ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области рекомбинантного получения белков. Более подробно, настоящее изобретение относится к рекомбинантному получению белков шелка паука. В настоящем изобретении предоставляются новые выделенные спидроиновые белки главной ампулярной железы и слитые белки главной ампулярной железы, а также способы получения таких белков и молекулы полинуклеиновых кислот для их получения. Кроме того, предоставляются полимеры спидроиновых белков главной ампулярной железы и способы получения таких полимеров.
УРОВЕНЬ ТЕХНИКИ
Паучьи шелка представляют собой природные полимеры с высокими характеристиками, обладающие исключительной крепкостью благодаря комбинации прочности и эластичности. Пауки имеют до семи специализированных желез, которые вырабатывают множество видов шелковой нити с различными механическими свойствами и функциями. Канатный шелк, вырабатываемый главной ампулярной железой, представляет собой очень прочное волокно и в весовом соотношении оно превосходит созданные человеком материалы, такие как высокопрочная сталь и кевлар. Свойства канатного шелка являются привлекательными для создания новых материалов для медицинских или технических целей.
Канатный шелк состоит из двух основных полипептидов, обычно называемых спидроинами главной ампулярной железы (MaSp) 1 и 2, но полипептиды из Araneus diadematus называются ADF-3 и ADF-4. Указанные белки имеют кажущуюся молекулярную массу в диапазоне 200-720 кДа, в зависимости от возраста образца и условий анализа, но не было публикаций о полноразмерном гене канатного шелка паука. Свойства полипептидов канатного шелка описаны в Huemmerich, D. et al. Novel assembly properties of recombinant spider dragline silk proteins. Curr. Biol. 14, 2070-2074 (2004). Известные спидроины канатного шелка состоят из многократно повторяющихся блоков чередующихся сегментов, богатых аланином, которые образуют кристаллические β-листы в волокне, и сегментов, богатых глицином, которые являются более гибкими и в основном лишены упорядоченной структуры. С-концевой участок не имеет повторов, является высоко консервативным в разных видах и принимает конформацию α-спирали. N-концевой участок белков канатного шелка был охарактеризован совсем недавно, причем показано, что N-концевой участок является высоко консервативным в различных спидроинах и также у различных видов пауков (Rising, A. et al. N-terminal nonrepetitive domain common to dragline, flagelliform, and cylindriform spider silk proteins. Biomacromolecules 7, 3120-3124 (2006)).
Механические свойства канатного шелка отличаются у различных видов пауков; канатный шелк Euprosthenops sp. является более жестким, прочным (требует большего усилия для разрыва) и менее гибким, чем канатный шелк, например Araneus diadematus или Nephila clavipes. Канатный шелк Euprosthenops sp., по-видимому, содержит большую количественную долю кристаллической β-листовой структуры, чем канатный шелк Araneus diadematus, скорее всего по этой причине MaSp из Euprosthenops sp. имеет самое высокое содержание полиаланина среди всех исследованных до настоящего времени видов (Pouchkina-Stantcheva, N. N. & McQueen-Mason, S. J. Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp. (Pisauridae). Comp Biochem Physiol B Biochem Mol Biol 138, 371-376 (2004)).
В опытах по получению искусственных волокон паутины использовались фрагменты натуральных или синтетических генов, кодирующих белки канатного шелка, потому что полноразмерный ген еще не описан. Рекомбинантные белки канатного шелка экспрессированы в различных системах, включая бактерии, дрожжи, клетки млекопитающих, растения, клетки насекомых, трансгенного тутового шелкопряда и трансгенных коз. Для примера, см. Lewis, R. V. et al. Expression and purification of a spider silk protein: a new strategy for producing repetitive proteins. Protein Expr. Purif. 7, 400-406 (1996); Fahnestock, S. R. & Irwin, S. L. Synthetic spider dragline silk proteins and their production in Escherichia coli. Appl. Microbiol. Biotechnol. 47, 23-32 (1997); Arcidiacono, S. et al. Purification and characterization of recombinant spider silk expressed in Escherichia coli. Appl. Microbiol. Biotechnol. 49, 31-38 (1998); Fahnestock, S. R. & Bedzyk, L. A. Production of synthetic spider dragline silk protein in Pichia pastoris. Appl. Microbiol. Biotechnol. 47, 33-39 (1997); and Lazaris, A. et al. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 295, 472-476 (2002).
В международном патенте WO 2004/016651 (Университет Йорка) раскрыты последовательности нуклеиновых кислот, кодирующих внутренние повторяющиеся части белков MaSp1 из Euprosthenops sp. Белок не экспрессирован.
В публикации Huemmerich, D. et al. Primary structure elements of spider dragline silks and their contribution to protein solubility. Biochemistry 43, 13604-13612 (2004) раскрывается искусственный ген, "(AQ)12NR3", кодирующий повторяющиеся фрагменты, богатые Ala, и фрагменты, богатые Gly/Gln, и неповторяющийся фрагмент, которые получены из ADF3 Araneus. Ген экспрессирован с образованием растворимого белка (59,8 кДа, >528 aa), который агрегирует, но не образует полимеров или волокон. Содержание аланина в белке составляет 10-15%.
В международном патенте WO 03/057727 раскрыта экспрессия растворимых рекомбинантных полипептидов шелка в клеточных линиях млекопитающих и животных. Один экспрессированный полипептид шелка (ADF-3; 60 кДа, 652 aa) состоит из повторяющейся единицы и неповторяющегося гидрофильного домена. Другой экспрессированный полипептид шелка (ADF-3 His; 63 кДа, 677 aa) состоит из повторяющейся единицы, неповторяющегося гидрофильного домена, эпитопа c-myc и тага из шести гистидинов. Повторяющаяся единица имеет низкое содержание Ala (10-20%). Полученные полипептиды шелка имеют низкую растворимость в водных средах и/или образуют преципитаты. Поскольку полученные полипептиды шелка не полимеризуются самопроизвольно, то для получения полимеров или волокон необходимо формование.
Несколько факторов затрудняют экспрессию белков канатного шелка. Из-за многократной повторяемости генов, которая сопровождается ограниченным аминокислотным составом белков, происходят ошибки транскрипции и трансляции. Другой причиной может быть истощение пулов тРНК в микробных системах экспрессии с последующей разобщенной трансляцией, приводящей к преждевременной терминации синтеза белка. В качестве других причин прерывания синтеза белка обсуждаются строение вторичной структуры мРНК и рекомбинация генов. Показано, что нативные гены MaSp длиной больше чем 2,5 т.п.н. являются нестабильными в бактериальных хозяевах. Кроме того, существуют сложности в поддержании рекомбинантных белков шелка в растворимой форме, поскольку как фрагменты канатного шелка природного происхождения, так и созданные блочные сополимеры, особенно белки, полученные из MaSpl/ADF-4, легко образуют аморфные агрегаты, вызывающие осаждение и потерю белка. См. Huemmerich, D. et al. Primary structure elements of spider dragline silks and their contribution to protein solubility. Biochemistry 43, 13604-13612 (2004) and Lazaris, A. et al. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 295, 472-476 (2002).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предоставление нового белка шелка паука, из которого могут быть получены волокна шелка паука.
Другой целью настоящего изобретения является предоставление водорастворимого белка шелка паука, который при простом воздействии самополимеризуется в волокна. Это дает возможность для уникальных применений, таких как культивирование эукариотических клеток на волокнах. Кроме того, данное свойство делает возможным проведение всех последующих стадий при физиологических условиях, что снижает риск токсичности и денатурации белка.
Целью настоящего изобретения также является предоставление волокон нового белка шелка паука.
Одной целью настоящего изобретения является предоставление белков шелка паука в большом количестве, которые после простой обработки могут легко самополимеризоваться с образованием волокна.
Также целью изобретения является предоставление способов получения белков шелка и волокон белков шелка паука.
Для данных и других целей, которые становятся очевидными из последующего раскрытия, в настоящем изобретении по одному аспекту предоставляется выделенный спидроиновый белок главной ампулярной железы, где белок состоит из 150-420 аминокислотных остатков и описывается формулой REP-CT, где REP представляет собой фрагмент белка, имеющий 80-300 аминокислотных остатков, где указанный фрагмент выбран из группы L(AG) n L, L(AG) n AL, L(GA) n L, L(GA) n GL, где n представляет собой целое число от 4 до 8; каждый индивидуальный сегмент A представляет собой аминокислотную последовательность из 8-18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala; каждый индивидуальный сегмент G является аминокислотной последовательностью из 12-30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков являются Gly; и каждый индивидуальный сегмент L является линкерной аминокислотной последовательностью из 0-20 аминокислотных остатков; и CT представляет собой фрагмент белка, имеющий от 70 до 120 аминокислотных остатков, фрагмент которого является C-концевым фрагментом, полученным из спидроинового белка главной ампулярной железы или из его производных.
Настоящее изобретение основано на идентификации белкового мотива, которого достаточно для формирования волокон, подобных шелку, и на применении указанного мотива для построения рекомбинантных белков MaSp, которые возможно вырабатывать в подходящих хозяевах, таких как бактерии, предпочтительно E. coli.
В определенных воплощениях согласно изобретению каждый индивидуальный сегмент А имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 7-19, 43-56, 71-83, 107-120, 135-147, 171-183, 198-211, 235-248, 266-279, 294-306, 330-342, 357-370, 394-406, 421-434, 458-470, 489-502, 517-529, 553-566, 581-594, 618-630, 648-661, 676-688, 712-725, 740-752, 776-789, 804-816, 840-853, 868-880, 904-917, 932-945, 969-981, 999-1013, 1028-1042 и 1060-1073 SEQ ID NO: 3; аминокислотных остатков 31-42, 61-75, 90-104, 122-135 и 153-171 SEQ ID NO: 9; аминокислотных остатков 12-25, 46-60, 75-88, 112-119, 150-158 и 173-180 SEQ ID NO: 13; аминокислотных остатков 31-42 SEQ ID NO: 14; и аминокислотных остатков 122-135 SEQ ID NO: 15. В специфических воплощениях каждый индивидуальный сегмент А является аминокислотной последовательностью, выбранной из данной группы аминокислотных последовательностей.
В некоторых воплощениях согласно изобретению каждый индивидуальный сегмент G имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 20-42, 57-70, 84-106, 121-134, 148-170, 184-197, 212-234, 249-265, 280-293, 307-329, 343-356, 371-393, 407-420, 435-457, 471-488, 503-516, 530-552, 567-580, 595-617, 631-647, 662-675, 689-711, 726-739, 753-775, 790-803, 817-839, 854-867, 881-903, 918-931, 946-968, 982-998, 1014-1027, 1043- 1059 и 1074-1092 SEQ ID NO: 3; SEQ ID NO: 5; SEQ ID NO: 6; SEQ ID NO: 7; аминокислотных остатков 11-30, 43-60, 76-89, 105-121 и 136-152 SEQ ID NO: 9; и аминокислотных остатков 1-11, 26-45, 61-74, 89-111, 120-149 и 159-172 SEQ ID NO: 13. В специфических воплощениях каждый индивидуальный сегмент G является идентичным аминокислотной последовательности, выбранной из данной группы аминокислотных последовательностей.
В определенных воплощениях согласно изобретению указанный фрагмент СТ имеет по меньшей мере 50% идентичности с SEQ ID NO: 8 или по меньшей мере 80% идентичности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4, аминокислотных остатков 172-269 SEQ ID NO: 9, аминокислотных остатков 181-276 SEQ ID NO: 13, и аминокислотных остатков 172-269 SEQ ID NO: 16, а также с любой аминокислотной последовательностью фигуры 3, особенно с последовательностями MaSp1 фигуры 3. В определенных воплощениях указанный фрагмент СТ представляет собой аминокислотную последовательность, выбранную из данной группы аминокислотных последовательностей.
В определенных воплощениях согласно настоящему изобретению содержание липополисахаридов (LPS) и других пирогенов в выделенном спидроиновом белке главной ампулярной железы составляет 1 единицу эндотоксина (ЭЕ)/мг белка или меньше.
Согласно другому аспекту в настоящем изобретении предоставляется выделенный слитый белок, состоящий из первого белкового фрагмента, который представляет собой спидроиновый белок главной ампулярной железы и второго белкового фрагмента, отличающегося тем, что указанный второй белковый фрагмент содержит партнер по слитию и участок узнавания расщепляющего агента, где указанный первый белковый фрагмент соединяется через указанный участок узнавания расщепляющего агента с указанным партнером по слитию.
В настоящем изобретении предоставляется выделенный слитый белок, выбранный из группы X-REP-CT, и REP-CT-X, где REP и CT являются белковыми фрагментами согласно изобретению; и X является белковым фрагментом, содержащим партнер по слитию и участок узнавания расщепляющего агента; где комбинированный белковый фрагмент REP-CT соединяется через указанный участок узнавания расщепляющего агента с указанным партнером по слитию.
В определенных воплощениях согласно изобретению содержание LPS и других пирогенов в выделенном спидроиновом белке составляет 1 ЭЕ/мг белка или меньше.
Согласно еще одному аспекту в настоящем изобретении предоставляется способ получения спидроинового белка главной ампулярной железы согласно изобретению, включающий в себя стадии: (i) предоставления раствора слитого белка согласно изобретению в жидкой среде, (ii) добавления к указанной жидкой среде подходящего расщепляющего агента для расщепления слитого белка в участке узнавания расщепляющего агента и получения благодаря этому спидроинового белка главной ампулярной железы; и, если надо, (iii) выделения спидроинового белка главной ампулярной железы, полученного на стадии (ii) из указанной жидкой среды.
Настоящее изобретение также предоставляет способ получения полимера спидроинового белка главной ампулярной железы согласно изобретению, включающий в себя стадии: (i) предоставления раствора слитого белка согласно изобретению в жидкой среде, (ii) добавления к указанной жидкой среде подходящего расщепляющего агента для расщепления слитого белка в участке узнавания расщепляющего агента, и получения благодаря этому спидроинового белка главной ампулярной железы; (iii) вызывания полимеризации спидроинового белка главной ампулярной железы, полученного на стадии (ii) в жидкой среде; и, если надо, (iv) выделения полимера, полученного на стадии (iii) из указанной жидкой среды. В предпочтительном способе указанная стадия (iii) далее включает в себя создание границы раздела между указанной жидкой средой и другой фазой, выбранной из группы, состоящей из газовой фазы, жидкой фазы и твердой фазы, где указанная полимеризация вызывается в указанной границе раздела или в области, окружающей границу раздела. В предпочтительном способе указанная жидкая среда является водной средой и указанную другую фазу выбирают из группы, состоящей из органических растворителей, несмешиваемых с воздухом и водой.
Согласно другому аспекту в настоящем изобретении предоставляется выделенная молекула полинуклеиновой кислоты, включающая в себя последовательность нуклеиновой кислоты, которая кодирует спидроиновый белок главной ампулярной железы согласно изобретению или комплементарную ей последовательность нуклеиновой кислоты.
Согласно еще одному аспекту в настоящем изобретении предоставляется выделенная молекула полинуклеиновой кислоты, включающая в себя последовательность нуклеиновой кислоты, которая кодирует слитый белок согласно изобретению или комплементарную ей последовательность нуклеиновой кислоты.
Другой аспект изобретения относится к способу получения растворимого слитого белка согласно изобретению, который включает в себя стадии: (i) экспрессии молекулы полинуклеиновой кислоты, кодирующей растворимый слитый белок согласно изобретению, в подходящем хозяине; и (ii) выделения растворимого слитого белка, полученного на стадии (i). Если надо, указанная стадия (ii) выделения растворимого слитого белка включает в себя удаление LPS и других пирогенов.
В настоящем изобретении также предоставляется способ производства спидроинового белка главной ампулярной железы согласно изобретению, включающий в себя стадии: (i) экспрессии молекулы полинуклеиновой кислоты, кодирующей растворимый слитый белок согласно изобретению, в подходящем хозяине; и (ii) выделения растворимого слитого белка, полученного на стадии (i); (iii) предоставления раствора указанного растворимого слитого белка, полученного на стадии (ii) в жидкой среде, (iv) добавления к указанной жидкой среде подходящего расщепляющего агента для расщепления слитого белка в сайте узнавания расщепляющего агента, и получения благодаря этому спидроинового белка главной ампулярной железы; и, если надо, (v) выделения спидроинового белка главной ампулярной железы, полученного на стадии (iv) из указанной жидкой среды. Далее по желанию указанная стадия (ii) выделения растворимого слитого белка и по желанию указанная стадия (v) выделения спидроинового белка главной ампулярной железы включают в себя удаление LPS и других пирогенов.
