Область техники, к которой относится изобретение
Настоящее изобретение относится к области рекомбинантных слитых белков, и, более конкретно, к слитым белкам, содержащим части, происходящие из белков шелка пауков (спидроинов). Настоящее изобретение относится к способам предоставления белковой структуры, которая представляет собой полимер, содержащий рекомбинантный слитый белок, который содержит части, происходящие из спидроинов. Также предусмотрены новые белковые структуры для связывания с органической мишенью.
Уровень техники
В прикладной химии белков частой проблемой является то, как создать или предоставить биологически активный пептид или белок в соответствующую область активности, как правило, к органической мишени, такой как нуклеиновая кислота, белок, комплекс белков, или комплекс белка(ов) и/или липидов и/или углеводов и/или нуклеиновой кислоты(кислот). Наиболее простым решением является предоставление просто водного раствора биологически активного пептида или белка. Однако многие применения требуют некоторых других средств для достижения желаемой цели. Например, пептиды/белки можно связывать со смесью липидов или химически иммобилизовывать на подложку.
Применения для пептидов/белков, иммобилизованных на подложку, включают методики препаративного и аналитического разделения, такие как биопроцессы, хроматография, улавливание и культура клеток, активные фильтры и диагностические способы. Структуры на основе белков внеклеточного матрикса, например коллагена, описаны в EP 704 532 и EP 985 732.
Также было предложено использовать белки шелка пауков в подложках. Шелк пауков представляет собой высококачественные природные полимеры, достигающие необычайной прочности и растяжимости вследствие комбинации сопротивления разрыву и упругости. Пауки имеют вплоть до семи различных желез, которые продуцируют различные типы шелка с различными механическими свойствами и функциями. Каркасная нить, продуцируемая большой ампуловидной железой, является наиболее прочной нитью. Она состоит из двух основных полипептидов, как правило, называемых спидроинами большой ампуловидной железы (MaSp) 1 и 2, но, например, ADF-3 и ADF-4 в Araneus diadematus. Эти белки имеют молекулярную массу в диапазоне 200-720 кДа. Белки каркасной нити пауков, или MaSp, имеют трехкомпонентный состав: неповторяющийся N-концевой домен, центральную повторяющуюся область, содержащую множество повторяющихся сегментов поли-Ala/Gly, и неповторяющийся C-концевой домен. Главным образом, полагают, что повторяющаяся область образует межмолекулярные контакты в волокнах шелках, в то время как точные функции концевых доменов менее понятны. Также полагают, что в сочетании с образованием волокон повторяющаяся область претерпевает структурное преобразование от случайной спирали и α-спиральной конформации до структуры β-слоя. C-концевая область спидроинов, главным образом, консервативна среди видов пауков и типов шелка.
В WO 07/078239 и Stark, M. et al., Biomacromolecules 8: 1695-1701, (2007) описан уменьшенный белок шелка пауков, состоящий из повторяющегося фрагмента с высоким содержанием Ala и Gly и C-концевого фрагмента белка, а также растворимые слитые белки, содержащие белок шелка пауков. Волокна белка шелка пауков образуются самопроизвольно при освобождении белка шелка пауков от его партнера по слиянию.
В Rising, A. et al., CMLS 68(2): 169-184 (2010) рассмотрены достижения в получении белков шелка пауков.
В US 2009/0263430 описано химическое присоединение фермента β-галактозидазы к пленкам укороченного белка шелка пауков. Однако химическое присоединение может требовать условий, которые неблагоприятны для стабильности и/или функции белка. Были разработаны белки, содержащие многократные повторы сегмента, происходящего из повторяющейся области белков шелка пауков, включающие сегмент связывания клеток RGD (Bini, E et al., Biomacromolecules 7:3139-3145 (2006)) и/или пептид R5 (Wong Po Foo, C et al., Proc Natl Acad Sci 103 (25): 9428-9433 (2006)) или другие белковые сегменты, вовлеченные в минерализацию (Huang, J et al., Biomaterials 28: 2358-2367 (2007); WO 2006/076711). В этих документах уровня техники пленки формируют путем растворения слитых белков в денатурирующем органическом растворителе гексафторизопропаноле (HFIP) и высушивания.
В US 2005/261479 A1 описан способ очистки рекомбинантных белков шелка, имеющих аффинную метку, вовлекающий магнитное выделение по аффинности индивидуальных белков шелка из комплексных смесей без образования волокон белка шелка или других полимерных структур.
Известные положки и ассоциированные с ними способы имеют определенные недостатки в отношении, например, экономичности, эффективности, стабильности, способности к регенерации, биоактивности и биосовместимости.
Сущность изобретения
Задачей настоящего изобретения является предоставление новой белковой структуры, которая способна к селективному взаимодействию с органической мишенью.
Также задачей настоящего изобретения является предоставление белковой структуры, которая способна к селективному взаимодействию с органической мишенью, где структура образована без жестких растворителей, которые имеют непредсказуемый эффект на вторичную структуру или активность белка и/или остаются в белковой структуре.
Одной из задач настоящего изобретения является предоставление стабильной белковой структуры, которая способна к селективному взаимодействию с органической мишенью, которую можно легко регенерировать после применения, например, с помощью химической обработки.
Другой задачей настоящего изобретения является предоставление стабильной белковой структуры, которая является биосовместимой и подходящей для клеточной культуры и в качестве имплантата.
Другой задачей изобретения является предоставление белковой структуры с высокой плотностью находящихся на равных расстояниях функциональных групп, которые способны селективно взаимодействовать с органической мишенью.
Кроме того, задачей изобретения является предоставление белковой структуры, которая сохраняет ее способность селективного связывания при хранении при +4°C или при комнатной температуре в течение месяцев.
Также задачей изобретения является предоставление белковой структуры, которая поддается автоклавированию, т.е. сохраняет ее способность к селективному связыванию после стерилизующей тепловой обработки.
Для этих и других задач будет очевидно из представленного ниже описания, что, согласно первому аспекту, настоящее изобретение относится к слитому белку и белковой структуре, состоящей из полимеров, содержащих в качестве повторяющегося структурного элемента слитый белок, как указано в формуле изобретения.
В соответствии с родственным аспектом, настоящее изобретение относится к выделенной полинуклеиновой кислоте, кодирующей слитый белок, и к способу получения слитого белка, как указано в формуле изобретения.
Согласно другому аспекту, настоящее изобретение относится к способу предоставления белковой структуры, как указано в формуле изобретения.
Согласно следующему аспекту, настоящее изобретение относится к аффинной среде, как указано в формуле изобретения.
Согласно одному аспекту, настоящее изобретение относится к каркасному материалу для клеток, как указано в формуле изобретения. Согласно родственному аспекту, настоящее изобретение также относится к комбинации клеток и каркасного материала для клеток в соответствии с формулой изобретения.
Согласно одному аспекту, настоящее изобретение относится к новым применениям белковой структуры и слитого белка, как указано в формуле изобретения.
Согласно другому аспекту, настоящее изобретение относится к способу выделения органической мишени из образца, как указано в формуле изобретения.
Согласно следующему аспекту, настоящее изобретение относится к способу иммобилизации и необязательно культивирования клеток, как указано в формуле изобретения.
Краткое описание чертежей
На фиг. 1 представлено выравнивание последовательностей C-концевых доменов спидроинов.
На фиг. 2 представлено выравнивание последовательностей N-концевых доменов спидроинов.
На фиг. 3 представлено макроскопическое волокно слитого белка, содержащего Z-домен.
На фиг. 4 представлен гель SDS-PAGE после очистки и анализа слитого белка, содержащего Z-домен.
На фиг. 5 представлены волокна, изготовленные из слитых белков, и контрольные волокна, которые иллюстрируют функциональность Z-домена в слитом белке.
На фиг. 6 представлена часть сформированной пленки, изготовленной из слитых белков, на дне планшета для культивирования тканей.
На фиг. 7-12 представлены невосстанавливающие гели SDS-PAGE, иллюстрирующие функциональность Z-домена в структурах слитых белков.
На фиг. 13-15 представлены невосстанавливающие гели SDS-PAGE, иллюстрирующие способность Z-домена в структурах слитых белков связывать IgG по сравнению с коммерческой матрицей с белком A.
На фиг. 16-17 представлены невосстанавливающие SDS-PAGE гели при процессах очистки на месте (CIP) структур слитых белков по сравнению с матрицей с белком A.
На фиг. 18 представлена интенсивность флуоресценции белковых пленок, смоченных биотинилированным Atto-565.
На фиг. 19 представлен график и линейная аппроксимация величин интенсивности флуоресценции для различных концентраций биотинилированного Atto-565 при связывании с пленкой слитого белка.
На фиг. 20 представлены графики величин интенсивности флуоресценции до (-) и после (+) добавления биотинилированного Atto-565 в лунки с пленками, изготовленными из слитого белка или контроля.
На фиг. 21 представлен график, на котором показана стандартная кривая и линейная аппроксимация полученных скоростей реакции при катализе свободной биотинилированной HRP в растворе.
На фиг. 22 представлен график, демонстрирующий скорость реакции при катализе биотинилированной HRP, иммобилизованной на пленке слитого белка, по сравнению с контролем.
На фиг. 23 представлен восстанавливающий гель SDS-PAGE с растворенными структурами слитых белков.
На фиг. 24-26 представлены графики, иллюстрирующие связывание IgG-HRP с пленкой слитого белка, содержащего Z-домены.
На фиг. 27 представлены графики, иллюстрирующие связывание IgG-Alexa Fluor 633 с пленкой слитого белка, содержащего Z-домены.
На фиг. 28 представлен невосстанавливающий гель SDS-PAGE, иллюстрирующий функциональность Z-домена в слитом белке после автоклавирования.
На фиг. 29 представлен гель SDS-PAGE продуктов расщепления при обработке протеазой 3C волокна слитого белка, содержащего Z-домены.
На фиг. 30 представлен невосстанавливающий гель SDS-PAGE, иллюстрирующий функциональность Z-домена в структурах слитого белка, образованных в присутствии органической мишени (IgG).
На фиг. 31-32 представлены невосстанавливающие гели SDS-PAGE, иллюстрирующие функциональность Abd-домена в структурах слитых белков.
На фиг. 33-34 представлены невосстанавливающие гели SDS-PAGE, иллюстрирующие функциональность C2-домена в структурах слитых белков.
Список прилагаемых последовательностей
SEQ ID NO
1 4Rep
2 4RepCT
3 NT4Rep
4 NT5Rep
5 NT4RepCTHis
6 NT
7 CT
8 консенсусная последовательность NT
9 консенсусная последовательность CT
10 повторяющаяся последовательность из MaSp1 Euprosthenops australis
11 консенсусная последовательность 1 G-сегмента
12 консенсусная последовательность 2 G-сегмента
13 консенсусная последовательность 3 G-сегмента
14 HisZQG4Rep4CT
15 HisZQG4Rep4CT (ДНК)
16 HisAbdQG4RepCT
17 HisAbdQG4RepCT (ДНК)
18 HisC2QG4RepCT
19 HisC2QG4RepCT (ДНК)
20 4RepCT 2
21 4RepCT 2 (ДНК)
22 M44RepCT
23 M44RepCT (ДНК)
24 modM44RepCT
25 modM44RepCT (ДНК)
26 4RepCTM4
27 4RepCTM4 (ДНК)
Подробное описание изобретения
Настоящее изобретение, главным образом, основано на идее, что можно получать твердые белковые структуры, способные селективно взаимодействовать с органической мишенью, в форме полимеров рекомбинантного слитого белка в качестве повторяющейся структурной единицы. Слитый белок содержит по меньшей мере одну неспидроиновую часть из более чем 30 аминокислотных остатков, которая способна селективно взаимодействовать с органической мишенью, и части, соответствующие по меньшей мере повторяющемуся и C-концевому фрагментам белка шелка пауков. Неожиданно, можно индуцировать структурную реорганизацию частей, происходящих из белка шелка пауков, и в результате образование полимерных твердых структур, в то время как неспидроиновая часть не подвергается структурной реорганизации, но сохраняет ее желаемую структуру и функцию, т.е. способность к селективному взаимодействию с органической мишенью. Белковые структуры можно получать без стадии химического присоединения или стадии способа денатурации, которая облегчает процесс и повышает вероятность получения слитого белка с сохраненной функциональностью его частей, в частности, когда функции зависят от вторичной структуры частей. Образование этих полимеров слитого белка можно тщательно контролировать, и в результате развития этой идеи были достигнуты представленные ниже новые белковые структуры, способы получения белковых структур и применения белковых структур в различных применениях и способах.
Таким образом, слитый белок по изобретению обладает как желаемой активностью селективного взаимодействия, так и присущей ему активностью твердой подложки, которые реализуются в белковой структуре в физиологических условиях. Необходимо учитывать, что является неожиданным, что активность связывания слитого белка сохраняется, несмотря на ковалентное присоединение неспидроиновой части к спидроиновой части, когда последняя структурно реорганизуется с образованием полимерных твердых структур. В действительности, стабильность при нагревании и/или химическая стабильность и/или связывающая активность части, обеспечивающей активность селективного взаимодействия, могут увеличиваться при встраивании в структуру слитого белка по изобретению. Белковая структура также обеспечивает высокую и предсказуемую плотность активности селективного взаимодействия с органической мишенью. Утрата ценных белковых частей с активностью селективного взаимодействия минимизируется, поскольку все экспрессированные белковые части связаны с твердой подложкой.
Полимеры, которые образованы из слитых белков по изобретению, представляют собой твердые структуры и пригодны по их физическим свойствам, особенно пригодна комбинация высокого сопротивления разрыву, упругости и легкой массы. Особенно пригодным признаком является то, что происходящие из спидроина части слитого белка являются биохимически стабильными и пригодными для регенерации, например, кислотой, основанием или хаотропными агентами, и пригодны для стерилизации нагреванием, например, автоклавирования при 120°C в течение 20 мин. Полимеры также пригодны вследствие их способности обеспечивать прикрепление и рост клеток. Свойства каркасной нити являются привлекательными для разработки новых материалов для медицинских или технических целей. В частности, белковые структуры по изобретению пригодны для препаративных и аналитических процессов разделения, таких как хроматография, улавливание клеток, селекция и культивирование, активные фильтры и диагностические способы. Белковые структуры по изобретению также пригодны в медицинских устройствах, таких как имплантаты и медицинские продукты, такие как системы закрытия раны, лейкопластыри, швы, повязки на рану и каркасы для иммобилизации клеток, культивирования клеток, инженерии ткани и направленной регенерации клеток.
Настоящее изобретение относится к рекомбинантному слитому белку, который способен к селективному взаимодействию с органической мишенью, причем этот слитый белок содержит части B, REP и CT, и необязательно NT. Настоящее изобретение также относится к белковой структуре, которая способна к селективному взаимодействию с органической мишенью, где указанная белковая структура представляет собой полимер, содержащий, и необязательно состоящий из, рекомбинантный слитый белок согласно изобретению, т.е. содержащий, и необязательно состоящий из, части B, REP и CT, и необязательно NT.
Хотя части REP и CT слитых белков в примерах всегда относятся к конкретным белкам, например белкам, происходящим из большого спидроина 1 (MaSpl) из Euprosthenops australis, считается, что настоящее описание применимо к любым структурно сходным частям для цели получения структур слитых белков по изобретению. Более того, хотя часть B, которая обеспечивает активность селективного взаимодействия слитых белков, в примерах всегда относится к конкретным белковым частям, например, частям, происходящим из белка A, белка G и стрептавидина, считается, что настоящее изобретение применимо к любой структурно и/или функционально сходной части B для цели получения структур слитых белков по изобретению, способных к селективному взаимодействию с органической мишенью.
Конкретные слитые белки согласно изобретению определяются формулами Bx-REP-By-CT-Bz и Bx-CT-By-REP-Bz, где x, y и z представляют собой целые числа от 0 до 5; и x+y+z≥1, необязательно дополнительно содержащими одну часть NT на любом конце слитого белка или между любыми двумя белковыми частями в слитом белке. Если x+y+z>1, т.е. если существует две или более частей B, они могут быть идентичными или могут отличаться. Две или более частей B могут обладать способностью селективного взаимодействия с одной и той же органической мишенью или с различными органическими мишенями. Предпочтительно, чтобы две или более части B были по существу идентичными, каждая из которых обладает способностью к селективному взаимодействию с той же органической мишенью.
В предпочтительных слитых белках по изобретению x, y и z представляют собой целые числа от 0 до 2, предпочтительно от 0 до 1. В определенных предпочтительных слитых белках по изобретению y=0. В более предпочтительных конкретных слитых белках по изобретению y=0 и либо x, либо z, равны 0, т.е. слитые белки определяются формулами BX-REP-CT, BX-CT-REP, REP-CT-BZ и CT-REP-BZ, где x и z представляют собой целые числа от 1 до 5. В предпочтительных слитых белках по изобретению y=0, x и z представляют собой целые числа от 0 до 1; и x+z=1. Таким образом, определенные предпочтительные слитые белки по изобретению определяются формулами B-REP-CT, B-CT-REP, REP-CT-B и CT-REP-B. В предпочтительных слитых белках по изобретению необязательная часть NT отсутствует.
Термин "слитый белок" в рамках настоящего изобретения означает что его получают путем экспрессии рекомбинантной нуклеиновой кислоты, т.е. ДНК или РНК, которая создана искусственно путем комбинирования двух или более последовательностей нуклеиновых кислот, которые в норме не встречаются вместе (генетическая инженерия). Слитые белки по изобретению представляют собой рекомбинантные белки, и, таким образом, они не идентичны встречающимся в природе белкам. В частности, спидроины дикого типа не являются слитыми белками по изобретению, поскольку они не экспрессируются рекомбинантной нуклеиновой кислотой, как указано выше. Комбинированные последовательности нуклеиновых кислот кодируют различные белки, неполные белки или полипептиды с определенными функциональными свойствами. Полученный слитый белок или рекомбинантный слитый белок представляет собой единый белок с функциональными свойствами, происходящими из каждого из исходных белков, неполных белков или полипептидов. Более того, слитый белок по изобретению и соответствующие гены являются химерными, т.е. белковые/генные части происходят из по меньшей мере двух различных видов. Части REP и CT, а также необязательная часть NT происходят из белка шелка пауков. Для устранения сомнений, часть B согласно изобретению представляет собой неспидроиновый белок или полипептид, т.е. она не происходит из белка шелка пауков. В частности, часть B по изобретению не происходит из C-концевого, повторяющегося или N-концевого фрагментов белка шелка пауков.
Слитый белок, как правило, состоит из от 170 до 2000 аминокислотных остатков, как например, от 170 до 1000 аминокислотных остатков, как например, от 170 до 600 аминокислотных остатков, предпочтительно от 170 до 500 аминокислотных остатков, как например, от 170 до 400 аминокислотных остатков. Небольшой размер является преимущественным, поскольку более длинные белки, содержащие фрагменты белка шелка пауков, могут образовывать аморфные агрегаты, для растворения и полимеризации которых требуется применение жестких растворителей. Рекомбинантный слитый белок может содержать более 2000 остатков, в частности, в случаях, где белок шелка пауков содержит более одной части B и/или когда он содержит часть NT.
Термины "спидроины" и "белки шелка пауков" используют взаимозаменяемо на протяжении описания, и они охватывают все известные белки шелка пауков, включая белки шелка пауков большой ампуловидной железы, которые, как правило, сокращенно обозначают "MaSp", или "ADF" в случае Araneus diadematus. Эти белки шелка пауков большой ампуловидной железы, главным образом, бывают двух типов: 1 и 2. Эти термины, более того, включают неприродные белки с высокой степенью идентичности и/или сходства с известными белками шелка пауков.
Следовательно, термин "неспидроин" подразумевает белки, которые не происходят из белка шелка пауков, т.е. с низкой степенью (или отсутствием) идентичности и/или сходства с белками шелка пауков.
Белковая структура по изобретению способна к селективному взаимодействию с органической мишенью. Эта способность является присущей слитому белку по изобретению, и, более конкретно, части B слитого белка. Никакие взаимодействия частей REP и CT, а также необязательной части NT, с органическими молекулами не охватываются термином "способен к селективному взаимодействию с органической мишенью". Для устранения сомнений, термин "способен к селективному взаимодействию с органической мишенью" не охватывает димеризацию, олигомеризацию или полимеризацию слитых белков по изобретению, которые основаны на взаимодействиях, вовлекающих части REP и CT, а также необязательную часть NT.
Термин "органическая мишень" охватывает все химические молекулы, содержащие углерод, за исключением того, что специалист в данной области традиционно считает неорганическими молекулами, например, карбонатов, простых оксидов углерода, цианидов, алмаза и графита. Для устранения сомнений, неорганические молекулы, соли и ионы, такие как диоксид кремния и хлорид кальция, не являются органическими. Органическая мишень может представлять собой комплекс, содержащий или состоящий из органических молекул, например, рецепторный комплекс на клеточной поверхности. Органическая мишень может представлять собой мономер, димер, олигомер или полимер одного или нескольких типов органических молекул, которые могут удерживаться вместе ковалентными связями или другими типами связей. Также она, безусловно, может представлять собой просто единичную органическую молекулу. Предпочтительные органические мишени по изобретению включают, но не ограничиваются ими, нуклеиновые кислоты, белки и полипептиды, липиды и углеводы, а также их комбинации. Кроме того, предпочтительные органические мишени по изобретению включают, но не ограничиваются ими, иммуноглобулины, молекулы, содержащие иммуноглобулин или его производные, альбумин, молекулы, содержащие альбумин или его производные, биотин и молекулы, содержащие биотин или его производные или аналоги.
В контексте настоящего изобретения, "специфическое" или "селективное" взаимодействие лиганда, например, части B слитого белка по изобретению, с его мишенью означает, что взаимодействие является таким, что отличие между специфическим и неспецифическим, или между селективным и неселективным, взаимодействием становится значимым. Взаимодействие между двумя белками иногда измеряют с помощью константы диссоциации. Константа диссоциации описывает прочность связывания (или аффинность) между двумя молекулами. Как правило, константа диссоциации между антителом и его антигеном составляет от 10-7 до 10-11 M. Однако высокая специфичность не обязательно требует высокой аффинности. Было показано, что молекулы с низкой аффинностью (в молярном диапазоне) к их партнеру, являются настолько же специфичными, как и молекулы со значительно более высокой аффинностью. В случае настоящего изобретения, специфическое или селективное взаимодействие относится к степени, с которой конкретный способ можно использовать для определения присутствия и/или количестве конкретного белка, белка-мишени или его фрагмента, в данных условиях в присутствии других белков в образце природной или переработанной биологической или биохимической жидкости. Иными словами, специфичность или селективность представляет собой способность различать родственные белки. В настоящем описании термины “специфичный” и “селективный” иногда используют взаимозаменяемо.
Слитый белок по изобретению также может содержать один или несколько линкерных пептидов. Линкерный пептид(ы) может быть расположен между любыми частями слитого белка, например, между частями CT и REP, между двумя частями B, между частями B и CT, и между частями B и REP, или он может быть расположен на любом конце слитого белка. Если слитый белок содержит две или более частей B, линкерный пептид(ы) также может быть расположен между двумя частями B. Линкер(ы) может обеспечивать спейсер между функциональными единицами слитого белка, но также может помочь в идентификации и очистке слитого белка, как например, His-метка и/или Trx-метка. Если слитый белок содержит два или более линкерных пептида для идентификации и очистки слитого белка, предпочтительно, чтобы они были разделены спейсерной последовательностью, например His6-спейсер-His6-. Линкер также может представлять собой сигнальный пептид, такой как частица, распознающая сигнал, которая направляет слитый белок к мембране и/или обеспечивает секрецию слитого белка из клетки-хозяина в окружающую среду. Слитый белок также может включать участок расщепления в его аминокислотной последовательности, который обеспечивает расщепление и удаление линкер(ов) и/или других соответствующих частей, как правило, части или частей B. Различные участки расщепления известны специалисту в данной области, например, участки расщепления для химических агентов, таких как CNBr после остатков Met, и гидроксиламин между остатками Asn-Gly, участки расщепления для протеаз, таких как тромбин или протеаза 3C, и последовательности самосплайсинга, такие как последовательности самосплайсинга интеинов.
Части REP, CT и B связаны друг с другом прямо или непрямо. Прямая связь подразумевает прямое ковалентное связывание между частями без встроенных между ними последовательностей, таких как линкеры. Непрямое связывание также подразумевает, что части связаны ковалентными связями, но что существуют встроенные последовательности, такие как линкеры и/или одна или несколько дополнительных частей, например часть NT.
Часть или части B могут быть расположены внутри или на любом конце слитого белка, т.е. расположены на C-конце или на N-конце. Предпочтительно, чтобы часть или части B были расположены на N-конце слитого белка. Если слитый белок содержит один или несколько линкерный пептид(ов) для идентификации и очистки слитого белка, например, His-метку(и) или Trx-метку(и), предпочтительно, чтобы он был расположен на N-конце слитого белка.
Предпочтительный слитый белок имеет форму расположенной на N-конце части B, присоединенный линкерным пептидом из 1-30 аминокислотных остатков, как например, 1-10 аминокислотных остатков, к расположенным на C-конце частям REP и CT. Линкерный пептид может содержать участок расщепления. Необязательно, слитый белок имеет N-концевой или C-концевой линкерный пептид, который может содержать метку для очистки, такую как His-метка, и участок расщепления.
Другой предпочтительный слитый белок имеет форму расположенной на N-конце части B, связанной непосредственно с расположенными на C-конце частями REP и CT. Необязательно, слитый белок имеет N-концевой или C-концевой линкерный пептид, который может содержать метку для очистки, такую как His-метка, и участок расщепления.
Белковая структура по изобретению представляет собой полимер, содержащий в качестве повторяющейся структурной единицы рекомбинантные слитые белки по изобретению, что подразумевает, что она содержит упорядоченное множество слитых белков по изобретению, как правило, значительно больше 100 элементов слитого белка, например 1000 элементов слитого белка или более. Необязательно, полимер может содержать в качестве дополнительной повторяющейся структурной единицы дополнительные белки без части B, предпочтительно белки, происходящие из шелка пауков. Это может быть преимущественным, если часть B слитого белка является крупной и/или объемной. Эти дополнительные белки, как правило, содержат часть REP и часть CT, и необязательно часть NT. Предпочтительные дополнительные белки по изобретению могут иметь любые структуры, указанные в настоящем описании, с удаленной частью B. Предпочтительно, чтобы дополнительный слитый белок был по существу идентичным слитому белку с удаленной частью B. Однако предпочтительно, чтобы белковая структура по изобретению представляла собой полимер, состоящий из рекомбинантных слитых белков по изобретению в качестве повторяющегося структурного элемента, т.е. чтобы белковая структура по изобретению представляла собой полимер из рекомбинантного слитого белка по изобретению.
Количество слитых элементов в полимере предполагает, что белковая структура достигает значительного размера. В предпочтительном варианте осуществления белковая структура имеет размер по меньшей мере 0,1 мкм по меньшей мере по двум измерениям. Таким образом, термин "белковая структура", как используют в рамках изобретения, относится к полимерам слитого белка, имеющим толщину по меньшей мере 0,1 мкм, предпочтительно, к макроскопическим полимерам, видимым для человеческого глаза, т.е. имеющим толщину по меньшей мере 1 мкм. Термин "белковая структура" не охватывает неструктурные агрегаты или преципитаты. Хотя мономеры слитого белка растворимы в воде, понятно, что белковые структуры по изобретению представляют собой твердые структуры, т.е. не растворимы в воде. Белковые структуры представляют собой полимеры, содержащие в качестве повторяющихся структурных элементов мономеры рекомбинантных слитых белков по изобретению.
Предпочтительно, чтобы белковая структура по изобретению имела физическую форму, выбранную из группы, состоящей из волокна, пленки, пены, сети, решетки, сферы и капсулы.
Предпочтительно, чтобы белковая структура по изобретению представляла собой волокно или пленку с толщиной по меньшей мере 0,1 мкм, предпочтительно по меньшей мере 1 мкм. Предпочтительно, чтобы волокно или пленка имели толщину в диапазоне 1-400 мкм, предпочтительно 60-120 мкм. Предпочтительно, чтобы волокна имели длину в диапазоне 0,5-300 см, предпочтительно 1-100 см. Другие предпочтительные диапазоны представляют собой 0,5-30 см и 1-20 см. Волокно обладает способностью оставаться неизмененным в ходе физического манипулирования, т.е. его можно использовать для прядения, тканья, скручивания, вязки и сходных процессов. Пленка является преимущественной в том, что она является липкой и прилипает к твердым структурам, например, к пластмассе микропланшетов для титрования. Это свойство пленки облегчает процессы промывания и регенерации и в высокой степени пригодно для целей разделения. Особенно пригодной белковой структурой является пленка или волокно, где часть B представляет собой Z-домен, происходящий из белка A стафилококков, или белкового фрагмента, имеющего по меньшей мере 70% идентичность с ним, см. например примеры 1-6.
Также предпочтительно, чтобы белковая структура по изобретению имела прочность на растяжение выше 1 МПа, предпочтительно выше 2 МПа, более предпочтительно 10 Мпа или выше. Предпочтительно, чтобы белковая структура по изобретению имела прочность на растяжение выше 100 МПа, более предпочтительно 200 Мпа или выше.
