По настоящей заявке испрашивается приоритет Китайской патентной заявки No.201910711540.3, поданной ранее в Китайское патентное ведомство 2 августа 2019 г., озаглавленной «A method for constructing an animal model and the use of the corresponding model», и Китайской патентной заявки No.202010038537.2, поданной ранее в Китайское патентное ведомство 14 января 2020 г., озаглавленной «Use of TPK as a target in Alzheimer’s disease», полное содержание которых приведено в настоящем описании посредством ссылки.
Область техники, к которой относится изобретение
Изобретение относится к области медицины, в частности, к применению TPK в качестве мишени при болезни Альцгеймера.
Уровень техники
Болезнь Альцгеймера (AD) является наиболее распространенным типом деменции, в основном характеризующимся хронической и прогрессирующей регрессией когнитивных функций и памяти, и изменением личности. К 2015 г., во всем мире было более 50 миллионов пациентов с AD, и заболеваемость увеличивалась с возрастом людей более 65 лет. Вместе с непрерывным развитием прогресса глобального старения, AD приводит к тяжелой экономической и ментальной нагрузке на общество и семьи.
AD представляет собой заболевание с различными патофизиологическими изменениями, включая потерю нейронов, активацию глиальных клеток, характерные сенильные бляшки, сформированные посредством внеклеточного накопления белка бета-амилоида (Aβ), и нейрофибриллярные клубки, вызванные гиперфосфорилированием внутриклеточного белка Tau и т.д. Кроме того, потеря синапсов, нарушения церебрального метаболизма глюкозы, окислительный стресс и т.п. также представляют собой постоянные патологические изменения в головном мозге при AD, и уменьшение церебрального метаболизма глюкозы у пациентов является тесно связанным с когнитивным нарушением.
Поскольку патогенез является неясным, все еще отсутствует эффективный способ лечения AD. Классическая «гипотеза каскада Aβ» всегда является наиболее популярной теорией для объяснения патогенеза AD. На этом основании, разработаны различные модели на трансгенных животных, и проведены эксперименты для разработки лекарственных средств, все из которых, однако, в настоящее время были остановлены на фазе II/III клинических исследований или потерпели неудачу. Другие новые лекарственные средства, такие как ингибирующие аномальное накопление белка Tau, также не достигли желательных результатов. Существует множество факторов, ограничивающих прогресс исследований применительно к механизму и лекарственному средству для AD, первым из которых является выбор мишени для лекарственного средства. AD представляет собой заболевание с различными патофизиологическими изменениями, лекарственное средство против одиночной мишени не может полностью остановить прогрессирование заболевания, и является необходимой разработка новых терапевтических мишеней. Кроме того, существующие модели на животных для исследования AD в основном разработаны на основании генетического фона, и чтобы быть более точными, они представляют собой модели гипотезы каскада Aβ, а не модели AD на животных, и не могут полностью симулировать множественные патофизиологические изменения в головном мозге при AD; и для лекарственных средств, подтвержденных как эффективные в существующих моделях на животных, не могли достигнуть ожидаемых результатов в клинических исследованиях. В настоящее время является неотложной проблемой нахождение подходящих моделей на животных для AD.
Сущность изобретения
В свете этого, конкретный вариант осуществления изобретения относится к применению TPK в качестве новой мишени в лечении или предотвращении болезни Альцгеймера, конкретные технические решения этого являются следующими:
Применение гена или белка TPK в качестве мишени в лечении или предотвращении болезни Альцгеймера.
Необязательно, с использованием гена или белка TPK в качестве мишени, стимулируют активность киназы и/или уровень экспрессии белка TPK в головном мозге.
Применение гена или белка TPK в качестве мишени в лекарственном средстве для предотвращения или лечения болезни Альцгеймера, или в скрининге или получении лекарственного средства.
Необязательно, лекарственное средство, с использованием гена или белка TPK в качестве мишени, стимулирует активность киназы и/или уровень экспрессии белка TPK в головном мозге.
Применение реагента, стимулирующего активность киназы и/или уровень экспрессии белка TPK в головном мозге, в получении лекарственного средства для предотвращения или лечения болезни Альцгеймера.
Необязательно, реагент, стимулирующий активность киназы и/или уровень экспрессии белка TPK в головном мозге, представляет собой реагент, регулирующий белок TPK и/или реагент, регулирующий ген TPK.
Лекарственное средство для предотвращения или лечения болезни Альцгеймера, которое, с использованием гена или белка TPK в качестве мишени, стимулирует активность киназы и/или уровень экспрессии белка TPK в головном мозге.
Применение гена или белка TPK в качестве мишени в конструировании модели на животных болезни Альцгеймера.
Способ конструирования модели на животных, где модель на животных конструируют посредством нокаута гена TPK у животного-мишени.
Необязательно, животное-мишень представляет собой грызуна.
Необязательно, грызун представляет собой мышь.
Необязательно, экзон 4 в гене TPK подвергают нокауту.
Необязательно, способ включает следующие стадии: конструирование нацеливающего вектора TPK-loxp для нокаута гена, введение нацеливающего вектора в животное-мишень для получения TPK-loxP+/+ животного-мишени.
Необязательно, последовательность нацеливающего вектора указана в SEQ ID NO. 1.
Необязательно, способ дополнительно включает стадии: скрещивания TPK-loxP+/+ животного-мишени с CamK2α-Cre/ERT2+/- животным-мишенью, получения CamK2α-Cre/ERT2+/-; TPK-loxP+/+ животного-мишени с условным нокаутом гена TPK.
Применение модели на животных, полученной посредством вышеуказанного способа, для конструирования модели на животных в исследовании нейродегенеративного заболевания с отличной от лечения заболевания целью.
Необязательно, нейродегенеративное заболевание представляет собой болезнь Альцгеймера.
Способ конструирования нацеливающего вектора для модели на животных, где бактериальную искусственную хромосому используют в качестве вектора, прямые последовательности loxP вставляют на обоих концах последовательности, гомологичной экзону гена TPK, вместе с последовательностью гена устойчивости к неомицину.
Необязательно, экзон представляет собой экзон 4.
Необязательно, последовательность нацеливающего вектора указана в SEQ ID NO. 1.
Нацеливающий вектор сконструирован посредством вышеупомянутого способа для конструирования нацеливающего вектора для модели на животных.
Изобретение относится к новой терапевтической мишени для AD, конкретный вариант осуществления изобретения может, посредством стимуляции активности киназы и/или уровня экспрессии белка TPK в головном мозге, лечить или предотвращать симптомы AD, вызванные посредством ингибирования на уровне белка TPK, такие как нарушение метаболизма тиамина, нарушение метаболизма глюкозы, вызывающие когнитивную регрессию, потерю нейронов, активацию глиальных клеток, дисфункцию синапсов, увеличенное накопление Aβ, увеличенное аномальное фосфорилирование белка Tau и т.д. Применительно к ограничению существующих моделей на животных для исследования AD и недостаткам современных моделей на животных с недостаточностью тиамина, способом конструирования модели на животных из конкретного варианта осуществления изобретения конструируют новую модель на животных с церебральной недостаточностью тиамина посредством нокаута гена TPK, в которой можно индуцировать снижение церебрального метаболизма глюкозы, вызывающее когнитивную регрессию, потерю нейронов, активацию глиальных клеток, дисфункцию синапсов, увеличенное накопление Aβ, увеличенное аномальное фосфорилирование белка Tau и т.д. Модель на животных может симулировать подобные AD хронические, множественные патофизиологические изменения, и может быть использована для исследования роли недостаточности TDP и нарушения метаболизма глюкозы при возникновении и развитии AD, так же как корреляции между различными патофизиологическими изменениями при AD, имеет очень важную научную значимость и практическую ценность для расширения исследования AD, изучения патогенеза AD, поиска новых молекулярных маркеров для диагностики заболевания и терапевтических мишеней для лекарственных средств, и т.д.
Краткое описание чертежей
Для более ясной иллюстрации примеров по изобретению или технических решений из предшествующей области техники, чертежи, предназначенные для использования в примерах, кратко представлены следующим образом.
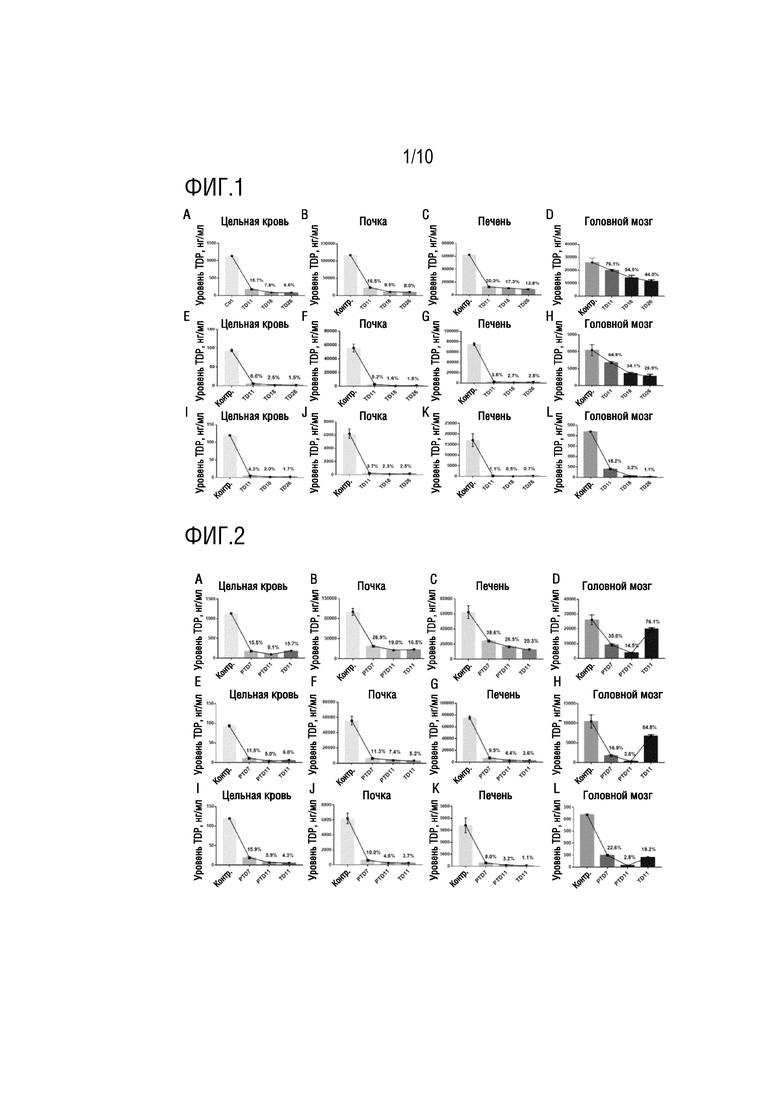
На фиг. 1 показано сравнение уровней TDP, TMP и TM в головном мозге, крови, печени и почке у мышей из модели TD в примере 1.
На фиг. 2 показано сравнение уровней TDP, TMP и TM в головном мозге, крови, печени и почке у мышей из модели PTD в примере 1.
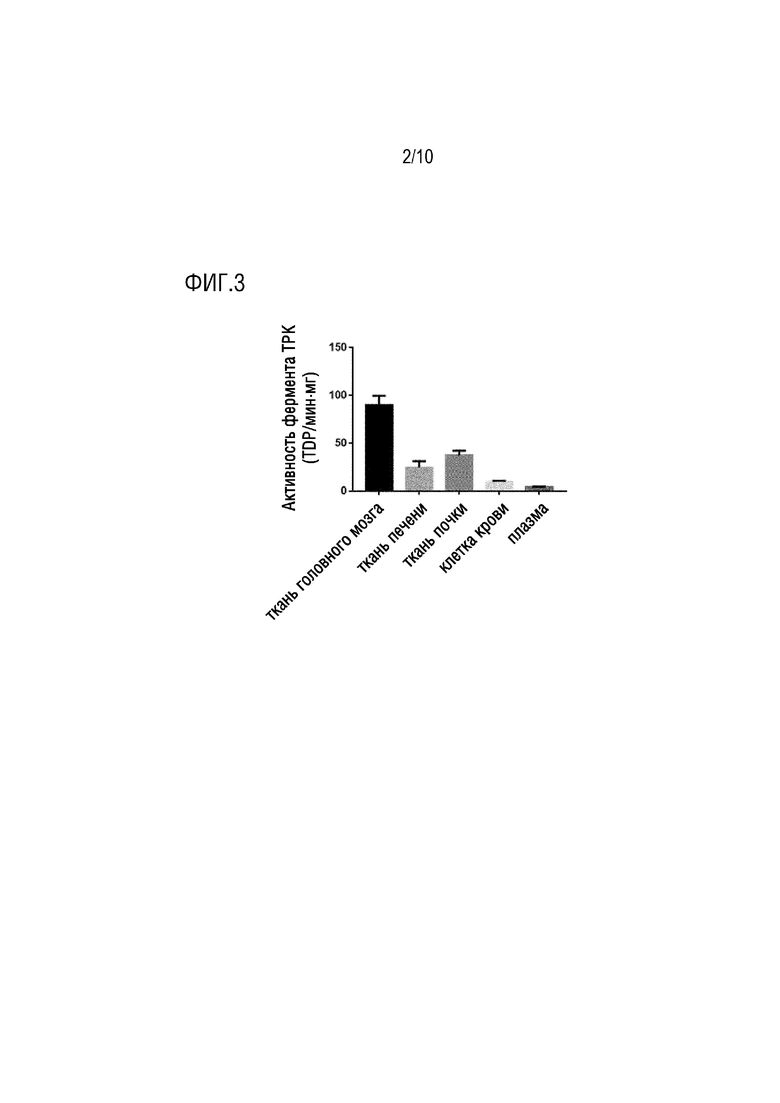
На фиг. 3 показано сравнение активности фермента TPK в головном мозге и крови, печени и почке у нормальных мышей в примере 1.
