Настоящее изобретение относится к применению РНК и водного инъекционного буфера, содержащего натриевую соль, кальциевую соль, необязательно калиевую соль и необязательно также лактат, для приготовления раствора для инъекции РНК, предназначенного для увеличения переноса РНК и/или трансляции РНК в организм(е)-хозяин(е).
Медицинские методы, основанные на молекулярной биологии, такие как генная терапия и генетическая вакцинация, играют главную роль в лечении и предупреждении многочисленных заболеваний. Такие методы основаны на интродукции нуклеиновых кислот в клетки или ткани пациента и последующей обработке информации, кодируемой интродуцированными нуклеиновыми кислотами, т.е. трансляции в требуемые полипептиды или белки. В качестве предназначенных для интродукции нуклеиновых кислот можно применять как ДНК, так и РНК.
Генетические вакцинации, которые заключаются в инъекции «оголенной» плазмидной ДНК, впервые были осуществлены в начале 1990-х годов на мышах. Однако при проведении фазы I/II клинических исследований на человеке стало ясно, что эта технология не может удовлетворить ожидания, которые пробудили исследования на мышах1. С тех пор были разработаны многочисленные методы генетических вакцинаций, основанных на использовании ДНК, и методы интродукции ДНК в клетки (среди прочего такие методы, как трансфекция с использованием фосфата кальция, трансфекция с использованием полипрена, слияние протопластов, электропорация, микроинъекция, липофекция, применение ДНКовых вирусов в качестве носителей ДНК).
15 лет назад Wolff с соавторами продемонстрировали, что инъекция «оголенной» генетической информации в форме плазмидной ДНК (пДНК) или мРНК мышам может приводить к экспрессии белка2. После получения этих результатов были проведены исследования, в которых было установлено, что «оголенную» ДНК можно использовать для вакцинации3-5. Однако применению мРНК для вакцинации уделялось мало внимания вплоть до конца 1990-х годов, когда было продемонстрировано, что перенос мРНК в дендритные клетки запускает иммунные ответы6. Однако непосредственной инъекции «оголенной» мРНК с целью вакцинации уделялось мало внимания, и она была обсуждена только в четырех статьях, опубликованных тремя группами исследователей7-10. Одной из главных причин этого была нестабильность мРНК, обусловленная быстрым расщеплением рибонуклеазами, и связанная с этим недостаточная эффективность мРНК в качестве генетического инструмента in vivo. Между тем из существующего уровня техники известны многочисленные методы стабилизации мРНК, описанные, например, в ЕР-А-1083232, WO 99/14346, US 5580859 и US 6214804.
Однако как нуклеиновая кислота РНК в качестве носителя генетической информации обладает многочисленными преимуществами по сравнению с ДНК, к которым относятся следующие:
(I) РНК, интродуцированная в клетку, не интегрируется в геном (в то время как ДНК в определенной степени интегрируется в геном и тем самым может встраиваться в интактный ген генома клетки-хозяина, в результате чего этот ген может подвергаться мутации и это может приводить к частичной или полной потере генетической информации или появлению ошибочной информации),
(II) для эффективной транскрипции РНК не требуется никаких вирусных последовательностей, таких как промоторы и т.д. (в то время как для экспрессии интродуцированной в клетку ДНК требуется сильный промотор (например, промотор CMV (вирус мозаики цветной капусты)). Интеграция таких промоторов в геном клетки-хозяина может приводить к нежелательным изменениям регуляции экспрессии гена,
(III) расщепление интродуцированной РНК происходит в течение ограниченного периода времени (несколько часов)11,12, поэтому можно достигать кратковременной экспрессии гена, которую можно прерывать после требуемого периода обработки (в то время как это невозможно в случае ДНК, интегрированной в геном),
(IV) РНК не приводит к индукции патогенных антител к РНК в организме пациента (в то время как известно, что индукция антител к ДНК вызывает нежелательный иммунный ответ),
(V) РНК находит широкое применение, поскольку для вакцинации можно очень быстро получать любую требуемую РНК для любого представляющего интерес белка, даже для индивидуального пациента.
В целом, следует отметить, что мРНК представляет собой кратковременную копию кодируемой генетической информации во всех организмах, служит моделью при синтезе белков и в отличие от ДНК обладает всеми предпосылками для получения пригодного вектора для переноса экзогенной генетической информации in vivo.
Наиболее пригодной процедурой для описанного переноса нуклеиновых кислот в организм-хозяин, прежде всего организм млекопитающего, является их инъекция. В то время как ДНК для таких инъекций, как правило, разбавляют в воде, NaCl или ЗФР в качестве инъекционного буфера, РНК, как правило, разбавляют только в инъекционном буфере. В качестве буферов для инъекции РНК применяют стандартные буферы, такие как забуференные фосфатом физиологические растворы, прежде всего ЗФР и забуференный HEPES физиологический раствор (HBS). В случае переноса мРНК такой раствор для инъекции РНК предпочтительно нагревают в течение короткого промежутка времени для нарушения вторичных структур мРНК. Недостатком использования таких стандартных буферов для растворов, предназначенных для инъекции РНК, является очень малая эффективность внутрикожного переноса РНК. Еще одним недостатком является очень низкая скорость трансляции переносимой РНК. Следующим недостатком является то, что при использовании таких стандартных буферов РНК часто образует вторичную структуру (например, так называемую структуру «шпильки»), которая может сильно снижать эффективность проникновения РНК в цитозоль.
Таким образом, целью настоящего изобретения является создание системы, позволяющей, с одной стороны, улучшать внутрикожный перенос РНК в организм-хозяин, а с другой стороны, повышать скорость трансляции перенесенной РНК.
Эта цель достигается с помощью вариантов осуществления изобретения, описанных в формуле изобретения.
Один из вариантов осуществления настоящего изобретения относится к применению РНК и водного инъекционного буфера, содержащего натриевую соль, предпочтительно натриевую соль в концентрации по меньшей мере 50 мМ, кальциевую соль, предпочтительно кальциевую соль в концентрации по меньшей мере 0,01 мМ, и необязательно калиевую соль, предпочтительно калиевую соль в концентрации по меньшей мере 3 мМ, для приготовления раствора для инъекции РНК с целью усиления переноса РНК и/или трансляции РНК в организм(е)-хозяин(е). Следующий объект настоящего изобретения относится также к полученному таким путем инъекционному раствору. Таким образом, инъекционный раствор получают из инъекционного буфера и РНК, которую растворяют в инъекционном буфере.
Согласно предпочтительному варианту осуществления изобретения натриевые соли, кальциевые соли и необязательно калиевые соли, содержащиеся в инъекционном буфере, находятся в форме галогенидов, например хлоридов, йодидов или бромидов, в форме их гидроксидов, карбонатов, бикарбонатов или сульфатов. В качестве примеров следует отметить: для натриевой соли - NaCl, NaI, NaBr, Na2CO3, NaHCO3, Na2SO4; для необязательно присутствующей калиевой соли - KCl, KI, KBr, K2СО3, KНСО3, K2SO4; и для кальциевой соли - СаСl2, CaI2, СаВr2, СаСО3, CaSO4, Са(ОН)2. Инъекционный буфер может содержать также органические анионы указанных выше катионов.
В наиболее предпочтительном варианте осуществления предлагаемого в изобретении применения РНК и инъекционного буфера инъекционный буфер, предлагаемый в изобретении, содержит в качестве солей хлорид натрия (NaCl), хлорид кальция (CaCl2) и необязательно хлорид калия (KCl), причем в дополнение к хлоридам он может содержать также другие анионы. Указанные соли, как правило, присутствуют в инъекционном буфере в концентрации, составляющей по меньшей мере 50 мМ для хлорида натрия (NaCl), по меньшей мере 3 мМ для хлорида калия (KCl) и по меньшей мере 0,01 мМ для хлорида кальция (CaCl2).
Инъекционный буфер, предлагаемый в изобретении, может представлять собой как гипертонический, так и изотонический или гипотонический инъекционный буфер. В контексте настоящего изобретения инъекционный буфер является гипертоническим, изотоническим или гипотоническим по отношению к соответствующей среде, с которой производится сравнение. Это означает, что инъекционный буфер, предлагаемый в изобретении, имеет более высокое, одинаковое или более низкое содержание солей по отношению к соответствующей среде, с которой производится сравнение, применяемые концентрации указанных выше солей предпочтительно должны быть такими, чтобы они не приводили к повреждению клеток, вызываемому осмосом или другими зависящими от концентрации эффектами. Среды, с которыми производится сравнение, представляют собой, например, жидкости, участвующие в процессах in vivo, такие, например, как кровь, лимфатическая жидкость, цитозольные жидкости или другие жидкости, присутствующие в организме, или жидкости или буферы, которые обычно применяют в процессах, осуществляемых in vitro. Такие жидкости и буферы известны специалисту в данной области.
Инъекционный буфер может содержать другие компоненты, например сахара (моно-, ди-, три- или полисахариды), прежде всего глюкозу или маннит. Однако в предпочтительном варианте осуществления изобретения сахара не присутствуют в инъекционном буфере, который предназначен для применения, предлагаемого в изобретении. Предпочтительно также, чтобы буфер, предлагаемый в изобретении, не содержал каких-либо незаряженных компонентов, таких, например, как сахара. Буфер, предлагаемый в изобретении, как правило, содержит только катионы металлов, прежде всего из группы щелочных или щелочно-земельных металлов, и анионы, прежде всего указанные выше анионы.
Значение рН инъекционного буфера, предлагаемого в настоящем изобретении, предпочтительно составляет от 1 до 8,5, предпочтительно от 3 до 5, более предпочтительно от 5,5 до 7,5, наиболее предпочтительно от 5,5 до 6,5. Инъекционный буфер необязательно может содержать также буферную систему, которая фиксирует инъекционный буфер при забуференном значении рН. Такой системой может служить, например, фосфатная буферная система, HEPES или Na2HPO4/NaH2PO4. Однако наибольшее предпочтение следует отдавать инъекционному буферу, применяемому согласно изобретению, который не содержит никаких из указанных выше буферных систем или совсем никакой буферной системы.
Инъекционный буфер, применяемый согласно изобретению, содержит, как указано выше, соли натрия, кальция и необязательно калия, причем натрий и калий, как правило, присутствуют в инъекционном буфере в форме одновалентных катионов (Na+, K+), а кальций присутствует в форме двухвалентного катиона (Са2+). Согласно предпочтительному варианту осуществления изобретения в дополнение к ним или в качестве альтернативы одновалентным и двухвалентным катионам, содержащимся в инъекционном буфере, который предназначен для применения, предлагаемого в изобретении, могут присутствовать двухвалентные катионы металлов, прежде всего из группы щелочно-земельных металлов, таких, например, как магний (Mg2+) или также железо (Fe2+), и одновалентные катионы металлов, прежде всего из группы щелочных металлов, таких, например, как литий (Li+). Такие одновалентные катионы предпочтительно присутствуют в форме их солей, например в форме галогенидов, например хлоридов, йодидов или бромидов, в форме их гидроксидов, карбонатов, бикарбонатов или сульфатов. В качестве примеров следует отметить: для литиевой соли - LiCl, LiI, LiBr, Li2CO3, LiHCO3, Li2SO4; для магниевой соли - MgCl2, MgI2, MgBr2, MgCO3, MgSO4 и Mg(OH)2; и для соли железа - FeCl2, FeBr2, FeI2, FeF2, Fe2O3, FeCO3, FeSO4, Fe(OH)2. Таким образом, можно применять также все комбинации двухвалентных и/или одновалентных катионов, как указано выше. Таким образом, под объем изобретения подпадают инъекционные буферы, которые содержат только двухвалентные, только одновалентные или и одновалентные и двухвалентные катионы. Под объем изобретения подпадают также инъекционные буферы, которые содержат только один тип двухвалентных или одновалентных катионов, наиболее предпочтительно, например, только катионы Са2+ или его соли, например CaСl2.
Предпочтительно следует также учитывать молярности, указанные выше для Са2+ (в качестве двухвалентного катиона) и Na1+ (в качестве одновалентного катиона) (а именно, типичные концентрации, составляющие по меньшей мере 50 мМ для Na+, по меньшей мере 0,01 мМ для Ca2+ и по меньшей мере 3 мМ для необязательно присутствующего K+), в инъекционном буфере, когда вместо некоторых из или всех ионов Ca2+ или Na1+, в инъекционном буфере, который применяют согласно изобретению при приготовлении инъекционного раствора, используют другой двухвалентный или одновалентный катион или другие двухвалентные или одновалентные катионы, прежде всего другие катионы металлов из группы щелочно-земельных металлов или щелочных металлов. Хотя, как указано выше, Са2+ и Na1+ в инъекционном буфере, который предназначен для применения, предлагаемого в изобретении, можно полностью заменять другими двухвалентными или одновалентными катионами, например также на комбинацию других двухвалентных катионов (вместо Са2+) и/или на комбинацию других одновалентных катионов (вместо Na1+) (прежде всего, на комбинацию других двухвалентных катионов из группы щелочно-земельных металлов или на комбинацию других одновалентных металлов из группы щелочных металлов), предпочтительно осуществлять частичную замену Са2+ или Na1+, т.е. это означает, что по меньшей мере 20%, предпочтительно по меньшей 40%, более предпочтительно по меньшей мере 60% и еще более предпочтительно по меньшей мере 80% соответствующих общих молярностей одновалентных или двухвалентных катионов в буфере для инъекций приходится на долю Ca2+ или Na1+. Однако, наиболее предпочтительно, чтобы инъекционный буфер, который предназначен для применения, предлагаемого в изобретении, содержал только Ca2+ в качестве двухвалентного катиона и Na1+ в качестве одновалентного катиона, это означает, что на долю Са2+ приходится 100% общей молярности двухвалентных катионов, а на долю Na1+ приходится 100% общей молярности одновалентных катионов.
Приготовление инъекционного буфера предпочтительно осуществляют при комнатной температуре (25°С) и атмосферном давлении. Приготовление можно осуществлять с помощью любого пригодного процесса, известного из существующего уровня техники. Предпочтительно ионы или содержащие их соли разбавляют в водном растворе, при этом соотношения концентраций следует выбирать в соответствии с конкретными условиями (организм-хозяин, представляющий, прежде всего, млекопитающее, в который требуется инъецировать содержащий РНК раствор для инъекции, состояние здоровья, возраст и т.д. организма-хозяина и условия растворимости и взаимодействия компонентов, температура реакции, время реакции и т.д.).
Концентрации компонентов, представляющих собой ионы натрия, кальция и хлорида и необязательно ионы калия и необязательно лактат (см. приведенное ниже описание варианта осуществления изобретения), содержащиеся в водном инъекционном буфере, зависят, прежде всего, от их растворимости в воде, взаимодействия компонентов друг с другом, а также от температуры реакции и давления в реакционной смеси в процессе приготовления инъекционного буфера или раствора для инъекций РНК.
Основой инъекционного буфера, применяемого согласно настоящему изобретению, является водный раствор, т.е. раствор, состоящий из воды и солей, который применяют согласно изобретению при приготовлении инъекционного раствора, и необязательно лактат. Соли указанных выше одновалентных или двухвалентных катионов необязательно могут быть слаборастворимыми или даже нерастворимыми в таком водном растворе. Степень растворимости солей можно рассчитывать на основе произведения растворимостей. Методы точного определения растворимости и произведения растворимостей известны специалисту в данной области. Такой водный раствор может содержать вплоть до 30 мол.% растворенных солей, предпочтительно вплоть до 25 мол.%, предпочтительно вплоть до 20 мол.%, предпочтительно также вплоть до 15 мол.%, более предпочтительно вплоть до 10 мол.%, еще более предпочтительно вплоть до 5 мол.%, более предпочтительно также вплоть до 2 мол.%, нерастворимых или слаборастворимых солей. Соли, для которых произведение растворимостей составляет <10-4, рассматриваются в контексте настоящего изобретения как слаборастворимые. Соли, для которых произведение растворимостей составляет >10-4, рассматриваются как легкорастворимые.
Растворимость соли, или иона, или содержащего ионы соединения в воде зависит от энергии его кристаллической решетки и энергии гидратации, при этом необходимо принимать во внимание обусловленные энтропией эффекты, которые имеют место. Применяют также понятие «произведение растворимостей», более точно равновесие, которое устанавливается, когда соль, или ион, или содержащее ионы соединение растворяется в воде. Произведение растворимостей обычно определяют как произведение концентраций ионов в насыщенном растворе электролита. Например, соли щелочных металлов (такие, например, как содержащие Na+, K+ соли) растворимы в воде в более высоких концентрациях, чем соли щелочно-земельных металлов (такие, например, как содержащие Ca2+ соли), это означает, что они имеют более высокое значение произведения растворимостей. Это означает, что калиевые и натриевые соли, содержащиеся в водном растворе инъекционного буфера, предлагаемого в изобретении, более легко растворимы, чем присутствующие в нем кальциевые соли. Следовательно, при определении концентрации этих ионов необходимо принимать во внимание, среди прочего, взаимодействие между калиевыми, натриевыми и кальциевыми солями.
Предпочтительным является применение, предлагаемое в изобретении, в котором инъекционный буфер содержит хлорид натрия (NaCl) в концентрации от 50 до 800 мМ, предпочтительно от 60 до 500 мМ, более предпочтительно от 70 до 250 мМ, наиболее предпочтительно от 60 до 110 мМ, хлорид кальция (CaCl2) в концентрации от 0,01 до 100 мМ, предпочтительно от 0,5 до 80 мМ, более предпочтительно от 1,5 до 40 мМ, и необязательно хлорид калия (KCl) в концентрации от 3 до 500 мМ, предпочтительно от 4 до 300 мМ, более предпочтительно от 5 до 200 мМ.
Помимо указанных выше неорганических анионов, например галогенидов, сульфатов или карбонатов, в качестве дополнительных анионов могут присутствовать также органические анионы. Среди них можно отметить сукцинат, лактобионат, лактат, малат, малеонат и т.д., которые могут присутствовать также в виде комбинаций. Инъекционный буфер, предназначенный для применения, предлагаемого в изобретении, предпочтительно содержит лактат, наиболее предпочтительно такой инъекционный буфер, в котором присутствует органический анион, содержит в качестве органического аниона только лактат. В контексте изобретения лактат может представлять собой любой требуемый лактат, например L-лактат и D-лактат. Применительно к настоящему изобретению в качестве солей молочной кислоты (лактатов), как правило, применяют лактат натрия и/или лактат кальция, прежде всего в том случае, когда инъекционный буфер содержит в качестве одновалентного катиона только Na+ и в качестве двухвалентного катиона только Са2+.
