ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет по предварительной патентной заявке США 61/611485, поданной 15 марта 2012 года. Описание предыдущей заявки рассматривается в качестве части описания данной заявки (и включено в нее в качестве ссылки).
ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к выделенным молекулярным комплексам, обладающим активностью кислой альфа-глюкозидазы и, более подробно, к молекулярным комплексам, включающим, по меньшей мере, два полипептида, полученных протеолизом из прекурсорной молекулы, при этом молекулярный комплекс включает, по меньшей мере, одну модификацию, которая приводит к увеличению способности молекулярного комплекса транспортироваться внутрь клетки млекопитающего.
УРОВЕНЬ ТЕХНИКИ
Болезнь Помпе (также называемая болезнью накопления гликогена II типа или дефицита кислой мальтазы) представляет собой аутосомное рецессивное заболевание, которое приводит к накоплению гликогена в лизосоме вследствие дефицита альфа-глюкозидазы (GAA). Накопление гликогена вызывает прогрессирующее ослабление мышц (миопатию) по всему телу и воздействует на различные ткани тела, включая сердце, скелетные мышцы, печень и нервную систему.
Болезнь Помпе широко подразделяется по времени развития на инфантильную и взрослую форму. При инфантильной форме в рамках раннего детства (4-8 месяцев) дети обычно обладают слабостью и адинамией, не способны держать голову и демонстрировать иную моторную активность, характерную для их возраста, такую как переворачивание. Без лечения дети, страдающие болезнью Помпе, обычно умирают до возраста 12 месяцев вследствие сердечной недостаточности и респираторной слабости. См. United Pompe Foundation. Поздним формам (включая подростковые и взрослые формы) присуще позднее возникновение и более медленное прогрессирование, чем детским формам. Для лечения болезни Помпе применяют рекомбинантную GAA человека (Myozyme® или Lumizyme®). Однако стоимость Myozyme® или Lumizyme® является очень высокой и превышает $300000 в год. Таким образом, существует потребность в усовершенствованных лекарствах для лечения болезни Помпе.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте данное изобретение относится к выделенному молекулярному комплексу, обладающему активностью кислой альфа-глюкозидазы (GAA) и включающему, по меньшей мере, два полипептида (например, по меньшей мере, три или, по меньшей мере, четыре полипептида), при этом каждый полипептид обладает, по меньшей мере, 85% (например, по меньшей мере, 90%, 95%, 99% или 100%) идентичностью последовательности относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65). Молекулярный комплекс включает, по меньшей мере, одну модификацию, которая приводит к увеличению способности молекулярного комплекса транспортироваться внутрь клетки млекопитающего. Протеолиз аминокислотной последовательности, указанной в SEQ ID ΝΟ:1, также может включать расщепление в одном или нескольких сайтах между аминокислотой 719 и аминокислотой 746 или расщепление в одном или нескольких сайтах между аминокислотой 137 и аминокислотой 151 аминокислотной последовательности, указанной в SEQ ID NO:1. Протеолиз также может включать расщепление в одном или нескольких сайтах между аминокислотой 719 и аминокислотой 746 аминокислотной последовательности, указанной в SEQ ID NO:1, и расщепление в одном или нескольких сайтах между аминокислотой 137 и аминокислотой 151 аминокислотной последовательности, указанной в SEQ ID NO:1.
В любых описанных здесь молекулярных комплексах, по меньшей мере, один полипептид может включать один или несколько фосфорилированных N-гликанов, при этом модификация может включать декэпирование и деманнозилирование, по меньшей мере, одного фосфорилированного N-гликана. По меньшей мере, 40% (например, по меньшей мере, 60%, 80%, 90%, 95% или 99%) N-гликанов на, по меньшей мере, одном полипептиде могут быть декэпированными и деманнозилированными.
В любом описанном здесь молекулярном комплексе в одном из, по меньшей мере, двух полипептидов сегмент включает аминокислоты от 22 до 57 SEQ ID NO:1, при этом в одном из, по меньшей мере, двух полипептидов сегмент включает аминокислоты от 66 до 896 SEQ ID NO:1.
В любом описанном здесь молекулярном комплексе, содержащем, по меньшей мере, три полипептида, в одном из, по меньшей мере, трех полипептидов сегмент включает аминокислоты от 22 до 57 SEQ ID NO:1, при этом в одном из, по меньшей мере, трех полипептидов сегмент включает аминокислоты от 66 до 726 SEQ ID NO:1, при этом в одном из, по меньшей мере, трех полипептидов сегмент включает аминокислоты от 736 до 896 SEQ ID NO:1.
В любом описанном здесь молекулярном комплексе, содержащем, по меньшей мере, четыре полипептида, в одном из, по меньшей мере, четырех полипептидов сегмент включает аминокислоты от 22 до 57 SEQ ID NO:1, при этом в одном из, по меньшей мере, четырех полипептидов сегмент включает аминокислоты от 66 до 143 SEQ ID ΝΟ:1, при этом в одном из, по меньшей мере, четырех полипептидов сегмент включает аминокислоты от 158 до 726 SEQ ID NO:1, при этом в одном из, по меньшей мере, четырех полипептидов сегмент включает аминокислоты от 736 до 896 SEQ ID NO:1.
В любом описанном здесь аминокислотном комплексе, по меньшей мере, одна модификация может включать любой из указанных ниже, слитый, по меньшей мере, с одним полипептидом в молекулярном комплексе: лиганд для внеклеточного рецептора, таргетирующий домен, который связывает внеклеточный домен рецептора на поверхности клетки-мишени, плазминогенный рецептор урокиназного типа или распознающий домен инсулинподобного фактора роста II человека. Данное изобретение также относится к композициям, которые включают любой из описанных здесь молекулярных комплексов, при этом молекулярный комплекс является лиофилизированным. Композиция может быть упакована во флакон для одноразового применения.
Данное изобретение также относится к фармацевтической композиции, которая включает любой из описанных здесь молекулярных комплексов и фармацевтически приемлемый носитель. Композиция может быть составлена для внутривенного или подкожного введения. Композиция может быть составлена для внутривенной инфузии.
В другом аспекте изобретение относится к способу лечения болезни Помпе. Способ включает введение любой из описанных здесь композиций пациенту с диагностированной болезнью Помпе. Пациенту может быть диагностирована инфантильная форма болезни Помпе или взрослая форма болезни Помпе.
Данное изобретение также относится к способу получения молекулярного комплекса. Способ включает контактирование полипептида с, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:1, с протеазой с, по меньшей мере, 85% (например, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 99% или 100%) идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8, при этом протеаза расщепляет полипептид в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65). Этап контактирования может быть осуществлен in vitro.
Данное изобретение также относится к способу получения молекулярного комплекса, который включает декэпированные и деманнозилированные фосфорилированные N-гликаны. Способ включает контактирование молекулярного комплекса с маннозидазой, способной (i) гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата и (ii) гидролизовать терминальные связи с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой, при этом молекулярный комплекс обладает GAA активностью и включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65), при этом перед контактированием, по меньшей мере, один полипептид включает фосфорилированные N-гликаны, содержащие один или несколько маннозо-1-фосфо-6-маннозных остатков. Маннозидаза может представлять собой гликозил-гидролазу семейства 38 (например, Canavalia ensiformis mannosidase или Yarrowia lipolytica mannosidase). Контактирование может осуществляться в рекомбинантной грибковой клетке, экспрессирующей маннозидазу.
Данное изобретение также относится к способу получения молекулярного комплекса, который включает декэпированные и деманнозилированные фосфорилированные N-гликаны. Способ включает контактирование молекулярного комплекса с маннозидазой, способной гидролизовать терминальные связи с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой, при этом молекулярный комплекс обладает GAA активностью и включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно аминокислотного сегмента, указанного в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65), при этом перед контактированием, по меньшей мере, один полипептид включает фосфорилированные N-гликаны, включающие декэпированные маннозо-6-фосфатные остатки. Маннозидаза может представлять собой гликозил-гидролазу семейства 47 (например, маннозидазу Aspergillus satoi), гликозил-гидролазу семейства 92 (например, маннозидазу Cellulosimicrobium cellulans) или гликозил-гидролазу семейства 38 (например, маннозидазу Canavalia ensiformis). Контактирование может осуществляться в рекомбинантной грибковой клетке, экспрессирующей маннозидазу.
Данное изобретение также относится к способу получения молекулярного комплекса, который включает декэпированные и деманнозилированные фосфорилированные N-гликаны. Способ включает контактирование молекулярного комплекса с маннозидазой, способной гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата, при этом молекулярный комплекс обладает GAA активностью и включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно аминокислотного сегмента, указанного в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65), при этом перед контактированием, по меньшей мере, один полипептид включает один или несколько маннозо-1-фосфо-6-маннозных остатков. Маннозидаза может представлять собой гликозил-гидролазу семейства 38 (например, маннозидазу Canavalia ensiformis или маннозидазу Yarrowia lipolytica).
В другом аспекте данное изобретение относится к способу получения молекулярного комплекса, который включает декэпированные и деманнозилированные фосфорилированные N-гликаны. Способ включает а) контактирование молекулярного комплекса с маннозидазой, способной гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата для декэпирования маннозо-6-фосфатных остатков в, по меньшей мере, одном полипептиде молекулярного комплекса, при этом молекулярный комплекс обладает GAA активностью и включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно аминокислотного сегмента, указанного в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65); и b) контактирование молекулярного комплекса с маннозидазой, способной гидролизовать терминальные связи с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой. Этап (а) и этап (b) могут быть катализированы двумя различными ферментами или одним ферментом. Связанные с контактированием этапы могут быть осуществлены совместно или по отдельности и в любом порядке. Контактирование может осуществляться в рекомбинантной грибковой клетке-хозяине, при этом грибковая клетка-хозяин экспрессирует маннозидазу, способную катализировать этап (а), и маннозидазу, способную катализировать этап (b). Контактирование может осуществляться в рекомбинантной грибковой клетке-хозяине, при этом грибковая клетка-хозяин экспрессирует маннозидазу, способную катализировать этапы (а) и (b).
Любой из описанных здесь молекулярных комплексов, который включает, по меньшей мере, один декэпированный и деманнозилированный N-гликан, может применяться для контактирования клетки млекопитающего, при этом после контактирования молекулярный комплекс транспортируется внутрь клетки млекопитающего с увеличенной эффективностью. Клетка млекопитающего может представлять собой клетку человека.
Данное изобретение также относится к способу транспортирования молекулярного комплекса с GAA активностью внутрь клетки. Способ включает контактирование клетки млекопитающего с молекулярным комплексом, при этом молекулярный комплекс включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно аминокислотного сегмента, указанного в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65); при этом фосфорилированные N-гликаны в, по меньшей мере, одном полипептиде декэпированы и деманнозилированы как указано далее в описании способов. Клетка млекопитающего может быть in vitro или может находиться в субъекте, представляющем собой млекопитающее. Клетка млекопитающего может представлять собой клетку человека.
В другом аспекте, данное изобретение относится к способу транспортирования молекулярного комплекса с GAA активностью внутрь клетки. Способ включает контактирование клетки млекопитающего с молекулярным комплексом, который включает, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно аминокислотного сегмента, указанного в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65), при этом молекулярный комплекс включает, по меньшей мере, одну модификацию, которая приводит к увеличению способности молекулярного комплекса транспортироваться внутрь клетки млекопитающего. Клетка млекопитающего может быть in vitro или находится в субъекте, представляющем собой млекопитающее. Клетка млекопитающего может представлять собой клетку человека. Модификация может включать любой из указанных ниже, слитый, по меньшей мере, с одним полипептидом в молекулярном комплексе: лиганд для внеклеточного рецептора, таргетирующий домен, который связывает внеклеточный домен рецептора на поверхности клетки-мишени, плазминогенный рецептор урокиназного типа или распознающий домен инсулин подобного фактора роста II человека.
В другом аспекте данное изобретение относится к выделенной грибковой клетке, которая включает экзогенную нуклеиновую кислоту, кодирующую щелочную протеазу, обладающую, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8.
Данное изобретение также относится к выделенной грибковой клетке, включающей нуклеиновую кислоту, кодирующую GAA аминокислотную последовательность, указанную в SEQ ID NO:1, и нуклеиновую кислоту, кодирующую щелочную протеазу, обладающую, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8. Грибковая клетка продуцирует молекулярный комплекс, обладающий GAA активностью и включающий, по меньшей мере, два полипептида, при этом каждый полипептид обладает, по меньшей мере, 85% идентичностью последовательности относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65) щелочной протеазой. В некоторых вариантах осуществления грибковая клетка также включает нуклеиновую кислоту, кодирующую маннозидазу, при этом маннозидаза способна гидролизовать маннозо-1-фосфо-маннозный остаток с образованием маннозо-6-фосфата. В некоторых вариантах осуществления грибковая клетка также включает нуклеиновую кислоту, кодирующую маннозидазу, при этом маннозидаза способна гидролизовать терминальную связь с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой. В некоторых вариантах осуществления грибковая клетка также может включать нуклеиновую кислоту, кодирующую маннозидазу, при этом маннозидаза способна (i) гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата и (ii) гидролизовать терминальную связь с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой. Любая из подобных грибковых клеток также может включать нуклеиновую кислоту, кодирующую полипептид, способный промотировать маннозил-фосфорилирование и/или быть генетически модифицированным для устранения ОСН1 активности.
Если не указано иное, все используемые здесь технические и научные термины обладают тем же значением, которое обычно понимается средним специалистом в данной области, к которой принадлежит данное изобретение. Несмотря на то, что способы и материалы, аналогичные или эквивалентные тем, которые указаны здесь, могут применяться при применении на практике или тестировании настоящего изобретения, примерные способы и материалы описаны ниже. Все публикации, патентные заявки, патенты, каталожные номера Genbank® и другие приведенные здесь ссылки приведены для ссылки в их полноте. В случае конфликта настоящее изобретение, включая определения, обладает приоритетом. Материалы, способы и примеры приведены для иллюстрирования и не должны рассматриваться в качестве ограничивающих.
Другие признаки и преимущества изобретения будут очевидны исходя из нижеследующего подробного описания и пунктов формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 представляет собой изображение аминокислотной последовательности (SEQ ID NO:1) кислой альфа-глкжозидазы (GAA) человека после расщепления сигнальной последовательности.
Фиг. 2А представляет собой изображение нуклеотидной последовательности открытой рамки считывания (ORF) OsbA-Cellulosimicrobium cellulans маннозидазы 5 (CcMan5) (SEQ ID NO:2).
Фиг. 2В представляет собой изображение аминокислотной последовательности полипептида СсМап5 с выделенной сигнальной последовательностью (SEQ ID NO:3).
Фиг. 2С представляет собой изображение аминокислотной последовательности полипептида CcMan5 без сигнальной последовательности (SEQ ID NO:4). Предполагаемая молекулярная масса полипептида CcMan5 без сигнальной последовательности составляет 173 кДа.
Фиг. 3А и 3В представляют собой серию электрофореграмм, изображающих анализ N-гликанов rhGAA, подвергаемой действию CcMan5 и JbMan. Анализ осуществляли с помощью углеводного электрофореза с применением секвенирования ДНК и флюорофора (DSA-FACE). Ось Y соответствует относительному количеству единиц флюоресценции в качестве показателя количества каждой структуры N-гликанов. Ось X соответствует относительной капиллярной мобильности каждой структуры N-гликанов. На Фиг. 3А и Фиг. 3В изображение А представляет собой сравнительный образец, содержащий N-гликаны, высвобожденные из RNaseB с помощью PNGaseF. На Фиг. 3А изображения В и С содержат анализ N-гликанов из huGAA (76 кДа вариант) до и после обработки, соответственно, CcMan5 и JbMan.
На Фиг. 3В изображения В, С и D содержат анализ N-гликанов из 76 кДа формы huGAA, 95 кДа формы и 110 кДа формы, соответственно.
Фиг. 4 представляет собой линейный график количества глюкозы, образующейся в минуту с применением Myozyme (⋅), 76 кДа GAA (▲), 95 кДа GAA (▼) и 110 кДа GAA (♦) с применением гликогена печени кролика в качестве субстрата.
Фиг. 5А содержит два изображения уровней гликогена (мкг/мг белка) в сердце отдельной мыши. Фиг. 5В содержит два изображения уровней гликогена (мкг/мг белка) отдельной мыши в скелетной мышце. Красные точки означают самок, черные точки - самцов. Линии означают медиану в каждой группе.
Фиг. 6 содержит изображение аминокислотной последовательности маннозидазы AMS1 Yarrowia lipolytica (SEQ ID NO:5).
Фиг. 7 содержит изображение аминокислотной последовательности маннозидазы Aspergillus satoi (SEQ ID NO:6).
Фиг. 8 содержит изображение аминокислотной последовательности маннозидазы 4 Cellulosimicrobium cellulans (CcMan4, SEQ ID NO:7) с выделенной сигнальной последовательностью. Предполагаемая молекулярная масса полипептида CcMan4 без сигнальной последовательности составляет 184 кДа.
Фиг. 9 содержит изображение аминокислотной последовательности щелочной протеазы Aspergillus oryzae, включающей сигнальный пептид (21 аминокислоты), про-пептид (100 аминокислот) и зрелый белок (282 аминокислоты) (SEQ ID NO:9).
Фиг. 10 содержит изображение нуклеотидной последовательности слитого конструкта, содержащего кодон-оптимизированную последовательность Y. lipolytica, кодирующую щелочную протеазу A. oryzae (SEQ ID NO:10). Применяющиеся для клонирования сайты рестрикции подчеркнуты. Нуклеотидная последовательность, кодирующая линкер, выделена, при этом нуклеотидная последовательности, кодирующая метку His (10 His остатков) указана курсивом.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В целом данное изобретение относится к выделенным молекулярным комплексам, обладающим активностью кислой альфа-глюкозидазы (GAA) и, по меньшей мере, одной модификацией, которая приводит к увеличению способности транспортироваться внутрь клетки млекопитающего. GAA синтезируется в виде прекурсора массой 110 кДа, содержащего N-связанные гликаны. Прекурсор подвергается протеолитической обработке для удаления сигнальной последовательности и затем подвергается протеолитической обработке с образованием крупных образцов массой 95 кДа, 76 кДа и 70 кДа. Однако, по меньшей мере, некоторые пептиды, которые высвобождаются из прекурсора, остаются связанными с крупными образцами. См., например, Moreland et al., J. Biol. Chem., 280:6780-6791 (2005). Таким образом, описанные здесь обладающие GAA активностью молекулярные комплексы включают, по меньшей мере, два полипептида (по меньшей мере, два, три или четыре полипептида), которые получены с помощью протеолитического расщепления прекурсорной молекулы в одном или нескольких сайтах. По меньшей мере, два полипептида в молекулярном комплексе являются полученными в результате протеолитического расщепления в одном или нескольких сайтах в прекурсоре. Например, протеолиз аминокислотной последовательности, указанной в SEQ ID NO:1, может осуществляться между аминокислотой 50 и аминокислотой 74, например между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65 для продуцирования, по меньшей мере, двух полипептидов. Молекулярный комплекс, содержащий два полипептида, обозначен здесь как форма массой 95 кДа.
В некоторых вариантах осуществления, по меньшей мере, три полипептида в молекулярном комплексе являются полученными в результате протеолитического расщепления в двух или нескольких сайтах в прекурсоре.
Например, протеолиз аминокислотной последовательности, указанной в SEQ ID NO:1, может включать, помимо расщепления между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 50 и аминокислотой 74 или между аминокислотой 60 и аминокислотой 65), расщепления в одном или нескольких сайтах между аминокислотой 719 и аминокислотой 746 или расщепление в одном или нескольких сайтах между аминокислотой 137 и аминокислотой 151 аминокислотной последовательности, указанной в SEQ ID NO:1. Молекулярный комплекс, содержащий три полипептида, обозначен здесь как форма массой 76 кДа.