Часть REP представляет собой белковый фрагмент, содержащий от 70 до 300 аминокислотных остатков, и происходит из повторяющегося фрагмента белка шелка пауков. Это подразумевает, что часть REP имеет повторяющийся характер с чередованием между богатыми аланином участками и богатыми глицином участками. Часть REP, как правило, содержит более 70, как например, более 140, и менее 300, предпочтительно менее 240, как например, менее 200, аминокислотных остатков, и сама по себе может быть подразделена на несколько L (линкерных) сегментов, A (богатых аланином) сегментов и G (богатых глицином) сегментов, как более подробно объяснено ниже. Как правило, указанные линкерные сегменты, которые являются необязательными, расположены на концах части REP, в то время как остальные сегменты в свою очередь являются богатыми аланином и богатыми глицином. Таким образом, часть REP, главным образом, может иметь следующие структуры, где n представляет собой целое число:
L(AG)nL, как например, LA1G1A2G2A3G3A4G4A5G5L;
L(AG)nAL, как например, LA1G1A2G2A3G3A4G4A5G5A6L;
L(GA)nL, как например, LG1A1G2A2G3A3G4A4G5A5L; или
L(GA)nGL, как например, LG1A1G2A2G3A3G4A4G5A5G6L.
Следовательно, некритично то, что богатый аланином или богатый глицином сегмент находится рядом с N-концевым или C-концевым линкерными сегментами. Предпочтительно, чтобы n представляло собой целое число от 2 до 10, предпочтительно от 2 до 8, предпочтительно от 4 до 8, более предпочтительно, от 4 до 6, т.е. n=4, n=5 или n=6.
В предпочтительных вариантах осуществления содержание аланина в части REP согласно изобретению составляет выше 20%, предпочтительно выше 25%, более предпочтительно выше 30% и ниже 50%, предпочтительно ниже 40%, более предпочтительно ниже 35%. Это является преимущественным, поскольку предусматривается, что более высокое содержание аланина обеспечивает более жесткую и/или более прочную и/или менее растяжимую структуру.
В определенных вариантах осуществления часть REP лишена остатков пролина, т.е. в части REP отсутствуют остатки пролина.
Далее, обращаясь к сегментам, которые составляют часть REP по изобретению, следует подчеркнуть, что каждый сегмент является индивидуальным, т.е. любые два сегмента A, любые два сегмента G или любые два сегмента L конкретной части REP могут быть идентичными или могут не быть идентичными. Таким образом, не является главным признаком изобретения то, что каждый тип сегмента идентичен в конкретной части REP. Вместо этого, в описании далее представлено квалифицированному специалисту руководство, как сконструировать индивидуальные сегменты и собрать их в часть REP, которую, тем самым, считают происходящей из повторяющегося фрагмента белка шелка пауков, и которая составляет часть функционального слитого белка по изобретению.
Каждый индивидуальный сегмент A представляет собой аминокислотную последовательность, имеющую от 8 до 18 аминокислотных остатков. Предпочтительно, чтобы каждый индивидуальный сегмент A содержал от 13 до 15 аминокислотных остатков. Также возможно, чтобы большинство или более двух сегментов A содержало от 13 до 15 аминокислотных остатков, и чтобы меньшинство, такое как один или два сегмента A, содержало от 8 до 18 аминокислотных остатков, как например, 8-12 или 16-18 аминокислотных остатков. Широкое большинство этих аминокислотных остатков представляют собой остатки аланина. Более конкретно, от 0 до 3 аминокислотных остатков не являются остатками аланина, а остальные аминокислотные остатки являются остатками аланина. Таким образом, все аминокислотные остатки в каждом индивидуальном сегменте A представляют собой остатки аланина без исключения, или за исключением одного, двух или трех аминокислотных остатков, которые могут представлять собой любую аминокислоту. Предпочтительно, чтобы замещающая аланин аминокислота(ы) представляла собой природную аминокислоту, предпочтительно индивидуально выбранную из группы из серина, глутаминовой кислоты, цистеина и глицина, более предпочтительно серина. Безусловно, возможно, чтобы один или несколько из сегментов A представляли собой полностью аланиновые сегменты, в то время как оставшиеся сегменты A содержали 1-3 неаланиновых остатка, таких как серин, глутаминовая кислота, цистеин или глицин.
В предпочтительном варианте осуществления каждый сегмент A содержит 13-15 аминокислотных остатков, включая 10-15 остатков аланина и 0-3 неаланиновых остатка, как описано выше. В более предпочтительном варианте осуществления каждый сегмент A содержит 13-15 аминокислотных остатка, включая 12-15 остатков аланина и 0-1 неаланиновый остаток как описано выше.
Предпочтительно, чтобы каждый индивидуальный сегмент A имел по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичность с аминокислотной последовательностью, выбранной из группы из аминокислотных остатков 7-19, 43-56, 71-83, 107-120, 135-147, 171-183, 198-211, 235-248, 266-279, 294-306, 330-342, 357-370, 394-406, 421-434, 458-470, 489-502, 517-529, 553-566, 581-594, 618-630, 648-661, 676-688, 712-725, 740-752, 776-789, 804-816, 840-853, 868-880, 904-917, 932-945, 969-981, 999-1013, 1028-1042 и 1060-1073 SEQ ID NO: 10. Каждая последовательность в этой группе соответствует сегменту встречающейся в природе последовательности белка MaSp1 Euprosthenops australis, которая установлена путем клонирования соответствующей кДНК, см. WO 2007/078239. Альтернативно каждый индивидуальный сегмент A имеет по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичность с аминокислотной последовательностью, выбранной из группы из аминокислотных остатков 143-152, 174-186, 204-218, 233-247 и 265-278 SEQ ID NO: 3. Каждая последовательность этой группы соответствует сегменту экспрессированных неприродных белков шелка пауков, которые способны образовывать шелковые структуры в соответствующих условиях. Таким образом, в определенных вариантах осуществления изобретения каждый индивидуальный сегмент A идентичен аминокислотной последовательности, выбранной из упомянутых выше аминокислотных сегментов. Без связи с какой-либо конкретной теорией, предусматривается, что сегменты A по изобретению образуют спиральные структуры или бета-слои.
Термин "% идентичность", как используют на протяжении описания и в прилагаемой формуле изобретения, вычисляют следующим образом. Последовательность запроса выравнивают с последовательностью-мишенью с использованием алгоритма CLUSTAL W (Thompson, J.D., Higgins, D.G. и Gibson, T.J., Nucleic Acids Research, 22: 4673-4680 (1994)). Сравнение проводят на протяжении окна, соответствующего наиболее короткой из выравниваемых последовательностей. Аминокислотные остатки в каждом положении сравнивают и процент положений в последовательности запроса, которые имеют идентичные соответствия в последовательности-мишени, сообщают как % идентичность.
Термин "% сходство", как используют на протяжении описания и в прилагаемой формуле изобретения, вычисляют, как описано для "% идентичности", за исключением того, что гидрофобные остатки Ala, Val, Phe, Pro, Leu, Ile, Trp, Met и Cys являются сходными; основные остатки Lys, Arg и His являются сходными; кислотные остатки Glu и Asp являются сходными; и гидрофильные незаряженные остатки Gln, Asn, Ser, Thr и Tyr являются сходными. Оставшаяся природная аминокислота Gly не является сходной ни с одной другой аминокислотой в этом контексте.
На протяжении настоящего описания альтернативные варианты осуществления изобретения соответствуют, вместо указанного процента идентичности, соответствующему проценту сходства. Другие альтернативные варианты осуществления соответствуют указанному проценту идентичности, а также другому, более высокому, проценту сходства, выбранному из группы предпочтительных процентов идентичности для каждой последовательности. Например, последовательность может быть на 70% сходной с другой последовательностью; или она может быть на 70% идентична другой последовательности; или она может быть на 70% идентична и 90% сходна с другой последовательностью.
Более того, из экспериментальных данных было сделано заключение, что каждый индивидуальный сегмент G представляет собой аминокислотную последовательность от 12 до 30 аминокислотных остатков. Предпочтительно, чтобы каждый индивидуальный сегмент G состоял из от 14 до 23 аминокислотных остатков. По меньшей мере 40% аминокислотных остатков каждого сегмента G представляют собой остатки глицина. Как правило, содержание глицина в каждом индивидуальном сегменте G находится в диапазоне 40-60%.
Предпочтительно, чтобы каждый индивидуальный сегмент G имел по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичность с аминокислотной последовательностью, выбранной из группы их аминокислотных остатков 20-42, 57-70, 84-106, 121-134, 148-170, 184-197, 212-234, 249-265, 280-293, 307-329, 343-356, 371-393, 407-420, 435-457, 471-488, 503-516, 530-552, 567-580, 595-617, 631-647, 662-675, 689-711, 726-739, 753-775, 790-803, 817-839, 854-867, 881-903, 918-931, 946-968, 982-998, 1014-1027, 1043-1059 и 1074-1092 SEQ ID NO: 10. Каждая последовательность этой группы соответствует сегменту встречающейся в природе последовательности белка MaSp1 Euprosthenops australis, которая установлена при клонировании соответствующей кДНК, см. WO 2007/078239. Альтернативно каждый индивидуальный сегмент G имеет по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно 95%, наиболее предпочтительно 100% идентичность с аминокислотной последовательностью, выбранной из группы из аминокислотных остатков 153-173, 187-203, 219-232, 248-264 и 279-296 SEQ ID NO: 3. Каждая последовательность этой группы соответствует сегменту экспрессированных неприродных белков шелка пауков, которые способны образовывать шелковые структуры в соответствующих условиях. Таким образом, в определенных вариантах осуществления изобретения каждый индивидуальный сегмент G идентичен аминокислотной последовательности, выбранной из упомянутых выше аминокислотных сегментов.
В определенных вариантах осуществления первые два аминокислотных остатка каждого сегмента G по изобретению не являются -Gln-Gln-.
Имеется три подтипа сегмента G по изобретению. Эта классификация основана на тщательном анализе белковой последовательности MaSp1 Euprosthenops australis (WO 2007/078239), и эту информацию использовали и подтвердили при конструировании новых неприродных белков шелка пауков.
Первому подтипу сегмента G по изобретению соответствует консенсусная последовательность, обозначаемая однобуквенным обозначением аминокислот GQG(G/S)QGG(Q/Y)GG(L/Q)GQGGYGQGA GSS (SEQ ID NO: 11). Этот первый и, как правило, наиболее длинный, подтип сегмента G, главным образом, содержит 23 аминокислотных остатка, но может содержать всего 17 аминокислотных остатков и лишен заряженных остатков или содержит один заряженный остаток. Таким образом, предпочтительно, чтобы этот первый подтип сегмента G содержал 17-23 аминокислотных остатка, однако предусматривается, что он может содержать только 12 или вплоть до 30 аминокислотных остатков. Без связи с какой-либо конкретной теорией, предусматривается, что этот подтип формирует структуры витков или 31-спиральные структуры. Типичные сегменты G этого первого подтипа представляют собой аминокислотные остатки 20-42, 84-106, 148-170, 212-234, 307-329, 371-393, 435-457, 530-552, 595-617, 689-711, 753-775, 817-839, 881-903, 946-968, 1043-1059 и 1074-1092 SEQ ID NO: 10. В определенных вариантах осуществления первые два аминокислотных остатка каждого сегмента G этого первого подтипа согласно изобретению не являются -Gln-Gln-.
Второму подтипу сегмента G по изобретению соответствует консенсусная последовательность, обозначаемая однобуквенным обозначением аминокислот GQGGQGQG(G/R)YGQG(A/S)G(S/G)S (SEQ ID NO: 12). Этот второй, как правило, среднего размера, подтип сегмента G, главным образом, содержит 17 аминокислотных остатков и лишен заряженных остатков или содержит один заряженный остаток. Предпочтительно, чтобы этот второй сегмент G содержал 14-20 аминокислотных остатков, однако предусматривается, что он может содержать только 12 или вплоть до 30 аминокислотных остатков. Без связи с какой-либо конкретной теорией, предусматривается, что этот подтип формирует структуры витков. Типичными сегментами этого второго подтипа G являются аминокислотные остатки 249-265, 471-488, 631-647 и 982-998 SEQ ID NO: 10; и аминокислотные остатки 187-203 SEQ ID NO: 3.
Третьему подтипу сегмента G по изобретению соответствует консенсусная последовательность, обозначаемая однобуквенным обозначением аминокислот G(R/Q)GQG(G/R)YGQG(A/S/V)GGN (SEQ ID NO: 13). Этот третий подтип сегмента G, как правило, содержит 14 аминокислотных остатков, и, главным образом, является наиболее коротким из подтипов сегмента G по изобретению. Предпочтительно, чтобы этот третий подтип сегмента G содержал 12-17 аминокислотных остатков, однако предусматривается, что он может содержать вплоть до 23 аминокислотных остатков. Без связи с какой-либо конкретной теорией, предусматривается, что этот подтип формирует структуры поворотов. Типичными сегментами G этого третьего подтипа являются аминокислотные остатки 57-70, 121-134, 184-197, 280-293, 343-356, 407-420, 503-516, 567-580, 662-675, 726-739, 790-803, 854-867, 918-931, 1014-1027 SEQ ID NO: 10; и аминокислотные остатки 219-232 SEQ ID NO: 3.
Таким образом, в предпочтительных вариантах осуществления каждый индивидуальный сегмент G имеет по меньшей мере 80%, предпочтительно 90%, более предпочтительно 95%, идентичность с аминокислотной последовательностью, выбранной из SEQ ID NO: 11, SEQ ID NO: 12 и SEQ ID NO: 13.
В предпочтительном варианте осуществления чередующейся последовательности сегментов A и G в части REP, каждый второй сегмент G представляет собой сегмент первого подтипа, в то время как остальные сегменты G представляют собой сегменты третьего подтипа, например, …A1GкороткийA2GдлинныйA3GкороткийA4GдлинныйA5Gкороткий… В другом предпочтительном варианте осуществления части REP, один сегмент G второго подтипа прерывает регулярность сегментов G путем встраивания, например, …A1GкороткийA2GдлинныйA3GсреднийA4GкороткийA5Gдлинный…
Каждый индивидуальный сегмент L представляет собой необязательную линкерную аминокислотную последовательность, которая может содержать от 0 до 20 аминокислотных остатков, как например, от 0 до 10 аминокислотных остатков. Хотя этот сегмент является необязательным и функционально некритичным для белка шелка пауков, его присутствие, тем не менее, допускает полностью функциональные слитые белки шелка пауков, образующие белковые структуры по изобретению. Также имеются линкерные аминокислотные последовательности, присутствующие в повторяющейся части (SEQ ID NO: 10) установленной аминокислотной последовательности белка MaSp1 из Euprosthenops australis. В частности, аминокислотная последовательность линкерного сегмента может напоминать любой из описанных сегментов A или G, однако обычно является недостаточной, чтобы удовлетворять их критериям, как определено в настоящем описании.
Как показано в WO 2007/078239, линкерному сегменту, расположенному в C-концевой области части REP, могут соответствовать консенсусные последовательности, обозначаемые однобуквенным обозначением аминокислот ASASAAASAA STVANSVS и ASAASAAA, которые являются богатыми аланином. В действительности, вторая последовательность может считаться сегментом A согласно изобретению, в то время как первая последовательность имеет высокую степень сходства с сегментами A по изобретению. Другой пример линкерного сегмента по изобретению имеет аминокислотную последовательность, обозначаемую однобуквенным обозначением GSAMGQGS, которая богата глицином и имеет высокую степень сходства с сегментами G по изобретению. Другим примером линкерного сегмента является SASAG.
Типичные сегменты L представляют собой аминокислотные остатки 1-6 и 1093-1110 SEQ ID NO: 10; и аминокислотные остатки 138-142 SEQ ID NO: 3, однако специалисту в данной области хорошо понятно, что существует множество подходящих альтернативных аминокислотных последовательностей для этих сегментов. В одном варианте осуществления части REP согласно изобретению, один из сегментов L содержит 0 аминокислот, т.е. один из сегментов L отсутствует. В другом варианте осуществления части REP по изобретению оба сегмента L содержат 0 аминокислот, т.е. оба сегмента L отсутствуют. Таким образом, эти варианты осуществления частей REP согласно изобретению могут быть схематично представлены следующим образом: (AG)nL, (AG)nAL, (GA)nL, (GA)nGL; L(AG)n, L(AG)nA, L(GA)n, L(GA)nG; и (AG)n, (AG)nA, (GA)n, (GA)nG. Любые из этих частей REP пригодны для применения с любой частью CT, как определено ниже.
Часть CT представляет собой фрагмент белка, содержащий от 70 до 120 аминокислотных остатков, и происходит из C-концевого фрагмента белка шелка пауков. Выражение "происходящий из" подразумевает, в контексте части CT по изобретению, что она имеет высокую степень сходства с C-концевой аминокислотной последовательностью белков шелка пауков. Как показано на фиг. 1, эта аминокислотная последовательность является высоко консервативной среди различных видов и белков шелка пауков, включая MaSp1 и MaSp2. Консенсусная последовательность C-концевых областей MaSpl и MaSp2 представлена в качестве SEQ ID NO: 9. На фиг. 1 выровнены следующие белки MaSp, обозначенные с помощью номеров доступа GenBank, где это применимо:

Не является критическим, какая конкретная часть CT присутствует в белках шелка пауков по изобретению, при условии что часть CT не отсутствует полностью. Таким образом, часть CT по изобретению может быть выбрана из любых аминокислотных последовательностей, представленных на фиг. 1 и в таблице 1, или последовательностей с высокой степенью сходства. В белке шелка пауков по изобретению можно использовать широкое множество C-концевых последовательностей.
Последовательность части CT по изобретению имеет по меньшей мере 50% идентичность, предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65% идентичность, или даже по меньшей мере 70% идентичность, с консенсусной аминокислотной последовательностью SEQ ID NO: 9, которая основана на аминокислотных последовательностях фиг. 1.
Типичной частью CT согласно изобретению является последовательность Euprosthenops australis SEQ ID NO: 7. Таким образом, согласно предпочтительному аспекту изобретения, часть CT имеет по меньшей мере 80%, предпочтительно по меньшей мере 90%, как например, по меньшей мере 95%, идентичность с SEQ ID NO: 7 или любой индивидуальной аминокислотной последовательностью на фиг. 1 и в таблице 1. В предпочтительных аспектах изобретения часть CT является идентичной SEQ ID NO: 7 или любой индивидуальной аминокислотной последовательности на фиг. 1 и в таблице 1.
Часть CT, как правило, состоит из 70-120 аминокислотных остатков. Предпочтительно, чтобы часть CT содержала по меньшей мере 70, или более 80, предпочтительно более 90, аминокислотных остатков. Также предпочтительно, чтобы часть CT содержала не более 120, или менее 110 аминокислотных остатков. Типичная часть CT содержит приблизительно 100 аминокислотных остатков.
Необязательная часть NT представляет собой белковый фрагмент, содержащий от 100 до 160 аминокислотных остатков, и происходит из N-концевого фрагмента белка шелка пауков. Выражение "происходящий из" подразумевает в контексте части NT согласно изобретению, что она имеет высокую степень сходства с N-концевой аминокислотной последовательностью белков шелка пауков. Как показано на фиг. 2, эта аминокислотная последовательность является высоко консервативной среди различных видов и белков шелка пауков, включая MaSp1 и MaSp2. На фиг. 2 выровнены следующие части NT спидроинов, обозначенные с помощью номеров доступа GenBank, где это применимо:

Для каждой последовательности представлена только часть, соответствующая N-концевой части, без сигнального пептида. Nc-flag и Nlm-flag транслируются и редактируются согласно Rising A. et al. Biomacromolecules 7, 3120-3124 (2006)).
Не является критичным, какая конкретная часть NT присутствует в белках шелка пауков согласно изобретению. Таким образом, часть NT согласно изобретению можно выбирать из любых аминокислотных последовательностей, представленных на фиг. 2, или последовательностей с высокой степенью сходства. В белке шелка пауков по изобретению можно использовать широкое множество N-концевых последовательностей. Исходя из гомологичных последовательностей на фиг. 2, следующая последовательность представляет собой консенсусную аминокислотную последовательность NT:
QANTPWSSPNLADAFINSF(M/L)SA(A/I)SSSGAFSADQLDDMSTIG(D/N/Q)TLMSAMD(N/S/K)MGRSG(K/R)STKSKLQALNMAFASSMAEIAAAESGG(G/Q)SVGVKTNAISDALSSAFYQTTGSVNPQFV(N/S)EIRSLI(G/N)M(F/L)(A/S)QASANEV (SEQ ID NO: 8).
Последовательность части NT согласно изобретению имеет по меньшей мере 50% идентичность, предпочтительно по меньшей мере 60% идентичность, с консенсусной аминокислотной последовательностью SEQ ID NO: 8, которая основана на аминокислотных последовательностях, представленных на фиг. 2. В предпочтительном варианте осуществления последовательность части NT согласно изобретению имеет по меньшей мере 65% идентичность, предпочтительно по меньшей мере 70% идентичность, с консенсусной аминокислотной последовательностью SEQ ID NO: 8. Более того, в предпочтительных вариантах осуществления часть NT согласно изобретению имеет 70%, предпочтительно 80%, сходство с консенсусной аминокислотной последовательностью SEQ ID NO: 8.
Типичная часть NT согласно изобретению представляет собой последовательность Euprosthenops australis SEQ ID NO: 6. Согласно предпочтительному варианту осуществления изобретения часть NT имеет по меньшей мере 80% идентичность с SEQ ID NO: 6 или любой индивидуальной аминокислотной последовательностью, представленной на фиг. 1. В предпочтительных вариантах осуществления изобретения часть NT имеет по меньшей мере 90%, как например, по меньшей мере 95%, идентичность с SEQ ID NO: 6 или любой индивидуальной аминокислотной последовательностью на фиг. 2. В предпочтительных вариантах осуществления изобретения часть NT идентична SEQ ID NO: 6 или любой индивидуальной аминокислотной последовательности, представленной на фиг. 1, в частности MaSp1 Ea.
Часть NT содержит от 100 до 160 аминокислотных остатков. Предпочтительно, чтобы часть NT содержала по меньшей мере 100 или более 110, предпочтительно более 120, аминокислотных остатков. Также предпочтительно, чтобы часть NT содержала не более 160 или менее 140 аминокислотных остатков. Типичная A часть NT содержит приблизительно 130-140 аминокислотных остатков.
Часть B представляет собой белковый или полипептидный фрагмент, содержащий более 30 аминокислотных остатков. Часть B предпочтительно содержит более 40 аминокислотных остатков, как например, более 50 аминокислотных остатков. Часть B предпочтительно содержит менее 500 аминокислотных остатков, как например, менее 200 аминокислотных остатков, более предпочтительно, менее 100 аминокислотных остатков, как например, менее 100 аминокислотных остатков. Она способна к селективному взаимодействию с органической мишенью и представляет собой часть B в слитом белке, которая обеспечивает способность селективного взаимодействия с органической мишенью.
Часть B представляет собой неспидроиновую часть. Это подразумевает, что она не происходит из белка шелка пауков, т.е. она имеет низкую степень (или не имеет) идентичности и/или сходства с белками шелка пауков. Последовательность части B согласно изобретению предпочтительно имеет менее чем 30% идентичность, такую как менее чем 20% идентичность, предпочтительно менее чем 10% идентичность, с любой из аминокислотных последовательностей спидроинов, описанных в настоящем описании, и, в частности, с любой из SEQ ID NO: 6-10.
Выбор части B находится в пределах квалификации специалистов в данной области. Тем не менее, примеры аффинных лигандов, которые могут оказаться полезными в качестве частей B, а также примеры форматов и условий для детекции и/или количественного определения, приведены ниже для иллюстрации.
Разнообразие биомолекул, требуемое для выбора аффинных лигандов, можно обеспечивать путем комбинаторной инженерии одной из множества возможных каркасных молекул, а затем специфические и/или селективные аффинные лиганды выбирают с использованием подходящей платформы для селекции. Неограничивающими примерами таких структур, пригодных для получения аффинных лигандов против органической мишени, являются белок A стафилококков и его домены и производные этих доменов, такие как Z-домен (Nord K ef al. (1997) Nat. Biotechnol. 15:772-777); липокалины (Beste G et al. (1999) Proc. Natl. Acad. Sci. U.S.A. 96: 1898-1903); домены анкириновых повторов (Binz HK et al. (2003) J. Mol. Biol. 332:489-503); связывающие целлюлозу домены (CBD) (Smith GP et al. (1998) J. Mol. Biol. 277:317-332; Lehtio J et al. (2000) Proteins 41:316-322); γ-кристаллины (Fiedler U and Rudolph R, WO01/04144); зеленый флуоресцентный белок (GFP) (Peelle B et al. (2001) Chem. Biol. 8:521-534); ассоциированный с цитотоксическим T-лимфоцитами антиген 4 человека (CTLA-4) (Hufton SE et al. (2000) FEBS Lett. 475:225-231; Irving RA et al. (2001) J. Immunol. Meth. 248:31-45); ингибиторы протеаз, такие как белки Knottin (Wentzel A et al. (2001) J. Bacteriol. 183:7273-7284; Baggio R et al. (2002) J. Mol. Recognit. 15:126-134) и домены Kunitz (Roberts BL et al. (1992) Gene 121:9-15; Dennis MS and Lazarus RA (1994) J. Biol. Chem. 269:22137-22144); домены PDZ (Schneider S et al. (1999) Nat. Biotechnol. 17:170-175); пептидные аптамеры, такие как тиоредоксин (Lu Z et al. (1995) Biotechnology 13:366-372; Klevenz B et al. (2002) Cell. Mol. Life Sci. 59:1993-1998); нуклеаза стафилококков (Norman TC et al. (1999) Science 285:591-595); тендамистаты (McConell SJ и Hoess RH (1995) J. Mol. Biol. 250:460-479; Li R et al. (2003) Protein Eng. 16:65-72); тринектины на основе домена фибронектина типа III (Koide A et al. (1998) J. Mol. Biol. 284:1141-1151; Xu L et al. (2002) Chem. Biol. 9:933-942); и цинковые пальцы (Bianchi E et al. (1995) J. Mol. Biol. 247:154-160; Klug A (1999) J. Mol. Biol. 293:215-218; Segal DJ et al. (2003) Biochemistry 42:2137-2148).
Упомянутые выше примеры включают каркасные белки, обеспечивающие единичную рандомизированную петлю, используемые для получения новой специфичности связывания, белковые каркасы с жесткой вторичной структурой, где боковые цепи, выступающие с поверхности белка, рандомизированы для получения новой специфичности связывания, и каркасы, обладающие не последовательно расположенной областью гипервариабельной петли, используемые для получения новой специфичности связывания.
Также в качестве аффинных лигандов можно использовать олигонуклеотиды. Одноцепочечные нуклеиновые кислоты, называемые аптамерами или ловушками, сворачиваются в строго определенные трехмерные структуры и связываются с их мишенью с высокой аффинностью и специфичностью. (Ellington AD and Szostak JW (1990) Nature 346:818-822; Brody EN и Gold L (2000) J. Biotechnol. 74:5-13; Mayer G and Jenne A (2004) BioDrugs 18:351-359). Олигонуклеотидные лиганды могут представлять собой либо РНК, либо ДНК, и могут связываться с широким диапазоном классов молекул-мишеней.
Для выбора лиганда с желаемой аффинностью из совокупности вариантов любой из каркасных структур, упомянутых выше, доступен ряд селекционных платформ для выделения специфического нового лиганда против выбранного белка-мишени. Селекционные платформы включают, но не ограничиваются ими, фаговый дисплей (Smith GP (1985) Science 228: 1315-1317), рибосомный дисплей (Hanes J и Plückthun A (1997) Proc. Natl. Acad. Sci. U.S.A. 94:4937-4942), дрожжевую двухгибридную систему (Fields S and Song O (1989) Nature 340:245-246), дрожжевой дисплей (Gai SA and Wittrup KD (2007) Curr Opin Struct Biol 17:467-473), дисплей мРНК (Roberts RW и Szostak JW (1997) Proc. Natl. Acad. Sci. U.S.A. 94:12297-12302), бактериальный дисплей (Daugherty PS (2007) Curr Opin Struct Biol 17:474-480, Kronqvist N et al. (2008) Protein Eng Des Sel 1-9, Harvey BR et al. (2004) PNAS 101 (25):913-9198), дисплей на микрогранулах (Nord O et al. (2003) J Biotechnol 106: 1-13, WO01/05808), SELEX (System Evolution of Ligands by Exponential Enrichment) (Tuerk C and Gold L (1990) Science 249:505-510) и анализы комплементации белковых фрагментов (PCA) (Remy I and Michnick SW (1999) Proc. Natl. Acad. Sci. U.S.A. 96:5394-5399). Предпочтительной группой частей B с аффинностью к иммуноглобулинам, альбумину или другим органическим мишеням, являются домены рецептина бактерий или их производные.
Группа предпочтительных частей B способна к селективному взаимодействию с иммуноглобулинами и молекулами, содержащими иммуноглобулин или его производные. Предпочтительной группой подклассов иммуноглобулинов являются подклассы, которые распознаются Z-доменом, происходящим из белка A стафилококков, т.е. IgG1, IgG2, IgG4, IgA и IgM человека, все подклассы Ig кролика и коровы, IgG1 и IgG2 морской свинки, и IgG1, IgG2a, IgG2b, IgG3 и IgM мыши (см. Hober, S. et al., J. Chromatogr B. 848:40-47 (2007)), более предпочтительно подклассы иммуноглобулинов IgG1, IgG2, IgG4, IgA и IgM человека. Другой предпочтительной группой подклассов иммуноглобулинов являются подклассы, которые распознаются C2-доменом белка G стрептококков; т.е. все подклассы IgG человека, включая IgG3, и IgG нескольких животных, включая мышь, кролика и овцу.