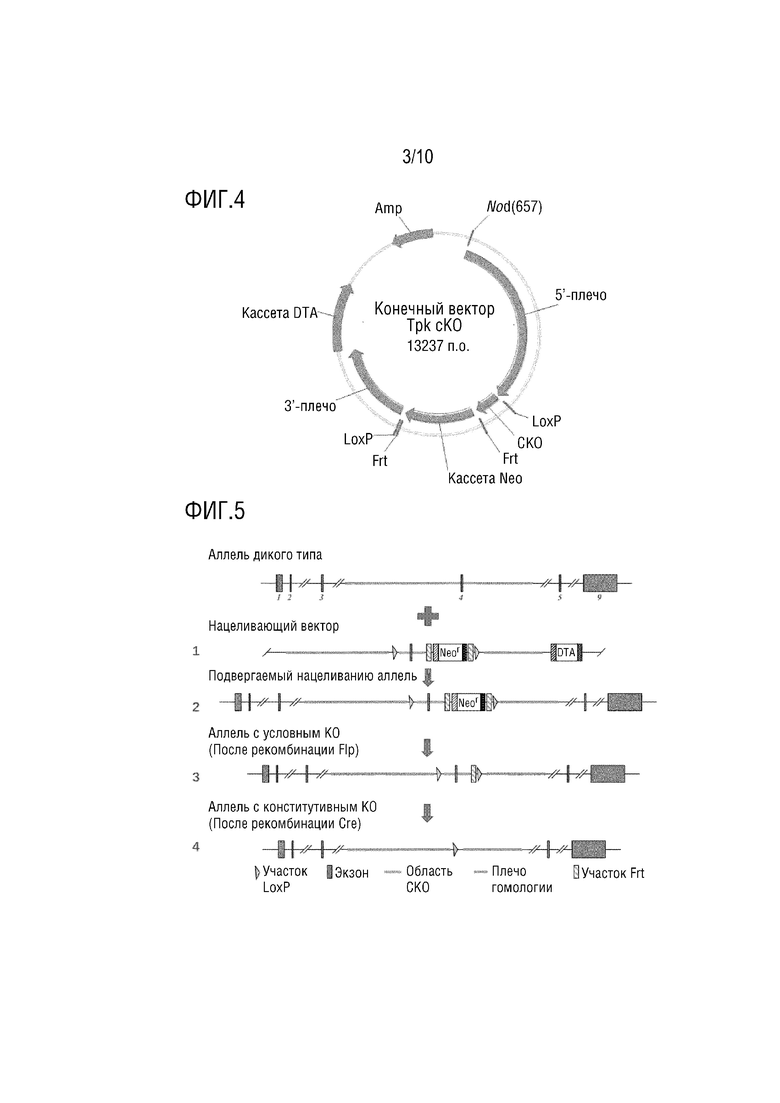
На фиг. 4 показана карта плазмиды для нацеливающего вектора, используемого в примере 2, на которой Amp представляет собой последовательность гена устойчивости к ампициллину, NotI представляет собой участок-мишень рестрикционной эндонуклеазы, 3’-плечо и 5’-плечо представляют собой гомологичные последовательности, фланкирующие экзон 4, кассета Neo представляет собой кодирующую последовательность гена устойчивости к неомицину, последовательность loxP предназначена для локализации и нацеливания на фрагмент, подлежащий нокауту, Frt представляет собой участок узнавания для Flp для разрезания последовательности Neo.
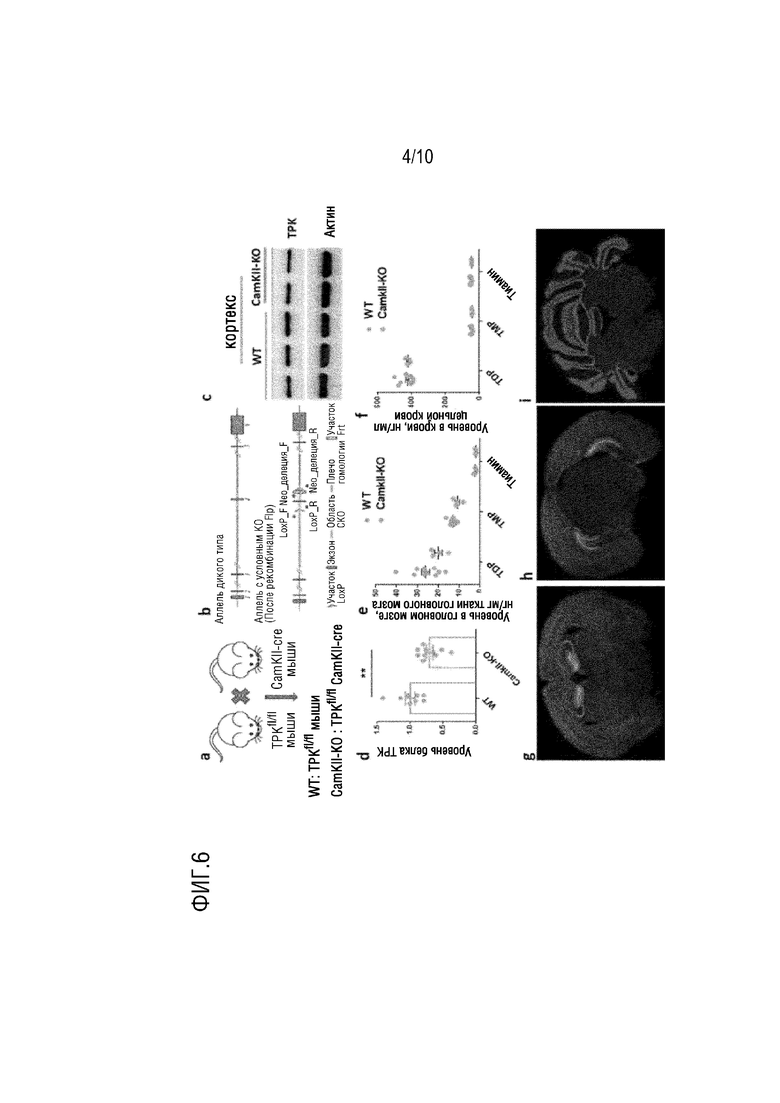
На фиг. 5 показана краткая блок-схема для получения CamK2α-Cre/ERT2+/-; TPK-loxP+/+ мышей в примере 2, в которой 1 представляет собой электротрансформацию полученным нацеливающим вектором эмбриональной стволовой клетки; 2 представляет собой отбор корректных положительных клонов; 3 представляет собой скрещивание с мышами Flp для удаления последовательности Neo и получения CamK2α-Cre/ERT2+/-; Tpk-loxP+/+ мышей.
На фиг. 6 показано влияние нокаута гена TPK на экспрессию TPK и TDP, в которой a показывает сравнительную модель на мышах в экспериментах, в которых мыши WT представляют собой TPKfl/fl мышей, т.е., TPK-loxP+/+ мышей, CamkII-KO мыши представляют собой CamK2α-Cre/ERT2+/-; Tpk-loxP+/+ мыши представляют собой мышей с нокаутом TPK; b показывает структуру гена с нокаутом TPK-flox мыши; c-d показывают сравнение уровней экспрессии фермента TPK у CamkII-KO мышей и мышей WT; e-f представляют собой сравнения уровней TDP, TMP и TM у CamkII-KO мышей и мышей WT; g-i показывают CamK2α-Cre/ERT2+/-, полученных посредством скрещивания CamK2α-Cre/ERT2+/- мышей с мышами Ai9 с репортером люциферазой; Cre в основном экспрессирован в кортексе и гиппокампе у мыши Ai9, но не в мозжечке и стволе головного мозга.
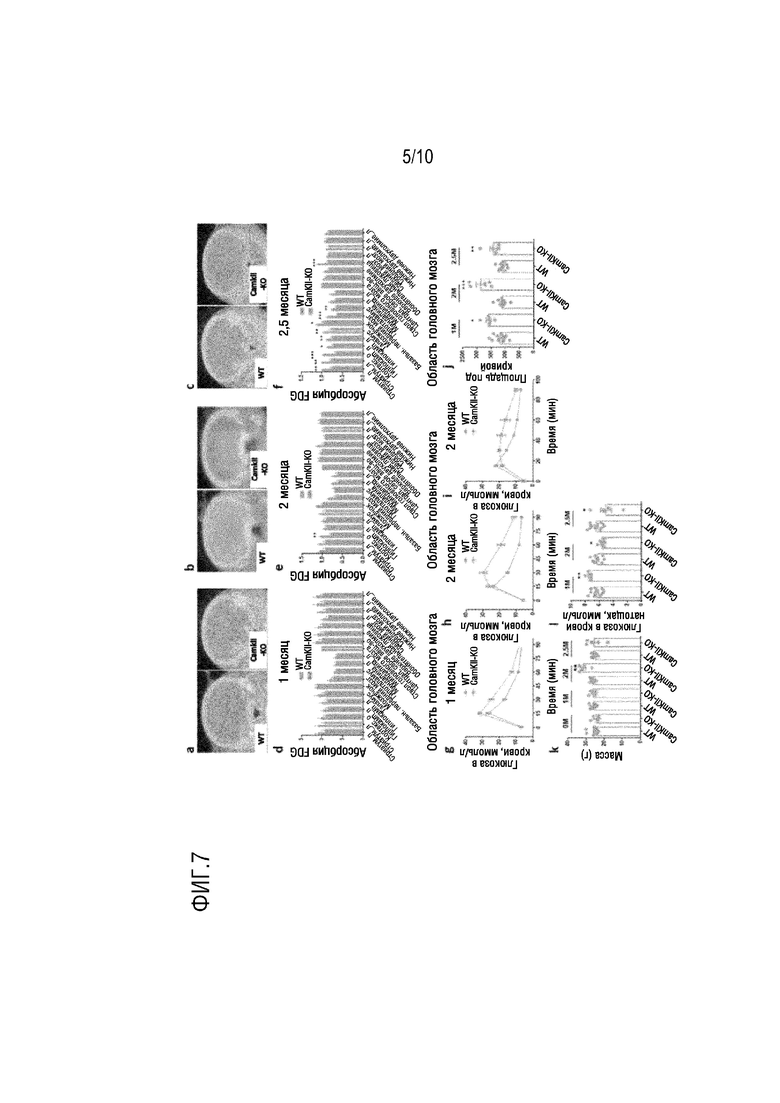
На фиг. 7 показаны результаты тестов FDG-PET, нарушенной толерантности к глюкозе, так же как изменений массы тела и уровня глюкозы в крови натощак, проведенных через 1, 2, 2,5 месяцев после индуцированного тамоксифеном нокаута гена TPK в примере 2, соответственно, в котором a-f представляют собой оценки FDG-PET изменений метаболизма глюкозы в областях головного мозга через 1, 2, 2,5 месяцев после индуцированного тамоксифеном нокаута гена TPK; g-j представляют собой нарушенную толерантность к глюкозе через 1, 2, 2,5 месяцев после индуцированного тамоксифеном нокаута гена TPK; k-l представляют собой изменения массы тела и уровня глюкозы в крови натощак через 1, 2, 2,5 месяцев после индуцированного тамоксифеном нокаута гена TPK.
