Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности к способу получения янтарной кислоты с использованием дрожжей, принадлежащих к роду Yarrowia, в которых снижена активность сукцинатдегидрогеназы.
Предшествующий уровень техники
Yarrowia lipolytica являются уникальными дрожжами в виду их способности к продукции широкого спектра органических кислот, включая интермедиаты цикла трикарбоновых кислот, такие как лимонная и изолимонная кислоты, и секреции их в среду. Установлено, что в ходе продолжительного выращивания Y.lipolytica N 1 потребность в кислороде для роста, а также синтез лимонной кислоты зависят от концентрации железа в питательной среде. Установлен связанный эффект влияния концентраций кислорода и железа на функционирование митохондриального транспорта электронов транспортной цепи в штамме Y lipolytica N 1. На основе результатов, полученных в ходе продолжительного выращивания, предложены условия для производства лимонной кислоты в дозированной среде с помощью Y.lipolytica N 1 (Kamzolova S.V. et al, FEMS Yeast Res.; 3(2):217-22(2003)).
Янтарная кислота, член семейства С4-дикарбоновых кислот, широко используется в производстве пищи, медикаментов и биодеградируемых пластмасс. Традиционно янтарную кислоту производят путем химического синтеза из нефтяного сырья, которое является невозобновимым, кроме того, процесс химического синтеза приводит к загрязнению окружающей среды. Поэтому большое внимание уделяется использованию эффективных природных продуцентов янтарной кислоты, таких как микроорганизмы.
Большинство описанных попыток увеличить продукцию промышленно значимой янтарной кислоты (сукцината) проводили в анаэробных условиях, в которых ферментация Escherichia coli приводит к получению смеси кислот. Разработаны стратегии использования системы аэробной продукции сукцината, позволяющие штаммам Е.coli эффективно производить и накапливать сукцинат, и такая система является базисом для увеличения аэробной продукции сукцината в Е.coli, основанной на создании нового аэробного центрального метаболизма (Lin Н. et al., Biotechnol Bioeng; 89(2): 148-56(2005)).
Различные мутантные штаммы Е.coli, сконструированные для продукции сукцината в аэробных условиях, были охарактеризованы в хемостате. Для того чтобы лучше понять метаболизм в этих мутантных штаммах изучали профили метаболитов, активности ферментов, и профили экспрессии генов. Наиболее эффективный продуцент сукцината, мутантный штамм HL27659k, содержал 5 следующих мутаций: sdhAB, (ackA-pta), рохВ, iclR, и ptsG. Было показано, что путь биосинтеза сукцината, сконструированный в штамме HL27659k, был высокоэффективным, что приводило к получению сукцината как основного продукта, получаемого в аэробных условиях (Lin Н. et al, Metab Eng.; 7(5-6):337-52(2005)).
Для установления и демонстрации способности к высокой продукции сукцината штамма HL27659k(pKK313) в аэробных условиях проводились эксперименты в реакторах с подпиткой. Результаты показали, что продукция сукцината в аэробных условиях с использованием сконструированного штамма HL27659k(pKK313) более практична по сравнению с традиционными анаэробными системами и обладает значительным потенциалом для производства сукцината в промышленных масштабах и для оптимизации процесса (Lin Н. et al, Biotechnol Bioeng; 90(6):775-9(2005)).
Для получения сукцината было осуществлено двухступенчатое культивирование штамма NZN111, двойного мутанта по pflB ldhA, у которого утрачена способность к анаэробной ферментации глюкозы вследствие дисбаланса редокс-системы. Было установлено, что после аэробного выращивания штамма NZN111 на среде с ацетатом, этот штамм восстанавливает способность к ферментации глюкозы с продукцией сукцината в качестве основного продукта в последующих анаэробных условиях. Анализ активностей ключевых ферментов выявил, что активности изоцитратлиазы, малатдегидрогеназы, малатного фермента (malic enzyme), фосфоенолпируваткарбоксикиназы (ФЕП-карбоксикиназы) были существенно повышены, в то время как активности пируваткиназы и ФЕП-карбоксилазы были снижены в клетках, выращенных на ацетате. Полученные результаты указывают на большой потенциал, который может дать выгоду в механизме клеточной регуляции для улучшения продукции сукцината с использованием штаммов Е.coli с модифицированным метаболизмом (Wu Н. et al., Appl Environ Microbiol.; 73(24):7837-43(2007)).
На основе данных о последовательности полного генома капнофильных («любящих CO2») сукцинат-продуцирующих бактерий из первого желудка жвачных животных, Mannheimia succiniciproducens, было проведено исследование по делециям генов (нокаут генов) для изучения метаболизма бактерий при анаэробной ферментации и последующей разработки метаболически измененных штаммов, способных к продукции сукцината без образования побочных продуктов. Среди трех метаболических реакций, протекающих с фиксацией СO2 катализируемых ФЕП-карбоксикиназой, ФЕП-карбоксилазой и малатным ферментом, ФЕП-карбоксикиназа была наиболее важной для анаэробного роста М. succiniciproducens и для продукции сукцината. Было установлено, что оксалоацетат, образующийся при карбоксилировании ФЕП, превращается в сукцинат в ходе трех последовательных реакций, катализируемых малатдегидрогеназой, фумаразой и фумаратредуктазой. Главные метаболические пути, приводящие к образованию побочных продуктов, были успешно удалены путем разрушения генов ldhA, pflB, pta и ackA. Полученный штамм LPK7 обладал способностью к продукции сукцината из глюкозы с незначительным накоплением уксусной, муравьиной или молочной кислот или вообще без него (Lee S.J. et al., Appl Environ Microbiol.; 72(3):1939-48(2006)).
Рост и продукция сукцината в сравнении с продукцией лактата из глюкозы с использованием Anaerobiospirillum succiniciproducens регулировали уровнем доступного оксида углерода и pH среды. Выход сукцината и выход АТФ в пересчете на моль глюкозы существенно повышался в условиях избытка СО2-HCO3 -, и было высказано предположение о том, что существует пороговый уровень СО2 для усиления продукции сукцината с использованием A.succiniciproducens. Было показано, что А.succiniciproducens, в отличие от других анаэробов, продуцирующих сукцинат и также образующих пропионат, могут расти быстро и продуцировать сукцинат с высоким выходом при рН 6.2 и избытке СО2-HCO3 - как следствие регулируемого метаболизма оттока электронов (Samuelov N.S. et al., Appl Environ Microbiol.; 57(10):3013-9(1991)).