В некоторых вариантах осуществления, по меньшей мере, четыре полипептида в молекулярном комплексе являются полученными в результате протеолитического расщепления в трех или нескольких сайтах в прекурсоре. Например, протеолиз аминокислотной последовательности, указанной SEQ ID NO:1, может включать, помимо расщепления между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65), расщепление в одном или нескольких сайтах между аминокислотой 719 и аминокислотой 746 аминокислотной последовательности, указанной в SEQ ID NO:1, и расщепление в одном или нескольких сайтах между аминокислотой 137 и аминокислотой 151 аминокислотной последовательности, указанной в SEQ ID NO:1. Молекулярный комплекс, содержащий четыре полипептида, обозначен здесь как форма массой 70 кДа.
Будет являться очевидным, что расщепление может осуществляться в одном или нескольких сайтах в одной молекуле и что сайты расщепления могут являться различными в разных молекулах.
Коммерчески доступная протеазная смесь, содержащая протеазы Aspergillus oryzae (например, от Sigma или NovozymesCorp) может применяться для расщепления аминокислотной последовательности, указанной в SEQ ID NO:1, между аминокислотами 50 и 74, например между аминокислотами 56 и 68 или между аминокислотами 60 и 65. Альтернативно, может применяться щелочная протеаза, обладающая, по меньшей мере, 85% (например, по меньшей мере, 95%, 97%, 98%, 99% или 100%) идентичностью последовательности относительно щелочной протеазы Aspergillus oryzae (SEQ ID NO:8). Например, как описано здесь, GAA полипептид, обладающий аминокислотной последовательностью, указанной в SEQ ID NO:1, может контактировать с протеазой, обладающей 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8 или SEQ ID NO:9. SEQ ID NO:8 представляет собой аминокислотную последовательность зрелой щелочной протеазы Aspergillus oryzae. SEQ ID NO:9 представляет собой аимнокислотную последовательность протеазы Aspergillus oryzae, включающей сигнальный пептид, про-пептид и зрелый белок. Контактирование может осуществляться in vitro с применением протеазы, которая была выделена из Aspergillus oryzae или была получена с помощью рекомбинантных способов. Альтернативно, хозяин, представляющий собой грибок, может быть модифицирован таким образом, что GAA полипептид и щелочная протеаза секретируются в культуральную среду, в которой щелочная протеаза может расщеплять аминокислотную последовательность, указанную в SEQ ID NO:1, между аминокислотой 50 и аминокислотой 74 (например, между аминокислотами 56 и 68 или между аминокислотой 60 и аминокислотой 65).
Описанные здесь выделенные молекулярные комплексы обладают, по меньшей мере, одной модификацией, которая приводит к увеличению способности транспортироваться внутрь клетки млекопитающего. Неограничивающие примеры модификаций, которые увеличивают способность комплекса транспортироваться внутрь клетки млекопитающего, включают декэпирование и деманнозилирование фосфорилированных N-гликанов или пептидных меток, которые обеспечивают транспорт. Способы и материалы описаны здесь для получения молекулярных комплексов, содержащих метки или декэпированные или деманнозилированные N-гликаны.
Выделенные описанные здесь молекулярные комплексы особенно применимы для лечения пациентов с болезнью Помпе, включая пациента с диагностированной инфантильной и взрослой формой болезни Помпе. Болезнь Помпе приводит к накоплению гликогена в лизосоме вследствие дефицита GAA. Накопление гликогена вызывает прогрессирующее ослабление мышц (миопатию) по всему телу и воздействует на различные ткани тела, включая сердце, скелетные мышцы, печень и нервную систему. Каждый полипептид в молекулярном комплексе обладает, по меньшей мере, 85% идентичностью последовательности (например, по меньшей мере, 90%, 95%, 97%, 98%, 99% или 100%о) относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65). Степень идентичности в процентах между определенной аминокислотной последовательностью и аминокислотной последовательностью, указанной в SEQ ID NO:1, может быть определена следующим образом. Сначала аминокислотные последовательности выравнивают с помощью программы BLAST 2 Sequences (Bl2seq) из выделенной версии BLASTZ, содержащей BLASTP версии 2.0.14. Данная выделенная версия BLASTZ может быть загружена с вебсайта Fish & Richardson (например, www.fr.com/blast/) или вебсайта Национального центра биотехнологической информации правительства США (National Center for Biotechnology Information) (www.ncbi.nlm.nih.gov). Инструкции, объясняющие использование программы Bl2seq, могут быть найдены в файле readme, сопровождающем BLASTZ. Bl2seq осуществляет сравнение между двумя аминокислотными последовательностями с применением алгоритма BLASTP. Для сравнения двух аминокислотных последовательностей опции Bl2seq установлены следующим образом: -i соответствует файлу, содержащему первую аминокислотную последовательность для сравнения (например, C:\seq1.txt); -j соответствует файлу, содержащему вторую аминокислотную последовательность для сравнения (например, C:\seq2.txt); -р соответствует blastp; -о соответствует любому желаемому имени файла (наппример, C:\output.txt); а все другие опции оставлены по умолчанию. Например, следующая команда может применяться для генерации выходного файла, содержащего сравнение между двумя аминокислотными последовательностями: C:\Bl2seq -i c:\seq1.txt -j c:\seq2.txt -p blastp -о c:\output.txt. Если две сравниваемые последовательности обладают гомологией, то полученный выходной файл будет включать данные участки гомологии в виде выравненных последовательностей. Если две сравниваемые последовательности не обладают гомологией, то полученный выходной файл не будет включать выравненные последовательности. Аналогичные процедуры могут применяться для последовательностей нуклеиновых кислот за исключением применения blastn.
После проведения выравнивания количество совпадений определяется путем подсчета количества позиций, в которых идентичные аминокислоты присутствуют в обеих последовательностях. Идентичность в процентах определяется путем деления количества совпадений на длину полноразмерной полипептидной аминокислотной последовательности с последующим умножением полученного результата на 100.
Следует отметить, что значение идентичности в процентах округляется до ближайшей одной десятой. Например, 78,11, 78,12, 78,13 и 78,14 округляются до 78,1, в то время как 78,15, 78,16, 78,17, 78,18, и 78,19 округляются до 78,2. Следует также отметить, что значение длины всегда будет представлять собой целое число.
Является очевидным, что ряд нуклеиновых кислот может кодировать полипептид с определенной аминокислотной последовательностью. Дегенерация генетического кода хорошо известна в данный области, то есть в отношении многих аминокислот существует более чем один триплет, который служит в качестве кодона для аминокислоты. Например, кодоны в кодирующей последовательности какого-либо GAA полипептида могут быть модифицированы таким образом, что достигается оптимальная экспрессия в определенном виде (например, бактериях или грибах) с применением подходящих таблиц смещения кодонов для данного вида.
В одном варианте осуществления молекулярный комплекс может включать, по меньшей мере, два полипептида, при этом один полипептид включает аминокислоты от 22 до 57 SEQ ID NO:1, а другой полипептид включает аминокислоты от 66 до 896 SEQ ID NO:1.
В одном варианте осуществления молекулярный комплекс может включать, по меньшей мере, три полипептида, при этом один полипептид включает аминокислоты от 22 до 57 SEQ ID NO:1, один полипептид включает аминокислоты от 66 до 726 SEQ ID NO:1 и один полипептид включает аминокислоты от 736 до 896 SEQ ID NO:1.
В одном варианте осуществления молекулярный комплекс может включать, по меньшей мере, четыре полипептида, при этом один полипептид включает аминокислоты от 22 до 57 SEQ ID NO:1, один полипептид включает аминокислоты от 66 до 143 SEQ ID NO:1, один полипептид включает аминокислоты от 158 до 726 SEQ ID NO:1 и один полипептид включает аминокислоты от 736 до 896 SEQ ID NO:1.
Биологически активные варианты GAA могут содержать добавления, делеции или замещения относительно последовательностей, указанных в SEQ ID NO:1. GAA белки с замещениями в целом будут обладать не более чем 10 (например, не более чем одним, двумя, тремя, четырьмя, пятью, шестью, семью, восемью, девятью или десятью) консервативными аминокислотными замещениями. Консервативное замещение представляет собой замещение одной аминокислоты на другую с теми же характеристиками. Консервативные замещения включают замещения в рамках следующих групп: валин, аланин и глицин; лейцин, валин и изолейцин; аспарагиновая кислота и глутаминовая кислота; аспарагин и глутамин; серии, цистеин и треонин; лизин и аргинин; и фенилаланин и тирозин. Неполярные гидрофобные аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин. Полярные нейтральные аминокислоты включают глицин, серии, треонин, цистеин, тирозин, аспарагин и глутамин. Положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин. Отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту. Любое замещение одного члена вышеупомянутых полярной, основной или кислой групп другим членом той же группы может рассматриваться в качестве консервативного замещения. С другой стороны, неконсервативное замещение представляет собой замещение одной аминокислоты на другую с иными характеристиками.
В некоторых вариантах осуществления GAA полипептид может представлять собой слитый белок с гетерологичной аминокислотной последовательностью, такой как последовательность, применяемая для очистки рекомбинантного белка (например, FLAG, полигистидин (например, гексагистидин), гемаглютенин (НА), гемаглютенин-S-трансфераза (GST) или связывающий мальтозу белок (МВР)).
В некоторых вариантах осуществления гетерологичная аминокислотная последовательность применяется для увеличения эффективности транспорта молекулярного комплекса в клетку млекопитающего. Например, по меньшей мере, один из полипептидов в комплексе может быть слит с лигандом для внеклеточного рецептора, таргетирующим доменом, который связывает внеклеточный домен рецептора на поверхности клетки-мишени, плазминогенным рецептором урокиназного типа или доменами инсулин подобного фактора роста II человека, который связывается с маннозо-6-фосфатным рецептором (например, аминокислоты 1-67 или 1-87; по меньшей мере, аминокислоты 48-55; по меньшей мере, аминокислоты 8-28 и 41-61; или, по меньшей мере, аминокислоты 8-87 инсулинподобного фактора роста человека; вариант последовательности инсулинподобного фактора роста II человека (например, R68A) или усеченной формы инсулинподобного фактора роста человека (например, усеченного на С-конце в положении 62)). Гетерологичная аминокислотная последовательность может быть слитой на N-конце или С-конце полипептида. В одном варианте осуществления пептидная метка слита с N- или С-концом полипептида с помощью спейсера (например, 5-30 аминокислотами или 10-25 аминокислотами). См., например, патент США 7785856. Гетерологичные аминокислотные последовательности также могут представлять собой белки, которые могут применяться в качестве диагностических или определяемых маркеров, например люциферазу, зеленый флюоресцентный белок (GFP) или хлорамфеникол-ацетилтрансферазу (CAT).
В определенных клетках-хозяевах (например, дрожжевых клетках-хозяевах) экспрессия и/или секреция белка-мишени может быть увеличена за счет применения гетерологичной сигнальной последовательности. В некоторых вариантах осуществления слитый белок может содержать носитель (например, KLH), который может применяться, например, при продуцировании иммунного ответа на генерацию антител) или эндоплазматический ретикулум или сигналы удержания аппарата Гольджи. Гетерологичные последовательности могут иметь различную длину и в некоторых случаях могут представлять собой более длинные последовательности, чем полноразмерные белки-мишени, с которыми соединяются гетерологичные последовательности.
Способы деманнозилированш или декэпирование и деманнозширование гликопротеинов Гликопротеины, содержащие фосфорилированный N-гликан, могут быть деманнозилированными, а гликопротеины, содержащие фосфорилированный N-гликан, содержащий маннозо-1-фосфо-6-маннозную связь или остаток, могут быть декэпированы и деманнозилированы путем контактирования гликопротеина с маннозидазой, способной (i) гидролизовать маннозо-1-фосфо-6-маннозную связь или остаток с образованием маннозо-6-фосфата и (ii) гидролизовать терминальную альфа-1,2-маннозную, альфа-1,3-маннозную и/или альфа-1,6-маннозную связь или остаток. Неограничивающие примеры подобных маннозидаз включают маннозидазу Canavalia ensiformis (канавалия мечевидная) и маннозидазу Yarroxvia lipolytica (например, AMS1). Маннозидазы канавалии мечевидной и AMS1 представляют собой гликозид-гидролазы семейства 38.
Маннозидаза канавалии мечевидной является коммерчески доступной, например, от Sigma-Aldrich (St. Louis, МО) в виде сульфат-аммонийной суспензии (каталожный номер М7257) и препарата протеомического класса (каталожный номер М5573). Данные коммерческие препараты могут быть далее очищены, например, с помощью гель-фильтрационной хроматографии для удаления загрязнителей, таких как фосфатазы. Маннозидаза канавалии мечевидной содержит сегмент со следующей аминокислотной последовательностью NKIPRAGWQIDPFGHSAVQG (SEQ ID NO:11). См. Howard et al., J. Biol. Chem., 273(4):2067-2072, 1998.
Маннозидаза AMS1 Yarrowia lipolytica может быть получена с помощью рекомбинантных способов. Аминокислотная последовательность AMS1 полипептида указана в see also FIG. 5 (см. также Фиг. 6).
В некоторых вариантах осуществления этапы декэпирования и деманнозилирования катализируются двумя различными ферментами. Например, декэпирование маннозо-1-фосфо-6-маннозной связи или остатка может быть осуществлено с применением маннозидазы из Cellulosimicrobium cellulans (например, CcMan5). Нуклеотидная последовательность, кодирующая СсМап5 полипептид, указана в SEQ ID NO:2 (см. Фиг. 2А). Аминокислотная последовательность CcMan5 полипептида, содержащего сигнальную последовательность, указана в SEQ ID NO:3 (см. Фиг. 2В). Аминокислотная последовательность CcMan5 полипептида без сигнальной последовательности указана в SEQ ID NO:4 (см. Фиг. 2С). В некоторых вариантах осуществления применяется биологически активный фрагмент CcMan5 полипептида. Например, биологически активный фрагмент может включать остатки 1-774 аминокислотной последовательности, указанной в SEQ ID NO:4. См. также WO 2011/039634. CcMan5 маннозидаза представляет собой гликозид-гидролазу семейства 92.
Деманнозилирование декэпированного гликопротеина или молекулярных комплексов гликопротеинов может быть катализировано с помощью маннозидазы из Aspergillus satoi (As) (также известного как Aspergillus phoenicis) или маннозидазы из Cellulosimicrobium cellulans (например, CcMan4). Маннозидаза Aspergillus satoi представляет собой гликозид-гидролазу семейства 47, а CcMan4 маннозидаза представляет собой гликозид-гидролазу семейства 92. Аминокислотная последовательность маннозидазы Aspergillus satoi указана на Фиг. 7 (SEQ ID NO:6) и под каталожным номером GenBank ВАА08634. Аминокислотная последовательность CcMan4 полипептида указана на Фиг. 8 (SEQ ID NO:7).
Деманнозилирование декэпированного гликопротеина или молекулярных комплексов гликопротеинов также может быть катализировано с помощью маннозидазы семейства 38 гликозид-гидролаз, такой как маннозидаза Canavalia ensiformis (канавалия мечевидная) или маннозидаза Yarrowia lipolytica (например, AMS1). Например, CcMan5 может применяться для декапирования маннозо-1-фосфо-6-маннозного остатка на гликопротеине (или молекулярном комплексе гликопротеинов), а маннозидаза канавалии мечевидной может применяться для деманнозилирования декэпированного гликопротеина (или молекулярных комплексов гликопротеинов).
Для получения деманнозилированных гликопротеинов (или молекулярных комплексов гликопротеинов) или декэпированных и деманнозилированных гликопротеинов (или молекулярных комплексов гликопротеинов) молекула-мишень (или молекулярный комплекс), содержащая маннозо-1-фосфо-6-маннозную связь или остаток контактирует при подходящих условиях с подходящей маннозидазой(ами) и/или клеточным лизатом, содержащим подходящую рекомбинантно полученную маннозидазу(ы). Подходящие маннозидазы описаны выше. Клеточный лизат может быть получен из любой генетически модифицированной клетки, включая грибковую клетку, растительную клетку или клетку животного. Неограничивающие примеры клеток животных включают клетки нематоды, насекомого, растения, птицы, рептилии и млекопитающих, таких как мышь, крыса, кролик, хомяк, песчанка, собака, кошка, коза, свинья, корова, лошадь, кит, обезьяна или человек.
При контактировании молекулы-мишени (например, молекулярного комплекса) с очищенными маннозидазами и/или клеточным лизатом маннозо-1-фосфо-6-маннозная связь или остаток могут быть гидролизованы с образованием фосфо-6-маннозы, при этом терминальная альфа-1,2-маннозная, альфа-1,3-маннозная и/или альфа-1,6-маннозная связь или остаток данного фосфата, содержащего гликан, могут быть гидролизованы для получения декэпированной и деманнозилированной молекулы-мишени. В некоторых вариантах осуществления применяется одна маннозидаза, которая катализирует этапы декэпирования и деманнозилирования. В некоторых вариантах осуществления применяется одна маннозидаза для катализирования этапа декэпирования и применяется иная маннозидаза для катализирования этапа деманнозилирования. После обработки с помощью маннозидазы молекула-мишень или молекулярный комплекс могут быть выделены.
Подходящие способы получения клеточных лизатов, которые сохраняют активность или целостность маннозидазной активности в лизате, могут включать применение подходящих буферов и/или ингибиторов, включая нуклеазные, протеазные и фосфатазные ингибиторы, которые сохраняют или минимизируют изменения активности N-гликозилирования в клеточном лизате. Данные ингибиторы включают, например, хелаты, такие как этилендиаминтетрауксусная кислота (ЭДТА), этиленгликольбис(Р-аминоэтиловый эфир)N,N,N1,N1-тетрауксусная кислота (EGTA), протеазные ингибиторы, такие как фенилметилсульфонилфторид (PMSF), апротинин, leu-пептин, антипаин и им подобные, и фосфатазные ингибиторы, такие как фосфат, фторид натрия, ванадат и им подобные. Подходящие буферы и условия для получения лизатов, обладающих ферментативной активностью, описаны, например, у Ausubel et al. Current Protocols in Molecular Biology (Supplement 47), John Wiley & Sons, New York (1999); Harlow and Lane, Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press (1988); Harlow and Lane, Using Antibodies: A Laboratory Manual, Cold Spring Harbor Press (1999); Tietz Textbook of Clinical Chemistry, 3-е изд. Burtis and Ashwood, eds. W.B. Saunders, Philadelphia, (1999).
Клеточный лизат может быть также обработан для удаления или минимизации присутствия нежелательных веществ, если это необходимо. В случае необходимости клеточный лизат может быть подвергнут фракционированию с помощью различных способов, хорошо известных специалистам в данной области, включая субклеточное фракционирование и хроматографические способы, такие как ионный обмен, гидрофобная и обратная фаза, эксклюзия на основе размера, аффинная, гидрофобная заряд-индуцирующая хроматография и им подобных.
В некоторых вариантах осуществления может быть получен клеточный лизат, в котором цельные клеточные органеллы остаются интактными и/или функциональными. Например, лизат может содержать одну или несколько органелл из интактного гранулярного эндоплазматического ретикулума, интактного гладкого эндоплазматического ретикулума или интактного аппарата Гольджи. Подходящие способы получения лизатов, содержащих интактные клеточные органеллы, и тестирования на предмет функциональности органелл описаны, например, у Moreau et al. (1991) J. Biol. Chem. 266(7):4329-4333; Moreau et al. (1991) J. Biol. Chem. 266(7):4322-4328; Rexach et al. (1991) J. Cell Biol. 114(2):219-229; и Paulik et al. (1999) Arch. Biochem. Biophys. 367(2):265-273.