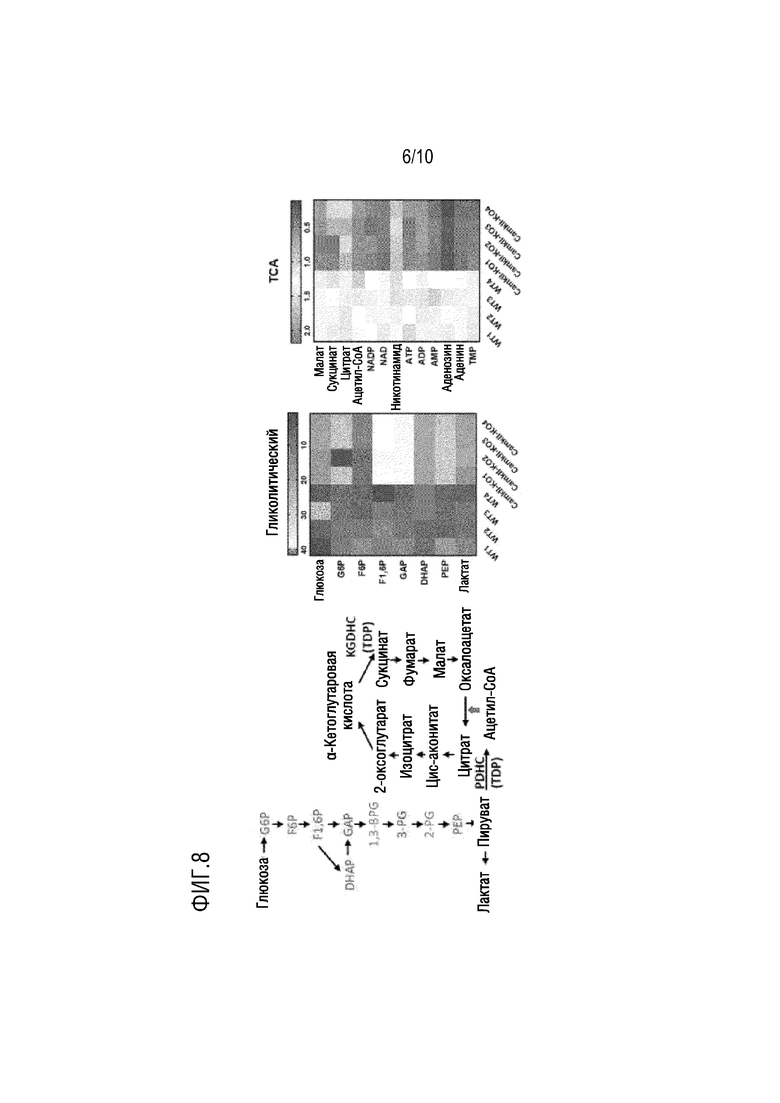
На фиг. 8 показаны результаты нарушения гликолиза и цикла трикарбоновых кислот после нокаута гена TPK в примере 2.
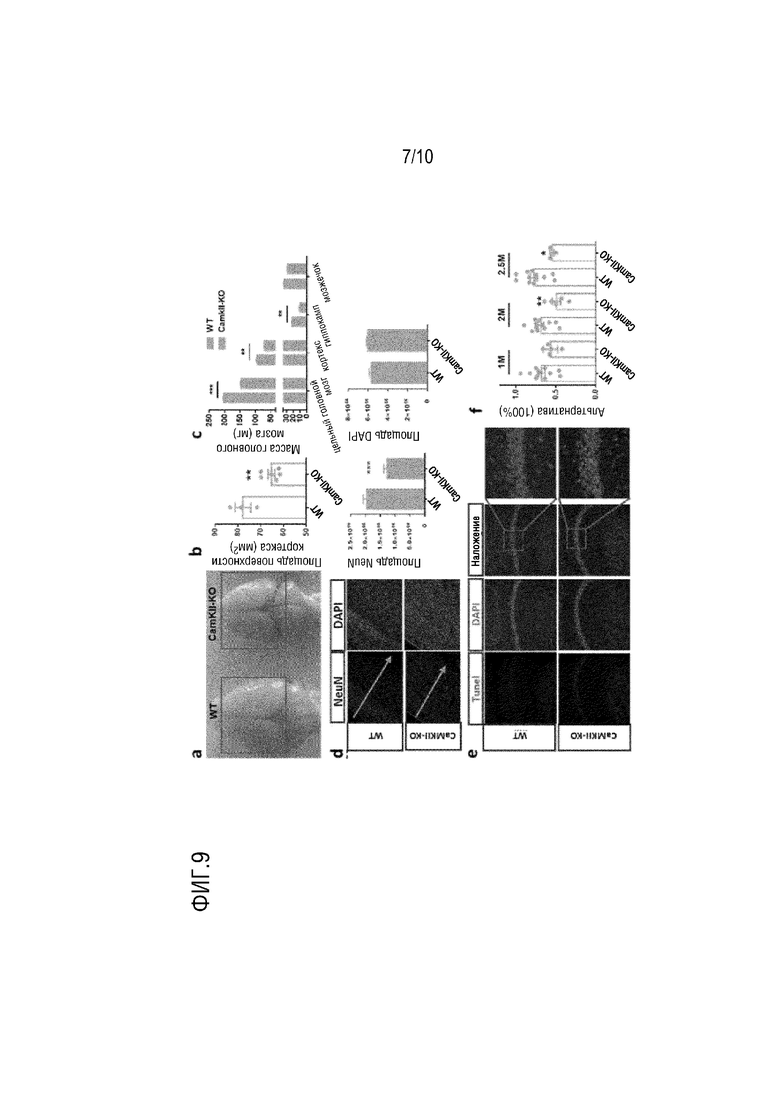
На фиг. 9 показаны результаты патологического тестирования до и после нокаута гена TPK в примере 2, в котором a-c показывают изменения в области площади поверхности кортекса и массы у мышей с нокаутом гена TPK; d показывает изменение в нейроне у мышей с нокаутом гена TPK; e показывает результат изменения положительных по Tunel клеток у мышей с нокаутом TPK; f показывает результаты оценки когнитивного нарушения в Y-образном лабиринте у мышей с нокаутом TPK.
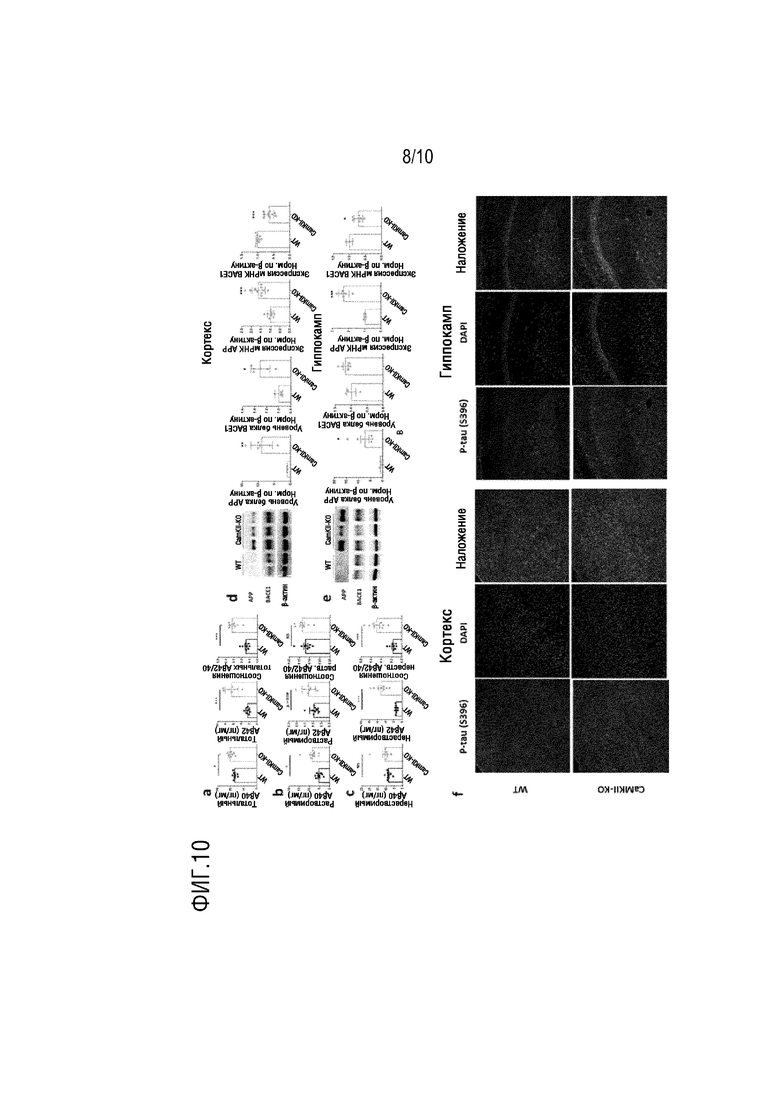
На фиг. 10 показаны результаты патологических изменений в Aβ и tau, вызванных до и после нокаута гена TPK в примере 2, в котором a-c показывают детекцию посредством ELISA уровней растворимого и нерастворимого Aβ40 и Aβ42, так же как соотношение Aβ40/Aβ42 до и после нокаута гена TPK; d-e показывают изменения в уровне экспрессии белка и уровне мРНК APP и Bacel в гиппокампе и кортексе до и после нокаута гена TPK при детекции посредством WB и RT-ПЦР; f показывает изменения уровня фосфорилированного Tau в кортексе и гиппокампе до и после нокаута гена TPK посредством детекции иммунофлуоресцентного окрашивания.
На фиг. 11 показан результат ответа нейровоспаления и дисфункции кровеносного сосуда, вызванного до и после нокаута гена TPK в примере 2, в котором a-d показывают активацию и пролиферацию клеток микроглии и астроцитов в кортексе и гиппокампе мыши до и после нокаута гена TPK посредством IHC детекции; e-f показывают уровни мРНК Ibal и GFAP в гиппокампе и кортексе мыши до и после нокаута гена TPK при детекции посредством RT-ПЦР; g-j показывают морфологию кровеносного сосуда и подтекание IgG из кровеносного сосуда в кортексе и гиппокампе мыши до и после нокаута гена TPK при детекции посредством иммунофлуоресцентного окрашивания.
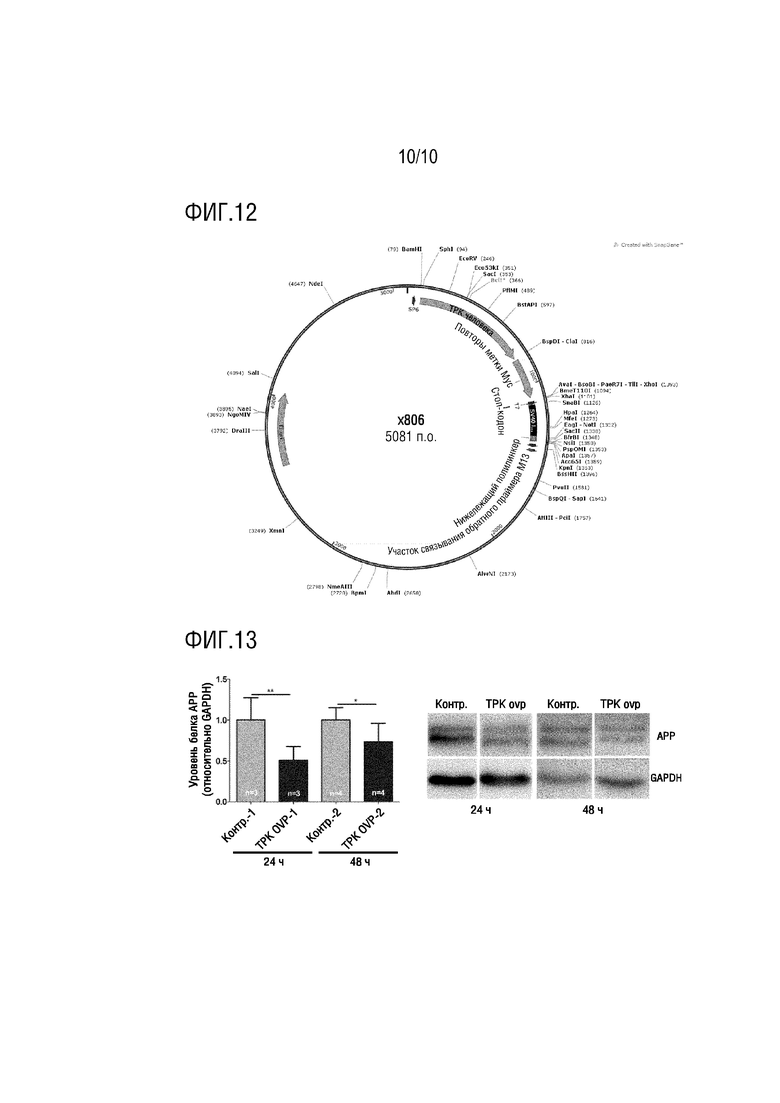
На фиг. 12 показана плазмида для сверхэкспрессии TPK в примере 3.
