Область техники, к которой относится изобретение
Данное изобретение относится к новым сериновым протеазам, родственным дипептидилпептидазе IV (DPPIV), и к выделенным нуклеиновым кислотам, кодирующим эти протеазы, все из которых применимы для обнаружения новых терапевтическиих средств, для измерения протеазной активности и для определения ингибиторной активности соединений против этих протеаз.
Уровень техники
Протеазы и пептидазы являются ферментами, которые катализируют гидролиз пептидных амидных связей. Протеазы играют важную роль в регуляции биологических процессов в почти любой форме жизни от бактерий до вирусов и до млекопитающих. Они выполняют критические функции, например, в пищеварении, свертывании крови, апоптозе, активации иммунных реакций, активации зимогенов, созревании вирусов, секреции белков и транспорте белков. Они могут быть классифицированы в соответствии с рядом критериев, таких как сайт действия, предпочтительность субстратов и механизм действия. Так, например, аминопептидазы действуют преимущественно на N-концевые остатки пептида, тогда как карбоксипептидазы действуют преимущественно на С-конце и эндопептидазы действуют по сайтам, удаленным от этих двух концов. Среди карбокси- и аминопептидаз, пептидилпептидазы отщепляют единственный аминокислотный остаток от субстрата, дипептидилпептидазы отщепляют дипептидную единицу (две аминокислоты) от субстрата и трипептидазы отщепляют три аминокислоты от субстрата. Предпочтительность субстрата часто выражается в терминах аминокислотного остатка, расположенного непосредственно в N-концевом направлении относительно сайта расщепления. Например, трипсиноподобные пептидазы будут предпочтительно расщеплять пептид по соседству с основной аминокислотой (аргинином или лизином), т.е. где гидролизуемая связь является связью Arg/Lys-Xaa. В качестве другого примера, семейство химотрипсиноподобных пептидаз преимущественно гидролизует пептиды по соседству с ароматическим остатком. Механистически, пептидазы классифицируются как серин-зависимые, цистеин-зависимые, аспаргиновая кислота-зависимые или цинк-зависимые.
Поскольку пептидазы и протеазы участвуют в регуляции многих физиологических процессов, они являются привлекательными мишенями для разработки терапевтических средств. Ингибиторы протеаз и пептидаз применимы, например, в лечении гипертонии, нарушений свертывания крови и вирусной инфекции.
Протеолитические ферменты, которые используют серин для своей каталитической активности, встречаются повсеместно, будучи обнаруживаемыми в вирусах, бактериях и эукариотах. Свыше 20 семейств (обозначаемых S1-S27) сериновых протеаз были идентифицированы; они сгруппированы в 6 кланов (SA, SB, SC, SE, SF и SG) на основе структурного сходства и других функциональных признаков. Структуры известны для четырех из этих кланов (SA, SB, SC и SE); они являются, по-видимому, совершенно неродственными, что предполагает по меньшей мере четыре эволюционных пути происхождения сериновых пептидаз и, возможно, гораздо большего их числа (Rawlings and Barret, Meth. Enzymol. 244:19-61 (1994)).
Семейство пролилолигопептидаз состоит из ряда эволюционно родственных пептидаз, каталитическая активность которых, по-видимому, обеспечивается системой переноса заряда, сходной с системой переноса заряда трипсинового семейства сериновых протеаз, которая, однако, развилась в результате независимой конвергентной эволюции. Экспериментально было показано, что консервативный остаток серина (в протеазе II Е. coli, а также в свиной и бактериальной РЕ) является необходимым в каталитическом механизме. Этот серин, который является частью каталитической триады (Ser, His, Asp), обычно расположен на расстоянии 150 остатков от С-конца этих ферментов (все из которых являются белками, содержащими около 700-800 аминокислот).
Одной из наиболее интенсивно исследуемых пролилолигопептидаз является дипептидилпептидаза IV (DPPIV, ЕС 3.414.5), гликопротеин типа II, которая является единственной хорошо охарактеризованной аминопептидазой, которая, как известно, локализована на наружной стороне плазматических мембран. Как указано выше, дипептидиламинопептидазы характеризуются их способностью отщеплять N-концевые дипептиды от различных небольших пептидов. Дипептидиламинопептидазы обладают различной субстратной специфичностью и клеточной локализацией, что предполагает различные функции каждой активности в процессинге пептидов. DPPIV характеризуется ее способностью отщеплять N-концевые дипептиды, содержащие пролин или аланин в качестве предпоследнего остатка. Ген DPPIV простирается на приблизительно 70 т.п.н. и содержит 26 экзонов в диапазоне размеров от 45 п.н. до 1,4 т.п.н. Нуклеотидная последовательность (3465 п.н.) кДНК содержит открытую рамку считывания, кодирующую полипептид, содержащий 766 аминокислот. Нуклеотиды, которые кодируют последовательность активного центра (G-W-S-Y-G), распределены между 2 экзонами. Это явно отличает геномную организацию семейства пролилолигопептидаз от геномной организации семейства классических сериновых протеаз.
DPPIV широко распространена в тканях млекопитающих и в большом количестве представлена в почках, эпителии кишечника и плаценте (Yaron, A. and Naider, F., Critical Reviews in Biochem. Mol. Biol. 1993 [1], 31). В иммунной системе человека этот фермент экспрессируется исключительно активированными Т-лимфоцитами CD4+-типа, где этот фермент, как было показано, является синонимом антигена клеточной поверхности CD26. Хотя точная роль DP-IV в физиологии человека все еще не выяснена полностью, недавнее исследование показало, что этот фермент явно играет важную роль в физиологии и патофизиологии человека.
На Т-клетках человека экспрессия DPPIV появляются на поздней стадии дифференцировки тимуса и ограничена преимущественно популяцией клеток CD4+ хелперов/памяти, a CD26 может доставлять мощный сигнал костимуляторной активации Т-клеток. Таким образом, фермент DPPIV, также известный как антиген CD26 активации Т-клеток, играет важную роль в иммунной реакции вследствие ассоциации с CD45 тирозинфосфатазой и, вследствие его способности связывать аденозиндезаминазу (ADA) с поверхностью Т-клетки, защищает Т-клетку от опосредованного аденозином ингибирования пролиферации. Кроме того, регуляция функции хемокинов CD26/DPPIV, по-видимому, является существенной для перемещения лимфоцитов и инфекционности штаммов ВИЧ. DPPIV связана с многочисленными функциями, включающими в себя участие в активации Т-клеток, адгезию клеток, расщепление пролинсодержащих пептидов в почках и кишечнике, инфекцию ВИЧ и апоптоз и регуляцию онкогенности в некоторых меланомных клетках (Pethiyagoda et al., Clin. Exp.Metastasis 2000: 18(5):391-400). Предполагается также, что DPPIV участвует в эндокринной регуляции и метаболической физиологии. Более конкретно, DPPIV отщепляет амино-концевой дипептид His-Ala от GLP-1, приводя к появлению антагониста GLP-1-рецептора, и, следовательно, снижает физиологическую реакцию на GLP-1. Глюкагоноподобный пептид (GLP-1), инкретин, который индуцирует зависимую от глюкозы секрецию инсулина, быстро разрушается DPPIV, и, поскольку полупериод расщепления DPPIV в организме является гораздо более коротким, чем полупериод для удаления GLP-1 из кровотока, существенное увеличение биологической активности GLP-1 (в 5-10 раз) ожидается в результате ингибирования DPPIV. Ингибиторы DPPIV исследуются в настоящее время в клинике в качестве потенциальных терапевтических средств для диабета типа 2 и нарушенной толерантности к глюкозе.
Различные ингибиторы DPPIV были известны в 1993 году. Одним из них является суицидный ингибитор N-Ala-Pro-O-(нитробензоил)гидроксиламин. Другим ингибитором является конкурентный ингибитор е-(4-нитро)бензоксикарбонил-Les-Pro и еще одним ингибитором является поликлональный кроличий иммуноглобулин против DPPIV почки свиньи. С тех пор были разработаны и другие и они описаны подробно в патентах США № 5939560; 6110949 6011155 и 5462928.
Помимо его каталитической активности серинового типа, но независимо от нее, фермент DPPIV прочно связывается с растворимым внеклеточным ферментом аденозиндезаминазой (ADA), действуя в качестве рецептора и, как предполагают, опосредует трансдукцию сигнала. Структура DPPIV характеризуется двумя внеклеточными доменами, α/β-складчатым гидролазным доменом и 7-лопастным бета-пропеллерным доменом, состоящим из повторяющихся бета-складок из приблизительно 50 аминокислот. Недавно было показано, что, наряду с селекцией субстратов по размеру, бета-пропеллерный домен, содержащий 10 из 12 высококонсервативных остатков цистеина, вносит вклад в катализ, осуществляемый пептидазным доменом. Кроме того, этот богатый цистеином домен является ответственным за связывание DPPIV с коллагеном I и с внеклеточной ADA. Сообщалось также, что DPPIV играет роль в опосредованных фибронектином взаимодействиях клеток с внеклеточным матриксом. Недавние исследования показали, что протеазная активность DPPIV не требуется для его антиинвазивной активности, так как мутанты DPPIV, которые не имеют внеклеточной активности сериновой протеазы, сохраняют такую активность.
Ряд белков, которые имеют сходство с DPPIV, были описаны в литературе. Несколько из этих белков клонировали, в том числе DPP-I, DPP-II, DPP-III, DPP-X и белок активации фибробластов (FAP). Они были идентифицированы и охарактеризованы либо молекулярным клонированием и функциональными исследованиями экспрессированных белков, либо по биохимическим активностям в экстрактах тканей. DPPIV-бета и другие новые пептидазы с функциональным сходством с DPPIV еще не были клонированы. Идентификация, характеристика и/или подходящая классификация дополнительных членов семейства пролилолигопептидаз, выяснение их физиологической (и, в особенности, патофизиологической роли) и применение этого знания для разработки новых терапевтических средств являются важными задачами.
Сущность изобретения
Данное изобретение обеспечивает белки с пролилолигопептидазными (расщепление после пролина) активностями, которые составляют три новых члена семейства белков, родственных DPPIV, включая полноразмерные белки, формы альтернативного сплайсинга, субъединицы и мутанты, а также кодирующие их нуклеотидные последовательности. Данное изобретение обеспечивает также способы скрининга, направленного на субстраты, взаимодействующие белки, агонисты, антагонисты или ингибиторы вышеуказанных белков и, кроме того, фармацевтические композиции, содержащие белки и/или их мутанты, производные и/или аналоги и/или лиганды к ним.
Эти новые белки, имеющие значительную гомологию последовательности с DPPIV, обозначены как белки 1, 2 и 3, родственные дипептидилпептидазе IV (DPRP-1, DPRP-2 и DPRP-3). Аминокислотные последовательности DPRP-1, DPRP-2 и DPRP-3 представлены последовательностями SEQ ID NO:1, 3 и 5, соответственно. Дополнительно описаны последовательности нуклеиновых кислот, кодирующие эти белки (SEQ ID NO:2, 4 и 6). Таблица 1 иллюстрирует гомологию (т.е. сходство) между новыми белками DPRP-1, DPRP-2 и DPRP-3 и другими известными сериновыми протеазами.
Наивысшая гомология между DPRP-1, DPRP-2 и DPPIV наблюдается в С-концевых последовательностях. На основе гомологии последовательности с DPPIV (см. фигуру 1) можно предсказать, что эти белки DPRP могут иметь функции, которые включают в себя функционирование в качестве ферментов, но не ограничиваются ими. Клонирование, экспрессия, биохимическая и молекулярная характеристика подтвердили эту гипотезу.
Профили экспрессии DPRP и локализация в специализированных эпителиальных клетках и плазматических клетках (клетках Лейдига, эпителиальных клетках предстательной железы, лимфоцитах, В-клетках) согласуются с ролью в дифференцировке, пролиферации и воспалении. Локализация гена DPRP-1 в чувствительных к гормонам раках (молочной железы, предстательной железы и яичек), тканях, регулируемых тестостероном, и обильная экспрессия в слабо дифференцированных раках демонстрируют, что DPRP-активирующие или ингибирующие молекулы будут иметь многочисленные терапевтические применения в лечении заболеваний, характеризующихся нарушенной регуляцией роста, дифференцировки и синтеза и деградации стероидных или полипептидных гормонов. Описанные здесь данные подтверждают гипотезу, что DPRP-1 и DPRP-2 участвуют в регуляции пролиферации в in vitro моделях рака предстательной железы и яичка, хорошо известных специалистам в данной области.
Активности DPRP-1 и DPRP-2, описанные здесь, и их профили экспрессии согласуются с тем, что они имеют функциональные роли в качестве физиологических регуляторов иммунной и нейроэндокринной систем, которые действуют посредством ферментативной модификации биохимических медиаторов, таких как пептиды и хемокины. Многочисленные функции, описанные ранее для DPPIV на основе применения ингибиторов, могут быть обусловлены отчасти его действием и действием родственных белков, подобных DPRP. Таким образом, обнаружение селективных и сильнодействующих ингибиторов DPPIV, DPRP и других родственных протеаз, подобных FAP, считается важнейшим для достижения эффективного и безопасного фармацевтического применения этих и любых недавно идентифицированных ингибиторов сериновых протеаз, а также других активных соединений, которые модифицируют функцию (функции) таких белков.
Таким образом, данное изобретение обеспечивает новые белки или полипептиды, кодирующие их нуклеиновые кислоты, клетки, которые были модифицированы такой нуклеиновой кислотой таким образом, что они экспрессируют эти белки, антитела к этим белкам, способ скрининга для обнаружения новых терапевтических средств, которые являются ингибиторами активности этих белков (или которые являются ингибиторами DPPIV и не являются ингибиторами этих белков), и терапевтические средства, обнаруженные такими способами скрининга. Новые белки и кодирующие их нуклеиновые кислоты могут быть использованы для обнаружения новых терапевтических средств для лечения определенных заболеваний, таких как, например, репродуктивные, воспалительные и метаболические нарушения, а также в получении антител с терапевтической или диагностической ценностью.
В соответствии с одним аспектом данного изобретения обеспечены новые, зрелые, биологически активные белки, в основном человеческого происхождения. Такие белки могут быть выделены в небольших количествах из ткани или биологических жидкостей подходящего животного (в том числе человека) стандартными способами; однако, большие количества более удобно получать в культурах клеток, генетически модифицированных таким образм, что они экспрессируют данный белок.
Согласно другому аспекту данного изобретения, обеспечиваются изолированные молекулы нуклеиновых кислот, кодирующие полипептиды данного изобретения, в том числе их мРНК, ДНК, кДНК, геномные ДНК.
Согласно следующему аспекту данного изобретения обеспечиваются также зонды на основе нуклеиновых кислот, содержащие молекулы нуклеиновых кислот достаточной длины для специфической гибридизации с последовательностью нуклеиновой кислоты данного изобретения.
Согласно еще одному аспекту данного изобретения обеспечиваются способы получения таких полипептидов на основе рекомбининтной технологии, применимые для научных исследований in vitro, например синтеза ДНК и получения ДНК-векторов. Способы получения таких полипептидов включают в себя культивирование рекомбинантных прокариотических и/или эукариотических клеток-хозяев, которые были трансфицированы ДНК-векторами, содержащими нуклеотидную последовательность, кодирующую такой полипептид и/или зрелый белок, в условиях, стимулирующих экспрессию такого белка, и последующее извлечение такого белка или фрагмента этого экспрессированного продукта.