Далее в настоящем изобретении предоставляется способ получения полимера спидроинового белка главной ампулярной железы согласно изобретению, включающий в себя стадии: (i) экспрессии молекулы полинуклеиновой кислоты, кодирующей растворимый слитый белок согласно изобретению, в подходящем хозяине; и (ii) выделения растворимого слитого белка, полученного на стадии (i); (iii) предоставления раствора указанного растворимого слитого белка, полученного на стадии (ii) в жидкой среде, (iv) добавления к указанной жидкой среде подходящего расщепляющего агента для расщепления слитого белка в сайте узнавания расщепляющего агента, и получения благодаря этому спидроинового белка главной ампулярной железы; (v) вызывание полимеризации спидроинового белка главной ампулярной железы, полученного на стадии (iv) в жидкой среде; и, если надо, (vi) выделение полимера, полученного на стадии (v) из указанной жидкой среды. В предпочтительном способе указанная стадия (v) далее включает в себя создание границы раздела между указанной жидкой средой и другой фазой, выбранной из группы, состоящей из газовой фазы, жидкой фазы и твердой фазы, где указанная полимеризация вызывается в указанной границе раздела или в области, окружающей границу раздела. В предпочтительном способе указанная жидкая среда является водной средой, и указанная другая фаза выбрана из группы, состоящей из органических растворителей, несмешиваемых с воздухом и с водой.
Согласно другому аспекту в настоящем изобретении предоставляется полимер спидроинового белка главной ампулярной железы согласно изобретению. В настоящем изобретении также предоставляется полимер спидроинового белка главной ампулярной железы, полученный по способу согласно изобретению. В предпочтительном воплощении указанный полимер является волокном. В других предпочтительных воплощениях указанный полимер образует структуру, выбранную из группы, состоящей из пены, геля, сетки или пленки.
Согласно еще одному аспекту в настоящем изобретении предоставляется новое применение белкового фрагмента, включающего в себя партнера по слитию и участок узнавания расщепляющего агента для производства слитого белка, включающего в себя указанный белковый фрагмент, соединенный через указанный участок узнавания расщепляющего агента с фрагментом белка шелка паука. В предпочтительных воплощениях указанный фрагмент белка шелка паука состоит из 150-420 аминокислотных остатков.
Согласно последнему аспекту в настоящем изобретении предоставляется выделенная молекула полинуклеиновой кислоты, включающая в себя последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 1 и последовательностей нуклеиновых кислот, кодирующих SEQ ID NO: 2-16, или комплементарных им последовательностей нуклеиновых кислот. В настоящем изобретении предоставляется применение выделенной молекулы полинуклеиновой кислоты для производства искусственного гена, кодирующего белок шелка паука.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 показано выравнивание сегментов внутри повторяющейся части белка MaSp1 Euprosthenops australis, т.e. SEQ ID NO: 3.
На фигуре 2A проиллюстрирована в виде схемы предсказанная структурная организация повторяющейся части белка MaSp1 Euprosthenops australis (SEQ ID NO: 3).
На фигуре 2B проиллюстрированы в виде схем предсказанные структурные организации спидроиновых белков, построенных согласно примерам 5-8 (SEQ ID NO: 9-13).
На фигуре 3 показано выравнивание C-концевых участков MaSp1 и MaSp2, которое иллюстрирует их консервативную природу.
На фигуре 4 проиллюстрирован макроскопический внешний вид волокон, сформированных из спидроиновых белков, которые построены согласно примерам 5-8. (A): волокна белка 6Gly/Ala-CThyb (SEQ ID NO: 13), полоска 0,5 см; (B): волокна белка 5Gly/Ala-CTnat (SEQ ID NO: 9), полоска 1 см; (C): волокна белка 5Gly/Ala-CTnat (SEQ ID NO: 9), полоска 1 см.
На фигуре 5 представлены полученные с помощью сканирующего электронного микроскопа (SEM) микрофотографии волокон, сформированных из спидроиновых белков, которые построены согласно примерам 5-8. Одиночные волокна (а) и гель-фаза (b, с) из 6Gly/Ala-CThyb (SEQ ID NO: 13). Волокна 5Gly/Ala-CTnat (SEQ ID NO: 9), вытянутые в 75% метаноле, высушенные на воздухе и нанесенные на целики SEM (d, e, f). Волокно, скрученное перед высушиванием на воздухе (е); конец волокна (f).
На фигуре 6 показан спектр кругового дихроизма (CD) волокна 6Gly/Ala-CThyb (SEQ ID NO: 13).
На фигуре 7 проиллюстрированы результаты цитотоксического исследования на тучных клетках мышей, которые показывают число живых и мертвых клеток через три дня культивирования в присутствии и в отсутствии шелковых волокон, полученных in vitro.
На фигуре 9 представлена кривая зависимости между напряжением и деформацией, которая показывает прочность при растяжении дважды вытянутых волокон из 5Gly/Ala-CTnat (SEQ ID NO: 9).
На фигуре 10 представлены SEM-микрофотографии рекомбинантных волокон из 5Gly/Ala-CTnat (SEQ ID NO: 9). (a, b) Спонтанно образованные волокна. На изображении (b) крупным планом показана фибриллярная субструктура. Тонкая выступающая фибрилла (отмечена стрелкой) имеет ширину около 300 нм. (c-f) Волокна после двух циклов растяжения-расслабления. На (с) и (d) показано одно и то же волокно в различных увеличениях. На (е) показан срезанный конец волокна, и на (f) показана точка разрыва после теста на растяжение.
ПОДРОБНОЕ РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом основано на идентификации мотива спидроинового белка, который является достаточным для рекомбинантного получения волокон шелка паука. Мотив основан на расшифрованной аминокислотной последовательности в результате клонирования и секвенирования частичной кДНК спидроина 1 (MaSp1) главной ампулярной железы Euprosthenops australis. Из этого следует, что выделенная кДНК MaSp1 является полезной в качестве точки старта для построения новых генов спидроинов, таких как гены, представленные в настоящем описании. Полимеры, которые созданы из белков, полученных из новых спидроиновых кДНК, являются полезными благодаря своим физическим свойствам, и особенно полезной является комбинация высокой прочности, эластичности и малого веса. Они также являются полезными благодаря своей способности поддерживать клеточную адгезию и рост. Свойства канатного шелка являются привлекательными для разработки новых материалов для медицинских и технических целей. В частности, паучьи шелка согласно изобретению могут быть полезны в медицинских устройствах, таких как импланты, и в медицинских продуктах, таких как системы для ушивания ран, пластыри, шовный материал, повязки на рану, и в каркасных структурах для инженеринга тканей и для направленной клеточной регенерации. Паучьи шелка согласно изобретению также являются особенно полезными для применения в качестве ткани или материала, такого как материал для парашютов, для пуленепробиваемой одежды, для ремней безопасности и т.п.
Термин "волокно", использованный в настоящем описании, относится к полимерам, имеющим толщину, по меньшей мере, 1 мкм, предпочтительными являются макроскопические полимеры, которые видимы человеческим глазом, например, имеющие толщину, по меньшей мере, 1 мкм, и которые имеют значительное растяжение в длину, по сравнению с их толщиной, предпочтительно, приблизительно 5 мм. Термин "волокно" не включает в себя неструктурированные агрегаты или преципитаты.
Термины "спидроиновые белки главной ампулярной железы", "спидроиновые белки" используются взаимозаменяемо в описании и включают в себя все известные спидроиновые белки главной ампулярной железы, обычно обозначаемые аббревиатурой "MaSp" или "ADF" в случае Araneus diadematus. Данные спидроиновые белки главной ампулярной железы обычно бывают двух типов, 1 и 2. Указанные термины, кроме того, включают в себя новые белки согласно изобретению, как описано в приложенной формуле изобретения, и другие искусственные белки с высокой степенью идентичности и/или сходства с известными спидроиновыми белками главной ампулярной железы.
Авторы настоящего изобретения использовали идентифицированный мотив спидроинового белка для создания новых генных конструкций, кодирующих искусственные спидроиновые белки. Было обнаружено, что спидроиновый белок главной ампулярной железы, состоящий из 150-420 аминокислотных остатков, т.е. больше или равно 150, предпочтительно больше или равно 220, предпочтительно больше или равно 250, и меньше или равно 420, предпочтительно меньше или равно 380 аминокислотных остатков, предпочтительно меньше или равно 320 аминокислотных остатков, предпочтительно меньше или равно 280 аминокислотных остатков, а именно 220-360 аминокислотных остатков, может быть получен рекомбинантно, например, в бактериях или в других подходящих продукционных организмах. Полученные спидроиновые белки спонтанно образуют макроскопические шелковые волокна согласно изобретению. Это удивительный результат, природные спидроиновые белки и ранее известные рекомбинантные, образующие волокна спидроиновые белки являются значительно более длинными, чем белки согласно изобретению. Кроме того, природные спидроиновые белки и ранее известные рекомбинантные, образующие волокна спидроиновые белки, как правило, содержат большое число внутренних повторов и требуют центрифугирования и/или жестких растворителей для полимеризации.
В настоящем описании впервые показано, что спидроиновые белки могут спонтанно образовывать волокна in vitro. Данные, представленные в настоящем описании, также показывают, что для запуска образования волокон необходимо только присутствие фракции последовательности спидроина. Кроме того, видовой гибрид, содержащий повторяющийся домен Euprosthenops и неповторяющийся C-концевой домен Nephila (перенесенный в пример 6C) также образует волокна, проявляя сильный потенциал к образованию волокон у данного мотива.
В общем виде спидроиновый белок главной ампулярной железы согласно изобретению описывается формулой REP-CT. Белковый фрагмент REP и белковый фрагмент CT соединены ковалентно, обычно с помощью пептидной связи.
Белковый фрагмент REP имеет повторяющийся характер, с чередованием участков, богатых аланином и богатых глицином. Фрагмент REP обычно содержит более чем 80, а именно более чем 140 и менее чем 300, предпочтительно менее чем 240, а именно менее чем 200, аминокислотных остатков, и может быть разделен на несколько сегментов L (линкерных), сегментов A (богатых аланином) и сегментов G (богатых глицином), как будет подробно объяснено далее. Обычно указанные линкерные сегменты, которые являются произвольными, расположены на концах фрагмента REP, в то время как остальные сегменты представляют собой чередующиеся богатые аланином и богатые глицином сегменты. Таким образом, фрагмент REP может иметь в целом любую из следующих структур, где n является целым числом:
L(AG) n L, а именно LA 1 G 1 A 2 G 2 A 3 G 3 A 4 G 4 A 5 G 5 L;
L(AG) n AL, а именно LA 1 G 1 A 2 G 2 A 3 G 3 A 4 G 4 A 5 G 5 A 6 L;
L(GA) n L, а именно LG 1 A 1 G 2 A 2 G 3 A 3 G 4 A 4 G 5 A 5 L; или
L(GA) n GL, а именно LG 1 A 1 G 2 A 2 G 3 A 3 G 4 A 4 G 5 A 5 G 6 L.
Из этого следует, что не имеет особой важности, какой из двух сегментов, богатый аланином или богатый глицином, граничит с N-концевыми или C-концевыми линкерными сегментами. Предпочтительно n является целым числом от 4 до 8, более предпочтительно от 4 до 6, например n=4, n=5 или n=6.
В предпочтительных воплощениях содержание аланина в фрагменте REP согласно изобретению составляет выше 20%, предпочтительно выше 25%, наиболее предпочтительно выше 30%, и ниже 50%, предпочтительно ниже 40%, более предпочтительно ниже 35%. Это является преимуществом, поскольку предполагается, что более высокое содержание аланина дает более жесткое, и/или более крепкое, и/или менее растяжимое волокно. Причиной этого явления по-видимому является то, что более высокое содержание аланина ассоциировано с более высоким содержанием β-листовых структур в волокне. Таким образом, в предпочтительном воплощении, содержание β-листовых структур в полимере, таком как волокно, спидроинового белка главной ампулярной железы согласно изобретению составляет более 50%, т.е. более чем 50% вторичной структуры белка имеет β-листовую форму.
В определенных воплощениях фрагмент REP лишен остатков пролина, т.е. во фрагменте REP нет остатков Pro.
Далее, обращаясь к сегментам, из которых построен фрагмент REP согласно изобретению, следует подчеркнуть, что каждый сегмент является индивидуальным, т.е. любые два сегмента A, любые два сегмента G или любые два сегмента L специфического фрагмента REP могут быть идентичными или могут не быть идентичными. Таким образом, общей особенностью изобретения является то, что каждый тип сегмента не является одинаковым внутри специфического фрагмента REP. Скорее, последующее раскрытие обеспечивает специалиста в данной области техники рекомендациями, как создать индивидуальные сегменты и собрать их во фрагмент REP, который является частью функционального спидроинового белка согласно изобретению.
Из представленных в настоящем описании экспериментальных данных было сделано заключение, что каждый индивидуальный сегмент A представляет собой аминокислотную последовательность, имеющую от 8 до 18 аминокислотных остатков. Предпочтительно, каждый индивидуальный сегмент A содержит от 13 до 15 аминокислотных остатков. Также возможно, что большинство или более чем два сегмента A содержат от 13 до 15 аминокислотных остатков, и что меньшинство, а именно один или два сегмента A, содержат от 8 до 18 аминокислотных остатков, а именно 8-12 или 16-18 аминокислотных остатков. Значительное большинство аминокислотных остатков являются остатками аланина. Более подробно, от 0 до 3 аминокислотных остатков являются неаланиновыми остатками и остальные аминокислотные остатки являются остатками аланина. Таким образом, все аминокислотные остатки в каждом индивидуальном сегменте A являются остатками аланина, без исключения или за исключением одного, двух или трех аминокислотных остатков, которые могут быть любой аминокислотой. Предпочтительно, аланин-замещающая аминокислота (аминокислоты) является (являются) природными аминокислотами, предпочтительно индивидуально выбранными из группы из серина, глутаминовой кислоты, цистеина и глицина, более предпочтителен серин. Конечно, возможно, что один или более сегментов A являются полностью аланиновыми сегментами, в то время как остальные сегменты A содержат 1-3 неаланиновых остатка, таких как серин, глутаминовая кислота, цистеин или глицин.
В предпочтительном воплощении каждый сегмент A содержит 13-15 аминокислотных остатков, включая 10-15 остатков аланина и 0-3 неаланиновых остатка, как описано выше. В более предпочтительном воплощении каждый сегмент A содержит 13-15 аминокислотных остатков, включая 12-15 остатков аланина и 0-1 неаланиновых остатков, как описано выше.
Предпочтительно, каждый индивидуальный сегмент A имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 7-19, 43-56, 71-83, 107-120, 135-147, 171-183, 198-211, 235-248, 266-279, 294-306, 330-342, 357-370, 394-406, 421-434, 458-470, 489-502, 517-529, 553-566, 581-594, 618-630, 648-661, 676-688, 712-725, 740-752, 776-789, 804-816, 840-853, 868-880, 904-917, 932-945, 969-981, 999-1013, 1028-1042 и 1060-1073 SEQ ID NO: 3. Каждая последовательность данной группы соответствует сегменту природной последовательности белка MaSp1 Euprosthenops australis, которая расшифрована с помощью клонирования соответствующей кДНК, см. примеры 1-2 и фигуры 1-2A. Альтернативно каждый индивидуальный сегмент A имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 31-42, 61-75, 90-104, 122-135 и 153-171 SEQ ID NO: 9, аминокислотных остатков 12-25, 46-60, 75-88, 112-119, 150-158 и 173-180 SEQ ID NO: 13, аминокислотных остатков 31-42 SEQ ID NO: 14 и аминокислотных остатков 122-135 SEQ ID NO: 15. Каждая последовательность данной группы соответствует сегменту экспрессированных, искусственных спидроиновых белков согласно изобретению, белки которого обладают способностью образовывать шелковые волокна при определенных условиях. См. примеры 5-8, 12 и фигуру 2B. Вне связи с какой-либо теорией, авторы изобретения предусматривают, что сегменты A согласно изобретению образуют спиральные структуры или бета-листы.
Термин "% идентичности", применяемый в настоящем описании и в прилагаемой формуле изобретения, вычисляется следующим образом. Тестируемую последовательность выравнивают с целевой последовательностью с помощью алгоритма CLUSTAL W (Thompson, J. D., Higgins, D. G. and Gibson, T.J., Nucleic Acids Research, 22: 4673-4680 (1994)). Сравнивают аминокислотные остатки в каждом положении и процентное содержание положений в тестируемой последовательности, которые имеют идентичные соответствия в исследуемой последовательности, описывают как % идентичности.
Термин "% сходства", применяемый в настоящем описании и в прилагаемой формуле изобретения, рассчитывается так же, как описано для "% идентичности", за исключением того, что сходными являются гидрофобные остатки Ala, Val, Phe, Pro, Leu, Ile, Trp, Met и Cys; сходными являются основные остатки Lys, Arg и His; сходными являются кислые остатки Glu и Asp; и сходными являются гидрофильные, незаряженные остатки Gln, Asn, Ser, Thr и Tyr. Оставшаяся природная аминокислота Gly является не сходной ни с одной аминокислотой в данном контексте.
