Область изобретения
Настоящее изобретение относится к новому классу моноклонального антитела, которое специфически связывается с альфа-синуклеином, а также к способам применения этих молекул и их связывающих альфа-синуклеин фрагментов для лечения и диагностики синуклеопатий.
Ссылка на перечень последовательностей
Настоящая заявка содержит один или несколько перечней последовательностей в соответствии с 37 CFR 1.821 и далее, которые раскрыты на машиночитаемых носителях (название файла: 0992_ST25.txt, создан 22 июня 2016 года и имеет размер 44 кБ), при этом данный файл включен в данный документ посредством ссылки во всей своей полноте.
Предпосылки изобретения
Синуклеопатии, также известные как болезни телец Леви (LBD), характеризуются отложением внутриклеточных белковых агрегатов, которые видны под микроскопом в виде телец Леви (LB) и/или нейритов Леви, где основным компонентом является белок альфа-синуклеин (Jellinger, Mov Disord. 2012 Jan; 27(1):8-30; McKeith et al., Neurology (1996) 47:1113-24). Синуклеопатии включают болезнь Паркинсона (PD) (в том числе идиопатическая и наследственная формы болезни Паркинсона) и болезнь диффузных телец Леви (DLB) (также известная как деменция с тельцами Леви (DLB), вариант болезни Альцгеймера с тельцами Леви (LBV), комбинированная болезнь Альцгеймера и Паркинсона (CAPD), истинная вегетативная недостаточность (PAF) и множественная системная атрофия (MSA, например, оливопонтоцеребеллярная атрофия, стрионигральная дегенерация и синдром Шая-Дрейджера)). Синуклеопатии зачастую характеризуются дегенерацией дофаминергической нигростриатной системы, ответственной за основные двигательные нарушения при паркинсонизме (ригидность, брадикинезия, тремор в покое), но также широко распространено появления телец Леви и дистрофических нейритов Леви в центральной, периферической и вегетативной нервной системе и областях головного мозга, и других органах, ассоциированных с функциональными нарушениями, отличными от двигательных, например, при деменции и нарушениях вегетативной нервной системы. Полагают, что некоторые из признаков и симптомов, отличных от двигательных, предшествуют двигательным симптомам при болезни Паркинсона и других синуклеопатиях. Такие ранние признаки включают, например, поведенческое расстройство фазы быстрого сна (RBD) и утрату обоняния и обстипацию (Mahowald et al., Neurology (2010) 75:488-489). Синуклеопатии по-прежнему являются общей причиной двигательных нарушений и нарушений когнитивных функций у стареющего населения (Galasko et al., Arch. Neurol. (1994) 51:888-95).
Альфа-синуклеин является членом семейства белков, в которое входит бета- и гамма-синуклеин и синоретин. В нормальном состоянии альфа-синуклеин экспрессируется ассоциированный с синапсами, и полагают, что он играет некоторую роль в регуляции высвобождения синаптических везикул и за счет этого влияет на нейронную связь, пластичность, способность к обучению и память.
В нескольких исследованиях было показано, что альфа-синуклеин задействован в патогенезе PD и играет в нем центральную роль. При патологических состояниях этот белок может агрегироваться с образованием внутриклеточных нерастворимых фибрилл. Например, синуклеин накапливается в LB (Spillantini et al., Nature (1997) 388:839-40; Takeda et al., J. Pathol. (1998) 152:367-72; Wakabayashi et al., Neurosci. Lett. (1997) 239:45-8). Мутации в гене альфа-синуклеина, а также дупликации и трипликации этого гена, совместно сегрегируют с редкими наследственными формами паркинсонизма (Kruger et al., Nature Gen. (1998) 18:106-8; Polymeropoulos, et al., Science (1997) 276:2045-7). Важным открытием было то, что альфа-синуклеин может секретироваться во внеклеточную жидкость и присутствовать в плазме крови и спинномозговой жидкости (CSF). По результатам нескольких исследований, например, проведенных Pacheco et al. (2015), и других (Pacheco et al J Neurochem. 2015 Mar; 132(6):731-4; Conway et al., Proc Natl Acad Sci USA (2000) 97:571-576; Volles et al., J. Biochem. 42:7871-7878, 2003) было сделано предположение, что внеклеточный синуклеин играет патогенную роль в головном мозге. Они продемонстрировали, что внеклеточные олигомеры альфа-синуклеина обладают нейротоксичностью в отношении плазматических мембран нейронов головного мозга. Другая интересная гипотеза, основанная на данных по секреции синуклеина, заключается в том, что в основе прогрессирования болезни Паркинсона и других синуклеопатий лежит прионоподобное распространение альфа-синуклеина (Lee et al. 2014, Nat Rev Neurol. 2014 Feb;10(2):92-8; Hansen and Li 2012, Trends Mol Med. 2012 May; 18(5):248-55). Эти результаты дали надежду, что внеклеточный синуклеин может выступать в качестве мишени для иммунотерапии (Vekrellis et al. 2011, Lancet Neurol. 2011 Nov; 10(11): 1015-25).
Было показано, что встечающиеся в природе аутоантитела к альфа-синуклеину присутствуют как у пациентов с PD, так и у здоровых контролей (Smith et al. 2012, PLoS One. 2012;7(12):e52285; Maetzler et al. 2014, PLoS One. 2014 Feb 21;9(2):e88604, Papachroni et al. 2007 J Neurochem. 2007 May; 101(3):749-56 и Woulfe et al. 2002, Neurology. 2002 May 14;58(9): 1435-6), иногда сообщалось о повышенных уровнях аутоантител к альфа-синуклеину при PD (Gruden et al. 2011, J Neuroimmunol. 2011 Apr;233(l-2):221-7, Gruden et al.
2012, Neuroimmunomodulation. 2012;19(6):334-42 и Yanamandra 2011, PLoS One. 2011 Apr 25;6(4):el8513) или пониженных уровнях аутоантител к альфа-синуклеину у пациентов с PD по сравнению со здоровыми контролями (Besong-Agbo et al 2013, Neurology. 2013 Jan 8;80(2): 169-75).. Возможность того, что циркулирующие аутоантитела к альфа-синуклеину могут выполнять защитную роль в отношении агрегирования альфа-синуклеина, была предположена почти сразу после обнаружения аутоантител (Woulfe et al. 2002, Neurology. 2002 May 14;58(9):1435-6).
Сверхэкспрессия альфа-синуклеина у трансгенных мышей имитирует некоторые патологические аспекты болезни телец Леви. За последние десять лет было создано несколько различных трансгенных линий мышей, сверхэкспрессирующих альфа-синуклеин мышей (описано в обзорах: Koehler et al 2014, PLoS One. 2013 May 31;8(5):e64649; Fleming and Chesselet, 2006, Behav Pharmacol. 2006 Sep; 17(5-6):383-91; Springer and Kahle 2006, Curr Neurol Neurosci Rep.2006 Sep; 6(5):432-6). У мышиных линий с промоторами Thy-1 и PDGF-бета развиваются двигательные нарушения и когнитивные нарушения, и они были использованы для демонстрации нейропротекторного действия антител к альфа-синуклеину in vivo. Тем не менее, ни одна из трансгенных линий не характеризуется робастной дегенерацией дофаминергических нейронов, и зачастую двигательные фенотипы управляются экспрессией в двигательных нейронах, которые обычно не дегенерируют при болезни Паркинсона. Поэтому неясно, опосредуется ли положительный результат потенциальной симптом-модифицирующей терапии эффектами, оказываемыми на дофаминергические нейроны или другие нейроны центральной нервной системы.
Одним из устойчивых результатов, полученных на трансгенных мышиных моделях, было то, что хроническая сверхэкспрессия альфа-синуклеина человека ухудшает синаптическую функцию. С помощью исследований как в in vitro, так и в in vivo системах было показано, что сверхэкспрессия альфа-синуклеина дикого типа (wt) человека нарушала синаптическую передачу в гиппокампе (Nemani et al. 2010, Neuron. 2010 Jan 14;65(l):66-79; Paumier et al. 2013, PLoS One. 2013 Aug 1;8(8):е70274). Это было показано в области СА1 гиппокампа, где в обоих исследованиях была обнаружена сниженная базальная синаптическая передача. Была сделано предположение, что лежащий в основе этого механизм заключается во внутриклеточном накоплении альфа-синуклеина, что приводит к нарушенному высвобождению в синапс. Тем не менее, последние полученные результаты по секреции альфа-синуклеина во внеклеточное пространство в синапсах и по токсическим эффектам, которые оказывают олигомеры альфа-синуклеина на функцию синапса, раскрывают возможность некоторой роли внеклеточного альфа-синуклеина в функциональном нарушении синапса и, соответственно, возможность терапевтических антител устранить это нарушение.
Применение вирусных векторов для сверхэкспрессии альфа-синуклеина представляет собой важный способ моделирования PD у грызунов, поскольку с помощью этого подхода получают относительно быструю прогрессирующую дегенерацию нигростриальных нейронов, признак, который все еще не был воспроизведен с помощью генетических мутаций у мышей или крыс (Kirik and Bjorklund, 2003, Trends Neurosci. 2003 Jul; 26(7):386-92). Кроме того, вирусная доставка генов выявила способность альфа-синуклеина wt индуцировать развитие нигростриальной патологии (Kirik et al. 2002, J Neurosci. 2002 Apr 1;22(7):2780-91), результат, согласующийся с данными, наблюдаемыми при наследственных формах PD с дупликациями или трипликациями альфа-синуклеина (Lee and Trojanowski, 2006, Neuron. 2006 Oct 5;52(1):33-8). В одном исследовании было показано, что пул антител козы к N-концу альфа-синуклеина защищал от гибели дофаминергические клетки и уменьшал поведенческие нарушения в крысиной модели болезни Паркинсона на основе AAV-альфа-синуклеина (Shahaduzzaman et al 2015, PLoS One. 2015 Feb 6;10(2):e0116841).
Недавно было показано, что прионоподобное распространение альфа-синуклеиновой патологии приводит к развитию альфа-синуклеиновой патологии, а также приводит к гибели дофаминергических клеток (Luk et al. 2012, Science. 2012 Nov 16:338(6109):949-53). Эта модель была использована для демонстрации, что антитела к альфа-синуклеину способны облегчать патологию (Tran et al. 2014, Cell Rep.2014 Jun 26;7(6):2054-65). В этой модели обработка антителами позволяла уменьшить накопление фосфорилированного альфа-синуклеина в нескольких областях головного мозга, в том числе в дофаминергических нейронах в черном веществе, и уменьшить развитие двигательного нарушения.
Помимо мутаций, в результате альтернативного сплайсинга гена альфа-синуклеина и посттрансляционных модификаций белка, таких как фосфорилирование, убиквитинирование, нитрование и усечение, могут быть получены формы белка альфа-синуклеина, которые обладают повышенной способностью образовывать агрегированные и/или токсичные формы альфа-синуклеина (Beyer and Ariza, Mol Neurobiol. 2013 Apr; 47(2):509-24). Тем не менее, конкретные патологические разновидности альфа-синуклеина остаются неизвестными. С токсичностью была установлена ассоциация для различных неправильно уложенных/агрегированных/секретируемых разновидностей, варьирующих от олигомеров до фибрилл, и различных посттрансляционных модификаций, но все еще отсутствует единое мнение по вопросу, какая из них является наиболее важной, даже если фактически есть одна единственная токсичная разновидность.
В целом, накопление альфа-синуклеина со схожими морфологическими и неврологическими изменениями в животных моделях, которые характеризуются обширным разнообразием, например, люди, мыши и мухи, позволяет предположить, что эта молекула является центральной в патогенезе болезней телец Леви.
Было показано, что несколько различных антител к альфа-синуклеину оказывают терапевтический эффект в доклинических животных моделях. Было показано, что как антитело, нацеленное на эпитоп, включающий остатки 91-99 альфа-синуклеина, так и антитела, нацеленные на эпитоп, включающий остатки 118-126 альфа-синуклеина, оказывают влияние на двигательные и когнитивные нарушения у трансгенных мышей (Games et al. 2014, J Neurosci. 2014 Jul 9;34(28):9441-54). Наиболее совершенным из этих антител является гуманизированное антитело на основе моноклонального антитела 9Е4 мыши, которое нацелено на эпитоп, который включает остатки 118-126 альфа-синуклеина, и которое сейчас находится на I фазе клинических испытаний. Также было показано, что С-концевое антитело 274, которое нацелено на эпитоп, который включает остатки 120-140 альфа-синуклеина (Bae et al. 2012, J Neurosci. 2012 Sep 26;32(39): 13454-69), проявляет свой эффект в доклинической модели на распространение патологии среди клеток. Кроме этого, было показано, что антитела, нацеленные на такие конформационные разновидности, как олигомеры и фибриллы, альфа-синуклеина, способны по меньшей мере снижать уровни этих предположительно токсичных разновидностей альфа-синуклеина ( et al. 2014, Neurobiol Dis. 2014 Sep; 69:134-43 и Spencer et al. 2014, Mol Ther. 2014 Oct; 22(10): 1753-67). Также было показано, что эти конформационные антитела, которые снижают уровни олигомеров альфа-синуклеина in vivo, такие как mab47, нацелены на эпитопы на С-конце альфа-синуклеина, охватывающие аминокислоты 121-125 (US 20120308572). Другие конформационные, специфические к фибриллам и олигомерам антитела также нацелены на С-концевые последовательности (Vaikath et al. Neurobiol Dis. 2015;79:81-99).
et al. 2014, Neurobiol Dis. 2014 Sep; 69:134-43 и Spencer et al. 2014, Mol Ther. 2014 Oct; 22(10): 1753-67). Также было показано, что эти конформационные антитела, которые снижают уровни олигомеров альфа-синуклеина in vivo, такие как mab47, нацелены на эпитопы на С-конце альфа-синуклеина, охватывающие аминокислоты 121-125 (US 20120308572). Другие конформационные, специфические к фибриллам и олигомерам антитела также нацелены на С-концевые последовательности (Vaikath et al. Neurobiol Dis. 2015;79:81-99).
Поскольку токсичная форма альфа-синуклеина является неизвестной, терапевтическое антитело в идеале должно быть способно связываться с большинством разновидностей альфа-синуклеина, которые образуются в результате альтернативного сплайсинга или посттрансляционных модификаций, таких как усечения, а также с олигомерными и фибриллярными формами. Одна проблема с существующими антителами, которые были протестированы в качестве терапевтических средств в доклинических моделях, как рассмотрено выше, заключается в том, что многие из них нацелены на С-концевые эпитопы, которые не встречаются у некоторых основных усеченных форм альфа-синуклеина. Например, аминокислотами, которые важны для связывания 9Е4, являются аспарагин 122 и тирозин 125 (по результатам аланинового сканирования, представленным в патенте US 20140127131), и это означает, что это антитело не может связывать разновидности альфа-синуклеина, которые усечены по аминокислотам 119 и 122 и являются одними из основных усеченных разновидностей в ткани головного мозга, пораженной болезнью Паркинсона (Kellie et al. Sci Rep. 2014;4:5797). Аналогичная ситуация обстоит с антителом 274 и антителом mab47 (US 8632776). Также, аминоконцевые антитела могут не быть способны связываться с некоторыми из основных усеченных разновидностей альфа-синуклеина, у которых отсутствуют первые аминокислоты, такими как альфа-синуклеин, усеченный до аминокислот 5-140. В случае антитела 9Е4, одним из предложенных механизмов действия является предотвращение усечения по аминокислотам 119-122 во внеклеточном пространстве, поскольку антитело будет связываться с той же областью, что и протеаза, которая будет расщеплять альфа-синуклеин (Games et al. 2014, J Neurosci. 2014 Jul 9;34(28):9441-54). Схожий механизм действия можно также обнаружить в случае антител в непосредственной близости от этого сайта, и поэтому можно ожидать, что вокруг этой области такой активностью будут обладать многие антитела.
Существуют некоторые полученные на животных моделях подтверждения о токсичной роли усеченных разновидностей альфа-синуклеина. Было показано, что экспрессия усеченного альфа-синуклеина под управлением тирозин-гидроксилазного промотора приводит к развитию нигростриальной патологии, которая обычно не наблюдается в моделях с трансгенным альфа-синуклеином (Tofaris et al. 2006, J Neurosci. 2006 Apr 12;26(15):3942-50; Wakamatsu et al. 2006, Neurobiol Aging. 2008 Apr; 29(4):574-85). Например, экспрессия аминокислот 1-130 белка альфа-синуклеина человека, имеющего мутацию А53Т, вызывала эмбриональную утрату дофаминергических нейронов в компактном слое черного вещества, тогда как экспрессия полноразмерного белка не вызывала (Wakamatsu et al. 2006, Neurobiol Aging. 2008 Apr; 29(4):574-85). Экспрессия молекулы альфа-синуклеина размером в 120 аминокислот под управлением промотора кальций/кальмодулин-зависимой протеинкиназы II альфа (CamKII-альфа) была ассоциирована с агрегированием альфа-синуклеина и прогрессирующим нарушением в тестах памяти кортикально-гиппокампальных областей, включая лабиринт Барнеса и распознавание новых объектов (Hall et al. 2015, Exp Neurol. 2015 Feb; 264:8-13). Также, в крысиной AAV-модели совместная экспрессия усеченного на С-конце альфа-синуклеина увеличивала индуцированную полноразмерным альфа-синуклеином патологию (Ulusoy et al. 2010, Eur J Neurosci. 2010 Aug; 32(3):409-22).
В настоящем изобретении были созданы антитела (такие как "GM37" и "GM285", описанные примерах), которые могут связываться с токсичным фрагментом альфа-синуклеина 1-119/122 и нейтрализовать эту усеченную форму альфа-синуклеина. Антитела по настоящему изобретению, такие как GM37 и GM285, могут связываться с другими олигомерными формами альфа-синуклеина и изменять их поглощение другими резидентными клетками ЦНС таким образом, чтобы в результате уменьшалось распространение заболевания. Кроме того, неожиданно было обнаружено, что антитела по настоящему изобретению, такие как GM37 и 285, превосходят антитела, известные из уровня техники, такие как 9Е4, в связывании с различными разновидностями альфа-синуклеина в головном мозге человека и оказывают неожиданно превосходный эффект на устранение внеклеточного альфа-синуклеина и нормализацию нарушенной синаптической передачи, индуцированной присутствием аномального альфа-синуклеина in vivo. В качестве дополнительной иллюстрации их терапевтических возможностей можно привести, что антитела по настоящему изобретению, такие как GM37 и 285, могут предотвращать появление связанного с заболеванием двигательного фенотипа в крысиной модели болезни Паркинсона. Наконец, антитела GM37 и GM285 могут подавлять затравочное действие в отношении агрегирования и фосфорилирования эндогенного альфа-синуклеина, индуцированное внеклеточными затравочными частицами рекомбинантного патологического альфа-синуклеина, в первичных нейронах мышей. Антитела, такие как GM37 и 285, также могут подавлять затравочное действие в отношении развития альфа-синуклеиновой патологии в дофаминергических нейронах in vivo в мышиной модели болезни Паркинсона, что еще больше подтверждает терапевтическую способность этих антител предотвращать распространение патологии от клетки к клетки. Вместе взятые эти данные убедительно подтверждают возможность применения этих новых антител GM37 и GM285 в качестве новых терапевтических средств, способных модифицировать заболевание путем подавления механизма распространения патологических изменений, связанных с заболеванием, среди нейронов у пациентов с болезнью Паркинсона.
Согласно дополнительному аспекту настоящего изобретения предусмотрены 3 аминокислотных варианта антитела GM37. Все варианты имеют схожие функциональные показатели с исходным антителом GM37, но с улучшенными свойствами для возможности производства. Эти варианты уменьшают риск посттрансляционной модификации, происходящей в связывающем домене антитела GM37, и обеспечивают некоторое облегчение получения антитела. Это является преимущественным, поскольку крупномасштабное клиническое или коммерческое изготовление антител является сложным и дорогостоящим, а обеспечение однородного продукта в фармацевтических лекарственных препаратах имеет решающее значение, в частности, в случае иммуноглобулинов и белков.
Краткое описание изобретения
Настоящее изобретение относится к новым моноклональным антителам и их антигенсвязывающим фрагментам, которые способны специфически связываться с эпитопом в пределах аминокислот 112-117 в альфа-синуклеине (SEQ ID NO: 9 (ILEDMP)). Эпитоп, связываемый антителами или антигенсвязывающими фрагментами по настоящему изобретению, такими как иллюстративное антитело "GM37" или "GM285", в данном документе называют "эпитопом 112-117". Антитела по настоящему изобретению специфически связываются с эпитопом в эпитопе 112-117 и могут, в соответствии с одним вариантом осуществления, конкурировать с антителом GM37 или GM285 за связывание с эпитопом в пределах аминокислот 112-117. Например, антитела или их антигенсвязывающие фрагменты по настоящему изобретению могут конкурировать за связывание с эпитопом в пределах аминокислот 112-117 альфа-синуклеина человека, при этом тяжелая цепь состоит из вариабельного домена с SEQ ID NO: 7, а легкая цепь состоит из вариабельного домена с SEQ ID NO: 8. Такое конкурентное ингибирование связывания можно определить с помощью анализов и способов, хорошо известных из уровня техники, например, с помощью анализа немеченого связывания, такого как поверхностный плазмонный резонанс (SPR). Например, иммобилизация альфа-синуклеина человека на поверхности и инкубирование с эталонным антителом "GM37" или без него до инкубирования с антителом или связывающим фрагментом, подлежащими тестированию. В качестве альтернативы, можно применять подход попарного картирования, при котором эталонное антитело "GM37" иммобилизируют на поверхности, антиген альфа-синуклеин человека связывают с иммобилизированным антителом, а затем тестируют второе антитело в отношении способности одновременно связываться с альфа-синуклеином человека (см. 'BIAcore® Assay Handbook', GE Healthcare Life Sciences, 29-0194-00 AA 05/2012, раскрытия которого включены в данный документ посредством ссылки).
Более конкретно, антитело GM285 связывает эпитоп в пределах остатков 112-117 альфа-синуклеина, содержащий остатки 112-115 альфа-синуклеина (ILED; SEQ ID NO: 19).
Согласно одному варианту осуществления настоящее изобретение относится к моноклональному антителу GM37, его вариантам (например, варианту 1 GM37, варианту 2 GM37 и варианту 3 GM37), или GM285.
В частности, настоящее изобретение предусматривает моноклональное антитело GM37, его варианты (например, вариант 1 GM37, вариант 2 GM37 и вариант 3 GM37), или GM285, и охватывает такие антитела, а также их производные, которые обладают достаточным количеством (например, 1, 2 или 3) CDR легкой цепи, и достаточным количеством (например, 1,2 или 3) CDR тяжелой цепи для образования сайта связывания, способного специфически связываться с синуклеином человека. Предпочтительно такие антитела будут обладать тремя CDR легкой цепи и тремя CDR тяжелой цепи, как определено ниже. Нумерация аминокислотных остатков в этой области соответствует IMGT®, международной информационной системе ImMunoGeneTics® или Kabat, Е.A., Wu, Т.Т., Perry, Н.М., Gottesmann, К. S. & Foeller, С. (1991). Sequences of Proteins of Immunological Interest, 5-е издание, U.S. Department of Health and Human Services, публикация NIH №91-3242; Chothia, C. & Lesk, A. M. (1987). Canonical structures For The Hypervariable domains Of Immunoglobulins. J. Mol. Biol. 196, 901-917.
Согласно одному варианту осуществления моноклональное антитело или его антигенсвязывающие фрагменты обладают связывающим синуклеиновый антиген фрагментом, содержащим или состоящим из
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 1; и/или
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 2; и/или
(c) CDR3 тяжелой цепи с аминокислотной последовательностью SEQ ID NO: 3; и/или
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4; и/или
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и/или
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6;
который может специфически связываться с альфа-синуклеином человека.
Согласно другому варианту осуществления моноклональное антитело или его антигенсвязывающие фрагменты обладают связывающим синуклеиновый антиген фрагментом, содержащим или состоящим из
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: l;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 33, 34 или 35;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 3;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6;
который может специфически связываться с альфа-синуклеином человека.
Согласно еще одному варианту осуществления моноклональное антитело или его антигенсвязывающие фрагменты обладают связывающим синуклеиновый антиген фрагментом, содержащим или состоящим из
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 20; и/или
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 21; и/или
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 22; и/или
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 23; и/или
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 24; и/или
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 25.
который может специфически связываться с альфа-синуклеином человека.
Согласно одному варианту осуществления моноклональное антитело или его антигенсвязывающие фрагменты обладают связывающим синуклеиновый антиген фрагментом, содержащим аминокислотную последовательность (в его CDR, его вариабельных доменах, его каркасных остатках или в его константных доменах), которая отличается от последовательности встречающихся в природе антител к альфа-синуклеину и которая характеризуется (относительно таких встречающихся в природе антител к альфа-синуклеину)
(i) различием в аффинности связывания (KD) к альфа-синуклеину;
(ii) различием в способности подавлять усечение протеазами фибрилл альфа-синуклеина;
(iii) различием в способности обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
(iv) различием в способности снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo; и/или
(v) различием в способности, при длительном введении, восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
(vi) различием в способности предотвращать затравочное действие альфа-синуклеина (такое как накопление нерастворимого фосфорилированного альфа-синуклеина in vitro и/или в мышиной модели болезни Паркинсона); и/или
(vii) различием в способности связывать усеченный альфа-синуклеин в головном мозге человека.
Антитела и их антигенсвязывающие фрагменты по настоящему изобретению можно применять в способе лечения, диагностики или визуализации синуклеопатий, таких как болезнь Паркинсона ((PD), в том числе идиопатическая и наследственная формы болезни Паркинсона), болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBV), болезнь Гоше (GD), комбинированная болезнь Альцгеймера и Паркинсона (CAPD), истинная вегетативная недостаточность и множественная системная атрофия.
Краткое описание графических материалов
На фиг. 1 показаны протоколы иммунизации для получения гибридом. В таблице указаны различия иммуногенов и линий мышей, которые были использованы для выявления GM37 и GM285. Различных мышей HCo17-Balb/c и HCo12/Balb/c иммунизировали независимо (описание этих мышей приведено ниже). Гибридома, экспрессирующая GM37, была выявлена у мышей, иммунизированных фибриллами полноразмерного альфа-синуклеина, содержащего аминокислоты 1-140, и стимулированных усеченными фрагментами альфа-синуклеина 1-60 и 1-119 от полноразмерного (FL) альфа-синуклеина (SEQ ID NO 10). Гибридома, экспрессирующая антитело GM285, получена согласно протоколу иммунизации, в котором мышей HCo12-Balb/c иммунизировали полноразмерным мономерным альфа-синуклеином, с аминокислотами 1-140, с последующим стимулированием полноразмерным фибриллярным альфа-синуклеином (пример 1).
На фиг. 2 (секции А-С) показаны результаты скрининга GM37 в отношении связывания с альфа-синуклеином, гомологами и ортологами альфа-синуклеина.
A) Связывание антитела GM37 с альфа-синуклеином по результатам ELISA без промывочного раствора (FMAT).
B) По результатам SPR (Fortebio) связывание антитела GM37 является специфическим к альфа-синуклеину (секция "альфа"), и оно не связывает другие родственные белки семейства синуклеинов, бета-синуклеин (секция "бета") и гамма-синуклеин (секция "гамма"). Измерения проводили с помощью SPR (Fortebio Octetred). GM37 демонстрирует схожее связывание с альфа-синуклеином от яванского макака (секция Cyno) и мыши (секция "мышь"). (Пример 1).
С) По результатам SPR (Fortebio Octetred) связывание антитела GM285 является специфическим к альфа-синуклеину, и оно не связывает другие родственные белки семейства синуклеинов, бета-синуклеин и гамма-синуклеин. Измерения проводили с помощью SPR (Fortebio Octetred), которое продемонстрировало схожее связывание GM285 с альфа-синуклеином от яванского макака (Cyno) и мыши ("мышь") (пример 1).
На фиг. 3 (секции А-С) показана аффинность связывания GM37 в режиме реального времени.
A) Связывание антитела GM37 с альфа-синуклеином, измеренное в RU (относительных единицах) (ось у), с течением времени (ось X), определенное с помощью SPR (BIAcore® 3000). IgG козы к антителам человека был иммобилизирован на чипе СМ5. GM37 захватывали на чипе с иммобилизированными IgG козы к антителам человека и тестировали серию концентраций альфа-синуклеина человека (3,125, 6,25, 12,5, 25, 50, 100 нМ) в отношении связывания с поверхностью. В каждом промежутке между циклами восстанавливали поверхность сенсора.
B) Сигнал от связывания в различных концентрациях, преобразованный в кривую связывания.
C) Рассчитанные константы связывания антитела GM37 (обозначенного hlgG1-6004-037-C106S) (пример 2).
На фиг. 4 (секции А-С) показана аффинность связывания GM285 в режиме реального времени.
А) Связывание антитела GM285 с альфа-синуклеином, измеренное в RU (ось у), с течением времени (ось X), определенное с помощью SPR (BIAcore® 3000). IgG козы к антителам человека был иммобилизирован на чипе СМ5. GM285 захватывали на чипе с иммобилизированными IgG козы к антителам человека и тестировали серию концентраций альфа-синуклеина человека (3,125, 6,25, 12,5, 25, 50, 100 нМ) в отношении связывания с поверхностью. В каждом промежутке между циклами восстанавливали поверхность сенсора.
B) Сигнал от связывания в различных концентрациях, преобразованный в кривую связывания.
C) Рассчитанные константы связывания антитела GM285 (обозначенного hlgG1-6004-285) (пример 2).
На фиг. 5 (секции А-С) показано связывание антитела сравнения 9Е4 в режиме реального времени.
A) Показано связывание 9Е4 с альфа-синуклеином, измеренное в RU (ось у), с течением времени (ось X), определенное с помощью SPR (BIAcore® 3000). IgG козы к антителам человека был иммобилизирован на чипе СМ5. 9Е4 захватывали на чип путем его связывания с IgG козы к антителам человека, который был иммобилизирован на чипе. Тестировали серию концентраций альфа-синуклеина человека (3,125, 6,25, 12,5, 25, 50, 100 нМ) в отношении связывания с поверхностью. В каждом промежутке между циклами восстанавливали поверхность сенсора.
B) Сигнал от связывания в различных концентрациях, преобразованный в кривую связывания.
C) Рассчитанные константы связывания для антитела 9Е4. (Пример 2).
На фиг. 6 показана аминокислотная последовательность альфа-синуклеина. Основные сайты усечения (обозначенные стрелками), идентифицированные посредством масс-спектрометрии в ткани головного мозга человека (Kellie JF, Higgs RE, Ryder JW, Major A, Beach TG, Adler CH, Merchant K, Knierman MD. Quantitative measurement of intact alpha-synuclein proteoforms from post-mortem control and Parkinson's disease brain tissue by mass spectrometry. Sci Rep. 2014 Jul 23;4:5797. doi: 10.1038/srep05797)
На фиг. 7 (секции A-B) показаны результаты картирования эпитопа для антитела GM37 и GM285. Данные ELISA, демонстрирующие относительные уровни связывания антител с последовательными пептидами (из 20 мономеров), полученными из аминокислотной последовательности 95-132 альфа-синуклеина (другие несвязывающиеся пептиды не показаны).
A) Эпитопу GM37 для полного связывания необходима последовательность ILEDMP (SEQ ID NO: 9).
B) GM285 для полного связывания необходим пептид ILED (SEQ ID NO: 19). (Пример 3).
На фиг. 8 (секции А-В) показано схематическое представление усеченных форм альфа-синуклеина.
A) Эпитопы связывания для GM37/285 (ILEDMP; SEQ ID NO: 9) и 9E4 (NEAYE; SEQ ID NO: 36) показаны жирным на аминокислотной последовательности альфа-синуклеина (SEQ ID NO: 10). Стрелки указывают С-концевые сайты усечения, показанные на фиг. 6.
B) Основные усеченные формы альфа-синуклеина, которые были идентифицированные из материала головного мозга человека. Размер, исходя из количества аминокислот указан справа. Полноразмерный синуклеин составляет 140 аминокислот. Как можно высчитать из эпитопов, GM37, его варианты 1-3 и GM285 должны связывать полноразмерную молекулу и фрагменты 1-119/122, 1-135. Антитело 9Е4 будет связываться только с полноразмерной молекулой и фрагментом 1-135. Специфичность меньших С-концевых фрагментов, оставшихся после усечений, не показана.
На фиг. 9 показано, что антитела GM37 и GM285 образуют иммунопреципитат с полноразмерным альфа-синуклеином, а также с усеченным альфа-синуклеином из головного мозга человека. Неочищенные гомогенаты головного мозга человека с DLB инкубировали с тестируемыми антителами (гранулы (без Ab), с контрольным антителом IgG1 человека В12, которое не связывается с альфа-синуклеином, GM-37, вариантом 2 GM37, GM-285 и (m)9Е4) мыши и на SDS-PAGE разделяли иммуноистощенный супернатант и иммунопреципитированный материал. На вестерн-блоте видны полосы, представляющие собой полноразмерную форму и различные усеченные формы альфа-синуклеина, истощаемые из супернатанта и образующие иммунопреципитат с антителами (IP). Как можно видеть, антитело GM37, GM37v2 и GM285 истощало основные разновидности альфа-синуклеина из супернатанта, и IP демонстрирует эти разновидности: усеченные разновидности 1-135, 1-119/122 и полноразмерный альфа-синуклеин. 9Е4 не влияет на разновидности 1-119/122, а только лишь IP с полноразмерным и 1-135 (пример 4).
На фиг. 10 показано схематическое изображение протеолиза фибрилл альфа-синуклеина, расщепляемых кальпаином по аминокислоте 119/122. Фибриллы альфа-синуклеина (PFF) добавляют к культуре с тестируемым антителом (PFF+) или без него (PFF). Присутствие GM-37/285 подавляет образование усеченного альфа-синуклеина в клетках и его секрецию в клеточную среду.
На фиг. 11А показано, что GM37 подавляет образование полосы усеченной формы (12 кДа) как в средах, так и в клеточных лизатах первичных культур клеток коры мышей, обработанных с помощью PFF. Белки разделяли с помощью SDS-PAGE и подвергали вестерн-блоттингу для обнаружения различных разновидностей альфа-синуклеина. В клетках, обработанных только PFF или контрольным антителом (В12), обнаруживается две полосы мономерного альфа-синуклеина на уровне 12 и 14 кДа, представляющие собой соответственно усеченный и полноразмерный альфа-синуклеин. В присутствии GM-37 имеет место только слабая полоса на уровне 12 кДа, что указывает на то, что большая часть продукта расщепления заблокирована. Этот эффект также отражается в средах клеток. Относительные уровни накопления также могут быть ингибированы присутствием GM-37, что отражается в уменьшении относительной интенсивности полосы на уровне 14 кДа. В качестве альтернативы может уменьшаться количество полосы 14 кДа, доступное для поглощения клетками. (Пример 5).
На фиг. 11В показано дозозависимое подавление протеолиза фибрилл альфа-синуклеина антителами GM37, вариантом 2 GM37 и GM285. В клеточных лизатах из первичных культур клеток коры мыши с низкой концентрацией антител (0,1 мкг/мл) присутствует как полоса, представляющая собой полноразмерный альфа-синуклеин (FL), так и полоса, представляющая усеченный на С-конце (СТ) альфа-синуклеин (показано стрелками). Увеличение концентрации антител до 1, 5 и 10 мкг/мл приводит к снижению протеолиза фибрилл альфа-синуклеина в клетках. Это наблюдается как с антителом GM37, так и с GM37v2 и с антителом GM285. Контрольные образцы обрабатывают антителом IgG1 человека В12, которое не распознает альфа-синуклеин. Также присутствует контроль без добавления антител (без Ab), а также клетки, не содержащие фибриллы альфа-синуклеина (без Asyn). Общее количество альфа-синуклеина также снижается в образцах, обработанных 37, 37v2 и 285, по сравнению с контролем В12 или "без антитела", что указывает на то, что все три антитела уменьшают накопление альфа-синуклеина в клетках зависимым от концентрации образом. Полоса актина на верхней части геля демонстрирует равную загрузку образцов (пример 5).
На фиг. 12 показано влияние GM37 и GM285 на затравочное действие в отношении агрегирования альфа-синуклеина и фосфорилирования альфа-синуклеина в первичных кортикальных нейронах мыши.
12А) Пример изображений первичных нейронов, окрашенных по фосфорилированному альфа-синуклеину, который проявляется в виде пятен или точечного окрашивания в клетках, при затравочном действии на клетки либо 1 нг чистых затравочных молекул, либо неочищенными затравочными частицами альфа-синуклеина.
