют из отношения ЕС50 при R2 к ЕС50 при R3. Большинство из следующих полипептидов конструируют на основе VIP, для которого было показано отсутствие активности в отношении R1 (D.Vaudry et al., 2000, Pharmacological Reviews, 52, 269-324). Следовательно, считается, что указанные полипептиды не будут проявлять какую-либо активность в отношении R1. Пептиды, сформированные по параметрам VIP, включают SEQ ID NO 6-53, 62-65 и 70-175.
Пример 17 Получение поликлональных антител
Синтез пептида Ac-CRKQVAAKKYLQSIKNKRY-COOH осуществляли с помощью пептидного синтезатора Applied Biosystems 430A с использованием Fmoc-химии при активации аминокислот действием HBTU. Пептид отщепляли с использованием смеси 84,6% ТФК, 4,4% фенола, 4,4% воды, 4,4% тиоанизола и 2,2% этандитиола. Неочищенный пептид очищали с использованием колонки С18 с обращенной фазой с градиентом 0,1% ТФК в СН3CN. Оценку чистоты проводили с помощью масс-спектрометра PerSeptive V Biosystems Voyager DE Pro MALDI. Остаток цистеина присоединяли к KLH с использованием набора реактивов Pierce Imject Maleimide Activated mcKLH kit и соответствующей прописи (Pierce, Rockford, IL). Кроликов иммунизовали с использованием следующей схемы получения поликлональной антисыворотки:
День 0 - 10 мл предварительной пробы крови для оценки нормальных параметров сыворотки. 250 мкг каждого пептида в 1 мл эмульсии полного адъюванта Фройнда: 0,1 мл на сайт на 10 сайтов, подкожно.
День 14 - бустинг 250 мкг каждого пептида в 1 мл эмульсии неполного адъюванта Фройнда: 0,1 мл на сайт на 10 сайтов, подкожно.
День 21 - 35-мл проба крови.
День 35 - бустинг 250 мкг каждого пептида в 1 мл эмульсии неполного адъюванта Фройнда: 0,1 мл на сайт на 10 сайтов, подкожно.
День 42 - 35-мл проба крови.
День 56 - бустинг 250 мкг каждого пептида в 1 мл эмульсии неполного адъюванта Фройнда: 0,1 мл на сайт на 10 сайтов, подкожно.
День 63 - 35-мл проба крови.
День 77 - бустинг 250 мкг каждого пептида в 1 мл эмульсии неполного адъюванта Фройнда: 0,1 мл на сайт на 10 сайтов, подкожно.
День 84 - окончательная проба крови.
Антитела анализировали с использованием следующей методики.
Планшет Immulon-III (Dynatech Laboratories Inc., Chantilly, VI) покрывали пептидом-Р66 (диапазон 0,3-100 нг) в 100 мкл EIA-покрывающего буфера (1,6 л 0,1 М NaHCO3+0,4 л 0,1 М Na2СО3, рН 9,5) в течение 3 часов при комнатной температуре. Планшет блоттировали 100 мкл 5% молока в TBS Твин-20 (10 мМ Трис, рН 8,0; 150 мМ NaCl, 0,05% Твин-20 (Sigma, P-1379)) в течение 1 часа при комнатной температуре и трижды промывали в TBS Твин. Антитело к R3P66 добавляли в лунку в 100 мкл 5% "bloto" (TBS Твин-20 + 5% молока) в течение 2 часов при комнатной температуре с последующей промывкой в TBS Твин (промывку повторяли пятикратно). Вторичное антитело (конъюгат козьего антикроличьего антитела с щелочной фосфатазой: BioRad) добавляли в лунку в разведении 1:1000 в 100 мкл 5% "bloto" на 1 час. Планшет 5 раз промывали в TBS Твин. 0,5 мг/мл р-нитрофенилфосфата (Sigma 104-105) в 100 мкл субстратного буфера (1 М диэтаноламина, 0,5 мкМ MgCl2•6H2O, рН 9,8; w/HCl) инкубировали при комнатной температуре в течение 1 часа до O/N. Планшет считывали при OD405 с помощью SPECTRAmax-250 (Molecular Devices Corp., Sunnyvale, CA).