В предпочтительном варианте применения, предлагаемого в изобретении, инъекционный буфер, предлагаемый в изобретении, содержит лактат предпочтительно в концентрации от 15 до 500 мМ, более предпочтительно от 15 до 200 мМ и еще более предпочтительно от 15 до 100 мМ.
При создании изобретения было установлено, что применение инъекционного буфера, содержащего указанные выше компоненты, в котором необязательно присутствует или отсутствует лактат (ниже в настоящем описании обозначен как «инъекционный буфер RL», когда компонент, представляющий собой лактат, отсутствует, или как «инъекционный буфер RL с лактатом», когда присутствует компонент, представляющий собой лактат), для приготовления растворов для инъекции РНК (т.е. инъекционных растворов, которые содержат РНК и пригодны для инъекции указанной РНК) существенно усиливает как перенос, так и трансляцию РНК в клетки(ах)/ткани(ях) организма-хозяина (млекопитающего) по сравнению с инъекционными буферами, обычно применяемыми согласно существующему уровню техники.
Раствор, содержащий указанные выше компоненты, представляющие собой хлорид натрия (NaCl), хлорид кальция (CaCl2), лактат, прежде всего в форме лактата натрия, и необязательно также хлорид калия (KCl), обозначают также как «раствор Рингера» или «лактат Рингера». Лактат Рингера представляет собой кристалловидный полный электролитный раствор, который применяют в качестве раствора для возмещения объема и раствора-носителя, например, для совместимых с ним лекарственных средств. Например, лактат Рингера применяют в качестве основного агента для возмещения объема в случае потери жидкости и электролита (в результате рвоты, диареи, непроходимости кишечника или ожогов), прежде всего у младенцев и маленьких детей, и для поддержания открытого доступа к периферическим и/или центральным венам. Однако предлагаемое в изобретении применение лактата Рингера в качестве инъекционного буфера в растворах для инъекции РНК неизвестно из существующего уровня техники.
В контексте настоящего изобретения РНК представляет собой любую требуемую РНК, например мРНК, тРНК, рРНК, siRNA, одно- или двухцепочечную РНК, гетеродуплексную РНК и т.д. В качестве РНК можно применять РНК, кодирующую любой представляющий интерес белок. РНК, которую используют для применения, предлагаемого в изобретении, предпочтительно представляет собой «оголенную» РНК. Особенно предпочтительно она представляет собой мРНК, более предпочтительно «оголенную» мРНК.
В контексте изобретения подразумевается, что «оголенная» РНК, прежде всего «оголенная» мРНК, представляет собой РНК, которая не находится в составе комплекса, например, с поликатионными молекулами. «Оголенная» РНК может находиться в одноцепочечной форме, а также в двухцепочечной форме, т.е. в виде вторичной структуры, например в виде так называемой структуры «шпильки». Такие двухцепочечные формы встречаются, прежде всего, у «оголенных» РНК, прежде всего «оголенных» мРНК, когда в молекуле присутствуют комплементарные рибонуклеотидные последовательности.
Однако согласно изобретению РНК, прежде всего мРНК, могут присутствовать также в составе комплекса. В результате такой комплексации/конденсации РНК, прежде всего мРНК, предлагаемой в изобретении, эффективный перенос РНК в предназначенные для обработки клетки или предназначенную для обработки ткань организма, предназначенного для обработки, может быть усилен благодаря ассоциации или связыванию РНК с (поли)катионным полимером, пептидом или белком. Такую РНК (мРНК) предпочтительно комплексируют или конденсируют по меньшей мере с одним катионным или поликатионным агентом. Такой катионный или поликатионный агент предпочтительно представляет собой агент, выбранный из группы, включающей протамин, поли-L-лизин, поли-L-аргинин, нуклеолин, спермин и гистоны или производные гистонов или протаминов. В качестве поликатионного связывающегося с нуклеиновой кислотой белка наиболее предпочтительно применяют протамин. Такая процедура стабилизации РНК описана, например, в ЕР-А-1083232, соответствующее содержание которой во всей его полноте включено в настоящее изобретение.
РНК, предлагаемую в изобретении, можно подвергать дополнительным модификациям. Эти модификации служат, прежде всего, для повышения стабильности РНК. Предпочтительно РНК имеет одну или несколько (встречающихся в естественных условиях или не встречающихся в естественных условиях) модификаций, прежде всего химических модификаций, которые, например, способствуют увеличению времени полужизни РНК в организме или улучшают эффективность трансляции мРНК в цитозоле по сравнению с эффективностью трансляции немодифицированной мРНК в цитозоле. Предпочтительно эффективность трансляции улучшают с помощью модификации, предлагаемой в изобретении, по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, также предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 60%, также более предпочтительно по меньшей мере на 75%, наиболее предпочтительно по меньшей мере на 85%, наиболее предпочтительно по меньшей мере на 100%, по сравнению с эффективностью трансляции немодифицированной мРНК в цитозоле.
Например, можно увеличивать содержание G/C в кодирующей области модифицированной мРНК по сравнению с содержанием G/C в кодирующей области соответствующей мРНК дикого типа, при этом аминокислотная последовательность, кодируемая модифицированной мРНК, предпочтительно остается неизмененной по сравнению с аминокислотной последовательностью, кодируемой мРНК дикого типа. Такая модификация основана на том факте, что последовательность области мРНК, предназначенной для трансляции, важна для эффективной трансляции мРНК. В этом случае имеет важное значение состав и последовательность расположения различных нуклеотидов. В частности, последовательности, которые имеют высокое содержание G (гуанозин)/С (цитозин), являются более стабильными, чем последовательности, имеющие высокое содержание А (аденозин)/U (урацил). Поэтому целесообразно при сохранении транслируемой аминокислотной последовательности изменять кодоны по сравнению с мРНК дикого типа таким образом, чтобы они имели более высокое содержание G/C-нуклеотидов. Благодаря тому факту, что несколько кодонов кодируют одну и ту же аминокислоту (так называемая «вырожденность генетического кода»), оказывается возможным определять кодоны, которые полезны с точки зрения стабильности, предпочтительно имеющие максимальное содержание G/C. В результате этого РНК в буфере для инъекций предпочтительно имеет содержание G/C, увеличенное предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 70%, более предпочтительно на 80%, в пересчете на максимальное содержание G/C (т.е. содержание G/C после модификации всех потенциальных триплетов в кодирующей области последовательности, встречающейся в естественных условиях, с использованием вырожденности генетического кода для максимизации содержания G/C без изменения кодируемой аминокислотной последовательности), и наиболее предпочтительно максимальное содержание G/C, где максимальное содержание G/C присутствует в последовательности, в которой содержание G/C максимизировано без изменения при этом кодируемой аминокислотной последовательности.
В зависимости от аминокислотной последовательности, которая должна кодироваться модифицированной мРНК, существуют различные возможности для модификации последовательности мРНК по сравнению с последовательностью дикого типа. В том случае когда аминокислоты кодируются кодонами, которые содержат только нуклеотиды G или С, не требуется осуществлять модификацию кодонов. Примерами таких кодонов являются для Pro (ССС или CCG), Arg (CGC или CGG), Ala (GCC или GCG) и Gly (GGC или GGG).
С другой стороны, кодоны, которые содержат нуклеотиды А и/или U, можно изменять путем замены на различные кодоны, которые кодируют ту же самую аминокислоту, но которые не содержат А и/или U. Такими примерами являются:
- кодоны для Pro можно изменять с CCU или ССА на ССС или CCG;
- кодоны для Arg можно изменять с CGU, или CGA, или AGA, или AGG на CGC или CGG;
- кодоны для Аlа можно изменять с GCU или GCA на GCC или GCG;
- кодоны для Gly можно изменять с GGU или GGA на GGC или GGG.
В определенных случаях, хотя и нельзя исключать из кодонов нуклеотиды А и U, оказывается возможным уменьшать содержание А и U путем использования кодонов, которые имеют меньшее содержание нуклеотидов А и/или U. Такими примерами являются:
- кодоны для Phe можно изменять с UUU на UUC;
- кодоны для Leu можно изменять с UUA, UUG, CUU или CUA на CUC или CUG;
- кодоны для Ser можно изменять с UCU, или UCA, или AGU на UCC, UCG или AGC;
- кодон для Тyr можно изменять с UAU на UAC;
- кодон для Cys можно изменять с UGU на UGC;
- кодон для His можно изменять с CAU на САС;
- кодон для Gln можно изменять с САА на CAG;
- кодоны для Ilе можно изменять с AUU или AUA на AUC;
- кодоны для Thr можно изменять с ACU или АСА на АСС или ACG;
- кодон для Asn можно изменять с AAU на ААС;
- кодон для Lys можно изменять с ААА на AAG;
- кодон для Val можно изменять с GUU или GUA на GUC или GUG;
- кодон для Asp можно изменять с GAU на GAC;
- кодон для Glu можно изменять с GAA на GAG,
- стоп-кодон UAA можно изменять на UAG или UGA.
Перечисленные выше замены можно применять индивидуально или во всех возможных комбинациях для повышения содержания G/C в модифицированной мРНК по сравнению с мРНК дикого типа (исходная последовательность). Предпочтительно, например, использовать следующие комбинации возможных замен:
- замену всех кодонов, кодирующих Thr в исходной последовательности (мРНК дикого типа), на АСС (или ACG) и замену всех кодонов, кодирующих Ser в исходной последовательности, на UCC (или UCG, или AGC);
- замену всех кодонов, кодирующих Ilе в исходной последовательности, на AUC, и замену всех кодонов, кодирующих Lys в исходной последовательности, на AAG, и замену всех кодонов, кодирующих Тyr в исходной последовательности, на UAC;
- замену всех кодонов, кодирующих Val в исходной последовательности, на GUC (или GUG), и замену всех кодонов, кодирующих Glu в исходной последовательности, на GAG, и замену всех кодонов, кодирующих Аlа в исходной последовательности, на GCC (или GCG), и замену всех кодонов, кодирующих Arg в исходной последовательности, на CGC (или CGG);
- замену всех кодонов, кодирующих Val в исходной последовательности, на GUC (или GUG), и замену всех кодонов, кодирующих Glu в исходной последовательности, на GAG, и замену всех кодонов, кодирующих Аlа в исходной последовательности, на GCC (или GCG), и замену всех кодонов, кодирующих Gly в исходной последовательности, на GGC (или GGG), и замену всех кодонов, кодирующих Asn в исходной последовательности, на ААС;
- замену всех кодонов, кодирующих Val в исходной последовательности, на GUC (или GUG), и замену всех кодонов, кодирующих Phe в исходной последовательности, на UUC, и замену всех кодонов, кодирующих Cys в исходной последовательности, на UGC, и замену всех кодонов, кодирующих Leu в исходной последовательности, на CUG (или CUC), замену всех кодонов, кодирующих Gln в исходной последовательности, на CAG, и замену всех кодонов, кодирующих Pro в исходной последовательности, на ССС (или CCG), и т.д.
В том случае когда изменение содержания G/C осуществляют в области модифицированной мРНК, которая кодирует белок, то это содержание следует увеличивать по меньшей мере на 7%, более предпочтительно по меньшей мере на 15%, также более предпочтительно по меньшей мере на 20%, еще более предпочтительно по меньшей мере на 30% по сравнению с содержанием G/C области мРНК дикого типа, кодирующей белок. В этом случае наиболее предпочтительно увеличивать содержание G/C в модифицированной мРНК, прежде всего в области, кодирующей белок, до максимального уровня по сравнению с последовательностью дикого типа.
Кроме того, предпочтительно увеличивать содержание A/U в области сайта связывания рибосомы в модифицированной мРНК по сравнению с содержанием A/U в области сайта связывания рибосомы в соответствующей мРНК дикого типа. Эта модификация повышает эффективность связывания рибосом с мРНК. В свою очередь, эффективное связывание рибосом с сайтом связывания рибосомы (последовательность Козака (Kozak): GCCGCCACCAUGG, AUG представляет собой стартовый кодон) позволяет осуществлять эффективную трансляцию мРНК. Увеличение заключается в том, что интродуцируют по меньшей мере одну дополнительную единицу A/U, как правило, по меньшей мере 3 единицы A/U, в область сайта связывания, т.е. область, простирающуюся от положения -20 до положения +20 относительно А стартового кодона AUG.
Модификация, которая также является предпочтительной, относится к мРНК, в которой кодирующая область и/или 5'- и/или 3'-нетранслируемая область модифицированной мРНК изменена по сравнению с мРНК дикого типа таким образом, что она не содержит никаких элементов, дестабилизирующих последовательность, при этом кодируемая аминокислотная последовательность модифицированной РНК предпочтительно не имеет изменений по сравнению с мРНК дикого типа. Известно, что элементы, дестабилизирующие последовательность (DSE), встречаются, например, в последовательностях мРНК эукариотических организмов, с которыми связываются сигнальные белки, представляющие собой элементы, дестабилизирующие последовательность, и осуществляют регуляцию ферментативного расщепления мРНК in vivo. Таким образом, для дополнительной стабилизации модифицированной мРНК, предлагаемой в изобретении, необязательно можно осуществлять одно или несколько таких изменений в области, кодирующей белок, по сравнению с соответствующей областью мРНК дикого типа, в результате которых она не будет совсем иметь или практически не будет иметь никаких элементов, дестабилизирующих последовательность. Путем таких изменений можно также согласно изобретению исключать из мРНК DSE, присутствующие в нетранслируемых областях (3'- и/или 5'-UTR).
Такими дестабилизирующими последовательностями являются, например, последовательности, богатые AU («AURES»), которые встречаются в 3'-UTR-участках многочисленных нестабильных мРНК (Caput и др., Proc. Natl. Acad. Sci. USA, 83, 1986, c.1670-1674), а также мотивы последовательности, которые распознаются эндонуклеазами (см., например. Binder и др., EMBO J., 13, 1994, c.1969-1980).
Предпочтительной является также модифицированная мРНК, которая имеет стабилизирующую структуру, представляющую собой кэп, расположенный с 5'-конца (5'-кэп-структуру). Примерами кэп-структур, которые можно применять согласно изобретению, являются m7G(5')ppp, 5'(А,G(5')ррр(5')А и G(5')ppp(5')G.
Предпочтительно также, чтобы модифицированная мРНК имела хвост поли-(А), предпочтительно состоящий по меньшей мере из 25 нуклеотидов, более предпочтительно по меньшей мере из 50 нуклеотидов, еще более предпочтительно по меньшей мере из 70 нуклеотидов, также более предпочтительно по меньшей мере из 100 нуклеотидов, наиболее предпочтительно по меньшей мере из 200 нуклеотидов.
Предпочтительно также, чтобы модифицированная мРНК имела по меньшей мере один IRES и/или по меньшей мере одну 5'- и/или 3'-стабилизирующую последовательность. Согласно изобретению в модифицированную мРНК можно интродуцировать один или несколько так называемых IRES («внутренний рибосомальный сайт проникновения»). Таким образом, IRES может функционировать в качестве единственного сайта связывания рибосомы, но может также служить для того, чтобы получать мРНК, кодирующую несколько белков, пептидов или полипептидов, которые должны транслироваться рибосомами независимо друг от друга («мультицистронная мРНК»). Примерами последовательностей IRES, которые можно применять согласно изобретению, являются последовательности IRES пикорнавирусов (например, вирус ящура (FMDV)), вирусов чумы (CFFV), вирусов полиомы (PV), вирусов энцефаломиокардита (ECMV), вирусов ящура (FMDV), вирусов гепатита С (HCV), вирусов классической лихорадки свиней (CSFV), вируса мышиного лейкоза (MLV), вируса иммунодефицита обезьян (SIV) или вирусов паралича сверчков (CrPV).
Предпочтительно также, чтобы модифицированная мРНК имела по меньшей мере одну 5'- и/или 3'-стабилизирующую последовательность. Указанные стабилизирующие последовательности, локализованные в 5'- и/или 3'-нетранслируемых областях, приводят к увеличению времени полужизни мРНК в цитозоле. Такие стабилизирующие последовательности могут быть на 100% гомологичны встречающимся в естественных условиях последовательностям, которые присутствуют в вирусах, бактериях и эукариотических организмах, но могут быть также частично или полностью синтетическими. В качестве примера стабилизирующих последовательностей, которые можно применять в настоящем изобретении, можно отметить нетранслируемые последовательности (UTR) гена глобина, например, Homo sapiens или Xenopus laevis. Другим примером стабилизирующей последовательности является последовательность общей формулы (C/U)CCANxCCC(U/A)PyxUC(C/U)CC, которая содержится в 3'-UTR обладающей очень большой стабильностью мРНК, кодирующей α-глобин, (I)-коллаген, 15-липоксигеназу или тирозингидроксилазу (см. Holcik и др., Рrос. Natl. Acad. Sci. USA, 94, 1997, c.2410-2414). Естественно, такие стабилизирующие последовательности можно применять индивидуально, или в сочетании друг с другом, или также в сочетании с другими стабилизирующими последовательностями, известными специалисту в данной области.
В предпочтительном варианте осуществления настоящего изобретения модифицированная мРНК содержит по меньшей мере один аналог встречающихся в естественных условиях нуклеотидов. Этот/эти аналог/аналоги служит/служат для дополнительной стабилизации модифицированной мРНК, что основано на том факте, что расщепляющие РНК ферменты, встречающиеся в клетках, распознают в качестве субстрата преимущественно встречающиеся в естественных условиях нуклеотиды. Таким образом, интродукция нуклеотидных аналогов в РНК затрудняет расщепление РНК, однако интродукция этих аналогов, прежде всего, в кодирующую область мРНК может оказывать как позитивное, так и негативное влияние на эффективность трансляции. В качестве примеров нуклеотидных аналогов, которые можно применять согласно изобретению, следует отметить (но никоим образом не ограничиваясь ими) фосфорамидаты, фосфоротиоаты, пептидные нуклеотиды, метилфосфонаты, 7-деазагуанозин, 5-метилцитозин и инозин. Методы получения таких аналогов известны специалисту в данной области, например описаны в патентах US 4373071, US 4401796, US 4415732, US 4458066, US 4500707, US 4668777, US 4973679, US 5047524, US 5132418, US 5153319, US 5262530 и US 5700642. Такие аналоги могут встречаться как в нетранслируемых, так и в транслируемых областях модифицированной мРНК.