При контактировании клетки млекопитающего с молекулярным комплексом, содержащим декэпированные и деманнозилированные фосфорилированные N-гликаны молекулярный комплекс может быть транспортирован внутрь клетки млекопитающего (например, клетки человека). Молекулярный комплекс, обладающий декэпированным, но не деманнозилированным фосфорилированным N-гликаном в значительной степени не связывает маннозо-6-фосфатные рецепторы на клетках млекопитающего, и, таким образом, не подвергается эффективному транспорту внутрь клетки. В соответствии с используемым здесь значением, "в значительной степени не связывает" означает, что менее чем 15% (например, менее чем 14%, 12%, 10%, 8%, 6%, 4%, 2%, 1%, 0,5% или менее или 0%) молекул гликопротеина связывается с маннозо-6-фосфатными рецепторами на клетке млекопитающего. Тем не менее, если данный молекулярный комплекс контактирует с маннозидазой, способной гидролизовать терминальную альфа-1,2-маннозную связь или остаток, когда нижележащая манноза является фосфорилированной, продуцируется деманнозилированный гликопротеин, который в значительной степени связывается с маннозо-6-фосфатным рецептором на клетках млекопитающего и эффективно транспортируется внутрь клетки. В соответствии с используемым здесь значением, "в значительной степени связывается" означает, что 15% или более (например, более чем 16%, 18%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%), 75%, 80%, 85%), 90% или 95%) молекулярного комплекса связывается с маннозо-6-фосфатными рецепторами на клетках млекопитающего. Является очевидным, что препарат (например, препарат на основе рекомбинантных клеток-хозяев или препарат без клеток), содержащий фермент, который декэпирует, но не деманнозилирует фосфорилированные N-гликаны, может быть загрязнен ферментом, который деманнозилирует фосфорилированные N-гликаны. Образец белка-мишени после контакта с подобным препаратом может содержать белковые молекулы с некоторыми фосфорилированными N-гликанами, которые являются лишь декэпированными, и другие, которые являются декэпированными и деманнозилированными. Данные белковые молекулы, содержащие декэпированные и деманнозилированные фосфорилированные N-гликаны могут естественным образом в значительной степени связываться с маннозо-6-фосфатными рецепторами. Приведенное выше определение "в значительной степени не связывает" не применимо к подобному образцу белка-мишени, поскольку фосфорилированные N-гликаны на белковых молекулах не могут быть охарактеризованы в качестве декэпированных, но не деманнозилированных.
Таким образом, данное изобретение относится к способам конвертации молекулярного комплекса из первой формы, которая не связывает маннозо-6-фосфатный рецептор на клетки млекопитающего во вторую форму, которая связывает маннозо-6-фосфатный рецептор на клетке млекопитающего. В первой форме молекулярный комплекс, в котором, по меньшей мере, один из полипептидов в комплексе включает один или несколько N-гликанов, содержащих один или несколько маннозных остатков, которые связаны в 1 положении с маннозным остатком, который содержит фосфатный остаток в 6 положении. В рамках данных способов первая форма молекулярного комплекса контактирует с маннозидазой, которая деманнозилирует терминальные маннозные остатки, что приводит к тому, что манноза, содержащая фосфат в 6 положении, становится терминальной маннозой. В некоторых вариантах осуществления маннозидаза обладает декэпирующей и деманнозилирующей активностью (например, маннозидаза Canavalia ensiformis (канавалия мечевидная) или маннозидаза AMS1 Yarrowia lipolytica). В некоторых вариантах осуществления маннозидаза не обладает декэпирующей активностью (например, маннозидаза из Aspergillus satoi или маннозидаза из Cellulosimicrobium cellulans (например., CcMan4)).
Транспорт гликопротеина или молекулярного комплекса внутрь клетки может быть проанализирован с помощью анализа их включения клеткой. Например, клетки млекопитающего и молекулярный комплекс, содержащий декэпированные и деманнозилированные фосфорилированные N-гликаны, может быть культивирован с последующей промывкой и лизисом клеток. Клеточные лизаты могут быть проанализированы на предмет присутствия GAA комплекса (например, с помощью вестерн-блота) или на основе активности GAA в клеточном лизате. Например, включение может быть проанализировано в фибробластах с дефицитом активности кислой альфа-глюкозидазы. Внутриклеточная активность альфа-глюкозидазы может быть проанализирована с помощью анализа с применением 4-метилумбеллиферид-альфа-D-глюкопиранозида (4-MUG). Расщепление субстратного 4-MUG с помощью глюкозидазы ведет к образованию фторогенного продукта 4-MU, который может быть визуализирован или определен с помощью облучения УФ излучением.
Рекомбинантное получение полипептидов
Выделенные молекулы нуклеиновой кислоты, кодирующие полипептиды (например, маннозидазу, щелочную протеазу или GAA или ее фрагмент), могут быть получены с помощью стандартных способов. Термины "нуклеиновая кислота" или "полинуклеотид" используются здесь взаимозаменяемо и означают РНК и ДНК, включая кДНК, геномную ДНК, синтетическую ДНК и ДНК (или РНК), содержащую аналоги нуклеиновой кислоты. Полинуклеотиды могут иметь любую трехмерную структуру. Нуклеиновая кислота может быть одноцепочечной или двухцепочечной (например, смысловой спиралью или антисмысловой спиралью). Неограничивающие примеры полинуклеотидов включают гены, фрагменты генов, экзоны, интроны, информационную РНК (мРНК), транспортную РНК, рибосомную РНК, малую интерферирующую РНК, микро-РНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенную ДНК любой последовательности, выделенную РНК любой последовательности, нуклеотидные зонды и праймеры, а также аналоги нуклеиновой кислоты.
"Выделенная нуклеиновая кислота" означает нуклеиновую кислоту, которая отделена от других молекул нуклеиновой кислоты, которые присутствуют во встречающемся в природе геноме, включая нуклеиновые кислоты, которые обычно фланкируют одну или обе стороны нуклеиновой кислоты во встречающемся в природе геноме (например, геноме дрожжей). Термин "выделенный" в соответствии с используемым здесь значением в отношении нуклеиновых кислот также включает любую не являющуюся встречающейся в природе последовательность нуклеиновой кислоты, поскольку данные не встречающиеся в природе последовательности не обнаружены в природе и не обладают непосредственно фланкирующими последовательностями во встречающемся в природе геноме.
Выделенная нуклеиновая кислота может представлять собой, например, молекулу ДНК в случае, если одна из последовательностей нуклеиновой кислоты, обычно непосредственно фланкирующая данную ДНК молекулу во встречающемся в природе геноме, удалена или отсутствует.Таким образом, выделенная нуклеиновая кислота включает, без ограничения, ДНК молекулу, которая существует в виде отдельной молекулы (например, химически синтезированной нуклеиновой кислоты или кДНК или фрагмента геномной ДНК, продуцируемой с помощью ПЦР или обработки эндонуклеазой рестрикции), являющейся независимой от других последовательностей, а также ДНК, которая встроена в вектор, автономно реплицирующуюся плазмиду, вирус (например, любой парамиксовирус, ретровирус, лентивирус, аденовирус или вирус герпеса) или в геномную ДНК прокариота или эукариота. Кроме того, выделенная нуклеиновая кислота может включать модифицированную нуклеиновую кислоту, такую как ДНК молекула, которая является частью гибридной или слитой нуклеиновой кислоты. Нуклеиновая кислота, существующая среди от сотен до миллионов других нуклеиновых кислот, например в библиотеках кДНК или геномных библиотеках или гелевых слайсах, содержащих рестрикционное картирование геномных ДНК, не рассматривается в качестве выделенной нуклеиновой кислоты.
Термин "экзогенный" в соответствии с используемым здесь значением в отношении нуклеиновой кислоты и определенных клеток-хозяев означает любую нуклеиновую кислоту, которая не встречается (и не может быть получена) из данной определенной клетки во встречающемся в природе виде. Таким образом, не встречающиеся в природе нуклеиновые кислоты рассматриваются в качестве экзогенных по отношению к клетке-хозяину при введении в клетку-хозяина. Важно отметить, что не встречающиеся в природе нуклеиновые кислоты могут содержать субпоследовательности нуклеиновой кислоты или фрагменты последовательностей нуклеиновой кислоты, которые встречаются в природе при условии, если нуклеиновая кислота в целом виде не существует в природе. Например, молекула нуклеиновой кислоты, содержащая последовательность геномной ДНК в векторе экспрессии, представляет собой не встречающуюся в природе нуклеиновую кислоту и, таким образом, является экзогенной по отношению к клетке-хозяину при введении в клетку-хозяина, поскольку молекула нуклеиновой кислоты в целом (геномная ДНК плюс векторная ДНК) не существует в природе. Таким образом, любой вектор, автономно реплицирующаяся плазмида или вирус (например, ретровирус, аденовирус или вирус герпеса), которые в целом не существуют в природе, рассматриваются в качестве не встречающейся в природе нуклеиновой кислоты. Таким образом, фрагменты геномной ДНК, полученные с помощью ПЦР или обработки эндонуклеазой рестрикции, а также кДНК рассматриваются в качестве не встречающейся в природе нуклеиновой кислоты, поскольку они существуют в виде отдельных молекул, которые не встречаются в природе. Таким же образом любая нуклеиновая кислота, содержащая промоторную последовательность и кодирующую полипептид последовательность (например, кДНК или геномная ДНК) и не встречающаяся в природе представляет собой не встречающуюся в природе нуклеиновую кислоту. Нуклеиновая кислота, которая представляет собой встречающуюся в природе, может быть экзогенной по отношению к определенной клетке. Например, цельная хромосома, выделенная из клетки дрожжей х, является экзогенной нуклеиновой кислотой в отношении клетки дрожжей у при введении данной хромосомы в клетку дрожжей.
Для получения выделенной нуклеиновой кислоты, содержащей описанную здесь нуклеотидную последовательность могут применяться способы с использованием полимеразной цепной реакции (ПЦР). ПЦР может применяться для амплификации определенных последовательностей из ДНК, а также РНК, включая последовательности из полной геномной ДНК или полной клеточной РНК. В целом, информация о последовательностях из концов представляющего интерес участка или далее применяется для конструирования олигонуклеотидных праймеров, которые идентичны или аналогичны в плане их последовательности обратным спиралям подвергаемого амплификации шаблона. Также доступны различные ПЦР стратегии, с помощью которых сайт-специфичные модификации нуклеотидных последовательностей могут быть введены в матричную нуклеиновую кислоту.
Выделенные нуклеиновые кислоты также могут быть синтезированы химически, либо в виде отдельной молекулы нуклеиновой кислоты (например, с применением автоматизированного синтеза ДНК в от 3' до 5' направлении с применением фосфорамидитного способа) или в виде серий олигонуклеотидов. Например, может быть синтезирована одна или несколько пар длинных нуклеотидов (например, длиной >100 нуклеотидов), содержащих желаемую последовательность, при этом каждая пара содержит короткий сегмент комплементарности (например, длиной около 15 нуклеотидов), так что при отжиге олигонуклеотидной пары образуется дуплекс.Для удлинения олигонуклеотидов применяется ДНК полимераза, что приводит к получению отдельной двухцепочечной молекулы нуклеиновой кислоты на одну олигонуклеотидную пару, которая затем может быть лигатирована в вектор. Выделенные нуклеиновые кислоты также могут быть получены с помощью мутагенеза, например, встречающейся в природе ДНК.
Для рекомбинантного получения полипептида (например, маннозидазы, щелочной протеазы или GAA или ее фрагмента) применяется вектор, который содержит промотор, функционально связанный с нуклеиновой кислотой, кодирующей полипептид. В соответствии с используемым здесь значением, термин "промотор" означает ДНК последовательность, обеспечивающую возможность транскрипции гена. Промотор распознается РНК полимеразой, которая затем инициирует транскрипцию. Таким образом, промотор содержит ДНК последовательность, которая либо связывается непосредственно, либо принимает участие в привлечении РНК полимеразы. Промоторная последовательность может также включать "энхансерные участки", которые представляют собой один или несколько участков ДНК, которые могут быть связаны с белками (а именно транс-действующие факторы, подобно набору транскрипционных факторов) для увеличения уровней транскрипции генов (отсюда название) в генном кластере. Энхансер, который обычно расположен на 5' конце кодирующего участка, также может быть отделен от промоторной последовательности и может быть расположен, например, в интронном участке гена или на 3' конце кодирующего участка гена.
В соответствии с используемым здесь значением, "функционально связан" означает включение в генетический конструкт (например, вектор), так что контролирующие экспрессию последовательности эффективно контролируют экспрессию представляющей интерес кодирующей последовательности.
В клетки-хозяева могут быть введены векторы экспрессии (например, путем трансформации или трансфекции) для экспрессии кодируемого полипептида, который затем может быть очищен. Системы экспрессии, которые могут применяться для получения полипептидов в малом или крупном масштабе (например, маннозидазы, щелочной протеазы или GAA или ее фрагментов) включают, без ограничения, микроорганизмы, такие как бактерии (например, E. coli), трансформированные векторами экспрессии рекомбинантной ДНК бактериофагов, плазмидной ДНК или космидной ДНК, содержащими молекулы нуклеиновой кислоты, и грибки (например, Yarrowia lipolytica, Arxula adeninivorans, Pichia pastoris, Hansenula polymorpha, Ogataea minuta. Pichia methanolica, Aspergillus niger, Trichoderma reesei, and Saccharomyces cerevisiae), трансформированные рекомбинантными грибковыми векторами экспрессии, содержащими молекулы нуклеиновой кислоты. Применимые системы экспрессии также включают клеточные системы насекомых, инфицированные векторами экспрессии рекомбинантных вирусов (например, бакуловируса), содержащими молекулы нуклеиновой кислоты, и клеточные системы растений, инфицированные векторами экспрессии рекомбинантных плазмид (например, Ti плазмиды), содержащими молекулы нуклеиновой кислоты.
Полипептиды маннозидаза или щелочная протеаза также могут быть получены с помощью систем экспрессии млекопитающих, которые включают клетки (например, иммортализированные линии клеток, таких как COS клетки, клетки яичника китайского хомячка, HeLa клетки, эмбриональные почечные клетки 293 человека и 3Т3 L1 клетки), включающие рекомбинантные конструкты экспрессии, содержащие промоторы, полученные из генома клеток млекопитающего (например, металлотионеиновый промотор) или из вирусов млекопитающих (например, аденовирусный поздний промотор и цитомегаловирусный промотор).
Рекомбинантные полипептиды, такие как маннозидаза, могут быть помечены гетерологичной аминокислотной последовательностью, такой как FLAG, полигистидин (например, гексагистидин), гемаглуттинин (НА), глутатион-8-трансфераза (GST) или связывающим мальтозу белком (МВР) для облегчения очистки белка. Другие способы очистки белков включают хроматографические способы, такие как ионный обмен, гидрофобная и обратная фаза, эксклюзия на основе размера, аффинная, гидрофобная заряд-индуцирующая хроматография и им подобные (см., например, Scopes, Protein Purification: Principles and Practice, third edition, Springer-Verlag, New York (1993); Burton and Harding, J. Chromatogr. A 814:71-81 (1998)).
In Vivo способы декэпирования и деманнозилирования гликопротеинов
Описанные здесь генетически модифицированные клетки могут применяться для получения молекулярных комплексов, обладающих GAA активностью. Например, генетически модифицированные клетки могут применяться для получения молекулярных комплексов, обладающих GAA активностью и включающих, по меньшей мере, два полипептида, при этом каждый обладает, по меньшей мере, 85% идентичностью последовательности относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO:1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO:1, в одном или нескольких сайтах между аминокислотой 50 и аминокислотой 74 (например, между аминокислотой 56 и аминокислотой 68 или между аминокислотой 60 и аминокислотой 65). Например, грибковая клетка может быть модифицирована для включения нуклеиновой кислоты, кодирующей аминокислотную последовательность, указанную в SEQ ID NO:1, и нуклеиновой кислоты, кодирующей щелочную протеазу, обладающей, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8, так что каждый из кодируемых полипептид секретируется в культуральную среду, в которой щелочная протеаза может расщеплять аминокислотную последовательность, указанную в SEQ ID NO:1. Как описано в Примере 12, когда рекомбинантная GAA секретировалась в культуральную среду с щелочной протеазой, переход прекурсора массой кДа 110 в форму массой 95 кДа завершался, то есть прекурсор массой 110 кДа не обнаруживался.
Описанные здесь генетически модифицированные клетки также могут применяться для получения декэпированных и деманнозилированных молекулярных комплексов. Данные генетически модифицированные клетки могут включать нуклеиновую кислоту, кодирующую полипептид с аминокислотной последовательностью, указанной в SEQ ID NO:1, нуклеиновую кислоту, кодирующую маннозидазу, как описано здесь, и необязательно нуклеиновую кислоту, кодирующую щелочную протеазу, обладающую, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8.
Способ получения декэпированных и деманнозилированных молекулярных комплексов на основе клеток может включать введение в грибковую клетку, являющуюся генетически модифицированной для включения нуклеиновой кислоты, кодирующей маннозидазу, которая способна гидролизовать маннозо-1-фосфо-6-маннозную связь или остаток с образованием фосфо-6-маннозы, нуклеиновой кислоты, кодирующей полипептид с аминокислотной последовательностью, указанной в SEQ ID NO:1, и необязательно нуклеиновой кислоты, кодирующей щелочную протеазу с, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8, при этом клетка продуцирует описанный здесь молекулярный комплекс, содержащий декэпированные фосфорилированные N-гликаны. Данные фосфорилированные N-гликаны могут быть деманнозилированы как описано выше. В некоторых вариантах осуществления нуклеиновые кислоты, кодирующие маннозидазу и GAA, содержат секреционную последовательность, так что маннозидаза и GAA секретируются совместно. В генетически модифицированных клетках, которые включают нуклеиновую кислоту, кодирующую щелочную протеазу, молекулярные комплексы могут быть конвертированы в форму массой 95 кДа.
Другой способ на основе клеток может включать этапы введения в грибковую клетку, являющуюся генетически модифицированной для включения нуклеиновой кислоты, кодирующей маннозидазу, которая способна (i) гидролизовать маннозо-1-фосфо-6-маннозную связь или остаток с образованием фосфо-6-маннозы и (ii) гидролизовать терминальную альфа-1,2-маннозную, альфа-1,3-маннозную и/или альфа-1,6-маннозную связь или остаток фосфатсодержащего гликана, нуклеиновой кислоты, кодирующей полипептид, обладающий аминокислотной последовательностью, указанной в SEQ ID NO:1, и, необязательно, нуклеиновой кислоты, кодирующей щелочную протеазу с, по меньшей мере, 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO:8, при этом клетка продуцирует декэпированные и деманнозилированные молекулярные комплексы. В некоторых вариантах осуществления нуклеиновые кислоты, кодирующие маннозидазу и GAA, содержат секреционную последовательность, так что маннозидаза и молекула-мишень секретируются одновременно. В генетически модифицированных клетках, которые включают нуклеиновую кислоту, кодирующую щелочную протеазу, молекулярные комплексы могут быть конвертированы в форму массой 95 кДа.