Одна группа предпочтительных частей B выбрана из группы, состоящей из Z-домена, происходящего из белка A стафилококков, белка A стафилококков и его доменов, предпочтительно доменов E, D, A, B и C, белка G стрептококков и его доменов, предпочтительно доменов C1, C2 и C3; и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность, с любой из этих аминокислотных последовательностей. Предпочтительно, часть B выбрана из группы, состоящей из Z-домена, происходящего из белка A стафилококков, B-домена белка A стафилококков, и C2-домена белка G стрептококков; и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность, с любой из этих аминокислотных последовательностей. Предпочтительно, часть B выбрана из группы, состоящей из Z-домена, происходящего из белка A стафилококков и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность, с этой аминокислотной последовательностью. Предпочтительно, чтобы часть B была выбрана из группы, состоящей из Z-домена, происходящего из белка A стафилококков, и C2-домена белка G стрептококков, см. например, примеры 1-6 и 8. Предпочтительной группой частей B с аффинностью к иммуноглобулинам являются домены рецептина бактерий и их производные.
Другая группа предпочтительных частей B способна к селективному взаимодействию с альбумином и молекулами, содержащими альбумин или его производные. Предпочтительной группой частей B с аффинностью к альбумину являются домены рецептина бактерий или их производные. Предпочтительные части B выбирают из белка G стрептококков, альбумин-связывающего домена белка G стрептококков, GA-модулей из Finegoldia magna; и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность или по меньшей мере 90% идентичность, с любой из этих аминокислотных последовательностей. Предпочтительно, часть B выбирают из альбумин-связывающего домена белка G стрептококков и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность с ними. Предпочтительно, чтобы часть B представляла собой альбумин-связывающий домен белка G стрептококков, см. например, пример 7.
Следующая группа предпочтительных частей B способна к селективному взаимодействию с биотином и молекулами, содержащими биотин или его производные или аналоги. Предпочтительные части B выбраны из группы, состоящей из стрептавидина, мономерного стрептавидина (M4); и белковых фрагментов, имеющих по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность с любой из этих аминокислотных последовательностей. Предпочтительно, чтобы часть B представляла собой мономерный стрептавидин (M4), см. например, примеры 10-12.
Конкретные слитые белки и белковые структуры по изобретению представлены в примерах. Эти предпочтительные слитые белки образуют группу, состоящую из SEQ ID NO: 14, 16, 18, 22, 24 и 26. Следующие предпочтительные слитые белки имеют по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, идентичность с любой из этих последовательностей.
Кроме того, настоящее изобретение относится к выделенным полинуклеиновым кислотам, кодирующим слитый белок по изобретению. В частности, конкретные полинуклеиновые кислоты представлены в примерах и прилагаемом списке последовательностей, например SEQ ID NO: 15, 17, 19, 23, 25 и 27. Следующие предпочтительные полинуклеиновые кислоты кодируют слитые белки, имеющие по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, идентичность с любой из SEQ ID NO: 14, 16, 18, 22, 24 и 26.
Полинуклеиновые кислоты по изобретению пригодны для получения слитых белков по изобретению. Настоящее изобретение относится к способу получения слитого белка. Первая стадия вовлекает экспрессию в подходящем хозяине слитого белка по изобретению. Подходящие хозяева хорошо известны специалисту в данной области и включают, например, бактерии и эукариотические клетки, такие как дрожжи, клеточные линии насекомых и клеточные линии млекопитающих. Как правило, эта стадия вовлекает экспрессию молекулы полинуклеиновой кислоты, которая кодирует слитый белок в E. coli.
Вторая стадия способа вовлекает получение смеси, содержащей слитый белок. Смесь можно получать, например, лизисом или механическим разрушением клеток-хозяев. Также смесь можно получать путем сбора клеточной культуральной среды, если слитый белок секретируется клеткой-хозяином. Полученный таким образом белок можно выделять с использованием стандартных методик. Если желательно, эту смесь можно подвергать центрифугированию, и можно собирать соответствующую фракцию (преципитат или супернатант). Смесь, содержащую слитый белок, также можно подвергать гель-фильтрации, хроматографии, например, анионообменной хроматографии, диализу, разделению фаз или фильтрации для обеспечения разделения. Необязательно, на этой стадии активно удаляются липополисахариды и другие пирогены. Если желательно, на этой стадии можно удалять линкерные пептиды путем расщепления.
Белковые структуры по изобретению собираются самопроизвольно из слитых белков по изобретению в подходящих условиях, и сборка в полимеры стимулируется присутствием сдвиговых усилий и/или поверхности контакта между различными фазами, например, между твердой и жидкой фазой, между воздушной и жидкой фазой, или поверхности контакта гидрофобной/гидрофильной среды, например поверхности контакта минеральное масло-вода. Присутствие полученной поверхности контакта стимулирует полимеризацию на поверхности контакта или в области, окружающей поверхность контакта, где эта область распространяется в жидкую среду, так чтобы указанная полимеризация начиналась на указанной поверхности контакта или в указанной области поверхности контакта. Можно получать различные белковые структуры путем адаптации условий в процессе сборки. Например, если сборке позволяют произойти в контейнере, который осторожно качают из стороны в сторону, на поверхности контакта воздух-вода образуется волокно. Если смеси позволяют стоять, на поверхности контакта воздух-вода образуется пленка. Если смесь упаривают, пленка образуется на дне контейнера. Если сверху водной смеси добавляют масло, на поверхности контакта масло-вода образуется пленка, либо если ей позволяют стоять, либо если ее качают. Если смесь пенится, например, путем образования пузырьков воздуха или взбивания, пена является стабильной и застывает, если ей позволить высохнуть.
Таким образом, настоящее изобретение относится к способу предоставления белковой структуры, проявляющей активность связывания органической мишени. На первой стадии способа предусмотрен рекомбинантный слитый белок по изобретению. Слитый белок, например, может быть предоставлен путем экспрессии его в подходящем хозяине с полинуклеиновой кислотой по изобретению. На второй стадии способа слитый белок подвергают воздействию условий, обеспечивающих образование полимера, содержащего рекомбинантный слитый белок. Примечательно, хотя самопроизвольно собранные белковые структуры можно растворять в гексафторизопропаноле, растворенные слитые белки затем неспособны самопроизвольно обратно собираться, например, в волокна.
Белковая структура пригодна в качестве части аффинной среды для иммобилизации органической мишени, где часть B способна к селективному взаимодействию с органической мишенью. Образец, например, биологический образец, можно наносить на слитый белок или белковую структуру по изобретению, которые способны связываться с органической мишенью, присутствующей в биологическом образце, а затем слитый белок или белковая структура пригодны для выделения органической мишени из образца. Биологический образец, такой как кровь, сыворотка или плазма, который может быть извлечен из индивидуума, можно подвергать детекции, выделению и/или количественному определению органической мишени.
Таким образом, настоящее изобретение относится к способу выделения органической мишени из образца. Предусмотрен образец, например, биологический образец, такой как кровь, сыворотка или плазма, содержащий органическую мишень. Биологический образец может представлять собой ранее полученный образец. Если использовать ранее полученный образец в способе, никакие стадии способа не осуществляют в организме человека или животного.
Предусмотрена аффинная среда по изобретению, содержащая слитый белок или белковую структуру по изобретению. В определенных вариантах осуществления аффинная среда состоит из слитого белка или белковой структуры по изобретению. Аффинная среда способна к селективному взаимодействию с органической мишенью посредством части B в слитом белке по изобретению. Аффинную среду контактируют с образцом в подходящих условиях для достижения связывания между аффинной средой и органической мишенью. Несвязанный образец удаляют в подходящих условиях для поддержания селективного связывания между аффинной средой и органической мишенью. Этот способ приводит к органической мишени, иммобилизованной на аффинной среде и, в частности, слитом белке по изобретению.
В предпочтительном способе в аффинной среде в качестве белковой структуры по изобретению присутствует по изобретению слитый белок, когда аффинную среду контактируют с образцом для достижения связывания между аффинной средой и органической мишенью.
Особенно пригодной белковой структурой в этом отношении является пленка или волокно, где часть B представляет собой Z-домен, происходящий из белка A стафилококков или белкового фрагмента, имеющего по меньшей мере 70% идентичность, как например, по меньшей мере 80% идентичность, или по меньшей мере 90% идентичность с ним, см. например, пример 1-6. Пленка является преимущественной в том, что она прикрепляется к твердым структурам, например, к пластмассе в микропланшетах для титрования. Это свойство пленки облегчает промывание и регенерацию и в высокой степени пригодно для целей выделения.
Неожиданно было выявлено, что устойчивость к щелочам Z-домена может быть даже усилена, когда он является частью слитого белка по изобретению в белковой структуре по изобретению. Это свойство может быть пригодным для промывания и регенерации, например, допуская высокие концентрации NaOH, такие как 0,1 M, 0,5 M, 1 M или даже выше 1 M, например 2 M, и/или высокие концентрации мочевины, например 6-8 M. Химическая стабильность также может быть пригодна для обеспечения повторяющихся циклов применения Z-домена для аффинной очистки. Эту устойчивость к щелочам можно далее увеличивать с использованием стабилизированного мутанта Z-домена. Более того, было преимущественной показано, что слитые белки по изобретению, включая Z-домен, являются термостабильными. Это обеспечивает стерилизацию нагреванием с сохранением аффинности связывания.
Известной проблемой с традиционными аффинными матрицами с Z-доменами является утечка Z-домена из аффинной матрицы. Благодаря стабильному встраиванию Z-домена посредством пептидной связи в слитый белок по изобретению, предусматривается, что нежелательная утечка Z-домена из белковых структур по изобретению является низкой или отсутствует. Другое преимущество слитых белков по изобретению состоит в том, что полученная белковая структура имеет высокую плотность Z-доменов (или других частей B). Предусматривается, что эта высокая плотность обеспечивает высокую связывающую способность. Взятые вместе, эти свойства слитых белков являются в высокой степени привлекательными для различных частей B, и, в частности, для аффинной очистки с использованием белка Z с высокой экономией при продукции. Эти свойства также пригодны в форматах, отличных от традиционных аффинных колонок с гелевыми гранулами, например в форматах фильтров.
Иммобилизованная органическая мишень способна к селективному взаимодействию со второй органической мишенью. Следовательно, способ, кроме того, включает стадию контактирования указанной аффинной среды и иммобилизованной органической мишени со второй органической мишенью, которая способна к селективному взаимодействию с первой органической мишенью в подходящих условиях для достижения связывания между первой и второй органическими мишенями.
Иммобилизованная органическая мишень поддается детекции и/или количественному определению. Детекцию и/или количественное определение органической мишени можно проводить любым известным специалисту в данной области способом детекции и/или количественного определения связывающих реагентов в анализах на основе различных биологических или небиологических взаимодействий. Сами органические мишени можно метить различными маркерами или в свою очередь их можно выявлять с помощью вторичных меченых аффинных лигандов для обеспечения детекции, визуализации и/или количественного определения. Это можно осуществлять с использованием любой одной или нескольких из множества меток, которые могут быть конъюгированы с органической мишенью или с любым вторичным аффинным лигандом с использованием любого одного или нескольких из множества способов, известных квалифицированному специалисту, и, по существу, не вовлекая никакого излишнего экспериментирования. Неограничивающие примеры меток, которые можно конъюгировать с органическими мишенями и/или вторичными аффинными лигандами, включают флуоресцентные красители или металлы (например, флуоресцеин, родамин, фикоэритрин, флуорескамин), хромофорные красители (например, родопсин), хемилюминесцентные соединения (например, люминал, имидазол) и биолюминесцентные белки (например, люциферин, люцифераза), гаптены (например, биотин). Различные другие подходящие флуоресцирующие вещества и хромофоры описаны в Stryer L (1968) Science 162:526-533 и Brand L and Gohlke JR (1972) Annu. Rev. Biochem. 41:843-868. Органические мишени и/или вторичные аффинные лиганды также можно метить ферментами (например, пероксидаза хрена, щелочная фосфатаза, бета-лактамаза), радиоизотопами (например, 3H, 14C, 32P, 35S или 125I) и частицами (например, золото). В контексте настоящего описания "частицы" относятся к частицам, таким как металлические частицы, подходящие для мечения молекул. Кроме того, аффинные лиганды также можно метить флуоресцентными полупроводниковыми нанокристаллами (квантовые точки). Квантовые точки имеют более высокий квантовый выход и являются более фотостабильными по сравнению с органическими флуорофорами и, таким образом, их легче выявлять (Chan et al. (2002) Curr Opi Biotech. 13: 40-46). Различные типы меток можно конъюгировать с органической мишенью или вторичным аффинным лигандом с использованием различных химических реакций, например, реакции аминов или реакции тиолов. Однако, помимо аминов и тиолов, можно использовать другие реакционноспособные группы, например, альдегиды, карбоновые кислоты и глутамин.
Если детекция/или количественное определение вовлекает воздействие второй органической мишени или вторичного аффинного лиганда, аффинную среду промывают еще один раз буферами для удаления не связавшихся вторичных аффинных лигандов. В качестве примера, вторичный аффинный лиганд может представлять собой антитело или его фрагмент или производное. После этого, органические мишени можно подвергать детекции и/или количественному определению общепринятыми способами. Связывающие свойства для вторичного аффинного лиганда могут варьировать, однако специалисты в данной области должны быть способны определить действующие и оптимальные условия анализа для каждого определения путем стандартного экспериментирования.
Детекция, определение локализации и/или количественное определение меченой молекулы могут вовлекать способы визуализации, такие как световая микроскопия или иммунофлуоресцентная микроскопия. Другие способы могут вовлекать детекцию с помощью цитометрии или люминометрии. Способ визуализации меток может включать, но не ограничиваться ими, флуориметрический, люминометрический и/или ферментативный способы. Детекцию и/или количественное определение проводят путем воздействия на флуоресцентные метки света определенной длины волны, и, после этого, детекции и/или количественного определения испускаемого света в конкретной области длин волн. Детекцию и/или количественное определение присутствия люминесцентно меченой молекулы можно проводить с помощью люминесценции, появляющейся в ходе химической реакции. Детекцию ферментативной реакции проводят по цветовому сдвигу в образце, полученном при химической реакции. Специалистам в данной области известно, что множество различных протоколов можно модифицировать для надлежащей детекции и/или количественного определения.
Одним доступным способом детекции и/или количественного определения органической мишени является связывание ее с вторичным аффинным лигандом или ферментом, который затем можно подвергать детекции и/или количественному определению в иммуноферментном анализе (таком как EIA или ELISA). Такие способы являются общепринятыми и их осуществление не представляет никаких излишних трудностей для квалифицированного специалиста. В таких способах биологический образец контактируют с белковой структурой по изобретению, которая связывается с органической мишенью, которую затем подвергают детекции и/или количественному определению с меченным ферментом вторичным аффинным лигандом. После этого соответствующий субстрат подвергают реакции в соответствующих буферах с ферментной меткой с получением химической части, которую, например, подвергают детекции и/или количественному определению с использованием спектрофотометра, флуориметра, люминометра или визуальными способами.
Органическую мишень или вторичные аффинные лиганды можно метить радиоизотопами для обеспечения детекции и/или количественного определения. Неограничивающими примерами подходящих радиоактивных меток в рамках настоящего изобретения являются 3H, 14C, 32P, 35S или 125I. Удельная активность меченого аффинного лиганда зависит от времени полужизни радиоактивной метки, изотопной чистоты и того, как метка включается в аффинный лиганд. Аффинные лиганды предпочтительно метят с использованием хорошо известных способов (Wensel TG and Meares CF (1983): Radioimmunoimaging and Radioimmunotherapy (Burchiel SW and Rhodes BA eds.) Elsevier, New York, pp 185-196). Радиоактивно меченный таким путем аффинный лиганд можно использовать для визуализации органической мишени с помощью детекции радиоактивности. Радиоизотопное сканирование можно проводить, например, с помощью гамма-камеры, спектроскопии магнитного резонанса, эмиссионной томографии, гамма/бета-счетчиков, сцинтилляционных счетчиков и радиографии.
Таким образом, образец можно наносить на белковую структуру для детекции, выделения и/или количественного определения органической мишени. Эта методика обеспечивает не только детекцию органической мишени, но, кроме того, может демонстрировать распределение и относительный уровень ее экспрессии. Необязательно, органическую мишень можно высвобождать из аффинной среды и собирать. Таким образом, применение может включать аффинную очистку на аффинной среде, на которой иммобилизована органическая мишень. Белковая структура, например, может быть расположена в колонке или в планшетах с лунками (таких как 96-луночные планшеты), или на магнитных гранулах, агарозных гранулах или гранулах из сефарозы. Кроме того, применение может включать использование белковых структур на растворимой матрице, например, с использованием декстрановой матрицы, или использование в устройстве поверхностного плазмонного резонанса, таком как устройстве BiacoreTM, где анализ, например, может включать мониторинг аффинности к иммобилизованной органической мишени или ряду потенциальных аффинных лигандов.
Белковые структуры по изобретению можно промывать и регенерировать с помощью различных очищающих агентов, включая кислоту, основание и хаотропные агенты. Особенно пригодные очищающие агенты включают NaOH, такой как 0,1, 0,5 или 1 M NaOH, и мочевину, такую как 6-8 M мочевина. Поскольку белковые структуры по изобретению являются неожиданно устойчивыми к химической обработке и/или стерилизующей обработке нагреванием, способы по изобретению, вовлекающие применение белковых структур, могут включать конечную стадию регенерации белковой структуры. Способы предпочтительно включают конечную стадию регенерации аффинной среды посредством химической обработки и/или стерилизующей обработки нагреванием. Предпочтительно, чтобы химическая обработка включала обработку NaOH, таким как 0,1, 0,5 или 1 M NaOH, и/или мочевиной, такой как 6-8 M мочевина.
Слитым белкам по изобретению также можно позволять связываться с органической мишенью в растворе, т.е. до того, как позволить слитому белку полимеризоваться и образовать белковую структуру, такую как пленка, пена или волокно. Как происходящие из спидроина части (например REP-CT) как таковые, так и соответствующие слитые белки, включающие часть B, полимеризуются в твердые структуры, даже в присутствии контаминирующих белков, без поддающегося оценке включения примесей в материал, и функциональные части (B) сохраняют их ожидаемые связывающие свойства. Таким образом, предусматривается, что связывающие свойства части B можно использовать для улавливания соединения или клеток из окружающего раствора и включения уловленных соединений или клеток в белковую структуру или на белковую структуру по изобретению.
Таким образом, в другом предпочтительном способе по изобретению слитый белок в аффинной среде присутствует в растворе при контактировании аффинной среды с образцом для достижения связывания между аффинной средой и органической мишенью. Затем комплексу слитого белка с органической мишенью позволяют сформировать структуру слитого белка по изобретению.
Этот способ может быть особенно пригодным, когда целью является "вылавливание" конкретных молекул или клеток из раствора, например, для получения молекул-мишеней из среды в крупномасштабных системах продуцирования из эукариотических клеток, когда белки-мишени секретируются. Поскольку связывание молекул-мишеней и образование твердых структур происходящими из спидроина частями может происходить при физиологических условиях и поскольку происходящие из спидроина части являются совместимыми с клетками, способ можно применять неоднократно для непрерывного процесса продукции.
Белковая структура по изобретению также пригодна для выделения, иммобилизации и/или культивирования клеток. Особенно пригодной белковой структурой в этом отношении является пленка, волокно или пена, см. например, пример 14 и 23. Пленка является преимущественной тем, что она прикрепляется к твердым структурам, например, к пластмассе в микропланшетах для титрования. Это свойство пленки способствует промыванию и регенерации и в высокой степени пригодно для целей выделения.
Таким образом, настоящее изобретение относится к каркасному материалу для клеток, чтобы культивировать клетки, имеющие органическую мишень на клеточной поверхности. Каркасный материал для клеток содержит белковую структуру по изобретению. В определенных вариантах осуществления каркасный материал для клеток состоит из белковой структуры по изобретению.
Авторы настоящего изобретения открыли, что каркасный материал для клеток, содержащий полимер, содержащий и необязательно состоящий из слитого белка по изобретению, относится к благоприятной окружающей среде для культивирования клеток, и предпочтительно эукариотических клеток, во множестве различных условий. Более того, эта окружающая среда обеспечивает установление культур клеток, которые в ином случае очень трудно, с высокими затратами или даже невозможно культивировать в лаборатории, и получение содержащих клетки материалов, пригодных для инженерии тканей и/или трансплантации.
Также изобретение относится к комбинации клеток, предпочтительно эукариотических клеток, и каркасного материала для клеток по изобретению. Такая комбинация по изобретению может присутствовать во множестве различных форматов, и она может быть подобрана так, чтобы удовлетворять потребности данной конкретной ситуации. Например, предусматривается, что комбинация по изобретению может быть пригодна в качестве содержащего клетки имплантата для замены клеток в поврежденной или пораженной заболеванием ткани.
Каркасный материал для клеток можно использовать для улавливания клеток прямо или непрямо. При прямом культивировании часть B способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности. Альтернативно часть B способна к селективному взаимодействию и связывается с промежуточной органической мишенью, и эта промежуточная органическая мишень способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности. Таким образом, при непрямом улавливании каркасный материал для клеток, кроме того, содержит промежуточную органическую мишень, и часть B способна к селективному взаимодействию и связывается с указанной промежуточной органической мишенью. Промежуточная органическая мишень, в свою очередь, способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности.
В одном варианте осуществления каркасных материалов для клеток, описанных в настоящем описании, слитый белок, кроме того, содержит олигопептидный связывающийся с клеткой мотив. Применительно к культивированию определенных клеток в определенных ситуациях, было обнаружено, что присутствие олигопептидных связывающихся с клеткой мотивов повышает или сохраняет жизнеспособность клеток, и полагают, что включение такого мотива в каркасный материал для клеток в качестве части белка шелка пауков обеспечит дополнительные преимущества. Связывающийся с клеткой мотив представляет собой олигопептид, присоединенный к остальной части слитого белка через по меньшей мере одну пептидную связь. Например, он может быть присоединен к N-концу или C-концу остальной части слитого белка или в любом положении в аминокислотной последовательности остальной части белка шелка пауков. Что касается выбора олигопептидных связывающихся с клеткой мотивов, квалифицированному специалисту известно несколько альтернатив. Указанный олигопептид может, например, содержать аминокислотную последовательность, выбранную из группы, состоящей из RGD, IKVAV, YIGSR, EPDIM и NKDIL. RGD, IKVAV и YIGSR являются общими связывающимися с клеткой мотивами, в то время как EPDIM и NKDIL известны в качестве кератиноцит-специфических мотивов, которые могут быть особенно пригодными в контексте культивирования кератиноцитов. Другие пригодные связывающиеся с клеткой мотивы включают GRKRK из тропоэластина, KYGAASIKVAVSADR (происходящий из ламинина), NGEPRGDTYRAY (из сиалопротеина костей), PQVTRGDVFTMP (из витронектина) и AVTGRGDSPASS (из фибронектина). Присоединение олигопептидного связывающегося с клеткой мотива к остальной части белка шелка пауков легко может быть без труда осуществлено квалфицированным специалистом с использованием стандартных способов генетической инженерии или химического присоединения. Таким образом, в некоторых вариантах осуществления связывающийся с клеткой мотив вносят путем генетической инженерии, т.е. путем формирования части генетической слитой конструкции между нуклеиновой кислотой, кодирующей слитый белок, и связывающимся с клеткой мотивом. В качестве дополнительной благоприятной характеристики таких вариантов осуществления связывающийся с клеткой мотив присутствует в соотношении 1:1 с мономерами слитого белка в полимере, составляющем каркасный материал для клеток.
Полимер в каркасном материале для клеток, используемом в способах или комбинации, описанных в настоящем описании, может принимать множество физических форм, и использование конкретной физической формы может обеспечить дополнительные преимущества в различных конкретных ситуациях. Например, в одном варианте осуществления способов или комбинации, указанный каркасный материал для клеток находится в физической форме, выбранной из группы, состоящей из пленки, пены, волокна и волокнистого сита.
Таким образом, настоящее изобретение относится к способу иммобилизации клеток. Предусмотрен образец, например, биологический образец, такой как кровь, содержащий представляющие интерес клетки. Биологический образец может представлять собой полученный ранее образец. Если в способе используют ранее полученный образец, никакие стадии способа не осуществляют в организме человека или животного.
Образец наносят на каркасный материал для клеток по изобретению в подходящих условиях для обеспечения селективного взаимодействия между каркасным материалом для клеток и органической мишенью, которая присутствует на поверхности представляющих интерес клеток. Клеткам позволяют иммобилизоваться на указанном каркасном материале для клеток путем связывания между органической мишенью на клеточной поверхности и указанным каркасным материалом для клеток. Не связавшийся образец удаляют в подходящих условиях для сохранения селективного связывания между каркасным материалом для клеток и органической мишенью. Этот способ обеспечивает клетки, имеющие органическую мишень, иммобилизованную на каркасном материале для клеток и, в частности, на белковой структуре по изобретению.
Как указано выше, каркасный материал для клеток можно использовать для улавливания клеток, либо прямо, либо непрямо. При прямом улавливании часть B способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности. Альтернативно часть B способна к селективному взаимодействию и связывается с промежуточной органической мишенью, и эта промежуточная органическая мишень способна к селективному взаимодействию c органической мишенью, которая присутствует на клеточной поверхности. Таким образом, при непрямом улавливании каркасный материал для клеток, кроме того, содержит промежуточную органическую мишень, и часть B способна к селективному взаимодействию и связывается с указанной промежуточной органической мишенью. Промежуточная органическая мишень, в свою очередь, способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности.
Независимо от способа улавливания, уловленные клетки можно освобождать от слитого белка путем расщепления слитого белка для высвобождения части, вовлеченной в улавливание клеток, из каркасного материала для клеток. Как упоминалось в настоящем описании выше, слитый белок может включать участок расщепления в его аминокислотной последовательности, который позволяет расщепление и удаление соответствующей части, как правило, части B или связывающегося с клеткой мотива. Специалисту в данной области различные участки расщепления, например, участки расщепления для химических агентов, таких как CNBr после остатков Met и гидроксиламин между остатками Asn-Gly, участки расщепления для протеаз, таких как тромбин или протеаза 3C, и последовательности самосплайсинга, такие как последовательности самосплайсинга интеинов.
Также настоящее изобретение относится к способу культивирования клеток. Представляющие интерес клетки иммобилизуют на каркасном материале для клеток с использованием способа, описанного в настоящем описании выше. Комбинацию каркасного материала для клеток и иммобилизованных клеток поддерживают в условиях, пригодных для культивирования клеток.
В контексте настоящего изобретения, термины "культивирование" клеток, "клеточная культура" и т.д. интерпретируются широко, так что они охватывают, например, ситуации, в которых клетки делятся и/или пролиферируют, ситуации, в которых клетки поддерживаются в различном дифференцированном состоянии с сохранением по меньшей мере одной функциональной характеристики, проявляемой типом клеток, когда он находится в естественных окружающих условиях, и ситуации, в которых стволовые клетки поддерживают в недифференцированном состоянии.
Настоящее изобретение далее проиллюстрировано с помощью следующих неограничивающих примеров.
Примеры
Пример 1 - Клонирование, экспрессия и образование волокон связывающего IgG слитого белка
Для подтверждения концепции слитого белка, получали белок Rep4CT (часть REP с 4 внутренними повторами и часть CT) в виде слитой конструкции с Z-доменом белка (часть B). Z-домен представляет собой полученную способами инженерии версию связывающего иммуноглобулин G (IgG) B-домена белка A стафилококков, и он представляет собой трехспиральный мотив длиной 58 аминокислот, который связывает область кристаллизующегося фрагмента (Fc) IgG. Целью авторов настоящего изобретения было исследование того, возможно ли получить структуры, такие как волокна, пленки и мембраны, из слитого белка, состоящего из Z-домена, слитого с Rep4CT (обозначаемый как His6ZQGRep4CT, SEQ ID NO: 14) и будут ли они все еще сохранять способность Z-домена связывать IgG, а также свойства Rep4CT в отношении формирования структуры. Для этого клонировали слитый белок, состоящий из Z-домена на N-конце Rep4CT.
Клонирование
Ген, кодирующий слитый белок His6ZQGRep4CT (SEQ ID NO: 14-15) конструировали следующим образом. Праймеры конструировали для получения ПЦР-фрагментов Z-домена из вектора, содержавшего такую последовательность Z. Также праймеры содержали участок распознавания для расщепления протеазой 3C (LEALFQGP, обозначаемый как QG) между Z и Rep4CT. Затем полученные продукты ПЦР обрабатывали эндонуклеазами рестрикции NdeI и EcoRI, также как и вектор-мишень (обозначаемый как pT7His6TrxHis6QGRep4CT, содержащий ген устойчивости к канамицину). При рестрикционном расщеплении вектора-мишени часть TrxHis6QG отщеплялась. Расщепленные ПЦР-фрагменты и вектор-мишень связывали вместе с помощью ДНК-лигазы T4, после чего полученный правильно лигированный вектор (pT7His6ZQGRep4CT) трансформировали в химически компетентные клетки Escherichia coli (E. coli) BL21 (DE3), которым позволяли расти на чашках с агаром, дополненных канамицином (70 мкг/мл). После этого колонии отбирали и подвергали скринингу ПЦР в отношении правильной вставки, а затем также секвенировали для подтверждения встроенной последовательности ДНК ZQG в вектор-мишень.
Продуцирование
Клетки E. coli BL21 (DE3), имеющие вектор pT7His6ZQGRep4CT, выращивали в среде Луриа-Бертани (всего 6 литров), дополненной канамицином (70 мкг/мл) до величины OD600 1-1,5 при 30°C, с последующей индукцией экспрессии His6ZQGRep4CT с помощью 300 мкМ IPTG (изопропил-β-D-1-тиогалактопиранозид) и инкубацией при 20°C в течение приблизительно 2 ч. Далее клетки собирали центрифугированием в течение 20 мин при 4700 об./мин. и полученный клеточный осадок растворяли в 20 мМ Tris (pH 8,0).