Еще в одном предпочтительном варианте осуществления настоящего изобретения модифицированная мРНК дополнительно содержит последовательность, кодирующую сигнальный пептид. Эта последовательность, кодирующая сигнальный пептид, предпочтительно имеет длину от 30 до 300 оснований и кодирует от 10 до 100 аминокислот. Более предпочтительно последовательность, кодирующая сигнальный пептид, имеет длину от 45 до 180 оснований и кодирует от 15 до 60 аминокислот. Для модификации РНК, применяемой согласно изобретению, можно использовать, например, последовательности, перечисленные ниже в таблице 1. Можно применять также последовательности, полученные из указанных в таблице 1, в которых от 1 до 20, предпочтительно от 1 до 10 и наиболее предпочтительно от 1 до 5 оснований заменены на А, Т, С или G по сравнению с последовательностями, указанными в таблице 1.
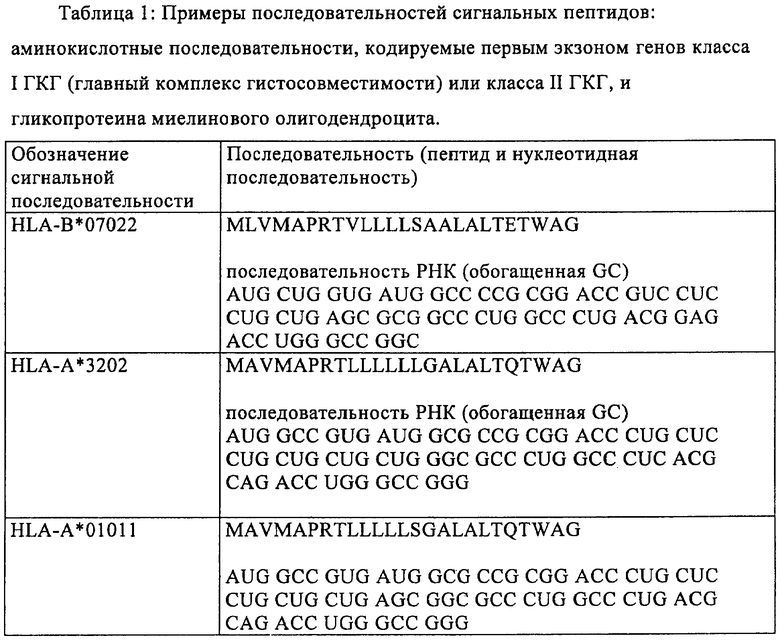
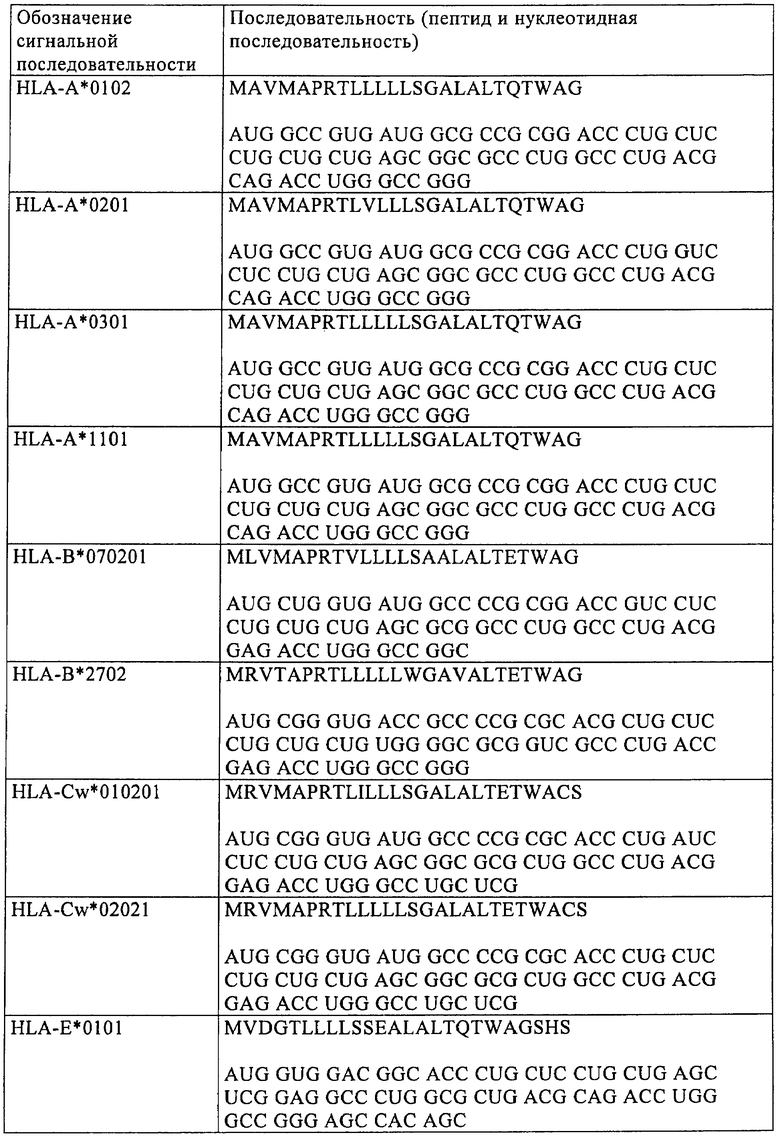
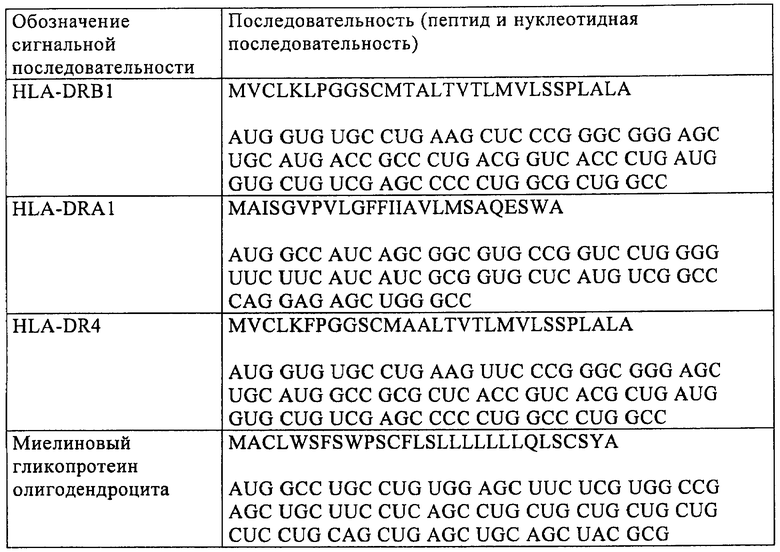
Для осуществления указанных выше модификаций можно применять различные процессы, известные специалисту в данной области. Например, для замены кодонов в модифицированной мРНК, предлагаемой в изобретении, можно в случае относительно коротких кодирующих областей синтезировать полную мРНК химическим путем с помощью стандартных методов. Однако замены, добавления или удаления оснований предпочтительно осуществляют с использованием ДНК-матрицы для получения модифицированной мРНК с помощью обычных методов мишень-направленного мутагенеза (см., например, Maniatis и др., Molecular Cloning: A Laboratory Manual, 3-е изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 2001). При таком процессе для получения мРНК осуществляют транскрипцию in vitro соответствующей молекулы ДНК. Эта ДНК-матрица имеет пригодный промотор, например промотор Т7 или SP6, для осуществления транскрипции in vitro, после которого расположена требуемая предназначенная для получения нуклеотидная последовательность мРНК и сигнал терминации для транскрипции in vitro. Молекулу ДНК, представляющую собой матрицу для конструкции РНК, которую требуется получить, можно создавать путем размножения в ферментативной среде и последующего выделения в виде фрагмента плазмиды, которая может реплицироваться в бактериях. Так, требуемую нуклеотидную последовательность можно клонировать в пригодной плазмиде с помощью методов молекулярной биологии, известных специалисту в данной области, с использованием коротких синтетических ДНК-олигонуклеотидов, которые имеют короткие одноцепочечные транзиции в образовавшихся сайтах расщепления, или с использованием генов, полученных путем химического синтеза (см. Maniatis и др., выше). После этого молекулу ДНК вырезают с помощью расщепления рестриктазами из плазмиды, в которой она может присутствовать в виде одной копии или в виде нескольких копий.
Под объем изобретения подпадают описанные выше модификации РНК, прежде всего мРНК, которые могут присутствовать как индивидуально, так и в комбинации друг с другом. Аналогичным образом можно объединять одну или несколько модификацию(й) с описанным выше включением в состав комплекса РНК, прежде всего мРНК.
Цель изобретения заключается в увеличении переноса РНК и/или трансляции РНК в организм(е)-хозяина(е). В контексте изобретения организм-хозяин представляет собой любой организм, в клетки или ткань которого можно переносить РНК, а затем осуществлять ее трансляцию. Организм-хозяин, подпадающий под объем изобретения, представляет собой, прежде всего, млекопитающее, выбранное из группы, включающей мышь, крысу, свинью, корову, лошадь, собаку, кошку, обезьяну и, прежде всего, человека.
При создании настоящего изобретения было установлено, что РНК, кодирующая люциферазу, прежде всего мРНК, разбавленная в инъекционном буфере RL, предлагаемом в изобретении (содержащем или не содержащем лактат), обеспечивает существенно более высокую скорость трансляции по сравнению с мРНК, которую разбавляют в стандартных буферах, обычно применяемых для РНК, таких как HBS или ЗФР (см. фиг.1). Кроме того, установлено, что эффективность переноса и трансляции инъецируемой мРНК в большой степени зависит от присутствия ионов кальция. В соответствующих сравнительных опытах с использованием инъекционного буфера RL (содержащего или не содержащего лактат), в котором присутствуют или не присутствуют ионы кальция, было установлено, что при отсутствии ионов кальция эффективность переноса РНК существенно снижается до уровня, сопоставимого с уровнем, достигаемым при использовании стандартных буферов ЗФР и HBS (см. фиг.2).
Таким образом, было установлено, что, во-первых, инъекционный буфер RL, предлагаемый в изобретении (содержащий или не содержащий лактат), существенно повышает перенос РНК и, во-вторых, этот увеличенный перенос РНК еще более возрастает при использовании инъекционного буфера RL, предлагаемого в изобретении (содержащего или не содержащего лактат), который имеет высокую концентрацию ионов кальция, составляющую вплоть до 100 мМ.
Инъекционный буфер, предлагаемый в изобретении, предпочтительно применяют в комбинации с РНК в растворе для инъекции РНК. Таким образом, в изобретении предложен также раствор для инъекции РНК, содержащий РНК и инъекционный буфер, который содержит хлорид натрия (NaCl) в концентрации по меньшей мере 50 мМ, хлорид кальция (CaCl2) в концентрации по меньшей мере 0,01 мМ и необязательно хлорид калия (KCl) в концентрации по меньшей мере 3 мМ, для увеличения переноса РНК и/или трансляции РНК в клетки(ах). Предпочтительно применяют раствор для инъекций РНК, предлагаемый в изобретении, в котором инъекционный буфер содержит хлорид натрия (NaCl) в концентрации по меньшей мере от 50 до 800 мМ, предпочтительно по меньшей мере от 60 до 500 мМ, более предпочтительно по меньшей мере от 70 до 250 мМ, наиболее предпочтительно от 60 до 110 мМ, хлорид кальция (CaCl2) в концентрации по меньшей мере от 0,01 до 100 мМ, предпочтительно по меньшей мере от 0,5 до 80 мМ, более предпочтительно по меньшей мере от 1,5 до 40 мМ и необязательно хлорид калия (KCl) в концентрации по меньшей мере от 3 до 500 мМ, предпочтительно по меньшей мере от 4 до 300 мМ, более предпочтительно по меньшей мере от 5 до 200 мМ.
Инъекционный буфер, входящий в раствор для инъекции РНК, предлагаемый в изобретении, предпочтительно дополнительно содержит лактат. Такой инъекционный буфер, входящий в раствор для инъекции РНК, предлагаемый в изобретении, предпочтительно содержит лактат в концентрации по меньшей мере 15 мМ. Кроме того, предпочтительным является также раствор для инъекций РНК, предлагаемый в изобретении, в котором инъекционный буфер содержит лактат в концентрации от 15 до 500 мМ, предпочтительно от 15 до 200 мМ, более предпочтительно от 15 до 100 мМ.
Раствор для инъекции РНК можно приготавливать с помощью любого требуемого метода, известного из существующего уровня техники. Предпочтительно РНК разбавляют в инъекционном буфере RL или в инъекционном буфере RL с лактатом. Можно применять также РНК в форме безводной РНК (например, полученной с помощью сушки вымораживанием) и к ней можно добавлять инъекционный буфер RL или инъекционный буфер RL с лактатом, необязательно при повышенной температуре, перемешивании, облучении ультразвуком и т.д. для ускорения растворения. Соотношения концентраций следует выбирать с учетом конкретных условий (организм-хозяин, прежде всего млекопитающее, которому осуществляют инъекцию РНК, состояние здоровья, возраст и т.д. организма-хозяина и т.д.).
РНК в растворе для инъекции РНК, предлагаемом в изобретении, предпочтительно представляет собой «оголенную» РНК, более предпочтительно мРНК, предпочтительно «оголенную» мРНК, как это уже указано выше.
Как указано, раствор для инъекции РНК, предлагаемый в изобретении, можно применять прежде всего для увеличения переноса РНК и трансляции РНК в организм(е)-хозяин(е).
Таким образом, в настоящем изобретении предложено также применение указанного выше раствора для инъекции РНК для увеличения переноса РНК и трансляции РНК в организм(е)-хозяин(е).
При создании изобретения были также проведены исследования дозы (прежде всего, с точки зрения количества и продолжительности действия при клинических применениях) РНК, предназначенной для переноса в инъекционном буфере RL (с лактатом или без него). Исследования позволили выявить повышение экспрессии люциферазы при увеличении количеств мРНК вплоть до 0,1 мкг (в объеме 100 мкл инъецируемого раствора) для мышей и вплоть до 3 мг (в объеме 150 мкл инъецируемого раствора) для людей. Трансляция мРНК происходит кратковременно, и поэтому для осуществления продолжительной равномерной экспрессии чужеродной молекулы (белка) в зависимости от различных факторов, таких как конкретная подлежащая экспрессии чужеродная молекула и требуемое действие, организм, которому осуществляют инъекцию, а также его состояние (здоровья), ее следует регулировать путем повторных инъекций примерно через каждые три дня, или даже каждые два дня, или ежедневно. Количество РНК, также в зависимости от различных факторов, среди прочего, от указанных выше факторов, может составлять от 0,01 до 1000 мкг, предпочтительно от 1 до 800 мкг, также предпочтительно от 2 до 500 мкг, более предпочтительно от 5 до 100 мкг, еще более предпочтительно от 10 до 90 мкг, наиболее предпочтительно от 20 до 80 мкг, в объеме 100 мкл инъецируемого раствора. Наиболее предпочтительно количество РНК составляет 60 мкг в объеме 100 мкл инъецируемого раствора.
Таким образом, вариантами применения, предлагаемыми в изобретении, как РНК и инъекционного буфера RL или инъекционного буфера RL с лактатом, так и раствора для инъекций РНК, предлагаемого в настоящем изобретении, являются, например, применение для лечения и/или профилактики или для приготовления лекарственного средства, предназначенного для лечения и/или профилактики раковых или опухолевых заболеваний, например меланомы, такой как злокачественная меланома, кожная меланома, карциномы, такой как карцинома ободочной кишки, карцинома легкого, такая как мелкоклеточная карцинома легкого, аденокарцинома, карцинома предстательной железы, карцинома пищевода, карцинома молочной железы, карцинома почек, саркомы, миеломы, лейкоза, прежде всего AML (острый миелоидный лейкоз), глиомы, лимфом и бластом, аллергий, аутоиммунных заболеваний, таких как рассеянный склероз, вирусных и/или бактериальных инфекций.
Например, под объем настоящего изобретения подпадает применение как РНК и инъекционного буфера RL или инъекционного буфера RL с лактатом, так и раствора для инъекций РНК, предлагаемого в настоящем изобретении, среди прочего, для генной терапии и для вакцинации, например для противовирусной или противоопухолевой вакцинации с целью предупреждения указанных выше заболеваний.
В контексте настоящего изобретения понятие «генная терапия» обозначает, прежде всего, восстановление недостающей функции организма или клетки путем интродукции функционирующего гена в заболевшие клетки или ингибирование нарушенной функции с помощью соответствующей генетической информации. Например, в случае гена-супрессора опухоли, например р53, если он отсутствует или экспрессируется лишь в небольших количествах, его можно интродуцировать в клетку в форме его мРНК и встраивать в ДНК, в результате чего белок, экспрессируемый первоначально в недостаточных количествах, может снова продуцироваться в имеющих физиологическое значение количествах. Примерами генов-супрессоров опухолей в контексте настоящего изобретения являются р53 ТР53, RB1, АРС, WT1, NF1, NF2, VHL, BRCA1, BRCA2, DCC, MEN 1, MEN 2, PTCH, p57/KIP2, MSH2, MLH1, FMS1, FMS2, MET, p16/INK4a/CDKN2, CDK4, RET, EXT1, EXT2, EXT3, PTEN/MMAC1, ATM, BLM, XPB, XPD, XPA, XPG, FACC, FACA, SMAD4/DPC4, p14Art(p19Art), DPC4, E-CAD, LKB1/STK1, TSC2, PMS1, PMS2, MSH6, TGF-β типа II R, BAX, α-CAT, MADR2/SMAD2, CDX2, MKK4, PP2R1B, MCC и т.д.