На фиг. 13 показаны результаты детекции посредством WB для плазмиды для сверхэкспрессии TPK и ее контрольной группы на линии клеток HEK293-APP через 24 час и 48 час в примере 3.
Конкретные варианты осуществления
Пациенты с AD имеют множество каскадных патофизиологических изменений в головном мозге, где нарушение церебрального метаболизма глюкозы является их постоянным патофизиологическим признаком, который является тесно связанным с когнитивной регрессией, и появляется за много лет до клинических симптомов. Глюкоза является первичным энергетическим субстратом для клеток в головном мозге, и промежуточные продукты метаболизма глюкозы также предоставляют субстраты для синтеза нейротрансмиттеров. Церебральный метаболизм глюкозы в основном включает два процесса, которые представляют собой трансмембранный транспорт и внутриклеточный метаболизм глюкозы, соответственно. Нарушение внутриклеточной утилизации глюкозы у пациентов с AD в основном вызвано уменьшенной активностью ключевых ферментов в метаболизме глюкозы, т.е., уменьшенной активностью транскетолазы, пируватдегидрогеназы и α-кетоглутаратдегидрогеназы. Тиаминдифосфат (TDP), первичная активная форма тиамина, является общим коферментом для этих трех ключевых ферментов и играет важную роль в метаболизме глюкозы. Уменьшение активности трех вышеуказанных ключевых ферментов и значительная потеря TDP являются специфическими и универсальными у пациентов с AD, которые не присутствуют у пациентов с сосудистой деменцией, лобно-височной деменцией и болезнью Паркинсона. Тиаминпирофосфаткиназа (TPK) является не только ключевым ферментом для синтеза TDP, но также критическим фактором для поддержания гомеостаза уровня TDP в головном мозге. Авторы настоящего изобретения, посредством дополнительных исследований, обнаружили, что снижение церебрального метаболизма глюкозы, вызванное недостаточностью TDP, является ассоциированным с возникновением AD, и AD является ассоциированным с экспрессией белка TPK. Кроме того, ингибирование уровня белка TPK в головном мозге посредством нокаута гена TPK может явно приводить к нарушению метаболизма тиамина, нарушению метаболизма глюкозы, и более важно, может неожиданно индуцировать подобное AD нейродегенеративное заболевание, энцефалатрофию, патологические изменения в Aβ и tau, нейровоспаление и нейроваскулярное нарушение. Таким образом, ген или белок TPK играет очень важную роль при AD, и таким образом, является новой мишенью для лечения и предотвращения AD. AD, вызванную посредством ингибирования белка TPK, можно предотвращать посредством стимуляции активности киназы и/или уровня экспрессии белка TPK в головном мозге с геном или белком TPK в качестве мишени, таким образом, предотвращения. Модель на животных, полученную с использованием способа конструирования модели на животных посредством нокаута гена TPK у животного-мишени, можно использовать в исследовании нейродегенеративных заболеваний, в частности, исследовании болезни Альцгеймера, включая исследования с целью лечения заболевания и с отличной от лечения заболевания целью. Исследования с отличной от лечения заболевания целью в модели на животных могут конкретно включать, например, 1) исследование и/или изучение механизма нейродегенеративных заболеваний; 2) получение и/или скрининг продуктов, способных лечить нейродегенеративные заболевания; 3) поиск и подтверждение молекулярных маркеров, способных диагностировать и/или предполагать нейродегенеративные заболевания.
В конкретном варианте осуществления изобретения, стимуляция уровня экспрессии белка TPK в головном мозге включает, но без ограничения, предотвращение ингибирования экспрессии белка TPK, увеличение уровня экспрессии белка TPK, увеличение активности белка TPK или увеличение стабильности белка TPK.
В конкретном варианте осуществления изобретения, реагент, стимулирующий уровень экспрессии белка TPK в головном мозге, может представлять собой реагент, регулирующий белок TPK, и может также представлять собой реагент, регулирующий ген TPK. Реагент включает, но без ограничения, ускоритель, агонист, активатор для стимуляции уровня экспрессии белка TPK в головном мозге. Реагент может представлять собой соединение, химическую малую молекулу, биомолекулу и т.п., конкретно, например, реагенты, регулирующие белок TPK, такие как связывающий TPK белок, химический агонист для TPK или ферментов для модификации TPK и т.п., реагенты, регулирующие ген TPK, такие как вещество, активирующие репликацию или транскрипцию гена TPK, вещество, увеличивающее экспрессию гена TPK, осуществляющее повышающую регуляцию средство, стимулирующее промотор гена TPK, белок для специфической сверхэкспрессии гена TPK.
В конкретном варианте осуществления изобретения, лекарственное средство может содержать безопасную и эффективную дозу реагента, стимулирующего уровень экспрессии белка TPK в головном мозге, и фармацевтически приемлемый носитель. Фармацевтический препарат обычно соответствует введению, лекарственная форма препарата может представлять собой препарат для инъекции, пероральный препарат (таблетку, капсулу, пероральный раствор), чрескожный препарат или препарат с замедленным высвобождением.
В конкретном варианте осуществления изобретения, животное-мишень представляет собой не относящееся к человеку животное, включая примата или грызуна и т.д., и кроме того, мышь. Экзон 4 в гене TPK подвергают нокауту в этом способе, для мыши, ген TPK имеет всего 9 экзонов, ATG локализован в экзоне 2, терминирующий кодон локализован в экзоне 9, и экзон 4 выбирают в качестве мишени для нокаута, с одной стороны, подвергнутая нокауту последовательность экзона не является целым кратным трем, и является способной приводить к мутациям сдвига рамки считывания в нижележащей рамке считывания; с другой стороны, чем ближе выбранный экзон локализован к участку инициации трансляции, тем меньшая функция гена может быть сохранена.
Конкретный вариант осуществления изобретения, в котором модифицированную систему рекомбиназы Cre-loxP можно использовать для получения модели на животных для условного нокаута гена TPK, включает стадии: конструирования нацеливающего вектора для нокаута гена TPK-loxP, введения нацеливающего вектора в животное-мишень для получения TPK-loxP+/+ животного-мишени, скрещивания TPK-loxP+/+ животного-мишени с CamK2α-Cre/ERT2+/- животным-мишенью, получения CamK2α-Cre/ERT2+/-; TPK-loxP+/+ животного-мишени с условным нокаутом гена TPK. В частности, бактериальную искусственную хромосому можно использовать в качестве вектора, и прямые последовательности loxP и последовательность гена устойчивости к неомицину можно вставлять фланкирующими гомологичную последовательность подходящего экзона; нацеливающим вектором трансформируют эмбриональную стволовую (ES) клетку посредством электротрансформации; клетку ES, трансформированную с использованием корректной последовательности, локализованной в корректном участке, идентифицируют посредством скрининга по устойчивости к антибиотику, ПЦР и Саузерн-блоттинга; положительный клон микроинъецируют в бластоцисту животного-мишени; и бластоцисту после успешной инъекции имплантируют в матку животного-мишени для получения химерного животного-мишени с интегрированной клеткой ES; полученное химерное животное-мишень скрещивают с животным-мишенью с инструментом Flp, последовательность гена устойчивости к неомицину вырезают посредством индукции, и TPK-loxP+/+ животное-мишень с потенциалом к нокауту можно получать после очистки; животное-мишень с инструментом Cre с CamK2α в комбинации с мутантным рецепторным элементом эстрогена (LBD-ER) отбирают для скрещивания с вышеуказанным TPK-loxP+/+ животным-мишенью; и после идентификации, получают имеющее стабильное наследование CamK2α-Cre/ERT2+/-; TPK-loxP+/+ животное-мишень.
В конкретном варианте осуществления изобретения, настоящее изобретение относится к способу конструирования нацеливающего вектора для модели на животных, в котором бактериальную искусственную хромосому используют в качестве вектора, и прямые последовательности loxP вставляют на обоих концах последовательности, гомологичной экзону гена TPK, вместе с последовательностью гена устойчивости к неомицину.
В конкретном варианте осуществления способа конструирования нацеливающего вектора для модели на животных по изобретению, экзон представляет собой экзон 4, и конструируют следующую последовательность-мишень: «5’-гомологичное плечо-loxP-экзон 4-frt-Neo-frt-loxP-гомологичное плечо-3’».
В конкретном варианте осуществления способа конструирования нацеливающего вектора для модели на животных по изобретению, последовательность нацеливающего вектора указана в SEQ ID NO. 1.
В конкретном варианте осуществления, настоящее изобретение относится также к нацеливающему вектору для модели на животных, полученному посредством способа конструирования нацеливающего вектора для вышеуказанной модели на животных.
Изобретение далее проиллюстрировано в связи со следующими примерами.
Пример 1
Эксперименты проводили в простой модели диеты с недостаточностью тиамина (TD) и диеты с недостаточностью тиамина в комбинации с моделью инъекции пиритиамина (PTD), соответственно. Отслеживали динамические изменения в моделях TD и PTD. Шесть групп (n=4), включая TD 11 суток, 18 суток, 26 суток и PTD 7 суток, 11 суток, организовали для сравнения. Выбирали мышей C57 в возрасте 3 месяцев, рожденных в одни и те же сутки и имеющих сходную массу тела. Различные временные точки для начала получения моделей выбирали в зависимости от группировки, и получения моделей окончательно завершали на одни и те же сутки, так же как отбор образцов. Модели TD: получение моделей в течение 11 суток, 18 суток и 26 суток, соответственно, с использованием нормальной питьевой воды и диеты с недостаточностью тиамина; модели PTD: получение моделей в течение 7 суток и 11 суток, соответственно, с использованием нормальной питьевой воды, диеты с недостаточностью тиамина, и ежесуточной внутрибрюшинной инъекции 0,1 мг/мл PT (пиритиамина) в дозе 500 мкг/(кг∙сут.); контрольная группа: нормальная питьевая вода, нормальный корм, и внутрибрюшинная инъекция нормального солевого раствора. После завершения получения моделей, мышей подвергали анестезии посредством ингаляции изофлурана, глаза извлекали, образцы крови отбирали в пробирки с антикоагулянтом гепарином, которые затем быстро переворачивали для перемешивания. Равный объем 7,5% PCA добавляли к 150 мкл цельной крови, что перемешивали на встряхивателе. Мышей быстро умерщвляли посредством смещения шейных позвонков на льду, и отбирали образцы тканей головного мозга, почки, печени и т.п., и взвешивали. Массу тканей регистрировали. Ткани смешивали с KH2PO4 в соотношении ткань (мг):KH2PO4 (мкл)=100:900, измельчали в течение 90 с использованием измельчителя ткани, при 70 Гц, подвергали воздействию ледяной бани после полного измельчения в течение 30 мин, и сохраняли при -80°C до использования. Уровни TDP, TMP и TM (тиамина) определяли посредством высокоэффективной жидкостной хроматографии. В моделях TD и PTD, сравнения активности TDP, TMP и TM в головном мозге, крови, печени и почке у мышей показаны на фиг. 1 и фиг. 2, и сравнение активности TPK в головном мозге, крови, печени и почке у нормальных мышей показано на фиг. 3.
В соответствии с вышеуказанными экспериментами, обнаружено на моделях TD, что, по сравнению с кровью, печенью и почкой, признак значимо более медленного уменьшения уровня TDP проявлялся в головном мозге, указывая на защиту против потери TDP в головном мозге (см. фиг. 1). В моделях PTD, PT, специфический ингибитор для TPK, может нарушать гомеостаз TDP в организме, особенно уровень TDP в головном мозге (см. фиг. 2). Обнаружено посредством дополнительной детекции активности тиаминпирофосфокиназы (TPK) в различных тканях нормальных мышей, что активность TPK являлась наивысшей в головном мозге (фиг. 3), и таким образом, можно предположить, что высокая активность TPK, специфическая для ткани головного мозга, являлась ключевой для поддержания гомеостаза TDP в головном мозге.
Пример 2
1. Конструирование нацеливающего вектора
Бактериальную искусственную хромосому (BAC) использовали в качестве вектора, и последовательность-мишень «5’-гомологичное плечо-loxP-экзон 4-frt-Neo-frt-loxP-гомологичное плечо-3’», сконструированную с использованием интронов, фланкирующих экзон 4, в качестве гомологичных плеч, вставляют для конструирования нацеливающего вектора для нокаута гена (см. фиг. 4).