Подходящие клетки для in vivo получения молекул-мишеней или молекулярных комплексов могут быть грибкового происхождения, включая Yarrowia lipolytica, Arxula adeninivorans, метилотрофные дрожжи (такие как метилотрофные дрожжи вида Candida, Hansenula, Oogataea, Pichia или Torulopsis) или нитевидные грибы видов Aspergillus, Trichoderma, Neurospora, Fusarium или Chrysosporium. Примерные виды грибов включают, без ограничения, Pichia anómala, Pichia bovis, Pichia canadensis, Pichia carsonii, Pichia farinose, Pichia fermentons, Pichia fluxuum, Pichia membranaefaciens, Pichia membranaefaciens, Candida valida, Candida albicans, Candida ascalaphidarum, Candida amphixiae, Candida Antarctica, Candida atlantica, Candida atmosphaerica, Candida blattae, Candida carpophila. Candida cerambycidarum, Candida chauliodes, Candida corydalis, Candida dosseyi, Candida dubliniensis, Candida ergatensis, Candida fructus, Candida glabrata, Candida fermentati, Candida guilliermondii, Candida haemulonii, Candida insectamens, Candida insectorum, Candida intermedia, Candida jeffresii, Candida kefyr, Candida krusei, Candida lusitaniae, Candida lyxosophila, Candida maltosa. Candida membranifaciens, Candida milleri, Candida oleophila, Candida oregonensis, Candida parapsilosis, Candida quercitrusa, Candida shehatea, Candida temnochilae, Candida tenuis, Candida tropicalis, Candida tsuchiyae, Candida sinolaborantium, Candida sojae, Candida viswanathii, Candida utilis, Oogataea minuta, Pichia membranaefaciens, Pichia silvestris, Pichia membranaefaciens, Pichia chodati, Pichia membranaefaciens, Pichia menbranaefaciens, Pichia minuscule, Pichia pastoris, Pichia pseudopolymorpha, Pichia quercuum, Pichia robertsii, Pichia saitoi, Pichia silvestrisi, Pichia strasburgensis, Pichia terricola, Pichia vanriji, Pseudozyma Antarctica, Rhodosporidium toruloides, Rhodotorula glutinis, Saccharomyces bayanus, Saccharomyces bayanus, Saccharomyces momdshuricus, Saccharomyces uvarum, Saccharomyces bayanus, Saccharomyces cerevisiae, Saccharomyces bisporus, Saccharomyces chevalieri, Saccharomyces delbrueckii, Saccharomyces exiguous, Saccharomyces fermentati, Saccharomyces fragilis, Saccharomyces marxianus, Saccharomyces mellis, Saccharomyces rosei, Saccharomyces rouxii, Saccharomyces uvarum, Saccharomyces willianus, Saccharomycodes ludwigii, Saccharomycopsis capsularis, Saccharomycopsis fibuligera, Saccharomycopsis fibuligera, Endomyces hordei, Endomycopsis fobuligera. Saturnispora saitoi, Schizosaccharomyces octosporus, Schizosaccharomyces pombe, Schwanniomyces occidentalis, Torulaspora delbrueckii, Torulaspora delbrueckii, Saccharomyces dairensis, Torulaspora delbrueckii, Torulaspora fermentati, Saccharomyces fermentati, Torulaspora delbrueckii, Torulaspora rosei, Saccharomyces rosei, Torulaspora delbrueckii, Saccharomyces rosei, Torulaspora delbrueckii, Saccharomyces delbrueckii, Torulaspora delbrueckii, Saccharomyces delbrueckii, Zygosaccharomyces mongolicus, Dorulaspora globosa, Debaryomyces globosus, Torulopsis globosa, Trichosporon cutaneum, Trigonopsis variabilis, Williopsis californica, Williopsis saturnus, Zygosaccharomyces bisporus, Zygosaccharomyces bisporus, Debaryomyces disporua. Saccharomyces bisporas, Zygosaccharomyces bisporus, Saccharomyces bisporus, Zygosaccharomyces mellis, Zygosaccharomyces priorianus, Zygosaccharomyces rouxiim, Zygosaccharomyces rouxii, Zygosaccharomyces barkeri, Saccharomyces rouxii, Zygosaccharomyces rouxii, Zygosaccharomyces major, Saccharomyces rousii, Pichia anomala, Pichia bovis, Pichia Canadensis, Pichia carsonii, Pichia farinose, Pichia fermentons, Pichia fluxuum, Pichia membranaefaciens, Pichia pseudopolymorpha, Pichia quercuum, Pichia robertsii, Pseudozyma Antarctica, Rhodosporidium toruloides, Rhodosporidium toruloides, Rhodotorula glutinis, Saccharomyces bayanus, Saccharomyces bayanus, Saccharomyces bisporus, Saccharomyces cerevisiae, Saccharomyces chevalieri, Saccharomyces delbrueckii, Saccharomyces fermentati, Saccharomyces fragilis, Saccharomycodes ludwigii, Schizosaccharomyces pombe, Schwanniomyces occidentalis, Torulaspora delbrueckii, Torulaspora globosa, Trigonopsis variabilis, Williopsis californica, Williopsis saturnus, Zygosaccharomyces bisporus, Zygosaccharomyces mellis или Zygosaccharomyces rouxii. Примерные нитевидные грибы включать различные виды Aspergillus включая, без ограничения, Aspergillus caesiellus, Aspergillus candidus, Aspergillus carneus, Aspergillus clavalus, Aspergillus deflectus, Aspergillus flavus, Aspergillus fumigatus, Aspergillus glaucus, Aspergillus nidulans, Aspergillus niger, Aspergillus ochraceus, Aspergillus oryzae, Aspergillus parasiticus, Aspergillus penicilloides, Aspergillus restrictus, Aspergillus sojae, Aspergillus sydowi, Aspergillus tamari, Aspergillus terreus, Aspergillus ustus или Aspergillus versicolor. Данные клетки до генетической модификации, как описано здесь, могут быть получены из множества коммерческих источников и исследовательских учреждений, таких как, например Американская Коллекция Типовых Культур (Rockville, MD).
Генетическая модификация клеток может включать, помимо экзогенной нуклеиновой кислоты, кодирующей маннозидазу, GAA и/или щелочную протеазу, одну или несколько генетических модификаций, таких как: делеция эндогенного гена, кодирующего белок удлинения внешней цепи (OCH1); (ii) введение рекомбинантной нуклеиновой кислоты, кодирующей полипептид, способный промотировать маннозил-фосфорилирование (например, MNN4 полипептид из Yarrowia lipolytica, S. cerevisiae, Ogataea minuta, Pichia pastoris или С. albicans, или PNO1 полипептид из P. pastoris) для увеличения фосфорилирования маннозных остатков; (iii) введение или экспрессия РНК молекулы, которая препятствует функциональной экспрессии ОСН1 белка; (iv) введение рекомбинантной нуклеиновой кислоты, кодирующей белок дикого типа (например, эндогенный или экзогенный), обладающий связанной с N-гликозилированием активностью (например, экспрессией белка со связанной с N-гликозилированием активностью); или (v) изменение промоторных или энхансерных элементов одного или нескольких эндогенных генов, кодирующих белки со связанной с N-гликозилированием активностью для, таким образом, изменения экспрессии их кодируемых белков. РНК молекулы включают, например, малые интерферирующие РНК, малые образующие шпильки РНК, антисмысловые РНК или микроРНК. Генетическая модификация также включает изменение эндогенного гена, кодирующего белок, обладающий связанной с Ν-гликозилированием активностью, для получения белка с добавлениями (например, гетерологичной последовательностью), делециями или замещениями (например, мутациями, такими как точечные мутации; консервативные или неконсервативные мутации). Мутации могут быть введены специфично (например, с помощью сайт-направленного мутагенеза или гомологической рекомбинации) или могут быть введены случайным образом (например, клетки могут быть подвергнуты химическому мутагенезу, как описано, например, у Newman and Ferro-Novick (1987) J. Cell Biol. 105(4):1587.
Описанные здесь генетические модификации могут приводить к одному или нескольким эффектам: (i) повышению одной или нескольких активностей в генетически модифицированной клетке, (ii) понижению одной или нескольких активностей в генетически модифицированной клетке или (iii) изменению локализации или внутриклеточного распределения одной или нескольких активностей в генетически модифицированной клетке. Является известным, что увеличение степени определенной активности (например, промотирование маннозного фосфорилирования) может быть вызвано сверхэкспрессией одного или нескольких белков, способных промотировать маннозил-фосфорилирование, увеличение количества копий эндогенного гена (например, генной дупликации) или изменение промотора или энхансера эндогенного гена, который стимулирует увеличение экспрессии белка, кодируемого геном. Понижение одной или нескольких определенных активностей может быть связано со сверхэкспрессией мутантной формы (например, доминантной отрицательной формы), введением или экспрессией одной или нескольких интерферирующих РНК молекул, которые понижают экспрессию одного или нескольких белков, обладающих определенной активностью, или делецией одного или нескольких эндогенных генов, которые кодируют белок, обладающий определенной активностью.
Для изменения гена с помощью гомологичной рекомбинации может быть сконструирован "генозаместительный" вектор для включения селектируемого маркерного гена. Селектируемый маркерный ген может быть функционально связан на 5' и 3' конце с частями гена достаточной длины для опосредования гомологичной рекомбинации. Селектируемый маркер может представлять собой один из любого ряда генов, которые комплементируют ауксотрофы клеток-мишеней или обеспечивают резистентность к антибиотикам, включая URA3, LEU2 и HIS3 гены. Другие подходящие селектируемые маркеры включают CAT ген, который обеспечивает хлорамфеникольную резистентность клеток дрожжей, или lacZ, который приводит к возникновению колоний синего цвета вследствие экспрессии β-галактозидазы. Линеаризированные ДНК фрагменты генозаместительного вектора затем вводятся в клетки с применением способов, хорошо известных в данной области (см. ниже). Степень интеграции линейных фрагментов в геном и изменения гена может быть определена на основе селектируемого маркера и может быть проверена, например, с помощью саузерн-блот анализа. Селектируемый маркер может быть удален из генома клетки-хозяина с помощью, например, Cre-loxP систем (см. ниже).
Альтернативно, генозаместительный вектор может быть сконструирован для включения части изменяемого гена, чья часть лишена какой-либо последовательности эндогенного генного промотора и не кодирует или кодирует неактивный фрагмент кодирующей последовательности гена. "Неактивный фрагмент" представляет собой фрагмент гена, который кодирует белок, обладающий, например, менее чем 10% (например, менее чем около 9%, менее чем около 8%, менее чем около 7%, менее чем около 6%, менее чем около 5%, менее чем около 4%, менее чем около 3%, менее чем около 2%, менее чем около 1% или 0%) активности белка, полученного от полноразмерной кодирующей последовательности гена. Данная часть гена инсертируется в вектор таким образом, что ни одна известная промоторная последовательность не является функционально связанной с генной последовательностью, но так, что стоп-кодон и последовательность терминации транскрипции являются функционально связанными с частью генной последовательности. Данный вектор может быть далее линеаризован в части генной последовательности и трансформирован в клетку. С помощью разовой гомологичной рекомбинации данный линеаризованный вектор затем интегрируется в эндогенную копию гена.
Векторы экспрессии могут быть автономными или интегративными. Рекомбинантная нуклеиновая кислота (например, та, которая кодирует маннозидазу, GAA или щелочную протеазу) может быть введена в клетку в форме вектора экспрессии, такого как плазмида, фаг, транспозон, космида, или вирусная частица. Рекомбинантная нуклеиновая кислота может оставаться экстрахромосомной или быть интегрирована в хромосомную ДНК клетки дрожжей. Векторы экспрессии могут содержать селектируемые маркерные гены, кодирующие белки, необходимые для жизнеспособности клеток при выбранных условиях (например, URA3, который кодирует фермент, необходимый для урацильного биосинтеза или TRP1, который кодирует фермент, необходимый для триптофанового биосинтеза) для обеспечения определения и/или отбора клеток, трансформированных требуемыми нуклеиновыми кислотами (см., например, патент США 4704362). Векторы экспрессии могут также включать последовательность автономной репликации (ARS). Например, в патенте США 4837148 описаны последовательности автономной репликации, которые обеспечивают подходящие условия для поддержания плазмин в Pichia pastoris.
Интегративные векторы описаны, например, в патенте США 4882279. Интегративные векторы в целом включают серийно расположенную последовательность, по меньшей мере, первого инсертируемого ДНК фрагмента, селектируемый маркерный ген и второй инсертируемый ДНК фрагмент. Первый и второй инсертируемые ДНК фрагменты включают около 200 (например, около 250, около 300, около 350, около 400, около 450, около 500 или 1000 или более) нуклеотидов в длину и обладают нуклеотидными последовательностями, которые являются гомологичными по отношению к частям геномных ДНК трансформируемых видов. Нуклеотидная последовательность, содержащая представляющий интерес ген (например, ген, кодирующий GAA) для экспрессии, инсертируется в данный вектор между первым и вторым инсертируемым ДНК фрагментом в зависимости от положения (до или после) относительного маркерного гена. Интегративные векторы могут быть линеаризованы перед трансформацией дрожжей для обеспечения интеграции вызывающей интерес нуклеотидной последовательности в геном клетки-хозяина.
Вектор экспрессии может включать рекомбинантную нуклеиновую кислоту под контролем промотора дрожжей (например, Yarrowia lipolytica, Arxula adeninivorans, P. pastoris или других подходящих видов грибов), который обеспечивает возможность ее экспрессии в грибковых клетках. Подходящие дрожжевые промоторы включают, например ADC1, TPI1, ADH2, hp4d, POX, и Gal10 промоторы (см., например, Guarente et al (1982) Proc. Natl. Acad. Sci. USA 79(23):7410). Дополнительные подходящие промоторы описаны, например, у Zhu and Zhang (1999) Bioinformatics 15(7-8):608-611 и в патенте США 6265185.
Промотор может являться конститутивным или индуцируемым (кондициональным). Считается, что конститутивный промотор представляет собой промотор, чья экспрессия является постоянной при стандартных условиях культивирования. Индуцируемые промоторы представляют собой промоторы, которые отвечают на один или несколько индукционных стимулов. Например, индуцируемый промотор может являться химически регулируемым (например, промотором, транскрипционная активность которого регулируется присутствием или отсутствием химического индуцирующего агента, такого как спирт, тетрациклин, стероид, металл или иная малая молекула) или физически регулируемым (например, промоторам, транскрипционная активность регулируется присутствием или отсутствием физического индуктора, такого как свет или высокая или низкая температура). Индуцируемый промотор также может косвенно регулироваться одним или несколькими транскрипционными факторами, которые сами по себе напрямую регулируются химическими или физическими стимулами.
Считается, что другие генетически модифицированные модификации также могут являться кондициональными. Например, ген может быть кондиционально удален с помощью, например, сайт-специфичной ДНК рекомбиназы, такой как Cre-loxP система (см., например, Gossen et al. (2002) Ann. Rev. Genetics 36:153-173 и публикацию патентной заявки США 20060014264).
Рекомбинантная нуклеиновая кислота может быть введена в описанную здесь клетку с применением различных способов, таких как сферопластный способ или способ с использованием цельноклеточной трансформации дрожжей хлоридом лития. Другие способы, которые могут применяться для трансформации плазмид или линейных векторов нуклеиновой кислоты в клетки, описаны, например, в патенте США 4929555; Hinnen et al. (1978) Proc. Nat. Acad. Sci. USA 75:1929; Ito et al. (1983)J. Bacteriol. 153:163; патенте США 4879231; и у Sreekrishna et al. (1987) Gene 59:115, описание которых приведено здесь в их полноте для ссылки. Также могут применяться электропорация и процедуры с использованием цельноклеточной PEG 1000 трансформации, как описано у Cregg and Russel, Methods in Molecular Biology: Pichia Protocols, гл.3, Humana Press, Totowa, N.J., стр. 27-39 (1998).
Трансформированные грибковые клетки могут быть отобраны с применением подходящих способов, включая, без ограничения, культивирование ауксотрофных клеток после трансформации при отсутствии требуемого биохимического продукта (вследствие ауксотрофов клетки), отбор и определение нового фенотипа или культивирование в присутствии антибиотика, который является токсичным по отношению к дрожжам при отсутствии обеспечивающего резистентность гена, содержащегося в трансформантах. Трансформанты могут быть также отобраны и/или проверены с помощью интеграции экспрессионной кассеты в геном, которая может быть исследована с помощью, например, саузерн-блота или ПЦР-анализа.
До введения векторов в представляющую интерес клетку-мишень векторы могут быть выращены (например, амплифицированы) в бактериальных клетках, таких как Escherichia coli (Е. coli), как описано выше. Векторная ДНК может быть выделена из бактериальных клеток с помощью любого из способов, известных в данной области, которые приводят к очистке векторной ДНК от бактериальной среды. Очищенная векторная ДНК может быть подвергнута интенсивной экстракции фенолом, хлороформом и эфиром для обеспечения отсутствия белков Е. coli в продукте с плазмидной ДНК, поскольку данные белки могут быть токсичными по отношению к клеткам млекопитающих.
В некоторых вариантах осуществления генетически модифицированная грибковая клетка не включает ОСН1 ген или его генные продукты (например, мРНК или белок) и не обладает ОСН1 активностью. В некоторых вариантах осуществления генетически модифицированная клетка экспрессирует полипептид, способный промотировать маннозил-фосфорилирование (например, MNN4 полипептид из Yarrowia lipolytica, S. cerevisiae, Ogataea minuta, Pichia pastoris или С. albicans или PNO1 полипептид из P. pastoris). Например, грибковая клетка может экспрессировать MNN4 полипептид из Y. lipolytica (каталожный номер Genbank®: ХМ_503217, каталожный номер Genolevures: YALI0D24101g). В некоторых вариантах осуществления генетически модифицированная клетка не обладает ОСН1 активностью и экспрессирует полипептид, способный промотировать маннозил-фосфорилирование.
После декэпирования и деманнозилирования молекулярный комплекс может быть выделен. В некоторых вариантах осуществления молекулярный комплекс содержится в клетке дрожжей и высвобождается при лизисе клетки. В некоторых вариантах осуществления молекулярный комплекс секретируется в культуральную среду с помощью механизма, обеспечиваемого кодирующей последовательностью (нативной относительно экзогенной нуклеиновой кислоты или искусственно встроенной в вектор экспрессии), который направляет секретирование молекулы из клетки. Присутствие декэпированного и деманнозилированного молекулярного комплекса в клеточном лизате или культуральной среде может быть проверено с помощью различных стандартных протоколов для определения присутствия молекул, например иммуноблоттинга или радиоиммунопреципитации с антителом, специфичным к GAA, или тестирования на предмет определенной ферментативной активности (например, деградации гликогена). В некоторых вариантах осуществления после выделения декэпированная и деманнозилированная молекула-мишень или молекулярный комплекс могут быть связаны с гетерологичным остатком, например с помощью ферментативных или химических способов. "Гетерологичный остаток" означает любой компонент, который связывается (например, ковалентно или нековалентно) с измененной молекулой-мишенью, чей компонент отличается от компонента, первоначально присутствующего на измененной молекуле-мишени. Гетерологичные остатки включают, например, полимеры, носители, адъюванты, иммунотоксины или определяемые (например, флуоресцентные, люминесцентные или радиоактивные) остатки. В некоторых вариантах осуществления дополнительный N-гликан может быть добавлен к целевой молекуле-мишени.