В контексте изобретения понятие «вакцинация» обозначает введение генетической информации в форме РНК, прежде всего мРНК, в организм, прежде всего, в одну/несколько клеток или ткань организма. Введенная таким путем мРНК транслируется в организме в молекулу-мишень (например, пептид, полипептид, белок), так что молекула-мишень, кодируемая мРНК, экспрессируется и стимулирует иммунный ответ. Известно, что антигенпрезентирующие клетки (АПК) играют необходимую имеющую решающее значение роль в процессе стимуляции иммунного ответа, поскольку они представляют собой единственный тип клеток, в которых после активации происходит стимуляция всех сигналов, необходимых для инициации пролиферации антигенспецифических иммунных клеток. В объеме настоящего изобретения вакцинацию можно осуществлять, например, с использованием РНК, прежде всего мРНК, кодирующей антиген, где антиген представляет собой опухолевый антиген в случае противоопухолевой вакцинации или чужеродный антиген в случае вакцинации против чужеродных патогенов. Примерами опухолевых антигенов, предлагаемых в настоящем изобретении, являются Т-клеточные опухолевые антигены, такие, например, как антигены «рака/яичка», например MAGE, RAGE, NY-ESO-1, дифференцировочные антигены, например MART-1/Melan-A, тирозиназа, gp100, PSA, CD20, антигенные эпитопы мутантных генов, например: CDK4, каспаза-8, или онкоэмбриональные антигены, например СЕА, AF. Другими опухолевыми антигенами являются, например, опухолевые антигены CD5 и CAMPATH-1(CDw52), которые присутствуют в Т-клеточных и В-клеточных лимфомах, CD20, который присутствует в неходжскинских В-клеточных лимфомах, опухолевые антигены СЕА (онкоэмбриональный антиген), муцин, СА-125 и FAP-a, который присутствует в плотных опухолях, прежде всего, в эпителиальных опухолях (молочной железы, кишечника и легкого), тенасцин и металлопротеиназы, которые присутствуют, кроме того, в глиобластомных опухолях. Другими опухолевыми антигенами являются, например, опухолевые антигены EGF (эпидермальный фактор роста), p185HER2 и рецептор IL-2, который присутствует в опухолях легкого, молочной железы, головы и шеи, а также в Т- и В-клеточных опухолях, например опухолевый антиген SV40, и т.д.
Можно применять РНК, прежде всего мРНК, которая кодирует несколько таких антигенов. В результате этого можно эффективно контролировать меланому, карциному, AML или глиому, поскольку комбинация различных антигенов, специфических в отношении конкретной опухоли, обладает очень широким спектром действия. Кроме того, РНК, прежде всего мРНК, предлагаемая в изобретении, может кодировать иммуногенный белок. Такой иммуногенный белок может опосредовать реактивацию иммунного ответа. Такая реактивация основана на открытии того факта, что почти каждый организм обладает так называемым «вторичным иммунным ответом» на определенные чужеродные молекулы, например белки, прежде всего, вирусные белки, антигены. Это означает, что организм уже был инфицирован ранее такой чужеродной молекулой и что иммунный ответ на такую чужеродную молекулу, например вирусный белок, уже был стимулирован этой инфекцией, и этот ответ сохраняется в «памяти», т.е. запасается. Когда организм снова инфицируется этой же самой чужеродной молекулой, то иммунный ответ реактивируется. В соответствии с изобретением такую реактивацию иммунного ответа можно осуществлять путем вакцинации с помощью РНК, прежде всего мРНК, которая содержит по меньшей мере одну область, кодирующую по меньшей мере один иммуногенный белок. Предпочтительной является РНК, прежде всего мРНК, кодирующая как один или более антигенов, так и один или более иммуногенных белков.
В контексте изобретения иммуногенные белки предпочтительно представляют собой структурные белки вирусов, прежде всего матричные белки, капсидные белки и поверхностные белки липидной мембраны. Другими примерами таких вирусных белков являются белки аденовирусов, риновирусов, коронавирусов, ретровирусов. Наиболее предпочтительными в данном случае являются белки поверхностного антигена вируса гепатита В (ниже обозначен как «HBS-антиген») и матричные белки вируса гриппа, прежде всего матричный белок M1 вируса гриппа.
Настоящее изобретение относится также к применению РНК и вышеописанного инъекционного буфера RL или инъекционного буфера RL с лактатом или вышеописанного раствора для инъекции РНК, для увеличения переноса РНК и/или трансляции РНК в процессах in vitro, например, для анализов экспрессии генов или процессов скрининга in vitro, например, с помощью HTS (высокопроизводительный скрининг).
Кроме того, настоящее изобретение относится к способу увеличения переноса РНК и/или трансляции РНК в организм(е)-хозяин(е), например, для лечения и/или профилактики раковых или опухолевых заболеваний, например меланомы, такой как злокачественная меланома, кожная меланома, карциномы, такой как карцинома ободочной кишки, карциномы легкого, такой как мелкоклеточная карцинома легкого, аденокарциномы, карциномы предстательной железы, карциномы пищевода, карциномы молочной железы, карциномы почек, саркомы, миеломы, лейкоза, прежде всего AML (острый миелоидный лейкоз), глиомы, лимфом и бластом, аллергий, аутоиммунных заболеваний, таких как рассеянный склероз, вирусных и/или бактериальных инфекций, и для генной терапии и/или вакцинации, необязательно для антивирусной вакцинации, с целью предупреждения вышеуказанных заболеваний, который заключается в том, что:
а) приготавливают раствор для инъекции РНК, предлагаемый в настоящем изобретении, и
б) вводят раствор для инъекции РНК, полученный на стадии а), в организм-хозяин.
Приготовление раствора для инъекции РНК на стадии а) можно осуществлять как описано выше, т.е. с помощью любого процесса, известного из существующего уровня техники, предпочтительно путем разбавления РНК в инъекционном буфере RL или инъекционном буфере RL с лактатом. В этом случае также соотношения концентраций следует выбирать в зависимости от вышеуказанных условий (например, организма-хозяина, в частности млекопитающего, которому инъецируют раствор для инъекции РНК, состояния здоровья, возраста и т.д. организма-хозяина и т.д.). Раствор для инъекции РНК можно вводить, например, с помощью инъекции с использованием шприца (например, типа Sub-Q, фирма Becton Dickinson, Гейдельберг, Германия) любым пригодным путем, например внутрикожно, внутриэпителиально, подкожно, внутривенно, внутрисосудисто, внутриартериально, интраабдоминально, внутрибрюшинно, интранодально (т.е. в лимфатические узлы) и т.д.
Организм-хозяин в способе, предлагаемом в изобретении, предпочтительно представляет собой млекопитающее, выбранное из группы, включающей мышь, крысу, свинью, корову, лошадь, собаку, кошку, обезьяну и, прежде всего, человека.
Инъекционный раствор, который приготавливают согласно настоящему изобретению, можно, однако, применять также для трансфекции in vitro клеток РНК, прежде всего мРНК. Такую трансфекцию in vitro можно использовать для лабораторного применения, или она может представлять собой часть генной терапии ex vivo, которая заключается в том, что клетки выделяют из организма пациента, осуществляют трансфекцию ех vivo РНК, содержащейся в инъекционном растворе, предлагаемом в изобретении, и затем ретрансплантируют в организм пациента. Трансфекцию можно осуществлять с помощью процесса электропорации, необязательно также путем приложения импульсов напряжения с напряженностью поля не более 2-10 кВсм-1, продолжительностью импульсов от 10 до 200 мкс и плотностью тока, составляющей по меньшей мере 2 Асм-2. Хотя это не требуется для трансфекции, можно использовать также более продолжительные импульсы, составляющие от 1 до 100 мс. Если инъекционный раствор, предлагаемый в изобретении, используют для лабораторных целей, то таким методом можно трансфектировать любые возможные лабораторные линии клеток. Для генной терапии ех vivo можно рассматривать многочисленные пригодные для трансфекции типы клеток, прежде всего первичные человеческие гемоциты, мультипотентные предшественники гемоцитов, а также фибробласты, нейроны, эндотелиальные клетки или мышечные клетки, причем этот перечень приведен в качестве примера и не направлен на ограничение объема изобретения.
Все литературные ссылки, процитированные в настоящем описании, включены в настоящее описание во всей их полноте.
Ниже настоящее изобретение объяснено и проиллюстрировано на чертежах и примерах, не ограничивающих его объем.
Описание чертежей
В экспериментах, результаты которых представлены на фиг.1-5, 100 мкл буфера, указанного для каждого конкретного случая (составы буферов приведены ниже в разделе «Материалы 1. Инъекционные буферы»), который содержит мРНК (фиг.4, пДНК в 100 мкл ЗФР), кодирующую люциферазу Photinus pyralis, вводили путем внутрикожной инъекции в ушную раковину мышей линии BALB/c13. Анализировали люциферазную активность во всем мышином ухе. Ее выражали в миллионах молекул люциферазы. Предел обнаружения показан на диаграммах жирной линией, на которой указано число. Каждая точка на диаграммах соответствует экспрессии люциферазы в одном ухе. Короткие прямоугольники с числами обозначают средние значения для различных групп. Значения p приведены для групп, для которых имеются достоверные различия между их средними значениями (согласно критерию Манна-Уитни). В экспериментах, результаты которых приведены на фиг.1, 2 и 5, уши удаляли через 15 ч после инъекции. Данные представляют собой результаты, полученные по меньшей мере в трех независимых экспериментах для каждой группы.
На чертежах показаны:
на фиг.1 - сравнение различных буферов для инъекции мРНК: забуференного фосфатом физиологического раствора (ЗФР) и забуференного HEPES физиологического раствора (HBS) и инъекционного буфера RL с лактатом (RL). мРНК lacZ использовали в качестве отрицательного контроля. При создании изобретения установлено, что мРНК, разведенная в инъекционном буфере RL с лактатом, приводит к достоверно (p<0,001) более высокому уровню экспрессии люциферазы, чем мРНК, разведенная в HBS или ЗФР (фиг.1А).
На фиг.2 проиллюстрировано влияние отсутствия кальция (-CaCl), калия (-KCl) или лактата натрия (-NaLa) в инъекционном буфере RL (с лактатом и без лактата, а также с кальцием или калием или без них) на эффективность поглощения мРНК. Основное отличие ЗФР и HBS от инъекционного буфера RL или инъекционного буфера RL с лактатом (с кальцием или без него) состоит в отсутствии лактата и кальция (в HBS или ЗФР). Поэтому были проведены исследования, в которых сравнивали перенос и трансляцию мРНК, кодирующей люциферазу, при использовании, с одной стороны, полного инъекционного буфера RL (инъекционный буфер RL с лактатом) и, с другой стороны, состава инъекционного буфера RL без кальция, или без калия, или без лактата. Эти исследования продемонстрировали, что при отсутствии лактата уровень экспрессии люциферазы сопоставим с уровнем экспрессии, достигаемым с использованием полного инъекционного буфера RL (инъекционный буфер RL с лактатом). В отличие от этого отсутствие кальция в инъекционном буфере RL или инъекционном буфере RL с лактатом приводит к достоверному (р=0,004) снижению эффективности переноса РНК до уровня, сопоставимого с уровнем, достигаемым при использовании ЗФР и HBS.
На фиг.3 и 4 проиллюстрирована кинетика трансляции мРНК непосредственно in vivo. Проводили параллельные эксперименты по изучению кинетики с использованием РНК в RL с лактатом, предлагаемом в изобретении, и ДНК в стандартном ЗФР-буфере и сравнивали результаты. Регистрировали трансляцию мРНК (при использовании инъекционного буфера RL с лактатом) (фиг.3) или пДНК (в ЗФР) (фиг.4) в течение десяти дней после инъекции, результаты представлены на диаграммах. При проведении обоих тестов (с использованием РНК и ДНК) регистрировали люциферазную активность в живых мышах. Представлены результаты, полученные для репрезентативного уха. Экспрессия люциферазы, которую обнаруживали после инъекции мРНК (кодирующей люциферазу) в инъекционном буфере RL с лактатом, очень быстро достигала своего максимума (через 17 ч), и по истечении девяти дней ее уровень становился ниже предела обнаружения (фиг.3). В отличие от этого инъекция пДНК (кодирующей люциферазу) в ЗФР приводила к более поздней экспрессии белка, которая достигала своего максимума через три дня после инъекции и продолжалась в течение более девяти дней (фиг.4). Эти результаты еще раз подтверждают не только эффективность предлагаемого в изобретении инъекционного буфера RL с лактатом, но и то, что РНК является более пригодной в качестве носителя для осуществления кратковременной экспрессии гена в организмах-хозяевах, прежде всего млекопитающих, чем ДНК. Экспрессия РНК происходила, с одной стороны, более быстро, и, с другой стороны, кратковременно, это означает, что экспрессию требуемого гена можно стимулировать раньше и в течение ограниченного промежутка времени и, следовательно, более направленным и дифференцированным образом. С помощью этих исследований удалось продемонстрировать возможность как увеличенного успешного переноса РНК, так и эффективной последующей трансляции.
На фиг.5 проиллюстрировано влияние различных количеств мРНК на экспрессию люциферазы. Эти эксперименты были проведены, в частности, для того, чтобы более точно определять дозу РНК, которую требуется переносить в инъекционном буфере RL с лактатом для клинических применений, с точки зрения количества и продолжительности. Для этой цели нескольким мышам вводили путем инъекций возрастающие количества РНК. Усиление экспрессии люциферазы обнаруживали при увеличении количеств мРНК вплоть до 5 мкг (в объеме 100 мкл раствора для инъекций). Дозы, превышающие 5 мкг, не приводили к дальнейшему усилению экспрессии. В соответствующих экспериментах на людях (данные не приведены) использовали мРНК в количестве 120 мкг, которое увеличивали до 200 мкг, что приводило к усилению экспрессии. Количество мРНК, составляющее для человека 200 мкг, которое сопоставимо с 5 мкг для мыши, можно рассчитывать, среди прочего, по размеру области инъекции, которая у человека примерно в 40 раз больше. Эксперименты на людях осуществляли на здоровых добровольцах после объяснения причин и возможных последствий исследований и после получения согласия.
Касательно зависимости доз от времени следует отметить, что трансляция мРНК происходит в течение короткого промежутка времени (как продемонстрировано на фиг.3, она достигала максимума через 12 ч и снижалась до не обнаруживаемого уровня через девять дней) и, следовательно, требуется ее регулировать. Таким образом, для обеспечения продолжительной равномерной трансляции чужеродной молекулы (белка) можно осуществлять повторную инъекцию примерно через каждый день, каждые два дня или даже каждые три дня (в зависимости от различных факторов, таких, например, как чужеродная молекула или организм, в который осуществляют инъекцию мРНК).
На фиг.6 представлено еще раз влияние CaCl2 на люциферазную активность. Для этой цели приготавливали серийные разведения CaCl2 в буфере для лизиса люциферазы (конечные концентрации приведены на диаграмме) и ко всем образцам добавляли рекомбинантный белок люциферазы в одинаковом определенном количестве (конечная концентрация примерно 4,7 пМ). Испускание света смесями оценивали с помощью люминометра (после добавления АТФ и люцеферина). После этого рассчитывали влияние концентрации CaCl2 на люциферазную активность согласно следующей формуле:
% относительной люциферазной активности (RLA)=(RLA образца с определенной концентрацией CaCl2 - RLA чистого буфера для лизиса) / (RLA образца без СаСl2 - RLA чистого буфера для лизиса)×100%.
Присутствие ионов Ca2+ в относительно высокой концентрации (начиная примерно от 2 мМ) не приводило к увеличению ферментативной активности люциферазы.
На фиг.7 проиллюстрировано еще раз влияние концентрации CaCl2 на перенос мРНК in vivo. Использовали различные концентрации инъекционного буфера RL с лактатом, для приготовления растворов для инъекции РНК (100 мкл), содержащих одинаковое количество мРНК, кодирующей люциферазу Photinus pyralis (20 мкг), но имеющих различные осмолярности (осмол.). Растворы для инъекции РНК вводили путем инъекции в ушные раковины мышей линии BALB/c. Через 15 ч мышей умерщвляли и приготавливали лизаты ушей. Представлены рассчитанное общее количество молекул люциферазы, продуцированных в одном ухе, среднее значение для различных групп (прямоугольники с числами), размер каждой группы (n) и предел обнаружения (жирная линия с числом). В результате исследования было установлено, что для эффективного переноса и, следовательно, для эффективной последующей трансляции мРНК in vivo минимальная концентрация ионов составляет 170 мОсм.
На фиг.8А-Е - характеризация клеток, которые экспрессируют введенную мРНК in vivo. Вводили путем инъекции 20 мкг мРНК, которая кодирует β-галактозидазу Escherichia coli, разведенной в общем объеме раствора для инъекции РНК, содержащего 100 мкл инъекционного буфера RL с лактатом. Через 14 ч после инъекции мышей умерщвляли, удаляли уши и приготавливали поперечные криосрезы. Срезы, представленные на фиг.8А и 8В-8Д, охарактеризованы с помощью цвета.
Кроме того, исследовали направленную экспрессию гена с использованием РНК в инъекционном буфере RL (с лактатом и без лактата). Для этой цели определяли, в каких типах клеток происходит поглощение и трансляция экзогенной РНК, переносимой в инъекционном буфере RL (с лактатом и без него) (см. также пример 5, фиг.8 А-Д и также фиг.11, 12, 16 и 17). После этого анализировали, каким образом в рамках основанной на использовании мРНК вакцинации, предлагаемой в изобретении, можно стимулировать иммунный ответ с помощью трансляции экзогенной РНК, переносимой в инъекционном буфере RL (с лактатом и без него) в определенные клетки-мишени иммунной системы.
В экспериментах, результаты которых представлены на фиг.8А, каждый пятый индивидуальный срез окрашивали раствором, содержащим X-gal. В последовательных криосрезах толщиной 20 мкм было выявлено до 10 позитивных в отношении β-галактозидазы клеток. Область, в которой происходила экспрессия, т.е. в которой присутствовали позитивные в отношении β-галактозидазы клетки (обозначенные стрелками), простиралась на один-два миллиметра в продольном направлении и сагиттальном направлении уха и была локализована в узком слое между эпидермисом и хрящом ушной мышцы.
Кроме того, при создании изобретения было проведено исследование того, обнаруживают ли АПС чужеродный антиген путем непосредственного поглощения и самотрансляции перенесенной мРНК или путем поглощения продукта трансляции перенесенной РНК другими клетками (так называемая «перекрестная презентация»). На основе локализации клеток, их формы и их фенотипа ГКГ класса II можно заключить, что клетки, которые поглощают и экспрессируют экзогенную «оголенную» мРНК в области инъекции, в основном представляют собой мышечные клетки и/или фибробласты (фиг.8А). Результаты согласуются с вышеуказанной «перекрестной презентацией» антигенов, которые транслируются другими клетками. Такая процедура, по-видимому, может объяснить образование антител к белкам, кодируемым вакцинами на основе нуклеиновых кислот. Таким образом, при создании изобретения впервые было установлено, что стимуляция иммунного ответа происходит посредством так называемого «перекрестного примирования», при котором мышечные клетки или клетки дермы (фибробласты) поглощают и экспрессируют перенесенную РНК и АПС активируются этими клетками.
На гистограмме, представленной на фиг.8Б, показано количество позитивных в отношении β-галактозидазы клеток в последовательных срезах. Каждый столбец соответствует одному срезу.