В настоящем описании альтернативные воплощения согласно изобретению соответствуют вместо указанного процентного значения идентичности, соответствующему процентному значению сходства. Другие альтернативные воплощения отвечают соответствующему процентному значению идентичности, а также другому, более высокому процентному значению сходства, выбранному из группы предпочтительных процентных значений идентичности для каждой последовательности. Например, последовательность может быть на 70% сходной с другой последовательностью; или она может быть на 70% идентична другой последовательности; или она может быть на 70% идентичной и на 90% сходной с другой последовательностью.
В предпочтительных воплощениях согласно изобретению каждый индивидуальный сегмент A имеет, по меньшей мере, 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 7-19, 43-56, 71-83, 107-120, 135-147, 171-183, 198-211, 235-248, 266-279, 294-306, 330-342, 357-370, 394-406, 421-434, 458-470, 489-502, 517-529, 553-566, 581-594, 618-630, 648-661, 676-688, 712-725, 740-752, 776-789, 804-816, 840-853, 868-880, 904-917, 932-945, 969-981, 999-1013, 1028-1042 и 1060-1073 SEQ ID NO: 3; аминокислотных остатков 31-42, 61-75, 90-104, 122-135 и 153-171 SEQ ID NO: 9; аминокислотных остатков 12-25, 46-60, 75-88, 112-119, 150-158 и 173-180 SEQ ID NO: 13; аминокислотных остатков 31-42 SEQ ID NO: 14; и аминокислотных остатков 122-135 SEQ ID NO: 15. Таким образом, в определенных воплощениях согласно изобретению каждый индивидуальный сегмент A является идентичным аминокислотной последовательности, выбранной из упомянутых выше аминокислотных сегментов.
Кроме того, из представленных в настоящем описании экспериментальных данных было сделано заключение, что каждый индивидуальный G сегмент представляет собой аминокислотную последовательность из 12-30 аминокислотных остатков. Предпочтительно каждый индивидуальный сегмент G состоит из 14-23 аминокислотных остатков. По меньшей мере, 40% аминокислотных остатков каждого сегмента G представляют собой остатки глицина. Обычно содержание глицина в каждом индивидуальном сегменте G находится в диапазоне 40-60%.
Предпочтительно каждый индивидуальный сегмент G имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 20-42, 57-70, 84-106, 121-134, 148-170, 184-197, 212-234, 249-265, 280-293, 307-329, 343-356, 371-393, 407-420, 435-457, 471- 488, 503-516, 530-552, 567-580, 595-617, 631-647, 662-675, 689-711, 726-739, 753-775, 790-803, 817-839, 854-867, 881-903, 918-931, 946-968, 982-998, 1014-1027, 1043-1059 и 1074-1092 SEQ ID NO: 3. Каждая последовательность данной группы соответствует сегменту природной последовательности белка MaSp1 Euprosthenops australis, которая установлена с помощью клонирования соответствующей кДНК, см. примеры 1-2 и фигуру 1-2A. Альтернативно каждый индивидуальный сегмент G имеет, по меньшей мере, 80% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 11-30, 43-60, 76-89, 105-121 и 136-152 SEQ ID NO: 9 и аминокислотных остатков 1-11, 26-45, 61-74, 89-111, 120-149 и 159-172 SEQ ID NO: 13. Каждая последовательность данной группы соответствует сегменту экспрессированных искусственных спидроиновых белков согласно изобретению, белки которого обладают способностью образовывать шелковые волокна при определенных условиях. См. примеры 5-8, 12 и фигуру 2B.
В предпочтительных воплощениях согласно изобретению каждый индивидуальный сегмент G имеет, по меньшей мере, 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичности с аминокислотной последовательностью, выбранной из группы аминокислотных остатков 20-42, 57-70, 84-106, 121-134, 148-170, 184-197, 212-234, 249-265, 280-293, 307-329, 343-356, 371-393, 407-420, 435-457, 471-488, 503-516, 530-552, 567-580, 595-617, 631-647, 662-675, 689-711, 726-739, 753-775, 790-803, 817-839, 854-867, 881-903, 918-931, 946-968, 982-998, 1014-1027, 1043-1059 и 1074-1092 SEQ ID NO: 3; аминокислотных остатков 11-30, 43-60, 76-89, 105-121 и 136-152 SEQ ID NO: 9; и аминокислотных остатков 1-11, 26-45, 61-74, 89-111, 120-149 и 159-172 SEQ ID NO: 13. Таким образом, в определенных воплощениях согласно изобретению, каждый индивидуальный сегмент G является идентичным аминокислотной последовательности, выбранной из упомянутых выше аминокислотных сегментов.
В определенных воплощениях первые два аминокислотных остатка каждого сегмента G согласно изобретению не являются -Gln-Gln-.
В определенных воплощениях положение, соответствующее консервативному остатку Tyr (а именно соответствующее положению 16 в SEQ ID NO: 5, положению 10 в SEQ ID NO: 6 и положению 7 в SEQ ID NO: 7) не является Phe в любом сегменте G согласно изобретению.
В определенных воплощениях положение, соответствующее консервативному остатку Tyr (а именно соответствующее положению 16 в SEQ ID NO: 5, положению 10 в SEQ ID NO: 6 и положению 7 в SEQ ID NO: 7) является Tyr в каждом сегменте G согласно изобретению.
Из этого следует, что определенные воплощения белков согласно изобретению демонстрируют комбинацию упомянутых выше ограничений, т.е. первые два аминокислотных остатка каждого сегмента G согласно изобретению не являются -Gln-Gln- и консервативный остаток Tyr (а именно соответствующий положению 16 в SEQ ID NO: 5, положению 10 в SEQ ID NO: 6 и положению 7 в SEQ ID NO: 7) является Tyr в каждом сегменте G согласно изобретению. В определенных воплощениях упомянутые выше ограничения, взятые отдельно или в любой возможной комбинации, далее могут сочетаться с тем ограничением, что фрагмент REP не имеет остатков пролина, как обсуждалось выше.
На фигурах 1-2 и в примерах 3-4 представлены три подтипа сегмента G согласно изобретению. Данная классификация основана на тщательном анализе последовательности белка MaSp1 Euprosthenops australis (фигуры 1-2A), и полученная информация была использована и проверена при построении новых искусственных белков шелка паука (фигура 2B).
Первый подтип сегмента G согласно изобретению представлен однобуквенной аминокислотной консенсусной последовательностью GQG(G/S)QGG(Q/Y)GG (L/Q)GQGGYGQGA GSS, как показано на фигуре 2A и SEQ ID NO: 5. Первый, и, в целом, самый длинный подтип сегмента G обычно содержит 23 аминокислотных остатка, но может содержать только 17 аминокислотных остатков, и не содержит заряженных остатков или содержит один заряженный остаток. Таким образом, предпочтительно указанный первый подтип сегмента G содержит 17-23 аминокислотных остатков, но предполагается, что он может содержать минимально 12 или до 30 аминокислотных остатков. Вне связи с какой-либо теорией предусматривается, что данный подтип образует спиральные структуры или 31-спиральные структуры. Типичные сегменты G указанного первого подтипа представляют собой аминокислотные остатки 20-42, 84-106, 148-170, 212-234, 307-329, 371-393, 435-457, 530-552, 595-617, 689-711, 753-775, 817-839, 881-903, 946-968, 1043-1059 и 1074-1092 SEQ ID NO: 3; аминокислотные остатки 11-30, 105-121 и 136-152 SEQ ID NO: 9; и аминокислотные остатки 26-45 и 89-111 SEQ ID NO: 13. Альтернативно сегменты G указанного первого подтипа представляют собой аминокислотные остатки 120-149 и 159-172 SEQ ID NO: 13. В определенных воплощениях первые два аминокислотных остатка каждого сегмента G данного первого подтипа согласно изобретению не являются -Gln-Gln-.
Второй подтип сегмента G согласно изобретению представлен однобуквенной аминокислотной консенсусной последовательностью GQGGQGQG (G/R)YGQG(A/S)G (S/G)S, как показано на фигуре 2A и SEQ ID NO: 6. Указанный второй подтип сегмента G, как правило, имеет средний размер, обычно содержит 17 аминокислотных остатков и не содержит заряженных остатков или содержит один заряженный остаток. Предпочтительно указанный второй подтип сегмента G содержит 14-20 аминокислотных остатков, но предусматривается, что он может содержать минимально 12 или до 30 аминокислотных остатков. Вне связи с какой-либо теорией предусматривается, что данный подтип образует спиральные структуры. Типичными сегментами G указанного второго подтипа являются аминокислотные остатки 249-265, 471-488, 631-647 и 982-998 SEQ ID NO: 3; и аминокислотные остатки 43-60 SEQ ID NO: 9.
Третий подтип сегмента G согласно изобретению представлен однобуквенной аминокислотной консенсусной последовательностью G(R/Q)GQG(G/R)YGQG (A/S/V)GGN, как показано на фигуре 2A и SEQ ID NO: 7. Указанный третий подтип сегмента G обычно содержит 14 аминокислотных остатков, и, как правило, является самым коротким среди подтипов сегмента G согласно изобретению. Предпочтительно указанный третий подтип сегмента G содержит 12-17 аминокислотных остатков, но предусматривается, что он может содержать до 23 аминокислотных остатков. Вне связи с какой-либо теорией предусматривается, что данный подтип образует изогнутые структуры. Показательными сегментами G указанного третьего подтипа являются аминокислотные остатки 57-70, 121-134, 184-197, 280-293, 343-356, 407-420, 503-516, 567-580, 662-675, 726-739, 790-803, 854-867, 918-931, 1014-1027 SEQ ID NO: 3; аминокислотные остатки 76-89 SEQ ID NO: 9; и аминокислотные остатки 61-74 SEQ ID NO: 13. Альтернативно сегмент G указанного третьего подтипа представляет собой аминокислотные остатки 1-11 SEQ ID NO: 13.
Таким образом, в предпочтительных воплощениях, каждый индивидуальный сегмент G имеет, по меньшей мере, 80%, предпочтительно 90%, более предпочтительно 95%, идентичности c аминокислотной последовательностью, выбранной из SEQ ID NO: 5, SEQ ID NO:6 и SEQ ID NO:7.
В предпочтительном воплощении последовательности из сменяющих друг друга сегментов A и G фрагмента REP каждый второй сегмент G относится к первому подтипу, в то время как остальные сегменты G относятся к третьему подтипу, например, ...A 1 G short A 2 G long A 3 G short A 4 G long A 5 G short... В другом предпочтительном воплощении фрагмента REP регулярность сегментов G прерывается вставкой одного сегмента G второго подтипа, например, ...A 1 G short A 2 G long A 3 G mid A 4 G short A 5 G long...
Каждый индивидуальный сегмент L представляет собой произвольную линкерную аминокислотную последовательность, которая может содержать от 0 до 20 аминокислотных остатков, как, например, от 0 до 10 аминокислотных остатков. Несмотря на то, что данный сегмент является произвольным и не является функционально критическим в спидроиновом белке, тем не менее его присутствие предусматривается в полностью функциональных спидроиновых белках, образующих волокна шелка паука согласно изобретению. Линкерные аминокислотные последовательности также представлены в повторяющейся части (SEQ ID NO: 3) в расшифрованной аминокислотной последовательности белка MaSp1 Euprosthenops australis. В частности, аминокислотная последовательность линкерного сегмента может иметь сходство с любым из описанных сегментов A или G, но обычно указанная последовательность недостаточно соответствует их критериям, описанным в настоящем документе.
Как показано на рисунке 2A, линкерный сегмент, расположенный в C-концевой части фрагмента REP может быть представлен однобуквенными аминокислотными консенсусными последовательностями ASASAAASAA STVANSVS и ASAASAAA, богатыми аланином. В действительности, вторая последовательность может рассматриваться как сегмент A согласно изобретению, тогда как первая последовательность имеет высокую степень сходства с сегментами A согласно изобретению. Другой пример линкерного сегмента согласно изобретению имеет однобуквенную аминокислотную последовательность GSAMGQGS, которая богата глицином и имеет высокую степень сходства с сегментами G согласно изобретению.
Показательные сегменты L представляют собой аминокислотные остатки 1-6 и 1093-1110 SEQ ID NO: 3; аминокислотные остатки 1-10 и 153-171 SEQ ID NO: 9; и аминокислотные остатки 173-180 SEQ ID NO: 13, однако специалист в данной области техники легко обнаружит, что существует много подходящих альтернативных аминокислотных последовательностей для данных сегментов. В одном воплощении фрагмента REP согласно изобретению один из сегментов L содержит 0 аминокислот, т.е. один из сегментов L ликвидируется. В другом воплощении фрагмента REP согласно изобретению оба сегмента L содержат 0 аминокислот, т.е. оба сегмента L ликвидируются. Таким образом, данные воплощения фрагментов REP согласно изобретению могут быть схематично представлены следующим образом: (AG) n L, (AG) n AL, (GA) n L, (GA) n GL; L(AG) n , L(AG) n A, L(GA) n , L(GA) n G; и (AG) n , (AG) n A, (GA) n , (GA) n G. Любой из указанных фрагментов REP является подходящим для применения с любым фрагментом CT, описанным далее.
C-концевой (CT) фрагмент спидроинового белка согласно изобретению имеет высокую степень сходства с C-концевой аминокислотной последовательностью спидроиновых белков. Как показано на фигуре 3, данная аминокислотная последовательность хорошо сохранилась в различных видах и в спидроиновых белках, включая MaSp1 и MaSp2. В следующих примерах показано, что не важно, какой конкретно фрагмент CT представлен в спидроиновых белках согласно изобретению, если только фрагмент CT не отсутствует полностью. Таким образом, фрагмент CT согласно изобретению может быть выбран из любой из аминокислотных последовательностей, показанных на фигуре 3 или из последовательностей с высокой степенью сходства. Примечательно, что фрагмент CThyb SEQ ID NO: 13 имеет 96% идентичности с консенсусной аминокислотной последовательностью SEQ ID NO: 8, в то время как фрагмент CTnat SEQ ID NO: 9 демонстрирует только 59% идентичности с консенсусной аминокислотной последовательностью SEQ ID NO: 8. Эти данные показывают, что в спидроиновом белке согласно изобретению можeт быть применен широкий ряд C-концевых последовательностей.
Последовательность фрагмента CT согласно изобретению имеет по меньшей мере 50% идентичности, предпочтительно по меньшей мере 60% идентичности, с консенсусной аминокислотной последовательностью SEQ ID NO: 8, которая основана на аминокислотных последовательностях фигуры 3. В предпочтительном воплощении последовательность фрагмента CT согласно изобретению имеет по меньшей мере 65% идентичности, предпочтительно по меньшей мере 70% идентичности с аминокислотными остатками 1-71 консенсусной аминокислотной последовательности SEQ ID NO: 8. В предпочтительных воплощениях фрагмент CT согласно изобретению имеет, кроме того, 70%, предпочтительно 80%, сходства с консенсусной аминокислотной последовательностью SEQ ID NO: 8, или ее аминокислотными остатками 1-71.
Показательные фрагменты CT согласно изобретению представляют собой последовательность SEQ ID NO: 4 Euprosthenops australis, полученные из Euprosthenops australis аминокислотные остатки 172-269 SEQ ID NO: 9 и аминокислотные остатки 181-276 SEQ ID NO: 13, которые, как считают, происходят из Euprosthenops sp. (Pouchkina-Stantcheva, N. N. & McQueen-Mason, S. J. Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp. (Pisauridae). Comp Biochem Physiol B Biochem Mol Biol 138, 371-376 (2004)), но которые имеют высокую степень сходства с MaSp1 из Nephila clavipes и из Nephila senegalensis. Таким образом, согласно предпочтительному аспекту изобретения фрагмент CT имеет, по меньшей мере, 80% идентичности с SEQ ID NO: 4, с аминокислотными остатками 172-269 SEQ ID NO: 9, с аминокислотными остатками 181-276 SEQ ID NO: 13, с аминокислотными остатками 172-269 SEQ ID NO: 16 или с любой индивидуальной аминокислотной последовательностью MaSp1/ADF-4 фигуры 3 и примера 4. В предпочтительных аспектах изобретения фрагмент CT имеет по меньшей мере 90%, а именно по меньшей мере 95% идентичности с SEQ ID NO: 4, с аминокислотными остатками 172-269 SEQ ID NO: 9, аминокислотными остатками 181-276 SEQ ID NO: 13, аминокислотными остатками 172-269 SEQ ID NO: 16 или с любой индивидуальной аминокислотной последовательностью MaSpl/ADF-4 фигуры 3 и примера 4. В предпочтительных аспектах изобретения фрагмент CT является идентичным SEQ ID NO: 4, аминокислотным остаткам 172-269 SEQ ID NO: 9, аминокислотным остаткам 181-276 SEQ ID NO: 13, аминокислотным остаткам 172-269 SEQ ID NO: 16 или любой индивидуальной аминокислотной последовательности MaSpl/ADF-4 фигуры 3 и примера 4.