Для определения того, распознает ли антитело, выработанное в кроликах к пептиду Ac-CRKQVAAKKYLQSIKNKRY-COOH в соответствии с примером 17, пептид R3P66 (SEQ ID NO 72), проводили твердофазный иммуноферментный анализ (ТИФА). Если антитела распознавали пептид, то выявляется сигнал OD405. На фигуре 10 показано, что такие антитела распознают R3P66, но не взаимодействуют с гомологичными пептидами РАСАР-27 или VIP вплоть до концентрации пептида 30 мкг.
Пример 18. Гиперчувствительность дыхательных путей в модели острой астмы у приматов
Самцов макак (Масаса fascicularis), использованных в данном анализе, содержали в условиях постоянной температуры и влажности при 12-часовом световом цикле. Их кормили дважды в сутки за исключением дня эксперимента, когда их оставляли без пищи в ночь перед процедурой. Все время эксперимента воду давали без ограничений.
Гиперчувствительность дыхательных путей (ГДП) (основную) измеряли за 1 неделю до контроля (без воздействия), при первом введении антигена и опять через 24 часа после введения антигена. Индуцированную антигеном гиперчувствительность дыхательных путей измеряли по снижению РС100 (концентрации метахолина, необходимой для того, чтобы обусловить 100%-ное повышение легочной сопротивляемости) через 24 часа по сравнению с основным значением. После 2-недельного отдыха проводили другое измерение основной ГДП. Еще через неделю пептиды по настоящему изобретению вводили аэрозольным путем за 10 минут до введения антигена. Через 24 часа вновь измеряли ГДП, и снижение РС100 в результате обработки сравнивали с величиной без обработки.
Экспериментальная процедура. В каждый день эксперимента животных обездвиживали смесью кетамина/ксилазина (70:12 мг/кг при 0,1 мл/кг) непосредственно в вольере. Под наркозом их переносили в лабораторию, где их помещали в положение "на спине" на подогретое водяное покрывало на тележке. В каждый глаз наносили глазную мазь и 0,2 мл лидокаина (2%) распыляли в гортань и по задней части глотки. Челюсти удерживали раздвинутыми с помощью челюстного расширителя и снабженную манжетой эндотрахейную трубку калибра 5,0 (конец которой обильно смазан ксилокаиновым гелем, 2%) вводили для проведения ларингоскопического анализа. Затем животных помещали в специально сконструированное кресло-ограничитель таким образом, что животное оказывалось в положении небольшого отклонения назад, но в вертикальном сидячем положении, причем удерживалось только ошейником за шею. Животное было окружено водонагреваемым покрывалом.
Эндотрахейную трубку присоединяли к аппарату искусственного дыхания Harvard, настроенному на 30-35 дыхательных актов в минуту. Поток воздуха измеряли с помощью пневмотахографа Fleisch, и грудное давление определяли с помощью датчика давления "validyne" (как разность между давлением на дальнем конце трубки и комнатным давлением).
Пневмотахограф и Validyne присоединяли к предварительному усилителю и затем к респираторному анализатору MI2. С использованием первичных сигналов потока и давления анализатор подсчитывал сопротивляемость и эластичность дыхательных путей (а также ряд других параметров дыхания). Исходное 5-6-минутное измерение проводили для того, чтобы убедиться в стабильности сигнала и в том, что значения сопротивляемости и эластичности находятся в измеряемых пределах.
Введение антигена. Проводили ингаляционное введение свиной аскариды Ascarium suum. Имеющийся экстракт свиной аскариды разводили в 10 раз ФСБ с получением раствора 1000 мкг/мл. Аэрозоль вводили под давлением с помощью пульверизатора Rainbow (Puritan-Bennett), присоединенного к аппарату искусственного дыхания Bird mark 7A, установленному на 15 дыхательных актов в минуту. Проводили 30 дыхательных актов с введением антигеном, после чего в течение 15 минут контролировали острое сокращение бронхов.
По окончании введения животное отключали от аппарата искусственного дыхания, и после того, как оно могло дышать самостоятельно, его высвобождали из кресла-ограничителя и клали на спину на тележку. По восстановлении нормальных рефлексов (моргание глаз, глотательный рефлекс) наряду с мышечным тонусом конечностей животных возвращали в виварий.
