Настоящее изобретение направлено на способ модификации белка шелка пауков и на белок шелка пауков, получаемый указанным способом. Данное изобретение, кроме того, относится к последовательности нуклеиновой кислоты, кодирующей модифицированный белок шелка пауков, к вектору, содержащему указанные последовательности, и к клеткам-хозяевам, трансформированным этим вектором. Данное изобретение, кроме того, направлено на фармацевтическую или косметическую композицию, содержащую модифицированный белок шелка пауков, как здесь определено, и на применение указанных модифицированных последовательностей в разных областях, конкретно в областях медицины, косметики и в промышленном применении.
Шелка пауков представляют собой белковые полимеры, которые демонстрируют удивительные физические свойства. Среди разных типов шелков пауков канатные шелка исследованы наиболее интенсивно. Канатные шелка используются пауками-кругопрядами для создания остова и радиусов их паутин и в качестве страховочных канатов, которые постоянно тянутся позади них. Для этих целей требуются высокая прочность на разрыв и эластичность. Комбинация таких свойств приводит к прочности, которая выше, чем прочность большинства других известных материалов. Канатные шелка обычно состоят из двух главных белков, первичные структуры которых имеют общую структуру с повторяющимися последовательностями.
Ловчая спираль круглой паутины частично состоит из клейкого шелка, образуемого жгутиковидной железой, который, следовательно, называется жгутиковидным шелком, является эластичным и может утраиваться в длину до разрыва, но обеспечивает только половину прочности на разрыв канатного шелка.
Вариации одиночной повторяющейся единицы, которая может содержать вплоть до 60 аминокислот, повторяются несколько раз, представляя самую большую часть последовательности шелка пауков. Эти повторяющиеся единицы включают ограниченный набор отдельных аминокислотных мотивов. Один мотив, обнаруженный во всех повторяющихся единицах канатного шелка, представляет собой блок, обычно состоящий из 6-9 остатков аланина. В шелковых нитях несколько полиаланиновых мотивов образуют кристаллические стопки β-складчатых конформаций, приводящие к прочности на разрыв.
Обогащенные глицином мотивы, такие как GGX или GPGXX, принимают форму гибких спиральных структур, которые соединяют кристаллические участки и придают нити эластичность.
Сборка шелка in vivo представляет собой примечательный процесс. Белки канатного шелка пауков хранятся в концентрациях вплоть до 50% (масс./об.) в так называемой главной ампуловидной железе. Несмотря на то что для белков в главной ампуловидной железе предложили «динамическую рыхлую спиральную структуру», более новые данные говорят о случайной спиральной конформаций для белков так называемой А-зоны, которая представляет собой самую большую часть данной железы. Высококонцентрированный белковый раствор образует шелковую пасту (прядильный раствор), которая проявляет свойства жидкого кристалла.
Сборка нити инициируется на протяжении прохождения данной пасты через прядильный канал, что сопровождается экстракцией воды, натрия и хлорида. В то же время концентрации более лиотропных ионов калия и фосфата увеличиваются, и рН падает от 6,9 до 6,3. Наконец, сборка запускается механическим напряжением, которое вызвано выталкиванием нити из брюшка паука.
Для нескольких целей нити природного шелка не могут использоваться непосредственно, а должны быть растворены и повторно собраны в другие структуры, такие как пленки, пены, сферы, нанофибриллы, гидрогели и тому подобное.
В то время как были выявлены некоторые структурные аспекты белков шелка пауков, все еще мало известно о вкладе отдельных белков шелка и их первичных структурных элементов в процесс сборки. Сравнительные исследования двух главных белков канатного шелка паука-крестовика Araneus diadematus, ADF-3 и ADF-4, выявили, что хотя их аминокислотные последовательности и являются довольно сходными, они показывают примечательно разную растворимость и особенности сборки: в то время как ADF-3 является растворимым даже при высоких концентрациях, ADF-4 является фактически нерастворимым и подвергается самосборке в нитевидные структуры в конкретных условиях (неопубликованные результаты).
Научный и коммерческий интерес инициировал исследование производства шелка пауков в промышленном масштабе. Получение природного шелка паука не является практичным из-за каннибализма пауков, а искусственное получение столкнулось с проблемами достижения как достаточного выхода белка, так и качества сборки нитей. Бактериальная экспрессия давала низкие уровни белка, что, вероятно, было вызвано различным использованием кодонов у бактерий и у пауков. Синтетические гены с использованием кодонов, адаптированных к экспрессии в хозяине, приводили к более высоким выходам, но белки, синтезированные на их основе, показывали отличные характеристики по сравнению с природными шелками пауков. Экспрессия частичных кДНК канатного шелка в линиях клеток млекопитающих действительно давала белки шелка (например, ADF-3), которые могли быть искусственно сплетены в «шелковые» нити, хотя они до сих пор имеют плохое качество.
Авторы данного изобретения ранее разработали системы для рекомбинантной продукции белков шелка пауков в Е.coli. В качестве примера дается ссылка на WO 2006/008163 (имеющей приоритет предварительной заявки на патент США №60/590196). В этой системе экспрессии можно свободно варьировать одиночными строительными блоками (= модулями), и, таким образом, можно проводить адаптацию к требованиям конкретного случая. Модули этого типа также раскрыты в Hümmerich, D., Helsen, C.W., Oschmann, J., Rudolph, R. & Scheibel, Т. (2004): "Primary structure elements of dragline silks and their contribution to protein solubility and assembly, Biochemistry 43, 13604-13612".
Одной весьма важной задачей, в частности для применений белков шелка пауков в области медицины, является ковалентное связывание лекарственных средств, белков, химических реактивов и тому подобного с этими белками шелка пауков. Однако, вплоть до настоящего времени, не известно удовлетворительной методики для связывания, которая, с одной стороны, обеспечивает связывание этих веществ с белками шелка пауков в заданном количестве и, с другой стороны, заданные положения связывания в белке шелка пауков.
Следовательно, задачей, лежащей в основе настоящего изобретения, является предложение способа изготовления модифицированных белков шелка пауков, которые можно использовать для направленного связывания с этими белками шелка пауков таких веществ, как лекарственные средства, металлы, полипептиды, полисахариды, маркерные молекулы, квантовые точки, нуклеиновые кислоты, липиды и так далее. Также в задачу данного изобретения входит предложение таких модифицированных последовательностей шелка пауков, которые можно использовать для переноски и доставки точного количества этих веществ, и где эти вещества связаны в последовательности белка шелка пауков в заданных положениях.
Эта задача достигается объектом по независимым пунктам формулы изобретения. Предпочтительные воплощения содержатся в зависимых пунктах формулы изобретения.
Весьма интересным в этом отношении является включение в модули шелка пауков аминокислот, которые имеют химически специфичную боковую цепь аминокислоты, в настоящем случае тиольную группу цистеина или аминогруппу лизина. Ни один из вышеупомянутых модулей белков шелка пауков, которые были описаны вплоть до настоящего времени, не содержит цистеина или лизина и, таким образом, специфический мутагенез соответствующих последовательностей нуклеиновых кислот позволяет включить желательные аминокислоты в последовательность модулей контролируемым способом. Модули, которые модифицировали этим способом, можно собрать в новые конструкции и, следовательно, путем комбинирования одиночных модулей в одной одиночной конструкции также можно объединить более чем один химически активный агент или лекарственное средство.
Следовательно, впервые становится возможным специфичное множественное связывание реагентов с рекомбинантными белками шелка пауков. Помимо модификации основных модулей дополнительно существует возможность связывания химически реакционноспособных аминокислот с существующими конструкциями посредством меток (TAG) для того, чтобы активировать или модифицировать их.
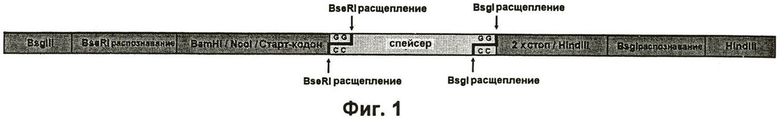
Как упомянуто выше, сам автор данного изобретения создал эффективный способ получения белков, аналогичных белкам шелка пауков и имеющих характеристики, на которые можно оказать специфическое воздействие, путем стратегии клонирования, которая позволяет собрать одиночные модули последовательности ДНК контролируемым путем с образованием синтетического гена (Hümmerich et al., 2004). Данные одиночные модули не разделены последовательностями чужеродных ДНК, как это происходило в системах клонирования из предшествующего уровня техники. В используемой в настоящее время системе клонирования, в качестве примера, можно использовать вектор для клонирования pAZL (разработанный автором данного изобретения), который содержит определенную кассету клонирования (Фиг.1). Самыми важными элементами этой кассеты клонирования являются последовательности распознавания для двух эндонуклеаз рестрикции (BseRI и Bsgl), причем их сайты рестрикции располагаются, соответственно, на расстоянии 8 и 14 нуклеотидов от соответствующих последовательностей распознавания. Это обеспечивает организацию кодонов начала и завершения трансляции и интеграцию дополнительных сайтов рестрикции непосредственно перед или после данного синтетического гена.
Между сайтами рестрикции BseRI и Bsgl в кассете клонирования присутствует спейсерная область, которая будет заменена на следующих стадиях сначала одиночными модулями последовательности и затем синтетическим геном. Организация одиночных элементов будет поддерживаться на следующих стадиях (см. Фиг.1).
Основой мономерных модулей последовательности, которые образуют отправную точку настоящего изобретения, являются гены ADF3 и ADF4 паука Araneus diadematus, а также ген FLAG паука Nephila clavipes. Варианты используемых последовательностей ADF3 и ADF4 являются общедоступными (доступны под номерами доступа U47855 и U47856). Первые два гена (ADF3 и ADF4) кодируют белки, образующие нить канатного шелка пауков, третий кодирует белок жгутиковидного шелка. На основе аминокислотной последовательности этих белков сконструировали несколько модулей:

Из этих аминокислотных модулей были собраны конструкции синтетических белков шелка пауков. Эти модули и белки шелка пауков, происходящие из них, наряду с другими, образуют исходное вещество в настоящем способе модификации белков шелка пауков.