Фрагмент CT обычно состоит из 70-120 аминокислотных остатков. Предпочтительно фрагмент CT содержит, по меньшей мере, 70 или более чем 80, предпочтительно более чем 90 аминокислотных остатков. Также предпочтительно фрагмент CT содержит, самое большее, 120 или менее чем 110 аминокислотных остатков. Типичный фрагмент CT содержит приблизительно 100 аминокислотных остатков.
Согласно другому аспекту в настоящем изобретении предоставляется выделенный слитый белок, состоящий из первого белкового фрагмента, который является спидпроиновым белком главной ампулярной железы, предпочтительно состоящим из 150-420 аминокислотных остатков, и второго белкового фрагмента, который включает в себя партнера по слитию и участок узнавания расщепляющего агента. Первый белковый фрагмент соединяется через участок узнавания расщепляющего агента с партнером по слитию, т.е. партнер по слитию может быть удален при обработке слитого белка подходящим расщепляющим агентом при подходящих условиях, с получением спидроинового белка главной ампулярной железы, предпочтительно состоящего из 150-420 аминокислотных остатков. Преимуществом данного слитого белка является то, что он может быть получен в больших количествах в растворе, предпочтительно в физиологической среде, обычно в буферной водной среде, такой как 10-100 мМ Tris-HCl буфер, pH 6-9, без преципитации и других проблем получения, если его получают в подходящих хозяевах, таких как бактерии, предпочтительно E. coli. Растворимые слитые белки могут находиться в растворе в течение долгого периода времени, обычно в течение дней или недель, что способствует их массовому производству и снижает риск белковой агрегации. Термины "растворимый" и "в растворе" означают, что нет видимой агрегации белка, и белок не осаждается из растворителя при 60000 x g. По желанию слитые белки в растворе могут быть подвергнуты расщеплению подходящим расщепляющим агентом, с получением спидроинового белка главной ампулярной железы, который спонтанно образует шелковые волокна.
В предпочтительном аспекте в настоящем изобретении предоставляется выделенный слитый белок, выбранный из группы X-REP-CT и REP-CT-X, предпочтительно X-REP-CT. REP и CT являются белковыми фрагментами согласно изобретению, что означает, что итоговый белок MaSp формы REP-CT является белком MaSp согласно изобретению. X является белковым фрагментом, включающим в себя партнера по слитию и участок узнавания расщепляющего агента, как описано выше. Комбинированный белковый фрагмент REP-CT соединяется через участок узнавания расщепляющего агента с партнером по слитию.
Партнеры по слитию согласно изобретению включают в себя любой белковый фрагмент, который улучшает растворимость и/или стабильность его белкового фрагмента - партнера, в настоящем описании белка MaSp согласно изобретению. Партнер по слитию также включает в себя подходящую основу для аффинной очистки. Примеры партнеров по слитию согласно изобретению включают в себя без ограничения тиоредоксин, мальтоза-связывающий белок, глутатион-S-трансфераза (GST), MTB32-C, Gb1, ZZ и Nus A. Специалисту в данной области техники хорошо известны альтернативные подходящие партнеры по слитию. В предпочтительном воплощении изобретения партнер по слитию представляет собой тиоредоксиновую часть (ThrX) в комбинации с His-тагом или с S-тагом. В одном предпочтительном воплощении изобретения партнер по слитию представляет собой часть ThrX в комбинации с двумя His-тагами, а именно His-tag/ThrX/His-tag. В другом предпочтительном воплощении изобретения партнер по слитию представляет собой тиоредоксиновую часть (ThrX).
Участок узнавания расщепляющего агента располагается в том конце белкового фрагмента X, который соединяется с белковым фрагментом MaSp, так что расщепление в участке узнавания дает в результате белок MaSp и партнер по слитию. Примеры участка узнавания расщепляющего агента согласно изобретению включают в себя без ограничения участок узнавания тромбина, имеющий аминокислотную последовательность LVPRGS (расщепляется между R и G); участок узнавания энтерокиназы, имеющий аминокислотную последовательность DDDK (расщепляется после K); участок узнавания гидроксиламина, имеющий аминокислотную последовательность NG (расщепляется между N и G); участок узнавания протеазы HRV 3C, имеющий аминокислотную последовательность LGVLFQGP (расщепляется между Q и G); участок узнавания фактора Xa, имеющий аминокислотную последовательность I(E/D)GR (расщепляется после R); участок узнавания TEV, имеющий аминокислотную последовательность EXXYXQ (G/S), общий ENLYFQG (расщепляется между Q и G/S), участок узнавания Ac-TEV, имеющий аминокислотную последовательность EDNLYFQG (расщепляется между Q и G); и участок узнавания PreScission имеющий аминокислотную последовательность LEVLFQGP (расщепляется между Q и G). Другие подходящие участки узнавания представляют собой участки расщепления для трипсина, эндопротеиназы, протеазы V8, пепсина и CNBr. Другие примеры подходящих участков узнавания для расщепления являются доступными специалисту в данной области техники. В предпочтительном воплощении изобретения участок узнавания расщепляющего агента является участком узнавания тромбина.
В предпочтительном воплощении фрагмент X согласно изобретению имеет структуру ThrX/His-tag/S-tag/тромбин-расщепляемый участок, и фрагмент X соединяется с N-концом белкового фрагмента REP-CT согласно изобретению.
В одном предпочтительном воплощении фрагмент X согласно изобретению имеет структуру His-tag/ThrX/His-tag/тромбин-расщепляемый участок, и фрагмент X соединяется с N-концом белкового фрагмента REP-CT согласно изобретению.
Согласно другому аспекту в настоящем изобретении предоставляется способ получения спидроинового белка главной ампулярной железы согласно изобретению. На первой стадии получают раствор слитого белка согласно изобретению в жидкой среде. Предпочтительно слитый белок не агрегирует и, следовательно, не требуется процедуры повторной солюбилизации. Слитый белок может быть получен рекомбинантным путем и очищен с помощью аффинной основы в слитом белке, такой как His-таг или любой подходящий эпитоп в слитом белке. Жидкая среда может быть любой подходящей средой, предпочтительно физиологической средой, обычно буферной водной средой, такой как 10-100 мM Tris-HCl буфер, pH 6-9. На второй стадии расщепляющий агент согласно изобретению добавляют к жидкой среде для расщепления слитого белка в участке узнавания расщепляющего агента. Как было раскрыто выше, при этом получают спидроиновый белок главной ампулярной железы согласно изобретению. На третьей необязательной стадии, полученный таким образом спидроиновый белок главной ампулярной железы выделяют из жидкой среды с помощью подходящих техник выделения, таких как хроматография и/или фильтрация.
Согласно еще одному аспекту в настоящем изобретении предоставляется способ получения полимера спидроинового белка главной ампулярной железы согласно изобретению. На первой стадии получают раствор слитого белка согласно изобретению в жидкой среде. Предпочтительно, слитый белок не агрегирует и, следовательно, не требуются процедуры повторной солюбилизации. Слитый белок может быть получен рекомбинантным путем и очищен с помощью аффинной основы в слитом белке, такой как His-таг или любой подходящий эпитоп в слитом белке. Жидкая среда может быть любой подходящей средой, предпочтительно физиологической средой, обычно буферной водной средой, такой как 10-100 мM Tris-HCl буфер, pH 6-9. На второй стадии расщепляющий агент согласно изобретению добавляют к жидкой среде для расщепления слитого белка в участке узнавания расщепляющего агента. Как было раскрыто выше, при этом получают спидроиновый белок главной ампулярной железы согласно изобретению. На третьей стадии проводят полимеризацию полученного таким образом спидроинового белка главной ампулярной железы в жидкой среде. Полимеризация обычно инициируется на границе раздела между двумя различными фазами, такими как границы раздела жидкость/воздух, жидкость/твердая фаза, и вода/масло. Таким образом, данная третья стадия также может далее включать в себя создание границы раздела между жидкой средой и другой фазой. Другая фаза выбирается из группы, состоящей из газовой фазы, жидкой фазы и твердой фазы. Как подробно описано выше, жидкая среда обычно является водной средой, и другие подходящие фазы представляют собой, например, органические растворители, не смешиваемые с воздухом и водой, такие как масло, например минеральное масло, подходящее для реакций ПЦР. Присутствие полученной границы раздела стимулирует полимеризацию в самой границе раздела или в окружающей ее области, и данная область распространяется в жидкую среду, так что указанная полимеризация начинается в указанной границе раздела или в указанной области. Предпочтительные границы раздела включают в себя границы раздела вода/воздух и вода/масло. Полимеризация обычно происходит спонтанно в течение минут или нескольких часов, а именно от 1 мин до 5 часов, при инкубации при комнатной температуре. На необязательной четвертой стадии полученный таким образом полимер спидроинового белка главной ампулярной железы выделяют из жидкой среды с помощью подходящих техник выделения.
Как обсуждалось выше, образование волокон вызывается протеолитическим высвобождением спидроина небольшого размера из слитого белка. Если реакцию расщепления проводят в пробирке, которую осторожно покачивают из стороны в сторону, то волокно образуется на границе раздела воздух/вода вдоль по пробирке. Пробирка может быть сделана из любого подходящего материала, такого как пластик или стекло. Если смесь, в которой происходит реакция расщепления, не перемешивают, то, на границе раздела воздух/вода образуется пленка. Если поверх водной смеси для реакции расщепления добавляют масло, то на границе раздела масло/вода образуется пленка и при перемешивании, и без него. Если смесь, в которой проходит реакция расщепления, вспенивают, например, барботированием воздуха или вибрацией, то получают устойчивую пену, которая затвердевает при высыхании.
Применяя способ(ы) настоящего изобретения, можно получить рекомбинантным путем большие количества слитых белков согласно изобретению, которые можно расщеплять и проводить их полимеризацию по желанию. Это обеспечивает лучший контроль над процессом полимеризации и позволяет оптимизировать параметры получения шелковых волокон с желаемыми свойствами.
Спидроиновый белок главной ампулярной железы согласно изобретению обычно получают рекомбинантным путем, используя ряд подходящих хозяев. Согласно другому аспекту в настоящем изобретении предоставляется выделенная полинуклеотидная молекула, включающая в себя последовательность нуклеиновой кислоты, которая кодирует спидроиновый белок главной ампулярной железы согласно изобретению, или комплементарную ей последовательность нуклеиновой кислоты, а именно SEQ ID NO: 9-13, предпочтительно SEQ ID NO: 9, 12 и 13. Данные молекулы полинуклеиновых кислот, а также молекулы полинуклеиновых кислот, кодирующие различные белки, раскрытые в настоящем документе (SEQ ID NO: 2-16) также могут быть полезны в дальнейших разработках искусственных спидроиновых белков или систем для их получения.
Слитый белок согласно изобретению обычно получают рекомбинантным путем, с помощью ряда подходящих хозяев, таких как бактерии, дрожжи, клетки млекопитающих, растения, клетки насекомых и трансгенные животные. Предпочтительно слитый белок согласно изобретению получают в бактериях.
Согласно другому аспекту в настоящем изобретении в силу вышесказанного предоставляется выделенная молекула полинуклеотидной кислоты, включающая в себя последовательность нуклеиновой кислоты, которая кодирует слитый белок согласно изобретению, или комплементарную ему последовательность нуклеиновой кислоты. Молекула полинуклеиновой кислоты также может быть применена в создании других искусственных спидроиновых белков или систем для их получения.
Молекулы полинуклеиновых кислот согласно изобретению могут быть молекулами ДНК, включая молекулы кДНК, или молекулами РНК. Специалисту в данной области техники хорошо известно, что последовательность нуклеиновой кислоты также может быть описана с помощью комплементарной ей последовательности нуклеиновой кислоты. Поэтому последовательности нуклеиновых кислот, которые комплементарны последовательностям нуклеиновых кислот согласно изобретению, также входят в охраняемый объем изобретения.
Согласно одному аспекту в настоящем изобретении предоставляется способ получения растворимого слитого белка согласно изобретению. На первой стадии молекулу полинуклеиновой кислоты, которая кодирует слитый белок согласно изобретению, экспрессируют в подходящем хозяине. На второй стадии полученный таким образом растворимый слитый белок выделяют, например, с помощью хроматографии и/или фильтрации. По желанию указанная вторая стадия выделения растворимого слитого белка включает в себя удаление LPS и других пирогенов.
В настоящем изобретении далее предоставляется способ получения спидроинового белка главной ампулярной железы согласно изобретению. На первой стадии молекулу полинуклеиновой кислоты, которая кодирует слитый белок согласно изобретению, экспрессируют в подходящем хозяине. На второй стадии полученный таким образом растворимый слитый белок выделяют, например, с помощью хроматографии и/или фильтрации. На третьей стадии получают раствор выделенного слитого белка, на четвертой стадии добавляют подходящий расщепляющий агент в жидкую среду. При этом происходит расщепление слитого белка в участке узнавания расщепляющего агента, и таким образом получают спидроиновый белок главной ампулярной железы. На необязательной пятой стадии полученный таким образом спидроиновый белок главной ампулярной железы выделяют из жидкой среды. Далее по желанию указанная вторая стадия выделения растворимого слитого белка и по желанию пятая стадия выделения спидроинового белка главной ампулярной железы включают в себя удаление LPS и других пирогенов.
В настоящем изобретении также предоставляется способ получения полимера спидроинового белка главной ампулярной железы согласно изобретению. На первой стадии молекулу полинуклеиновой кислоты, которая кодирует слитый белок согласно изобретению, экспрессируют в подходящем хозяине. На второй стадии полученный таким образом растворимый слитый белок выделяют, например, с помощью хроматографии и/или фильтрации. На третьей стадии получают раствор выделенного слитого белка, на четвертой стадии добавляют подходящий расщепляющий агент в жидкую среду. При этом происходит расщепление слитого белка в участке узнавания расщепляющего агента, и таким образом получают спидроиновый белок главной ампулярной железы. На пятой стадии проводят полимеризацию полученного таким образом спидроинового белка главной ампулярной железы в жидкой среде. Полимеризация обычно инициируется на границе раздела между двумя различными фазами, такими как границы раздела жидкость/воздух, жидкость/твердая фаза и вода/масло. Таким образом, данная пятая стадия также может далее включать в себя создание границы раздела между жидкой средой и другой фазой. Другая фаза выбирается из группы, состоящей из газовой фазы, жидкой фазы и твердой фазы. Как подробно описано выше, жидкая среда обычно является водной средой, и другие подходящие фазы представляют собой, например, органические растворители, не смешиваемые с воздухом и водой, такие как масло, например минеральное масло, подходящее для реакций ПЦР. Присутствие полученной границы раздела стимулирует полимеризацию в самой границе раздела или в окружающей ее области, и данная область распространяется в жидкую среду, так что указанная полимеризация начинается в указанной границе раздела или в указанной области. Предпочтительные границы раздела включают в себя границы раздела вода/воздух и вода/масло. Полимеризация обычно происходит спонтанно в течение минут или нескольких часов, а именно от 1 мин до 5 часов, при инкубации при комнатной температуре. На необязательной шестой стадии полученный таким образом полимер железы выделяют из жидкой среды.
Для получения белка с низким содержанием пирогенов, что является обязательным при применении биоматериала in vivo, был разработан оптимизированный протокол очистки для удаления липополисахаридов (LPS). Чтобы избежать контаминации высвобождаемыми LPS, продуцирующие бактериальные клетки подвергали стадиям отмывки с заменой CaCl2 и EDTA. После лизиса клеток все последующие стадии очистки проводят в буферах с низкой электропроводностью, чтобы минимизировать гидрофобные взаимодействия между получаемым белком и LPS. Затем содержание LPS минимизируют путем пропускания раствора белка через колонку Endotrap, которая имеет лиганд, который специфически адсорбирует LPS. Чтобы гарантировать постоянное, низкое содержание LPS и других пирогенов, все партии проверяют с помощью теста на пирогенность in vitro (IPT) и/или с помощью кинетического теста с лизатом Limulus amebocyte (LAL). Хотя произведенные в Грам-отрицательном бактериальном хозяине рекомбинантные спидроиновые белки могут быть очищены так, чтобы остаточные уровни LPS и других пирогенов были ниже предельных значений, требуемых в тестах на животных, т.е. ниже 25 ЭЕ/имплант. В определенных воплощениях согласно изобретению содержание LPS и других пирогенов в выделенном слитом белке составляет 1 ЭЕ/мг белка или меньше. В определенных воплощениях согласно изобретению содержание LPS и других пирогенов в выделенном спидроиновом белке главной ампулярной железы составляет 1 ЭЕ/мг белка или меньше, предпочтительно 0,25 ЭЕ/мг белка или меньше.
Согласно другому аспекту в настоящем изобретении предоставляется полимер спидроинового белка главной ампулярной железы согласно изобретению. В предпочтительном воплощении полимер данного белка представляет собой полимер, получаемый любым из способов согласно изобретению.