2. Конструирование CamK2α-Cre/ERT2+/-; TPK-loxP+/+ мышей (см. фиг. 5)
Получение и скрининг TPK-floxP+/- эмбриональных стволовых клеток
(1) Получение эмбриональных стволовых клеток (ES) мышей
Первичные эмбриональные фибробласты мыши, в форме трофобластов, инокулировали в чашки, обработанные 0,1% желатином на сутки перед сутками размораживания клеток ES, и культивировали в культуральном бульоне с LIF, добавленным в концентрации 1000 ед./мл.
(2) Электротрансформация клеток ES
1. Клетки ES расщепляли панкреатином, с последующим ресуспендированием в PBS (приблизительно 2*10^7/мл), и помещали на лед;
2. Линеаризованный нацеливающий вектор (45 мкг) смешивали с 1 мл клеток, и загружали в резервуар для электропорации, и электротрансформировали с параметрами 600 В, 25 мкФ, 10 мс;
3. Клетки после электротрансформации инокулировали в чашки с трофобластами с полной конфлюэнтностью, и помещали в инкубатор при 37°C, бульон для скрининга клеток, содержащий G418 (280 мкг/мл), добавляли для замены через 24 час.
(3) Отбор и амплификация положительного клона ES
Отбор клонов начинали через несколько суток культивирования, отдельные недифференцированные клоны ES отбирали под микроскопом с небольшим увеличением и помещали в 96-луночный планшет для последующего культивирования; каждый клон разделяли на два, один подвергали криоконсервированию, и другой переносили в 24-луночный планшет для амплификации и пассирования.
(4) Идентификация подвергнутых нацеливанию клеток ES посредством ПЦР и Саузерн-блоттинга
Положительные клоны, устойчивые к G418, отбирали и амплифицировали, и геномную ДНК выделяли, ферментно расщепляли, и анализировали полосы после электрофореза в агарозном геле. Затем Саузерн-блоттинг проводили с использованием зондов, нацеленных на 5’- и 3’-концы экзона 4 Tpk, идентификацию проводили в соответствии с различиями между полосами между контрольными и подвергнутыми нацеливанию клетками ES. Где,
праймеры для идентификации посредством ПЦР представляли собой:
Праймеры для получения 5’- и 3’-зондов для Саузерн-блоттинга представляли собой:
Конструирование TPK-floxP химерной мыши посредством микроинъекции бластоцисты
(1) Получение бластоцисты мыши
Отбирали самок мышей C57BL/6 в возрасте 4 недель, и внутрибрюшинно инъецировали с использованием 10 IU гонадотропина в полдень. 10 IU хорионического гонадотропина человека инъецировали через 48 час, и самок мышей помещали в клетки вместе с самцами мышей. На утро следующих суток, у самок мышей проверяли копулятивные пробки (регистрировали как 0,5 суток), и самок мышей с копулятивными пробками умерщвляли через 3 суток, и матки извлекали, и бластоцисты вымывали с использованием бульона BMOC-3. Бластоцисты переносили в капли бульона, накапанные на чашку 60 мм, которые покрывали минеральным маслом. Микроинъекцию проводили после размножения бластоцеля.
(2) Микроинъекция бластоцисты
Подготавливали удерживающие пипетки и инъекционные пипетки для микроинъекции. Свежий бульон добавляли для замены к клеткам ES для инъекции за 3 час до инъекции. Суспензию отдельных клеток получали при расщеплении панкреатином. Небольшие и круглые клетки ES с блестящей поверхностью отбирали для инъекции, каждый эмбрион инъецировали с использованием 12~15 клеток ES, инъецированные бластоцисты, как правило, восстанавливали свою нормальную морфологию после 1 час культивирования, бластоцисты, имеющие интактную морфологию, отбирали для трансплантации.
(3) Трансплантация бластоцисты
Получение псевдобеременных мышей: самок мышей белой породы Kunming скрещивали с подвергнутыми вазолигированию самцами мышей и проверяли по образованию копулятивных пробок на следующие сутки (регистрировали как 0,5 суток). Псевдобеременных мышей можно использовать для трансплантации эмбриона через двое суток. Псевдобеременных мышей подвергали анестезии с использованием изофлурана, операцию на спинном отделе проводили для переноса 10 подвергнутых инъекции бластоцист в одну сторону матки. Если хирургическая трансплантация была успешной, новорожденные мыши могут быть рождены через 17 суток, возможно визуально определять, в соответствии с цветом шерсти через несколько суток, были ли получены TPK-floxP химерные мыши с интегрированными подвергнутыми нацеливанию ES клетками, и степень химерности оценивали в соответствии с соотношением коричневой шерсти ко всей шерсти.
Получение TPK-loxP+/+ химерной мыши
Полученных TPK-floxP химерных мышей скрещивали с мышами с инструментом Flp, фермент инструмента Flp может индуцировать расщепление последовательности Neo в участке Frt, и Tpk-loxP+/+ гомозиготных мышей с потенциалом к нокауту можно получать после очистки.
Получение CamK2α-Cre/ERT2+/-; Tpk-loxP+/+ мышей
(1) Tpk-loxP+/+ мышей получали посредством вышепомянутой операции и подтверждали возможность достижения нокаута Tpk;
(2) Tpk-loxP+/+ мышей скрещивали с CamK2α-Cre/ERT2+/- мышами для получения CamK2α-Cre/ERT2+/-; Tpk-loxP+/+ мышей, которых генетически идентифицировали, и затем разводили.
Праймеры для идентификации:
Фенотип мышей, имеющих условный нокаут гена TPK
Нокаута гена TPK специфически в кортексе и гиппокампе достигали посредством скрещивания с CamkII-Cre/ERT2+/- мышами, и мышам в возрасте 3 месяцев внутрибрюшинно инъецировали тамоксифен для индукции нокаута гена TPK. Проводили фенотипирование мышей с нокаутом гена TPK, результаты которого показаны на фиг. 6-11.
Со ссылкой на фиг. 6, церебральный уровень белка TPK и уровень TDP можно было значимо уменьшать через 2,5 месяца после индукции тамоксифеном нокаута гена TPK, без влияния на уровень тиамина. Со ссылкой на фиг. 7, FDG-PET проводили через 1, 2, 2,5 месяцев после индукции тамоксифеном нокаута гена TPK, соответственно, обнаружено посредством анализа глюкозы в крови и теста толерантности к глюкозе, что мыши с нокаутом TPK имели значительное нарушение церебрального метаболизма глюкозы, дисгомеостаз периферической глюкозы и нарушение толерантности к глюкозе. Со ссылкой на фиг. 8, исследование метаболома показало, что нокаут гена TPK вызывал нарушение гликолиза и цикла трикарбоновых кислот. Со ссылкой на фиг. 9, патологическая детекция показала, что нокаут гена TPK может значительно вызывать церебральную атрофию, большую потерю синапсов и нейронов, увеличение количества положительных по Tunel клеток, конденсацию хромосом и кариопикноз. Со ссылкой на фиг. 10, патологические исследования Aβ и tau, которые являются необходимыми для AD, показали, что нокаут гена TPK может значимо увеличивать продукцию Aβ40 и Aβ42, увеличения в основном достигали посредством увеличения экспрессии APP и уровня белка Bace1, и увеличения фосфорилирования белка Tau. Со ссылкой на фиг. 11, воспалительный ответ играл важную роль в возникновении и развитии AD, и также обнаружено что присутствовали значительная активация астроцитов и микроглиальных клеток и увеличенная экспрессия фактора воспаления в кортексе и гиппокампе головного мозга у мышей с нокаутом гена TPK; в то же время, нокаут гена TPK приводил к нейроваскулярному нарушению и увеличенной проницаемости сосудов. В заключение, нокаут гена TPK, ключевого фермента для регуляции гомеостаза TDP в головном мозге, может приводить к значительному нарушению метаболизма тиамина, нарушению метаболизма глюкозы, и более важно, может неожиданно индуцировать подобное AD нейродегенеративное заболевание, энцефалатрофию, патологические изменения в Aβ и tau, нейровоспаление и нейроваскулярное нарушение.
Пример 3
Информация об антителах в этом примере являлась следующей:
Информация о реагентах в этом примере являлась следующей:
1. Конструирование плазмиды для сверхэкспрессии TPK
Последовательности TPK человека амплифицировали посредством ПЦР с кДНК, полученной посредством обратной транскрипции из клеток HEK 293 человеческого происхождения, и продукт амплификации ПЦР расщепляли с использованием рестрикционных эндонуклеаз (5': BamHI-HF (Буфер 4), 3': ClaI (Буфер 4+BSA)), и целевого фрагмента последовательности гена TPK1 732 п.о. (NM_022445) (SEQ ID NO.14). Восемь повторов метки Myc вставляли с C-конца кодирующей последовательности для получения TPK1-Myc, целевой фрагмент клонировали в вектор pCS2, и таким образом, конструировали плазмиду для сверхэкспрессии TPK (см. фиг. 12).
Праймеры для амплификации ПЦР:
Прямой праймер: AT-GGATCC(BamHI)-ACCATGGAGCATGCCTTTACCCCG (SEQ ID NO.15);
Обратный праймер: AT-ATCGAT(ClaI)-GGCTTTTGATGGCCATGGTCCA (SEQ ID NO.16).
2. Клетки HEK293-APP и культивирование
1) Клетки HEK293-APP культивировали в 10 см чашках, и среду в чашке удаляли при по меньшей мере 90% конфлюэнтности адгерентных клеток. Адгерентные клетки осторожно промывали с использованием 1 мл предварительно нагретого до 37°C буфера 1XPBS, с последующим удалением PBS, и затем, 1 мл предварительно нагретого до 37°C трипсина-ЭДТА с масс-объемной процентной концентрацией 0,25% добавляли в чашку, которую затем осторожно встряхивали, для полного контакта расщепляющего раствора с адгерентными клетками. Клетки HEK293-APP помещали при комнатной температуре, только приблизительно 2-4 мин было необходимо для расщепления.
2) 4 мл предварительно нагретой до 37°C среды DMEM, содержащей 10% FBS (объемная процентная концентрация) добавляли для остановки расщепления, с последующим осторожным пипетированием вверх и вниз. Суспензию клеток HEK293-APP собирали, и центрифугировали при 1000 об./мин в течение 5 мин.
3) Супернатант отбрасывали, и клетки HEK293-APP осторожно пипетировали вверх и вниз для диспергирования с использованием 1 мл наконечника, соответственно.
4) В соответствии с требованиями эксперимента, клетки HEK293-APP из 3) инокулировали в 12-луночные планшеты при 150000 клеток/лунку, после 2-суточного культивирования, клетки адгерентно вырастали до 75-85% конфлюэнтности, и затем проводили следующий эксперимент липосомной трансфекции.
3. Эксперимент липосомной трансфекции
1) Плазмиду извлекали с -20°C, растворяли при нормальной температуре, и центрифугировали при 13000 об./мин, комнатной температуре, в течение 3 мин.
2) Составление раствора плазмиды: X806 (плазмиду для сверхэкспрессии TPK (1,218 мкг/мкл)) и контр. плазмиду (пустую контрольную плазмиду (1,21 мкг/мкл)) предварительно смешивали со средой opti-MEM, соответственно, при масс-объемном соотношении 1:50.
3) Получение раствора липосом: липофектамин 2000 смешивали со средой opti-MEM в соответствии с масс-объемным соотношением 1:50.
4) Равные объемы раствора плазмиды и раствора липосом смешивали, и помещали при комнатной температуре на 20 мин для формирования комплекса плазмиды и липосомы. Смесь являлась стабильной в течение 6 часов.
5) Смесь добавляли в 12-луночный культуральный планшет, всего 150 мкл на лунку. Смесь пипетировали вверх и вниз, или планшет встряхивали вперед и назад, для избегания скопления.
6) Клетки HEK293-APP с добавленной смесью плазмиды-липосомы помещали в инкубатор с диоксидом углерода на 24 час и 48 час, с последующим сбором, обработкой и тестированием образцов белка из клеток.
4. Сбор и обработка образцов белка из клеток HEK293-APP
Клетки HEK293-APP, трансфицированные с использованием липосом, культивировали в течение 24 час и 8 час, с последующей промывкой клеток 3 раза с использованием предварительно нагретого до 37°C 1x PBS. PBS отбрасывали, и 100 мкл раствора для лизиса RIPA (содержащего ингибитор протеаз PMSF) добавляли в каждую лунку 12-луночного культурального планшета. Раствор для лизиса RIPA полностью приводили в контакт с клетками, с последующим помещением на лед на 5 мин. Клетки соскребали с использованием скребка для клеток или наконечника и затем собирали в пробирку, которую центрифугировали при 4°C, 13000 об./мин в течение 15 мин. Преципитат отбрасывали, супернатант белка собирали, концентрацию белка в нем определяли с использованием набора BCA, и затем экспрессию целевого белка детектировали посредством Вестерн-блоттинга.