Использование питательных сред с определенным химическим составом позволяет проводить различные исследования метаболизма, которые невозможны со средами с неопределенным составом. Среда с определенным составом АМ3 была создана для расширения экспериментальных возможностей при изучении метаболизма при ферментации Actinobacillus succinogenes, продуцирующих сукцинат. Было произведено сравнение тенденций роста A.succinogenes и набор конечных продуктов при ферментации в среде АМ3 и в богатой среде. Неспособность к синтезу α-кетоглутарата из глюкозы указывает на то, что активности по крайней мере двух ферментов цикла трикарбоновых кислот отсутствуют в A.succinogenes (McKinlay J.B. et al., Appl Environ Microbiol; 71(11):6651-6(2005)).
Описан способ получения больших количеств сукцината в анаэробных условиях с использованием мутантных штаммов Е.coli, в которых разрушены гены adhE, ldhA, iclR, arcA и ack-pta genes или их комбинация (US 20060046288 A1).
Описан способ получения янтарной кислоты из промышленных гидролизатов, включающий стадии: выращивание микроорганизмов (Е.coli, Klebsiela, Erwinia, Lactobacillus), содержащих мутации в генах ptsG, pflB, and ldhA с целью накопления биомассы; и предоставление микроорганизмам возможности метаболизировать гидролизаты (US 20030017559 A1).
Раскрыт ферментативный способ получения янтарной кислоты, включающий выбор штамма бактерии, который не продуцирует янтарную кислоту в большом количестве, разрушение нормальной регуляции метаболизма сахара в такой бактерии, и объединения такого мутантного штамма бактерии с выбранных сахаром в анаэробных условиях для осуществления получения янтарной кислоты. Также раскрыт способ превращения бактерии, продуцирующей янтарную кислоту с низким выходом, в бактерию, продуцирующую янтарную кислоту с высоким выходом, включающий отбор штамма бактерий, обладающего фосфотрансферазной системой, и изменение этой фосфотрансферазной системы таким образом, что штамм бактерии становится способным метаболизировать различные сахара одновременно (US 6159738).
Сукцинатдегидрогеназа (SDH) из Saccharomyces cerevisiae состоит из четырех субъединиц, кодирующихся генами SDH1, SDH2, SDH3 и SDH4. Было установлено, что разрушение двух генов SDH1 и SDH2 или SDH1b (гомолог SDH1) необходимо для полной утраты активности SDH и что ген SDH1b компенсирует функцию гена SDH1. Мутанты с разрушенными генами SDH1 и SDH1B показали небольшое увеличение продукции сукцината (до 1.9 раза) наряду с уменьшением продукции малата в сравнении с исходными штаммами в условиях культивирования со встряхиванием (Kubo Y. et al., J Biosci Bioeng; 90(6):619-24(2000)).
Путь, приводящий к накоплению сукцината, был изучен в жидкой культуре в присутствии высокой концентрации глюкозы (15%) в аэробных и анаэробных условиях с использованием серии штаммов Saccharomyces cerevisiae, в которых были разрушены различные гены, кодирующие ферменты, требующиеся в цикле трикарбоновых кислот. Полученные результаты свидетельствуют о том, что сукцинат может быть синтезирован двумя путями, а именно окислением α-кетоглутарата через цикл трикарбоновых кислот и восстановлением фумарата в анаэробных условиях (Arikawa Y. et al., J Biosci Bioeng; 87(1):28-36(1999)).
Описан способ получения янтарной кислоты с использованием микроорганизма, трансформированного рекомбинантным вектором, содержащим гены, кодирующие малатный фермент В (maeB)/ фумаратгидратазу С (fumC)/фумаратдегидрогеназы D и Е (fdhD and fdhE) (US 20070042476 A1, US 20070042477 A1 and US 20080020436 A1 соответственно).
Описан набор метаболических модификаций, связывающих необходимую и обязательную продукцию сукцината с ростом микроорганизма, включающий разрушение одного или нескольких генов, выбранных из группы, состоящей из генов: (a) adhE, ldhA; (b) adhE, ldhA, acka-pta; (c)pfl, ldhA; (d)pfl, ldhA, adhE; (e) acka-pta, pykF, atpF, sdhA; (f) acka-pta, pykF, ptsG, или (g) acka-pta, pykF, ptsG, adhE, ldhA (US 20070111294).
В настоящее время отсутствуют сообщения о снижении активности сукцинатдегидрогеназы в дрожжах, принадлежащих к роду Yarrowia, для получения янтарной кислоты.
Краткое описание изобретения
Целью настоящего изобретения является увеличение продукции янтарной кислоты с использованием дрожжей, принадлежащих к роду Yarrowia, и предоставление способа получения янтарной кислоты.
Цель настоящего изобретения была достигнута установлением того факта, что снижение активности сукцинатдегидрогеназы может существенно увеличить продукцию янтарной кислоты.
Целью настоящего изобретения является предоставление дрожжей, принадлежащих к роду Yarrowia, которые продуцируют янтарную кислоты и у которых снижена активность сукцинатдегидрогеназы.
Также целью настоящего изобретения является предоставление дрожжей, описанных выше, которые модифицировны таким образом, что экспрессия одного или нескольких генов, выбранных из группы, состоящей из генов SDH1 (YALI0D11374g), SDH2 (YALI0D23397g), SDH3 (YALI0E29667g) и SDH4 (YALI0A14784g), ослаблена.
Также целью настоящего изобретения является предоставление дрожжей, описанных выше, в которых экпрессия гена SDH1 (YALI0D11374g) ослаблена за счет введения температурочувствительной мутации в ген SDH1 (YALI0D11374g).
Также целью настоящего изобретения является предоставление дрожжей, описанных выше, в которых экпрессия гена SDH2 (YALI0D23397g) ослаблена за счет инактивации гена SDH2 (YALI0D23397g).
Также целью настоящего изобретения является предоставление дрожжей, описанных выше, при этом указанные дрожжи принадлежат к виду Yarrowia lipolytica.
Также целью настоящего изобретения является предоставление дрожжей, описанных выше, при этом указанные дрожжи представлены штаммом Yarrowia lipolytica VKPMY-3314.
Также целью настоящего изобретения является предоставление способа получения янтарной кислоты, который включает выращивание дрожжей, описанных выше, в питательной среде и выделение янтарной кислоты из культуральной жидкости.
Также целью настоящего изобретения является предоставление способа, описанного выше, в котором по крайней мере часть процесса осуществляется при рН менее 4.
Также целью настоящего изобретения является предоставление способа, описанного выше, в котором питательная среда содержит глицерин.
Также целью настоящего изобретения является предоставление способа получения полимера, содержащего янтарную кислоту, включающий стадии получения янтарной кислоты способом, описанным выше, и полимеризации полученной янтарной кислоты.
Далее настоящее изобретение описано более подробно.
Подробное описание изобретения
В настоящем изобретении «янтарная кислота» может быть в форме свободной кислоты или ее соли в зависимости от рН или присутствия ионов. Термины «янтарная кислота» и «сукцинат» являются здесь взаимозаменяемыми.
1. Дрожжи согласно настоящему изобретению.