В предпочтительных воплощениях содержание β-листовой формы полимера спидроинового белка главной ампулярной железы согласно изобретению составляет свыше 50%, т.е. более чем 50% вторичной структуры полимера данного белка находится в β-листовой форме. Это является преимуществом, поскольку предполагается, что более высокое содержание β-листовых структур дает более жесткое и/или более крепкое и/или менее растяжимое волокно.
Предпочтительно полимер спидроинового белка согласно изобретению представляет собой волокно макроскопического размера, т.е. с диаметром больше 1 мкм, предпочтительно больше 10 мкм и длиной более 5 мм. Предпочтительно волокно имеет диаметр в диапазоне 10-400 мкм, предпочтительно 60-120 мкм, и длину в диапазоне 0,5-300 см, предпочтительно 1-100 см. Другими предпочтительными диапазонами являются 0,5-30 см и 1-20 см. Также предпочтительно полимер спидроинового белка согласно изобретению имеет предел прочности больше 1 MПa, предпочтительно больше 2 MПa, более предпочтительно 10 MПa или больше. Предпочтительно полимер спидроинового белка согласно изобретению имеет предел прочности больше 100 MПa, более предпочтительно 200 MПa или больше. Волокно обладает способностью оставаться неповрежденным при физическом воздействии, т.е. его можно применять в прядении, ткачестве, кручении, вязании и в подобных процедурах.
В других предпочтительных воплощениях полимер спидроинового белка согласно изобретению образует пену, гель, сетку или пленку.
Согласно еще одному аспекту в настоящем изобретении предоставляется новое применение белкового фрагмента, включающего в себя партнера по слитию и участок узнавания расщепляющего агента, для производства слитого белка. Слитый белок состоит из указанного белкового фрагмента и из белка шелка паука согласно изобретению, и два фрагмента соединяются через указанный участок узнавания расщепляющего агента. Белковый фрагмент шелка паука предпочтительно состоит из 150-420 аминокислотных остатков.
Далее настоящее изобретение проиллюстрировано в следующих неограничивающих примерах.
ПРИМЕРЫ
Пример 1. Клонирование и секвенирование кДНК MaSp1 Euprosthenops australis
Главные ампулярные железы приблизительно из 100 взрослых самок пауков Euprosthenops australis, собранных в Южной Африке, использовали для создания по особому заказу библиотеки кДНК на основе pDONR222 (Invitrogen, Paisley, UK). Клоны кДНК, кодирующие белок MaSp1, получили путем скрининга библиотеки с помощью зонда кДНК, кодирующего богатый аланином и глицином фрагмент, полученный из неизвестных подвидов пауков вида Euprosthenops. Блоттинг колоний и обнаружение осуществляли с помощью ЭХЛ-направленного мечения и системы обнаружения (Amersham Biosciences, Uppsala, Sweden) согласно инструкции производителя.
Для дальнейшего исследования выбирали один единичный клон. Для получения полноразмерной последовательности вставки кДНК из данного клона делали внутригенные делеции с помощью системы Erase-a-Base System (Promega, Southampton, UK) и осуществляли секвенирование на MegaBase 1000 instrument (Amersham Biosciences).
Полученная кДНК из 3,8 т.п.н. (SEQ ID NO: 1) кодирует белок MaSp1 (SEQ ID NO: 2) из 1207 аминокислотных остатков, содержащий повторяющийся фрагмент из 34 аланин- и глицин-богатых сегментов (SEQ ID NO: 3) и C-концевой неповторяющийся фрагмент из 97 аминокислотных остатков (SEQ ID NO: 4).
Пример 2. Анализ последовательности повторяющегося фрагмента белка MaSp1 Euprosthenops australis
Белковую последовательность повторяющегося фрагмента белка MaSp1 Euprosthenops australis примера 1 (SEQ ID NO: 3) далее анализировали путем выравнивания повторяющихся сегментов фрагмента, см. фигуру 1. Результаты выравнивания были тщательно исследованы, и были сделаны следующие выводы о структуре.
Аланин-богатые сегменты белка MaSp1 Euprosthenops australis имеют длину 13-15 аминокислотных остатков и состоят только из остатков аланина или из остатков аланина и одного остатка, который представляет собой остаток серина, глутамата или глицина.
Повторяющийся фрагмент белка MaSp1 Euprosthenops australis содержит три родственных, но различных типа глицин-богатых сегментов, перенесенных на фигуру 2A. Два глицин-богатых сегмента отличаются только по длине и встречаемости; самый распространенный глицин-богатый сегмент содержит 23 аминокислотных остатка, в то время как наименее распространенный вариант содержит 17 аминокислотных остатков. Оба указанных глицин-богатых сегмента обычно не имеют заряженных остатков или содержат один заряженный остаток. В отличие от этого самый короткий глицин-богатый сегмент, состоящий из 14 аминокислотных остатков, единственный содержит последовательность GRGQG или GQGQG в N-концевой области, и GN в C-концевой области.
Самый длинный глицин-богатый сегмент представлен однобуквенной аминокислотной консенсусной последовательностью
GQG(G/S)QGG(Q/Y)GG (L/Q)GQGGYGQGA GSS (SEQ ID NO: 5), и не имеет заряженных остатков. Предсказывается, что данный сегмент образует спиральные структуры или 31-спиральные структуры. Богатый глицином сегмент среднего размера представлен однобуквенной аминокислотной консенсусной последовательностью GQGGQGQG(G/R)Y GQG(A/S)G(S/G)S (SEQ ID NO: 6), и не имеет заряженных остатков или содержит один заряженный остаток. Как предсказывают, данный сегмент образует спиральные структуры. Самый короткий глицин-богатый сегмент представлен однобуквенной аминокислотной консенсусной последовательностью G(R/Q)GQG(G/R)YGQG (A/S/V)GGN (SEQ ID NO: 7). Предсказывается, что данный сегмент образует изогнутые структуры.
Повторяющийся фрагмент белка MaSp1 Euprosthenops australis строится из чередующихся богатых аланином и богатых глицином сегментов, например,
...A 1 G 1 A 2 G 2 A 3 G 3 A 4 G 4 A 5 G 5...
Отмечается, что каждый из идентифицированых выше самый короткий и самый длинный глицин-богатые сегменты обычно встречаются в качестве каждого второго глицин-богатого сегмента, например,
...A 1 G short A 2 G long A 3 G short A 4 G long A 5 G short...
И напротив, менее распространенный, богатый глицином фрагмент среднего размера обычно находится между глицин-богатым сегментом более длинного типа и глицин-богатым сегментом более короткого типа, например,
...A 1 G short A 2 G long A 3 G mid A 4 G short A 5 G long...
Пример 3. Предсказание вторичной и третичной структуры повторяющегося фрагмента белка MaSp1 Euprosthenops australis
Спидроиновые полипептиды в растворе обычно сворачиваются с образованием структуры шпильки, которая служит прообразом антипаралелльной β-складчатой структуры готового волокна. Чтобы определить возможные типы сворачивания повторяющегося фрагмента (SEQ ID NO: 3) белка MaSp1 Euprosthenops australis из примеров 1-2, идентифицируют участки белка, которые являются сходными с участками, образующими шпильку или изогнутые структуры. Аланин-богатые сегменты являются маловероятными кандидатами для изогнутого образования, поскольку, согласно расчетам, они образуют спиральные структуры, и, что более важно, указанные сегменты обычно образуют β-слои в волокне.
При применении недавно описанного алгоритма предсказания формы (Fuchs, PF & Alix, AJ, High accuracy prediction of beta-turns and their types using propensities and multiple alignments. Proteins 59, 828-839 (2005)), самые короткие глицин-богатые сегменты показывают высокую вероятность формирования β-изгибов типа II, в то время как для двух более длинных глицин-богатых сегментов предсказывают образование спиральных структур. Высокое содержание триплетов GGX в длинных Gly-богатых сегментах предполагает, что они могут образовывать 31-спиральные структуры.
Повторяющийся характер аминокислотных последовательностей спидроина предполагает таким же образом повторяющийся характер типа укладки. Данные наблюдения, взятые вместе, дают в результате укладку повторяющегося фрагмента белка MaSp1 Euprosthenops australis, которая показана на фигуре 2A. Примечательно, что положительно заряженные остатки почти всегда расположены в предполагаемых изогнутых структурах.
Из типа укладки повторяющегося фрагмента белка MaSp1 Euprosthenops australis может быть выделен мотив следующего состава: аланин-богатый сегмент/(длинный) глицин-богатый спиральный сегмент/аланин-богатый сегмент/(короткий) глицин-богатый изогнутый сегмент/аланин-богатый сегмент/(длинный) глицин-богатый спиральный сегмент/аланин-богатый сегмент, (проиллюстрировано в виде схемы на фигуре 2A).
Пример 4. Анализ последовательности неповторяющегося С-концевого фрагмента белков MaSp1
Производили выравнивание первичной структуры C-концевого неповторяющегося фрагмента (SEQ ID NO: 4) белка MaSp1 Euprosthenops australis, полученного в примере 1, с рядом известных C-концевых фрагментов белков MaSp1 и MaSp2, в частности из вида Euprosthenops sp. (Pouchkina-Stantcheva, NN & McQueen-Mason, SJ, Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp. (Pisauridae). Comp Biochem Physiol B Biochem Mol Biol 138, 371-376 (2004)), Nephila clavipes P19837-5 (Xu, M & Lewis, RV, Structure of a protein superfiber: spider dragline silk. Proc Natl Acad Sci USA 87, 7120-7124 (1990)) и из других видов.
Из показанного на фигуре 3 выравнивания, которое начинается с последнего Ser в повторяющемся фрагменте, становится очевидным, что C-концевые участки MaSp1 и MaSp2 являются высоко консервативными. Виды Euprosthenops sp и Nephila clavipes имеют 95% идентичных остатков; Euprosthenops australis и Nephila clavipes имеют 54% идентичных остатков; и Euprosthenops australis и Euprosthenops sp имеют 55% идентичных остатков. Консенсусная последовательность C-концевых участков MaSp1 и MaSp2 предоставляется в виде SEQ ID NO: 8. На фигуре 3 показано выравнивание следующих белков MaSp, обозначенных номерами доступа GenBank:


Пример 5. Построение генов MaSp1
Последовательность ДНК, кодирующую белок 5Gly/Ala-CTnat (SEQ ID NO: 9), полученный из Euprosthenops australis, амплифицировали методом ПЦР с набором Advantage GC2 kit (BD Biosciences, San Jose, CA, USA) с применением в качестве матрицы клона MaSp1 из библиотеки кДНК примера 1. Участки узнавания ферментов рестрикции BamHI и HindIII вводили в 5'- и 3'-концы, соответственно, и стоп-кодон вводили перед участком HindIII, с применением желаемых праймеров. Затем конструкцию BamHI-5GIy/Ala-CTnat-HindIII субклонировали в модифицированный вектор pET32 (Merck Biosciences, Darmstadt, Germany), приготовленный, как описано в примере 6(C) ниже.
Пример 6. Построение химерных генов MaSp1
(A) фрагменты гена REP
Последовательности ДНК, кодирующие неполные повторяющиеся фрагменты (REP), обозначаемые 3Gly/Ala и 4Gly/Ala, были ампифицированы методом ПЦР с LA Taq (TaKaRa Bio; Saint Germain-en-laye, France) в присутствии бетаина (Henke W et al, Betaine improves the PCR amplification of GC-rich DNA sequences. Nucleic Acids Res 25, 3957-3958 (1997)), с применением в качестве матрицы клона неполной кДНК, кодирующей повторяющейся участок белка MaSp1 вида Euprosthenops sp. (Pouchkina-Stantcheva, NN & McQueen-Mason, SJ, Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp. (Pisauridae). Comp Biochem Physiol B Biochem Mol Biol 138, 371-376 (2004)) (номер Генбанка CQ974358 или CQ816656). Участки узнавания фермента рестрикции вводили в 5'- и 3'-концы с получением следующих конструкций: NcoI-3Gly/Ala-NheI и NcoI-4GIy/Ala-NheI, которые соединялись с фрагментом CT (см. ниже); и клон NcoI-4Gly/Ala-XhoI, который экспрессировался индивидуально, где стоп-кодон вставляли непосредственно перед участком XhoI.
(B) фрагменты гена CT
Последовательность ДНК, кодирующая неповторяющийся С-концевой домен A вида Euprosthenops sp (но с высокой степенью сходства с MaSp1 из Nephila clavipes и Nephila senegalensis) амплифицировали методом ПЦР с применением клона геномной ДНК, кодирующего C-концевой домен MaSp1 (Pouchkina-Stantcheva, NN & McQueen-Mason, SJ, Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp. (Pisauridae). Comp Biochem Physiol B Biochem Mol Biol 138, 371-376 (2004)). Участки узнавания ферментов рестрикции вводили в 5'- и 3'-концы с получением NheI-2Gly/Ala-CT hyb -XhoI, которые соединяли с клонами неполных REP 3Gly/Ala и 4Gly/Ala (см. выше), и NhoI-2Gly/Ala-CT hyb -XhoI, который экспрессировали индивидуально.
(C) Построение генов REP-CT гибрида MaSp1
Клоны REP 3Gly/Ala и 4Gly/Ala соединяли склонами CT с помощью вектора pCR®2.1-TOPO® (Invitrogen). Затем полученные гибридные клоны 5Gly/Ala-CThyb и 6Gly/Ala-CThyb вырезали NcoI и XhoI и субклонировали в модифицированный вектор pET32 (Novagen), где участок расщепления тромбина был удален и новый тромбиновый участок был введен после участка расщепления энтерокиназы.
Пример 7. Экспрессия слитых белков MaSp1
Белки MaSp1, кодированные генами, построенными в примерах 5-6, экспрессировали в качестве слитых белков (типа X-REP-CT), как изложено ниже, с применением модифицированного вектора pET32: тиоредоксин-таг/His-таг/S-таг/участок расщепления тромбина/ген MaSp1, кодирующий тиоредоксин/HiS6/S-таг/участок расщепления тромбина/белок MaSp1, и ген устойчивости к ампициллину под контролем промотора T7.
Различные конструкции MaSp1 в векторах экспрессии pET32 трансформировали в клетки Escherichia coli BL21(DE3) (Merck Biosciences). Клетки выращивали при 30°C в среде Лурия-Бертани, содержащей ампициллин, до OD600 1,0-1,5, вызванной IPTG, и затем инкубировали 4 часа при комнатной температуре. Клетки собирали центрифугированием и разрушали ДНКазой I и лизозимом в 20 мМ Tris-HCl, pH 8,0, 20 мМ имидазоле с 0,5 M NaCl и затем очищали аффинной хроматографией с His-таг на агарозе Ni-NTA (Qiagen, West Sussex, UK). Связанные слитые белки элюировали из колонки с Ni-NTA 200 мМ имидазолом в 20 мМ Tris-HCl, pH 8,0, с 0,5 M NaCl, и проводили диализ против 20 мМ Tris-HCl, pH 8,0. Полученные слитые белки имели чистоту >90%, согласно результатам окраски по Кумасси SDS-полиакриламидных гелей, и их растворяли в 20 мМ Tris-HCl, pH 8,0. Данный процесс давал приблизительно 40 мг/л культуры слитого белка, который оставался стабильным в течение недель без заметного образования осадка.
В другом эксперименте слитые белки экспрессировали в виде белков His6/тиоредоксин/His6/участок расщепления тромбина/MaSp1 из плазмиды, содержащей соответствующий ген, и ген устойчивости к канамицину под контролем промотора T7.
Пример 8. Образование волокон из белков MaSp1
Расщепление тагов слитых белков, полученных в примере 7, проводили в 20 мМ Tris-HCl, pH 8, с соотношением тромбин:слитый белок 1:1000 (об./об.), при очень осторожном покачивании при комнатной температуре. Расщепление тромбина происходило в течение 30-60 мин, согласно результатам электрофореза в SDS-полиакриламидном геле. Полученные белки MaSp1 (фигура 2B, SEQ ID NOS: 9-13) спонтанно полимеризовались в макроскопические волокна различных размеров, показанных в таблице 1. Волокна исходно формировались на границе раздела вода/воздух. Образование можно было наблюдать невооруженным глазом с 1 часа инкубации (см. фигуры 4A, 4B) и после 5 часов не происходило дальнейшего роста волокон. Волокна 6Gly/Ala-CThyb имели длину приблизительно 2 см и волокна 5Gly/Ala-CTnat имели длину ≥10 см. В повторных экспериментах получили волокна 5Gly/Ala-CTnat, которые имели длину ≥20 см (см. фигуру 4C), и даже ≥2 м. Образование волокон можно было наблюдать невооруженным глазом после 10 мин инкубации.
Волокна выделяли и отмывали буфером и затем проводили анализ N-концевой аминокислотной последовательности, который показывал только последовательность белка MaSp1. Анализ показал отсутствие расщепленных тагов в волокнах.