Согласно еще одному аспекту данное изобретение обеспечивает способы применения полипептидов и полинуклеотидов DPRP, в том числе для лечения инфекций, таких как бактериальные, грибковые, вызываемые простейшими и вирусные инфекции, в частности инфекций, вызываемых ВИЧ-1 или ВИЧ-2, боли, диабета, преждевременного полового созревания, бесплодия, ожирения, анорексии, булимии, болезни Паркинсона, острой сердечной недостаточности, гипотонии, гипертонии, задержки мочи, остеопороза, стенокардии, инфаркта миокарда, инсульта, язв, астмы, аллергий, доброкачественной гипертрофии предстательной железы, раков, в том числе чувствительных к гормонам, и андроген-независимых раков, мигреней, рвоты, психотических и неврологических нарушений, в том числе тревожности, шизофрении, маниакальной депрессии, депрессии, слабоумия и тяжелой умственной отсталости и дискинезий, далее называемых общим термином "заболевания".
Согласно еще одному аспекту данного изобретения обеспечивается способ применения таких полипептидов или полинуклеотидов, кодирующих такие полипептиды, для обнаружения соединений, которые ингибируют биологическую активность этих зрелых белков, например, отщеплением N-концевого дипептида, и, следовательно, обеспечиваются также такие ингибиторы.
Согласно еще более конкретному аспекту данное изобретение обеспечивает изолированную нуклеиновую кислоту, которая кодирует: (а) полипептид, который включает в себя аминокислотную последовательность одной из последовательностей SEQ ID NO:1, 3 и 5, или (b) полипептид, имеющий аминокислотную последовательность, которая является на по меньшей мере 70% сходной с указанной аминокислотной последовательностью и проявляет ту же самую биологическую функцию или которая является вариантом альтернативного сплайсинга одной из последовательностей SEQ ID NO:2, 4 и 6, или которая является зондом, содержащим по меньшей мере 14 следующих друг за другом нуклеотидов из указанной нуклеиновой кислоты, кодирующей (а) или (b), или которая комплементарна любой из предыдущих нуклеотидных последовательностей.
Согласно другому конкретному аспекту данное изобретение обеспечивает полипептид, который может быть необязательно гликозилированным, и который: (а) имеет аминокислотную последовательность зрелого белка, представленную любой из последовательностей SEQ ID NO:1, 3 и 5; (b) имеет аминокислотную последовательность зрелого белка, имеющего по меньшей мере 70% сходство с одним из зрелых белков (а), и который проявляет ту же самую биологическую функцию; (с) имеет аминокислотную последовательность зрелого белка, имеющего по меньшей мере приблизительно 90% идентичность со зрелым белком любой из последовательностей SEQ ID NO:1, 3 и 5; или (d) является иммунологически реактивным фрагментом (а).
Согласно еще одному конкретному аспекту данное изобретение обеспечивает способ скрининга, направленного на соединение, способное ингибировать ферментативную активность по меньшей мере одного зрелого белка данного изобретения, причем этот способ включает инкубирование указанного зрелого белка и подходящего субстрата для указанного зрелого белка в присутствии одного или нескольких тестируемых соединений или их солей, измерение ферментативной активности указанного зрелого белка, сравнение указанной активности с активностью, определенной в отсутствие тестируемого соединения, и отбор тестируемого соединения или тестируемых соединений, которые уменьшают эту ферментативную активность, и оно обеспечивает также способ скрининга, направленного на соединение, способное ингибировать ферментативную активность DPPIV, которое не ингибирует ферментативную активность по меньшей мере одного зрелого белка изобретения, причем этот способ включает инкубирование указанного зрелого белка и подходящего субстрата в присутствии одного или нескольких ингибиторов DPPIV или их солей, измерения ферментативной активности указанного зрелого белка, сравнения указанной активности с активностью, определенной в отсутствие ингибитора DPPIV, и отбора соединения, которое не уменьшает ферментативную активность указанного зрелого белка.
Эти и другие аспекты данного изобретения должны быть очевидными для специалистов с квалификацией в данной области из следующего ниже подробного описания.
Перечень фигур
Фиг.1А и 1 В показывают линейное выравнивание DPRP-1, DPRP-2, DPRP-3 и DPPIV, причем затенение использовано для указания идентичных (черный цвет) или сходных (серый цвет) аминокислотных остатков в конкретном положении.
Фиг.2 сходна с Фиг.1 и показывает линейное выравнивание DPRP-2 человека и мыши.
Фиг.3 является графиком, который показывает действие различных тетрапептидных амидных ингибиторов на дипептидилпептидазную ферментативную активность.
Фиг.4а-4с показывают действие трех ингибиторных соединений на пролиферацию клеточных линий рака предстательной железы РС3 в различных дозах.
Сведения, подтверждающие возможность осуществления изобретения
Согласно аспекту данного изобретения обеспечиваются изолированные нуклеотидные последовательности (полинуклеотиды), которые кодируют зрелые полипептиды, имеющие предсказанные аминокислотные последовательности трех DPRP (SEQ ID NO:1, 3 и 5).
Полинуклеотиды данного изобретения были обнаружены с использованием библиотеки кДНК яичка человека (DPRP-1), библиотеки толстой кишки человека (DPRP-2) и библиотеки кДНК гипоталамуса человека (DPRP-3). Изолированная нуклеиновая кислота для DPRP-1 содержит открытую рамку считывания, кодирующую белок длиной приблизительно 882 аминокислоты, который является структурно родственным DPPIV человека, обнаруживая 26% идентичность и 41% сходство на протяжении всей последовательности белка DPPIV человека. Изолированная нуклеиновая кислота для DPRP-2 содержит открытую рамку считывания, кодирующую белок длиной приблизительно 864 аминокислоты, который имеет 39% сходство со всей аминокислотной последовательностью DPPIV. Анализ первичной аминокислотной последовательности DPRP-1 и DPRP-2 с использованием графиков гидрофобности предсказывает, что эти два белка не имеют трансмембранного домена. Несмотря на этот факт, возможно, что эти внутриклеточные сериновые протеазы секретируются при клеточной активации. Пролиндипептидаза покоящихся клеток (QPP) является сериновой протеазой, мишенью которой являются внутриклеточные везикулы, которые отличаются от лизосом (Chiravuri М, et al., J. Immunol. 2000 Nov 15; 165(10):5695-702). Эта гипотеза расширяет потенциальный сайт (сайты) и масштабы вовлеченности DPRP-1 и DPRP-2 в механизмы посттрансляционной регуляции хемокинов, цитокинов, пептидов и полипептидов. Полноразмерная последовательность DPRP-3 содержит 796 аминокислот, сигнальный пептид с 1 по 48 аминокислоты и трансмембранный домен между 34 и 56 аминокислотами. Предсказывается, что зрелый белок является мембранным белком типа II и может быть расщеплен с образованием растворимой формы. Аминокислотная последовательность представлена в SEQ ID NO:5, которая была предсказана на основе SEQ ID NO:6 и имеет 54% сходство с DPPIV.
Выравнивание аминокислотных последовательностей этих полипептидов с членами подсемейства S9B ферментов пролилолигопептидаз показывают, что все три белка DPRP обладают гомологией полной последовательности и структуры гомологии с DPPIV и FAP. Предсказывается, что DPRP являются членами клана SC ферментов (серин как нуклеофил) с каталитическими остатками в следующем порядке Ser, Asp, His и последовательностью активного центра (G-W-S-Y-G).
DPRP-1, DPRP-2 и DPRP-3 не обнаруживают сходства последовательности с любыми членами семейств классических сериновых протеаз, химотрипсином и субтилизином. Порядок остатков каталитической триады является различным в этих трех основных родственных семействах клана SC: His-Asp-Ser в химотрипсине, Asp-His-Ser в субтилизине и Ser-Asp-His в пролилолигопептидазах.
Как показано в таблице 2, DPRP-3 имеет наивысшую гомологию с DPPIV (68% гомологию и 51% идентичность). Wada et al. выделили кДНК-клоны для DPPVI, DPPIV-родственного белка, из библиотек головного мозга коровы, крысы (Wada et al., Proc. Natl. Acad. Sci. 89:197-201 (1992)) и человека (Yokotani et al., Hum. Molec. Genet. 2:1037-1039 (1993)). Они показали, что, в противоположность DPPIV, каталитическая триада в DPPVI не содержит в качестве первого остатка серин. В DPRP-3 две аминокислоты в каталитической триаде, характерной для семейства сериновых протеаз, являются консервативными. Однако сам остаток серина заменен глицином. Хотя отсутствие остатка серина, вероятно, препятствует протеазной активности в этом центре, возможно, что множество других функций, опосредованных другими функциональными доменами этого белка, остаются интактными.
Как описано вкратце выше, DPPIV является мультифункциональной молекулой, которая проявляет важные функции в зависимости от экспрессирующих клеток и тканей, наряду с ее каталитической активностью в качестве пептидазы. DPRP-3 и DPPIV также, по-видимому, сохраняют множественные функции, несмотря на отсутствие интактной каталитической триады. Например, предполагалось, что DPPIV участвует в регуляции пластичности нейронов. DPPVI экспрессируется на высоком уровне в гиппокампе, таламусе, гипоталамусе и полосатом теле. Кроме того, считается, что задержка развития и эмбриональная смертность эмбрионов rump white Rw/Rw обусловлена разрушением гена DPPIV. Мутация Rw связана с инверсией сегмента хромосомы, простирающейся на 30 сМ проксимальной части хромосомы 5 мыши. Геномный анализ гена DPPIV на хромосоме Rw помещает точку разрыва инверсии в кодирующую область, что приводит к потере значительной части С-концевой области (Hough R.B. et al., Proc. Natl. Acad. Sci., 95, 13800-13805 (1998)).
Ген DPRP-1 человека с предсказанной длиной 32668 п.н. имеет по меньшей мере 22 экзона и восемь транскриптов. Он картируется на хромосоме 15 (NT_010265) в положении 15q21.1-15q22.1. Длины предсказанных транскриптов вариантов альтернативного сплайсинга варьируют между 602 п.н. и 4523 п.н. (см. SEQ ID NO:7-22). Это согласуется с множественными транскриптами, наблюдаемыми с использованием Нозерн-блот-анализа (см. пример 2). EST, представляющие эти транскрипты, были обнаружены в многочисленных тканях, в том числе стареющих фибробластах, Т-лимфоцитах, В-клетках зародышевого центра, семиноме зародышевых клеток, яичке, меланоцитах, матке, яичнике, молочной железе, поражениях при множественном склерозе, поджелудочной железе и плаценте.
Ген DPRP-2 человека относится к гену с по меньшей мере 27 экзонами и девятью вариантами сплайсинга (см. SEQ ID NO:23-40). Один точечный нуклеотидный полиморфизм (SNP) наблюдали в 3'-нетранслируемой области (3'UTR) (88% (37) С против 12% (5) Т). Ген DPRP-2 картируется в районе 19р13.3 хромосомы 19. В этом месте располагается целый ряд маркеров заболеваний и оно связано с различными нарушениями, в том числе гипокальциурической гиперкалиемией, мозжечковой атаксией типа II, мышечной дистрофией, судорогами, подверженностью атеросклерозу, псориазом, эктодермальной дисплазией и острым миелоидным лейкозом. В соответствии с повсеместным распространением этой мРНК, наблюдаемым при Нозерн-блот-анализе (см. пример 2), DPRP-2 экспрессировался в большом разнообразии тканей при исследовании представленности EST (например, более 64 EST экспрессировались в печени, селезенке, мышцах, меланоцитах, сердце, легком, плаценте, коже, поджелудочной железе, желудке, головном мозге, паращитовидной железе).
Ген DPRP-3 человека относится к гену с по меньшей мере 23 экзонами и двумя вариантами сплайсинга (см. SEQ ID NO:41-44). Этот ген картируется на хромосоме 2 (NT_005445) в положении 2q12.3-2q14.1. Транскрипты DPRP-3 не обнаруживают такого широкого распространения, какое обнаруживают DPRP-1 и DPRP-2. Как показано Нозерн-блот-анализом в примере 2, экспрессия DPRP-3 ограничивается головным мозгом и поджелудочной железой. EST, представляющие мРНК DPRP-3, были представлены в ткани, полученной из поражений множественного склероза, гипоталамуса, всего мозга и нервов, причем небольшое число транскриптов обнаруживали в матке и толстой кишке.
Родство среди протеаз человека и грызунов в клане SC, в том числе DPRP-1, DPRP-2 и DPRP-3, анализировали с использованием способа Neighbor Joining (NJ), см. Saitou and Nei, Mol. Biol. Evol., 4, 406-525 (1987). Филогенетический анализ показывает, что среди протеаз S9, DPRP-1 и DPRP-2, обе не имеющие трансмембранного домена, отличаются от DPPIV и близкородственных ему белков, таких как FAP. Однако показано сходство между DPPIV и FAP и между DPRP-3 и DPPIV, которые все являются мембранными белками типа II.
Поиск в базах данных дополнительных родственных DPRP генов выявил присутствие мышиной последовательности, родственной DPRP-1. Выравнивание этой мышиной последовательности с новыми протеазами человека показывает, что mDPRP-1 проявляет существенную гомологию с его человеческим аналогом (фиг.2). Специалисту в данной области будет понятно, что ген новой мышиной протеазы может быть выделен с использованием информации о последовательности, описанной здесь, и легко включен в состав одной из рутинно применяемых экспрессионных конструкций, которые хорошо известны в данной области. Применение этой описанной последовательности специалистами в данной области для создания трансгенной мышиной модели будет включать разработку векторов для таргетинга генов, например таких, которые приводят к гомологичной рекомбинации в мышиных эмбриональных стволовых клетках. Применение мышей с нокаутом генов является ценным инструментом в последующем анализе функции генов DPRP.
Полинуклеотиды данного изобретения могут быть в форме РНК или в форме ДНК; должно быть понятно, что ДНК включает в себя кДНК, геномную ДНК и синтетическую ДНК. Эта ДНК может быть двухцепочечной или одноцепочечной и, если она является одноцепочечной, она может быть кодирующей цепью или некодирующей (антисмысловой) цепью. Кодирующая последовательность, которая кодирует зрелый полипептид, может быть идентичной кодирующей последовательности, показанной в последовательностях SEQ ID NO:2, 4 и 6, соответственно, или она может быть отличающейся кодирующей последовательностью, кодирующей тот же самый зрелый полипептид, в результате избыточности или вырожденности генетического кода или точечного нуклеотидного полиморфизма. Например, она может быть РНК-транскриптом, который включает в себя полную длину любой из последовательностей SEQ Ю NO:2, 4 и 6.
Полинуклеотиды, которые кодируют зрелые белки SEQ ID NO:1, 3, 5, соответственно, могут включать в себя, но не ограничиваются ими, кодирующую последовательность только для зрелого белка; кодирующую последовательность для зрелого полипептида плюс дополнительную кодирующую последовательность, такую как лидерная или секреторная последовательность или последовательность пробелка (предшественника белка); и кодирующую последовательность для зрелого белка (и необязательно дополнительную кодирующую последовательность) плюс некодирующую последовательность, такую как интроны или некодирующая последовательность, расположенная в 5'- и/или 3'-направлении от кодирующей последовательности для зрелого белка.
Таким образом, термин "полинуклеотид, кодирующий полипептид" или термин "нуклеиновая кислота, кодирующая полипептид" должны пониматься как охватывающие полинуклеотид или нуклеиновую кислоту, которые включают в себя только кодирующую последовательность для зрелого белка, а также полинуклеотид или нуклеиновую кислоту, которые включают в себя дополнительную кодирующую и/или некодирующую последовательность. Термины полинуклеотид и нуклеиновая кислота используются взаимозаменяемо.