Дрожжами согласно настоящему изобретению являются дрожжи, принадлежащие к роду Yarrowia, которые производят янтарную кислоту и у которых снижена активность сукцинатдегидрогеназы.
Фраза "дрожжи, принадлежащие к роду Yarrowia" означает, что дрожжи относят к дрожжам рода Yarrowia в соответствии с классификацией, известной специалисту в данной области микробиологии. Примером дрожжей, принадлежащих к роду Yarrowia, используемых в рамках настоящего изобретения, являются дрожжи Yarrowia lipolytica (Y.lipolytica). В настоящем изобретении фраза "указанные дрожжи продуцируют янтарную кислоту" означает, что такие дрожжи обладают способностью производить янтарную кислоту и экскретировать янтарную кислоту в среду, в условиях, когда дрожжи выращивают в питательной среде.
В настоящем изобретении фраза «указанные дрожжи продуцируют янтарную кислоту» также означают, что дрожжи обладают способностью к продукции и накоплениию янтарной кислоты в количестве большем, чем природный или родительский штамм, и предпочтительно означает, что дрожжи способны накапливать в среде не менее чем 1.0 г/л, более предпочтительно не менее чем 5.0 г/л и наиболее предпочтительно не менее чем 10.0 г/л янтарной кислоты.
Фраза «(указанные дрожжи) у которых снижена активность сукцинатдегидрогеназы» означает, что дрожжи модифицированы таким образом, что активность сукцинатдегидрогеназы в клетке снижена по сравнению с немодифицированным, природным или родительским штаммом.
Сукцинатдегидрогеназа (SDH) является компонентом комплеса II цепи дыхания, который катализирует окисление сукцината в фумарат в цикле Кребса и насыщает пул убихинона электронами. Указанный комплекс, являющийся высококонсервативным в ходе эволюции, локализован во внутренней мембране и состоит из двух каталитических и двух структурных доменов, все из которых кодируются генами, находящимися в ядре. В Saccharomyces cerevisiae были обнаружены и охарактеризованы четыре гена (с SDH1 по SDH4), кодирующие SDH. Флавопротеиновая субъединица, ответственная за окисление сукцината в фумарат кодируется двумя генами-паралогами, SDH1 и SDH1b, при этом только SDH1 необходим для роста с дыханием на источниках углерода. SDH2 кодирует железозависимый белковую субъединицу, которая содержит три различных FeS-центра и вместе с белком Sdh1p составляет каталитический каркас комплекса SDH, который переносит электроны из ковалентно связанного флавинадениндинуклеотида (FAD) белка Sdh1p сначала на FeS центр, а затем на убихинон. Гены SDH3 и SDH4 кодируют два малых гидрофобных пептида, которые прикрепляют комплекс в внутренней мембране митохондрий (Saliola М. et al., Eukaryot Cell; 3(3):589-97(2004)).
Авторы настоящего изобретения проанализировали последовательность генома штамма Y.lipolytica CLIB122 (http://www.ncbi.nlm.nih.gov/sites/entrez) и обнаружили, что открытые рамки считывания (ОРС) YALI0D11374g, YALI0D23397g, YALI0E29667g и YALI0A14784g гомологичны генам SDH1, SDH2, SDH3 и SDH4 из S.cerevisiae соответственно. Указанные ОРС были названы SDH1, SDH2, SDH3 и SDH4 соответственно. Нуклеотидная последовательность гена SDH1 и аминокислотная последовательность белка SDH1, кодируемого геном SDH1, приведены в Перечне последовательностей под номерами SEQ ID NO:1-й SEQ ID NO:2 соответственно. Нуклеотидная последовательность гена SDH2 и аминокислотная последовательность белка SDH2, кодируемого геном SDH2, приведены в Перечне последовательностей под номерами SEQ ID NO:3 и SEQ ID NO:4 соответственно. Нуклеотидная последовательность гена SDH3 и аминокислотная последовательность белка SDH3, кодируемого геном SDH3, приведены в Перечне последовательностей под номерами SEQ ID NO:5 и SEQ ID NO:6 соответственно. Нуклеотидная последовательность гена. SDH4 и аминокислотная последовательность белка SDH4, кодируемого геном SDH4, приведены в Перечне последовательностей под номерами SEQ ID NO:7 и SEQ ID NO:8 соответственно.
В связи с тем, что могут существовать отличия в последовательности ДНК между штаммами Yarrowia, гены SDH1, SDH2, SDH3 и SDH4, которые предполагается модифицировать не ограничены генами, приведенными под номерами SEQ ID No:1, SEQ ID No:3, SEQ ID No:5 и SEQ ID No:7, но также могут включать в себя гены, гомологичные генам SEQ ID No:1, SEQ ID No:3, SEQ ID No:5 и SEQ ID No:7, которые кодируют варианты белков SDH1, SDH2, SDH3 и SDH4 соответственно. Фраза"вариант белка", использующаяся в настоящем изобретении означает белок, в котором присутствуют замены в последовательности, будь то делеции, вставки, добавления, или замены аминокислот, но при этом обладает активностью продуктов, таких как белки SDH1/ SDH2/ SDH3/ SDH4. Количество изменений в варианте белка зависит от положения или типа аминокислотного остатка в трехмерной структуре белка. Это может быть от 1 до 30, предпочтительно от 1 до 15 и наиболее предпочтительно от 1 до 5 в последовательности SEQ ID No:2, SEQ ID No:4, SEQ ID No:6 и SEQ ID No:8. Эти изменения в варианте белка могут быть в участках белка, которые не являются критичными для функционирования белка. Это происходит потому, что некоторые аминокислоты обладают большой гомологией по отношению к другим аминокислотам и трехмерная структура и активность не меняются при таких изменениях. Поэтому вариантом белка, кодируемого геном SDH1, SDH2, SDH3 или SDH4, может быть белок, обладающий гомологией не менее 80%, предпочтительно не менее 90%, еще более предпочтительно не менее 95% и наиболее предпочтительно не менее 98% по отношению к полной последовательности аминокислот, приведенной под номером SEQ ID No:2, SEQ ID No:4, SEQ ID No:6 или SEQ ID No:8, при условии, что активность белка SDH1/ SDH2/ SDH3/ SDH4, не подверженного модификации согласно настоящему изобретению, сохраняется.
Гомология между двумя аминокислотными последовательностями может быть определена с помощью хорошо известных методов, например компьютерной программы BLAST 2.0, рассчитывающей три параметра: счет (score), идентичность (identity) и сходство (similarity).
