Область изобретения
Изобретение относится к способу определения, является ли соединение ингибитором нейротрипсина, к конкретным ингибиторам нейротрипсина, к применению данных ингибиторов для лечения и/или профилактики атрофии скелетных мышц и шизофрении и применению в качестве стимуляторов познавания.
Предшествующий изобретению уровень техники
Серинпротеазы принадлежат к группе протеолитических ферментов, которые совокупно имеют хорошо изученный каталитический механизм. Серинпротеазы обнаруживают в вирусах, бактериях и эукариотах. Они включают экзопептидазы, эндопетидазы и олигопептидазы. Имеется сходство механизмов реакции ряда пептидаз различного эволюционного происхождения. Геометрическая ориентация каталитических остатков очень близка, несмотря на тот факт, что в других случаях складки белка сильно отличаются. Каталитическая триада остатков серина, гистидина и аспартата в активном центре отвечает за эффективное гидролитическое расщепление пептидной связи. Примеры серинпротеаз включают тромбин, фактор XIIa, фактор IXa, фактор Ха, плазмин, tPA (тканевый активатор плазминогена), трипсин, химотрипсин и другие белки, такие как урокиназа, триптаза, эластаза, калликреин, комплемент С, протеаза А, серинкарбоксипептидаза II. Они участвуют в ряде важных процессов, таких как, например, свертывание крови и переваривание пищи. Показано, что ингибиторы серинпротеазы подавляют клеточные процессы, такие как адгезия, миграция, образование свободных радикалов и апоптоз. Введенные внутривенно, ингибиторы серинпротеазы обеспечивают защитный эффект в отношении повреждения ткани. Показано, что маленькие молекулы ингибиторов обладают высоким потенциалом в плане лечения различных заболеваний, связанных с гематологией, онкологией, астмой, воспалением, неврологией, пульмонологией и иммунологией. Подходящие ингибиторы серинпротеазы можно использовать при лечении дисфункций в области тромботических заболеваний, астмы, цирроза, артрита, карциномы, меланомы, рестеноза, атеромы, травмы, шока и реперфузионного повреждения.
Исследованный фермент нейротрипсин (WO 98/49322) принадлежит к семейству химотрипсина, члены которого почти полностью ограничены животными. Последовательность аминокислот нейротрипсина определяет мозаичный белок из 875 аминокислот, состоящий из Kringle домена, после которого следуют четыре богатых цистеином повтора фагоцитарного рецептора (у мыши три) и домен серинпротеазы (см. фиг.1, А и В). Нейротрипсин включает подобно тромбину, tPA, трипсину и ряду других ферментов остаток аспартата на дне своего кармана S1, демонстрируя вследствие этого специфичность в отношении основных аминокислот в данном центре связывания. Структурная близость нейротрипсина к протеазам каскада свертывания крови и фибринолитической системы, таким как фактор X, фактор IX, тромбин, тканевый активатор плазминогена и плазмин, предполагает, что он может представлять собой элемент направляемого протеазой внеклеточного механизма передачи сигнала в нервной системе (см. статьи Gschwend, Т.Р., et al., Molec. Cell Neurosci. 9: 207-219, 1997; Proba, К., et al., Biochim. Biophys. Acta 1396: 143-147, 1998).
Как будет показано ниже в данном контексте, нейротрипсин находится в пресинаптическом нервном окончании синапсов центральной нервной системы (ЦНС) и в нервно-мышечном соединении (НМС). Синапс представляет собой связь между нервными клетками (нейронами), в которых происходит передача информации в форме химических субстанций, называемых нейромедиаторами. Синапсы состоят из пресинаптического окончания, образованного генерирующей сигнал клеткой, и постсинаптической специализации принимающей сигнал клетки. Нейромедиаторы, высвобождаемые из пресинаптического окончания, пересекают синаптическую щель и связываются с рецепторами нейромедиаторов в постсинаптической специализации. При связывании нейромедиатора рецептор индуцирует генерацию электрического импульса в постсинаптической клетке. Передача сигнала между двумя нейронами является основой функции нейронов. Функции головного мозга являются результатом специфической сборки огромного числа нейронов в сети, обрабатывающие информацию.
Большинство синапсов находится в центральной нервной системе (ЦНС, головной мозг), где каждый синапс соединяет два нейрона. Посредством данных двухсторонних поточечных соединений каждый нейрон может связываться с тысячами других нейронов. Однако синапсы также связывают нейрон с железой или мышечной клеткой. Нервно-мышечное соединение (НМС, мышечная концевая пластинка (окончание двигательного нерва)) представляет собой синапс, который соединяет нервную клетку с клеткой поперечно-полосатой мышцы. Синапсы, находящиеся вне головного мозга, ствола головного мозга и спинного мозга, называют синапсами периферической нервной системы (ПНС). Синапсы ЦНС и синапсы ПНС проявляют много структурных и функциональных общих свойств и имеют много общего в плане молекулярных компонентов (синаптических молекул). Вследствие этого синаптические молекулы-мишени можно использовать для получения направленности на синаптические функции как ЦНС, так и ПНС.
Атрофия скелетных мышц (саркопения), определяемая как потеря мышечной массы и силы, играет главную роль в патогенезе хрупкости и функционального нарушения, которое происходит при старении. Она играет главную роль в потере мышечной силы, пониженной скорости метаболизма, постепенном уменьшении плотности костей и пониженной аэробной производительности (см. статью Doherty, T.J., J. Appl. Physiol. 95: 1717-1727, 2003). Потеря мышечной массы проявляется как уменьшение площади поперечного среза мышцы с возрастом, которое, как определяют, является результатом комбинированного эффекта уменьшения числа мышечных волокон и толщины отдельных оставшихся волокон.
В последние годы достигнут существенный прогресс в идентификации и характеризации факторов, делающих вклад в деградацию мышечной массы. Важные гены, ассоциированные с данными процессами, кодируют лигазы белка убиквитина, уровень которых, как обнаружено, повышен в атрофической мышце. Обнаружено, что среди факторов, которые обладают гипертрофической активностью, и, таким образом, блокируют атрофию, инсулиноподобный фактор роста 1 (IGF-1) играет основную роль. Интенсивно исследован данный и ряд других регуляторных путей, контролирующих массу скелетной мускулатуры (в качестве обзора см. статью Glass, D.J., Nature Cell Biol. 5: 87-90, 2003). Несмотря на значительный прогресс в характеризации как молекулярных механизмов, которые контролируют деградацию мышц, приводящую к атрофии, так и гипертрофических эффектов инсулиноподобного фактора роста, и, несмотря на тот факт, что ряд фирм работает над созданием лекарственных препаратов, способных стимулировать повышение мышечной массы, до настоящего времени не одобрены никакие лекарственные препараты.
Морфологическим признаком атрофии скелетных мышц, обнаруживаемой в старческом возрасте (саркопении), является существенное уменьшение числа мышечных волокон. Достаточно фактов, полученных в многочисленных независимых исследованиях, подтверждает, что вход нерва во фракцию мышечных волокон разрушается с возрастом, приводя к последующей атрофии и, в конечном счете, исчезновению денервированных волокон (см. статью Kamal, H.K., Nutrition Reviews 61: 157-167, 2003). Другим характерным признаком атрофии скелетной мышцы, обнаруживаемом в старческом возрасте, является одновременное появление мышечной атрофии, существенное уменьшение числа двигательных нейронов (см. статью Welle, S., Can. J. Appl. Physiol. 27: 19-41, 2002) и заметное структурное изменение нервно-мышечной связи (см. статью Tapia, J.C. et al., Abstract Viewer/Itinerary Planner, Washington DC: Society for Neuroscience). Данные характеристики показывают, что существенное, связанное с возрастом ухудшение структуры и функции нервно-мышечной связи является основным фактором, способствующим процессу, который, в конечном счете, приводит к структурной и функциональной денервации. Денервированные мышечные волокна, которые не получают компенсаторной реиннервации в течение нескольких недель, постепенно становятся атрофическими и со временем исчезают.
Шизофрения представляет собой хроническое тяжелое и инвалидизирующее головной мозг заболевание. У приблизительно 1% населения планеты на протяжении жизни развивается шизофрения. Люди, у которых развивается шизофрения, испытывают сильные страдания. Приблизительно 10% совершают самоубийство. Хотя шизофрения поражает мужчин и женщин с равной частотой, нарушение чаще раньше проявляется у мужчин, обычно в конце подросткового возраста или вскоре после двадцатилетия, чем у женщин, которые, как правило, заболевают в возрасте от двадцати лет до вскоре после тридцати. Больные шизофренией часто испытывают ужасные симптомы, например слышат внутренние голоса, которые не слышат другие, или считают, что другие люди читают их мысли, контролируют их мысли или замышляют причинить им вред. Данные симптомы могут пугать их и делать замкнутыми. Их речь и поведение могут быть настолько дезорганизованными, что они могут становиться непонятными или пугающими для окружающих. Доступные в настоящее время лекарственные препараты от шизофрении существенно уменьшают страдания, но приблизительно 2/3 больных шизофренией требуется государственная помощь в течение всего нескольких лет после начала заболевания. Большинство из них не способны вернуться к работе или учебе и имеют относительно низкий уровень или не имеют социальных взаимосвязей, и большинство больных шизофренией продолжают страдать от ряда симптомов на протяжении всей жизни. По приблизительной оценке не больше чем один из пяти больных выздоравливает полностью. Вследствие этого шизофрения является одной из наиболее важных проблем здравоохранения во всем мире, и затраты общества исчисляются в миллиардах долларов.
В настоящее время наиболее постоянным нейропатологическим событием в головном мозге пациентов с шизофренией является уменьшение числа синапсов в сером веществе центральной нервной системы, что отражается в снижении объема нейропилей (синаптической площади). Не наблюдают никаких доказательств дегенерации нейронов. Как правило, число подсчитываемых нейронов/площадь ткани сильно увеличено, факт объясняют избирательным уменьшением числа синапсов на площади нейропиля между нейронами, тогда как число тел нервных клеток остается постоянным. Феномен описан в течение двух последних десятилетий в ряде независимых исследований на посмертном материале и показано, что он наиболее обширен в предлобной коре. Литературное документирование данного наблюдения внимательно рассмотрено в обзоре Selemon, L.D. и Goldman-Rakic, P.S. (Psychiatry 45: 17-25, 1999). McGlashan, Т.Н. и Hoffman, R.E. (Arch. Gen. Psychiatry 57: 637-648, 2000), где подводят итоги основным морфологическим, связанным с развитием, электрофизиологическим и метаболическим наблюдениям при шизофрении в свете гипотезы "избыточного прунинга синапсов" и приходят к заключению о том, что "избыточный прунинг синапсов" или "сниженный в период развития уровень синаптических соединений" представляет собой все более привлекательную патофизиологическую модель шизофрении. Основываясь на данной модели, шизофрения возникает при критически пониженной связанности синапсов в результате связанных с развитием нарушений синаптогенеза во время беременности и раннего детства и/или избыточного прунинга синапсов в период полового созревания. В модели учитывают феноменологию нарушения, симптоматические состояния, начало, нейродефициты, связанные с развитием, окно ухудшения, половые различия в клиническом представлении, течение, определяемое возрастом начала, и сохранение шизофренического генотипа в популяции, несмотря на пониженную фенотипическую плодовитость.
Стимуляторы познавания представляют собой лекарственные препараты, направленные на предупреждение, улучшение или лечение когнитивных дефицитов на обоих, клиническом и субклиническом, уровнях. Данные лекарственные препараты дают положительный эффект при лечении нарушений памяти у пожилых людей, у которых не развивается болезни Альцгеймера (мягкое когнитивное нарушение). Однако данные лекарственные препараты, кроме того, дают положительный эффект в плане улучшения когнитивных функций у пациентов с установленным диагнозом болезни Альцгеймера или другими заболеваниями, ассоциированными с деменцией, или в плане улучшения когнитивных функций при посттравматической когнитивной дисфункции, а также для улучшения связанного со старением нарушения когнитивных функций, которое считают нормальным признаком процесса старения.
Мягкое когнитивное нарушение представляет собой широко приводимое представление в клиническом исследовании, связанном со старением когнитивных нарушений (см. статью Ritchie, K. и Touchon, J., The Lancet 355: 225-228, 2000). Оно в основном относится к субклиническим нарушениям функционирования памяти у пожилых людей, у которые, как считают, имеется большая вероятность развития болезни Альцгеймера. Выявление людей с возможным риском развития деменции в свете раннего терапевтического вмешательства важно, поскольку это может уменьшить страдания как для больного, так и для семьи, свести к минимуму риск несчастных случаев, увеличить период независимости и, возможно, даже, в конечном счете, предупреждает начало процесса, приводящего к самой деменции.
Нарушение когнитивных функций без деменции настолько обычно у пожилых людей, что многие рассматривают его как неизбежный признак процесса старения. Тем не менее, оно приобретает клиническую значимость вследствие трудностей, с которыми пациенты могут сталкиваться в повседневной деятельности. Хотя круг нарушений, наблюдаемых в популяциях, не имеющих деменции, чрезвычайно широк, предложен ряд клинических меток для описания данного последнего нормального когнитивного круга. Одним из наиболее ранних является доброкачественная старческая забывчивость. Ее клинические признаки включают неспособность вспомнить мелкие детали, забывание отдаленных событий в противоположность недавним и осознание проблем с памятью. Термин ассоциированное со старением снижение когнитивной способности относится к более широкому кругу когнитивных функций (внимание, память, обучение, мышление, речь и зрительно-пространственная функция) и диагностируется относительно норм для пожилых людей. Назначение стимуляторов познавания может продлить способность пораженных болезнью осуществлять повседневную деятельность и, таким образом, увеличивать период их независимости. Другие нарушения, ассоциированные по меньшей мере у части больных с когнитивными нарушениями, которые могут в конечном счете привести к деменции, включают болезнь Паркинсона, рассеянный склероз, инсульт и травма головы. Назначение лекарственных препаратов стимуляторов познавания может также улучшить когнитивные функции у данных пациентов.
Раскрытие изобретения
Изобретение относится к способу измерения каталитической активности нейротрипсина, отличающемуся тем, что нейротрипсин, его вариант или фрагмент, включающий домен протеазы нейротрипсина, и белок либо пептид, включающий агрин, его вариант или фрагмент, включающий α- или β-сайт расщепления агрина, совместно инкубируют в водном буферном растворе, и количественно измеряют расщепление агрина. Более того, изобретение относится к способу определения, является ли соединение ингибитором нейротрипсина, отличающемуся тем, что соединение инкубируют вместе с нейротрипсином, его вариантом или фрагментом, включающим домен протеазы нейротрипсина, и с белком либо пептидом, включающим агрин, его вариант или фрагмент, включающий α- или β-сайт расщепления агрина, в водном буферном растворе, и количественно измеряют расщепление агрина.
Кроме того, изобретение относится к ингибиторам нейротрипсина, обнаруженным с помощью данного способа, в частности к соединениям формулы
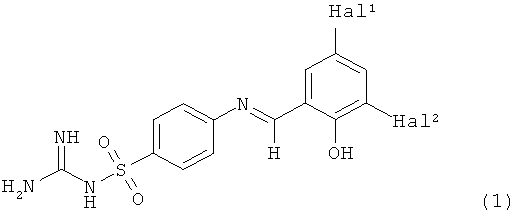
в которой Hal1 и Hal2 независимо друг от друга представляют собой фтор, хлор или бром и их фармацевтически приемлемым аддитивным солям.
Изобретение, кроме того, относится к применению данных ингибиторов как лекарственных средств, в частности, для лечения и/или профилактики заболеваний, вызываемых дефицитом синапсов, например атрофии скелетных мышц, шизофрении и когнитивного нарушения, и к применению данных ингибиторов для изготовления лекарственного средства для лечения и/или профилактики атрофии скелетных мышц, шизофрении и когнитивного нарушения.
Краткое описание чертежей
Фиг.1. Структура домена нейротрипсина.
(A) hNt: человеческий нейротрипсин.
(B) mNt: мышиный нейротрипсин.
Нейротрипсин состоит из богатого пролином основного домена (РВ), kringle-домена (KR), трех (mNt) или четырех (hNt) богатых цистеином доменов фагоцитарного рецептора (SRCR1, SRCR2, SRCR3 и SRCR4) и домена протеазы (PROT).
Фиг.2. Опосредованное нейротрипсином расщепление агрина: Анализ вестерн-блот агрина из клеток HEK293, сотрансфицированных агрином и нейротрипсином.
Полуконфлюэнтные клетки HEK293T временно трансфицируют либо pcDNA3.1-нейротрипсин либо pcDNA3.1-агрин либо обоими. Образцы разделяют SDS-PAGE (электрофорезом в полиакриламидном геле с использованием додецилсульфата натрия). Мембрану инкубируют с поликлональным антителом к агрину, направленным на С-концевую группу агрина, с последующим инкубированием с вторичным антителом, связанным с пероксидазой.
(Дорожка 1, Ag) Экстракт детергентом клеток, однократно трансфицированных агрином.
(Дорожка 2, Ag+hNt) Экстракт детергентом клеток, дважды трансфицированных агрином и нейротрипсином. Отмечают, что уровень агрина сильно понижен.
(Дорожка 3, Ag+hNt) Культуральная среда клеток, которые дважды трансфицированы агрином и нейротрипсином. Полосу 100-кД определяют с помощью антитела к агрину, направленного на С-концевую группу агрина.
(Дорожка 4, Ag) Культуральная среда клеток, которые однократно трансфицированы агрином.
Продукцию нейротрипсина во всех условиях подтверждают после повторного зондирования блота антителами к нейротрипсину. Анализ культуральной жидкости показывает, что иммунореактивность агрина, которая утрачена в клеточном экстракте дважды трансфицированных клеток, высвобождается в среду супернатанта. В среде супернатанта клеток HEK293T, трансфицированных агрином и каталитически неактивным нейротрипсином, не обнаруживают никакого сигнала.
Фиг.3. Временный характер расщепления агрина in vivo соответствует временному характеру экспрессии нейротрипсина.
Гомогенаты спинного мозга мышей различного возраста подвергают SDS-PAGE и анализу вестерн-блот и затем зондируют в отношении нейротрипсина и С-концевого фрагмента агрина 100 кД, используя специфические антитела SZ 177 к нейротрипсину и R132 к С-концевому фрагменту агрина 100 кД. β-актин зондируют в качестве контроля в отношении равных количеств тканевого гомогената в различных образцах.
Фиг.4. Трансгенная сверхэкспрессия нейротрипсина в двигательных нейронах приводит в результате к повышенному уровню расщепления агрина.
Вестерн-блоты экстрактов спинного мозга зондируют антителами к человеческому (hNt) и мышиному (mNt) нейротрипсину, а также антителами к С-концевому фрагменту агрина 100 кД. Результаты демонстрируют повышенное распространение С-концевого фрагмента агрина 100 кД у мышей со сверхэкспрессией нейротрипсина.
Фиг.5. Нейротрипсин удаляет агрин из нервно-мышечного соединения (НМС).
НМС диафрагмы мышей, иммуноокрашенной на агрин в постнатальные дни 0 (Р0), 4 (Р4) и 8 (Р8). У трансгенных мышей со сверхэкспрессией нейротрипсина в двигательных нейронах аргин исчезает из НМС в период, исчисляемый часами-днями после начала сверхэкспрессии. Р4: переходное состояние. Частичная потеря агрина из НМС. Стрелки указывают на отдельные хорошо сформированные НМС. Звездочки указывают частично диспергированные НМС.Р8: почти полная потеря агрина из НМС. Стрелки указывают на отдельные хорошо сформированные НМС. Звездочки указывают частично диспергированные НМС.
Фиг.6. Нейротрипсин-зависимое удаление агрина из НМС осуществляется путем рассасывания постсинаптического аппарата.
НМС диафрагмы тех же мышей, что представлены на фиг.7, окрашивают на ацетилхолиновые рецепторы α-бунгаротоксином с флуоресцентной меткой (α-Btx). Ацетилхолиновые рецепторы исчезают за период, исчисляемый часами-днями после начала сверхэкспрессии. Р4: переходное состояние. Частичная потеря НМС. Стрелки указывают на отдельные хорошо сформированные НМС. Звездочки указывают частично диспергированные НМС.Р8: почти полная потеря НМС. Стрелки указывают на отдельные хорошо сформированные НМС. Звездочки указывают частично диспергированные НМС.
Фиг.7. Фрагментация НМС в камбаловидной мышце мышей со сверхэкспрессией Nt.
(А-С) Окрашивание α-бунгаротоксином (α-Btx) НМС мышей дикого типа показывает типичную Претцель-структуру.
(D-F) Окрашивание α-Btx НМС мышей со сверхэкспрессией Nt показывает выраженную фрагментацию постсинаптического аппарата.
(G-I) НМС трансгенных мышей со сверхэкспрессией каталитически неактивной формы Nt (Нейротрипсин Ser711Ala) не изменяются.
Фиг.8. Поперечные срезы через камбаловидную мышцу мышей дикого типа и со сверхэкспрессией нейротрипсина.
(A) Мышь дикого типа
(B) Мышь со сверхэкспрессией нейротрипсина.
По сравнению с мышами дикого типа мышцы мышей со сверхэкспрессией Nt включают меньше мышечных волокон. Стрелки в (А) и (В) указывают на одно мышечное волокно.
Фиг.9. Количественная оценка числа синапсов/объем ткани в нейропиле радиального слоя области СА1 гиппокампа.
У всех экспериментальных животных число синапсов/объем ткани определяют в электронно-микроскопических срезах, взятых из одного и того же участка радиального слоя участка СА1 гиппокампа.
wt: дикий тип;
ЦМВ-Cre: трансгенная линия, экспрессирующая рекомбиназу Cre под контролем промотора ЦМВ;
491(неакт.Nt): трансгенная линия 491, несущая неактивный трансген, включающий транскрипционный стоп-сегмент;
494(неакт.Nt): трансгенная линия 494, несущая неактивный трансген, включающий транскрипционный стоп-сегмент;
ДТГ(Nt491/cre): дважды трансгенная мышь, полученная из линии 491, у которой неактивный трансген нейротрипсина активирован скрещиванием с рекомбиназой Cre;
ДТГ(Nt494/cre): дважды трансгенная мышь, полученная из линии 494, у которой неактивный трансген нейротрипсина активирован скрещиванием с рекомбиназой Cre.
** p<0,01.
Фиг.10. Шипики на вторичных дендритных ответвлениях пирамидальных нейронов СА1.
Шипики на вторичных дендритных ответвлениях пирамидальных нейронов СА1 мышей дикого типа (А и В) и дважды трансгенных мышей со сверхэкспрессией нейротрипсин (С и D). Пирамидальные клетки СА1 ионтофоретически наполняют биоцитином во время электрофизиологических исследований in vitro и визуализируют, используя гистохимический метод с применением авидин-биотин-пероксидазы. Дендриты мышей дикого типа имеют много длинных хорошо развитых шипиков (большие стрелки); кроме того, обнаруживают также много коротких плотных шипиков (маленькие размерные стрелки). Среди дендритов мышей со сверхэкспрессией нейротрипсина (животные одного помета) преобладают короткие плотные шипики (маленькие размерные стрелки); длинные хорошо развитые шипики (большие стрелки) встречаются очень редко. Общая густота шипиков (число шипиков/единицу длины дендрита) заметно ниже у мышей со сверхэкспрессией нейротрипсина (С и D).
Фиг.11. Очищенный человеческий нейротрипсин полной длины.
SDS-PAGE с последующим окрашиванием серебром (А) и вестерн-блоттингом (В) показывает одну полосу человеческого нейротрипсина полной длины (указано стрелками), мигрирующую в положении, соответствующем приблизительно 75 кД в невосстанавливающих условиях. Иммунодетекцию (В) проводят, используя антитело к нейротрипсину. Молекулярные массы стандартов (кД) указаны на полях слева.
Фиг.12. Очищенный агрин-EGFP.
Очищенный инженерный агрин-EGFP (указано стрелками) показан на окрашенном серебром SDS-геле (А) и на вестерн-блоте, определенный с помощью антитела, образующегося к С-концевой части агрина (В). Молекулярные массы стандартов (кД) указаны на полях слева. Отмечают, что EGFP используют только в качестве указателя места в данной конструкции, которая разработана так, чтобы включала только центр расщепления α, но не центр расщепления β.
Фиг.13. Очищенный фрагмент агрин-С45.
(A) Окрашенный серебром гель SDS-PAGE, показывающий очищенный фрагмент агрин-С45 (указано стрелками), мигрирующий ниже 50 кД. Числа указывают молекулярные массы стандарта точности и белка (BIORAD).
(B) Вестерн-блот, определяющий очищенный фрагмент агрин-С45 (указано стрелками), с использованием StrepTactin для детекции С-концевой метки strep. Числа указывают молекулярные массы стандарта точности и белка (BIORAD).
Фиг.14. Анализ активности нейротрипсина с использованием агрин-EGFP в качестве субстрата.
Для тестирования активности очищенного нейротрипсина на центр расщепления α субстрат, включающий только центр расщепления а (агрин-EGFP), инкубируют в виде монокомпонента (-) и совместно с нейротрипсином (+) и затем подвергают SDS-PAGE с последующим вестерн-блоттингом с использованием антитела к С-концевому фрагменту расщепления агрина (см. пример 22). Дорожка 1 показывает агрин-EGFP (указано стрелкой, обозначенной Ag-EGFP) без обработки нейротрипсином в качестве контроля. Дорожка 2 показывает агрин-EGFP (указано стрелкой, обозначенной Ag-EGPF) и С-концевой фрагмент приблизительно 150 кД, генерированный активностью нейротрипсина (указано стрелкой, обозначенной Ag-CF). Маркер молекулярной массы в кД (килодальтонах).
Фиг.15. Анализ на активность нейротрипсина с использованием агрин-С45 в качестве субстрата.
Для тестирования активности очищенного нейротрипсина на центр расщепления β агрина субстрат, включающий только центр расщепления β (агрин-С45), инкубируют в виде монокомпонента (-) и совместно с нейротрипсином (+) и затем подвергают SDS-PAGE.
(A) Окрашенный серебром гель SDS-PAGE, показывающий 250 нг агрина-С45, инкубируют в буфере для анализа в течение 3 часов без нейротрипсина (-) и 250 нг агрина-С45 инкубируют в буфере для анализа в течение 3 часов с добавлением нейротрипсина (+). Стандарт точности и белка (BIORAD) показан слева, числа указывают молекулярные массы (кД). Агрин-С45 (указано стрелкой) можно видеть ниже 50 кД. Продукты расщепления агрина-С45 обнаруживают между 20 и 25 кД (указано стрелками). Ag-C45-NF: N-концевой фрагмент расщепления агрина-С45; Ag-C45-CF: С-концевой фрагмент расщепления агрина-С45.
(B) Вестерн-блот тех же образцов, что в (А), в котором нерасщепленный агрин-С45 и расщепленный С-концевой фрагмент агрина-С45 определяют с помощью их концевой Strep-метки, используя StrepTactin (IBA GmbH). Ag-C45-CF: С-концевой фрагмент расщепления агрина-С45 (указано стрелкой).