12В) Вестерн-блот белков из первичных кортикальных нейронов, разделенных на растворимую и нерастворимую фракции. Блоты окрашивали специфическим к альфа-синуклеину человека антителом (4 В12/Н a-syn), специфическим кфосфо-Ser-129-альфа-синуклеину антителом (ab51253/pS-a-Syn) и специфическим к альфа-синуклеину мыши антителом (D37A2/M a-syn), и было видно, что добавление неочищенных затравочных частиц в первичных нейронах приводит к накоплению эндогенного альфа-синуклеина мыши и фосфорилированного альфа-синуклеина и мультимеров альфа-синуклеина с более высокой молекулярной массой в нерастворимой фракции.
12С) GM37, вариант 2 GM37 и GM285 подавляют появление фосфорилированного альфа-синуклеина, количественно определяемого как количество положительных пятен альфа-синуклеинового фосфосерина 129 в клетках с помощью автоматизированного микроскопа ARRAYSCAN™ от Cellomics. GM37, GM37v2 и GM285 уменьшают количество пятен фосфорилированного альфа-синуклеина в клетках дозозависимым образом.
12D) Из вестерн-блота гомогенатов из первичных кортикальных нейронов, обработанных наиболее высокой дозой антитела (133 нМ) и окрашенных по актину, альфа-синуклеину человека, фосфорилированному альфа-синуклеину и альфа-синуклеину мыши, видно, что антитела 37, 37v2 и 285 подавляют усечение неочищенных затравочных частиц альфа-синуклеина, поглощаемых клетками, в нерастворимой фракции. Все антитела также подавляют накопление фосфорилированных, эндогенных мышиных и высокомолекулярных мультимеров фосфорилированного альфа-синуклеина мыши в нерастворимой фракции. Полоса актина на верхней части геля демонстрирует равную загрузку образцов (пример 6).
На фиг. 13 показана базальная синаптическая передача в синапсе СА1 коллатерали Шеффера в гиппокампе трансгенных F28-snca и контрольных мышей соответствующего возраста. Полевые возбуждающие постсинаптические потенциалы (fEPSP) вызывали одним стимулом, прилагаемым к коллатералям Шеффера, а базальную синаптическую передачу оценивали путем измерения градиента fEPSP в зависимости от интенсивности стимуляции. Краткосрочную синаптическую пластичность оценивали по стимуляции индукции двойных импульсов. Различные интенсивности стимуляции составляли 0, 25, 50, 75, 100, 150, 200, 300, 400 и 500 мкА, и их последовательно прилагали в порядке возрастания с 2-3 повторами для каждой интенсивности. Было обнаружено, что базальная синаптическая передача значительно ухудшается у трансгенных мышей F28-snca, которые сверхэкспрессируют альфа-синуклеин дикого типа, по сравнению с контрольными мышами соответствующего возраста (пример 7).
На фиг 14 показано влияние системного введения однократной дозы 9Е4 человека (15 мг/кг, i.p.) на нарушение базальной синаптической передачи в синапсе СА1 коллатерали Шеффера в гиппокампе трансгенных мышей F28-snca. Полевые возбуждающие постсинаптические потенциалы (fEPSP) вызывали одним стимулом, прилагаемым к коллатералям Шеффера, а базальную синаптическую передачу оценивали путем измерения градиента fEPSP в зависимости от интенсивности стимуляции. Кратковременная обработка посредством h9E4 индуцировала значительное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (Tg-snca+h9E4 по сравнению с Tg-snca+PBS, р=0,002). Тем не менее, обращение при обработке h9E4 было лишь частичным, о чем свидетельствовала значимо более низкая базальная синаптическая передача по сравнению с однопометниками, которых обрабатывали посредством PBS (р=0,007) (пример 7).
На фиг 15 показано влияние системного введения однократной дозы GM37 человека (15 мг/кг, i.p.) или изотипического контрольного антитела (В12) на нарушение базальной синаптической передачи в синапсе СА1 коллатерали Шеффера в гиппокампе трансгенных мышей F28-snca. Полевые возбуждающие постсинаптические потенциалы (fEPSP) вызывали одним стимулом, прилагаемым к коллатералям Шеффера, а базальную синаптическую передачу оценивали путем измерения градиента fEPSP в зависимости от интенсивности стимуляции. Кратковременная обработка посредством GM37 индуцировала полное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (Tg-snca+GM37 по сравнению с Tg-snca+В12, р=0,004) (пример 7).
На фиг 16 показано влияние системного введения однократной дозы GM285 человека (15 мг/кг, i.p.) на нарушения базальной синаптической передачи в синапсе СА1 коллатерали Шеффера в гиппокампе трансгенных мышей F28-snca. Полевые возбуждающие постсинаптические потенциалы (fEPSP) вызывали одним стимулом, прилагаемым к коллатералям Шеффера, а базальную синаптическую передачу оценивали путем измерения градиента fEPSP в зависимости от интенсивности стимуляции. Кратковременная обработка посредством GM285 индуцировала полное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (Tg-snca + GM285 по сравнению с Tg-snca + PBS, р=0,001) (пример 7).
На фиг. 17 (секции А-В) показано влияние системного введения (15 мг/кг, i.p.) 9Е4 человека, GM37 или изотипического контрольного антитела (антитела к HEL) на уровни альфа-синуклеина человека в интерстициальной жидкости (isf) в гиппокампе свободно двигающихся трансгенных мышей F28-snca. Среднее значение двух-трех исходных значений (4 ч. - 6 ч.) до обработки антителом принимали в качестве исходного уровня и принимали за 100% для каждого животного. Различия анализировали с помощью двухфакторного дисперсионного анализа (ANOVA) с повторными измерениями. Исходные уровни альфа-синуклеина человека в гиппокампе составляли 8,1±1,1 нг/мл (среднее ± SEM, n=25, не скорректированные по восстановлению диализного зонда in vitro). Введение GM37 индуцировало большее уменьшение альфа-синуклеина человека в гиппокампе мышей F28 по сравнению как с антителом сравнения, 9Е4 человека, так и контрольным изотопическим антителом к HEL. Моменты времени, в которых наблюдали значимые различия в уровнях альфа-синуклеина среди животных, обрабатываемых GM37 или контрольным антителом, обозначены звездочкой. (Пример 8).
На фиг. 18 показано схематическое представление временного графика для обработки антителами (стрелки вниз), вирусных инъекций и поведенческих оценок в крысиной AAV-модели альфа-синуклеина человека, показанной на фиг. 19 (пример 9).
На фиг. 19 показано, что антитело GM37 может уменьшать паркинсонические двигательные нарушения после длительной обработки у крысиной AAV-модели. Влияние длительной обработки посредством GM37 или PBS у AAV-крыс с альфа-синуклеином человека на двигательную асимметрию оценивают в тесте передних конечностей на асимметричность. Каждую крысу тестировали в отношении использования передних лап путем наблюдения в течение 5 минут. Процент использования правой передней лапы (ипсилатеральной по отношению к инъекции) и использования левых (передних лап, контралатеральных к правым) рассчитывали для каждого животного (показано на оси у) *, ** р<0,05 и 0,01 по сравнению с крысами GFP-PBS. Крысы, обрабатываемые PBS, по-прежнему имеют значительную асимметрию в использовании лап, тогда как животные, обрабатываемые антителом GM37, уже не имеют значительного нарушения. (Пример 9).
На фиг. 20А-20С показано, что длительная обработка антителом GM37 может уменьшить патологическое фосфорилирование альфа-синуклеина, индуцированное введением фибриллярных затравочных частиц патологического альфа-синуклеина в полосатое тело мышей. На фиг. 20А представлено схематическое представление, показывающее относительное время обработки относительно введения затравочных частиц и подсчета клеток. Антитело GM37 вводили за один день до введения фибриллярных затравочных частиц рекомбинантного альфа-синуклеина в заднюю часть полосатого тела мышей, а затем еженедельно в течение шести недель. Режим дозирования представлял собой либо 15 мг/кг i.v., либо 30 мг/кг i.p.На фиг. 20В показан уровень концентрации GM37 в плазме крови, исходя из места введения и дозы. Еженедельно забирали образцы перед введением новой дозы антитела. На фиг. 20С показан уровень концентрации GM37 в csf, исходя из дозы и места введения в конце исследования. На фиг. 20D приведено сравнение количества клеток с положительными по фосфорилированному альфа-синуклеину включениями, подсчитанными с каждого шестого среза в черном веществе, после обработки GM37 или PBS-контролем. Мыши, обработанные GM37 как 15 мг/кг i.v., так и 30 мг/кг i.p., имели значимое уменьшение количества клеток с включениями фосфорилированного альфа-синуклеина по сравнению с мышами, обработанными PBS (пример 10).
На фиг. 21 показаны результаты выравнивания белков α- (SEQ ID NO: 10), β (SEQ ID NO: 37) и γ- (SEQ ID NO: 38) синуклеина человека. Выделены аминокислотные остатки, отличающиеся от α-синуклеина. Гэпы указаны точкой. Номера по SwissProt приведены в скобках.
На фиг. 22 показаны результаты выравнивания ортологов альфа-синуклеина (яванского макака, SEQ ID NO: 39; крысы, SEQ ID NO: 40; мыши, SEQ ID NO: 41). Выделены аминокислотные остатки, отличающиеся от альфа-синуклеина человека (SEQ ID NO: 10). Номера по SwissProt показаны в скобках.
На фиг. 23 показана временная экспрессия GM37 (под названием GM37 дикого типа (wt) и 3 варианта GM37 под названием var 1, 2 и 3 GM37. Звездочка указывает, что данные определены после очистки и нейтрализации белка.  указывает, что данные рассчитаны из выхода, полученного после белка А и нейтрализации, по отношению к масштабу экспрессирующей культуры (0,4 л).
указывает, что данные рассчитаны из выхода, полученного после белка А и нейтрализации, по отношению к масштабу экспрессирующей культуры (0,4 л).
На фиг. 24 показан конкурентный ELISA, измеряющий связывание четырех антител GM37 wt, var 1 GM37, var 2 GM37 и var 3 GM37 с альфа-синуклеином человека. Планшеты, покрытые альфа-синуклеином, применяют для обнаружения количества антитела, оставшегося после предварительного инкубирования в растворе каждого антитела (0,3 мкг/мл), с увеличением концентрации альфа-синуклеина (0-1000 нМ). Все четыре антитела проявляют схожее связывание с альфа-синуклеином.
На фиг. 25 показана таблица, в которой приведено сравнение кинетических параметров скорости связывания GM37wt и вариантов 1-3 с иммобилизированным рекомбинантным альфа-синуклеином человека. Связывание измеряли с помощью SPR, а скорости определяли с использованием алгоритма связывания 1:1 (BIAcore® Т200).
На фиг. 26 приведено сравнение эффекта антител к альфа-синуклеину, оказываемого на уровни фосфорилированного альфа-синуклеина в первичных нейронах мышей, обработанных фибриллярными затравочными частицами патологического альфа-синуклеина. Первичные нейроны обрабатывали затравочными частицами (10 нг) в присутствии или в отсутствие четырех антител: GM37, var 1 GM37, var 2 GM37 и var 3 GM37 (2 мкг). Нейроны фиксировали и окрашивали спустя 3 недели и анализировали с помощью ARRAYSCAN™ от Cellomics в отношении пятен, положительных по фосфорилированному по серину 129 альфа-синуклеину. Клетки, обработанные затравочными частицами отдельно или затравочными частицами плюс изотипическое контрольное антитело (В12), демонстрируют значимо повышенные уровни фосфорилирования. Клетки, обработанные антителом GM37wt и 3 вариантами, могут подавлять фосфорилирование альфа-синуклеина, все они демонстрируют тот же уровень фосфорилирования, что и клетки, которые не получали затравочных частиц. Данные показаны как среднее ± SD, как определено из семи изображений на лунку для пяти лунок. N=2.
На фиг. 27 приведено сравнение зависимого от температуры агрегирования wt GM37, var1, var2 и var3. Образец каждого из антител подвергали постоянному повышению температуры с течением времени и одновременно измеряли уровень агрегирования по многоугловому рассеянию света (Prometheus NT.48, NanoTemper Technologies). Температура для начала агрегирования является схожей для GM37 и вариантов GM37, однако наиболее низкий уровень агрегирования наблюдали для GM37-Var2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ ОПРЕДЕЛЕНИЯ
Используемый в данном документе термин "альфа-синуклеин" является синонимом "белка альфа-синуклеина" и относится к любой из изоформ белка альфа-синуклеина (идентифицируемого, например, в UniProt под номером Р37840, 1-3). Нумерация аминокислот альфа-синуклеина приведена относительно SEQ ID NO: 10, показанной ниже, причем метионин (М) является аминокислотным остатком 1:
SEQ ID NO: 10:
MDVFMKGLSK AKEGVVAAAE KTKQGVAEAA GKTKEGVLYV GSKTKEGVVH GVATVAEKTK EQVTNVGGAV VTGVTAVAQK TVEGAGSIAA ATGFVKKDQL GKNEEGAPQE GILEDMPVDP DNEAYEMPSE EGYQDYEPEA
Настоящее изобретение относится к антителам и к фрагментам антител, которые способны специфически связываться с альфа-синуклеином и, в частности, с альфа-синуклеином человека. В частности, антитела и их фрагменты характеризуются способностью специфически связываться с эпитопом в пределах 112-117 альфа-синуклеина человека.
Термин "антитело" (Ab) в контексте настоящего изобретения относится к молекуле иммуноглобулина или, в соответствии с некоторыми вариантами осуществления настоящего изобретения, фрагменту молекулы иммуноглобулина, которая обладает способностью специфически связываться с эпитопом молекулы ("антигена"). Встречающиеся в природе антитела обычно представляют собой тетрамер, который обычно состоит по меньшей мере из двух тяжелых (Н) цепей и по меньшей мере из двух легких (L) цепей. Каждая тяжелая цепь состоит из вариабельного домена тяжелой цепи (сокращенно обозначенного в данном документе как VH) и константного домена тяжелой цепи, обычно состоящего из трех доменов (CH1, СН2 и СН3). Тяжелые цепи могут относиться к любому изотипу, в том числе IgG (подтипы IgG1, IgG2, IgG3 и IgG4), IgA (подтипы IgA1 и IgA2), IgM и IgE. Каждая легкая цепь состоит из вариабельного домена легкой цепи (сокращенно обозначенного в данном документе как VL) и константного домена легкой цепи (CL). Легкие цепи включают каппа-цепи и лямбда-цепи. Вариабельный домен тяжелой и легкой цепей обычно отвечает за распознавание антигена, тогда как константный домен тяжелой и легкой цепей может опосредовать связывание иммуноглобулина с тканями или факторами хозяина, в том числе с различными клетками иммунной системы (например, эффекторными клетками) и первым компонентом (C1q) классической системы комплемента. VH-и VL-области можно дополнительно подразделить на области гипервариабельности, называемые "определяющими комплементарность областями", которые чередуются с областями с более консервативной последовательностью, называемыми "каркасными областями " (FR). Каждый VH и VL состоит из трех CDR-доменов и четырех FR-доменов, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Вариабельные домены тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Особого внимания заслуживают антитела и их антигенсвязывающие фрагменты, которые были "выделены" так, чтобы они существовали в физической среде, отличной от среды, в которой они могут встречаться в природе, или которые были модифицированы так, чтобы они отличались от встречающегося в природе антитела по аминокислотной последовательности.
Термин "эпитоп" означает антигенную детерминанту, способную специфически связываться с антителом. Эпитопы обычно состоят из поверхностных групп молекул, таких как аминокислоты или сахарные боковые цепи, и обычно имеют специфические характеристики трехмерных структур, а также специфические характеристики заряда. Конформационные и линейные эпитопы отличаются тем, что связывание с первым, но не с последним, всегда утрачивается в присутствии денатурирующих растворителей. Эпитоп может содержать аминокислотные остатки, непосредственно участвующие в связывании, и другие аминокислотные остатки, которые не принимают непосредственного участия в связывании, такие как аминокислотные остатки, которые эффективно блокируются пептидом, специфически связывающимся с антигеном (другими словами, аминокислотный остаток находится в пределах области узнавания пептида, специфически связывающегося с антигеном). Термин "эпитоп 112-117" относится к области альфа-синуклеина человека, который содержит по меньшей мере 4 из 6 аминокислотных остатков 112-117 альфа-синуклеина человека, причем эпитоп не включает ни одного остатка из 1-111 (в том числе любой остаток из 106-111) альфа-синуклеина человека или ни одного остатка из 118-140 (в том числе остаток 118-120) альфа-синуклеина человека. В контексте данного документа считают, что антитело способно специфически связываться с эпитопом в пределах эпитопа 112-117, если оно способно специфически связываться с альфа-синуклеином человека путем связывания по меньшей мере с 4 из 6 аминокислотных остатков эпитопа 112-117.
Применяемый в данном документе термин "антигенсвязывающий фрагмент антитела" означает фрагмент, часть, область или домен антитела (независимо от того, как он получен (например, посредством расщепления, рекомбинантно, синтетически и т.д.)), который может специфически связываться с эпитопом, и, таким образом, термин "антигенсвязывающий" означает то же самое, что и "эпитопсвязывающий", так что, например, "антигенсвязывающий фрагмент антитела" подразумевают как то же, что и "эпитопсвязывающий фрагмент антитела". Антигенсвязывающий фрагмент может содержать 1, 2, 3, 4, 5 или все 6 CDR-доменов такого антитела и, несмотря на то, что он может специфически связываться с таким эпитопом, он может проявлять специфичность, аффинность или селективность в отношении такого эпитопа, который отличается от эпитопа такого антитела. Предпочтительно, тем не менее, чтобы антигенсвязывающий фрагмент содержал все 6 CDR-доменов такого антитела. Антигенсвязывающий фрагмент антитела может представлять собой часть одной полипептидной цепи (например, scFv), или содержать ее, или может представлять собой часть двух или более полипептидных цепей, или содержать их, причем каждая из них имеет аминоконец и карбоксильный конец (например, диатело, Fab-фрагмент, Fab2-фрагмент и т.д.). Фрагменты антител, которые проявляют антигенсвязывающую способность, можно получить, например, в результате протеазного расщепления интактных антител. Более предпочтительно, чтобы, несмотря на то, что два домена Fv-фрагмента, VL и VH, в естественных условиях кодируются отдельными генами или полинуклеотидами, которые кодируют такие последовательности генов (например, их кодирующей кДНК), можно было соединить с помощью рекомбинантных способов гибким линкером, который обеспечивает возможность их получения в виде одной белковой цепи, в которой VL- и VH-области ассоциируются с образованием одновалентных антигенсвязывающих молекул (известных как одноцепочечные Fv (scFv), см., например, Bird et al., (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. (U.S.A.) 85:5879-5883). В качестве альтернативы, при использовании гибкого линкера, который является слишком коротким (например, менее чем приблизительно 9 остатков), для того, чтобы обеспечить возможность ассоциации VL- и VH- областей одной полипептидной цепи друг с другом, можно получить биспецифическое антитело, диатело или подобную молекулу (в которой две таких полипептидных цепи ассоциируют друг с другом с образованием двухвалентной антигенсвязывающей молекулы) (для описания диател см., например, PNAS USA 90(14), 6444-8 (1993)). Примеры антигенсвязывающих фрагментов, охватываемых настоящим изобретением, включают (i) Fab'- или Fab-фрагмент - одновалентный фрагмент, состоящий из VL-, VH-, CL- и СН1-доменов, или одновалентное антитело, описанное в WO 2007059782; (ii) Р(ab')2-фрагменты - двухвалентные фрагменты, содержащие два Fab-фрагмента, соединенные дисульфидным мостиком в шарнирном домене; (iii) Fd-фрагмент, по сути, состоящий из VH- и СН1-доменов; (iv) Fv-фрагмент, по сути, состоящий из VL- и VH-доменов, (v) dAb-фрагмент (Ward et al., Nature 341, 544-546 (1989)), который, по сути, состоит из VH-домена и также называется доменными антителами (Holt et al; Trends Biotechnol. 2003 Nov; 2i(ll): 484-90); (vi) антитела верблюдовых или нанотела (Revets et al; Expert Opin Biol Ther. 2005 Jan;5_(l): l ll-24) и (vii) выделенная определяющая комплементарность область (CDR). Кроме того, несмотря на то, что два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с помощью рекомбинантных способов синтетическим линкером, который обеспечивает возможность их получения в виде одной белковой цепи, в которой VL- и VH-области находятся в паре с образованием одновалентных молекул (известных как одноцепочечные антитела или одноцепочечные Fv (scFv), см., например, Bird et al., Science 242, 423-426 (1988) и Huston et al., PNAS USA 85, 5879-5883 (1988)). Эти и другие фрагменты антител, применимые в контексте настоящего изобретения, дополнительно обсуждаются в данном документе. Также следует понимать, что термин антитело, если не указано иное, также включает антителоподобные полипептиды, такие как химерные антитела и гуманизированные антитела, и фрагменты антител, сохраняющие способность специфически связываться с антигеном (антигенсвязывающие фрагменты), получаемые с помощью любой известной методики, такой как ферментативное расщепление, синтез пептидов и рекомбинантные методики. Полученное антитело может иметь любым изотип. Используемый в данном документе термин "изотип" обозначает класс иммуноглобулина (например, IgG1, IgG2, IgG3 или IgG4), который кодируется генами константного домена тяжелой цепи. Такие фрагменты антител получают с помощью традиционных методик, известных специалистам в данной области техники; подходящие фрагменты, способные связываться с желаемым эпитопом, можно без труда подвергнуть скринингу в отношении полезности таким же образом, как и интактное антитело.
Термин "биспецифическое антитело" обозначает антитело, содержащее два независимых антигенсвязывающих фрагмента, каждый из которых нацеливается на независимые мишени. Эти мишени могут представлять собой эпитопы, присутствующие в различных белках, или различные эпитопы, присутствующие в одной и той же мишени. Молекулы биспецифических антител можно получить с помощью компенсаторных аминокислотных изменений в константных доменах НС исходных молекул моноспецифических двухвалентных антител. Полученное в результате гетеродимерное антитело содержит один Fab, вклад в образование которого вносят два разных исходных моноспецифических антитела. Аминокислотные изменения в Fc-домене приводят к повышенной стабильности гетеродимерного антитела с биспецифичностью, которая стабильна с течением времени. (Ridgway et al., Protein Engineering 9, 617-621 (1996), Gunasekaran et al., JBC 285, 19637-1(2010), Moore et al., MAbs 3:6 546-557 (2011), Strop et al., JMB 420, 204-219 (2012), Metz et al., Protein Engineering 25:10 571-580 (2012), Labrijn et al., PNAS 110:113, 5145-5150 (2013), Spreter Von Kreudenstein et al., MAbs 5:5 646-654 (2013)). Биспецифические антитела также могут включать молекулы, которые получены с помощью слияний ScFv. Затем два моноспецифических scfv независимо соединяют с Fc-доменами, которые могут образовывать стабильные гетеродимеры, с получением одной биспецифической молекулы (Mabry et al., PEDS 23:3 115-127 (2010). Биспецифические молекулы обладают способностями к двойному связыванию. Например, нацеливание, как на терапевтическую мишень, так и на трансцитозный поверхностный рецептор с целью доставки терапевтического антитела через гематоэнцефалический барьер для лечения заболевания ЦНС.
Термины GM37, GM-37, GM37 дикого типа (wt), mab37 и 6004-37 используют в данном документе взаимозаменяемо, и все они относятся к одному и тому же антителу.
Под термином антитело GM37 подразумевают антитело или его антигенсвязывающий фрагмент, содержащий тяжелую цепь, которая приведена в CDR1-3 SEQ ID NO: 1-3, и CDR1-3 легкой цепи, которые приведены в SEQ ID NO: 4-6, или состоящий из них. Согласно одному варианту осуществления антитело GM37 или его антигенсвязывающий фрагмент может содержать вариабельный домен тяжелой цепи с SEQ ID NO: 7 и/или вариабельный домен легкой цепи с SEQ ID NO: 8 или состоять из них. Например, антитело GM37 может представлять собой антитело IgG, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 7 и константного домена с SEQ ID NO: 18, вместе с легкой цепью, состоящей из вариабельного домена с SEQ ID NO: 8 и константного каппа-домена с SEQ ID NO: 17.
Дезаминирование белков, и в этих случаях антител, может происходить спонтанно в ходе изготовления и хранения, но также и in vivo, и оно затрудняет контроль качества конечного фармацевтического лекарственного препарата. Дезаминирование также в некоторых случаях может влиять на активность молекулы. Дезаминирование происходит на аспарагиновых остатках, но локализацию соответствующего аспарагина может быть трудно с уверенностью спрогнозировать, но в некоторых случаях на нее может оказывать влияние аспарагин-глициновый мотив. На антителе GM37 обнаружено несколько возможных мотивов дезаминирования, тем не менее, было обнаружено, что один вероятный сайт дезаминирования находится на остатке 54 тяжелой цепи. Последующая замена аспарагина другой аминокислотой не является непосредственным следствием, но было обнаружено, что 3 варианта GM37 (вариант (var) 1, 2 и 3 GM37) сохраняют активность исходного GM37 (GM37 дикого типа (wt)).
Термин варианты GM37 обозначает дезаминированные варианты 1, 2 или 3, где вариант 1 содержит замену N54S, вариант 2 содержит замену N54Q, а вариант 3 содержит N54H по сравнению с антителом GM37, описанным в данном документе выше.
Таким образом, подразумевают, что вариант (var) 1, 2 и 3 антитела GM37 включает антитело или его антигенсвязывающий фрагмент, содержащий тяжелую цепь, которая приведена в CDR1 и 3 с SEQ ID NO: l и 3 из GM 37 и CDR1-3 легкой цепи из GM37, которые приведены в SEQ ID NO: 4-6, или состоящий из них, но различающийся по их CDR2 тяжелой цепи, так что вариант 1 содержит CDR 2 с SEQ ID NO: 33, вариант 2 содержит CDR 2 с SEQ ID NO: 34, а вариант 3 содержит CDR 2 с SEQ ID NO: 35.
Согласно одному варианту осуществления варианты антитела GM37 или их антигенсвязывающие фрагменты могут содержать вариабельный домен тяжелой цепи с SEQ ID NO: 30, 31 и 32 соответственно для варианта 1, 2 и 3 и вариабельный домен легкой цепи с SEQ ID NO: 8 или состоять из них. Антитело GM37 может представлять собой антитело IgG, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 30, 31 или 32 и константного домена с SEQ ID NO: 18, вместе с легкой цепью, состоящей из вариабельного домена с SEQ ID NO: 8 и константного каппа-домена с SEQ ID NO: 17.
Термины GM285, GM-285, mab285 и 6004-285 используют в данном документе взаимозаменяемо, и все они относятся к одному и тому же антителу.
Под термином антитело GM285 подразумевают антитело или его антигенсвязывающий фрагмент, содержащий тяжелую цепь, которая приведена в CDR1-3 SEQ ID NO: 20-22, и CDR1-3 легкой цепи, которые приведены в SEQ ID NO: 23-25, или состоящий из них. Согласно одному варианту осуществления антитело GM37 или его антигенсвязывающий фрагмент может содержать вариабельный домен тяжелой цепи с SEQ ID NO: 26 и/или вариабельный домен легкой цепи с SEQ ID NO: 27 или состоять из них. Например, антитело GM37 может представлять собой антитело IgG, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 26 и константного домена с SEQ ID NO: 28, вместе с легкой цепью, состоящей из вариабельного домена с SEQ ID NO: 27 и константного каппа-домена с SEQ ID NO: 29.
Антитело GM285 специфически связывает эпитоп в последовательности 112-115 (ILED, SEQ ID NO: 19) альфа-синуклеина человека (SEQ ID NO: 10).
Если в данном документе не указано иное, то нумерация аминокислотных остатков в этой области соответствует IMGT®, международной информационной системе ImMunoGeneTics®, или Kabat, Е.A., Wu, Т.Т., Perry, Н.М., Gottesmann, К. S. & Foeller, С. (1991). Sequences of Proteins of Immunological Interest, 5th edit., NIH Publication no. 91-3242 U.S. Department of Health and HumanServices. Chothia, C. & Lesk, A. M. (1987). Canonical structures for the hypervariable domains of immunoglobulins. J. Mol. Biol. 196, 901-917).
Выражение "антитело к альфа-синуклеину" или "антитело против альфа-синуклеина" (используемое в данном документе взаимозаменяемо в зависимости от контекста, в котором оно написано) представляет собой антитело или его антигенсвязывающий фрагмент, которые специфически связываются с альфа-синуклеином или фрагментом альфа-синуклеина, как определено в данном документе выше, в частности, с последовательностью альфа-синуклеина, соответствующей SEQ ID NO: 9 и/или 19.
Используемый в данном документе термин "антитело человека" (который может быть сокращен до "humAb" или "HuMab") подразумевают как включающий антитела, имеющие вариабельные и константные домены, полученные из последовательностей иммуноглобулина зародышевой линии человека. Антитела человека по настоящему изобретению могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные с помощью неспецифического или сайт специфического мутагенеза in vitro или во время перестройки гена или с помощью соматической мутации in vivo).
Используемые в данном документе термины "моноклональное антитело" или "композиция на основе моноклональных антител" относятся к препарату из молекул антител единого молекулярного состава. Традиционная композиция на основе моноклональных антител характеризуется единой специфичностью связывания и аффинностью к конкретному эпитопу. Согласно определенным вариантам осуществления моноклональное антитело может состоять из более чем одного Fab-домена, за счет чего повышается специфичность более чем к одной мишени. Термины "моноклональное антитело" или "композиция на основе моноклональных антител" не предназначены для ограничения до какого-либо конкретного способа получения (например, рекомбинантного, трансгенного, гибридомного и т.д.).
Термин "гуманизированный" относится к молекуле, обычно получаемой с использованием рекомбинантных методик, имеющей антигенсвязывающий сайт, полученный из иммуноглобулина от вида, отличного от человека, и остальную часть структуры иммуноглобулина, в основе которой лежит структура и/или последовательности иммуноглобулина человека. Антигенсвязывающий сайт может содержать либо полные вариабельные домены антитела, отличного от человеческого, слитые с константными доменами человека, либо только определяющие комплементарность области (CDR) таких вариабельных доменов, привитые на соответствующие человеческие каркасные области вариабельных доменов человека. Остатки каркасных областей таких гуманизированных молекул могут быть дикого типа (например, полностью человеческими), или они могут быть модифицированы так, чтобы содержать одну или несколько аминокислотных замен, которые не встречаются в антителе человека, последовательность которого послужила основой для гуманизации. Гуманизация уменьшает или устраняет вероятность того, что константный домен молекулы будет выступать в роли иммуногена у людей, но сохраняется возможность иммунного ответа на чужеродный вариабельный домен (LoBuglio, A.F. et al. (1989) "Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response," Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход фокусируется не только на получении константных доменов человеческого происхождения, но также и на модификации вариабельных доменов, с тем чтобы реконструировать их как можно более близко к человеческой форме. Известно, что вариабельные домены как тяжелой, так и легкой цепей содержат три определяющие комплементарность области (CDR), которые различаются по ответу на рассматриваемые антигены и определяют способность к связыванию, фланкированные четырьмя каркасными областями (FR), которые являются относительно консервативными у данного вида и которые предположительно исполняют роль остова для CDR. Если отличные от человеческих антитела получают к определенному антигену, вариабельные домены можно "реконструировать" или "гуманизировать" путем прививки CDR, полученных из антитела, отличного от человеческого, на FR, присутствующем в подлежащем модификации антителе человека. О применение такого подхода к различным антителам сообщалось в Sato, К. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) "Reshaping Human Antibodies for Therapy," Nature 332:323-327; Verhoeyen, M. et al. (1988) "Reshaping Human Antibodies: Grafting An Antilysozyme Activity," Science 239:1534-1536; Kettleborough, C. A. et al. (1991) "Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation," Protein Engineering 4:773-3783; Maeda, H. et al. (1991) "Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity," Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) "Reshaping A Therapeutic CD4 Antibody," Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) "Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo," Bio/Technology 9:266-271; Co, M. S. et al. (1991) "Humanized Antibodies For Antiviral Therapy," Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) "Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy," Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) "Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen," J. Immunol. 148:1149-1154. Согласно некоторым вариантам осуществления гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное антитело мыши, которое содержит все шесть CDR антител мыши). Согласно другим вариантам осуществления гуманизированные антитела содержат один или несколько CDR (один, два, три, четыре, пять, шесть), которые изменены относительно исходного антитела и которые также называются одним или несколькими CDR, "полученными из" одного или нескольких CDR исходного антитела. Возможность гуманизации антигена хорошо известна (см., например, патенты США №№5225539, 5530101, 5585089, 5859205, 6407213, 6881557).
В контексте данного документа считают, что антитело или его антигенсвязывающий фрагмент "специфически" связываются с областью другой молекулы (т.е. эпитопом), если они вступают в реакцию или ассоциируются с этим эпитопом чаще, быстрее, с большей продолжительностью и/или с большей аффинностью или авидностью по сравнению с альтернативными эпитопами. Согласно одному варианту осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению связываются по меньшей мере в 10 раз сильнее со своей мишенью (альфа-синуклеином человека), чем с другой молекулой; предпочтительно по меньшей мере в 50 раз сильнее и более предпочтительно по меньшей мере в 100 раз сильнее. Предпочтительно, антитело или его антигенсвязывающий фрагмент связываются в физиологических условиях, например, in vivo. Таким образом, антитело, которое может "специфически связываться" с эпитопом в пределах остатков 112-117 (ILEDMP (SEQ ID NO: 9)) альфа-синуклеина человека, охватывает антитело или его антигенсвязывающие фрагменты, которые могут связываться с эпитопом в пределах остатков 112-117 альфа-синуклеина человека с такой специфичностью и/или в таких условиях. Способы, подходящие для определения такого связывания, будут известны специалистам в данной области техники, а иллюстративные способы описаны в сопровождающих примерах. Используемый в данном документе термин "связывание" в контексте связывания антитела с предопределенным антигеном обычно обозначает связывание с аффинностью, соответствующей KD, составляющей приблизительно 10-7 М или менее, как, например, приблизительно 10-8 М или менее, как, например, приблизительно 10-9 М или менее при определении, например, с помощью технологии поверхностного плазмонного резонанса (SPR) либо на BIAcore® 3000, либо на T200instrument с применением антигена в качестве лиганда и антитела в качестве анализируемого вещества, и которое связывается с предопределенным антигеном с аффинностью, соответствующей KD, которая по меньшей мере в десять раз ниже, как, например, по меньшей мере в 100 раз ниже, например по меньшей мере в 1000 раз ниже, как, например, по меньшей мере в 10000 раз ниже, например, по меньшей мере в 100000 раз ниже, чем его аффинность при связывании с неспецифическим антигеном (например, BSA, казеином), отличным от предопределенного антигена, или близкородственным антигеном. Степень, на которую аффинность является более низкой, зависит от KD антитела, так что если KD антитела является очень низкой (то есть антитело является высокоспецифическим), то степень, на которую аффинность к антигену является более низкой, чем аффинность к неспецифическому антигену, может быть по меньшей мере 10000-кратной.
Используемый в данном документе термин "kd" (с-1 или 1/с) относится к константе скорости диссоциации для конкретного взаимодействия антитела и антигена. Указанное значение также называют значением koff.
Используемый в данном документе термин "ka" (М-1 х с-1 или 1/М с) относится к константе скорости ассоциации для конкретного взаимодействия антитела и антигена.
Используемый в данном документе термин "KD" (М) относится к равновесной константе диссоциации для конкретного взаимодействия антитела и антигена, и ее получают путем деления kd на ka.
Используемый в данном документе термин "KA" (М-1 или 1/М) относится к равновесной константе ассоциации для конкретного взаимодействия антитела и антигена, и ее получают путем деления ka на kd.
Согласно одному варианту осуществления настоящее изобретение относится к антителу или его антигенсвязывающим фрагментам, которые характеризуются одним или несколькими из следующих свойств:
i. аффинностью связывания (KD) к альфа-синуклеину в диапазоне 0,5-10 нМ, например 1-5 нМ или 1-2 нМ;
ii. способностью подавлять усечение протеазами фибрилл альфа-синуклеина;
iii. способностью обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
iv. способностью снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo;
v. способностью, при длительном введении, восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
vi. способностью предотвращать затравочное действие альфа-синуклеина (такое как накопление нерастворимого фосфорилированного альфа-синуклеина in vitro и/или в мышиной модели болезни Паркинсона); и/или
vii. способностью связывать усеченный альфа-синуклеин в головном мозге человека.