5. Детекция посредством Вестерн-блоттинга
Сверхэкспрессию плазмиды TPK в линии клеток HEK293-APP в течение 24 час (TPK OVER-1) и 48 час (TPK OVER-1) детектировали посредством WB. Конкретный способ детекции являлся следующим.
1) Получение геля: верхний гель (масс-объемная процентная концентрация составляла 5%); нижний гель (масс-объемная процентная концентрация составляла 10%).
2) Электрофорез: 1 X рабочий буфер, прогон при 90 В в течение 20-30 мин и затем прогон при 120 В в течение 60-90 мин.
3) Перенос на мембрану: мембрану PVDF помещали в метанол для активации, с последующим переносом белка из полиакриламидного геля на твердую подложку, мембрану PVDF, при постоянной силе тока 400 мА в течение 100 мин.
4) Реакция с антителом: PVDF блокировали с использованием 5% BSA (масс-объемная процентная концентрация в TBST) в течение 2 часов, и добавляли первичное антитело, с последующим помещением при 4°C на ночь. После 3 промывок с использованием 1 X TBST в течение 10 мин, мембрану PVDF инкубировали с вторичным антителом, которое разводили с использованием 5% BSA (в TBST) в течение 2 часов при комнатной температуре, с последующими 3 промывками с использованием 1 X TBST в течение 10 мин.
5) Раствор для проявления ECL: проявление проводили с использованием люминесцентного устройства CLiNX (CLiNX, Chemi scopeseries 6000).
T-критерий использовали для статистического анализа. Результаты теста WB показали, что экспрессия белка APP в клетках HEK293-APP была значимо уменьшена, и значимые различия наблюдали при всех сравнениях (*p < 0,05; **p < 0,01) (см. фиг. 13).
Можно наблюдать, что сверхэкспрессия TPK может уменьшать уровни экспрессии белка APP в клетках HEK293-APP, уровни белка APP были уменьшены с значимым различием в группах как 24 час, так и 48 час, по сравнению с нулевой контрольной группой, что показывает, что плазмида для сверхэкспрессии TPK может ингибировать уровень экспрессии белка APP в клетках HEK293-APP. Таким образом, сверхэкспрессия TPK может уменьшать уровень экспрессии белка APP в клетках HEK293-APP.
Вышеупомянутое представляет собой просто предпочтительные варианты осуществления изобретения. Следует отметить, что специалист в данной области может дополнительно модифицировать и совершенствовать настоящее изобретение, не выходя за рамки принципа изобретения, что также можно считать находящимся в рамках объема изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SHANGHAI RAISING BIOTECHNOLOGY CO., LTD.
<120> Применение TPK в качестве мишени при болезни Альцгеймера
<130> OP200755
<141> 2020-07-20
<150> 201910711540.3
<151> 2019-08-02
<150> 202010038537.2
<151> 2020-01-14
<160> 16
<170> SIPOSequenceListing 1.0
<210> 1
<211> 13237
<212> ДНК
<213> Искусственная последовательность
<400> 1
ctaaattgta agcgttaata ttttgttaaa attcgcgtta aatttttgtt aaatcagctc 60
attttttaac caataggccg aaatcggcaa aatcccttat aaatcaaaag aatagaccga 120
gatagggttg agtgttgttc cagtttggaa caagagtcca ctattaaaga acgtggactc 180
caacgtcaaa gggcgaaaaa ccgtctatca gggcgatggc ccactacgtg aaccatcacc 240
ctaatcaagt tttttggggt cgaggtgccg taaagcacta aatcggaacc ctaaagggag 300
cccccgattt agagcttgac ggggaaagcc ggcgaacgtg gcgagaaagg aagggaagaa 360
agcgaaagga gcgggcgcta gggcgctggc aagtgtagcg gtcacgctgc gcgtaaccac 420
cacacccgcc gcgcttaatg cgccgctaca gggcgcgtcc cattcgccat tcaggctgcg 480
caactgttgg gaagggcgat cggtgcgggc ctcttcgcta ttacgccagc tggcgaaagg 540
gggatgtgct gcaaggcgat taagttgggt aacgccaggg ttttcccagt cacgacgttg 600
taaaacgacg gccagtgaat tgtaatacga ctcactatag ggcgaattgg gtacgcggcc 660
gcgtttgggg gatgggagtc agtgaggaca tcctcttaga aacaggggaa gaggagtgga 720
ctgaggaaca gttggaggac agaacaaact ggagggggat agcgactggg ctggaaaaaa 780
gaagagatta aagtcaaaaa taatgactaa agccaaatgt agtgacacgt ttctgtagtc 840
ctataatgta ggaggctgat tctggaagat cacatattca aacaagtagc atactagtgt 900
atatagaaac aacttccttg gtggtaggat atgaggccta ttgaatagtc atcagtgttt 960
acatgatagt catgtatgca tgctatgttt ttttctatta ctatatgtat gcaataacaa 1020
ttatccttat gcttaccatc ttgtctatgt agacaagaat ttatattgtc tatctaggat 1080
ttctgcatca cagaggaagc acagtttaaa atactgtaat gtacagttgt ctgttcattt 1140
acacatattc ctcggcatct cccctcttgc tttatctaca ttgcctttct gtcatttctg 1200
gatatttatg ttaattggtg gcatacacca tagtaatttt ggtgattttc cattaaaatc 1260
agagctgtag ggattagttt aaaatcattg tgctagtcag aattcataga tactgggacc 1320
aaaatcgaac tccagtgttc ttttaagtca ctaaaatgtc gagatctttt ttttcttcct 1380
ctctacctcc tcctcttttt tttttcatat aaatacttgg aattacttta ataatatact 1440
ttgcagtaag tgaaattatt tctcttcagt tttattcaac agaagatttt gaaatagcca 1500
atttacctgt agttgtaagt acagttcttt tttttgttgt tgttaatatt gctaaatgct 1560
gagttaaaaa acaacactta agatatcttt aacattctca gagttccttc gatagaacag 1620
ttgttccagt ggtttttagt gtgttttagt tttcacatgt ttatcaacct gaagcacagt 1680
aaataattct tcaaccatta ccagacaatc tgcaaaaaat ttgctgtata agctaatgaa 1740
catttgacat taagagctaa ctcatgcagt aatgcttagg acaatgtaaa ttatgatgaa 1800
tggatatttt gggaccaatt tattgtgatt tatctgacgc ttcctttagc atgccctcca 1860
tttcggggtc tgttacatca tctttctctt tattggttat atgtattgca tgttttttaa 1920
attgcctcgt gtgaacctgt gctatcactg cttgctatat tcttcaaata ttttattttg 1980
caattaattt tatttatatg tttgtaaaat ttgaaaatag ttttttgaat aaagttttta 2040
aaaagtaaag aaaattttta tggtacctta taggaatcaa attgttctga aagcgaattc 2100
ttttcccttc agaaggaata tatccacatt tctaacttat gtgtaacaag aggatagtct 2160
gtacatttcc ttagttatta ttctcagttt ctacttataa ccttgataat atggataaat 2220
ttagttactg tttgtctttc ataagtttcc ctaaatctat tacataaaat atgttagaaa 2280
taatgtcttc aatttttttt gattgaaaaa ataatgggaa tgttatcaga gatgggtgtg 2340
ttgagtgcct tccggtgtct tcctttgatg ctgcctcttc tgctcattgc aaatgaagct 2400
tttgttagtc tagaggacat acatacattt gcttctcact ccatttcctt ccacagcaac 2460
gtgtgcttgc ataacaacta aacgaaaata acctgtgtct tgtttatttt gttttgtttg 2520
tttgtctgtg atatcactgg ggacaaaacc atgcttcaca aatgctaggt cagcacttta 2580
tcacacatct ttacccccgg ctcttaaaat tacttaaacg ttgaattcag ctgatctttg 2640
ctatgtaata tagtatggaa attgatatca ttatacttgt taatttttac cagttaaaca 2700
ctatttcttt cattttatag tgctttagta aataaatttt tacataaaag atctcaaaaa 2760
cttgttaatt ttgtttatgg agaattactt tttatactct gagggtaatt actgatcgtt 2820
taccatctgt ttctattaat cagtcttttt atgctcccca atcagtgata tttgctattt 2880
tatgtgagtt catttttccc tctccactga tgatgatttt gatatgttgg aaatgtgcca 2940
caaattcaat gaacttctgt tgtattccta gcgtaatatg aacgtgggtt ctagtcatta 3000
taaagtgttt agggtggtta atgaaactag ccacccctct gagttgtttt tatataaact 3060
ctaatttcat ttaatatttt gtagtagctt gggctcttct tttctaaaat gtaaacttct 3120
catattttga aaaattctaa caaaaagcag agaaaactgt ttactgaatg cccagtgtgt 3180
tcatgtgttg tgactagatg taattactaa attactaatt gtttaatgac cctactctgt 3240
agtatagaag ctcaagctaa aacttagtgt tttaccccct tagcctgatt ttgtttcata 3300
tatgacaggg atcacagaat agatagactt tgatgagctt ttcagtctgt ccatagagtt 3360
tgatagctaa ccattgcatt ctctcttctg atgttagcat atcaaaaatt agagttgaat 3420
gttcgaaatc tacttaaata taggtaaatc tgcacacacg atggtgatag gatagttttg 3480
aaaatgaaga attaggtcaa cagagtagtt tgtttcttct gaagctattt ctgtcttagg 3540
catttgctca tgctatgact tagtgtggaa gacgttgaaa ttttggacta ggaaatcagt 3600
tgaatgctgt gagtggaact tattgggctg ttctatttgg agcctggaag agagtgctgt 3660
tgagagtaat acagacagtg gaaacccaca gataggaata aagactctat tagcatttgg 3720
gatacacgct attcatgaaa tattttggaa aaaaaccaat ctggctatat ttttccactt 3780
gcttgagatg taagtaatga actaatttat ttggtggagg aaatttcaag atagcatgaa 3840
tttggggttg tggcatcatt actaccgatt acacttgtat aaatctacaa tgaaaaagta 3900
accaatgtac ttaatagctt ggatcttaat agttgtatag gggcagaaga caagacattg 3960
gtctgttaaa gataaagaaa ttgccatgat atcctgttaa atcaccaata tcgtgcaacc 4020
attcctacac cataggataa aaaaattgat tattcttaac tcctagggca aggatttaaa 4080
aaaaaattaa acgaggccaa ggctctctaa atcaacatga tcgaatctca catgaactct 4140
cagagaacca ggcgctgttc tcagttctgc acaaggtttt ttacatatac attatgactt 4200
tcagtttagt gtttttaatg gaattcctaa gtgtaagaac tagtgggcct tttttcttgt 4260
gccttctctg gggcattttc tttctctctc tctttttttt ttgtctagct ccaatatatt 4320
aatttttgtt ttatctcatt ttattttatt ctaaaaacga agcattaaaa agtagcaaca 4380
aaatgccata aagatccttt cttgaaaatt gatttaatag tttaaaaaca gtaatcttaa 4440
ctactaatat taacacccaa ttattaacat aatattgatg tttgacataa ttgaagagag 4500
gttttataaa ttgcaagaga gttgaaggaa ggtcacatac agaagaaaga caagaatgaa 4560
acatctggag aaaggtcgag ggaaggtaag actgtcccca gaaatgtttc agtggaattc 4620
ccagagacag acaggtaaac aggacagagg atggaaagta attgacttaa gatgacttag 4680
attcatgaga acttgagaga tccgcttcct cagctgtggg gagcacagtc tgcttaggca 4740
taattttgaa gttcaagaca tcagagaaaa ggaagatctt caatgtttct agagggaaga 4800
caaacagagg cacagaactg tgaaacaggc tgtcacttga atataccaca gttcccagta 4860
gaattccaaa gccagcatgt taacatagtc aaagcaactg gctgtcagaa gctgtagcct 4920
gccgtcttta taagcctgaa cccagcttcg gtaataaaag agggagctag cgtcgacacg 4980
tataacttcg tataatgtat gctatacgaa gttattactg cgtacgaaaa ggtagaatgt 5040
agagcaggta gctccatatc ttctccacct ctgttcagat ggactggaga agatgcaaag 5100
ggtctgtgag tggttctgtc ttccaatgta gagtcacctg taataacaat agacacgcca 5160
taaggtaaat gtcatgatcc agggtagtgt gccaatatgc tctgagtgtt tgctctttag 5220
tgatgctcac atgacacact gtgctttgct atctctactg acataagatt tgtattcttt 5280
ttgtttgaag ctctcttaag agcctgtgct gatgggggtg ccaaccactt atatgatctc 5340
actgaaggag agagagaaag gtaagctcca gcgagtcagg gtctacatgg tcctcacttc 5400
tgctgctcct tgcttttaat tattctgtgg tgactagttc tttttcttca tattcatcat 5460