Фиг.16. Анализ на основе вестерн-блоттинга, направленный на скрининг ингибиторов нейротрипсина, с помощью детекции антителами субстрата агрина и С-концевого продукта.
Верхняя полоса показывает белок агрин-EGFP (стрелка, обозначенная Ag-EGFP) с молекулярной массой от 250 до 600 кД, используемый в качестве субстрата. Нижняя полоса показывает С-концевой фрагмент агрина-EGFP, генерированного нейротрипсином, с молекулярной массой приблизительно 150 кД (стрелка обозначена Ag-CF), который появляется с различной интенсивностью в соответствии с ингибирующей активностью тестируемых молекул ингибиторов No. 7, 47, 48, 49, 50 и 51. Гистограмма показывает относительные интенсивности (I) полосы 150 кД (Ag-CF), генерированной опосредованным нейротрипсином расщеплением агрина-EGFP, относительно набора положительного контроля до 100% и набора отрицательного контроля до 0%. Отрицательный контроль (-): только агрин-EGFP без нейротрипсина. Положительный контроль (+): агрин-EGFP с добавлением нейротрипсина.
No. 7: N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламид
No. 47: 4-хлорциклогекс-4-ен-1,2-дикарбоновая кислота N1-амидиносульфаниламид
No. 48: N1-амидино-N4-(4-диметиламинобензилиден)-сульфаниламид
No. 49: N1-амидино-N4-бензилиден-сульфаниламид
No. 50: N1-амидино-N4-(2,4-дихлорбензилиден)-сульфаниламид
No. 51: N1-амидино-N4-(4-метоксибензилиден)-сульфаниламид
Фиг.17. Дозозависимое ингибирование активности нейротрипсина соединением No.7.
(A) Определение с помощью вестерн-блота С-концевого фрагмента агрина (Ag-CF) 150 кД, генерированного опосредованным нейротрипсином расщеплением агрина-EGFP в зависимости от концентрации соединения No. 7, N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламида.
Дорожка 1: Агрин-EGFP
Дорожка 2: Агрин+мышиный нейротрипсин
Дорожка 3: Агрин+мышиный нейротрипсин + 25 мкМ соединения No. 7
Дорожка 4: Агрин+мышиный нейротрипсин + 37,5 мкМ соединения No. 7
Дорожка 5: Агрин+мышиный нейротрипсин + 50 мкМ соединения No. 7
Дорожка 6: Агрин+мышиный нейротрипсин + 75 мкМ соединения No. 7
Дорожка 7: Агрин+мышиный нейротрипсин + 100 мкМ соединения No. 7
(B) График данных интенсивности, полученных в (А) относительно концентрации ингибитора с I=100%, интенсивность агрина в сочетании с мышиным нейротрипсином без добавления соединения ингибитора No. 7.
Фиг.18-24. Тесты специфичности соединения No.7 (N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламида): Отсутствие ингибирования тестируемых протеаз Ха, трипсина, tPA, тромбина, урокиназы, калликреина и плазмина.
Графики показывают исходные скорости реакции (Vini) тестируемых протеаз Ха (см. фиг.18), трипсина (см. фиг.19), tPA (см. фиг.20), тромбина (см. фиг.21), урокиназы (см. фиг.22), калликреина (см. фиг.23) и плазмина (см. фиг.24), построенных относительно концентраций субстрата (мкМ) в отсутствие (белые квадраты) и в присутствии (белые треугольники) соединения No. 7. В качестве положительного контроля конкурентного ингибирования в измерениях для tPA (см. фиг.20) и урокиназы (см. фиг.22) добавляют бензамидин (ВА) и в анализах Ха (см. фиг.18) и плазмина (см. фиг.24) добавляют пара-аминобензамидин (рАВА) в указанных концентрациях (белые ромбы).
Детальное описание изобретения
Изобретение основано на том факте, что ингибирование нейротрипсина позволяет усилить просинаптическую (синапс-формирующую, синапс-дифференцирующую, синапс-организующую, синапс-защищающую, синапс-усиливающую) активность. Ген нейротрипсина экспрессируется во многих нейронах центральной нервной системы (см. статьи Gschwend, T.P., et al., Molec. Cell Neurosci. 9: 207-219, 1997; Wolfer, D.P. et al., Molec. Cell. Neurosci. 18: 407-433, 2001), включая двигательные нейроны спинного мозга (см. пример 1), и белок нейротрипсина обнаруживают во многих синапсах ЦНС (см. статью Molinari, F. et al., Science 298: 1779-1781, 2002), а также в нейромышечном соединении. Нейротрипсин играет существенную роль в развитии и/или поддержании хорошо сбалансированной синаптической функции. Слишком большое количество нейротрипсина (сверхэкспрессия) коррелирует со слишком маленьким числом синаптических связей. Трансгенные мыши со сверхэкспрессией нейротрипсина в нейронах ЦНС демонстрируют уменьшенное число синапсов в коре головного мозга и гиппокампе, двух структурах головного мозга, которые очень важны для когнитивных функций, таких как память и обучение. Аналогичным образом трансгенные мыши со сверхэкспрессией нейротрипсина в спинальных двигательных нейронах показывают уменьшение нейромышечных соединений (НМС), синапсов, которые опосредуют нейроконтроль мышечной активности.
Изменения в нейромышечных соединениях диафрагмы трансгенных мышей, которые способны к сверхэкспрессии нейротрипсина в двигательных нейронах, напоминают обусловленные направленной инактивацией гена агрина. Протеогликан агрин представляет собой очень хорошо охарактеризованный просинаптический (синапс-формирующий, синапс-дифференцирующий, синапс-организующий, синапс-защищающий, синапс-усиливающий) агент (см. статью Sanes, J.R. и Lichtman, J., Nature Rev. Neurosci. 2: 791-805, 2001). Он имеет белок ядра массой приблизительно 220 кД. Агрин существует в нескольких изоформах. Они кодируют как секретируемые белки внеклеточного матрикса, так и трансмембранные белки типа II, которые несут очень короткий N-концевой цитоплазматический сегмент. Участок агрина, который несет просинаптическую активность, находится в С-концевой группе агрина, в частности в 3 домене ламинина G (см. статью Bezakova, G. и Ruegg, M.A., Nature Rev. Molec. Cell Biol. 4: 295-308, 2003). Агрин является субстратом нейротрипсина (см. пример 2). Нейротрипсин расщепляет агрин по двум центрам (см. пример 25). Один центр (называемый α-центром) находится между аргинином 995 (R995) и аланином 996 (А996). Другой центр (называемый β-центром) находится между лизином 1754 (K1754) и серином 1755 (S1755). Номера аминокислот относятся к заякоренному в мембране агрину (сплайс-вариант А4В0) крысы (NP_786930). Однако как центр расщепления α, так и центр расщепления β достаточно консервативны в агрине млекопитающих, включая агрин человека. Расщепление агрина нейротрипсином генерирует фрагмент приблизительно 100 кД (килодальтон), расположенный от А996 до K1754 и фрагмент приблизительно 22 кД, расположенный от S1755 до С-конца. Расщепление обоих, α- и β-центра, отделяет активность, организующую агриновый синапс, от N-концевой группы агрина. Расщепление агрина происходит также in vivo. Обнаружено, что у мышей дикого типа фрагмент агрина 100 кД наиболее часто встречается во время первых постнатальных недель, в период, когда связанная с развитием экспрессия нейротрипсина достигает максимума (см. пример 3). Преобладание фрагмента агрина 100 кД заметно повышено у трансгенных мышей со сверхэкспрессией нейротрипсина в двигательных нейронах (см. примеры 4 и 5).
Агрин является естественным субстратом нейротрипсина как в НМС (см. пример 6), так и в ЦНС (см. пример 14). Расщепляя агрин, нейротрипсин противодействует просинаптической активности агрина. Избыточный нейротрипсин в нейромышечном соединении трансгенных мышей контролирует исчезновение ранее образованных нейромышечных соединений в течение меньше чем трех дней (см. примеры 4-7). Данные наблюдения позволяют оценить нейротрипсин как дестабилизирующий синапс или антисинаптический агент.
Совместное существование про- и антисинаптических агентов поддерживает представление о том, что цепь нейронов нервной системы является динамичной, а не фиксировано связанной системой. Сбалансированное соответствие между просинаптическими и антисинаптическими факторами приводит в результате к гомеостазу. Адаптивные изменения, которые требуются, например, когда необходимо изменить синаптическое соединение с целью удовлетворения изменившихся потребностей, сдвигают баланс между просинаптическими и антисинаптическими силами контролируемым образом. Это тонкое жестко контролируемое взаимодействие между про- и антисинаптическими силами подвержено нарушению регуляции, приводящему к несоответствующему синаптическому гомеостазу или несоответствующей адаптации к функциональным требованиям. Синаптическое заболевание может произойти, когда степень нарушения регуляции превышает пороговое значение.
Фармацевтическое регулирование активности нейротрипсина обеспечивает беспрецедентный доступ к регуляторному механизму синаптической функции. Ингибирование протеолитической активности нейротрипсина будет сдвигать синаптическое равновесие в сторону усиления просинаптических активностей при сокращении антисинаптических активностей и, таким образом, в сторону увеличения числа и/или размера, и/или силы синапсов.
Эксперименты с трансгенными мышами, способными к сверхэкспрессии нейротрипсина в спинальных двигательных нейронах, показывают корреляцию между атрофией скелетных мышц (см. примеры 8 и 10) и нарушением синаптических связей (см. пример 9). Ингибиторы нейротрипсина будут уравновешивать результат избытка нейротрипсина и обеспечат возможность лечения и профилактики атрофии скелетных мышц, вызываемой потерей синаптических связей, например атрофии скелетных мышц у пожилых пациентов.
В линиях трансгенных мышей, способных к сверхэкспрессии нейротрипсина в двигательных нейронах, наблюдают сильную атрофию скелетных мышц, которая в основном обусловлена заметным уменьшением числа мышечных волокон (см. пример 10). Количественную оценку эффекта избыточной продукции нейротрипсина двигательными нейронами приводят в табл.1. Избыточные уровни нейротрипсина, продуцируемые двигательными нейронами, приводят в результате к уменьшению числа мышечных волокон камбаловидной мышцы взрослых мышей в интервале от 18 до 48% в зависимости от уровня сверхэкспрессии нейротрипсина. Поскольку сверхэкспрессия нейротрипсина ограничена двигательными нейронами (в плане экспериментальных деталей см. пример 4), данные результаты показывают, что нейротрипсин, экспрессируемый двигательными нейронами, действует местно через нейромышечное соединение на мышечном волокне-мишени. Данный местный атрофический эффект жестко зависит от протеолитической активности нейротрипсина, поскольку мышцы мышей, способных к сверэкспрессии каталитически неактивной формы нейротрипсина, демонстрируют нормальные числа волокон.
Мыши со сверхэкспрессией нейротрипсина в двигательных нейронах проявляют в существенной мере повышенную фрагментацию нейромышечных соединений (см. пример 9). Фрагментация в сочетании с пониженным числом волокон является характерным признаком, наблюдаемым в скелетных мышцах пожилых людей и животных. Как отмечено выше, нарушение нейромышечных соединений и потеря мышечных волокон не вызываются сверхэкспрессией каталитически неактивного нейротрипсина. Это характеризует выделенный из двигательного нейрона нейротрипсин как фактор, который снижает уровень иннервации мышечных волокон и, в конечном счете, приводит к их потере. Предполагают, что агенты с активностью снижения уровня иннервации играют роль во время стадии, связанной с развитием элиминации синапсов как в нейромышечном соединении, так и в центральной нервной системе. Возможно, активность снижения уровня синапсов существует на протяжении взрослой жизни и играет роль в поддержании равновесия между пресинаптическими и постсинаптическими элементами. Временный характер экспрессии нейротрипсина поддерживает данную возможность, поскольку он достигает максимума в период, связанный с развитием элиминации синапсов (первые две постнатальные недели у мышей и крыс), и затем продолжает экспрессироваться на более низком уровне на протяжении взрослой жизни.
Сверхэкспрессия нейротрипсина в двигательных нейронах трансгенных мышей приводит в результате в разрушению нейромышечного соединения (см. пример 7). Систематические анализы роли нейротрипсина для поддержания нейромышечного соединения в различных типах мышц, включая диафрагму и камбаловидную мышцу, показывают, что сверхэкспрессия нейротрипсина в двигательных нейронах уменьшает размер нейромышечного соединения. Сильная сверхэкспрессия нейротрипсина в двигательных нейронах приводит в результате к полному рассасыванию ранее образованных нейромышечных соединений. Вследствие разрушающего синапсы эффекта нейротрипсина двигательные нервы, оставшиеся без постсинаптической специализации и/или со структурно и функционально пониженной постсинаптической специализацией, начинают расти, выходя их центра прежнего НМС. Нервы, теперь растущие над поверхностью мышечного волокна, образуют маленькие эктопические синапсы, которые кажутся незрелыми при электронно-микроскопическом исследовании, как заключают на основании отсутствия вторичных складок в постсинаптической мембране.
Сверхэкспрессия нейротрипсина в двигательных нейронах трансгенных мышей приводит в результате к расщеплению протеогликана агрина (см. пример 5). В результате С-концевая группа агрина исчезает из НМС (см. пример 6). Участок агрина, который несет НМС-консервативную и НМС-стимулирующую активность агрина, находится в С-концевой группе агрина, в частности в 3 домене ламинина G (см. статью Bezakova, G., Nature Reviews Molecular Cell Biology 4: 295-308, 2003). Вследствие этого удаление С-концевого домена агрина оставляет НМС незащищенным от так называемого разрушающего фактора, и НМС разрушается и исчезает в течение нескольких дней. Повышающая регуляция нейротрипсина в двигательных нейронах в период, когда промотор Thy-1 начинает управлять экспрессией трансгена нейротрипсина (через 2-5 дней после рождения), приводит в результате к исчезновению агрина из НМС в течение нескольких дней (см. пример 6). Вскоре после исчезновения агрина исчезает также постсинаптический ацетилхолиновый рецептор (см. пример 7). В заключение данные наблюдения показывают цепь событий, которые начинаются с повышающей регуляции нейротрипсина в двигательных нейронах. Избыточный нейротрипсин, в свою очередь, расщепляет агрин в НМС и удаляет С-концевую группу агрина из НМС. Поскольку С-концевая группа включает активный центр, связанный с НМС-защищающей и НМС-стимулирующей способностью агрина, НМС теперь остается незащищенным от разрушающего фактора и, в свою очередь, разлагается.
Наблюдение заметного снижения числа нервных волокон в скелетных мышцах мышей, которые способны к избирательной сверхэкспрессии нейротрипсина в двигательных нейронах, показывает, что нейротрипсин причинно связан с конечным разрушением, приводящим в результате к денервации с последующей атрофией и, в конечном счете, потере денервированных мышечных волокон. Данные наблюдения согласуются с наблюдениями, сделанными на мышцах и нейромышечных соединениях человека и животных с возрастной скелетной мышечной атрофией. В случае, когда зависимая от возраста потеря мышечных волокон происходит вследствие однонаправленного действия множества факторов, контролируемое и тонкое частичное ингибирование нейротрипсина может нарушить процесс конечного разрушения, денервации и потери мышечных волокон. Вследствие этого ожидают, что ингибирование нейротрипсина, будет иметь положительный эффект на зависимую от возраста денервацию мышечных волокон и атрофию скелетных мышц.
Атрофия скелетных мышц сопровождается существенной потерей мышечной силы и играет главную роль в патогенезе хрупкости и функционального нарушения, которое происходит при старении. Слабость нижних конечностей включает ряд функциональных нарушений, таких как затруднения, в том, чтобы подняться со стула или встать с постели, замедленная походка и другие движения и трудности в поддерживании равновесия, приводящие в результате к падениям и повреждениям. Потеря волокон скелетных мышц оказывает отрицательное воздействие как на абсолютную силу, которую может развить мышца, так и на скорость, с которой мышца может развить силу.
Старение связано с прогрессирующим снижением скорости метаболизма, которое, в свою очередь, имеет существенные физиологические последствия, включая пониженную переносимость тепла и холода, а также повышенную вероятность развития ожирения. Скелетные мышцы включают приблизительно 40% не содержащей жира массы тела и играют важную гомеостатическую роль в метаболизме организма. Вследствие этого уменьшение массы скелетных мышц с возрастом является основным фактором, обусловливающим пониженную скорость метаболизма. Предупреждая прогрессирующую потерю волокон, ингибирование нейротрипсина воздействует на данные метаболические и физиологические последствия.
Прогрессирующую потерю массы и силы скелетных мышц с возрастом считают основным фактором, обусловливающим постепенное уменьшение плотности костей, наблюдаемое при старении. Напротив, хорошо известно, что силы, воздействующие на кости при мышечной активности, стимулируют формирование костей. Таким образом, силы, генерируемые при сокращении мышц, представляют собой важный фактор, определяющий качество костей. Предупреждение потери мышечных волокон путем ингибирования периферической активности нейротрипсина может, таким образом, препятствовать или уменьшать неблагоприятные эффекты на качество скелетных мышц и опосредованно оказывать антагонистическое действие на развитие остеопороза.
Благоприятные эффекты ингибирования нейротрипсина можно также ожидать при атрофиях скелетных мышц, которые имеют место в многочисленных клинических ситуациях, при которых ослабление мышц является сопутствующей проблемой, включая рак, СПИД и сепсис.
Нейротрипсин обладает также антисинаптической функцией в центральной нервной системе (ЦНС). мРНК нейротрипсина экспрессируется нейронами серого вещества ЦНС (см. статью Gschwend, Т.Р., et al., Molec. Cell Neurosci. 9: 207-219, 1997) и белок нейротрипсина встречается в изобилии на богатых синапсами участках во многих областях головного мозга (см. статью Molinari, F. et al., Science 298: 1779-1781, 2002). Особенно высокие концентрации белка нейротрипсина обнаруживают на богатых синапсами участках коры головного мозга, гиппокампе и миндалевидной железы. Однако другие богатые синапсами участки также демонстрируют обильную экспрессию нейротрипсина. При большом увеличении белок нейротрипсина обнаруживают в мембране пресинаптического окончания, в частности в области мембраны, выстилающей синаптическую щель (см. статью Molinari, F. et al., Science 298: 1779-1781, 2002). Наиболее интенсивное иммуноокрашивание на нейротрипсин обнаруживают над синаптическими активными зонами пресинаптического окончания. Иногда иммунореактивность в отношении нейротрипсина наблюдают также в пузырьках пресинаптического окончания. Однако заслуживает внимания, что большинство пресинаптических пузырьков лишено иммунореактивности в отношении нейротрипсина. Независимое доказательство синаптической локализации нейротрипсина получают с помощью биохимического подхода, т.е. путем анализа синаптосом. Иммуноцитохимическая локализация нейротрипсина в мембране пресинаптической активной зоны идентична у человека и мышей. Подводя итог, результаты показывают, что нейротрипсин находится в пресинаптическом окончании, в частности в пресинаптической мембране, выстилающей синаптическую щель в пресинаптической активной зоне.
Вследствие локализации в синапсе, в частности в пресинаптической активной зоне, нейротрипсин находится в стратегическом положении в плане контроля синаптической структуры и функции. Роль нейротрипсина как модулятора синаптической структуры и функции, в частности, его функции как антисинаптического агента в ЦНС, продемонстрирована в экспериментах с трансгенными мышами, способными к сверхэкспрессии нейротрипсина в нейронах ЦНС (см. пример 11). Избыточные количества нейротрипсина, продуцируемые нейронами ЦНС, вызывают существенное снижение числа и размера синапсов в центральной нервной системе. Доказательство структурных изменений получают как морфологическими, так и электрофизиологическими методами.
Подсчет синапсов на участках нейропилей показывает уменьшение числа синапсов/площадь (см. пример 12). Проверка дендритных шипиков, расположенных вдоль наполненных красителем дендритов, показывает уменьшение размера и числа шипиков у мышей, способных к сверхэкспрессии нейротрипсина (см. пример 13). Данные результаты взаимосогласуются, поскольку многие синапсы заканчиваются на дендритных шипиках. Вследствие этого уменьшение числа синапсов и уменьшение числа дендритных шипиков представляют два проявления данного феномена. Вместе взятые данные наблюдения ясно демонстрируют антисинаптическую функцию нейротрипсина.
В ЦНС нейротрипсин также опосредует расщепление агрина (см. примеры 14 и 15). Протеогликан агрин присутствует как в нейромышечном соединении (см. статью Sanes, J.R. и Lichtman, J., Nat. Rev. Neurosci. 2: 791-805, 2001), так и в синапсах центральной нервной системы (см. статьи Smith, M.A. и Hilgenberg, L.G., Neuroreport 13: 1485-1495, 2002; Kroger, S. и Schroder, J.E., News Physiol. Sci. 17: 207-212, 2002). Показано, что в гомогенатах ЦНС мышей дикого типа фрагмент агрина 100 кД наиболее часто встречается в первые постнатальные недели, время, когда связанная с развитием экспрессия агрина максимальна (см. пример 14). Преобладание фрагмента агрина 100 кД заметно повышено у трансгенных мышей со сверхэкспрессией нейротрипсина в нейронах ЦНС (см. пример 15). В присутствии нейротрипсина, который инактивирован заменой серина активного центра аланином, расщепление агрина не происходит. Таким образом, протеолитическая активность нейротрипсина явно опосредует расщепление агрина и генерацию фрагмента агрина 100 кД в ЦНС. Форма агрина, используемая в данных экспериментах, представляет собой форму, заякоренную в мембране. Данная связанная по N-концу форма агрина в основном обнаружена в центральной нервной системе, и, как показано, она контролирует дифференцировку синапсов в ЦНС (см. статью Bose, C.M. et al., J. Neurosci. 20: 9086-9095, 2000).
В заключение, нейротрипсин обладает антисинаптической функцией не только в НМС, но также в ЦНС. Слишком большое количество нейротрипсина (сверхэкспрессия) коррелирует со слишком маленьким количеством синаптических связей в коре головного мозга и в гиппокампе, двух структурах головного мозга, которые очень важны для когнитивных функций, таких как память и обучение. Организующий синапс протеогликан агрин представляет собой также физиологический субстрат нейротрипсина в ЦНС. Посредством расщепления агрина нейротрипсин противодействует просинаптической активности агрина.

Совместное существование и взаимодействие просинаптического агента агрина и антисинаптического агента нейротрипсина в ЦНС поддерживает представление о том, что цепь нейронов нервной системы является динамичной, а не фиксированно связанной системой. Гомеостаз поддерживается сбалансированным соответствием между просинаптическими и антисинаптическими факторами. Когда требуются адаптивные изменения, например, когда необходимо изменить цепь для сохранения воспоминаний, равновесие между просинаптическими и антисинаптическими силами сдвигается контролируемым образом. Когда достигается изменение в цепи, равновесие между просинаптическими и антисинаптическими силами восстанавливается.
Тонкое жестко контролируемое взаимодействие между про- и антисинаптическими силами подвержено нарушению регуляции, приводящему к несоответствующему синаптическому гомеостазу или несоответствующей адаптации к функциональным требованиям. Синаптическое заболевание может произойти, когда степень нарушения регуляции превышает пороговое значение.
Фармацевтическое регулирование активности нейротрипсина обеспечивает беспрецедентный доступ к регуляторному механизму синаптической функции. Ингибирование протеолитической активности нейротрипсина приводит в результате к более длительному существованию и, вследствие этого, повышенной концентрации агрина. Вследствие данной связи между нейротрипсином и агрином пониженная антисинаптическая активность, индуцируемая ингибированием нейротрипсина, повышает просинаптическую активность агрина. Следовательно, равновесие сдвигается в сторону усиления просинаптической активности, приводя в результате к увеличению числа, размера и силы синапсов.
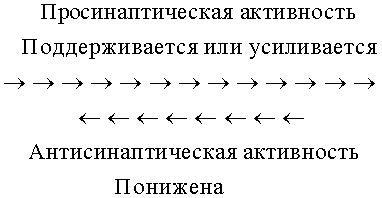
Представление о регуляции синапса путем снижения антисинаптической активности нейротрипсина и вследствие этого усиления просинаптических активностей при истощении антисинаптичеких активностей предлагает широкий круг применений в области нарушенных когнитивных функций головного мозга. В частности, ингибирование нейротрипсина благоприятно при заболеваниях и субклинических состояниях, при которых требуется формирование синапсов и увеличение размера и силы имеющихся синапсов.
Ингибирование нейротрипсина используют при лечении шизофрении. Избыточный нейротрипсин в синапсе направляет синаптический прунинг и, таким образом, генерирует синаптический фенотип, который соответствует синаптическому фенотипу, обнаруживаемому в головном мозге пациентов с шизофренией. Данное экспериментальное наблюдение определяет нейротрипсин как один из факторов, который направляет прунинг синапсов. В случае, когда избыточный синаптический прунинг происходит вследствие однонаправленного действия множества факторов, стимулирующих прунинг, контролируемое и тонкое частичное ингибирование нейротрипсина снижает до минимума направленность на синаптический прунинг. Это дает возможность выхода из "шизофренического синаптического фенотипа" и приводит в результате к ослаблению шизофренических симптомов. Уменьшение числа синапсов в ЦНС мышей, способных к сверхэкспрессии нейротрипсина, показывает, что ингибирование нейротрипсина дает в результате более низкую степень синаптического прунинга и, таким образом, повышенное число синапсов и повышенную способность нейронов к связыванию и коммуникации. Соединения, представленные в изобретении, ингибирующие ферментную функцию нейротрипсина, вследствие этого используют для реверсии синаптических изменений при шизофрении и для восстановления нормальной синаптической структуры и функции и, таким образом, для блокирования или укорочения шизофренических эпизодов и защиты от новых эпизодов.
Ингибирование нейротрипсина также поддерживает усиление когнитивной функции при слабом когнитивном нарушении и других клинических и субклинических состояниях, связанных с пониженными когнитивными функциями. Слабое когнитивное нарушение, а также другие клинические и субклинические состояния поврежденных когнитивных функций, как обнаружено, ассоциированы с очевидностью атрофии ткани головного мозга в некоторых областях ЦНС. Уменьшение числа синапсов в ЦНС мышей, способных к сверхэкспрессии нейротрипсина, показывает, что ингибирование нейротрипсина дает в результате увеличенное число синапсов и повышенную способность нейронов к связыванию и коммуникации. Соединения, представленные в изобретении, ингибирующие ферментную функцию нейротрипсина, вследствие этого используют для реверсии синаптических изменений при всех клинических и субклинических нарушениях, при которых включены уменьшенное число синапсов или пониженная функция синапсов, и для восстановления нормальной синаптической структуры и функции. Тем самым фармацевтическое ингибирование нейротрипсина может улучшать когнитивные функции при различных состояниях с пониженными когнитивными функциями гетерогенного происхождения.