Пример 9. Анализ белковых волокон MaSp1
A. Измерения предела прочности
Предел прочности волокон 6Gly/Ala-CThyb (SEQ ID NO: 13) и 5Gly/Ala-CTnat (SEQ ID NO: 9) примера 8 устанавливали следующим образом. Для манипуляций с короткими (1-2 см) волокнами 6Gly/Ala-CThyb в ходе измерения пределов прочности их инкубировали в течение короткого времени в 15% растворе глицерина в воде перед высушиванием на воздухе. Длинные волокна (10 см) 5Gly/Ala-CTnat либо не обрабатывали, либо инкубировали в течение короткого времени в 15% глицерине, либо вытягивали рукой в 75% метаноле перед высушиванием на воздухе. Предел прочности высушенных на воздухе волокон измеряли путем растягивания волокон с помощью прибора Zwick Material Tester со скоростью 10 мм/мин. См. таблицу 1.
Предел прочности обработанных глицерином, высушенных на воздухе, длиной 1-2 см волокон из 6Gly/Ala-CThyb (SEQ ID NO: 13) составлял 2 MПа, и прочность волокон длиной 10 см из 5Gly/Ala-CTnat (SEQ ID NO: 9) составляла 4-5 MПа. Десятисантиметровые волокна 5Gly/Ala-CTnat, вытянутые в обезвоженном растворителе метаноле перед высушиванием на воздухе имели предел прочности 2-3 MПа, что немного меньше, чем предел прочности у обработанных глицерином волокон такой же длины. Самый высокий измеренный в эксперименте предел прочности составлял 10 MПа, и он был получен для волокна 5Gly/Ala-CTnat длиной 10 см, высушенного на воздухе без другой обработки.
Диапазон предела прочности (2-10 MПа) сравним с более низкими значениями, опубликованными для регенерированных волокон шелка паука (2-320 MПа). Самые длинные спонтанно образованные волокна получены из конструкции 5Gly/Ala-CTnat, и такие высушенные на воздухе волокна также имеют наибольший предел прочности. Потенциально, указанный предел прочности может быть связан с наличием в данном белке поли-Ala сегментов из 12-15 остатков, по сравнению с поли-Ala сегментами из 8-14 остатков в 6Gly/Ala-CThyb, что могло бы давать большую долю кристаллической конформации β-листа в первом белке.
Способность к образованию волокон у белков MaSp1
(пример 11)
n.d. = не определен
B. Сканирующая электронная микроскопия
Микроскопическую архитектуру волокон 6Gly/Ala-CThyb и 5Gly/Ala-CTnat анализировали с помощью сканирующего электронного микроскопа (SEM) (фигура 5). Кратко, образцы помещали на целики SEM и покрывали в вакууме 6 нм слоем золота и палладия. Образцы просматривали и фотографировали с помощью микроскопа SEM LEO 1550 FEG с применением ускоряющего напряжения 10 кV.
Данным методом выявлены диаметры одиночных волокон 10-30 мкм, при этом индивидуальные волокна демонстрировали довольно однородные диаметры (показаны на фигуре 5a 6Gly/Ala-CThyb, SEQ ID NO: 13). В дополнение к макроскопическим волокнам были обнаружены гелеподобные частицы. После высушивания на воздухе данные частицы 6Gly/Ala-CThyb помещали непосредственно на целик SEM, и были видны волокна с диаметром, приблизительно, 10-15 мкм (фигуры 5b, c). Диаметр макроскопических волокон 5Gly/Ala-CTnat (SEQ ID NO: 9), вытянутых в 75% метаноле и высушенных на воздухе, составлял 60-120 мкм, и они, очевидно, содержали несколько выровненных волокон (фигуры 5d-f). Волокно, скрученное перед высушиванием на воздухе (фигура 5e), конец волокна (фигура 5f).
С. Спектроскопия кругового дихроизма
Волокна, состоящие из белка 6Gly/Ala-CThyb (SEQ ID NO: 13) или 5Gly/Ala-CTnat (SEQ ID NO: 9), в примере 8, отмывали в 20 мМ фосфатном буфере, рН 7, и суспендировали в 2% SDS в том же самом буфере. Спектры кругового дихроизма от 250 до 190 нм регистрировали при 22°С в кварцевой кювете с длиной пути 0,1 см с помощью спектрополяриметра Jasco J-810. Скорость сканирования составляла 50 нм/мин, время ответа 2 с, интервал поглощения 0,1 нм, и ширина полосы 1 нм.
Спектр, представленный на фигуре 6, представляет собой совокупность трех сканирований волокон белка 6Gly/Ala-CThyb (SEQ ID NO: 13). Спектр имеет минимум при 220 нм и максимум при 195 нм, т.е. параметры, которые характерны для антипаралелльных β-листовых структур. Очень похожие спектры были получены для волокон 5Gly/Ala-CTnat (не показаны). Таким образом, спонтанно образованные волокна демонстрируют сходную морфологию и структуру как, например, у нативных и регенирированных волокон шелка паука.
Пример 10. Биосовместимость рекомбинантного шелка паука
Поскольку желательно применение волокон шелка паука в биомедицинских областях, биосовместимость волокон оценивали путем выяснения эффектов рекомбинантно полученного шелка на различных типах клеток.
Белок MaSpl 5Gly/Ala-CTnat (SEQ ID NO: 9) экспрессировали в бактериях, как описано в примерах 7-8. Очищенный белок применяли для получения волокон искусственного шелка длиной >10 см, и даже >20-200 см, и диаметром примерно 100 мкм.
А. Эмбриональные мышиные тучные клетки
Эмбриональные (12,5-дневные) мышиные тучные клетки (пролиферация in vitro в течение восьми недель с применением IL-8 и фактора тучных стволовых клеток) в двух различных клеточных плотностях, при этом высокая клеточная плотность была в четыре раза больше низкой плотности. Указанные клетки не прилипали к пластиковой поверхности и росли в суспензии. Кусочки шелкового волокна, каждый длиной, примерно, 0,5 см, добавляли в лунки. Тучные клетки инкубировали в течение трех дней в присутствии или в отсутствие шелкового волокна, и затем подсчитывали живые и мертвые клетки после окраски трипановым синим (фигура 7). Полосками показаны средние значения со стандартной ошибкой среднего, n=2, каждую пробу подсчитывали в триплете.
Тучные клетки не подвергались изменениям в присутствии шелковых волокон. После трех дней роста не наблюдали значительных различий в клеточной смерти или пролиферации, по сравнению с отрицательными контролями, которые выращивали без шелковых волокон.
Тестированные тучные клетки не подвергались изменениям в присутствии рекомбинантных шелковых волокон, даже при сравнительно высоких количествах шелка. Указанные результаты показывают, что тестируемые волокна искусственного шелка имеют сходство с диким типом канатного шелка Euprosthenops australis и являются нетоксичными и биосовместимыми. Таким образом, волокна искусственного шелка представляются подходящими для биомедицинских применений.
Пример 11. Механические свойства и структура волокон из белка MaSpl
Механические свойства волокон из 5Gly/Ala-CTnat (SEQ ID NO: 9) проверяли с помощью тестов на определение предела прочности, при осуществлении которых получали кривые зависимости деформации от напряжения (фигура 9). Свойства прочности при растяжении определяли с помощью прибора Zwick Roell Z2.5 material tester (Zwick, Ulm, Germany). Тесты осуществляли в воздухе при комнатных условиях (20°С и 52% относительной влажности), используя скорость нагрузки 10 мм/мин. Кусочки волокон брали непосредственно из буфера, закрепляли в приборе и подвергали двум циклам растяжения-расслабления. Для получения однородной шелковой нити, подходящей для теста на растяжение, волокна растягивали с помощью циклов растяжения-расслабления. Сначала волокна удлиняли путем вытягивания с силой 0,1 Н. После расслабления волокна продолжали вытягивать до приложения силы в 0,25 Н.
В результате такой обработки получали вытянутые однородные волокна с диаметром приблизительно 80 мкм, как определяли по измерениям высоты с помощью прибора Mitutoyo IDC-112B instrument (Mitutoyo Corp, Tokyo, Japan) и подтверждали сканирующей электронной микроскопией (SEM), как описано далее. До и после циклов вытягивания-расслабления кусочки волокон наносили на целики SEM и высушивали на воздухе в течение ночи. Образцы покрывали под вакуумом 6 нм слоем золота и палладия. Образцы просматривали и фотографировали с помощью микроскопа LEO 1550 FEG (Carl Zeiss, Oberkochen, Germany) с применением ускоряющего напряжения 10 kV.
Вытянутые волокна были нарезаны на кусочки, концы которых закрепляли клеем между плотной бумагой (Loctite 420, Loctite, Göteborg, Sweden). Затем образцы волокон закрепляли в губках тисков прибора для испытания материалов и растягивали до их разрыва. Кривые зависимости деформации от напряжения построены с применением начальной площади поперечного сечения невытянутого волокна, исходя из круглого поперечного сечения. Значения напряжения нормализованы относительно начальной площади сечения волокна. Деформация соответствует dL/L0, где L0 представляет собой начальную длину волокна и dL представляет собой изменение длины волокна. На фигуре 9 показаны кривые зависимости напряжение-деформация для трех различных образцов дважды вытянутых волокон 5Gly/Ala-CTnat (SEQ ID NO: 9), и их пределы прочности, оцененные приблизительно в 0,2 ГПа.
Микроскопическую архитектуру волокон анализировали с помощью SEM (фигура 10). Спонтанно сформированные волокна имеют однородный сглаженный внешний вид и ширину до нескольких сотен микрометров, в то время как высота оценивается в несколько десятков микрометров (фигура 10а, b).
После того как волокна подвергали циклам вытягивания-расслабления, их поперечное сечение принимало более округлую форму с компактной субструктурой из сильно выровненных фибрилл (фигура 10с-f). Внешний вид разрезанных или разрушенных поверхностей (фигура 10е, f) дополнительно подтверждает компактность полученного волокна.
В заключение следует отметить, что спонтанно образованные волокна и без формования демонстрируют морфологию и механические свойства, сходные с нативными или регенерированными волокнами шелка паука.
Пример 12. Варианты спидроинового белка
Предполагают, что сильные межмолекулярные взаимодействия вносят свой вклад в необычайно высокий предел прочности шелка паука. Поэтому были созданы варианты небольших спидроинов, которые позволяют внутримолекулярное ковалентное перекрестное сшивание в волокнах. Два различных мутантных спидроиновых белка были сконструированы путем сайт-направленного мутагенеза для введения двух остатков цистеина в первый (SEQ ID NO: 14, положения 36 и 37) и в четвертый (SEQ ID NO: 15, положения 128 и 129) аланиновые блоки, соответственно. Данные варианты были экспрессированы и выделены с помощью того же протокола, который описан в примерах 7-8 для генов, сконструированных в примерах 5-6. Данные варианты (SEQ ID NOS: 14-15) образуют волокна таким же образом, как 5Gly/Ala-CTnat (SEQ ID NO: 9).
Чтобы выяснить значение димеризации С-концевого домена, был построен вариант, в котором остаток цистеина в С-концевом домене заменяли остатком серина (SEQ ID NO: 16, положение 222). Несмотря на это данный вариант (SEQ ID NO: 16) формировал волокна таким же образом, как 5Gly/Ala-CTnat (SEQ ID NO: 9).
Пример 13. Удаление LPS и других пирогенов из экспрессированных спидроиновых белков
Клетки Е.coli, экспрессирующие желаемый спидроиновый слитый белок, отмывают следующими буферами:
А: 100 мМ Tris, pH 8,
В: 5 мМ СаСl2, 100 мМ Tris, pH 8,
С: 10 мМ EDTA, 100 мМ Tris, pH 8,
D: 100 мМ Tris, pH 8, и
Е: 100 мМ Tris, pH 8.
Затем клетки разрушали в 20 мМ Tris, pH 8 дополненном лизозимом и ДНКазой I. Затем пробу белка помещали на Ni-сефарозный матрикс и отмывали 20 мМ Tris, 10-100 мМ имидазолом, рН 8 перед элюированием 20 мМ Tris, 100-300 мМ имидазолом, рН 8. Соответствующие фракции собирали и проводили диализ против 20 мМ Tris, pH 8 в течение ночи. Пробу белка затем дополняют 100 мкМ СаСl2 и в заключение пропускают через колонку EndoTrap Blue, предварительно уравновешенную 20 мМ Tris, 100 мкМ СаСl2, рН 8. Таким образом, могут быть получены пробы белка с содержанием пирогена 1 ЭЕ/мг белка согласно оценке IPT и кинетического теста LAL.
Затем слитый белок протеолитически расщепляют тромбином в соотношении 1:1000 (масс./масс.) тромбин:слитый белок, что вызывает образование волокон (как описано выше). Волокна отмывали 3 раза в 20 мМ Tris, pH 8 и в заключение 3 раза в воде. Получают волокна с содержанием пирогенов 0,25 ЭЕ/мг волокна.
Структурные характеристики волокон не изменяются под действием автоклавирования при 125°С и 1,5 бар в течение 10 мин, что обеспечивает эффективную стерилизацию материала. Волокна являются химически стабильными и не растворяются в 8 М мочевине, 6 М GuaHCl или в чистом НАс. Однако волокна могут растворяться в чистом HFIP или в муравьиной кислоте.