Аффинность связывания (KD) к альфа-синуклеину можно определить с помощью способов, хорошо известных из уровня техники, например, описанных в примере 2.
Термин "способность подавлять усечение протеазами фибрилл альфа-синуклеина" включает в себя способность подавлять индуцированное кальпаином-1 образование фрагмента 1-119-122 альфа-синуклеина человека в первичных кортикальных нейронах (см. пример 5).
Термин "способность обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca" включает способность обращать нарушение синаптической передачи и пластичности в области СА1 гиппокампа у трансгенных мышей F28-snca, например, как показано с помощью градиента fEPSP, измеренного электрофизиологическими способами (см. пример 6).
Термин "способность снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo", включает способность снижать уровни альфа-синуклеина человека в гиппокампе бодрствующих, свободно перемещающихся трансгенных мышей F28-snca, по результатам изменения с помощью микродиализа in vivo (см. пример 7).
Термин "способность, при длительном введении, восстанавливать двигательную функцию в крысиной модели болезни Паркинсона", включает способность уменьшать или устранять двигательную асимметрию в крысиной модели болезни Паркинсона с использованием вектора на основе рекомбинантного аденоассоциированного вирусного вектора (rAAV) (см. пример 8).
В некоторых антителах для сохранения связывания в гуманизированном антителе необходима лишь часть CDR, а именно подмножество остатков CDR, необходимых для связывания, называемых SDR. Остатки CDR, не контактирующие с соответствующим эпитопом и не содержащиеся в SDR, можно идентифицировать на основании предыдущих исследований (например, остатки Н60-Н65 в Н2 CDR зачастую не являются необходимыми) из областей CDR по Kabat, расположенных вне гипервариабельных петель по Chothia (см, Kabat et al. (1992) SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST, National Institutes of Health Publication No. 91-3242; Chothia, C. et al. (1987) "Canonical Structures For The Hypervariable domains Of Immunoglobulins," J. Mol. Biol. 196:901-917), с помощью молекулярного моделирования, и/или эмпирическим путем, или как описано в Gonzales, N.R. et al. (2004) "SDR Grafting Of A Murine Antibody Using Multiple Human Germline Templates To Minimize Its Immunogenicity," Mol. Immunol. 41:863-872. У таких гуманизированных антител в положениях, в которых отсутствует один или несколько остатков донорного CDR или в которых отсутствует весь донорный CDR, аминокислота, занимающая такое положение, может представлять собой аминокислоту, занимающую соответствующее положение (согласно нумерации по Kabat) в акцепторной последовательности антитела. Количество таких замен акцепторных аминокислот на донорные в CDR, которые нужно включить, отражает баланс альтернативных мнений. Такие замены потенциально являются преимущественными для снижения количества аминокислот мыши в гуманизированном антителе и, следовательно, снижения потенциальной иммуногенности. Тем не менее, замены также могут вызывать изменения аффинности, и предпочтительно избегать значительных уменьшений аффинности. Положения для замены в пределах CDR и подлежащие замене аминокислоты также можно выбрать эмпирическим путем.
Тот факт, что изменение одной аминокислоты в остатке CDR может привести к утрате функционального связывания (Rudikoff, S. etc. (1982) "Single Amino Acid Substitution Altering Antigen-Binding Specificity," Proc. Natl. Acad. Sci. (USA) 79(6): 1979-1983), обеспечивает возможность систематической идентификации альтернативных функциональных последовательностей CDR. В одном предпочтительном способе получения таких вариантов CDR полинуклеотид, кодирующий CDR, подвергают мутации (например, посредством случайного мутагенеза или с помощью сайт-направленного способа (например, с амплификацией, опосредованной полимеразной цепной реакцией, с праймерами, которые кодируют мутантный локус)) с получением CDR, имеющий замененный аминокислотный остаток. Путем сравнения идентичности исходной (функциональной) последовательности CDR по соответствующему остатку с идентичностью варианта последовательности CDR с заменой (нефункционального) для этой замены можно определить вес замены согласно BLOSUM62.iij. В системе BLOSUM представлена матрица аминокислотных замен, создаваемая путем анализа базы данных последовательностей в отношении достоверных выравниваний (Eddy, S.R. (2004) "Where Did The BLOSUM62 Alignment Score Matrix Come From?," Nature Biotech. 22(8):1035-1036; Henikoff, J.G. (1992) "Amino acid substitution matrices from protein blocks," Proc. Natl. Acad. Sci. (USA) 89:10915-10919; Karlin, S. et al. (1990) "Methods For Assessing The Statistical Significance Of Molecular Sequence Features By Using General Scoring Schemes," Proc. Natl. Acad. Sci. (USA) 87:2264-2268; Altschul, S.F. (1991) "Amino Acid Substitution Matrices From An Information Theoretic Perspective," J. Mol. Biol. 219, 555-565. В настоящее время наиболее совершенной базой данных BLOSUM является база данных BLOSUM62 (BLOSUM62.iij). В таблице 1 представлены значения веса замен согласно BLOSUM62.iij (чем больше вес, тем более консервативной является замена и, таким образом, тем более вероятно, что замена не будет влиять на функцию). Если, например, антигенсвязывающий фрагмент, содержащий полученный в результате CDR, не может связываться с альфа-синуклеином, то вес замены согласно BLOSUM62.iij считают недостаточно консервативной и выбирают и производят новую кандидатную замену, имеющую больший вес замены. Так, например, если исходный остаток представлял собой глутамат (Е), а нефункциональный заменяющий остаток представлял собой гистидин (Н), то вес замены согласно BLOSUM62.iij будет равен 0, а более консервативные изменения (например, аспартат, аспарагин, глутамин или лизин) являются предпочтительными.
Таким образом, настоящее изобретение предусматривает применение случайного мутагенеза для идентификации улучшенных CDR. В контексте настоящего изобретения консервативные замены могут быть определены как замены в пределах классов аминокислот, отраженных в одной или нескольких из следующих трех таблиц.
Классы аминокислотных остатков для консервативных замен

Альтернативные классы аминокислотных остатков для консервативных замен
Альтернативные физические и функциональные классификации аминокислотных остатков

Группы более консервативных замен включают валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин и аспарагин-глутамин.
Дополнительные группы аминокислот также могут быть составлены с помощью принципов, описанных, например, в Creighton (1984) Proteins: Structure and Molecular Properties (2d Ed. 1993), W.H. Freeman and Company.
В качестве альтернативы можно применять технологию фагового дисплея для повышения (или снижения) аффинности CDR. Эта технология, называемая созреванием аффинности, предусматривает использование мутагенеза или "прогулки по CDR", а повторный отбор предусматривает применение целевого антигена или фрагмента антигена, с которым происходит связывание, для выявления антител, имеющих CDR, которые связываются с более высокой (или более низкой) аффинностью с антигеном по сравнению с первоначальным или исходным антителом (см., например, Glaser et al. (1992) J. Immunology 149:3903). Мутагенез целых кодонов, а не отдельных нуклеотидов, приводит к получению полуслучайного набора аминокислотных мутаций. Можно конструировать библиотеки, состоящие из пула вариантов клонов, каждый из которых отличается одним изменением аминокислоты в одном CDR и который содержит варианты, представляющие каждую возможную аминокислотную замену для каждого остатка CDR. Мутанты с повышенной (или пониженной) аффинностью связывания с антигеном можно подвергнуть скринингу путем приведения в контакт иммобилизированных мутантов с меченым антигеном. Для идентификации мутантных антител с повышенной или пониженной аффинностью к антигену можно применять любой известный из уровня техники способ скрининга (например, ELISA) (см. Wu et al. 1998, Proc. Natl. Acad. Sci. (U.S.A.) 95:6037; Yelton et al., 1995, J. Immunology 155:1994). Возможно применение методики "прогулки по CDR", с помощью которой рандомизируют легкую цепь (см. Schier et al., 1996, J. Mol. Bio. 263:551).
Способы осуществления такого созревания аффинности описаны, например, в Krause, J.C. et al. (2011) "An Insertion Mutation That Distorts Antibody Binding Site Architecture Enhances Function Of A Human Antibody," MBio. 2(1) pii: e00345-10. doi: 10.1128/mBio.00345-10; Kuan, C.T. et al. (2010)"Affinity-Matured Anti-Glycoprotein NMB Recombinant Immunotoxins Targeting Malignant Gliomas And Melanomas" Int. J. Cancer 10.1002/ijc.25645; Hackel, B.J. et al. (2010) "Stability And CDR Composition Biases Enrich Binder Functionality Landscapes," J. Mol. Biol. 401(1):84-96; Montgomery, D.L. et al. (2009) "Affinity Maturation And Characterization Of A Human Monoclonal Antibody Against HIV-1 gp41," MAbs 1(5):462-474; Gustchina, E. et al. (2009) "Affinity Maturation By Targeted Diversification Of The CDR-H2 Loop Of A Monoclonal Fab Derived From A Synthetic  Human Antibody Library And Directed Against The Internal Trimeric Coiled-Coil Of Gp41 Yields A Set Of Fabs With Improved HIV-1 Neutralization Potency And Breadth," Virology 393(1):112-119; Finlay, W.J. et al. (2009) "Affinity Maturation Of A Humanized Rat Antibody For Anti-RAGE Therapy: Comprehensive Mutagenesis Reveals A High Level Of Mutational Plasticity Both Inside And Outside The Complementarity-Determining Regions" J. Mol. Biol. 388(3):541-558; Bostrom, J. et al. (2009) "Improving Antibody Binding Affinity And Specificity For Therapeutic Development," Methods Mol. Biol. 525:353-376; Steidl, S. et al. (2008) "In Vitro Affinity Maturation Of Human GM-CSF Antibodies By Targeted CDR-Diversification," Mol. Immunol. 46(1):135-144; и Barderas, R. et al. (2008) "Affinity Maturation Of Antibodies Assisted By In Silico Modeling," Proc. Natl. Acad. Sci. (USA) 105(26):9029-9034.
Human Antibody Library And Directed Against The Internal Trimeric Coiled-Coil Of Gp41 Yields A Set Of Fabs With Improved HIV-1 Neutralization Potency And Breadth," Virology 393(1):112-119; Finlay, W.J. et al. (2009) "Affinity Maturation Of A Humanized Rat Antibody For Anti-RAGE Therapy: Comprehensive Mutagenesis Reveals A High Level Of Mutational Plasticity Both Inside And Outside The Complementarity-Determining Regions" J. Mol. Biol. 388(3):541-558; Bostrom, J. et al. (2009) "Improving Antibody Binding Affinity And Specificity For Therapeutic Development," Methods Mol. Biol. 525:353-376; Steidl, S. et al. (2008) "In Vitro Affinity Maturation Of Human GM-CSF Antibodies By Targeted CDR-Diversification," Mol. Immunol. 46(1):135-144; и Barderas, R. et al. (2008) "Affinity Maturation Of Antibodies Assisted By In Silico Modeling," Proc. Natl. Acad. Sci. (USA) 105(26):9029-9034.
Таким образом, последовательность вариантов CDR охватываемых антител или их антигенсвязывающих фрагментов, может отличаться от последовательности CDR исходного антитела GM37, GM37 var 1-3 или 285 по заменам; например, по замененному 4 аминокислотному остатку, 3 аминокислотному остатку, 2 аминокислотному остатку или 1 из аминокислотных остатков. Согласно варианту осуществления настоящего изобретения дополнительно предусматривается, что аминокислоты в CDR-областях можно заменять посредством консервативных замен, определенных в 3 таблицах выше.
Используемый в данном документе термин "лечение" или "осуществление лечения" означает облегчение, замедление, ослабление или обращение развития или тяжести заболевания или нарушения или облегчение, замедление, ослабление или устранение одного или нескольких симптомов или побочных эффектов такого заболевания или нарушения. Для целей настоящего изобретения "лечение" или "осуществление лечения" дополнительно означает подход для получения благоприятных или желаемых клинических результатов, где "благоприятные или желаемые клинические результаты" включают без ограничения облегчение симптома, снижение степени нарушения или заболевания, стабилизированное (т.е. не ухудшающееся) состояние заболевания или нарушения, задержку или замедление прогрессирования состояния заболевания или нарушения, облегчение или смягчение состояния заболевания или нарушения и ремиссию заболевания или нарушения, либо частично, либо полностью, поддающаяся обнаружению или не поддающаяся обнаружению.
"Эффективное количество", применяемое в отношении антитела или его антигенсвязывающих фрагментов по настоящему изобретению, относится к количеству, которое при введении в необходимых дозах и в течение необходимых периодов времени является достаточным для достижения намеченного биологического эффекта или желаемого терапевтического результата, в том числе без ограничения клинических результатов. Фраза "терапевтически эффективное количество", применяемая в отношении антитела или его антигенсвязывающих фрагментов по настоящему изобретению, подразумевается как означающая количество антитела или его антигенсвязывающих фрагментов, которое является достаточным для облегчения, смягчения, стабилизации, устранения, замедления, ослабления или задержки прогрессирования нарушения или болезненного состояния или симптома нарушения или заболевания. Согласно одному варианту осуществления способ по настоящему изобретению предусматривает введение антитела или его антигенсвязывающего фрагмента в комбинациях с другими соединениями. В таком случае "эффективное количество" представляет собой количество комбинации, достаточное для того, чтобы вызывать намеченный биологический эффект.
Терапевтически эффективное количество антитела к альфа-синуклеину или его антигенсвязывающего фрагмента по настоящему изобретению может варьировать в зависимости от таких факторов, как болезненное состояние, возраст, пол и вес индивидуума, а также способность антитела к альфа-синуклеину вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество также является количеством, при котором любые токсические или пагубные эффекты антитела или части антитела перевешиваются терапевтически благоприятными эффектами.
Как указано выше, настоящее изобретение, в частности, относится к моноклональному антителу, способному специфически связываться с эпитопом в пределах аминокислот 112-117 альфа-синуклеина человека (SEQ ID NO: 9 (ILEDMP)). Согласно одному варианту осуществления антитело может конкурировать с антителом GM37 за связывание с эпитопом в пределах аминокислот 112-117 альфа-синуклеина.
Антитела по настоящему изобретению, проиллюстрированные с помощью GM37, его вариантов var 1-3 GM37 и GM285, и их связывающие альфа-синуклеин фрагменты могут связывать фрагмент токсичного альфа-синуклеина, состоящий из остатков 1-119/122 альфа-синуклеина, и нейтрализовать его токсичность (например, путем внеклеточного связывания с фрагментом альфа-синуклеина и за счет этого предотвращая его поглощение клетками. К удивлению, антитела по настоящему изобретению, которые могут связываться с эпитопом в пределах аминокислот 112-117 альфа-синуклеина, обладают превосходящими свойствами по сравнению с антителами из предшествующего уровня техники, такими как антитело 9Е4, в связывании с токсичными разновидностями альфа-синуклеина в головном мозге человека и оказывают превосходные эффекты на устранение внеклеточного альфа-синуклеина и нормализацию нарушенной синаптической передачи, индуцированной альфа-синуклеином in vivo. Антитела по настоящему изобретению также могут облегчать проявление соответствующего двигательного фенотипа в крысиной модели болезни Паркинсона.
Антитела по настоящему изобретению предпочтительно представляют собой человеческие или гуманизированные антитела.
Настоящее изобретение также предусматривает способ уменьшения образования агрегатов альфа-синуклеина у пациента, предусматривающий введение пациенту, нуждающемуся в таком лечении, терапевтически эффективного количества антитела по настоящему изобретению.
Дополнительно антитела могут быть представлены в композиции вместе с фармацевтически приемлемым носителем, разбавителем и/или стабилизатором. Антитела по настоящему изобретению можно применять в терапии. В частности, антитела по настоящему изобретению можно применять при лечении синуклеопатий, таких как болезнь Паркинсона (в том числе идиопатическая наследственная формы болезни Паркинсона), болезнь Гоше, болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBV), комбинированная болезнь Альцгеймера и Паркинсона, истинная вегетативная недостаточность и множественная системная атрофия.
Лечение, предусматриваемое настоящим изобретением, может быть длительным, и пациент может получать лечение по меньшей мере в течение 2 недель, например, по меньшей мере в течение 1 месяца, 6 месяцев, 1 года или дольше.
Антитела антигенсвязывающего фрагмента по настоящему изобретению можно получить в различных линиях клеток, таких как линия клеток человека, линия клеток млекопитающего, отличного от человека, и линия клеток насекомого, например линия клеток СНО, линия клеток НЕК, линия клеток BHK-21, линия клеток мыши (такая как линия клеток миеломы), линия клеток фибросаркомы, линия клеток PER.C6, линия клеток HKB-11, линия клеток CAP и линия клеток HuH-7 человека (Dumont et al, 2015, Crit Rev Biotechnol. Sep 18:1-13., содержание, которого включено в данный документ посредством ссылки).
Антитела по настоящему изобретению могут представлять собой, например, моноклональные антитела, получаемые с помощью гибридомного способа, впервые описанного в Kohler et al., Nature 256, 495 (1975), или могут быть получены способами с применением рекомбинантной ДНК. Моноклональные антитела также можно выделить из фаговых библиотек антител с помощью методик, описанных, например, в Clackson et al., Nature 352, 624-628 (1991) и Marks et al., J. Mol. Biol. 222, 581-597 (1991). Моноклональные антитела можно получить из любого подходящего источника. Так, например, моноклональные антитела можно получить из гибридом, полученных из В-лимфоцитов селезенки мыши, полученных от мышей, иммунизированных представляющим интерес антигеном, например, в виде клеток, экспрессирующих антиген на поверхности, или нуклеиновой кислоты, кодирующей представляющий интерес антиген. Моноклональные антитела также могут быть получены из гибридом, полученных из экспрессирующих антитела клеток от иммунизированных людей или отличных от человека млекопитающих, таких как крысы, кролики, собаки, овцы, козы, приматы и т.д.
Согласно одному варианту осуществления антитело по настоящему изобретению представляет собой антитело человека. Моноклональные антитела человека, направленные против альфа-синуклеина, можно получать с применением трансгенных или трансхромосомных мышей, несущих части иммунной системы человека вместо системы мыши. Такие трансгенные и трансхромосомные мыши включают мышей, называемых в данном документе соответственно мышами HuMAb и мышами KM.
Мышь HuMAb содержит минилокус гена иммуноглобулина человека, который кодирует неперестроенные последовательности вариабельных и константных доменов тяжелых цепей (μ и Y) и вариабельных и константных доменов легких (κ) цепей иммуноглобулина человека вместе с целевыми мутациями, которые инактивируют эндогенные локусы μ- и κ-цепи (Lonberg, N. et al., Nature 368, 856-859 (1994)). Соответственно, мыши характеризуются сниженной экспрессией IgM или IgK мыши, и в ответ на иммунизацию введенные трансгены человеческой тяжелой и легкой цепи подвергаются переключению класса и соматической мутации с образованием высокоаффинных моноклональных антител IgG человека κ-изотипа (Lonberg, N. et al. (1994), выше; обзор которых приведен в Lonberg, N., Handbook of Experimental Pharmacology 113, 49-101 (1994), Lonberg, N. and Huszar, D., Intern. Rev. Immunol. Vol. 13 65-93 (1995) и Harding, F. and Lonberg, N., Ann. N. Y. Acad. Sci 764 536-546 (1995)). Получение мышей HuMAb подробно описано в Taylor, L. et al., Nucleic Acids Research 20, 6287-6295 (1992), Chen, J. et al., International Immunology 5, 647-656 (1993), Tuaillon et al., J. Immunol. 152, 2912-2920 (1994), Taylor, L. et al., International Immunology 6, 579-591 (1994), Fishwild, D. et al., Nature Biotechnology 14, 845-851 (1996). См. также US 5545806, US 5569825, US 5625126, US 5633425, US 5789650, US 5877397, US 5661016, US 5814318, US 5874299, US 5770429, US 5545807, WO 98/24884, WO 94/25585, WO 93/1227, WO 92/22645, WO 92/03918 и WO 01/09187.
Мыши НСо7, НСо12, НСо17 и НСо20 имеют нарушение JKD в своих эндогенных генах легкой цепи (каппа) (описанное в Chen et al., EMBO J. 12, 811-820 (1993)), нарушение CMD в своих эндогенных генах тяжелой цепи (описанное в примере 1 WO 01/14424) и трансген KCo5 человеческой легкой каппа-цепи (описанный в Fishwild et al., Nature Biotechnology 14, 845-851 (1996)). Кроме того, мыши НСо7 имеют трансген НСо7 человеческой тяжелой цепи (описанный в US 5770429), мыши НСо12 имеют трансген НСо12 человеческой тяжелой цепи (описанный в примере 2 WO 01/14424), мыши НСо17 имеют трансген НСо17 человеческой тяжелой цепи (описанный в примере 2 WO 01/09187), а мыши НСо20 имеют трансген НСо20 человеческой тяжелой цепи. Полученные в результате мыши экспрессируют трансгены тяжелой и легкой каппа-цепи иммуноглобулина человека в генетическом окружении, гомозиготном в отношении нарушения эндогенных локусов мышиной тяжелой и легкой каппа-цепи.
У мышей линии КМ эндогенный ген мышиной легкой каппа-цепи был подвергнут гомозиготному разрушению, как описано в Chen et al., EMBO J. 12, 811-820 (1993), а эндогенный ген мышиной тяжелой цепи был подвергнут гомозиготному разрушению, как описано в примере 1 WO 01/09187. Эта линия мышей несет трансген KCo5 человеческой легкой каппа-цепи, описанный в Fishwild et al., Nature Biotechnology 14, 845-851 (1996). Эта линия мышей также несет трансхромосому человеческой тяжелой цепи, состоящую из фрагмента hCF (SC20) хромосомы 14, описанного в WO 02/43478. Мышей НСо12-Balb/с, НСо17-Balb/c и НСо20-Balb/с можно получить путем скрещивания НСо12, НСо17 и НСо20 с KCo5[J/K](Balb), как описано в WO 09/097006.
У мышей линии KM эндогенный ген мышиной легкой каппа-цепи был подвергнут гомозиготному разрушению, как описано в Chen et al., EMBO J. 12, 811-820 (1993), а эндогенный ген тяжелой цепи мыши был подвергнут гомозиготному разрушению, как описано в примере 1 WO 01/09187. Эта линия мышей несет трансген KCo5 человеческой легкой каппа-цепи, описанный в Fishwild et al., Nature Biotechnology 14, 845-851 (1996). Эта линия мышей также несет трансхромосому человеческой тяжелой цепи, состоящую из антигенсвязывающего фрагмента hCF (SC20) хромосомы 14, описанного в WO 02/43478.
Спленоциты от этих трансгенных мышей можно применять для получения гибридом, которые секретируют моноклональные антитела человека, в соответствии с хорошо известными методиками. Моноклональные или поликлональные антитела человека по настоящему изобретению, или антитела по настоящему изобретению, происходящие от других видов, также можно получить трансгенным путем посредством получения другого млекопитающего, отличного от человека, или растения, являющегося трансгенным по представляющим интерес последовательностям тяжелых и легких цепей иммуноглобулина и получения из него антитела в извлекаемой форме. Что касается получения антител у млекопитающих трансгенным путем, антитела могут быть получены в молоке коз, коров или других млекопитающих и извлечены из него. См., например, US 5827690, US 5756687, US 5750172 и US 5741957.
Антитело по настоящему изобретению может относиться к любому изотипу. Выбор изотипа обычно будет определяться желаемыми эффекторными функциями, такими как индукция ADCC. Иллюстративными изотипами являются IgG1, IgG2, IgG3 и IgG4. Можно использовать константные домены любой из человеческих легких цепей каппа- или лямбда-типа. При необходимости, класс антитела к альфа-синуклеину по настоящему изобретению можно переключить с помощью известных способов. Например, антитело по настоящему изобретению, которое изначально представляло собой IgM, можно подвергнуть переключению класса на антитело IgG по настоящему изобретению. Дополнительно, методики переключения класса можно применять для превращения одного подкласса IgG в другой, например из IgG1 в IgG2. Таким образом, эффекторная функция антител по настоящему изобретению может быть изменена путем переключения изотипа, например, на антитело IgG1, IgG2, IgG3 или IgG4 для различных терапевтических путей применения. Согласно одному варианту осуществления антитело по настоящему изобретению представляет собой антитело IgG1, например IgG1, κ. Считают, что антитело относится к конкретному изотипу, если его аминокислотная последовательность наиболее гомологична такому изотипу по сравнению с другими изотипами.
Согласно одному варианту осуществления антитело по настоящему изобретению представляет собой полноразмерное антитело, предпочтительно антитело IgG, в частности, антитело IgG1, κ. Согласно другому варианту осуществления антитело по настоящему изобретению представляет собой фрагмент антитела или одноцепочеченое антитело.
Антитела и их антигенсвязывающие фрагменты могут быть, например, получены путем фрагментации на антигенсвязывающие фрагменты с применением традиционных методик, а антигенсвязывающие фрагменты подвергнуты скринингу в отношении полезности таким же образом, как описано в данном документе для полных антител. Например, антигенсвязывающие F(ab')2-фрагменты могут быть получены путем обработки антитела пепсином. Полученный в результате антигенсвязывающий F(ab')2-фрагмент можно обработать для восстановления дисульфидных мостиков с получением антигенсвязывающих фрагментов Fab'. Антигенсвязывающие Fab-фрагменты можно получить путем обработки антитела IgG папаином; антигенсвязывающие Fab'-фрагменты можно получить с помощью расщепления пепсином антитела IgG. Антигенсвязывающий F(ab')-фрагмент также может быть получен путем связывания описанного ниже Fab' с помощью тиоэфирной связи или дисульфидной связи. Антигенсвязывающий Fab'-фрагмент представляет собой антигенсвязывающий фрагмент антитела, полученный путем разрезания дисульфидной связи в шарнирном домене F(ab')2. Антигенсвязывающий Fab'-фрагмент может быть получен путем обработки антигенсвязывающего F(ab')2-фрагмента восстановителем, таким как дитиотреитол. Антигенсвязывающий фрагмент антитела также может быть получен путем экспрессии нуклеиновых кислот, кодирующих такие антигенсвязывающие фрагменты, в рекомбинантных клетках (см., например, Evans et al., J. Immunol. Meth. 184, 123-38 (1995)). Например, химерный ген, кодирующий часть антигенсвязывающего F(ab')2-фрагмента, может включать последовательности ДНК, кодирующие область СН1 и шарнирный домен цепи Н, за которыми расположен трансляционный стоп-кодон с получением такой усеченной молекулы антигенсвязывающего фрагмента антитела.
Согласно одному варианту осуществления антитело к альфа-синуклеину представляет собой одновалентное антитело, предпочтительно одновалентное антитело, описанное в WO 2007059782 (которое включено в данный документ посредством ссылки во всей своей полноте), имеющее делецию в шарнирном домене. Соответственно, согласно одному варианту осуществления антитело представляет собой одновалентное антитело, где указанное антитело к альфа-синуклеину сконструировано с помощью способа, предусматривающего i) обеспечение конструкции нуклеиновой кислоты, кодирующей легкую цепь указанного одновалентного антитела, при этом указанная конструкция содержит нуклеотидную последовательность, кодирующую VL-область выбранного антиген-специфического антитела к альфа-синуклеину, и нуклеотидную последовательность, кодирующую константную CL-область Ig, где указанная нуклеотидная последовательность, кодирующая VL-область выбранного антиген-специфического антитела, и указанная нуклеотидная последовательность, кодирующая CL-область Ig, функционально связаны друг с другом, и где в случае подтипа IgG1 нуклеотидная последовательность, кодирующая CL-область, была модифицирована так, чтобы CL-область не содержала каких-либо аминокислот, способных образовывать дисульфидные связи или ковалентные связи с другими пептидами, содержащими идентичную аминокислотную последовательность CL-области, в присутствии поликлонального IgG человека или при введении животному или человеку; ii) обеспечение конструкции нуклеиновой кислоты, кодирующей тяжелую цепь указанного одновалентного антитела, причем указанная конструкция содержит нуклеотидную последовательность, кодирующую VH-область выбранного антиген-специфического антитела, и нуклеотидную последовательность, кодирующую константную СН-область Ig человека, где нуклеотидная последовательность, кодирующая СН-область, была модифицирована так, чтобы область, соответствующая шарнирному домену, и, как того требует подтип Ig, другие области СН-области, такие как СН3-область, не содержали каких-либо аминокислотных остатков, которые участвуют в образовании дисульфидных связей или ковалентных или стабильных нековалентных связей между тяжелой цепью и другими пептидами, содержащими идентичную аминокислотную последовательность СН-области Ig человека, в присутствии поликлонального IgG человека или при введении животному человеку, где указанная нуклеотидная последовательность, кодирующая VH-область выбранного антиген-специфического антитела, и указанная нуклеотидная последовательность, кодирующая СН-область указанного Ig, функционально связаны друг с другом; iii) обеспечение клеточной системы экспрессии для получения указанного одновалентного антитела; iv) получение указанного одновалентного антитела путем совместной экспрессии конструкций нуклеиновых кислот из (i) и (ii) в клетках клеточной системы экспрессии из (iii).
Аналогично, согласно одному варианту осуществления антитело к альфа-синуклеину представляет собой одновалентное антитело, которое содержит
(i) вариабельный домен антитела по настоящему изобретению, описанный в данном документе, или антигенсвязывающую часть указанной области, и
(ii) СН-домен иммуноглобулина или его домен, содержащий домены СН2 и СН3, где СН-домен или его домен был модифицирован таким образом, чтобы домен, соответствующий шарнирному домену, и, если иммуноглобулин не относится к подтипу IgG4, другие домены СН-домена, такие как домен СН3, не содержали каких-либо аминокислотных остатков, которые могут образовывать дисульфидные связи с идентичным доменом СН, или другие ковалентные или стабильные нековалентные связи между тяжелой цепью и идентичным СН-доменом в присутствии поликлонального IgG человека.
Согласно дополнительному варианту осуществления тяжелая цепь одновалентного антитела к альфа-синуклеину была модифицирована таким образом, чтобы весь шарнирный домен была подвергнут делеции.
Согласно еще одному дополнительному варианту осуществления последовательность указанного одновалентного антитела была модифицирована таким образом, чтобы она не содержала каких-либо акцепторных сайтов N-связанного гликозилирования.
Настоящее изобретение также включает "биспецифические антитела", где область связывания с альфа-синуклеином (например, область связывания с альфа-синуклеином моноклонального антитела к альфа-синуклеину) является частью двухвалентного или поливалентного биспецифического остова, который нацеливается на более чем один эпитоп (например, второй эпитоп может предусматривать эпитоп рецептора, участвующего в активном транспорте, так чтобы биспецифическое антитело могло проявлять улучшенный трансцитоз через биологический барьер, такой как гематоэнцефалический барьер). Таким образом, согласно еще одному дополнительному варианту осуществления одновалентный Fab антитела к синуклеину может быть соединен с дополнительным Fab или scfv, которые нацеливаются на другой белок, с получением биспецифического антитела. Биспецифическое антитело может иметь двойную функцию, например терапевтическую функцию, придаваемую связывающимся с синуклеином доменом, и транспортную функцию, которая может обеспечивать связывание с молекулой рецептора для усиления переноса через биологический барьер, такой как гематоэнцефалический барьер.
Антитела к альфа-синуклеину и их антигенсвязывающие фрагменты по настоящему изобретению также включают одноцепочечные антитела. Одноцепочечные антитела представляют собой пептиды, в которых соединены Fv-области тяжелой и легкой цепи. Согласно одному варианту осуществления настоящее изобретение предусматривает одноцепочечный Fv (scFv), где тяжелые и легкие цепи в Fv антитела к альфа-синуклеину по настоящему изобретению соединены с помощью гибкого пептидного линкера (обычно из приблизительно 10, 12, 15 или более аминокислотных остатков) в одну пептидную цепь. Способы получения таких антител описаны, например, в US 4946778, Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994), Bird et al., Science 242, 423-426 (1988), Huston et al., PNAS USA 85, 5879-5883 (1988) и McCafferty et al., Nature 348, 552-554 (1990). Одноцепочечное антитело может быть одновалентным, если используются только один VH и VL, двухвалентным, если используются два VH и VL, или поливалентным, если используется более двух VH и VL.
Описываемые в данном документе антитела к альфа-синуклеину и их антигенсвязывающие фрагменты могут быть модифицированными путем включения любого подходящего количества модифицированных аминокислот и/или ассоциаций с такими конъюгированными заместителями. Пригодность в этом контексте обычно определяется способностью по меньшей мере в значительной степени сохранять селективность в отношении альфа-синуклеина и/или специфичность в отношении альфа-синуклеина, ассоциируемую с недериватизированным исходным антителом к альфа-синуклеину. Включение одной или нескольких модифицированных аминокислот может быть преимущественным, например, для увеличения периода полужизни полипептида в сыворотке, уменьшения антигенности полипептида или повышения стабильности полипептида при хранении. Аминокислота(аминокислоты) поддаются модификации, например, котрансляционно или посттрансляционно в ходе получения рекомбинантным путем (например, N-связанное гликозилирование в мотивах N-X-S/T в ходе экспрессии в клетках млекопитающих) или поддаются модификации синтетическими средствами. Неограничивающие примеры модифицированной аминокислоты включают гликозилированная аминокислота, сульфатированная аминокислота, пренилированная (например, фарнезилированная, геранилгеранилированная) аминокислота, ацетилированная аминокислота, ацилированная аминокислота, пегилированная аминокислота, биотинилированная аминокислота, карбоксилированная аминокислота, фосфорилированная аминокислота и т.д. В литературе представлено множество источников, подходящих в качестве руководства для специалиста по модификации аминокислот. Примеры протоколов находятся в Walker (1998) Protein Protocols On CD-Rom, Humana Press, Totowa, NJ. Модифицированную аминокислоту можно выбрать, например, из гликозилированной аминокислоты, пегилированной аминокислоты, фарнезилированной аминокислоты, ацетилированной аминокислоты, биотинилированной аминокислоты, аминокислоты, конъюгированной с липидным фрагментом, или аминокислоты, конъюгированной с органическим дериватизирующим средством.
Антитела к альфа-синуклеину также можно модифицировать химическим путем посредством ковалентного конъюгирования с полимером, например, для увеличения периода полужизни в кровотоке. Иллюстративные полимеры и способы присоединения их к пептидам приведены, например, в US 4766106, US 4179337, US 4495285 и US 4609546. Дополнительные иллюстративные полимеры включают полиоксиэтилированные полиолы и полиэтиленгликоль (PEG) (например, PEG с молекулярной массой от приблизительно 1000 до приблизительно 40000, например, от приблизительно 2000 до приблизительно 20000, например, приблизительно 3000-12000 г/моль).
Антитела по настоящему изобретению можно дополнительно применять в диагностическом способе или в качестве диагностического визуализирующего лиганда.
Согласно одному варианту осуществления предусматриваются антитела к альфа-синуклеину, содержащие одну или несколько меченных радиоактивным изотопом аминокислот. Меченное радиоактивным изотопом антитело к альфа-синуклеину можно применять как в диагностических, так и в терапевтических целях (еще одним возможным признаком является конъюгирование с мечеными радиоактивным изотопом молекулами). Неограничивающие примеры таких меток включают без ограничения висмут (213Bi), углерод (11С, 13С, 14С), хром (51Cr), кобальт (57Со, 60Со), медь (64Cu), диспрозий (165Dy), эрбий (169Er), фтор (18F), гадолиний (153Gd, 159Gd), галлий (68Ga, 67Ga), германий (68Ge), золото (198Au), гольмий (166Но), водород (3Н), индий (111In, 112In, 113In, 115In), йод (121I, 123I, 125I, 131I), иридий (192Ir), железо (59Fe), криптон (81mKr), лантан (140La), лютеций (177Lu), магний (54Mn), молибден (99Мо), азот (13N, 15N), кислород (15О), палладий (103Pd), фосфор (32Р), калий (42K), празеодим (142Pr), прометий (149Pm), рений (186Re, 188Re), родий (105Rh), рубидий (81Rb, 82Rb), рутений (82Ru, 97Ru), самарий (153Sm), скандий (47Sc), селен (75Se), натрий (24Na), стронций (85Sr, 89Sr, 92Sr), серу (35S), технеций (99Tc), таллий (201Tl), олово (113Sn, 117Sn), ксенон (133Хе), иттербий (169Yb, 175Yb, 177Yb), иттрий (90Y) и цинк (65Zn). Способы получения аминокислот, меченных радиоактивными изотопами, и соответствующих производных пептидов известны из уровня техники (см., например, Junghans et al., в Cancer Chemotherapy and Biotherapy 655-686 (2-е издание, под редакцией Chafner и Longo, Lippincott Raven (1996)) и US 4681581; US 4735210; US 5101827; US 5102990 (US RE35500), US 5648471 и US 5697902. Например, радиоактивный изотоп можно конъюгировать с помощью способа с применением хлорамина Т (Lindegren, S. et al. (1998) "Chloramine-T In High-Specific-Activity Radioiodination Of Antibodies Using N-Succinimidyl-3-(Trimethylstannyl)Benzoate As An Intermediate," Nucl. Med. Biol. 25(7):659-665; Kurth, M. et al. (1993) "Site-Specific Conjugation Of A Radioiodinated Phenethylamine Derivative To A Monoclonal Antibody Results In Increased Radioactivity Localization In Tumor," J. Med. Chem. 36(9):1255-1261; Rea, D.W. et al. (1990) "Site-specifically radioiodinated antibody for targeting tumors," Cancer Res. 50(3 Suppl):857s-861s).