Очистка
Клеточные осадки, растворенные в 20 мМ Tris (pH 8,0), дополняли лизоцимом и ДНКазой I для лизиса бактериальных клеток, после чего клеточные лизаты выделяли после центрифугирования при 15000 об./мин. в течение 30 мин. Далее выделенные клеточные лизаты разделяли и наносили всего на четыре колонки Chelating Sepharose Fast Flow Zn2+, поддерживая белок His6ZQGRep4CT связанным с матрицей колонки через His6-метку. После промывания связанные белки элюировали 20 мМ Tris/300 мМ имидазолом (pH 8,0). Объединенные фракции элюата содержали 27 мг белка His6ZQGRep4CT согласно измерению при A280. Далее, объединенную жидкость элюата разделяли на две равные половины (13,5 мг His6ZQGRep4CT в каждой), где первую половину подвергали диализу против 5 литров 20 мМ Tris (pH 8,0) в течение ночи, концентрировали до 1,07 мг/мл и, наконец, позволяли образовать волокна. На фиг. 3 представлено макроскопическое волокно His6ZQGRep4CT. Образование волокон из этого слитого белка (SEQ ID NO: 14) иллюстрирует, что Z-домен (часть B) не препятствует свойствам образования волокон Rep4CT (части REP и CT). Количество белка His6ZQGRep4CT перед расщеплением протеазой 3C составляло 10 мг после диализа и концентрирования (фиг. 4).
Вторую половину совокупного элюата расщепляли 1,34 мг протеазы 3C, дополненной 1 мМ дитиотреитолом (DTT), отщепляя His6Z от Rep4CT. Расщепление проводили в течение ночи при диализе против 20 мМ Tris (pH 8,0), после чего раствор белка пропускали через колонку с агарозой Ni-NTA, и прошедшую фракцию, содержавшую Rep4CT, собирали, концентрировали до 0,79 мг/мл и позволяли образовывать волокна. Конечное количество Rep4CT после расщепления составляло 6 мг (фиг. 4).
На фиг. 4 представлен гель SDS-PAGE после очистки слитого белка His6ZQGRep4CT (SEQ ID NO: 14) и его продукта Rep4CT после последующего расщепления протеазой 3C (остатки 81-339 SEQ ID NO: 14). Гель наносили в следующем порядке:
(1) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(2) Клеточный лизат
(3) Прошедшая фракция клеточного лизата, нанесенного на колонку Chelating Sepharose Fast Flow Zn2+
(4) Совокупный элюат His6ZQGRep4CT из колонки Chelating Sepharose Fast Flow Zn2+
(5) His6ZQGRep4CT, расщепленный протеазой 3C
(6) Прошедшая фракция расщепленного His6ZQGRep4CT, нанесенного на колонку с агарозой Ni-NTA
(7) Регенерация колонки Ni-NTA 5 мл смеси 20 мМ Tris/500 мМ имидазол (pH 8,0).
Молекулярные массы His6ZQGRep3CT, His6Z, Rep4CT и протеазы 3C составляют 32 кДа, 9 кДа, 23 кДа и 30 кДа, соответственно.
Тот факт, что макроскопические волокна His6ZQGRep4CT (SEQ ID NO: 14) можно получить, несмотря на то, что Rep4CT слит с другим белком, т.е. Z-доменом длиной 58 аминокислот с аффинностью связывания к IgG, демонстрирует, что Rep4CT все еще сохраняет его свойства образования волокон, несмотря на то, что он слит с Z-доменом (остатки 13-70 SEQ ID NO: 14). Более того, Z-домен слитого белка, по-видимому, обеспечивает хорошую растворимость His6ZQGRep4CT.
Пример 2 - Связывание биотинилированного IgG с волокнами слитого белка
Для дальнейшего подтверждения идеи слитого белка проводили исследование того, сохраняет ли часть B в слитой белковой структуре ее способность к селективному взаимодействию с органической мишенью. В этом исследовании оценивали способность Z-домена (части B) в волокнах слитого белка His6ZQGRep4CT (SEQ ID NO: 14) связывать IgG. Раствор биотинилированного IgG кролика инкубировали с волокнами His6ZQGRep4CT, после чего те же волокна инкубировали в растворе с функционализированными стрептавидином гранулами, а затем волокна визуализировали под световым микроскопом. Выбор использования IgG, образовавшегося в кролике, основан на том факте, что IgG из кролика связывается с Z-доменом с высокой аффинностью.
Волокно His6ZQGRep4CT длиной приблизительно 50 мм, полученное, как описано в примере 1, погружали в раствор для связывания, содержащий 50 мкл 1×PBS/0,5% бычьего сывороточного альбумина и 10 мкл 0,5 мг/мл биотинилированного IgG, происходящего из кролика (адсорбированное антитело мыши против IgG (H+L) крысы, Vector Laboratories, Inc.), и инкубировали в течение 75 мин при комнатной температуре при легком встряхивании. Супернатант удаляли и волокно промывали три раза 60 мкл 1×PBS/0,07% Tween 20. Далее, волокно погружали в раствор, содержавший 40 мкл 1×PBS/0,5% бычьего сывороточного альбумина и 20 мкл 10 мг/мл Dynabeads M-280 Streptavidin (Dynal AS), и вновь инкубировали в течение 75 мин при комнатной температуре при легком встряхивании. Супернатант удаляли и волокно три раза промывали в 60 мкл 1×PBS/0,07% Tween 20.
Чтобы получить индикатор неспецифического связывания Dynabeads с волокном, другое волокно His6ZQGRep4CT погружали только в раствор Dynabeads, как описано выше, без предшествующей инкубации с биотинилированным IgG. Ту же методику, как описано выше для His6ZQGRep4CT, проводили с волокнами типа Rep4CT, и все волокна визуализировали под микроскопом USB с фиксированным увеличением 500×.
На фиг. 5 представлена визуализация волокон His6ZQGRep4CT и Rep4CT после связывания биотинилированного IgG кролика, а затем функционализированных стрептавидином Dynabeads. На панелях (A, B) представлено два типичных изображения волокна His6ZQGRep4CT, полученных в различных положениях вдоль волокна, которое сначала инкубировали с биотинилированным IgG (образовавшимся у кролика), а затем инкубировали с Dynabeads M-280 Streptavidin (Ø 2,8 мкм). На панелях (C, D) представлено два типичных изображения другого волокна His6ZQGRep4CT, которое погружали только в раствор Dynabeads M-280 Streptavidin, без предшествующей инкубации с IgG. Соответствующие изображения волокон Rep4CT представлены на панелях (E, F) и (G, H), соответственно. Все изображения получали при фиксированном увеличении 500× под микроскопом USB, и Dynabeads выглядят на изображениях как темно-серые точки.
На фиг. 5 на панелях A и B видно, что видны практически только Dynabeads, причем на обоих из этих изображений представлено волокно His6ZQGRep4CT, подвергнутое воздействию биотинилированного IgG, а затем функционализированных стрептавидином Dynabeads. Этот результат подразумевает, что значительная часть Z-доменов в слитом белке сохраняет их способность связывать IgG в His6ZQGRep4CT после формирования волокна.
Пример 3 - Связывание чистого IgG и сывороточного IgG с волокнами и пленками слитого белка
Для дальнейшего исследования способности части B в слитой белковой структуре к селективному взаимодействию с органической мишенью исследовали способность Z-домена в слитом белке His6ZQGRep4CT (SEQ ID NO: 14) связывать IgG. Волокна и пленки этого слитого белка использовали для связывания очищенного IgG и IgG из сыворотки с последующим элюированием и анализом на SDS-PAGE, где IgG при невосстанавливающих условиях выглядит как полоса, соответствующая массе ~146 кДа. Сыворотка представляет собой остающуюся жидкую фазу после свертывания крови и двумя основными составляющими сыворотки являются альбумин и IgG. В сыворотке кролика, например, концентрация IgG составляет 5-10 мг/мл и концентрация альбумина является еще более высокой. Источником как очищенного IgG, так и сыворотки, был кролик.
Пленки His6ZQGRep4CT получали путем высушивания воздухом 100 мкл раствора белка (0,96 мг/мл) в течение ночи при комнатной температуре на дне индивидуальных ячеек 24-луночного планшета для культивирования тканей. Сформированные пленки затем хранили при +4°C в течение 18 суток, либо погруженными в 20 мМ Tris (pH 8,0), что обозначают как "T-пленки", либо без погружения в какую-либо жидкость, что обозначают как "A-пленки". На фиг. 6 представлена часть сформированной пленки His6ZQGRep4CT, полученной на дне ячейки гидрофильного планшета для культивирования тканей с 24 ячейками. Для получения изображения использовали инвертационный световой микроскоп при увеличении 2×. Волокна слитого белка получали, как описано в примере 1, и хранили при +4°C в 20 мМ Tris (pH 8,0) до применения.
Выполняли два параллельных экспериментальных протокола. В первом протоколе T-пленки, A-пленки и волокна, изготовленные из His6ZQGRep4CT, в трех экземплярах погружали в 500 мкл очищенного IgG кролика в концентрации 50 мкг/мл (очищенного из совокупной сыворотки кролика, Vector Laboratories, Inc.) в течение 1 ч при комнатной температуре при мягком встряхивании. В другом протоколе пленки и волокна His6ZQGRep4CT того же типа в трех экземплярах альтернативно погружали в 500 мкл пятикратного разведения инактивированной нагреванием центрифугированной сыворотки кролика (National Veterinary Institute, Uppsala, Швеция), также в течение 1 ч при комнатной температуре при мягком встряхивании. Супернатант удаляли из всех волокон и пленок, а затем промывали три раза в 500 мкл 20 мМ Tris (pH 8,0). Связанный IgG из очищенного IgG или из сыворотки элюировали путем инкубации в течение 30 мин в 500 мкл 0,5 M уксусной кислоты/1 M мочевины/100 мМ NaCl (pH 2,7). Ту же методику, как описано выше для волокон His6ZQGRep4CT и A-пленок, также проводили для пленок и волокон His6TrxHis6QGRep4CT и Rep4CT в качестве контролей. Элюированные фракции анализировали с помощью SDS-PAGE в невосстанавливающих условиях (фиг. 7-9).
На фиг. 7 представлен невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили на дорожки следующим образом:
(1-3) His6TrxHiS6QGRep4CT, A-пленка, инкубированная с IgG кролика
(4-6) His6TrxHis6QGRep4CT, A-пленка, инкубированная с сывороткой кролика
(7-8) His6TrxHis6QGRep4CT, волокно, инкубированное с IgG кролика
(9) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(10-11) His6TrxHis6QGRep4CT, волокно, инкубированное с сывороткой кролика
(12-14) His6ZQGRep4CT, T-пленка, инкубированная с IgG кролика
(15-17) His6ZQGRep4CT, T-пленка, инкубированная с сывороткой кролика.
На фиг. 8 представлен другой невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили в соответствии с:
(1-3) His6ZQGRep4CT, A-пленка, инкубированная с IgG кролика
(4-6) His6ZQGRep4CT, A-пленка, инкубированная с сывороткой кролика
(7-9) His6ZQGRep4CT, волокно, инкубированное с IgG кролика
(10) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(11-13) His6ZQGRep4CT, волокно, инкубированное с сывороткой кролика
(14-16) Rep4CT, A-пленка, инкубированная с IgG кролика.
На фиг. 9 представлен следующий невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили в соответствии с:
(1-3) Rep4CT, A-пленка, инкубированная с сывороткой кролика
(4-6) Rep4CT, волокно, инкубированное с IgG кролика
(7) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(8-10) Волокно Rep4CT, инкубированное с сывороткой кролика
(11) Очищенный IgG кролика (50 мкг/мл), использованный при инкубации
(12) Сыворотка кролика (разведение 1:50), используемая при инкубации.
Результаты на фиг. 7-9 демонстрируют, что все типы матриц His6ZQGRep4CT, т.е. пленок (как A-типа, так и T-типа) и волокон способны связывать IgG (либо чищенный, либо из сыворотки) через Z-домен. Более того, матрицы His6ZQGRep4CT, которые подвергали воздействию сыворотки кролика, по-видимому, не связывали ничего из фракции сыворотки, кроме IgG. Использованные матрицы двух других белковых вариантов (т.е. His6TrxHis6QGRep4CT и Rep4CT), по-видимому, не связывали вообще ничего, за исключением волокон Rep4CT, подвергнутых воздействию сыворотки, и это демонстрирует слабые полосы в области IgG (~146 кДа) и альбумина (~70 кДа). Этот подход для оценки способности Z-домена связывать IgG в слитом белке His6ZQGRep4CT (SEQ ID NO: 14) строго указывает на то, что Z-домен является активным как в версии волокна, так и в версии пленки, слитого белка. Не было обнаружено никакой другой фракции сыворотки кролика, отличной от IgG, связывающейся с His6ZQGRep4CT.
Пример 4 - Воспроизводимость связывания чистого и сывороточного IgG с волокнами и пленками слитого белка
Для исследования воспроизводимости связывания IgG с пленками и волокнами слитого белка His6ZQGRep4CT (SEQ ID NO: 14), вновь проводили эксперименты, представленные в примере 3. Вновь использовали те же волокна и пленки His6ZQGRep4CT, His6TrxHis6QGRep4CT и Rep4CT, которые использовали в примере 3. Все материалы волокон и пленок погружали на 70 суток в 20 мМ Tris (pH 8,0) при +4°C после проведения предшествующих экспериментов. Эксперименты проводили, как описано в примере 3. Элюированные фракции анализировали с помощью SDS-PAGE в невосстанавливающих условиях (фиг. 10-12).
На фиг. 10 представлен невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили в соответствии с:
(1-3) His6TrxHis6QGRep4CT, A-пленка, инкубированная с IgG кролика
(4-6) His6TrxHis6QGRep4CT, A-пленка, инкубированная с сывороткой кролика
(7-8) His6TrxHis6QGRep4CT, волокно, инкубированное с IgG кролика
(9) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(10-11) His6TrxHis6QGRep4CT, волокно, инкубированное с сывороткой кролика
(12-14) His6ZQGRep4CT, T-пленка, инкубированная с IgG кролика
(15-17) His6ZQGRep4CT, T-пленка, инкубированная с сывороткой кролика.
На фиг. 11 представлен другой невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили в соответствии с:
(1-3) His6ZQGRep4CT, A-пленка, инкубированная с IgG кролика
(4-6) His6ZQGRep4CT, A-пленка, инкубированная с сывороткой кролика
(7-9) His6ZQGRep4CT, волокно, инкубированное с IgG кролика
(10) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(11-13) His6ZQGRep4CT, волокно, инкубированное с сывороткой кролика
(14-16) Rep4CT, A-пленка, инкубированная с IgG кролика.
На фиг. 12 представлен невосстанавливающий гель SDS-PAGE. Элюированные фракции наносили в соответствии с:
(1-3) Rep4CT, пленка A-типа, инкубированная с сывороткой кролика
(4-6) Rep4CT, волокно, инкубированное с IgG кролика
(7) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(8-10) Rep4CT, волокно, инкубированное с сывороткой кролика
(11) Пустая лунка
(12) Очищенный IgG кролика (50 мкг/мл), используемый для инкубации
(13) Сыворотка кролика (разведение 1:50), используемая для инкубации.
Профили связывания IgG со всеми тремя белковыми матрицами (т.е. His6ZQGRep4CT, His6TrxHis6QGRep4CT и Rep4CT) соответствуют тому, что было выявлено в примере 3, хотя для волокон His6ZQGRep4CT, инкубированных с сывороткой кролика, вместо этого наблюдают слабые полосы, соответствующие альбумину (~70 кДа). Это первоначальное исследование воспроизводимости связывания IgG демонстрирует, что как волокна, так и пленки His6ZQGRep4CT, можно использовать по меньшей мере два раза для связывания и элюирования очищенного и сывороточного IgG. Следующее исследование воспроизводимости описано в примере 19.
Пример 5 - Связывание IgG с матрицами слитого белка и с коммерческой матрицей белка A
Способность связывать IgG слитых белковых структур His6ZQGRep4CT оценивали путем сравнения с коммерчески доступной матрицей белка A (Protein A Sepharose CL-4B, GE Healthcare). Волокна и пленки His6ZQGRep4CT (SEQ ID NO: 14) получали внутри центрифужных колонок. Матрицу белка A также добавляли к центрифужным колонкам так, чтобы общее количество Z молекул белка A, связанных с матрицей, было равно общему количеству молекул в пленках His6ZQGRep4CT. Позволяли происходить связыванию очищенного IgG и IgG из сыворотки с пленками и волокнами His6ZQGRep4CT, а также с матрицей белка A, с последующим элюированием и анализом на SDS-PAGE.
Пленки His6ZQGRep4CT получали путем высушивания воздухом 100 мкл раствора белка (1,05 мг/мл) в течение трех суток при комнатной температуре на дне полиэтиленовой фритты внутри центрифужных колонок (SigmaPrep™ Spin Columns, Sigma), с учетом всего 3×10-9 моль His6ZQGRep4CT на пленку. Также волокна His6ZQGRep4CT помещали на фритты внутри того же типа центрифужных колонок. Ту же методику выполняли для пленок и волокон Rep4CT, где пленки содержали всего 4×10-9 моль Rep4CT на пленку. Для коммерческой матрицы с белком A, отжатый объем матрицы, соответствующий 3×10-9 моль белка A, переносили во фритту на каждую центрифужную колонку, где матрицу промывали 1×500 мкл плюс 2×150 мкл деионизированной воды центрифугированием центрифужных колонок при 400 оцс в течение 1,5 мин.
Выполняли два параллельных экспериментальных протокола для волокон и пленок His6ZQGRep4CT и Rep4CT, а также для матрицы белка A. В первом протоколе все три различных матрицы в двух экземплярах погружали в 500 мкл 50 мкг/мл очищенного IgG кролика (IgG, очищенный из сыворотки кролика, Sigma) в течение 1 ч при комнатной температуре. В другом протоколе те же три типа матриц в двух экземплярах вместо этого погружали в 500 мкл пятикратного разведения центрифугированной сыворотки кролика (нормальная сыворотка кролика, Invitrogen), также в течение 1 ч при комнатной температуре.
Супернатант удаляли из всех волокон и пленок путем простого пипетирования и для матрицы белка A центрифугированием (400 оцс, 1,5 мин), а затем промывали три раза в 500 мкл 20 мМ Tris (pH 8,0). Связанный IgG из очищенного IgG или из сыворотки элюировали путем инкубации в течение 30 мин в 500 мкл 0,5 M уксусной кислоты/1 M мочевины/100 мМ NaCl (pH 2,7). Элюированные фракции анализировали на SDS-PAGE в невосстанавливающих условиях (фиг. 13-15), и их обозначали как полученные при прогоне 1. Сразу после элюирования все матрицы промывали 3×500 мкл 20 мМ Tris (pH 8,0), после чего описанный выше эксперимент повторяли еще один раз для оценки воспроизводимости связывания IgG. Элюированные фракции из повторного эксперимента обозначены как полученные при прогоне 2. Примечание: в невосстанавливающих условиях SDS-PAGE молекулярная масса IgG составляет приблизительно 146 кДа.
На фиг. 13 представлен невосстанавливающий гель SDS-PAGE элюированных фракций из прогона 1, нанесенных с соответствии с:
(1) Очищенный IgG кролика (50 мкг/мл), используемый при инкубации
(2) Сыворотка кролика (разведение 1:50), используемая при инкубации
(3-4) His6ZQGRep4CT, пленка, инкубированная с IgG кролика
(5-6) His6ZQGRep4CT, пленка, инкубированная с сывороткой кролика
(7-8) Rep4CT, пленка, инкубированная с IgG кролика
(9) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(10-11) Rep4CT, пленка, инкубированная с сывороткой кролика
(12-13) Матрица Protein A Sepharose CL-4B, инкубированная с IgG кролика
(14-15) Матрица Protein A Sepharose CL-4B, инкубированная с сывороткой кролика
(16-17) His6ZQGRep4CT, волокно, инкубированное с IgG кролика.
На фиг. 14 представлен невосстанавливающий гель SDS-PAGE элюированных фракций, полученных как в прогоне 1, так и в прогоне 2, нанесенных в соответствии с:
(1-2) His6ZQGRep4CT, волокно, инкубированное с сывороткой кролика, прогон 1
(3-4) Rep4CT, волокно, инкубированное с IgG кролика, в двух экземплярах, прогон 1
(5-6) Rep4CT, волокно, инкубированное с сывороткой кролика, в двух экземплярах, прогон 1
(7) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(8) Очищенный IgG кролика (50 мкг/мл), использованный при инкубации
(9) Сыворотка кролика (разведение 1:50), использованная при инкубации
(10-11) His6ZQGRep4CT, пленка, инкубированная с IgG кролика, прогон 2
(12-13) His6ZQGRep4CT, пленка, инкубированная с сывороткой кролика, прогон 2
(14-15) Rep4CT, пленка, инкубированная с IgG кролика, прогон 2
(16-17) Rep4CT, пленка, инкубированная с сывороткой кролика, прогон 2.
На фиг. 15 представлен невосстанавливающий гель SDS-PAGE элюированных фракций, все из которых получены в прогоне 2, и нанесенных в соответствии с:
(1-2) Матрица Protein A Sepharose CL-4B, инкубированная с IgG кролика
(3-4) Матрица Protein A Sepharose CL-4B, инкубированная с сывороткой кролика
(5-6) His6ZQGRep4CT, волокно, инкубированное с IgG кролика
(7) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(8-9) His6ZQGRep4CT, волокно, инкубированное с сывороткой кролика
(10-11) Rep4CT, волокно, инкубированное с IgG кролика
(12-13) Rep4CT, волокно, инкубированное с сывороткой кролика.
Результаты на фиг. 13-15 демонстрируют, что матрицы селективно связывают IgG из сыворотки. Способность всех типов матриц His6ZQGRep4CT, т.е. пленок и волокон, связывать IgG находится в том же диапазоне, что и у коммерческой матрицы белка A. Аналогично коммерческой матрице белка A, структуры слитых белков можно регенерировать с сохранением связывающей способности.
Пример 6 - Очистка на месте (CIP) матриц слитого белка и коммерческой матрицы белка A
Для оценки того, остаются ли преципитированные или денатурированные вещества связанными с белковыми структурами, изготовленными из His6ZQGRep4CT (SEQ ID NO: 14) и Rep4CT, и с коммерческой матрицей белка A (Protein A Sepharose CL-4B, GE Healthcare) после элюирования, проводили очистку на месте (CIP) с 8 M мочевиной для всех матриц, подвергнутых воздействию сыворотки кролика, использованной в экспериментах согласно примерам 4-5.
Волокна и пленки, изготовленные из His6ZQGRep4CT и Rep4CT из примера 4 и примера 5, и коммерческую матрицу белка A примера 5, все из которых ранее подвергали воздействию сыворотки кролика, погружали в 200 мкл 8 M мочевины при комнатной температуре в течение 20 мин перед удалением супернатанта и последующим анализом фракций мочевины на SDS-PAGE в невосстанавливающих условиях (фиг. 16-17).
На фиг. 16 представлен невосстанавливающий гель SDS-PAGE после очистки на месте с помощью 8 M мочевины матриц, дважды подвергнутых воздействию сыворотки кролика. На гель проводили нанесение в соответствии с:
(1) Сыворотка кролика (разведение 1:50)
(2) Пустая лунка
(3-5) His6ZQGRep4CT, T-пленка, из примера 4
(6-8) His6ZQGRep4CT, A-пленка, из примера 4
(9) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(10-12) His6ZQGRep4CT, волокно, из примера 4
(13-15) Rep4CT, A-пленка, из примера 4
(16) Rep4CT, волокно, из примера 4
(17) Пустая лунка.
На фиг. 17 представлен второй невосстанавливающий гель SDS-PAGE после очистки на месте с помощью 8 M мочевины матриц, дважды подвергнутых воздействию сыворотки кролика. Нанесение на гель проводили в соответствии с:
(1) Сыворотка кролика (разведение 1:50)
(2) Пустая лунка
(3-4) Rep4CT, волокно, из примера 4
(5-6) His6ZQGRep4CT, пленка, из примера 5
(7) Rep4CT, пленка, из примера 5
(8) Маркер молекулярной массы белка Spectra Multicolor Broad Range Protein Ladder, Fermentas
(9) Rep4CT, пленка, из примера 5
(10-11) His6ZQGRep4CT, волокно, из примера 5
(12-13) Rep4CT, волокно, из примера 5
(14-15) Матрица Protein A Sepharose CL-4B, из примера 5
(16-17) Пустые лунки.
Результаты на фиг. 16-17 указывают на то, что только низкие количества преципитированных или денатурированных веществ остаются связанными со слитыми белковыми структурами после элюирования и очистки. В частности, только низкие количества преципитированных или денатурированных веществ, в том же диапазоне, что и для коммерческой матрицы белка A, остаются связанными с пленками слитого белка His6ZQGRep4CT.
Пример 7 - Клонирование, экспрессия и образование волокон связывающего альбумин слитого белка
Чтобы далее подтвердить идею слитого белка, Rep4CT получали в слитой конструкции со связывающим альбумин доменом (Abd) из белка G стрептококков. Abd представляет собой трехспиральный мотив массой 5 кДа, который связывает альбумин. Для этого клонировали слитый белок, состоящий из домена Abd, находящегося на N-конце Rep4CT (обозначаемый как His6AbdQGRep4CT) (SEQ ID NO: 16-17).
Клонирование
Конструировали праймеры для получения ПЦР-фрагментов Abd из вектора, содержащего такую последовательность. Также праймеры содержали участок расщепления протеазой 3C, обозначаемый QG, между Abd и Rep4CT. Затем полученные продукты ПЦР обрабатывали эндонуклеазами рестрикции NdeI и EcoRI, как и заданный вектор, обозначаемый pT7His6TrxHis6QGRep4CT (содержащий ген устойчивости к канамицину). При рестрикционном расщеплении заданного вектора часть TrxHis6QG отщеплялась. Отщепленные ПЦР-фрагменты и заданный вектор связывали с помощью ДНК-лигазы T4, после чего полученный правильно лигированный вектор (pT7His6AbdQGRep4CT) трансформировали в химически компетентные клетки E. coli BL21 (DE3), которым позволяли расти на чашках с агаром, дополненных канамицином. После этого колонии отбирали и подвергали скринингу ПЦР в отношении правильной вставки, а затем также секвенировали.
Продуцирование
Клетки E. coli BL21 (DE3), имеющие вектор pT7His6AbdQGRep4CT, выращивали в среде Луриа-Бертани (всего 6 литров), дополненной канамицином до величины OD600 1-1,5 при 30°C, с последующей индукцией экспрессии His6AbdQGRep4CT с помощью IPTG и дальнейшей инкубацией при 20°C в течение приблизительно 2 ч. Далее клетки собирали центрифугированием и полученный клеточный осадок растворяли в 20 мМ Tris (pH 8,0).
Очистка
Клеточные осадки, растворенные в 20 мМ Tris (pH 8,0), дополняли лизоцимом и ДНКазой I для лизиса бактериальных клеток, после чего супернатанты выделяли после центрифугирования при 15000 об./мин. Далее выделенные супернатанты наносили на колонку Ni IMAC или на колонку Chelating Sepharose Fast Flow ZN, поддерживая белок His6AbdQGRep4CT связанным с матрицей колонки через His6-метку. После промывания связанные белки элюировали 20 мМ Tris/300 мМ имидазолом (pH 8,0). Объединенные фракции элюата, содержавшие His6AbdQGRep4CT (SEQ ID NO: 16), подвергали диализу против 5 литров 20 мМ Tris (pH 8,0), концентрировали и получали конечное количество 4 мг белка.
Образование волокна и пленки
Из очищенного растворимого белка Abd-Rep4CT, как волокна, так и пленки, успешно получали при концентрации белка 0,87 мг/мл.
Тот факт, что макроскопические волокна и пленки Abd-Rep4CT получали, несмотря на то, что Rep4CT был слит с другим белком, а именно связывающим альбумин доменом (Abd), демонстрирует, что Rep4CT сохраняет его свойства образования волокна, несмотря на то, что он слит с Abd-доменом. Далее, целью было выявить, сохранял ли Abd-домен его способность связывать альбумин, когда он был слит с Rep4CT, см. примеры 24 и 25.
Пример 8 - Клонирование, экспрессия и образование волокон связывающего IgG слитого белка
Чтобы далее подтвердить идею слитого белка, Rep4CT получали в слитой конструкции с связывающим IgG C2-доменом из белка G стрептококков. C2 содержит 55 аминокислот и его структуру составляют две β-шпильки, которые ассоциированы с образованием четырехцепочечного смешанного антипараллельного/параллельного β-слоя с единичной α-спиралью, лежащей поперек одной поверхности слоя. Для этого клонировали слитый белок, состоящий из C2-домена, находящегося на N-конце Rep4CT (обозначаемый как His6C2QGRep4CT) (SEQ ID NO: 18-19).
Клонирование
Конструировали праймеры для получения ПЦР-фрагментов C2 из вектора, содержащего такую последовательность. Также праймеры содержали участок расщепления протеазой 3C, обозначаемый QG, между С2 и Rep4CT. Затем полученные продукты ПЦР обрабатывали эндонуклеазами рестрикции NdeI и EcoRI, как и заданный вектор, обозначаемый pT7His6TrxHis6QGRep4CT (содержащий ген устойчивости к канамицину). При рестрикционном расщеплении заданного вектора часть TrxHis6QG отщеплялась. Отщепленные ПЦР-фрагменты и заданный вектор связывали с помощью ДНК-лигазы T4, после чего полученный правильно лигированный вектор (pT7His6C2QGRep4CT) трансформировали в химически компетентные клетки E. coli BL21 (DE3), которым позволяли расти на чашках с агаром, дополненных канамицином. После этого колонии отбирали и подвергали скринингу ПЦР в отношении правильной вставки, а затем также секвенировали.