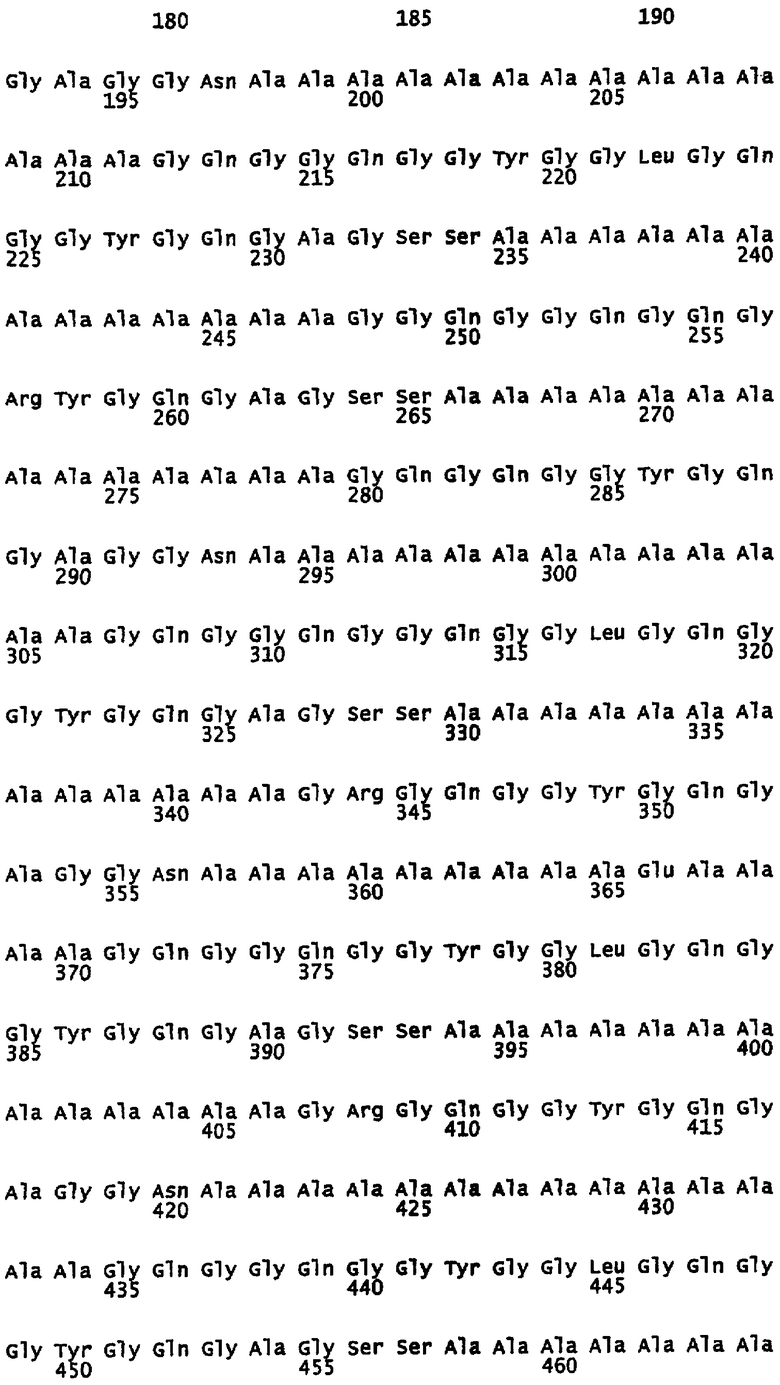
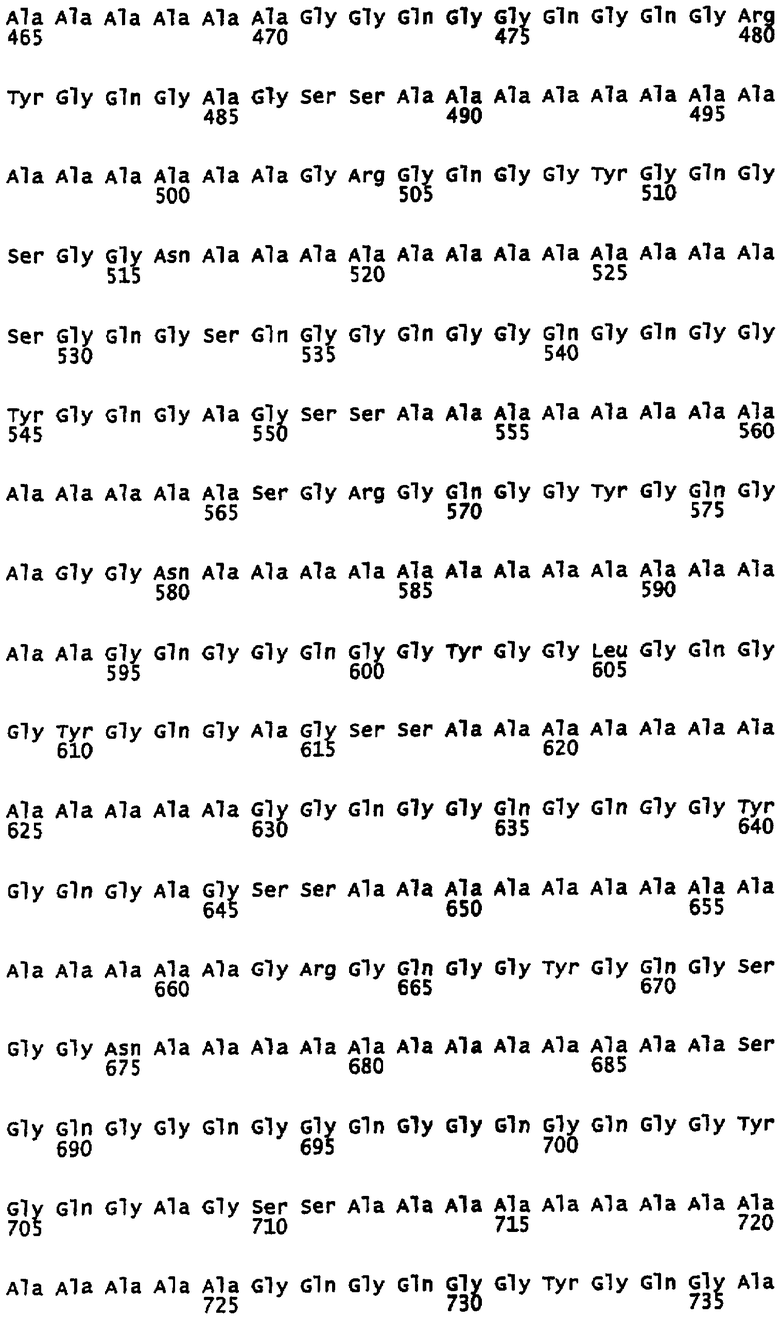






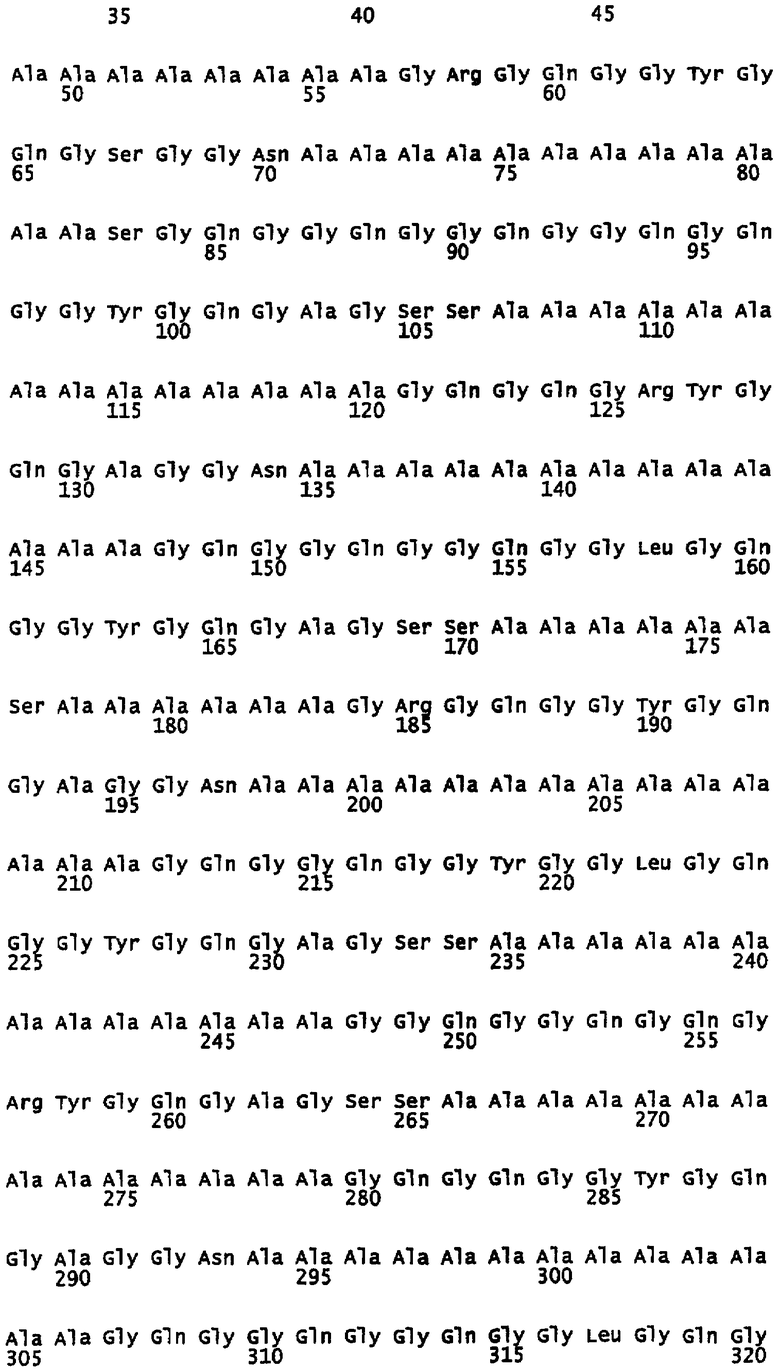

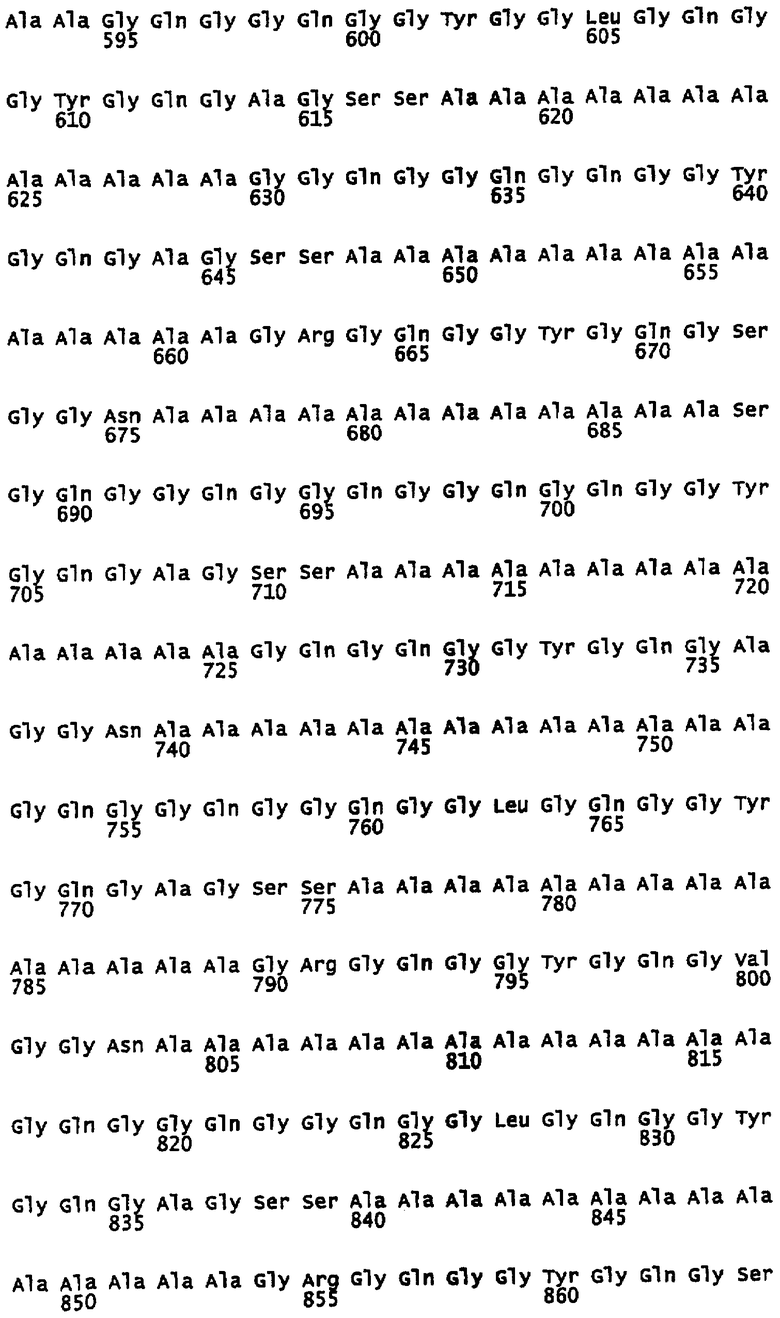
















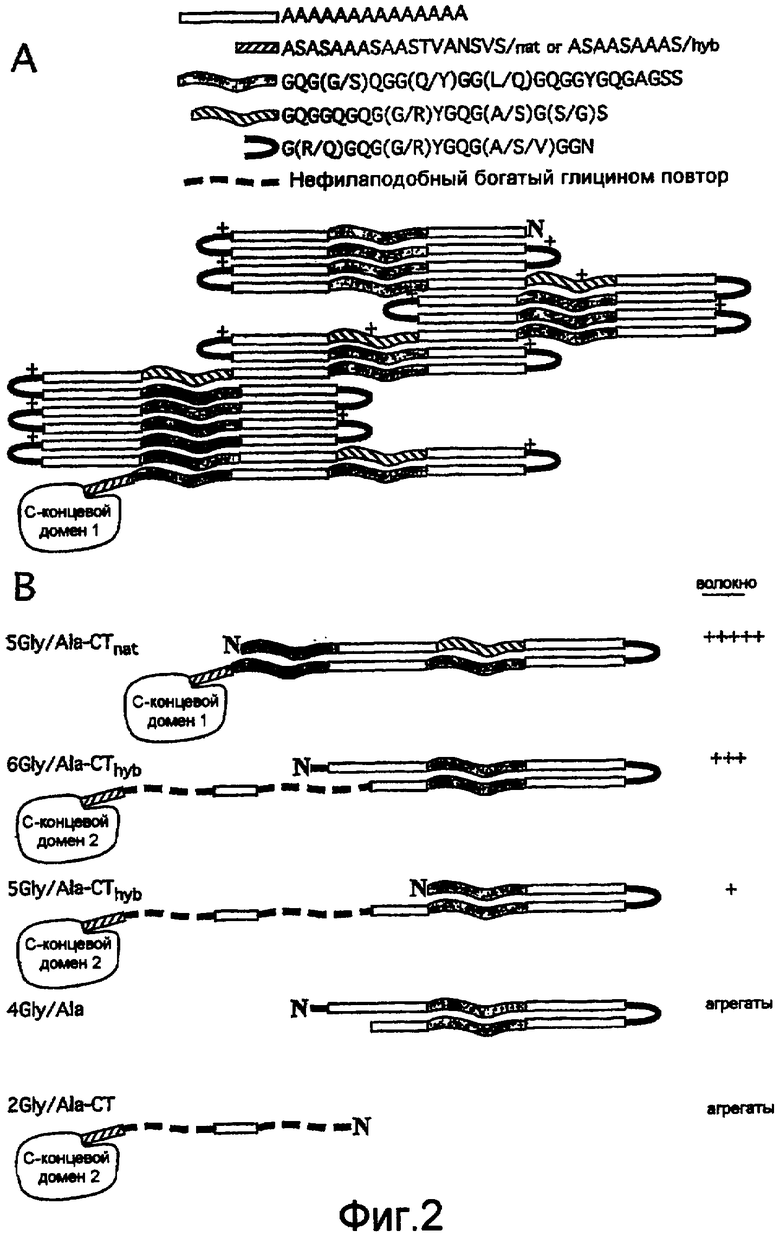

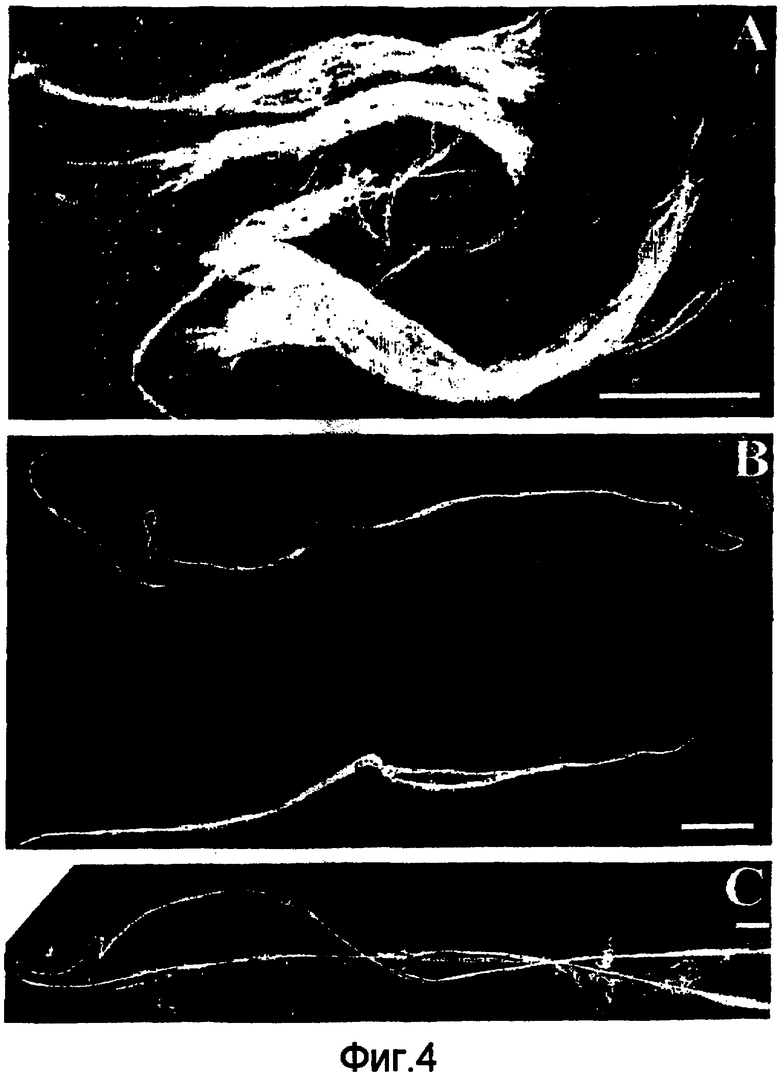
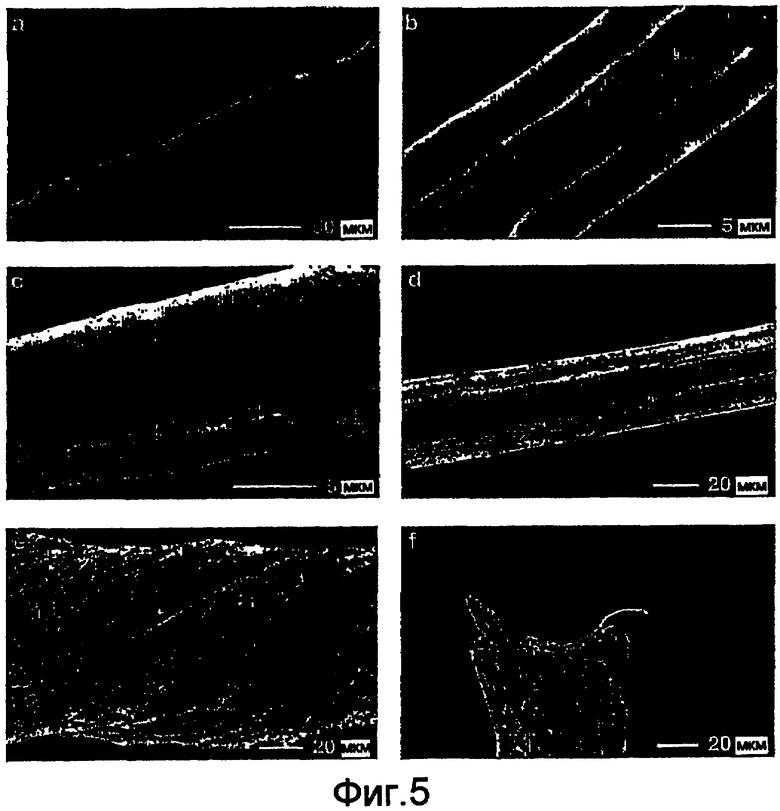
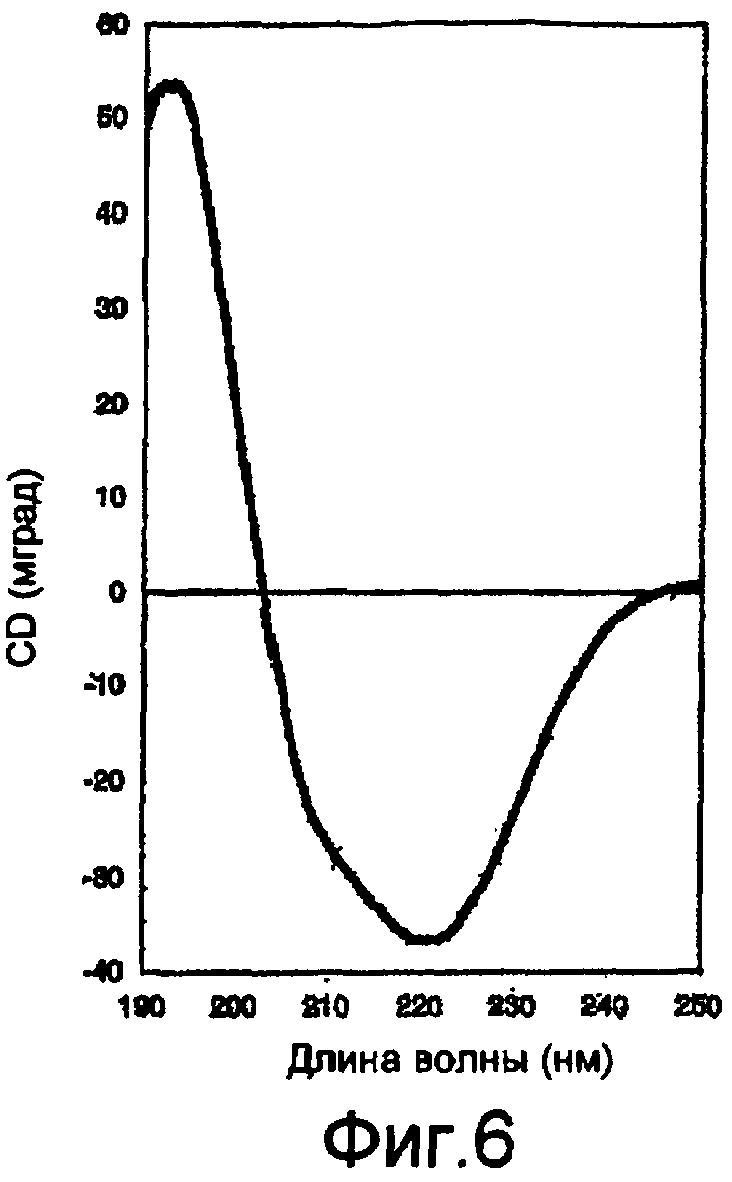
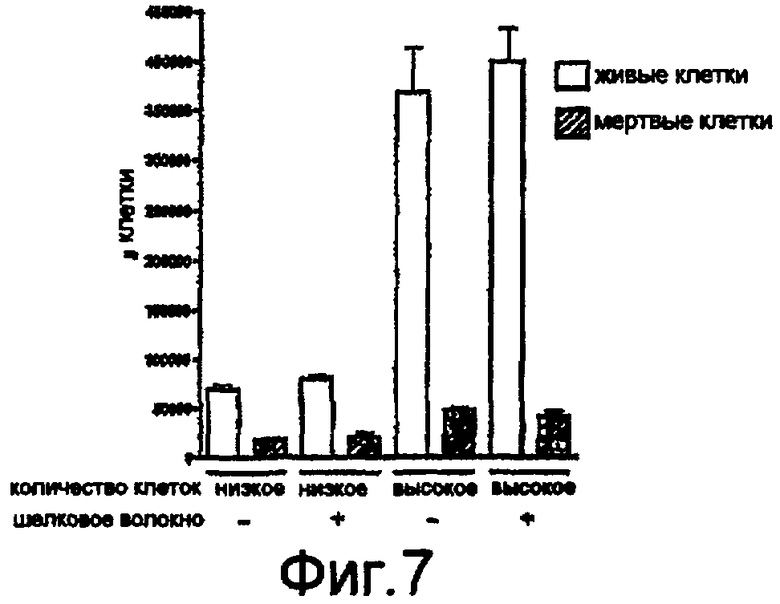
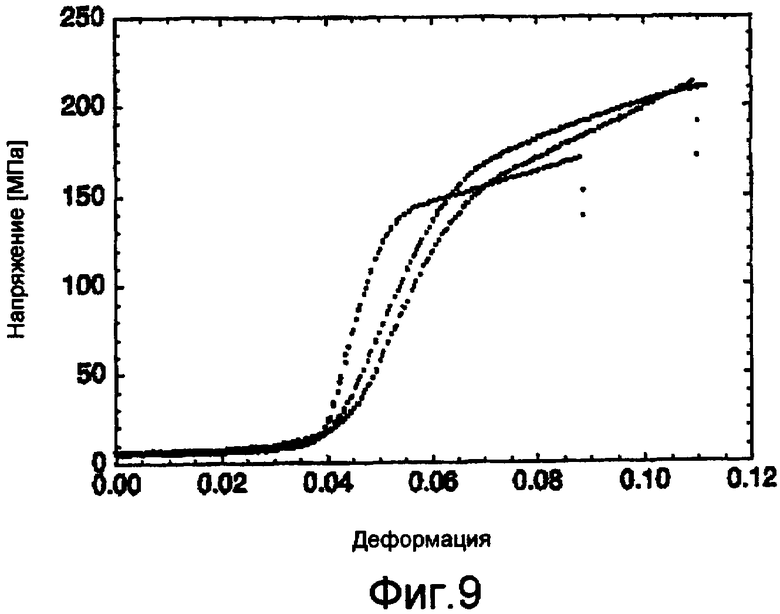

Изобретение относится к области биотехнологии, конкретно к получению белков паука, и может быть использовано в медицине. Рекомбинантным путем получают спидроиновый белок главной ампулярной железы, который состоит из 150-420 аминокислотных остатков и описывается формулой REP-CT, где REP - это фрагмент белка из 80-300 аминокислотных остатков, выбранный из L(AG)nL, L(AG)nAL, L(GA)nL или L(GA)nGL, где n - целое число от 4 до 8. Каждый индивидуальный сегмент А состоит из 8-18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, а остальные - представляют собой Ala. Каждый отдельный сегмент G состоит из 12-30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков являются Gly. Каждый индивидуальный сегмент L является линкерной аминокислотной последовательностью из 0-20 аминокислотных остатков. СТ - это фрагмент от 70 до 120 аминокислотных остатков С-концевого фрагмента спидроинового белка главной ампулярной железы. Изобретение позволяет получить спидроиновый белок главной ампулярной железы, способный к полимеризации, а также на его основе получить слитый белок, способный к управляемой полимеризации. 9 н. и 6 з.п. ф-лы, 1 табл., 9 ил., 13 пр.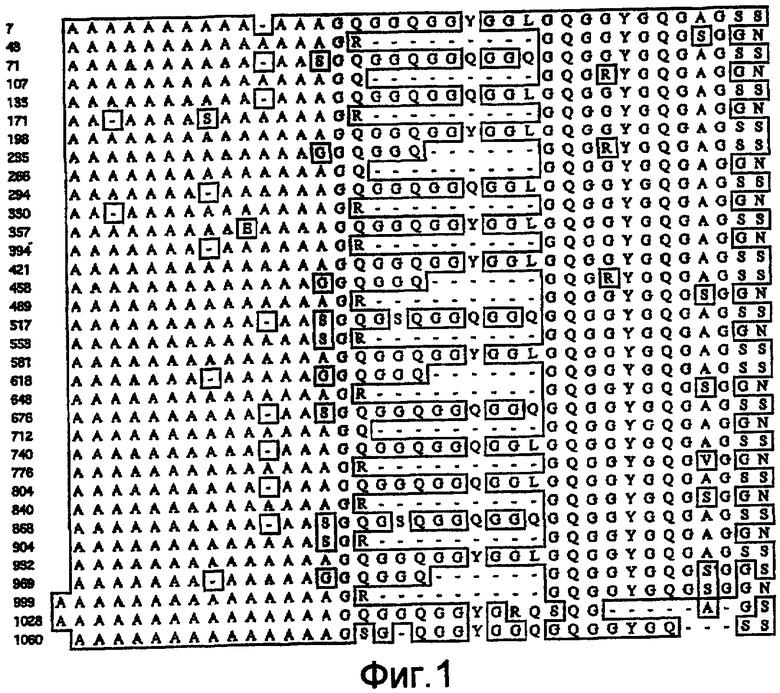
1. Рекомбинантный спидроиновый белок главной ампулярной железы, способный к полимеризации, отличающийся тем, что указанный белок состоит из 150-420 аминокислотных остатков и описывается формулой REP-CT, где
REP представляет собой фрагмент белка, имеющий 80-300 аминокислотных остатков, где указанный фрагмент выбран из группы L(AG)nL, L(AG)nAL, L(GA)nL, L(GA)nGL, где n представляет собой целое число от 4 до 8; каждый индивидуальный сегмент А представляет собой аминокислотную последовательность из 8-18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala; каждый отдельный сегмент G является аминокислотной последовательностью из 12-30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков являются Gly; и каждый индивидуальный сегмент L является линкерной аминокислотной последовательностью из 0-20 аминокислотных остатков; и
СТ представляет собой фрагмент белка, имеющий от 70 до 120 аминокислотных остатков, являющийся С-концевым фрагментом, полученным из спидроинового белка главной ампулярной железы.
2. Рекомбинантный белок по п.1, в котором каждый индивидуальный сегмент А идентичен или, по меньшей мере, на 80% идентичен аминокислотной последовательности, выбранной из группы аминокислотных остатков 7-19, 43-56, 71-83, 107-120, 135-147, 171-183, 198-211, 235-248, 266-279, 294-306, 330-342, 357-370, 394-406, 421-434, 458-470, 489-502, 517-529, 553-566, 581-594, 618-630, 648-661, 676-688, 712-725, 740-752, 776-789, 804-816, 840-853, 868-880, 904-917, 932-945, 969-981, 999-1013, 1028-1042 и 1060-1073 SEQ ID NO: 3; аминокислотных остатков 31-42, 61-75, 90-104, 122-135 и 153-171 SEQ ID NO: 9; аминокислотных остатков 12-25, 46-60, 75-88, 112-119, 150-158 и 173-180 SEQ ID NO: 13; аминокислотных остатков 31-42 SEQ ID NO: 14; и аминокислотных остатков 122-135 SEQ ID NO: 15;
каждый индивидуальный сегмент G идентичен или, по меньшей мере, на 80% идентичен аминокислотной последовательности, выбранной из группы аминокислотных остатков 20-42, 57-70, 84-106, 121-134, 148-170, 184-197, 212-234, 249-265, 280-293, 307-329, 343-356, 371-393, 407-420, 435-457, 471-488, 503-516, 530-552, 567-580, 595-617, 631-647, 662-675, 689-711, 726-739, 753-775, 790-803, 817-839, 854-867, 881-903, 918-931, 946-968, 982-998, 1014-1027, 1043-1059 и 1074-1092 SEQ ID NO: 3; SEQ ID NO: 5-7; аминокислотных остатков 11-30, 43-60, 76-89, 105-121 и 136-152 SEQ ID NO: 9; и аминокислотных остатков 1-11, 26-45, 61-74, 89-111, 120-149 и 159-172 SEQ ID NO: 13; и
указанный фрагмент СТ идентичен или, по меньшей мере, на 50% идентичен SEQ ID NO: 8; или где указанный фрагмент СТ идентичен или, по меньшей мере, на 80% идентичен аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 4, аминокислотных остатков 172-269 SEQ ID NO: 9, аминокислотных остатков 181-276 SEQ ID NO: 13, и аминокислотных остатков 172-269 SEQ ID NO: 16.