Настоящее изобретение также предусматривает антитела к альфа-синуклеину и их антигенсвязывающие фрагменты, которые помечены с помощью выявляемой флуоресцентной метки (такой как хелат редкоземельного элемента (например, хелат европия)), метки флуоресцеинового типа (например, флуоресцеин, флуоресцеинизотиоцианат, 5-карбоксифлуоресцеин, 6-карбоксифлуоресцеин, дихлортиазиниламинфлуоресцеин), метки родаминового типа (например, ALEXA FLUOR® 568 (Invitrogen), TAMRA® или данзилхлорид), VIVOTAG 680 XL FLUOROCHROME™ (Perkin Elmer), фикоэритрина; умбеллиферона, лиссамина; цианина; фикоэритрина, техасского красного, BODIPY FL-SE® (Invitrogen) или его аналога, все из которых подходят для оптического выявления. Можно использовать хемилюминесцентные метки (например, люминол, люциферазу, люциферин и экворин). Такую диагностику и выявление также можно осуществить путем связывания диагностической молекулы по настоящему изобретению с выявляемыми веществами, в том числе без ограничения различные ферменты, при этом ферменты включают без ограничения пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу, или с комплексами с простетическими группами, такие как без ограничения стрептавидин/биотин и авидин/биотин.
Можно использовать хемилюминесцентные метки (например, люминол, люциферазу, люциферин и экворин). Такую диагностику и выявление также можно осуществить путем связывания диагностической молекулы по настоящему изобретению с выявляемыми веществами, в том числе без ограничения различные ферменты, при этом ферменты включают без ограничения пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу, или с комплексами с простетическими группами, такие как без ограничения стрептавидин/биотин и авидин/биотин. Также можно использовать парамагнитные метки, и их предпочтительно выявляют с помощью позитронно-эмиссионной томографии (PET) или однофотонной эмиссионной компьютерной томографии (SPECT). Такие парамагнитные метки включают без ограничения соединения, содержащие парамагнитные ионы алюминия (Al), бария (Ва), кальция (Са), церия (Се), диспрозия (Dy), эрбия (Er), европия (Eu), гадолиния (Gd), гольмия (Но), иридия (Ir), лития (Li), магния (Mg), марганца (Mn), молибдена (М), неодимия (Nd), осмия (Os), кислорода (О), палладия (Pd), платины (Pt), родия (Rh), рутения (Ru), самария (Sm), натрия (Na), стронция (Sr), тербия (Tb), тулия (Tm), олова (Sn), титана (Ti), вольфрама (W), и циркония (Zi), и, в частности, Со+2, CR+2, Cr+3, Cu+2, Fe+2, Fe+3. Ga+3, Mn+3, Ni+2, Ti+3, V+3 и V+4, позитронно-активные металлы с использованием различных видов позитронно-эмиссионной томографии и нерадиоактивных парамагнитных ионов металлов.
Таким образом, согласно одному варианту осуществления антитело к альфа-синуклеину по настоящему изобретению может быть помечено флуоресцентной меткой, хемилюминесцентной меткой, парамагнитной меткой, радиоизотопной меткой или ферментной меткой. Меченое антитело можно применять для выявления наличия или измерения количества указанного альфа-синуклеина в головном мозге субъекта. Этот способ может предусматривать обнаружение или измерение количества антитела к альфа-синуклеину, связанного с указанным альфа-синуклеином, посредством визуализации in vivo и может предусматривать визуализацию ех vivo указанного антитела к альфа-синуклеину, связанного с указанным альфа-синуклеином.
Согласно дополнительному аспекту настоящее изобретение относится к вектору экспрессии, кодирующему одну или несколько полипептидных цепей антитела по настоящему изобретению или его антигенсвязывающего фрагмента. Такие векторы экспрессии можно применять для получения антител и антигенсвязывающих фрагментов по настоящему изобретению рекомбинантным путем.
Вектор экспрессии в контексте настоящего изобретения может представлять собой любой подходящий ДНК- или РНК-вектор, в том числе хромосомные, нехромосомные и синтетические векторы нуклеиновых кислот (последовательность нуклеиновой кислоты, содержащая подходящий набор элементов контроля экспрессии). Примеры таких векторов включают производные SV40, бактериальные плазмиды, фаговую ДНК, бакуловирус, дрожжевые плазмиды, векторы, полученные в результате объединения плазмид и фаговой ДНК, и векторы на основе вирусной нуклеиновой кислоты (РНК или ДНК). Согласно одному варианту осуществления нуклеиновая кислота, кодирующая антитело к альфа-синуклеину, содержится в векторе на основе "голой" ДНК или РНК, включающем, например, линейный элемент экспрессии (описанный, например, в Sykes and Johnston, Nat Biotech 12, 355-59 (1997)), вектор на основе компактизированной нуклеиновой кислоты (описанный, например, в US 6077835 и/или WO 00/70087), плазмидный вектор, такой как pBR322, pUC 19/18 или pUC 118/119, вектор минимального размера MIDGE на основе нуклеиновой кислоты (описанный, например, в Schakowski et al., MoI Ther 3, 793-800 (2001)), или представлена в виде осажденной векторной конструкции нуклеиновой кислоты, такой как осажденная в CaPO4 конструкция (описанная, например, в WO 00/46147, Benvenisty and Reshef, PNAS USA 83, 9551-55 (1986), Wigler et al., Cell 14, 725 (1978), и Coraro and Pearson, Somatic Cell Genetics 2, 603 (1981)). Такие векторы на основе нуклеиновых кислот и их применение хорошо известны из уровня техники (см., например, US 5589466 и US 5973972).
Согласно одному варианту осуществления вектор подходит для экспрессии антител к альфа-синуклеину или их антигенсвязывающих фрагментов в бактериальной клетке. Примеры таких векторов включают векторы экспрессии, такие как BlueScript (Stratagene), векторы pIN (Van Heeke & Schuster, J Biol Chem 264, 5503-5509 (1989), векторы рЕТ (Novagen, Мэдисон, Висконсин) и т.п.).
Вектор экспрессии может также или в качестве альтернативы представлять собой вектор, подходящий для экспрессии в дрожжевой системе. Можно использовать любой вектор, подходящий для экспрессии в дрожжевой системе. Подходящие векторы включают, например, векторы, содержащие конститутивные или индуцируемые промоторы, такие как промотор гена фактора альфа, алкогольоксидазы и PGH (обзор которых приведен в F. Ausubel et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley InterScience New York (1987), Grant et al., Methods in Enzymol 153, 516-544 (1987), Mattanovich, D. et al. Methods Mol. Biol. 824, 329-358 (2012), Celik, E. et al. Biotechnol. Adv. 30(5), 1108-1118 (2012), Li, P. et al. Appl. Biochem. Biotechnol. 142(2), 105-124 (2007),  , E. et al. Appl. Microbiol. Biotechnol. 77(3), 513-523 (2007), van der Vaart, J.M. Methods Mol. Biol. 178, 359-366 (2002), и Holliger, P. Methods Mol. Biol. 178, 349-357 (2002)).
, E. et al. Appl. Microbiol. Biotechnol. 77(3), 513-523 (2007), van der Vaart, J.M. Methods Mol. Biol. 178, 359-366 (2002), и Holliger, P. Methods Mol. Biol. 178, 349-357 (2002)).
В векторе экспрессии по настоящему изобретению нуклеиновые кислоты, кодирующие антитело к альфа-синуклеину, могут содержать любой подходящий промотор, энхансер и другие элементы, способствующие экспрессии, или быть связанными с ними. Примеры таких элементов включают сильные промоторы экспрессии (например, IE промотор/энхансер CMV человека, а также LTR промоторы RSV, SV40, SL3-3, MMTV и HIV), эффективные поли-(А)-последовательности терминации, точку начала репликации для продукта плазмиды в E.coli, ген устойчивости к антибиотикам в качестве селективного маркера и/или подходящий сайт клонирования (например, полилинкер). Нуклеиновые кислоты также могут содержать индуцируемый промотор, а не конститутивный промотор, такой как IE CMV (специалист в данной области техники поймет, что такие термины фактически описывают степень экспрессии гена в определенных условиях).
Антитела антигенсвязывающего фрагмента по настоящему изобретению можно получить в различных линиях клеток, таких как линия клеток человека, линия клеток млекопитающего, отличного от человека, и линия клеток насекомого, например линия клеток СНО, линия клеток НЕК, линия клеток BHK-21, линия клеток мыши (такая как линия клеток миеломы), линия клеток фибросаркомы, линия клеток PER.C6, линия клеток HKB-11, линия клеток CAP и линия клеток HuH-7 человека (Dumont et al, 2015, Crit Rev Biotechnol. Sep 18:1-13., содержание, которого включено в данный документ посредством ссылки).
Согласно еще одному дополнительному аспекту настоящее изобретение относится к рекомбинантной эукариотической или прокариотической клетке-хозяину, такой как трансфектома, которая продуцирует антитело или его антигенсвязывающий домен по настоящему изобретению, определенные в данном документе, или биспецифическую молекулу по настоящему изобретению, определенную в данном документе. Примеры клеток-хозяев включают клетки дрожжей, бактерий и млекопитающих, такие как клетки СНО или HEK. Например, согласно одному варианту осуществления настоящее изобретение предусматривает клетку, содержащую нуклеиновую кислоту, стабильно интегрированную в клеточный геном, которая содержит последовательность, кодирующую продукт экспрессии в виде антитела к альфа-синуклеину по настоящему изобретению или его антигенсвязывающий фрагмент. Согласно другому варианту осуществления настоящее изобретение предусматривает клетку, содержащую неинтегрированную нуклеиновую кислоту, такую как плазмида, космида, фагмида или линейный элемент экспрессии, которая содержит последовательность, кодирующую продукт экспрессии в виде антитела к альфа-синуклеину по настоящему изобретению.
Согласно дополнительному аспекту настоящее изобретение относится к способу получения антитела к альфа-синуклеину по настоящему изобретению, причем указанный способ предусматривает стадии а) культивирования гибридомы или клетки-хозяина по настоящему изобретению, описанных в данном документе выше, и b) очистку антитела по настоящему изобретению от культуральной среды.
Согласно одному варианту осуществления настоящее изобретение относится к препарату, который, в том смысле, в котором данный термин используют в данном документе, содержит антитело к альфа-синуклеину, определенное в данном документе, и который практически не содержит антител, образующихся в естественных условиях, которые либо не способны связываться с альфа-синуклеином или которые существенным образом не изменяют функциональные характеристики антитела к альфа-синуклеину в препарате. Таким образом, такой препарат не охватывает сыворотку крови, образующуюся в естественных условиях, или очищенное производное такой сыворотки крови, которое содержит смесь антител к альфа-синуклеину и другого антитела, которое не изменяет функциональные характеристики антитела к альфа-синуклеину в препарате; где такие функциональные характеристики выбраны из группы, состоящей из
(i) аффинности связывания (KD) антитела к альфа-синуклеину в отношении альфа-синуклеина;
(ii) способности антитела к альфа-синуклеину подавлять усечение протеазами фибрилл альфа-синуклеина;
(iii) способности антитела к альфа-синуклеину обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
(iv) способности антитела к альфа-синуклеину снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo; и
(v) способности, при длительном введении, антитела к альфа-синуклеину восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;.
(vi) способности предотвращать затравочное действие альфа-синуклеина (такое как накопление нерастворимого фосфорилированного альфа-синуклеина in vitro и/или в мышиной модели болезни Паркинсона); и/или
(vii) способности связывать усеченный альфа-синуклеин в головном мозге человека.
Настоящее изобретение, в частности, относится к препаратам такого антитела к альфа-синуклеину, имеющего структурное изменение в своей аминокислотной последовательности (в любых своих CDR, вариабельных доменах, остатках каркасной области и/или константных доменах) по сравнению со структурой встречающегося в природе антитела к альфа-синуклеину, где указанное структурное изменение обуславливает значительное изменение проявляемых антителом к альфа-синуклеину функциональных характеристик (т.е. различие более чем на 20%, различие более чем на 40%, различие более чем на 60%, различие более чем на 80%, различие более чем на 100%, различие более чем на 150%, различие более чем в 2 раза, различие более чем в 4 раза, различие более чем в 5 раз или различие более чем в 10 раз в функциональных характеристиках) по сравнению с функциональными характеристиками, проявляемыми указанным встречающимся в природе антителом к альфа-синуклеину; где такие функциональные характеристики представляют собой:
(i) аффинность связывания (KD) моноклонального антитела к альфа-синуклеину в отношении альфа-синуклеина;
(ii) способность моноклонального антитела к альфа-синуклеину подавлять усечение протеазами фибрилл альфа-синуклеина;
(iii) способность моноклонального антитела к альфа-синуклеину обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
(iv) способность моноклонального антитела к альфа-синуклеину снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo; и/или
(v) способность, при длительном введении, моноклонального антитела к альфа-синуклеину восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
(vi) способность предотвращать затравочное действие в отношении агрегирования альфа-синуклеина (такую как накопление нерастворимого фосфорилированного альфа-синуклеина in vitro и/или в мышиной модели болезни Паркинсона); и/или
(vii) способность связывать усеченный альфа-синуклеин в головном мозге человека.
особенно в тех случаях, где такие измененные функциональные характеристики являются результатом структурного изменения и, следовательно, неотделимы от нее.
Термин "практически не содержащий" антител, образующихся в естественных условиях, относится к полному отсутствию таких антител, образующихся в естественных условиях, в таких препаратах или к включению таких антител, образующихся в естественных условиях, в такие препараты в концентрации, в которой они существенным образом не влияют на свойства связывания с альфа-синуклеином данных препаратов. Говорят, что антитело является "выделенным", если оно не имеет эквивалента, образующегося в естественных условиях, или было отделено или очищено от компонентов, которые в естественных условиях сопутствуют ему.
Термин "антитела, образующиеся в естественных условиях", используемый в отношении таких препаратов, относится к антителам (в том числе аутоантителам, образующимся в естественных условиях), образование которых вызывается в организме живых людей или других животных в качестве естественного следствия функционирования их иммунных систем.
Таким образом, препараты по настоящему изобретению не исключают, а в действительности явным образом охватывают препараты, которые содержат антитело к альфа-синуклеину и специально добавленное дополнительное антитело, которое может связываться с эпитопом, который отсутствует у альфа-синуклеина. Такие препараты, в частности, включают варианты их осуществления, где данный препарат характеризуются повышенной эффективностью при лечении синуклеопатий, таких как болезнь Паркинсона (в том числе идиопатическая и наследственная форма болезни Паркинсона), болезнь Гоше, болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBV), комбинированная болезнь Альцгеймера и Паркинсона, истинная вегетативная недостаточность и множественная системная атрофия.
В еще одном дополнительном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей
(i) антитело к альфа-синуклеину или его антигенсвязывающий фрагмент, оба из которых определены в данном документе, или препарат, в том смысле, в котором этот термин определен в данном документе, который содержит такое антитело к альфа-синуклеину или его антигенсвязывающий фрагмент, и
(ii) фармацевтически приемлемый носитель.
Фармацевтические композиции можно составлять с фармацевтически приемлемыми носителями или разбавителями, а также с любыми другими известными вспомогательными средствами и наполнителями согласно общепринятым методикам, таким как, например, методики, раскрытые в Remington: The Science and Practice of Pharmacy, 22nd Edition, Gennaro, Ed., Mack Publishing Co., Easton, PA, 2013.
Фармацевтически приемлемые носители или разбавители, а также любые другие известные вспомогательные средства и наполнители должны быть пригодными для выбранного соединения по настоящему изобретению и выбранного способа введения. Пригодность носителей и других компонентов фармацевтических композиций определяют на основании отсутствия значительного отрицательного влияния на желаемые биологические свойства выбранного соединения или фармацевтической композиции по настоящему изобретению (например, менее чем существенного влияния (10% или меньшего относительного ингибирования, 5% или меньшего относительного ингибирования и т.д.) на связывание с эпитопом.
Фармацевтическая композиция по настоящему изобретению также может содержать разбавители, заполнители, соли, буферы, детергенты (например, неионный детергент, такой как Tween-20 или Tween-80), стабилизаторы (например, сахара или белковые свободные аминокислоты), консерванты, фиксаторы тканей, солюбилизаторы и/или другие материалы, подходящие для включения в фармацевтическую композицию. Разбавитель выбирают так, чтобы он не влиял на биологическую активность комбинации. Примерами таких разбавителей являются дистиллированная вода, физиологический фосфатно-солевой буферный раствор, растворы Рингера, раствор декстрозы и раствор Хэнка. Кроме того, фармацевтическая композиция или состав также могут содержать другие носители или нетоксичные нетерапевтические неиммуногенные стабилизаторы и т.п. Композиции также могут содержать крупные, медленно метаболизируемые макромолекулы, такие как белки, полисахариды, такие как хитозан, полимолочные кислоты, полигликолевые кислоты и сополимеры (например, функционализированная латексом сефароза, агароза, целлюлоза и т.п.), полимеры аминокислот, сополимеры аминокислот и агрегаты липидов (например, масляные капли или липосомы).
Фактические уровни доз активных ингредиентов в фармацевтических композициях по настоящему изобретению можно изменять для того, чтобы получить количество активного ингредиента, которое является эффективным для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, в том числе активности конкретных используемых композиций по настоящему изобретению или их амида, пути введения, времени введения, скорости выведения конкретного используемого соединения, продолжительности лечения, других лекарственных средств, соединений и/или материалов, применяемых в комбинации с конкретными используемыми композициями, возраста, пола, веса, состояния, общего состояния здоровья и анамнеза подвергаемого лечению пациента, а также аналогичных факторов, хорошо известных в области медицины.
Фармацевтическую композицию можно применять любым подходящим путем и способом, в том числе парентеральным, местным, пероральным или интраназальным способом, для профилактического и/или терапевтического лечения. Согласно одному варианту осуществления фармацевтическую композицию по настоящему изобретению вводят парентерально. Используемые в данном документе фразы "парентеральное введение" и "вводимый парентерально" означают способы введения, отличные от энтерального и местного введения, обычно путем инъекции, и включают эпидермальную, внутривенную, внутримышечную, внутриартериальную, интратекальную, внутрикапсулярную, внутриглазничную, внутрисердечную, внутрикожную, внутрибрюшинную, внутрисухожильную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, подкапсулярную, субарахноидальную, интраспинальную, внутричерепную, интраторакальную, эпидуральную и внутригрудинную инъекцию и инфузию. Дополнительные подходящие пути введения соединения по настоящему изобретению in vivo и in vitro хорошо известны из уровня техники и могут быть выбраны специалистами в данной области техники. Согласно одному варианту осуществления эту фармацевтическую композицию вводят посредством внутривенной или подкожной инъекции или инфузии.
Фармацевтически приемлемые носители включают все без исключения подходящие растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, средства, придающие изотоничность, антиоксиданты и средства, замедляющие абсорбцию, и т.п., которые являются физиологически совместимыми с соединением по настоящему изобретению.
Примеры подходящих водных и неводных носителей, которые можно использовать в фармацевтических композициях по настоящему изобретению, включают воду, солевой раствор, фосфатно-солевой буферный раствор, этанол, декстрозу, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.), а также их пригодные смеси, растительные масла, такие как оливковое масло, кукурузное масло, арахисовое масло, хлопковое масло и кунжутное масло, коллоидные растворы карбоксиметилцеллюлозы, трагакантовую камедь и пригодные для инъекций сложные органические эфиры, такие как этилолеат, и/или различные буферы. В области фармацевтики хорошо известны другие носители.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для индивидуального получения стерильных инъекционных растворов или дисперсий. Применение таких сред и средств для фармацевтически активных веществ известно из уровня техники. За исключением случаев, когда какие-либо традиционные среда или средство являются несовместимыми с активным соединением, предполагается их применение в фармацевтических композициях по настоящему изобретению.
Надлежащую текучесть можно поддерживать, например, путем применения материалов для покрытия, таких как лецитин, путем поддержания требуемого размера частиц в случае дисперсий и путем применения поверхностно-активных веществ.
Фармацевтические композиции по настоящему изобретению также могут содержать фармацевтически приемлемые антиоксиданты, например, (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; (2) жирорастворимые антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (ВНА), бутилированный гидрокситолуол (ВНТ), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) металлохелаторы, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и т.п.
Фармацевтические композиции по настоящему изобретению также могут содержать средства, обеспечивающие изотоничность, такие как сахара, многоатомные спирты, такие как маннит, сорбит, глицерин, или хлорид натрия, в композициях.
Фармацевтические композиции по настоящему изобретению также могут содержать одно или несколько вспомогательных веществ, подходящих для выбранного пути введения, такие как консерванты, смачивающие вещества, эмульгаторы, диспергирующие вещества, консерванты или буферы, которые могут увеличить срок хранения или эффективность фармацевтической композиции. Соединения по настоящему изобретению можно получать с носителями, которые будут защищать соединение от быстрого высвобождения, например, в виде состава с контролируемым высвобождением, в том числе имплантатов, трансдермальных пластырей и микроинкапсулированных систем доставки. Такие носители могут включать желатин, глицерилмоностеарат, глицерилдистеарат, биоразлагаемые, биосовместимые полимеры, такие как сополимер этилена и винилацетата, полиангидриды, полигликолевую кислоту, коллаген, сложные полиортоэфиры и полимолочную кислоту отдельно или вместе с воском, или другие материалы, хорошо известные из уровня техники. Способы получения таких составов в целом известны специалистам в данной области техники. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Согласно одному варианту осуществления соединения по настоящему изобретению можно составить для обеспечения надлежащего распределения in vivo. Фармацевтически приемлемые носители для парентерального введения включают стерильные водные растворы или дисперсии и стерильные порошки для индивидуального получения стерильных инъекционных растворов или дисперсий. Применение таких сред и средств для фармацевтически активных веществ известно из уровня техники. За исключением случаев, когда какие-либо традиционные среда или средство являются несовместимыми с активным соединением, предполагается их применение в фармацевтических композициях по настоящему изобретению. В композиции также могут быть включены дополнительные активные соединения.
Фармацевтические композиции для инъекций как правило должны быть стерильными и стабильными в условиях изготовления и хранения. Композиция может быть составлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может представлять собой водный или неводный растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и подходящие их смеси, растительные масла, такие как оливковое масло, и пригодные для инъекций сложные органические эфиры, такие как этилолеат. Надлежащую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ, о многих случаях будет предпочтительным включение средств, обеспечивающих изотоничность, например сахара, многоатомные спирты, такие как глицерин, маннит, сорбит или хлорид натрия, в композицию. Длительного всасывания инъекционных композиций можно достичь путем включения в композицию средства, замедляющего всасывание антитела, например моностеаратных солей и желатина. Стерильные инъекционные растворы можно получить путем помещения активного соединения в требуемом количестве в соответствующий растворитель с одним ингредиентом или комбинацией ингредиентов, например, перечисленных выше, при необходимости с последующей стерилизующей микрофильтрацией. Обычно дисперсии получают путем помещения активного соединения в стерильную основу, которая содержит основную дисперсионную среду и другие необходимые ингредиенты, например, из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов примерами способов получения являются вакуумная сушка и сублимационная сушка (лиофилизация), в результате которой получают порошкообразный активный ингредиент плюс любой дополнительный желаемый ингредиент из их раствора, предварительно подвергнутого стерилизующей фильтрации.
Стерильные инъекционные растворы можно получить путем помещения активного соединения в требуемом количестве в соответствующий растворитель с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости с последующей стерилизующей микрофильтрацией. Обычно дисперсии получают путем помещения активного соединения в стерильный основу, которая содержит основную дисперсионную среду и другие требуемые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов примерами способов получения являются вакуумная сушка и сублимационная сушка (лиофилизация), в результате которой получают порошкообразный активный ингредиент плюс любой дополнительный желаемый ингредиент из их раствора, предварительно подвергнутого стерилизующей фильтрации.
Схемы приема в вышеуказанных способах лечения и путях применения, описанных в данном документе, корректируют для получения оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить однократную болюсную дозу, можно вводить несколько разделенных доз в течение некоторого времени, или дозу можно пропорционально уменьшать или увеличивать, как определяется потребностями терапевтической ситуации. Композиции для парентерального введения можно составлять в виде единичной лекарственной формы для простоты введения и равномерности дозирования. Единичная лекарственная форма, используемая в данном документе, относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; при этом каждая единица содержит предварительно определенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, совместно с требуемым фармацевтическим носителем. Технические требования к единичным лекарственным формам по настоящему изобретению продиктованы (а) уникальными характеристиками активного соединения и конкретным терапевтическим эффектом, которого необходимо достичь, и (b) ограничениями, присущими области составления такого активного соединения для лечения чувствительности у индивидуумов, и непосредственно зависят от них.
Эффективные дозы и схемы приема для антител к альфа-синуклеину зависят от подлежащего лечению заболевания или состояния и могут быть определены специалистами в данной области техники. В любой заданный день, когда дают дозу, доза может находиться в диапазоне от приблизительно 0,0001 до приблизительно 100 мг/кг и чаще от приблизительно 0,01 до приблизительно 5 мг/кг веса тела реципиента. Например, дозы могут составлять 1 мг/кг веса тела или 10 мг/кг веса тела или находиться в диапазоне 1-10 мг/кг веса тела. Таким образом, иллюстративные дозы включают от приблизительно 0,1 до приблизительно 10 мг/кг/веса тела, от приблизительно 0,1 до приблизительно 5 мг/кг/веса тела, от приблизительно 0,1 до приблизительно 2 мг/кг/веса тела, от приблизительно 0,1 до приблизительно 1 мг/кг/веса тела, например, приблизительно 0,15 мг/кг/веса тела, приблизительно 0,2 мг/кг/веса тела, приблизительно 0,5 мг/кг/веса тела, приблизительно 1 мг/кг/веса тела, приблизительно 1,5 мг/кг/веса тела, приблизительно 2 мг/кг/веса тела, приблизительно 5 мг/кг/веса тела или приблизительно 10 мг/кг/веса тела.
Врач со стандартной квалификацией в данной области легко может определить и назначить эффективное количество требуемой фармацевтической композиции. Например, врач может начать с доз антитела к альфа-синуклеину, используемого в фармацевтической композиции, на более низких уровнях, чем требуется для достижения желаемого терапевтического эффекта, и постепенно повышать дозу до достижения желаемого эффекта. В целом, подходящей суточной дозой композиции по настоящему изобретению будет такое количество соединения, которое является наиболее низкой дозой, эффективной для получения терапевтического эффекта. Такая эффективная доза обычно будет зависеть от описанных выше факторов. Введение может быть, например, внутривенным, внутримышечным, внутрибрюшинным или подкожным. При желании эффективную суточную дозу фармацевтической композиции можно вводить в виде двух, трех, четырех, пяти, шести или более частей дозы, вводимых по отдельности с соответствующими интервалами в течение дня, необязательно в виде единичных лекарственных форм. Хотя соединение по настоящему изобретению можно вводить отдельно, предпочтительно вводить соединение в виде фармацевтической композиции, описанной выше.
Меченые антитела по настоящему изобретению можно применять в диагностических целях для выявления, диагностики или мониторинга заболеваний или нарушени. Настоящее изобретение предусматривает выявление или диагностику нейродегенеративного или когнитивного заболевания или нарушения, в том числе без ограничения болезни Паркинсона, идиопатической болезни Паркинсона, болезни Паркинсона семейного происхождения, болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBV), комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии, которые предусматривают (а) проведение анализа наличия разновидностей альфа-синуклеина и фрагментов в клетках или образцах ткани субъекта с применением одного или нескольких антител, которые специфически связываются с альфа-синуклеином; и (b) проведение сравнения уровня антигена с контрольным уровнем, например, уровнями в образцах нормальной ткани, при этом повышение анализируемого уровня антигена по сравнению с контрольным уровнем антигена свидетельствует о заболевании или нарушении или свидетельствует о тяжести заболевания или нарушения.
Антитела по настоящему изобретению можно применять для анализа альфа-синуклеинового мономера, олигомеров, фибриллярных форм или фрагментов альфа-синуклеина в биологическом образце с помощью иммуногистохимических способов, хорошо известных из уровня техники. Другие способы с использованием антител, применимые для выявления белка, включают иммунологические анализы, такие как твердофазный иммуноферментный анализ (ELISA), радиоиммунологический анализ (RIA) и анализы с использованием платформы для мезомасштабных исследований (MSD). В таких наборах и способах можно применять подходящие метки для антител, и метки, известные из уровня техники, включают ферментные метки, такие как щелочная фосфатаза и глюкозооксидаза; радиоизотопные метки, такие как йод (125I, 131I), углерод (14С), сера (35S), тритий (3H), индий (121In) и технеций (99mTc); а также люминесцентные метки, такие как люминол и люцифераза; и флуоресцентные метки, такие как флуоресцеин и родамин.
Наличие меченых антител к альфа-синуклеину или их связывающих альфа-синуклеин фрагментов, можно выявлять in vivo в диагностических целях. Согласно одному варианту осуществления диагностика предусматривает а) введение субъекту эффективного количества такой меченой молекулы; b) ожидание в течение некоторого промежутка времени после введения для обеспечения концентрирования меченой молекулы в местах отложения Аβ (при наличии таковых) и для обеспечения очищения от несвязанной меченой молекулы до фонового уровня; с) определение фонового уровня; и d) выявление меченой молекулы у субъекта, таким образом, что выявление меченой молекулы на уровне, превышающем фоновый, свидетельствует о том, что субъект имеет заболевание или нарушение, или свидетельствует о тяжести заболевания или нарушения. В соответствии с таким вариантом осуществления молекулу метят визуализирующим фрагментом, подходящим для выявления с помощью конкретной системы визуализации, известной специалистам в данной области техники. Фоновые уровни можно определить с помощью различных способов, известных из уровня техники, в том числе путем сравнения количества выявленного меченого антитела со стандартным значением, предварительно определенным для конкретной системы визуализации. Способы и системы, которые можно применять в диагностических способах по настоящему изобретению, включают без ограничения компьютерную томографию (СТ), сканирование всего тела, такое как позитронно-эмиссионная томография (PET), магнитно-резонансная визуализация (MRI) и ультразвуковое исследование.
Согласно дополнительному аспекту настоящее изобретение относится к антителу или его антигенсвязывающим фрагментам по настоящему изобретению для применения в медицине.
Согласно дополнительному аспекту настоящее изобретение относится к антителу или его антигенсвязывающим фрагментам по настоящему изобретению для применения при лечении, диагностике или визуализации синуклеопатии.
Согласно одному варианту осуществления моноклональное антитело или его антигенсвязывающий фрагмент предназначены для применения при лечении болезни Паркинсона, идиопатической болезни Паркинсона, семейных форм болезни Паркинсона, болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBV), комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
Согласно дополнительному аспекту настоящее изобретение относится к применению антитела или его антигенсвязывающего фрагмента по настоящему изобретению при изготовлении лекарственного препарат для лечения, диагностики или визуализации синуклеопатии.
Согласно дополнительному аспекту настоящее изобретение относится к лечению, диагностике или визуализации болезни Паркинсона или других синуклеопатии, предусматривающему введение эффективной дозы антитела по настоящему изобретению или его антигенсвязывающего фрагмента.
Предпочтительно, в путях применения и способах по данным аспектам настоящего изобретения лечение является длительным и предпочтительно осуществляется по меньшей мере в течение 2 недель, например, по меньшей мере в течение 1 месяца, 6 месяцев, 1 года или дольше.
Согласно дополнительному аспекту настоящее изобретение предусматривает набор, содержащий антитело или его антигенсвязывающий фрагмент по настоящему изобретению.
ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Как будет очевидно из текста и примеров, настоящее изобретение дополнительно относится к приведенным ниже вариантам осуществления.
1. Моноклональное антитело или его антигенсвязывающий фрагмент которые способны специфически связываться с эпитопом в пределах аминокислот 112-117 в альфа-синуклеине (SEQ ID NO: 9 (ILEDMP)).
2. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 1, которые конкурируют с антителом GM37 за связывание с указанным эпитопом.
3. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 1, которые представляет собой GM37, вариант 1 GM37, вариант 2 GM37 или вариант 3 GM37.
4. Моноклональное антитело или его антигенсвязывающий фрагмент, которые способны специфически связываться с эпитопом в пределах аминокислот 112-115 в альфа-синуклеине (SEQ ID NO: 19 (ILED)).
5. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 1 или 4, которые представляют собой GM285.
6. Моноклональное антитело или его антигенсвязывающий фрагмент согласно предыдущим вариантам осуществления, где антитело содержит интактное антитело или состоит из него.
7. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, содержащие антигенсвязывающий фрагмент, выбранный из группы, состоящей из Fv-фрагментов (например, одноцепочечного Fv и связанного дисульфидной связью Fv), Fab-подобных фрагментов (например, Fab-фрагментов, Fab'-фрагментов и F(ab)2-фрагментов) и доменных антител (например, отдельных вариабельных VH-доменов или вариабельных VL-доменов), или состоящие из него.
8. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, где моноклональное антитело выбрано из группы, состоящей из антител подтипа IgG1, IgG2, IgG3 и IgG4.
9. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, где антитело или антигенсвязывающий фрагмент характеризуется одним или несколькими из следующих свойств:
(i) аффинностью связывания (KD) к альфа-синуклеину в диапазоне 0,5-10 нМ, например 1-5 нМ или 1-2 нМ;
(ii) способностью подавлять усечение протеазами фибрилл альфа-синуклеина;
(iii) способностью обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
(iv) способностью снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo;
(v) способностью, при длительном введении, восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
(vi) способностью предотвращать затравочное действие альфа-синуклеина (такое как накопление нерастворимого фосфорилированного альфа-синуклеина in vitro и/или в мышиной модели болезни Паркинсона); и/или
(vii) способностью связывать усеченный альфа-синуклеин в головном мозге человека.
10. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, которые являются человеческими или гуманизированными.
11. Моноклональное антитело или моноклональное антитело согласно вариантам осуществления 1-3 и 6-10, или их антигенсвязывающий фрагмент, содержащие вариабельный домен тяжелой цепи, содержащий следующие CDR:
a) GFTFSSYAMT (SEQ ID NO: 1) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
b) AIRS(N/S/Q/H) GDRTD YADSVKG (SEQ ID NO: 2, 33, 34, 35) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; или
c) AKNWAPFDS (SEQ ID NO: 3) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
12. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 11, содержащие вариабельный домен тяжелой цепи, содержащий CDR с SEQ ID NO: 1 и 3 и CDR с SEQ ID NO: 2 и 33, 34 или 35.
13. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 11, содержащие вариабельный домен тяжелой цепи, или состоящие из него, выбранный из группы, состоящей из
а) 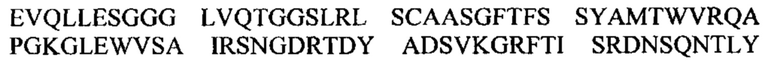
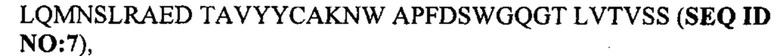
b) 
с)  или
или
d) 
14. Моноклональное антитело или моноклональное антитело согласно вариантам осуществления 1-3 и 6-13, или их антигенсвязывающий фрагмент, содержащие вариабельный домен легкой цепи, содержащий следующие CDR:
a) ASQSVSSSYLA (SEQ ID NO: 4) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
b) GASSRAT (SEQ ID NO: 5) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; или
c) QQYGSSPWT (SEQ ID NO: 6) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
15. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 14, содержащие вариабельный домен легкой цепи, содержащий CDR с SEQ ID NO: 4, 5 и 6.
16. Антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 14, содержащие вариабельный домен легкой цепи, содержащий аминокислотную последовательность:
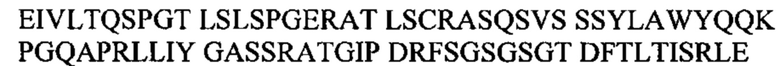
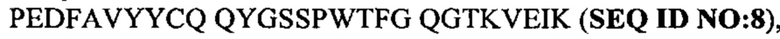
или состоящий из нее.
17. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 14, содержащие легкую цепь, содержащую аминокислотную последовательность:
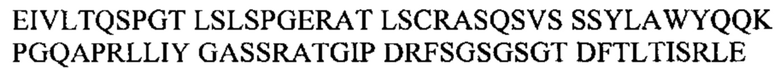
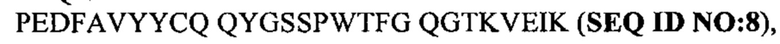
или состоящую из нее.
18. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 1-3 и 6-17, содержащие вариабельный домен легкой цепи, содержащий аминокислотную последовательность с SEQ ID NO: 8 или состоящие из нее, и вариабельный домен тяжелой цепи, содержащий аминокислоты, приведенные в любом из SEQ ID NO: 7, 33, 34 или 35, или состоящий из них.
19. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 1-3 и 6-18, содержащие легкую цепь, содержащую аминокислотную последовательность с SEQ ID NO: 8 или состоящую из нее, и вариабельный домен тяжелой цепи, содержащий аминокислоты, приведенные в SEQ ID NO: 34, или состоящий из них, имеющие повышенную термостабильность, такую как повышенная стабильность, предотвращающая агрегирование и развертывание, как показано на фиг. 27, являющиеся на 2%-10% более стабильными при значениях температуры выше 65°C по сравнению с GM37 wt, на 2%-8% более стабильными при значениях температуры выше 65°C по сравнению с GM37 wt или на 2%-5% более стабильными при значениях температуры выше 65°C по сравнению с GM37 wt.
20. Моноклональное антитело или моноклональное антитело согласно вариантам осуществления 1-10, или их антигенсвязывающий фрагмент, содержащие вариабельный домен тяжелой цепи, содержащий следующие CDR:
a) AASGFTFSRFTMT (SEQ ID NO: 20) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
b) AISGSGGGTS YADSVKG (SEQ ID NO: 21) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; или
c) AKNWAPFDY (SEQ ID NO: 22) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
21. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 20, содержащие вариабельный домен тяжелой цепи, содержащий CDR с SEQ ID NO: 20, 21 и 22.
22. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 20, содержащие вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность:

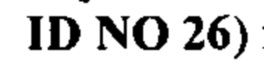 или состоящий из нее.
или состоящий из нее.
23. Моноклональное антитело или моноклональное антитело по любому из вариантов осуществления 1-10 и 20-22, или их антигенсвязывающий фрагмент, содержащие вариабельный домен легкой цепи, содержащий следующие CDR:
d) RASQSVSRSYLA (SEQ ID NO: 23) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах;
e) GASSRAT (SEQ ID NO: 24) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах; или
f) QQYGSSPWT (SEQ ID NO: 25) или аминокислотную последовательность, имеющую не более 4 отличий в аминокислотах, или не более 3 отличий в аминокислотах, или не более 2 отличий в аминокислотах, или не более 1 отличия в аминокислотах.
24. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 23, содержащие вариабельный домен легкой цепи, содержащий CDR с SEQ ID NO: 23, 24 и 25.
25. Антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 24, содержащие вариабельный домен легкой цепи, содержащий аминокислотную последовательность:
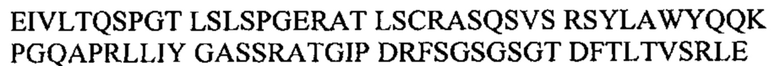
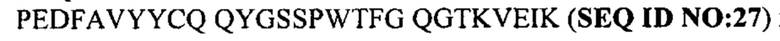 или
или
состоящий из него.
26. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, содержащие вариабельный домен легкой цепи, содержащий аминокислотную последовательность с SEQ ID NO: 27 или состоящий из нее, и вариабельный домен тяжелой цепи, содержащий аминокислоты, приведенные в любом из SEQ ID NO: 26, или состоящий из них.
27. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, содержащие Fc-область.
28. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, дополнительно содержащие фрагмент для увеличения периода полужизни in vivo данного средства.
29. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 28, где фрагмент для увеличения периода полужизни in vivo выбран из группы, состоящей из полиэтиленгликоля (PEG), сывороточного альбумина человека, гликозилирующих групп, жирных кислот и декстрана.
30. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, где полипептид антитела дополнительно содержит выявляемый фрагмент.
31. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 30, где выявляемый фрагмент представляет собой флуоресцентную метку, хемилюминесцентную метку, парамагнитную метку, радиоизотопную метку или ферментную метку.
32. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 30 или 31, где выявляемый фрагмент содержит радиоактивный изотоп или состоит из него.
33. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 32, где радиоактивный изотоп выбран из группы, состоящей из 99mTc, 111In, 67Ga, 68Ga, 72As, 89Zr, 123I и 201Tl.
34. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 30, где выявляемый фрагмент содержит парамагнитный изотоп или состоит из него.
35. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 34, где парамагнитный изотоп выбран из группы, состоящей из 157Gd, 55Mn, 162Dy, 52Cr и 56Fe.
36. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из вариантов осуществления 30-35, где выявляемый фрагмент выявляется с помощью методики визуализации, такой как SPECT, PET, MRI, оптическая или ультразвуковая визуализация.
37. Моноклональное антитело или его антигенсвязывающий фрагмент согласно любому из вариантов осуществления 30-36, где выявляемый фрагмент опосредованно соединен с антителом, или его антигенсвязывающим фрагментом, с помощью соединяющего фрагмента.
38. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 37, где соединяющий фрагмент выбран из группы, состоящей из производных 1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусной кислоты (DOTA), дефероксамина (DFO), производных диэтилентриаминпентауксусного авида (DTPA), производных S-2-(4-изотиоцианатобензил)-1,4,7-триазациклононан-1,4,7-ацетилацетоуксусной кислоты (NOTA) и производных 1,4,8,11-тетраазациклододекан-1,4,8,11-тетрауксусной кислоты (ТЕТА).
39. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент согласно любому из предыдущих вариантов осуществления, или входящую в их состав полипептидную цепь.
40. Молекула нуклеиновой кислоты согласно варианту осуществления 39, где молекула представляет собой молекулу кДНК.
41. Молекула нуклеиновой кислоты согласно варианту осуществления 30 или 31, кодирующая тяжелую цепь антитела или ее вариабельный домен.
42. Молекула нуклеиновой кислоты согласно любому из вариантов осуществления 39-41, кодирующая легкую цепь антитела или ее вариабельный домен.
43. Вектор, содержащий молекулу нуклеиновой кислоты согласно любому из вариантов осуществления 39-42.
44. Рекомбинантная клетка-хозяин, содержащая молекулу нуклеиновой кислоты согласно любому из вариантов осуществления 39-42 или вектор согласно варианту осуществления 43.
45. Способ получения антитела или антигенсвязывающего фрагмента согласно любому из вариантов осуществления 1-27, при этом способ предусматривает культивирование клетки-хозяина согласно варианту осуществления 44 в условиях, обеспечивающих возможность экспрессии кодируемого антитела или его антигенсвязывающего фрагмента.
46. Фармацевтическая композиция, содержащая моноклональное антитело или антигенсвязывающий фрагмент согласно любому из вариантов осуществления 1-35 и фармацевтически приемлемый носитель.
47. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 1-35 для применения в медицине.
48. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 1-35 для применения при лечении, диагностике или визуализации синуклеопатии.
49. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 48 для применения при лечении болезни Паркинсона (в том числе идиопатической и наследственной форм болезни Паркинсона), болезни Гоше, болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBV), комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности и множественной системной атрофии.
50. Применение моноклонального антитела или его антигенсвязывающего фрагмента согласно вариантам осуществления 1-35 при изготовлении лекарственного препарата для лечения, диагностики или визуализации синуклеопатии.
51. Применение моноклонального антитела или его антигенсвязывающего фрагмента согласно варианту осуществления 50 при изготовлении лекарственного препарата для лечения болезни Паркинсона (в том числе идиопатической и наследственной форм болезни Паркинсона), болезни Гоше, болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBV), комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности и множественной системной атрофии.
52. Способ лечения, диагностики или визуализации синуклеопатии у субъекта, при этом указанный способ предусматривает введение фармацевтической композиции согласно варианту осуществления 46 указанному субъекту в эффективном количестве.
53. Антитело или его антигенсвязывающий фрагмент для применения согласно варианту осуществления 48, или применения согласно варианту осуществления 50, или способ согласно варианту осуществления 52 для лечения болезни Паркинсона (в том числе идиопатической и наследственной формы болезни Паркинсона), болезни Гоше, болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBV), комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности и множественной системной атрофии.
54. Антитело или его антигенсвязывающий фрагмент для применения; или применение; или способ согласно варианту осуществления 52 или 53, где лечение является длительным.
55. Антитело или его антигенсвязывающий фрагмент для применения; или применение; или способ согласно варианту осуществления 52, где длительное лечение продолжается в течение по меньшей мере 2 недель, например, в течение по меньшей мере 1 месяца, 6, месяцев, 1 года или дольше.
56. Антитело или его антигенсвязывающий фрагмент для применения; или применение; или способ согласно любому из вариантов осуществления 47-55, где субъектом является человек.
57. Набор, содержащий антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 1-35.
58. Набор согласно варианту осуществления 57 для применения в медицине.
59. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 30-35 для применения при выявлении наличия или измерении количества указанного альфа-синуклеина в головном мозге или жидкости организма субъекта.
60. Моноклональное антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 59, где указанное выявление или измерение предусматривает визуализацию in vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
61. Моноклональное антитело или его антигенсвязывающий фрагмент согласно вариантам осуществления 30-35, где указанное выявление или измерение предусматривает визуализацию ex vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
ПРИМЕРЫ
Пример 1. Скрининг антител
1. Получение иммуногена и лиганда
Для применения в качестве иммуногенов получали и продуцировали следующие белки, показанные на фиг. 1. Мышей иммунизировали тремя иммуногенами: фибриллами полноразмерного рекомбинантного альфа-синуклеина человека; рекомбинантным белком альфа-синуклеином человека, содержащим аминокислоты 1-60 (Rpeptide, Богарт, Джорджия), и рекомбинантным белком альфа-синуклеином человека, содержащим аминокислоты 1-119. Для получения фибрилл из полноразмерного альфа-синуклеина применяли лиофилизированный продукт от Rpeptide, Богарт, Джорджия (номер по каталогу S-1001-2). Его растворяли в буфере на основе 20 мМ Tris и 300 мМ NaCl до концентрации белка 1 мг/мл. Для получения фибрилл раствор белка инкубировали в виде 170 мкл аликвот в 96-луночном планшете с керамической гранулой диаметром 70 мкм в каждой лунке при 200 об./мин. в инкубаторе с шейкером Vortemp 56 (Labnet International, Эдисон, Нью-Джерси, США) при 37°C в течение 7 дней, а после образования фибрилл добавляли тиофлавин Т и измеряли флуоресценцию в одной из лунок. Рекомбинантный альфа-синуклеин, содержащий аминокислоты 1-60, растворяли в воде с получением концентрации 1 мг/мл.
Рекомбинантный альфа-синуклеин, содержащий аминокислоты 1-119, получали с использованием приведенной далее конструкции. Синтетический ген, кодирующий гистидиновую метку из 6 аминокислот, за которой следовал сайт расщепления фактором Ха и последовательность, кодирующая аминокислоты 1-119 альфа-синуклеина человека:

был синтезирован с помощью Genscript и клонирован в сайт NdeI-XhoI в векторе экспрессии рЕТ24а(+) (Novagen).
Вектор экспрессии вводили с помощью трансформации в Е. coli штамма BL21 и отдельную колонию забирали для экспрессии с помощью системы аутоиндукции экспрессии в течение ночи от компании Novagen (пользовательский протокол ТВ383, рев. Н1005). Масштаб конечного объема культуры составлял 500 мл. Клетки собирали посредством центрифугирования в течение 10 мин. при 6000 g, а затем лизировали с помощью реагента для экстракции белка BugBuster (пользовательский протокол ТВ245, рев. 0304). После лизиса образец осветляли путем центрифугирования, а супернатант использовали для последующего очищения.
Белок с His-меткой очищали на 5 мл колонке HisTrap (GE healthcare), уравновешенной в 20 мМ фосфате натрия, рН 7,5, 1 М NaCl (А-буфер). После внесения образца и промывания с помощью А-буфера белок элюировали в градиенте до 0,25 М имидазола в А-буфере в течение 20 объемов колонки. Фракции по 5 мл собирали и анализировали с помощью SDS-PAGE. Фракции с представляющим интерес белком объединяли, концентрировали и вносили в колонку для эксклюзионной хроматографии S200 (26/60) (GE healthcare) в 10 мМ Tris, рН 7,4, 300 мМ NaCl. Фракции еще раз объединяли согласно наличию в SDS-PAGE полосы с ожидаемым размером.
Для удаления N-концевой метки очищенный альфа-синуклеин 1-119 с His-меткой инкубировали с фактором Ха в соотношении 1:50 с использованием набора от Novagen (69037-3FRX). После инкубирования в течение ночи фактор Ха удаляли порциями с помощью агарозы Xarrest. Расщепленный альфа-синуклеин 1-119 окончательно очищали с помощью пермиссивной хроматографии HisTrap, как описано выше. Из элюата получали очищенный альфа-синуклеин 1-119 и концентрировали его до ~400 мкг/мл с помощью концентрирующих устройств Centricon.
Альфа-синуклеин (Rpeptide) регидратировали в PBS при 2 мг/мл и при перемешивании по каплям добавляли пероксинитрит (100 мкл/мг белка). Затем нитрозилированный альфа-синуклеин подвергали диализу в 5 л PBS и хранили при -20°C.
Для окисления альфа-синуклеина использовали дофамин. Объединяли равные объемы 200 мкМ раствора дофамина-HCL (Sigma Р5244), приготовленного в 10 мМ PBS, рН 7,4, и 28 мкМ раствора альфа-синуклеина (Rpeptide) в 10 мМ PBS, рН 7,4. Полученный в результате 14 мкМ альфа-синуклеин/100 мкМ дофамина инкубировали при 37°C О/N (в течение ночи). Затем окисленный альфа-синуклеин подвергали диализу в PBS и хранили при -20°C.
Для осуществления скрининга разнообразной библиотеки антител к альфа-синуклеину получали различные нативные и химерные версии белков синуклеина. Конструкции для скрининга включали следующее: альфа-синуклеин человека, мыши, крысы и яванского макака, бета-синуклеин человека, гамма-синуклеин человека (фиг. 21 и 22) и, наконец, производное альфа-синуклеина, у которого отсутствовали остатки 120-140 альфа-синуклеина. Кроме того, получали серию из 4 конструкций, полученных перетасовкой: A-Syn-AAKK-BAP, A-Syn-ВААК-ВАР, A-Syn-BBAA-BAP, A-Syn-BBKK-BAP (SEQ ID NO: 11-14). Эти конструкции содержали линейные отрезки альфа-синуклеина человека (А), бета-синуклеина человека (В) и альфа-синуклеина курицы (К). Клонировали гены, содержащие метку в виде биотин-акцепторного пептида (ВАР), слитую с С-концом лигандов для облегчения сайт-специфического биотинилирования каждого из лигандов. Биотинилирование обеспечивало присоединение лигандов к гранулам, используемым в формате ELISA в растворе. Конструировали векторы экспрессии млекопитающих, несущие различные слитые конструкции альфа-синуклеина с ВАР-меткой (ASynBAP). Лиганды экспрессировали в клетках HEK 293 с помощью временной трансфекции (Genmab A/S).
2. Иммунизация
Антитела HuMab-Synuclein получали в результате иммунизации мышиной линии HuMAb HCo17-BALB/c и HCo12-BALB/c, мышей с двойным нокаутом по легкой каппа-цепи и тяжелой цепи иммуноглобулина (Ig) мыши, что предотвращает экспрессию антител, которые являются полностью мышиными (моноклональное антитело мыши, Medarex Inc., Сан-Хосе, Калифорния, США). Различные мышиные линии были сделаны трансгенными путем вставки локусов тяжелой цепи Ig человека и локусов легкой каппа-цепи Ig человека, и они отличались по количеству генов VH человека (вариабельного домена тяжелой цепи) и VL (вариабельного домена легкой цепи).
48 мышей поочередно иммунизировали внутрибрюшинно (IP) с помощью 20 мкг антигенов и подкожно (SC, в основание хвоста) тем же иммуногеном с интервалом 14 дней. Осуществляли максимум восемь иммунизации: 4 IP и 4 SC.
Первую иммунизацию проводили альфа-синуклеиновыми иммуногенами в полном адъюванте Фрейнда (CFA, Difco Laboratories, Детройт, Мичиган, США), следующие иммунизации - в неполном адъюванте Фрейнда (IFA). После обнаружения, что сывороточные титры являются достаточными (разведение сыворотки 1/50 или ниже было обнаружено положительным в анализе антиген-специфического скрининга, описанного выше, по меньшей мере в двух последовательных, проводимых два раза в неделю событиях скрининга), мышей дополнительно дважды стимулировали внутривенно (IV) с помощью 10 мкг иммуногенного белка альфа-синуклеина в 100 мкл PBS, за четыре и три дня до слияния.
Протоколы иммунизации показаны на фиг 1.
Антитело 37 было получено по протоколу иммунизации, согласно которому использовали фибриллы полноразмерного α-синуклеина человека, чередующиеся с усеченными на С-конце формами альфа-синуклеина с аминокислотами 1-60 и 1-119.
Антитело 285 было получено по протоколу иммунизации, согласно которому α-синуклеиновый мономер 1-140 человека использовали для первых 4 иммунизации. Если титр отсутствовал, иммунизацию продолжали фибриллами (ip/sc), в противном случае ее продолжали мономером.
3. Создание HuMab-гибридомы
HuMAb-мышей с выработкой достаточного антигенспецифического титра, как определено выше, умерщвляли и собирали селезенку и лимфатические узлы, прилегающие к брюшной аорте и полой вене. Слияние спленоцитов и клеток лимфатических узлов с линией мышиных клеток миеломы проводили путем электрослияния с использованием системы для электрослияния CEEF 50 (Cyto Pulse Sciences, Глен Берни, Мэриленд, США), по сути, в соответствии с инструкциями производителя. Слитые клетки высевали в среду для культивирования слитых клеток, содержащую 10% бычьей сыворотки FetalClone I (Perbio), 1 мМ пирувата натрия (Cambrex), 0,5 ед./мл пенициллина, 0,5 ед./мл стрептомицина (Cambrex), 50 мкМ 2-меркаптоэтанола (Invitrogen), 600 нг/мл интерлейкина 6 (IL-6) (Strathmann), 1 x HAT (Sigma) и 0,5 мг/мл канамицина (Invitrogen) в HyQ mADCF-Mab (Perbio). Через десять дней собирали супернатант и клетки обновляли средой для сбора, содержащей 10% бычьей сыворотки FetalClone I, 0,5 ед./мл пенициллина, 0,5 ед./мл стрептомицина, 600 нг/мл IL-6 и 1 x proHT (Cambrex) в HyQ mADCF-Mab. Супернатанты культур гибридомы подвергали скринингу с помощью первичных скрининговых исследований. Супернатанты охарактеризовывали в отношении связывания с восемью различными лигандами. В их число входили 4 ортолога: человека, мыши, крысы и яванского макака, альфа-синуклеин человека бета-синуклеин и гамма-синуклеин человека (SEQ ID NO: 37-41), и, наконец, их тестировали в отношении их способности связываться с производным альфа-синуклеина человека, у которого отсутствовали остатки 120-140 альфа-синуклеина.
Скрининг антител к альфа-синуклеину проводили с использованием формата высокоэффективной ELISA в суспензии с помощью автоматизированных систем обработки жидкости (Genmab A/S). Считывание планшетов выполняли с помощью двух систем: FMAT 8200 от Applied Biosystems использовали для считывания 384-луночных планшетов, а цитометр ImageXpress Velos Cytometer от Molecular Devices использовали для считывания 1536-луночных планшетов.
В ходе первичного скрининга клоны охарактеризовывали по их способности связываться с 8 различными лигандами. В их число входила серия из 4 конструкций, полученных перетасовкой: A-Syn-AAKK-BAP, A-Syn-BAAK-BAP, A-Syn-BBAA-BAP, A-Syn-BBKK-BAP (SEQ ID NO: 11-14), альфа-синуклеин-ВАР с делецией по 120-140, нитрованный альфа-синуклеин-ВАР человека и окисленный альфа-синуклеин-ВАР человека.
Вкратце, сыворотки или супернатант, потенциально содержащие специфические в отношении альфа-синуклеина антитела, добавляли к гранулам для обеспечения связывания с альфа-синуклеином и/или конструкциями, полученными на основе альфа-синуклеина. Связывание антител к альфа-синуклеину выявляли с помощью флуоресцентного конъюгата, DyLight649, конъюгированного с IgG козы к антителам человека, Fc-специфического. В скрининговые исследования в качестве положительных контролей были включены два известных антитела мыши к альфа-синуклеину, LB509 и Syn211. Для обеспечения специфического выявления антител к альфа-синуклеину при скрининге титра в 384-луночном формате в качестве отрицательного контроля использовали сывороточный пул антител к альфа-синуклеину, в то время как в анализе на основе 8 гранул в 1536-луночном формате использовали ChromPure IgG человека.
Клетки гибридомы из определенных в ходе первичного скрининга лунок с наилучшим результатом высевали в полутвердую среду, изготовленную из 40% CloneMedia (Genetix, Гэмпшир, Великобритания) и 60% полной среды HyQ 2х (Hyclone, Уолтем, США). В случае каждой лунки из первичного скрининга ее содержимое высевали в лунку 6-луночного черного планшета Genetix. Из каждой лунки забирали 25 субклонов с помощью системы ClonePix (Genetix). Субклоны забирали в среду для сбора. Спустя семь дней супернатанты субклонов снова подвергали скринингу в отношении специфического к синуклеину связывания IgG человека и измеряли концентрацию IgG человека с помощью платформы Octet (Fortebio, Менло-Парк, США). Из каждой лунки первичного скрининга выбирали наилучший субклон и размножали его в среде для размножения, содержащей только 600 нг/мл IL-6, 0,5 ед./мл пенициллина, 0,5 ед./мл стрептомицина и 1 х proHT. Субклоны размножали в следующем порядке: одна лунка 96-луночного планшета - одна лунка 24-луночного планшета - четыре лунки 24-луночного планшета - шесть лунок 6-луночного планшета. Клоны, полученные в результате этого процесса, были обозначены как первичные клоны (PC).
Дополнительные исследования по связыванию антитела проводили с использованием Octet 384RED (Fortebio, Менло-Парк, США). Растворы антител HuMab с концентрацией 2 мкг/мл получали путем разведения в разбавителе образцов (ForteBio, арт. №18-5028). Чувствительные к аминогруппам сенсоры (ForteBio, арт. №18-0008) использовали для иммобилизации HuMab. Перед связыванием с чувствительными к аминогруппам сенсорами HuMabs разводили в буфере MES, рН 6,0 (18-5027). Связывание осуществляли при 30°C и 1000 об./мин. следующим образом. Чувствительные к аминогруппам сенсоры предварительно смачивали в PBS и затем активировали с помощью раствора для активации EDC/NHS (ForteBio. арт. №18-1033/18-1034) (в соответствии с инструкцией производителя) в течение 300 секунд. На активированные сенсоры иммобилизировали HuMabs в течение 600 секунд.
Связывание 37 и 285 в платформе Octet с рекомбинантным альфа-синуклеином человека, яванского макака и мыши и отсутствие связывания с бета- или гамма-синуклеином человека показано на фигуре 2.
4. Анализ последовательностей специфических в отношении синуклеина вариабельных доменов HuMab и клонирование в векторы экспрессии
Из 0,2-5×106 клеток гибридомы получали общую РНК и из 100 нг общей РНК получали 5'-RACE-комплементарную ДНК (кДНК) с помощью набора для амплификации кДНК SMART RACE (Clontech) в соответствии с инструкциями производителя. Области, кодирующие VH и VL, амплифицировали с помощью ПЦР и непосредственно клонировали в рамке считывания в векторы экспрессии p33G1f и p33Kappa (содержащие последовательности, кодирующие константные домены IgG1 человека/каппа) путем независящего от лигирования клонирования (Aslanidis, С. and PJ de Jong, Nucleic Acids Res 1990; 18 (20): 6069-74). Для каждого антитела секвенировали 16 клонов VL и 16 клонов VH. Для дополнительного изучения и экспрессии отбирали клоны с корректной открытой рамкой считывания (ORF). Осуществляли временную экспрессию векторов всех комбинаций тяжелых цепей и легких цепей в клетках FreestyleTM 293-F с использованием 293fectin.
В случае секвенирования GM37 в VH-области был выявлен дополнительный цистеин в домене CDR3 в положении 106. Для исключения возможности неправильного сворачивания и потенциальной потери активности антител из-за образования дисульфидной связи в положении 106 осуществляли мутацию цистеина в серии.
Антитело сравнения 9Е4 создавали на основе последовательности VH и VL, полученной от гибридомы РТА-8221 (патент США 20080175838) (SEQ ID NO 42 и 43)
5. Экспрессия/очистка антител
Антитела получали путем введения трансфекцией в клетки HEK293 6Е с использованием векторов рТТ5 и PEIpro в качестве средства для временной трансфекции (Канадский национальный совет по научным исследованиям). Вкратце, тяжелые и легкие цепи вводили трансфекцией в клетки HEK293 с использованием PEIpro (VWR) и спустя 24 часа после трансфекции клетки дополняли TN1 (Sigma). Клетки выращивали до тех пор, пока жизнеспособность не достигала 50%, и выход антител измеряли с помощью набора easy IgG titre (Thermo). Культуральный супернатант фильтровали через 0,2 мкм тупиковые фильтры, загружали на 5 мл колонки с белком A (rProtein A FF, Amersham Bioscience) и элюировали с помощью 0,1 М лимонной кислоты-NaOH, рН 3. Элюат немедленно нейтрализовали посредством 2 М Tris-HCl, рН 9 и диализировали к 12,6 мМ NaH2PO4, 140 мМ NaCl, рН 7,4 (B. Braun), O/N. После диализа образцы подвергали стерилизующей фильтрации через 0,2 мкм тупиковые фильтры. Чистоту определяли с помощью SDS-PAGE, а концентрацию измеряли с помощью нефелометрии и поглощения при 280 нм. Очищенные антитела разделяли на аликвоты и хранили при -80°C.
Пример 2. Характеристика антител с помощью поверхностного плазменного резонанса
Связывание антител с альфа-синуклеином в режиме реального времени измеряли с помощью BIAcore® 3000. Поверхность для захвата получалипутем аминного связывания поликлонального антитела кролика к антителам мыши (часть набора для захвата антител мыши, GE Healthcare, № по кат. BR-1008-38) в первой проточной ячейке (Fc1) и второй проточной ячейке (Fc2) чипа СМ5 (BIAcore®). Антитело мыши захватывали в Fc2 при концентрации, необходимой для достижения уровня лиганда около 500RU. Обеспечивали стабилизацию исходного уровня в течение 10 мин. перед введением аналита (ASynBAP) в Fc1-2 при 30 мкл/мин. ASynBAP прогоняли соответственно при 100-3200 нМ и 25-3200RU. Наивысшую концентрацию в каждой серии титрования прогоняли в двух повторностях. Поверхность восстанавливали посредством 10 мМ глицин-HCl, рН 1,7 (30 с инъекция) для удаления захваченного антитела мыши и аналита в конце каждого цикла. HBS-EP (GE Healthcare, № по кат. BR-1001-88) использовали в качестве подвижного буфера и разбавителя образца во всех экспериментах, а анализ проводили при 25°C. Все образцы до получения хранили при 4°C.
Реакцию, зарегистрированную в Fc1, где было иммобилизировано иммобилизованное антитело, но отсутствовало иммобилизованное антитело к альфа-синуклеину, вычитали из реакции в Fc2. Алгоритм связывания 1:1 или 2:1 использовали для приближения к набору данных с помощью программного обеспечения BIAevaluation версии 4.1.1. Результаты можно видеть на фиг. 3, 4 и 5, на которых показано связывание антитела 37, 285 и 9Е4 с альфа-синуклеином человека.
Пример 3. Картирование эпитопов
Картирование эпитопов антител к альфа-синуклеину проводили с помощью матриц перекрывающихся линейных пептидов в Pepscan (Pepscan Zuidersluisweg 2 8243 RC, Лелистад, Нидерланды). Связывание антитела с каждым из синтезированных пептидов из 20 звеньев тестировали в ELISA на основе Pepscan. Матрицу с линейными пептидами, охватывающую всю кодирующую последовательность альфа-синуклеина, а также все пептиды с окисленными метионинами или нитрозилированными тирозинами, инкубировали с раствором первичного антитела (в течение ночи при 4°C). После промывания матрицы с пептидами инкубировали с 1/1000 разведением конъюгата пероксидазы и антитела (SBA, № по кат. 2010-05) в течение одного часа при 25°C. После промывания добавляли субстрат для пероксидазы 2,2'-азиноди-3-этилбензтиазолинсульфонат (ABTS) и 2 мкл/мл 3-процентного Н2О2. Через один час измеряли проявление цвета. Проявление цвета оценивали количественно с помощью камеры с полупроводниковой светочувствительной матрицей (CCD) и системы обработки изображений. Для обработки данных значения получали с CCD-камеры в диапазоне от 0 до 3000 mAU аналогично со стандартным ELISA-считывающим устройством для 96-луночных планшетов. Результаты количественно оценивали и сохраняли в базе данных Peplab. Изредка лунка содержала воздушный пузырек, что приводило к ложноположительному значению, тогда снимки проверяли вручную, а любые значения, вызванные воздушным пузырьком, оценивали как 0. Данные связывания для антитела 37 и 285 с пептидами, содержащими соответственно последовательность ILEDMP или ILED, можно увидеть на фигуре 7.
Пример 4. Иммунопреципитация альфа-синуклеина из гомогенатов передней поясной коры головного мозга человека от пациентов с деменцией с тельцами Леви
Способность антител связываться и осаждать альфа-синуклеин из неочищенных гомогенатов поясной коры от человека с DLB или здорового контроля (отмеченного *) анализировали с помощью иммунопреципитации. Замороженный образец из поясной коры человека (полученный с помощью Tissue Solutions Ltd, Шотландия) рассекали на части в криостате и 100 мг образца добавляли к 1600 мкл реагента для лизиса клеток cellytic М (Sigma С2978), содержащему ингибиторы протеаз и ингибиторы фосфатаз (Roche). Ткань головного мозга гомогенизировали до полного растворения образца с помощью бисерного гомогенизатора Precellys (Bertin technologies, Франция) 4×30 с при 5000 об./мин. Раствор центрифугировали при 3000×g и супернатант использовали в качестве неочищенного гомогената для иммунопреципитации.
Для иммунопреципитации 10 мкг антитела смешивали с магнитными гранулами Dynabeads protein G с использованием инструкций производителя (Life Technologies, Пейсли, Великобритания). Неочищенный гомогенат головного мозга разводили в 30 раз в буфере для лизиса (Sigma). Связанные с антителами гранулы Dynabeads смешивали с 500 мкл разведенного гомогената и инкубировали 90 минут при комнатной температуре при непрерывном перемешивании во встряхивателе. После инкубирования гранулы промывали в промывочном буфере и связанные антигены элюировали с использованием неденатурирующего буфера для элюирования в соответствии с инструкциями производителя (протокол Dynabeads G, Life Technologies, Пейсли, Великобритания). Выход иммунопреципитации визуализировали с помощью вестерн-блоттинга с обнаружением моноклонального антитела мыши к альфа-синуклеину человека (4В12, Thermo Scientific). Паттерны полос, представляющих собой осаждаемые формы альфа-синуклеина с различной молекулярной массой, различались у антител 37, варианта 2 37 и 285 и антитела сравнения 9Е4 в том, что антитело 37, 37v2 и 285 может иммунопреципитировать основную разновидность альфа-синуклеина, полноразмерный альфа-синуклеин (FL asyn 1-140) и усеченные на С-концевом конце кразновидности (1-135 и 1-119/122), тогда как антитело 9Е4 не могло иммунопреципитировать усеченные разновидности 1-119/122. Фигура 9.
Пример 5. Подавление усечения протеазами фибрилл альфа-синуклеина антителами в клеточной культуре
Мономеры и фибриллы рекомбинантного альфа-синуклеина могут поглощаться первичными нейронами в культуре. Как показано схематически на фигуре 10, после поглощения альфа-синуклеина в нейронах он может быть обработан внутриклеточными протеазами, такими как кальпаин I, при этом основной сайт чувствительности к протеазе приходится на аминокислоту 119/122. Для изучения усечения альфа-синуклеина протеазами получали первичные кортикальные нейроны мышей, как описано в Elvang et al. 2009 (Elvang et al. J Neurochem. 2009; 110(5): 1377-87), и обрабатывали цитарабином на DIV3 (3-й день in vitro) для подавления роста астроцитов. На DIV4 (4-й день in vitro) нейроны обрабатывали посредством обработанных ультразвуком (5 мин. при 50% мощности в устройстве для обработки ультразвуком с чашеобразным звукоприемником) предварительно сформировавшихся фибрилл альфа-синуклеина (PFF) с конечной концентрацией 0,7 мкМ отдельно или вместе с антителами при указанных концентрациях. После 24 часов инкубирования собирали среду и лизировали клетки. Вестерн-блоттинг проводили как на средах, так и на клеточном лизате с использованием антитела 4В12 (фиг. 11А) (Pierce MA1-90346) и вторичного антитела к антителам мыши. После зондирования посредством 4В12 + антител к антителам мыши блоты очищали и вторично зондировали с помощью антитела к IgG человека. На блоте с 4В12 можно видеть, что в средах от клеток, обработанных только PFF, были интенсивные полосы на уровне 14 и 12 кДа, где 14 кДа представляет собой полноразмерный альфа-синуклеин (FL-asyn), а 12 кДа представляет собой усеченный на С-конце фрагмент 1-119/122 (CT-asyn). В дополнение к этому присутствовали полосы с более высокой молекулярной массой, которые, скорее всего, представляли собой устойчивые к SDS олигомерные разновидности. Совместная обработка изотопическим контрольным антителом В12 не изменяла данный паттерн протеолиза или поглощения.
В средах от клеток, обработанных фибриллами вместе с 37, в основном присутствовал полноразмерный альфа-синуклеин (14 кДа) и лишь небольшие количества полосы усеченной на конце разновидности (12 кДа). В клеточных лизатах от клеток, обработанные фибриллами вместе с 37, присутствовал только полноразмерный альфа-синуклеин, что свидетельствовало о том, что 37 предотвращает расщепление FL-альфа-синуклеина. Более того, общее количество FL-альфа-синуклеина было сниженным по сравнению с клетками, обработанными только PFF или контрольным антителом В12. Несколькими группами (Games et al, Am J of Pathol, Vol. 182, No. 3, March, 2013; Ritchie et al, Health, Vol. 4, Special Issue, 1167-1177, 2012; Mishizen-Eberz, Biochemistry, 2005, 44, 7818-7829; Dufty et al, Am J of Pathol, Vol. 170, No. 5, May 2007) было показано, что альфа-синуклеин может расщепляться кальпаином-1. Было обнаружено, что сайт расщепления кальпаином-1 у фибриллированного альфа-синуклеина находится в области 114-122 (Mishizen-Eberz, J of Neurochem, 86, 836-847, 2003). In vivo у трансгенных животных и в головном мозге человека 1-119/122, по-видимому, является основным продуктом расщепления, в альфа-синуклеине расщепление, судя по всему, имеет место после аспарта 119 или аспарагина 122, который дезамидирован до аспартата и расщепляется кальпаином или другой протеазой с аналогичной специфичностью расщепления. Эти результаты свидетельствуют, что антитело 37 способно подавлять С-концевое усечение альфа-синуклеина. Эпитоп антитела 37 перекрывается с сайтом связывания фермента кальпаин-1, поэтому связывание 37 с альфа-синуклеином может непосредственно подавлять связывание и расщепление, опосредованное кальпаином-1 (фиг. 10 и 11).