Основываясь на данных фактах, изобретение далее относится к применению ингибиторов нейротрипсина формулы (1), как описано выше и ниже, для лечения и/или профилактики заболеваний, вызываемых недостатком синапсов, например атрофии скелетных мышц, шизофрении и когнитивного расстройства. Атрофия скелетных мышц, которую предполагают лечить, представляет собой, в частности, так называемую саркопению, т.е. атрофию скелетных мышц, обусловленную старением, атрофию скелетных мышц, сопровождающуюся остеопорозом, и атрофию скелетных мышц, обусловленную истощением, ассоциированным с рядом заболеваний, таких как рак, СПИД и сепсис, или, кроме того, атрофию скелетных мышц как следствие иммобилизации и/или постельного режима вследствие тяжелого повреждения или тяжелой болезни. Шизофрения, которую предполагают лечить, представляет собой расстройство из всей области шизофрении и расстройств, подобных шизофрении, включая хроническую шизофрению, хронические шизоаффективные расстройства, неспецифические нарушения, острую и хроническую шизофрению различной симптоматики, как, например, тяжелая неремиссионная „Крепелиновая" шизофрения или прототип DSM-III-R расстройств, подобных шизофрении, эпизодические шизофренические расстройства, бредовые расстройства, подобные шизофрении, расстройства личности, подобные шизофрении, такие как, например, расстройства личности, подобные шизофрении с мягкой симптоматикой, шизотипические расстройства личности, латентные формы шизофренических и подобных шизофрении расстройств и неорганические психотические расстройства. Более того, ингибиторы нейротрипсина, как описано в данном контексте, могут быть использованы в качестве стимуляторов познавания, для улучшения работы головного мозга и для облегчения обучения и функций памяти. Когнитивными недостаточностями, которые предполагают лечить, являются мягкое когнитивное расстройство, например, при потенциальной ранней стадии болезни Альцгеймера, нарушение когнитивной функции без деменции у пожилых людей и нарушение когнитивных функций у пациентов с болезнью Альцгеймера, болезнью Паркинсона, рассеянным склерозом, инсультом и травмой головы.
Аналогичным образом изобретение относится к применению данных ингибиторов формулы (1) для изготовления лекарственного препарата для лечения и/или профилактики заболеваний, вызываемых недостатком синапсов, например атрофии скелетных мышц, шизофрении и когнитивного расстройства. Кроме того, изобретение относится к лечению и/или профилактике заболеваний, вызываемых недостатком синапсов, например, атрофии скелетных мышц, шизофрении и когнитивного расстройства, которые включают введение соединения формулы (1) или его фармацевтически приемлемой соли в количестве, эффективном в отношении указанного заболевания, теплокровному животному, которому требуется данное лечение. Соединения формулы (1) можно вводить как таковые или главным образом в форме фармацевтических композиций профилактически или терапевтически, предпочтительно в количестве, эффективном в отношении указанных заболеваний, теплокровному животному, например человеку, которому требуется данное лечение. В случае пациента, имеющего массу тела приблизительно 70 кг, вводимая дневная доза составляет от приблизительно 0,05 г до приблизительно 5 г, предпочтительно от приблизительно 0,25 г до приблизительно 1,5 г соединения, представленного в настоящем изобретении.
Нейротрипсин состоит из богатого пролином основного домена (РВ), kringle-домена (KR), трех (мышиный нейротрипсин, mNt) или четырех (человеческий нейротрипсин, hNt) богатых цистеином доменов фагоцитарного рецептора (SRCR1, SRCR2, SRCR3 и SRCR4) и домена протеазы (PROT) (см. фиг.1). Центр активации зимогена (ZA) представляет собой центр расщепления на N-конце домена протеазы нейротрипсина. Протеолитическое расщепление в центре ZA превращает белок нейротрипсина из каталитически неактивной в каталитически активную форму. Посредством данного расщепления в случае мышиного нейротрипсина генерируются фрагмент приблизительно 55 кД, включающий некаталитический участок, и фрагмент приблизительно 30 кД, включающий домен протеазы. В случае человеческого нейротрипсина генерируются фрагменты, имеющие молекулярную массу 67 кД и 30 кД, соответственно.
Биохимический анализ человеческого нейротрипсина и поиск ингибиторов нейротрипсина требует количеств белка в интервале от миллиграмма до грамма. Для оптимальной продукции и секреции нейротрипсина протестирован ряд эукариотических экспрессионных систем, включая стабильную экспрессию в клетках мышиной миеломы, опосредованную бакуловирусом экспрессию в клетках насекомых и временную экспрессию в человеческих эмбриональных почечных клетках (НЕК), временную экспрессию в клетках яичников китайского хомячка (СНО) и стабильную экспрессию в Picchia pastoris. Данные системы имеют преимущество в том, что они могут быть легко адаптированы к бессывороточным условиям с целью уменьшения количества примесных белков в супернатанте и с целью создания системы для широкомасштабной продукции. Экспрессию нейротрипсина можно осуществить во всех данных экспрессионных системах. Однако наиболее эффективную продукцию и секрецию нейротрипсина получают в клетках миеломы, как описано в примерах 16 и 17.
Экспрессия в эукариотических клетках альтернативно может быть достигнута с помощью ряда эукариотических экспрессирующих векторов (коммерчески доступных или сделанных своими силами). Аналогично может быть использован ряд эукариотических клеточных линий, в том числе клетки НЕК293Т и HEK293-EBNA, клетки COS, клетки СНО, клетки HeLa, клетки Н9, клетки Jurkat, клетки NIH3T3, клетки С 127, клетки CV1 или клетки Sf.
Продукция нейротрипсина может быть также основана на клеточных линиях млекопитающих, проявляющих эндогенную экспрессию нейротрипсина. Экспрессию эндогенного человеческого нейротрипсина наблюдают на уровне РНК в клеточной линии человеческих мастоцитов НМС-1 (см. статью Poorafshar, М. и Hellman, L., Eur. J. Biochem. 261: 244-250, 1999). Клеточная линия НМС-1 представляет собой природный источник надлежащим образом процессированного и, вследствие этого, с большой вероятностью активного человеческого нейротрипсина. Данные клетки могут быть выращены в суспензионной культуре, и они конститутивно экспрессируют человеческий нейротрипсин. Белок, экспрессируемый клетками НМС-1, можно определить как полосу 97 кД с помощью специфического поликлонального антитела, возникающего к kringle-домену в экспериментах с использованием вестерн-блота.
Для очистки стандарта белка нейротрипсина используют способы очистки (см. примеры 18 и 19). Предпочтительно, когда используют аффинную очистку на гепариновой колонке, затем колонку гидрофобного взаимодействия и хроматографическую колонку с иммобилизованным хелатом металла. Элюированный белок затем очищают ионообменной хроматографией на колонке Mono S. В зависимости от требований эксперимента для очистки нейротрипсина используют также дополнительные или альтернативные хроматографические стадии на ионообменных колонках (DEAE или Mono Q) или гель-фильтрацию.
Изобретение относится к способу измерения каталитической активности нейротрипсина, отличающемуся тем, что нейротрипсин, его вариант или фрагмент, включающий домен протеазы нейротрипсина, и белок или пептид, включающий агрин, его вариант или фрагмент, включающий центр расщепления α или центр расщепления β агрина (см. примеры 20 и 21), инкубируют совместно в водном буферном растворе и подсчитывают количество расщепления агрина (см. пример 24).
Как используют в данном контексте, выражение "белок или пептид, включающий агрин, его вариант или фрагмент" означает агрин человека или другого млекопитающего либо птицы, слитый белок данного агрина с одним или более, например двумя или тремя, другими пептидами или белками, в частности с маркерным белком, например, зеленым флуоресцентным белком (GFP), белком с повышенной зеленой флуоресценцией (EGFP) или также с коротким маркерным пептидом, таким как 8 × гистидин, вариант агрина, в котором одна или более, например, одна, две, три или четыре аминокислоты удалены или заменены другими аминокислотами, слитый белок данного варианта агрина, как указано выше в данном контексте, или фрагмент агрина, включающий по меньшей мере 6, в частности по меньшей мере 8 аминокислот агрина, например от 8 до 20 или от 400 до 600 аминокислот агрина, либо как такового, либо слитого с маркерным пептидом или белком и где вариант агрина или фрагмент агрина сохраняет центр расщепления α, центр расщепления β или оба центра расщепления, в особенности, где фрагмент агрина включает консенсусную последовательность центра расщепления α и/или центра расщепления β, как определено ниже в данном контексте. Данный "белок или пептид, включающий агрин, его вариант или фрагмент" могут включать дополнительные непептидные маркеры, например, для спектроскопической детекции, как описано ниже в данном контексте.
В частности, способ измерения каталитической активности нейротрипсина относится к применению нейротрипсина человека, другого млекопитающего или птицы полной длины или домена протеазы нейротрипсина и агрина полной длины или его фрагмента, например инженерного варианта, связанного с мембраной агрина, например, агрин-EGFP последовательности SEQ ID NO:9, или, например, С-концевого фрагмента агрина, агрина-С45 последовательности SEQ ID NO:12.
Особенно предпочтительные условия реакции для измерения каталитической активности нейротрипсина представляют собой буферный раствор с рН 7 - рН 8, включающий ионы Са2+, например, буферный раствор 10 мМ MOPS, рН 7,5 или также 50-100 мМ Трис-HCl, включающий 100-200 мМ NaCl, 1-20 мМ CaCl2, в частности 2-5 мМ CaCl2, и необязательно до 0,5% полиэтиленгликоля, например, полиэтиленгликоля 6000, температуру реакции от 20 до 40°С, например около 25°С, и время реакции от 1 до 48 часов, например, от 2 до 16 часов, например около 3 часов. Нейротрипсин используют при концентрации, которая приводит в результате к расщеплению приблизительно 80% субстрата в течение 3 часов. Предпочтительная концентрация нейротрипсина составляет 0,1-10 нМ, например около 1 нМ, и агрин используют в концентрации, в 10-5000 раз превышающей концентрацию нейротрипсина, например в 1000 раз превышающей данную концентрацию.
Длина полученного из агрина субстрата может варьировать от агрина полной длины до маленького пептида, включающего по меньшей мере один из центров расщепления для нейротрипсина. Нейротрипсин расщепляет агрин в двух различных эволюционно консервативных центрах (см. пример 25). Консенсусная последовательность аминокислот, включающая первый центр расщепления (центр расщепления α), представляет собой …P-P/A-I/V-E-R-A-S/T-C-Y…, где расщепление происходит между остатками R995 и А996. Консенсусная последовательность аминокислот, включающая второй центр расщепления (центр расщепления β) представляет собой …G/A-L/I/T-I/V-E-K-S-V/A-G…, где расщепление происходит между остатками K1754 и S1755.
Анализ измеряет высвобождение фрагмента белка или пептида из субстратного белка или пептида. Если пептид, включающий последовательность аминокислот, непосредственно N-концевую относительно центра расщепления α или β, используют в сочетании с хромогенным или флуорогенным субстратом, который ковалентно связан с С-концом пептида, измеряют высвобождаемый хромогенный или флуорогенный субстрат. Короткие пептидные субстраты, перекрывающие центр расщепления α или β, можно также использовать для измерения протеолитической активности нейротрипсина методом гашения флуоресценции (см. статью Le Bonniec, B.F. et al., Biochemistry 35: 7114-7122, 1996).
Детекцию продуктов расщепления белковых субстратов, включающих центры расщепления α или β, проводят с помощью специфических антител к одному или другому из фрагментов, генерированных посредством протеолитической активности нейротрипсина (см. примеры 22 и 23) или посредством связывания флуоресцентной, хромогенной или другой метки с одним или другим концом агрина, или фрагмента агрина. Для детекции продуктов расщепления применим любой метод детекции белков или пептидов и продуктов их расщепления. Например, расщепление агрина полной длины и более крупных фрагментов агрина (больше, чем приблизительно 10 кД) определяют посредством SDS-PAGE с последующим вестерн-блоттингом, или непосредственной визуализацией в геле с помощью флуоресцентно, или иным образом помеченных белков по появлению фрагментов меньшего размера при расходовании исходного субстрата. Более мелкие фрагменты агрина (меньше, чем приблизительно 10 кД) могут быть иммобилизованы путем связывания с пластиковой поверхностью или частицей, и расщепление визуализируют по солюбилизации фрагмента, который можно либо определить путем специфического распознавания с помощью антитела, либо с помощью флуоресцентного или иным образом определяемого соединения (см. статью Patel, D. et al., BioTechniques 31: 1194-1203, 2001).
Одним из полученных из агрина субстратов для изучения расщепления агрина нейротрипсином в центре расщепления α является, например, инженерный агрин-EGFP, в основе которого лежит трансмембранная форма агрина. Растворимую форму данной молекулы генерируют заменой трансмембранной части последовательностью секретирующегося сигнального пептида человеческого кальсинтенина-1 и меткой 8 × гистидин. Данный белок можно очистить из супернатантов клеточных культур, используя металл-аффинную хроматографию. Для делеции центра расщепления β домен LG3 и петля, связывающая домен EGF4 и домен LG3, включающий β-центр, заменяют EGFP или другим белковым доменом, связанным с коротким линкером (см. пример 20). Используемые рабочие концентрации для анализа активности нейротрипсина составляют от низких наномолярных до микромолярных, например 1 нМ-10 мкМ. Данный белковый субстрат и продукты его расщепления можно измерить с использованием вестерн-блот анализов с помощью антител, определяющих С-концевой продукт расщепления агрина, который генерируют расщеплением в центре расщепления α (см. пример 22) или в анализах ELISA посредством связывания белка через его N-концевую полигистидиновую метку и мониторирования высвобождения С-концевого фрагмента посредством соответствующих антител (см. пример 22) или измерением высвобождающейся флуоресценции группы EGFP в ридере для планшетов.
Другой полученный из агрина субстрат для мониторирования протеолитической активности нейротрипсина состоит из С-концевого фрагмента агрина приблизительно 45 кД, включающего центр расщепления β агрина. Он состоит из доменов LG2-EGF4 и LG3 агрина, включающих центр расщепления β между доменами EGF4 и LG3. Для получения белка в секретируемой форме в клеточных культурах секретируемый сигнальный пептид человеческого кальсинтенина-1 сливают с N-концом, а затем с меткой 8 × гистидин для простоты очистки и связывания с планшетами Ni-NTA. Для детекции и дальнейшей очистки может быть введена С-концевая метка Streptag II. Данный субстрат (см. пример 21) подходит для измерения активности нейротрипсина от низких наномолярных до микромолярных концентраций, например 1 нМ-10 мкМ. Активность нейротрипсина можно мониторировать окрашиванием нерасщепленного субстрата и продуктов расщепления красителями для белков, такими как Кумасси синий или Sypro ruby, либо посредством вестерн-блота, определяющего С-конец продукта расщепления при использовании StrepTactin (IBA, Göttingen) или подходящего антитела, определяющего домен LG3 (см. пример 23). Дополнительные конструкции, включающие С-концевой EGFP или SNAP-Tag (Covalys), меченные подходящими красителями или другими дающими сигнал белками, можно использовать для высокоэффективных анализов в тестах на планшетах, определяющих высвобождение флуоресценции при расщеплении нейротрипсином, путем связывания N-концевой части белкового субстрата с планшетом Ni-NTA или частицами Ni-NTA (см., например, статью Patel, D., BioTechniques 31: 1194-1203, 2001).
Хромогенные протеолитические субстраты, как правило, включают природные или искусственные пептиды, состоящие из 3-5 природных или синтетических аминокислот. Они могут быть защищены по N-концу для снижения нежелательного разложения аминопептидазами. На С-концах они модифицированы так, что хромогенная или флуорогенная группа высвобождается при расщеплении амидной связи. Детекция зависит от типа уходящей группы и может лежать в интервале от УФ- до видимой области света. Другие генерируют флуоресцентный сигнал. Наиболее часто используемыми группами являются п-нитроанилин (pNA), который поглощает свет длины волны 405 нм (см. статью Nail, T.A. et al., J. Biol. Chem. 279: 48535-48542, 2004), и флуорогенный 7-амино-4-метилкумарин (АМС) с длиной волны возбуждения 342 нм и длиной волны эмиссии 440 нм (см. статью Niyomrattanakit, P. et al., J. Virol., 78: 13708-13716, 2004). Для детекции активности нейротрипсина можно использовать короткий трипептид IER-pNA в концентрации 20-50 мкМ в буфере для анализов (150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ 6000, 20 мМ MOPS, pH 7,5) при 25-37°С. При расщеплении нейротрипсином в концентрации 1-5 нМ усиление интенсивности сигнала при 410 нм можно проследить с помощью спектрофотометра.
Субстраты FRET широко используют в протеолитических анализах, поскольку они дают возможность провести однородный и чувствительный анализ, легко адаптируемый для высокоэффективного скрининга (ВСА). Способ особенно подходит для скрининга библиотек органических соединений на конкурентные ингибиторы нейротрипсина в формате высокоэффективного анализа. В анализе FRET пептидный субстрат синтезируют с двумя флуорофорами, флуоресцентным донором (орто-аминобензойная кислота, „Abz") и гасящим акцептором (этилен-диамин-2,4-динитрофенил, „ED-DNP"). Расстояние между данными двумя группами можно выбрать так, чтобы при возбуждении светом акцептор (ED-DNP) в существенной мере гасил энергию флуоресценции донора (Abz) посредством квантового механического феномена, известного как перенос энергии резонанса флуоресценции (FRET), который происходит без испускания света. При расщеплении субстратного пептида протеазой флуорофор отделяется от гасящей группы, восстанавливая полный выход флуоресценции донора. Повышение уровня флуоресценции на коэффициент 7-100 линейно связан с уровнем протеолиза (см. статью Le Bonniec, B.F. et al., Biochemistry 35: 7114-7122, 1996).
Одним из субстратов FRET на основе агрина, используемым для детекции активности нейротрипсина является нонапептид с последовательностью аминокислот Abz-PIERASCY-ED-DNP, включающий нейротрипсиновый центр распознавания агрина α (см. статью Jerini AG, Berlin, Germany). Данный центр расщепления находится между аминокислотами четыре (R) и пять (А) субстрата. При расщеплении пептида при концентрации 1-40 мкМ 1-10 нМ нейротрипсина в буфере для анализа (150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ 6000, 20 мМ MOPS, pH 7,5) при 25-37°С, активность можно определить спектрофотометрически по повышению интенсивности сигнала при длине волны возбуждения 320 нм и длине волны эмиссии 430 нм на флуоресцентном спектрофотометре.
Основой другого субстрата FRET являются два флуоресцентных белка, циан-флуоресцентный белок (CFP) и усиленный желтый флуоресцентный белок (EYFP). Между двумя белками введен линкер из 16 аминокислот, включающий последовательность распознавания нейротрипсина α (PIERASCY) и два спейсера из 4 аминокислот, один ниже и один выше последовательности распознавания нейротрипсина (последовательность линкера: GAGSPIERASCYGSST). Альтернативно можно также использовать соответствующую последовательность аминокислот, фланкирующую центр расщепления β. Расщепление чувствительной линкерной последовательности нейротрипсином разделяет два флуорофора и приводит к потере энергии переноса. Сбор данных осуществляют в буфере для анализа (150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ 6000, 20 мМ MOPS, pH 7,5) при 25-37°C с использованием 1-10 нМ нейротрипсина и 0,1-1 мкМ субстрата. Таким образом гидролиз субстрата можно оценить измерением повышающейся интенсивности флуоресценции донора (эмиссия при 485 нм) и одновременно снижающейся флуоресценции акцептора (эмиссия при 528 нм) после возбуждения при 400-450 нм (см. статью Pollock, В.A. et al., Trends in Cell Biol. 9: 57-60, 1999).
Изобретение, кроме того, относится к способу определения, является ли соединение ингибитором нейротрипсина, отличающемуся тем, что соединение инкубируют вместе с нейротрипсином, в частности человеческим нейротрипсином, его вариантом или фрагментом, включающим домен протеазы, и с белком или пептидом, включающим агрин, его вариант или фрагмент, включающий центр расщепления α или центр расщепления β агрина в водном буферном растворе, и измеряют количество расщепления агрина.
В частности, способ определения, является ли соединение ингибитором нейротрипсина, относится к использованию человеческого нейротрипсина полной длины или домена протеазы человеческого нейротрипсина и агрина полной длины или его фрагмента, например инженерного варианта связанного с мембраной агрина, например агрин-EGFP последовательности SEQ ID NO:9, или, например С-концевого фрагмента агрина, агрин-С45, последовательности SEQ ID NO:12.
Ингибиторы нейротрипсина обнаруживают, используя анализ на протеолитическую активность нейротрипсина (см. пример 24) и тестируя на эффект снижения активности потенциального соединения (см. примеры 26 и 27). Для измерения протеолитической активности нейротрипсина очищенный нейротрипсин (см. примеры 18 и 19) и очищенный агрин или очищенный фрагмент агрина, включающий по меньшей мере один центр расщепления (см. пример 20 и 21), инкубируют совместно с тестируемым соединением в соответствующих условиях. В конце периода инкубирования измеряют продукты расщепления, которые были генерированы с помощью протеолитической активности нейротрипсина. Путем сравнения количества генерированного продукта расщепления в реакциях, включающих потенциальные ингибиторы нейротрипсина, с контрольной реакцией без добавления органических соединений определяют ингибирующие эффекты соединений (см. пример 27). Дозозависимость ингибирующей активности соединения, показанного в качестве ингибитора нейротрипсина, определяют, как описано в примере 28. Специфичность соединения, показанного в качестве ингибитора нейротрипсина, исследуют путем тестирования на ингибирующую активность на других серинпротеазах, как описано в примере 29.
Особенно предпочтительными условиями реакции для определения, является ли соединение ингибитором нейротрипсина, представляют собой буферный раствор с рН около 7, включающий ионы Са2+, например 100-200 мМ NaCl, 5-20 мМ CaCl2, 20 мМ MOPS, рН 7,5 и необязательно до 0,5% полиэтиленгликоля, температура реакции от 20 до 40°С, например около 25°С, и время реакции от 1 до 48 часов, например 3 часа. Предпочтительная концентрация нейротрипсина составляет 0,1-10 нМ, например приблизительно 1 нМ, и агрин используют в концентрации в 10-5000 раз больше концентрации нейротрипсина, например в 1000 раз больше данной концентрации. Тестируемое соединение добавляют в возрастающих концентрациях, предпочтительно в концентрациях от 0,001 до 500 мкМ. Можно добавить до 30% ДМСО (диметилсульфоксида) с целью повышения растворимости тестируемых соединений.
Далее изобретение относится к способам детекции активности нейротрипсина и ингибирующего эффекта на нейротрипсин маленьких молекул органических соединений в высокоэффективных системах анализа (ВСА) так что нейротрипсин, его вариант или фрагмент, включающий домен протеазы нейротрипсина и белок или пептид, включающий агрин, его вариант или фрагмент, включающий последовательность центра расщепления α или центра расщепления β агрина, или любой другой белок, включающий последовательность, гомологичную последовательности центра расщепления α или β агрина, инкубируют совместно в водном буферном растворе и измеряют количество продукта расщепления в способах, подходящих для ВСА.
В частности, все вышеописанные способы, относящиеся к способам измерения каталитической активности нейротрипсина, можно также использовать в мультипланшетных анализах или дот-блотах с подходящими методами считывания ВСА.
Субстраты в виде маленьких пептидов, включающие либо центр расщепления α, либо центр расщепления β с С-концевыми хромогенными или флуорогенными уходящими группами, можно легко считывать в мультипланшетном ридере посредством измерения повышения уровня флуоресценции или поглощения при соответствующих длинах волн. Данный способ применим также к белковым субстратам с N-концевой или С-концевой аффинными метками, например полигистидиновой меткой или Streptag II либо белковой метками, и слитыми по С-концу или N-концу дающими сигнал белками или белковыми доменами, например флуоресцентными белками или белками, которые могут быть помечены хромофорами, или флуорофорами, или даже ферментами, катализирующими хромогенные или флуорогенные реакции, такими как β-галактозидаза или другие, путем фиксации части белка на поверхности лунки и детекции генерации флуоресценции или поглощения либо ферментной активности в супернатанте лунки при соответствующих длинах волны. Генерацию продуктов расщепления при активности нейротрипсина в ВСА можно также мониторировать в вариантах ELISA (твердофазного иммуноферментного анализа) путем детекции количества продуктов расщепления в супернатанте лунок, покрытых белковыми субстратами, подходящими для расщепления нейротрипсином, или остаточного количества нерасщепленного субстрата с помощью подходящих антител, связанных с ферментами или группами, дающими сигнал (см. статью Gutierrez, О.A. et al., Anal. Biochem. 307: 18-24, 2002).
Изобретение, кроме того, относится к другим непланшетным анализам, подходящим для подобной ВСА детекции продуктов расщепления субстратов на основе маленьких пептидов или белковых субстратов любого описанного типа методами, подобными ВЭЖХ (высокоэффективной жидкостной хроматографии) (см. статью Betageri, R. et al., J. Biochem. Biophys. Methods 27: 191-197, 1993), FPLC (жидкостной экспресс-хроматографии), масс-спектрометрии (см. статью Mathur, S. et al., J. Biomol. Screen. 8: 136-148, 2003), SELDI (Cyphergen) или другим подобным вариантам.
Дополнительно изобретение относится к ингибиторам нейротрипсина, обнаруженным данным способом, в частности, к соединениям формулы
в которой Hal1 и Hal2 независимо друг от друга представляют собой фтор, хлор или бром, в особенности бром, и их фармацевтически приемлемые аддитивные соли. Предпочтительными также являются соединения, в которых одно из Hal1 и Hal2 представляет собой бром и другое представляет собой хлор или фтор, и их фармацевтически приемлемые аддитивные соли. Особенно предпочтительными являются фармацевтически приемлемые аддитивные соли соединения формулы (1), в котором Hal1 и Hal2 представляют собой бром.
Данные фармацевтически приемлемые соли образуются, например, с органическими или неорганическими кислотами. Подходящими неорганическими кислотами являются, например, галогеновые кислоты, такие как хлористоводородная кислота, серная кислота или фосфорная кислота. Подходящими органическими кислотами являются, например, карбоновая, фосфоновая, сульфоновая или сульфаминовая кислоты, например уксусная кислота, пропионовая кислота, октановая кислота, декановая кислота, додекановая кислота, гликолевая кислота, молочная кислота, фумаровая кислота, янтарная кислота, адипиновая кислота, пимелиновая кислота, убериновая кислота, азелаиновая кислота, яблочная кислота, винная кислота, лимонная кислота, аминокислоты, такие как глутаминовая кислота или аспарагиновая кислота, малеиновая кислота, гидроксималеиновая кислота, метилмалеиновая кислота, циклогексанкарбоновая кислота, адаментанкарбоновая кислота, бензойная кислота, салициловая кислота, 4-аминосалициловая кислота, фталевая кислота, фенилуксусная кислота, миндальная кислота, коричная кислота, метан- или этансульфоновая кислота, 2-гидроксиэтансульфоновая кислота, этан-1,2-дисульфоновая кислота, бензолсульфоновая кислота, 2-нафталинсульфоновая кислота, 1,5-нафталин-дисульфоновая кислота, 2-, 3- или 4-метилбензолсульфоновая кислота, метилсерная кислота, этилсерная кислота, додецилсерная кислота, N-циклогексилсульфаминовая кислота, N-метил-, N-этил- или N-пропил-сульфаминовая кислота или другие протонные кислоты, такие как аскорбиновая кислота.