3. Рекомбинантный белок по любому из пп.1-2, выбранный из группы, состоящей из SEQ ID NO: 9 и SEQ ID NO: 12-16.
4. Рекомбинантный слитый белок, способный к управляемой полимеризации, выбранный из группы X-REP-CT и REP-CT-X, где REP и СТ являются фрагментами белка по любому из пп.1-3; и
X является фрагментом белка, содержащим партнера по слитию и участок узнавания расщепляющего агента;
где комбинированный фрагмент белка REP-CT соединяется через указанный участок узнавания расщепляющего агента с указанным партнером по слитию.
5. Полимер спидроинового белка главной ампулярной железы по любому из пп.1-3 для применения в качестве материала для медицинских или технических целей.
6. Композиция для управляемой полимеризации, включающая в себя эффективное количество рекомбинантного белка по п.4, в которой содержание LPS и других пирогенов составляет 1 ЭЕ/мг выделенного белка или менее.
7. Рекомбинантная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, которая кодирует рекомбинантный спидроиновый белок главной ампулярной железы по любому из пп.1-3, или последовательность нуклеиновой кислоты, комплементарную ей.
8. Рекомбинантная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, которая кодирует слитый белок по п.4, или последовательность нуклеиновой кислоты, комплементарную ей.
9. Способ получения растворимого слитого белка, описанного в п.4, включающий в себя стадии:
(i) экспрессии молекулы нуклеиновой кислоты, которая кодирует слитый белок по п.4, в подходящем хозяине; и
(ii) выделения растворимого слитого белка, полученного на стадии (i), если надо, включающая в себя удаление LPS и других пирогенов.
10. Способ получения полимера спидроинового белка главной ампулярной железы, описанного в любом из пп.1-3, включающий в себя стадии:
(i) предоставления раствора слитого белка по п.4 в жидкой среде,
(ii) добавления к указанной жидкой среде подходящего расщепляющего агента для расщепления слитого белка в участке узнавания расщепляющего агента, и получения благодаря этому спидроинового белка главной ампулярной железы;
(iii) вызывания полимеризации спидроинового белка главной ампулярной железы, полученного на стадии (ii) в жидкой среде; и, если надо, (iv) выделения полимера, полученного на стадии (iii) из указанной жидкой среды, если надо, включающее в себя удаление LPS и других пирогенов.
11. Способ по п.10 получения полимера спидроинового белка главной ампулярной железы, описанного в пп.1-3, в котором стадия (i) предоставления раствора слитого белка по п.4 состоит в:
(a) экспрессии молекулы нуклеиновой кислоты, которая кодирует слитый белок по п.4, в подходящем хозяине;
(b) выделении растворимого слитого белка, полученного на стадии (i), если надо, включающего в себя удаление LPS и других пирогенов; и
(c) предоставлении раствора указанного растворимого слитого белка, полученного на стадии (и) в жидкой среде.
12. Способ по любому из пп.10 и 11, в котором указанная стадия (iii) вызывания полимеризации в жидкой среде спидроинового белка главной ампулярной железы, полученного на стадии (ii), далее включает в себя создание границы раздела между указанной жидкой средой и другой фазой, выбранной из группы, состоящей из газовой фазы, жидкой фазы и твердой фазы, где указанная полимеризация инициируется на указанной границе раздела или в области, окружающей указанную границу раздела.
13. Способ по п.12, где указанная жидкая среда представляет собой водную среду, и указанная другая фаза выбрана из группы, состоящей из органических растворителей, несмешивающихся с воздухом и водой.
14. Способ культивирования эукариотических клеток, включающий стадию выращивания указанных эукариотических клеток, за исключением эмбриональных клеток человека, на материале, который включает полимер по п.5.
15. Способ по п.14, где указанный материал включает волокна указанного полимера.
| WO 03020916 А2, 13.03.2003 | |||
| LEWIS R.V | |||
| et al | |||
| Expression and purification of a spider silk protein: a new strategy for producing repetitive proteins, Protein Expr | |||
| Purif., 1996, v.7, n.4, p.400-406 | |||
| POUCHKINA-STANTCHEVA N.N | |||
| et al | |||
| Molecular studies of a novel dragline silk from a nursery web spider, Euprosthenops sp | |||
| (Pisauridae), Соmр | |||
| Biochem. |