Введение пептида. Анализируемые пептиды доставляли с помощью ингаляции в соответствии с описанным выше. Базовый раствор пептидов разбавляли стерильной водой до достижения концентрации 0,5 мкг на 4 мкл. Исходя из предыдущих измерений пульверизатор, как было показано, доставлял по 4 мкл на один акт дыхания, и число дыхательных актов для каждого животного корректировали так, чтобы доставить точную конечную концентрацию из пульверизатора. Для каждого пептида конечные доставляемые концентрации были следующими:
R3P0 (SEQ ID NO 5) - 0,6 мкг/кг;
R3P76 (SEQ ID NO 82) - 3 мкг/кг;
R3P82 (SEQ ID NO 88) - 1,8 мкг/кг.
Введение метахолина. Кривые реакции на дозы метахолина выстраивали для оценки гиперчувствительности дыхательных путей. В модели острого ответа измерение проводили через 24 часа и сравнивали с чувствительностью за 7 дней до эксперимента. Аэрозольный фосфатно-солевой буфер (ФСБ) доставляли с помощью пульверизатора в соответствии с описанным выше. Аэрозоль вводили за 15 дыхательных актов и затем сопротивляемость легких контролировали в течение 10 минут. Метахолин (Sigma) доводили до концентрации 100 мг/мл в ФСБ и такой маточный раствор разбавляли ФСБ до конечного диапазона концентраций от 0,1 мг/мл до 100 мг/мл. Через 10 минут после введения метахолина (0,1 мг/мл, 15 дыхательных актов) проводили дополнительный анализ. Последовательные дозы метахолина вводили с повышением дозы с полулогарифмическим шагом либо до момента удвоения легочной сопротивляемости, либо до максимальной дозы метахолина (100 мг/кг). Основная сопротивляемость (0%) была принята как сопротивляемость, достигаемая в ответ на введение