Эпитоп 285 перекрывается с эпитопом 37, и ожидается, что он будет подавлять расщепление протеазами. Аминокислотная последовательность 37v2 отличается от 37 только по одной аминокислоте в CDR и имеет сходное с 37 связывание, поэтому также ожидается, что оно, аналогично 37, будет подавлять расщепления протеазами. Для изучения того, является ли эффект антител дозозависимым, был поставлен 24-часовой эксперимент с совместным добавлением PFF и антител к первичным корковым нейронам. Концентрация PFF была стабильной (10 мкг/мл), тогда как концентрации контрольного антитела В12 и антител 37, 37v2 и 285, которые были протестированы, составляли 10, 5,1 и 0,1 мкг/мл. Выявление альфа-синуклеина на вестерн-блотах проводили с помощью антитела 1904/4В12 (Abcam), которое имеет эпитоп в области 103-108 и, следовательно, связывается как с FL, так и с усеченным на С-конце альфа-синуклеином (фиг. 11В). Как можно видеть из фигуры 11В, как GM37, так и 37v2, так и GM285 оказывают дозозависимое подавление расщепления протеазами с почти полным подавлением расщепления при высокой концентрации антитела.
Пример 6. Антитело-опосредованное подавление затравочного действия в отношении агрегирования альфа-синуклеина в клеточной культуре
В нескольких исследованиях было показано, что экзогенное добавление фибриллярных агрегатов рекомбинантного альфа-синуклеина может привести к их проникновению в клетки, и мобилизации эндогенного альфа-синуклеина, и индукции агрегирования и фосфорилирования альфа-синуклеина in vitro и in vivo в агрегаты, которые имеют сходство с LB. (Volpicelli-Daley et al. 2011, Luk et al. 2012a, Luk et al. 2012b, Recasens et al. 2013, Peelaerts et al. 2015). Для исследования затравочного действия в отношении эндогенного альфа-синуклеина мыши с использованием затравочных частиц рекомбинантного альфа-синуклеина первичные корковые нейроны мыши, полученные, как указано выше, высевали в 96-луночные планшеты (15000 клеток на лунку). На 5-й день культивирования in vitro (DIV) 50% среды заменяли и дополняли арабинозидом цитозина (конечная конц. 1 мкМ). На DIV 6 половину среды заменяли на кондиционированную глией среду с фибриллярным материалом альфа-синуклеина, либо неочищенными затравочными частицами фибрилл, либо чистыми затравочными частицами. Неочищенные затравочные частицы фибрилл получали из рекомбинантного мономерного альфа-синуклеина человека, который был выделен из бактерий, а мономеры фильтровали через отсекающий фильтр Amicon Ultra 100000 (Millipore, № по кат. UFC510096) и доводили до концентрации 1 мг/мл в PBS, рН 7,4. Для получения неочищенных затравочных частиц фибрилл раствор мономера инкубировали в термомиксере при 37С с непрерывным перемешиванием (800 об./мин.) до достижения плато (оценивали путем ежедневных измерений с тиофлавином S). Для минимизации испарения добавляли каплю минерального масла для покрытия раствора. Общее время для инкубирования составляло 5-7 дней. Чистые затравочные частицы получали из неочищенных фибриллярных затравочных частиц, которые центрифугировали для их очищения, а образовавшийся осадок ресуспендировали в свежем PBS и обрабатывали ультразвуком. Антитела добавляли один раз на DIV 6 вместе с неочищенными затравочными частицами альфа-синуклеина. Половину среды в первичных нейронах каждую неделю заменяли кондиционированной глией средой, поддерживая их до DIV21. Нейроны фиксировали и окрашивали по фосфосинуклеину с помощью антитела кролика, специфичного в отношении фосфорилирования альфа-синуклеина по аминокислоте S129 (abcam 51253), с последующим добавлением флуоресцентно меченого антитела к антителу кролика, флуоресценцию количественно оценивали с помощью автоматизированной флуоресцентной микроскопии на Cellomics Arrayscan. Ядра были выявлены в одном канале, и по ним определяли количество пригодных клеток. Пятна фосфорилированного альфа-синуклеина были обнаружены в другом канале в предопределенной кольцеобразной области, окружающей ядро, таким образом представляя собой цитоплазму клеток. Рассчитывали среднее количество пятен на клетку. Пример окрашивания клеток показан на фиг. 12А. Пятна фосфорилированного альфа-синуклеина не наблюдали у необработанных нейронов. Нейроны, инкубированные с неочищенными или чистыми затравочными частицами (1-10 нг на лунку), индуцировали фосфорилирование альфа-синуклеина (фиг. 12А). В нейритах фосфорилированный синуклеин имел вид пятен или точек, а некоторая часть фосфо-синуклеина в нейритах выглядела удлиненной.
Для исследований разделения на фракции клетки собирали в фосфатно-солевой буферный раствор (PBS) и центрифугировали. Осадок ресуспендировали в 1% буфере на основе Тритона с ингибиторами протеаз. Образцы выдерживали на льду в течение 15 мин. с последующей обработкой ультразвуком. Образцы центрифугировали при 100000×g в течение 30 мин. при 4С. Супернатант собирали и помечали как растворимую фракцию. Осадок промывали один раз в буфере на основе Тритона и ресуспендировали в 1% SDS-буфере с последующей обработкой ультразвуком. Образцы снова центрифугировали при 100000×g в течение 30 мин. Супернатант собирали в виде нерастворимой фракции. Измеряли концентрации белка и образцы прогоняли на 4-12% геле SDS_PAGE, промакивали на мембраны и осуществляли выявление альфа-синуклеина и фосфорилированного альфа-синуклеина (S129P) с помощью соответственно антитела 4В12/1904 (Thermo scientific: МА1-90346-синуклеин человека), антитела S129P-asyn (abcam 51253) и антитела к синуклеину мыши (передача сигналов в клетке - D37A6).
На фиг. 12В показаны результаты вестерн-блоттинга растворимой и нерастворимой фракции из первичных нейронов с неочищенными затравочными частицами и без них. Как можно видеть из фигуры 12В, добавление затравочных частиц приводило к накоплению эндогенного альфа-синуклеина мыши и p-S129-альфа-синуклеина, а также мультимеров фосфорилированного альфа-синуклеина мыши в нерастворимой фракции клеток.
Для проверки, могут ли антитела подавлять затравочное действие, использовали затравочные частицы альфа-синуклеина синуклеина в конц. 6,6 нМ (10 нг/лунка). Для получения дозозависимого эффекта на DIV 6 совместно добавляли антитела и затравочные частицы альфа-синуклеина в различных концентрациях (начиная с наиболее высокой конц. антитела на уровне 133 нМ и до 133 пМ). Нейроны снова фиксировали и окрашивали по фосфо-синуклеину (abcam 51253) и флуоресценцию от клеток количественно оценивали с помощью автоматизированной флуоресцентной микроскопии на Cellomics arrayscan. На Cellomics arrayscan подсчитывали пятна/точки на клетку. Как можно видеть из фигуры 12С, как антитело 37, так и 37v2 и антитело 285 уменьшали фосфорилирование альфа-синуклеина в нейронах дозозависимым образом со схожим максимальным ингибированием для 37, 37v2 и 285 (около 70-75%) и ЕС50 около 5 нМ. Фракционирование клеточных белков на растворимую и нерастворимую фракцию после обработки антителом в наиболее высокой концентрации (133 нМ) демонстрировало, что все антитела 37, 37v2 и 285 подавляли усечение рекомбинантных неочищенных затравочных частиц и накопление усеченного на С-конце фрагмента (СТ a-syn) и уменьшали накопление фосфорилированного эндогенного альфа-синуклеина мыши и агрегированных форм альфа-синуклеина мыши в нерастворимой фракции, что видно на фигуре 12D.
Пример 7. Сильные электрофизиологические эффекты антител к альфа-синуклеину in vivo
Высокие уровни экспрессии альфа-синуклеина человека присутствуют в гиппокампе трансгенных мышей F28-snca, модели, у которой сверхэкспрессируется альфа-синуклеин дикого типа под контролем промотора альфа-синуклеина мыши (Westerlund М, et al. Mol Cell Neurosci. 2008 Dec; 39 (4):586-91). Оценку синаптической передачи и пластичности в СА1-зоне гиппокампа у 4-6-месячных самцов трансгенньгх мышей F28-snca и контрольных мышей соответствующего возраста проводили с помощью электрофизиологических методов in vivo. Из данных видно, что базальная синаптическая передача значительно снижалась у трансгенных мышей F28-snca по сравнению с контрольными мышами соответствующего возраста (фиг. 13).
Самцов трансгенных мышей F28-snca и контрольных мышей соответствующего возраста (CRO breeding, Taconic Europe A/S) в возрасте от 4 до 6 месяцев размещали по одному в условиях с контролируемой температурой (22±1,5°C) и влажности (55-65%) и содержали при цикле чередования света и темноты 12:12 часов (свет включали в 06:00 ч). Пища и вода были доступны без ограничений.
Животных анестезировали путем внутрибрюшинной (i.p.) инъекции уретана (1,2 г/кг). Затем мышей размещали в стереотаксической рамке, их температуру доводили до 37,5°C с помощью электрической грелки, и обнажали череп. На лобную кость помещали платиновую проволоку, которая выступала в качестве контроля, и просверливали дополнительное отверстие для введения регистрирующих и стимулирующих электродов в гиппокамп по следующим координатам в соответствии с атласом Paxinos и Franklin (Paxinos and Franklin's the Mouse Brain in Stereotaxic Coordinates, 4th Edition, 2001): регистрирующий - на 1,5-1,7 мм кзади от брегмы, на 1,0-1,2 мм латеральнее от срединной линии, на 1,4-1,7 мм ниже поверхности головного мозга; стимулирующий - на 1,8-2,0 мм кзади от брегмы, на 1,5-1,7 мм латеральнее от срединной линии, на 1,5-1,7 мм ниже поверхности головного мозга. Животных оставляли в стереотаксической рамке в течение всего периода регистрации данных и регулярно проверяли уровень их анестезии.
Потенциалы поля (fEPSP) вызывали в СА1 путем электрической стимуляции коллатерали Шеффера каждые 30 с, а глубину расположения регистрирующего электрода регулировали до тех пор, пока в ответ на монополярный прямоугольный импульс не был зарегистрирован отрицательный fEPSP. Градиент вызванного fEPSP измеряли в диапазоне 30-70% от максимальной амплитуды fEPSP.
После индукции оптимального fEPSP базальную синаптическую передачу оценивали по зависимости между интенсивностью стимуляции и градиентом вызванного fEPSP (зависимости "вход-выход"). Различные интенсивности стимуляции составляли 0,25, 50, 75,100, 150, 200, 300, 400 и 500 мкА, и их последовательно прилагали в порядке возрастания с 2-3 повторами для каждой интенсивности. Было обнаружено, что базальная синаптическая передача значительно ухудшалась у трансгенных мышей F28-snca по сравнению с контрольными мышами соответствующего возраста.
Установленные нарушения базальной синаптической передачи у трансгенных мышей F28-snca использовали для тестирования GM37, GM285 и антитела сравнения h9E4 в отношении их способности блокировать опосредованный альфа-синуклеином эффект.
Регистрации данных проводили во всех экспериментах через 3-6 ч. после введения однократной дозы антитела дозой 15 мг/кг (i.p.). Базальную синаптическую передачу регистрировали у каждого животного в гиппокампе при наличии возможности и регистрировали их как отдельные эксперименты.
Кратковременная обработка посредством h9E4 индуцировала значительное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (Tg-snca + h9E4 по сравнению с Tg-snca + PBS, р=0,002, фиг. 14). Тем не менее, обращение при обработке h9E4 было лишь частичным, о чем свидетельствовало значительное различие с базальной синаптической передачей у однопометников, обработанных PBS (р=0,007).
Кратковременная обработка посредством GM37 индуцировала значительное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (Tg-snca + GM37 по сравнению с Tg-snca + PBS, р=0,004, фиг. 15). Базальная синаптическая передача у обработанных посредством GM37 трансгенных мышей значительно не отличалась от базальной синаптической передачи у обработанных посредством PBS однопометников, что свидетельствовало о полном обращении нарушения (фиг. 15).
GM285 также индуцировало значительное обращение нарушения базальной синаптической передачи у трансгенных мышей F28-snca (фиг. 16). Базальная синаптическая передача у обработанных посредством GM285 трансгенных мышей значительно не отличалась от базальной синаптической передачи у обработанных посредством PBS однопометников, что свидетельствовало о полном обращении нарушения.
Пример 8. Микродиализ для оценки альфа-синуклеина человека в головном мозге бодрствующих свободно двигающихся животных
Для оценки уровней альфа-синуклеина человека в интерстициальной жидкости головного мозга (ISF) использовали двухтактный способ микродиализа. Мышей по одной размещали в условиях с контролируемой температурой (22±1,5°C) и влажностью (55-65%) и содержали при цикле чередования света и темноты 12:12 часов (свет включали в 06:00 ч). Пища и вода были доступны без ограничений. Настоящее исследование проводили на гиппокампе трансгенных мышей F28-snca (возрастом 50-54 недели). Для осуществления микродиализа в гиппокампе мышей анестезировали изофлураном и в головном мозг стереотаксически имплантировали внутрицеребральную направляющую канюлю (СМА), располагая микродиализный зонд в гиппокампе (координаты наконечника зонда: на 3,1 мм кзади и на 2,8 мм латеральнее от брегмы и на 1,3 мм относительно твердой мозговой оболочки) в соответствии с атласом Paxinos и Franklin 2001. Для фиксации направляющих канюль применяли анкерные винты и акриловый цемент. После имплантации канюли обеспечивали восстановление мышей после операции в течение 2-3 дней перед диализом.
В день эксперимента через направляющую канюлю вводили 2-миллиметровый СМА-зонд с пределом обнаружения 1000 кДа. Зонд подсоединяли к перистальтическому насосу для микродиализа с двумя каналами (МАВ20, Microbiotech) и запускали в двухтактном режиме. Впускную трубку микродиализного зонда соединяли с перистальтическим насосом, перфузирующим в зонд искусственную спинномозговую жидкость (aCSF, в мМ: 147 NaCl, 2,7 KCl, 1,2 CaCl2, 0,85 MgCl2). Перистальтический насос также подсоединяли к выпускной трубке для предотвращения потери перфузионной жидкости из зонда при выталкивании жидкости через трубку. В качестве перфузионного буфера 25% фракцию бычьего альбумина V (Sigma) разводили до 0,2% искусственной CSF в день применения и фильтровали через 0,1-мкм мембрану. Фактическую скорость потока у насоса определяли без присоединенного зонда. Пробирки для образцов взвешивали до и после забора образцов в течение заданного периода времени и рассчитывали скорость потока. Затем насос устанавливали на постоянный поток 1 мкл/мин. На протяжении всего периода эксперимента использовали 120-минутный режим забора образцов. Во избежание препятствия в виде повреждения ткани промежуток проведения эксперимента устанавливали на 14-48 ч. после имплантации зонда. Через 14-16 ч после начала экспериментов GM37, 9Е4 человека или изотопический контроль (антитело к HEL) вводили i.p. при 15 мг/кг и собирали дополнительные 6 образцов (12 ч сбора). Диализаты хранили при -80°C. Концентрацию альфа-синуклеина человека определяли с помощью ELISA (набора для ELISA Covance).
Среднее значение двух-трех исходных значений (4 ч.-6 ч.) до обработки антителом принимали в качестве исходного уровня и принимали за 100% для каждого животного. Данные оценивали с помощью двухфакторного дисперсионного анализа (ANOVA) с повторными измерениями для оценки статистической значимости. Исходные уровни альфа-синуклеина человека в гиппокампе составляли 8,1±1,1 нг/мл (среднее ± SEM, n=25, не скорректированные по восстановлению диализного зонда in vitro). Введение GM37 индуцировало большее уменьшение уровня альфа-синуклеина человека в гиппокампе мышей F28 по сравнению как с антителом сравнения, 9Е4 человека, так и с изотипическим контролем (антителом к HEL). (Фиг. 17).
Пример 9. Длительные эффекты антител к альфа-синуклеину in vivo. Антитело GM37 улучшает двигательный фенотип в крысиной модели болезни Паркинсона.
Целенаправленной сверхэкспрессии альфа-синуклеина человека в дофаминергических нейронах в среднем мозге крысы можно достичь с помощью вектора на основе рекомбинантного аденоассоциированного вируса (rAAV), и она ассоциирована с прогрессирующей утратой дофаминергических клеток в черном веществе, а также с двигательными нарушениями.
Взрослых самок крыс линии Sprague-Dawley (225-250 г) использовали для экспрессии альфа-синуклеина человека в черном веществе (SN) путем введения им аденоассоциированного вируса серотипа AAV2/5, содержащего промотор бета-актина курицы с энхансерными элементами из промотора цитомегаловируса, после которого идет кДНК альфа-синуклеина человека и WPRE-элемент, как описано ранее (Xu L, Daly Т, Gao С, Flotte TR, Song S, Byrne BJ, Sands MS, Ponder KP (2001). В этой модели было показано, что экспрессия альфа-синуклеина человека приводит к нейродегенерации дофаминергических нейронов. Maingay М, et al. CNS Spectr. 2005 Mar; 10(3):235-44). Для проверки влияния терапевтического антитела к альфа-синуклеину в этой модели обработку антителом начинали за 2-4 дня до введений вируса и продолжали до конца исследования (фиг. 18). Введение PBS в том же объеме (5 мл/кг: IP) использовали в качестве контроля. GM37 вводили дважды в неделю дозой 15 мг/кг (IP). Вирусные частицы (rAAV2/5), содержащие ген альфа-синуклеина человека или зеленый флуоресцентный белок (GFP), вводили односторонней инъекцией в SN. Животных анестезировали комбинацией Hypnorm® и Dormicum® в количестве 2,0 мл/кг s.c. и помещали в стереотаксическую рамку. Их температуру регулировали до 37,5°C с помощью нагревательной подушки и обнажали череп. Просверливали отверстие над правым SN по следующим координатам, в соответствии с атласом Paxinos и Watson (Paxinos & Watson, 1998): 5,5 мм кзади и 2,0 мм латеральнее от брегмы. Однократную инъекцию 3 мкл rAAV2/5-alpha-syn или rAAV2/5-GFP производили на глубине 7,2 мм ниже твердой мозговой оболочки и при скорости потока 0,2 мкл/мин. с помощью шприца Гамильтона, соединенного со стереотаксическим инжектором. Игла оставалась на месте на дополнительные 5 минут для обеспечения диффундирования вектора в SN. После операции животных возвращали в их собственную клетку и помещали в нагретую среду, где обеспечивали их восстановление после анестезии. Оценку двигательной асимметрии в тесте передних конечностей на асимметричность проводили до инъекций AAV, а также через 3, 7 и 10 недель после инъекций AAV. Представленные данные соответствовали соотношению между использованием правой передней лапы по сравнению с общим использованием левой и правой передних лап. Результативность каждого животного в тесте передних конечностей на асимметричность регистрировали на камеру в течение итого 5 минут, и была произведена ручная оценка количества касаний с использованием левой и правой передних лап в течение 5 минут в заключительный день тестирования через 10 недель после введения вируса. На 10-й неделе значительное нарушение присутствовало у AAV-syn по сравнению с крысами, которым вводили AAV-GFP (р=0,012). Показана тенденция к обращению у животных, обрабатываемых GM37, поскольку их характеристики отличались от GFP-крыс (соответственно р=0,163 и р=0,407 для gm37). Эти результаты свидетельствовали, что антитело GM37 могло улучшать паркинсонический двигательный фенотип в этой крысиной модели (фиг. 18 и 19).
Пример 10. Длительные эффекты антител к альфа-синуклеину in vivo. Антитело GM37 подавляет затравочное действие в отношении агрегирования и фосфорилирования эндогенного альфа-синуклеина мыши
Введение предварительно образованных фибрилл альфа-синуклеина, полученных из рекомбинантного белка, в заднюю часть полосатого тела мышей дикого типа мобилизировала эндогенный альфа-синуклеин мыши и индуцировала образование фосфорилированных агрегатов Ser-129 внутри нейронов в коре, миндалине и черном веществе (Luk et al. 2012, Science. 2012 Nov 16;338(6109): 949-53). Для проверки того, что специфическое в отношении альфа-синуклеина моноклональное антитело GM37 может уменьшать начало образования индуцированных фибриллами альфа-синуклеина включений фосфорилированного альфа-синуклеина in vivo, было использовано в общей сложности 45 мышей. Мышам вводили GM 37 при 30 мг/кг i.p, GM 37 при 15 мг/кг i.v. или основу ip (PBS). Спустя один день мышей анестезировали и стереотаксическим методом вводили в одно полушарие 2 мкл неочищенных затравочных частиц рекомбинантного альфа-синуклеина человека, полученных, как описано ранее (пример 6) (итого 2 мкг неочищенных затравочных частиц на животное). Для введения неочищенных затравочных частиц вскрывали череп, просверливая отверстие, и вставлялиодну стеклянную пипетку (координаты: +0,5 мм кпереди от брегмы, +2,0 мм латеральнее от срединной линии) в правый передний мозг для целенаправленного введения инокулята в заднюю часть полосатого тела (+2,6 мм под твердой мозговой оболочкой). После восстановления мыши получали еженедельные i.p. или i.v. инъекции антител до умерщвления через 45 дней. Группам из 15 мышей/группа раз в неделю вводили либо iv GM37 15 мг/кг, либо ip GM37 30 мг/кг, либо PBS (10 мл/кг) ip.
Для измерения концентрации антител в плазме крови один раз в неделю брали кровь из щек непосредственно перед следующей инъекцией, т.е. через 7 дней после последней инъекции. Плазму крови получали путем центрифугирования при 2000g, инкубирования в течение 15 мин. при RT, после этого супернатант замораживали при -20°C. В конце исследования забирали образец CSF и замораживали его при -20°C. Образцы плазмы крови и CSF анализировали для определения концентрации IgG человека с помощью MSD. Вкратце, антитело мыши к IgG человека (клон МН16-1 (M1268) использовали для захвата, плазму крови или CSF инкубировали в лунке с последующим добавлением антитела козы с сульфо-меткой к антителам человека в качестве антитела выявления (MSD № по кат. R32AJ-1). Планшеты анализировали в отношении электрохемилюминесценции с помощью MSD.
Уровни антител в плазме крови показаны на фигуре 20В, и из них видно дозозависимое увеличение концентрации антител в плазме крови и накопление антител в плазме в течение шести недель. Уровни антител в csf показаны на фигуре 20С, и из них видно, что около 0,1% уровня антител в плазме крови можно измерить в csf.
На 45-й день с момента инъекции затравочных частиц в виде фибрилл альфа-синуклеина мышей анестезировали, перфузировали транскардиально PBS с последующей перфузией забуференным до нейтрального параформальдегидом (4%). Удаляли головной мозг и инкубировали его в течение ночи для последующей фиксации в забуференном до нейтрального параформальдегиде. Иммуногистохимию проводили на последовательных срезах толщиной 45 мкм в Neuroscience associates. Вкратце, с помощью технологии MultiBrain® до 25 штук головного мозга мышей совместно заливали в один блок, итого 3 блока, замораживали срезы толщиной 45 мкм во фронтальной плоскости и собирали в чашки, содержащие антиген-консервирующий раствор. Каждый шестой срез окрашивали антителом кфосфорилированному альфа-синуклеину Ser-129 (антителом к альфа-синуклеину (фосфо-8129) [Psyn/81A]ab184674) для выявления реакционных структур фосфорилированного альфа-синуклеина Ser129.
Количественное определение патологии pSyn осуществляли путем ручного подсчета положительных по иммунореактивности клеток на изображениях с 10-кратным увеличением 5-7 срезов, охватывающих все черное вещество от каждого шестого среза. Подсчет осуществляли слепым методом. Количество клеток в миндалине и черном веществе анализировали с помощью однофакторного ANOVA с последующим анализом с помощью t-критерия Бонферрони, где эффект антитела GM37 сравнивали с обработкой PBS.
Как можно видеть из фиг. 20С, обработка антителом GM37 значимо уменьшала количество внутриклеточных включений в черном веществе по сравнению с контролем PBS при либо ip, либо iv обработке. Из данных видно, что антитело GM37 может оказывать терапевтический эффект при PD путем блокировки проникновения внеклеточного патологического альфа-синуклеина в нейроны, блокировки его распространение между нейронами и/или облегчения его устранения из ISF в результате поглощения в микроглию. Поскольку такое появление включений было связано с утратой дофаминергических нейронов и развитием паркинсонических двигательных нарушений в животных моделях, обработка антителом GM37 может оказать терапевтический эффект на утрату дофаминергических клеток и развитие двигательных нарушений при PD.
Пример 11. Возможность изготовления GM37 и вариантов GM37
Антитела к альфа-синуклеину получали в культуре клеток млекопитающих в условиях, которые имитируют условия получения, которые будут использованы для получения клинического материала для применения на пациентах. Хорошо известно, что белки, полученные таким образом, подвергаются посттрансляционным модификациям, которые могут влиять как на терапевтическую эффективность антитела, так и на биофизические характеристики, которые влияют на стабильность антитела с течением времени. Эмпирические знания, полученные в течение десятилетий исследований, позволили выявить набор посттрансляционных модификаций, которые известны как придающие риск возможности разработки конкретной молекулы. Было показано, что эти посттрансляционные модификации коррелируют с аминокислотными цепями, присутствующими в первичной последовательности белков тяжелых и легких цепей. Были созданы алгоритмы, которые могут выявить эти последовательности и определить потенциальный риск, который они будут оказывать на возможность изготовления и возможность разработки терапевтического антитела.
In silico анализ первичной последовательности антитела можно применять для снижения риска для молекулы в отношении возможности ее разработки в качестве терапевтического средства. В частности, подробный анализ VH и VL-областей может выявить уникальные аминокислоты, которые считаются важными для активности молекул, но также могут нести потенциальный риск для их стабильности с течением времени. Специфическое дезамидирование последовательностей было идентифицировано как потенциальный риск для белковых структур. Дезамидирование белков может происходить на амидных боковых цепях глутаминов или аспарагиновых остатков и превращать их в карбоксилатную группу (Lorenzo et al. PLOSone, DOI:10.1371, Dec. (2015)). Неферментативное дезамидирование при нейтральном рН происходит быстрее для аспарагина, и поэтому его считают более высоким риском, нежели глутамин. На активность дополнительно оказывает влияние последующая аминокислота в последовательности, и такое изменение активности может происходить со скоростью, которая исчисляется днями или годами. Фактическую судьбу белка, который подвергается дезамидированию, необходимо оценивать экспериментально для определения влияния такого изменения как на его стабильность, так и на его активность.
Авторами настоящего изобретения был идентифицирован сайт для дезамидирования в VH-домене GM37. Аминокислотным остатком 54 является аспарагин (N), после которого следует глицин (G) в положении 55. N54 подвержен высокому риску спонтанного дезамидирования. Для снижения такого риска авторами настоящего изобретения был создан набор из 3 вариантов, в которых аспарагин (N) заменен серином (S), глутамином (Q) или гистидином (Н). Все 3 варианта были получены в культуре клеток млекопитающих с помощью способов временной трансфекции (пример 1.5). Все 3 варианта демонстрировали схожие свойства экспрессии и очистки, как и GM37wt. (Фиг. 23).
Для каждого из восьми продуктов объемом 400 мл осуществляли временные трансфекции с применением клеток CHOK1SV GS-KO, которые до этого находились в культуре в течение минимум 2 недель. Клетки пересевали за 24 часа до трансфекции. Все трансфекции проводили посредством электропорации с помощью Gene Pulse XCell (Bio-Rad). Для каждой трансфекции жизнеспособные клетки ресуспендировали в предварительно нагретой среде CD-CHO, дополненной 6 мМ L-глутамина, до 2,86×107 клеток/мл. 40 мкг ДНК с каждой установленной SGV, содержащей соответствующие тяжелые и легкие цепи, вносили аликвотой в каждую кювету (кювета Bio-Rad, GenePulser, зазор 0,4 см, 165-2088) и добавляли 700 мкл клеточной суспензии. Клетки подвергали электропорации с напряжением 300 В, 900 мкФ. Подвергнутые трансфекции клетки переносили в повторно нагретую среду в колбах Эрленмейера и также переносили в колбы содержимое кювет, дважды промытых предварительно нагретой средой. Культуры трансфектантов инкубировали во встряхивающем термостате при 36,5°C, 5% CO2, 85% влажности, 140 об/мин в течение 6 дней. Жизнеспособность клеток измеряли во время сбора с помощью автоматизированного устройства для подсчета клеток Cedex HiRes (Rosche).
Для оценки важности остатка 54 в связывании с альфа-синуклеином человека авторами настоящего изобретения была проанализирована способность вариантов связываться в двух различных экспериментах. С помощью конкурентного формата ELISA авторами настоящего изобретения было оценено, будет ли оказывать влияние изменение по остатку 54 на способность GM37 связывать альфа-синуклеин в растворе. Путем оценки концентрации синуклеина, которая может подавлять связывание антитела с синуклеином, нанесенным на планшеты для ELISA, авторами настоящего изобретения было показано, что все три варианта сохраняли такое же связывание, что и GM37wt, и связывались с альфа-синуклеином с высокой аффинностью, что в результате давало IC50 1-2 нМ (фиг. 24). Конкурентный анализ проводили с помощью предварительной инкубации фиксированной концентрации (0,3 мкг/мл) каждого из следующих антител: GM37 (называемого GM 37wt), варианта 1 GM37, варианта 2 GM37 и варианта 3 GM37 с диапазоном 0-1000 нМ альфа-синуклеина человека в течение 60 минут при комнатной температуре. Оставшееся несвязанное антитело захватывали и измеряли на планшетах для ELISA, покрытых 100 нг/мл рекомбинантного альфа-синуклеина человека, с применением антитела для обнаружения антител человека путем электрохемилюминесценции (MSD, Гейтерсберг, Мэриленд). Значения IC50 взаимодействия составляли 1,9 нМ, 1,6 нМ, 2,1 нМ и 1,4 нМ для соответственно GM37 wt, варианта 1 GM37, варианта 2 GM37 и варианта 3 GM37 (по результатам определения с помощью Prism Graphpad®).
С помощью поверхностного плазмонного резонанса (SPR) авторами настоящего изобретения была оценена кинетика связывания в режиме реального времени GM37 wt (2 партии) и трех вариантов (пример 2). Альфа-синуклеин человека захватывали на микроскопический препарат (лиганд) и каждое из антител тестировали в нескольких концентрациях в качестве аналитов. Анализ кривых связывания в присутствии антитела в нескольких концентрациях показал, что скорости ассоциации были одинаковыми для всех четырех антител, аналогично, при удалении антитела из буфера, измеренные скорости диссоциации не демонстрировали статистической разницы между антителами. При использовании алгоритма связывания 1:1 все 4 антитела имели почти идентичные константы связывания (фиг. 25).
Для оценки влияние изменений по N54 на функциональную активность GM37 авторами настоящего изобретения была проанализирована способность антител блокировать затравочную активность синуклеина в культуре первичных нейронов (пример 6). Уровень затравочного действия измеряли с помощью антитела, специфичного в отношении фосфо-синуклеина. Все 3 антитела были способны блокировать затравочное действие, по результатам измерения с помощью сигнала от фосфо-синуклеина (фиг. 26). Более того, уровень подавления был одинаковым для всех 4 антител. Данные на основе такого сигнала дополнительно подтверждают данные по связыванию, что аминокислота 54 в VH-домене не является необходимой для аффинности связывания с альфа-синуклеином человека или для ингибирования затравочного действия в анализе на первичных клетках. Более того, авторами настоящего изобретения было обнаружено, что все три этих антитела можно было получить с помощью стандартных способов экспрессии и очистки. Что интересно, для одного из вариантов N54Q наблюдали увеличение выхода при получении по сравнению с другими вариантами, что имеет большое значение, когда антитело необходимо будет получать в больших промышленных масштабах. Эти данные подтверждают возможность снижения потенциального риска дезамидирования путем замены аспарагина (N) на другую аминокислоту без проблемы утраты активности.