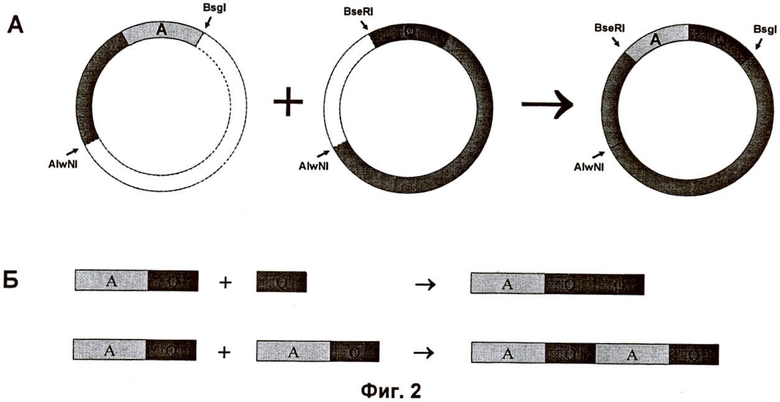
Структура кассеты клонирования в каждом случае обеспечивает произвольную сборку двух модулей или мультимеров модулей. Смотрите в связи с этим Фиг.2, на которой показан пример мультимеризации ДНК-модулей.
В настоящем изобретении предложена система связывания нескольких реагентов с белками шелка пауков, которая делает возможным проведение разных реакций связывания в одно и то же самое время и без больших трудозатрат. Это является ключевым требованием для потенциальных промышленных применений и производства связанных белков шелка пауков.
Для достижения этой задачи выбранные модули белков шелка пауков модифицировали для того, чтобы ввести в выбранные положения аминокислот аминокислоты с химически отличными боковыми цепями. Вновь введенными аминокислотами являются лизин и цистеин.
Настоящее изобретение, в частности, направлено на следующие аспекты и воплощения:
Согласно первому аспекту предложен способ модификации белка шелка пауков, включающий стадии:
а) получения нуклеиновой кислоты, кодирующей белок шелка пауков или его фрагмент, не содержащий остатков лизина или цистеина;
б) замены нуклеиновых кислот, кодирующих один или более чем один аминокислотный остаток в указанном белке шелка пауков, последовательностью нуклеиновой кислоты, кодирующей лизин или цистеин, и/или добавления к указанной последовательности последовательности нуклеиновой кислоты, содержащей нуклеиновые кислоты, кодирующие лизин и/или цистеин;
в) проведения экспрессии модифицированной последовательности нуклеиновой кислоты, полученной в (б), в подходящем хозяине и
г) выделения экспрессированного модифицированного белка шелка пауков.
Приведенный выше способ является самым эффективным способом продукции модифицированных белков шелка пауков по настоящему изобретению. Однако также можно продуцировать те же самые белки, например, путем получения белка шелка пауков или его фрагмента, не содержащего остатка лизина или цистеина (в форме белка), и химически связывать (или присоединять) аминокислотную метку, содержащую лизин и/или цистеин наряду с другими аминокислотами указанного белка шелка пауков.
Кроме того, приведенный выше способ также включает возможность конструирования последовательностей, кодирующих модифицированные белки шелка пауков, путем замещения и последующего добавления к указанной последовательности последовательности нуклеиновой кислоты, содержащей нуклеиновые кислоты, кодирующие лизин и/или цистеин.
Тип и происхождение белка шелка пауков, используемого на стадии (а), не ограничиваются, пока он удовлетворяет требованию не содержать остатков лизина или цистеина. Не играет никакой роли, являются ли они последовательностями, имеющими природное происхождение, или искусственными последовательностями.
Термин «фрагмент» в том виде, как он здесь используется, направлен на части белков шелка пауков (либо искусственные/синтетические, либо природного происхождения), имеющие длину примерно 5-50 аминокислотных остатков, предпочтительно 10-40, например от 20 до 30 аминокислотных остатков.
Согласно предпочтительному воплощению настоящее изобретение дополнительно включает связывание других веществ с указанными молекулами лизина и/или цистеина в модифицированном белке шелка пауков. Как упомянуто выше, это будет приводить к контролируемой и направленной картине связывания указанных веществ с белками шелка пауков.
Одна или более чем одна аминокислота, заменяемая на стадии (б), предпочтительно выбрана из группы, состоящей из глицина, аланина, серина, глутамата, аспартата и треонина. Они обычно не изменяют образующийся модифицированный белок шелка пауков ненадлежащим образом в том, что касается характера сборки и так далее.
Вещество, подлежащее связыванию с указанными остатками лизина и/или цистеина, содержащимися в модифицированном белке шелка пауков, предпочтительно выбрано из группы, состоящей из полипептидов, полисахаридов, маркерных молекул, квантовых точек, металлов, нуклеиновых кислот, липидов и низкомолекулярных лекарственных средств.
Например, частицы нанозолота могут быть связаны с цистеиновыми остатками через химический линкер. В этом случае достигается ковалентное связывание с тиольной группой цистеина через малеимидную или йодацетамидную группу линкера. В основном можно связать все вещества, которые способны ковалентно связываться с аминогруппой лизина или с тиольной группой цистеина.
Предпочтительно низкомолекулярные лекарственные средства выбраны из лекарственных средств, содержащих карбоксильную, карбонильную, имидо или тиольную группу. Неограниченный выбор лекарственных средств представляет собой диклофенак, индометацин, толметин, ибупрофен, флурбипрофен, фенопрофен, напроксен, кетопрофен, пенициллины или цефалоспорины.
Предпочтительно белок шелка пауков, предложенный в (а), основан на канатном и/или жгутиковидном белке. Последовательности шелка пауков, например, могли бы происходить из пауков-кругопрядов (Araneidae и Araneoids).
Более предпочтительно белки шелка пауков происходят из одного или более чем одного из следующих пауков: Arachnura higginsi, Araneus circulissparsus, Araneus diadematus, Argiope picta, Banded Garden Spider (Argiope trifasciata), Batik Golden Web Spider (Nephila antipodiana), Beccari's Tent Spider (Cyrtophora beccarii), Bird-dropping Spider (Celaenia excavata), Black-and-White Spiny Spider (Gasteracantha kuhlii), Black-and-yellow Garden Spider (Argiope aurantia), Bolas Spider (Ordgahus furcatus), Bolas Spiders - Magnificent Spider (Ordgarius rnagnificus), Brown Sailor Spider (Neoscona nautica), Brown-Legged Spider (Neoscona rufofemorata), Capped Black-Headed Spider (Zygiella calyptrata), Common Garden Spider (Parawixia dehaani), Common Orb Weaver (Neoscona oxancensis), Crab-like Spiny Orb Weaver (Gasteracantha cancriformis (elipsoides)), Curved Spiny Spider (Gasteracantha arcuata), Cyrtophora moluccensis, Cyrtophora parnasia, Dolophones conifera, Dolophones turrigera, Doria's Spiny Spider (Gasteracantha doriae), Double-Spotted Spiny Spider (Gasteracantha mammosa), Double-Tailed Tent Spider (Cyrtophora exanthematica), Aculeperia ceropegia, Eriophora pustulosa, Flat Anepsion (Anepsion depressium), Four-spined Jewel Spider (Gasteracantha quadrispinosa), Garden Orb Web Spider (Eriophora transmahna), Giant Lichen Orbweaver (Araneus bicentenarius), Golden Web Spider (Nephila maculata), Hasselt's Spiny Spider (Gasteracantha hasseltii), Tegenaria atrica, Heurodes turrita, Island Cyclosa Spider (Cyclosa insulana), Jewel or Spiny Spider (Astracantha minax), Kidney Garden Spider (Araneus mitificus), Laglaise's Garden Spider (Eriovixia lagtaisei), Long-Bellied Cyclosa Spider (Cyclosa bifida), Malabar Spider (Nephilengys malabarensis), Multi-Coloured St Andrew's Cross Spider (Argiope versicolor), Ornamental Tree-Trunk Spider (Herennia ornatissima), Oval St. Andrew's Cross Spider (Argiope aemula), Red Tent Spider (Cyrtophora unicolor), Russian Tent Spider (Cyrtophora hirta), Saint Andrew's Cross Spider (Argiope keyserlingi), Scarlet Acusilas (Acusilas coccineus), Silver Argiope (Argiope argentata), Spinybacked Orbweaver (Gasteracantha cancriformis), Spotted Orbweaver (Neoscona domiciliorum), St. Andrews Cross (Argiope aetheria), St. Andrew's Cross Spider (Argiope Keyserlingi), Tree-Stump Spider (Poltys illepidus), Triangular Spider (Arkys clavatus), Triangular Spider (Arkys lancearius), Two-spined Spider (Poecilopachys australasia), виды рода Nephila, например Nephila clavipes, Nephila senegalensis, Nephila madagascariensis и многие другие (относительно других видов пауков также смотрите ниже).
Наиболее предпочтительно канатные белки происходят из Araneus diadematus, и жгутиковидные белки происходят из Nephila clavipes.
Предпочтительными канатными последовательностями являются ADF-3 и ADF-4. Термин ADF-3/-4 используют в контексте белков MaSp, продуцируемых Araneus diadematus (фиброин-3/-4 Araneus diadematus). Оба белка ADF-3 и -4 принадлежат к классу белков MaSp II (спидроин II главной ампуловидной железы).
Согласно другому воплощению фрагмент представляет собой модуль, где данный модуль содержит одну или более чем одну консенсусную последовательность, содержащую полиаланин. Эта консенсусная последовательность, содержащая полиаланин, предпочтительно происходит из ADF-3 и имеет аминокислотную последовательность SEQ ID NO: 1 (модуль А) или ее вариант.
Согласно еще одному воплощению данный фрагмент представляет собой модуль, происходящий из ADF-3, и содержит аминокислотную последовательность SEQ ID NO: 2 (модуль Q) или ее вариант. Также рассматриваются комбинированные последовательности из последовательностей, приведенных выше (и все другие упомянутые здесь модули). Например, на стадии (а) предложен фрагмент, который содержит один или более чем один из (AQ) и/или (QAQ). Предпочтительно белок шелка пауков в этом случае содержит (AQ)12, (AQ)24, (QAQ)8 или (QAQ)16.