ФСБ. Повышение сопротивляемости легких (%) и дозы метахолина вносили в таблицу значений, и величину РС100 (доза метахолина, обусловливающая повышение сопротивляемости на 100%) подсчитывали по графику зависимости сопротивляемости от дозы. Полученные величины преобразовывали в величины log10. Разность РС100 (значение на 24-м часу минус основное значение) для эксперимента с воздействием сравнивали с данными контроля.
В таблице 4 показаны параметры РС100 для трех пептидов. Как видно из приведенных данных, пептиды эффективны против вызванной антигеном гиперчувствительности дыхательных путей и поэтому являются потенциальными средствами для лечения астмы.
Все публикации и патенты, упоминавшиеся выше в данной заявке, включены здесь для сведения в виде библиографических ссылок. Различные модификации и изменения описанных композиций и способов по настоящему изобретению будут ясны специалистам в данной области техники без отклонения от сущности и объема настоящего изобретения. Хотя изобретение было описано в связи с конкретными предпочтительными вариантами, должно быть понятно, что заявленное изобретение не должно без оснований ограничиваться такими конкретными вариантами. На самом деле различные модификации описанных выше путей воплощения изобретения, которые будут ясны специалистам в области молекулярной биологии или родственных областей техники, призваны попасть в объем нижеследующей формулы изобретения. Специалистам в данной области техники станут ясны, или в результате постановки только стандартных экспериментов ими будут определены многочисленные эквиваленты конкретных вариантов воплощения настоящего изобретения, описанных в данном тексте. Такие эквиваленты рассматриваются как удовлетворяющие нижеследующей формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕЙРОПЕПТИДЫ ДЛЯ КУЛЬТУРЫ ВОДНЫХ ОРГАНИЗМОВ | 2006 |