Более того, гены SDH1, SDH2, SDH3, и SDH4 могут быть вариантами, которые гибридизуются в жестких условиях с последовательностями нуклеотидов, приведенными под номерами SEQ ID No:1, SEQ ID No:3, SEQ ID No:5 и SEQ ID No:7 соответственно, или с зондами, которые могут быть приготовлены на основе этих последовательностей, при условии, что они кодируют функциональные белки. "Жесткие условия" означают условия, при которых специфический гибрид, например гибрид, обладающий гомологией не менее чем 70%, предпочтительно не менее чем 80%, более предпочтительно не менее 90%, еще более предпочтительно не менее 95% и наиболее предпочтительно не менее 98%, образуется, а неспецифический гибрид, например гибрид, обладающий гомологией меньшей, чем указано выше, не образуется. Например, примером жестких условий являются условия с отмывкой один или более раз, предпочтительно два или три раза при концентрации солей 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS при 60°C. Продолжительность отмывки зависит от типа мембраны, использующейся для блоттинга, и, как правило, должна соответствовать рекомендации производителя. Например, рекомендуемая продолжительность отмывки нейлоновой мембраны Hybond™ N+ (Amersham) в жестких условиях составляет 15 минут. Предпочтительно, чтобы отмывку проводили 2 или 3 раза. Длина зонда может быть выбрана в зависимости от условий гибридизации и составляет обычно от 100 до 1 тысячи оснований.
Активность сукцинатдегидрогеназы может быть снижена в дрожжах путем ослабления экспрессии одного или нескольких генов, выбранных из группы, состоящей из генов SDH1, SDH2, SDH3 и SDH4. Для того чтобы ослабить экспрессию гена дрожжи могут быть модифицированы таким образом, что клетки дрожжей содержат количество белка, кодируемого указанным геном, пониженное по сравнению с немодифицированными клетами, или клетки неспособны к синтезу белка, кодируемого этим геном. Такая модификация дрожжей может быть осуществлена путем изменения последовательности, регулирующей экспрессию гена, такой как промотор, последовательность Shine-Dalgarno (AD)h т.п. В дополнение, экспрессия гена SDH1, SDH2, SDH3 или SDH4 может быть ослаблена внесением мутаций в ген на хромосоме таким образом, что внутриклеточная активность белка, кодируемого этим геном, понижена в сравнении с немодифицированным штаммом. Такой мутацией в гене может быть замена одного или нескольких оснований, приводящих к аминокислотным заменам в белке, кодируемом этим геном, введение стоп-кодона, делеция одного или двух оснований, приводящих к сдвигу рамки считывания, вставка гена, кодирующего устойчивость к антибиотику, или делеция части или всего гена.
Например, следующие методы могут быть использованы для внесения мутаций генной рекомбинацией. Синтезируется мутантный ген, кодирующий мутантный белок, обладающий пониженной активностью, дрожжи трансформируются фрагментом ДНК, содержащим мутантный ген, и затем после замены природного гена в хромосоме мутантным геном путем гомологичной рекомбинации отбирают полученный штамм, содержащий мутантный ген, заместивший природный ген в хромосоме. Такой метод внесения сайт-специфических мутаций путем генной замены с использованием гомологичной рекомбинации разработан. В разделе Примеры, приведенном ниже, температурочувствительная мутация была внесена в ген SDH1 в хромосоме штамма Y.lipolytica Po1f (АТСС MYA-2613). Вследствие указанной мутации экспрессия гена SDH1 ослаблена и активность сукцинатдегидрогеназы снижена при повышенной температуре (32°C) в полученных штаммах Po1f (SDH1-ts-0134) и Po1f (SDH1-ts-2007).
Экспрессия генов SDH1, SDH2, SDH3, или SDH4 также может быть ослаблена путем инактивации гена. Фраза «инактивация гена» означает, что модифицированный ген кодирует полностью неактивный белок. Также возможно, что модифицированный фрагмент ДНК не способен к природной экспрессии гена из-за частичной или полной делеции гена, сдвига рамки считывания, введения мутаций или модификации регионов, прилегающих к гену, включающих последовательности, контролирующие экспрессию гена, такие как промотор, энхансер, аттенуатор, сайт связывания рибосомы и т.п..
Вышеупомянутая замена гена может быть произведена следующим образом. А именно, дрожжи трансформируют рекомбинантной ДНК, содержащей мутантный ген, для последующей рекомбинации между мутантным геном и соответствующим хромосомным геном. В этом случае в рекомбинантную ДНК вводят ген-маркер с определенной характеристикой, например ауксотрофия, для удобства работы с клеткой-хозяином. Более того, использование вышеупомянутой рекомбинантной ДНК в линейной форме, например расщепление ферментом рестрикции, и затем удаление участка, контролирующего репликацию в дрожжах, из этой рекомбинантной ДНК tc высокой эффективностью приводит к получению штамма, в котором указанная рекомбинантная ДНК интегрирована в хромосому. В Примерах, приведенных ниже ген SDH2 в хромосоме штамма Y.lipolytica Po1f (АТСС MYA-2613) был инактивирован с получением штамма Po1f(dSDH2), который продуцировал янтарную кислоту и у которого снижена активность сукцинатдегидрогеназы. Среди мутантов штамма Po1f(dSDH2) был выбран штамм Y.lipolytica VKPM Y-3314, обладающий наибольшей способностью к продукции янтарной кислоты.
Экспрессия гена может быть ослаблена или ген может быть также полностью инактивирован путем введения транспозона или IS-фактора в кодирующую часть гена (патент США 5175107), или традиционными методами, такими как мутагенная обработка с использованием УФ-излучения и обработка с помощью гуанидина (N-метил-N'-нитро-N-нитрозогуанидин) или метоксиламина.
Методы получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, выбора олигонуклеотидов в качестве праймеров и т.д. являются стандартными методами, хорошо известными специалисту в данной области техники. Такие методы описаны, например, в книге Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989). Для трансформации дрожжей могут быть использованы традиционные методы, такие как метод протопластов, электропорация и другие.
2. Способ согласно настоящему изобретению
Способом согласно настоящему изобретению является способ получения янтарной кислоты, включающий выращивание дрожжей согласно настоящему изобретению в питательной среде с целью продукции и экскреции янтарной кислоты в среду и выделения янтарной кислоты из культуральной жидкости.
В настоящем изобретении выращивание, выделение и очистка янтарной кислоты могут быть осуществлены способом, традиционно использующимся для получения янтарной кислоты методом ферментации с использованием микроорганизмов.
Среда, используемая для выращивания, может быть как синтетической, так и природной при условии, что такая среда содержит источники углерода и азота, минералы и, если необходимо, подходящее количество питательных веществ, необходимых дрожжам для роста. Источники углерода включают различные углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от вида ассимиляции используемого микроорганизма могут быть использованы спирты, такие как этанол и глицерин. В качестве источников азота могут быть использованы различные соли аммония, такие как сульфат аммония, водный аммиак, другие азотистые соединения, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минералов используют фосфат калия, сульфат магния, хлорид натрия сульфат железа, сульфат марганца, хлорид кальция и подобные соединения. В качестве витаминов используют тиамин, дрожжевой экстракт и другие соединения.