Таким образом, конкретные модули для применения в настоящем изобретении также можно объединять друг с другом, то есть настоящим изобретением также охвачены модули (повторяющиеся единицы), объединяющие А и Q, Q и С и так далее. Несмотря на то что число модулей, подлежащих введению в белок шелка пауков, не ограничено, для каждого рекомбинантного белка предпочтительно использовать число модулей синтетической повторяющейся последовательности, которое предпочтительно находится в интервале 5-50 модулей, более предпочтительно 10-40, и наиболее предпочтительно 15-35 модулей.
Другой предпочтительный модуль происходит из ADF-4 и содержит аминокислотную последовательность SEQ ID NO: 3 (модуль С) или ее вариант. Объединенные последовательности, предложенные в (а), предпочтительно могут содержать C16 или С32.
Предпочтительными модулями, происходящими из жгутиковидного белка, являются модуль К (SEQ ID NO: 4), модуль sp (SEQ ID NO: 5), модуль Х (SEQ ID NO: 6) и модуль Y (SEQ ID NO: 7).
Предпочтительные комбинации включают Y8, Y16, X8, X16, K8, K16 или Y6X2sp1K2Y2.
При помощи способа по изобретению были генерированы следующие новые модули, и они включены в конкретные предпочтительные воплощения:
Модули канатного шелка пауков

Модули жгутиковидного шелка пауков

Эти модули можно целенаправленно объединять с другими модулями/белками шелка пауков для того, чтобы добиться конкретной модификации. В качестве примера возможностей комбинаций используется конструкция С16. В этом отношении возможными конструкциями с использованием модуля СС могли бы быть следующие конструкции:
C16CC, CCC16, C8CCC8, C16 C, C8CC 8, CC 8C8, C4CC 8C4, CC 4C8CC 4 и так далее.
В этих конструкциях можно добиться контролируемого и направленного связывания через тиольную группу цистеина, а также через аминогруппы лизина. Например, за счет комбинации СC с модулем СK1 существует возможность связать подходящие вещества с тиольными группами цистеина и с аминогруппами боковых цепей лизина.
Предпочтительные конструкции могли бы быть сконструированы следующим образом: например, обе аминокислоты вводят в один одиночный модуль (модуль СKC), что раскрывает дополнительные возможности (имеет место огромное число вариантов конструкций).
Поскольку нельзя исключить то, что в одиночных случаях обмен аминокислотами может приводить к изменению характеристик сборки или к модифицированным характеристикам конструкции, в качестве дополнительной предпочтительной альтернативы, данное изобретение направлено на применение специфичных меток (TAG). Эти метки (например, TAG, как раскрыто в SEQ ID NO: 20-28 ниже) содержат цистеин или лизин, как упомянуто выше. Последовательность TAG выбрана так, что взаимодействие с остальным белком и влияние на характеристики сборки могут быть исключены в максимально возможной степени.
Таким образом, согласно предпочтительному воплощению модифицированный белок шелка пауков, выделенный на стадии (г), содержит один или более чем один модуль SEQ ID NO: 8-19.
Для предпочтительного использования в конструкциях шелка пауков были сконструированы следующие TAG:
аминоконцевые TAG

карбоксиконцевые TAG

Также, как было упомянуто выше, в качестве примера, можно использовать следующие разные варианты:
NHCYS1C16, C16CHCYS1, NHCYS1C16CHLYS1, NHLYS1C16CHCYS1 и так далее.
Замена нуклеиновых кислот, которые кодируют одну или более чем одну аминокислоту в белках шелка пауков, нуклеиновыми кислотами, кодирующими лизин или цистеин, может приводить к изменениям характеристик образующегося модифицированного белка шелка пауков. Для того чтобы избежать нежелательных модификаций, специалист в данной области знает, как выбрать конкретное положение для реакции замещения, чтобы избежать этих нежелательных изменений или для того, чтобы ввести в последовательность белка шелка пауков дополнительные желательные характеристики. Следовательно, особенно предпочтительно использовать негидрофобные аминокислоты, которые являются нейтральными, например не несущие каких-либо зарядов в боковых цепях аминокислот. Аминокислоты, подлежащие замене в исходной последовательности белка шелка пауков, дополнительно должны иметь сопоставимый размер для того, чтобы избежать стерических препятствий из-за вновь введенных аминокислот. Следовательно, для замещения лизином или цистеином особенно предпочтительно использовать серин, аланин, глицин, глутамат, аспартат или треонин.
Таким образом, к модифицированному белку шелка пауков, выделенному на стадии (г), или к белку шелка пауков, предложенному на стадии (а), можно добавить нуклеиновую кислоту, кодирующую аминоконцевой TAG согласно SEQ ID NO: 20-24 и/или карбоксиконцевой TAG согласно SEQ ID NO: 25-28.
Как было описано выше, раскрытые здесь аминокислотные последовательности не ограничиваются точными последовательностями, предложенными в SEQ ID NO. Указанные здесь аминокислотные последовательности также включают варианты. Таким образом, аминокислотные последовательности белков по настоящему изобретению также охватывают все последовательности, отличающиеся от раскрытых здесь последовательностей аминокислотными вставками, делениями и заменами.
Предпочтительно аминокислотные «замены» представляют собой результат замены одной аминокислоты другой аминокислотой, имеющей сходные структурные и/или химические свойства, то есть консервативные аминокислотные замены. Аминокислотные замены можно сделать на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы участвующих остатков. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту.
«Вставки» или «делеции» обычно имеют протяженность от примерно 1 до 5 аминокислот, предпочтительно примерно 1, 2 или 3 аминокислот. Добавления аминокислот обычно содержат не более 100, предпочтительно не более 80, более предпочтительно не более 50, наиболее предпочтительно не более 20 аминокислот, которые добавляют и/или вставляют в белки по настоящему изобретению. Следует отметить, что в данном изобретении рассматриваются только те добавления, которые не оказывают негативного воздействия на желательные характеристики раскрытых здесь белков.
Разрешенные вариации можно экспериментально установить путем систематического создания вставок, делеций или замен аминокислот в белке, используя технологии рекомбинантных ДНК и анализируя образующиеся рекомбинантные варианты на активность. Для специалиста в данной области это не требует проведения экспериментов, более сложных, чем обычные.
Следует отметить, что настоящий способ может дополнительно включать стадию прядения указанных белков, полученных на стадии (г), в филаменты, нановолокно и нити подходящим способом.
Для этой цели можно использовать способы прядения, которые известны в данной области как таковые. Например, раствор пасты белка шелка пауков выдавливают через прядильный механизм с образованием биофиламента. Образующийся биофиламент можно вытягивать или растягивать. Всякий раз, когда в биофиламентах существуют как кристаллические, так и аморфные структуры молекул, вытягивание или растягивание будут налагать напряжение сдвига, достаточное для ориентации молекул таким образом, чтобы сделать их более параллельными стенкам данного филамента и увеличить прочность на разрыв и прочность биофиламента.
Предпочтительно раствор пасты содержит по меньшей мере 1%, 5%, 10%, 15% масс./об. модифицированного белка шелка. Более предпочтительно, раствор пасты содержит 20%, 25%, 30%, 35%, 40%, 45% или 50% масс./об. белка шелка. В предпочтительных воплощениях раствор пасты содержит по существу чистый модифицированный белок шелка пауков. В предпочтительных воплощениях паста имеет рН приблизительно 6,9.
Под «раствором пасты» подразумевают любую жидкую смесь, которая содержит белок шелка и поддается выдавливанию для образования биофиламента или отливки пленки. Растворы пасты также могут содержать, помимо белковых мономеров, агрегаты более высокого порядка, включающие, например, димеры, тримеры и тетрамеры. Обычно растворы пасты представляют собой водные растворы с рН 4,0-12,0 и имеющие менее 40% органических веществ или разобщающих агентов (масс./об.). Предпочтительно растворы пасты не содержат каких-либо органических растворителей или разобщающих агентов, однако могут включать добавки для увеличения сохранности, стабильности или технологичности раствора.
Под «филаментом» подразумевают волокно неограниченной длины, варьирующей от наномасштаба и микроскопической длины до длин в милю (1609 м) или более. Шелк представляет собой природный филамент, тогда как нейлон и полиэстер представляют собой, в качестве примера, синтетические филаменты.
Дополнительную информацию относительно того, как прясть волокна белка шелка пауков, можно найти в WO 03060099 (Karatzas et al.), опубликованной 24 июля 2003, которая включена в данную заявку на патент посредством ссылки.
Кроме того, модифицированные белки шелка пауков по настоящему изобретению можно получить в виде пленок или тому подобного, то есть как продукт белка шелка пауков, для которого не требуется стадия прядения.
Согласно второму аспекту в настоящем изобретении предложен модифицированный белок шелка пауков, который можно получить приведенным выше способом.
Предпочтительный модифицированный белок шелка пауков дополнительно содержит один или более чем один модуль согласно SEQ ID NO: 8-28.
Согласно третьему аспекту предложена последовательность нуклеиновой кислоты, кодирующая модифицированный белок шелка пауков, полученный на стадии (г) способа по изобретению, или модифицированный белок шелка пауков согласно формуле данного изобретения, как упомянуто выше.
Термин «последовательность нуклеиновой кислоты» относится к гетерополимеру нуклеотидов или к последовательности этих нуклеотидов. Термины «нуклеиновая кислота» и «полинуклеотид» используются здесь взаимозаменяемо, чтобы назвать гетерополимер из нуклеотидов.
Термин «строгость гибридизации», как он здесь используется, относится к условиям, при которых полинуклеотидные дуплексы являются стабильными. Как известно специалистам в данной области, стабильность дуплекса является функцией концентрации ионов натрия и температуры (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd Ed. (Cold Spring Harbor Laboratory, (1989)). Уровни строгости, используемые для гибридизации, легко могут быть изменены специалистами в данной области.