|
RU2409027C2 |
| КЛЕТКИ-ХОЗЯЕВА И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2014 |
|
RU2663587C2 |
| ЭКСПРЕССИЯ МНОЖЕСТВА ГЕНОВ, ВКЛЮЧАЯ sORF-КОНСТРУКЦИИ, И СПОСОБЫ ЭКСПРЕССИРОВАНИЯ ИММУНОГЛОБУЛИНА | 2006 |
|
RU2478709C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИАНГИОГЕННОГО ПОЛИПЕПТИДА | 2012 |
|
RU2488635C1 |
| СИГНАЛЬНЫЕ ПЕПТИДЫ ТАТ ДЛЯ ПРОДУКЦИИ БЕЛКОВ В ПРОКАРИОТАХ | 2008 |
|
RU2487937C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЦЕЛЕВОГО СЕКРЕТИРУЕМОГО БЕЛКА В ДРОЖЖАХ Saccharomyces cerevisiae | 2012 |
|
RU2502805C1 |
| КОМПОЗИЦИИ И СПОСОБЫ, СВЯЗАННЫЕ С РАСТВОРИМЫМИ СОПРЯЖЕННЫМИ С G-БЕЛКОМ РЕЦЕПТОРАМИ (SGPCR) | 2006 |
|
RU2423378C2 |
| ПОЛИПЕПТИД ОЖИРЕНИЯ (ОВ)(ВАРИАНТЫ), ЕГО АНАЛОГ (ВАРИАНТЫ) И СЛИТЫЙ БЕЛОК (ВАРИАНТЫ), ИЗОЛИРОВАННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, МОЛЕКУЛА ДНК, РЕКОМБИНАНТНЫЙ ВЕКТОР КЛОНИРОВАНИЯ, РЕКОМБИНАНТНЫЙ ВЕКТОР ЭКСПРЕССИИ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, МОНОКЛОНАЛЬНОЕ И ПОЛИКЛОНАЛЬНОЕ АНТИТЕЛО | 1995 |
|
RU2273645C9 |
| ГЕН НОВОЙ СЕРИНОВОЙ ПРОТЕАЗЫ, РОДСТВЕННОЙ DPPIV | 2001 |
|
RU2305133C2 |
| СЛИТЫЕ КОНСТРУКЦИИ ЛЕКАРСТВЕННОГО СРЕДСТВА И КОНЪЮГАТЫ | 2005 |
|
RU2428431C2 |






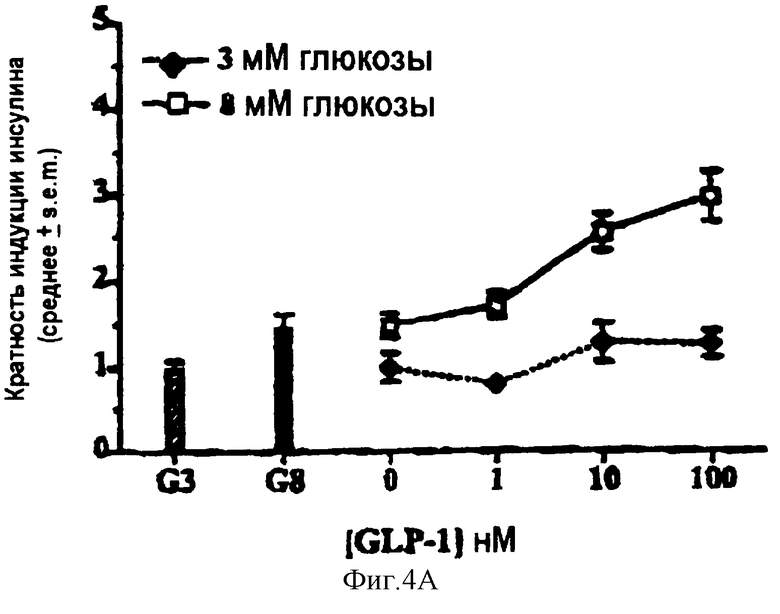
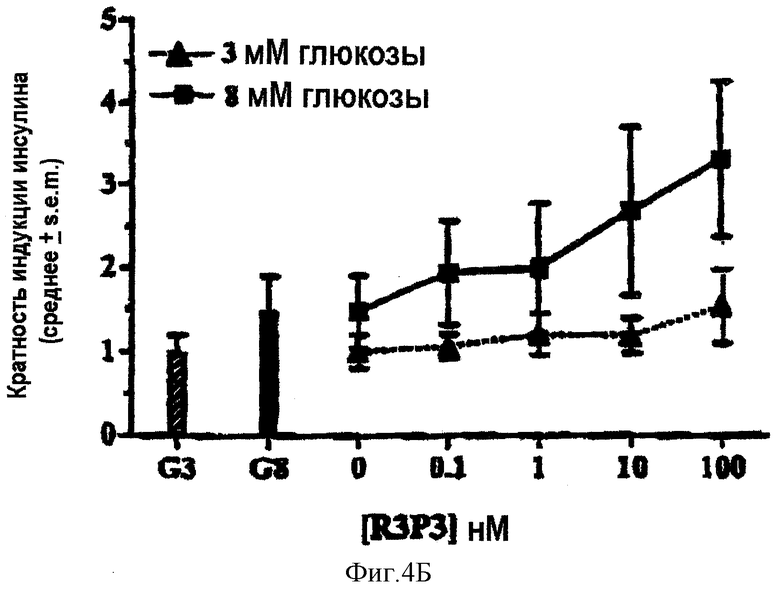
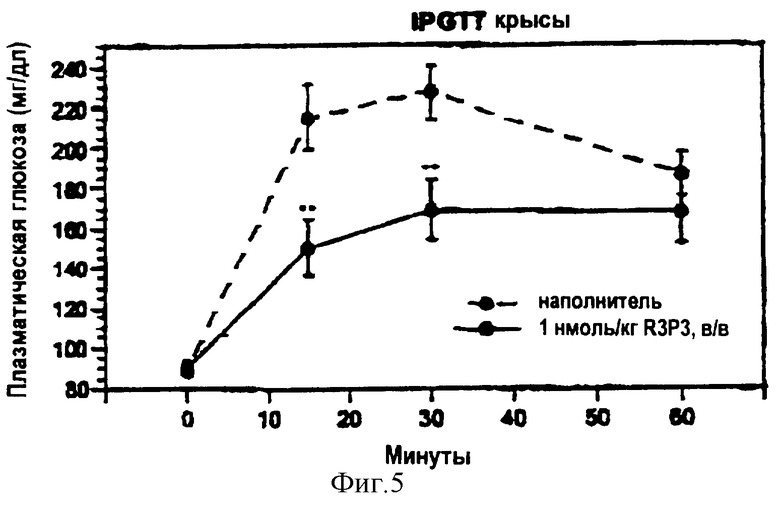
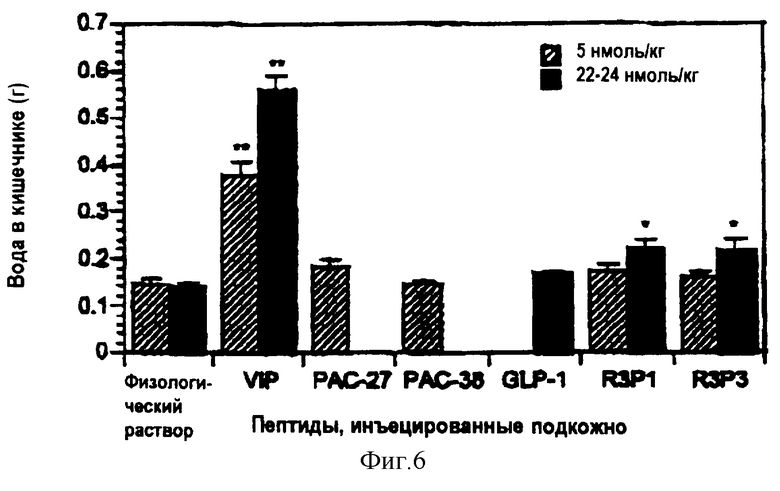
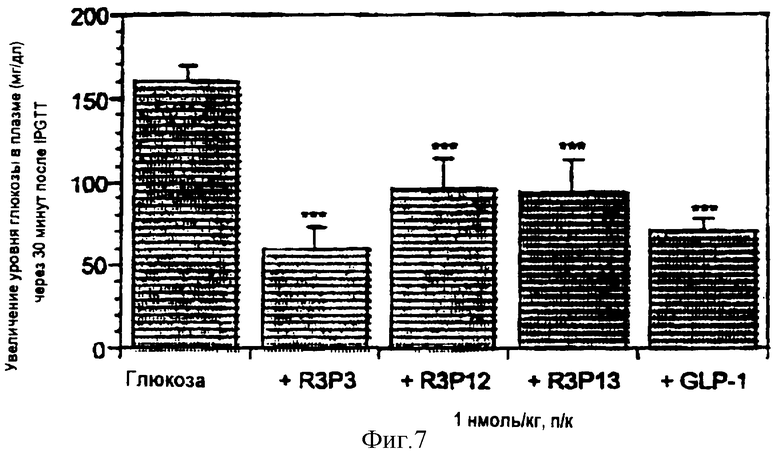