Выращивание предпочтительно проводят в аэробных условиях, таких как культивирование с перемешиванием и перемешивание с аэрацией, при температуре от 20 до 40°C, предпочтительно от 24 до 32°C. рН среды обычно поддерживают в районе от 2 до 9, предпочтительно от 3 до 7.5. рН среды поддерживают добавлением аммония, карбоната кальция, различными основаниями и буферами. Также, выращивание может проводиться без контроля за рН, и после того, как рН среды снизится, выращивание может быть продолжено в кислых условиях, в которых и происходит основное накопление янтарной кислоты. Предпочтительно, чтобы по крайней мере часть процесса проводилась при рН меньше 4 для более высокой продукции и накопления янтарной кислоты. Обычно выращивание в течение от 1 до 7 дней приводит к накоплению янтарной кислоты в питательной среде.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости центрифугированием или фильтрацией через мембрану, а затем янтарная кислота может быть выделена и очищена с помощью ионообменной хроматографии, концентрированием и/или методом перекристаллизации.
Далее, после получения янтарной кислоты способом согласно настоящему изобретению, описанным выше, может быть проведена реакция полимеризации с использованием полученной янтарной кислоты в качестве сырья для получения полимера, содержащего янтарную кислоту. Несмотря на то, что в последние годы увеличилось количество промышленных продуктов, полученных без нанесения вреда окружающей среде, полимеры, полученные с использованием растительного сырья, все равно обращают на себя особое внимание. Янтарная кислота, полученная согласно настоящему изобретению, может быть использована при производстве таких полимеров, как полиэфирные и полиамидные полимеры. Частными примерами полимера, содержащего янтарную кислоту, являются полиэфир янтарной кислоты, полученный сополимеризацией диолов, таких как бутандиол или этиленгликоль, и янтарной кислоты, и полиамид янтарной кислоты, полученный сополимеризацией диаминов, таких как гескаметилендиамин, и янтарной кислоты. Также, янтарная кислота и композиции, содержащие янтарную кислоту, полученную согласно настоящему изобретению, могут быть использованы в пищевых добавках, фармацевтике, косметике и других областях.
Краткое описание чертежей
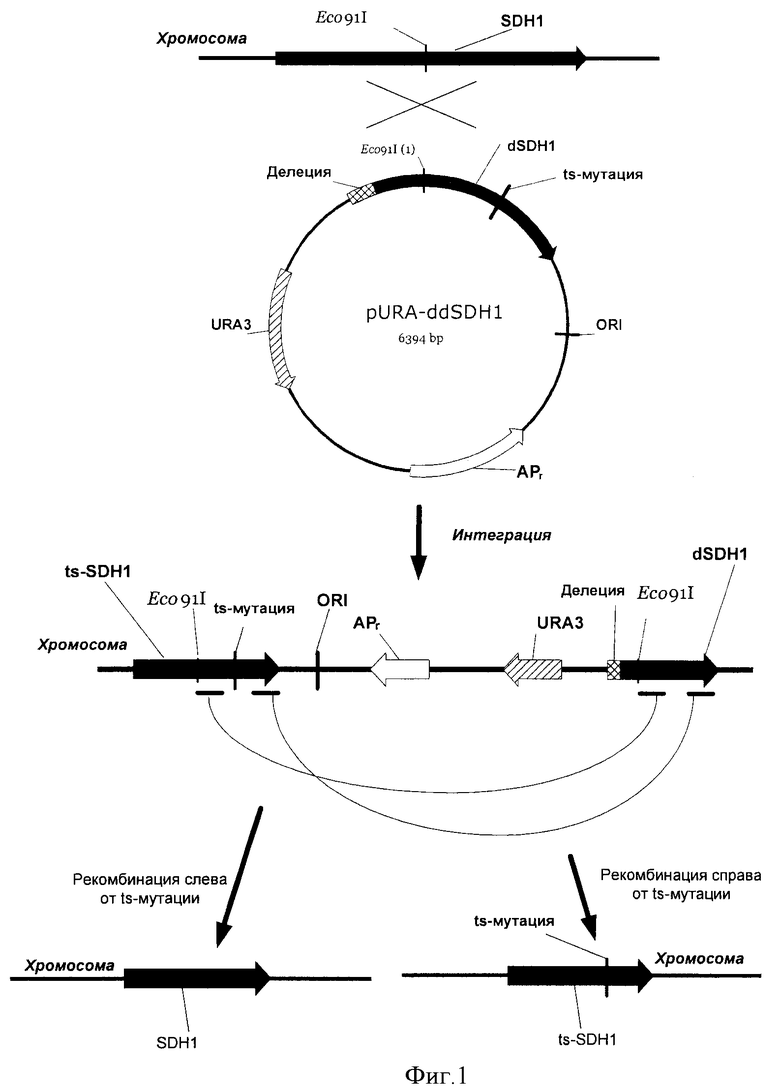
На Фиг.1 показана схема конструирования штамма, содержащего температурочувствительную мутацию (ts-) в гене SDH1.
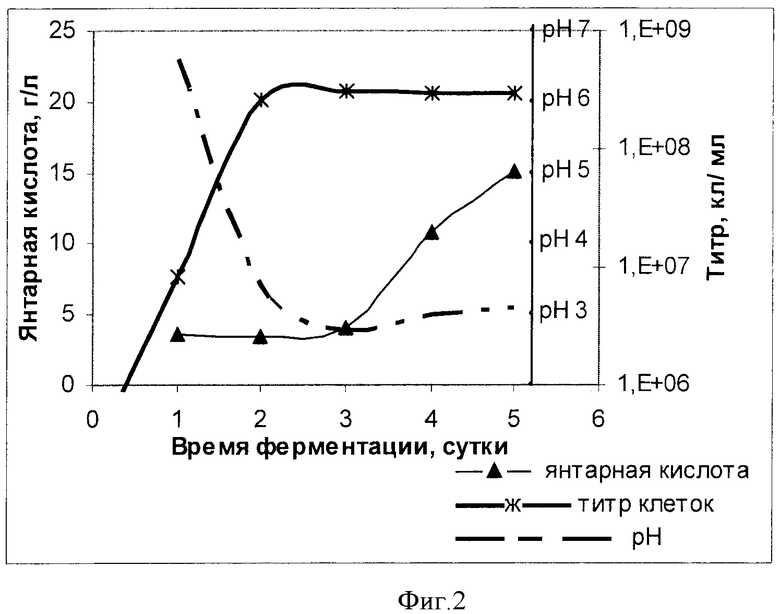
На Фиг.2 показаны кривые роста, рН и концентрация янтарной кислоты в процессе выращивания штамма Y.lipolytica VKPM Y-3314.