Данное изобретение включает в себя также полинуклеотиды, где кодирующая последовательность для зрелого белка может быть слита в одной и той же рамке считывания с полинуклеотидной последовательностью, которая способствует экспрессии и секреции полипептида из клетки-хозяина; например, может быть слита лидерная последовательность, которая функционирует в качестве секреторной последовательности для регуляции транспорта полипептида из клетки. Полипептид, имеющий такую лидерную последовательность, называется пребелком или препробелком и может иметь лидерную последовательность, отщепляемую клеткой-хозяином с образованием зрелой формы белка. Эти полинуклеотиды могут иметь 5'-удлиненный район, так что он кодирует пробелок, который является зрелым белком плюс дополнительные аминокислотные остатки на N-конце. Продукт экспрессии, имеющий такую пропоследовательность, называют пробелком, который является неактивной формой зрелого белка; однако, как только эта пропоследовательность отщепляется, остается зрелый белок. Таким образом, например, полинуклеотиды данного изобретения могут кодировать зрелые белки или белки, имеющие пропоследовательность, или белки, имеющие как пропоследовательность, так и препоследовательность (лидерную последовательность).
Полинуклеотиды данного изобретения могут также иметь кодирующую последовательность, слитую в рамке считывания с маркерной последовательностью, которая обеспечивает очистку полипептидов данного изобретения. Маркерная последовательность может быть полигистидиновой меткой (tag), гемагглютининовой (НА) меткой, с-myc-меткой или V5-меткой при использовании хозяина-млекопитающего, например клеток COS-1. НА-метка соответствует эпитопу, происходящему из белка гемагглютинина вируса гриппа (Wilson, I., et al., Cell, 37:767 (1984)), а c-myc-метка может быть эпитопом из белка Мус человека (Evans, G.I. et al., Mol. Cell. Biol. 5:3610-3616 (1985)).
Термин "ген" означает сегмент ДНК, участвующий в синтезе полипептидной цепи; он включает в себя районы, предшествующие кодирующему участку и следующие за кодирующим участком (лидер и трейлер), а также промежуточные последовательности (интроны) между отдельными кодирующими сегментами (экзонами). Термин "значительная гомология последовательности" означает, что по меньшей мере 25%, предпочтительно по меньшей мере 40% аминокислотных остатков являются консервативными и что из неконсервативных остатков по меньшей мере 40% являются консервативными заменами.
Фрагменты полноразмерных генов данного изобретения могут быть использованы в качестве гибридизационного зонда для библиотеки кДНК для выделения полноразмерной кДНК, а также для выделения других кДНК, которые имеют значительную гомологию последовательности с данным геном и будут кодировать белки или полипептиды, имеющие сходную биологическую активность и функцию. Под сходной биологической активностью или функцией, для целей этого изобретения, имеют в виду способность отщеплять N-концевой дипептид, имеющий в качестве предпоследнего Ala или Pro остатка или другие аминокислоты. Такой зонд этого типа имеет по меньшей мере 14 оснований (по меньшей мере 14 следующих друг за другом нуклеотидов из одной из последовательностей SEQ ID NO:2, 4 или 6), предпочтительно по меньшей мере 30 оснований, и такой зонд может содержать, например, 50 или более оснований. Такой зонд может быть также использован для идентификации кДНК-клона, соответствующего полноразмерному транскрипту, и/или геномного клона или клонов, которые содержат полный ген, включающий в себя регуляторные и промоторные области, экзоны и интроны. Меченые олигонуклеотиды, имеющие последовательность, комплементарную последовательности гена данного изобретения, применимы для скрининга библиотеки кДНК человека, геномной ДНК или мРНК для обнаружения членов этой библиотеки, с которыми гибридизуется этот зонд. В качестве примера, известная последовательность ДНК может быть использована для синтеза зонда, который затем используется в скрининге библиотеки для выделения кодирующего района представляющего интерес гена.
Считается, что данное изобретение дополнительно обеспечивает полинуклеотиды, которые гибридизуются с вышеописанными последовательностями, причем идентичность или сходство между последовательностями составляет по меньшей мере 70%, предпочтительно по меньшей мере 90% и более предпочтительно по меньшей мере 95% и, таким образом, кодируют белки, имеющие сходную биологическую активность. Кроме того, как известно в данной области, между двумя полипептидами имеется сходство, когда эти аминокислотные последовательности содержат одинаковые аминокислоты или консервативные аминокислотные замены для каждого индивидуального остатка в данной последовательности. Идентичность и сходство могут быть определены с использованием программного обеспечения для анализа последовательностей (например, Sequence Analysis Software Package, разработанной Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, WI 53705). В частности, данное изобретение обеспечивает такие полинуклеотиды, которые гибридизуются в строгих условиях с вышеописанными полинуклеотидами. В применении здесь термин "строгие условия" означает условия, которые делают возможной гибридизацию между полинуклеотидными последовательностями и полинуклеотидными последовательностями SEQ ID NO:2, 4 и 6, при наличии по меньшей мере приблизительно 70% идентичности. Подходящие строгие условия могут определяться, например, концентрацией соли или формамида в предгибридизационных или гибридизационных растворах, или температурой гибридизации и являются хорошо известными в данной области. В частности, строгость может быть увеличена уменьшением концентрации соли, увеличением концентрации формамида и/или повышением температуры гибридизации.
Например, при гибридизации в строгих условиях можно использовать приблизительно 50% формамида при приблизительно 37°С-42°С, тогда как при гибридизации в условиях меньшей строгости можно использовать приблизительно 35%-25% формамида при приблизительно 30-35°С. В одном конкретном наборе условий для гибридизации в условиях высокой строгости используют 42°С, 50% формамид, 5х SSPE, 0,3% SDS и 200 мкг/мл фрагментированной и денатурированной ДНК спермы лосося. Для гибридизации при условиях пониженной строгости могут быть использованы условия, подобные описанным выше, в присутствии 35% формамида при пониженной температуре 35°С. Диапазон температур, соответствующий конкретному уровню строгости, может быть дополнительно сужен расчетом отношения пуринов к пиримидинам представляющей интерес нуклеиновой кислоты и соответствующей коррекцией температуры. Варьирование указанных выше диапазонов и условий хорошо известны в данной области. Предпочтительно, гибридизация должна иметь место только в том случае, если имеется по меньшей мере 95% или, более предпочтительно, по меньшей мере 97% идентичность между последовательностями. Полинуклеотиды, которые гибридизуются с вышеописанными полинуклеотидами, в предпочтительном варианте кодируют полипептиды, которые проявляют по существу такую же биологическую функцию или активность, что и зрелый белок, кодируемый одной из кДНК последовательностей SEQ ID NO:2, 4 и 6.
Как упоминалось, подходящий полинуклеотидный зонд может содержать по меньшей мере 14 оснований, предпочтительно 30 оснований и более предпочтительно по меньшей мере 50 оснований, и будет гибридизоваться с полинуклеотидом данного изобретения, который имеет идентичность с ним, как описано выше, и который может обладать или может не обладать активностью. Например, такие полинуклеотиды могут быть использованы в качестве зонда для гибридизации с полинуклеотидами SEQ ID NO:2, 4 и 6, соответственно, например, для выделения такого полинуклеотида, или в качестве диагностического зонда, или в качестве ПЦР-праймера. Таким образом, данное изобретение включает в себя полинуклеотиды, имеющие по меньшей мере 70% идентичность, предпочтительно по меньшей мере 90% идентичность и более предпочтительно по меньшей мере 95% идентичность с полинуклеотидом, который кодирует полипептиды SEQ ID NO:1, 3 и 5, соответственно, а также их фрагменты, причем эти фрагменты предпочтительно имеют по меньшей мере 30 оснований и более предпочтительно по меньшей мере 50 оснований, и полипептиды, кодируемые такими полинуклеотидами.
Как хорошо известно в данной области, генетический код является избыточным, что проявляется в том, что определенные аминокислоты кодируются более чем одним триплетом нуклеотидов (кодоном), и данное изобретение включает в себя такие полинуклеотидные последовательности, которые кодируют одни и те же аминокислоты с использованием кодона, отличающегося от кодона, конкретно указанного в качестве примера в последовательностях данного изобретения. Такую полинуклеотидную последовательность называют здесь "эквивалентной" полинуклеотидной последовательностью. Кроме того, данное изобретение включает в себя варианты вышеописанных полинуклеотидов, которые кодируют фрагменты, такие как часть зрелого белка или весь зрелый белок, аналоги и производные одного из полипептидов, имеющих предсказанную аминокислотную последовательность SEQ ID NO:1, 3 и 5, соответственно. Вариантные формы полинуклеотидов могут быть встречающимся в природе аллельным вариантом этих полинуклеотидов или не встречающимся в природе вариантом полинуклеотидов. Например, вариант нуклеиновой кислоты может просто отличаться по последовательности кодона для аминокислоты, что обусловлено вырожденностью генетического кода, или это могут быть делеционные варианты, варианты с заменами и варианты с добавлениями или инсерциями. Как известно в данной области, аллельный вариант представляет собой альтернативную форму полинуклеотидной последовательности, которая может иметь замену, делецию или добавление одного или нескольких нуклеотидов, которые существенно не изменяют биологическую функцию кодируемого полипептида.
Далее, данное изобретение включает в себя полипептиды, которые имеют предсказанную аминокислотную последовательность SEQ ID NO:1, 3 и 5, а также фрагменты, аналоги и производные таких полипептидов. Термины "фрагмент", "производное" и "аналог", при ссылке на полипептиды SEQ ID NO:1, 3 и 5, обозначают полипептиды, которые сохраняют по существу ту же самую биологическую функцию или активность, что и такие полипептиды. Например, аналог может включать в себя пробелок, который может быть активирован отщеплением части пробелка с получением активного зрелого белка. Полипептиды данного изобретения могут быть рекомбинантными полипептидами, природными полипептидами или синтетическим полипептидом, однако, они предпочтительно являются рекомбинантными полипептидами, гликозилированными или негликозилированными.
Фрагмент, производное или аналог полипептида SEQ ID NO:1, 3 и 5, соответственно, может быть: (i) фрагментом, производным или аналогом, в котором один или несколько аминокислотных остатков заменены консервативным или неконсервативным аминокислотным остатком (предпочтительно консервативным аминокислотным остатком), и такой имеющий замену аминокислотный остаток может кодироваться или может не кодироваться генетическим кодом; или (ii) фрагментом, производным или аналогом, в котором один или несколько аминокислотных остатков имеет группу заместителя; или (iii) фрагментом, производным или аналогом, в котором дополнительные аминокислоты слиты со зрелым белком, такие как лидерная или секреторная последовательность или последовательность, которая используется для очистки зрелого полипептида или последовательности пробелка. Предполагается, что такие фрагменты, производные и аналоги могут быть обеспечены специалистами в данной области на основе приведенного здесь описания.
Полипептиды и полинуклеотиды данного изобретения должны быть в изолированной форме, и предпочтительно их очищают до существенной гомогенности или чистоты. Под существенной гомогенностью имеют в виду по меньшей мере приблизительно 85% чистоту.
Термин "изолированный" обозначает, что материал был извлечен из его исходной среды (например, природной среды, если он является встречающимся в природе). Например, встречающийся в природе полинуклеотид или полипептид в живом животном не считается изолированным, но тот же самый полинуклеотид или полипептид при отделении от по существу всех сосуществующих материалов в природной системе считается изолированным. В случае ДНК этот термин включает в себя, например, рекомбинантную ДНК, которая включена в вектор, в автономно реплицирующуюся плазмиду или вирус или в геномную ДНК прокариота или эукариота, или которая существует в виде отдельной молекулы (например, кДНК или фрагмент геномной ДНК или кДНК, полученный полимеразной цепной реакцией (ПЦР) или расщеплением эндонуклеазами рестрикции) независимо от других последовательностей. Он включает в себя также рекомбинантную ДНК, которая является частью гибридного гена, кодирующего дополнительную полипептидную последовательность, например слитой белок. Дополнительно в этот термин включена рекомбинантная ДНК, которая включает в себя часть нуклеотидов, показанных в одной из последовательностей SEQ ID NO:2, 4 или 6, которая кодирует вариант альтернативного сплайсинга DPRP. Различные варианты альтернативного сплайсинга приводятся в качестве примеров в последовательностях SEQ ID NO:8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44 и 46.
Полипептиды данного изобретения включают в себя любой из полипептидов SEQ ID NO:1, 3 и 5 (в частности, зрелый белок), а также полипептиды, которые имеют по меньшей мере 70% сходство (например, предпочтительно по меньшей мере 60% или, более предпочтительно, по меньшей мере 70% идентичность) с одним из полипептидов SEQ ID NO:1, 3 и 5, более предпочтительно по меньшей мере 90% сходство (например, предпочтительно по меньшей мере 90% идентичность) с одним из полипептидов SEQ ID NO:1, 3 и 5, и, наиболее предпочтительно по меньшей мере 95% сходство (например, предпочтительно по меньшей мере 95% идентичность) с одним из полипептидов SEQ ID NO: 1, 3 и 5. Кроме того, они должны предпочтительно включать в себя точно соответствующие фрагменты таких полипептидов, содержащие по меньшей мере 30 аминокислот или, более предпочтительно, по меньшей мере 50 аминокислот.
Фрагменты или части полипептидов данного изобретения могут быть использованы в качестве промежуточных продуктов для получения соответствующих полноразмерных полипептидов посредством пептидного синтеза. Фрагменты или части полинуклеотидов данного изобретения могут быть также использованы для синтеза полноразмерных полинуклеотидов данного изобретения.
Данное изобретение включает в себя также векторы, которые включают в себя такие полинуклеотиды, клетки-хозяева, которые являются генетически модифицированными такими векторами, и получение полипептидов рекомбинантными методами с их использованием. Клетки-хозяева являются генетически модифицированными (трансдуцированными, трансформированными или трансфицированными) такими векторами, которые могут быть, например, клонирующим вектором или экспрессионным вектором. Вектор может быть, например, в виде плазмиды, вирусной частицы, фага и т.д. Сконструированные клетки-хозяева могут культивироваться в общепринятой питательной среде, модифицированной подходящим образом для активации промоторов, отбора трансформантов или амплификации генов данного изобретения. Условия культивирования, такие как температура, рН и т.п., являются условиями, обычно используемыми для клетки-хозяина, выбранной для экспрессии, как хорошо известно специалисту с обычной квалификацией в данной области.
Полинуклеотиды данного изобретения могут быть использованы для получения полипептидов рекомбинантными способами. Так, например, эти полинуклеотиды могут быть включены в любой из различных экспрессионных векторов для экспрессии полипептидов. Такие векторы включают в себя хромосомные, нехромосомные и синтетические ДНК-последовательности, например производные SV40; бактериальные плазмиды; ДНК фага; бакуловирус; дрожжевые плазмиды; векторы, происходящие из комбинаций плазмид и ДНК фага, вирусной ДНК, такой как ДНК вируса осповакцины, аденовируса, вируса оспы птиц и псевдобешенства. Однако любой другой вектор может быть использован при условии, что он является реплицируемым и жизнеспособным в данном хозяине.
Подходящая ДНК-последовательность может быть встроена в вектор любой из различных процедур. Обычно ДНК-последовательность встраивается в подходящий сайт (сайты) эндонуклеаз рестрикции процедурами, хорошо известными в данной области, которые, как предполагается, находятся в пределах компетенции специалистов в данной области.
ДНК-последовательность в экспрессионном векторе функционально связана с подходящей регулирующей экспрессию последовательностью (последовательностями) (промотором) для управления синтезом мРНК. В качестве репрезентативных примеров таких промоторов могут быть упомянуты: промотор LTR или SV40, lac или trp E.coli, промотор фага лямбда P.sub.L и другие промоторы, которые, как известно, регулируют экспрессию генов в прокариотических и эукариотических клетках или их вирусах. Экспрессионный вектор должен также содержать сайт связывания рибосом для инициации трансляции и терминатор транскрипции. Вектор может также включать в себя подходящие последовательности для амплификации экспрессии. Кроме того, экспрессионные векторы предпочтительно содержат один или несколько селективных маркеров генов для обеспечения фенотипического признака для отбора трансформированных клеток-хозяев, такого как дигидрофолатредуктаза или устойчивость к неомицину, в случае культуры эукариотических клеток или такого, как устойчивость к тетрациклину или ампициллину, в случае E.coli.