Для целей выделения и очистки можно также использовать фармацевтически неприемлемые соли, например пикраты или перхлораты. Для терапевтического применения используют только фармацевтически приемлемые соли или свободные соединения (если применимо, в форме фармацевтических препаратов), и, вследствие этого, они предпочтительны.
Соединения формулы (1) получают методами, известными в области техники, например, путем конденсации амина формулы (2) или его предшественника, в котором амидиновая функция находится в защищенной форме или присутствует как функциональная группа, легко превращающаяся в амидиновую функцию, с альдегидом формулы (3), и необязательную защитную группу отщепляют после реакции конденсации либо функциональную группу превращают в амидиновую функцию.
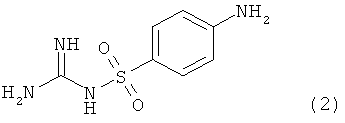
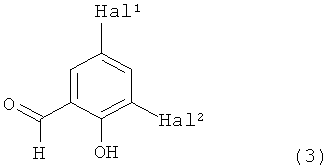
В частности, изобретение относится к фармацевтическим композициям, включающим соединения формулы (1), как описано выше в данном контексте, и к применению соединений формулы (1) в качестве лекарственных средств.
Настоящее изобретение относится также к фармацевтическим композициям, которые включают соединение формулы (1) в качестве активного ингредиента и которые могут быть использованы, в частности, при лечении упомянутых заболеваний. Предпочтительными являются композиции для энтерального введения, такого как назальное, защечное, ректальное или, в особенности, пероральное введение, и для парентерального введения, такого как внутривенное, внутримышечное или подкожное введение, теплокровным животным, особенно человеку. Композиции включают активный ингредиент в виде монокомпонента или, предпочтительно, совместно с фармацевтически приемлемым носителем. Доза активного ингредиента зависит от заболевания, которое лечат, и биологического вида пациента, его возраста, массы и индивидуального состояния, индивидуальных фармакокинетических данных и способа введения.
Фармацевтические композиции включают от приблизительно 1% до приблизительно 95% активного ингредиента и представляют собой, например, покрытые и непокрытые таблетки, ампулы, флаконы, суппозитории или мази, кремы, пасты, пены, настойки, капли, спреи, дисперсии и т.п. Примерами являются капсулы, включающие от приблизительно 0,05 г до приблизительно 1,0 г активного ингредиента. Фармацевтические композиции, представленные в настоящем изобретении, получают методом, известным, как таковой, например, посредством традиционных процессов смешивания, грануляции, покрытия, растворения или лиофилизации.
Предпочтение отдают применению растворов активного ингредиента, а также суспензий или дисперсий, особенно изотонических водных растворов, дисперсий или суспензий, которые, например, в случае лиофилизированных композиций, включающих активный ингредиент в виде монокомпонента или вместе с носителем, например маннитом, могут быть приготовлены перед применением. Фармацевтические композиции могут быть стерилизованными и/или могут включать наполнители, например, консерванты, стабилизаторы, смачивающие агенты и/или эмульгаторы, солюбилизаторы, соли для регуляции осмотического давления и/или буферы, и их получают способом, известным как таковой, например, посредством традиционных процессов растворения и лиофилизации. Указанные растворы или суспензии могут включать агенты, повышающие вязкость, как правило, натрий карбоксиметилцеллюлоза, карбоксиметилцеллюлоза, декстран, поливинилпирролидон или желатины либо также солюбилизаторы, например Твин 80® (полиоксиэтилен(20)сорбитанмоноолеат).
Суспензии в масле включают в качестве масляного компонента растительное, синтетическое или полусинтетическое масла, принятые для инъекционных целей.
Подходящие носители, в частности, представляют собой наполнители, такие как сахара, например лактозу, сахарозу, маннит или сорбит, препараты целлюлозы и/или фосфаты кальция, например трикальцийфосфат или гидроксифосфат кальция, а также связующие агенты, такие как крахмалы, например, кукурузный, пшеничный, рисовый или картофельный крахмал, метилцеллюлоза, гидроксипропилметилцеллюлоза, натрий карбоксиметилцеллюлоза и/или поливинилпирролидон и/или, при необходимости, разрыхлители, такие как вышеупомянутые крахмалы, а также карбоксиметилкрахмал, поперечно-сшитый поливинилпирролидон, альгиновая кислота или ее соль, такая как альгинат натрия. Дополнительными наполнителями являются, в частности, кондиционеры сыпучести и смазывающие компоненты, например кремневая кислота, тальк, стеариновая кислота или ее соли, такие как стеарат магния или кальция, и/или полиэтиленгликоль или его производные.
Ядра таблеток могут быть представлены с подходящими, необязательно энтеросолюбильным, покрытиями, известными в области техники.
Фармацевтические композиции для перорального применения включают также твердые капсулы, состоящие из желатина и пластификатора, такого как глицерин или сорбит. Фармацевтические композиции, пригодные для ректального применения, представляют собой, например, суппозитории, которые состоят из комбинации активного ингредиента и основы суппозитория. Подходящими основами суппозиториев являются, например, природные или синтетические триглицериды, предельные углеводороды, полиэтиленгликоли или высшие спирты.
Примеры
Пример 1: Нейротрипсин в сильной степени экспрессируется двигательными нейронами спинного мозга.
Характер гибридизации in situ на поперечных срезах замороженных тканей спинного мозга взрослых мышей показывает сильную клеточную экспрессию мРНК нейротрипсина в сером веществе (см. статью Gschwend, Т.Р., et al., Molec. Cell Neurosci. 9: 207-219, 1997). Самую сильную экспрессию нейротрипсина в спинном мозге обнаруживают в двигательных нейронах в вентральном роге серого вещества.
Для иммуногистохимической локализации белка нейротрипсина в спинном мозге генерируют антитела к каталитическому домену нейротрипсина. Для генерации антигена с целью иммунизации каталитический домен человеческого нейротрипсина, включающий His-метку на С-конце, получают в Е.coli, очищают на колонке Ni-NTA и переукладывают. Для иммунизации козы используют порции по 50 мкг (первичные иммунизации в полных адъювантах Фрейнда и бустерные инъекции в неполных адъювантах Фрейнда). Из иммунной сыворотки IgG выделяют аффинной хроматографией на иммобилизованном белке G. Аффинно очищенный IgG получают аффинной хроматографией на иммобилизованном домене нейротрипсина.
Взрослым (в возрасте 2 месяцев) мышам проводят глубокую анестезию пентобарбиталом натрия (80 мг/кг, Abbot) и транскардиальную перфузию 10 мл 0,9% NaCl, а затем 150 мл фиксирующего раствора, включающего 4% параформальдегида (Merck), 0,05% глутаральдегида (Merck) и 0,2% пикриновой кислоты (BDH) в 0,1 М фосфатном буфере, рН 7,4 (РВ). Венечные срезы головного мозга делают на вибрационном микротоме толщиной 60 мкм. Для усиления проникновения иммунореагентов срезы уравновешивают в 30% сахарозе в РВ, быстро замораживают в жидком азоте и затем оттаивают в РВ и предварительно обрабатывают 0,3% Тритон-Х100 в течение 10 минут. После предварительного инкубирования в 20% нормальной козьей сыворотке (NGS; Vector Labs, Servion, Switzerland) в 0,05 М Трис-забуференном солевом растворе (TBS; рН 7,4) в течение 45 минут при комнатной температуре (КТ), срезы инкубируют в течение 36-48 час при 4°C с первичным антителом к нейротрипсину (1 мкг/мл). Для световой микроскопии иммунопероксидазы срезы инкубируют в биотинилированном козьем IgG к кролику (1:200, Vector Labs) в течение 12 час при 4°C с последующим инкубированием в течение 3 час в комплексе авидин-биотин-пероксидаза (Elite ABC; 1:100, Vector Labs) при КТ. Антигенные центры визуализируют инкубированием в 3,3'-диаминобензидине (Sigma; 0,05% в TBS, рН 7,6) в присутствии 0,004% Н2О2. Срезы монтируют на желатинизированных предметных стеклах, сушат на воздухе, обезвоживают и закрывают покровными стеклами в Entelan (Merck).
Характер иммуногистохимического окрашивания, определяемый таким образом на поперечных срезах спинного мозга взрослых мышей, показывает сильные сигналы присутствия нейротрипсина в сером веществе. Самый высокий уровень экспрессии обнаруживают в двигательных нейронах вентрального рога. Контрольные срезы, обработанные тем же способом, но при исключении первого или второго антитела, не показывают никакого окрашивания.
В заключение, данные эксперименты ясно демонстрируют, что нейротрипсин сильно экспрессируется в сером веществе спинного мозга. Особенно сильную экспрессию обнаруживают в двигательных нейронах, которые находятся в вентральном роге и иннервируют скелетные мышцы.
Пример 2: Нейротрипсин расщепляет агрин.
Протеолитический эффект нейротрипсина в отношении агрина тестируют путем соэкспрессии двух белков в клетках HEK293T. Фрагмент KpnI-HindIII из 2310 пар осн., включающий кодирующую последовательность мышиного нейротрипсина, клонируют в эукариотический экспрессирующий вектор pcDNA3.1(-) (Invitrogen) посредством KpnI и HindIII. Клон кДНК, кодирующий крысиный агрин (см. статью Rupp, F. et al., Neuron 6: 811-823, 1991; GenBank Nr. M64780), состоящий из трансмембранной изоформы, включающей сплайс-варианты Y4 и Z8, вводят в полилинкер pcDNA3 (Invitrogen) посредством KpnI и EcoRI. Клетки HEK293T культивируют в среде DMEM/10% FCS (сыворотка телячьих эмбрионов) при 37°С в увлажненном воздухе с 10% СО2. Для трансфекции клетки высевают в 3 мл среды DMEM/10% FCS на стеклянные покровные стекла, помещенные в чашки диаметром 3 см. На следующий день после посева при конфлюэнтности 40-60% клетки временно трансфицируют кДНК, кодирующими нейротрипсин и агрин (5 мкл каждой ДНК), используя преципитацию фосфатом кальция. Через 4 часа после трансфекции среду тщательно удаляют и заменяют 3 мл свежей среды DMEM/10% FCS.
Метаболический путь агрина при соэкспрессии с нейротрипсином анализируют вестерн-блоттингом. Через 48 часов после трансфекции клетки промывают PBS и лизируют добавлением 250 мкл буфера (20 мМ Трис-HCl, рН 7,4, 150 мМ NaCl, 1 мМ EDTA (этилендиаминтетрауксусная кислота), 1% Тритон Х-100, коктейль ингибиторов протеазы). Экстракт инкубируют при 4°С в течение 20 минут и затем центрифугируют в течение 20 минут с 15000×g при 4°С. Собирают супернатант. После определения концентрации белка супернатант смешивают с 5 × буфер для нагрузки Laemmli, кипятят в течение 3 минут, центрифугируют и используют для анализа. Белки разделяют SDS-PAGE, используя 7,5% акриламид. После электрофореза белки переносят на нитроцеллюлозную мембрану. Качество переноса проверяют окрашиванием Ponceau S. Затем мембрану блокируют TBS, включающим 0,1% Твин-20 и 5% (масс./об.) блокирующий реагент (Amersham). Все последующие стадии осуществляют в TBS с 0,1% Твин-20. Мембрану инкубируют с первичным антителом (SZ177, 1:1000; AGR540, 1:1000; K-17, поликлональное антитело к агрину (Santa Cruz), 1:1000) в течение 60 минут. После сильного промывания мембрану инкубируют с вторичными связанными с пероксидазой антителами в течение 45 минут. Детекцию проводят с использованием ChemiGlow (Alpha Innotech) в соответствии с инструкцией производителя. Изображения получают с помощью ChemiImager (Alpha Innotech).
Агрин ясно идентифицируют в детергентных экстрактах одинарных трансфектантов (см. фиг.2, дорожка 1). В экстрактах двойных трансфектантов уровень агрина сильно понижен (см. фиг.2, дорожка 2). Никакого сигнала агрина не обнаруживают в клетках, трансфицированных ненагруженным вектором. Продукцию нейротрипсина во всех условиях подтверждают после повторного зондирования блота антителами к нейротрипсину. Анализ культуральрной среды данных различных условий показывает, что иммунореактивность, которая утрачена в клеточном экстракте дважды трансфицированных клеток, высвобождается в среду супернатанта. В 200 мкл культуральной среды дважды трансфицированных клеток HEK293T определяют полосу 100 кД с помощью антител к агрину (см. фиг.2, дорожка 3). Данный сигнал не обнаруживают в среде одинарных трансфектантов (см. фиг.2, дорожка 4). Аналогично никакой сигнал не определяют в среде клеток HEK293T, трансфицированных агрином и каталитически неактивным нейротрипсином.
Подводя итог, результаты показывают, что (1) нейротрипсин, продуцируемый в клетках HEK293T, имеет каталитическую активность, что (2) агрин, внеклеточно присутствующий компонент нейромышечного соединения и синапсов центральной нервной системы, можно расщепить путем нейротрипсин-зависимого протеолиза, и (3) что данный нейротрипсин-зависимый протеолиз приводит к формированию укороченной и высвобождающейся формы агрина. Высвобождающаяся часть агрина имеет приблизительную молекулярную массу приблизительно 100 кД. Поскольку антитела, используемые для детекции агрина, направлены на эпитопы в С-концевой части агрина, солюбилизированный фрагмент включает С-концевую сторону агрина.
Пример 3: Расщепление агрина in vivo происходит одновременно с экспрессией нейротрипсина в спинном мозге в период развития нейронов.
Для тестирования расщепления агрина in vivo гомогенаты спинного мозга растущих и взрослых мышей анализируют вестерн-блотом, используя специфические антитела к Nt и С-концевому фрагменту 100 кД агрина. Гомогенаты тканей получают из спинного мозга мышей в постнатальные дни 4, 7, 9, 10, 15 и 25, а также старых мышей в возрасте 3, 6 и 12 месяцев. Как демонстрируют на фиг.3, Nt наиболее сильно экспрессируется в первые три постнатальные недели при наиболее высоких уровнях экспрессии между днями 7 и 10. Вестерн-блоттинг с антителами к С-концевой группе агрина показывает в большой степени близкий временной характер, указывающий на то, что расщепление агрина нейротрипсином также имеет место in vivo.
Пример 4: Избирательная сверхэкспрессия нейротрипсина в двигательных нейронах с использованием технологии с трансгенными мышами.
Сверхэкспрессия нейротрипсина происходит под контролем промотора гена Thy-1. Ген Thy-1 экспрессируется в нейронах нервной системы мыши относительно поздно (постнатальный день 4-10, в зависимости от положения; см. статью Gordon, J.W. et al., Cell 50: 445-452, 1987). Вследствие этого экспрессия нейротрипсина под контролем промотора Thy-1 обеспечивает то, что присутствие избыточных количеств нейротрипсина не нарушает более ранние стадии развития.
Генерируют мышей, способных к сверхэкспрессии нейротрипсина, с условным трансгеном, который требует активации. Для данной цели удаляемую транскрипционную стоп-последовательность, фланкированную сайтами loxP, интродуцируют перед кДНК нейротрипсина. Таким образом, стоп-последовательность можно было бы удалить с помощью рекомбиназы Cre/системы рекомбинации loxP (см. статью Sauer В. et al., Proc. Natl. Acad. Sci. (USA) 85: 5166-5170, 1988). Белок Cre (Cre-рекомбиназа) кодируется бактериофагом P1 E.coli и эффективно стимулирует как внутри-, так и межмолекулярную рекомбинацию ДНК. Рекомбинация происходит в специфическом сайте, называемом loxP (см. статью Hamilton, D.L. и Abremski, K., J. Mol. Biol. 178: 481-486, 1984). Данный характерный признак Cre-рекомбиназы обеспечивает возможность делеции и инсерции специально отмеченных нитей ДНК между последовательностями loxP. Его можно использовать, чтобы генерировать специфические функциональные мутации in vivo (см. статью Chen S. et al., Cell 51:7-19, 1987).
Мышей, несущих конструкцию для условной сверхэкспрессии нейротрипсина, скрещивают с гетерозиготными мышами, несущими ДНК Cre-рекомбиназы в гене фактора транскрипции НВ9, с целью генерации мышей со сверхэкспрессией активного нейротрипсина в двигательных нейронах. Промотор НВ9 активен in vivo при спецификации двигательных нейронов, и, таким образом, все двигательные нейроны экспрессируют направляемую НВ9 Cre-рекомбиназу, приводя в результате к удалению транскрипционной стоп-последовательности из неактивного трансгена.
Трансгенных мышей генотипируют с помощью ПЦР (полимеразной цепной реакции). ДНК для ПЦР экстрагируют из хвоста мышей. Положение ПЦР-праймеров выбирают так, чтобы препятствовать детекции нативного мышиного гена нейротрипсина. 3'-Праймер соответствует последовательности ДНК в промоторе Thy-1 и 5'-праймер - последовательности в кДНК нейротрипсина. Данный фрагмент ДНК является уникальным для трансгена нейротрипсина. Оба праймера для детекции инсерта Cre эквивалентны последовательностям ДНК, выделенным из гена Cre, поскольку Cre обычно не присутствует у мышей. Данным способом получают три мышиных линии со сверхэкспрессией человеческого нейротрипсина и четыре линии со сверхэкспрессией мышиного нейротрипсина. Экспрессию трансгена проверяют на уровне мРНК нозерн-блотингом и гибридизацией in situ и на уровне белка - вестерн-блоттингом. Типичная сверхэкспрессия составляет порядка 2-10×.
Зависимость опосредованного нейротрипсином изменения от его каталитического домена проверяют, генерируя трансгенных мышей со сверхэкспрессией каталитически неактивной формы нейротрипсина под контролем того же промотора, т.е. Thy-1. Неактивный нейротрипсин легко можно генерировать путем мутации серина 711 основного активного центра (соответствующего серину 195 химотрипсина) в аланин. Поскольку во всех серинпротеазах серин активного центра участвует в ковалентном промежуточном продукте протеолитической реакции, его мутация приводит в результате к полной потере каталитической функции. Трансгенные мыши со сверхэкспрессией неактивной формы нейротрипсина здоровы и не проявляют каких-либо патологий.
Пример 5: Трансгенная сверхэкспрессия нейротрипсина повышает уровень расщепления агрина in vivo.
Гомогенаты тканей получают из спинного мозга взрослых мышей со сверхэкспрессией либо человеческого нейротрипсина (hNt), либо мышиного нейротрипсина (mNt). Данных мышей получают скрещиванием мышей с неактивными трансгенами (линии 497, 489 и 533 для mNt и линии 493 и 494 для hNt) с мышами, экспрессирующими рекомбиназу Cre под контролем промотора НВ9, специфического в отношении двигательных нейронов. В качестве контроля используют мышей дикого типа. Гомогенаты спинного мозга подвергают SDS-PAGE и вестерн-блоттингу. Вестерн-блоты зондируют антителами к hNt и mNt, а также к С-концевому фрагменту агрина 100 кД. Как показано на фиг.4, уровень С-концевого фрагмента агрина 100 кД сильно повышен у трансгенных мышей со сверхэкспрессией нейротрипсина. Увеличение количества С-концевого фрагмента агрина 100 кД хорошо коррелирует с уровнем сверхэкспрессии в различных линиях трансгенных мышей. Человеческий и мышиный Nt демонстрируют одинаковый протеолитический эффект в отношении агрина. Данные результаты показывают, что нейротрипсин расщепляет агрин in vivo зависимым от концентрации образом.
Пример 6: Трансгенная сверхэкспрессия нейротрипсина в двигательных нейронах приводит в результате к удалению агрина из НМС в период, исчисляемый часами-днями.
Эффект трансгенной сверхэкспрессии нейротрипсина (Nt) в спинальных двигательных нейронах исследуют иммуногистологическими анализами нейромышечного соединения (НМС). Особый интерес представляет первая постнатальная неделя, поскольку известно, что промотор Thy-1, используемый, чтобы направлять экспрессию трансгенного Nt, становится активным во время первой постнатальной недели (с некоторыми вариациями в зависимости от положения). Для данного анализа используют диафрагму, поскольку поверхностная локализация HMC делает данную мышцу хорошей моделью для данных сравнительных анализов. Визуализацию агрина осуществляют иммуногистологическим окрашиванием с использованием аффинно очищенных антител к С-концевому фрагменту агрина 100 кД. Как продемонстрировано на фиг.5, иммунореактивность агрина в Р0, точке времени перед началом активации трансгена, идентична у мышей дикого типа и трансгенных мышей (см. фиг.5: Р0). В обоих случаях иммунореактивность агрина бесспорно соответствует сигналам α-Btx HMC, показанного на фиг.6 для тех же мышц. В Р8 обнаруживают резкое снижение иммунореактивности агрина в HMC диафрагмы мышей со сверхэкспрессией Nt по сравнению с мышами дикого типа (см. фиг.5: Р8). Конечная полоса, которая плотно насыщена агрин-положительными HMC у соответствующих по возрасту мышей дикого типа, показывает только очень немного крупных подобных HMC агрин-положительных структур. Иммунореактивность агрина мышей Р4 показывает переходную стадию, характеризующуюся смешанным характером со структурами различного размера, варьирующими от вполне консервативных HMC до мелких структур, отражающие остатки HMC (см. фиг.5: Р4). Большинство иммунореактивных структур агрина на фиг.5 точно соответствуют α-Btx-положительным структурам, приведенным на фиг.6. Однако в существенной части синапсов переходной стадии мышей со сверхэкспрессией Nt соотношение между сигналом агрина и сигналом α-Btx меньше, чем в Р0 (см. пример 7).
Данные результаты демонстрируют, что нейротрипсин расщепляет агрин in vivo в HMC и что С-концевая часть агрина исчезает из HMC в период, исчисляемый часами-днями, после нейротрипсин-зависимого расщепления. Это особенно примечательно, поскольку С-концевая часть включает домен, ответственный за просинаптическую активность агрина.
Пример 7: Индуцированное нейротрипсином удаление агрина из HMC приводит в результате к разрушению HMC в период от часов до дней.
Эффект сверхэкспрессии нейротрипсина (Nt) в двигательных нейронах и последующее удаление агрина из HMC исследуют путем визуализации постсинаптического аппарата. Визуализацию постсинаптического аппарата осуществляют окрашиванием ацетилхолиновых рецепторов флуоресцентным α-бунгаротоксином (α-Btx). Как показано на фиг.6, при сравнении мышей со сверхэкспрессией Nt (Nt) с мышами дикого типа (wt), постсинаптический аппарат хорошо развит при рождении (постнатальный день 0, Р0). В конце первой постнатальной недели (Р8) большинство постсинаптических аппаратов НМС фактически исчезают у мышей со сверхэкспрессией Nt. Можно выявить только несколько остаточных НМС в так называемой конечной полосе, тогда как НМС обнаруживают с высокой плотностью у мышей дикого типа. В день 4 постнатального развития (Р4) обнаруживают гетерогенный характер, включающий смесь все еще хорошо сформированных НМС и частично рассосавшихся НМС. Уменьшение плотности НМС в конечной полосе трансгенных мышей по сравнению с мышами дикого типа позволяет предположить, что на данной стадии часть НМС уже полностью исчезла у мышей со сверхэкспрессией Nt.
НМС переходного состояния характеризуются отсутствием или по меньшей мере сильно уменьшенным присутствием агрина в α-Btx-декорированной структуре. Данные НМС переходного состояния обнаруживают исключительно у мышей со сверхэкспрессией Nt. У мышей дикого типа в противоположность мышам со сверхэкспрессией Nt иммуноокрашивание на агрин всегда хорошо соответствует окрашиванию α-Btx ацетилхолиновых рецепторов постсинаптического аппарата.
В заключение, данные эксперименты демонстрируют, что повышенный уровень экспрессии Nt в двигательных нейронах приводит в результате к рассасыванию уже сформированных НМС. Рассасывание НМС следует за трансгенной повышающей регуляцией Nt с короткой задержкой (оцениваемой самое большее как период от часов до нескольких дней). Анализ синапсов переходного состояния демонстрирует, что рассасывание постсинаптического аппарата следует за расщеплением агрина и удалением С-концевого фрагмента агрина 100 кД из НМС. Это показывает, что Nt играет антисинаптическую роль в НМС посредством противодействия просинаптической роли агрина. Если антисинаптическая функция в двигательных нейронах избыточно повышена посредством сильной сверхэкспрессии нейротрипсина, просинаптический агент агрин подавляется и, таким образом, НМС разрушается.
Пример 8: Взрослые мыши с повышенной экспрессией нейротрипсина в двигательных нейронах спинного мозга проявляют выраженный нейромышечный фенотип с пониженной мышечной силой.
Трансгенных мышей, которые способны к сверхэкспрессии Nt в двигательных нейронах, генерируют скрещиванием мышей, несущих условный трансген Nt (описаны в примере 4), с мышами, экспрессирующими рекомбиназу Cre под контролем промотора НВ9. Промотор НВ9 направляет сверхэкспрессию рекомбиназы Cre в спинальных двигательных нейронах и, таким образом, активирует неактивный трансген Thy1-Nt в двигательных нейронах путем эксцизии транскрипционного стоп-сегмента. Дважды трансгенные мыши, полученные при данных скрещиваниях, проявляют двигательный фенотип. Они передвигаются медленно, неуверенной походкой и маленькими шагами. Они также демонстрируют значительно пониженную мышечную силу.
В заключение, сверхэкспрессия Nt в двигательных нейронах дает в результате периферический двигательный фенотип, характеризующийся главным образом пониженной силой скелетных мышц.
Пример 9: Нейромышечные соединения взрослых мышей с повышенной экспрессией нейротрипсина проявляют выраженную фрагментацию пре- и постсинаптического аппарата.
Сравнение нейромышечных соединений трансгенных мышей со сверхэкспрессией Nt и мышей дикого типа одного и того же возраста (молодые взрослые) показывает заметную фрагментацию НМС мышей со сверхэкспрессией Nt. Типичная структура Прецела НМС, которая развивается в первые три постнатальные недели у мышей дикого типа, не обнаружена у мышей со сверхэкспрессией Nt (см. фиг.7D, Е и F). Хотя НМС мышей со сверхэкспрессией Nt занимает приблизительно такую же площадь на поверхности их целевых мышечных волокон, их постсинаптические, а также пресинаптические контакты не образуют прилегающую структуру, но она разорвана на многочисленные маленькие центры контакта. Фрагментацию НМС, наблюдаемую у мышей со сверхэкспрессией Nt, обнаруживают также в НМС пожилых людей, страдающих саркопенией, формой мышечной атрофии, обнаруживаемой у человека в пожилом возрасте.