Примеры
Настоящее изобретение более подробно описано ниже со ссылкой на последующие Примеры, которые не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Конструирование штамма, содержащего температурочувствительную мутацию (ts-) в гене SDH1
1. Конструирование плазмиды pURA-ddSDH1
Сначала с помощью ПЦР получали фрагмент ДНК, содержащий ген URA3. Использовали праймеры P1(SEQ ID NO:9) и P2(SEQ ID NO:10). Тотальную геномную ДНК выделяли из штамма Y.lipolytica W29 (CLIB (Collection de Levures d'Interet Biotechnologique, Thiverval Grignon, France (Коллекция дрожжей биотехнологического применения), номер CLIB89) методом, описанным Kaiser et., al. (1994 Methods in Yeast Genetics. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory), и использовали в качестве матрицы в ПЦР.
Продукт ПЦР длиной 1972 пар оснований был получен и очищен в 1% агарозном геле. Продукт ПЦР (0.5 мкг) лигировали в вектор pUC19 (0,2 мкг), предварительно обработанный эндонуклеазой Ecl136II. Полученную плазмиду использовали для электропорации штамма Е.coli XL-1 (Blue) (Stratagene, каталожный номер #200228) как описано в Sambrook et al. (Molecular Cloning, second edition, Cold Spring Harbor Laboratory, Plainview, N.Y. (1989)). Клоны, содержащие целевую вставку, отбирали на чашках с L-агаром, содержащим 100 мкг/мл ампициллина, и проверяли на отсутствие активности β-галактозидазы. Плазмидную ДНК выделяли из отобранных клонов и проверяли с помощью рестрикционного анализа. Полученная плазмида (4658 пар оснований) содержала ген URA3 из Y.lipolytica и его регуляторные элементы. Плазмиду назвали pUC-URA3.
Для получения фрагмента ДНК, содержащего ген SDH1 с делецией, с помощью ПЦР синтезировали два фрагмента ДНК. ПЦР проводили с использованием тотальной геномной ДНК из штамма Y.lipolytica W29 в качестве матрицы и праймеров P3(SEQ ID NO:11), P4(SEQ ID NO:12) и P5(SEQ ID NO:13), P6(SEQ ID NO:14) с получением фрагментов ДНК размером 1223 пар оснований и 1768 пар оснований соответственно. Для ПЦР использовали Pfu полимеразу. Полученные продукты ПЦР очищали в 1% агарозном геле, а затем полученные продукты (0.05 мкг каждого) использовали в качестве матриц для ПЦР с Pfu полимеразой и праймерами Р3 и Р6. Продукт ПЦР размером 2968 пар оснований был получен, продукт очищали в 1% агарозном геле. Очищенный ПЦР продукт (0.5 мкг) обрабатывали рестриктазами XbaI и HindIII, затем лигировали с плазмидой pUC-URA3, предварительно обработанной этими же рестриктазами XbaI и HindIII с последующим дефосфорилированием с использованием фосфатазы CIAP. Полученную плазмиду использовали для электропорации штамма Е.coli XL-1 (Blue). Трансформанты отбирали на чашках с L-агаром, содержащим 100 мкг/мл ампициллина. Плазмидную ДНК выделяли из выбранных клонов. Полученная плазмида (7608 пар оснований) содержала ген SDH1 из Y.lipolytica с делецией 195 пар оснований и регуляторной области. Плазмиду назвали pUC-URA3-dSDH1.
Плазмиду pUC-URA3-dSDH1 (0,05 мкг) обрабатывали эндонуклеазами XbaI и XmaJI. Больший фрагмент очищали в 1% агарозном геле. Полученный фрагмент ДНК (6394 пар оснований), обработанный лигазой Т4, использовали для электропорации штамма Е.coli XL-1 (Blue). Трансформанты отбирали на чашках с L-агаром, содержащим 100 мкг/мл ампициллина. Плазмидную ДНК выделяли из отобранных клонов и проверяли с помощью рестрикционного анализа. Полученная плазмида (6394 пар оснований) содержала большую часть ОРС гена SDH1 из Y.lipolytica и его терминатор. Плазмиду назвали pURA-ddSDH1.
2. Конструирование штаммов Po1f/pURA-ddSDH-ts № 2007 и Po1f/pURA-ddSDH-ts № 0134.
Плазмиду pURA-ddSDH1 (15 мкг) обрабатывали in vitro метоксиламином как описано в Kadonaga J.T and Knowles J.R., N.A.R., 13(5), 1733-45 (1985). После диализа ДНК осаждали из раствора этанолом, промывали и использовали для электропорации штамма Е.coli XL-1 (Blue). Трансформанты отбирали на чашках с L-агаром, содержащим 100 мкг/мл ампициллина. 107 трансформантов выращивали в течение ночи в 200 мл среды LB, содержащей ампициллин (100 мкг /мл). Выросшие культуры использовали для выделения ДНК. Полученную библиотеку мутантных плазмид pURA-ddSDH1 обрабатывали эндонуклеазой Eco91I и использовали для трансформации штамма Y.lipolytica Po1f (АТСС MYA-2613) как описано в Current Genetic, 1989, vol 16, рр.253-260. Трансформанты отбирали на чашках с минимальной агаризованной средой YNB, содержащей глюкозу (2%), лейцин (0.01%) и казаминовые кислоты (0.2%). 3000 трансформантов выращивали в течение 3-х дней в жидкой минимальной среде YNB, содержащей глюкозу (0.1%), лейцин (0.01%) сукцинат натрия (2%) при 20°C, рН 6.8 с аэрацией на качалке (1200 об/мин). Выросшие клоны помещали на чашки (каждый клон на две разные чашки) с агаризованной средой YNB, содержащей лейцин (0.01%) и сукцинат натрия (1%), рН 6,8. Две серии чашек параллельно выращивали в течение 3-х дней при 20°C и 32°C. Клоны, росшие существенно лучше при 20°C, чем при 32°C проверяли таким же способом еще один раз для того, чтобы подтвердить их чувствительность к температуре. Как видно из Фиг.1, после интеграции плазмиды pURA-ddSDH в хромосому только левая часть гена SDH1 может экспрессироваться. Часть этой копии (справа от сайта Eco91I) присутствует в плазмиде и, следовательно, содержит некие мутации. Таким образом, полученные трансформанты соответствуют библиотеке мутантного гена SDH1.
Для подтверждения интеграции плазмиды pURA-ddSDH в правильное место хромосомы проводили ПЦР с использованием Taq-полимеразы. Для ПЦР использовали локус-специфические праймеры Р7 (SEQ ID NO:15) и Р8 (SEQ ID NO:16) и тотальную геномную ДНК, выделенную из отобранных трансформантов. В результате для дальнейших исследований выбрали два трансформанта: Po1f/pURA-ddSDH-ts № 0134 и Po1f/pURA-ddSDH-ts №2007, содержащие мутации Ser675Phe и Glu483Lys соответственно в гене SDH1. Указанные штаммы назвали Po1f (SDHl-ts-0134) и Po1f (SDH1-ts-2007) соответственно.