Вектор, содержащий подходящую ДНК-последовательность, как описано выше, а также подходящую промоторную или регуляторную последовательность, может быть использован для трансформации подходящего хозяина, чтобы обеспечить хозяину возможность экспрессировать белок. В качестве репрезентативных примеров подходящих хозяев могут быть упомянуты: бактериальные клетки, такие как E.coli, Streptomyces, Salmonella typhimurium; клетки грибов, такие как дрожжи; клетки насекомых, такие как Drosophila S2 и Spodoptera Sf9; клетки животных, такие как СНО, COS или меланома Bowes; аденовирусы; клетки растений и т.д. Предполагается, что выбор подходящего хояина находится в пределах компетенции специалиста в данной области при использовании приведенного здесь описания.
Синтетическое получение нуклеотидных последовательностей является хорошо известным в данной области, как очевидно из каталога CLONTECH 95/96 Catalogue, pages 215-216, CLONTECH, 1020 East Meadow Circle, Palo Alto, Calif. 94303. Таким образом, данное изобретение включает в себя также экспрессионные векторы, применимые для получения белков данного изобретения.
Данное изобретение включает в себя дополнительно рекомбинантные конструкции, содержащие одну или несколько из последовательностей, описанных в широком смысле выше. Эти конструкции могут включать вектор, такой как плазмидный или вирусный вектор, в который была встроена последовательность данного изобретения, в прямой или обратной ориентации. В предпочтительном аспекте этого варианта эта конструкция содержит дополнительно регуляторные последовательности, в том числе, например, промотор, функционально связанный с этой последовательностью. Множество подходящих векторов и промоторов известны специалистам в данной области и являются коммерчески доступными. Следующие векторы приводятся в качестве примера: бактериальные: pQE70, pQE60, pQE-9 (Qiagen), pBS, pD10, phagescript, psiX174, pbluescript SK, pbsks, pNYSA, pNH16a, pNH18A, pNH46A (Stratagene), ptrc99a, pKK223-3, pKK233-3, pDR540 и pRIT5 (Pharmacia); и эукариотические: pWLNEO, pSV2CAT, pOG44, pXT1, pSG (Stratagene), pSVK3, pBPV, pMSG и pSVL (Pharmacia). Однако любые другие подходящие плазмида или вектор могут быть использовны при условии, что они являются реплицируемыми и жизнеспособными в хозяине.
Промоторные районы могут быть выбраны из любого желаемого гена с использованием CAT (хлорамфениколацетилтрансфераза)-векторов или других векторов, несущих селектируемые маркеры. Двумя подходящими векторами являются рКК232-8 и рСМ7. Конкретные бактериальные промоторы включают в себя lacI, lacZ, Т3, Т7, gpt, лямбда P.sub.R, P.sub.L и trp.Эукариотические промоторы включают в себя немедленно ранний промотор CMV, промотор тимидинкиназы HSV, ранний и поздний промоторы SV40, LTR из ретровируса и промотор металлотионеина-I мыши. Выбор подходящего вектора и промотора находится в пределах компетенции среднего специалиста в данной области.
Компоненты экспрессионного вектора обычно могут включать в себя: 1) ген неомицинфосфотрансферазы (G418) или ген гигромицин В-фосфотрансферазы (hyg) в качестве селектируемого маркера, 2) сайт инициации репликации Е. coli, 3) последовательность промотора фага Т7 и SP6, 4) последовательности lac-оператора, 5) ген репрессора лактозного оперона (laclq) и 6) линкерный участок сайта множественного клонирования. Такой сайт инициации репликации (oriC) может быть получен из pUC19 (LTI, Gaithersburg, Md.).
Нуклеотидную последовательность, кодирующую один из полипептидов SEQ ID NO:2, 4 и 6, содержащую подходящие рестрикционные сайты, получают, например, в соответствии с протоколом ПЦР, описанным в примере 1 ниже, с использованием ПЦР-праймеров, содержащих рестрикционные сайты для Kpnl (в качестве 5'-праймера) и NotI или Sad (в качестве 3'-праймера) для DPRP-1 или сайты для HindIII (в качестве 5'-праймера) и NotI или BamHI (в качестве 3'-праймера) для DPRP-2. ПЦР-фрагменты очищают электрофорезом в геле и расщепляют подходящими рестрикционными ферментами. Вставку и вектор лигируют в соответствии со стандартными протоколами.
В следующем варианте данное изобретение обеспечивает клетки-хозяева, содержащие вышеописанные конструкции. Клетка-хозяин может быть высшей эукариотической клеткой, такой как клетка млекопитающего, или низшей эукариотической клеткой, такой как клетка дрожжей, или клетка-хозяин может быть прокариотической клеткой, такой как бактериальная клетка. Введение этой конструкции в клетку-хозяина может выполняться трансфекцией с использованием фосфата кальция, ДЭАЭ-декстран-опосредованной трансфекцией, липофекцией или электропорацией (Davis, L., Dibner, M., Battey, L., Basic Methods in Molecular Biology (1986)).
Такие конструкции в клетках-хозяевах предпочтительно используют общепринятым образом для получения продукта гена, кодируемого рекомбинантной последовательностью. Альтернативно, полипептиды данного изобретения могут быть получены синтетически с использованием обычных пептидных синтезаторов или химическим лигированием полученных таким образом подходящих фрагментов.
Зрелые белки могут экспрессироваться в клетках млекопитающих, дрожжах, бактериях или других клетках под контролем подходящих промоторов. Бесклеточные системы трансляции могут быть также использованы для получения таких белков с использованием РНК, полученных из ДНК-конструкций данного изобретения. Подходящие клонирующие и экспрессионные векторы для применения с прокариотическими и эукариотическими хозяевами описаны Sambrook, et al.. Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Spring Harbor, N.Y. (1989).
Транскрипция ДНК, кодирующей полипептиды данного изобретения, высшими эукариотами увеличивается введением в вектор энхансерной последовательности. Энхансеры включают в себя цис-действующие элементы ДНК, обычно около 10-30 п.н., которые действуют на промотор, увеличивая его транскрипции. Примеры включают в себя энхансер SV40 на "поздней" стороне сайта инициации репликации, нуклеотиды 100-270, энхансер раннего промотора цитомегаловируса, энхансер полиомы на "поздней" стороне сайта инициации репликации и энхансеры аденовирусов.
Обычно рекомбинантные экспрессионные векторы содержат сайты инициации репликации и селектируемые маркеры, позволяющие трансформацию клетки-хозяина, например ген устойчивости к ампициллину Е. coli и ген TRP1 S. cerevisiae, и промотор, происходящий из высокоэкспрессируемого гена, для регуляции транскрипции находящейся по ходу транскрипции (downstream) справа от структурной последовательности. Такие промоторы могут быть получены из оперонов, кодирующих гликолитические ферменты, такие как 3-фосфоглицераткиназа (PGK), альфа-фактор, кислая фосфатаза или белки теплового шока, среди прочих. Гетерологичную структурную последовательность состыковывают в подходящей фазе с последовательностями инициации и терминации трансляции и предпочтительно лидерной последовательностью, способной направлять секрецию транслируемого белка в периплазматическое пространство или внеклеточную среду. Необязательно гетерологичная последовательность может кодировать слитый белок, включающий в себя N-концевой идентификационный пептид, обеспечивающий желаемые характеристики, например стабилизацию или упрощенную очистку экспрессируемого рекомбинантного продукта.
Экспрессионные векторы, применимые для бактериального использования, конструируют встраиванием структурной ДНК-последовательности, кодирующей желаемый белок, в функциональной фазе считывания вместе с подходящими сигналами инициации и терминации трансляции под функциональный промотор. Этот вектор будет содержать один или несколько фенотипических селектируемых маркеров и сайт инициации репликации для обеспечения поддержания вектора и, если желательно, для обеспечения амплификации в хозяине. Подходящие прокариотические хозяева для трансформации включают в себя Е. coli, Bacillus subtilis. Salmonella typhimurium и различные виды родов Pseudomonas, Streptomyces и Staphylococcus, хотя и другие могут также использоваться по выбору.
В качестве репрезентативного, но не ограничительного примера, применимые экспрессионные векторы для бактериального использования могут содержать селектируемый маркер и бактериальный сайт инициации репликации, полученные из коммерчески доступных плазмид, содержащих генетические элементы хорошо известного клонирующего вектора pBR322 (ATCC 37017). Такие коммерческие векторы включают в себя, например, рКК223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden) и GEM1 (Promega Biotec, Madison, Wis., U.S.A.). Эти "каркасные" участки pBR322 комбинируют с подходящим промотором и подлежащей экспрессии структурной последовательностью.
После трансформации подходящего штамма-хозяина и выращивания этого штамма-хозяина до подходящей плотности клеток выбранный промотор индуцируют подходящими способами (например, смещением температуры или химической индукцией), и клетки культивируют в течение дополнительного периода времени. Клетки обычно собирают центрифугированием и затем разрушают физическим или химическим способом, причем полученный неочищенный экстракт сохраняют для дальнейшей очистки. Микробные клетки, используемые в экспрессии белков, могут быть разрушены любым подходящим способом, в том числе циклическим замораживанием-оттаиванием, обработкой ультразвуком, механическим разрушением и с использованием лизирующих клетки агентов; такие способы хорошо известны специалистам в данной области.
Для экспрессии рекомбинантного белка могут также использоваться различные системы культур клеток млекопитающих. Примеры систем экспрессии млекопитающих включают в себя линии фибробластов почек обезьяны COS-7, описанные Gluzman, Cell, 23:175 (1981). Другие клеточные линии, способные экспрессировать совместимый вектор, включают в себя, например, клеточные линии С127, 3Т3, СНО, HeLa и ВНК. Экспрессирующие векторы млекопитающих обычно содержат сайт инициации репликации, подходящий промотор и энхансер, а также любые необходимые сайты связывания рибосом, сайт полиаденилирования, донорный и акцепторный сайты сплайсинга, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности. ДНК-последовательности, происходящие из сплайсинга SV40, и сайты полиаденилирования могут быть использованы для обеспечения требуемых нетранскрибируемых генетических элементов.
Полипептиды могут быть выделены и очищены из культур рекомбинантных клеток способами, включающими в себя осаждение сульфатом аммония или этанолом, экстракцию кислотой, анионо- или катионообменную хроматографию, хроматографию на фосфоцеллюлозе, хроматографию гидрофобного взаимодействия, аффинную хроматографию, хроматографию на гидроксилапатите и хроматографию на пектинах. Выделение может облегчаться, если полипептид экспрессируется на поверхности клеток, но это не является обязательным условием. Может быть также желательным выделение продуктов расщепления, которые отщепляются после экспрессии более длинной формы полипептида. Могут использоваться стадии повторной укладки белка, известные в данной области, если необходимо, для завершения образования полной конфигурации зрелого белка. Для конечных стадий очистки может применяться жидкостная хроматография высокого разрешения (ВЭЖХ).
Полипептиды данного изобретения могут быть очищенными природными продуктами или полученными рекомбинантными способами из прокариотического или эукариотического хозяина (например, бактериальных, дрожжевых клеток, клеток высших растений, насекомых или млекопитающих в культуре). В зависимости от хозяина, используемого при получении рекомбинантным способом, полипептиды данного изобретения могут быть гликозилированными или могут не быть гликозилированными. Полипептиды данного изобретения могут также содержать начальный остаток аминокислоты метионина.
В предпочтительном варианте белки данного изобретения выделяют и очищают таким образом, чтобы они по существу не содержали загрязнений другими белками. Например, белки данного изобретения должны составлять по меньшей мере 80% по весу от общего белка, присутствующего в пробе, более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95% и наиболее предпочтительно по меньшей мере 98% по весу от общего белка.
Эти белки могут быть в форме раствора в воде, другом подходящем растворителе, таком как диметилсульфоксид (ДМСО) или этанол, или смеси подходящих растворителей. Примеры смесей растворителей включают в себя 10% (по весу) этанол в воде и 2% (по весу) ДМСО в воде. Раствор может дополнительно содержать соли, буферящие агенты, хаотропные агенты, детергенты, консерванты и т.п. Альтернативно, эти белки могут быть в форме твердого вещества, такого как лиофилизированный порошок или кристаллическое твердое вещество, которое может также содержать остаточный растворитель, соль или т.п.
В контексте данного изобретения "антитела" включают в себя поликлональные антитела, аффинно-очищенные поликлональные антитела, моноклональные антитела и антигенсвязывающие фрагменты, такие как протеолитические F(ab')2- и Fab'-фрагменты. В этот термин включены также генетически сконструированные интактные антитела или фрагменты, такие как химерные антитела, Fv-фрагменты, одноцепочечные антитела и т.п., а также синтетические антигенсвязывающие пептиды и полипептиды. Антитела, происходящие не от человека, могут быть гуманизированы прививкой CDR нечеловеческого происхождения на каркасные и константные участки человека или путем встраивания целиком вариабельных доменов нечеловеческого происхождения (необязательно "обволакиванием" их "человекоподобной" поверхностью посредством замены экспонированных остатков, результатом чего является "облицованное" антитело). В некоторых случаях гуманизированные антитела могут сохранять остатки нечеловеческого происхождения в каркасных доменах вариабельной области для усиления характеристик специфического связывания. Посредством гуманизации антител может быть увеличен полупериод существования и может быть уменьшена потенциальная возможность появления вредных побочных иммунных реакций при введении людям.
Альтернативные способы для получения или отбора антител, применимых здесь, включают в себя подвергание in vitro лимфоцитов действию белка человеческого прогормона DPRP или его пептида и отбор из библиотек фагового дисплея антител или подобных векторов (например, посредством применения иммобилизованного или меченого белка или пептида DPRP человека). Гены, кодирующие полипептиды, имеющие потенциальные домены связывания полипептида DPRP человека, могут быть получены скринингом библиотек случайных пептидов, представленных на фаге (фаговый дисплей) или на бактериях, таких как Е. coli. Нуклеотидные последовательности, кодирующие такие полипептиды, могут быть получены рядом способов, хорошо известных в данной области.
Как должно быть очевидно специалисту с обычной квалификацией в данной области, поликлональные антитела могут быть получены инокуляцией различных теплокровных животных, таких как лошади, коровы, козы, овцы, собаки, куры, кролики, мыши и крысы, полипептидом DPRP человека или его фрагментом. Иммуногенность полипептида человеческого прогормона DPRP может быть увеличена путем использования адъюванта, такого как квасцы (гидроксид алюминия) или полный или неполный адъювант Фрейнда, или поверхностно-активными веществами, такими как лизолецитин полиолы, плуроника (pluronic), полианионы, пептиды, масляные эмульсии, гемоцианин Megathura crenulata (KLH) или динитрофенол. Среди адъювантов, применяемых на людях, особенно предпочтительными являются BCG (бациллы Calmette-Guerin) и Corinebacterium parvum. Полипептиды, применимые для иммунизации, включают в себя слитые белки, Например, соединяющие DPRP или его часть с полипептидом иммуноглобулина или с мальтозосвязывающим белком. Полипептидный иммуноген может быть полноразмерной молекулой или ее частью. Если полипептидная часть является "гаптен-подобной", такая часть может быть предпочтительно соединена или связана с макромолекулярным носителем, таким как гемоцианин Megathura crenulata (KLH), бычий сывороточный альбумин (БСА) или столбнячный токсоид, для иммунизации. Антитела к DPRP могут быть также получены с использованием способов, которые хорошо известны в данной области. Такие антитела могут включать в себя, но не ограничиваются ими, поликлональные, моноклональные, химерные и одноцепочечные антитела, Fab-фрагменты и фрагменты, полученные из экспресионной библиотеки Fab-фрагментов. Нейтрализующие антитела (т.е. такие, которые блокируют или модифицируют взаимодействия в активных центрах) являются особенно предпочтительными для терапевтического применения.