В заключение, повышенная продукция нейротрипсина в двигательных нейронах приводит в результате к фрагментации НМС, которая близко напоминает фрагментацию НМС, которая описана в исследованиях людей, страдающих саркопенией.
Пример 10: Мышцы взрослых мышей с повышенной экспрессией нейротрипсина имеют значительно уменьшенное число мышечных волокон.
Мышцы молодых взрослых мышей со сверхэкспрессией Nt в двигательных нейронах анализируют относительно числа мышечных волокон. Отдельные мышцы, например камбаловидную мышцу, выделяют и делают среды ткани перпендикулярно длинной оси мышцы через средний сегмент мышцы. Срез окрашивают гематоксилин-эозином и подсчитывают волокна. Фиг.8 показывает сравнение камбаловидной мышцы мыши дикого типа (см. фиг.8,А) и мыши со сверхэкспрессией Nt (см. фиг.8,В). Мышца мыши со сверхэкспрессией Nt значительно тоньше, чем мышца мыши дикого типа, и имеет заметно более низкое число мышечных волокон. Число мышечных волокон подсчитывают в камбаловидной мышце четырех различных линий мышей со сверхэкспрессией Nt (см. табл.1).
Результаты показывают заметное уменьшение числа волокон у мышей со сверхэкспрессией Nt всех четырех независимых линий трансгенных мышей.
В заключение, количественная оценка чисел волокон у мышей демонстрирует, что повышенный уровень Nt в двигательных нейронах приводит в результате к существенному уменьшению числа мышечных волокон.
Пример 11: Сверхэкспрессия нейротрипсина в нейронах ЦНС трансгенных мышей.
Согласно способу, представленному в примере 4, получают трансгенных мышей для условной сверхэкспрессии нейротрипсина. Данных мышей скрещивают с гетерозиготными мышами, экспрессирующим Cre-рекомбиназу под контролем промотора цитомегаловируса (ЦМВ). Промотор ЦМВ постоянно активен in vivo, и, следовательно, Cre-рекомбиназа способствует рекомбинации в двух последовательностях loxP во все периоды времени. Данный способ удаляет транскрипционную стоп-последовательность из неактивного трансгена и обеспечивает транскрипцию кДНК нейротрипсина. Генотипирование с помощью ПЦР и гибридизацию с помощью саузерн-блота проводят, как описано в примере 4.
Аналогично трансгенных мышей со сверхэкспрессией каталитически неактивной формы нейротрипсина под контролем промотора Thy-1 генерируют посредством мутации серина активного центра (серина 711) нейротрипсина в аланин.
Пример 12: Повышенные уровни нейротрипсина в нейронах ЦНС приводят в результате к уменьшению числа синапсов.
Для количественной оценки числа синапсов/объем ткани богатого синапсами участка и для измерения размерных параметров синапсов, включая площадь пресинаптических окончаний аксонов, площадь постсинаптических шипиков и длину синапсов (как измеряют по длине присоединения пре- и постсинаптических мембран) исследуют две независимые линии мышей со сверхэкспрессией нейротрипсина (Nt491/cre и Nt494/cre) и несколько линий контрольных мышей (мыши дикого типа, мыши ЦМВ-Cre и трансгенные родительские линии, несущие неактивный трансген нейротрипсина Nt491-инакт.Nt и №494-инакт.Nt). Мышам в возрасте 28 дней проводят глубокую анестезию метиофаном (Schering-Plough) и перфузируют через сердце 0,9% хлоридом натрия, а затем фиксирующим агентом, состоящим из 2% параформальдегида и 1% глутаральдегида в 0,1 М фосфатном буфере (РВ), рН 7,4. Головной мозг удаляют из черепа и делают серийные срезы толщиной 100 мкм с помощью вибратом. Срезы затем фиксируют в 1% тетраоксиде осмия РВ, обрабатывают 2% уранилацетатом, обезвоживают в этаноле и пропиленоксиде и заключают в смолу Durcupan ACM (Fluka). Для электронно-микроскопического анализа из срезов, включающих участок СА1 гиппокампа на антериокаудальном уровне Брегма -2 мм и медиолатерально 1,5 мм, делают ультрасрезы. Взятие образцов синапсов состоит из получения 15-23 образцов ЕМ нейропиля радиального слоя участка СА1 гиппокампа из трех несоседних областей, отстоящих друг от друга на расстояние по меньшей мере 50 мкм при исходном увеличении в 27500 раз. Электронные микрофотографии печатают при конечном увеличении в 80000 раз, что представляет 90-135 мкм2 ткани. Синапс определяют как две находящиеся друг перед другом утолщенные мембраны пресинаптического и постсинаптического профиля, причем пресинаптический профиль включает по меньшей мере три синаптических пузырька с тесной ассоциации с дифференцированными мембранами. Синапсы разделяют на аксодендритические и аксоспинические синапсы согласно ультраструктурным критериям. Дендритные стволы идентифицируют по их размеру и присутствию митохондрии и микротрубочек. Дендритные шипики имеют меньший диаметр, в них отсутствуют митохондрии и микротрубочки и они иногда включают аппарат шипика. Аксодендритические синапсы составляют несущественно малую долю во всех образцах и, вследствие этого, исключены из дальнейшей статистической оценки. Все аксоспинические синапсы подсчитывают на каждой микрофотографии за исключением затрагивающих ограничительные линии. Площади поперечных срезов окончаний аксонов и постсинаптических шипиков и длины синаптических соединений всех аксоспинических синапсов измеряют непосредственно на отпечатках, используя магнитный блокнот (Kurta) и программу анализа Macstereology 2.8 (Ranfurly Microsystems). Количественную плотность синапсов получают, используя метод частоты размера и формулу NV=NA/d (в которой NA означает число синаптических профилей/единицу площади и d означает среднюю длину синаптических соединений; см. статью DeFelipe, J., et al., Cereb. Cortex 9:722-732, 1999).
Подсчитывают число синапсов/мм3. Результаты показаны на фиг.9. Число синапсов/мм3 существенно уменьшено у мышей со сверхэкспрессией нейротрипсина. Напротив, числа синапсов у контрольных мышей, т.е. у родительских линий, используемых для генерации дважды трансгенных (ДТГ) мышей со сверхэкспрессией нейротрипсина (491-неакт.Nt, 494-неакт.Nt и ЦМВ-Cre), такие же, как у мышей дикого типа. Вследствие этого данные результаты указывают на существенное уменьшение синапсов у мышей со сверхэкспрессией нейротрипсина.
Пример 13: Повышенные уровни нейротрипсина в нейронах ЦНС приводят в результате к уменьшенному числу дендритных шипиков (постсинаптических элементов).
Парасагиттальные срезы гиппокампа (300 мкм) мышей со сверхэкспрессией нейротрипсина и дикого типа в возрасте от 17 до 32 дней делают, используя бритвенное лезвие из нержавеющей стали (Electron Microscopy Sciences), переносят в инкубационную камеру, наполненную нагретым до 34°С и оксигенированной искусственной спинномозговой жидкостью (ACSF), и инкубируют в течение одного часа с целью представления ткани головного мозга достаточного времени для восстановления после повреждения при разрезании. Затем срезы хранят при комнатной температуре до дальнейшего использования в экспериментах.
Для регистрации пэтч-клампа целых клеток срезы помещают в стандартную погруженную камеру, орошаемую также ACSF. Отдельные нейроны визуализируют с использованием микроскопа Axioscope (Zeiss), оборудованного контрастной оптикой с дифференциальной интерференцией. Экспериментальную камеру поддерживают при температуре 35-36°С, которая близка к физиологической температуре. Скорость потока ACSF через камеру составляет от 1 до 2 мл/мин. ASCF оксигенируют оксиуглеродом перед вхождением в регистрационную камеру. Регистрации осуществляют, используя пипетку для регистрации целых клеток (3-5 МΩ), вынимаемую пинцетом Флеминга/Брауна и наполненную тем же раствором, что камера. Для морфологической реконструкции нейроны инъецируют с раствором 115 мМ КОН, 20 мМ K-глюконата, 10 мМ KCl, 10 мМ HEPES (буфера Гуда), 10 мМ фосфокреатина, 4 мМ ATP-Mg, 0,3 мМ ГТФ и 13,4 мМ биоцитина. Каждый срез с меченной биоцитином пирамидной клеткой СА1 помещают между двух кусков фильтровальной бумаги Millipore, чтобы сделать его плоским во время фиксации в течение 2-3 часов при комнатной температуре путем погружения в фиксирующий агент, включающий 1% глутаральдегид, 2% параформальдегид и приблизительно 0,2% пикриновой кислоты в 0,1 М фосфатного буфера (РВ), рН 7,4. Срезы хранят в 0,5% параформальдегиде в РВ при 4°С. После нескольких промываний в РВ срезы обрабатывают 2% пероксидом водорода в течение 15 минут и затем предварительно инкубируют в 20% нормальной козьей сыворотке в 0,05 М Трис-забуференном солевом растворе (рН 7,4), включающем 0,5% Тритон Х-100 (TBST) в течение 30 минут при комнатной температуре. Затем их подвергают инкубированию в течение ночи в реагенте Vectastain Elite ABC (авидин-биотин-пероксидаза) (1:100; Vector Labs) в TBST при 4°С. После промывания 5 раз по 15 минут в TBST и Трис буфере (ТВ, рН 7,6), содержащие биоцитин клетки визуализируют путем инкубирования в 3,3'-диминобензидине (0,05% в ТВ) в присутствии 0,0048% Н2О2. Реакцию останавливают несколькими промываниями в ТВ. Срезы монтируют на предметных стеклах и покрывают покровными стеклами в Mowiol (Hoechst).
Как демонстрируют на фиг.10, пирамидные нейроны СА1 гиппокампа трансгенных мышей со сверхэкспрессией Nt в нейронах ЦНС проявляют заметное уменьшение как числа, так и размера дендритных шипиков. Поскольку дендритные шипики представляют собой постсинаптическую сторону синапсов в данных нейронах, данные результаты независимым способом подтверждают уменьшение числа синапсов у мышей со сверхэкспрессией нейротрипсина.
Пример 14: Расщепление агрина in vivo происходит также в ЦНС и совпадает с экспрессией нейротрипсина в нейронах ЦНС.
Для тестирования расщепления агрина в ЦНС in vivo гомогенаты головного мозга развивающихся и взрослых мышей анализируют вестерн-блотом, используя специфические антитела к Nt и С-концевому фрагменту агрина 100 кД. Гомогенаты тканей получают из спинного мозга мышей в постнатальные дни 4, 7, 9, 10, 15 и 25, а также мышей в возрасте 3, 6 и 12 месяцев. Обнаруживают, что Nt наиболее сильно экспрессируется в первые три постнатальные недели при самых высоких уровнях экспрессии в дни от 7 до 10. Вестерн-блоттинг с антителами к С-концевой группе агрина показывает высокую близость временного характера, показывая, что расщепление агрина нейротрипсином также происходит in vivo в ЦНС.
Пример 15: Трансгенная сверхэкспрессия нейротрипсина в нейронах ЦНС усиливает расщепление агрина in vivo.
Гомогенаты тканей получают из головного мозга взрослых мышей со сверхэкспрессией либо человеческого нейротрипсина (hNt), либо мышиного нейротрипсина (mNt), генерированных скрещиванием мышей, несущих неактивные трансгены (линии 497, 489 и 533 для mNt и линии 493 и 494 для hNt) с мышами, экспрессирующими рекомбиназу Cre под контролем промотора ЦМВ, и для сравнения у мышей дикого типа. Гомогенаты головного мозга подвергают SDS-PAGE и вестерн-блоттингу. Вестерн-блоты зондируют антителами к hNt и mNt, а также к С-концевому фрагменту агрина 100 кД. Обнаружено, что уровень С-концевого фрагмента агрина 100 кД сильно повышен у трансгенных мышей со сверхэкспрессией нейротрипсина. Увеличение количества С-концевого фрагмента агрина 100 кД хорошо коррелирует с уровнем сверхэкспрессии в различных линиях трансгенных мышей. Человеческий и мышиный Nt демонстрируют одинаковый протеолитический эффект в отношении агрина. Данные результаты показывают, что Nt расщепляет агрин in vivo зависимым от концентрации образом.
Пример 16: Получение рекомбинантного нейротрипсина
Нейротрипсин представляет собой секретируемый мультидоменный белок длиной 875 аминокислот и приблизительно оцененного размера 97 кД для человеческого нейротрипсина и 761 аминокислот и размера 85 кД для мышиного нейротрипсина (см. фиг.1, А и В). Экспрессия данной серинпротеазы как активного белка зависит от правильной укладки и, весьма вероятно, посттрансляционных модификаций, например N-гликозилирования, который, как полагают, имеет два центра в случае человеческого и три центра для мышиного белка (см. статьи Gschwend, T.P. et al., Mol. Cell. Neurosci. 9: 207-219, 1997; Proba, К. et al., Biochim. Biophys. Acta 1396: 143-147, 1998). Кроме того, нейротрипсин включает сигнальный пептид, направляющий белок в эндоплазматический ретикулюм, из которого он секретируется. Нейротрипсин не является интегральным мембранным белком, поскольку в нем отсутствует трансмембранный домен, как определяют по графику гидрофобности (см. статью Kyte, J. и Doolittle, R.F., J. Mol. Biol. 157: 105-132, 1982). Центр активации зимогена нейротрипсина показывает высокий уровень близости к одному из tPA (активатор плазминогена тканевого типа; см. статью Tate, K.M. et al., Biochemistry 26: 338-343, 1987). Расщепление данного центра протеазой приводит к двум фрагментам: одному, включающему некаталитические домены с приблизительной молекулярной массой приблизительно 55 кД (для мышиного нейротрипсина) или приблизительно 67 кД (для человеческого нейротрипсина) и одному, включающему только домен протеазы приблизительно 30 кД.
Получение нейротрипсина в клетках миеломы. Для стабильной трансфекции клеток миеломы кодирующие участки мышиного и человеческого нейротрипсина вводят в специально сконструированный вектор (см. статью Traunecker et al., Biotechnol. 9: 109-113, 1991). Экспрессию с помощью данного вектора направляют промотор и энхансер Ig κ. 3'-конец транскрипта, представляющего интерес, сплайсируют в экзон, кодирующий константный домен Ig κ, чтобы имитировать стабильные транскрипты Ig. Вектор включает ген гистидинолдегидрогеназы, который дает возможность селекции стабильных трансфектантов в присутствии L-гистидинола. L-гистидинол является предшественником L-гистамина и ингибитором синтеза белка. Вектор стабильно трансфицируют в клеточную линию мышиной миеломы J558L (ЕСАСС #88032902; Европейская коллекция клеточных культур) для продукции рекомбинантного нейротрипсина. Другие стабильные линии, предназначенные для стабильной трансфекции путем слияния протопластов или электропорации, включают мышиную P3-X63Ag8.653, мышиную Sp2/0-Ag14, мышиную NSO и крысиную YB2/0 (см. статьи Gillies et al., Biotechnology 7: 799-804, 1989; Nakatani et al., Biotechnology 7: 805-810, 1989; Bebbington et al., Biotechnology 10: 169-175, 1992; Shitara et al., J. Immunol. Meth. 167: 271-278, 1994).
Стабильная трансфекция клеток J558L может быть достигнута при использовании электропорации. Всего 106 клеток смешивают в 10 мкг линеаризованного или сверхспирального вектора в PBS в кювете 1 см. Электропорацию осуществляют с использованием генного генератора импульсов Bio-Rad Gene Pulser (Bio-Rad Chemical Division) с импульсом 960 мкФ и 170-230 В. Клетки переносят в 50 мл DMEM, включающей 10% FCS (сыворотка телячьих эмбрионов) и помещают на пять 96-луночных планшетов путем добавления 100 мкл/лунку с использованием многоканального дозатора.
Липосомальную трансфекцию клеток J558L осуществляют с использованием липофектамина и реагента PLUS (Invitrogen). 1×107 клеток миеломы центрифугируют при 500×g в течение 3 минут и однократно промывают бессывороточной средой DMEM. После второго центрифугирования клетки ресуспендируют в 3 мл бессывороточной среды DMEM. Для трансфекции 40 мкг плазмидной ДНК, кодирующей нейротрипсин, смешивают с 320 мкл бессывороточной DMEM и 80 мкл реагента PLUS (Invitrogen). После инкубирования в течение 15 мин при КТ 60 мкл трансфекционного реагента липофектамина, предварительно смешанные с 340 мкл бессывороточной DMEM добавляют к реакции и инкубируют в течение 15 мин при КТ. К смеси ДНК-липосома добавляют 1 мл бессывороточной DMEM перед добавлением к клеткам J558L. Трансфицированные клетки инкубируют в чашке для клеточной культуры 6 см при 37°C с 10% СО2. Через 4 часа клетки разводят в 45 мл DMEM, включающей 10% FCS, и помещают в пять 96-луночных планшетов путем добавления 100 мкл/лунку при использовании многоканального дозатора.
Для получения протопластов глицериновую исходную суспензию клона штамма 803 Е.coli, включающего экспрессирующий вектор млекопитающих, высевают штрихом на чашку с агаром LB/ампициллином и выращивают в течение ночи при 37°С (штамм 803 доступен в АТСС #35581). Одной отдельной колонией инокулируют 2 мл предварительно нагретой (37°С) среды LB, содержащей 50 мкг/мл ампициллина. Через 4 часа встряхивания при 250 об/мин и 37°С 100 мкл культуры переносят в 100 мл свежей среды. После того, как культура достигнет оптической плотности приблизительно 0,6 (OD при 600 нм), добавляют хлорамфеникол до конечной концентрации 120 мкг/мл и выращивают в течение ночи при 250 об/мин и 37°С. Плазмиды, несущие точку начала репликации colE1 можно амплифицировать в присутствии хлорамфеникола (см. статью Hershfield et al., Proc. Natl. Acad. Sci. USA 71, 4355-3459, 1974). Суточную культуру центрифугируют при 2500×g в течение 10 мин при 4°С. Пеллету ресуспендируют в 2,5 мл ледяного 20% (масс./об.) раствора сахарозы в 50 мМ Трис-HCl, рН 8,0. 500 мкл ледяного лизоцима 1 мг/мл в 250 мМ Трис-HCl, рН 8,0 добавляют перед инкубированием на льду в течение 5 минут. После добавления 1 мл ледяной 250 мМ EDTA, рН 8,0 и инкубирования на льду в течение 5 минут добавляют 1 мл ледяной 50 мМ Трис-HCl, рН 8,0 и инкубируют препарат протопластов при КТ в течение 10 минут. Во время данного периода инкубирования можно наблюдать образование сферических протопластов из бактерий, обычно имеющих форму палочек, используя микроскоп с 1000-кратным увеличением. Приблизительно 90% протопластов должно сформироваться к концу периода инкубирования. К суспензии протопластов добавляют 20 мл DMEM с добавками 10% (масс./об.) сахарозы, 10 мМ MgCl2 и 40 мкл 10 мг/мл ДНКазыI. После инкубирования в течение 15 мин при КТ препарат протопластов центрифугируют при 2500×g в течение 30 мин при КТ. В это время клетки миеломы J558L готовят для слияния. Клетки миеломы выращивают в DMEM с добавлением 10% (об./об.) FCS, и они должны достигать высокой густоты клеток приблизительно 1×106 клеток/мл в день трансфекции. Для слияния протопластов 5×106 клеток центрифугируют при 500×g в течение 10 мин при КТ. Клетки ресуспендируют в 5 мл предварительно нагретой DMEM (37°С) и медленно наслаивают на поверхность пеллеты протопластов после последнего центрифугирования. Для смешивания протопластов и клеток миеломы пробирку центрифугируют при 500×g в течение 10 мин при КТ. После удаления супернатанта клетки перемешивают резким встряхиванием пробирки. Для слияния добавляют 2 мл ПЭГ (полиэтиленгликоля) 1500 в DMEM с добавкой 10% ДМСО (диметилсульфоксида) и пеллету ресуспендируют, несколько раз набирая и выдувая из пипетки. Через 1-2 мин после добавления раствора ПЭГ добавляют 10 мл предварительно нагретого DMEM с добавкой 10% (об./об.) FCS (37°С). Клетки центрифугируют при 500×g в течение 10 мин при КТ. Супернатант удаляют отсасыванием и пеллету ресуспендируют в 10% (об./об.) FCS и 100 мкл мг/мл канамицина. Наконец, клетки помещают в пять 96-луночных планшетов для культур тканей путем добавления 100 мкл/лунку, используя многоканальный дозатор.
Для селекции трансфицированных клеток добавляют L-гистидинол в конечной концентрации 5 мМ через 48 часов. Только трансфицированные клетки миеломы будут выживать при обработке L-гистидинолом. Клоны становятся видимыми приблизительно через 12-14 дней после начала селекции. В среднем на слияние протопластов и липосомальную трансфекцию получают 40-50 клонов. Все клоны анализируют на экспрессию вестерн-блотом с помощью антител, специфических к нейротрипсину. Хотя большинство клонов клеток миеломы не экспрессирует или экспрессирует только умеренные количества нейротрипсина, небольшая доля, 5-10%, показывает очень высокий уровень экспрессии. Клоны с высоким уровнем экспрессии субклонируют в процессе трех циклов одноклеточных разведений, чтобы удостовериться в стабильности экспрессии нейротрипсина. Из стабильно экспрессирующихся клонов собирают клеточные экстракты и супернатанты и разделяют на 10% SDS PAGE. Белки переносят на нитроцеллюлозную мембрану. Детекцию нейротрипсина осуществляют либо с помощью нейротрипсин-специфического антитела, распознающего некаталитический сегмент, и вторичного козьего антитела к кролику, связанного с пероксидазой, или нейротрипсин-специфического антитела, распознающего домен протеазы, и кроличьего антитела к козе, связанного с пероксидазой. Поскольку нейротрипсин полной длины преимущественно определяют в клеточном экстракте, полосу 65 кД, соответствующую некаталитическому фрагменту, и полосу 30 кД протеолитического домена определяют в супернатанте.
Пример 17: Крупномасштабная продукция нейротрипсина через промежуточные продукты.
Используемым источником нейротрипсина является кондиционированный супернатант клеточной культуры, полученный в результате культивирования клеточной линии миеломы, экспрессирующей нейротрипсин. Данные клетки адаптированы для роста в бессывороточной среде (см. статьи Stoll, T.S. et al., J. Biotechnol. 45: 111-123, 1996; Ackermann, G.E. и Fent, К., Marine Environmental Research 46: 363-367, 1998) в ферментере TechnoMouse (Integra Biosciences). Начиная со среды, состоящей из DMEM (Gibco, No. 41966-029), включающей 2 мМ глутамин и 10% FCS, клетки постепенно адаптируют к росту в данной среде с 1% FCS. Адаптацию осуществляют в 24-луночных планшетах и среду заменяют приблизительно каждый второй день. Когда клетки достигают конфлюэнтности, их отделяют в другую лунку. Во время всей процедуры клетки поддерживают при густоте, близкой к конфлюэнтности. Затем адаптированные клетки, хорошо растущие в среде DMEM, включающей 1% FCS, переносят в бессывороточную, но включающую белок среду HL-1 (Bio-Whittacker, No. 77201) с добавлением 0,5% FCS. Затем в среде HL-1 клетки постепенно адаптируют к росту только в среде HL-1 (без FCS). Для того чтобы адаптировать клетки к безбелковой среде TurboDoma (Cell Culture Technologies GmbH, Zurich, No. THP) среду HL-1 постепенно заменяют TurboDoma. Стадии адаптации от HL-1 к среде TurboDoma осуществляют аналогично уменьшению FCS.
Пример 18: Очистка нейротрипсина полной длины.
20 литров супернатанта из предшествующего примера фильтруют через оптический фильтр Polygard CR 1 мкм (Millipore) и концентрируют до 5 литров фильтрацией с поперечным потоком (SKAN AG).
Гепарин-аффинная хроматография. NaCl добавляют к концентрированному супернатанту для достижения конечной концентрации 0,3 М перед его нагрузкой на гепариновую колонку 120 мл (колонка Гепаринсефароза 6 Fast Flow XK 50/20 column; Amersham Biosciences). Колонку уравновешивают 20 мМ MOPS, 300 мМ NaCl, pH 7,2 (буфер А). Образец нагружают на колонку при скорости потока 1 мл/мин, при 20°С на устройстве Aekta Purifier (Amersham Biosciences Europe GmbH). Колонку промывают четырьмя объемами колонки буфера А. Связанные белки элюируют градиентом в 20 мМ MOPS, 1 M NaCl, pH 7,2 (буфер В). Используют следующий градиент: В 1 CV от 0 до 43% В, 3 CV при 43% В, в 2 CV от 43 до 100% В и 3 CV со 100% В. Нейротрипсин начинает элюироваться при концентрации хлорида натрия 450 мМ. Элюционные фракции, содержащие нейротрипсин (белок полной длины и домен протеазы) объединяют, берут аликвоты и хранят при -20°С.
Хроматографию на основе гидрофобного взаимодействия проводят на бутил-замещенном полимерном матриксе (Бутилсефароза 4 Fast Flow, Amersham Biosciences Europe GmbH). Для того чтобы подвести образец для условий нагрузки, медленно добавляют сухой хлорид натрия при постоянном перемешивании. После подведения концентрации хлорида натрия образец центрифугируют на центрифуге Sorvall RC-5B в течение 30 мин при 12000 об/мин в роторе SS34 при 4°С. Супернатант нагружают при скорости потока 1 мл/мин на уравновешенную колонку объемом 25 мл (20 мМ MOPS, 1,5 М хлорида натрия, pH 7,2) на хроматографической системе Aekta Purifier (Amersham Biosciences Europe GmbH) при 20°С. Связанные белки элюируют, применяя линейный градиент снижающейся концентрации хлорида натрия (1,5 М - 0,05 М) в 20 мМ MOPS, pH 7,2, при скорости 1 мл/мин. Нейротрипсин полной длины начинает элюироваться при концентрации 900 мМ хлорида натрия. Элюционные фракции, содержащие белок полной длины, объединяют и хранят при -20°С.
Аффинная хроматография с иммобилизованным металлом (IMAC). Ионы Cu2+ связывают с сефарозой (Хелатирующая сефароза Fast Flow, Amersham Biosciences Europe GmbH) согласно инструкциям изготовителя. К образцу добавляют твердый хлорид натрия для повышения концентрации до больше чем 0,5 М. Затем образец центрифугируют на центрифуге Sorvall RC-5B в течение 30 мин при 12000 об/мин в роторе SS34 при 29°С. Полученный в результате супернатант наносят на медь-сефарозную колонку объемом 1 мл при скорости потока 1 мл/мин на хроматографической системе Ettan (Amersham Biosciences Europe GmbH) при 4°С. Белки элюируют градиентом имидазола от 10 до 250 мМ в 20 мМ MOPS, 0,5 М хлориде натрия, рН 7,2. Используют следующий градиент: В 15 CV от 0 до 10% В, в 5 CV от 10 до 100% В и 10 CV при 100% В. Нейротрипсин полной длины начинает элюироваться при 150 мМ имидазола. Фракции, содержащие нейротрипсин полной длины, объединяют и хранят при 4°С.