Пример 2. Конструирование штамма с инактивированным геном SDH2
Сначала получали три фрагмента ДНК, содержащих промоторную область гена SDH2 (1125 пар оснований), ген URA3 (1229 пар оснований) и терминаторную область гена SDH2 (1018 пар оснований), с помощью ПЦР с использованием Pfu полимеразы, тотальной геномной ДНК штамма Y.lipolytica W29 (АТСС 20460) и набора праймеров Р9 (SEQ ID NO:17) и P10 (SEQ ID NO:18), P11 (SEQ ID NO:19) и P12 (SEQ ID NO:20), и P13 (SEQ ID NO:21) и P14 (SEQ ID NO:22) соответственно. Полученные продукты ПЦР очищали в 1% агарозном геле. Продукты ПЦР, содержащие промоторную область гена SDH2 (1125 пар оснований), ген URA3 (1229 пар оснований), использовали в качестве матрицы для ПЦР с Pfu полимеразой и праймерами Р9 и Р12. Полученный продукт ПЦР (2333 пар оснований) очищали в 1% агарозном геле. Затем этот продукт ПЦР (2333 пары оснований) и продукт ПЦР, содержащий терминаторную область гена SDH2 (1018 пар оснований), использовали в качестве матрицы для ПЦР с Pfu полимеразой и праймерами Р9 и Р14. Полученный продукт ПЦР (3330 пар оснований) очищали в 1% агарозном геле и 1 мкг полученного продукта ПЦР использовали для трансформации штамма Y.lipolytica Po1f с помощью метода, использующего ацетат лития. Трансформанты отбирали на минимальной агаризованной среде YNB, содержащей глицерин (5% v/v), лейцин (0.01%) и казаминовые кислоты (0.2%). Отобранные трансформанты проверяли на способность к росту в питательной среде, содержащей сукцинат в качестве единственного источника углерода. Наличие вставки в локус SDH2 проверяли с помощью ПЦР. Для ПЦР использовали Taq-полимеразу, тотальную геномную ДНК, выделенную из трансформантов, в качестве матрицы, и локус-специфические праймеры Р15 (SEQ ID NO:23) и Р16 (SEQ ID NO:24). Один трансформант, названный Po1f(dSDH2), выбрали для дальнейших исследований.
Пример 3. Получение янтарной кислоты штаммами Y.lipolytica Po1f(SDH1-ts-0134), Po1f(SDH1-ts-2007) и Po1f(dSDH2)
Для проверки эффекта ослабления экспрессии генов SDH1 (YALI0D11374g) или SDH2 (YALI0D23397g) на продукцию сукцината сравнивали продуктивность штаммов Po1f(SDH1-ts-0134), Po1f(SDH1-ts-2007), Po1f(dSDH2) и Po1f.
Штаммы Po1f(SDH1-ts-0134), Po1f(SDH1-ts-2007) и Po1f (наальный титр клеток - 1-5·105 клеток/мл) выращивали в 5 мл минимальной среды YNB (Himedia), содержащей глицерин (5%(v/v)), урацил и лейцин (0.01%), рН 6.8 (поддерживали с использованием 50 мМ фосфатного буфера) в пробирках объемом 50 мл при 24°C. После 2-х дней выращивания в среду добавляли 2,5% CaCO3 для поддержания рН. Когда титр клеток достиг уровня (0.8-1)·107 несколько пробирок поместили в температуре 32°C, остальные оставили при 24°C. После 5 дней выращивания с помощью ВЭЖХ определяли количества янтарной кислоты, α-кетоглутаровой кислоты, лимонной кислоты, пирувиновой кислоты и уксусной кислоты, накопленных в культуральной жидкости, а также концентрацию глицерина. Результаты двух независимых пробирочных ферментации приведены в Таблице 1. Как видно из Таблицы 1, Po1f(SDH1-ts-0134), Po1f(SDH1-ts-2007) продуцировали большее количество янтарной кислоты в условиях, когда температура, при которой выращивали дрожжи, была изменена до 32°C. В таких же условиях штамм Po1f продуцировал только незначительные количества янтарной кислоты.
Штаммы Po1f(dSDH2) и Po1f (105 клеток/мл) выращивали в 10 мл минимальной среды YNB (Himedia), содержащей глицерин (5%(v/v)), лейцин (0.01%), урацил (0.01%) (только для штамма Po1f), рН 6.8 (поддерживали 50 мМ фосфатным буфером) в пробирках объемом 50 мл при 30°C. После 3-х дней выращивания в среду добавляли 2.5% СаСО3 для поддержания рН. После 7 дней выращивания с помощью ВЭЖХ определяли количества янтарной кислоты, α-кетоглутаровой кислоты, лимонной кислоты, пирувиновой кислоты и уксусной кислоты, накопленных в культуральной жидкости, а также концентрацию глицерина. Результаты пробирочных ферментаций приведены в Таблице 1. Как видно из Таблицы 1, штамм Po1f (dSDH2) продуцировал большее количество янтарной кислоты по сравнению со штаммом Po1f.
Штаммы Po1f(dSDH2) и Po1f выращивали в 50 мл среды YPG (1% дрожжевого экстракта, 2% пептона, 5%(v/v) глицерина) в колбах объемом 750 мл при 30°C. После 2 дней выращивания добавляли 2,5% CaCO3. После 5 дней выращивания с помощью ВЭЖХ определяли количества янтарной кислоты, α-кетоглутаровой кислоты, лимонной кислоты, пирувиновой кислоты и уксусной кислоты, накопленных в культуральной жидкости, а также концентрацию глицерина. Результаты ферментации в колбах приведены в Таблице 2. Как видно из Таблицы 2, штамм Po1f(dSDH2) продуцировал большее количество янтарной кислоты по сравнению со штаммом Po1f.
Пример 4. Отбор штамма Y.lipolytica VKPM Y-3314
Для улучшения ростовых характеристик штамма Po1f(dSDH2) проводили мутагенез следующим образом. Клетки штамма Po1f (dSDH2) выращивали в среде YPG (см. Пример 3) до достижения титра 109 клеток/мл, клетки осаждали центрифугированием, три раза промывали физиологическим раствором и ресуспендировали в 50 мМ калий фосфатном буфере (рН 6.8) до титра 1-3·108 клеток/мл. 1 мл полученной суспензии обрабатывали N-метил-N-нитро-N-нитрозогуанидином (40 мкг/мл) в течение 2 часов при 30°C. Затем клетки три раза промывали физиологическим раствором и наносили (титр клеток 108 или 109 клеток/мл) на чашки со средой YNB, содержащей лейцин (0,1 г/л) и глицерин (2% v/v). Процент выживших клеток составил 2.8%. После 5 дней выращивания среди всех колоний нормального размера появились колонии большого размера (1-2 из 104). Выбрали 50 колоний большого размера и поместили на такую же среду.