В экспериментах, представленных на фиг.8В-8Д, окрашивали каждый пятый срез, а именно для выявления экспрессии ГКГ класса II (обнаруживается в результате иммунофлуоресцентного окрашивания с помощью Alexa 546, зеленый цвет) и экспрессии β-галактозидазы (обнаруживается с помощью окрашивания фуксином-gal, фиолетовый цвет). На картинках в левой колонке представлены результаты окрашивания с помощью только фуксина, где темные области, окрашенные в фиолетовый цвет, соответствуют позитивным в отношении β-галактозидазы клеткам. На картинках, расположенных в средней и правой колонке, представлены результаты суперпозиции окрашивания с помощью фуксина-gal (показаны представляющие интерес области в средней и правой колонке) и окрашивания для выявления только ГКГ класса II (оранжевый цвет = светлые области в средней колонке) (зеленый цвет = светлые области в правой колонке). Можно видеть, что большинство позитивных в отношении β-галактозидазы клеток являются выраженно негативными в отношении ГКГ класса II.
На фиг.9А-Б представлены результаты, иллюстрирующие перенос in vivo «оголенной» мРНК в организм мыши и человека. Получали мРНК, кодирующую люциферазу, и растворяли в инъекционном растворе RL, содержащем инъекционный буфер RL. Предел обнаружения показан на диаграммах жирной линией, на которой указано число.
В экспериментах, результаты которых представлены на фиг.9А, в ушные мышцы мышей вводили путем инъекции раствор в общем объеме 100 мкл, содержащий 20 мкг мРНК. Через 14 ч после инъекции мышей умерщвляли, клетки ушей лизировали и лизат анализировали в отношении экспрессии люциферазы. Рассчитывали количество молекул люциферазы на одно ухо (в качестве стандарта использовали рекомбинантную люциферазу). Представлены данные, полученные по меньшей мере в трех независимых экспериментах для каждой группы.
В экспериментах, результаты которых представлены на фиг.9Б, ту же самую мРНК (120 мкг) в общем объеме 200 мкл вводили путем инъекции в кожу человека (в голень добровольцев). Через 16 ч отбирали образцы диаметром 2 мм для биопсии (пункционная биопсия) под местной анестезией, а именно из центра области инъекции («мРНК») и на определенном расстоянии от области инъекции («контроль»). Люциферазную активность можно было обнаружить только в средней части области инъекции. Представлены результаты одного из двух независимых экспериментов. Эти результаты демонстрируют наличие непосредственного переноса «оголенной» мРНК in vivo в кожу человека. Таким образом, изобретение позволяет осуществлять эффективную направленную вакцинацию людей с использованием мРНК.
На фиг.10 А-Г проиллюстрирована целостность и способность к трансляции инъецируемой мРНК в инъекционном буфере RL с лактатом. Целостность тестировали с помощью электрофореза на формальдегид-агарозном геле (1,2% (мас./об.)). Для этой цели выделяли 1 мкг мРНК, кодирующей либо люциферазу Photinus pyralis (luc, 1,9 т.п.н., фиг.10А), либо β-галактозидазу Escherichia coli (lacZ, 3,5 т.п.н., фиг.10В). Не было обнаружено никакого различия в целостности мРНК (до инъекции) перед разведением в соответствующем инъекционном буфере (маточный раствор) и после разведения в соответствующем инъекционном буфере. В отличие от этого было выявлено видимое полное расщепление мРНК (после инъекции) в том случае, когда остатки раствора для инъекции РНК собирали с помощью шприца для инъекций. Очевидно, что эти остатки были загрязнены рибонуклеазами при контакте шприца для инъекций с мышиной или человеческой тканью.
Способность к трансляции инъецированной мРНК тестировали с помощью электропорации клеток линии ВНК21 с использованием 10 мкг мРНК. В качестве контроля использовали либо 10 мкг не имеющей отношения к данному вопросу мРНК, либо опыт проводили без использования мРНК («контроль»). Затем клетки либо лизировали и анализировали их люциферазную активность с помощью люминометра (фиг.10Б), либо окрашивали с помощью X-gal и анализировали их люциферазную активность с помощью светового микроскопа (фиг.10Г).
На фиг.11 проиллюстрирована идентификация переноса мРНК на клеточном уровне. На диаграмме представлено схематическое изображение мыши. На диаграмме наружная (дорсальная) сторона обращена непосредственно к (видна) наблюдателю. мРНК в RL-буфере для инъекций, содержащем лактат, вводили путем инъекции в ушную мышцу мыши. Получали различные наборы последовательных поперечных срезов уха (1, 2, 3, 4). Срезы объединяли в различные наборы (1, 2, 3, 4), сушили на воздухе и хранили при -20°С до осуществления процедур окрашивания.
На фиг.12А-В проиллюстрирован перенос мРНК на клеточном уровне. 5 мкг мРНК, кодирующей β-галактозидазу Escherichia coli, в инъекционном буфере RL с лактатом, вводили путем инъекции в ухо мыши. Через 15 ч после инъекции ухо погружали в среду TissueTek О.С.Т и получали криосрезы толщиной 60 мкм. Срезы окрашивали в течение ночи X-gal. На фиг.12А показаны криосрезы уха, в которое не осуществляли перенос мРНК. В этом случае не выявлено наличие lacZ-позитивных клеток. На фиг.12Б изображен общий вид среза уха. На фиг.12В представлено подробное изображение уха, в которое осуществляли перенос мРНК. lacZ-позитивные клетки окрашены в синий цвет и обозначены стрелками.
На фиг.13 представлено сопоставление Alexa Fluor 546-сигнала с цветом фуксин-gal-позитивных клеток. Для того чтобы определить, можно ли обнаруживать Alexa Fluor 546 в фуксин-gal-позитивных клетках, ВНК-клетки трансфектировали комбинациями мРНК eGFP (eGFP = усиленный зеленый флуоресцентный белок) или мРНК lacZ. Анализировали следующие комбинации трансфектированных клеток:
- клетки, трансфектированные по отдельности либо только мРНК eGFP, либо только мРНК lacZ,
- смесь таких индивидуально трансфектированных клеток (eGFP/lacZ),
- клетки, подвергнутые двойной трансфекции (eGFP+lacZ).
Клетки окрашивали антителом к eGFP и проводили обнаружение с использованием Alexa Fluor 546 и затем фуксином-gal. Окрашенные фуксином-gal-позитивные клетки (которые экспрессировали lacZ) выявляли с помощью широкопольного оптического микроскопа (верхний ряд), а окрашенные Alexa Fluor 546-позитивные клетки (которые экспрессировали eGFP) выявляли с помощью флуоресцентной микроскопии (средний ряд). Результаты, полученные в двух опытах, накладывали друг на друга (нижний ряд) для того, чтобы получить точные результаты о локализации клеток относительно друг друга, хотя Alexa 546-сигнал на этой диаграмме перекрывает изображение, полученное с помощью оптического микроскопа. Нельзя исключать то, что имеет место непосредственное поглощение и самотрансляция интродуцированной мРНК в АПС и она является достаточной для стимуляции иммунного ответа. В некоторых АПС могут иметь место процессы слабой или неполной необнаруживаемой трансляции, и (в случае неполной трансляции) они могут оказывать влияние на процессинг и презентацию чужеродного антигена.
На фиг.14 А-Б продемонстрирована специфичность окрашивания ГКГ класса II в криосрезах. Осуществляли инъекцию 20 мкг мРНК, кодирующей β-галактозидазу Escherichia coli, в общем объеме 100 мкл инъекционного буфера RL с лактатом. Через 14 ч после инъекции мышей умерщвляли и удаляли уши. Приготавливали поперечные криосрезы. Криосрезы сначала окрашивали антителом к ГКГ класса II (фиг.14А) или контрольным антителом соответствующего изотипа (фиг.14Б) и проводили обнаружение с помощью иммунофлуоресцентного окрашивания Alexa 546. Затем криосрезы окрашивали фуксином-gal (для выявления экспрессии β-галактозидазы). На чертежах представлены результаты окрашивания фуксином-gal (левая колонка), окрашивания ГКГ класса II (средняя колонка, положения lacZ-позитивных клеток выделены контуром) и суперпозиция обоих результатов окрашивания (правая колонка, lacZ-позитивные клетки выделены контуром, ГКГ класса II-позитивные клетки представлены на чертеже светлыми областями).
На фиг.15 представлено взаимное соответствие клеток, позитивных к красителю X-gal и позитивных к красителю АЭК (3-амино-9-этилкарбазол). Для того чтобы определить, соответствует ли образование преципитата X-gal обнаруженным АЭК-позитивными клетками, ВНК-клетки подвергали совместной трансфекции с использованием мРНК eGFP и мРНК lacZ. Клетки окрашивали путем иммунного анти-eGFP окрашивания с использованием АЭК (красный цвет: позитивные клетки, экспрессирующие eGFP), раствором X-gal (сине-зеленый цвет: позитивные клетки, экспрессирующие lacZ) или комбинации АЭК и X-gal. Окрашенные клетки анализировали с помощью широкопольной оптической микроскопии. Дважды позитивные клетки окрашены в черный цвет (черные стрелки). Трудно различать индивидуально окрашенные позитивные клетки (зеленые и красные стрелки), когда индивидуальное окрашивание является сильным и поэтому выглядит черным.
На фиг.16 А-Б показано совмещение локализации ГКГ класса II-позитивных клеток и позитивных в отношении переноса мРНК клеток. Вводили путем инъекции 20 мкг мРНК, кодирующей β-галактозидазу, в полном объеме 100 мкл инъекционного буфера RL с лактатом. Через 14 ч мышей умерщвляли и удаляли уши. Приготавливали поперечные криосрезы и окрашивали сначала антителом к ГКГ класса II (фиг.16А+Б) или контрольным антителом соответствующего изотипа (фиг.16В) (обнаруживаемым с помощью окрашивания Alexa 546), а затем с использованием X-gal (для выявления экспрессии β-галактозидазы). Клетки, позитивные в отношении переноса мРНК, окрашивались в сине-зеленый цвет, клетки, позитивные в отношении ГКГ класса II, окрашивались в красный цвет, а дважды позитивные клетки окрашивались в черный цвет. Позитивные в отношении переноса мРНК клетки обозначены стрелками независимо от наличия экспрессии ГКГ класса II.
На фиг.17 проиллюстрированы перенос мРНК и морфология ушной мышцы. Вводили путем инъекции 20 мкг мРНК, кодирующей β-галактозидазу, в общем объеме 100 мкл инъекционного буфера RL с лактатом. Через 14 ч после инъекции мышей умерщвляли и удаляли уши. Приготавливали поперечные криосрезы и окрашивали сначала с помощью X-gal (для выявления экспрессии β-галактозидазы), затем гематоксилином и эозином. Клетки, позитивные в отношении переноса мРНК, указаны стрелками, они расположены вблизи слоя паренхимальных клеток.
Примеры
Материалы
1. Буферы для инъекций
Применяли следующие буферы:
- 2-кратный (2×) забуференный фосфатом физиологический раствор (ЗФР) (ЗФР: 274 мМ хлорид натрия, 5,4 мМ хлорид калия, 20 мМ вторичный кислый фосфат натрия, 4 мМ вторичный кислый фосфат калия, рН 7,3 при 20,8°С),
- 2×забуференный HEPES физиологический раствор (HBS) (HBS: 300 мМ хлорид натрия, 20 мМ Hepes, рН 7,4 при 20,8°С) и
- 1×инъекционный буфер RL (без лактата) (82,2 мМ хлорид натрия, 4,3мМ хлорид калия, 1,44 мМ хлорид кальция), если не указаны другой состав и концентрация,
- 1×инъекционный буфер RL с лактатом (102,7 мМ хлорид натрия, 5,4 мМ хлорид калия, 1,8 мМ хлорид кальция, 20 мМ лактат натрия), если не указаны другой состав и концентрация,
- 1×инъекционный буфер RL с лактатом, не содержащий хлорид натрия (4,3 мМ хлорид калия, 1,44 мМ хлорид кальция, 22,4 мМ лактат натрия), если не указаны другой состав и концентрация,
- 1×буфер RL с лактатом, не содержащий хлорид калия (82,2 мМ хлорид натрия, 1,44 мМ хлорид кальция, 22,4 мМ лактат натрия), если не указаны другой состав и концентрация,
- 1×инъекционный буфер RL с лактатом, не содержащий хлорид кальция (82,2 мМ хлорид натрия, 4,3 мМ хлорид калия, 22,4 мМ лактат натрия), если не указаны другой состав и концентрация.
Когда применяли 2×ЗФР и 2×HBS, то все компоненты растворяли в воде и регулировали значение рН. Затем добавляли диэтилпирокарбонат (DEPC, фирма Sigma, Шнелльдорф, Германия) до достижения концентрации 0,1% (мас./об.). Буферы инкубировали в течение 1 ч при 37°С. Затем буферы автоклавировали.
1×инъекционный буфер RL с лактатом приготавливали из 20×маточного раствора четырех различных солей (хлорид натрия, хлорид калия, хлорид кальция и лактат натрия). Аналогичным образом 1×инъекционный буфер RL приготавливали из 20×маточного раствора трех различных солей (хлорид натрия, хлорид калия и хлорид кальция). В других экспериментах исключали хлорид натрия, или хлорид калия, или хлорид кальция без компенсации пониженной осмолярности. RL-буферы, содержащие лактат, не содержащие NaCl, KCl или CaCl2, приготавливали также из 2-кратного маточного раствора. За исключением рацемического раствора лактата натрия (фирма Fluka, Шнелльдорф, Германия) каждый из этих компонентов обрабатывали DEPC и автоклавировали, как описано для 2×ЗФР и 2×HBS.
Все буферы и компоненты буферов проверяли на наличие рибонуклеазной активности путем инкубации 1 мкг мРНК в 1×буфере в течение более чем двух часов при 37°С. При анализе мРНК с помощью электрофореза на формальдегид-агарозном геле использовали буферы, в которых не было выявлено расщепления.
2. Мыши
Все эксперименты на животных проводили в соответствии с утвержденными и национальными руководствами. Самок мышей линии BALB/c возрастом от 8 до 15 недель получали от фирмы Charles River (Шульцфельд, Германия).
Перед осуществлением внутрикожной инъекции мышей анестезировали и ушные мышцы обрабатывали изопропанолом. Для анализа поглощения и трансляции мРНК мышей по истечении определенного периода времени умерщвляли и удаляли уши, которые обривали с помощью бритвенного лезвия для удаления мешающих волос.
3. Люди
Эксперименты на людях проводили с участием здоровых мужчин-добровольцев, которым объясняли причины и возможные последствия исследований и которые давали согласие.
Пример 1: Получение нуклеиновых кислот
мРНК
«Кэппированную» (метилированную) мРНК получали путем «сбегающей» транскрипции in vitro с использованием РНК-полимеразы Т7 (наборы для мРНК T7-Opti, фирма Cure Vac, Тюбинген, Германия).
Кодирующую последовательность этой мРНК (либо β-галактозидазы Escherichia coli [lacZ], клонированной из плазмиды с регистрационным номером U02445, либо люциферазы Photinus pyralis [luc], клонированной из плазмиды с регистрационным номером U47295) фланкировали на ее 3'-концах с помощью нетранслируемой области альфа-глобина и искусственного хвоста поли-А (n=70). Для экспериментов на мышах мРНК экстрагировали фенолом/хлороформом/изоамиловым спиртом и осаждали хлоридом лития. Затем мРНК ресуспендировали в воде и оценивали выход с помощью спектрофотометрии при 260 нм. В завершение мРНК осаждали ацетатом аммония и ресуспендировали в стерильных условиях в воде.
пДНК
ДНК pCMV-luc, не содержащую эндотоксин, получали с использованием набора EndoFree Plasmid Maxi (фирма Qiagen, Гильден, Германия). пДНК осаждали ацетатом аммония и окончательно ресуспендировали в стерильных условиях в воде. Плазмиду pCMV-luc модифицировали путем инсерции Хbа I-(затупленного с помощью фрагмента Кленова) Hind III-фрагмента из pGL3 (регистрационный номер U47295) в расщепленную с помощью Nsi I-(затупленный с помощью фрагмента Кленова) Hind III плазмиду pCMV-HB-S (регистрационный номер А44171). Репортерный ген пДНК находился под контролем промотора CMV.
Маточные растворы
Маточные растворы получали путем разведения мРНК или ДНК в стерильной воде и определения их концентрации и чистоты спектрофотометрическим методом (при 260, 280 и 320 нм).
Контроль качества
Для всех образцов нуклеиновой кислоты концентрацию определяли спектрофотометрически, а целостность проверяли с помощью электрофореза на формальдегид-агарозном геле (мРНК) или с помощью расщепления рестриктазами и электрофореза на ТВЕ-агарозном геле (ДНК) (фиг.10). Помимо целостности анализировали способность к трансляции всех образцов нуклеиновой кислоты путем электропорации клеток линии ВНК21. Для этой цели 1-3 миллиона клеток подвергали электропорации в 200 мкл ЗФР с использованием 10 мкг нуклеиновой кислоты при 300 В и 150 мкФ в кюветах размером 0,4 см. Через 8-24 ч после осуществления электропорации трансфектированные клетки анализировали в отношении экспрессии белка с помощью пригодного метода обнаружения (окрашивание с помощью X-gal или обнаружение люминесценции) (фиг.10). В экспериментах in vivo осуществляли инъекцию только тех образцов нуклеиновой кислоты, которые обладали способностью к экспрессии белка в клетках линии ВНK21 и приемлемой целостностью по результатам гель-электрофореза.
Пример 2: Приготовление растворов для инъекции РНК
При использовании HBS и ЗФР мРНК разбавляли в 1×буфере. Для инъекционного буфера RL с лактатом и без лактата и его определенных вариантов (отсутствие одного из ионов Са2+, K+, Na+) (составы и концентрации приведены в разделе «Материалы, 1. Инъекционные буферы для инъекций») мРНК разбавляли в 0,8-кратном буфере. Если не указано иное, для одного мышиного уха использовали 20 мкг мРНК в 100 мкл инъекционного буфера. Для нарушения вторичных структур, присутствующих в мРНК, растворы для инъекции РНК выдерживали в течение 5 мин при 80°С. Затем растворы помещали на лед еще на 5 мин. В завершение раствор для инъекции РНК набирали в шприцы для инъекции типа Sub-Q (фирма Becton Dickinson, Гейдельберг, Германия). Для каждой инъекции использовали отдельные шприцы для инъекций. Плазмидную ДНК (пДНК) разбавляли в 1×ЗФР.