Способы определения гликозилирования молекул включают масс-спектрометрию с применением ДНК-секвенирования (DSA), электрофореза углеводов с использованием фторофора (FACE) или времяпролетную масс-спектрометрию с матрично-активированной лазерной десорбцией/ионизацией (SELDI-TOF MS). Например, анализ может включать применение DSA-FACE, при которой, например, гликопротеины подвергаются денатурированию с последующей иммобилизацией, например, на мембране. Гликопротеины затем могут быть восстановлены подходящим восстановителем, таким как дитиотреитол (DTT) или ϑ-меркаптоэтанол. Сульфгидрильные группы белков могут быть карбоксилированы с помощью кислоты, такой как иодоуксусная кислота. Далее, N-гликаны могут быть высвобождены из белка с помощью фермента, такого как N-гликозидаза F. N-гликаны необязательно могут быть восстановлены и дериватизированы с помощью восстановительного аминирования. Например, высвобожденные N-гликаны могут быть помечены фторофором, таким как APTS (8-аминопирен-1,3,6-трисульфоновая кислота), на восстанавливающем конце с помощью восстановительного аминирования. Стехиометрия метки является такой, что лишь одна APTS молекула соединяется с каждой молекулой олигосахарида. Дериватизированные N-гликаны затем могут быть подвергнуты концентрированию. Аппаратура, подходящая для анализа N-гликанов, включает, например, ДНК секвенсор ABI PRISM® 377 (Applied Biosystems). Анализ данных может быть осуществлен с помощью, например, программного обеспечения GENESCAN® 3.1 (Applied Biosystems). Выделенные маннопротеины могут быть подвергнуты дальнейшей обработке одним или несколькими ферментами, такими как фосфатаза кишечника теленка, для подтверждения их статуса в качестве N-гликанов. Дополнительные способы анализа N-гликанов включают, например, масс-спектрометрию (например, MALDI-TOF-MS), жидкостную хроматографию высокого давления (ВЭЖХ) на нормальной фазе, обратной фазе и ион-обменную хроматографию (например, с импульсным амперометрическим детектированием когда гликаны не помечены и с поглощением УФ или флуоресценцией, если гликаны помечены подходящим образом). См. также Callewaert et al. (2001) Glycobiology 11(4):275-281 и Freire et al. (2006) Bioconjug. Chem. 17(2):559-564.
Культуры модифицированных клеток
Данное изобретение также относится к по существу чистой культуре любых описанных здесь генетически модифицированных клеток. В соответствии с используемым здесь значением, "по существу чистая культура" генетически модифицированной клетки представляет собой культуру данной клетки, в которой менее чем около 40% (то есть менее чем около 35%; 30%; 25%; 20%; 15%; 10%; 5%; 2%; 1%; 0.5%; 0.25%; 0.1%; 0.01%; 0.001%; 0.0001%»; или даже меньше) общего количества жизнеспособных клеток в культуре представляют собой жизнеспособные клетки, отличные от генетически модифицированных клеток, например, бактериальные, грибковые (включая дрожжевые), микоплазматические клетки или клетки простейших. Термин "около" в данном контексте означает, что соответствующая доля может быть на 15% больше или меньше указанной доли. Таким образом, например, около 20% может означать от 17% до 23%. Данная культура генетически модифицированных клеток включает клетки и среду для культивирования, хранения или транспортировки. Среда может быть жидкой, полутвердой (например, гелеподобной средой) или замороженной. Культура включает клетки, растущие в среде или в/на полутвердой среде или подвергаемые хранению или транспортировке в среде для хранения или транспортировки, включая замороженную среду для хранения или транспортировки. Культуры находятся в культуральном сосуде или сосуде для хранения или на субстрате (например, на культуральном диске, в колбе, или пробирке, или флаконе, или пробирке для хранения).
Описанные здесь генетически модифицированные клетки могут храниться, например, в виде замороженных клеточных суспензий, например в буфере, содержащем криопротектор, такой как глицерин или сахароза, в виде лиофилизированных клеток. Альтернативно, они могут храниться, например, в виде высушенных клеточных препаратов, полученных, например, с помощью сушки с применением псевдоожиженного слоя, или сушки распылением, или любого иного подходящего способа сушки.
Фармацевтические композиции и способы лечения
Описанные здесь GAA молекулы и молекулярные комплексы, например, молекулярные комплексы, содержащие, по меньшей мере, одну модификацию, которая увеличивает степень транспорта внутрь клетки млекопитающего, могут быть включены в фармацевтическую композицию, содержащую терапевтически эффективное количество молекулы и один или несколько адъювантов, эксципиентов, носителей и/или разбавителей. Подходящие разбавители, носители и эксципиенты обычно не оказывают неблагоприятного воздействия на гомеостаз реципиента (например, баланс электролитов). Подходящие носители включают биосовместимые, инертные или биоабсорбируемые соли, буферирующие агенты, олиго- или полисахариды, полимеры, увеличивающие вязкость агенты, консерванты и им подобные. Один примерный носитель представляет собой физиологический раствор (0,15 M NaCl, pH от 7,0 до 7.4). Другой примерный носитель представляет собой 50 мМ фосфат натрия, 100 мМ хлорид натрия. Подробности относительно способов составления и введения фармацевтических композиций можно найти, например, в Remington's Pharmaceutical Sciences (Maack Publishing Co., Easton, Pa.). В композиции также могут быть встроены дополнительные активные соединения.
Введение фармацевтической композиции, содержащей молекулярные комплексы с одной или несколькими описанными здесь модификациями, может быть системным или местным. Фармацевтические композиции могут быть составлены таким образом, что они являются подходящими для парентерального и/или непарентерального введения. Конкретные способы введения включают подкожное, внутривенное, внутримышечное, внутрибрюшинное, чрескожное, интратекальное, оральное, ректальное, буккальное, местное, назальное, глазное, внутрисуставное, внутриартериальное, субарахноидальное, бронхиальное, лимфатическое, вагинальное и внутриматочное введение.
Введение может осуществляться с помощью периодических инъекций болюса фармацевтической композиции или может быть непрерывным и продолжительном с помощью внутривенного или внутрибрюшинного введения из резервуара, являющегося внешним (например, в/в контейнером) или внутренним (например, биорастворимым имплантом, биоискусственным органом или колонией имплантированных измененных продуцирующих N-гликозилирующие молекулы клеток). См., например, патенты США 4407957, 5798113 и 5800828. Введение фармацевтической композиции может быть достигнуто с помощью подходящих средств доставки, таких как: помпа (см., например Annals of Pharmacotherapy, 27:912 (1993); Cancer, 41:1270 (1993); Cancer Research, 44:1698 (1984); микроинкапсуляция (см., например, патенты США 4352883; 4353888 и 5084350); непрерывное высвобождение полимерных имплантов (см., например, Sabel, патент США 4883666); макроинкапсуляция (см., например, патенты США 5284761, 5158881, 4976859 и 4968733 и опубликованные РСТ заявки W092/19195, WO 95/05452); инъекция, либо подкожно, внутривенно, внутриартериально, внутримышечно или в другой подходящий участок; или оральное введение в виде капсул, жидкости, таблетки, пилюли или с продолжительным высвобождением.
Примеры парентеральных систем доставки включают частицы на основе сополимера этиленвинилацетата, осмотические помпы, имплантируемые инфузионные системы, доставку с помощью помп, доставку на основе инкапсулированных клеток, липосомальную доставку, инъекцию с применением иглы, инъекцию без применения иглы, распыление, аэрозольное распыление, электропорацию и чрескожный пластырь. Составы, подходящие для парентерального введения, обычно содержат стерильный водный препарат на основе измененной N-гликозилирующей молекулы, которая предпочтительно является изотонической по отношению к крови реципиента (например, физиологический раствор). Составы могут быть представлены в форме с единичной дозой или множеством доз.
Составы, подходящие для орального введения, могут быть представлены в виде дискретных форм, таких как капсулы, таблетки или леденцы, каждая из которых содержит предварительно определенное количество измененной - гликозилирующей молекулы; или в виде суспензии в водном растворе или неводном растворе, таком как сироп, эликсир, эмульсия или микстура.
Молекулярный комплекс, содержащий, по меньшей мере, одну модификацию, которая увеличивает степень транспорта комплекса внутрь клетки млекопитающего, являющийся подходящим для местного введения, может быть введен млекопитающему (например, пациенту, представляющему собой человека) в виде, например, крема, спрея, пены, геля, мази или средства для растирания. Средство для растирания может быть подвергнуто гидратации в месте введения. Данные молекулы также могут быть подвергнуты инфузии (например, размочены и высушены) непосредственно в бинт, марлю или пластырь, которые затем могут быть введены топически. Данные молекулы также могут поддерживаться в полужидком, гелеобразном или полностью жидком состоянии в бинте, марле или пластыре для топического введения (см., например, патент США 4307717). Терапевтически эффективные количества фармацевтической композиции могут быть введены нуждающемуся в этом субъекту согласно схеме приема, определенной специалистом в данной области. Например, композиция может быть введена субъекту, например, системно в дозировке от 0,01 мкг/кг до 10,000 мкг/кг массы тела субъекта в расчете на одну дозу. В другом примере дозировка составляет от 1 мкг/кг до 100 мкг/кг массы тела субъекта в расчете на одну дозу. В другом примере дозировка составляет от 1 мкг/кг до 30 мкг/кг массы тела субъекта в расчете на одну дозу, например от 3 мкг/кг до 10 мкг/кг массы тела субъекта в расчете на одну дозу.
В целях оптимизации терапевтической эффективности описанный здесь молекулярный комплекс может быть сначала введен при различных схемах дозирования. Единичная доза и схема зависят от факторов, которые включают, например, вид млекопитающего, его иммунный статус, массу тела млекопитающего. Как правило, уровни данного молекулярного комплекса в ткани могут отслеживаться с помощью подходящих скрининговых анализов в качестве части процедуры клинического тестирования, например для определения эффективности рассматриваемого курса лечения. Частота приема описанного здесь молекулярного комплекса определяется навыками и клиническим заключением медицинских специалистов (например, докторов или медсестер). Как правило, схема введения определяется с помощью клинических испытаний, которые могут установить оптимальные параметры введения. Тем не менее, лечащий специалист может вносить изменения в данные курсы введения в соответствии с возрастом субъекта, здоровьем, весом, полом и медицинским статусом. Частота приема может изменяться в зависимости от того, является ли лечение профилактическим или терапевтическим.
Токсичность и терапевтическая эффективность данных молекулярных комплексов или фармацевтических композиций, таким образом, может быть определена на основе известных фармацевтических процедур, например на клеточных культурах или экспериментальных животных. Данные процедуры могут применяться, например, для определения LD50 (дозы, являющейся летальной для 50% популяции) и ED50 (дозы, являющейся терапевтически эффективной для 50% популяции). Соотношение значения дозировки между токсичным и терапевтическим эффектом представляет собой терапевтический индекс и может быть выражено в виде отношения LD50/ED50. Фармацевтические композиции, которые демонстрируют высокие терапевтические индексы, являются предпочтительными. В то время как могут применяться фармацевтические композиции, которые демонстрируют токсичные побочные эффекты, разработка системы доставки, которая таргетирует данные соединения в место поврежденной ткани, должна осуществляться с осторожностью в целях минимизации потенциального повреждения нормальных клеток (например, клеток, не представляющих собой мишени) и, таким образом, снижения побочных эффектов.
Для определения диапазона дозировок для применения у подходящих субъектов (например, пациентов, представляющих собой людей) могут применяться данные, полученные с помощью анализов клеточной культуры и исследований на животных. Дозировка данных фармацевтических композиций в целом находится в диапазоне значений концентрации в крови, которые включают ED50 с небольшой или отсутствующей токсичностью. Дозировка может различаться в рамках диапазона в зависимости от применяемой лекарственной формы и способа введения. Для фармацевтической композиции, применяемой как описано здесь (например, для лечения метаболического расстройства у субъекта), терапевтически эффективная доза может быть определена на основе анализов с применением клеточной культуры. Дозировка может быть определена на моделях с животными для определения диапазона концентрации в плазме, который включает IC50 (то есть концентрации фармацевтической композиции, которая позволяет достичь ингибирование половины симптомов) как в случае применения клеточной культуры. Данная информация может применяться для более точного определения дозировок для людей. Уровни в плазме могут быть определены, например, с помощью высокоэффективной жидкостной хроматографии. В соответствии с используемым здесь значением, "терапевтически эффективное количество" молекулярного комплекса представляет собой количество комплекса, которое способно продуцировать желаемый с медицинской точки зрения результат (например, уменьшение одного или нескольких симптомов болезни Помпе) у подвергаемого лечению субъекта. Терапевтически эффективное количество (то есть эффективная доза) может включать количества комплекса в миллиграммах или микрограммах на килограмм веса субъекта или образца (например, от около 1 микрограмма на килограмм до около 500 миллиграммов на килограмм, от около 100 микрограмм на килограмм до около 5 миллиграмм на килограмм или от около 1 микрограмм на килограмм до около 50 микрограмм на килограмм).
Субъект может представлять собой любой млекопитающее, например человека (например, пациента, представляющего собой человека) или не являющегося человеком примата (например, шимпанзе, бабуина или мартышку), мышь, крысу, кролика, гвинейскую свинью, песчанку, хомяка, лошадь, сельскохозяйственное животное (например, корову, свинью, овцу или козу), собаку, кошку или кита.
Описанный таким образом здесь молекулярный комплекс или фармацевтическая композиция могут быть введены субъекту в рамках комбинированной терапии с другими видами лечения, применяемыми для лечения болезни Помпе. Например, комбинированная терапия может включать введение субъекту (например, пациенту, представляющему собой человека) одного или нескольких дополнительных агентов, которые обеспечивают терапевтическую пользу субъекту, который страдает или обладает риском развития болезни Помпе (например, вследствие мутации гена, кодирующего GAA). Таким образом, соединение или фармацевтическая композиция и один или несколько дополнительных агентов могут вводиться одновременно. Альтернативно, сначала может вводиться молекулярный комплекс и затем один или несколько дополнительных агентов или наоборот.
Любой из описанных здесь молекулярных комплексов может быть лиофилизированным. Любая из описанных здесь фармацевтических композиций может содержаться в контейнере, пакете или диспенсере наряду с инструкциями по введению. В некоторых вариантах осуществления композиция упакована во флакон для одноразового применения.
Далее следуют примеры реализации изобретения. Они не должны рассматриваться в качестве ограничивающих в какой-либо мере область, охватываемую изобретением.
ПРИМЕРЫ
ПРИМЕР 1
Декэпирование и деманнозилирование рекомбинантной huGAA маннозидазой CcMan5 и α-маннозидазой канавалии мечевидной
Рекомбинантную GAA человека (rhGAA) получали как описано в WO 2011/039634 с применением продуцирующего штамма Y. lipolytica OXYY1589, который содержит три копии гена альфа-глюкозидазы человека (также известной как кислая альфа-глюкозидаза или кислая мальтаза ЕС3.2.1.3) и две копии гена MNN4 Y. lipolytica. Аминокислотная последовательность GAA человека указана на Фиг. 1. Генотип штамма OXY1589: MatA, leu2-958, ura3-302, xpr2-322,
gut2-744, ade2-844
POX2-Lip2pre-huGAA:URA3Ex::zeta
POX2-Lip2pre-huGAA:LEU2Ex::zeta
POX2-Lip2pre-hGM-CSF:GUTEx::zeta
Y1MNN4-POX2-hp4d-YLMNN4:ADE2::PT таргетированный
RhGAA декэпировали и деманнозилировали маннозидазой Cellulosimicrobium cellulans (CcMan5) и альфа-маннозидазой канавалии мечевидной (JbMan) (Sigma Product М7257, 3,0 M суспензия сульфата аммония). CcMan5 получали рекомбинантным способом путем клонирования нуклеиновой кислоты, кодирующей CcMan5 полипептид (Фиг. 2А) в вектор pLSAH36, который содержит DsbA сигнальную последовательность и приводит к экспрессии белка с N-терминальной гистидиновой меткой. Фиг. 2В и 2С содержат аминокислотную последовательность CcMan5 полипептида с/без сигнальной последовательности, соответственно. Плазмиду pLSAH36 клонировали в В21 клетки Е. coli, при этом белки в периплазме выделяли и очищали с помощью колонки Talon. Перед применением суспензии сульфата аммония JbMan ее также очищали с помощью гель-фильтрации на колонке Superdex 200 для удаления нежелательной фосфатазной активности.
RhGAA (концентрация около 5 мг/мл в буфере на основе 20 мМ ацетата натрия (NaOAc), pH 5,0) декэпировали и деманнозилировали путем инкубирования с CcMan5 (около 0,15-0,30 мг/мл в фосфатном буфере (PBS)) и JbMan (около 0,5-1 мг/мл в PBS) в об/об соотношении huGAA:CcMan5:JbMan 100:5:10. Общий реакционный объем разбавляли 500 мМ NaOAc буфером, pH 5.0 и 100 мМ CaCl2 для получения конечных концентраций 100 мМ NaOAc и 2 мМ CaCl2. Реакционную смесь инкубировали при 30°С в течение 16 часов.
Для оценки процесса декэпирования и анализа профиля N-гликанов очищенной huGAA N-гликаны из 5 мкг гликопротеина выделяли N-гликозидазой F (PNGaseF), метили APTS (8-амино-1,3,6-пирентрисульфоновая кислота; тринатриевая соль) и далее анализировали с помощью DSA-FACE (углеводный электрофорез с применением секвенирования ДНК и флюорофора). Способ по существу соответствует протоколу, описанному у Laroy et al, Nature Protocols, 1:397-405 (2006).
DSA-FACE электрофореграммы N-гликанов из huGAA (76 кДа форма) до (изображение В) и после (изображение С) обработки CcMan5 и JbMan приведены на Фиг. 3А. Изображение А представляет собой контрольный образец, содержащий N-гликаны, высвобожденные из RNaseB с помощью PNGaseF. Смесь N-гликанов, высвобожденная из декэпированной huGAA в основном состоит из ManP-Man8GlcNAc2 и (ManP)2-Man8GlcNAc2 (Фиг. 3А, изображение В). Немного более быстрый пик по сравнению с ManP-Man8GlcNAc2 может быть отнесен к ManP-Man7GlcNAc2. Основные пики, наблюдаемые после декэпирования и деманнозилирования, могут быть отнесены к подвергнутому двойному фосфорилированию P2-Man6GlcNAc2 и монофосфорилированным P-Mar4GlcNAc2, P-Man5GlcNAc2 и P-Man6GlcNAc2 (изображение С).
Декэпирование различных обработанных форм huGAA приводит к получению тех же профилей N-гликанов (Фиг. 3В) для 76 кДа формы (изображение В), 95 кДа формы (изображение С) и 110 кДа формы (изображение D).
ПРИМЕР 2
Очистка 110 кДа rhGAA
110 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора жидкую среду центрифугировали и фильтровали с помощью мембраны Durapore (Merck Millipore). Добавляли сульфат аммония (AMS) до концентрации 1 М, при этом раствор фильтровали перед загрузкой в колонку для гидрофобной хроматографии (HIC), уравновешенной с помощью 20 мМ фосфата натрия pH 6, 1 M сульфата аммония.
Продукт элюировали 20 мМ фосфатом натрия pH 6.
Перед загрузкой во вторую колонку для хроматографии продукт сначала концентрировали с помощью тангенциальной поточной фильтрации (TFF) на восстановленной целлюлозной мембране и затем подвергали ионному обмену из буфера на 20 мМ ацетата натрия pH 4,5. Данный материал загружали в колонку для катионобменной хроматографии (СЕХ), уравновешивали 20 мМ ацетатом натрия 4,5. После загрузки в колонку его промывали уравновешивающим буфером до достижения сигналом УФ абсорбции базового уровня и затем промывали 20 мМ ацетатом натрия pH 4,5, 50 мМ NaCl. Продукт элюировали в 20 мМ ацетат натрия рН 4,5, 150 мМ NaCl и концентрировали и подвергали ионному обмену из буфера на 20 мМ ацетат натрия pH 5 (см. ниже).