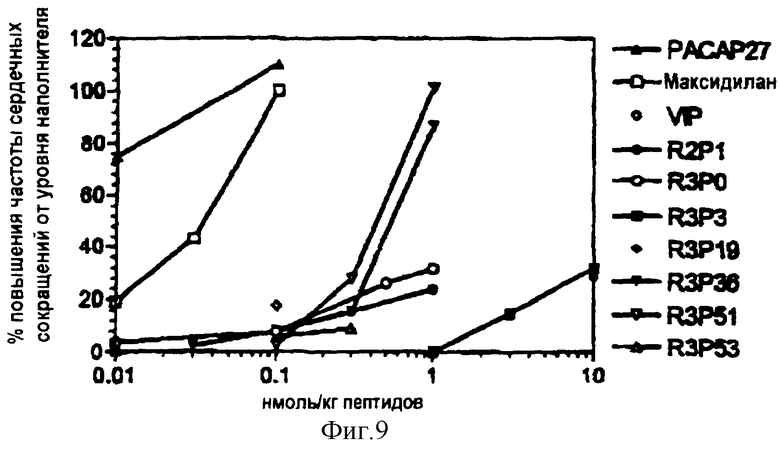
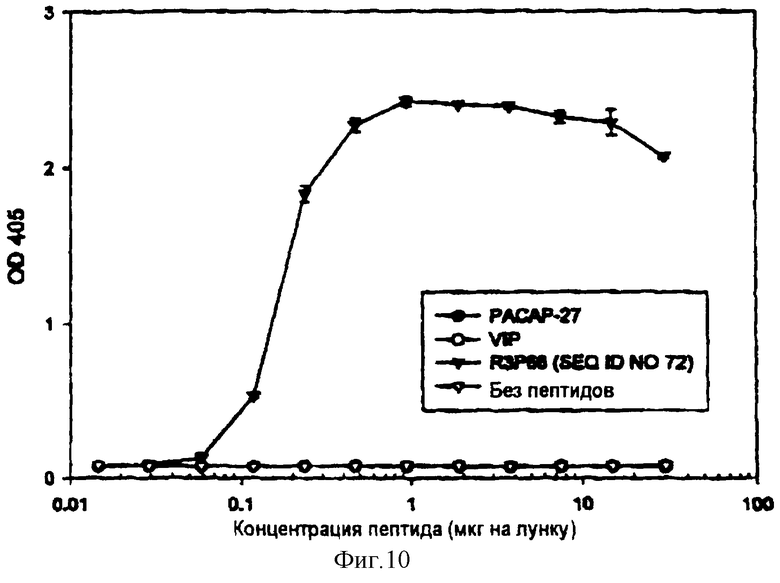
Изобретение представляет новые пептиды, которые функционируют in vivo как стимуляторы секреции инсулина бета-клетками поджелудочной железы в глюкозо-зависимом режиме. Показано, что такие пептиды-усилители секреции инсулина стимулируют секрецию инсулина островковыми клетками крыс in vitro и in vivo. Пептиды по настоящему изобретению представляют новый путь лечения больных со сниженной секрецией эндогенного инсулина, в частности в случае диабета 2-го типа. В частности, изобретение представляет полипептид, выбранный из конкретной группы VIP/PACAP - родственных полипептидов или их функциональных эквивалентов. Также заявляются способы получения как рекомбинантных, так и синтетических пептидов. Преимущество изобретения заключается в новых пептидах, которые могут применяться как стимуляторы секреции инсулина. 13 н. и 34 з.п. ф-лы, 4 табл., 10 ил.
(a) культивирование клетки-хозяина, содержащей вектор по п.30, в условиях, подходящих для экспрессии указанного полипептида, и
(b) выделение полипептида из культивированной клетки-хозяина.
(a) культивирование культуры клеток по п.33 в условиях, подходящих для экспрессии указанного полипептида, и
(b) выделение полипептида из клеточной культуры.
Приоритет по пунктам:
| US 5677419 А, 14.10.1997 | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| GOURLET P | |||
| et al., Vasoactive intestinal peptide modification at position 22 allows discrimination between receptor sybtypes, Eur | |||
| j, Pharmacol., 1998, v.348 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