Для получения антител, связывающих белков или пептидов, которые специфически связываются с DPRP, библиотеки одноцепочечных антител, Fab-фрагментов, других фрагментов антител, доменов белков, не являющихся антителами, или пептидов могут быть подвергнуты скринингу. Эти библиотеки могут быть получены с использованием фагового дисплея, другими методами рекомбинантных ДНК или синтезом пептидов (Vaughan, T.J. et al. Nature Biotechnology 14:309-314 (1966)). Такие библиотеки обычно анализируют с использованием способов, которые хорошо известны в данной области, для идентификации последовательностей, которые демонстрируют специфическое связывание с DPRP.
Предпочтительно, олигопептиды, пептиды или фрагменты, используемые для индукции антител к DPRP, имеют аминокислотную последовательность, состоящую из по меньшей мере приблизительно 5 аминокислот и более предпочтительно из по меньшей мере приблизительно 10 аминокислот. Предпочтительно также, чтобы эти олигопептиды, пептиды или фрагменты были идентичными части аминокислотной последовательности природного белка. Короткие фрагменты аминокислот DPRP могут быть также слиты с короткими фрагментами другого белка, такого как KLH, и антитела могут быть получены к этой химерной молекуле.
Моноклональные антитела к DPRP могут быть получены с использованием любого хорошо известного способа, который обеспечивает выработку молекул антител непрерывными клеточными линиями в культуре. Они включают в себя, но не ограничиваются ими, гибридомную технологию, метод гибридом В-клеток человека и метод EBV-гибридом, хотя могут быть предпочтительными моноклональные антитела, продуцируемые гибридомными клетками.
Кроме того, могут быть использованы способы, разработанные для получения "химерных антител", такие как сплайсинг генов мышиных антител в гены антител человека для получения молекулы с подходящей антигенной специфичностью и биологической активностью, см. Neuberger, M.S. et al. Nature 312:604-608 (1984). Альтернативно, способы, описанные для получения одноцепочечных антител, могут быть приспособлены, с использованием известных в данной области способов, для получения DPRP-специфических одноцепочечных антител. Антитела с родственной специфичностью, но с отличающимся идиотипическим составом, могут быть получены перетасовкой цепей (chain shuffling) из случайных комбинаторных библиотек иммуноглобулинов (Burton D.R. Proc. Natl. Acad. Sci. 88:11120-11123 (1991)).
Антитела могут быть также получены индукцией образования in vivo в популяции лимфоцитов или скринингом библиотек иммуноглобулинов или панелей высокоспецифических связывающих реагентов, как описано в литературе (Orlandi, R. et al. Proc. Natl. Acad. Sci. 86:3833-3837 (1989)).
Фрагменты антител, которые содержат сайты специфического связывания для DPRP, могут быть также получены. Например, такие фрагменты включают в себя, но не ограничиваются ими, F(ab')2-фрагменты, получаемые пепсином молекулы антитела, и Fab-фрагменты, генерируемые восстановлением дисульфидных мостиков F(ab')2-фрагментов. Альтернативно, могут быть сконструированы экспрессионные библиотеки Fab-фрагментов для быстрой и легкой идентификации моноклональных Fab-фрагментов с желаемой специфичностью (Huse, W.D. et al. Science 254:1275-1281 (1989)).
Различные иммуноанализы могут быть использованы для идентификации антител, имеющих желаемую специфичность. Многочисленные протоколы для конкурентного связывания или иммунорадиометрических анализов с использованием либо поликлональных, либо моноклональных антител с установленными особенностями хорошо известны в данной области. Такие иммуноанализы обычно включают в себя измерение образования комплексов между DPRP и специфическим антителом к нему. Двухсайтовый иммуноанализ на основе моноклональных антител, использующий моноклональные антитела, реагирующие с двумя непересекающимися эпитопами, является предпочтительным, но может также использоваться анализ конкурентного связывания.
Как упоминалось выше, DPRP могут использоваться в лечении заболеваний. Фармацевтические композиции, пригодные для применения в данном аспекте изобретения, включают в себя композиции, в которых активные ингредиенты содержатся в эффективном количестве для достижения предполагаемой цели, связанной с одним из этих заболеваний. Определение терапевтически эффективной дозы находится вполне в компетенции специалиста в данной области и может быть определено на начальной стадии либо при помощи анализа на клеточной культуре, например, неопластических клеток, или на животных моделях, обычно мышах, крысах, кроликах, собаках или свиньях. Животная модель может быть использована для определения подходящего диапазона концентраций и способа введения, причем эта информация затем обычно используется для определения применимых доз и способов для введения человеку.
Терапевтически эффективной дозой называется количество активного ингредиента, например DPRP или его фрагмента, антител к DPRP или агониста, антагониста или ингибитора DPRP, которое ослабляет конкретные симптомы или состояния заболевания. Например, количество, которое должно вводиться, может быть эффективным для расщепления желаемого субстрата-мишени при контакте с ним. Терапевтическая эффективность и токсичность могут быть также определены стандартными фармацевтическими процедурами на клеточных культурах или экспериментальных животных, например, статистическим расчетом ED50 (доза, терапевтически эффективная в 50% популяции) или LD50 (доза, летальная для 50% популяции). Отношение дозы токсического эффекта к дозе терапевтического эффекта является терапевтическим индексом, и она может выражаться как отношение LD50/ED50. Фармацевтические композиции, которые проявляют большие терапевтические индексы, являются предпочтительными. Данные, полученные в анализах на культурах клеток и исследованиях на животных, используют в определении диапазонов доз для применения в лечении человека. Дозировка, содержащаяся в таких композициях, находится предпочтительно в диапазоне концентраций в кровотоке, которые включают в себя ED50 при малой токсичности или без нее. Доза варьируется в этом диапазоне в зависимости от используемой формы дозы, чувсвительности пациента и способа введения.
Точная дозировка обычно должна определяться медицинским работником с учетом факторов, связанных с субъектом, нуждающимся в лечении, причем доза и введение корректируются для обеспечения достаточного уровня активного компонента или для поддержания желаемого эффекта. Факторы, которые должны учитываться, включают в себя тяжесть патологического состояния, общее состояние здоровья субъекта, возраст, вес и пол субъекта, диету, время и частоту введения, комбинацию (комбинации) лекарственных средств, чувствительность реакции и переносимость/реакцию на терапию. Фармацевтические композиции продолжительного действия могут вводиться каждые 3-4 дня, каждую неделю или даже один раз в две недели в зависимости от полупериода существования в организме и скорости клиренса конкретной формы.
Еще один аспект данного изобретения обеспечивает полинуклеотидные молекулы, имеющие последовательности, которые являются антисмысловыми относительно мРНК-транскриптов полинуклеотидов DPRP-1, DPRP-2 и DPRP-3. Введение молекулы антисмыслового полинуклеотида может блокировать выработку белка, кодируемого DPRP-1, DPRP-2 или DPRP-3. Способы получения молекул антисмысловых полинуклеотидов и введения таких молекул известны в данной области. Например, молекулы антисмысловых полинуклеотидов могут быть инкапсулированы в липосомы для слияния с клетками.
В частности, экспрессия DPRP-1, DPRP-2 и DPRP-3 в специализированных эпителиальных клетках, иммунных клетках (лимфоцитах и В-клетках), астроцитарных опухолях и в различных гормон-чувствительных раках служит доказательством потенциальной роли в патофизиологии рака, метаплазии и метастазировании. Таким образом, в следующем аспекте данное изобретение относится к диагностическим анализам для детектирования заболеваний, связанных с нежелательными уровнями активности или экспрессии DPRP. Антитела, которые специфически связывают DPRP, могут быть использованы для диагностики нарушений, характеризующихся экспрессией DPRP, или в анализах для мониторинга пациентов, получающих DPRP или агонисты или антагонисты (ингибиторы) DPRP для лечения. Антитела, применимые для диагностических целей, могут быть получены таким же образом, как антитела, описанные выше для терапевтических целей. Диагностические анализы на DPRP включают в себя способы, которые используют антитело и метку для детектирования DPRP в жидкостях тела человека или в экстрактах клеток или тканей. Эти антитела могут использоваться с модификациями или без модификаций, и они могут быть помечены ковалентным или нековалентным присоединением к репортерной молекуле. Большое разнообразие репортерных молекул известно в данной области. Рекомбинантные белки DPRP, которые были модифицированы таким образом, что они являются каталитически неактивными, могут быть также использованы в качестве доминантных негативных ингибиторов. Такие мутации включают в себя, например, мутацию в активном центре.
В данной области известны различные протоколы для измерения DPRP, в том числе ELISA, RIA и FACS, и они обеспечивают основу для диагностики измененных или ненормальных уровней экспрессии DPRP. Нормальные или стандартные уровни экспрессии DPRP установливают путем взаимодействия жидкостей тела или экстрактов клеток, взятых от здоровых субъектов - млекопитающих, предпочтительно людей, с антителом к DPRP в условиях, подходящих для образования комплекса. Способ для детектирования DPRP в биологической пробе предусматривает стадии: а) обеспечения биологической пробы; b) объединения этой биологической пробы и анти-DPRP-антитела в условиях, которые пригодны для образования комплекса между DPRP и антителом; и с) детектирования образования комплекса между DPRP и антителом с установлением посредством этого присутствия DPRP в данной биологической пробе. Затем образованный комплекс может быть определен количественно различными способами, предпочтительно фотометрическим способом. Количества DPRP, экспрессируемого в субъекте, контроле и биопсийных образцах тканей от больного, сравнивают со стандартными величинами. Отклонения между стандартными величинами и величинами у субъекта дают параметры для диагностики заболевания.
В другом варианте данного изобретения полинуклеотиды, кодирующие DPRP, используют для диагностических целей, причем эти полинуклеотиды могут включать в себя олигонуклеотидные последовательности, молекулы комплементарных РНК и ДНК и пептидонуклеиновых кислот (ПНК). Эти полинуклеотиды могут быть использованы для детектирования и количественного определения экспрессии генов в биоптатах тканей, экспрессия DPRP в которых может быть соотнесена с одним из заболеваний. Этот диагностический анализ может быть использован для дискриминации между отсутствием, присутствием экспрессии и избыточной экспрессией DPRP и для мониторинга регуляции уровней DPRP во время терапевтического вмешательства. Кроме того, анализ фармакогеномики, точечных нуклеотидных полиморфизмов (SNP) генов DPRP может быть использован в качестве способа скрининга, направленного на мутации, которые указывают на предрасположенность к заболеванию или модифицированную реакцию на лекарственные средства.
Полинуклеотидные и полипептидные последовательности DPRP, их фрагменты, антитела к DPRP и агонисты, антагонисты или ингибиторы DPRP могут использоваться в качестве исследовательских инстументов для идентификации событий молекулярного узнавания и, следовательно, белков, полипептидов и пептидов, которые взаимодействуют с белками DPRP. Характерным примером являются библиотеки фагового дисплея пептидов, где более 108 пептидных последовательностей могут быть подвергнуты скринингу за один раунд пэннинга (panning). Такие способы, а также другие, известны в данной области и могут быть использованы для идентификации соединений, которые ингибируют или усиливают активность DPRP-1, DPRP-2 или DPRP-3. Связанные звенья представляют функциональные взаимодействия, такие как комплексы или пути, и белки, которые взаимодействуют с DPRP, могут быть идентифицированы с использованием дрожжевой двухгибридной системы, протеомики (дифференциального двухмерного гель-анализа и масс-спектрометрии) и геномики (дифференциального анализа экспрессии генов с использованием микрочипов или серийного анализа экспрессии генов SAGE). Белки, идентифицированные как функционально связанные с DPRP, и способ взаимодействия образуют основу способов скрининга, направленного на ингибиторы, агонисты и антагонисты и модуляторы этих DPRP-белок-взаимодействий.
Термин "антагонист", в контексте данного изобретения, относится к ингибиторной молекуле, которая при связывании с DPRP уменьшает величину или продолжительность эффекта биологической или иммунологической активности DPRP, например, уменьшая ферментативную активность этой пептидазы, состоящую в отщеплении N-концевого дипептида. Антагонисты могут включать в себя белки, нуклеиновые кислоты, углеводы, антитела или любые другие молекулы, которые уменьшают действие DPRP; например, они могут включать в себя небольшие молекулы и органические соединения, которые связываются с DPRP и инактивируют их по конкурентному или неконкурентному механизму. Характерные примеры тетрапептидных ингибиторов фериентативной активности фермента DPRP пептидной природы описаны в примерах 6 и 7. Ингибиторы могут быть, например, ингибиторами протеазной активности DPRP или альтернативно ингибиторами активности связывания DPRP с белками, с которыми они взаимодействуют. Конкретные примеры таких ингибиторов могут включать в себя, например, анти-DPRP-антитела, пептиды, фрагменты белка или небольшие ингибиторы пептидилпротеазы или небольшие непептиды, ингибиторные органические молекулы, которые готовят в среде, допускающей введение в желаемый тип клеток. Альтернативно такие ингибиторы могут быть присоединены к лигандам, определяющим выбор мишени, для введения за счет клеточно-опосредованного эндоцитоза и других опосредованных рецепторами событий. Такие способы описаны дополнительно ниже и могут быть применены на практике специалистами в данной области с использованием описанных здесь нуклеотидных и аминокислотных последовательностей DPRP.
Следующим применением для DPRP является применение их для скрининга потенциальных антагонистов для использования в качестве терапевтических агентов, например для ингибирования связывания с DPRP, а также для скрининга, направленного на агонисты. DPRP, его иммуногенные фрагменты или его олигопептиды могут быть использованы для скрининга библиотек соединений, которые являются потенциальными агонистами или антагонистами, любым из различных способов скрининга лекарственных веществ. Фрагмент, используемый в таком скрининге, может быть в свободном виде в растворе, прикрепленным к твердому носителю, находящимся на поверхности клетки или локализованным внутри клетки. Затем измеряют образование комплексов между DPRP и испытуемым агентом. Другие способы анализа для обнаружения антагонистов, которые будут ингибировать DPRP, даются в описаниях Патентов США № 6011155, 6107317, 6110949, 6124305 и 6166063, которые описывают ингибиторы DPPIV. Другим полезным применением этих DPRP является скрининг ингибиторов DPPIV, чтобы показать, что они не будут иметь вредных побочных действий, вызываемых ингибированием также одного или нескольких из этих DPRP.
Способ, обеспеченный для скрининга библиотеки малых молекул, для идентификации молекулы, которая связывает DPRP, обычно предусматривает: а) обеспечение библиотеки малых молекул; b) смешивание библиотеки малых молекул либо с полипептидом SEQ ID NO:1, 3 или 5, либо с его фрагментом в условиях, которые подходят для образования комплекса; и с) детектирование образования комплекса, где присутствие такого комплекса идентифицирует малую молекулу, которая связывает DPRP.
Один способ для идентификации антагониста предусматривает добавление малой молекулы, которая связывает DPRP, в экстракты клеток, трансформированных вектором, экспрессирующим DPRP, вместе с хромогенным субстратом (например, Ala-Pro-AFC или Ala-Pro-AMC) в условиях, в которых могло бы нормально происходить расщепление, и затем анализ ингибирования расщепления этим ферментом путем регистрации изменений флуоресценции, или поглощения УФ-света, спектрофотометрическим методом для идентификации молекул, которые ингибируют расщепление. Уменьшенная скорость реакции или уменьшенная общая величина флуоресценции или поглощения УФ-света в присутствии этой молекулы означает, что эта малая молекула является антагонистом, который уменьшает каталитическую/ферментативную активность DPRP. После идентификации таких молекул они могут вводиться для уменьшения или ингибирования расщепления посредством DPRP.