Пример 3: Обнаружение люциферазной активности ex vivo
Для обнаружения люциферазной активности ех vivo приготавливали лизаты ткани. Для этой цели ткань измельчали в жидком азоте с использованием пестика и ступки и оставшиеся «комочки» гомогенизировали с помощью 800 мкл буфера для лизиса (25 мМ Трис-HCl, 2 мМ ЭДТК, 10% (мас./об.) глицерина, 1% (мас./об.) Triton Х-100 плюс свежедобавленный 2 мМ ДТТ (дитиотреитол) и 1 мМ ФМСФ (фенилметилсульфонил-фторид). Супернатант гомогената получали после центрифугирования (10 мин, 13000 об/мин, 4°С) в миницентрифуге. Аликвоты этого лизата объемом 110 мкл хранили при -80°С.
Для измерения люциферазной активности аликвоты оттаивали на льду и измеряли в течение 15 с испускание света от 50 мкл лизата с помощью люминометра (типа LB 9507, фирма Berthold, Бад-Вильдбад, Германия). Перед осуществлением измерений люминометр автоматически добавлял к лизату 300 мкл буфера А (25 мМ глицилглицин, 15 мМ сульфат магния, 5 мМ свежеприготовленная АТФ, рН 7,8) и 100 мкл буфера Б (250 мкМ люциферин в воде).
При всех измерениях для стандартизации использовали серийные разведения рекомбинантного белка люциферазы (QuantiLum®, фирма Promega, Мэдисон, США). С использованием этого стандарта рассчитывали количество молекул люциферазы для каждого индивидуального измерения. Для каждого лизата люциферазную активность измеряли дважды в два различных дня и рассчитывали среднее значение люциферазной активности. Коэффициент вариации (n=4) количества молекул люциферазы был ниже 10% для всех лизатов, обладающих люциферазной активностью, превышающей предел обнаружения. Этот предел обнаружения (обозначенный на всех диаграммах жирной линией с числом) рассчитывали как среднее значение по трем измерениям для образца, содержащего только буфер для лизиса, плюс три стандартных отклонения этих значений (n=80).
Пример 4: Обнаружение биолюминесценции in vivo
Для обнаружения инъецированной люциферазы в живых животных мышей анестезировали через определенный промежуток времени после инъекции нуклеиновой кислоты. Мышей разделяли на три различные группы: мышам в группе I вводили путем инъекции в левое ухо 100 мкл инъекционного буфера RL без лактата и вводили путем инъекции в правое ухо 20 мкг мРНК, кодирующей люциферазу, в 100 мкл инъекционного буфера RL. Мышам в группе II вводили путем инъекции и в левое и в правое ухо по 20 мкг мРНК, кодирующей люциферазу, в 100 мкл инъекционного раствора RL без лактата. Мышам в группе III вводили путем инъекции в правое ухо 100 мкл инъекционного буфера RL без лактата и в левое ухо по 20 мкг мРНК, кодирующей люциферазу, в 100 мкл инъекционного буфера RL без лактата. Затем мышам вводили путем внутрибрюшинной (i.p.) инъекции 200 мкл люциферина в концентрации 20 мг/мл (фирма Synchem, Кассель, Германия) в ЗФР (стерилизованный фильтрацией). Через 5 мин после инъекции люциферина регистрировали в течение 20-минутного периода времени испускание света от мышей. Для этой цели мышей помещали на предварительно нагретую (37°С) пластину в затемненный ящик (животных из группы I - слева, из группы II - посередине, из группы III - справа). Ящик был снабжен фотокамерой типа Aequoria Macroscopic Imaging (фирма Hamamatsu, Япония). Испускание света регистрировали в виде изображения в условном цвете, на котором на изображение мыши в сером цвете накладывали изображение мыши в обычном свете. Аналогичным образом осуществляли этот же эксперимент с использованием 20 мкг мРНК, кодирующей люциферазу, в инъекционном буфере RL с лактатом или инъекционном буфере RL с лактатом без хлорида натрия, инъекционном буфере RL с лактатом без хлорида калия и инъекционном буфере RL с лактатом без хлорида кальция.
Пример 5: β-Галактозидазная активность и гистология
Обритые мышиные уши разрезали, погружали в среду, содержащую соединение Tissue-Tek® О.С.Т™ (фирма Sakura, Зетервуде, Нидерланды), и хранили при -80°С. Из этих блоков 20 последовательных поперечных криосрезов толщиной 20 мкм помещали в виде 5 наборов (фиг.11) на держатели для образцов типа SuperFrost® plus (фирма Langenbrinck, Эммендинген, Германия) таким образом, чтобы расстояние по вертикали между двумя срезами в наборе составляло примерно 100 мкм. Затем срезы сушили на воздухе и хранили при -20°С до проведения окрашивания. Сначала для поиска области, в которой происходит поглощение и трансляция перенесенной мРНК (кодирующей β-галактозидазу Escherichia соli), 1 набор срезов окрашивали X-gal. Для этой цели держатели для образцов выдерживали при комнатной температуре и очерчивали контур с помощью ручки типа ImmEdge™ (фирма Vektor, Берлингейм, США). Затем срезы фиксировали в течение 15 мин с помощью 2%-ного раствора формалина в ЗФР. После этого держатели для образцов трижды отмывали ЗФР в течение 2 мин и затем окрашивали в течение ночи при 37°С в увлажняющей камере с помощью раствора красителя X-gal (1 мг/мл свежеприготовленного X-gal, 5 мМ феррицианид калия, 5 мМ ферроцианид калия, 1 мМ хлорид магния, 15 мМ хлорид натрия, 60мМ вторичный кислый фосфат натрия, 40 мМ первичный кислый фосфат натрия). Окрашивание прекращали путем двукратной отмывки держателей для образцов в течение 2 мин и обработки их средой Hydro-Matrix® (фирма Micro-Tech-Lab, Грац, Австрия, двукратно разведенная в воде).
Для получения информации о морфологии ткани для другого набора срезов проводили комбинированное окрашивание X-gal и окрашивание гематоксилином-эозином (НЕ). Для этой цели после окрашивания X-gal срезы трижды отмывали в ЗФР в течение 2 мин и затем еще в течение 5 мин в дважды дистиллированной воде, после чего в течение 2 с осуществляли окрашивание гемалюмом Майера (фирма Merck, Дармштадт, Германия). Окрашенные образцы проявляли в течение 10 мин в проточной водопроводной воде, после чего в течение 10 мин осуществляли контрастное окрашивание с помощью 0,1%-ного раствора эозина Y (фирма Sigma, Шнелльдорф, Германия) в воде. Окрашивание прекращали путем кратковременной отмывки в дважды дистиллированной воде, после чего осуществляли дегидратацию с использованием возрастающих концентраций спирта (2 мин - 80% этанолом, 2 мин - 95% этанолом, 2 мин - 100% этанолом, 5 мин - 100% ксилолом). В завершение высушенные срезы обрабатывали средой Roti®-Histokitt (фирма Roth, Карлсруэ, Германия).
Для того чтобы определить, являются ли клетки-мишени, трансфектированные мРНК, антигенпрезентирующими клетками, осуществляли двойное окрашивание молекул ГКГ класса II (экспрессируемых АРС) и перенос мРНК (который оценивали по экспрессии β-галактозидазы). Проводили как иммуногистохимическое, так и иммунофлуоресцентное обнаружение молекул ГКГ класса II. Для обоих протоколов срезы промывали трижды ЗФР в течение 2 мин между всеми тремя стадиями. Для иммуногистохимической процедуры срезы фиксировали с помощью 1% (мас./об.) формалина (фирма Fluka) в ЗФР. Затем липиды удаляли путем инкубации в течение 30 с в чистом ацетоне. Сразу после этого осуществляли блокирование в течение 30 мин при комнатной температуре с помощью 4% козьей крови (фирма Vektor Laboratories Inc., Берлингейм, шт.Калифорния) и 50 мкг/мл авидина D (фирма Vektor Laboratories Inc., Берлингейм, шт.Калифорния) в ЗФР. Оставшиеся сайты связывания биотина блокировали с помощью 50 мкг/мл биотина (фирма AppliChem, Дармштадт, Германия) и одновременно окрашивали молекулы ГКГ класса II с помощью моноклонального антитела 2G9 (фирма Becton Dickinson, Гейдельберг, Германия) или пригодного изотипа контрольного антитела (крысиный IgG 2a, R35-95, фирма Becton Dickinson, Гейдельберг, Германия), в каждом случае разбавленного до 1 мкг/мл (во всех случаях в ЗФР). После этого срезы инкубировали в течение 30 мин с вектором, содержащим биотинилированное козье антитело к IgG крысы (3 мкг/мл) и 2% мышиной сыворотки (фирма ССРrо, Нойштадт, Германия) в ЗФР. Затем добавляли в течение 30 мин при комнатной температуре АВС-комплекс, содержащий реагенты А и Б в соотношении 1:100 в ЗФР (фирма Vektor Laboratories Inc., Берлингейм, шт.Калифорния). После окрашивания ГКГ класса II проводили обнаружение с использованием свежеприготовленного раствора субстрата, 3-амино-9-этилкарбазола (АЭК, фирма Sigma) (0,5 мг/мл АЭС, 0,015% перекиси водорода, 50 мМ ацетат натрия, рН 5,5), который фильтровали через фильтр с размерами отверстий 0,45 мкм. Реакцию с субстратом прекращали путем двукратной отмывки в течение 5 мин водой и трехкратной отмывки в течение 5 минут ЗФР. Затем осуществляли окрашивание с помощью X-gal согласно описанному выше методу.
Аналогичный протокол окрашивания применяли для иммунофлуоресцентного обнаружения. После стадии, на которой использовали ацетон, срезы блокировали в течение 50 мин при комнатной температуре в блокирующем буфере (1% бычьего сывороточного альбумина в ЗФР). Затем срезы инкубировали в течение 40 мин с первичными антителами (2G9 или изотипами контрольных антител), разбавленными до концентрации 1 мкг/мл в блокирующем буфере. После этого осуществляли инкубацию в течение 40 мин при комнатной температуре с использованием красителя, представляющего собой козье антитело IgG крысы Alexa Fluor 546 (1:400; фирма Molecular Probes, Лейден, Нидерланды) в блокирующем буфере. Для этой цели X-gal в растворе для окрашивания заменяли на 0,1 мг/мл magenta-gal (фирма Peqlab, Эрланген, Германия).
Срезы анализировали с помощью микроскопа фирмы Zeiss (Оберкохен, Германия) типа Axioplan 2, снабженного фотокамерой типа Axiocam HRc с программным обеспечением Axiovision 4.0. Осуществляли линейную регулировку цветов и контрастности.
Пример 6: Перенос и трансляция РНК в организм(е) человека
Этот эксперимент проводили на здоровых мужчинах-добровольцах. Места инъекции выбривали, дезинфицировали и обрабатывали раствором RnaseZap (фирма Ambion, Остин, США). Затем осуществляли инъекцию 120 мкг мРНК в 0,8-кратном инъекционном буфере RL в виде одной порции в общем объеме 200 мкл. В дальнейшем вместо инъекционного буфера RL использовали такие же порции следующих буферов:
- инъекционный буфер RL с лактатом без хлорида натрия,
- инъекционный буфер RL с лактатом без хлорида калия и
- инъекционный буфер RL с лактатом без хлорида кальция.
Через 15 ч после инъекции брали образцы для биопсии диаметром 4 мм (пункционная биопсия) с использованием местной анестезии. Образцы для биопсии быстро замораживали в жидком азоте и приготавливали согласно описанному выше методу (пример 3). Измельченные образцы для биопсии ресуспендировали в 600 мкл буфера для лизиса.
Статистическая оценка
Средние значения, полученные для двух различных групп, сравнивали с использованием так называемого «непараметрического рангового критерия суммы Манна-Уитни». Принимали, что различие является статистически достоверным, если значение p, указанное на диаграммах, составляет <0,05.
Пример 7: Комментарии к различным применяемым методам окрашивания
Для идентификации типа клеток, которые поглощают и в которых происходит экспрессия мРНК in vivo, использовали гистологический метод, который позволяет проводить обнаружение переноса мРНК с помощью специфического для определенного типа клеток окрашивания.
Поскольку перенос мРНК нельзя обнаружить с помощью флуоресцентных зондов, осуществляли процесс, в котором использовали мРНК, кодирующую β-галактозидазу Escherichia coli в сочетании с различными красителями из класса индиго (X-gal или magenta-gal).
1. Некоторые особенности системы обнаружения с использованием β-галактозидазы
Для обеспечения того чтобы при осуществлении процесса окрашивания с помощью индиго все клетки присутствовали в виде одного слоя, приготавливали тонкие срезы мышиного уха. Принимая во внимание морфологию ушной мышцы мышей (тонкий слой, имеющий толщину примерно от 0,5 до 1 мм) и тот факт, что можно использовать только криосрезы (β-галактозидаза инактивируется тепловой обработкой при осуществлении стадий, необходимых для приготовления парафиновых срезов), требования к срезам и получению как можно большего количества субсрезов высокого качества, приготовление таких срезов оказалось очень трудным. Тем не менее, оказалось возможным приготовить несколько наборов срезов хорошего качества, имеющих толщину 20 мкм. Для обнаружения β-галактозидазной активности использовали два различных красителя. При использовании X-gal (позитивные клетки окрашивались в сине-зеленый цвет) достигали более контрастных изображений, чем при использовании magenta-gal (позитивные клетки окрашивались в фиолетовый цвет). В то же время, однако, при окрашивании X-gal очень заметно проявлялось неспецифическое фоновое окрашивание, вызываемое, например, волосяными фолликулами. Тем не менее, оказалось возможным четко выявлять перенос мРНК. При использовании magenta-gal неспецифическое окрашивание не было обнаружено.
2. Требования к комбинированному окрашиванию с помощью индиго и иммунного красителя
Процедура окрашивания для обнаружения переноса мРНК (краситель индиго) и окрашивания с использованием специфического для клетки маркера (специфические антитела) требует некоторой адаптации, касающейся фиксирующего агента и последовательности стадий комбинированного окрашивания (окрашивание с помощью индиго включает 14-часовую инкубацию при 37°С). Наилучшие результаты при окрашивании с использованием антитела к ГКГ класса II получали, когда сначала осуществляли фиксацию ацетоном, а затем окрашивание антителом. В отличие от этого наилучшие результаты при обнаружении β-галактозидазной активности получали в том случае, когда сначала осуществляли фиксацию с помощью смеси формальдегида и глутарового альдегида, а затем окрашивание с помощью индиго. Принимая во внимание эти различные обстоятельства, был выбран следующий процесс: фиксация с помощью формальдегида, но без глутарового альдегида, и окрашивание с помощью антитела. Глутаровый альдегид необходимо было исключить, поскольку он резко повышал автофлуоресценцию ткани, в то время как он оказывал лишь слабое действие, если оно вообще имело место, на качество окрашивания с помощью индиго. Фиксация с помощью формальдегида была выбрана по нескольким причинам:
1. обеспечивается более высокая защита морфологии ткани, чем при использовании ацетона,
2. фиксация с помощью формальдегида требуется для достижения более контрастного и интенсивного окрашивания с помощью индиго,
3. при использовании формальдегида все-таки достигается приемлемое качество окрашивание антителом к ГКГ класса II.
После этого осуществляли кратковременную инкубацию в чистом ацетоне для удаления липидов и жиров. Это позволяет достигать лучшего качества (меньшее количество или отсутствие воздушных пузырьков) при использовании водорастворимой среды. И, наконец, хорошего качества окрашивания антителом достигали лишь в том случае, когда это окрашивание осуществляли первым. Последовательность окрашиваний (при фиксации формальдегидом) оказывала лишь небольшое влияние на качество окрашивания с помощью индиго.
3. Совместимость красителей при комбинированном окрашивании с помощью индиго и иммунного красителя
Использование комбинации двух различных методов окрашивания требует не только совместимости различных стадий двух протоколов, но также совместимости зондов, применяемых для обнаружения. В принципе иммунное окрашивание можно осуществлять с использованием осаждающих красителей (ферментативный зонд) или флуоресцентных красителей (меченый зонд). Для объединения окрашивания с помощью индиго и осаждающего красителя использовали X-gal и АЕС. При таком окрашивании дважды позитивные клетки выглядят черными (фиг.15В). Может оказаться трудным различать интенсивно окрашенные индивидуальные позитивные клетки. Поэтому предпочтительно использовать комбинацию magenta-gal с флуоресцентным красителем Alexa Fluor 546. Применять magenta-gal вместо X-gal оказалось предпочтительным по двум причинам:
1. интенсивность окрашивания с помощью magenta-gal была намного более слабой (даже в том случае, когда краситель добавляли в насыщающих количествах),
2. цвет magenta-gal-позитивных клеток более соответствует длине волны испускания флуоресцентного красителя Alexa Fluor 546.
Оба фактора позволяют минимизировать гашение флуоресцентного сигнала Alexa Fluor 546. Такая комбинация красителей реально позволяет обнаруживать оба сигнала (фиг.13), по меньшей мере, в тех случаях, когда окрашивание с помощью индиго не является слишком интенсивным (как это происходило в случае позитивных в отношении переноса мРНК клеток в срезах).