Образцы каждого из антител wt GM37, var1, var2 и var3 подвергали постоянному повышению температуры с течением времени и одновременно измеряли уровень агрегирования по многоугловому рассеянию света (Prometheus NT.48, NanoTemper Technologies). Было обнаружено, что температура для начала агрегирования была схожей для GM37 и вариантов GM37, однако наиболее низкий уровень агрегирования наблюдали для GM37-Var2 (фиг. 27).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> H. Lundbeck A/S
<120> New Monoclonal Anti-Alpha-Synuclein Antibodies
<130> 0992
<160> 43
<170> PatentIn version 3.5
<210> 1
<211> 10
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR 1 Heavy Chain
<400> 1
Gly Phe Thr Phe Ser Ser Tyr Ala Met Thr
1 5 10
<210> 2
<211> 17
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR2 Heavy Chain
<400> 2
Ala Ile Arg Ser Asn Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 3
<211> 9
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR3 Heavy Chain
<400> 3
Ala Lys Asn Trp Ala Pro Phe Asp Ser
1 5
<210> 4
<211> 11
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR1 Light Chain
<400> 4
Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR 2 Light Chain
<400> 5
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 6
<211> 9
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR 3 Light Chain
<400> 6
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 7
<211> 116
<212> PRT
<213> Artificial
<220>
<223> GM37 CDR Heavy Chain
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Thr Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Arg Ser Asn Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Trp Ala Pro Phe Asp Ser Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 8
<211> 108
<212> PRT
<213> Artificial
<220>
<223> GM 37 Light Chain
<400> 8
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 9
<211> 6
<212> PRT
<213> Artificial
<220>
<223> Epitope 112-117
<400> 9
Ile Leu Glu Asp Met Pro
1 5
<210> 10
<211> 140
<212> PRT
<213> Artificial
<220>
<223> Alpha-synuclein
<400> 10
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 11
<211> 165
<212> PRT
<213> Artificial
<220>
<223> A-Syn-AAKK-BAP
<400> 11
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Asn Ile Ala Ala Ala Thr Gly Leu Val Lys
85 90 95
Lys Asp Gln Leu Ala Lys Gln Asn Glu Glu Gly Phe Leu Gln Glu Gly
100 105 110
Met Val Asn Asn Thr Asp Ile Pro Val Asp Pro Glu Asn Glu Ala Tyr
115 120 125
Glu Met Pro Pro Glu Glu Glu Tyr Gln Asp Tyr Glu Pro Glu Ala Gly
130 135 140
Ser Ala Gly Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
145 150 155 160
Ile Glu Trp His Glu
165
<210> 12
<211> 165
<212> PRT
<213> Artificial
<220>
<223> A-Syn-BAAK-BAP
<400> 12
Met Asp Val Phe Met Lys Gly Leu Ser Met Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Ala Lys Gln Asn Glu Glu Gly Phe Leu Gln Glu Gly
100 105 110
Met Val Asn Asn Thr Asp Ile Pro Val Asp Pro Glu Asn Glu Ala Tyr
115 120 125
Glu Met Pro Pro Glu Glu Glu Tyr Gln Asp Tyr Glu Pro Glu Ala Gly
130 135 140
Ser Ala Gly Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
145 150 155 160
Ile Glu Trp His Glu
165
<210> 13
<211> 162
<212> PRT
<213> Artificial
<220>
<223> A-Syn-BBAA-BAP
<400> 13
Met Asp Val Phe Met Lys Gly Leu Ser Met Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Arg Glu Gly Val
35 40 45
Val Gln Gly Val Ala Ser Val Ala Glu Lys Thr Lys Glu Gln Ala Ser
50 55 60
His Leu Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala Gly Ser Ala Gly
130 135 140
Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp
145 150 155 160
His Glu
<210> 14
<211> 165
<212> PRT
<213> Artificial
<220>
<223> A-Syn-BBKK-BAP
<400> 14
Met Asp Val Phe Met Lys Gly Leu Ser Met Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Arg Glu Gly Val
35 40 45
Val Gln Gly Val Ala Ser Val Ala Glu Lys Thr Lys Glu Gln Ala Ser
50 55 60
His Leu Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Asn Ile Ala Ala Ala Thr Gly Leu Val Lys
85 90 95
Lys Asp Gln Leu Ala Lys Gln Asn Glu Glu Gly Phe Leu Gln Glu Gly
100 105 110
Met Val Asn Asn Thr Asp Ile Pro Val Asp Pro Glu Asn Glu Ala Tyr
115 120 125
Glu Met Pro Pro Glu Glu Glu Tyr Gln Asp Tyr Glu Pro Glu Ala Gly
130 135 140
Ser Ala Gly Gly Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
145 150 155 160
Ile Glu Trp His Glu
165
<210> 15
<211> 141
<212> PRT
<213> Artificial
<220>
<223> A-Syn-120-140_Del-BAP
<400> 15
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Gly Ser Ala Gly Gly Ser Gly Gly Leu
115 120 125
Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
130 135 140
<210> 16
<211> 131
<212> PRT
<213> Artificial
<220>
<223> alpha-synuclein amino acids 1-119
<400> 16
Met Ala His His His His His His Ile Glu Gly Arg Met Asp Val Phe
1 5 10 15
Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val Ala Ala Ala Glu
20 25 30
Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys Thr Lys Glu Gly
35 40 45
Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val Val His Gly Val
50 55 60
Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr Asn Val Gly Gly
65 70 75 80
Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys Thr Val Glu Gly
85 90 95
Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys Lys Asp Gln Leu
100 105 110
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
115 120 125
Pro Val Asp
130
<210> 17
<211> 106
<212> PRT
<213> artificial
<220>
<223> kappa (LC constant region)
<400> 17
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 18
<211> 329
<212> PRT
<213> Artificial
<220>
<223> IgG1 (HC Constant region)
<400> 18
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 19
<211> 4
<212> PRT
<213> Artificial
<220>
<223> GM285 epitope 112-115
<400> 19
Ile Leu Glu Asp
1
<210> 20
<211> 13
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR1 Heavy Chain
<400> 20
Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe Thr Met Thr
1 5 10
<210> 21
<211> 17
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR2 Heavy Chain
<400> 21
Ala Ile Ser Gly Ser Gly Gly Gly Thr Ser Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 22
<211> 9
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR3 Heavy Chain
<400> 22
Ala Lys Asn Trp Ala Pro Phe Asp Tyr
1 5
<210> 23
<211> 12
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR1 Light Chain
<400> 23
Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr Leu Ala
1 5 10
<210> 24
<211> 7
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR2 Light Chain
<400> 24
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 25
<211> 9
<212> PRT
<213> Artificial
<220>
<223> GM285 CDR3 Light Chain
<400> 25
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 26
<211> 116
<212> PRT
<213> Artificial
<220>