Ионообменная хроматография. Для ионообменной хроматографии образец разводят в 2,5 раза до конечной концентрации хлорида натрия 0,2 М и центрифугируют на центрифуге Sorvall RC-5B в течение 30 мин при 12000 об/мин в роторе SS34 при 4°С. Прозрачный супернатант наносят при скорости потока 0,1 мл/мин на колонку MonoS PC 1,6/5 (Amersham Biosciences Europe GmbH), уравновешенную 20 мМ MOPS, 200 мМ NaCl, рН 7,2. Связанные белки элюируют линейным градиентом хлорида натрия (0,05 М - 1 М). Нейротрипсин полной длины начинает элюироваться при 300 мМ хлорида натрия. Фракции, включающие домен протеазы, объединяют и хранят при 20°С.
Таким образом нейротрипсин полной длины получают в электрофоретически чистой форме. Фиг.11 представляет очищенный нейротрипсин полной длины, как визуализируют окрашивание серебром на геле SDS-PAGE (А) и как визуализируют путем иммуноокрашивания с использованием нейротрипсин-специфического антитела на вестерн-блоте (В).
Пример 19: Очистка домена протеазы нейротрипсина.
Фракции, полученные при гепарин-аффинной хроматографии согласно предшествующему примеру, включающие белок полной длины и домен протеазы, подвергают хроматографии на основе гидрофобного взаимодействия, как описано выше. Супернатант центрифугированного раствора нагружают при скорости 1 мл/мин на уравновешенную колонку объемом 12 мл (20 мМ MOPS, 1,75 М хлорида натрия, рН 7,2). Связанные белки элюируют, применяя линейный градиент снижающейся концентрации хлорида натрия (1,75 М - 0,05 М) в 20 мМ MOPS, pH 7,2 при скорости 1 мл/мин. Домен протеазы начинает элюироваться при концентрации 1 М хлорида натрия. Фракции, включающие домен протеазы, объединяют и хранят при -20°С.
IMAC (Аффинную хроматографию с использованием иммобилизованного металла) проводят как для Nt полной длины.
Ионообменную хроматографию проводят как для Nt полной длины.
Таким образом выделенный каталитический домен нейротрипсина получают в электрофоретически чистой форме.
Пример 20: Клонирование, экспрессия и очистка инженерного белка агрина, пригодного в качестве субстрата, включающего центр расщепления α, но не центр расщепления β.
рсДНК-Агрин SN Y4Z19 кодирует связанную с мембраной форму агрина. Во-первых, секретируемый растворимый вариант агрина конструируют, используя праймеры 5'-Aaagttaacaaacctggaat ccacttcacaccagc-3' (SEQ ID NO:1) при интродукции сайта HpaI и 5'-AAAAGCGGCCGCTCATTTTTCGAACTGCGGGTGGCTCCAGGGAGTGGGGCA GGGTCTTAG-3' (SEQ ID NO:2) при интродукции сайта NotI. Полученный в результате ПЦР-продукт вырезают с помощью HpaI и NotI и клонируют в вектор рЕАК8, включающий кодирующую последовательность человеческого кальсинтенина-1, вырезанную с помощью тех же самых рестрикционных эндонуклеаз. Ранее дополнительный сайт HpaI в векторе удаляют, используя стратегию быстрой замены (Stratagene). Полученная в результате конструкция кодирует секретируемый агрин с сигнальной последовательностью человеческого кальсинтенина-1, который выщепляется при трансляции. Основываясь на данной конструкции, вводят N-концевую метку 8×His путем клонирования ПЦР-продукта с праймерами 5'-AAAAGTTAACCATCACCATCATCACCATCACCATAAACCTGGAATCCACTTC ACACCAG-3' (SEQ ID NO:3) и 5'-TTTATCATGACACAGTCGTTTTCATAG-3' (SEQ ID NO:4), используя сайты HpaI и BspHI в гене агрина. На третьей стадии домен LG3 на С-конце заменяют EGFP в SOE ПЦР с использованием праймеров 5'-GCTGGATATCAACAATCAGCAG-3' (SEQ ID NO:5) и 5'-GGTGAACAGCTCCTCGCCCTTGCTCACCATGGAGCCAACTAGCCCCTGTTCG CAGTGC-3' (SEQ ID NO:6) с вышеописанной конструкцией в качестве матрицы и 5'-GGTGAACAGCTC
CTCGCCCTTGCTCACCATGGAGCCAACTAGCCCCTGTTCGCAGTGC-3' (SEQ ID NO:7) и 5'-GGCTGCGGCCGCTCATTTTTCGAACTGCGGGTGGCTCCAGTTATCTAGATCCGGTGGATC-3' (SEQ ID NO:8) с pEGFP в качестве матрицы. Полученные в результате ПЦР-фрагменты комбинируют в SOE ПЦР и клонируют через сайт EcoRV и NotI в идентично разрезанный вектор. Полученный в результате секретируемый белок имеет последовательность SEQ ID NO:9.
Клетки HEK 293Т трансфицируют конструкцией инженерный агрин-EGFP и выращивают в среде DMEM с добавлением 10% FCS и культивируют в течение 6 часов. Среду заменяют DMEM без FCS и инкубируют в течение 50 час при 37°С. Двести мл супернатанта центрифугируют (4°С, 30 мин GS3, 5000 об/мин), чтобы избавиться от клеток и нерастворимого материала. Очищенный раствор диализуют два раза после разведения 1:25 против 50 мМ Трис-HCl, 150 мМ NaCl, 0,1% ПЭГ 6000, рН 8,0, при 4°С в течение ночи. После фильтрации (размер пор 0,45 мкм) раствор подвергают дальнейшей очистке посредством хроматографии.
IMAC-хроматография. После уравновешивания 5 CV 50 мМ Трис-HCl, 150 мМ NaCl, рН 8,0 образец белка нагружают со скоростью 10 мл/мин. Колонку промывают 10 CV 50 мМ Трис-HCl, 150 мМ NaCl, рН 8,0. Для элюции используют градиент от 0 до 500 мМ имидазола в буфере для промывания в 25 CV. Агрин-EGFP элюируют 20-50 мМ имидазолом. Фракции, включающие требуемый белок, как определяют вестерн-блоттингом и SDS-PAGE, объединяют, разводят 1:125 и диализуют против 20 мМ Трис-HCl, 150 мМ NaCl, 0,1% ПЭГ 6000, рН 8,0, при 4°С в течение ночи.
Анионообменная хроматография. После нагрузки диализата колонку POROS HQ20 объемом 8 мл промывают 2 CV 20 мМ Трис-HCl, 150 мМ NaCl, рН 8,0. Элюцию проводят градиентом 150-2000 мМ хлорида натрия в буфере для промывания. Агрин-EGFP элюируют хлоридом натрия в концентрации приблизительно 900-1100 мМ. Фракции, включающие требуемый белок, как определяют вестерн-блоттингом и SDS-PAGE, объединяют, разводят 1:25 и диализуют против 20 мМ Трис-HCl, 150 мМ NaCl, 0,1% ПЭГ 6000, рН 8,0 при 4°С в течение ночи. Полученный в результате нейротрипсин имеет чистоту 90% и пригоден для анализов активности in vitro. Белок замораживают в жидком азоте и хранят при -20°С.
Слитый белок агрин-EGFP, полученный данным образом, включает центр расщепления α, но не центр расщепления β. Агрин-EGFP имеет молекулярную массу больше чем 250 кД. Расщепление слитого белка агрин-EGFP нейротрипсином генерирует С-концевой фрагмент приблизительно 150 кД. На фиг.12 показан очищенный слитый белок агрин-EGFP, как визуализированный окрашиванием серебром на геле SDS-PAGE (А) и как визуализированный иммуноокрашиванием с использованием антитела к С-концевой половине агрина (см. пример 22) на вестерн-блоте (В). Отмечают, что EGFP не является основным для описанного применения, но скорее представляет собой заполнитель места. Замена EGFP другим белком или использование агрина полной длины, который является мутантным по центру расщепления β, представляют собой равноценные альтернативы описываемому продукту.
Пример 21: Клонирование, экспрессия и очистка маленького С-концевого фрагмента агрина (агрин-С45), пригодного в качестве субстрата, включающего центр расщепления β, но не центр расщепления α.
Используя праймеры 5'-GCGA GTTAACCACCATCACCATCACCATCACCATGGAAGCCTGGCTGACTTTAATGGCTTCTCCTACC-3' (SEQ ID NO:10), интродуцирующий сайт HpaI (HisBNterm) и 5'-ACCTGCGGCCGCTCATTTTTCGAACTGCGGGTGGCTCCAGCCAGAGCCAGAGCCGGGAGTGGGGCAGGGTCTTAGCTC-3' (SEQ ID NO:11), интродуцирующий сайт NotI (BStreplink), и pcDNA3.1-AgrinY0Z0 в качестве матрицы, амплифицируют фрагмент ДНК, кодирующий последние два домена LG и последний подобный EGF домен агрина. Используя данную стратегию, вводят последовательности ДНК, которые кодируют N-концевую метку 8×His и С-концевую метку Strep. Полученный в результате продукт ПЦР расщепляют рестрикционными ферментами NotI и HpaI (выделено жирным шрифтом в последовательностях праймеров) и клонируют в вектор рЕАК8, включающий кодирующую последовательность сигнального пептида человеческого кальсинтенина-1, разрезанную теми же самыми рестрикционными ферментами. Полученная в результате конструкция pEAK8-C45agrin включает кодирующий участок сигнальной последовательности человеческого кальсинтенина-1 как сигнал секреции С45-агрина. Клонирование осуществляют в Е.coli, а также путем амплификации плазмиды. Для экспрессии клетки HEK 293Т трансфицируют, используя метод с фосфатом кальция. Во время экспрессии в клетках HEK 293Т сигнальный пептид отщепляют. Полученный в результате секретируемый белок имеет последовательность (SEQ ID NO:12).
Клетки HEK 293Т культивируют до 80% конфлюэнтности в 7×500 см2 чашках для культур (CORNING) со 100 мл среды DMEM (GIBCO) с добавлением в каждую 10% FCS. Для трансфекции 35 мл 500 мМ CaCl2 и 35 мл буфера HBS (50 мМ HEPES, 140 мМ NaCl, 1,5 мМ Na2HPO4, pH 7,1) уравновешивают до КТ. Два мг ДНК pEAK8-agrin-C45 добавляют к раствору CaCl2 и смешивают с буфером HBS. Смесь для трансфекции инкубируют при КТ в течение 30 мин. Для трансфекции 500 см клеток НЕК 10 мл смеси для трансфекции по каплям добавляют к культуре и инкубируют в течение 4 час при 37°С в термостате при 37°С. Затем смесь для трансфекции удаляют однократным промыванием PBS и добавлением среды DMEM без FCS. Через 60 час. кондиционированную среду собирают и фильтруют, используя фильтр Steritop 0,22 мкм (MILLIPORE). Супернатнат непосредственно подвергают очистке IMAC, используя колонку Ni-NTA (8 мл, POROS) на перфузионной хроматографической системе BioCAD (Perseptive Biosystems) после подведения pH раствора до pH 8,5 1 М Трис буфером pH 8,5. Кондиционированную среду нагружают при скорости потока 10 мл/мин, колонку промывают 20 CV 100 мМ Трис-HCl, 150 мМ NaCl, pH 8,0. Используют элюцию линейным градиентом от 0 до 1 М имидазола в буфере для промывания с 10 CV. Фракции, включающие чистый фрагмент агрин-С45, объединяют и заменяют буфер с помощью колонки NAP 25 (Pharmacia) на 100 мМ Трис-HCl, 150 мМ NaCl, 10 мМ CaCl2, 0,1% ПЭГ 6000, pH 8,0. Очищенный белок замораживают в жидком азоте и хранят при -20°С. Фрагмент агрина, очищенный данным способом, является подходящим субстратом для нейротрипсина.
Фрагмент агрин-С45, генерированный данным способом, включает центр расщепления β, но не центр расщепления α. Агрин-С45 имеет молекулярную массу приблизительно 45 кД. Расщепление белка агрин-С45 нейротрипсином приводит к образованию N-концевого фрагмента приблизительно 23 кД и С-концевого фрагмента приблизительно 22 кД. На фиг.13 представлен очищенный белок агрин-С45, как визуализировано окрашиванием серебром на геле SDS-PAGE (А) и как визуализировано окрашиванием его С-концевой метки Strep с использованием StrepTactin на вестерн-блоте (В).
Пример 22: Генерация поликлонального антитела к С-концевой половине крысиного агрина.
Клетки HEK 293Т выращивают до 80% конфлюэнтности в 16×150 см2 колбах для культур тканей. Каждую колбу используют для инокуляции 4×500 см2 чашек. Чашки объемом 500 см2 содержат 80 мл среды для культивирования и покрыты поли-L-лизином. Клетки выращивают до 60-80% конфлюэнтности в течение двух дней. Клетки трансфицируют методом с использованием фосфата кальция 1 мкг каждого из pcDNA-agrinY4Z8 и pcDNA-hNT на мл среды для культивирования (DMEM/10%FCS). Среду заменяют DMEM без FCS на следующее утро. Клетки выращивают в течение 4 дней при 37°С/10% СО2. Супернатант собирают, центрифугируют в течение 30 мин при 3000 об/мин при 4°С и фильтруют (размер пор 0,45 мкм) при КТ. рН фильтрата подводят до 7,0 1 М буфером HEPES, но не превышая конечную концентрацию 20 мМ. Его нагружают со скоростью 1 мл/мин на гепариновой колонке (17 мл гепаринсефарозы; емкость приблизительно 2 мг/мл гелевой матрицы). Колонку уравновешивают 5 CV 20 мМ HEPES, 80 мМ NaCl, рН 7,5 и промывают 2 CV 20 мМ HEPES, 80 мМ NaCl, рН 7,5. Связанные белки элюируют линейным градиентом более 8 CV от 80 до 1000 мМ NaCl в 20 мМ HEPES, рН 7,5. Фрагмент 100 кД элюируют при 400-600 мМ NaCl. Фракции, которые включают целевой белок, объединяют, как мониторируют SDS-PAGE и вестерн-блотом. Объединенные фракции диализуют в течение ночи при 4°С против 20 мМ HEPES, рН 7,5 1:100, чтобы концентрация NaCl снизилась до меньше чем 5 мМ. Диализат центрифугируют в течение 30 мин со скоростью 12000 об/мин при 4°С и нагружают на колонку MonoQ (7,8 мл, колонка HQ POROS, емкость: матрица 10-20 мг/мл), уравновешенную 20 мМ Трис, рН 8,0. Связанные белки элюируют линейным градиентом 20 CV от 0 до 1000 мМ NaCl в 20 мМ Трис, рН 8,0. Фрагмент 100 кД элюируют при 100-200 мМ NaCl. Фракции, включающие целевой белок, как определяют с помощью SDS-PAGE и вестерн-блота, объединяют.
Иммунизация. Очищенный белок быстро замораживают, капая в пробирку, наполненную жидким азотом, и хранят при -80°С. Кроликов иммунизируют стандартными методами, используя порции белка по 50 мкг. Полученные в результате антитела подходят для детекции агрина полной длины, а также фрагментов агрина, включающих С-концевую часть агрина, в частности фрагментов агрина, включающих часть агрина, находящуюся между центрами расщепления α и β.
Пример 23: Генерация поликлонального антитела к домену LG3 крысиного агрина.
Клетки HEK 293Т культивируют до 80% конфлюэнтности в 5×500 см2 чашках для культур (CORNING) со 100 мл среды DMEM (GIBCO) с добавлением в каждую 10% FCS. Для трансфекции к 25 мл 500 мМ CaCl2 и 25 мл буфера HBS (50 мМ HEPES, 140 мМ NaCl, 1,5 мМ Na2HPO4, pH 7,1) добавляют 1,5 мг ДНК pEAK8-agrin-C45 и 15 мкг ДНК pcDNA-hNT. Смесь для трансфекции инкубируют при КТ в течение 45 мин. Для трансфекции 500 см2 клеток HEK 10 мл смеси для трансфекции по каплям добавляют к культуре и инкубируют в течение 4 час при 37°С. Смесь для трансфекции удаляют трехкратным промыванием PBS и добавлением среды DMEM без FCS. Через 60 час кондиционированную среду собирают и фильтруют, используя фильтр Steritop 0,22 мкм (MILLIPORE). Сотрансфекций клеток pcDNA-hNT фрагмент агрин-С45 расщепляют и высвобождают домен LG3 крысиного агрина. Для того чтобы избавиться от основных примесей, кондиционированную среду разводят 1:10, диализуют 5 раз против 50 мМ Трис-HCl, 50 мМ NaCl, pH 8,0 и подвергают анионообменной хроматографии. Диализуемую среду нагружают при скорости 10 мл/мин на колонку MonoQ 4 мл (собственная упаковка фирмы Uno Sphere MonoQ материалом фирмы BioRAD, 2×4 см), соединенную с хроматографической системой BioCAD (Perseptive Biosystems). Колонку промывают 20 CV 50 мМ Трис-HCl, 50 мМ NaCl, pH 8,0. Для элюции используют градиент от 50 мМ NaCl до 2000 мМ NaCl в 50 мМ Трис-HCl, pH 8,0. (Первую стадию можно необязательно заменить металл-аффинной хроматографией, используя колонку с Ni2+-хелатированной сефарозой). Целевые белки обнаруживают в потоке через фракцию и непосредственно подвергают аффинной хроматографии с использованием колонки StrepTactin объемом 10 мл, уравновешенной 50 мМ Трис-HCl, 150 мМ NaCl, pH 8,0. После связывания белков при безнапорном потоке колонку промывают 10 CV буфера для уравновешивания. Элюцию осуществляют, используя 6×0,5 CV буфера для уравновешивания с добавлением 2,5 мМ дезтиобиотина. Элюцию анализируют, используя SDS-PAGE, и фракции, включающие фрагмент агрин-С45 и домен LG3, концентрируют, используя концентраторы Centriprep 10000 (MILLIPORE), до объема 200 мкл. Полученный в результате концентрат нагружают на колонку для гель-фильтрации Superdex S75 (Amersham Pharmacia, 1,6×30). Хроматографию проводят при скорости потока 0,3 мл/мин, используя 50 мМ Трис-HCl, 250 мМ NaCl, pH 8,0. Элюцию анализируют, используя SDS-PAGE, фракции, включающие чистый домен LG3, объединяют и замораживают в жидком азоте.
Иммунизация. Для генерации поликлональных антител к домену LG3 агрина 50 мкг фрагмента используют для иммунизации кроликов. Полученные в результате антитела используют для детекции агрина полной длины, а также для детекции фрагментов агрина, включающих домен LG3 агрина.
Пример 24: Анализ на протеолитическую активность нейротрипсина.
Измерение активности нейротрипсина проводят в 150 мМ NaCl, 5 мМ CaCl2, 0,1% ПЭГ и 20 мМ MOPS, pH 7,5 в пробирках с низким уровнем связывания белка (Eppendorf). Измерения активности нейротрипсина можно провести, используя тот же буфер, включающий до 30% ДМСО. Человеческий нейротрипсин используют в концентрации, которая приводит в результате к расщеплению приблизительно 80% субстрата в течение 3 часов. В качестве субстрата используют 0,1-1 мкМ агрин-EGFP или 0,1-3 мкМ агрин-С45. Реакционную смесь инкубируют в течение 3 часов при 37°С. Затем реакцию останавливают добавлением принятого буфера для образцов SDS-PAGE и нагреванием при 70°С в течение 5 мин. Полученные продукты расщепления проверяют после SDS-PAGE.
На фиг.14 представлен пример анализа с использованием инженерного агрин-EGFP в качестве субстрата и антитела к С-концевой части агрина для детекции С-концевого продукта расщепления агрин-EGFP после SDS-PAGE и вестерн-блоттинга. Фиг.15 представляет пример анализа с использованием агрин-С45 в качестве субстрата. В данном случае Стрептактин (IBA GmbH) используют для детекции С-концевого продукта расщепления. Антитела к домену LG3 агрина (полученные, как описано в Примере 23) также могут быть использованы для данной цели.
Альтернативно гели SDS-PAGE окрашивают принятыми методами окрашивания белка, такими как окрашивание серебром (см. фиг.15,А), кумасси бриллиантовым синим или, для количественной оценки, Sypro ruby (BioRad).
Пример 25: Определение центров расщепления для нейротрипсина в агрине: Центры расщепления α и β.
Для того чтобы определить точное положение расщепления центра расщепления α, вариант связанного с мембраной агрина соэкспрессируют с человеческим нейротрипсином в клетках HEK293T. Полученный в результате продукт расщепления 100 кД, появляющийся в культуральном супернатанте, очищают (см. пример 22) и секвенируют N-конец с помощью разложения Эдмана на секвенаторе Precise 492 cLC (Applied Biosystems). Определяемая последовательность представляет собой ASXYNSPLGXXSGDK, в которое Х стоит на месте остатков цистеина. На основании изложенного можно заключить, что разложение находится в цепи последовательности VVTHGPPIERASCYNSPLGCCSDK после аргинина в положении 995.
Элайнменты последовательностей ряда последовательностей агрина млекопитающих и последовательности агрина кур (Gallus gallus) показывают высокую степень эволюционного консерватизма аминокислот, фланкирующих центр расщепления α агрина.
Пять аминокислот на N-концевой и четыре аминокислоты на С-концевой стороне центра расщепления α агрина (центр расщепления α агрина находится между Аргинином 995 и Аланином 996) определяют консенсусную последовательность P-P/A-I/V-E-R-A-S/T-C-Y для центра расщепления α агрина.
Для определения точного положения расщепления центра расщепления β агрин-С45 соэкпрессируют с человеческим нейротрипсином в клетках HEK293T. Полученный в результате продукт расщепления 21 кД очищают (см. пример 23) и секвенируют N-конец. Полученная в результате последовательность представляет собой SVGDLETLAF. Данную последовательность обнаруживают в цепи GLVEKSVGDLETLAFDGRT. На основании изложенного заключают, что агрин расщепляется нейротрипсином после лизина в положении 1754 между доменами EGF4 и LG3.
Элайнменты последовательностей ряда последовательностей агрина млекопитающих и последовательности агрина кур (Gallus gallus) показывают высокую степень эволюционного консерватизма аминокислот, фланкирующих центр расщепления β агрина.
Пять аминокислот на N-концевой и четыре аминокислоты на С-концевой стороне центра расщепления β агрина (центр расщепления β агрина находится между Лизином 1754 и Серином 1755) определяют консенсусную последовательность G/A-L/I/T-V/I-E-K-S-V/A-G для центра расщепления β агрина.
Вышеописанные результаты можно было бы подтвердить, используя анализы активности in vitro с очищенным человеческим нейротрипсином и вариантами очищенного агринового субстрата, путем переноса полученных в результате продуктов расщепления на мембрану PVDF и последующего секвенирования N-конца.
В заключение, агрин расщепляется нейротрипсином в двух положениях. Первый центр расщепления (центр расщепления α) обнаружен в положении R995-A996 (считая от регистрационного номера NP_786930, крысиный агрин), т.е. расщепление происходит на С-концевом аргинине в цепи последовательности PPIERASCY агрина крысы (Rattus norvegicus) между сегментом, богатым серин-треонином и доменом SEA. Сравнение последовательностей млекопитающих и птиц, фланкирующих центр расщепления а агрина, определяет консенсусную последовательность P-P/A-I/V-E-R-A-S/T-C-Y центра расщепления α агрина, причем расщепление нейротрипсином происходит на С-концевом остатке аргинина (R). Второе расщепление (центр расщепления β) находится в положении K1754-S1755 (считая от регистрационного номера NP_786930, крысиный агрин), т.е. расщепление происходит на С-концевом лизине в контексте последовательности LVEKSVGD в сегменте, связывающем EGF4 и домен LG3 агрина. Сравнение последовательностей млекопитающих и птиц, фланкирующих центр расщепления β агрина, определяют консенсусную последовательность G/A-L/I/T-V/I-E-K-S-V/A-G центра расщепления β агрина, причем расщепление нейротрипсином происходит на С-концевом остатке лизина (K).
Пример 26: Получение маленьких молекул соединений для анализа ингибирования.
Соединения растворяют в ДМСО до получения конечной концентрации 10 мМ. Для анализа раствор в ДМСО разводят 10 мМ MOPS, pH 7,5 до концентрации 500 мкМ и 5% ДМСО (разведение 1:20). Нерастворимый и осажденный материал удаляют центрифугированием (15 мин, 16 krcf, KT). Осветленный супернатант используют для анализа ингибирования, как описано в Примере 27.
Пример 27: Анализ для определения ингибирующей активности маленьких молекул соединений нейротрипсина.
Ингибирующую активность маленьких молекул соединений в отношении каталитической активности нейротрипсина измеряют в 150 мМ NaCl, 5 мМ CaCl2, 5% ДМСО, 0,1% ПЭГ 6000 и 20 мМ MOPS, pH 7,5 в пробирках Eppendorf с низким уровнем связывания белка объемом 0,5 мл в общем объеме 15 мкл. Человеческий или мышиный нейротрипсин или каталитически активные укороченные варианты нейротрипсина используют в концентрации, которая в результате обеспечивает расщепление 80% субстрата в течение 3 часов. В качестве субстрата используют инженерный растворимый агрин, например 0,1-1 мкМ агрин-EGFP (см. пример 20) или 0,1-3 мкМ инженерного агрин-С45 (см. пример 21). Раствор ингибитора в 10 мМ MOPS, pH 7,5, содержащий 5% ДМСО, добавляют до конечной концентрации: либо 25, либо 150 мкМ. Реакционную смесь инкубируют в течение 3 часов при 37°С. 5% ДМСО в реакционной смеси требуется для поддержания растворимости маленьких неорганических соединений ингибиторов. Реакцию начинают добавлением субстрата или фермента. В конце периода инкубирования реакцию останавливают добавлением принятого буфера для образцов SDS-PAGE и нагреванием до 70°С в течение 5 минут. Разложившийся образец разделяют с помощью SDS-PAGE и изучают после визуализации субстрата.
Одним способом визуализации субстрата является вестерн-блоттинг. Для анализа белки переносят на нитроцеллюлозные мембраны. По интенсивностям фрагмента разложения 150 кД, генерированного при расщеплении агрин-EGFP, или фрагмента разложения 22 кД, генерированного при разложении агрин-С45, оценивают ингибирующие активности подвергаемых скринингу маленьких молекул соединений на нейротрипсин. На фиг.16 показан типичный результат скрининга ингибиторов с помощью анализа вестерн-блот при анализе с использованием агрин-EGFP в качестве субстрата и антитела к С-концевой части агрина для детекции. Измеряют интенсивность фрагментов 150 кД агрин-EGFP, генерированных посредством нейротрипсин-опосредованного разложения в присутствии предполагаемых ингибирующих соединений (см. фиг.16,А) и строят график относительных интенсивностей (см. фиг.16,В).