Перечень ссылок
1. Ulmer J.B., Curr. Opin. Drug Discov. Devel., 4, 2001, c.192-197.
2. J.A.Wolff и др., Science 247, 1990, c.1465-1468.
3. H.L.Robinson, L.A.Hunt, R.G.Webster, Vaccine 11, 1993, c.957-960.
4. J.В.Ulmer и др., Science 259, 1993, c.1745-1749.
5. J.A.Wolff и др., Science 247, 1990, c.1465-1468.
6. D.Boczkowski, S.K.Nair, D.Snyder, E.Gilboa, J.Exp.Med. 184, 1996, c.465-472.
7. J.P.Carralot и др., Cell Mol. Life Sci., 2004.
8. R.M.Conry и др., Cancer Res. 55, 1995, c.1397-1400.
9. R.D.Granstein, W.Ding, H.Ozawa, J. Invest Dermatol 114, 2000, c.632-636.
10. I.Hoerr, R.Obst, H.G.Rammensee, G.Jung, Eur. J Immunol. 30, 2000, c.1-7.
11. J.Donnelly, K.Berry, J.В.Ulmer, Int J Parasitol. 33, 2003, c.457-467.
12. D.M.Klinman и др., Springer Semin. Immunopathol. 19, 1997, c.245-256.
13. P.Forg, P. von Hoegen, W. Dalemans, V. Schirrmacher, Gene Ther. 5, 1998, c.789-797.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПЛЕКСЫ НА ОСНОВЕ РНК И КАТИОННЫХ ПЕПТИДОВ ДЛЯ ТРАНСФЕКЦИИ И ИММУНОСТИМУЛЯЦИИ | 2008 |
|
RU2493256C2 |
| ПОЛИКОНЪЮГАТЫ ДЛЯ ВВЕДЕНИЯ IN VIVO ПОЛИНУКЛЕОТИДОВ | 2007 |
|
RU2430740C2 |
| НОВЫЕ МИНИМАЛЬНЫЕ UTR-ПОСЛЕДОВАТЕЛЬНОСТИ | 2017 |
|
RU2759737C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2678127C2 |
| АНТИТЕЛО ПРОТИВ ОПУХОЛЕСПЕЦИФИЧНЫХ АНТИГЕНОВ В КАЧЕСТВЕ МИШЕНИ | 2004 |
|
RU2345090C2 |
| СОСТАВЫ НА ОСНОВЕ МОДИФИЦИРОВАННОГО НУКЛЕОЗИДА, НУКЛЕОТИДА И НУКЛЕИНОВОЙ КИСЛОТЫ | 2012 |
|
RU2649364C2 |
| НУКЛЕИНОВАЯ КИСЛОТА, СОДЕРЖАЩАЯ ИЛИ КОДИРУЮЩАЯ ГИСТОНОВУЮ СТРУКТУРУ ТИПА"СТЕБЕЛЬ-ПЕТЛЯ" И ПОЛИ(А)-ПОСЛЕДОВАТЕЛЬНОСТЬ ИЛИ СИГНАЛ ПОЛИАДЕНИЛИРОВАНИЯ, ДЛЯ УВЕЛИЧЕНИЯ ЭКСПРЕССИИ КОДИРУЕМОГО ОПУХОЛЕВОГО АНТИГЕНА | 2013 |
|
RU2650795C2 |
| НУКЛЕИНОВАЯ КИСЛОТА, СОДЕРЖАЩАЯ ИЛИ КОДИРУЮЩАЯ ГИСТОНОВУЮ СТРУКТУРУ ТИПА "СТЕБЕЛЬ-ПЕТЛЯ" И ПОЛИ(А)-ПОСЛЕДОВАТЕЛЬНОСТЬ ИЛИ СИГНАЛ ПОЛИАДЕНИЛИРОВАНИЯ, ДЛЯ ПОВЫШЕНИЯ ЭКСПРЕССИИ КОДИРУЕМОГО БЕЛКА | 2011 |
|
RU2583295C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ФУНКЦИОНАЛЬНОГО БЕЛКА ИЗ ДНК, ИМЕЮЩЕЙ НОНСЕНС-МУТАЦИЮ, И ЛЕЧЕНИЯ НАРУШЕНИЙ, АССОЦИИРОВАННЫХ С НЕЙ | 2007 |
|
RU2462246C2 |
| Модифицированная РНК, кодирующая полипептиды VEGF-A, составы, содержащие ее, и пути их применения | 2017 |
|
RU2756313C2 |

















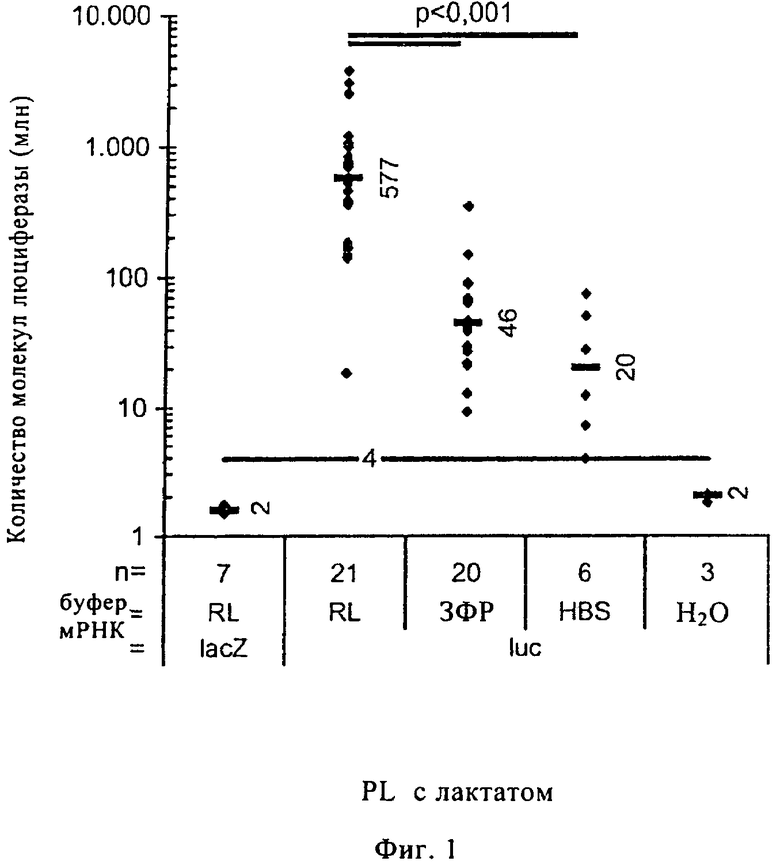
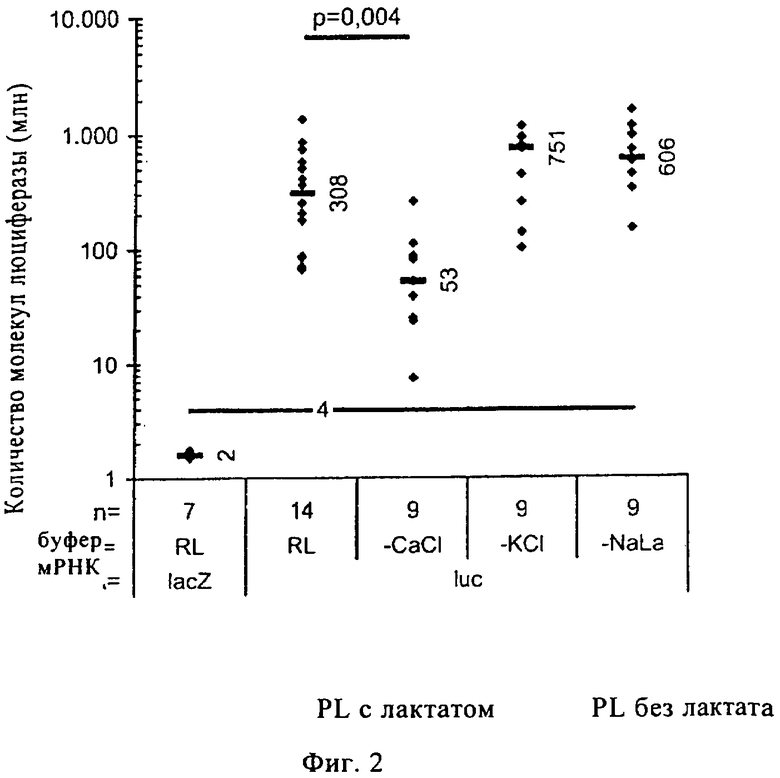
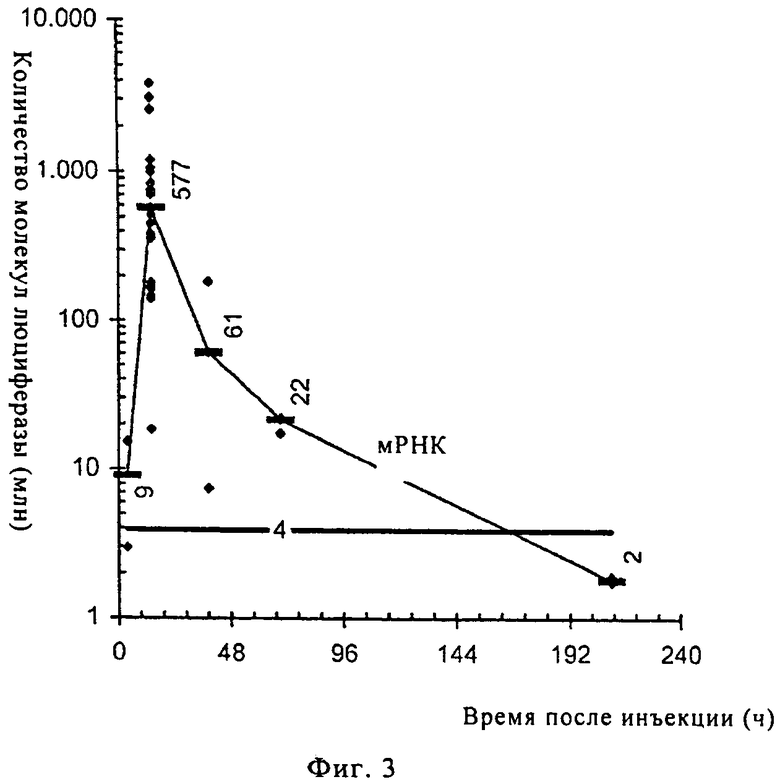
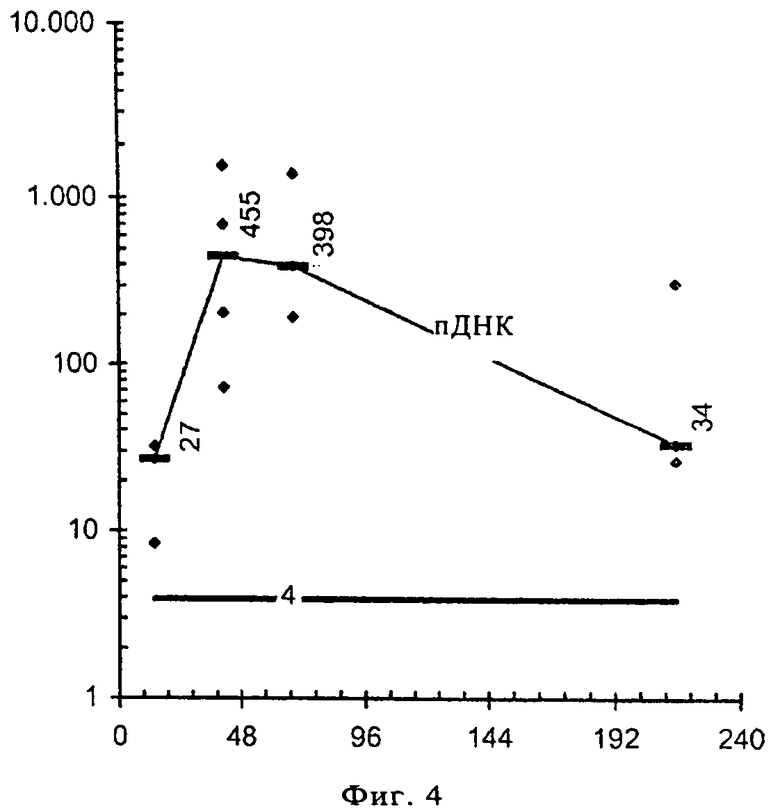
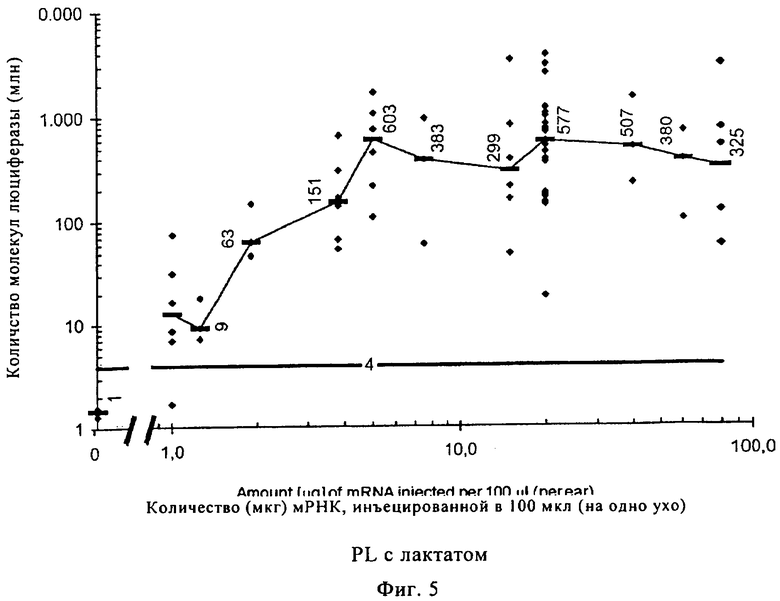
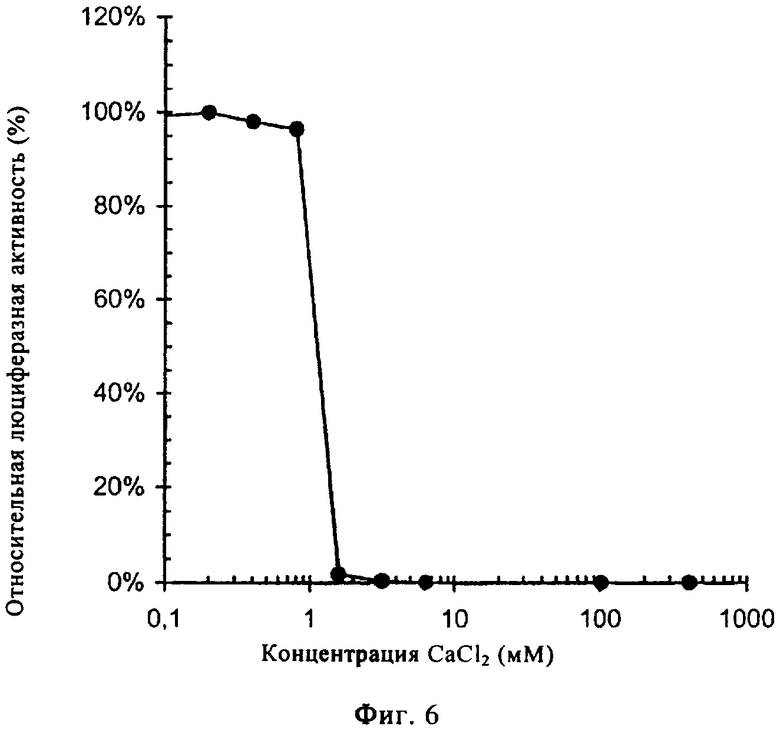
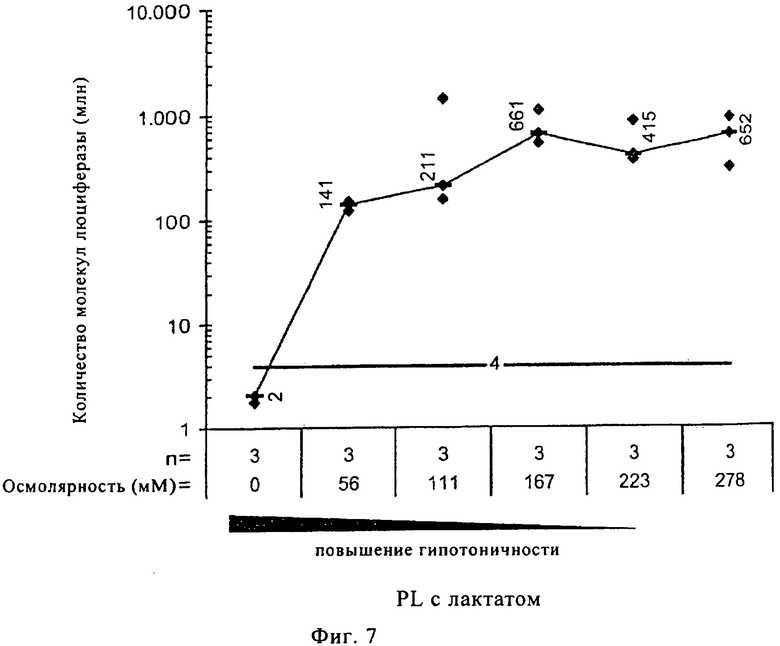
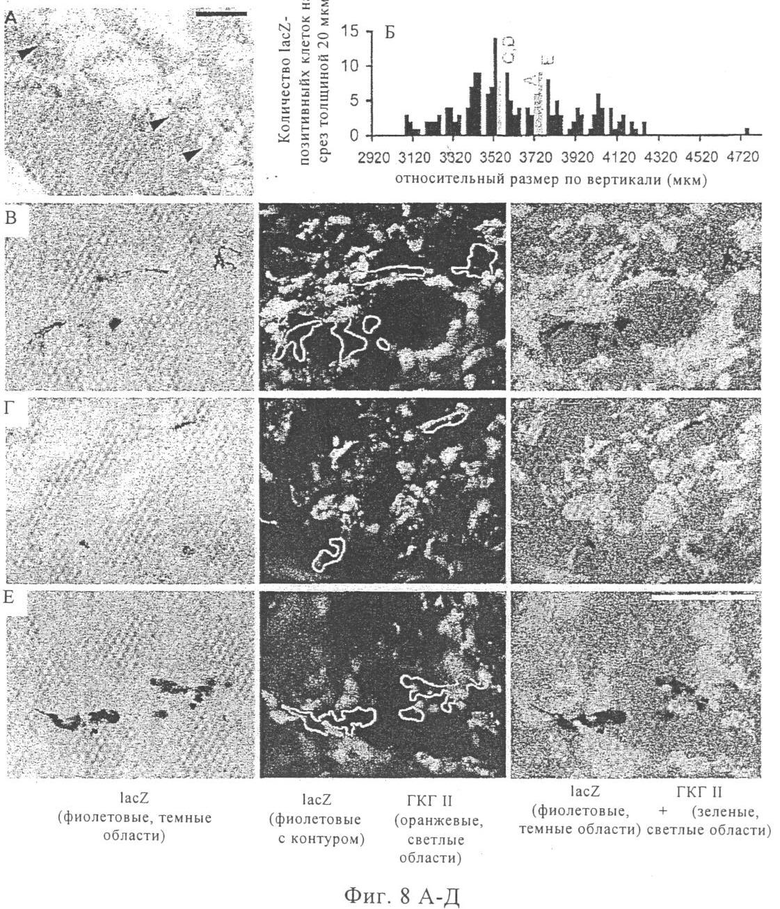
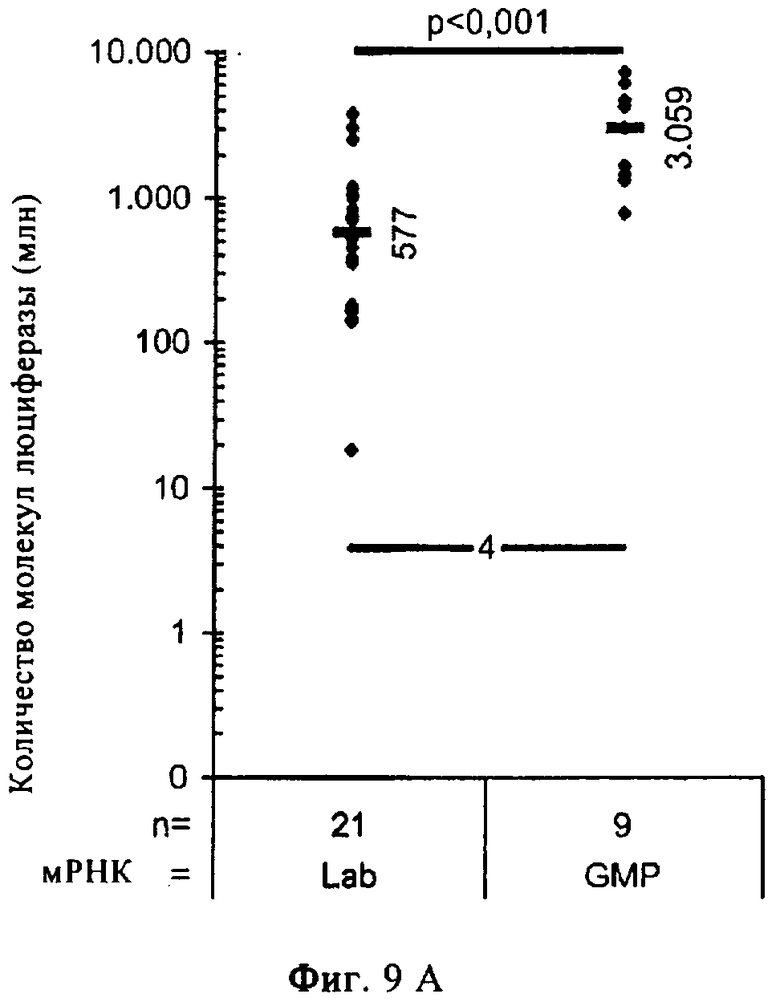
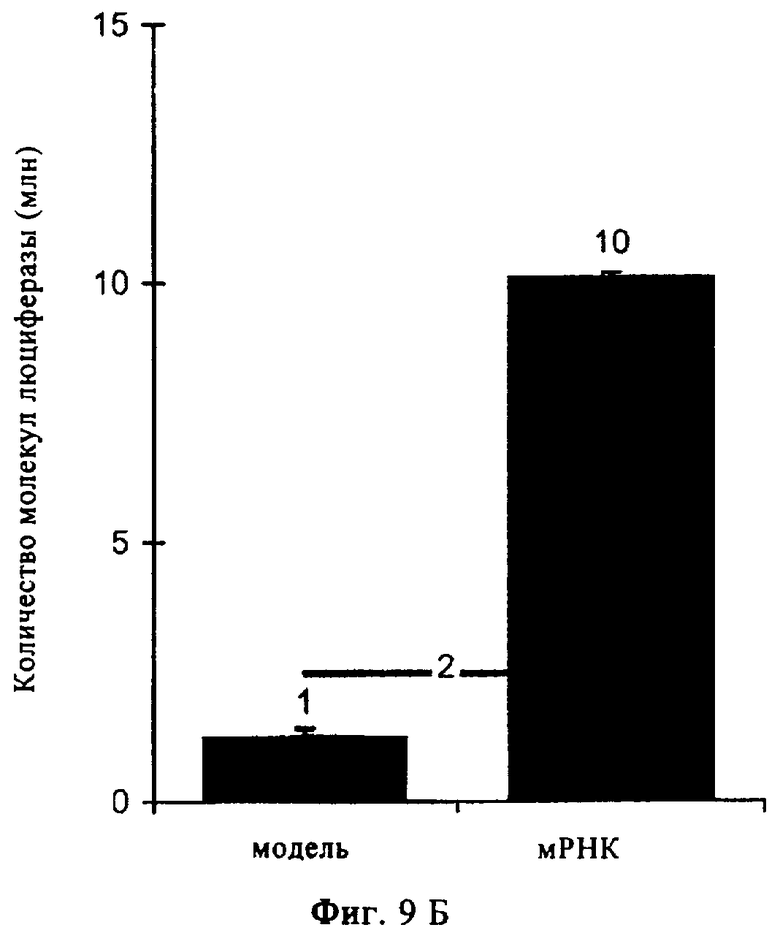
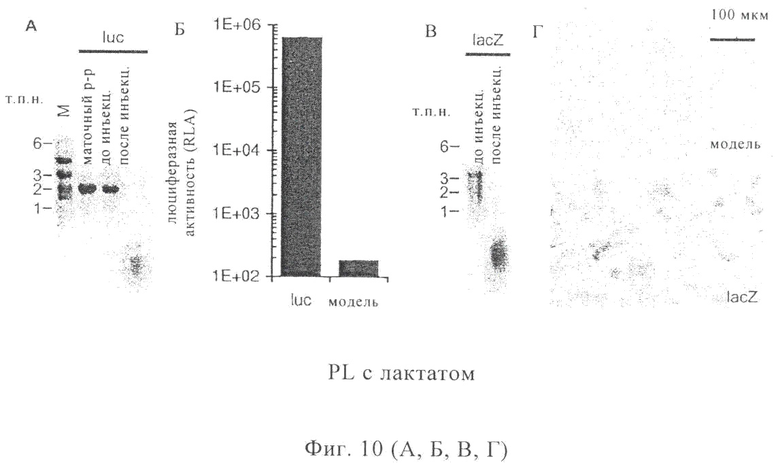
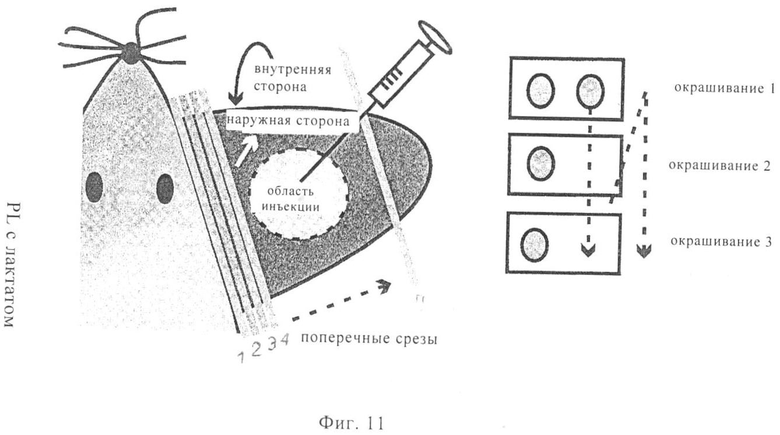
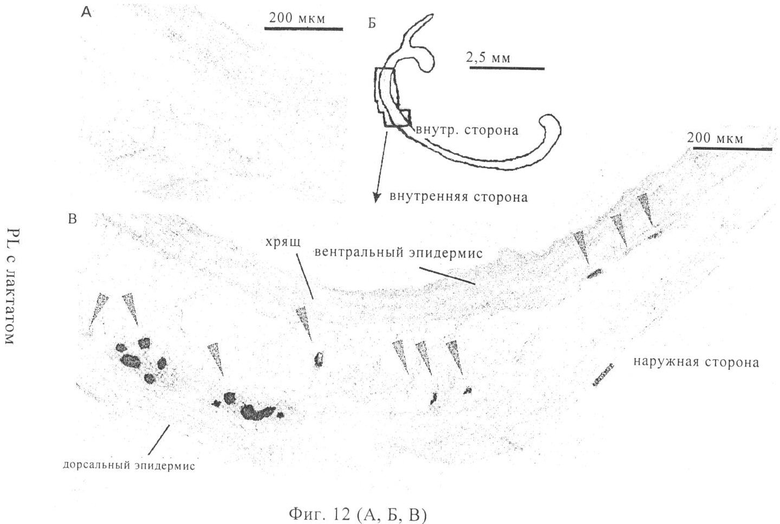
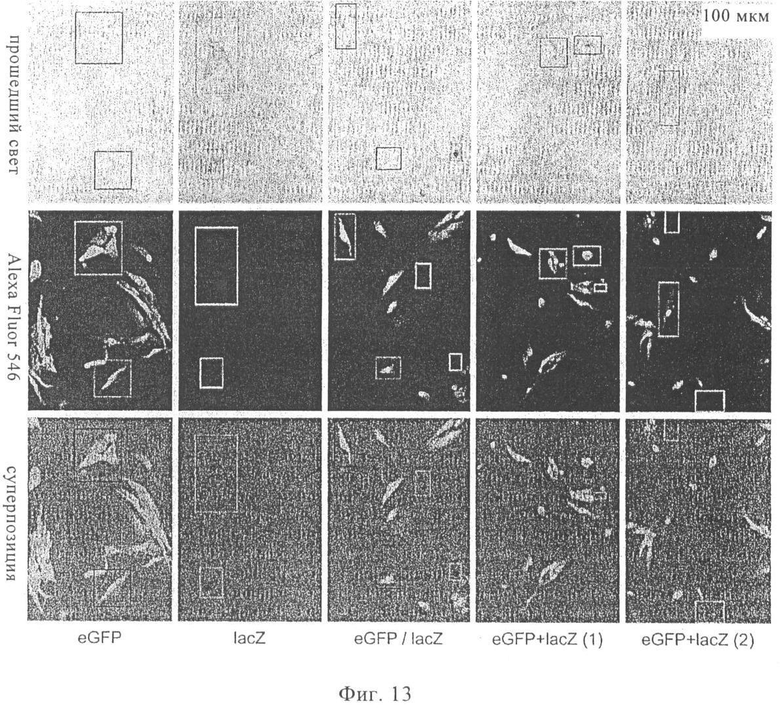
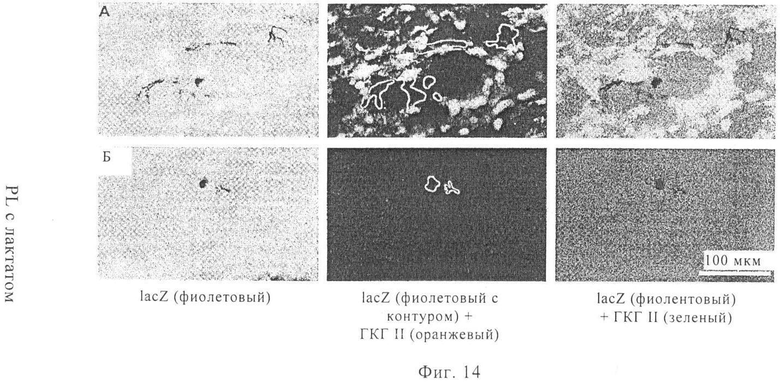
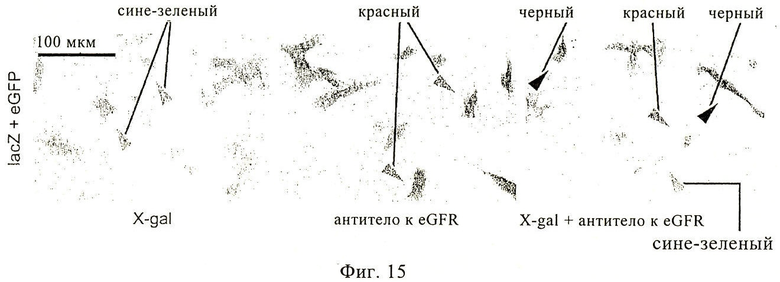

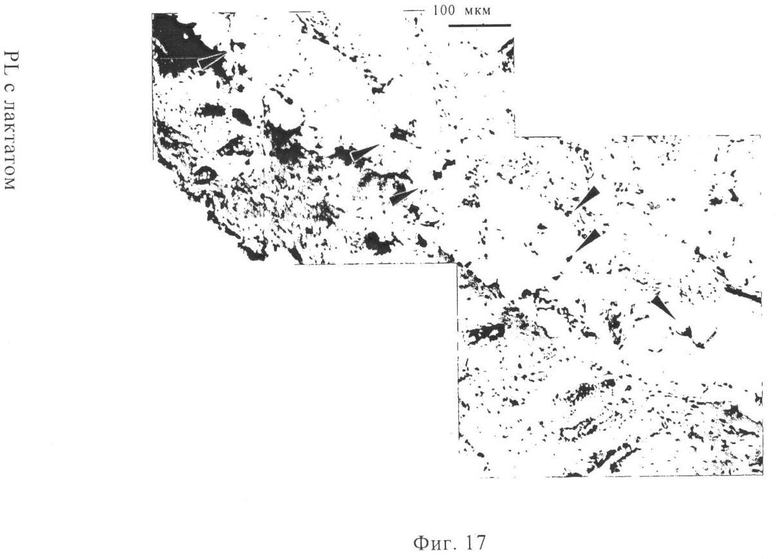
Изобретение относится к области медицины и касается оптимизированной композиции для инъекции РНК. Сущность изобретения включает применение мРНК и водного инъекционного буфера для приготовления раствора для инъекции РНК, предназначенного для увеличения переноса мРНК и/или для ее трансляции в организме хозяина, инъекционный раствор включает Na+ с содержанием 50 мМ, Са2+ по меньшей мере 0,01 мМ, содержание К+ - 3 мМ и необязательно лактат. Преимущество изобретения заключается в повышении стабильности мРНК. 4 н. и 15 з.п. ф-лы, 17 ил., 1 табл.
1. Применение мРНК и водного инъекционного буфера, содержащего натриевую соль, кальциевую соль и необязательно калиевую соль, для приготовления раствора для инъекции РНК, предназначенного для увеличения переноса мРНК и/или трансляции мРНК в организм(е)-хозяин(е), причем используемые концентрации для Nа+ составляют, по меньшей мере, 50 мМ, для Са2+ - по меньшей мере, 0,01 мМ и для К+ - по меньшей мере, 3 мМ.