Фраза «умеренно строгие условия», как она здесь используется, относится к условиям, которые позволяют ДНК связываться с комплементарной нуклеиновой кислотой, которая имеет примерно 60% идентичности, предпочтительно примерно 75% идентичности, более предпочтительно примерно 85% идентичности с данной ДНК; причем идентичность с указанной ДНК, большая чем примерно 90%, является особенно предпочтительной. Предпочтительно умеренно строгими условиями являются условия, эквивалентные гибридизации в 50%-ном формамиде, 5 × растворе Денхарта (Denhart), 5 × SSPE, 0,2%-ном SDS (додецилсульфате натрия) при 42°С, с последующей промывкой в 0,2 × SSPE, 0,2%-ном SDS при 65°С.
Четвертый аспект направлен на вектор экспрессии, который содержит определенную выше последовательность нуклеиновой кислоты и одну или более чем одну регуляторную последовательность. Этот вектор экспрессии предпочтительно содержит одну или более чем одну регуляторную последовательность. Термин «вектор экспрессии» обычно относится к плазмиде или фагу, или вирусу, или вектору, для экспрессии полипептида/белка с последовательности ДНК (РНК). Вектор экспрессии может содержать транскрипционную единицу, включающую совокупность: (1) генетического элемента или элементов, имеющих регуляторную роль в экспрессии генов, например, промоторов или энхансеров, (2) структурной или кодирующей последовательности, которая транскрибируется в мРНК и транслируется в белок, и (3) подходящих последовательностей инициации и терминации транскрипции. Структурные единицы, предназначенные для применения в дрожжевых или эукариотических системах экспрессии, предпочтительно включают лидерную последовательность, дающую возможность внеклеточной секреции транслированного белка клеткой-хозяином. В качестве альтернативы, когда рекомбинантный белок экспрессируется без лидерной или транспортной последовательности, он может включать аминоконцевой остаток метионина. Этот остаток затем может или не может быть отщеплен от экспрессируемого рекомбинантного белка с получением конечного продукта.
Данный вектор предпочтительно представляет собой плазмидный или вирусный вектор, предпочтительно бакуловирусную систему или векторную систему на основе вируса коровьей оспы. В данном изобретении также можно использовать дополнительные вирусные векторные системы. От случая к случаю может понадобиться модификация вектора. Примерами дополнительных вирусных векторов являются аденовирусы и все РНК-вирусы с минус цепью (negative-strand RNA-viruses), например вирус бешенства, кори, RSV (респираторно-синцитиальный вирус) и так далее.
Готовые к применению генетические конструкции можно клонировать в разных имеющихся в продаже векторах экспрессии, как, например, в системах рЕТ (Novagen, Madison, Wisconsin, США) или pQE (Qiagen GmbH, Hilden, Germany). При помощи конкретного выбора векторов или ферментов рестрикции, которые используются для клонирования, к белку можно присоединить разные белковые TAG (например, Т7-метку (Novagen, Madison, Wisconsin, США) или 6 × гистидиновую метку). Кроме того, можно сделать выбор между разными промоторами (например, между Т7 или Т5).
Предпочтительно данный вектор содержит приведенную выше последовательность нуклеиновой кислоты, кодирующую модифицированный белок шелка пауков, и предпочтительно происходит от вектора для клонирования согласно SEQ ID NO: 29 (вектор для клонирования pAZL) или от его варианта.



Пятый аспект данного изобретения относится к хозяину, которого трансформировали приведенным выше вектором. Данный хозяин может быть прокариотической клеткой, предпочтительно Е.соli или Bacillus subtilis. Экспрессию синтетического гена можно проводить, например, в клетках E.соli К12 или E.соli В. Выход экспрессии может составлять примерно 1 г очищенного белка на литр бактериальной культуры.
Хозяин также может быть эукариотической клеткой, например клеткой млекопитающего, растительной клеткой, дрожжевой клеткой или клеткой насекомого. Предпочтительно он может представлять собой клетку СНО (яичника китайского хомячка), COS, HeLa, 293T, НЕН или ВНК (почки детеныша хомяка), дрожжевую клетку (например, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris, Candida albicans, Hansenula polymorpha), клетку насекомого, выбранную из клеток насекомых Lepidoptera, предпочтительно из Spodoptera frugiperda и из Trichoplusia ni, более предпочтительно из Sf9, Sf21 или клетки "high five", или растительную клетку, предпочтительно происходящую из табака, картофеля, кукурузы, гороха и томата.
Одно преимущество системы экспрессии на основе клетки насекомого, например, по отношению к бактериальным системам, состоит в том, что продуцируемые белки являются гликозилированными, являясь, посредством этого, мишенью для деградации микроорганизмами. Это свойство может быть важным, например, в области медицины, всякий раз, когда белки шелка предназначены для применения in vivo, при котором биологическая деградация является желательной. Эта характеристика может, в частности, найти применение в шовных материалах и в системах ушивания и покрытия ран.
В шестом аспекте настоящее изобретение направлено на фибриллы/нити или филаменты, изготовленные из описанного выше модифицированного белка шелка пауков.
Белки, нити, филаменты, пленки, пены, сферы, нанофибриллы, гидрогели и тому подобное, такие как здесь определено, можно использовать в области биотехнологии и/или медицины, предпочтительно для изготовления систем ушивания или покрытия ран, шовных материалов для применения в нейрохирургии или глазной хирургии. Кроме того, белки/нити можно предпочтительно использовать для изготовления замещающих материалов, предпочтительно материалов искусственных хрящей или сухожилий.
Кроме того, нити/волокна по изобретению можно использовать в изготовлении медицинских приспособлений, таких как медицинские липкие пластыри, кожные трансплантаты, заменители связок и хирургическая сетка; и в широком диапазоне промышленных и коммерческих продуктов, таких как ткань для одежды, подкладка для пуленепробиваемого бронежилета, упаковочный материал, ремни для сумок и бумажников, канаты, тросы, адгезивное связующее вещество, неадгезивное связующее вещество, обвязочный материал, автомобильные покрытия и запчасти, материал для авиастроения, герметичный материал, гибкий перегородочный материал, спортивное оборудование; и, на самом деле, почти при любом применении волокна или ткани, желательными характеристиками для которых являются высокая прочность на разрыв и эластичность. Настоящим изобретением также рассматривается адаптируемость и применение стабильного волокнистого продукта в других формах, таких как сухое напыляемое покрытие, четковидные частицы или применение в смеси с другими композициями.
Как было упомянуто выше, фармацевтические вещества могут быть связаны с модифицированными белками шелка пауков по изобретению через остатки цистеина/лизина. Для приведенных выше целей могут быть использованы именно эти связанные белки. Например, предусмотренным применением такого связанного белка является изготовление шовного материала или системы покрытия ран, имеющих антибиотики или противовоспалительные лекарственные средства, прикрепленные к белкам/нитям, из которых они изготовлены.
Прямо отмечается, что наиболее предпочтительными применениями модифицированных белков шелка пауков по настоящему изобретению являются изготовление и обработка тканей для одежды (текстиль) и кожи, автомобильных покрытий и запчастей, материалов для авиастроения, а также изготовление и обработка бумаги.
Модифицированные белки шелка пауков по настоящему изобретению можно добавлять к целлюлозным, кератиновым и коллагеновым продуктам и, таким образом, настоящее изобретение также направлено на бумагу или продукт по уходу за кожей и волосами, содержащий целлюлозу, и/или кератин, и/или коллаген и белки шелка пауков по настоящему изобретению. Бумага и продукты по уходу за кожей и волосами, в которые включены белки по настоящему изобретению, показывают улучшенные характеристики, конкретно повышенную прочность на разрыв или прочность на отрыв.
Кроме того, модифицированные белки шелка пауков по изобретению можно использовать в качестве покрытия для текстильных и кожаных изделий, придавая, посредством этого, покрытому изделию стабильность и долговечность. Белки шелка, в частности, показывают применимость для покрытия кожаных продуктов, так как в этом случае можно избежать или по меньшей мере уменьшить дубление и его отрицательное влияние на окружающую среду.
Их также можно использовать в упаковке для еды или в элекронных устройствах, например в батареях. Эксперименты, проведенные с пленками, изготовленными из данных модифицированных белков шелка пауков, показали их устойчивость и стабильность к кислотам после погружения в электролит аккумулятора.
В седьмом аспекте согласно данному изобретению предложена фармацевтическая или косметическая композиция, содержащая модифицированный белок шелка пауков, как определено выше, и фармацевтически приемлемый носитель.