Продуцирование
Клетки E. coli BL21 (DE3), имеющие вектор pT7His6C2QGRep4CT, выращивали в среде Луриа-Бертани (всего 6 литров), дополненной канамицином до величины OD600 1-1,5 при 30°C, с последующей индукцией экспрессии His6C2QGRep4CT с помощью IPTG и дальнейшей инкубацией при 20°C в течение приблизительно 2 ч. Далее клетки собирали центрифугированием и полученный клеточный осадок растворяли в 20 мМ Tris (pH 8,0).
Очистка
Клеточные осадки, растворенные в 20 мМ Tris (pH 8,0), дополняли лизоцимом и ДНКазой I для полного лизиса бактериальных клеток, после чего супернатанты выделяли после центрифугирования при 15000 об./мин. Далее выделенные супернатанты наносили на колонку Ni IMAC, поддерживая белок His6C2QGRep4CT связанным с матрицей колонки через His6-метку. После промывания связанные белки элюировали 20 мМ Tris/300 мМ имидазолом (pH 8,0). Объединенные фракции элюата, содержавшие His6C2QGRep4CT, подвергали диализу против 5 литров 20 мМ Tris (pH 8,0), концентрировали и получали конечное количество 6 мг белка.
Образование волокна и пленки
Из очищенного растворимого белка C2-Rep4CT, как волокна, так и пленки, успешно получали при концентрации белка 0,87 мг/мл. Тот факт, что макроскопические волокна и пленки C2-Rep4CT получали, несмотря на то, что Rep4CT был слит с другим белком, а именно связывающим IgG C2-доменом, демонстрирует, что Rep4CT сохраняет его свойства образования волокна, несмотря на то, что он слит с С2-доменом. Далее, целью было выявить, сохранял ли С2-домен его способность связывать IgG, когда он был слит с Rep4CT, см. примеры 26 и 27.
Пример 9 - Клонирование, экспрессия и образование пленок и волокон связывающего биотин слитого белка
Стрептавидин представляет собой тетрамер четырех идентичных мономеров с одним участком связывания на мономер. Он демонстрируют высокую аффинность к биотину (витамин H), достигая константы диссоциации Kd~10-15 M, что делает связывание по существу необратимым событием. Стрептавидин также демонстрирует высокую стабильность в присутствии протеаз, при повышенных температурах и в денатурирующих агентах, и при крайних величинах pH (Wilchek, M. et al., Anal. Biochem. 171: 1-32 (1988)). Таким образом, это взаимодействие является привлекательным при многих применениях, включая мечение, разделение и нацеливание белка. На практике биотинилирование в настоящее время легко облегчать с использованием биотинилированных линкерных молекул, которые также содержат одну из нескольких реакционноспособных органических молекул, которые атакуют и связывают биотин с различными биологическими молекулами, например, белками и ДНК. Однако было показано, что получения высокоплотных функциональных поверхностей, покрытых стрептавидином, трудно достигнуть, см., например, таблицу 4. Для снижения прочности связывания в применениях, где является необходимой обратимость связывания (например в ходе очистки) и для того, чтобы иметь возможность успешно экспрессировать растворимый белок в E. coli, был разработан мономерный вариант стрептавидина, M4 (Wu. S.-C. et al., Protein Expres. Purif. 46, 268-273 (2006)). По сравнению с мономерами тетрамерного стрептавидина дикого типа, M4 имеет четыре аминокислотных замены (V55T, T76R, L109T и V125R), которые поддерживают M4 в активной мономерной форме.
M4 подвергали слиянию на N- или C-конце с Rep4CT (SEQ ID NO: 20-21) рекомбинантными способами. Полученные белки и гены, кодирующие их, были названы M4Rep4CT (SEQ ID NO: 22-23), modM4Rep4CT (SEQ ID NO: 24-25) и Rep4CTM4 (SEQ ID NO: 26-27), соответственно. Различие между M4Rep4CT и modM4Rep4CT состоит и замене Gly на Arg-Ala-Arg в линкерной области между M4 и Rep4CT. Все белки экспрессировали слитыми с меткой His6-Trx-His6, которую отщепляли и удаляли в ходе очистки.
Продуцирование и очистку всех белков проводили, по существу как описано в Stark, M. et al., Biomacromolecules 8, 1695-1701 (2007) и Hedhammar M. et al., Biochemistry 47, 3407-3417 (2008). Концентрацию белка Rep4CTM4, M4Rep4CT и modM4Rep4CT измеряли при 280 нм с использованием коэффициента молярной экстинкции 53860 M-1 см-1. Очищенный образец белка подвергали восстанавливающему SDS-PAGE, и чистоту белка определяли после окрашивания геля кумасси бриллиантовым синим R-250. Теоретические молекулярные массы всех из очищенных белков и другие имеющие значение физические свойства приведены в таблице 3.
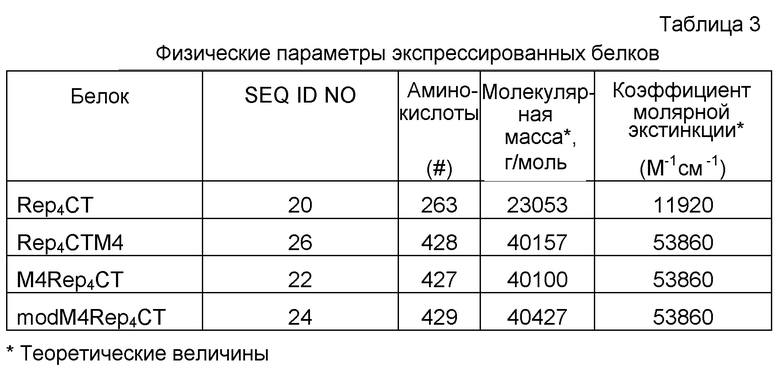
Из каждого из слитых белков Rep4CTM4, M4Rep4CT и modM4Rep4CT, формировались как пленки, так и волокна. Эти результаты подтверждают, что Rep4CT сохраняет его способность к самосборке в твердые структуры, несмотря на то, что он слит с M4. Это также подтверждает, что возможно получить волокна и пленки белков, где M4 слит с Rep4CT, с использованием линкеров различной длины.
Пример 10 - Связывание содержащей биотин мишени с волокнами и пленками из слитого белка
(A) Связывание биотинилированного Atto-565 с пленками Rep4CTM4.
Rep4CTM4 (SEQ ID NO: 26) и Rep4CT (SEQ ID NO: 20, контроль) позволяли образовывать пленки путем высушивания 25 мкл раствора белка при комнатной температуре на дне лунок в прозрачных или черных 96-луночных микропланшетах для титрования. Планшеты хранили при комнатной температуре в течение от одной до двух недель перед применением. Лунки инкубировали с 100 мкл 1% BSA в PBS (pH 7,4) в течение >1 часа при комнатной температуре для избежания неспецифического связывания. Фоновые величины получали путем измерения интенсивности флуоресценции в лунках с пленками из соответствующего белка, Rep4CT или Rep4CTM4, в 50 мкл фосфатно-солевого буфера (PBS) перед добавлением биотинилированного Atto-565. Лунки далее инкубировали с 50 мкл 80 мкМ раствора биотинилированного Atto-565 (Sigma Aldrich, Германия), растворенного в 1% BSA в PBS (pH 7,4). Смеси позволяли стоять при комнатной температуре от двух до трех часов перед промыванием два раза PBS, содержащим 0,05% Tween-20 (PBS-T) и один раз PBS. Полученную интенсивность флуоресценции регистрировали после добавления в лунки 50 мкл PBS в устройстве для считывания микропланшетов Tecan Infinite M200 (λex=565 нм, λem=590 нм).
Приготавливали серии разведений в PBS, и 50 мкл образцов с различными концентрациями биотинилированного Atto-565 добавляли в лунки с соответствующими пленками, в трех экземплярах, после чего регистрировали полученную интенсивность флуоресценции в лунках.
На фиг. 18 представлена интенсивность флуоресценции белковых пленок после смачивания биотинилированным Atto-565 и промывания, которую наблюдали при увеличении 2× с использованием флуоресцентного микроскопа Nicon Eclipse Ti-S (λex=509-550 нм и λem=570-614 нм). Различие в интенсивности флуоресценции между пленками Rep4CTM4 (панель A) и пленками Rep4CT (панель B) после добавления биотинилированного Atto-565 является очевидным.
Для установления общего количества биотинилированного Atto-565, связавшегося с пленками Rep4CTM4, регистрировали интенсивность флуоресценции с использованием серий разведений известных количеств биотинилированного Atto-565. Таким путем, интенсивность флуоресценции образцов с известными количествами (в моль) биотинилированного Atto-565 (в трех экземплярах, в лунках с Rep4CTM4) использовали для получения стандартной кривой. Полученные величины интенсивности флуоресценции, соответствующие фоновой величине (пленки Rep4CTM4 без биотинилированного Atto-565) и данные, коррелирующие с тремя концентрациями биотинилированного Atto-565, добавленного в лунки, представлены на графике на фиг. 19. В таблице ниже графика на фиг. 19 показаны величины, полученные в результате линейной регрессии величин интенсивности флуоресценции со стандартными отклонениями в измерениях при n=3 лунки с одной и той же концентрацией биотинилированного Atto-565.
Начиная с величин интенсивности флуоресценции, полученных в экспериментах по связыванию биотинилированного Atto-565 с пленками Rep4CTM4 (фиг. 20), эти величины использовали для вычисления количества молей биотинилированного Atto-565, которое соответствует полученной интенсивности флуоресценции. Полученные величины флуоресценции нанесены на график на фиг. 20, причем на панелях A и B представлены величины до (-) и после (+) добавления биотинилированного Atto-565 в лунки с пленками Rep4CT (A) и Rep4CTM4 (B). На панели C представлено сравнение полученной интенсивности флуоресценции после добавления биотинилированного Atto-565 к пленкам Rep4CT или Rep4CTM4. Не было значимых отличий (ns) между величинами до и после добавления для пленок Rep4CT (панель A). Значимые отличия величин до и после добавления биотинилированного Atto-565 для Rep4CTM4 (панель B; P<0,01), и между белками (панель C; P<0,0001), были подтверждены с помощью статистических тестов. Столбики на фиг. 20 указывают на стандартное отклонение между величинами интенсивности флуоресценции для n=10 пленок.
Вычисляли соотношение биотинилированный флуорофор/площадь поверхности из экспериментов по связыванию. На площади поверхности приблизительно 28 мм2, с пленками Rep4CTM4 связывалось 2,1 пмоль биотинилированного Atto-565. Это дает соотношение связанный биотин/площадь поверхности 0,073 пмоль/мм2 (таблица 5).
(B) Связывание биотинилированной пероксидазы хрена (HRP) с пленками Rep4CTM4
Вследствие относительно высокой стабильности и продуцирования хромогенных продуктов при преобразовании нехромогенного субстрата и пероксида, часто используют HRP, связанную с вторичным антителом или связывающей молекулой (например, биотин) в применениях, таких как ELISA, вестерн-блоттинг и иммуногистохимия. Для установления общего количества биотинилированной HRP, связывающегося с пленками, изготовленными из Rep4CTM4 (SEQ ID NO: 26) и Rep4CT (SEQ ID NO: 20; контроль), биотинилированной HRP позволяли связываться с каждой соответствующей пленкой. Скорость образования продукта регистрировали при 570 нм с использованием известных количеств субстрата. Коэффициент молярной экстинкции для продукта резоруфина получали от производителя (Invitrogen), и утверждалось, что он составляет 54000 см-1M-1.
Белковым растворам (25 мкл) Rep4CTM4 и Rep4CT позволяли полностью высохнуть на дне лунок в прозрачных 96-луночных микропланшетах для титрования, таким образом, формируя пленки. Лунки инкубировали с 100 мкл 1% BSA в PBS (pH 7,4) в течение >1 часа перед инкубацией с 50 мкл 0,3 мг/мл биотинилированной HRP (Invitrogen, Camarillo, CA) в 1% BSA в PBS (pH 7,4) в течение >1 часа. Затем лунки промывали два раза PBS-T и один раз PBS. Реакции начинали добавлением 50 мкл 50 мкМ раствора Amplex red (Invitrogen) с 2 мМ пероксидом водорода, растворенным в 0,2% BSA, 28 мМ NaCl, 0,54 мМ KCl, 0,3 мМ KH2PO4, 42 мМ Na2HPO4 (pH 7,4) при комнатной температуре. Измерения кинетики проводили на устройстве для считывания микропланшетов Tecan Infinite M200.
Для установления того, какое количество HRP приводило к той же скорости образования продукта, как при измерениях с биотинилированной HRP на пленках, использовали известные количества биотинилированной HRP (свободная в растворе). Полученные скорости реакции 50 мкМ Amplex red и 2 мМ пероксида водорода при катализе биотинилированной HRP, свободной в растворе (pH 7,4), до образования продукта, резоруфина, представлены на графике на фиг. 21. Данные коррелируют с измерениями в трех экземплярах в одной и той же концентрации биотинилированной HRP. В таблице ниже графика на фиг. 21 представлена величина, полученная путем линейной регрессии данных. Столбики на графике на фиг. 21 указывают на стандартные отклонений для измерений в n=3 лунки с одной и той же концентрацией биотинилированной HRP. Полученную стандартную кривую и наклон на фиг. 21 использовали для вычисления количества биотинилированной HRP, которая была связана с пленками слитого белка.
Скорости реакции 50 мкМ Amplex red и 2 мМ H2O2 при катализе в лунках, где пленки слитого белка и контроля инкубировали с биотинилированной HRP, представлены на фиг. 22. Скорости реакции на фиг. 21 и 22 выражены в качестве образования резоруфина в мкМ в минуту, вычисленного с использованием коэффициента экстинкции, предоставленного Invitrogen (54000 см-1M-1). Столбики указывают на стандартное отклонение для реакций, измеренных в n=8 лунки. На фиг. 22 представлено значимое отличие в скорости образования продукта (и, таким образом, также связанной биотинилированной HRP) между лунками, покрытыми Rep4CTM4, и контролями - покрытыми Rep4CT лунками. Было определено, что 0,2 пмоль HRP/мм2 связывалось с пленками Rep4CTM4, (таблица 5).
(C) Сравнение с коммерческими продуктами
Было показано, что трудно достигнуть получения высокоплотных функциональных поверхностей, покрытых стрептавидином. Способность связывания биотина для коммерчески доступных планшетов приведена в таблице 4.
Соотношение биотинилированный флуорофор/площадь поверхности, полученное для экспериментов связывания с биотинилированным Atto-565 в примере 10A и биотинилированной HRP в примере 10B, обобщенно представлено в таблице 5. Плотность двух различных биотинилированных молекул на пленках Rep4CTM4 (SEQ ID NO: 26) составляет 0,073-0,2 пмоль/мм2.
Способность связывания биотина пленок слитого белка M4
площадь поверхностси (пмоль/мм2)
Результаты в таблицах 4-5 указывают на то, что пленки, изготовленные из слитых белков с частью M4, обеспечивают плотность связывания биотина в тех же диапазонах, что и для коммерческих альтернатив, независимо от того, связан ли биотин с небольшой (флуорофор) или крупной (белок) молекулой.
Статистика
Для статистического анализа данных использовали GraphPad Prism 4.0 (GraphPad Software, San Diego, CA). В примере 10A непараметрический парный критерий Вилкоксона использовали при сравнении величин флуоресценции до и после добавления биотинилированного Atto-565 к пленкам, образованным либо из Rep4CT, либо из Rep4CTM4. Кроме того, непараметрический непарный U-критерий Манна-Уитни использовали для сравнения интенсивности флуоресценции в лунках после инкубации биотинилированного Atto-565 с использованием пленки Rep4CT или Rep4CTM4, и также для сравнения величин [резоруфин]/мин, полученных при измерениях на пленках, инкубированных с биотинилированной HRP, в примере 10B. Значимыми считали значения P<0,05.
Пример 11 - Связывание биотинилированного антитела и вторичного антитела с волокнами и пленками из слитого белка
Пленку и волокна Rep4CT (SEQ ID NO: 20, контроль) и modM4Rep4CT (SEQ ID NO: 24) исследуют в отношении способности связываться с биотинилированным антителом, происходящего из кролика. Пленки формируют в стрипы из 8× или 12×1 лунок (лунки равного размера с форматами 96-луночных микропланшетов).
После предварительной инкубации пленок и волокон с 1% BSA в PBS (pH 7,4), биотинилированное антитело (кролик) инкубируют с белковыми структурами (пленка/волокно) в течение >1 ч, с последующим добавлением вторичного антитела против антител кролика, радиоактивно меченого 125I. Пленки и волокна промывают. Детекцию гамма-излучения проводят на индивидуальных пленках или индивидуальных волокнах в гамма-счетчике. Приготавливают серии разведений известных количеств меченного 125I антитела и измеряют излучение с получением стандартной кривой, из которой можно вычислить количество биотинилированного антитела, связанного с пленками и волокнами из слитого белка.
Пример 12 - Получение чистой пленки из слитого белка
Пленки формировали с использованием 25 мкл RepCT4M4 (SEQ ID NO: 26) в концентрации 10-20 мкМ. 100 мкл PBS (pH 7,4) позволяли инкубироваться в лунках с пленкой в течение одного часа. Этот раствор удаляли и добавляли 50 мкл гексафторизопропанола (HFIP) для разрушения пленки и растворения белка. К дополнительным пленкам добавляли то же количество HFIP без предшествующего промывания PBS. HFIP позволяли воздействовать на четыре пленки в течение 3,5 ч, и полученный прозрачный раствор HFIP переносили в пробирки Eppendorf, содержавшие 50 мкл 20% SDS. Воду в пробирках выпаривали и оставшееся содержимое растворяли в 60 мкл 10 мМ Tris-HCl (pH 8,0) и подвергали восстанавливающему SDS-PAGE.
Результаты представлены на фиг. 23. Дорожка 1 соответствует белкам маркера молекулярной массы белка SpectraTM Multicolor Broad Range Protein Ladder (Fermentas). Числа соответствуют размерам белков. Дорожка 2 соответствует очищенным пленкам Rep4CTM4, которые не смачивали PBS перед добавлением HFIP. Дорожка 3 соответствует пленке Rep4CTM4, которую смачивали PBS в течение одного часа перед удалением этого раствора и после добавления HFIP.
Хотя некоторые белки совместно очищаются с белком Rep4CTM4 (дорожка 2, фиг. 23), инкубация с 100 мкл PBS в течение одного часа растворяет контаминирующие белки и остается только пленка, состоящая из белков Rep4CTM4, судя по восстанавливающему SDS-PAGE (дорожка 3, фиг. 23). Таким образом, большинство контаминирующих белков удаляются по существу с помощью процесса полимеризации, и любые остающиеся примеси в белковых структурах, образованных из слитых белков, можно легко удалять путем осторожного промывания водными буферами.
Пример 13 - Улавливание органических мишеней в растворе
Раствор белка ZRep4CT (~1 мг/мл) добавляют к образцу, состоящему из сыворотки, посредством чего молекулам IgG в образце позволяют связываться с частью Z слитого белка в растворе. Смесь помещают на поверхность контакта гидрофобная/гидрофильная среда, которая приводит к тому, что часть Rep4CT в комплексе слитый белок/IgG образует пленку или пену, оставляя другие сывороточные белки в растворе и улавливая IgG на твердой структуре.
Альтернативно, смеси позволяют высохнуть на твердой подложке, формируя пленку с иммобилизованным IgG. Эту пленку промывают буфером, например PBS, для растворения и удаления загрязняющих белков.
Альтернативно, волокна образуют путем предоставления поверхности контакта гидрофобная-гидрофильная среда и подвергая смесь сдвиговым усилиям. Часть Rep4CT комплекса слитый белок/IgG полимеризуется в макроскопические волокна, оставляя другие белки в растворе.
Сформированные твердые структуры (пленки, пены или волокна) можно собирать и IgG можно выделять в подходящем буфере для элюирования (например, путем снижения pH до 2,7). Элюированные белки идентифицируют на SDS-PAGE. Также см. пример 22.
Пример 14 - Каркасы из слитых белков для улавливания клеток
(A) Исследования in vivo не прикрепляющихся клеток в передней камере глаза
Волокнам/пленкам/пенам ZRep4CT позволяют инкубироваться с IgG против CD45 или CD34. Лейкоциты улавливают на каркасах ZRep4CT с IgG против CD45 и тучные клетки улавливают на каркасах ZRep4CT с IgG против CD34. Клеткам позволяют иммобилизоваться на каркасе. Клетки и каркасы из слитого белка трансплантируют в переднюю камеру глаза голых мышей. Клетки исследуют in vivo через глазное окно.
(B) Исследования in vitro не прикрепляющихся клеток
Тучные клетки и лейкоциты выращивают на каркасах ZRep4CT с IgG против CD34 и CD45, соответственно, а затем подвергают мониторингу in vitro. Некоторые клетки являются не прикрепляющимися в фазе развития, например, нервные стволовые клетки, которые могут расти кластерами. В кластерах нервных клеток клетки остаются в недифференцированной форме. Нервные стволовые клетки выращивают кластерами на каркасах ZRep4CT с IgG против клеточного рецептора на нервных стволовых клетках.
(C) Выбор конкретных клеток
Каркас ZRep4CT (пленка/пена/волокно) используют для выбора клеток с помощью специфических антител. Тучные клетки выбирают в двухстадийном процессе. Стадия 1: Связывание клеток с каркасами ZRep4CT с IgG против CD34. Стадия 2: Селекция тучных клеток на каркасах ZRep4CT с IgG против рецептора C-kit.
(D) Продуцирование белка в эукариотических клетках, выращенных на ZRep4CT
Многие клетки, используемые для продуцирования белка, происходят из неприкрепляющихся клеточных линий. Однако крупномасштабное продуцирование значительно облегчается, когда используют прикрепляющиеся клетки, поскольку упрощается физическое отделение. С использованием каркаса ZRep4CT с IgG против клеточного рецептора, можно осуществлять выращивание не прикрепляющихся клеток прикрепленным образом.
Пример 15 - Исследование влияния обработки мочевиной на связывание IgG с Z-Rep4CT
Оценивали эффект обработки мочевиной пленок и волокон Z-Rep4CT на связывание IgG. Пленки и волокна Z-Rep4CT обрабатывали мочевиной перед связыванием IgG из сыворотки кролика. Связанные IgG элюировали и анализировали с помощью SDS-PAGE.
Волокна и пленки Z-Rep4CT, которые, как показано в примере 6, связывают IgG из сыворотки кролика и которые обрабатывали 8 M мочевиной (200 мкл 8 M мочевины, 20 мин, комнатная температура), инкубировали с 500 мкл сыворотки кролика (разведение 1:5) в течение 1 ч при комнатной температуре. Пленки и волокна Rep4CT (SEQ ID NO: 20) использовали в качестве контрольного материала и обрабатывали аналогичным образом. После промывания три раза 600 мкл PBS, связанный IgG элюировали в 500 мкл путем снижения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не показано).
Исходя из геля, было сделано заключение, что пленки и волокна Z-Rep4CT сохраняют их связывающую способность в отношении IgG после обработки 8 M мочевиной. Контрольные пленки и волокна Rep4CT не показали никакого связывания IgG.
Пример 16 - Исследование влияния обработки NaOH на связывание IgG с Z-Rep4CT
Для дальнейшей оценки длительности воздействия условий очистки, оценивали эффект обработки NaOH пленок и волокон Z-Rep4CT (SEQ ID NO: 14) на связывание IgG. Волокна и пленки Z-Rep4CT из примера 4 и 5, которые, как показано в примере 6, связывают IgG из сыворотки кролика и которые обрабатывали 8 M мочевиной (200 мкл 8 M мочевины, 20 мин, комнатная температура), далее обрабатывали 1 M NaOH (500 мкл 1 M NaOH, 20 мин, комнатная температура). Пленки и волокна Rep4CT (SEQ ID NO: 20) использовали в качестве контрольного материала и обрабатывали аналогичным образом. После обработки NaOH, пленки и волокна инкубировали с 500 мкл сыворотки кролика (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный IgG элюировали в 500 мкл путем снижения pH приблизительно до 2,7 с помощью буфера для элюирования (т.е. 0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не показано).
Исходя из геля, было сделано заключение, что пленки и волокна Z-Rep4CT сохраняют их способность связывать IgG после обработки 1 M NaOH. Контрольные пленки и волокна из Rep4CT не показали никакого связывания IgG.
Пример 17 - Количественное определение связывания IgG-HRP с пленками Z-Rep4CT
(A) Связывание IgG-HRP с пленками Z-Rep4CT
Для количественного определения связывания IgG с Z-Rep4CT, конъюгированный с пероксидазой хрена (HRP) IgG (IgG-HRP) связывали с пленками Z-Rep4CT.
Пленки Z-Rep4CT (SEQ ID NO: 14) и Rep4CT (контроль, SEQ ID NO: 20) при различных концентрациях (0,011-890 пмоль) формировали в 96-луночных планшетах). Пленки блокировали 100 мкл 1% BSA в течение 1 ч при комнатной температуре. Затем пленки инкубировали в течение 1 ч при комнатной температуре в 50 мкл IgG-HRP (т.е. 34 пмоль IgG-HRP, источник IgG - кролик), и пленки промывали два раза 100 мкл 0,05% Tween, с последующим конечным промыванием 100 мкл PBS. Для измерения IgG-HRP, связанного с пленками, 50 мкл смеси 50 мкМ Amplex Red/2 мМ H2O2 добавляли к одной пленке за один раз, а затем проводили мониторинг поглощения при 570 нм в течение трех минут на устройстве для считывания планшетов Tecan Plate Reader. Также проводили измерения серии разведений растворимого IgG-HRP (0,05-0,5 пмоль) в том же типе планшета, что и для пленок, путем блокирования лунок 100 мкл 1% бычьего сывороточного альбумина (BSA) в течение 1 ч перед добавлением 20 мкл растворимого IgG-HRP, 20 мкл смеси 125 мкМ Amplex Red и 10 мкл 9,79 мМ H2O2. Для пленок и серий разведения измерения проводили в трех экземплярах.
Линейную регрессионную аппроксимацию проводили с использованием программного обеспечения Tecan для каждого отдельного измерения, соответствующего линейной области поглощения при 570 нм (Abs570/мин) против исходного графика данных. Наклон соответствует скорости преобразования HRP из бесцветного субстрата в цветной продукт, и он пропорционален количеству связанных молекул IgG-HRP.
Для каждого отдельного исследования в трех экземплярах вычисляли среднее значение и стандартное отклонение для количества связавшегося IgG-HRP (пмоль). На фиг. 24A представлено количество IgG-HRP, связавшегося с пленками Z-Rep4CT и Rep4CT при различных концентрациях белка, и на фиг. 24B представлены доли молекул Z-Rep4CT и Rep4CT в пленках при различных концентрациях белка, которые связывали IgG-HRP.
Для пленок, содержавших 1,1 пмоль белка и более, пленки Z-Rep4CT связывают значительно больше IgG-HRP, чем соответствующие контрольные пленки Rep4CT. Доля молекул Z-Rep4CT в этих пленках, которые связали IgG-HRP, составляет приблизительно ~7% или менее.
(B) Связывание IgG-HRP с пленками Z-Rep4CT после обработки NaOH
Для исследования эффекта обработки NaOH на связывание IgG с пленками Z-Rep4CT, IgG-HRP связывали с пленками, обработанными NaOH, и определяли количество связанного IgG-HRP.
Пленки Z-Rep4CT (SEQ ID NO: 14; 1,1-890 пмоль) и Rep4CT (SEQ ID NO: 20, 108 пмоль) со связанным IgG-HRP из примера 17 (A) инкубировали в 100 мкл буфера для элюирования (pH 2,7) в течение 1 ч при комнатной температуре для удаления связавшегося IgG-HRP. Далее пленки инкубировали в 100 мкл 1 M NaOH в течение 20-30 мин при комнатной температуре, а затем промывали два раза в 150 мкл PBS. Затем лунки, содержавшие пленки, блокировали 1% BSA, инкубировали с IgG-HRP (34 пмоль) и промывали три раза, затем добавляли 50 мкм Amplex Red/2 мМ H2O2, а затем проводили мониторинг поглощения при 570 нм, как указано выше в (A). Для каждого индивидуального измерения в трех экземплярах вычисляли среднее значение и стандартное отклонение для количества связавшегося IgG-HRP (пмоль).
На фиг. 25A представлено количество IgG-HRP, связавшегося с пленками Z-Rep4CT и Rep4CT при различных концентрациях белка, на фиг. 25B представлены доли молекул Z-Rep4CT и Rep4CT в пленках при различных концентрациях белка, которые связывали IgG-HRP. На фиг. 26 представлены визуализированные количества связавшегося IgG-HRP, до и после обработки NaOH, с пленками Z-Rep4CT и Rep4CT.
Пленки с 108 пмоль Z-Rep4CT показали значительно более высокое связывание, чем соответствующие пленки Rep4CT, и величина связывания IgG-HRP с обработанными NaOH пленками Z-Rep4CT показала тенденцию к увеличению по мере увеличению количества белка в пленке. Оказалось, что количество связавшегося IgG-HRP с пленками Z-Rep4CT снижается приблизительно в 2-4 раза при обработке жестким 1 M NaOH по сравнению с необработанными пленками.
Пример 18 - Количественное определение связывания конъюгата IgG-флуорофор с пленками Z-Rep4CT
Проводили связывание IgG, конъюгированного с флуорофором, с пленками Z-Rep4CT и Rep4CT, содержавшими различные количества белка.
Пленки Z-Rep4CT (SEQ ID NO: 14) и Rep4CT (контроль, SEQ ID NO: 20) при различных концентрациях (0,011-890 пмоль) получали и блокировали, как указано в примере 17. Затем пленки инкубировали в течение 1 ч при комнатной температуре в 50 мкл конъюгата IgG-флуорофор (100 пмоль конъюгата IgG-флуорофор, источник IgG - кролик, флуорофор: Alexa Fluor 633), после чего пленки промывали два раза 100 мкл 0,05% Tween, с последующим конечным промыванием 100 мкл PBS. Перед измерением флуоресценции к каждой пленке добавляли 100 мкл PBS.
Серии разведений растворимого конъюгата IgG-флуорофор (0-1 пмоль) также измеряли в планшете того же типа, что и для пленок, путем блокирования лунок 100 мкл 1% бычьего сывороточного альбумина (BSA) в течение 1 ч, перед добавлением 100 мкл растворимого конъюгата IgG-флуорофор. Флуоресценцию измеряли для пленок и серий разведения в трех экземплярах на устройстве для считывания планшетов Tecan Plate Reader (возбуждение: 632 нм, испускание: 660 нм, шаг: 200).
Для каждого индивидуального измерения в трех экземплярах вычисляли среднее значение и стандартное отклонение для количества связавшегося конъюгата IgG-флуорофор (пмоль) (фиг. 27A) для пленок с 0,011-55 пмоль белка. Также вычисляли долю молекул Z-Rep4CT и Rep4CT, связывающих конъюгат IgG-флуорофор (фиг. 27B).
Может быть сделано заключение, что связывание IgG-Alexa Fluor 633 с пленками Z-Rep4CT является значительно большим, чем с соответствующими контрольными пленками Rep4CT. Не было выявлено значимых отличий в связывании конъюгата IgG-флуорофор между пленками с 0,011-11 пмоль Z-Rep4CT, и это, по-видимому, не является следствием аутофлуоресценции пленок Z-Rep4CT при этой длине волны (данные не представлены). Связывание конъюгата IgG-флуорофор с пленками Z-Rep4CT, содержащими более 55 пмоль белка, нельзя было вычислить никаким надежным путем, поскольку его флоресцентные сигналы находились вне калибровочной кривой.
Путем сравнения количеств связавшегося конъюгата IgG-флуорофор с пленками Z-Rep4CT с соответствующим связыванием IgG-HRP согласно примеру 17, Z-Rep4CT, по-видимому, связывает больше конъюгата IgG-флуорофор, чем IgG-HRP (например отличие в ~4 и ~6 раз для пленок с 55 и 1,1 пмоль, соответственно), что может быть следствием различия размера между флуорофором и HRP. Более того, доля молекул Z-Rep4CT, связывающих IgG-флуорофор, также, по-видимому, возросла с долей молекул при связывании IgG-HRP.
Пример 19 - Связывание IgG из плазмы крови человека с пленками Z-Rep4CT
В этом эксперименте получали три пленки Z-Rep4CT (SEQ ID NO: 14), как указано в примере 3. Все пленки хранили, после их формирования, в течение восьми месяцев при +4°C без погружения в какую-либо жидкость в ходе хранения. Каждую ленку Z-Rep4CT инкубировали с 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связавшийся IgG элюировали в 500 мкл путем снижения pH приблизительно до 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не представлено). Пленки из Rep4CT (SEQ ID NO: 20) использовали в качестве контрольного материала и обрабатывали аналогичным образом.
Исходя из геля, очевидно, что в элюированных фракциях пленок Z-Rep4CT появляется IgG (~146 кДа), что указывает на то, что пленки Z-Rep4CT сохраняют способность связывать IgG из плазмы крови человека после хранения в течение восьми месяцев при +4°C без погружения в какую-либо жидкость. Контрольные пленки из Rep4CT не демонстрируют никакого IgG в элюированных фракциях. Эти данные далее подтверждают наблюдение об экспериментальной воспроизводимости, описанное в примере 4, при использовании структур по изобретению, и также демонстрируют, что белковые структуры могут связывать IgG человека.
Пример 20 - Связывание IgG из плазмы крови человека с автоклавированным волокном Z-Rep4CT
Исследовали способность Z-Rep4CT связывать IgG после стерилизации обработкой в автоклаве. После обработки в автоклаве волокна Z-Rep4CT, волокну позволяли связываться с IgG из плазмы крови человека. Связывание с IgG автоклавированного волокна сравнивали со связыванием неавтоклавированного волокна.
Два волокна Z-Rep4CT (SEQ ID NO: 14) приблизительно равного размера переносили в две пробирки, содержавшие 20 мМ Tris (pH 8). Затем одно из волокон автоклавировали в течение 20 мин при 121°C. В качестве контрольного материала использовали два волокна из Rep4CT (SEQ ID NO: 20), их обрабатывали аналогичным образом.
Волокна инкубировали с 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный IgG элюировали в 500 мкл путем снижения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (фиг. 28).
Нанесение на гель, представленный на фиг. 28 проводили в соответствии с:
(1) Плазма крови человека (1:5), нанесение 1,4 мкл
(2) Z-Rep4CT, неавтоклавированное волокно, нанесение 14 мкл
(3) Z-Rep4CT, автоклавированное волокно, нанесение 14 мкл
(4) Rep4CT, неавтоклавированное волокно, нанесение 14 мкл
(5) Rep4CT, автоклавированное волокно, нанесение 14 мкл
(6) Маркер молекулярной массы.
Как неавтоклавированное, так и автоклавированное волокно Z-Rep4CT, продемонстрировало четкую полосу IgG, соответствующую приблизительно 146 кДа. Нет очевидных отличий в интенсивности этих полос IgG, что указывает на то, что обработка в автоклаве имеет небольшой эффект на способность связывать IgG. Ни неавтоклавированные, ни автоклавированные волокна Rep4CT не показали никакого IgG в элюированных фракциях. Все волокна демонстрируют дополнительную, значительно более слабую, полосу альбумина (~50-60 кДа).
Пример 21 - Расщепление волокон Z-Rep4CT протеазой 3C
Белок Z-Rep4CT (полная структура белка представляет собой His6-Z-LEALFQGP-Rep4CT; SEQ ID NO: 14) содержит участок распознавания протеазой 3C (LEALFQGP, расщепление протеазой 3C между аминокислотами Q и G) между Z-доменом и сегментом Rep4CT.
Волокно Z-Rep4CT произвольного размера переносили в пробирку Eppendorf, и расщепление протеазой начинали путем добавления 9,6 мкг протеазы 3C и 0,35 мкл 1 M DTT (дитиотреитол) до общего объема 350 мкл. Расщеплению позволяли протекать в течение 24 ч при +4°C, после чего из супернатанта после расщепления отбирали образец для SDS-PAGE. Затем расщеплению позволяли протекать в течение дополнительных 24 ч (+4°C), когда второй образец для SDS-PAGE отбирали из супернатанта после расщепления. Затем два взятых образца анализировали с помощью SDS-PAGE (фиг. 29).
Нанесение на гель, представленный на фиг. 29, проводили в соответствии с:
(1) маркер молекулярной массы
(2) супернатант волокна Z-Rep4CT, расщепляемого протеазой 3C в течение 24 ч
(3) супернатант волокна Z-Rep4CT, расщепляемого протеазой 3C в течение 48 ч.
Как можно видеть на фиг. 29, оба супернатанта после расщепления волокна Z-Rep4CT протеазой 3C имели две отчетливых полосы. Первая полоса, соответствующая размеру немного менее 35 кДа, соответствует протеазе 3C (~31 кДа), в то время как вторая полоса расположена немного выше положения, соответствующего 10 кДа. Поскольку расщепление Z-Rep4CT протеазой 3C дает два олигопептидных сегмента, соответствующих (i) His6-Z-LEALFQ (~9 кДа) и (ii) GP-Rep4CT (~23 кДа), было сделано заключение, что вторая полоса соответствует отщепленному фрагменту HZ (9 кДа). Таким образом, было сделано заключение, что участок расщепления протеазой 3C доступен для расщепления в волокнах Z-Rep4CT и, таким образом, часть HZ может быть удалена.
Пример 22 - Образование волокна из растворимого Z-Rep4CT в присутствии IgG
При смешении растворимого Z-Rep4CT с IgG, кинетика связывания IgG с Z-доменами должна быть быстрее, чем образование волокон Z-Rep4CT. Исследовали возможность формирования волокон, даже если большинство Z-доменов оккупированы IgG.
Образование волокна в присутствии IgG
Очистку Z-Rep4CT (SEQ ID NO: 14) проводили аналогично тому, как описано выше, и очищенный раствор белка концентрировали до 2,2 мг/мл. Формирование волокна осуществляли в четырех различных условиях, все в общем объеме формирования волокна 3 мл, содержащем 71 нмоль растворимого белка Z-Rep4CT. Первые условия вовлекали только Z-Rep4CT; вторые условия представляли собой Z-Rep4CT, смешанный с очищенным IgG кролика (8-кратный избыток Z-Rep4CT относительно IgG); третьи условия представляли собой Z-Rep4CT, смешанный с сывороткой кролика (избыток IgG сыворотки в ~1,5 по сравнению с Z-Rep4CT); и четвертые условия представляли собой Z-Rep4CT, смешанный с сывороткой кролика (~7-кратный избыток Z-Rep4CT по сравнению с IgG сыворотки). Волокну позволяли образовываться в течение трех суток при комнатной температуре.
После формирования волокна в течение трех суток, волокна образовывались в условиях 1, 2 и 4. В дополнение к сформированному волокну в условиях 4, также образовывалось значительное количество агрегатов белка Z-Rep4CT. Для условий 3 абсолютно не было заметно ни волокна, ни агрегатов.
Одно заключение из этого может состоять в том, что образование волокна нарушается в присутствии больших количеств других биомолекул, как можно видеть в случае условий 3, которые препятствуют взаимодействию отдельных молекул Rep4CT друг с другом. Другим аспектом этого может быть то, что если присутствует слишком много IgG, как можно видеть в случае условий 3, многие из Z-доменов в Z-Rep4CT могут связывать IgG, и большая часть Z-Rep4CT со связанным IgG может препятствовать образованию волокна.
Удаление связанного IgG из волокон Z-Rep4СT
Волокна, полученные в условиях 1, 2 и 4, вместе с агрегатами, полученными в условиях 4, выделяли и промывали в 20 мМ Tris (pH 8). Далее, все волокна и агрегаты разделяли на две равные половины: одна половина для оценки связанного IgG при снижении pH, а другая половина для расщепления протеазой 3C.
Первую группу волокон и агрегатов переносили в пробирки Eppendorf и в каждую пробирку добавляли 144 мкл буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), pH 2,7. Элюированию IgG позволяли протекать в течение 30 мин при комнатной температуре, после чего супернатанты после элюирования выделяли и анализировали с помощью SDS-PAGE (фиг. 30).
К второй группе волокон и агрегатов добавляли 144 мкл протеазы 3C (т.е. 110 мкг протеазы 3C), содержавшей DTT. Расщеплению протеазой 3C позволяли протекать в течение ночи при +4°C, после чего подвергнутые расщеплению супернатанты выделяли и анализировали с помощью SDS-PAGE (фиг. 30).
На фиг. 30 представлен невосстанавливающий гель SDS-PAGE для IgG, удаленного из волокон Z-Rep4CT и агрегатов, образованных путем смешения растворимого Z-Rep4CT с IgG. Нанесение на гель проводили в соответствии с:
(1) Очищенный растворимый Z-Rep4CT (2,2 мг/мл)
(2) Элюирование при низком значении pH волокна Z-Rep4CT (условия 1)
(3) Элюирование при низком значении pH волокна Z-Rep4CT (условия 2)
(4) Элюирование при низком значении pH волокна Z-Rep4CT (условия 4)
(5) Элюирование при низком значении pH агрегатов Z-Rep4CT (условия 4)
(6) Маркер молекулярной массы
(7) Расщепление протеазой 3C волокна Z-Rep4CT (условия 1)
(8) Расщепление протеазой 3C волокна Z-Rep4CT (условия 2)
(9) Расщепление протеазой 3C волокна Z-Rep4CT (условия 4)
(10) Расщепление протеазой 3C агрегатов Z-Rep4CT (условия 4).
[Примечание: молекулярная масса His6Z, протеазы 3C и IgG кролика составляет 9, 30 и ~146 кДа, соответственно.]
Из фиг. 30 очевидно, что IgG выделяется из всех исследованных волокон и агрегатов, независимо от того, в каких условиях они образовывались. Также см. пример 13.
Пример 23 - Улавливание лимфоцитов на Z-Rep4CT с использованием связывания с антителом
Использование матриц Z-Rep4CT, например волокон и пленок, для связывания Fc-части IgG открывает возможность для дальнейшего связывания чего-либо, на что уловленные IgG специфично направлены. Одной из привлекательных мыслей является выделение определенного типа клеток из биологического образца, содержащего множество различных типов клеток, с использованием уловленного IgG на матрице Z-Rep4CT в качестве аффинного лиганда для клеток. Для исследования этого подхода для улавливания клеток, волокнам и пленкам Z-Rep4CT позволяли связывать IgG, которые специфично направлены на молекулу CD3, на клеточной поверхности T-лимфоцитов человека. Уловленные клетки анализировали с помощью флуоресцентной микроскопии.
Экспрессию и очистку белка Z-Rep4CT (SEQ ID NO: 14) проводили, как описано ранее. Очищенный белок концентрировали до ~1 мг/мл, после чего получали волокна и пленки в соответствии с ранее описанными методиками (пленки получали в планшетах для культивирования тканей из 24 ячеек). Кроме того, получали контрольные волокна и пленки Rep4CT (SEQ ID NO: 20) аналогичным путем из раствора белка ~1 мг/мл.
Для улавливания IgG, направленного против молекулы CD3 лимфоцитов человека, на матрицы, волокна и пленки погружали в 150 мкл конъюгированного с флуорофором IgG против CD3 человека (моноклональный IgG2a мыши против антигена CD3 человека, метка: Alexa Fluor 488) в разведении 1:20 в течение 1 ч при комнатной температуре. Волокна и пленки промывали три раза 300 мкл PBS (pH 7,4), после чего их анализировали в отношении связывания IgG с помощью инвертационного флуоресцентного микроскопа Nikon Eclipse Ti (возбуждение при 455-490 нм, детекция при 500-540 нм). Как волокно, так и пленка Z-Rep4CT, связывали конъюгированное с Alexa Fluor 488 IgG-антитело, таким образом, подтверждая способность Z-домена в Z-Rep4CT связывать IgG. Матрицы Z-Rep4CT, которые не подвергали воздействию IgG, не демонстрируют никакого флуоресцентного сигнала в этой определенной области, и контрольные волокна и пленки Rep4CT не демонстрируют никакой флуоресценции, даже если их подвергали воздействию IgG.
Мононуклеарные клетки (т.е. лимфоциты и моноциты) выделяли из свежеполученной периферической крови человека градиентным центрифугированием при комнатной температуре (30 мин, 400 × g) в среде для разделения с градиентом плотности Ficoll-Paque. После центрифугирования выделяли фракцию мононуклеарных клеток, а затем два раза промывали PBS, после чего клетки ресуспендировали в 20 мл среды RPMI/10 %FCS. Истощения моноцитов достигали путем переноса суспензии клеток в колбу для культивирования тканей T-75, а затем инкубировали в течение 90 мин при 37°C. После истощения моноцитов клетки в суспензии выделяли, и было определено общее количество лимфоцитов 11×106.
Для связывания лимфоцитов с матрицами Z-Rep4CT (SEQ ID NO: 14), лимфоциты (1 мл, ~0,37×106 клеток/мл) наносили на волокна и пленки с последующей инкубацией в течение 30 мин при +4°C (при осторожном качании). Далее волокна и пленки промывали три раза 3 мл PBS/2% FCS (pH 7,4). Связанные клетки фиксировали в 2% PFA (параформальдегид) в течение 15 мин при +4°C. Ядра клеток окрашивали путем погружения волокон и пленок со связавшимися клетками в 200 мкл раствора для окрашивания DAPI (1 мкг/мл) на 5 мин при комнатной температуре перед промыванием три раза с помощью 300 мкл PBS. PBS в объеме 300 мкл добавляли к каждому волокну или пленке до анализа флуоресцентной микроскопией с использованием инвертационного устройства Nikon Eclipse Ti (возбуждение при 380-395 нм, детекция при 415-475 нм).
Изображения волокон Z-Rep4CT (не представлены) демонстрируют мало связанных лимфоцитарных клеток в случае волокна, которое не было подвергнуто воздействию IgG-антител против CD3 человека, в то время как оказалось, что волокно, которое было подвергнуто воздействию IgG, имеет больше связанных лимфоцитов. В случае пленок Z-Rep4CT, окрашенные клетки являются отчетливо видимыми в случае пленки, подвергнутой воздействию IgG, но также в случае пленки, не подвергнутой воздействию IgG. Однако, также для пленок оказалось, что пленка, подвергнутая воздействию IgG против CD3 человека перед связыванием клеток, имеет больше связанных клеток, чем соответствующая пленка, не подвергнутая воздействию IgG перед связыванием клеток. Более того, контрольные волокна и пленки Rep4CT (SEQ ID NO: 20) вообще не демонстрируют никакого связывания лимфоцитов.
В этом эксперименте с помощью флуоресцентной микроскопии было показано, что волокна и пленки Z-Rep4CT, в противоположность волокнам и пленкам Rep4CT, способны связывать флуоресцентно меченное IgG-антитело, а именно IgG мыши против CD3 человека. Более того, как волокна, так и пленки Z-Rep4CT способны связывать лимфоциты, независимо от наличия IgG-антитела, специфично распознающего T-лимфоциты человека. Это может подразумевать, что сам по себе Z-домен обладает некоторой аффинностью к лимфоцитам человека. Однако количество связанных клеток с матрицами Z-Rep4CT оказалось немного более высоким, когда они были покрыты IgG перед связыванием клеток. Чтобы было возможно установить, являются ли связанные клетки лимфоцитами T-типа, необходимо было применить второе антитело, также направленное против молекулы CD3 человека, чтобы отличить T-лимфоциты от других типов лимфоцитов (например B-лимфоцитов и NK-клеток).
Пример 24 - Связывание альбумина из плазмы человека с пленками Abd-Rep4CT
Для оценки доступности домена Abd (остатки 13-58 в SEQ ID NO: 16) в пленках и способности пленок Abd-Rep4CT связывать альбумин, использовали плазму крови человека в качестве источника альбумина. Связанный альбумин элюировали и анализировали с помощью SDS-PAGE.
Шесть пленок Abd-Rep4CT (SEQ ID NO: 16), полученных согласно примеру 7, инкубировали с 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный альбумин элюлировали в 500 мкл путем снижения значения pH до приблизительно 2,7, с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl) после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (фиг. 31). Пленки Rep4CT (SEQ ID NO: 20), также полученные согласно примеру 7, использовали в качестве контрольного материала и обрабатывали аналогичным образом.
Нанесение на гель, представленный на фиг. 31 проводили в соответствии с:
(1) Плазма крови человека (1:50), нанесение 0,7 мкл
(2-7) Abd-Rep4CT, пленка, в шести экземплярах, нанесение 14 мкл
(8) Пустая лунка, инкубированная с плазмой крови человека, нанесение 14 мкл
(9) Маркер молекулярной массы
(10-12) Rep4CT, пленка, в трех экземплярах, нанесение 14 мкл.
Все шесть пленок Abd-Rep4CT связывали альбумин из плазмы крови человека (дорожки 2-7). Поскольку только одна полоса альбумина (~60 кДа) появлялась в элюированной фракции этих пленок Abd-Rep4CT, они, по-видимому, не связывали ничего неспецифично из плазмы крови человека. Пленки Rep4CT не демонстрируют никакого альбумина в элюированных фракциях (дорожки 10-12).
Для исследования стабильности пленок вновь исследовали способность пленок Abd-Rep4CT, которые использовали один раз ранее (см. выше), и хранили в течение 29 суток в PBS (+4°C), связывать альбумин. Шесть пленок Abd-Rep4CT инкубировали с 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный альбумин элюировали в 500 мкл путем снижения значения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не показано). Пленки Rep4CT использовали в качестве контрольного материала и обрабатывали аналогичным путем.
Все шесть пленок Abd-Rep4CT сохраняли способность связывать альбумин из плазмы крови человека после хранения в течение 29 суток в PBS. Пленки Rep4CT не продемонстрировали никакого альбумина в элюированных фракциях.
Пример 25 - Очистка пленок Abd-Rep4CT
(A) Обработка пленок Abd-Rep4CT мочевиной
Пленки Abd-Rep4CT (SEQ ID NO: 16) (всего шесть пленок, ранее использованных в примере 24) инкубировали с 500 мкл 8 M мочевины в течение 20 мин при комнатной температуре, после чего их промывали три раза 600 мкл PBS. Далее пленки инкубировали в 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный альбумин элюировали в 500 мкл путем снижения значения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не показано). Все шесть пленок Abd-Rep4CT, тем не менее, могут связывать альбумин из плазмы крови человека после обработки 8 M мочевиной.
(B) Обработка волокон и пленок Abd-Rep4CT NaOH
Волокно Abd-Rep4CT (SEQ ID NO: 16) сначала инкубировали в 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный альбумин элюировали в 500 мкл путем снижения значения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (не показано). Ту же методику выполняли для контрольного волокна Rep4CT (SEQ ID NO: 20).
Для обработки NaOH использовали три набора пленок Abd-Rep4CT (SEQ ID NO: 16) в трех экземплярах: (i) пленки, предварительно обработанные 8 M мочевиной (см. (A) выше), которые обрабатывают 1 M NaOH, с последующим связыванием альбумина; (ii) ранее неиспользованные пленки, которые обрабатывают 1 M NaOH, с последующим связыванием альбумина; и (iii) ранее неиспользованные пленки, которые анализировали только в отношении связывания альбумина. Волокна Abd-Rep4CT, использованные, как описано выше, для связывания альбумина, обрабатывают 1 M NaOH, с последующим повторным связыванием альбумина.
Для обработки NaOH волокно и пленки Abd-Rep4CT [наборы пленок (i) и (ii)] инкубировали с 500 мкл 1 M NaOH в течение ~20 мин при комнатной температуре, после чего их три раза промывали с помощью 600 мкл PBS. Далее волокно и все три набора пленок инкубировали в 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный альбумин элюировали в 500 мкл путем снижения значения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (фиг. 32).
Нанесение на гель, представленный на фиг. 32, проводили в соответствии с:
(1) Плазма крови человека (1:5), нанесение 1,4 мкл
(2-3, 5) Пленка Abd-Rep4CT в трех экземплярах, обработанная 8 M мочевиной и 1 M NaOH перед связыванием альбумина, нанесение 14 мкл
(4) Маркер молекулярной массы
(6-8) Пленка Abd-Rep4CT в трех экземплярах, необработанная перед связыванием альбумина, нанесение 14 мкл
(9-11) Пленка Abd-Rep4CT в трех экземплярах, обработанная 1 M NaOH перед связыванием альбумина, нанесение 14 мкл
(12) Волокно Abd-Rep4CT, необработанное перед связыванием альбумина, нанесение 14 мкл
(13) Волокно Abd-Rep4CT, обработанное 1 M NaOH перед связыванием альбумина, нанесение 14 мкл
(14) Волокно Rep4CT, необработанное перед связыванием альбумина, нанесение 14 мкл.
Волокно Abd-Rep4CT, очевидно, связывает альбумин (~60 кДа), до и после обработки 1 M NaOH (дорожка 12 и 13, соответственно), в то время как соответствующее необработанное волокно Rep4CT не демонстрирует никакого связывания альбумина (дорожка 14). Все пленки Abd-Rep4CT демонстрируют связывание альбумина без очевидного различия в интенсивности полос между необработанными пленками и пленками, обработанными 1 M NaOH перед связыванием альбумина (дорожки 6-8 и 9-11, соответственно). Однако пленки, обработанные как 8 M мочевиной, так и 1 M NaOH перед связыванием альбумина, демонстрируют снижение интенсивности полос элюированного альбумина (дорожки 2, 3 и 5) по сравнению с другими двумя наборами пленок.
Пример 26 - Связывание IgG кролика и мыши с пленками и волокнами C2-Rep4CT
Доступность C2-домена (остатки 13-67 в SEQ ID NO: 18) в волокнах и пленках C2-Rep4CT анализировали следующим образом. Две пленки и одно волокно C2-Rep4CT (SEQ ID NO: 18) инкубировали с 500 мкл сыворотки кролика (разведение 1:5), в то время как другие две пленки и одно волокно C2-Rep4CT (SEQ ID NO: 18) инкубировали с 500 мкл IgG1 мыши (моноклональные иммуноглобулины против антител кролика, изотип IgG1 мыши, асцитная жидкость мышей) в концентрации ~50 мкг/мл в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный IgG элюировали в 500 мкл путем снижения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (фиг. 33). Пленки и волокна Rep4CT (SEQ ID NO: 20) использовали в качестве контрольного материала и обрабатывали аналогичным путем.
Нанесение на гель, представленный на фиг. 33, проводили в соответствии с:
(1) Асцитная жидкость мыши (изотип IgG1)
(2) Сыворотка кролика (1:50)
(3-4) C2-Rep4CT в двух экземплярах, пленка, IgG1 мыши
(5,7) C2-Rep4CT в двух экземплярах, пленка, сыворотка кролика
(6) Маркер молекулярной массы
(8) C2-Rep4CT, волокно, IgG1 мыши
(9) C2-Rep4CT, волокно, сыворотка кролика
(10-11) Rep4CT в двух экземплярах, пленка, IgG1 мыши
(12) Rep4CT, пленка, сыворотка кролика
(13) Rep4CT, волокно, сыворотка кролика
(14) Rep4CT, волокно, IgG1 мыши
[Примечание: IgG кролика имеет массу ~146 кДа и IgG мыши имеет массу ~160 кДа в невосстанавливающих условиях SDS-PAGE.]
Связывание IgG1 из асцитной жидкости с пленками C2-Rep4CT не обеспечивало никакой поддающейся детекции полосы IgG в элюированных фракциях при SDS-PAGE (дорожки 3-4), однако волокно C2-Rep4CT, как оказалось, связывает IgG1 мыши (дорожка 8, IgG мыши имеет массу ~160 кДа). Однако как пленки (дорожки 5 и 7), так и волокно (дорожка 9) C2-Rep4CT продемонстрировали связывание IgG из сыворотки кролика. Поскольку источник IgG1 мыши в данном случае находится в форме асцитной жидкости, возможно, что что-либо из этой жидкости каким-либо образом препятствует связыванию между C2 и IgG1 в пленке. Контрольные пленки и волокна Rep4CT не продемонстрировали никакого IgG в элюированных фракциях (дорожки 10-14).
Пример 27 - Связывание IgG из плазмы крови человека с пленками C2-Rep4CT
Для дальнейшего исследования способности C2-Rep4CT связывать IgG, две пленки каждого из C2-Rep4CT (SEQ ID NO: 18), Z-Rep4CT (SEQ ID NO: 14) и Rep4CT (SEQ ID NO: 20) инкубировали с 500 мкл плазмы крови человека (разведение 1:5) в течение 1 ч при комнатной температуре. После промывания три раза 600 мкл PBS, связанный IgG элюировали в 500 мкл путем снижения значения pH до приблизительно 2,7 с помощью буфера для элюирования (0,5 M уксусная кислота, 1 M мочевина, 100 мМ NaCl), после чего элюированные фракции анализировали с помощью невосстанавливающего SDS-PAGE (фиг. 34).
Нанесение на гель, представленный на фиг. 34, проводили в соответствии с:
(1) Плазма крови человека (1:5), нанесение 1,4 мкл
(2) Маркер молекулярной массы
(3-4) C2-Rep4CT, пленка, в двух экземплярах, нанесение 14 мкл
(5-6) Z-Rep4CT, пленка, в двух экземплярах, нанесение 14 мкл
(7-8) Rep4CT, пленка, в двух экземплярах, нанесение 14 мкл.
На фиг. 34 отчетливо можно видеть, что пленки Z-Rep4CT имеют связанный IgG из плазмы крови человека (дорожки 5-6). Пленки C2-Rep4CT также демонстрируют более слабые полосы IgG в элюированных фракциях (дорожки 3-4). Контрольные пленки Rep4CT не демонстрируют связывания IgG из плазмы крови человека (дорожки 7-8).
Пример 28 - Сравнение вторичной структуры в волокнах и пленках Z-Rep4CT и Rep4CT с использованием ATR-FTIR
Вторичная структура Z-домена представляет собой α-спираль в ее активной конформации, в то время как вторичная структура волокон Rep4CT, главным образом, представляет собой структуру типа β-слоя. Если Z-домены в волокнах и пленках Z-Rep4CT правильно свернуты, можно ожидать более высокого содержания α-спиралей в этих матрицах по сравнению с волокнами и пленками Rep4CT. Для исследования отличий во вторичной структуре, волокна и пленки Z-Rep4CT и Rep4CT анализировали спектроскопическим способом ИК-спектроскопии нарушенного полного внутреннего отражения с Фурье-преобразованием (ATR-FTIR), посредством которого можно отличить структуру α-спирали (положение полосы: 1648-1657 см-1) от структуры β-слоя (положения полос: 1623-1641, 1674-1695 см-1).