Термин "агонист", в контексте изобретения, относится к молекуле, которая при связывании с DPRP увеличивает или пролонгирует эффект DPRP. Агонисты могут включать в себя белки, нуклеиновые кислоты, углеводы или любые другие молекулы, которые связываются с DPRP и модулируют действие DPRP. Хотя менее вероятно, что малые молекулы окажутся эффективными агонистами DPRP, способы идентификации такой малой молекулы, которая связывается с DPRP в качестве агониста, предусматривает доставку хромогенной формы малой молекулы, которая связывается с DPRP, в клетки, трансформированные вектором, экспрессирующим DPRP, и анализ изменения флуоресценции или поглощения УФ-света спектрофотометрическим методом. Увеличенное значение поглощения УФ-света или флуоресценции означало бы, что эта малая молекула является агонистом, который увеличивает активность DPRP.
Другой способ скрининга лекарственных средств, который может быть использован, обеспечивает высокопроизводительный скрининг соединений, имеющих подходящее сродство связывания с представляющим интерес белком, как описано в опубликованной заявке РСТ WO 84/03564. В этом способе большие количества различных малых тестируемых соединений синтезируют на твердом субстрате, таком как пластиковые штифты или какая-либо другая поверхность. Тестируемые соединения вводят в реакцию с DPRP или с их фрагментами и затем их промывают. Затем связанный DPRP детектируют способами, хорошо известными в данной области. Очищенные DPRP могут быть также нанесены непосредственно на пластины для применения в вышеописанных способах скрининга лекарственных средств. Альтернативно ненейтрализующие антитела могут быть использованы для захвата пептида и иммобилизации его на твердом носителе.
В другом варианте можно использовать конкурентный способ скрининга лекарственных средств, в котором нейтрализующие антитела, способные связывать DPRP, конкурируют с тестируемым соединением за связывание DPRP. Таким путем антитела могут быть использованы для детектирования присутствия любого пептида, который имеет одну или несколько общих антигенных детерминант с DPRP.
Как указано выше, при помощи исследования сайтов связывания могут быть сконструированы лиганды, которые, например, имеют более сильные взаимодействия с DPRP, чем его природные лиганды. Такие антагонистические лиганды будут связываться с DPRP с более высоким сродством и, следовательно, функционировать как конкурентные лиганды. Альтернативно могут быть сконструированы синтетические или рекомбинантные белки, гомологичные или аналогичные сайту связывания лиганда нативного DPRP, так же, как и другие молекулы, имеющие высокую аффинность в отношении DPRP. Такие молекулы должны быть также способны к вытеснению DPRP и обеспечению защитного действия.
Как указано выше, знание структур DPRP позволяет сконструировать синтетические гомологи и аналоги сайта связывания. Такие молекулы будут в значительной степени облегчать применение этих свойств связывания для нацеливания потенциальных терапевтических агентов на мишень, и они могут быть также использованы для скрининга потенциальных терапевтическиих агентов. Кроме того, они могут быть использованы в качестве иммуногенов при получении моноклональных антител, которые сами могут быть использованы в диагностике и/или терапии, как описано выше.
В связи с повсеместной экспрессией нескольких членов семейства S9B пролилолигопептидаз, клеточные линии, в которых произведено разрушение генов DPPIV, DPRP-1, DPRP-2, DPRP-3, FAP и DPPVI методом таргетинга для установления нуль-фенотипа, будут иметь большую ценность как содействующие скринингу, направленному на селективные и сильнодействующие соединения. Таким образом, данное изобретение обеспечивает такие клеточные линии, сконструированные с использованием элемента ДНК кассеты Lox-Neo IRES tk и GFP-IRES-Neo Knock-in/out для конструирования соматических векторов, для таргетинга генов.
Пример 1
Клонирование и экспрессия генов DPRP с использованием экспрессионной системы млекопитающих
ДНК-фрагменты, кодирующие полноразмерный полипептид DPRP-1, амплифицировали с использованием олигонуклеотидных ПЦР-праймеров, соответствующих 5'- и 3'-последовательностям этого гена, т.е. SEQ ID NO:45 и 46. Кроме того, ДНК-фрагменты, кодирующие полноразмерный полипептид DPRP-2, амплифицировали с использованием олигонуклеотидных ПЦР-праймеров, соответствующих 5'- и 3'-последовательностям этого гена, т.е. SEQ ID NO:50 и 51. Кроме того, ДНК-фрагменты, кодирующие полноразмерный полипептид DPRP-3, амплифицировали с использованием олигонуклеотидных ПЦР-праймеров, соответствующих 5'- и 3'-последовательностям этого гена, т.е. SEQ ID NO:55 и 56.
Эти три амплифицированные последовательности выделяли соответственно из 0,7% агарозного геля с использованием коммерчески доступного набора (GFX PCR DNA and Gel Band Purification Kit, Amersham Pharmacia Biotech Inc. Piscataway NJ, USA). Затем эти фрагменты лигировали в клонирующий вектор, pGEM-7Zf(-) (Promega Corporation, Madison WI, USA) и секвенировали. Соответствующие клонирующие конструкции были соответственно обозначены pGEM7-DPRP1, pGEM7-DPRP2 и pGEM7-DPRP-3. ДНК-последовательности, кодирующие укороченные DPRP-1, DPRP-2 и DPRP-3, амплифицировали с использованием pGEM7-DPRP1 или pGEM7-DPRP2 или pGEM7-DPRP-3 в качестве матрицы и олигонуклеотидных ПЦР-праймеров. SEQ ID NO:45 и 47 использовали для DPRP-1; SEQ ID NO:50 и 52 использовали для DPRP-2 и SEQ ID NO:57 и 58 использовали для DPRP-3. Амплифицированные последовательности вновь выделяли из 0,7% агарозного геля с использованием тех же самых наборов для очистки и субклонировали в pGEM-7Zf(-). Полученные конструкции были названы pGEM7-DPRP1f, pGEM7-DPRP2f и pGEM7-DPRP3f.
Для получения конструкции, экспрессирующей DPRP-1 в млекопитающих, pGEM7-DPRP1 расщепляли рестриктазами KpnI и NotI для высвобождения полноразмерного гена DPRP-1. ДНК-фрагмент, несущий ген DPRP-1, очищали из полосы геля с использованием того же самого набора и затем встраивали в экспрессионный вектор pcDNA3 (Invitrogen, Carlsbad CA, USA) для получения экспрессионной конструкции для нативного DPRP-1, которая была обозначена pcDNA-DPRP1. pGEM7-DPRP1f расщепляли рестриктазами XbaI и HindIII для высвобождения укороченного гена DPRP-1f. ДНК-фрагмент, несущий ген DPRP-1f, очищали из полосы геля с использованием указанного выше набора и затем встраивали в экспрессионный вектор pcDNA3.1(-)/myc-His A (Invitrogen, Carlsbad CA, USA) с получением экспрессионной конструкции pcDNA-MycHis-DPRP1 для меченого DPRP-1.
Для получения конструкции экспрессирующей DPRP-2 в млекопитающих, pGEM7-DPRP2 расщепляли рестриктазами HindIII и BamHI для высвобождения полноразмерного гена DPRP-2. ДНК-фрагмент, несущий ген DPRP-2, очищали из полосы геля с использованием того же самого набора и затем встраивали в экспрессионный вектор pcDNA3 (Invitrogen, Carlsbad CA, USA) для получения экспрессионной конструкции для нативного DPRP-2, которая была обозначена pcDNA-DPRP2. pGEM7-DPRP2f расщепляли рестриктазами EcoRI и BamHI для высвобождения укороченного гена DPRP-2f. ДНК-фрагмент, несущий ген DPRP-2f, очищали из полосы геля с использованием указанного выше набора и затем встраивали в экспрессионный вектор pcDNA3.1(-)/myc-His В (Invitrogen, Carlsbad CA, USA) с получением экспрессионной конструкции pcDNA-MycHis-DPRP2 для меченого DPRP-2.
Для получения конструкции, экспрессирующей DPRP-3 в млекопитающих, pGEM7-DPRP3 расщепляли рестриктазами EcoRI и XhoI для высвобождения полноразмерного гена DPRP-3. ДНК-фрагмент, несущий ген DPRP-3, очищали из полосы геля с использованием того же самого набора и затем встраивали в экспрессионный вектор pcDNA3 (Invitrogen, Carlsbad CA, USA) для получения экспрессионной конструкции для нативного DPRP-3, которая была обозначена pcDNA-DPRP3. pGEM7-DPRP3f расщепляли рестриктазами Nhel и Apal для высвобождения укороченного гена DPRP-3f. ДНК-фрагмент, несущий ген DPRP-3f, очищали из полосы геля с использованием указанного выше набора и затем встраивали в экспрессионный вектор pcDNA3.1(-)/myc-His В (Invitrogen, Carlsbad CA, USA) с получением экспрессионной конструкции pcDNA-MycHis-DPRP3 для меченого DPRP-3.
Пример 2
Профиль экспрессии генов DPRP в тканях человека
Количественный ПЦР-анализ проводили для определения уровней экспрессии мРНК для полипептидов данного изобретения в тканях человека. ОТ-ПЦР проводили также на ряде клеточных линий человека, в том числе, но не только, клетках рака предстательной железы (LNCaP, PC3, DU145), линии MLTC-1 (яичко мыши) и клетках MDA-MB231 (рак молочной железы). Полосы ожидаемых размеров для DPRP-1, DPRP-2 и DPPIV экспрессировались в различных линиях раковых клетках, причем FAP также экспрессируется на очень низких уровнях.
Нозерн-блот-анализ
Нозерн-блот-анализ проводили с 2 мкг поли(А)+ РНК, выделенной из восьми различных тканей с использованием DPRP-зондов. Конкретно, блот Human Multiple Tissue Northern (MTN) blot (Clontech, Palo Alto, Calif.) испытывали при помощи N-концевого фрагмента длиной 1 т.п.н., который был радиоактивно мечен методом случайной затравки в присутствии 32PdCTP (A.P.Feinberg et al., Anal. Biochem., 132, 6 (1983)). Гибридизацию проводили при 68°С в течение ночи в гибридизационном растворе ExpressHyb™ (Clontech, Palo Alto, Calif.). Блоты сначала промывали при комнатной температуре в 2х SSC и 0,05% SDS и затем промывали при 60°С (DPRP-1 и DPRP-2) и 50°С (DPRP-3) в 0,1х SSC и 0,1% SDS.
Нозерн-анализ выявил экспрессию DPRP-1 в нескольких тканях с наиболее представленным сигналом в яичке, предстательной железе, мышце и головном мозге. В яичке обнаружено 3 транскрипта длиной приблизительно 7,5, 4,5 и 2,5 т.п.о. Более короткие варианты мРНК присутствовали в большом количестве в яичке, но в пренебрежимо малом в других исследованных тканях. DPRP-2 экспрессировался повсеместно в каждой ткани с наиболее высокими уровнями в печени и мышце и преобладающим транскриптом размером 5 т.п.о. Экспрессия DPRP-3 ограничивалась головным мозгом и поджелудочной железой. Дополнительный анализ в отношении этих трех протеаз проводили в отдельных областях головного мозга (мозжечке, коре, продолговатом мозге, спинном мозге, затылочной доле головного мозга, лобной доле головного мозга, височной доле головного мозга и скорлупе). DPRP-1 экспрессировался во всех областях, причем низкие уровни были в спинном мозге, тогда как DPRP-2 экспрессировался во всех исследованных областях головного мозга.
Олигонуклеотидные праймеры SEQ ID NO:48 и 49 использовали для количественной ПЦР в случае DPRP-1, тогда как олигонуклеотидые праймеры SEQ ID NO:53 и 54 использовали для количественной ПЦР в случае DPRP-2. Панели кДНК Human Multiple Tissue cDNA (MTStm) Panel I и Panel II (Clontech, Palo Alto, CA, USA) использовали в качестве нормализованных кДНК-матриц. 0,5 нг каждой кДНК использовали в ПЦР (25 мкл) с каждым праймером в конечной концентрации 300 нМ. ПЦР проводили с использованием набора реагентов SYBR Green PCR Core Reagents Kit (Applied Biosystems, Foster City CA, USA) и детектировали при помощи системы детектирования последовательностей Applied Biosystems GeneAmp 5700. Использовали рекомендуемые изготовителем параметры проведения термических циклов, например 50°С в течение 2 мин, 95°С в течение 10 мин с последующими 40 циклами, состоящими из 95°С в течение 15 с и 60°С в течение 1 мин. Полученные данные показывают относительно высокие уровни экспрессии как для DPRP-1, так и для DPRP-2 в поджелудочной железе, яичнике и яичке и особенно высокий уровень для DPRP-2 в печени.
Пример 3 Получение поликлональных антител к DPRP и Вестерн-блоттинг
Аминокислотную последовательность, предсказанную по кДНК, кодирующей DPRP-1, анализировали с использованием программы DNASTAR (DNASTAR, Inc.) для определения районов высокой иммуногенности и соответствующий олигонуклеотид синтезировали и использовали для индукции анти-DPRP-1-антител. Эту процедуру повторяли для DPRP-2 и DPRP-3. Отбор подходящих пептидных последовательностей и способы получения антител являются способами, хорошо известными специалистам в данной области. Отбор подходящих эпитопов, таких как эпитопы вблизи С-конца или в гидрофильных областях, хорошо известен в данной области.
Обычно олигопептиды, которые имеют длину приблизительно 15-20 остатков, например, SEQ ID NO:59 для DPRP-1, SEQ ID NO:60 для DPRP-2 и SEQ ID NO:61 для DPRP-3, синтезировали с использованием пептидного синтезатора Applied Biosystems. Peptide Synthesizer Model 431 А. Использовали Fmoc-химию и пептиды из 19 или 15 остатков, соответственно, конъюгировали с гемоцианином Megathura crenulata (KLH, Sigma, St. Louis, МО) реакцией с N-малеимидобензоил-N-гидроксисукцинимидным эфиром (MBS). Кроликов иммунизировали комплексом олигопептид-KLH в полном адъюванте Фрейнда. Полученные антисыворотки испытывали на антипептидную активность, например, связыванием этого пептида с пластиком, блокированием 1% БСА, реакцией с кроличьими антисыворотками, промыванием и реакцией с меченым радиоактивным йодом козьим антителом против кроличьего IgG.
Вестерн-блоттинг проводили с использованием проб белка здорового человека (Protein Medley), полученных от Clontech (около 36 мкг общего белка). Белки фракционировали на 10% SDS-полиакриламидных гелях и переносили на нитроцеллюлозные мембраны 0,45 мкм). Мембраны блокировали в забуференном Трис солевом растворе (TBS) с 0,05% Твином 20 и 1% БСА. Анти-DPRP-1- или анти-ОРКР-2-специфичные антитела использовали в качестве первичных антител и их разводили 1:5000 в забуференном Трис солевом растворе с 0,05% Твином 20 (TBST), и конъюгированные со щелочной фосфатазой (АР) козьи антитела против кроличьего IgG (Promega) разводили 1:5000 в том же самом буфере перед использованием. Положительную реакцию визуализировали инкубированием мембраны в растворе субстрата для АР, Western Blue Stabilized Substrate (Promega), пока представляющие интерес полосы не достигали желаемой интенсивности. Белки DPRP-1 и DPRP-2 были обнаружены в тканях головного мозга, мышц, почек, предстательной железы, яичка и яичника. DPRP-1 и DPRP-3 синтезировались в виде форм с массой около 101 кДа и 100 кДа, соответственно, что хорошо согласуется с молекулярными массами, рассчитанными из их первичной структуры, как показано в таблице 3.
Наблюдали несколько дополнительных полос близкой молекулярной массы. Предполагается, что они обусловлены присутствием посттрансляционного гликозилирования этих белков. Таблица 3 показывает также число потенциальных сайтов N-гликозилирования для белков DPRP. Присутствие гликозилированных и негликозилированных форм этих белков оценивали с использованием туникамицина, ингибитора синтеза олигосахаридов. Очевидно, что меньшие формы были негликозилированными формами. Корреляция между количествами мРНК (Нозерн-анализ) и белка (Вестерн-анализ) для DPRP-1 показана в таблице 4.