aggaaaccac ctctgtgaga gggctgggct gtataaccat gatgcattac ttctttctat 5520
gaatgagatc tcttagaatt aggatcatgt tttgtcttgt ttttaaatct gtattctatt 5580
actttaaagc ctagctaagt cacattatca tgtattgaaa tttatcgata agcttgatat 5640
cgaattccga agttcctatt ctctagaaag tataggaact tcaggtctga agaggagttt 5700
acgtccagcc aagctagctt ggctgcaggt cgtcgaaatt ctaccgggta ggggaggcgc 5760
ttttcccaag gcagtctgga gcatgcgctt tagcagcccc gctgggcact tggcgctaca 5820
caagtggcct ctggcctcgc acacattcca catccaccgg taggcgccaa ccggctccgt 5880
tctttggtgg ccccttcgcg ccaccttcta ctcctcccct agtcaggaag ttcccccccg 5940
ccccgcagct cgcgtcgtgc aggacgtgac aaatggaagt agcacgtctc actagtctcg 6000
tgcagatgga cagcaccgct gagcaatgga agcgggtagg cctttggggc agcggccaat 6060
agcagctttg ctccttcgct ttctgggctc agaggctggg aaggggtggg tccgggggcg 6120
ggctcagggg cgggctcagg ggcggggcgg gcgcccgaag gtcctccgga ggcccggcat 6180
tctgcacgct tcaaaagcgc acgtctgccg cgctgttctc ctcttcctca tctccgggcc 6240
tttcgacctg cagcctgttg acaattaatc atcggcatag tatatcggca tagtataata 6300
cgacaaggtg aggaactaaa ccatgggatc ggccattgaa caagatggat tgcacgcagg 6360
ttctccggcc gcttgggtgg agaggctatt cggctatgac tgggcacaac agacaatcgg 6420
ctgctctgat gccgccgtgt tccggctgtc agcgcagggg cgcccggttc tttttgtcaa 6480
gaccgacctg tccggtgccc tgaatgaact gcaggacgag gcagcgcggc tatcgtggct 6540
ggccacgacg ggcgttcctt gcgcagctgt gctcgacgtt gtcactgaag cgggaaggga 6600
ctggctgcta ttgggcgaag tgccggggca ggatctcctg tcatctcacc ttgctcctgc 6660
cgagaaagta tccatcatgg ctgatgcaat gcggcggctg catacgcttg atccggctac 6720
ctgcccattc gaccaccaag cgaaacatcg catcgagcga gcacgtactc ggatggaagc 6780
cggtcttgtc gatcaggatg atctggacga agagcatcag gggctcgcgc cagccgaact 6840
gttcgccagg ctcaaggcgc gcatgcccga cggcgatgat ctcgtcgtga cccatggcga 6900
tgcctgcttg ccgaatatca tggtggaaaa tggccgcttt tctggattca tcgactgtgg 6960
ccggctgggt gtggcggacc gctatcagga catagcgttg gctacccgtg atattgctga 7020
agagcttggc ggcgaatggg ctgaccgctt cctcgtgctt tacggtatcg ccgctcccga 7080
ttcgcagcgc atcgccttct atcgccttct tgacgagttc ttctgagggg atcaattctc 7140
tagagctcgc tgatcagcct cgactgtgcc ttctagttgc cagccatctg ttgtttgccc 7200
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 7260
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 7320
gcaggacagc aagggggagg attgggaaga caatagcagg catgctgggg atgcggtggg 7380
ctctatggct tctgaggcgg aaagaaccag ctggggctcg actagagctt gcggaaccct 7440
tcgaagttcc tattctctag aaagtatagg aacttcatca gtcaggtaca taatataact 7500
tcgtataatg tatgctatac gaagttatta ggtggatcca ctaggcgcgc ctgaggactt 7560
gaagattggc taaataaacc taaaatggct atttatattg ggattaatac actttaagtg 7620
aaaaaaaaaa aaaaaagcaa ctggactcct agaaaagaaa tagtctgctg tagaaatgtg 7680
aggttttata ttagtgttga ataaatctta gtgtgactta ctgtttctta attacttctg 7740
agttacagac gtatattttt gagacttcat ctaaagaatt gctctctgca atatcttggt 7800
ttcctgacct ttggatggtg agatattttt cagtgagaga acacatttga tagagataga 7860
ttttcaaact agtttagatg tgaaggtgtt tgtggcccaa ctctttttgg aaacacaaga 7920
tttcatggta tagttttgtg tgcagtttgt gaaaactcag tgtttgtaaa tacactgttc 7980
agtcattctc attctcgtat gtgttagtag ctattttgaa agggctggta tttgagaaat 8040
cgcctacttg gcttccaaaa caaaaataaa aatattatgg cataacaggc cctgtgtggg 8100
cactaaacat atgtactttg aaatcatgta tttttctttg taacatcaga atgtctaaca 8160
tcagctttct aatactggaa gtgctgttga gagcagctgg ttgcacgttg tggagaagtc 8220
atgtttcctt tgagatcaga tgctattgct aatgtctgga gactggctgc agttatcaaa 8280
aaggcaataa tagtgacagc ctaggacagt taaatttcat ttgcttattt actgcacaac 8340
tttgcaagtt tctctaccga atacctcaca tggacaatgc ctgaaaaaca tgcaagttgt 8400
tttaaaaagt gccgtggcac atagttcaga acagggaggc atgccatggg gcccagatta 8460
taaagtgggc actgtttgac tcgtgcctac agcctggttg ctaccaagta gttggcactt 8520
aaacactttt tccataatgg tagacacagc tccggaacgt ggcagttgaa catgacctct 8580
ctaatggtag atacaactca agaacataat taaaagcagg acttagcaag cattaactca 8640
gcactttcca cctacttcat gattatgaga accgtctgaa atgattatga aaaattacac 8700
atctttatac tgggtatttt atttagttga gccaagcact ggtagcaaag aaaaaaggga 8760
acacttttat tcatatttca aagcaaaaca ttagtgattt tggaaacaag taacattagt 8820
tttgatatta ttttaattaa tttcaaaata acaatcacat tataaataat ttgttttttt 8880
tttttggctt tcttgttttg ttttttgaga caggttctca ctatatagcc ctggttgtcc 8940
tggaactcac tatgtagacc agtctgactt caaactcaca agctctctgc ctcctgagta 9000
ctgggatcac aggtgttatg cccagtcaaa tttacaaata atcttattga acgcattgag 9060
gaatgtagat aatgtgtctt acaattgttt gtctacacat gaagatgttt agtgaagcaa 9120
tgcagttact actcattgat tttcaggttg gaccgatgct tgcagtgatt cttgctctga 9180
ctcctttgga catggttggc ctggggaagg gtagagtccc tagaagtgac aagtgaaact 9240
aactgctttg catgggaata aaatcctttg aataaatctc attacttcta cgccctaacc 9300
agtaaaacta attggataca ggtgactaca cataattcct cttaattgta ttttttaaca 9360
aaaagccctc acaatgaaaa acaaaataaa caccaacaaa atcctgagct ttaaaaggcc 9420
aaattacaga ttgtgggctg aggctggtct ctcttttttt tttttttttc ttaaatttga 9480
ttttattttt ttcacattcc atattccatt ccccgccccc cccacatcca ccctccaact 9540
gatccacatc ccacacctcc taggatcgcc cgggttgatt cgaggctgct aacaaatcga 9600
gtcgagcatc gagcagtgtg gttttcaaga ggaagcaaaa agcctctcca cccaggcctg 9660
gaatgtttcc acccaatgtc gagcagtgtg gttttgcaag aggaagcaaa aagcctctcc 9720
acccaggcct ggaatgtttc cacccaatgt cgagcaaacc ccgcccagcg tcttgtcatt 9780
ggcgaattcg aacacgcaga tgcagtcggg gcggcgcggt cccaggtcca cttcgcatat 9840
taaggtgacg cgtgtggcct cgaacaccga gcgaccctgc agcgacccgc ttaacagcgt 9900
caacagcgtg ccgcagatct tggtggcgtg aaactcccgc acctcttcgg ccagcgcctt 9960
gtagaagcgc gtgccatgga tcctgatgat gttgttgatt cttctaaatc ttttgtgatg 10020
gaaaactttt cttcgtacca cgggactaaa cctggttatg tagattccat tcaaaaaggt 10080
atacaaaagc caaaatctgg tacacaagga aattatgacg atgattggaa agggttttat 10140
agtaccgaca ataaatacga cgctgcggga tactctgtag ataatgaaaa cccgctctct 10200
ggaaaagctg gaggcgtggt caaagtgacg tatccaggac tgacgaaggt tctcgcacta 10260
aaagtggata atgccgaaac tattaagaaa gagttaggtt taagtctcac tgaaccgttg 10320
atggagcaag tcggaacgga agagtttatc aaaaggttcg gtgatggtgc ttcgcgtgta 10380
gtgctcagcc ttcccttcgc tgaggggagt tctagcgttg aatatattaa taactgggaa 10440
caggcgaaag cgttaagcgt agaacttgag attaattttg aaacccgtgg aaaacgtggc 10500
caagatgcga tgtatgagta tatggctcaa gcctgtgcag gaaatcgtgt caggcgatct 10560
ctttgtgaag gaaccttact tctgtggtgt gacataattg gacaaactac ctacagagat 10620
ttaaagctct aaggtaaata taaaattttt aagtgtataa tgtgttaaac tactgattct 10680
aattgtttgt gtattttaga ttccaaccta tggaactgat gaatgggagc agtggtggaa 10740
tgcagatcct agagctcgct gatcagcctc gactgtgcct tctagttgcc agccatctgt 10800
tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca ctgtcctttc 10860
ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta ttctgggggg 10920
tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc atgctgggga 10980
tgcggtgggc tctatggctt ctgaggcgga aagaaccagc ccgggcggtg gagctccagc 11040
ttttgttccc tttagtgagg gttaatttcg agcttggcgt aatcatggtc atagctgttt 11100
cctgtgtgaa attgttatcc gctcacaatt ccacacaaca tacgagccgg aagcataaag 11160
tgtaaagcct ggggtgccta atgagtgagc taactcacat taattgcgtt gcgctcactg 11220
cccgctttcc agtcgggaaa cctgtcgtgc cagctgcatt aatgaatcgg ccaacgcgcg 11280
gggagaggcg gtttgcgtat tgggcgctct tccgcttcct cgctcactga ctcgctgcgc 11340
tcggtcgttc ggctgcggcg agcggtatca gctcactcaa aggcggtaat acggttatcc 11400
acagaatcag gggataacgc aggaaagaac atgtgagcaa aaggccagca aaaggccagg 11460
aaccgtaaaa aggccgcgtt gctggcgttt ttccataggc tccgcccccc tgacgagcat 11520
cacaaaaatc gacgctcaag tcagaggtgg cgaaacccga caggactata aagataccag 11580
gcgtttcccc ctggaagctc cctcgtgcgc tctcctgttc cgaccctgcc gcttaccgga 11640
tacctgtccg cctttctccc ttcgggaagc gtggcgcttt ctcatagctc acgctgtagg 11700
tatctcagtt cggtgtaggt cgttcgctcc aagctgggct gtgtgcacga accccccgtt 11760
cagcccgacc gctgcgcctt atccggtaac tatcgtcttg agtccaaccc ggtaagacac 11820
gacttatcgc cactggcagc agccactggt aacaggatta gcagagcgag gtatgtaggc 11880
ggtgctacag agttcttgaa gtggtggcct aactacggct acactagaag aacagtattt 11940
ggtatctgcg ctctgctgaa gccagttacc ttcggaaaaa gagttggtag ctcttgatcc 12000
ggcaaacaaa ccaccgctgg tagcggtggt ttttttgttt gcaagcagca gattacgcgc 12060
agaaaaaaag gatctcaaga agatcctttg atcttttcta cggggtctga cgctcagtgg 12120
aacgaaaact cacgttaagg gattttggtc atgagattat caaaaaggat cttcacctag 12180
atccttttaa attaaaaatg aagttttaaa tcaatctaaa gtatatatga gtaaacttgg 12240
tctgacagtt accaatgctt aatcagtgag gcacctatct cagcgatctg tctatttcgt 12300
tcatccatag ttgcctgact ccccgtcgtg tagataacta cgatacggga gggcttacca 12360
tctggcccca gtgctgcaat gataccgcga gacccacgct caccggctcc agatttatca 12420
gcaataaacc agccagccgg aagggccgag cgcagaagtg gtcctgcaac tttatccgcc 12480
tccatccagt ctattaattg ttgccgggaa gctagagtaa gtagttcgcc agttaatagt 12540
ttgcgcaacg ttgttgccat tgctacaggc atcgtggtgt cacgctcgtc gtttggtatg 12600
gcttcattca gctccggttc ccaacgatca aggcgagtta catgatcccc catgttgtgc 12660
aaaaaagcgg ttagctcctt cggtcctccg atcgttgtca gaagtaagtt ggccgcagtg 12720
ttatcactca tggttatggc agcactgcat aattctctta ctgtcatgcc atccgtaaga 12780
tgcttttctg tgactggtga gtactcaacc aagtcattct gagaatagtg tatgcggcga 12840
ccgagttgct cttgcccggc gtcaatacgg gataataccg cgccacatag cagaacttta 12900
aaagtgctca tcattggaaa acgttcttcg gggcgaaaac tctcaaggat cttaccgctg 12960
ttgagatcca gttcgatgta acccactcgt gcacccaact gatcttcagc atcttttact 13020
ttcaccagcg tttctgggtg agcaaaaaca ggaaggcaaa atgccgcaaa aaagggaata 13080
agggcgacac ggaaatgttg aatactcata ctcttccttt ttcaatatta ttgaagcatt 13140
tatcagggtt attgtctcat gagcggatac atatttgaat gtatttagaa aaataaacaa 13200
ataggggttc cgcgcacatt tccccgaaaa gtgccac 13237