Обнаруживают, что соединение No. 7 (идентификационный номер 1672-3440, ChemDiv, San Diego, CA, USA, N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламид, название IUPAC: амино{[(4-{[(1Е)-(3,5-дибромо-2-гидроксифенил)метилен]амино}фенил)сульфонил]амино}метаниминий) обладает существенной активностью ингибирования нейротрипсина.
Альтернативно визуализацию и количественное определение образца после разложения осуществляют непосредственно в геле посредством окрашивания принятыми методами окрашивания белка, такими как окрашивание серебром, окрашивание кумасси бриллиантовым синим или, для количественной оценки, путем окрашивания Sypro ruby (BioRad).
Пример 28: Дозозависимость ингибирующего соединения No. 7, N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламида.
Различные концентрации соединения No. 7 от 0 до 200 мкМ тестируют с помощью анализа, описанного в примере 27 с инженерным агрин-EGFP в качестве субстрата. Генерированный продукт определяют вестерн-блотом (см. фиг.17,А) и количественно определяют (см. фиг.17,В). Полумаксимальное количество продукта по сравнению с реакцией без соединения No. 7 обнаруживают при концентрации приблизительно 60 мкМ, таким образом, значение IC50 для соединения No. 7 лежит в интервале приблизительно 60 мкМ.
Пример 29: Определение специфичности ингибирования нейротрипсина для соединения No. 7, N1-амидино-N4-(3,5-дибромосалицилиден)-сульфаниламида.
Для того чтобы проверить специфичность соединения, которое, как показано, обладает ингибирующим эффектом на человеческий нейротрипсин, проводят стандартные кинетические измерения фермента с набором распространенных серинпротеаз. Используют стандартный фотометрический анализ с коммерчески доступными протеазами и субстратами в виде связанных с пара-нитроанилидом маленьких пептидов.
Протеазы:
Фактор Ха, активированный плазмой крупного рогатого скота (6,1 мг белка/мл; Sigma Aldrich Chemie GmbH, D-89552 Steinheim, Germany).
Трипсин из свиной поджелудочной железы (16099 ед./мг; Fluka Chemie AG, CH-9471 Buchs, Switzerland).
tPA: Актилиз (10 мг; Dr. Karl Thomae GmbH, Birkendorfer Straβe 65, D-88397 Biberach, Germany).
Тромбин из плазмы крупного рогатого скота (50 NIH (единица тромбина Национального Института здоровья)/мг, Merck, D-64271 Darmstadt, Germany).
Урокиназа HS medac (100000 1.Е.(межд. Ед.); medac Gesellschaft für klinische Spezialpräparate mbH, D-22880 Wedel, Germany).
Калликреин из свиной поджелудочной железы (43 ед./мг твердого вещества; Sigma Aldrich Chemie GmbH).
Плазмин из плазмы человека (3,2 ед./мг твердого вещества; Sigma Aldrich Chemie GmbH).
Субстраты:
Bz-IEGR-pNA: S-2222, Chromogenix-Instrumentation Laboratory SpA, 1-20128 Milano, Italy.
Bz-FVR-pNA: N-бензоил-Phe-Val-Arg-п-нитроанилид HCl, Bachem AG, CH-4416 Bubendorf, Switzerland.
IPR-pNA: S-2288, Chromogenix-Instrumentation Laboratory SpA.
Bz-VGR-pNA: N-бензоил-Val-Gly-Arg-п-нитроанилид, Sigma Aldrich Chemie GmbH.
N-тозил-GPK-pNA: N-тозил-Gly-Pro-Lys-п-нитроанилид, #90178, Fluka Chemie AG.
Условия анализа:
Анализ проводят в 100 мМ Трис-HCl, 150 мМ NaCl, 1 мМ CaCl2, 5% ДМСО, 0,1% ПЭГ 6000, рН 8,0 с соответствующим количеством протеазы с целью измерения исходных скоростей и варьирующих количеств пептид-р-нитроанилидных субстратов в интервале меньше чем 0,1 КМ. Для исследований ингибирования используют 100 мкМ соединение No. 7. Измерения проводят при 25°С на спектрофотометре Cary 50 (VARIAN). Исходные скорости определяют, используя различные концентрации субстрата ниже 0,1 КМ, чтобы они находились в интервале прямой зависимости исходной скорости от концентрации субстрата. Строят график зависимости исходных скоростей от концентраций субстрата в отсутствие или в присутствии потенциального ингибитора No. 7. Соединение используют в концентрации 100 мкМ.
Ни один из исследованных ферментов не показывает существенного ингибирования соединением No. 7 в концентрации 100 мкМ. На фиг.18-24 представлены результаты кинетических измерений ферментов в присутствии и в отсутствие соединения No. 7 для фактора Ха, трипсина, tPA, тромбина, урокиназы, калликреина и плазмина.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИЗОБРЕТЕНИЕ БЕЛКА PI4KIIIA И РОДСТВЕННОГО МЕМБРАННОГО БЕЛКОВОГО КОМПЛЕКСА В ЛЕЧЕНИИ БОЛЕЗНИ АЛЬЦГЕЙМЕРА | 2016 |
|
RU2724493C2 |
| ПЕПТИДЫ И ИХ ПРОИЗВОДНЫЕ, ВЗАИМОДЕЙСТВУЮЩИЕ С НИКОТИНОВЫМ АЦЕТИЛХОЛИНОВЫМ РЕЦЕПТОРОМ И ПРИГОДНЫЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КОСМЕТОЛОГИИ ПРОТИВ МИМИЧЕСКИХ И ВОЗРАСТНЫХ МОРЩИН | 2013 |
|
RU2524428C1 |
| Штамм клеток Escherichia coli BL21(DE3)/pET32v11-Cre, продуцирующих сайт-специфическую рекомбиназу Cre | 2020 |
|
RU2761637C1 |
| СРЕДСТВА, ПУТИ ПРИМЕНЕНИЯ И СПОСОБЫ ЛЕЧЕНИЯ СИНУКЛЕОПАТИИ | 2016 |
|
RU2765303C2 |