Одну пленку Z-Rep4CT (SEQ ID NO: 14) и одну пленку Rep4CT (SEQ ID NO: 20) получали, позволяя 15 мкл раствора белка высохнуть на воздухе при комнатной температуре в течение ночи. Получали волокна для Z-Rep4CT и Rep4CT и после этого высушивали их воздухом в течение ~30 мин при комнатной температуре при растяжении. Затем регистрировали ATR-FTIR с использованием платинового элемента ATR от Bruker. ИК-спектры как для волокна, так и для пленки (не представлено) демонстрируют, что Z-Rep4CT имеет более высокое содержание α-спиралей, чем Rep4CT, что указывает на присутствие правильно свернутого Z-домена. Это согласуется с сохраненной функциональностью Z-домена в структурах Z-Rep4CT по изобретению, см. например, примеры 2-5, 17-19 и 22-23.










































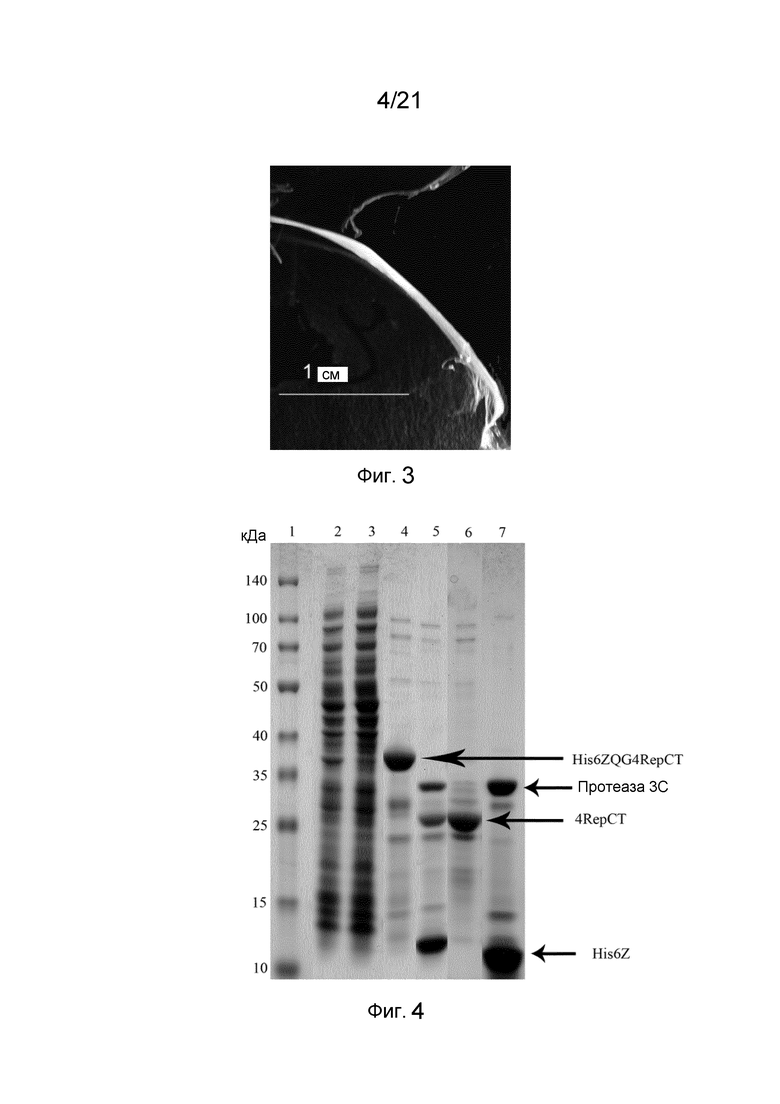


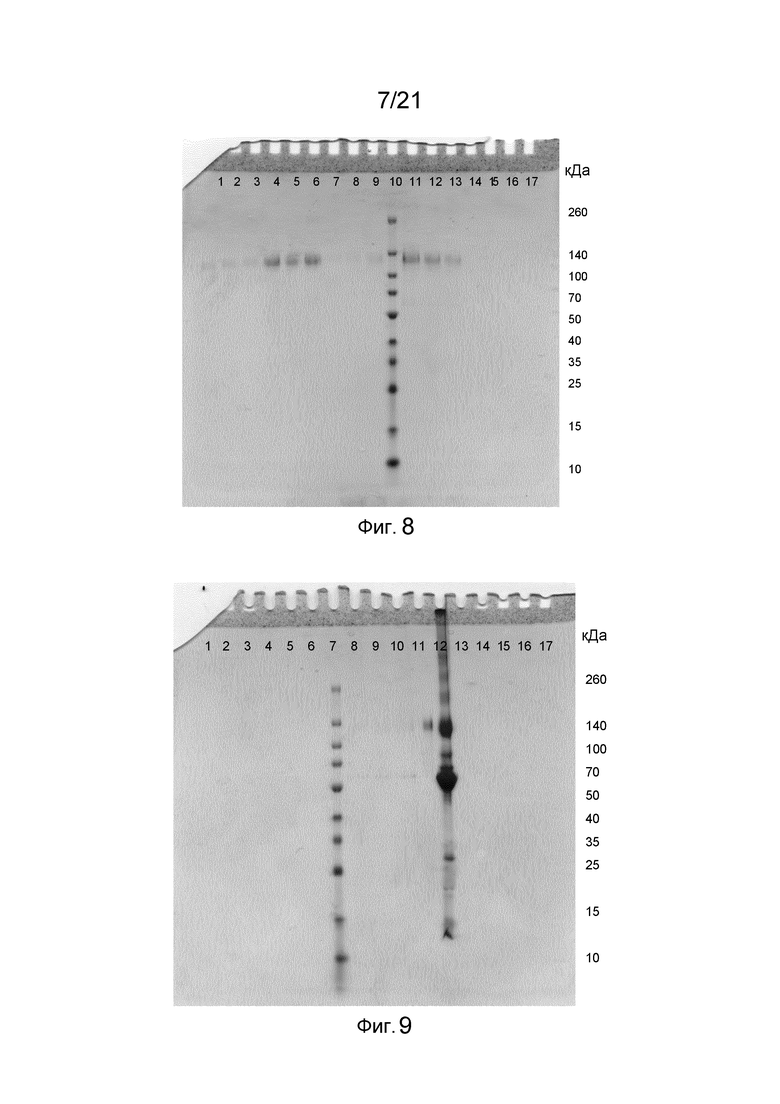

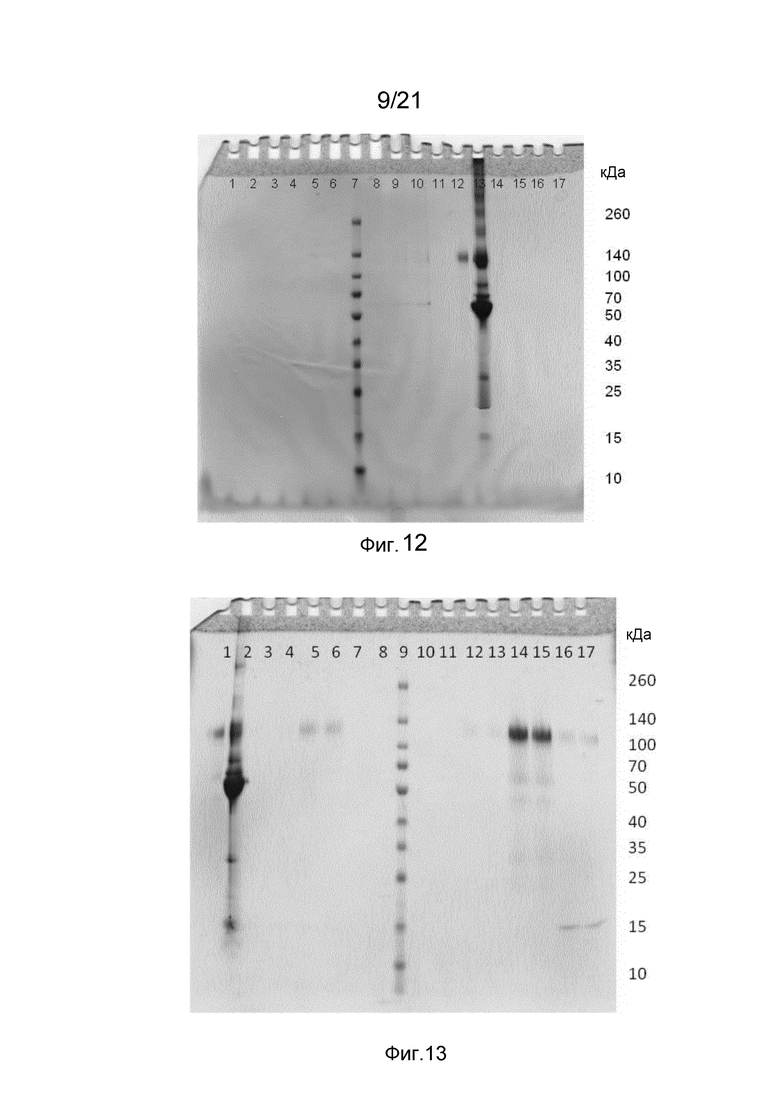
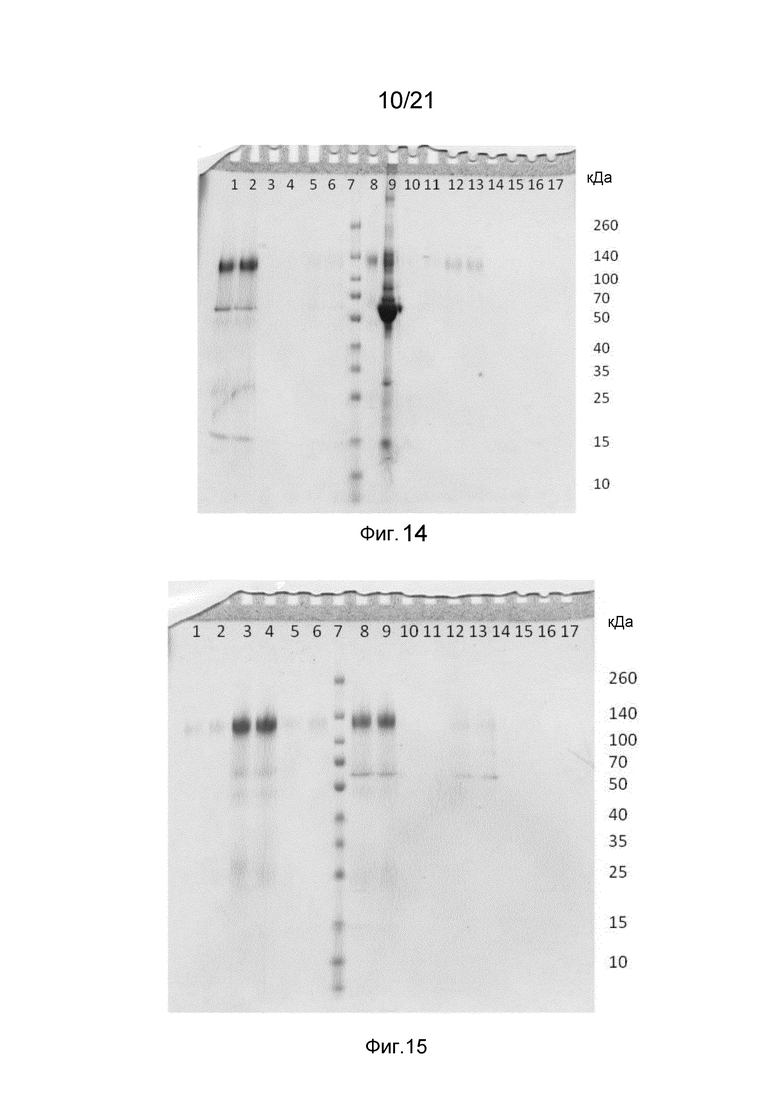
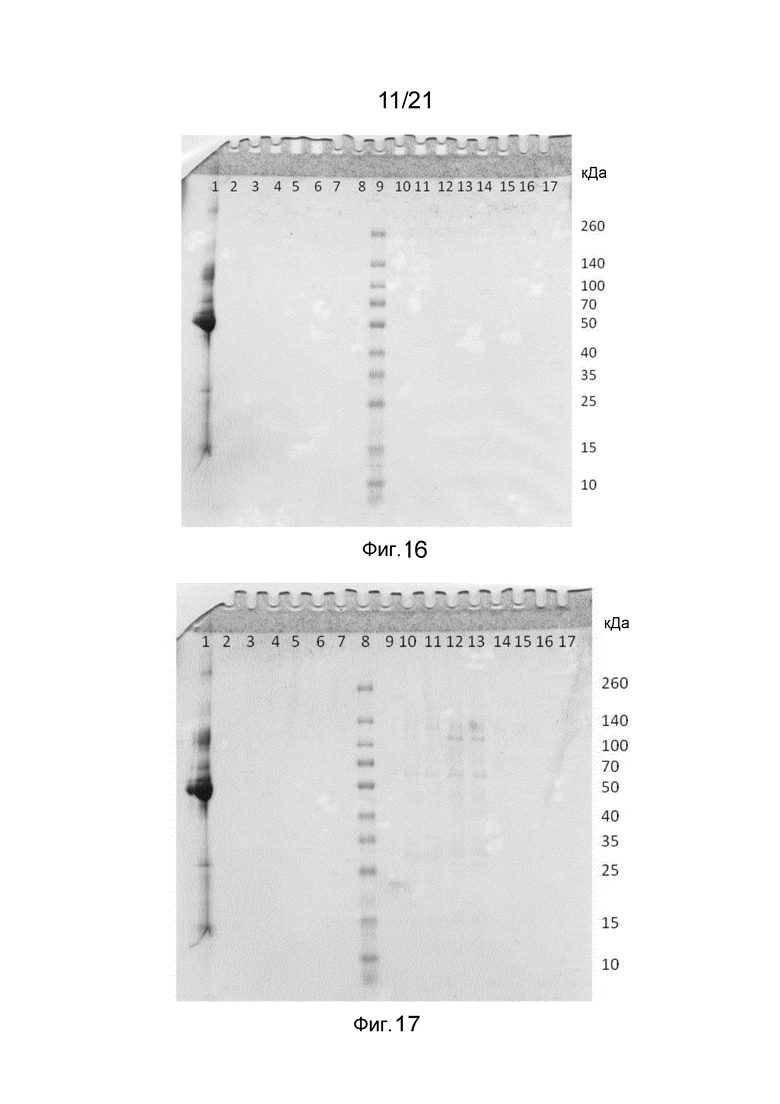
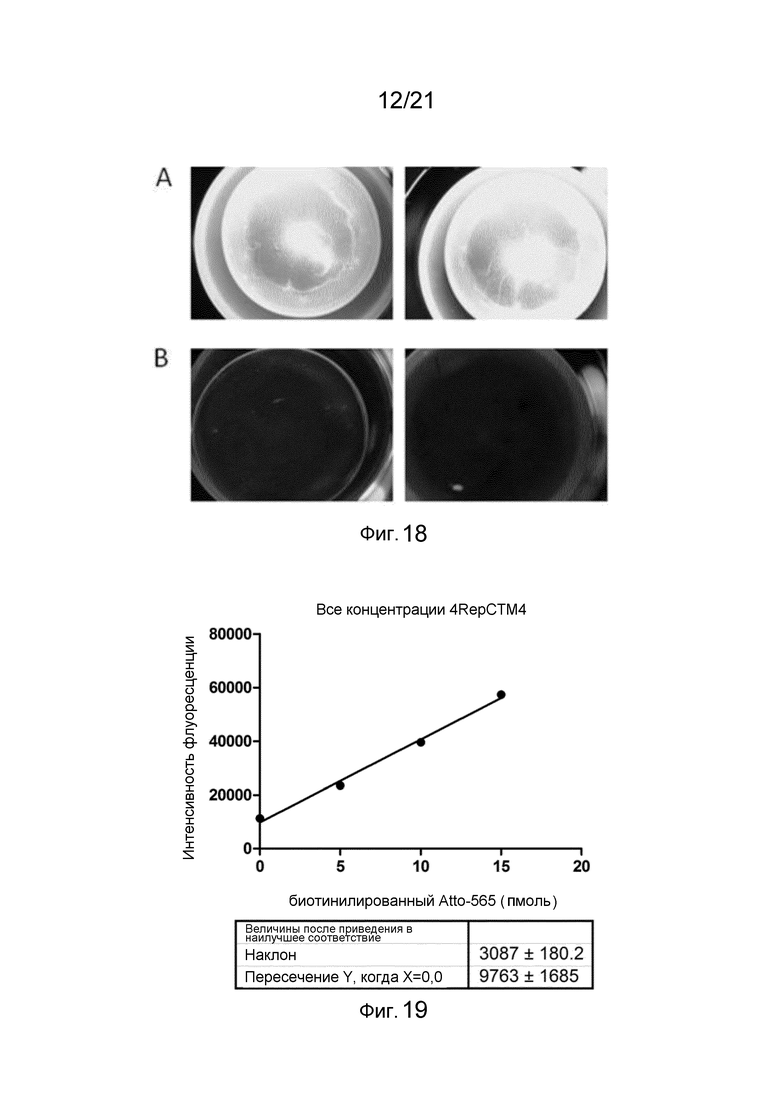
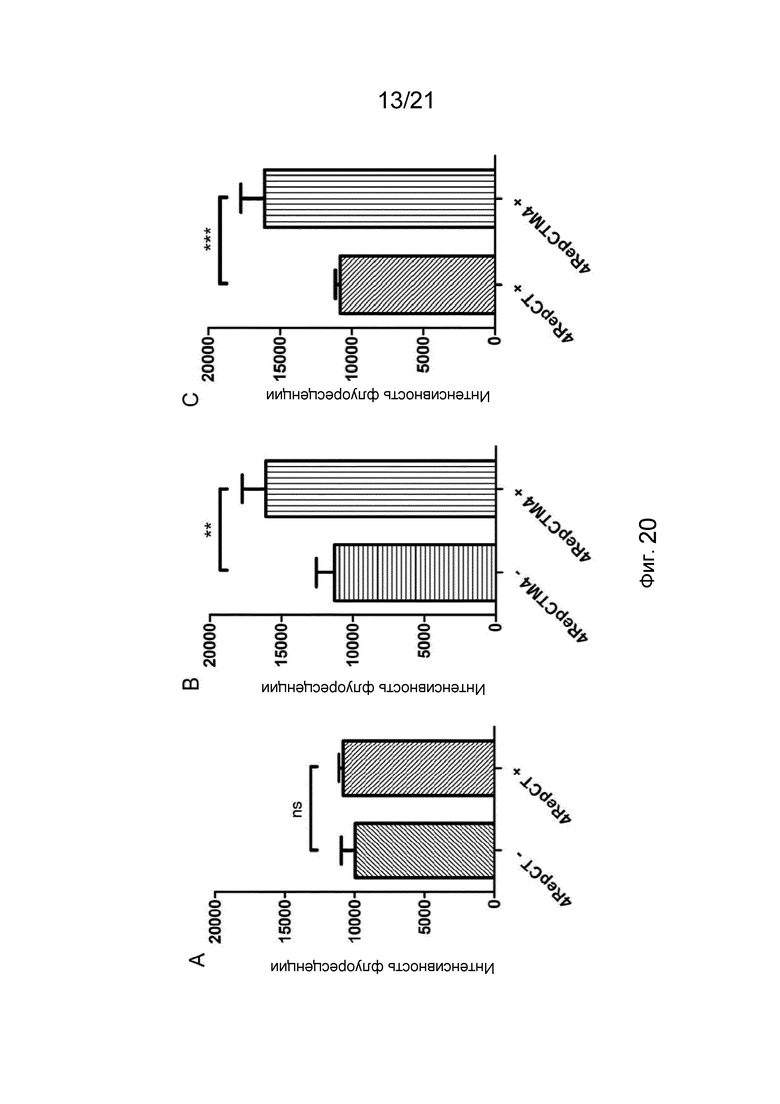
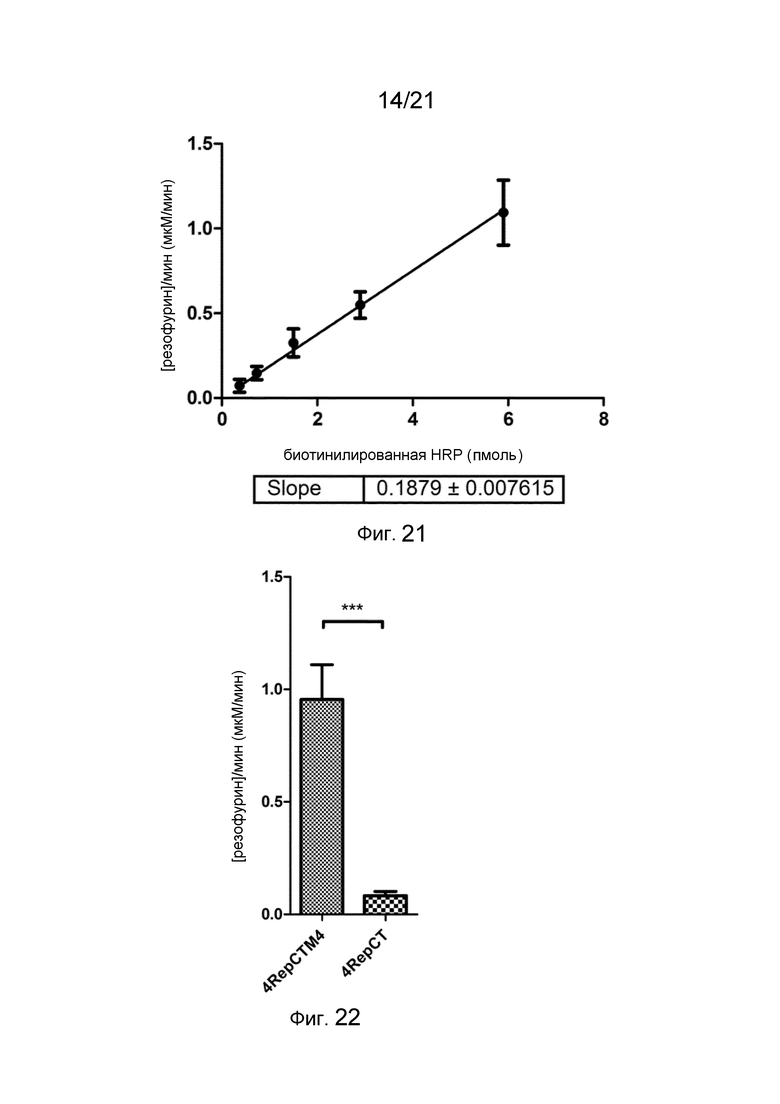
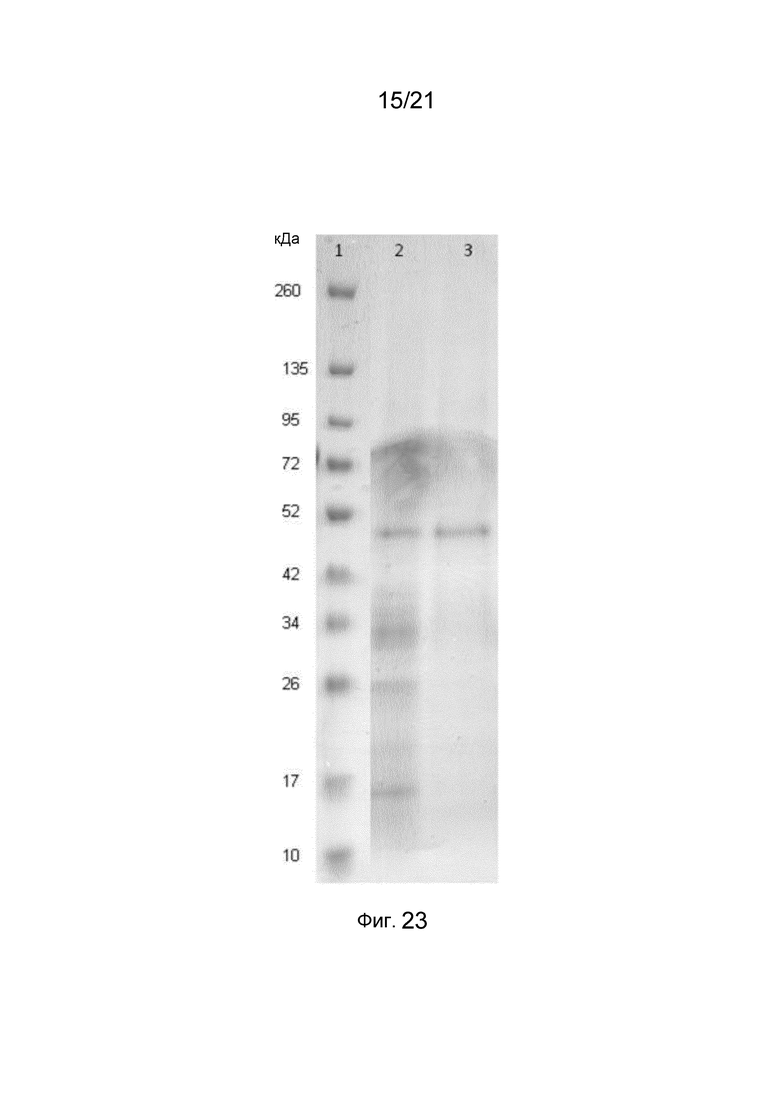
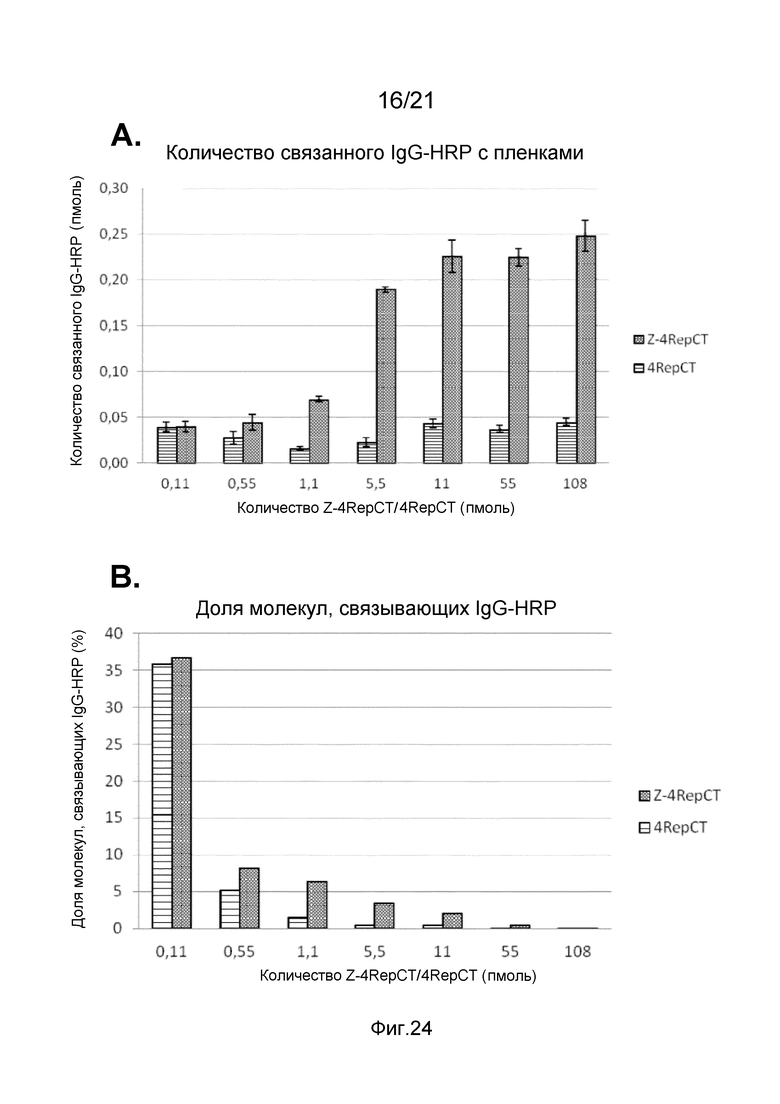
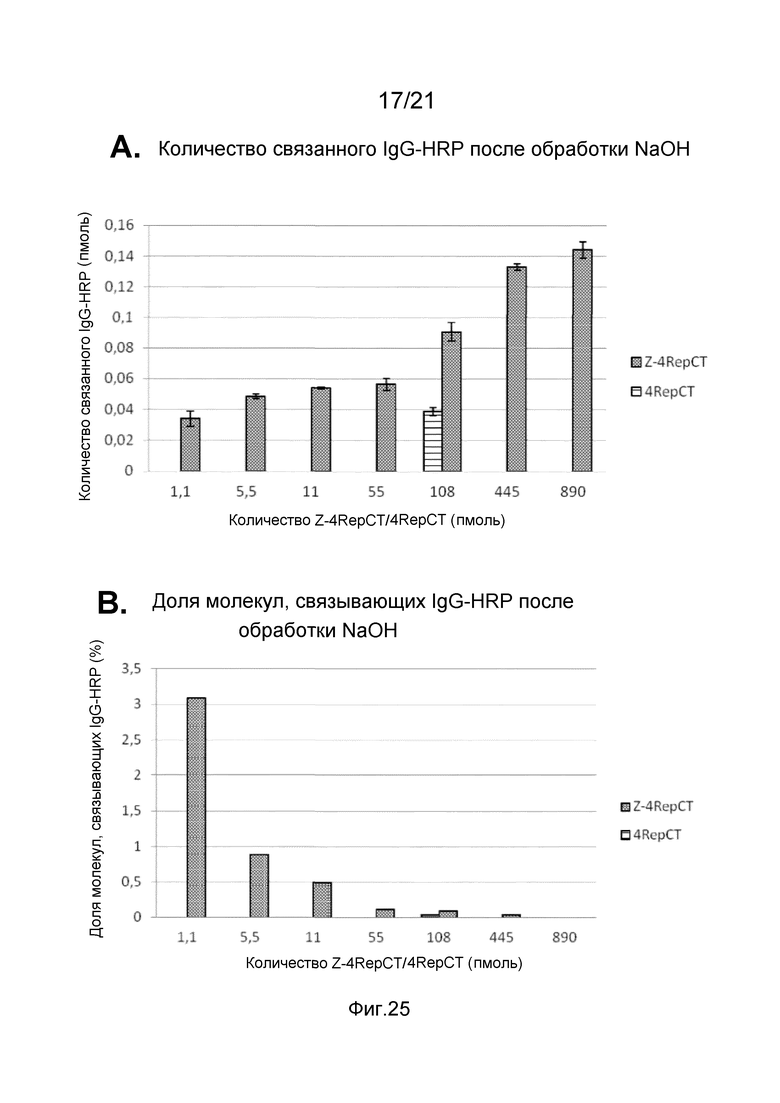
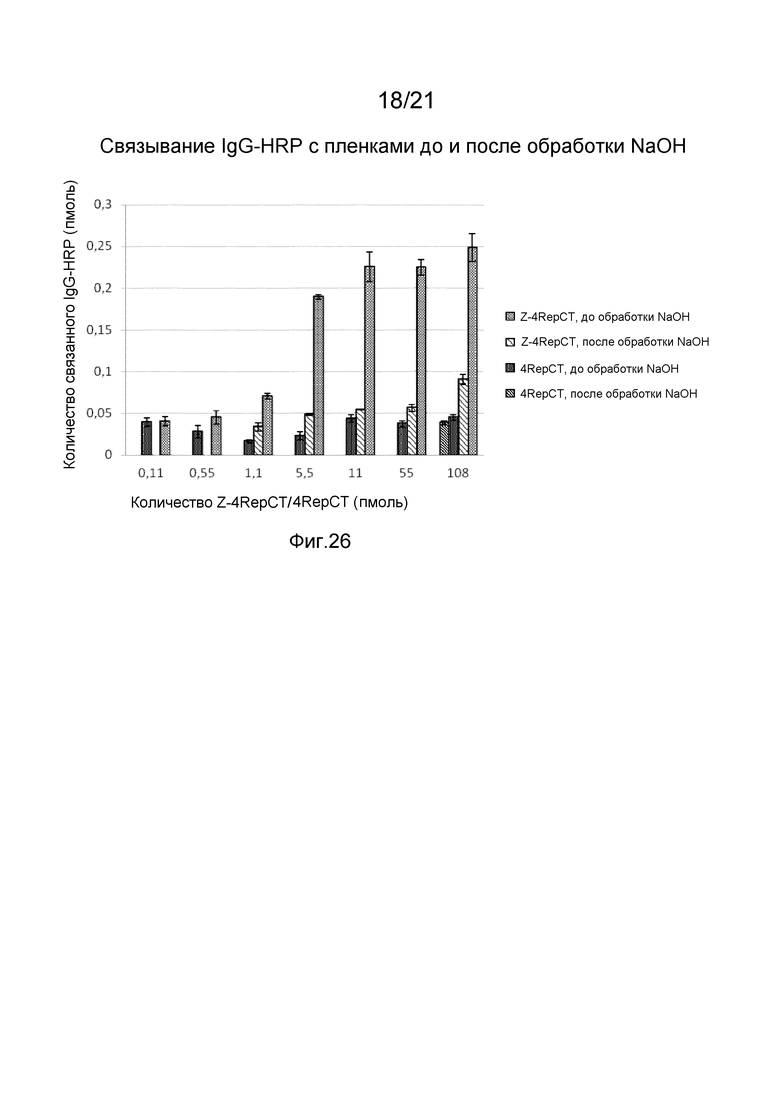
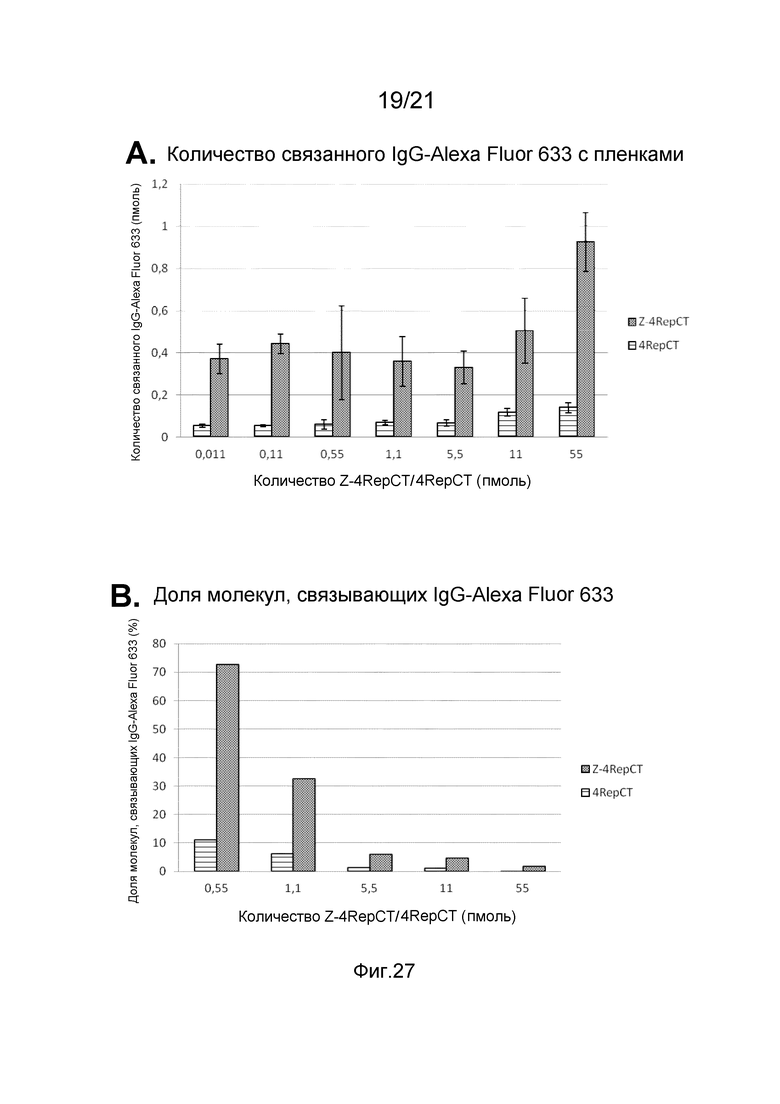
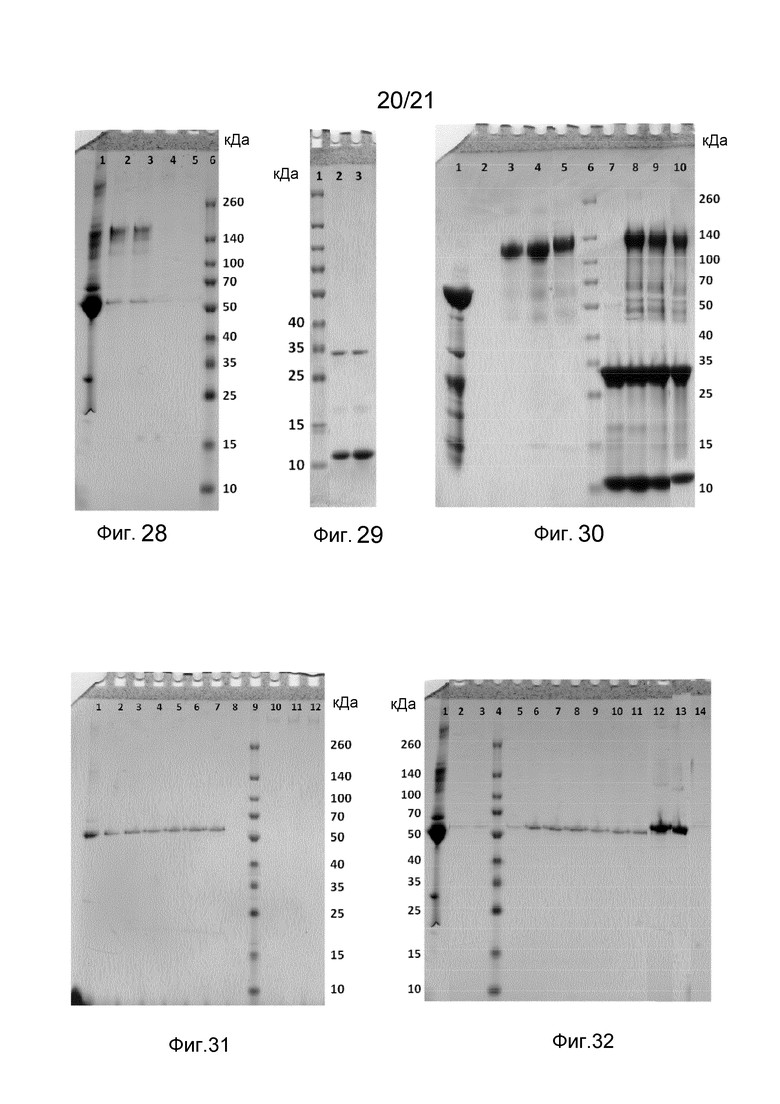
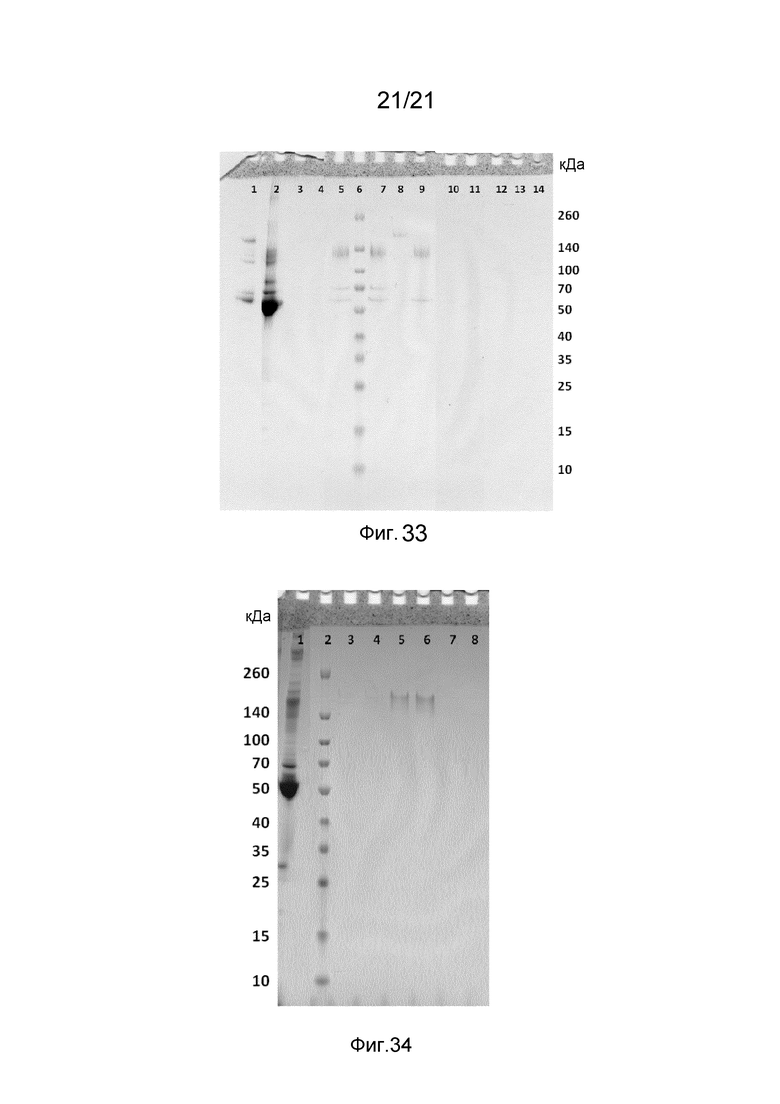
Изобретение относится к области биотехнологии, конкретно к получению полимерной белковой структуры, способной к селективному взаимодействию с органической мишенью, и может быть использовано в медицине. Белковая полимерная структура содержит в качестве повторяющегося структурного элемента рекомбинантный слитый белок, содержащий части B, REP и CT. Изобретение позволяет получить белковую структуру, пригодную в качестве аффинной среды и каркасного материала для клеток. 13 н. и 28 з.п. ф-лы, 34 ил., 5 табл., 28 пр.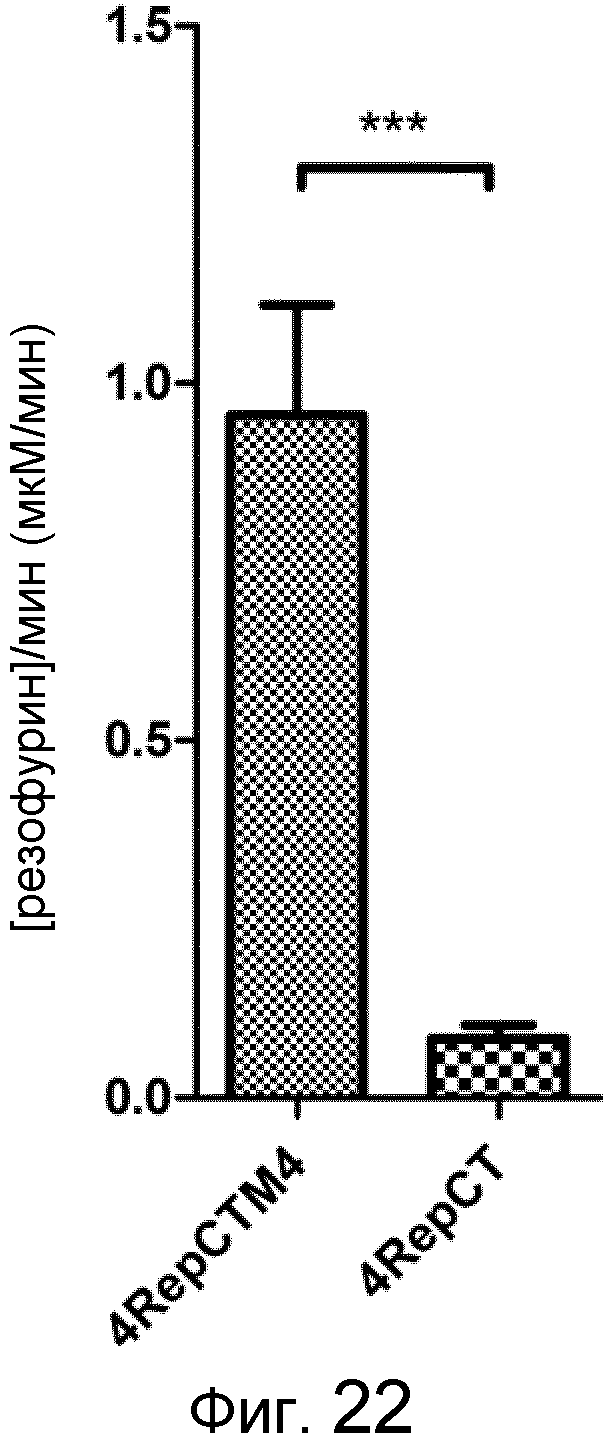
1. Белковая структура, способная к селективному взаимодействию с органической мишенью, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, где указанная белковая структура содержит упорядоченное множество слитых белков, где указанная белковая структура представляет собой полимер, содержащий в качестве повторяющегося структурного элемента рекомбинантный слитый белок, который способен к селективному взаимодействию с органической мишенью, и содержащий части В, REP и СТ, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами Bx-REP-CT, Bx-СТ-REP, REP-CT-Bz и CT-REP-Bz; где х и z представляют собой целые числа от 1 до 5, где полимер содержит более 100 структурных элементов слитого белка и где:
В представляет собой неспидроиновую часть из более чем 30 аминокислотных остатков, которая обеспечивает способность к селективному взаимодействию с органической мишенью, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков, белка А стафилококков и его доменов Е, D, А, В и С; и белковых фрагментов, имеющих по меньшей мере 70% идентичность с любой из этих аминокислотных последовательностей;
части REP и СТ обеспечивают способность образовывать полимер;
REP представляет собой часть из от 70 до 300 аминокислотных остатков, выбранных из группы, состоящей из L(AG)nL, L(AG)nAL, L(GA)nL, L(GA)nGL, где
n представляет собой целое число от 2 до 10;
каждый индивидуальный сегмент А представляет собой аминокислотную последовательность из от 8 до 18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala;
каждый индивидуальный сегмент G представляет собой аминокислотную последовательность из от 12 до 30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков представляют собой Gly; и
каждый индивидуальный сегмент L представляет собой линкерную аминокислотную последовательность из от 0 до 20 аминокислотных остатков; и
СТ представляет собой часть из от 70 до 120 аминокислотных остатков и имеет по меньшей мере 50% идентичность с SEQ ID NO: 9 или по меньшей мере 80% идентичность с SEQ ID NO: 7.
2. Белковая структура по п. 1, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков, В-домена белка А стафилококков и белковых фрагментов, имеющих по меньшей мере 70% идентичность с любой из этих аминокислотных последовательностей.
3. Белковая структура по п. 2, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков и белковых фрагментов, имеющих по меньшей мере 70% идентичность с Z-доменом белка А стафилококков.
4. Белковая структура по п. 3, где часть В представляет собой Z-домен белка А стафилококков.
5. Белковая структура по п. 4, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами
Bx-REP-By-CT-Bz и Bx-CT-By-REP-Bz,
где х, у и z представляют собой целые числа от 0 до 5;
и x+y+z≥1.
6. Белковая структура по п. 1, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами
B-REP-CT, B-CT-REP, REP-CT-B и CT-REP-B.
7. Белковая структура по п. 1, где часть В имеет менее чем 30% идентичность с любой из SEQ ID NO: 6-10.
8. Белковая структура по п. 1, где указанная белковая структура имеет размер по меньшей мере 0,1 мкм по меньшей мере по двум измерениям.
9. Белковая структура по п. 1, где указанная белковая структура находится в физической форме, выбранной из группы, состоящей из волокна, пленки и пены.
10. Белковая структура по п. 1, где часть СТ имеет по меньшей мере 50% идентичность с SEQ ID NO: 9 и по меньшей мере 80% идентичность с SEQ ID NO: 7.
11. Белковая структура по п. 1, где часть СТ имеет по меньшей мере 90% идентичность с SEQ ID NO: 7.
12. Белковая структура по п. 11, где часть СТ представляет собой SEQ ID NO: 7.
13. Белковая структура по п. 1, где слитый белок выбран из группы, состоящей из SEQ ID NO: 14, и белков, имеющих по меньшей мере 80% идентичность с SEQ ID NO: 14.
14. Способ получения белковой структуры по любому из пп. 1-13, проявляющей активность связывания с органической мишенью, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, включающий стадии:
(а) получения рекомбинантного слитого белка, который способен к селективному взаимодействию с органической мишенью, и содержащий части В, REP и СТ, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами Вх-REP-CT, Bx-CT-REP, REP-CT-Bz и CT-REP-Bz; где х и z представляют собой целые числа от 1 до 5, где
В представляет собой неспидроиновую часть из более чем 30 аминокислотных остатков, которая обеспечивает способность к селективному взаимодействию с органической мишенью, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков, белка А стафилококков и его доменов Е, D, А, В и С; и белковых фрагментов, имеющих по меньшей мере 70% идентичность с любой из этих аминокислотных последовательностей;
части REP и СТ обеспечивают способность образовывать полимер;
REP представляет собой часть из от 70 до 300 аминокислотных остатков, выбранных из группы, состоящей из L(AG)nL, L(AG)nAL, L(GA)nL, L(GA)nGL, где
n представляет собой целое число от 2 до 10;
каждый индивидуальный сегмент А представляет собой аминокислотную последовательность из от 8 до 18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala;
каждый индивидуальный сегмент G представляет собой аминокислотную последовательность из от 12 до 30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков представляют собой Gly; и
каждый индивидуальный сегмент L представляет собой линкерную аминокислотную последовательность из от 0 до 20 аминокислотных остатков; и
СТ представляет собой часть из от 70 до 120 аминокислотных остатков и имеет по меньшей мере 50% идентичность с SEQ ID NO: 9 или по меньшей мере 80% идентичность с SEQ ID NO: 7;
(b) воздействия на слитый белок условий для достижения образования полимера, содержащего указанный рекомбинантный слитый белок,
тем самым получают указанную белковую структуру.
15. Аффинная среда для иммобилизации органической мишени, необязательно содержащая указанную органическую мишень, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, причем указанная аффинная среда содержит слитый белок, который способен к селективному взаимодействию с органической мишенью, и содержащий части В, REP и СТ, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами Вх-REP-CT, Bx-CT-REP, REP-CT-Bz и CT-REP-Bz; где x и z представляют собой целые числа от 1 до 5, где
В представляет собой неспидроиновую часть из более чем 30 аминокислотных остатков, которая обеспечивает способность к селективному взаимодействию с органической мишенью, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков, белка А стафилококков и его доменов Е, D, А, В и С; и белковых фрагментов, имеющих по меньшей мере 70% идентичность с любой из этих аминокислотных последовательностей;
части REP и СТ обеспечивают способность образовывать полимер;
REP представляет собой часть из от 70 до 300 аминокислотных остатков, выбранных из группы, состоящей из L(AG)nL, L(AG)nAL, L(GA)nL, L(GA)nGL, где
n представляет собой целое число от 2 до 10;
каждый индивидуальный сегмент А представляет собой аминокислотную последовательность из от 8 до 18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala;
каждый индивидуальный сегмент G представляет собой аминокислотную последовательность из от 12 до 30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков представляют собой Gly; и