Штаммы всех 50 выбранных колоний и штамм Po1f(dSDH2) в качестве контроля выращивали в 5 мл среды YPG до получения титра клеток ~5·105 клеток/мл. После 7 дней выращивания при 30°C без использования буфера и добавления CaCO3 с помощью ВЭЖХ определяли количества янтарной кислоты, α-кетоглутаровой кислоты, лимонной кислоты, пирувиновой кислоты и уксусной кислоты, накопленных в культуральной жидкости. Результаты пробирочных ферментаций приведены в Таблице 3. Как видно из Таблицы 3, подавляющее большинство мутантнов продуцировали большее количество янтарной кислоты по сравнению со штаммами Po1f(dSDH2) и Po1f. Было обнаружено, что значение рН в культуральной жидкости достигало значения 3.2-3.5 после 3-4 дней выращивания. Наилучший продуцент янтарной кислоты мутантный штамм No. 18 выбрали для дальнейших исследований. Указанный штамм депонировали во Всероссийской коллекции промышленных микроорганизмов (Россия, 117545 Москва, 1-й проезд, 1) 15 ноября 2008 под номером ВКПМ Y-3314.
Пример 5. Получение янтарной кислоты с использованием штамма Y.lipolytica ВКПМ Y-3314.
Штамм Y.lipolytica ВКПМ Y-3314 выращивали в пробирках объемом 50 мл в 5 мл среды YPG, содержащей глицерин (5%,v/v) различных производителей. Начальный титр клеток составлял ~5·105 клеток/мл. Первую серию экспериментов проводили с добавлением CaCO3 (2.5%) на 3-й день выращивания. Вторую серию экспериментов проводили без добавления CaCO3. Результаты измерений pH среды, концентраций органических кислот и остаточного глицерина приведены в Таблице 4 для первой серии экспериментов и в Таблице 5 для второй серии экспериментов. Как видно из Таблиц 4 и 5, штамм Y.lipolytica ВКПМ Y-3314 продуцировал янтарную кислоту в обеих сериях экспериментов с глицерином от различных производителей.
Штамм Y.lipolytica ВКПМ Y-3314 также выращивали в колбах объемом 750 мл в 50 мл среды YPG, содержащей глицерин (5%,v/v), начальный титр клеток составлял ~2,5·105 клеток/мл. Каждые 24 часа в ходе выращивания отбирали образцы и определяли титр клеток, pH среды и коццентрацию янтарной кислоты. Результаты измерений приведены на Фиг.2. Как видно из Фиг.2, клетки штамма Y.lipolytica ВКПМ Y-3314 продуцировали янтарную кислоту при низких значениях pH, ниже рН4.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.













| название | год | авторы | номер документа |
|---|---|---|---|
| ИНТЕГРАТИВНАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pQ-SRUS ДЛЯ ПОЛУЧЕНИЯ ШТАММОВ ДРОЖЖЕЙ Yarrowia lipolytica, ОБЛАДАЮЩИХ СПОСОБНОСТЬЮ К ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ ГЕНОМА МИТОХОНДРИЙ ЗА СЧЁТ ЭКСПРЕССИИ ГЕНА БЕЛКА RecA БАКТЕРИАЛЬНОГО ПРОИСХОЖДЕНИЯ | 2014 |
|
RU2562869C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica ВКПМ Y-3753 - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ | 2012 |
|
RU2487931C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ КЛЕТОЧНО-СВЯЗАННОЙ ЛИПАЗЫ | 2012 |
|
RU2475532C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ЭКСПОЗИЦИИ БЕЛКА НА ПОВЕРХНОСТИ КЛЕТОЧНОЙ СТЕНКИ ДРОЖЖЕЙ Yarrowia lipolytica | 2010 |
|
RU2451749C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИНОЗИНА И ИНОЗИН 5'-МОНОФОСФАТА, ШТАММ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ BACILLUS - ПРОДУЦЕНТ ИНОЗИНА (ВАРИАНТЫ) | 2002 |
|
RU2260040C2 |
| Модифицированная нуклеотидная последовательность, кодирующая полипептид, обладающий активностью фитоен синтазы и геранилгеранилпирофосфат синтазы (варианты), ее использование при конструировании трансформантов дрожжей Yarrowia lipolytica, продуцирующих каротиноиды | 2021 |
|
RU2794980C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПУРИНОВЫХ РИБОНУКЛЕОЗИДОВ И РИБОНУКЛЕОТИДОВ | 2008 |
|
RU2422510C2 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
| МУТАНТНАЯ ФОСФОРИБОЗИЛПИРОФОСФАТСИНТЕТАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, СПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОЗИДОВ И CПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОТИДОВ | 2008 |
|
RU2403286C2 |
| СПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОЗИДОВ И НУКЛЕОТИДОВ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ Escherichia ИЛИ Bacillus | 2006 |
|
RU2365622C2 |
Изобретение относится к биотехнологии и генной инженерии и касается способа получения янтарной кислоты с использованием штамма дрожжей, Yarrowia lipolytica ВКПМ Y-3314. Данные дрожжи модифицированы таким образом, что у них снижена активность сукцинатдегидрогеназы. Способ включает стадию выращивания штамма дрожжей в питательной среде, содержащей глицерин, и выделения янтарной кислоты из культуральной жидкости. Использование штамма Yarrowia lipolytica ВКПМ Y-3314 привело к существенному увеличению продукции янтарной кислоты этим способом. 1 з.п. ф-лы, 2 ил., 5 табл.
1. Способ получения янтарной кислоты, включающий стадии выращивания штамма дрожжей Yarrowia lipolytica, обладающего способностью к продукции янтарной кислоты, в питательной среде, содержащей глицерин, и выделения янтарной кислоты из культуральной жидкости, отличающийся тем, что используют штамм дрожжей Yarrowia liроlуtiса ВКПМ Y-3314.
2. Способ по п.1, отличающийся тем, что, по крайней мере, часть процесса выращивания дрожжей осуществляют при рН ниже 4.
| WO 2009011974, 22.01.2009 | |||
| AHLERS.E.ET AL | |||
| Effects of a temperature-sensitive mutation in the immediate-early gene of pseudorabies virus on class II and class III gene transcription | |||
| Journal of Virology, 1987, v.61 | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| KAMZOLOVA S.V | |||
| ET AL | |||
| Chemically assisted microbial production of succinic acid by the yeast Yarrowia lipolytica | |||