Данное изобретение далее проиллюстрировано сопровождающим графическим материалом, где:
на Фиг.1 показана схематическая структура кассеты клонирования, использованной в настоящем изобретении;
на Фиг.2 проиллюстрирован пример мультимеризации модулей ДНК по настоящему изобретению;
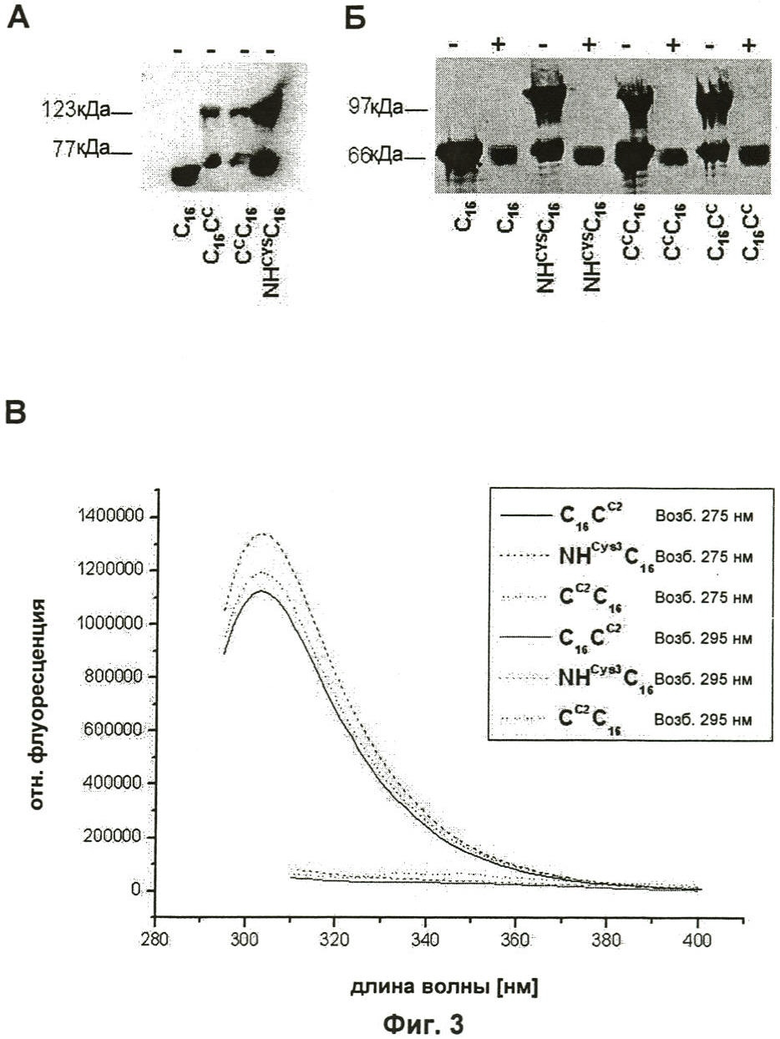
на Фиг.3 показан анализ модифицированных белков шелка пауков. Образцы анализировали с (+) или без (-) восстанавливающего агента (2-меркаптоэтанол). (А) Т7-метки рекомбинантных белков шелка были определены после вестерн-блоттинга с антителом против Т7-метки. (Б) Белки подвергали SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) с последующим окрашиванием серебром. (В) Показаны флуоресцентные спектры испускания очищенных С16СC (прямая линия), NHCYS3C16 (длинные штрихи) и CCC16 (пунктирная линия) с длинами волн возбуждения 275 нм или 295 нм, соответственно. Возбуждение при 295 нм показывает отсутствие флуоресценции триптофана. Следовательно, данные белковые пробы не демонстрируют какого-либо определимого загрязнения бактериальными белками;
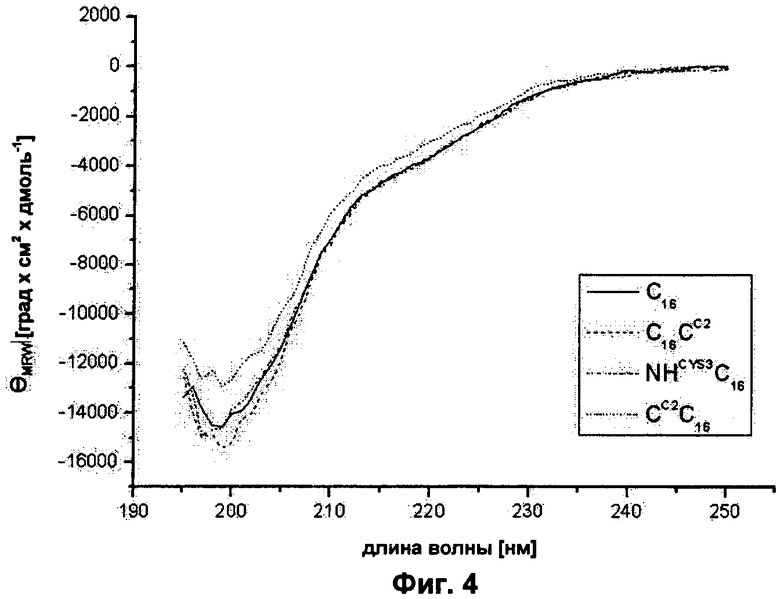
на Фиг.4 показан анализ вторичной структуры модифицированных белков шелка пауков. Спектры CD (циркулярный дихроизм), C16 (прямая линия), C16CC (длинные штрихи), NHCYS3C16 (точки и штрихи) и CCC16 (пунктирная линия) записывали при 20°С;
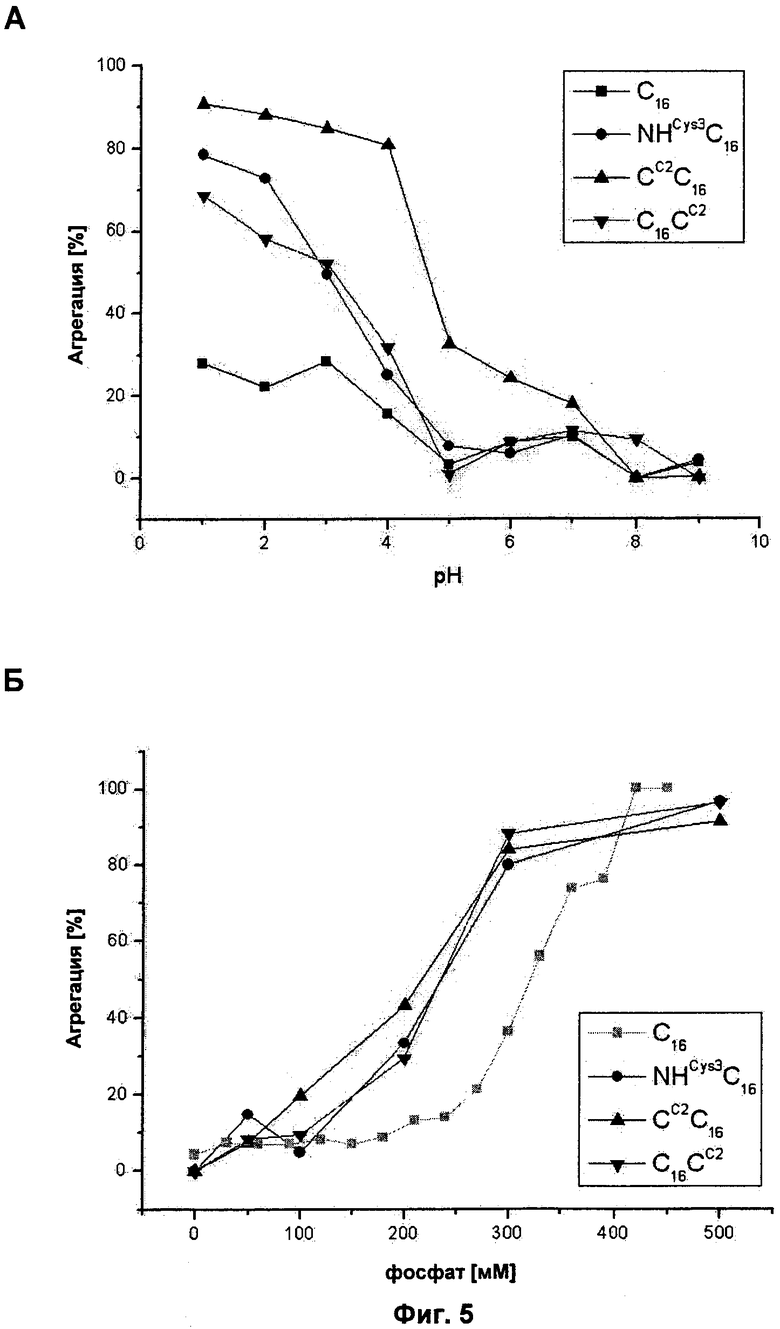
на Фиг.5 показана агрегация модифицированных синтетических белков шелка пауков. Агрегацию белков определяли после инкубации в течение 2 часов в буферах с (А) варьирующим рН или (Б) с варьирующими концентрациями фосфата калия (смотрите таблицу 2). Полученная кривая для C16 представлена квадратиками, кривая для NHCYS3C16 - кружками, для CC2C16 - треугольниками и кривая для C16CC2 - поворотными треугольниками;
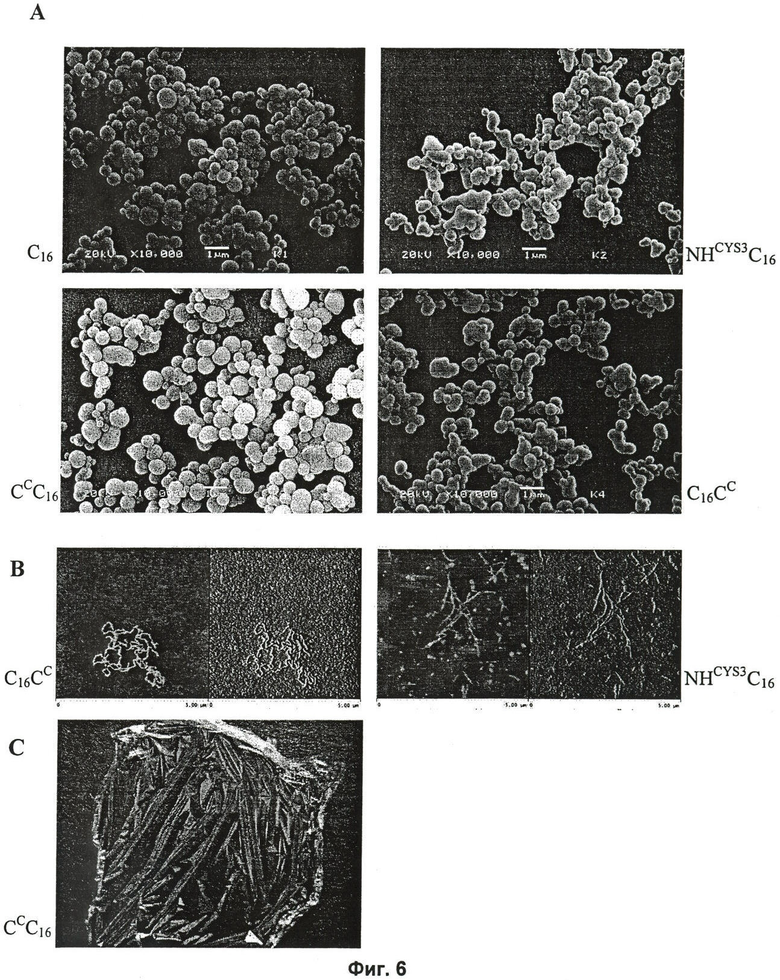
на Фиг.6 показаны формы сборки модифицированных белков шелка пауков. (А) Сферы, образованные С16 и модифицированными белками, визуализированные сканирующей электронной микроскопией (SEM). (Б) Нанофибриллы, визуализированные атомно-силовой микроскопией. (В) Пленка, отлитая из 1%-ного масс./об. раствора CC2C16 в HFIP (гексафторизопропанол);
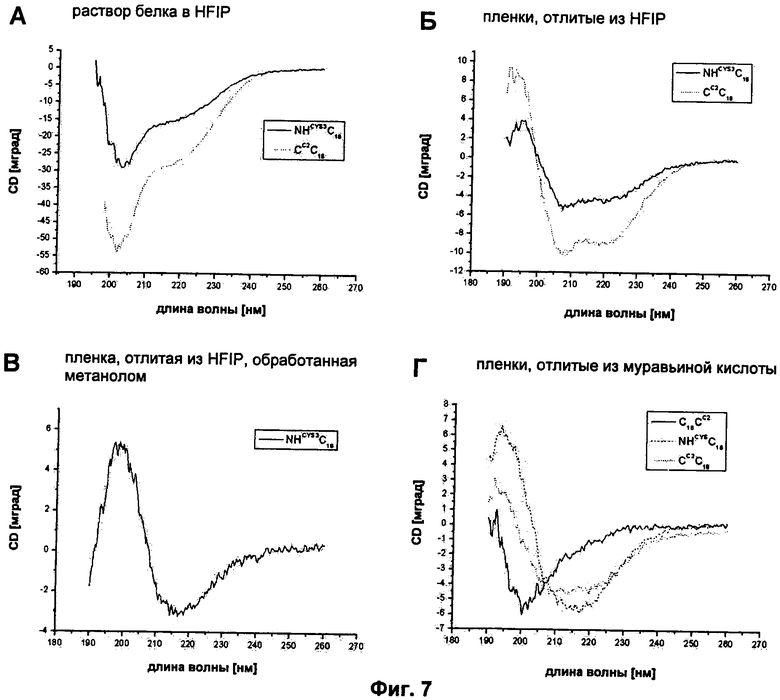
на Фиг.7 показаны спектры CD белковых пленок, изготовленных из модифицированных белков шелка пауков. (А) Белковый раствор в HFIP анализировали перед отливкой пленки. (Б) Пленки отливали из HFIP непосредственно на простом кварцевом стекле и анализировали CD-спектроскопией. (В) Анализ CD пленок, отлитых из HFIP, обработанных метанолом (в качестве примера показано для NHCYS3C16). (Г) Анализ пленок, отлитых из муравьиной кислоты, непосредственно на простом кварцевом стекле. Из-за неточностей в определении толщины пленок ΘMRW не мог быть определен;
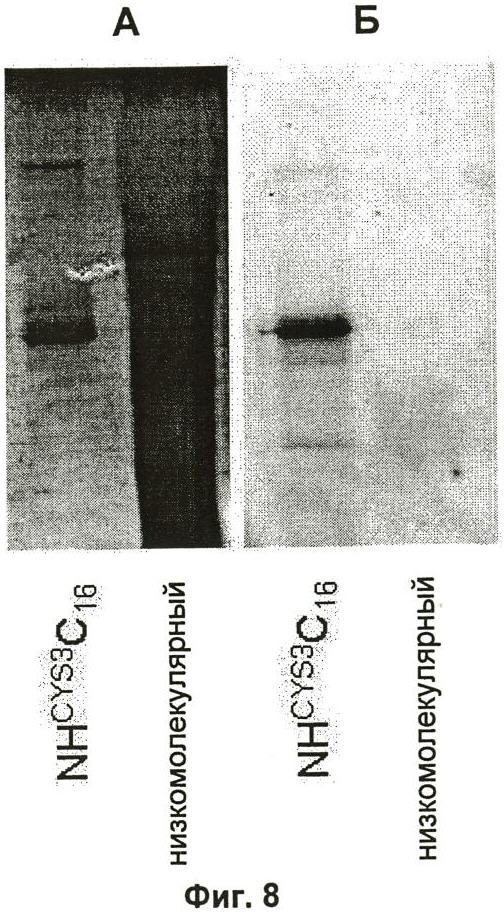
на Фиг.8 показано связывание родамина RedTM C2 малеимида с NHCYS3C16. Связанный белок анализировали SDS-PAGE. (А) Белки визуализировали окрашиванием серебром. (Б) Родамин визуализировали флуоресцентной визуализацией.
ПРИМЕРЫ
ЭКСПЕРИМЕНТАЛЬНЫЕ МЕТОДИКИ
Материалы. Химические реактивы приобретали у Merck KGaA (Darmstadt, Германия), если не определено иначе. Манипуляции с ДНК и модификацию ДНК проводили, как описано ранее (1). Ферменты рестрикции приобретали у New England Biolabs (Beverly, MA, США) и лигазу - у Promega Biosciences Inc. (San Luis Obispo, CA, США). Очистку ДНК проводили, используя наборы от Promega Biosciences Inc. (San Luis Obispo, CA, США). Синтетические олигонуклеотиды приобретали у MWG Biotech AG (Ebersberg, Германия). Все стадии клонирования проводили в штамме DH10B Е.coli от Novagen (Madison, WI, США).
Клонирование модулей модифицированного шелка и TAG в векторе pAZL
Модуль СC (SEQ ID NO: 10) создавали посредством ПЦР (полимеразная цепная реакция) мутагенеза с использованием модуля С (SEQ ID NO: 3) в качестве матричной нуклеотидной последовательности и праймеров pAZL-прямой (CACTGAGCGTCAGACCCCGTAGAAAAGA) (SEQ ID NO: 30) и pAZLмут-обратный (CTCTTAAGCTTTCATTAGCCTGGACCACCTGGACCGTAGCCGCCCGGGCCGCAAGGACCCTGG) (SEQ ID NO: 31). Для того чтобы получить оптимизированный праймер, некоторые кодоны исходного модуля С мутировали (ССА (Рro24) до ССТ (Pro), GGT (Gly28) до GGC (Gly), ССТ (Pro32) до ССА (Pro), GGC (Gly33) до GGT и CCG (Pro35) до ССА (Pro)). Продукт ПЦР и вектор pAZL (SEQ ID NO: 29) лигировали после расщепления AlwNI и HindIII. Модуль NHCYS3 (GCGGSGGGGSGGGG, ggt tgc ggt ggc tct ggt ggt ggc ggg tcc gga ggc ggt ggc) (SEQ ID NO: 22) создавали путем отжига двух синтетических олигонуклеотидов N1 (GATCCATGGGTTGCGGTGGCTCTGGTGGTGGCGGGTCCGGAGGCGGTGGCTAATGAA) (SEQ ID NO: 32) и N2 (AGCTTTCATTAGCCACCGCCTCCGGACCCGCCACCACCAGAGCCACCGCAACCCATG) (SEQ ID NO: 33). Отжиг проводили путем снижения температуры 50 пмоль/мкл (каждого) раствора олигонуклеотида от 95°С до 20°С с инкрементом 0,1°С/с. Ошибочно спаренные двойные цепи денатурировали при 70°С с последующим дополнительным снижением температуры до 20°С. После повторения цикла 20°С-70°С-20°С десять раз проводили десять дополнительных циклов с температурой денатурации 65°С. Образующуюся кассету клонирования лигировали с вектором pAZL (SEQ ID NO: 29), расщепленным ВаmНI и HindIII.
Конструирование модифицированных синтетических генов шелка пауков Соединение двух фрагментов гена, например одиночных модулей или модульных мультимеров представляло собой основную стадию стратегии клонирования. Для этой цели вектор pAZL, содержащий обозначенный 5'-концевой фрагмент гена, расщепляли Bsal и Bsgl, тогда как вектор, содержащий 3'-концевой фрагмент гена, расщепляли, соответственно, BseRI и Bsal (Фиг.1). Лигирование подходящих фрагментов плазмиды давало соединение двух фрагментов гена и приводило к воссозданию гена устойчивости к ампициллину (Ap') вектора pAZL, который облегчал идентификацию правильных конструктов.
Для конструирования гена модифицированные модули CC (SEQ ID NO: 10) или NHCYS3 (SEQ ID NO: 22) соединяли с повторяющимися единицами, подобными C16. Затем их вырезали из вектора pAZL с помощью BamHI и HindIII и лигировали с бактериальным вектором для экспрессии рЕТ21а (Novagen), расщепленным подобным образом, получая кодирующую последовательность Т7-метки (MASMTGGQQMGR) (SEQ ID NO: 34) (2). Правильность всех конструктов подтверждали секвенированием ДНК.
Экспрессия генов. Все гены шелка экспрессировали в штамме BLR [DE3] E.соli (Novagen). Клетки выращивали при 37°С в среде LB (Луриа-Бертани) до OD600=0,6-0,7. После индукции 1 мМ IPTG (изопропил-β-D-тиогалактозид) клетки переводили на 25°С, в случае NHCYS3C16 и C16CC и на 30°С, в случае СCС16, соответственно. Клетки, экспрессирующие NHCYS3C16, собирали после 3-4 часов индукции, тогда как клетки, экспрессирующие CCC16, собирали через 4 часа и клетки, экспрессирующие C16CC, - через 5 часов.
Очистка белка. Клетки ресуспендировали 5 мл/г буфера, содержащего 20 мМ N-(2-гидроксиэтил)пиперазин-N'-(2-этансульфоновую кислоту) (HEPES) рН 7,5, 100 мМ NaCl, 0,2 мг/мл лизоцима (Sigma-Aldrich, St. Louis, МО, США) и инкубировали в течение 30 мин при 4°С. Клетки разрушали высоким давлением с использованием французского жима (Basic Z Model, APV Deutschland GmbH, Lübeck, Германия). Геномную ДНК расщепляли путем инкубации лизатов клеток с 0,1 мг/мл ДНКазой I (Roche, Mannheim, Германия) и 3 мМ MgCl2 при комнатной температуре в течение 30 мин. Нерастворимые фрагменты клеток осаждали при 50000×g и 4°С в течение 30 мин. Растворимые белки лизатов E.соli осаждали путем денатурации нагреванием при 80°С в течение 20 мин. Осажденные белки удаляли седиментацией при 50000×g в течение 30 мин. Белки шелка, которые оставались растворимыми на протяжении денатурации нагреванием, осаждали 20%-ным сульфатом аммония (800 мМ) при комнатной температуре и собирали центрифугированием при 10000×g в течение 10 мин. Осадки промывали 8 М мочевиной и растворяли в 6 М гуанидиния тиоцианате (GdmSCN). Все белки диализировали против 10 мМ NH4HCO3. Осадки, образующиеся во время диализа, удаляли седиментацией при 50000×g в течение 30 мин и остальные растворимые белки шелка лиофилизировали. Перед анализом лиофилизированный белок растворяли в 6 М GdmSCN с последующим диализом против подходящих буферов. Агрегаты удаляли седиментацией при 125000×g в течение 30 мин. Концентрации белка определяли фотометрически в кюветах с длиной пути 1 см при 276 нм, используя рассчитанные коэффициенты экстинкции (таблица 1) (3). Идентичность белков подтверждали электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE; 10%-ные Tris-глициновые гели) с последующим блоттингом на поливинилиденфторидных (PVDF) мембранах (Millipore, Billerica, MA, США) и определением с использованием мышиного моноклонального антитела против Т7 (Novagen, 1:10000), в качестве первичного, и конъюгата антимышиный IgG-пероксидаза (Sigma-Aldrich, 1:5000), в качестве вторичного антитела. Пероксидазную активность визуализировали, используя набор для выявления вестерн блотов ECLplus от Amersham Biosciences (Piscataway, NJ, США).