2. Применение по п.1, в котором инъекционный буфер содержит соли, имеющие анионы, которые выбраны из группы, включающей хлориды, йодиды, бромиды, гидроксиды, карбонаты, бикарбонаты или сульфаты, прежде всего, инъекционный буфер содержит в качестве солей NaCl, CaCl2 и необязательно КСl.
3. Применение по п.2, в котором инъекционный буфер содержит хлорид натрия (NaCl) в концентрации, по меньшей мере, 50 мМ, хлорид кальция (CaCl2) в концентрации, по меньшей мере, 0,01 мМ и необязательно хлорид калия (КСl), в концентрации, по меньшей мере, 3 мМ.
4. Применение по п.2, в котором инъекционный буфер содержит хлорид натрия (NaCl) в концентрации от 50 до 800 мМ, предпочтительно от 60 до 500 мМ, более предпочтительно от 70 до 250 мМ, наиболее предпочтительно от 60 до 110 мМ; хлорид кальция (СаСl2) в концентрации от 0,01 до 100 мМ, предпочтительно от 0,5 до 80 мМ, более предпочтительно от 1,5 до 40 мМ; и необязательно хлорид калия (КСl) в концентрации от 3 до 500 мМ, предпочтительно от 4 до 300 мМ, более предпочтительно от 5 до 200 мМ.
5. Применение по п.1 или 2, где инъекционный буфер дополнительно содержит лактат.
6. Применение по п.1 или 2, в котором инъекционный буфер не содержит буферную систему, такую как HEPES, Трис-HCl, Na(K)2HPO4/Na(K)H2PO4.
7. Применение по п.1 или 2, в котором инъекционный буфер не содержит моно-, ди- или полисахариды.
8. Применение по п.1 или 2, в котором инъекционный буфер содержит хлорид натрия (NaCl) в концентрации, по меньшей мере, 50 мМ, хлорид кальция (CaCl2) в концентрации, по меньшей мере, 0,1 мМ, лактат в концентрации, по меньшей мере, 15 мМ и необязательно хлорид калия (КСl) в концентрации, по меньшей мере, 3 мМ.
9. Применение по п.5, где инъекционный буфер содержит лактат в концентрации от 15 до 500 мМ, предпочтительно от 15 до 200 мМ, более предпочтительно от 15 до 100 мМ.
10. Применение по п.1, в котором мРНК представляет собой «оголенную» мРНК или в частности мРНК, образующую комплекс с поликатионом.
11. Применение по п.10, в котором мРНК образует комплекс с протамином.
12. Применение по п.1, в котором мРНК имеет, по меньшей мере, одну встречающуюся в естественных условиях или не встречающуюся в естественных условиях модификацию, где модификация выбрана, прежде всего, из группы, включающей увеличение содержания G/C в кодирующей области мРНК без изменения кодируемой аминокислотной последовательности, интродукцию, по меньшей мере, одного рибонуклеотидного аналога, удаление дестабилизирующих последовательностей из последовательности дикого типа (WT) или увеличение содержания A/U в области сайта связывания рибосомы.
13. Раствор для инъекции мРНК, содержащий мРНК и инъекционный буфер, указанный в пп.1-12, предназначенный для увеличения переноса мРНК и/или экспрессии мРНК в организм(е)-хозяина(е).
14. Раствор для инъекций по п.13, где инъекционный буфер дополнительно содержит лактат.
15. Раствор для инъекций по п.14, где инъекционный буфер содержит лактат в концентрации, по меньшей мере 15 мМ, более предпочтительно от 15 до 100 мМ.
16. Раствор для инъекции мРНК по п.13, в котором мРНК представляет собой оголенную мРНК.
17. Способ увеличения переноса мРНК и/или трансляции мРНК в организм(е)-хозяин(е), заключающийся в том, что осуществляют следующие стадии:
а) приготавливают раствор для инъекции мРНК по любому из пп.13-16, и
б) вводят раствор для инъекции мРНК, полученный на стадии а), в организм-хозяин.
18. Способ по п.17, в котором организм-хозяин представляет собой млекопитающее, выбранное из группы, включающей мышь, крысу, свинью, корову, лошадь, собаку, кошку, обезьяну и, прежде всего, человека.
19. Способ трансфекции клеток in vitro, заключающийся в том, что вводят инъекционный раствор по одному из пп.13-16 в клетки, прежде всего, в лабораторные линии клеток или родительские клетки, необязательно с помощью электропорации.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