Пример 4
Иммуногистохимическая локализация белков DPRP в тканях человека
Срезы толщиной 4 микрона готовили из нескольких различных фиксированных формалином залитых в парафин тканей. Срезы тканей депарафинировали посредством 4 погружений в ксилол на 5 минут с последующей промывкой серией растворов спирта со ступенчато снижающейся концентрацией до дистиллированной воды. Индуцированное нагреванием паром извлечение эпитопа (SHIER) использовали с несколькими различными растворами для SHIER с ферментативным расщеплением ткани или без него в двух разных концентрациях (Ladner et al., Cancer Res., 60, p.3493-3503, 2000). Использованные обработки и разведения антител:
1. Блокирующий реагент в течение 15 мин (нормальная козья сыворотка).
2. Первичное антитело в течение 25, 60 мин или инкубирование в течение ночи.
3. Вторичное антитело в течение 25 мин (биотинилированные козьи антитела против кроличьего IgG).
4. Блокирование эндогенной пероксидазы в течение 3×1,5 мин.
5. АВС (комплекс авидин-биотин)/пероксидаза хрена в течение 25 мин.
6. Хромоген DAB в течение 3×5 мин (коричневый продукт реакции).
7. Окрашивание Light Hematoxylin Counter Stain 1 мин.
Положительные контроли ставили для того, чтобы гарантировать, что химия детектирования и предварительные обработки антигена срабатывали должным образом. Кроличий IgG использовали в качестве отрицательного контроля. Использовали систему окрашивания ткани на основе авидина-биотина для детектирования антитела к DPRP-1. Пероксидазу хрена (HRP) использовали в качестве репортерного фермента с DAB в качестве хромогена. После окрашивания препараты дегидратировали серией спиртовых промывок до абсолютного этанола с последующими промывками ксилолом. Препараты перманентно заключали при помощи покровных стекол и среды Permount. Цифровые изображения репрезентативного окрашивания, где положительное окрашивание отображалось темно-коричневым хромогеном (продукт реакции DAB-HRP), получали с использованием видеокамеры Olympus. Контрольное окрашивание гематоксилином обеспечивает синюю окраску ядер для оценки морфологии клеток и тканей.
Кроличье поликлональное антитело против DPRP-1 метит фиксированные формалином, залитые в парафин ткани человека, в том числе нормальные яичко, предстательную железу, эндометрий, миндалины и поджелудочную железу. Оно присутствует также в эндотелиальных клетках нормального яичника, мочевого пузыря и почки. Окрашивание локализовано в цитоплазме эпителиальных клеток и некоторых стромальных клеток, таких как фибробласты, эндотелиальные клетки и лимфоциты. Интересно, что в нормальном яичке, испытанном с антителами к DPRP-1, была явно выраженная экспрессия в клетках Лейдига и многоядерных макрофагах, обнаруженных в интерстициальной ткани, которая находится в пространстве, окружающем семенные канальцы. В-клетки миндалин окрашивались антителом против DPRP-1.
Пример 5
Экспрессия белков DPRP в клетках млекопитающих и насекомых и очистка
Плазмидную ДНК pcDNA-DPRP1 pcDNA-MycHis-DPRP1, pcDNA-DPRP2 или pcDNA-MycHis-DPRP2 трансфицировали в клетки PEAK (EdgeBioSystems, Gaithersburg MD, USA) или COS-1 (ATCC CRL-1650) с использованием способа с липофектамином (LipofectAmine) (Life Technologies, Gaithersburg MD, USA), рекомендованного изготовителем. Трансфицированные клетки поддерживали в среде DMEM с 5% фетальной телячьей сывороткой (ФТС) при 37°С с 5% CO2 в течение 48 часов. Затем клетки собирали и использовали для экстракции рекомбинантного белка. Клетки собирали спустя 48 часов после трансфекции, гомогенизировали и затем центрифугировали при 18000×g в течение 40 минут. Супернатанты собирали в качестве цитозольных фракций. Эту фракцию наносили на центрифужную колонку TALON (Clontech) и несущие His-метку белки элюировали 50 мМ PBS, 150 мМ имидазолом, рН 7. Затем рекомбинантные белки детектировали вестерн-блоттингом с анти-myc-антителом и визуализировали с использованием системы ProtoBlot II АР (Promega). Рекомбинантные аффинно очищенные слитые белки DPRP-1 и DPRP-2 детектировали вестерн-блоттингом, и DPRP-1 и DPRP-2 синтезировались в виде форм с массой 112 кДа и 109 кДа, как и предсказывалось.
Природные или рекомбинантные белки DPRP в значительной мере очищали иммуноаффинной хроматографией с использованием антител, специфичных в отношении DPRP-1, DPRP-2 или DPRP-3. Иммуноаффинную колонку получали ковалентным присоединением антител DPRP к активированной хроматографической смоле, такой как CNBr-активированная Сефароза (Pharmacia and Upjohn). После присоединения колонку блокировали и промывали в соответствии с инструкциями изготовителя.
Среды или клеточные экстракты, содержащие белки DPRP, пропускали через эту иммуноаффинную колонку и колонку промывали при условиях, которые обеспечивают предпочтительное связывание DPRP (например, буферами высокой ионной силы в присутствии детергента). Колонку элюировали при условиях, которые разрушают связывание антитело/DPRP (например, буфером рН 2-3 или высокой концентрацией хаотропа, такого как мочевина или тиоцианатный ион) и очищенный DPRP собирали.
Пример 6
Ферментативная активность белков DPRP и способы скрининга, направленного на ингибиторы
Кинетические характеристики рекомбинантых DPRP-1 и DPRP-2 определяли методом непрерывного флуорометрического анализа. Оптимизация зависимости от буфера, рН и температуры привела к следующим условиям проведения анализа: ферментативные анализы проводили в 50 мМ PBS, рН 7,4. 50 мкл (50 мкг/мл) очищенных ферментов смешивали с 1 мкл Ala-Pro-AMC (Enzyme Systems) в различных концентрациях. Затем планшеты инкубировали при 37°С в течение 30 минут и флуоресценцию детектировали с использованием флуориметра Wallac 1420 при λех 355 нм и λem 535 нм. Величины Кm DPRP-1 и DPRP-2 были сходными (208 и 161 мкМ, соответственно).
Дополнительная биохимическая характеризация показала, что DPRP-1 и DPRP-2 имеют сходные с DPPIV профили. Эти две очищенные протеазы и DPPIV предынкубировали с ингибиторами при комнатной температуре в течение 30 минут. Затем добавляли субстрат Ala-Pro-AMC (100 мкМ) и интенсивность флуоресценции регистрировали в виде 60 измерений в течение 60-минутного периода. Необратимый ингибитор сериновых протеаз AEBSF был единственным из испытанных ингибиторов, который показал сильное ингибирование всех трех ферментов (таблица 5). Это подтверждает предсказание, основанное на анализе структуры и доменов, что эти белки принадлежат к суперсемейству сериновых протеаз.
Кроме Ala-Pro-АМС, дополнительные испытанные субстраты также подтвердили, что DPRP-1 и DPRP-2 являются дипептидилпептидазами. Эти данные получали определением изменения флуоресценции после 30-минутного инкубирования субстратов (125 мкМ) с ферментами, выраженного в процентах от умеренной флуоресценции, и Ala-Pro-АМС и Gly-Pro-AMC были единственными хорошими субстратами среди испытанных.
Дополнительные ди-, три- и тетрапептиды, содержащие природные и неприродные аминокислоты, испытывали для определения оптимального субстрата для испытания каждого из белков DPRP, которые будут также показывать уменьшенную активность при инкубировании CDPPIV.
Описанный здесь способ определения фермента является одним из ряда способов, которые могут быть использованы для скрининга, направленного на пептидные и непептидные ингибиторы ферментов DPRP. Были испытаны библиотеки тетрапептидных ингибиторов для обнаружения ингибиторов ферментативной активности. Ингибиторы-кандидаты готовили в виде исходных растворов с концентрацией 10-20 мМ в ДМСО и хранили при -20°С. Разбавления делали в буфере для анализа. Ингибирование определяли сравнением изменений флуоресценции в присутствии ингибированного фермента с изменением флуоресценции в присутствии контрольного фермента (разбавитель). Выражение 100-(единицы фл. пробы/единицы фл. контроля ×100) дает величину ингибирования в процентах. Процент ингибирования и концентрация ингибитора, при которой фермент ингибировался на 50% (IC50), определяли построением графика зависимости процента ингибирования от концентрации ингибитора в логарифмическом масштабе. Как показано на фигуре 3, несколько амидов тетрапептидов ингибировали ферментативную активность, где данные выражены в процентах от активности в присутствии только разбавителя (0,02% ДМСО). Соединения добавляли в концентрации 1 мМ. Наиболее интересным было явно различное действие некоторых тетрапептидов в отношении DPRP-1 и DPRP-2 в сравнении с DPPIV. Хотя все три фермента ингибировались пептидом-1, только DPRP-1 и DPRP-2 значительно ингибировались пептидом-4 и пептидом-5. Это показывает, что селективное ингибирование очищенных ферментов является достижимым.
Способ анализа, описанный в этом примере, может быть также использован для скрининга дополнительных библиотек синтетических или природных соединений, включающих в себя макромолекулы, в отношении агентов, которые либо ингибируют, либо усиливают активность DPRP. Полипептиды DPRP-1 и DPRP-2 для использования в этом анализе могут быть получены, например, трансляцией in vitro, рекомбинантной экспрессией (см. пример 5) или биохимическими методами. Другие способы, отличающиеся от описанных здесь, могут быть также использованы для скрининга и идентификации соединений, ингибирующих DPRP-1, DPRP-2 или DPRP-3, и эти способы могут включать в себя, например, такие способы анализа связывания, как ELISA и RIAs.
Пример 7
Действие ингибиторов DPRP на пролиферацию раковых клеток человека in vitro
В попытке оценить действие, которое могут иметь некоторые ингибиторы активности DPRP-1 и DPRP-2 на пролиферацию раковых клеток человека, клетки LNCap, РС3 и Du145, линии рака мышиного яичка MLTC-1 и рака молочной железы MDA-MB231 высевали (104 на лунку) в 96-луночные планшеты для культуры ткани и давали им расти и прикрепляться в течение 24 часов при 37°С в СО2-инкубаторе. Затем соединения в разных разведениях (конечные разведения: 0,1 нМ - 10 мкМ) добавляли в лунки и выдерживали в течение различных периодов инкубации от 24 часов до 96 часов с ежедневной заменой соединения свежим соединением. Добавление одного разбавителя ДМСО служило в качестве контроля. После инкубирования с этими соединениями в трех повторностях измеряли пролиферацию этих клеток с использованием теста пролиферации клеток ХТТ (Roche 1-465-015). Снимали показания с планшетов при 490 и 650 нм спустя 5 часов после добавления смеси ХТТ. Увеличение пролиферации клеток наблюдали с тремя ингибиторами при концентрациях, равных 0,1, 1, 10 и 100×IC50, и эти результаты показаны на фигурах 4a, 4b и 4c для клеток РС3.
В целом, DPRP экспрессируются в разнообразных тканях, как было показано амплификацией мРНК, Вестерн-блоттингом и иммуногистохимией. DPRP-1 является наиболее представленным в яичке по данным Нозерн-блоттинга и Вестерн-блоттинга. Большое число маркерных экспрессируемых последовательностей (EST) в яичке как источнике кДНК, которые являются гомологичными с DPRP-1, также подтверждает обильную экспрессию DPRP-1 в яичке. Пример 4 описывает иммуногистохимическую локализацию белка DPRP-1 в яичке человека с использованием специфического для DPRP-1 антитела. DPRP-1 сильно экспрессируется в эпителоидных клетках Лейдига, и клетки Лейдига являются основным источником тестикулярных андрогенов (мужских стероидных гормонов) в мужских особях млекопитающих. В интерстиции яичка клетки Лейдига и макрофаги находятся в тесной связи, обусловленной тем, что выросты клеток Лейдига простираются в виде "пальцев" на поверхность макрофагов. Многоядерные клетки в непосредственной близости к клеткам Лейдига также окрашивались антителами к DPRP-1, что предполагает, что эта протеаза экспрессировалась также в макрофагах, и макрофаги в яичке играют важную роль в паракринной регуляции клеток Лейдига. Цитокины, секретируемые тестикулярными макрофагами, являются митогенными для клеток Лейдига и играют важную роль в дифференцировке мезенхимных клеток-предшественников в зрелые клетки Лейдига. Более ясное знание белков и путей, участвующих в созревании яичка, является важным для открытия новых способов лечения преждевременного полового созревания. Кроме того, клетки Лейдига индуцируют опухоли, такие как стромальные опухоли зародышевых тяжей, посредством продуцирования половых стероидных гормонов (преимущественно тестостерона). Тестостерон связан с несколькими неоплазиями и заболеваниями, такими как карцинома молочной железы и раки матки, карцинома яичников и андрогенная алопеция (потеря волос). Дальнейшее исследование локализации белков DPRP в других железах в теле (например, в надпочечниках), которые продуцируют тестостерон и другие андрогенные гормоны, находится в настоящее время в стадии исследования. Исследуется возможная связь DPRP-1 с функциями путей биосинтеза стероидных и полипептидных гормонов, и пример 7 важен для понимания роли белков DPRP в in vitro моделях клеток предстательной железы, яичка и молочной железы.
Иммуногистохимический анализ также показал локализацию DPRP-1 в эндометриальных железах матки (см. пример 4), дольках поджелудочной железы, почечных клубочках, плазматических клетках мочевого пузыря, субпопуляции В-клеток в миндалинах, столбчатых эпителиальных клетках предстательной железы и слабо дифференцированной чешуйчатой метаплазии предстательной железы, раке предстательной железы 4 степени по Gleason и гиперплазированных железах при доброкачественной гиперплазии предстательной железы. Положительное окрашивание рака молочной железы, а также семиномы и чешуйчатой метаплазии предстательной железы, предполагает общую связь DPRP-1 с гормон-чувствительными тканями, в частности, в клетках, которые становятся слабодифференцированными. Присутствие DPRP-1 в специализированных эпителиальных клетках и в воспалительных плазматических клетках (лимфоцитах) также представляет интерес. Воспалительный рак молочной железы имеет обилие инфильтрирующих лимфоцитов и общий плохой прогноз. Белки DPRP-1 и другие белки DPRP обнаруживаются в медуллярных раках, которые обычно имеют постоянный инфильтрирующий лимфоплазмацитарный компонент на периферии опухоли, который, как считают, представляет реакцию тканей хозяина на новообразование. Большинство этих лимфоцитов являются Т-клетками, а большинство плазмоцитов являются клетками IgG-продуцирующего типа. Несколько антигенов были широко представлены на В-клетках, подгруппе клеток рака молочной железы и других эпителиальных раковых клеток, и эти антигены являются мишенями для нового класса терапевтических моноклональных антител, причем явный успех был достигнут с гуманизированным моноклональным антителом против специфического для В-клеток антигена CD20. Таким образом, предполагается, что моноклональные антитела к белкам DPRP могут оказаться применимыми для диагностики и лечения заболеваний, в которые эти белки вовлечены, в том числе рака.
Экспрессия DPRP-1 в специализированных эпителиальных клетках ряда тканей позволяет предположить, что DPRP-1 и другие белки DPRP могут участвовать в росте и дифференцировке этих клеток. Испытание с использованием ингибиторов, описанных в примере 6, in vitro на моделях рака предстательной железы и яичка (пример 7), показывает, что ингибиторы DPRP-1/DPRP-2 вызывали 50-60% увеличение пролиферации клеток РС3 при наномолярных концентрациях, как показано на фигурах 4а-4с.