| ПРИМЕНЕНИЕ TPK В КАЧЕСТВЕ МИШЕНИ ПРИ БОЛЕЗНИ АЛЬЦГЕЙМЕРА | 2020 |
|
RU2804334C2 |
| ПЕРЕНОС ГЕНОВ, ОПОСРЕДОВАННЫЙ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, И ИХ ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2288742C2 |
| ГЕНЕТИЧЕСКИЕ НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ИСКУССТВЕННО МОДИФИЦИРОВАННОГО GDNF С ДЕЛЕТИРОВАННОЙ pro-ОБЛАСТЬЮ, ПРОДУКТ КОТОРЫХ ОБЛАДАЕТ ПОВЫШЕННЫМИ СВОЙСТВАМИ НЕЙРАЛЬНОГО ИНДУКТОРА И СТИМУЛЯТОРА ОБРАЗОВАНИЯ НЕЙРАЛЬНЫХ ОТРОСТКОВ, ПРИГОДНОГО ДЛЯ ТЕРАПИИ НЕЙРАЛЬНЫХ ТРАВМ, ИШЕМИЧЕСКИХ ИНСУЛЬТОВ И НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ, ВЕКТОР ЭКСПРЕССИИ, МОДИФИЦИРОВАННЫЙ GDNF | 2014 |
|
RU2595377C2 |
| ТРАНСГЕННОЕ ЖИВОТНОЕ, ОТЛИЧНОЕ ОТ ЧЕЛОВЕКА, И ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2579701C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИСПЫТАНИЙ НА ТОКСИКОГЕННОСТЬ | 2012 |
|
RU2616281C2 |
| МОДИФИЦИРОВАННЫЙ БИОСЕНСОР ДЛЯ ДЕТЕКЦИИ ВНУТРИКЛЕТОЧНОЙ РН | 2014 |
|
RU2603060C2 |
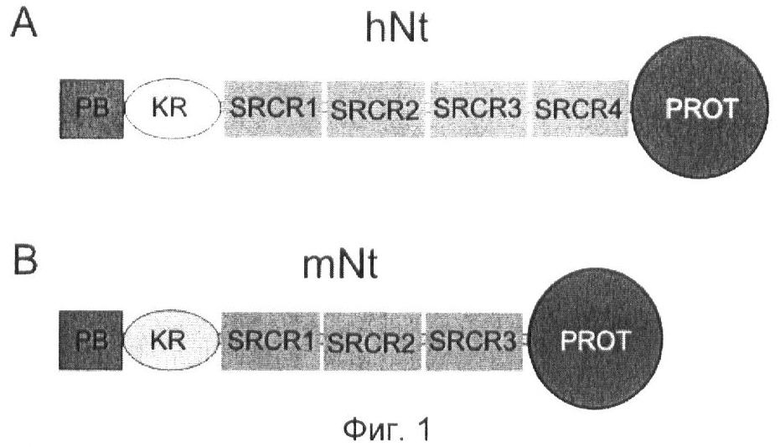
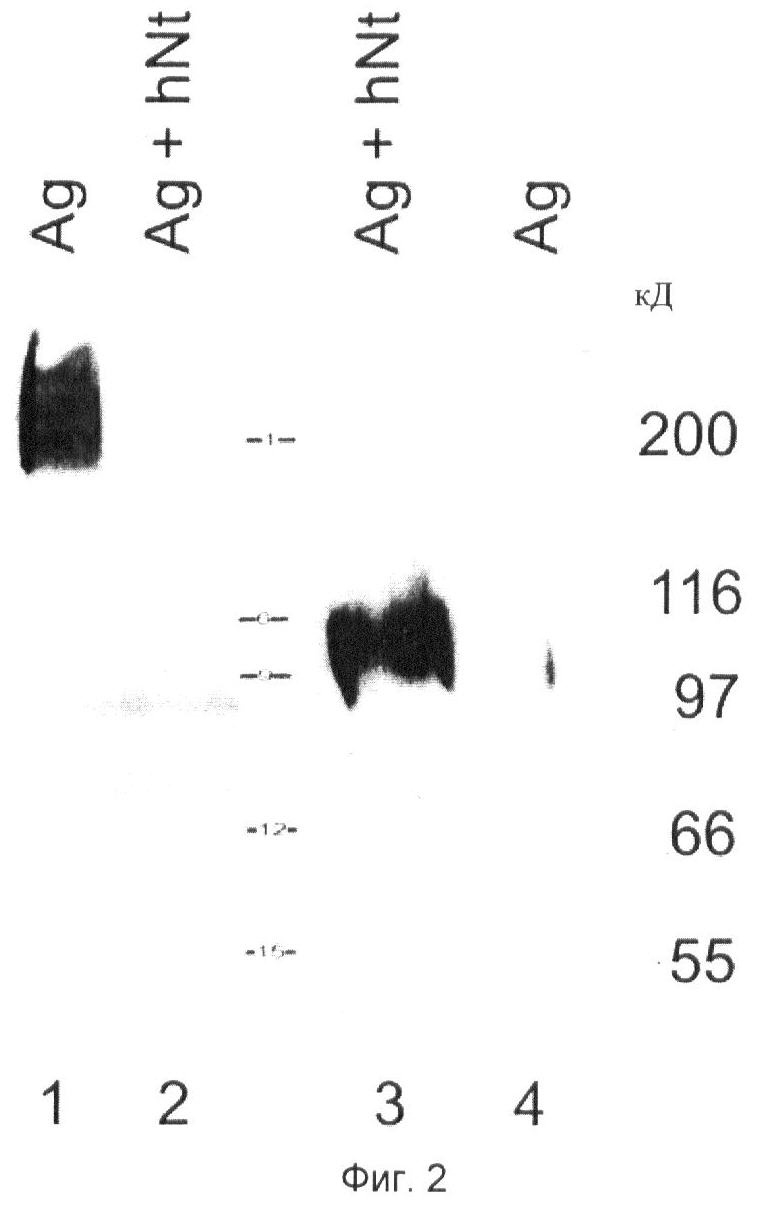
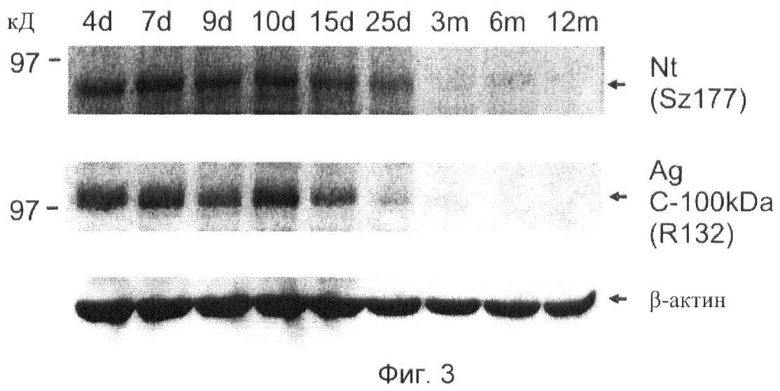
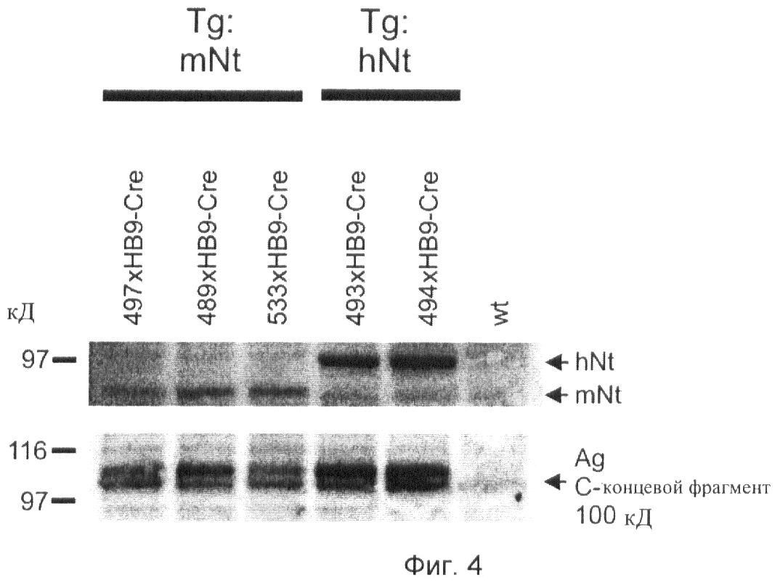
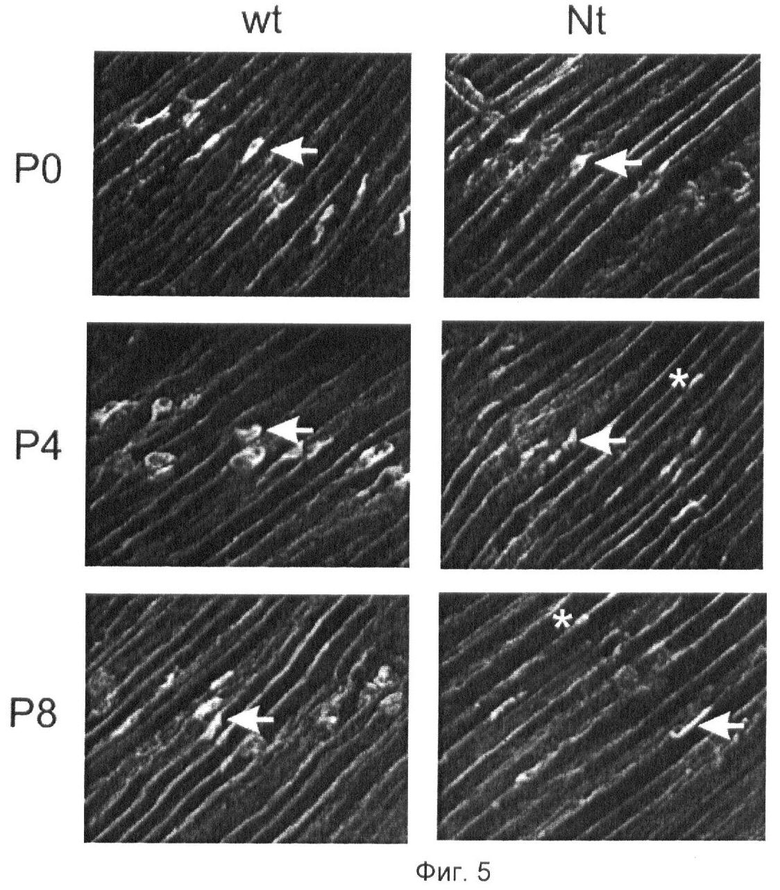
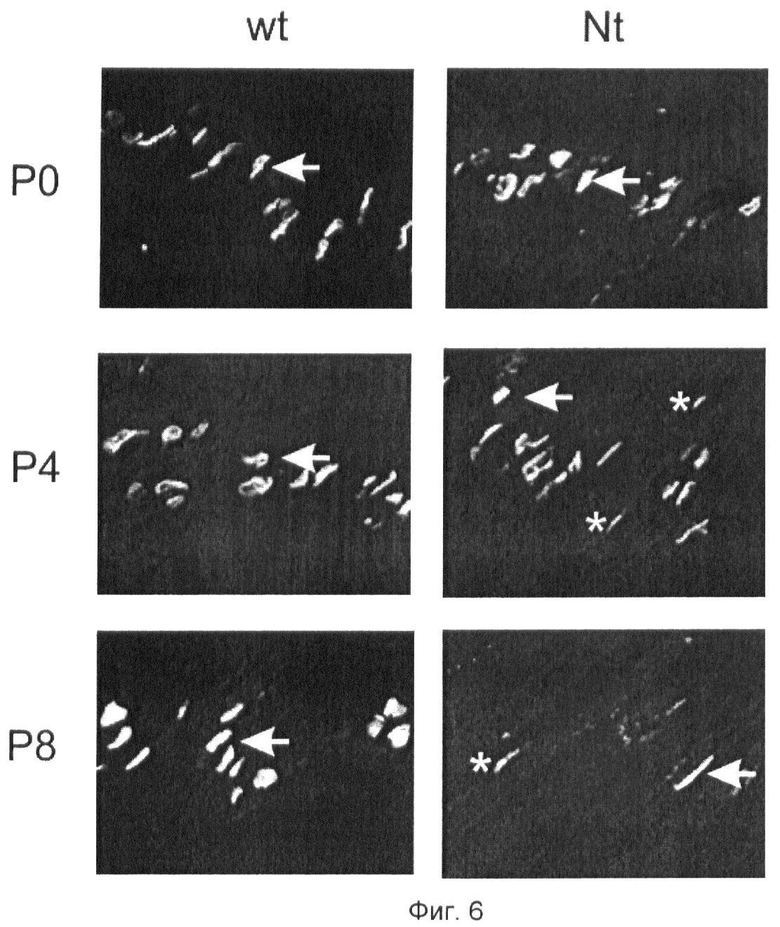
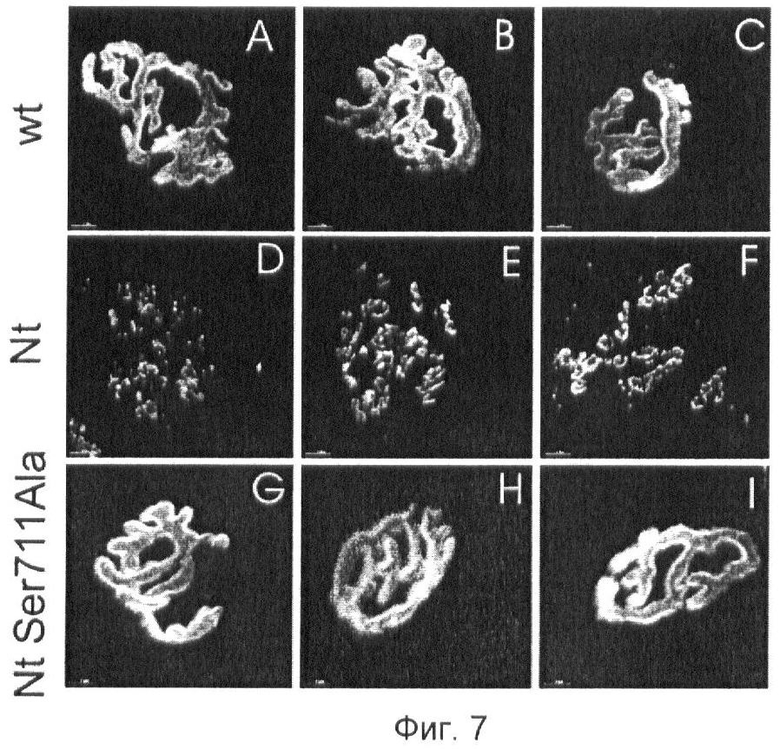
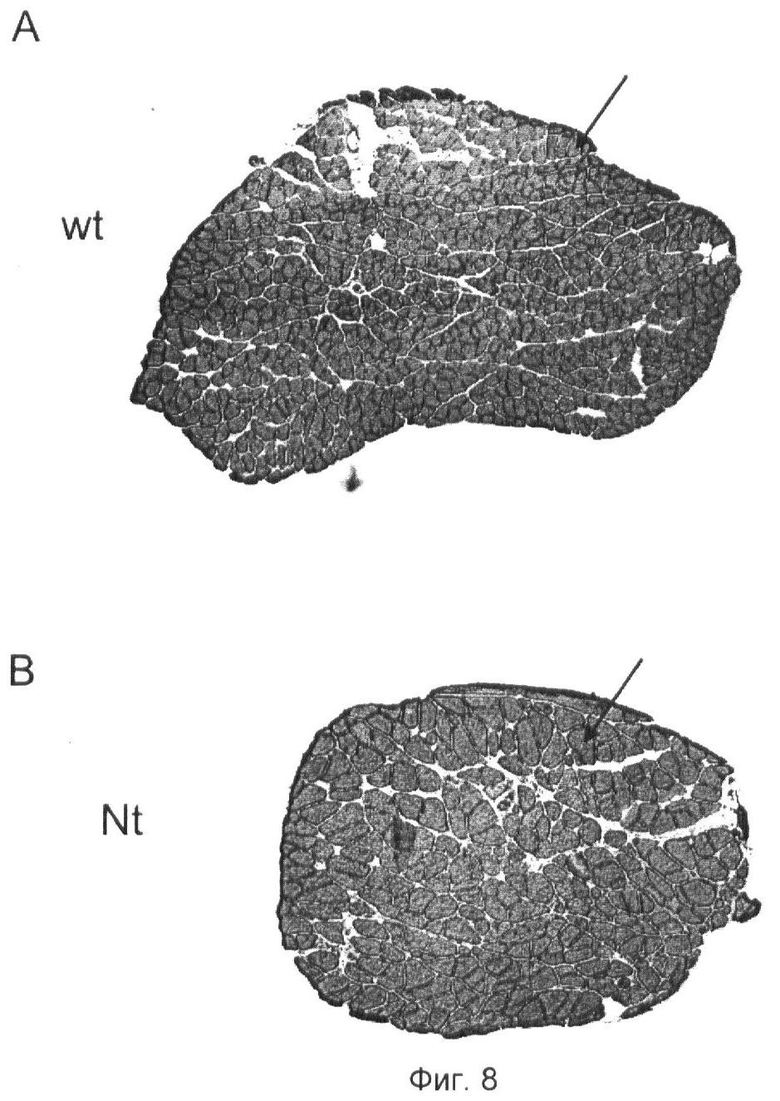
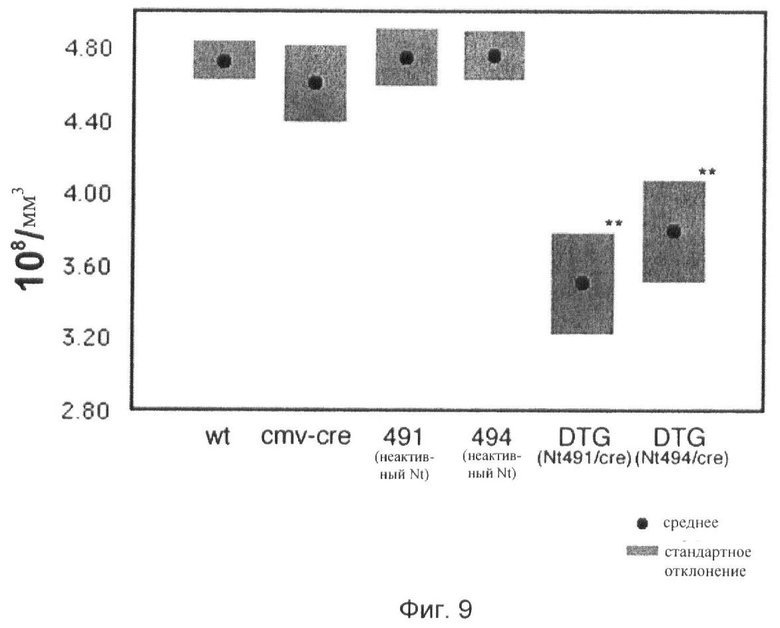
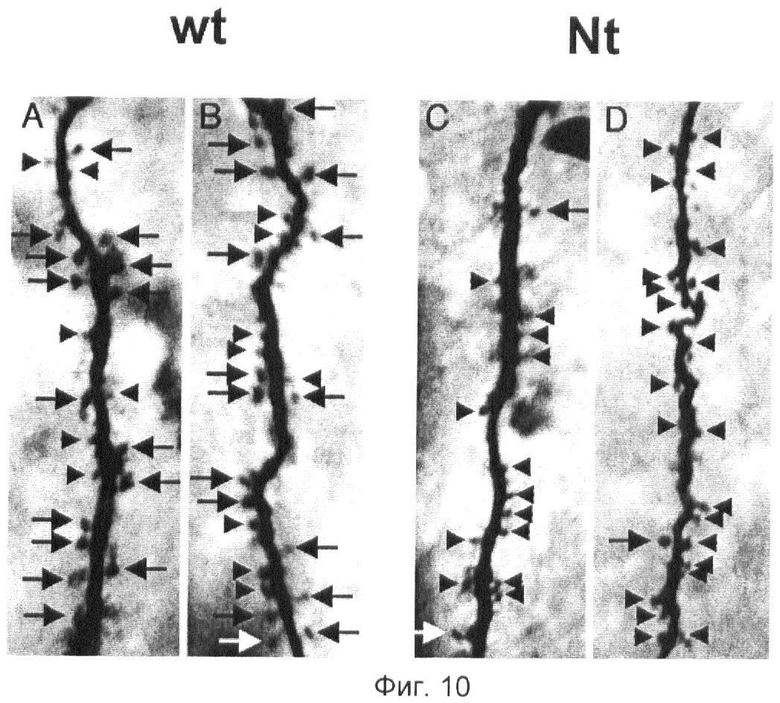
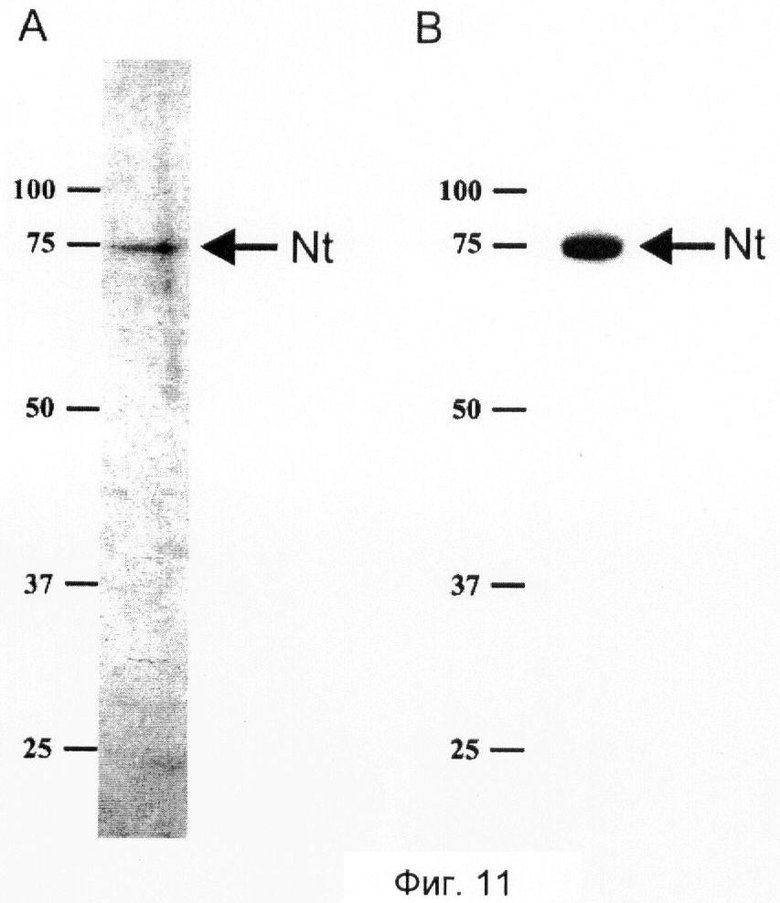
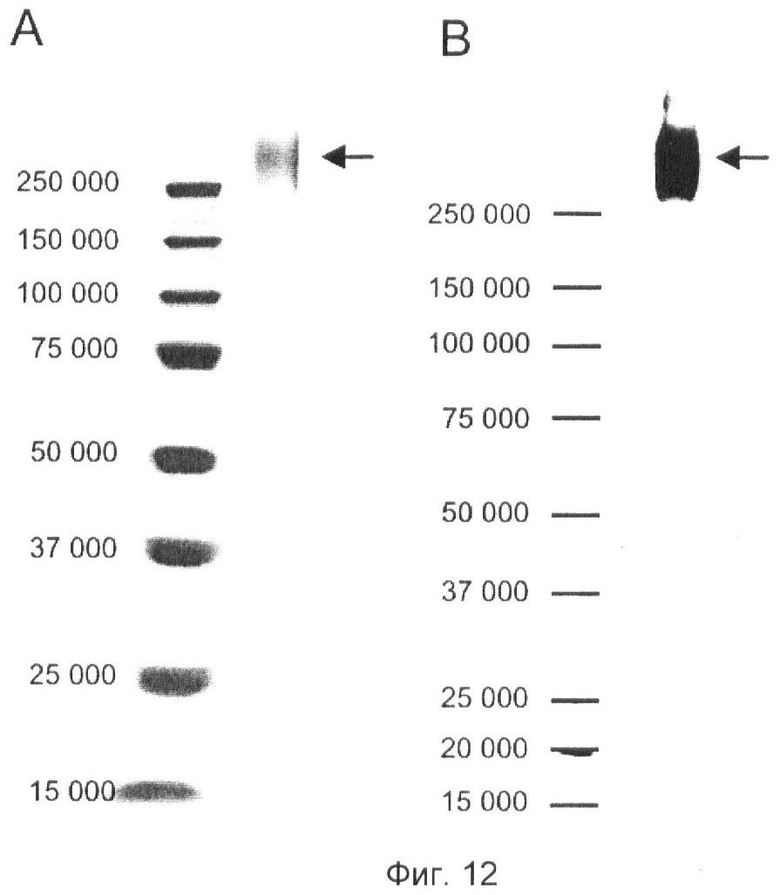
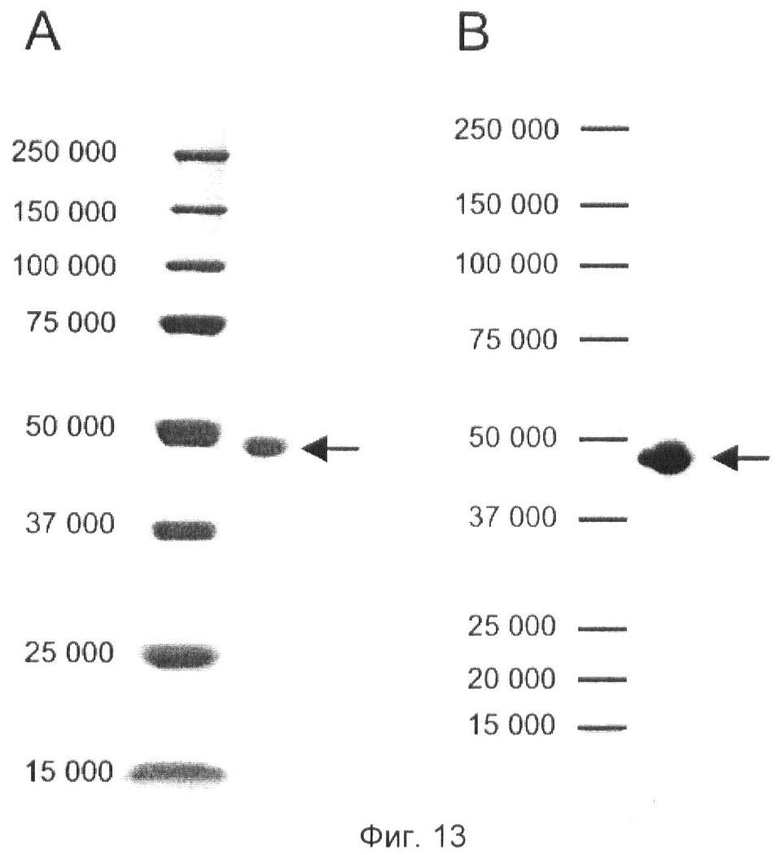
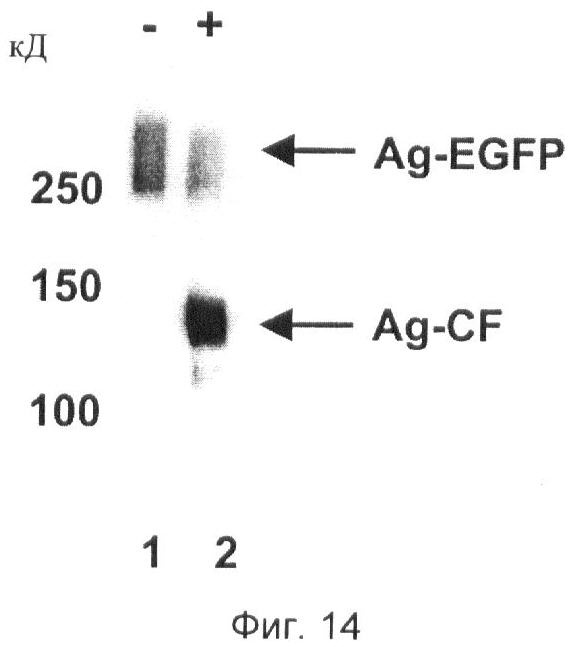
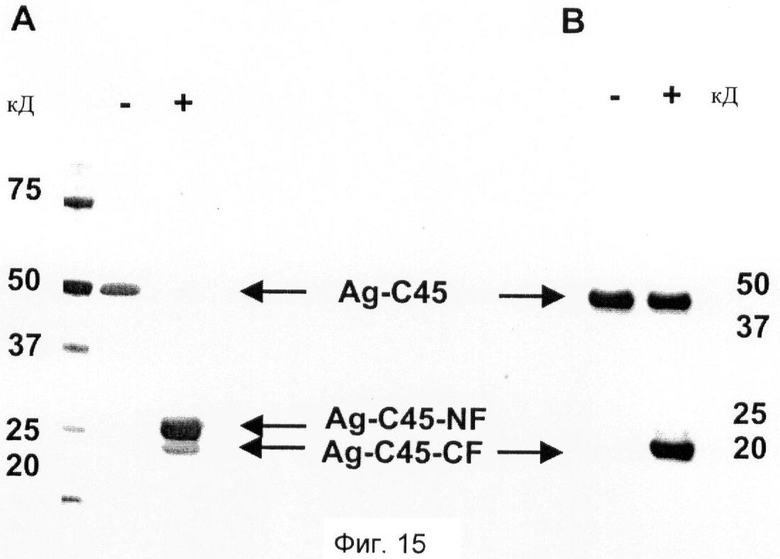
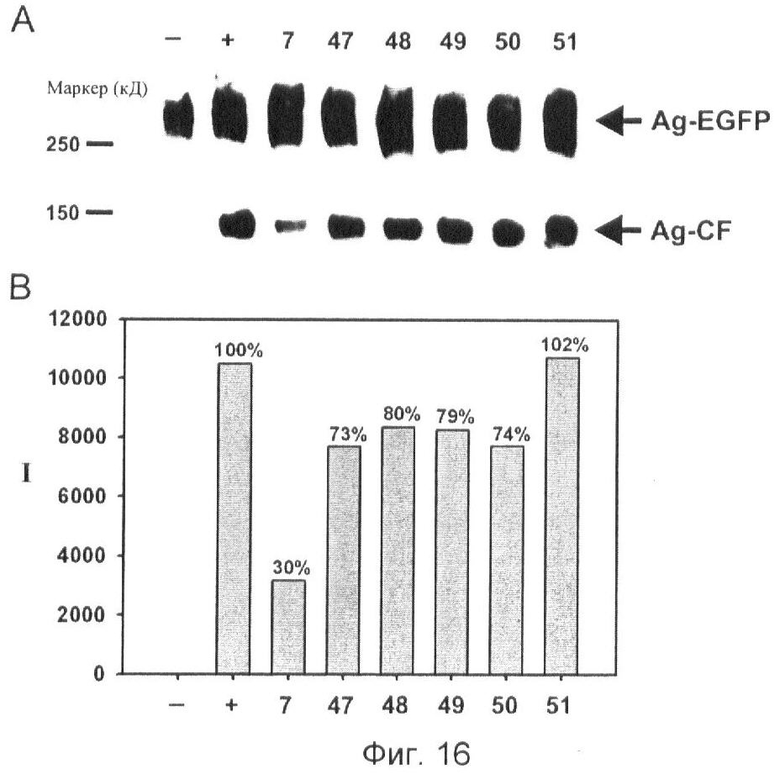
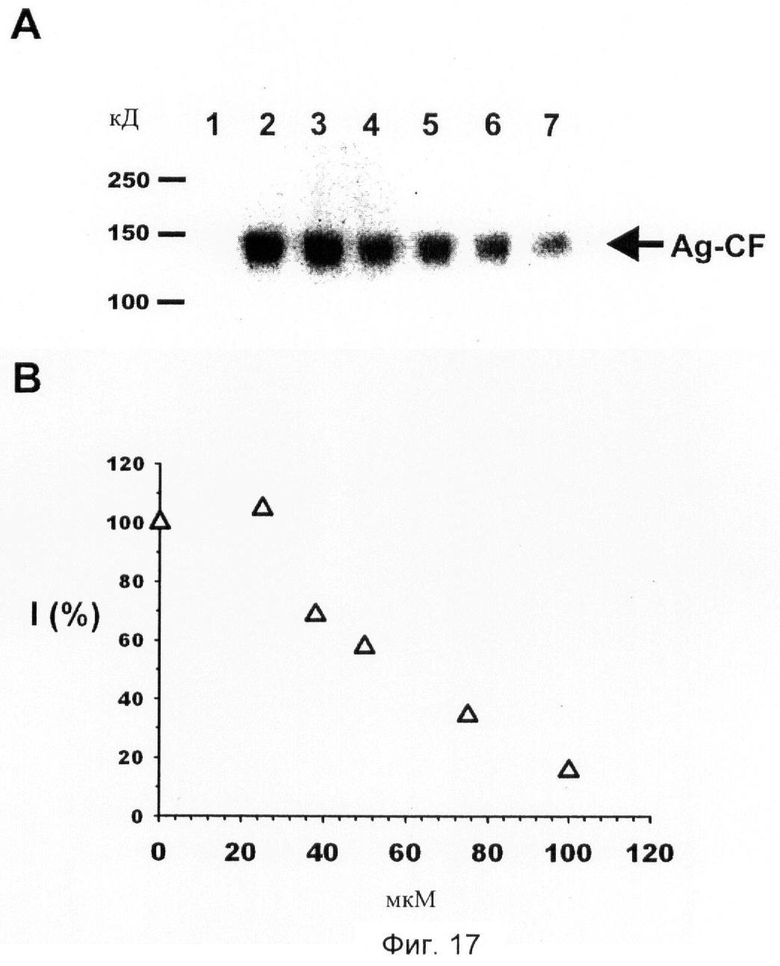
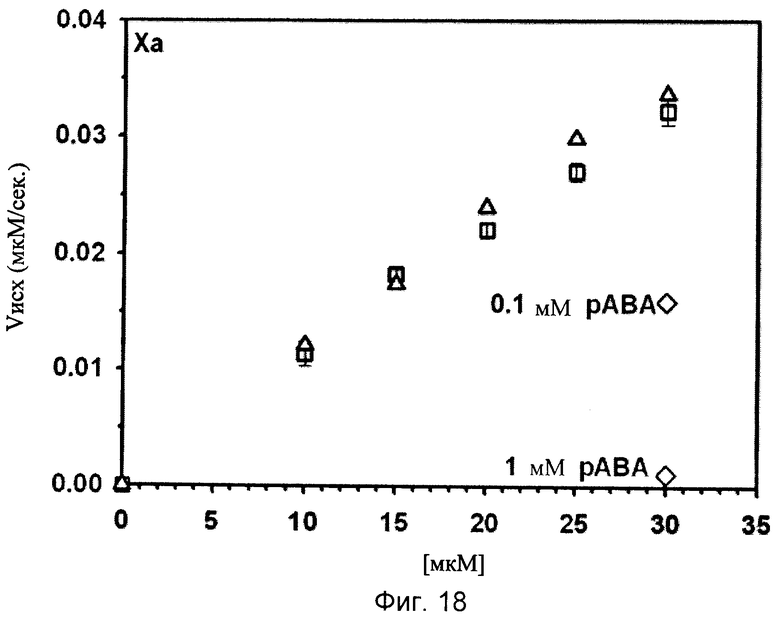
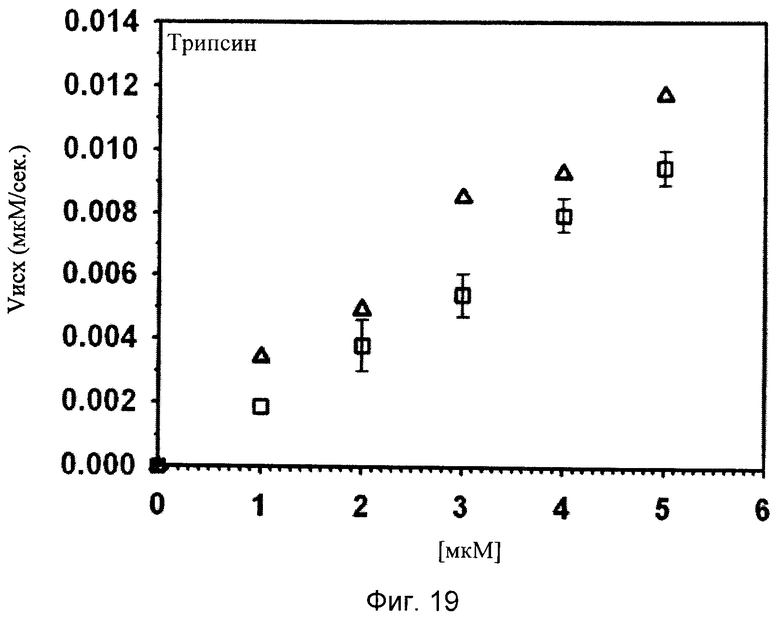
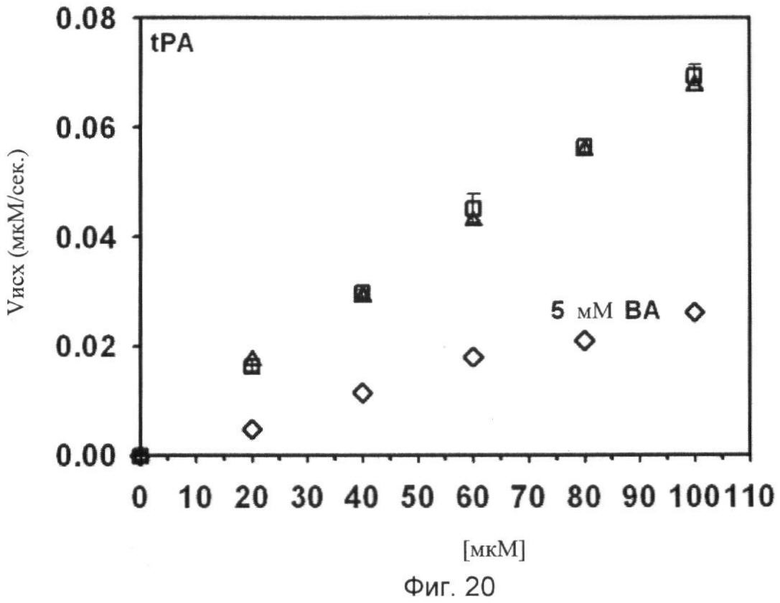
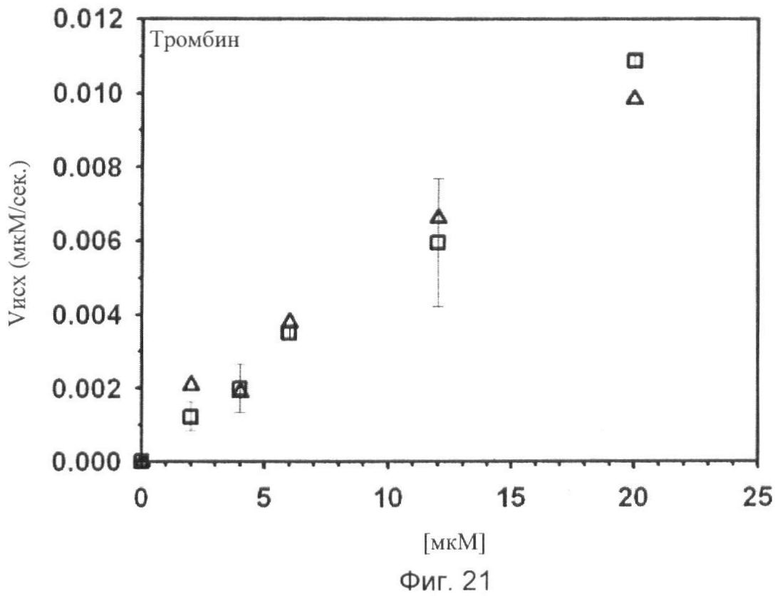
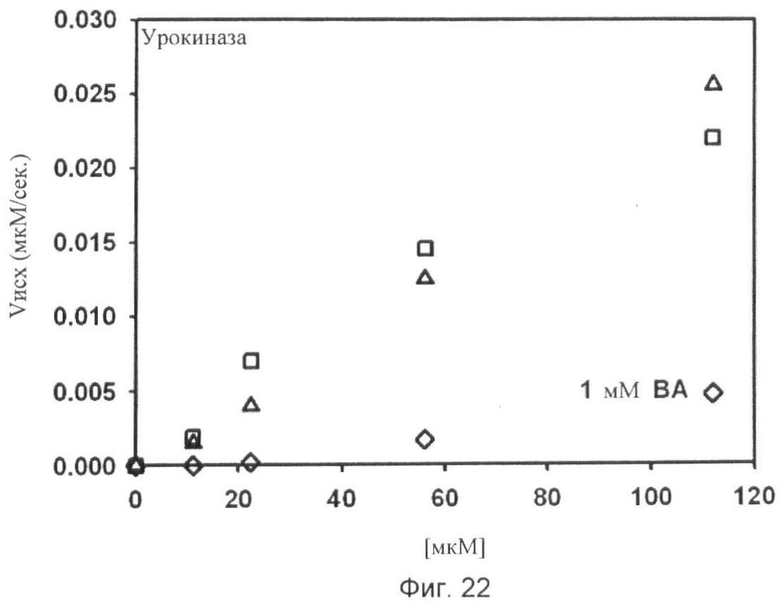
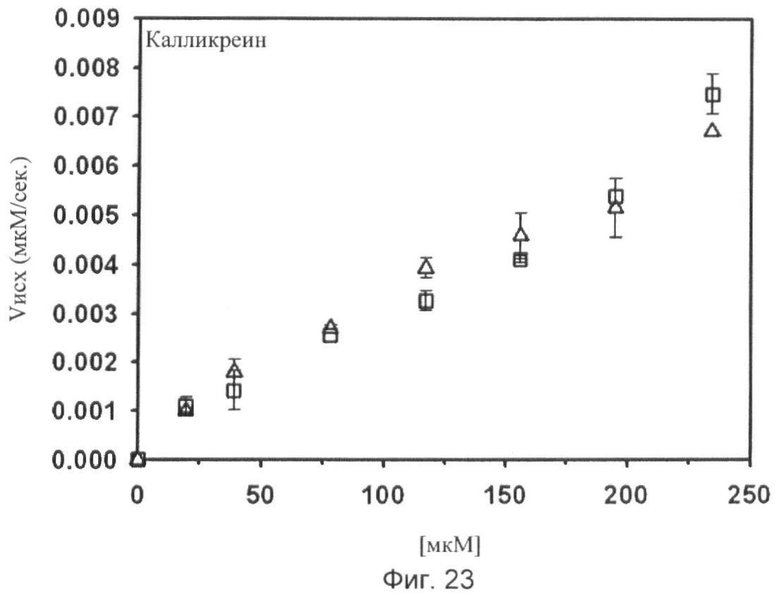
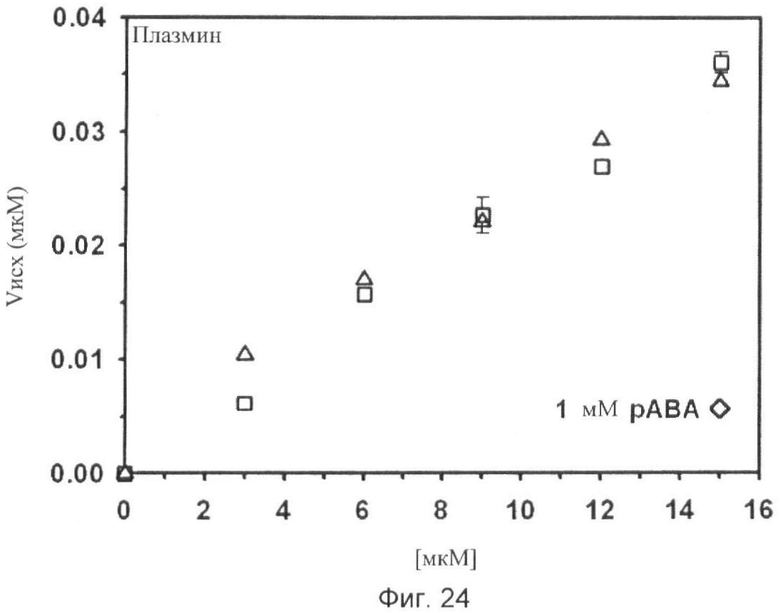
В изобретении описаны способ определения, является ли соединение ингибитором нейротрипсина, а также способ измерения каталитической активности нейротрипсина, где тестируемое соединение инкубируют вместе с человеческим или мышиным нейротрипсином или фрагментом человеческого или мышиного нейротрипсина, включающим домен протеазы нейротрипсина, и с полной длины или растворимым агрином, или агрином-С45, или флуоресцентным белком, или пептидом, включающим агрин, агрин-С45, в водном буферном растворе с рН 7,5 и измеряют количество расщепления агрина. 2 н. и 8 з.п. ф-лы, 24 ил., 1 табл.
1. Способ определения, является ли соединение ингибитором нейротрипсина, отличающийся тем, что соединение инкубируют вместе с человеческим или мышиным нейротрипсином или фрагментом человеческого или мышиного нейротрипсина, включающим домен протеазы нейротрипсина, и с полной длины или растворимым агрином, или агрином-С45, или флуоресцентным белком, или пептидом, включающим агрин, агрин-С45, в водном буферном растворе с рН 7,5 и измеряют количество расщепления агрина.
2. Способ по п.1, в котором используют человеческий нейротрипсин или его фрагмент, включающий домен протеазы человеческого нейротрипсина.
3. Способ по п.2, в котором используют человеческий нейротрипсин полной длины.
4. Способ по п.2, в котором используют фрагмент, включающий домен протеазы человеческого нейротрипсина.
5. Способ по любому из пп.1-4, в котором флуоресцентный белок или пептид, включающий агрин-С45, представляет собой белок, слитый с EGFP.
6. Способ по любому из пп.1-4, в котором используют флуоресцентный белок или пептид, включающий агрин полной длины.
7. Способ по любому из пп.1-4, в котором используют С-концевой фрагмент агрина-С45.
8. Способ измерения каталитической активности нейротрипсина, отличающийся тем, что человеческий или мышиный нейротрипсин или фрагмент человеческого или мышиного нейротрипсина, включающий домен протеазы нейротрипсина, инкубируют с полной длины или растворимым агрином, агрином-С45, или с флуоресцентным белком или пептидом, включающим агрин или агрин-С45 в водном буферном растворе с рН=7,5 и измеряют количество расщеплений агрина.
9. Способ по п.8, в котором флуоресцентный белок или пептид, включающий агрин-С45, представляет собой слитый белок с EGFP.
10. Способ по п.8, в котором используют С-концевой фрагмент агрина-С45.
| US 6783949 В1, 31.08.2004 | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Tsukamoto, Takeo и др | |||
| «Halosalicylaldehyde | |||
| I | |||
| Halosalicylaldehyde as reagent for primary amines», Kennosuke Yakugaku Zasshi, 78, 706-709, 1958 | |||
| Proba, К | |||
| et al | |||
| "Cloning and sequencing of the cDNA encoding human neurotrypsin", BIOCHIMICA ET BIOPHYSICA | |||