каждый индивидуальный сегмент L представляет собой линкерную аминокислотную последовательность из от 0 до 20 аминокислотных остатков; и СТ представляет собой часть из от 70 до 120 аминокислотных остатков и имеет по меньшей мере 50% идентичность с SEQ ID NO: 9 или по меньшей мере 80% идентичность с SEQ ID NO: 7.
16. Аффинная среда по п. 15, причем указанная аффинная среда содержит белковую структуру по любому из пп. 1-13, причем эта белковая структура представляет собой полимер, содержащий рекомбинантный слитый белок.
17. Аффинная среда по любому из пп. 15-16, дополнительно содержащая указанную органическую мишень, где часть В способна к селективному взаимодействию и связывается с указанной органической мишенью.
18. Аффинная среда по п. 17, где указанная органическая мишень способна к селективному взаимодействию со второй органической мишенью.
19. Аффинная среда по п. 16, где указанная белковая структура находится в физической форме пленки.
20. Каркасный материал для культивирования клеток, имеющих вторую органическую мишень, которая присутствует на клеточной поверхности, причем указанный каркасный материал для клеток содержит белковую структуру по любому из пп. 1-13,
где указанный каркасный материал для клеток дополнительно содержит органическую мишень, выбранную из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, где часть В способна к селективному взаимодействию и связывается с указанной органической мишенью, и где указанная органическая мишень способна к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности.
21. Каркасный материал для клеток по п. 20, где указанная белковая структура находится в физической форме пленки.
22. Применение белковой структуры по любому из пп. 1-13 для выделения органической мишени, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, из образца.
23. Применение белковой структуры по любому из пп. 1-13 для культивирования клеток.
24. Способ выделения органической мишени, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, из образца, включающий стадии:
получения образца, содержащего органическую мишень;
получения аффинной среды по любому из пп. 15-19, где указанная аффинная среда способна к селективному взаимодействию с указанной органической мишенью;
контактирования указанной аффинной среды с указанным образцом в подходящих условиях для достижения связывания между аффинной средой и указанной органической мишенью;
удаления несвязанного образца; и
необязательно включающий стадию контактирования указанной аффинной среды и указанной иммобилизованной органической мишени со второй органической мишенью.
25. Способ по п. 24, дополнительно включающий стадию контактирования указанной аффинной среды и иммобилизованной органической мишени со второй органической мишенью, которая способна к селективному взаимодействию с органической мишенью, в подходящих условиях для достижения связывания между органической мишенью и второй органической мишенью.
26. Способ по п. 24, где слитый белок в аффинной среде присутствует в качестве белковой структуры по любому из пп. 1-13 при контактировании указанной аффинной среды с указанным образцом для достижения связывания между аффинной средой и органической мишенью.
27. Способ по п. 24, где слитый белок в аффинной среде присутствует в растворе, когда указанную аффинную среду контактируют с указанным образцом для достижения связывания между аффинной средой и органической мишенью, и где комплексу слитого белка, связанного с органической мишенью, позволяют образовывать слитую белковую структуру по любому из пп. 1-13.
28. Способ по п. 24, дополнительно включающий стадию детекции, и необязательно количественного определения, присутствия иммобилизованной мишени на указанной аффинной среде.
29. Способ по п. 24, дополнительно включающий стадию высвобождения и сбора органической мишени из аффинной среды.
30. Способ по п. 24, где слитый белок в аффинной среде присутствует в качестве белковой структуры по любому из пп. 1-13 при контактировании указанной аффинной среды с указанным образцом для достижения связывания между органической мишенью и второй органической мишенью.
31. Способ по п. 24, где слитый белок в аффинной среде присутствует в растворе, когда указанную аффинную среду контактируют с указанным образцом для достижения связывания между органической мишенью и второй органической мишенью, и где комплексу слитого белка, связанного с органической мишенью и второй органической мишенью, позволяют образовывать слитую белковую структуру по любому из пп. 1-13.
32. Способ по п. 24, дополнительно включающий стадию высвобождения и сбора второй органической мишени из аффинной среды.
33. Способ по п. 24, дополнительно включающий конечную стадию регенерации аффинной среды путем химической обработки и/или стерилизующей обработки нагреванием.
34. Способ по п. 33, где химическая обработка включает обработку NaOH и/или мочевиной.
35. Способ по п. 24, где указанная белковая структура находится в физической форме пленки, волокна или пены.
36. Способ иммобилизации клеток, включающий
предоставление образца, содержащего представляющие интерес клетки;
нанесение указанного образца на каркасный материал для клеток по любому из пп. 20-21, где указанный каркасный материал для клеток способен к селективному взаимодействию с органической мишенью, которая присутствует на клеточной поверхности; и
обеспечение иммобилизации указанных клеток на указанный каркасный материал для клеток путем связывания между органической мишенью на клеточной поверхности и указанным каркасным материалом для клеток.
37. Способ культивирования клеток, включающий
иммобилизацию представляющих интерес клеток на каркасном материале для клеток в соответствии со способом по п. 36; и поддержание указанного каркасного материала для клеток, имеющего клетки, нанесенные на него, в условиях, подходящих для клеточной культуры.
38. Рекомбинантный слитый белок для формирования белковой структуры, способной к селективному взаимодействию с органической мишенью, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, состоящий из SEQ ID NO: 14.
39. Выделенная полинуклеиновая кислота, кодирующая белок для формирования белковой структуры, способной к селективному взаимодействию с органической мишенью, выбранной из группы, состоящей из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, выбранная из группы, состоящей из нуклеиновых кислот, кодирующих слитый белок по п. 38, и SEQ ID NO: 15.
40. Способ получения слитого белка, включающий следующие стадии:
a) экспрессия в подходящем хозяине слитого белка по п. 38; и
b) получение смеси, содержащей слитый белок, и выделение слитого белка.
41. Применение рекомбинантного слитого белка, который способен к селективному взаимодействию с органической мишенью, и содержащий части В, REP и СТ, где указанный рекомбинантный слитый белок выбран из группы белков, определяемой формулами Вх-REP-CT, Bx-CT-REP, REP-CT-Bz и CT-REP-Bz; где х и z представляют собой целые числа от 1 до 5, где
В представляет собой неспидроиновую часть из более чем 30 аминокислотных остатков, которая обеспечивает способность к селективному взаимодействию с органической мишенью, где часть В выбрана из группы, состоящей из Z-домена белка А стафилококков, белка А стафилококков и его доменов Е, D, А, В и С; и белковых фрагментов, имеющих по меньшей мере 70% идентичность с любой из этих аминокислотных последовательностей;
части REP и СТ обеспечивают способность образовывать полимер;
REP представляет собой часть из от 70 до 300 аминокислотных остатков, выбранных из группы, состоящей из L(AG)nL, L(AG)nAL, L(GA)nL, L(GA)nGL, где
n представляет собой целое число от 2 до 10;
каждый индивидуальный сегмент А представляет собой аминокислотную последовательность из от 8 до 18 аминокислотных остатков, где от 0 до 3 аминокислотных остатков не являются Ala, и остальные аминокислотные остатки представляют собой Ala;
каждый индивидуальный сегмент G представляет собой аминокислотную последовательность из от 12 до 30 аминокислотных остатков, где по меньшей мере 40% аминокислотных остатков представляют собой Gly; и каждый индивидуальный сегмент L представляет собой линкерную аминокислотную последовательность из от 0 до 20 аминокислотных остатков; и
СТ представляет собой часть из от 70 до 120 аминокислотных остатков и имеет по меньшей мере 50% идентичность с SEQ ID NO: 9 или по меньшей мере 80% идентичность с SEQ ID NO: 7,
для получения белковой структуры, способной к селективному взаимодействию с органической мишенью, выбранной из области кристаллизующегося фрагмента (Fc) IgG и молекул, содержащих IgG или его производные, где указанная белковая структура представляет собой полимер, содержащий рекомбинантный слитый белок.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| WO 2010097385 A1, 02.09.2010 | |||
| PRINCE J.T | |||
| et al., Construction, cloning, and expression of synthetic genes encoding spider dragline silk, Biochemistry, 1995, v.29, is.34, p.10879-10885 | |||
| RU 2007102846 A, 27.08.2008. | |||