Образец декэпировали и деманнозилировали как описано в Примере 1, затем добавляли D-маннитол до концентрации 100 мМ. Три четверти данного материала были уменьшены в объеме с помощью TFF с использованием восстановленной целлюлозной мембраны с порогом отсечения молекулярной массы 10 кДа (MWCO) и далее очищены с помощью эксклюзионной хроматографии (SEC) на колонке Superdex 200, уравновешенной при 4°С 25 мМ фосфатом натрия pH 6, 150 мМ NaCl, 100 мМ D-маннитол. Далее фракции подвергали скринингу на предмет чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях. Объединенные фракции концентрировали с помощью TFF и подвергали ультрацентрифугированию с помощью центрифужных фильтров Amicon 10 кДа MWCO (восстановленная целлюлозная мембрана, Merck Millipore).
ПРИМЕР 3
Очистка 110 кДа rhGAA
110 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора материал центрифугировали и фильтровали до повышения концентрации AMS до 1 М. Раствор снова фильтровали, при этом продукт улавливали на HIC колонке, уравновешенной 20 мМ фосфатом натрия pH 6, 1 M AMS и выделяли ступенчатым градиентом от 1 до 0 M AMS в буфере на основе 20 мМ фосфата натрия pH 6.
Элюат концентрировали и подвергали ионному обмену из буфера на 10 мМ B1S-TRIS, pH 6 с помощью TFF на Vivaflow 200 модуле (PES мембрана, 10 кДа MWCO, Sartorius). Обессоленный материал загружали в колонку для анионобменной хроматографии (АЕС). После промывки колонки до тех пор, пока УФ сигнал почти достиг базового уровня, применяли двухфазный непрерывный солевой градиент; первый градиент от 0 до 0,3 M NaCl, второй - от 0,3 до 1 M NaCl. Фракции собирали при градиенте и подвергали скринингу на предмет GAA с помощью качественного 4-метилумбеллиферил-а-D-глюкопиранозида (4-MUG). В рамках 4-MUG анализа реакции начинали путем добавления реакционного буфера, состоящего из 0,35 мМ 4-MUG, 0,1% BSA и 100 мМ ацетата натрия pH 4, в 10:1 объемном отношении к 10 мкл элюционной фракции. После инкубирования в течение от 30 минут до 1 часа при 37°С добавляли равный объем 10 мМ глицина pH 11 для остановки реакции. Выделение продукта флюорогенной реакции 4-метилумбеллиферона наблюдали при УФ освещении.
Фракции, содержащие GAA, были объединены и подвергнуты концентрированию с помощью TFF на Vivaflow 200 модуле (PES мембрана, 10 кДа MWCO, Sartorius) и ультрацентрифугированию с помощью центрифужных фильтров Amicon 10 кДа MWCO (восстановленная целлюлозная мембрана, Merck Millipore).
Концентрированный материал разделяли на две части и подвергали дальнейшей очистке на колонке Superdex 200, уравновешенной при 4°С 50 мМ фосфатом натрия pH 6, 250 мМ NaCl, 0,05% Tween-20. Фракции далее подвергали скринингу на предмет чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях, при этом фосфатазное содержимое определяли с помощью колориметрического теста с использованием пара-нитрофенилфосфата, который измеряет ферментативное высвобождение окрашенного в желтый п-нитрофенолатного реакционного продукта при длине волны 405 нм.
Из фракций, содержащих GAA, делали тестовые пулы. Общее количества белка в тестовых пулах определяли с помощью метода Бредфорда. Отобранные фракции объединяли для увеличения концентрации на Vivaflow 200 TFF модуле (PES мембрана, 10 кДа MWCO, Sartorius). Объем далее снижали с помощью 15 мл центрифужных фильтров Amicon 10 кДа MWCO (восстановленная целлюлозная мембрана, Merck Millipore).
Концентрированный материал подвергали второму этапу эксклюзионной хроматографии (SEC) с применением тех же условий, что и на первом SEC этапе. Фракции подвергали повторному скринингу на предмет чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях. Фракции объединяли в соответствии с выбранным тестовым пулом и концентрировали на 15 мл центрифужных фильтрах Amicon (10 кДа MWCO, восстановленная целлюлозная мембрана, Merck Millipore).
ПРИМЕР 4
Очистка 95 кДа rhGAA
95 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора жидкую среду центрифугировали и фильтровали с помощью мембраны Durapore (Merck Millipore). Продукт затем концентрировали с помощью TFF на модифицированной полиэфирсульфоновой (PES) мембране с порогом отсечения молекулярной массы (MWCO) 10 кДа. Добавляли AMS до концентрации 1 М, при этом раствор фильтровали перед загрузкой в HIC колонку, уравновешивали 20 мМ фосфатом натрия pH 6, 1 M AMS. Продукт элюировали водой, pH элюата корректировали путем добавления стандартного буфера 100 мМ BIS-TRIS рН 6 до концентрации 10 мМ и хранили при 4°С в течение 13 дней.
Перед загрузкой в АЕХ колонку продукт концентрировали с помощью TFF на восстановленной целлюлозной мембране с MWCO 10 кДа и подвергали ионному обмену из буфера на 10 мМ BIS-TRIS pH 6. Обессоленный материал далее подвергали АЕХ хроматографии, осуществляемой как описано в Примере 3. Фракции собирали при градиенте и подвергали скринингу на предмет GAA с помощью качественного 4-MUG анализа. Фракции, содержащие GAA, объединяли в пулы для дальнейшей очистки.
На третьем этапе хроматографии концентрацию AMS увеличивали до 1 M и, после фильтрации, материал подвергали дальнейшей очистке с помощью HIC. При сборе фракций при градиенте применяли непрерывный солевой градиент от 1 до 0 M AMS. Все фракции подвергали скринингу на предмет GAA с помощью качественного анализа, при этом содержащие GAA фракции объединяли в пулы для дальнейшей обработки.
Пул концентрировали с помощью ультрацентрифугирования с применением 15 мл центрифужных фильтров Amicon 10 кДа MWCO восстановленной целлюлозной мембраны и далее очищали с помощью SEC с применением тех же процедур, которые описаны в Примере 3. После этого фракции подвергали скринингу на предмет чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях. На 90% чистые GAA фракции объединяли в пулы и сначала концентрировали на модуле TFF Vivaflow 200 (PES мембрана, 10 кДа MWCO, Sartorius) и затем подвергали ультрацентрифугированию с помощью 15 мл центрифужных фильтров Amicon (10 кДа MWCO, восстановленная целлюлозная мембрана, Merck Millipore). Концентрированный материал подвергали второму этапу SEC с применением тех же условий, что и на первом SEC этапе. Фракции подвергали скринингу на предмет GAA с помощью качественного 4-MUG GAA анализа. Фракции, обладающие GAA активностью, объединяли в пулы и концентрировали.
После декэпирования и деманнозилирования, как указано в Примере 1, добавляли D-маннитол до концентрации 100 мМ, при этом объем снова снижали перед загрузкой в последнюю гель-фильтрационную колонку Superdex 200, уравновешенную при 4°С 25 мМ фосфатом натрия pH 6, 150 мМ NaCl, 100 мМ D-маннитолом. Фракции подвергали скринингу на предмет GAA с помощью количественного 4-MUG анализа, при этом содержащие продукт фракции объединяли в пулы и концентрировали.
ПРИМЕР 5
Очистка 95-110 кДа rhGAA смеси
110 кДа прекурсор и 95 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора материал обрабатывали в рамках второго этапа хроматографии как описано в Примере 2. После HIC этапа продукт концентрировали и подвергали ионному обмену из буфера на 10 мМ BIS-TRIS pH 6 с помощью TFF и загружали в АЕХ колонку. Продукт элюировали в рамках одного этапа от 0 до 300 мМ NaCl при рН 6 (10 мМ BIS-TRIS) и затем концентрировали с помощью TFF модуля Pellicon XL50 (восстановленная целлюлозная мембрана с 10 кДа MWCO). Половину материала подвергали дальнейшей очистке с помощью эксклюзионной хроматографии. Этап хроматографии осуществляли как описано в Примере 3, однако отбор фракций для дальнейшей обработки был осуществлен лишь на основе чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях.
Половину пула концентрировали и объединяли с остатком АЕХ материала. После декэпирования и деманнозилирования концентрация D-маннитола увеличивалась до 100 мМ, при этом последующие этапы концентрирования и SEC были осуществлены согласно тем же процедурам, которые описаны в Примере 2. Фракции объединяли в пулы на основе качественного 4-MUG анализа и объединяли в пулы с декэпированным продуктом из Примера 6.
ПРИМЕР 6
Очистка 95 кДа rhGAA
95 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора материал обрабатывали включительно до АЕХ этапа как описано в Примере 3. На АЕХ этапе значительное количество продукта осталось в подвижной фазе вследствие увеличения проводимости при загрузке.
Подвижную фазу, таким образом, подвергали повторной диафильтрации с подходящим буфером и подвергали второму этапу АЕХ хроматографии. С этого момента оба объема (объем А и объем В) обрабатывали по отдельности.
Объем А соединяли с остатком SEC пула из Примера 5 и остатком СЕХ пула из Примера 2, при этом пул далее подвергали концентрированию и диафильтрации в буфер, содержащий 10 мМ BIS-TRIS pH 6, 300 мМ NaCl. Материал инкубировали при 30°С в течение 65 часов. Затем pH понижали до pH 5 путем добавления 1 M стандартного буфера ацетата натрия pH до концентрации 125 мМ, при этом образец повторно инкубировали при 30°С. Спустя 24 часа продукт обрабатывали Flavourzyme (Novozymes Corp), протеазной смесью из Aspergillus oryzae, с использованием 40:1 в/в отношения общего содержащегося в продукте белка к протеазной смеси и инкубировали последний раз при 30°С. После инкубирования в течение ночи материал очищали с помощью первого SEC этапа, осуществляемого при тех же условиях, которые описаны в Примере 3. Фракции объединяли в пулы, которые оценивали на предмет содержания чистого продукта на основе окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях. После концентрирования и обмена с помощью буфера на 20 мМ ацетата натрия рН 5 материал был декэпирован и деманнозилирован как указано в Примере 1. D-маннитол добавляли до концентрации 100 мМ, при этом материал объединяли в пул с декэпированным и деманнозилированным материалом из Примера 5. Последний SEC этап и последующие манипуляции с образцом осуществляли как описано в Примере 2.
Объем В соединяли с конечным материалом из Примера 3, при этом пул подвергали концентрированию и диафильтрации в буфер, содержащий 10 мМ BIS-TRIS pH 6, 300 мМ NaCl. Затем продукт обрабатывали протеазной смесью A. oryzae в рамках инкубационного периода в течение ночи при 30°С с применением тех же массовых соотношений, которые описаны в Примере 5, и после этого очищали с помощью первого SEC этапа, осуществляемого при тех же условиях, которые описаны в Примере 3. Дальнейшая обработка продукта осуществлялась как описано в Примере 5. На последнем этапе продукт из объема А и объема В был смешан в 14:1 соотношении GAA содержимого.
ПРИМЕР 7
Очистка 76 кДа rhGAA
76 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора культуру подвергали двум этапам продолжительного центрифугирования. Супернатант объединяли в пул, при этом добавляли AMS до концентрации приблизительно 1 М. После растворения 1 объем HIC смолы, предварительно уравновешенный в 20 мМ фосфате натрия pH 6, 1 M AMS, добавляли к 50 объемам супернатанта при размешивании для связывания продукта в режиме серийного поглощения. Полученную кашицу хранили в течение ночи при 4°С без размешивания.. В течение данного период на поверхности раствора появился слой коричневого цвета, который был удален утром путем осторожной аспирации. Смолу промывали три раза тремя объемами основного буфера (20 мМ фосфат натрия pH 6, 1 M AMS) на каждом этапе перед загрузкой в колонку. Загруженную смолу промывали, пока УФ сигнал почти не достиг основного уровня, при этом продукт был далее элюирован водой. pH элюата корректировали путем добавления стандартного буфера 100 мМ BIS-TRIS pH 6 до концентрации 10 мМ. Материал затем подвергали стерильной фильтрации в контейнере и хранили в течение одиннадцати дней при 4°С.
Второй и третий этапы хроматографии и сопровождающие манипуляции с материалом осуществляли как описано в Примере 4. После третьего этапа хроматографии концентрация пула была повышена приблизительно в семь раз на двух TFF модулях Vivaflow 200, установленных параллельно (PES мембрана, 10 кДа MWCO, Sartorius), затем пул подвергали ультрацентрифугированию с применением 15 мл центрифужных фильтров Amicon 10 кДа MWCO (восстановленная целлюлозная мембрана, Merck Millipore). Концентрированный материал был разделен на две части и далее очищен с помощью SEC с использованием тех же условий, которые описаны в Примере 4. После этого фракции подвергали скринингу на предмет чистоты на окрашенных цибакроном синим полиакриламидных гелях при денатурационных условиях. Отобранные фракции объединяли для увеличения концентрации на Vivaflow 200 TFF модулях, установленных параллельно (PES мембрана, 10 кДа MWCO, Sartorius). Объем далее снижали с помощью 15 мл центрифужных фильтров Amicon 10 кДа MWCO (восстановленная целлюлозная мембрана, Merck Millipore).
После декэпирования и деманнозилирования добавляли D-маннитол до концентрации 100 мМ, при этом образец снова концентрировали на Vivaflow 50 TFF модуле (PES мембрана, 10 кДа MWCO, Sartorius) перед загрузкой в последнюю SEC колонку так же, как описано в Примере 4. Продукт, содержащий фракции, объединяли в пул и концентрировали.
ПРИМЕР 8
Ферментативная характеристика различных вариантов huGAA (76, 95 и 110 кДа варианты) с применением искусственного хромогенного субстрата п-нитрофенил-α-D-глюкопиранозид
Применяли искусственный хромогенный субстрат п-нитрофенил-α-D-глюкопиранозид (PNPG) для определения кинетических параметров необработанной huGAA (110 кДа), полученной в Примере 2, и обработанных huGAA вариантов, полученных в Примере 7 (76 кДа), Примере 6 (95 кДа) и Примере 4 (95 кДа). Также проводили сравнение с коммерческой альфа-глюкозидазой человека, Myozyme® (алглюкозидаза альфа, Genzyme).
Ферменты разбавляли до 20 мкг/мл в буфере на основе 100 мМ ацетата натрия pH 4,0, содержащем 0,1% BSA, и 100 мМ AMS (реакционный буфер). Десять мкл ферментативных растворов добавляли в 96-луночный планшет в трех экземплярах. PNPG субстрат (Sigma) разбавляли с получением различных концентраций субстрата (10, 6, 4, 2, 1.6, 1.2, 0,8, и 0,4 мМ) в реакционном буфере, при этом 90 мкл разбавленного субстрата добавляли в каждую лунку. Ферментативную реакцию инкубировали в течение 60 минут при 37°C с последующим добавлением 100 мкл 10% карбоната натрия, pH 12 для остановки реакции. Абсорбцию измеряли при 405 нм. Измеряли стандартную кривую с п-нитрофенолом (PNP) для подсчета количества образовывающегося в минуту продукта. Скорость, выраженная как мкМ/мин, была графически выражена в виде функции различных концентраций субстрата, образующих кривую Михаэлиса-Ментен. Применяли GraphPad Prism для подсчета Vmax и Km в соответствии с уравнением Михаэлиса-Ментен. Подсчитывали каталитическую константу kcat и каталитическую эффективность kcat/Km. Специфичная активность различных ферментов была указана в виде U/mg, где 1 единица определена как количество фермента, которое катализирует гидролиз 1 нмоль субстрата в минуту при концентрации субстрата 2 мМ в буфере на основе 100 мМ ацетата натрия, pH 4,0+0,1% BSA и 100 мМ AMS. Результаты показаны в Таблице 1.

Необработанные и обработанные формы huGAA и Myozyme обладают сравнимыми кинетическими параметрами по отношению к субстрату PNPG. Это соответствует данным из литературы в отношении кислой α-глюкозидазы человека из среды на основе молока мышей и яичника китайского хомячка (СНО) (Bijvoet et al (1998), Human Molecular Genetics, 7, 1815-1824). Сообщается, что необработанная (110 кДа) и обработанная (76 кДа) форма обладают теми же значениями Km и kcat в случае искусственного субстрата 4-метилумбеллиферил-α-D-глюкопиранозид.
ПРИМЕР 9
Ферментативная характеристика различных вариантов huGAA (76, 95 и 110 кДа варианты) с применением гликогена из печени кролика в качестве субстрата
Ферментативные параметры необработанной huGAA (110 кДа вариант; пример 2) и обработанных huGAA вариантов (76 кДа, Пример 7; и 95 кДа, Пример 6) анализировали с применением гликогена из печени кролика (лот № 099K37931V, Sigma). Сравнение проводили с коммерческой α-глюкозидазой человека, Myozyme® (алглюкозидаза альфа, Genzyme). Ферменты разбавляли до 500 нг/мл в буфере на основе 100 мМ ацетата натрия pH 4,0. 50 мкл ферментативных растворов добавляли в 96-луночный планшет в двух экземплярах. Гликогеновый субстрат разбавляли с получением различных концентраций субстрата (250, 200, 150, 100, 75, 50, 25 мг/мл) в ацетатном буфере, при этом 100 мкл разбавленного субстрата добавляли в каждую лунку. Ферментативную реакцию инкубировали в течение 60 минут при 37°С. Количество глюкозы измеряли с помощью оксидазного способа с применением субстрата на основе амплекс красного.
Измеряли стандартную кривую глюкозы для подсчета количества образовывающегося в минуту продукта. Скорость фермента, выраженная как мкМ/мин, была графически выражена в виде функции различных концентраций субстрата, образующих кривую Михаэлиса-Ментен. Применяли GraphPad Prism для подсчета Vmax и Km в соответствии с уравнением Михаэлиса-Ментен. Подсчитывали каталитическую константу kcat и каталитическую эффективность kcat/Km. Специфичная активность различных ферментов была указана в виде U/mg, где 1 единица определена как количество фермента, которое катализирует образование 1 мкмоль глюкозы в минуту при 50 мг/мл конечной концентрации субстрата в буфере на основе 100 мМ ацетата натрия, pH 4,0. Результаты показаны в Таблице 2.
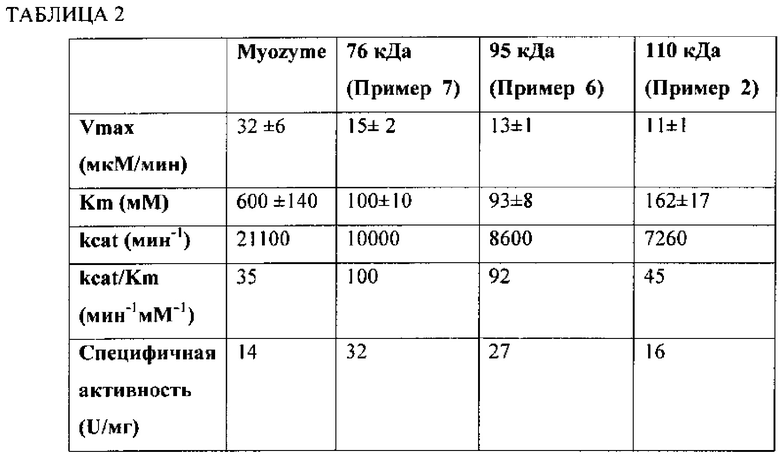
В данном эксперименте насыщение субстрата не может быть достигнуто вследствие ограниченной растворимости гликогена кролика (Фиг. 4). Для Myozyme были подсчитаны лишь очевидные значения Km и kcat. Для трех huGAA вариантов были определены нижние очевидные значения Km. Каталитическая эффективность обработанных форм в два раза превышает каталитическую эффективность необработанной huGAA и Myozyme.