Коэффициенты экстинкции синтетических шелков паука (рассчитанные согласно Gill & Hippel (3))
Флуоресценция. Спектры флуоресценции записывали на спектрофлуориметре FluoroMax 3 (Jobin Yvon Inc, Edison, NJ, США). Спектры получали в 10 мМ NН4НСО3 или в 10 мМ трис(гидроксиметил)аминометане (Tris)/HCl (рН 8,0) при 25°С. Время интегрирования составляло 1 с, размер шага составлял 0,5 нм, и ширина полос составляла 5 нм (возбуждение) и 5 нм (испускание), соответственно.
Анализ вторичной структуры. Спектры циркулярного дихроизма (CD) в дальнем УФ получали, используя спектрополяриметр Jasco 715, оснащенный термографом (Jasco International Co. Ltd., Tokyo, Япония). Спектры получали при концентрации белка 150 мкг/мл в 10 мМ Tris/HCl (рН 8,0) в кварцевой кювете с длиной пути 0,1 см при 20°С. Скорость сканирования составляла 20 нм/мин, размер шага составлял 0,2 нм, время интегрирования было установлено на 1 с, и ширина полосы составляла 1 нм. Четыре сканирования усредняли и исправляли для буфера.
Анализ агрегации. Для тестирования растворимости белков при разных значениях рН и при разных концентрациях фосфата, соответственно, лиофилизированный белок растворяли в 6 М GdmSCN и диализировали против 10 мМ Tris/HCl (рН 8,0) при 4°С. Все пробы инкубировали при комнатной температуре в течение 2 часов в разных буферах (смотрите ниже), белковые осадки удаляли из проб седиментацией при 125000×g в течение 30 мин, и количество остающегося растворимого белка определяли фотометрически. Поскольку сумма растворимого и агрегированного белка должна была быть равна исходному количеству растворимого белка, процентное содержание агрегированного белка могло быть рассчитано путем вычитания количества растворимого белка из первоначально использованного количества белка. В качестве контроля белки инкубировали в 10 мМ Tris/HCl (рН 8,0).
Влияние значений рН
Белковые растворы разводили 1:10 буферами, имеющими разные рН (Таблица 2). Конечные концентрации белка составляли 0,2 мг/мл в случае NHCYS3C16 и CCC16 и 0,174 мг/мл в случае С16СC.
Использованные буферные системы с разными значениями рН
Влияние концентрации фосфата
Белковые растворы разводили 1:5 в КхНхРO4 (рН 8,0). Конечные концентрации белка составляли 0,4 мг/мл и концентрации фосфата составляли 50 мМ, 100 мМ, 200 мМ, 300 мМ и 500 мМ.
Связывание родаминмалеимида с тиольными группами. Для того чтобы связать маленькое органическое соединение с модифицированными белками шелка пауков, флуоресцентный краситель родамин связывали с белком NHCYS3C16. Маточный раствор 1 мМ родамина RedTM C2 малеимида (Molecular Probes, Leiden, Нидерланды) в ДМСО добавляли к раствору белка в 10 мМ Tris/HCl (pH 7,5) с получением молярного избытка 20. Данную реакцию проводили в темноте в течение ночи при 4°С. Для инактивации оставшегося несвязанным флуоресцентного красителя к реакционной смеси до гель-фильтрации (колонки PD10, Sephadex G 25, Pharmacia Biotech, Uppsala, Швеция) добавляли 100 мМ 2-меркаптоэтанол. Фракции собирали, тестировали на присутствие белка УФ-спектрометрией и, наконец, анализировали SDS-PAGE. Визуализацию белков и флуорофора проводили окрашиванием серебром и флуоресцентной визуализацией с использованием установки для визуализации с переменным режимом Typhoon 9200 (Molecular Dynamics, Amersham Pharmacia Biotech, Uppsala, Швеция) с длиной волны возбуждения 532 нм и длиной волны испускания 580 нм, соответственно.
РЕЗУЛЬТАТЫ
Конструирование, синтез и очистка модифицированных синтетических шелков пауков. Создавали разные модифицированные модули, основанные на модуле С (SEQ ID NO: 3) синтетического шелка пауков, который получен из белка канатного шелка ADF-4 из паука-крестовика Araneus diadematus. Данные модификации содержат один цистеин в каждом из вариантов, либо в TAG, либо в модуле С. В нем серин в положении 25 мутировали до цистеина вследствие аналогичного размера и полярности обеих аминокислот. Дополнительно считали, что данное положение не оказывает слишком большого влияния на характеристики белка, так как прогнозы гидрофобности для этой мутации лишь слегка отличаются от модуля С. Образующийся модуль СC (SEQ ID NO: 10) получали с использованием ПЦР-мутагенеза (смотрите экспериментальные методики). На следующей стадии нуклеотидную последовательность модуля СC клонировали вверх по течению или вниз по течению относительно нуклеотидной последовательности, кодирующей C16, получая белки CCC16 и C16CC, используя вектор клонирования pAZL (SEQ ID NO: 29). Дополнительно олигонуклеотидную последовательность, кодирующую TAG, состоящий из глицина, серина и одного цистеина (NHCYS3, SEQ ID NO: 22), клонировали в 5'-концевой нуклеотидной последовательности, кодирующей C16, с получением NHCYS3C16.
После бактериального синтеза модифицированные белки шелка очищали стадией нагревания с последующим осаждением сульфатом аммония. Идентичность данных белков подтверждали иммуноблоттингом, используя антитела, направленные против последовательностей Т7 пептидных меток, присоединенных к аминотерминальному концу всех белков шелка. Для сравнения использовали немодифицированный белок C16. Помимо белков с полной длиной наблюдали следы белков с меньшей молекулярной массой. В отличие от C16 все модифицированные белки, каждый из которых содержал один цистеин, показывали дополнительную белковую полосу с большей молекулярной массой (Фиг.3А и Б «-»), которая могла быть удалена добавлением к пробам восстанавливающих агентов, подобных 2-меркаптоэтанолу (Фиг.3Б «+»). Следовательно, они представляют собой белковые димеры, связанные дисульфидными мостиками. Чистоту белка определяли путем измерения испускания флуоресценции. Падающий свет с длиной волны 275 нм приводит к возбуждению и испусканию флуоресценции остатками тирозина и триптофана. Свет с длиной волны 295 нм возбуждает исключительно остатки триптофана. Поскольку ни один из сконструированных белков шелка пауков не содержал остатков триптофана, испускание флуоресценции при возбуждении 295 нм указывало бы на загрязняющие белки Е.соli, которые в среднем содержат 1,5% триптофана (4). Измерения флуоресценции всех препаратов модифицированного белка шелка выявили спектры испускания, похожие на спектр тирозина, который в больших количествах присутствует в белках шелка. Напротив, не могли определить флуоресценцию триптофана, что указывает на высокую чистоту данных белковых препаратов (Фиг.3В). Выходы чистых белков варьировали от 12 до 30 мг на литр культуральной среды.
Модифицированные C16 шелка пауков показывают ту же самую вторичную структуру, что и немодифицированные C16. Вторичную структуру исследовали CD-спектроскопией. Все модифицированные белки показывали такие же спектры, как C16, который демонстрирует спектр, типичный для внутренне неструктурированных белков (Фиг.4).
Модифицированные C16 шелка пауков являются более чувствительными к фосфату и рН, чем немодифицированные C16. pH, ионы, такие как калий и фосфат, и механическое напряжение участвуют в сборке природного шелка. Авторы данного изобретения исследовали влияние разных концентраций фосфата калия и разных рН на характер агрегации модифицированных белков CCC16, C16CC и NHCYS3C16 по сравнению с C16. Все модифицированные белки показывали значительную агрегацию (>10%) при значениях рН менее 5,0, CCC16 - даже при значениях менее 7,0 и показывал более чем 70%-ную агрегацию при рН 1. C16, напротив, показывал только умеренную агрегацию при этих условиях (Фиг.5А). Аналогично, фосфат калия приводил к агрегации модифицированных белков при более низких концентрациях, чем концентрации, наблюдаемые в случае с C16 (Фиг.5Б). Эту большую чувствительность для агрегации модифицированных белков нельзя объяснить другим зарядом белков, так как их теоретическая изоэлектрическая точка является идентичной (NHCYS3C16) изоэлектической точке C16 или отличается очень слабо (3,45 для CCC16 и 3,48 для С16). Следовательно, этот эффект может быть обусловлен присутствием цистеина и возможностью образовать стабильные димеры.
Сборка модифицированных синтетических белков шелка пауков
Синтетические белки шелка пауков, происходящие из последовательностей шелка пауков ADF-3 и ADF-4, можно собрать в морфологически отличающиеся формы, подобные сферам, нанофибриллам, пенам и пленкам. Провели следующие эксперименты для демонстрации того, что модифицированные белки шелка пауков показывают те же самые свойства, касающиеся отличных характеристик сборки.