Хотя данное изобретение было описано в соответствии с его предпочтительными вариантами, которые представляют наилучший способ, известный в настоящее время авторам данного изобретения, должно быть понятно, что, как будет очевидно специалистам в данной области, могут быть произведены изменения и модификации без отклонения от его объема, который определен прилагаемой формулой изобретения. Например, хотя данное описание сосредоточено в определенных случаях на DPRP-1 и DPRP-2, DPRP-3 и его фрагменты должны рассматриваться как применимые сходным образом, как и кодирующие их нуклеиновые кислоты. Конкретные признаки данного изобретения подчеркиваются в формуле изобретения.
Краткий перечень последовательностей
SEQ ID NO:
1. Аминокислотная последовательность DPRP-1
2. ДНК-последовательность DPRP-1
3. Аминокислотная последовательность DPRP-2
4. ДНК-последовательность DPRP-2
5. Аминокислотная последовательность DPRP-3
6. ДНК-последовательность DPRP-3
7. Аминокислотная последовательность транскрипта 0 DPRP-1
8. ДНК-последовательность транскрипта 0 DPRP-1
9. Аминокислотная последовательность транскрипта 1 DPRP-1
10. ДНК-последовательность транскрипта 1 DPRP-1
11. Аминокислотная последовательность транскрипта 2 DPRP-1
12. ДНК-последовательность транскрипта 2 DPRP-1
13. Аминокислотная последовательность транскрипта 3 DPRP-1
14. ДНК-последовательность транскрипта 3 DPRP-1
15. Аминокислотная последовательность транскрипта 4 DPRP-1
16. ДНК-последовательность транскрипта 4 DPRP-1
17. Аминокислотная последовательность транскрипта 5 DPRP-1
18. ДНК-последовательность транскрипта 5 DPRP-1
19. Аминокислотная последовательность транскрипта 6 DPRP-1
20. ДНК-последовательность транскрипта 6 DPRP-1
21. Аминокислотная последовательность транскрипта 7 DPRP-1
22. ДНК-последовательность транскрипта 7 DPRP-1
23. Аминокислотная последовательность транскрипта 0 DPRP-2
24. ДНК-последовательность транскрипта 0 DPRP-2
25. Аминокислотная последовательность транскрипта 1 DPRP-2
26. ДНК-последовательность транскрипта 1 DPRP-2
27. Аминокислотная последовательность транскрипта 2 DPRP-2
28. ДНК-последовательность транскрипта 2 DPRP-2
29. Аминокислотная последовательность транскрипта 3 DPRP-2
30. ДНК-последовательность траснскрипта 3 DPRP-2
31. Аминокислотная последовательность транскрипта 4 DPRP-2
32. ДНК-последовательность транскрипта 4 DPRP-2
33. Аминокислотная последовательность транскрипта 5 DPRP-2
34. ДНК-последовательность транскрипта 5 DPRP-2
35. Аминокислотная последовательность транскрипта 6 DPRP-2
36. ДНК-последовательность транскрипта 6 DPRP-2
37. Аминокислотная последовательность транскрипта 7 DPRP-2
38. ДНК-последовательность транскрипта 7 DPRP-2
39. Аминокислотная последовательность транскрипта 8 DPRP-2
40. ДНК-последовательность транскрипта 8 DPRP-2
41. Аминокислотная последовательность транскрипта 0 DPRP-3
42. ДНК-последовательность транскрипта 0 DPRP-3
43. Аминокислотная последовательность транскрипта 1 DPRP-3
44. ДНК-последовательность транскрипта 1 DPRP-3
45. Прямой праймер, используемый для клонирования DPRP-1
46. Обратный праймер, используемый для клонирования полноразмерного гена DPRP-1
47. Обратный праймер, используемый для клонирования слитого гена DPRP-1
48. Прямой праймер, используемый для анализа профиля экспрессии DPRP-1
49. Обратный праймер, используемый для анализа профиля экспрессии DPRP-1
50. Прямой праймер, используемый для клонирования DPRP-2
51. Обратный праймер, используемый для клонирования полноразмерного гена DPRP-2
52. Обратный праймер, используемый для клонирования слитого гена DPRP-2
53. Прямой праймер, используемый для анализа профиля экспрессии DPRP-2
54. Обратный праймер, используемый для анализа профиля экспрессии DPRP-2
55. Прямой праймер, используемый для клонирования DPRP-3
56. Обратный праймер, используемый для клонирования полноразмерного гена DPRP-3
57. Прямой праймер, используемый для клонирования слитого гена DPRP-3
58. Обратный праймер, используемый для клонирования слитого гена DPRP-3
59. Пептидная последовательност антигена DPRP-1
60. Пептидная последовательность антигена DPRP-2
61. Пептидная последовательности антигена DPRP-3
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ИЗОФОРМЫ ИНГИБИТОРА РОСТА ВАСКУЛЯРНЫХ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК | 2002 |
|
RU2316591C2 |
| ИЗОЛИРОВАННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ПОЛИПЕПТИД TAG 7, ИЗОЛИРОВАННЫЙ ПОЛИПЕПТИД TAG 7, СПОСОБ ИНГИБИРОВАНИЯ РАЗВИТИЯ ОПУХОЛЕЙ У МЛЕКОПИТАЮЩИХ (ВАРИАНТЫ) И СПОСОБ ЛЕЧЕНИЯ РАКА У ЖИВОТНОГО(ВАРИАНТЫ) | 1998 |
|
RU2238976C2 |
| ПОЛИПЕПТИДЫ, ПРОИСХОДЯЩИЕ ИЗ ТРИПТОФАНИЛ-ТРНК-СИНТЕТАЗЫ, И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕГУЛЯЦИИ РАЗВИТИЯ КРОВЕНОСНЫХ СОСУДОВ | 2002 |
|
RU2297425C2 |
| ПОЛИНУКЛЕОТИДЫ И ПОЛИПЕПТИДЫ ФАГА φ-mru, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2520738C2 |
| ПОЛИПЕПТИД, ИМЕЮЩИЙ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ ФОСФОДИЭСТЕРАЗЫ | 1998 |
|
RU2272841C2 |
| ГЕН MTS, МУТАЦИИ ДАННОГО ГЕНА И СПОСОБЫ ДИАГНОСТИКИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ С ИСПОЛЬЗОВАНИЕМ ПОСЛЕДОВАТЕЛЬНОСТИ ГЕНА MTS | 1995 |
|
RU2164419C2 |
| ПРОНИКАЮЩИЕ В КЛЕТКУ ПЕПТИДЫ И ПОЛИПЕПТИДЫ ДЛЯ КЛЕТОК МИКРООРГАНИЗМОВ | 2008 |
|
RU2526511C2 |
| НОВЫЕ СЕЛЕКТИВНЫЕ ВЕКТОРЫ И СПОСОБЫ СЕЛЕКЦИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ХОЗЯЕВ | 2014 |
|
RU2716977C2 |
| ВАКЦИНЫ И КОМПОНЕНТЫ ВАКЦИН ДЛЯ ПОДАВЛЕНИЯ МИКРОБНЫХ КЛЕТОК | 2008 |
|
RU2528854C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |


































































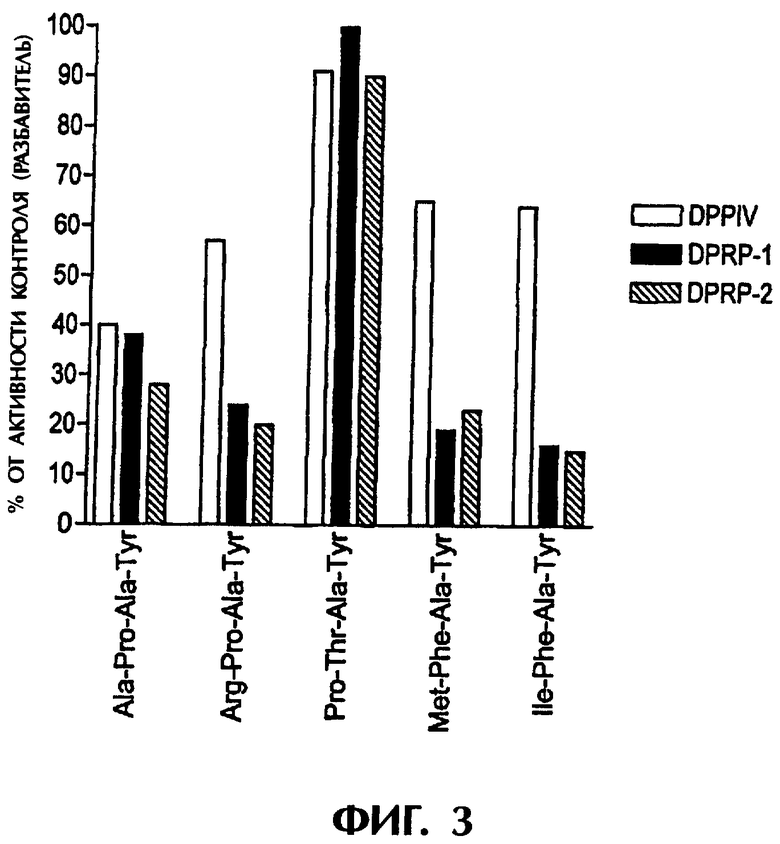
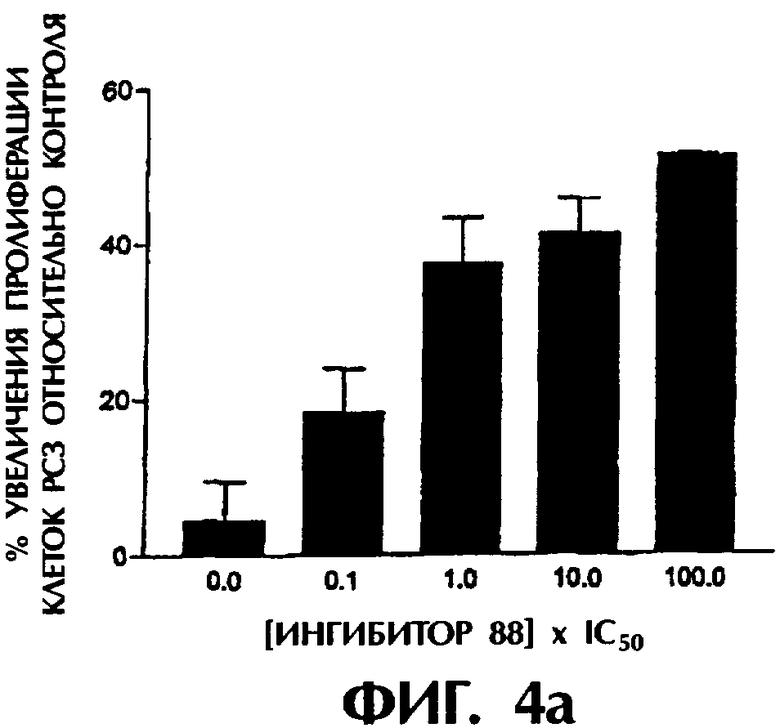
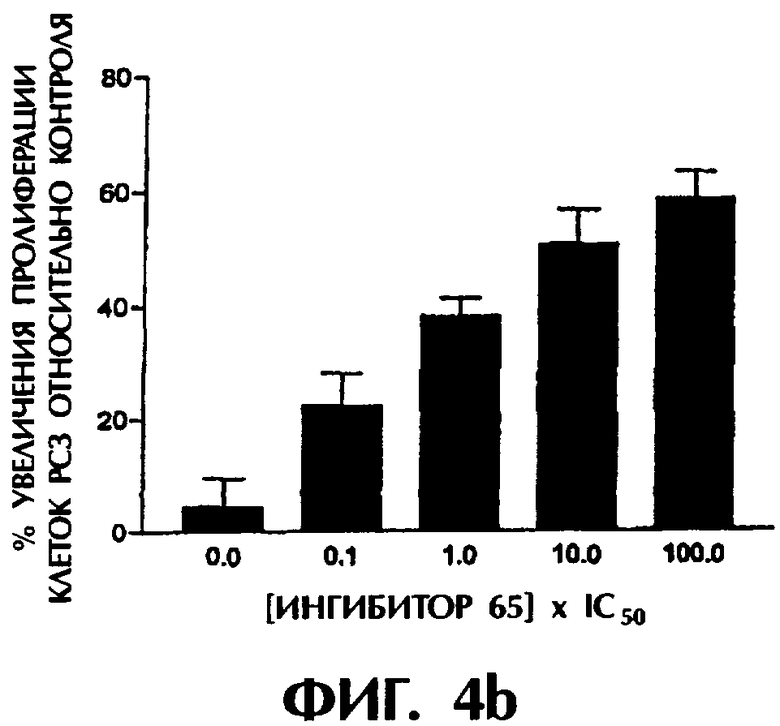
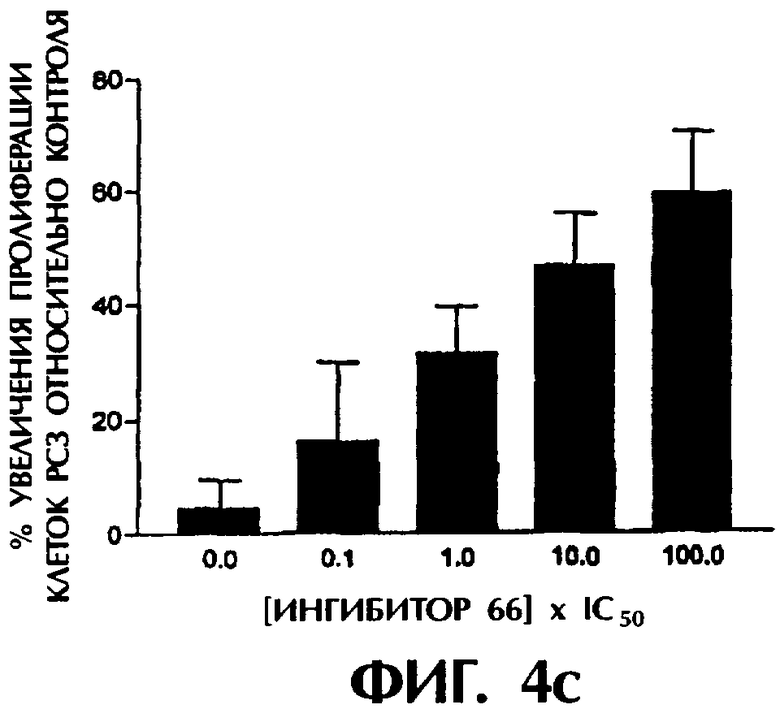
Изобретение относится к области биотехнологии и может быть использовано в фармакологии и медицине. Клонирована и определена последовательность ДНК, кодирующая новый вид сериновой протеазы человека (DPRP-2), родственной дипептидилолигопептидазе IV, гомология между которыми на уровне выведенной аминокислотной последовательности определена как 39%. С помощью технологии рекомбинантных ДНК получена рекомбинантная форма DPRP-2 и исследованы кинетические свойства нового фермента (субстратная специфичность, значения Km, ингибиторы активности и т.п.). Осуществление изобретения обеспечивает новые возможности при поиске и создании новых терапевтических средств для лечения широкого круга заболеваний человека. 2 н. и 3 з.п. ф-лы, 4 ил., 6 табл.
| АВВОТТ С | |||
| ет al | |||
| Eur | |||
| J | |||
| Biochem | |||
| Тепловой измеритель силы тока | 1921 |
|
SU267A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| DATABASE GenPept | |||
| Ac | |||
| no | |||
| Способ изготовления охотничьей дроби из чугуна | 1933 |
|
SU33801A1 |
| DATABASE GenPept | |||
| Ac | |||
| no | |||
| Головка для сифона | 1940 |
|
SU62840A1 |