<210> 2
<211> 27
<212> ДНК
<213> Искусственная последовательность
<400> 2
gtcacttgaa tataccacag ttcccag 27
<210> 3
<211> 23
<212> ДНК
<213> Искусственная последовательность
<400> 3
catattggca cactaccctg gat 23
<210> 4
<211> 23
<212> ДНК
<213> Искусственная последовательность
<400> 4
gctgaccgct tcctcgtgct tta 23
<210> 5
<211> 25
<212> ДНК
<213> Искусственная последовательность
<400> 5
gaccaccaaa cagcatacac cagct 25
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 6
atctttccag tgtctcattc 20
<210> 7
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 7
tacttcttgt ttcggtctt 19
<210> 8
<211> 18
<212> ДНК
<213> Искусственная последовательность
<400> 8
ggatgttggc attgaagc 18
<210> 9
<211> 18
<212> ДНК
<213> Искусственная последовательность
<400> 9
gattttagga gacgagca 18
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 10
gaattccaaa gccagcatgt 20
<210> 11
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 11
ccactcacag accctttgca 20
<210> 12
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 12
gacaggcagg ccttctctga a 21
<210> 13
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 13
cttctccaca ccagctgtgg a 21
<210> 14
<211> 732
<212> ДНК
<213> Искусственная последовательность
<400> 14
atggagcatg cctttacccc gttggagccc ctgctttcca ctgggaattt gaagtactgc 60
cttgtaattc ttaatcagcc tttggacaac tattttcgtc atctttggaa caaagctctt 120
ttaagagcct gtgccgatgg aggtgccaac cgcttatatg atatcaccga aggagagaga 180
gaaagctttt tgcctgaatt catcaatgga gactttgatt ctattaggcc tgaagtcaga 240
gaatactatg ctactaaggg atgtgagctc atttcaactc ctgatcaaga ccacactgac 300
tttactaagt gccttaaaat gctccaaaag aagatagaag aaaaagactt aaaggttgat 360
gtgatcgtga cactgggagg ccttgctggg cgttttgacc agattatggc atctgtgaat 420
accttgttcc aagcgactca catcactcct tttccaatta taataatcca agaggaatcg 480
ctgatctacc tgctccaacc aggaaagcac aggttgcatg tagacactgg aatggagggt 540
gattggtgtg gccttattcc tgttggacag ccttgtatgc aggttacaac cacaggcctc 600
aagtggaacc tcacaaatga tgtgcttgct tttggaacat tggtcagtac ttccaatacc 660
tacgacgggt ctggtgttgt gactgtggaa actgaccacc cactcctctg gaccatggcc 720
atcaaaagct aa 732
<210> 15
<211> 32
<212> ДНК
<213> Искусственная последовательность
<400> 15
atggatccac catggagcat gcctttaccc cg 32
<210> 16
<211> 30
<212> ДНК
<213> Искусственная последовательность
<400> 16
atatcgatgg cttttgatgg ccatggtcca 30
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ОТЛИЧНОГО ОТ ЧЕЛОВЕКА ЖИВОТНОГО С МУТИРОВАННЫМ НОКИН-ГЕНОМ, СПОСОБ ТЕСТИРОВАНИЯ ВЕЩЕСТВА НА ПРИМЕНИМОСТЬ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА (ВАРИАНТЫ), ПЛАЗМИДА (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПЕРВИЧНОЙ КУЛЬТУРЫ КЛЕТОК ИЛИ СУБКУЛЬТИВИРУЕМОЙ КЛЕТКИ | 1999 |
|
RU2266002C2 |
| ТРАНСГЕННЫЕ ЖИВОТНЫЕ, ИМЕЮЩИЕ ОСНОВНЫЕ РАССТРОЙСТВА, СВЯЗАННЫЕ С БОЛЕЗНЬЮ АЛЬЦГЕЙМЕРА | 2004 |
|
RU2373707C2 |
| Способ получения генно-модифицированных кроликов с нокаутом гена LEPR с помощью системы CRISPR/Cas9 | 2023 |
|
RU2836438C1 |
| ГРЫЗУНЫ С УСЛОВНЫМИ МУТАНТНЫМИ АЛЛЕЛЯМИ ACVR1 | 2014 |
|
RU2676961C2 |
| МОДЕЛЬ ПРОГРЕССИРУЮЩЕЙ ОССИФИЦИРУЮЩЕЙ ФИБРОДИСПЛАЗИИ НА ГРЫЗУНАХ | 2019 |
|
RU2795136C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |
| СПОСОБ СКРИНИНГА ВЕЩЕСТВ, ИМЕЮЩИХ РЕГУЛИРУЮЩЕЕ ВЕС ДЕЙСТВИЕ | 2012 |
|
RU2603745C2 |
| ЭКСПРЕССИЯ АНТИГЕНСВЯЗЫВАЮЩИХ БЕЛКОВ В НЕРВНОЙ СИСТЕМЕ | 2020 |
|
RU2831086C2 |
| НАЦЕЛИВАНИЕ НА SRC-3 В ИММУННЫХ КЛЕТКАХ В КАЧЕСТВЕ ИММУНОМОДУЛИРУЮЩЕГО ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ РАКА | 2021 |
|
RU2828447C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ДЕГЕНЕРАТИВНОГО ЗАБОЛЕВАНИЯ ГОЛОВНОГО МОЗГА, ВКЛЮЧАЮЩАЯ ПЕРЕНОСЧИК ГЛИЦИНА В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2021 |
|
RU2816256C1 |

Изобретение относится к области биотехнологии, в частности к применению гена или белка тиаминпирофосфаткиназы (TPK) в качестве мишени для лечения, предотвращения болезни Альцгеймера, в скрининге лекарственного средства для предотвращения или лечения болезни Альцгеймера, в качестве мишени для получения лекарственного средства для предотвращения или лечения болезни Альцгеймера, а также в конструировании модели на животных болезни Альцгеймера. Также раскрыто применение реагента, стимулирующего активность киназы белка TPK в головном мозге и/или уровень экспрессии белка TPK в головном мозге, в получении лекарственного средства для лечения или предотвращения болезни Альцгеймера. Изобретение также относится к лекарственному средству, которое содержит ген ТРК или белок ТРК. Изобретение эффективно для предотвращения или лечения болезни Альцгеймера. 12 н. и 13 з.п. ф-лы, 13 ил., 3 пр.
1. Применение гена или белка тиаминпирофосфаткиназы (TPK) в качестве мишени для предотвращения болезни Альцгеймера.
2. Применение гена или белка TPK в качестве мишени для лечения болезни Альцгеймера.
3. Применение по п.1 или 2, отличающееся тем, что предотвращение или лечение осуществляют с использованием гена или белка TPK в качестве мишени посредством стимуляции активности киназы белка TPK в головном мозге и/или уровня экспрессии белка TPK в головном мозге.
4. Применение гена или белка TPK в качестве мишени в скрининге лекарственного средства для предотвращения или лечения болезни Альцгеймера.
5. Применение гена или белка TPK в качестве мишени для получения лекарственного средства для предотвращения или лечения болезни Альцгеймера.
6. Применение по п.4 или 5, отличающееся тем, что лекарственное средство, с использованием гена или белка TPK в качестве мишени, стимулирует активность киназы белка TPK в головном мозге и/или уровень экспрессии белка TPK в головном мозге.
7. Применение реагента, стимулирующего активность киназы белка TPK в головном мозге и/или уровень экспрессии белка TPK в головном мозге, в получении лекарственного средства для предотвращения болезни Альцгеймера.
8. Применение реагента, стимулирующего активность киназы белка TPK в головном мозге и/или уровень экспрессии белка TPK в головном мозге, в получении лекарственного средства для лечения болезни Альцгеймера.
9. Применение по п.7 или 8, отличающееся тем, что реагент, стимулирующий активность киназы белка TPK в головном мозге и/или уровень экспрессии белка TPK в головном мозге, представляет собой реагент, регулирующий белок TPK, и/или реагент, регулирующий ген TPK.
10. Лекарственное средство для предотвращения болезни Альцгеймера, отличающееся тем, что лекарственное средство содержит ген ТРК или белок ТРК.
11. Лекарственное средство для лечения болезни Альцгеймера, отличающееся тем, что лекарственное средство содержит ген ТРК или белок ТРК.
12. Применение гена или белка TPK в качестве мишени в конструировании модели на животных болезни Альцгеймера.
13. Способ конструирования модели на животных, отличающийся тем, что модель на животных конструируют посредством нокаута гена TPK у животного-мишени.
14. Способ конструирования модели на животных по п.13, отличающийся тем, что животное-мишень представляет собой грызуна.
15. Способ конструирования модели на животных по п.14, отличающийся тем, что грызун представляет собой мышь.
16. Способ конструирования модели на животных по п.15, отличающийся тем, что экзон 4 в гене TPK подвергают нокауту.
17. Способ конструирования модели на животных по п.13, отличающийся тем, что способ включает стадии: конструирования нацеливающего вектора TPK-loxp для нокаута гена, введения нацеливающего вектора в животное-мишень для получения TPK-loxP+/+ животного-мишени.
18. Способ конструирования моделей на животных по п.17, отличающийся тем, что последовательность нацеливающего вектора указана в SEQ ID NO. 1.
19. Способ конструирования модели на животных по п.17, отличающийся тем, что способ дополнительно включает стадии: скрещивания TPK-loxP+/+ животного-мишени с CamK2α-Cre/ERT2+/- животным-мишенью, получения CamK2α-Cre/ERT2+/-; TPK-loxP+/+ животного-мишени с условным нокаутом гена TPK.
20. Способ конструирования нацеливающего вектора для модели на животных, отличающийся тем, что бактериальную искусственную хромосому используют в качестве вектора, прямые последовательности loxP вставляют на обоих концах последовательности, гомологичной экзону гена TPK, вместе с последовательностью гена устойчивости к неомицину.
21. Способ конструирования нацеливающего вектора по п.20, отличающийся тем, что экзон представляет собой экзон 4.
22. Способ конструирования нацеливающего вектора по п.21, отличающийся тем, что последовательность нацеливающего вектора указана в SEQ ID NO. 1.
23. Нацеливающий вектор, который представляет собой бактериальную искусственную хромосому, содержащую прямые последовательности loxP, вставленные на обоих концах последовательности, гомологичной экзону гена TPK, вместе с последовательностью гена устойчивости к неомицину.
24. Нацеливающий вектор по п.23, отличающийся тем, что экзон представляет собой экзон 4.
25. Нацеливающий вектор по п.24, отличающийся тем, что последовательность нацеливающего вектора указана в SEQ ID NO. 1.
| SHAOMING SANG et al., Thiamine diphosphate reduction strongly correlates with brain glucose hypometabolism in Alzheimer’s disease, whereas amyloid deposition does not, Alzheimers Res Ther., 2018, 10: 26 | |||
| CHANGPENG WANG et al., High thiamine diphosphate level as a protective factor for Alzheimer's disease, Neurol Res, 2018 Aug, 40(8), pp.658-665 |