1. Сферы
Белковые сферы, демонстрирующие диаметры, варьирующие от 0,3 до 1,5 мкм (Фиг.6А), генерировали путем добавления 2,4 М сульфата аммония к 1 мг/мл раствору СCС16, С16СC, NHCYS3C16 и C16 в 10 мМ Трис-(гидроксиметил)-аминометане (Tris) рН 8,0. Не наблюдали значимых различий между сферами, изготовленными из модифицированных белков или C16.
2. Нанофибриллы
Нанофибриллы формировали путем инкубации растворов С16СC и NHCYS3C16 в 10 мМ Трис рН 8,0 при 4°С в течение нескольких недель с последующей инкубацией при комнатной температуре в течение 3 суток (Фиг.6Б).
3. Пленки
Пленки, изготовленные из синтетических белков шелка пауков, происходящих из белка канатного шелка ADF-4 из паука-крестовика Araneus diadematus, могут быть отлиты из гексафтор-2-пропанола (HFIP) или из муравьиной кислоты (5). Лиофилизированные белки непосредственно растворяли в HFIP или в муравьиной кислоте. Как наблюдалось и с C16, HFIP индуцирует увеличение содержания вторичной структуры модифицированных белков CCC16, С16СC и NHCYS3C16. В то время как CD-спектры этих белков в 10 мМ Tris (pH 8,0) показывали только один минимум при длине волны менее 200 нм (Фиг.4), что свидетельствует главным образом о белке со случайной спиралью, CD-спектры растворов белков в HFIP демонстрировали минимум при 202-203 нм и дополнительное плечо при 220 нм, которое свидетельствует о повышенном содержании α-спирали (Фиг.7).
Из модифицированных белков шелка пауков можно было отлить пленки из HFIP, а также из муравьиной кислоты. Пленки отливали на полистирольной поверхности, откуда их можно было легко снять после выпаривания растворителя (Фиг.6В). Для того чтобы проанализировать вторичную структуру данных пленок, 0,01% масс./об. раствор белка отливали на кварцевом стекле. После выпаривания растворителя измеряли циркулярный дихроизм. Пленки, отлитые из HFIP, показывали спектры с двумя минимумами при 208 нм и 220 нм, свидетельствующие о высоком содержании α-спирали (Фиг.7). Для того чтобы сделать пленки, отлитые из HFIP, водонерастворимыми (литые пленки легко растворяются при контакте с водой), их обрабатывали метанолом (5). После этой обработки спектр показал одиночный минимум при 218 нм, который является типичным для структуры, обогащенной β-складками (в качестве примера показано для NHCYS3C16 на Фиг.7). Это изменение во вторичной структуре при обработке пленок уже было описано для пленок из синтетических шелков пауков (5). Интересно, что пленки, отлитые из муравьиной кислоты, отличались по их вторичной структуре в зависимости от используемого белка. В то время как NHCYS3C16 давал пленку, демонструющую структуру, обогащенную β-складками (с минимумом в CD спектре при 218 нм), подобно тому, как это было описано для C16 (5), спектр пленки из CCC16 показывал два минимума при 208 нм и 220 нм. Пленки из белка C16CC показывали CD-спектры с одним минимумом при 200 нм.
4. Мечение модифицированных синтетических шелков пауков
Модификация синтетических шелков пауков цистеином или лизином должна делать возможным специфическое связывание лекарственных средств, металлов, полипептидов, квантовых точек и так далее. Для того чтобы продемонстрировать связывание SH-группы цистеина с маленькими органическими молекулами, флуорофор родамин связывали с белком NHCYS3C16. Родамин использовали в форме, конъюгированной с малеимидом, который легко и очень специфично реагирует с SH-группами при рН 7,0-7,5. Эффективное связывание визуализировали SDS-PAGE с последующей флуоресцентной визуализацией и окрашиванием серебром (Фиг.8).
ССЫЛКИ
1. Sambrook, J. and Russell, D. (2001,) Molecular cloning.
2. Kroll, D.J., Abdel-Malek Abdel-Hafiz, H., Marcell, T., Simpson, S., Chen, C.Y., Gutierrez-Hartmann, A., Lustbader, J.W., and Hoeffler, J.P. (1993) A multifunctional prokaryotic protein expression system: overproduction, affinity purification, and selective detection, DNA Cell Biol. 12, 441-453.
3. Gill, S.C. and von Hippel, P.H. (1989) Calculation of Protein Extinction Coefficients from Amino-Acid Sequence Data, Analytical Biochemistry 182, 319-326.
4. Blattner, F.R, Plunkett, G., III, Bloch, C.A., Perna, N.Т., Burland, V., Riley, M., Collado-Vides, J., Glasner, J.D., Rode, C.K., Mayhew, G.F., Gregor, J., Davis, N.W., Kirkpatrick, H.A., Goeden, M.A., Rose, D.J., Маu, В., and Shao, Y. (1997) The complete genome sequence of Escherichia coli K-12, Science 277, 1453-1474.
5. Slotta, U., Tammer, M., Kremer, F., Koelsch, P., Scheibel, T. (2006) Structural analysis of spider silk films, Supromolecular chemistry 18, 465-471.











Изобретение относится к области биотехнологии и может быть использовано для получения рекомбинантных белков шелка пауков. Рекомбинантным путем получают белки, состоящие из модуля SEQ ID NO: 3, содержащего замены серина и/или глутамата на лизин и/или цистеин, и/или добавления аминоконцевого TAG согласно SEQ ID NO: 20-24 или карбоксиконцевого TAG согласно SEQ ID NO: 25-28, содержащих лизин или цистеин наряду с другими аминокислотами. Изобретение позволяет получить модифицированный белок шелка пауков, обладающий способностью связываться с другими веществами через остаток лизина и/или цистеина с сохранением структурной целостности белка шелка пауков. 5 н. и 8 з.п. ф-лы, 8 ил., 2 табл.
1. Способ модификации белка шелка пауков, включающий стадии:
а) получения последовательности нуклеиновой кислоты, кодирующей белок шелка пауков, который содержит один модуль SEQ ID NO: 3 или его вариант, где вариант содержит 1 аминокислотную замену, 1-2 аминокислотных делеций и/или 1-2 аминокислотных вставок, и который не содержит остатков лизина или цистеина;
б) замены последовательности нуклеиновой кислоты, кодирующей серии и/или глутамат в указанном модуле указанного белка шелка пауков, последовательностью нуклеиновой кислоты, кодирующей лизин и/или цистеин, и/или добавления к указанной последовательности последовательности нуклеиновой кислоты, кодирующей аминоконцевой TAG согласно SEQ ID NO:20-24, содержащий лизин или цистеин наряду с другими аминокислотами, или карбоксиконцевой TAG согласно SEQ ID NO:25-28, содержащий лизин или цистеин наряду с другими аминокислотами;
в) проведения экспрессии модифицированной последовательности нуклеиновой кислоты, полученной в (б), в подходящем хозяине и
г) выделения экспрессированного модифицированного белка шелка пауков.
2. Способ по п.1, который дополнительно включает связывание других веществ с указанной молекулой лизина и/или цистеина в модифицированном белке шелка пауков.
3. Способ по п.2, где вещество, подлежащее связыванию с указанным лизином и/или цистеином, представляет собой маркерную молекулу.
4. Способ по п.1, где белок шелка пауков, кодируемый последовательностью нуклеиновой кислоты, полученной в (а), содержит C16 или С32.
5. Способ по п.1, где модифицированный белок шелка пауков, выделенный на стадии (г), содержит один модуль с SEQ ID NO: 10-13.
6. Способ по п.1, где к модифицированному белку шелка пауков, выделенному на стадии (г), добавляют аминоконцевой TAG согласно SEQ ID NO: 20-24, содержащий лизин или цистеин наряду с другими аминокислотами, или карбоксиконцевой TAG согласно SEQ ID NO: 25-28, содержащий лизин или цистеин наряду с другими аминокислотами.
7. Модифицированный белок шелка пауков, подходящий для направленного связывания с веществами, получаемый способом по любому из пп.1-6.
8. Модифицированный белок шелка пауков по п.7, содержащий один модуль SEQ ID NO: 10-13.
9. Последовательность нуклеиновой кислоты, кодирующая модифицированный белок шелка пауков по п.7 или 8.
10. Вектор экспрессии, который содержит последовательность нуклеиновой кислоты по п.9 и одну и более регуляторных последовательностей.
11. Вектор экспрессии по п.10, который представляет собой плазмидный или вирусный вектор, предпочтительно бакуловирусную систему или векторную систему на основе вируса коровьей оспы.
12. Клетка-хозяин, трансформированная вектором экспрессии по п.10 или 11 и подходящая для продуцирования модифицированного белка шелка пауков по п.7 или 8.
13. Клетка-хозяин по п.12, которая представляет собой прокариотическую клетку.
14. Клетка-хозяин по п.13, которая представляет собой E.coli или Bacillus subtilis.
15. Клетка-хозяин по п.12, которая представляет собой эукариотическую клетку.
16. Клетка-хозяин по п.15, которая представляет собой клетку млекопитающего, растительную клетку, дрожжевую клетку или клетку насекомого.
17. Клетка-хозяин по п.16, где клетка млекопитающего представляет собой клетку СНО (яичника китайского хомячка), COS, HeLa, 293T, НЕН или ВНК (почки детеныша хомяка).
18. Клетка-хозяин по п.16, где дрожжевая клетка представляет собой Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris, Candida albicans или Hansenula polymorpha.
19. Клетка-хозяин по п.16, где клетка насекомого выбрана из клеток насекомых Lepidoptera, предпочтительно из Spodoptera frugiperda и из Trichoplasia ni.
20. Клетка-хозяин по п.19, где клетка насекомого представляет собой Sf9, Sf21 или клетку "high five".
21. Клетка-хозяин по п.16, где растительная клетка происходит из табака, картофеля, кукурузы, гороха и томата.
| SCHEIBEL Т | |||
| Spider silks: recombinant synthesis, assembly, spinning, and engineering of synthetic proteins, Microb | |||
| Cell Fact., 2004, v.3, p.1-10 | |||
| ZHOU Y., Genetically directed synthesis and spectroscopic analysis of a protein polymer derived from a flagelliform silk sequence, Biomacromolecules, 2001, v.2, n.1, p.111-125 | |||
| RU 95109941 A1, 10.01.1997. |