ПРИМЕР 10
Воздействие кислой альфа-глюкозидазы на клиренс гликогена на модели болезни Помпе с мышами
GAA продукты из Примера 7 (76 кДа, декэпированные и деманнозилированные), Примера 6 (95 кДа, декэпированные и деманнозилированные) и Примера 2 (110 кДа декэпированные и деманнозилированные) вводили в представляющую собой модель болезни Помпе мышь для определения клиренса гликогена из скелетной мышцы и сердца.
Эксперимент проводили с применением FVB GAA нокаутных мышей и FVB мышей дикого типа. Данная модель с животными была выбрана в качестве тестовой системы в связи с тем, что она хорошо соответствует ранней инфантильной форме заболевания. Начиная с рождения нокаутные мыши обладали генерализированным и прогрессирующим накоплением лизосомного гликогена (Bijvoet et al., 1998, supra). Самцы и самки FVB GAA нокаутных мышей были получены из университета Эразма Роттердамского. В начале исследования возраст мышей составлял 26-49 недель.
Анализируемые вещества или носитель вводили внутривенно с помощью медленного болюса в хвостовую вену, при этом объем дозы составлял 10 мл/кг массы тела (м.т.) один раз неделю в течение четырех недель. До некропсии мышей не кормили в течение 16 часов. Животных умерщвляли через четыре дня после последней инъекции. Данные об исследуемых группах показаны в Таблице 3.
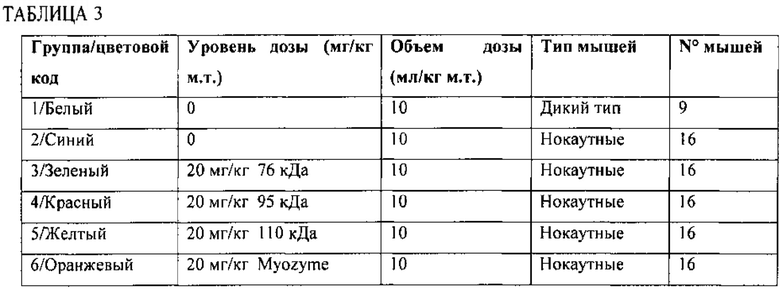
Перфузия и гомогенизация органов
Сердце и скелетные мышцы (четырехглавая мышца бедра, обе стороны) выделяли после перфузии PBS. Ткань гомогенизировали в 10 массовых объемах ледяного PBS с помощью ultra-turrax. После этого гомогенат разрушали ультразвуком при 16 микронах на льду два раза в течение 15 минут. После центрифугирования в течение 30 минут при 12000 g супернатант собирали для измерения уровня гликогена.
Биоанализ
Гликоген в сердце и скелетной мышце каждой отдельной мышцы измеряли с помощью валидированного количественного ферментативного анализа. После кипячения тканей добавляли смесь амилоглюкозидазы и α-амилазы in vitro для расщепления гликогена с образованием глюкозы. Количество глюкозы измеряли с помощью оксидазного способа с применением субстрата на основе амплекс красного. Количество гликогена указано в виде мкг гликогена/мг белка.
Статистический анализ
Гликоген в сердце из групп 2, 3, 4, 5 анализировали с помощью ANOVA с последующим сравнением с группой 6 (Myozyme) и с группой 2 (плацебо) с помощью теста Даннета. Группу 1 исключали из статистического анализа и применяли для качественной проверки на предмет дефицита накопленного гликогена в модели с мышами дикого типа. В связи с присутствием резко отличающихся наблюдений в данных по четырехглавой мышце применяли тест Крускала-Уоллиса для оценки потенциального дифференциального распределения данных по гликогену среди разных продуктов. Последующий анализ данных по четырехглавой мышце осуществляли с помощью теста Уилкоксона для суммарного ранга. Каждую продуктовую группу и группу Myozyme сравнивали с группой с плацебо (группа 2), при этом каждую продуктовую группу сравнивали с Myozyme.
Результаты
В Таблице 4 показаны средние уровни гликогена (мкг/мг белка) в сердце (А) и скелетной мышце (В) 16 мышей на одну группу.
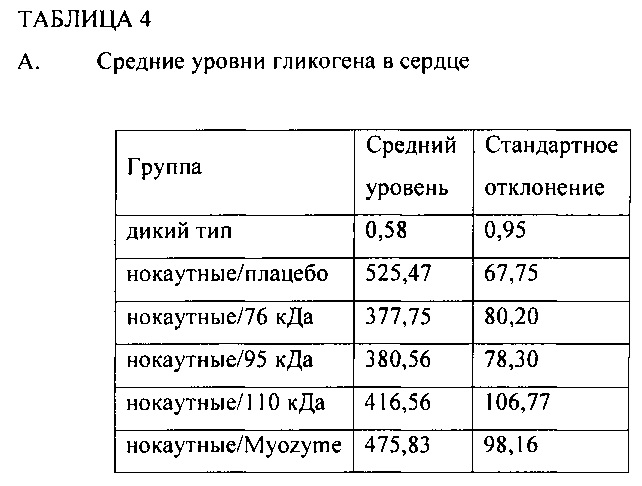
В. Средние уровни гликогена в скелетной мышце
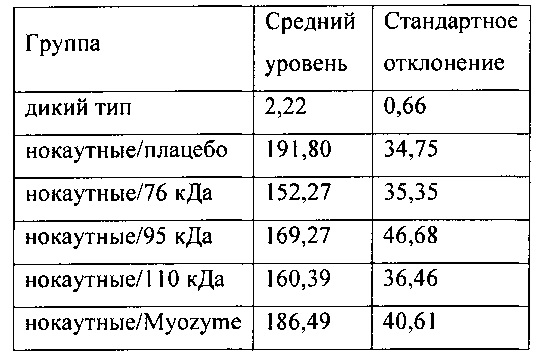
На Фиг. 5В показаны уровни гликогена (мкг/мг белка) отдельных мышей в сердце (5А) и скелетной мышце (5В). Результаты показывают, что полученные здесь GAA продукты (110 кДа, 95 кДа и 76 кДа) статистически снижают уровни гликогена в крови по сравнению с получавшими плацебо мышами после четырех внутривенных инъекций при 20 мг/кг. Та же доза Myozyme® не снижала количество гликогена в сердце. Уровни гликогена в получавших 76 кДа продукт и 95 кДа продукт группах были статистически отличными от получавшей Myozyme® группы. Разница между тремя различными полученными здесь GAA продуктами статически не существовала.
Полученный здесь 76 кДа продукт также статистически снижал количество гликогена в скелетной мышце по сравнению с получавшими плацебо или Myozyme® мышами. Уровни гликогена в случае 95 кДа и 110 кДа продукта не являлись статистическими отличными по сравнению с получавшими плацебо и Myozyme® мышами, скорее всего вследствие более высокой вариативности среди отдельных мышей. Myozyme® при 20 мг/кг не был способен снижать уровни гликогена в скелетной мышце по сравнению с плацебо.
ПРИМЕР 11
Идентификация протеазы из Aspergillus oryzae
GAA подвергается определенному протеолитическому расщеплению при инкубировании с низкими количествами Flavourzyme (Novozymes Corp), протеазной смеси из Aspergillus oryzae, при кислых значениях pH. Полученный GAA продукт обладал молекулярной массой приблизительно 95 кДа при SDS-PAGE при восстановительных условиях. Аналогичную протеолитическую активность наблюдали в определенных частично очищенных GAA продуктах, содержащих побочные белки из продуцирующего штамма (Yarrowia lipolytica).
Для анализа протеолиза N-гликаны GAA удаляли в один N-ацетильный глюкозамин на сайт N-гликозилирования с применением EndoH до протеолитической обработки. Это способствовало более точному анализу с помощью SDS-PAGE. GAA продукт затем инкубировали с протеазным коктейлем Flavourzyme или его очищенными образцами. Реакцию осуществляли при 30°С в буфере на основе 100 мМ ацетата натрия pH 5. Образцы отбирали в различные моменты времени и анализировали с помощью SDS-PAGE при восстановительных условиях. В каждую канавку загружали объемы, содержащие 0,5 мкг GAA.
Для определения семейства протеаз, ответственных за определенный протеолиз GAA в протеазном коктейле, в анализы включали протеазные ингибиторы, являющиеся специфичными по отношению к определенным семействам протеаз для осуществления идентификации протеазы. Реакцию осуществляли как описано выше за исключением того, что в реакционную смесь в данном случае были добавлены протеазные ингибиторы. Перед протеолитической реакцией необратимые ингибиторы PMSF (Sigma-Aldrich, номер продукта E5134-500G) и Е-64 (Calbiochem, номер продукта CALB324890-5) инкубировали с разбавленным протеазным коктейлем при концентрации 1 мМ и 10 мкМ, соответственно. Обратимые ингибиторы химостатин (Calbiochem, номер продукта CALB230790-5) и лейпептин (Calbiochem, номер продукта CALB108976-10MG) непосредственно добавляли в реакционную смесь до концентрации 60 мкг/мл, 50 мМ и 100 мкМ, соответственно.
Специфичный протеолиз GAA ингибировали с помощью PMSF и химостатина, протеазных ингибиторов, которые блокируют активность сериновых и цистеиновых протеаз. Необратимый ингибитор Е-64, который ингибирует цистеиновые протеазы, не блокирует протеолиз. Эти данные свидетельствуют о том, что специфический протеолиз представляет собой члена семейства сериновых протеаз. Еще одно доказательство, поддерживающее данную гипотезу, было представлено с помощью дополнительных анализов, в которых протеазный коктейль инкубировали с PMSF и окислительно-восстановительным агентом дитиотреитолом (DTT), которые восстанавливает дисульфидные связи. Добавление данного восстановителя восстанавливает ковалентную неактивную цистеиновую протеазу:PMSF аддукт, что восстанавливает активность цистеиновой протеазы. При ингибировании PMSF активность сериновых протеаз не может быть восстановлена с помощью DTT. Данную разницу в поведении применяли для дальнейшего различения сериновых и цистеиновых протеаз, воздействующих на GAA.
Инкубирование PMSF-ингибированной протеазы с DTT не восстанавливало GAA-специфичную протеолитическую активность протеазного коктейля. GAA-специфичный протеолиз также не ингибировался металло-протеазным ингибитором ЭДТА и ингибитором широкого спектра лейпептином. Все данные указывают на то, что сериновая протеаза является ответственной за данный протеолиз GAA.
В целях идентификации протеазы из смеси протеазу очищали с помощью серии этапов с применением хроматографии. На первом этапе хроматографии применяли анионобменную смолу для хроматографии (Q-Sepharose FF, GE healthcare). Материал протеазного коктейля разбавляли в буфере на основе 20 мМ TRIS-HCl pH 7 перед загрузкой. Собирали фильтрат и элюент при 100 мМ, 300 мМ и 500 мМ NaCl в буфере на основе 20 мМ TRIS-HCl. Весь фильтрантные и элюентные фракции анализировали с помощью анализа как описано выше. Протеаза, воздействующая на GAA, присутствовала в фильтрантной фракции данного запуска и была в значительной степени обогащенной по сравнению с исходным материалом.
Фильтрантный материал был далее обработан с помощью катионобменной хроматографии (SP сефароза XL (GE Heathcare) при pH 5 10 мМ ацетате натрия; элюирование 0-300 мМ NaCl). Элюентные фракции собирали и анализировали с помощью мгновенного SDS-PAGE с окрашиванием синим и анализировали на присутствие вызывающей интерес протеазы с применением анализа как описано выше.
Большая часть активности присутствовала в последних фракциях элюата СЕХ хроматографии. Последние две фракции были объединены в пул и подвергнуты анализу с помощью масс-спектрометрии следующим образом. Смесь белков обессоливали, восстанавливали и алкилировали перед расщеплением с помощью трипсина и далее подвергали ЖХ-МС/МС. Полученные спектры сравнивали с базой данных NCBI с помощью алгоритма Mascot. Применяли следующие параметры:
- Трипсин, Химотрипсин (до 4 неудачных расщеплений)
- Окисление (M, W), деамидирование (N, Q) (переменные модификации)
- Карбамидометилирование (фиксированная модификация)
- Таксономия: Эукариоты
- MS толерантность: 0,05 Да, MS/MS толерантность: 0,05 Да
На основе поиска идентифицировали щелочную протеазу из Aspergillus (каталожный номер GenBank ВАА00258.1; gi 217809). Последовательность зрелой протеазы: >gi|217809|dbj|BAA00258.1| щелочная протеаза [Aspergillus oryzae]
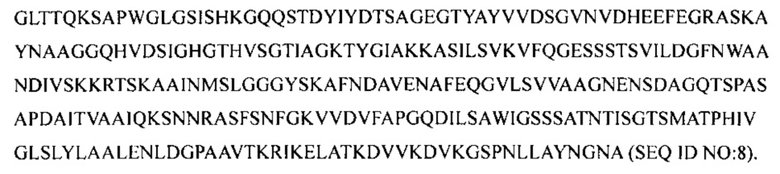
SDS-PAGE анализ очищенной протеазы из A. oryzae с применением геля показывает присутствие полосы при молекулярной массе около 30 кДа (зрелая протеаза) и несколько полос при молекулярной массе от 20 до 10 кДа. Полосы с малой молекулярной массой выделяли из геля, расщепляли трипсином и анализировали с помощью нано-ЖХ-МС/МС. Данные полосы идентифицировали в качестве продуктов и щелочной протеазы A. oryzae, что свидетельствует о том щелочная протеаза из A. oryzae подвержена аутопротеолизу.
ПРИМЕР 12
Экспрессия протеазы Aspergillus oryzae в Yarrowia lipolytica
Настоящий пример описывает конструкцию Y. lipolytica, экспрессирующей зрелый белок ALP. Ген, кодирующий щелочную протеазу (ALP) из Aspergillus oryzae (ЕС. 3.4.21.63), был оптимизирован на уровне кодонов для экспрессии Y. lipolytica и химически синтезирован в виде слитого конструкта. Слитый конструкт кодировал полную открытую рамку считывания (ORF) фермента, включая сигнальный пептид (21 аминокислота), про-пептид (100 аминокислот) и зрелый белок (282 аминокислот) и далее линкер (SGGG) и His метку (10х His остаток). См. Фиг. 9. Полная нуклеотидная последовательность слитого конструкта показана на Фиг. 10.
Синтетическую ORF ALP клонировали в серию векторов рРТ в виде BamHI/AvrII фрагментов для таргетированной интеграции в геном Y. lipolytica с применением различных локусов для стабильной интеграции экспрессионной кассеты. В pPT векторах бактериальный остаток получен из плазмидного pHSS6 и включает бактериальный репликатор (ori) и канамицин-резистивный ген, обеспечивающий резистентность к канамицину (KanR). Интеграционная кассета включает а) селектируемый маркер для трансформации в Y. lipolytica (URA3; LEU2; ADE2), b) экспрессионную кассету, составленную из промотора (РОХ2; Hp4d), с) множественный сайт клонирования (MCS) для инсерции синтетического ALP конструкта и d) терминатор гена YlLIP2. Интеграционная кассета фланкирована апстрим- (Р) и даунстрим- (Т) последовательностями специфичного локуса для стабильной таргетированной интеграции одной копии в геном Y. lipolytica с помощью гомологичной рекомбинации. Два NotI сайта рестрикции обеспечивают выделение экспрессионной кассеты до трансформации для препятствования интеграции бактериального остатка.
Применяемые для Y. lipolytica среда и способы описаны Barth and Gaillardin (FEMS Microbiol Rev., 19(4):219-37, 1997), клетки дрожжей трансформировали с помощью способа с применением ацетата лития, описанного Le Dall et al. (Curr Genet., 26(1):38-44, 1994), с применением 1 мкг очищенной интеграционной кассеты и стандартных способов, используемых для Е. coli.
Интеграцию экспрессионной кассеты ALP осуществляли на одном свободном локусе и 2 специфичных локусах на основе того, что инсерция обеспечивает устранение экспрессии секретируемых в больших количествах белков (липаза 2 и липаза 8), являющейся нежелательной при процессе ферментации. Итоговый штамм OXYY2184 содержит 3 экспрессионные кассеты ALP под контролем полуконститутивного Hp4D промотора.
OXYY2184 продуцирует рекомбинантную зрелую форму ALP Aspergillus oryzae (35 кДа) с от около 2 до около 2,5 г/л ферментативного бульона в среднем. Общий белок анализировали с помощью метода Бредфорда, при этом протеазную активность измеряли с помощью анализа с использованием азоказеина в качестве субстрата. Протеазы расщепляли азоказеин с образованием казеина и свободного азокрасителя. Преципитация и центрифугирование расщепленных белков обеспечивало возможность измерения свободного азокрасителя при щелочных условиях, что являлось индикатором протеолитической активности. Абсорбцию данного продукта измеряли при оптической плотности 440 нм. Количество расщепленного азоказеина может быть подсчитано на основе корреляции с сериями разбавления азоказеина с известными концентрациями, при которых абсорбция измеряется при оптической плотности 440 нм.
ALP в культуральном супернатанте штамма OXYY2184 анализировали с помощью SDS-PAGE и иммунодетектировали анти-His поликлональным антителом. Рекомбинантная ALP, продуцированная в Y. lipolytica, являлась активной и обладала теми же свойствами, что очищенный нативный фермент.Данные ферментные свойства рекомбинантной ALP обеспечивают возможность ее применения для обработки rhuGAA прекурсора для получения 95 кДа формы rhuGAA.
Штамм OXYY2122 модифицировали для коэкспрессии ALP и rhuGAA. Одну копию ALP экспрессионной кассеты интегрировали в штамм-реципиент, экспрессирующий rhuGAA (4 копии rhuGAA).
Оба гена, кодирующих huGAA и ALP, находятся под контролем индуцируемого РОХ2 промотора. Полученный штамм OXYY2122 продуцирует зрелую форму ALP наряду с rhuGAA прекурсором (110 кДа). Рекомбинантную huGAA в культуральном супернатанте штамма OXYY2122 анализировали с помощью SDS-PAGE с последующим иммуноблоттингом, что подтвердило, что rhGAA была преобразована в 95 кДа форму в супернатанте. Данное преобразование было завершено; 110 кДа форма не была обнаружена, в то время как при том же культивировании штамма без ALP данное преобразование не осуществлялось.
ПРИМЕР 13
Очистка 95 кДа rhGAA, полученной после обработки rhGAA ферментационного бульона щелочной протеазой Aspergillus oryzae, экспрессированной в Yarrowia lipolytica
95 кДа форму rhGAA выделяли из штамма OXYY1589 следующим образом. После сбора бульон очищали с помощью керамических мембран (Pall Corporation). Продукт концентрировали с помощью полых волоконных мембран с порогом отсечения молекулярной массы (MWCO) 10 кДа. Добавляли AMS до концентрации 1 М, при этом раствор нагревали до 30°С перед фильтрацией. Фильтрат обрабатывали щелочной протеазой A. oryzae, рекомбинантно экспрессированной в Yarrowia lipolytica (штамм OXYY2184), и применяли после очистки ферментативного бульона без какой-либо дальнейшей очистки. Об:об соотношение 200:1 всего белка:протеазы и инкубирование в течение 16 часов при 30°С приводило к полному протеолизу с получением 95 кДа продукта.
Анализ после дальнейшей очистки и после декэпирования и деманнозилирования фосфорилированных N-гликанов выявил наличие 95 кДа GAA продукта (при анализе с помощью SDS-PAGE) с аналогичной специфичной активностью по отношению к PNPG как указано в Таблице 1.
ПРИМЕР 14
Идентификация сайта протеолитического расщепления в rhGAA после обработки щелочной протеазой Aspergillus oryzae (ALP)
rhGAA обрабатывали ALP Aspergillus oryzae и далее очищали как описано в вышеприведенных примерах. Для проведения анализа последовательностей очищенный образец обрабатывали PNGaseF для дегликозилирования rhGAA, поскольку PNGase F деаминирует N-гликозилированные аспарагиновые остатки в последовательности с образованием аспартата.
Для подтверждения последовательности rhGAA дегликозилированный белок расщепляли с помощью трипсина с последующим восстановлением дисульфидных мостиков и алкилированием цистеиновых остатков. Полученную пептидную смесь подвергали ЖХ-МС и МС/МС, при этом данные сравнивали с последовательностью кодируемого геном белка для определения идентичности. Точная масса (<10 ppm) и фрагментационные спектры являлись критериями, применяемыми для абсолютной идентификации.
На основе пептидной смеси был получен почти полный охват последовательности (остатки 23-60, 65-535 и 538-898), при этом было определено, что сайт протеолитического расщепления находится между аминокислотами 60 и 65 (нумерация последовательности в соответствии с SEQ ID NO:1) Промежуток в последовательности rhGAA между остатками 60 и 65 может быть связан с протеолизом перед Gly62 и/или перед Gly65. В литературе сообщается, что щелочная протеаза из Aspergillus oryzae подвергает деградации синтетический пептид Ileu-Gln-Asn-Cys-Pro-Leu-Gly-NH2 между Leu и Gly (see Nakadai et al., 1973, Agr. Biol. Chem., 37, 2685-2694).
Сайт протеолитического расщепления определен в данном эксперименте в соответствии с протеолитическим преобразованием GAA, наблюдаемым в лизосомах. См., Moreland et al., 2005, J. Biol. Chem., 280, 6780-6791, где для 95 кДа полипептида сайт расщепления был определен между аминокислотой 59 и аминокислотой 68 (нумерация последовательности в соответствии с SEQ ID NO:1). Расщепленный N-терминальный пептид остается связанным через межцепочечную дисульфидную связь.
В то время как изобретение было описано на основе его подробного описания, вышеприведенное описание предназначено для иллюстрирования и не предназначено для ограничения охватываемой изобретением области, которая определена охватываемой областью формулы изобретения. Другие аспекты, преимущества и модификации находятся в рамках области, охватываемой приведенной далее формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |
| ВЫДЕЛЕННЫЙ ОБОЛОЧЕЧНЫЙ БЕЛОК HCV, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЛЕКАРСТВО, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) ЕГО СОДЕРЖАЩИЕ | 2002 |
|
RU2274643C2 |
| КЛЕТКА НИТЧАТЫХ ГРИБОВ С ДЕФИЦИТОМ ПРОТЕАЗ И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2013 |
|
RU2645252C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2011 |
|
RU2604811C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТЕАЗНОЙ АКТИВНОСТЬЮ, И НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ УКАЗАННЫЕ ПОЛИПЕПТИДЫ | 2002 |
|
RU2318018C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2512525C2 |
| КЛОНИРОВАНИЕ, ЭКСПРЕССИЯ И ПРИМЕНЕНИЕ КИСЛЫХ ФОСФОЛИПАЗ | 2010 |
|
RU2567659C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| СПОСОБ СНИЖЕНИЯ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ БЕЛКОВ, СПОСОБЫ И БЕЛКИ | 2010 |
|
RU2575607C2 |
| КЛЕТКИ-ХОЗЯЕВА И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2014 |
|
RU2663587C2 |


























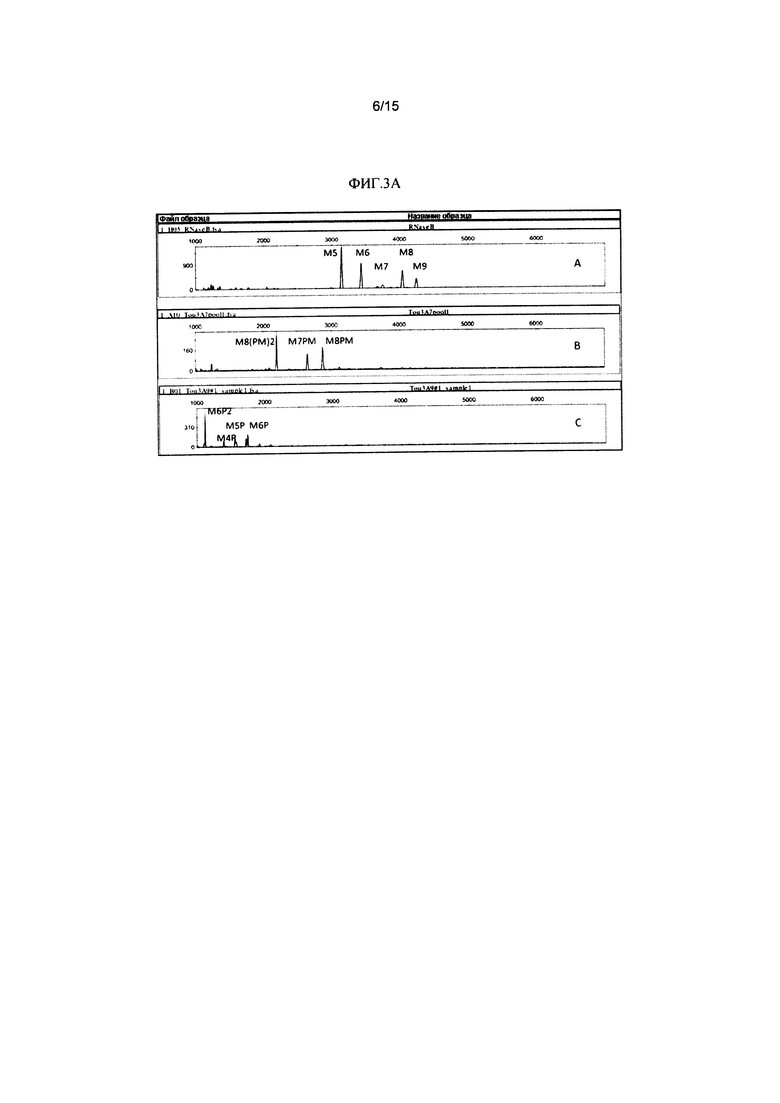

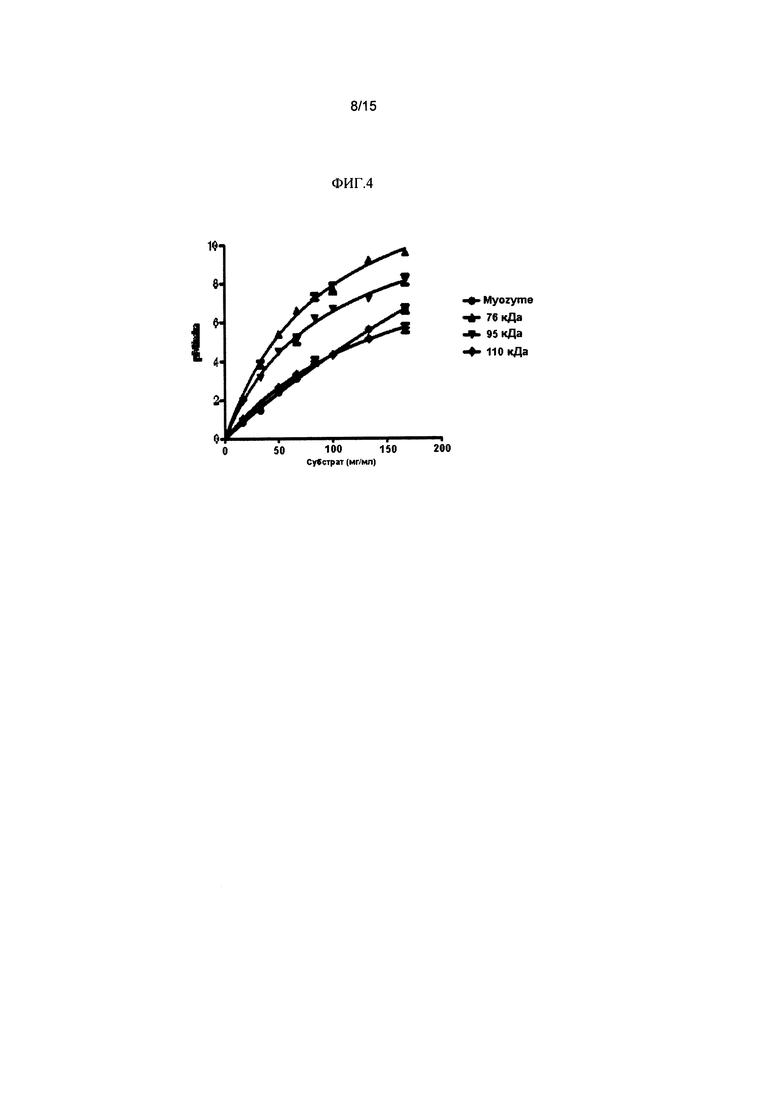

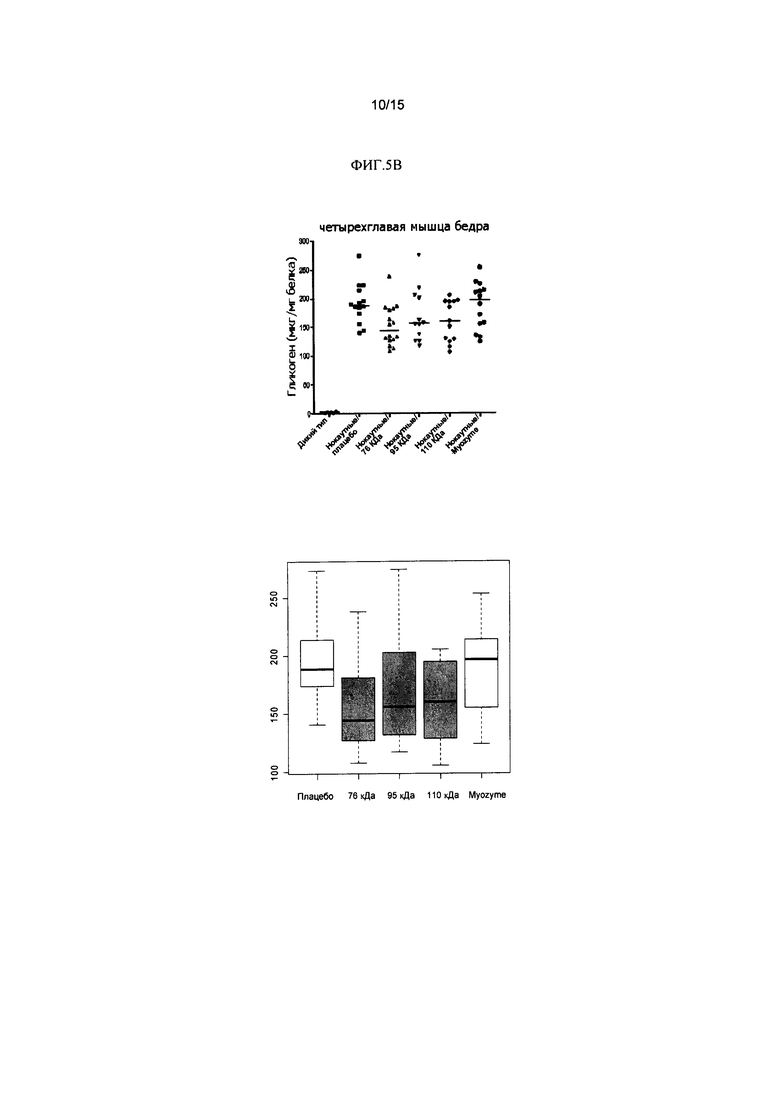





Группа изобретений относится к области биотехнологии. Представлен способ получения молекулярного комплекса, обладающего активностью GAA (кислой альфа-глюкозидазы), включающий контактирование полипептида с активностью GAA с протеазой из гриба вида Aspergillus oryzae, при этом указанная протеаза расщепляет указанный полипептид в одном или более сайтах между аминокислотой 50 и аминокислотой 74 последовательности указанного полипептида. Представлена выделенная грибковая клетка, включающая нуклеиновую кислоту, кодирующую аминокислотную последовательность указанного полипептида GAA, и нуклеиновую кислоту, кодирующую указанную щелочную протеазу, при этом указанная грибковая клетка продуцирует указанный молекулярный комплекс, обладающий GAA активностью и включающий по меньшей мере два полипептида. Группа изобретений позволяет получать более зрелые формы полипептида GAA, которые обладают увеличенной специфической активностью и улучшенной способностью к снижению уровня гликогена (в сердце, скелетной мышце) по сравнению с предшественником GAA и коммерческой GAA человека. 2 н. и 18 з.п. ф-лы, 14 ил., 5 табл., 14 пр.
1. Способ получения молекулярного комплекса, включающий контактирование полипептида, обладающего по меньшей мере 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO: 1, с протеазой, обладающей по меньшей мере 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO: 8, при этом указанная протеаза расщепляет указанный полипептид в одном или более сайтах между аминокислотой 50 и аминокислотой 74.
2. Способ по п.1, отличающийся тем, что полипептид обладает по меньшей мере 90% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO: 1, или отличающийся тем, что полипептид обладает по меньшей мере 95% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO: 1, или отличающийся тем, что полипептид обладает аминокислотной последовательностью, указанной в SEQ ID NO: 1.
3. Способ по п.1 или 2, отличающийся тем, что указанное контактирование осуществляют in vitro.
4. Способ по п.1 или 2, отличающийся тем, что контактирование осуществляется в рекомбинантной грибковой клетке.
5. Способ по п.4, отличающийся тем, что грибковая клетка представляет собой Yarrowia lipolytica, Arxula adeninivorans или метилотрофные дрожжи, такие как метилотрофные дрожжи вида, выбранного из группы, включающей Candida, Hansenula, Oogataea, Pichia или Torulopsis.
6. Способ по п.1 или 2, отличающийся тем, что указанная протеаза расщепляет указанный полипептид в одном или более сайтах между аминокислотой 60 и аминокислотой 65.
7. Способ по п.1 или 2, дополнительно включающий протеолиз полипептида в одном или более сайтах между аминокислотой 719 и аминокислотой 746 и/или в одном или более сайтах между аминокислотой 137 и аминокислотой 151.
8. Способ по п.1 или 2, отличающийся тем, что по меньшей мере один полипептид молекулярного комплекса включает один или более фосфорилированных N-гликанов.
9. Способ по п.1 или 2, дополнительно включающий изменение молекулярного комплекса с помощью по меньшей мере одной модификации, которая приводит к увеличению способности молекулярного комплекса транспортироваться внутрь клетки млекопитающего.
10. Способ по п.9, отличающийся тем, что по меньшей мере одна модификация включает лиганд для внеклеточного рецептора, или отличающийся тем, что по меньшей мере одна модификация включает распознающий домен инсулиноподобного фактора роста II человека.
11. Способ по п.9, отличающийся тем, что указанная по меньшей мере одна модификация влияет на связывание маннозо-6-фосфатного рецептора на клетке млекопитающего.
12. Способ по п.9, отличающийся тем, что по меньшей мере один полипептид молекулярного комплекса включает один или более фосфорилированных N-гликанов, при этом модификация включает декэпирование и деманнозилирование по меньшей мере одного фосфорилированного N-гликана.
13. Способ по п.12, отличающийся тем, что по меньшей мере 40% N-гликанов на указанном полипептиде являются декэпированными и деманнозилированными, или отличающийся тем, что по меньшей мере 60% N-гликанов на указанном полипептиде являются декэпированными и деманнозилированными, или отличающийся тем, что по меньшей мере 80% N-гликанов на полипептиде являются декэпированными и деманнозилированными, или отличающийся тем, что по меньшей мере 90% N-гликанов на полипептиде являются декэпированными и деманнозилированными.
14. Выделенная грибковая клетка, включающая нуклеиновую кислоту, кодирующую GAA аминокислотную последовательность, обладающую по меньшей мере 85% идентичностью последовательности относительно GAA аминокислотной последовательности, указанной в SEQ ID NO: 1, и нуклеиновую кислоту, кодирующую щелочную протеазу, обладающую по меньшей мере 85% идентичностью последовательности относительно аминокислотной последовательности, указанной в SEQ ID NO: 8, при этом указанная грибковая клетка продуцирует молекулярный комплекс, обладающий GAA активностью и включающий по меньшей мере два полипептида, при этом каждый полипептид обладает по меньшей мере 85% идентичностью последовательности относительно сегмента аминокислотной последовательности, указанной в SEQ ID NO: 1, при этом каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO: 1, в одном или более сайтах между аминокислотой 50 и аминокислотой 74 указанной щелочной протеазы.
15. Грибковая клетка по п.14, отличающаяся тем, что каждый сегмент получен протеолизом аминокислотной последовательности, указанной в SEQ ID NO: 1, в одном или более сайтах между аминокислотой 60 и аминокислотой 65 указанной щелочной протеазы.
16. Грибковая клетка по п.14 или 15, отличающаяся тем, что указанная грибковая клетка включает также нуклеиновую кислоту, кодирующую маннозидазу, при этом указанная маннозидаза способна (i) гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата и (ii) гидролизовать терминальную связь с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой.
17. Грибковая клетка по п.14 или 15, отличающаяся тем, что указанная грибковая клетка включает также нуклеиновую кислоту, кодирующую маннозидазу, при этом указанная маннозидаза способна гидролизовать маннозо-1-фосфо-6-маннозный остаток с образованием маннозо-6-фосфата.
18. Грибковая клетка по п.14 или 15, отличающаяся тем, что указанная грибковая клетка включает также нуклеиновую кислоту, кодирующую маннозидазу, при этом указанная маннозидаза способна гидролизовать терминальную связь с альфа-1,2-маннозой, альфа-1,3-маннозой и/или альфа-1,6-маннозой.
19. Грибковая клетка по п.14 или 15, отличающаяся тем, что указанная грибковая клетка также включает нуклеиновую кислоту, кодирующую полипептид, способный промотировать маннозил-фосфорилирование.
20. Грибковая клетка по п.14 или 15, отличающаяся тем, что указанная грибковая клетка является генетически модифицированной для устранения ОСН1 активности.
| MORELAND R | |||
| J | |||
| et al., Lysosomal Acid alpha-Glucosidase Consists of Four Different Peptides Processed from a Single Chain Precursor, The Journal of Biological Chemistry, Vol.280, No.8, pp.6780-6791, 2005 | |||
| WO 2011039634 A2, 07.04.2011 | |||
| HIROKI TATSUMI et al., Cloning and Sequencing of the Alkaline Protease cDNA from Aspergillus oryzae, Vol.52, Issue 7, pp.1887-1888, 1988 | |||
| Подвижной стол (герд) для воздушного обогащения угля | 1926 |
|
SU12617A1 |
| RU 2005123986 А, 27.02.2007 | |||
| ZHU Y | |||
| et al., Glycoengineered Acid α-Glucosidase With Improved Efficacy at Correcting the Metabolic Aberrations and Motor Function Deficits in a Mouse Model of Pompe Disease, Molecular Therapy, Vol.17, NO.6, pp.954-963, 2009 | |||
| BIJVOET A | |||
| G | |||
| A | |||
| et al., Recombinant human acid α-glucosidase: high level production in mouse milk, biochemical characteristics, correction of enzyme deficiency in GSDII KO mice, Human Molecular Genetics, Vol.7, NO.11, pp.1815-1824, 1998 | |||
| JOHAN L | |||
| K | |||
| VAN HOVE et al., High-level production of recombinant human lysosomal acid α-glucosidase in Chinese hamster ovary cells which targets to heart muscle and corrects glycogen accumulation in fibroblasts from patients with Pompe disease, Proceedings of the National Academy of Sciences, Vol.93, NO.1, pp.65-70, 1996. | |||