<223> GM285 VH
<400> 26
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Thr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Trp Ala Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 27
<211> 108
<212> PRT
<213> Artificial
<220>
<223> GM285 VL
<400> 27
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Val Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 28
<211> 329
<212> PRT
<213> Artificial
<220>
<223> GM285 IgG1 constant region
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 29
<211> 106
<212> PRT
<213> Artificial
<220>
<223> GM285 Kappa chain
<400> 29
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 30
<211> 116
<212> PRT
<213> Artificial
<220>
<223> GM37 Variant 1 heavy chain
<400> 30
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Thr Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Arg Ser Ser Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Trp Ala Pro Phe Asp Ser Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 31
<211> 116
<212> PRT
<213> Artificial
<220>
<223> GM 37 variant 2 heavy chain
<400> 31
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Thr Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Arg Ser Gln Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Trp Ala Pro Phe Asp Ser Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 32
<211> 116
<212> PRT
<213> Artificial
<220>
<223> GM 37 variant 3 heavy chain
<400> 32
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Thr Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Arg Ser His Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Trp Ala Pro Phe Asp Ser Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 33
<211> 17
<212> PRT
<213> Artificial
<220>
<223> GM37 variant 1 heavy chain CDR 2
<400> 33
Ala Ile Arg Ser Ser Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 34
<211> 17
<212> PRT
<213> Artificial
<220>
<223> GM37 variant 2 CDR 2 heavy chain
<400> 34
Ala Ile Arg Ser Gln Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 35
<211> 17
<212> PRT
<213> Artificial
<220>
<223> GM37 variant 3 CDR 2 heavy chain
<400> 35
Ala Ile Arg Ser His Gly Asp Arg Thr Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 36
<211> 5
<212> PRT
<213> Artificial
<220>
<223> 9E4 binding epitope
<400> 36
Asn Glu Ala Tyr Glu
1 5
<210> 37
<211> 134
<212> PRT
<213> Artificial
<220>
<223> HUMAN Beta-synuclein
<400> 37
Met Asp Val Phe Met Lys Gly Leu Ser Met Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Arg Glu Gly Val
35 40 45
Val Gln Gly Val Ala Ser Val Ala Glu Lys Thr Lys Glu Gln Ala Ser
50 55 60
His Leu Gly Gly Ala Val Phe Ser Gly Ala Gly Asn Ile Ala Ala Ala
65 70 75 80
Thr Gly Leu Val Lys Arg Glu Glu Phe Pro Thr Asp Leu Lys Pro Glu
85 90 95
Glu Val Ala Gln Glu Ala Ala Glu Glu Pro Leu Ile Glu Pro Leu Met
100 105 110
Glu Pro Glu Gly Glu Ser Tyr Glu Asp Pro Pro Gln Glu Glu Tyr Gln
115 120 125
Glu Tyr Glu Pro Glu Ala
130
<210> 38
<211> 127
<212> PRT
<213> Artificial
<220>
<223> HUMAN Gamma-synuclein
<400> 38
Met Asp Val Phe Lys Lys Gly Phe Ser Ile Ala Lys Glu Gly Val Val
1 5 10 15
Gly Ala Val Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Met Tyr Val Gly Ala Lys Thr Lys Glu Asn Val
35 40 45
Val Gln Ser Val Thr Ser Val Ala Glu Lys Thr Lys Glu Gln Ala Asn
50 55 60
Ala Val Ser Glu Ala Val Val Ser Ser Val Asn Thr Val Ala Thr Lys
65 70 75 80
Thr Val Glu Glu Ala Glu Asn Ile Ala Val Thr Ser Gly Val Val Arg
85 90 95
Lys Glu Asp Leu Arg Pro Ser Ala Pro Gln Gln Glu Gly Glu Ala Ser
100 105 110
Lys Glu Lys Glu Glu Val Ala Glu Glu Ala Gln Ser Gly Gly Asp
115 120 125
<210> 39
<211> 140
<212> PRT
<213> Artificial
<220>
<223> alpha-synuclein ortholog for Cynomolgus monkey
<400> 39
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Ile Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Gln Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 40
<211> 140
<212> PRT
<213> Artificial
<220>
<223> alpha-synuclein ortholog for Rat
<400> 40
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Thr Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Asn Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Met Gly Lys Gly Glu Glu Gly Tyr Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Ser Ser Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 41
<211> 140
<212> PRT
<213> Artificial
<220>
<223> alpha-synuclein ortholog for Mouse
<400> 41
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Thr Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Asn Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Met Gly Lys Gly Glu Glu Gly Tyr Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Gly Ser Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 42
<211> 446
<212> PRT
<213> artificial
<220>
<223> 9E4 HC
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 43
<211> 220
<212> PRT
<213> artificial
<220>
<223> 9E4 LC
<400> 43
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ АЛЬФА-СИНУКЛЕИНА | 2018 |
|
RU2787039C2 |
| АНТИТЕЛА | 2018 |
|
RU2798399C2 |
| Моноклональные антитела к альфа-синуклеину для предотвращения агрегации тау-белка | 2017 |
|
RU2760334C2 |
| КОМБИНАЦИИ АНТИТЕЛ К STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2804815C2 |
| АНТИТЕЛО, НАПРАВЛЕННОЕ ПРОТИВ ФАКТОРА СЛИПАНИЯ А (ClfA) S. aureus | 2019 |
|
RU2818805C2 |
| АНТИТЕЛО К АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2807060C1 |
| АНТИТЕЛА В7-Н4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2809243C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, КОТОРЫЕ РАСПОЗНАЮТ АЛЬФА-СИНУКЛЕИН | 2012 |
|
RU2743738C2 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| АНТИТЕЛО К PD-L1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2778085C2 |
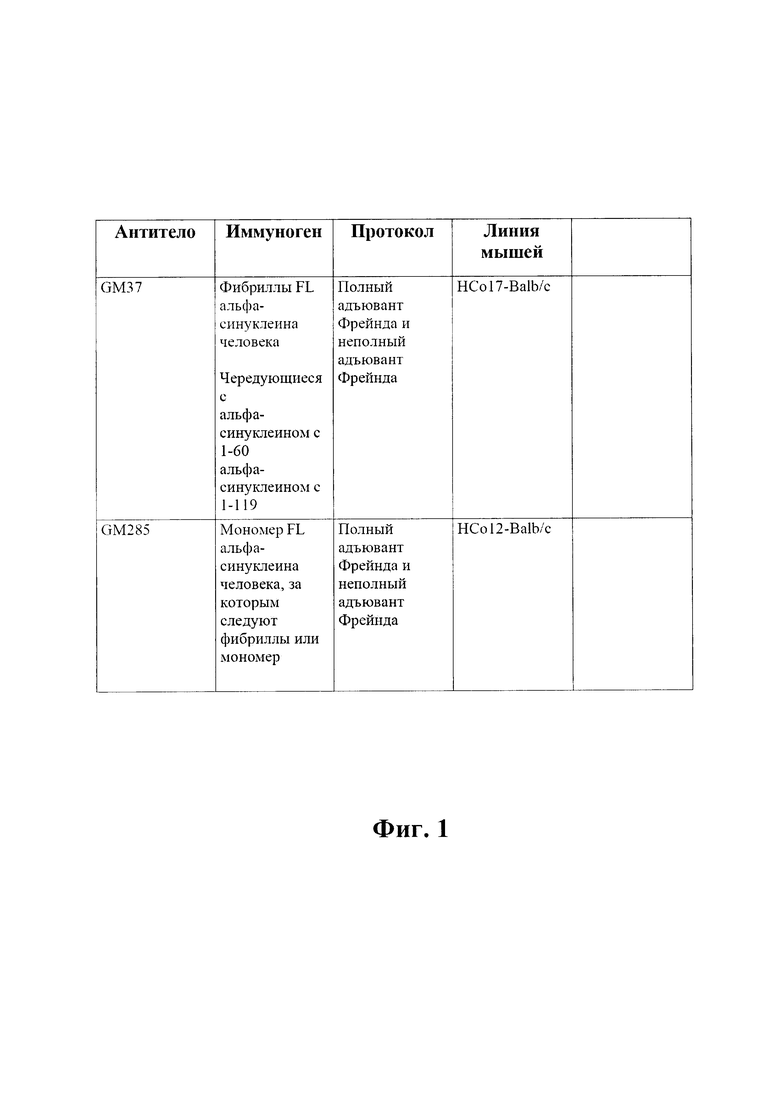
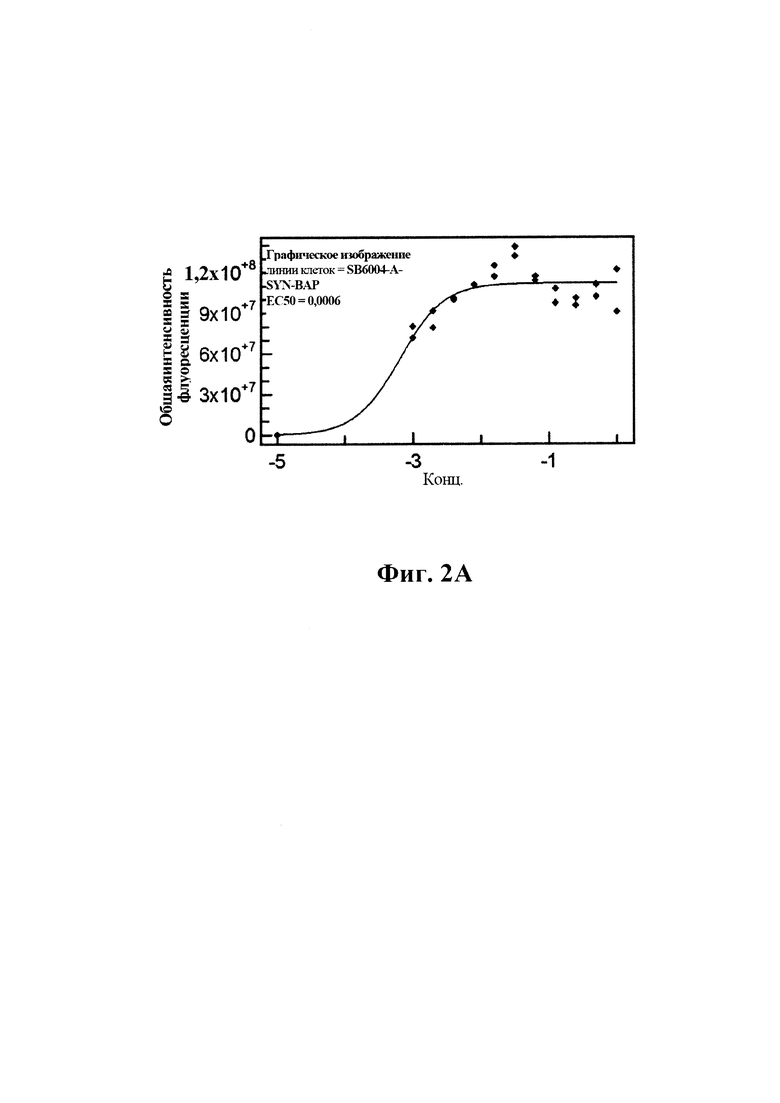
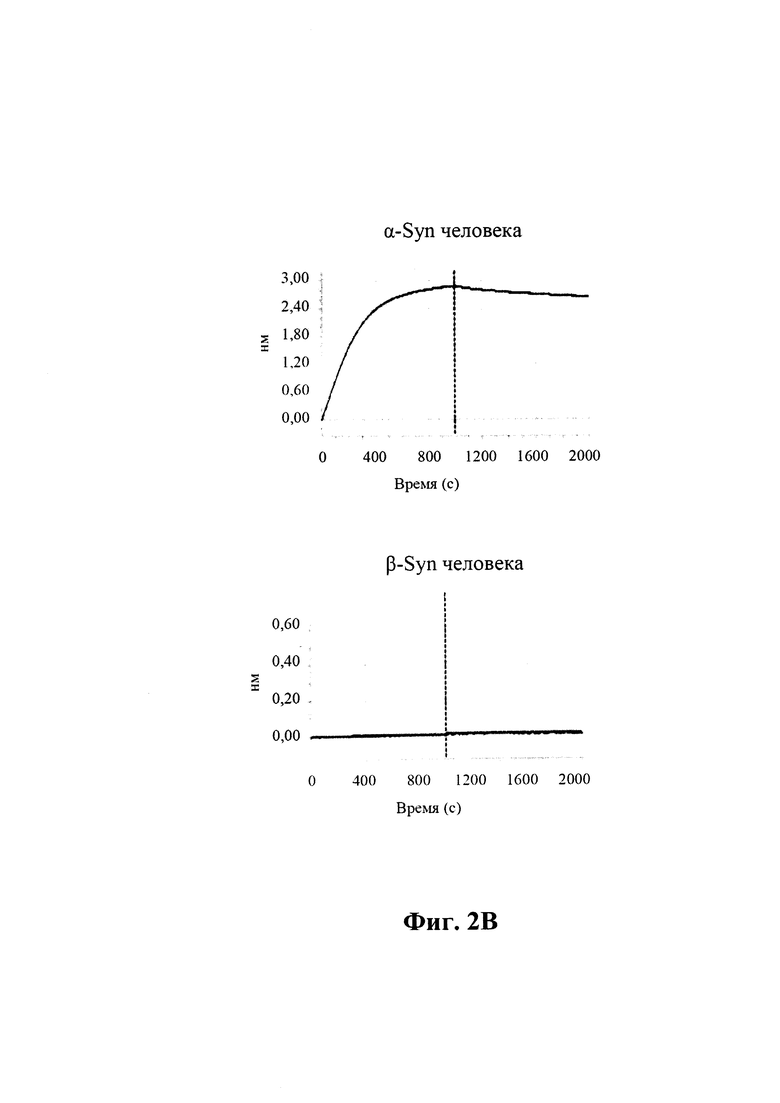
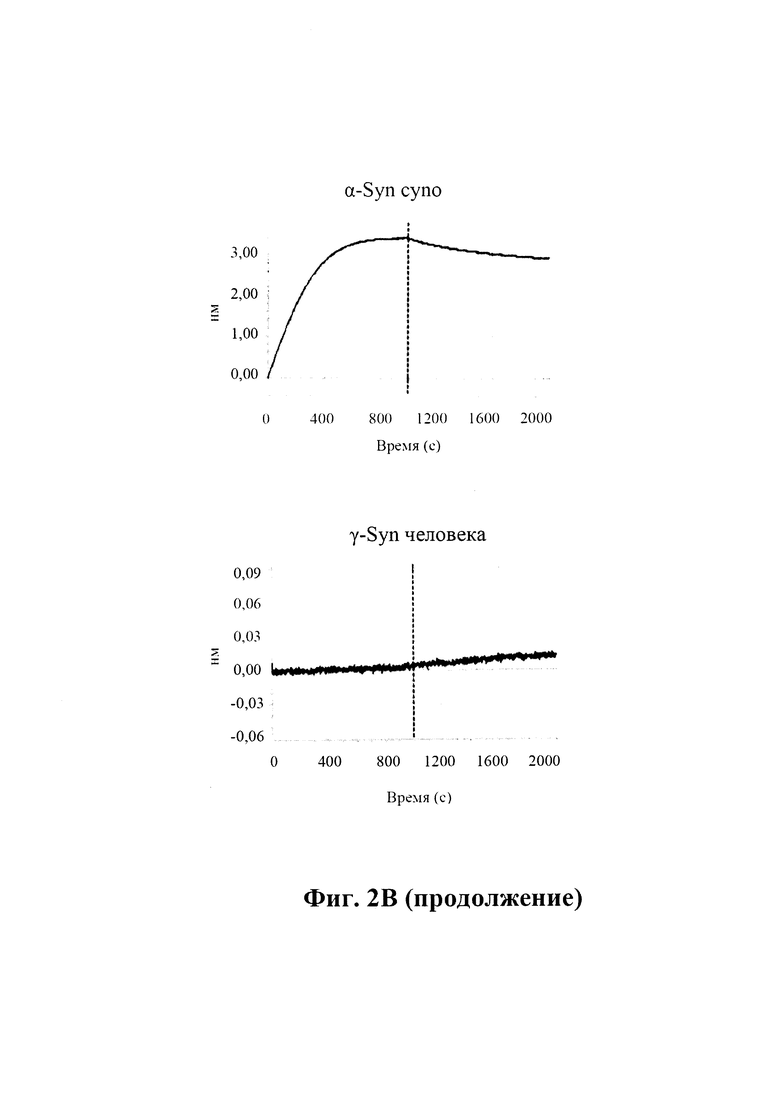
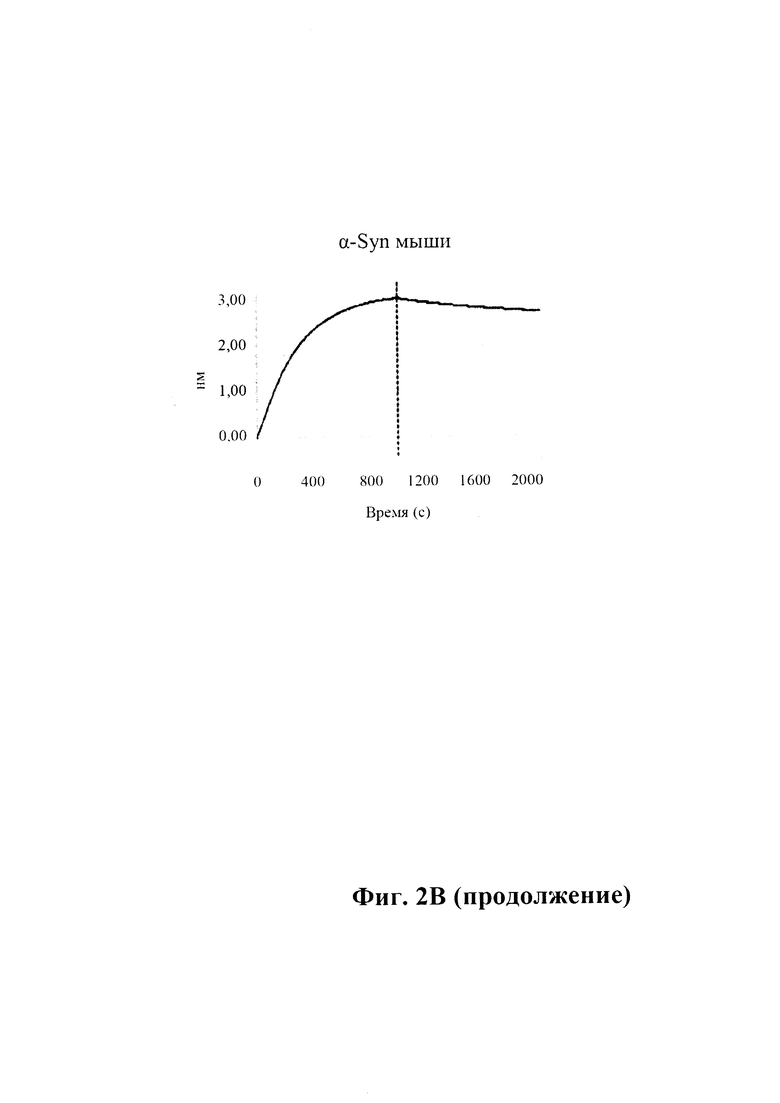
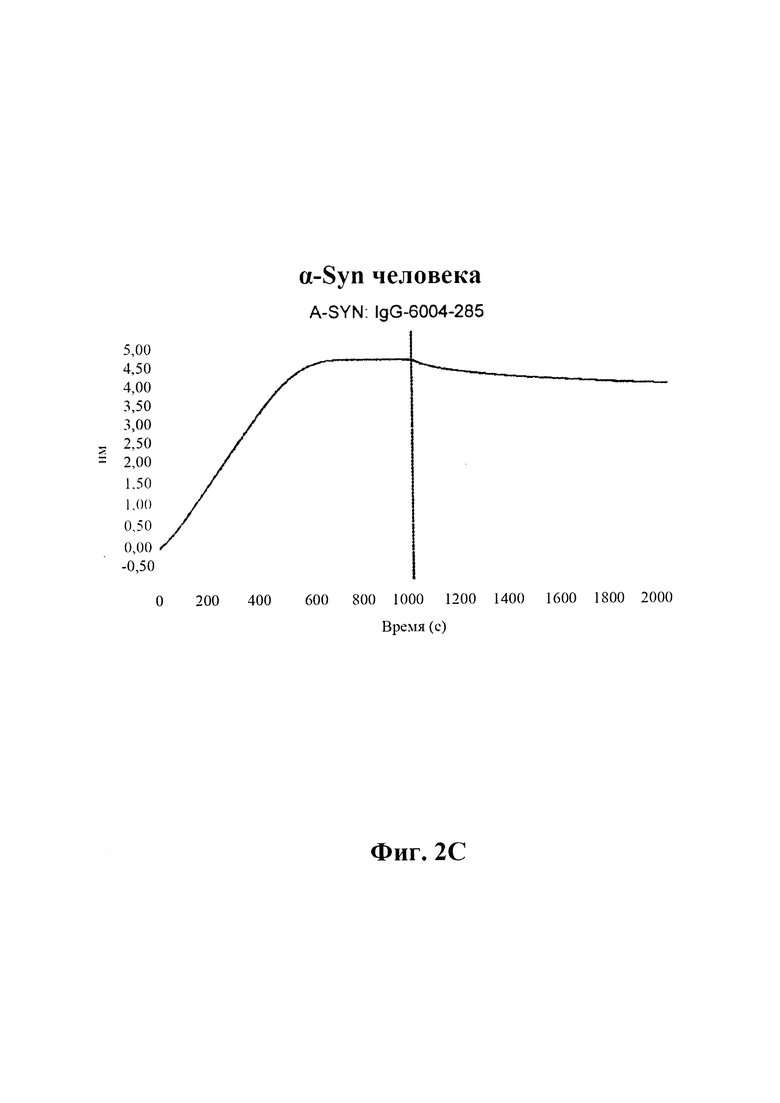
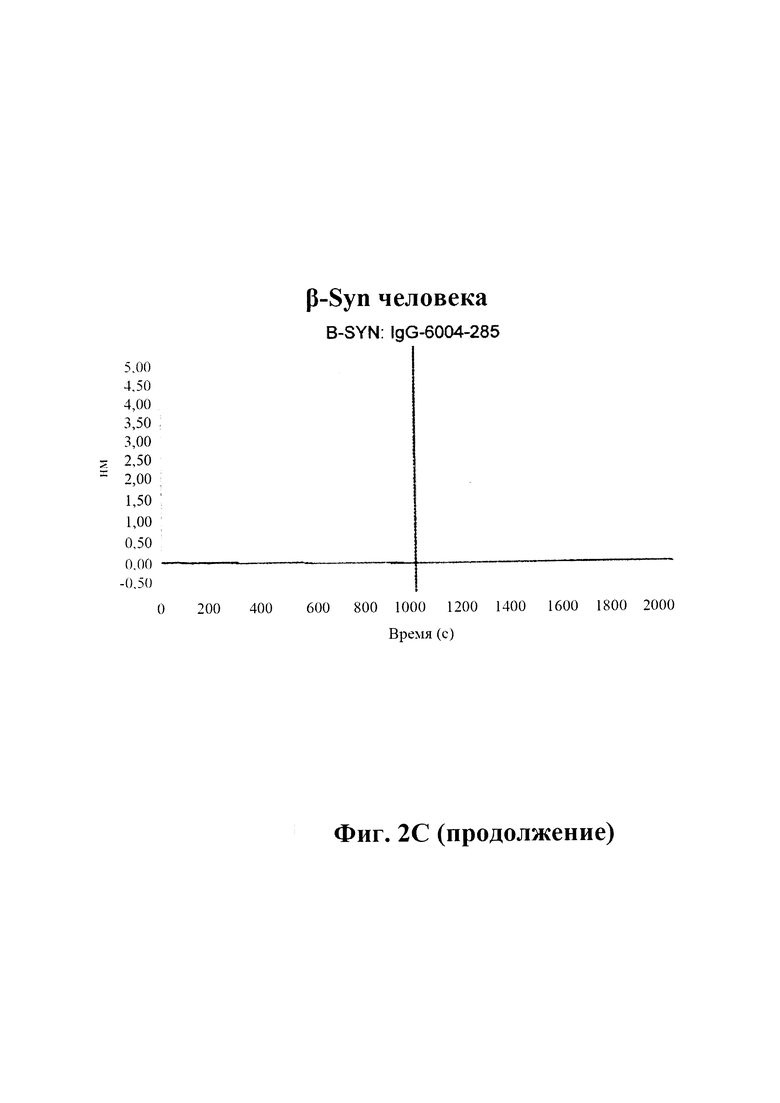
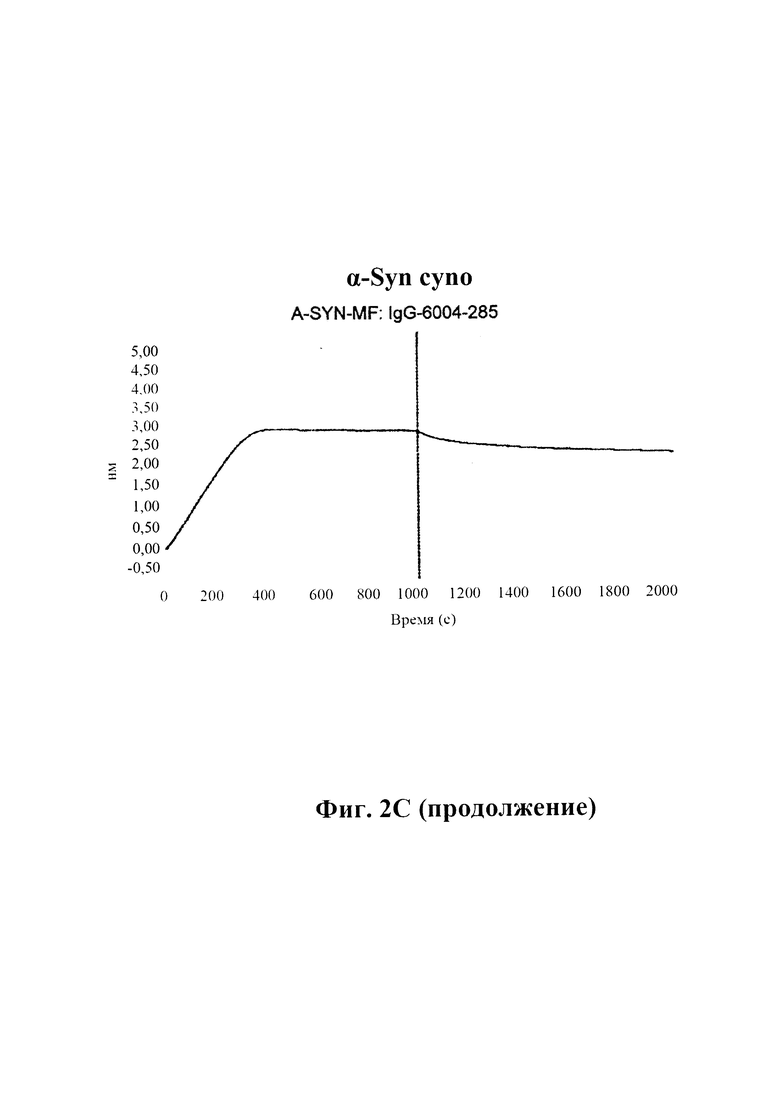
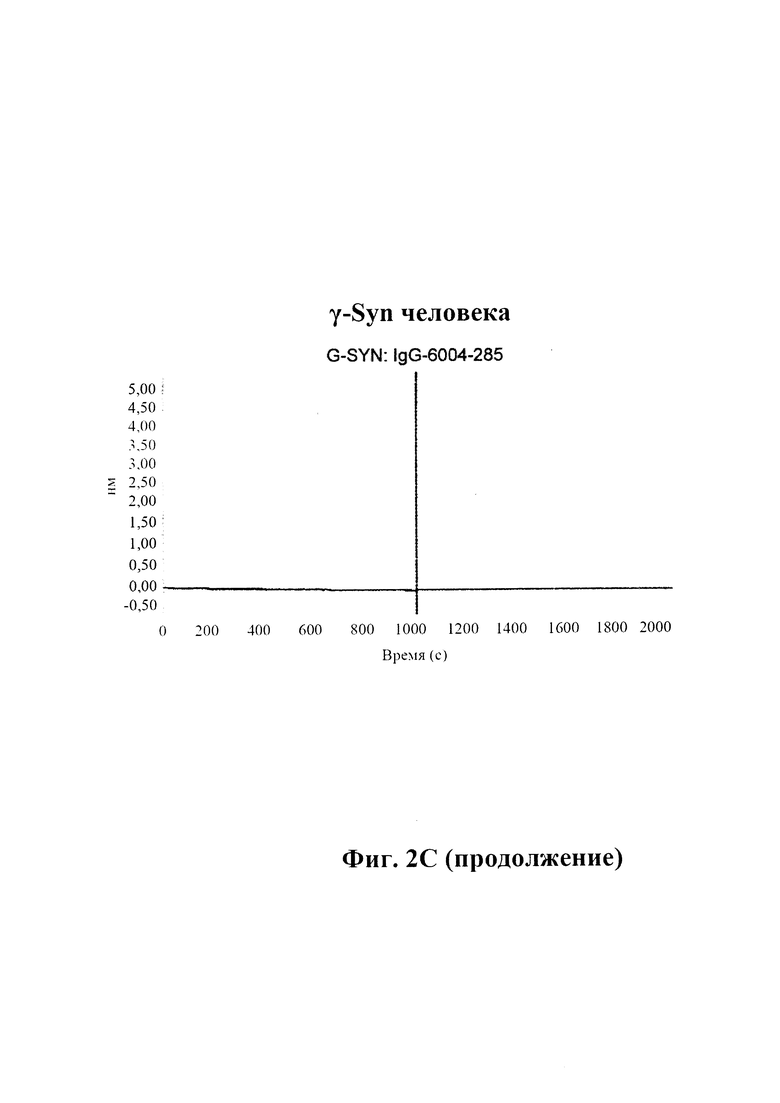
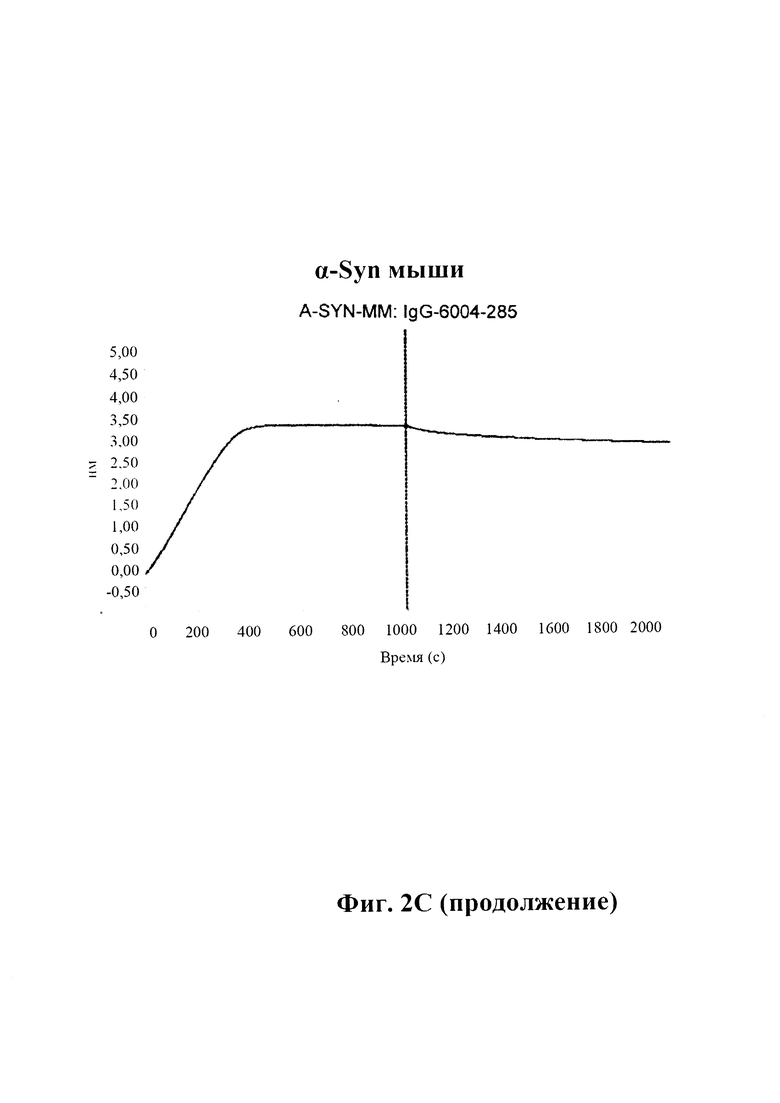
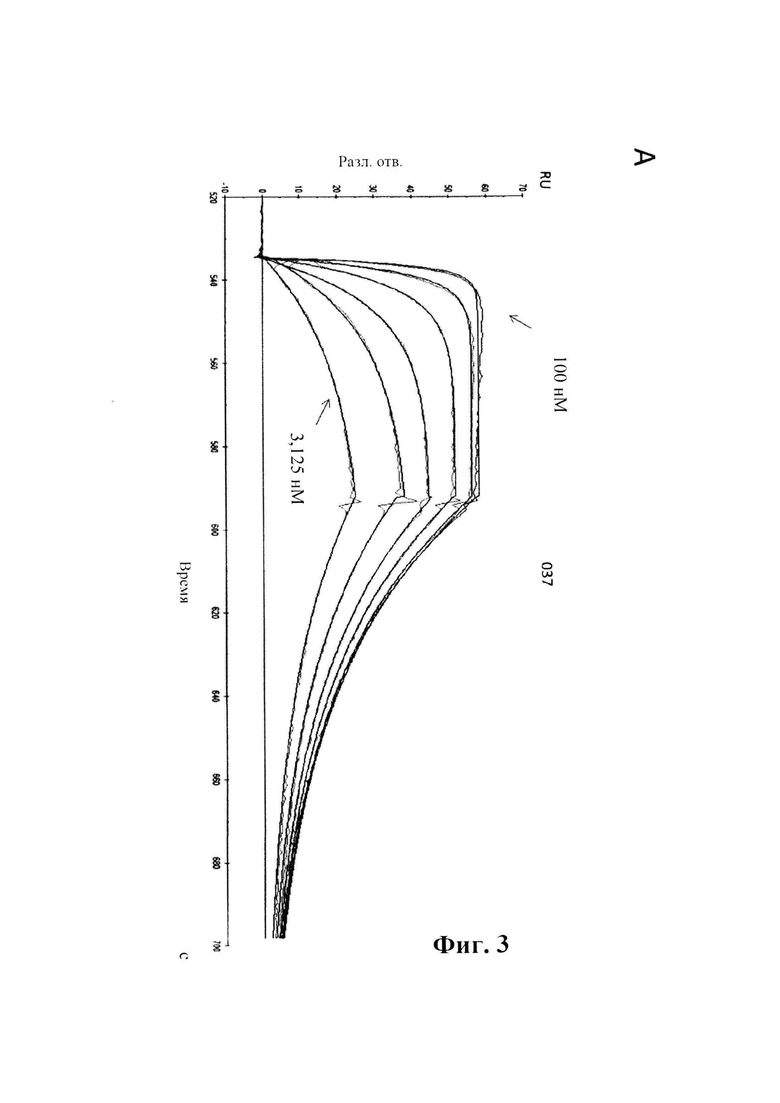
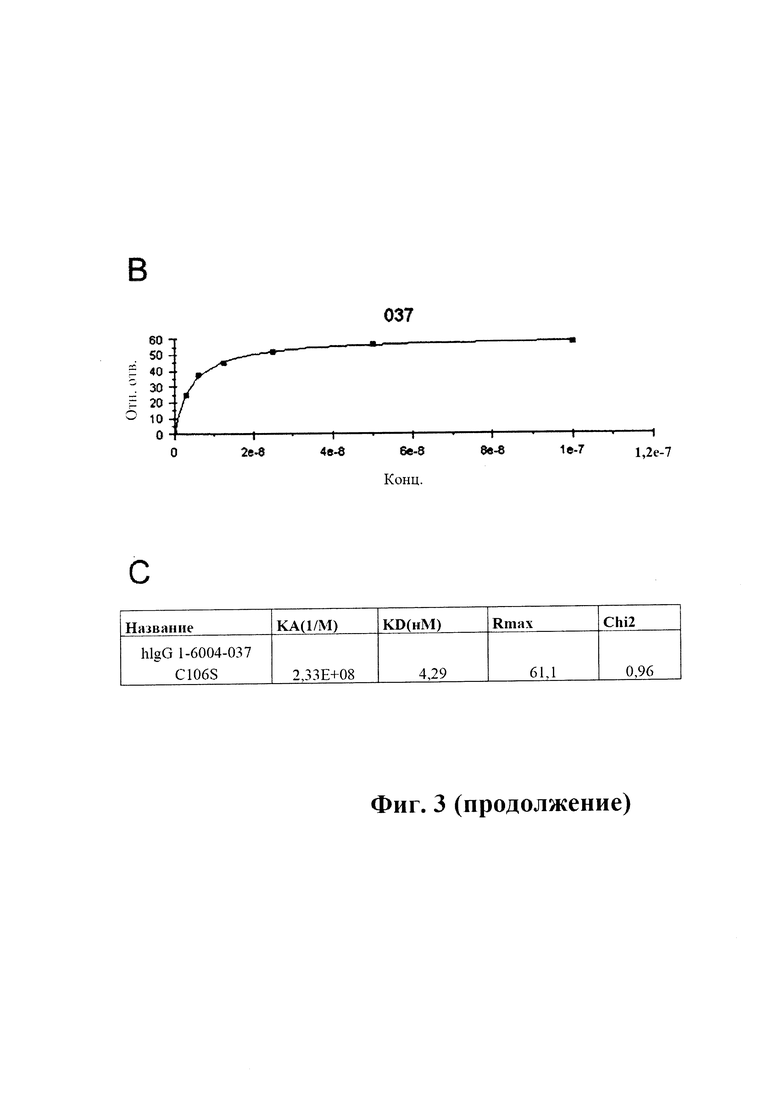
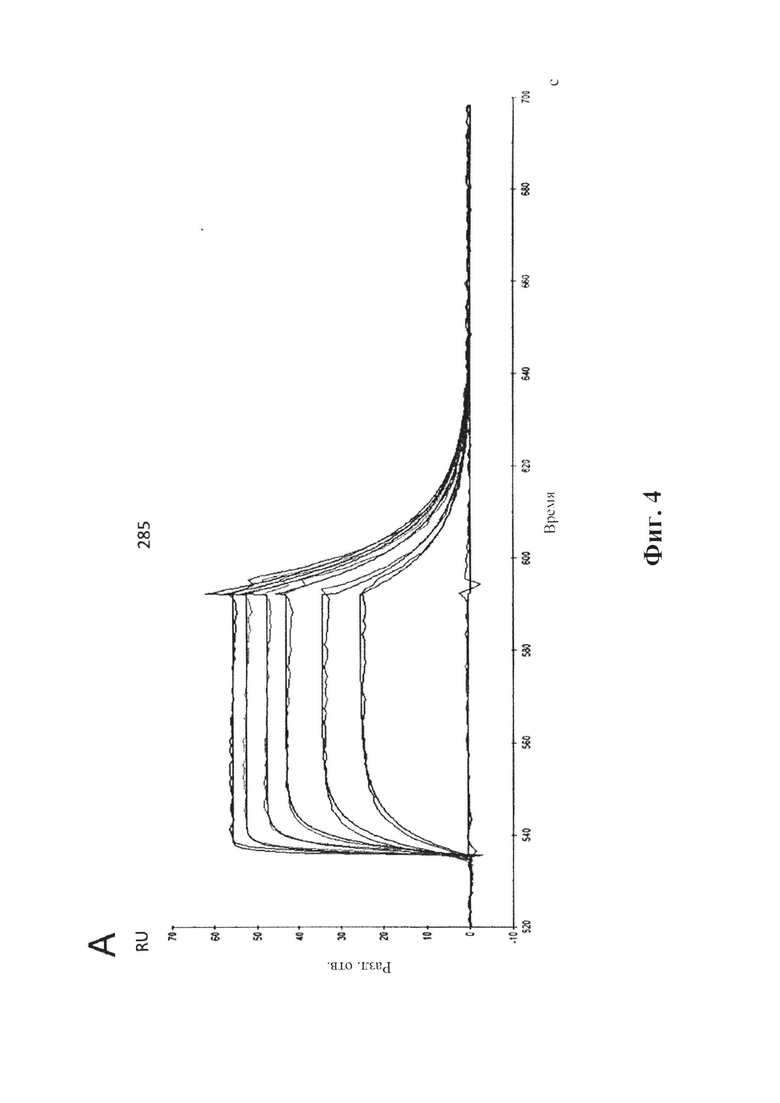
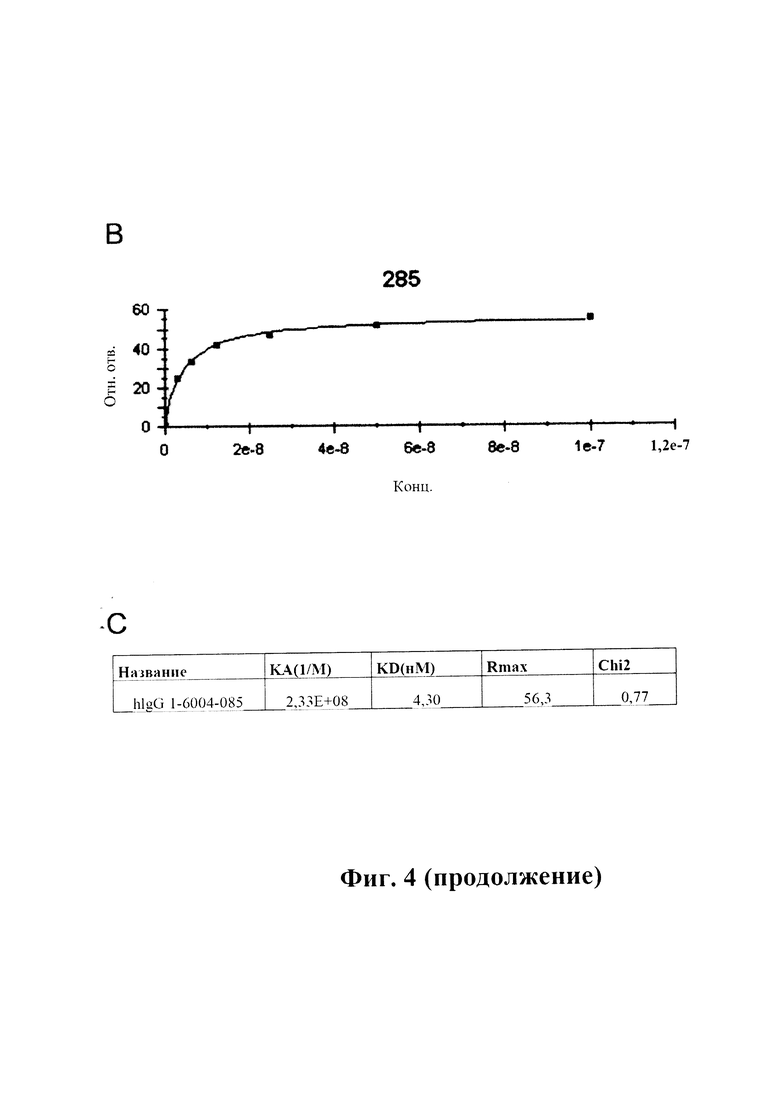
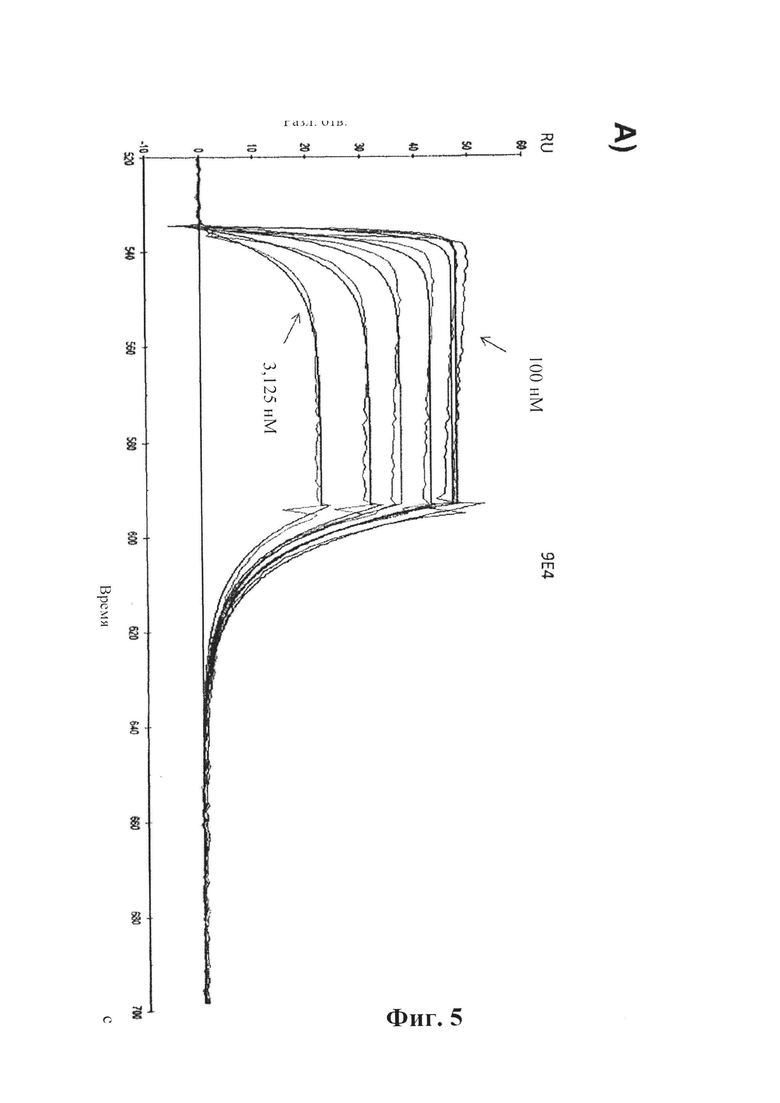
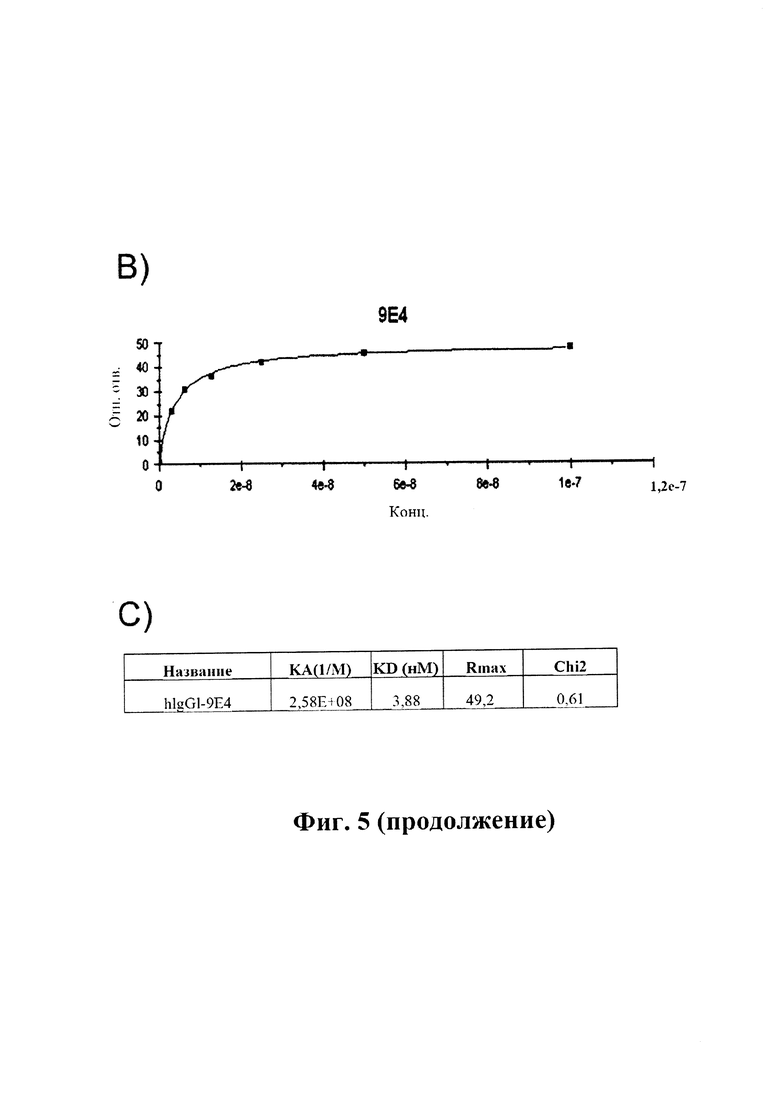

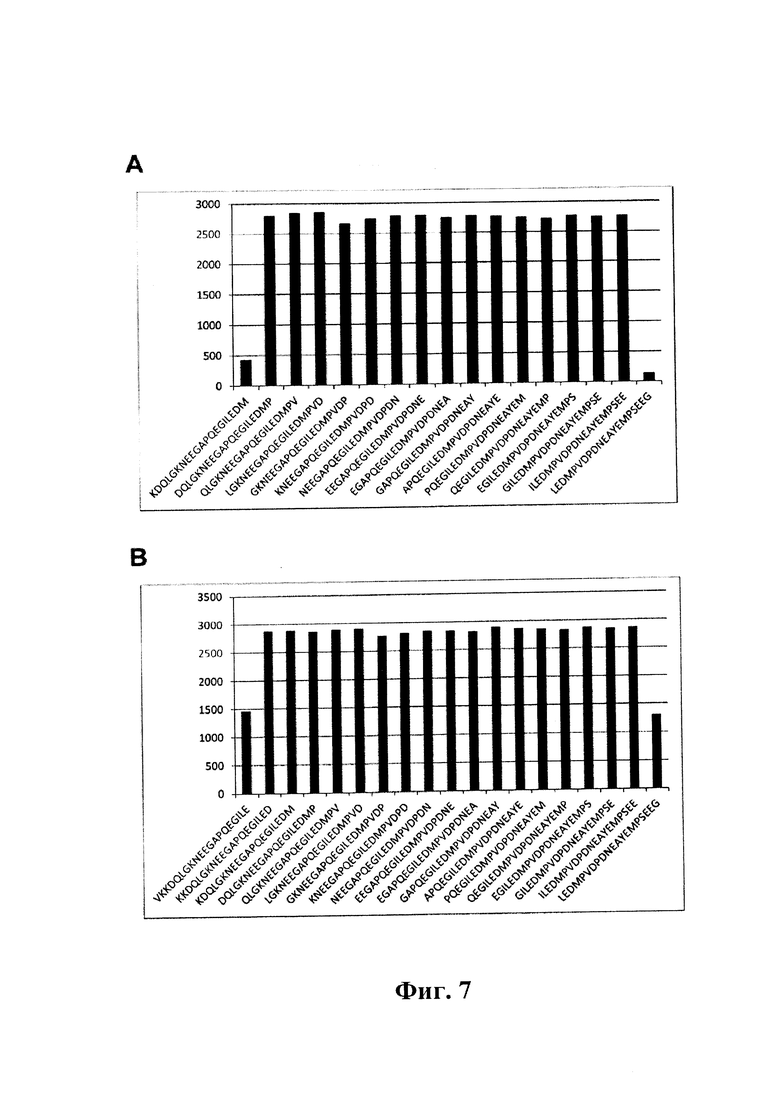

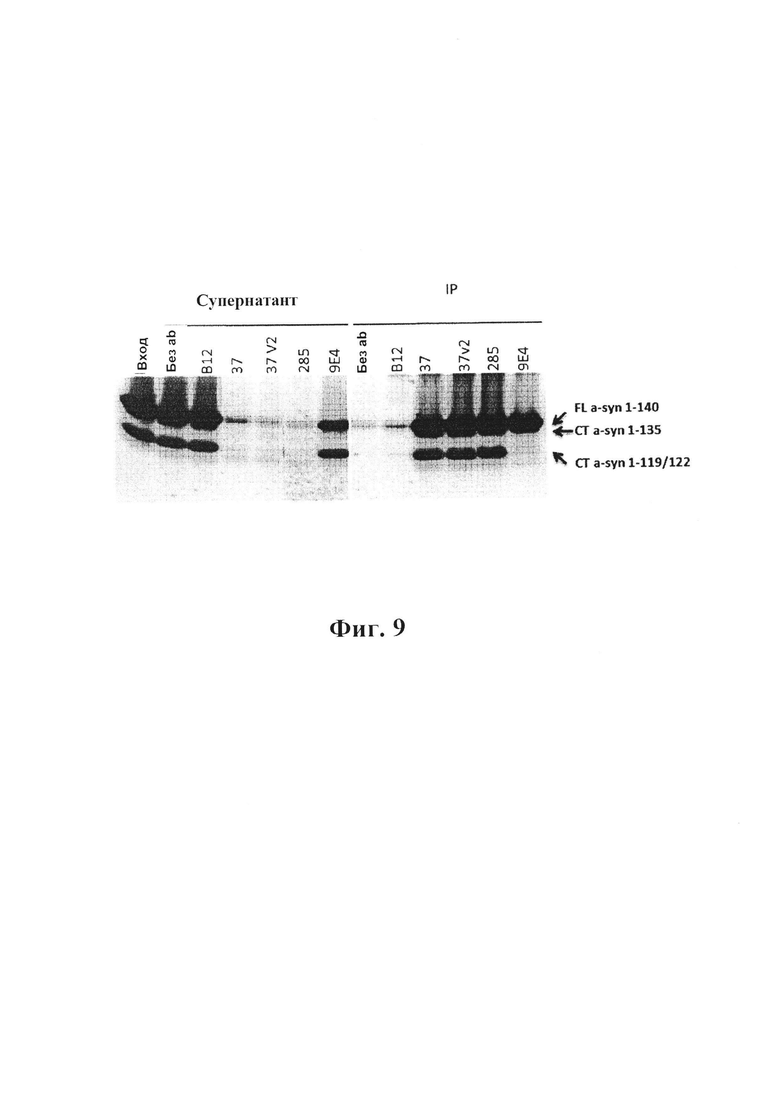
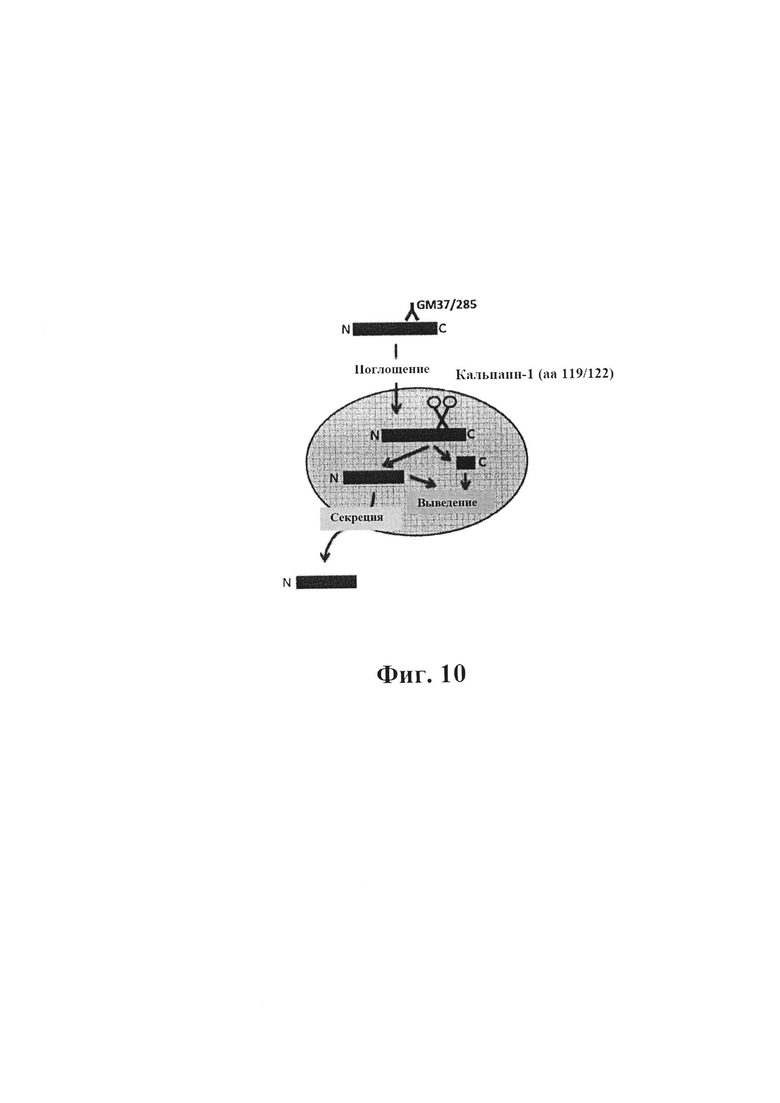
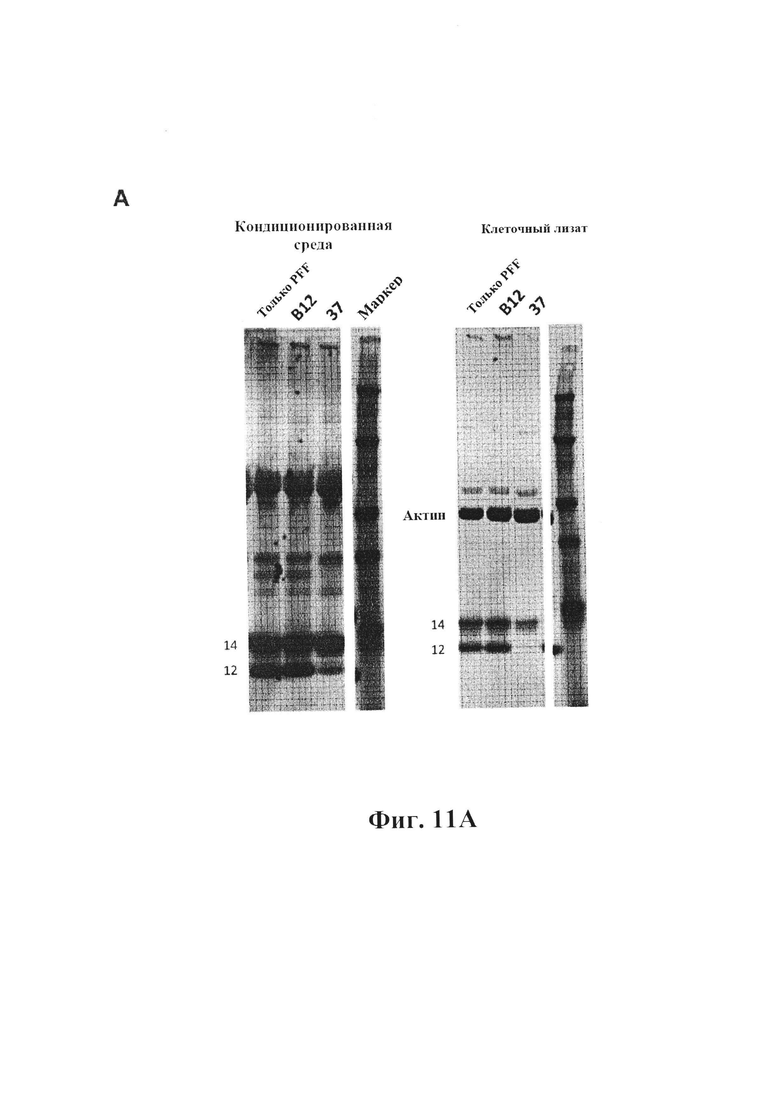
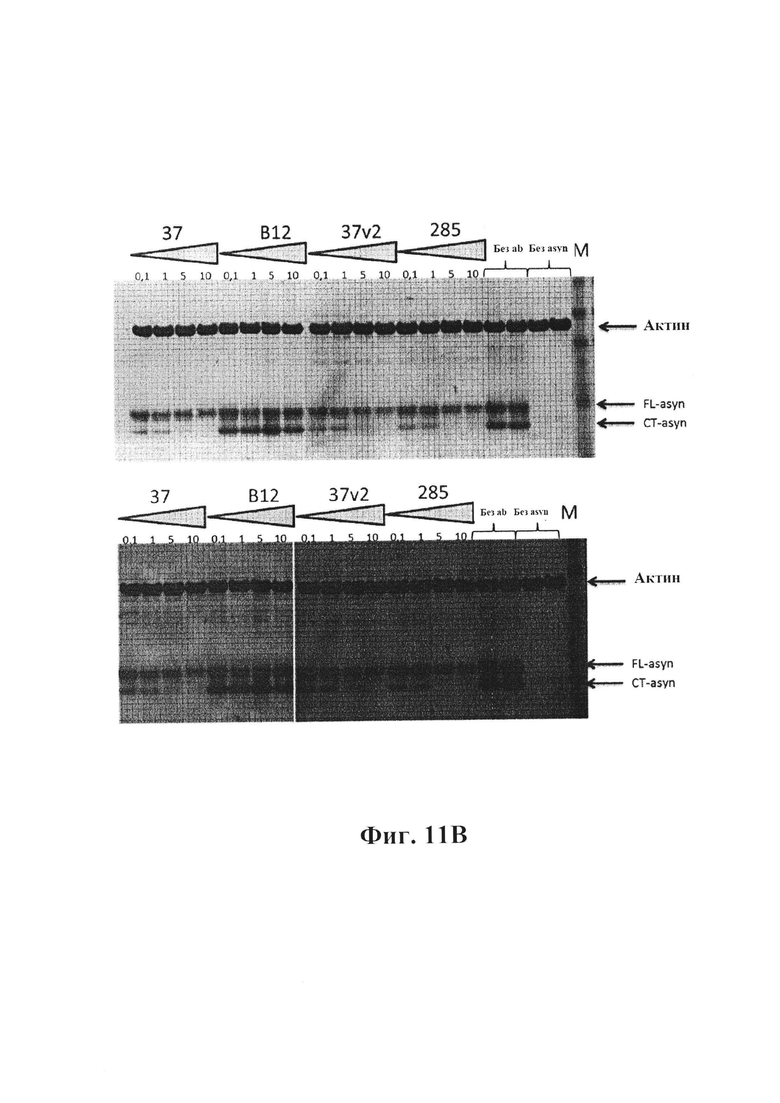

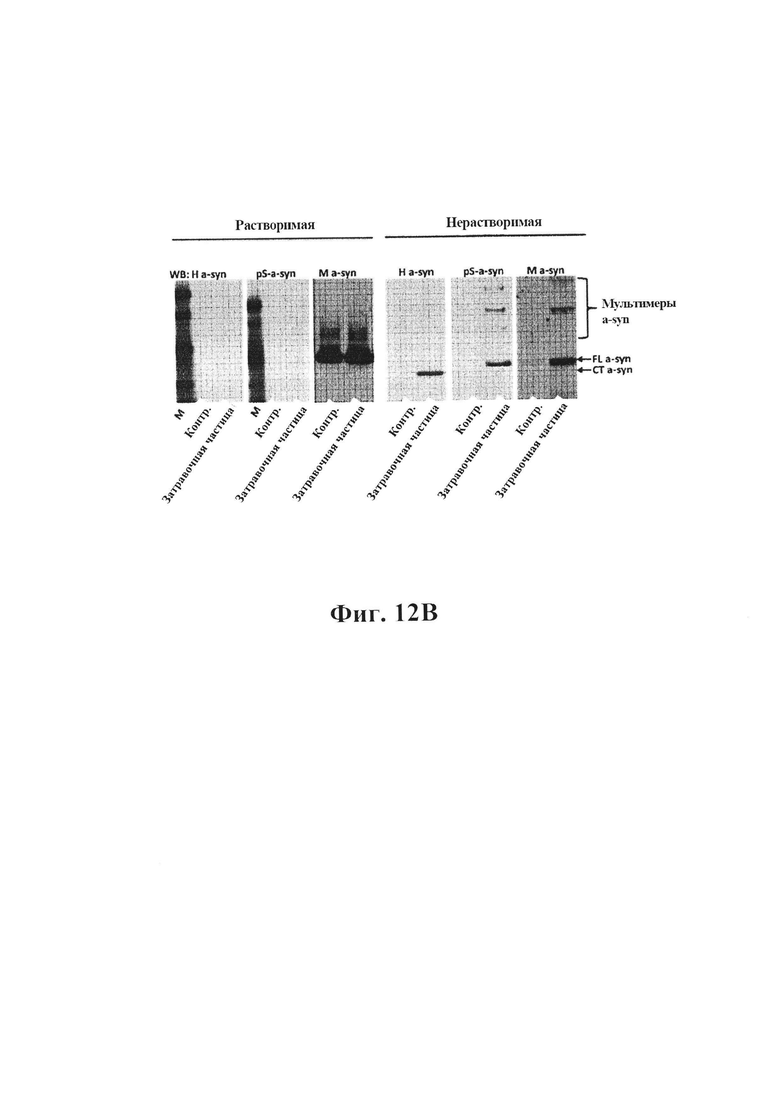
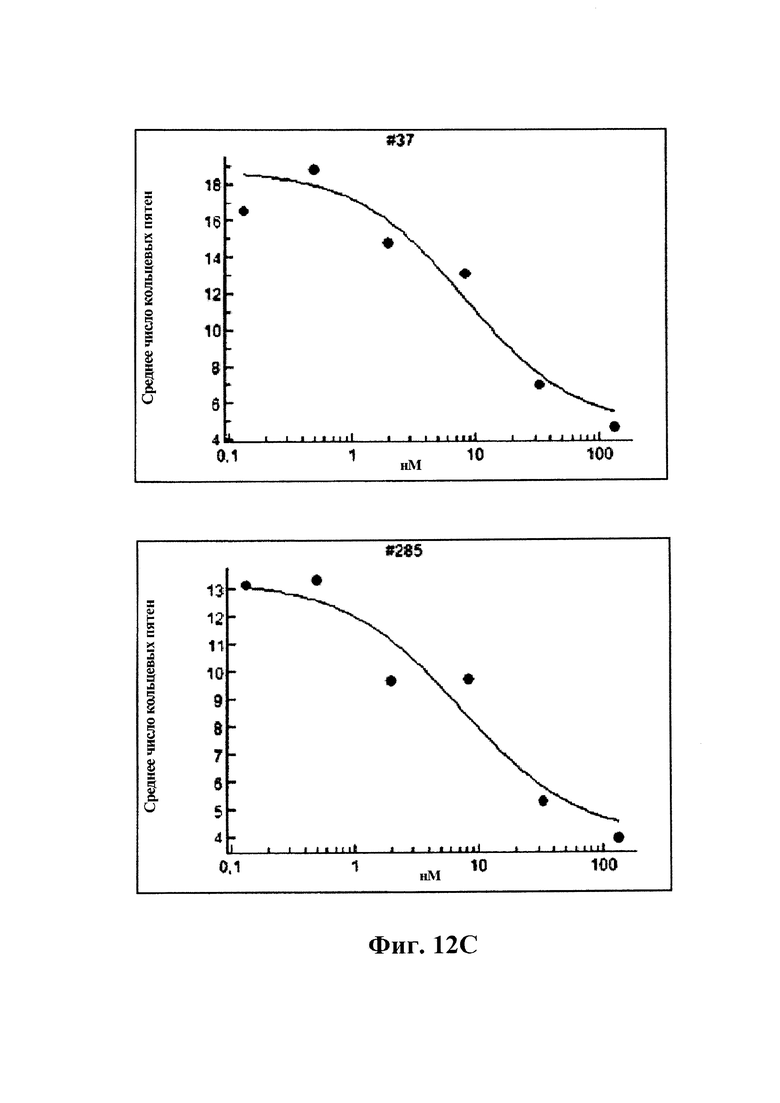
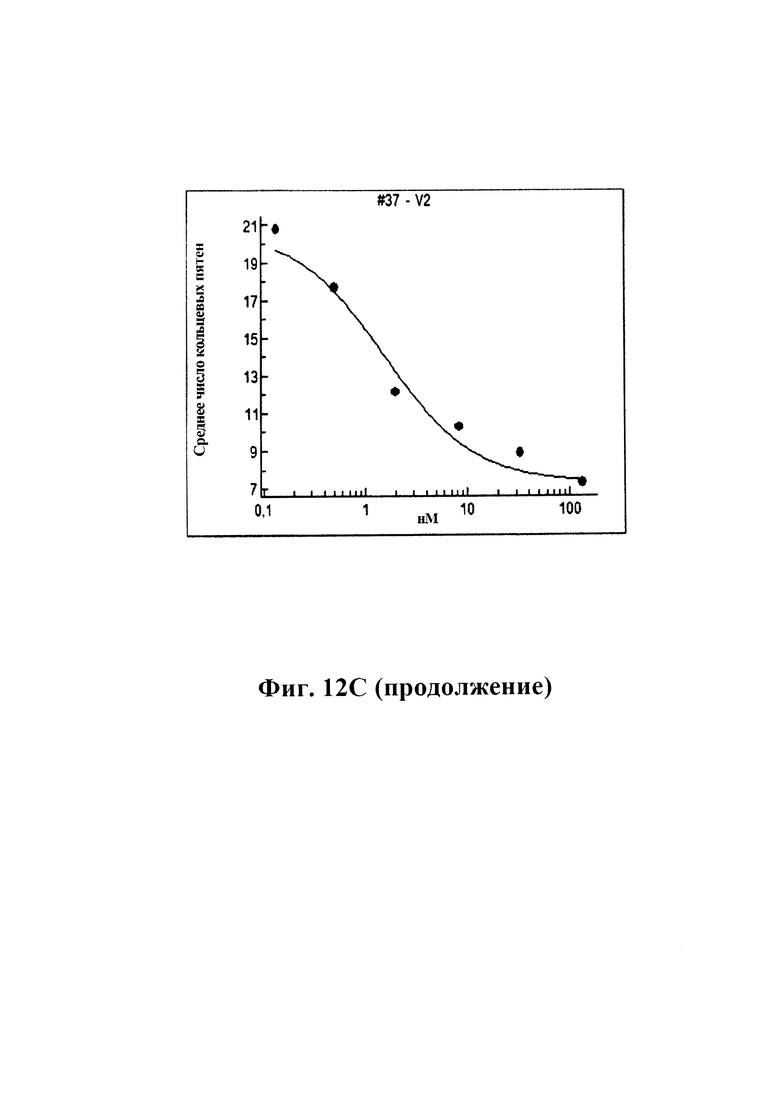

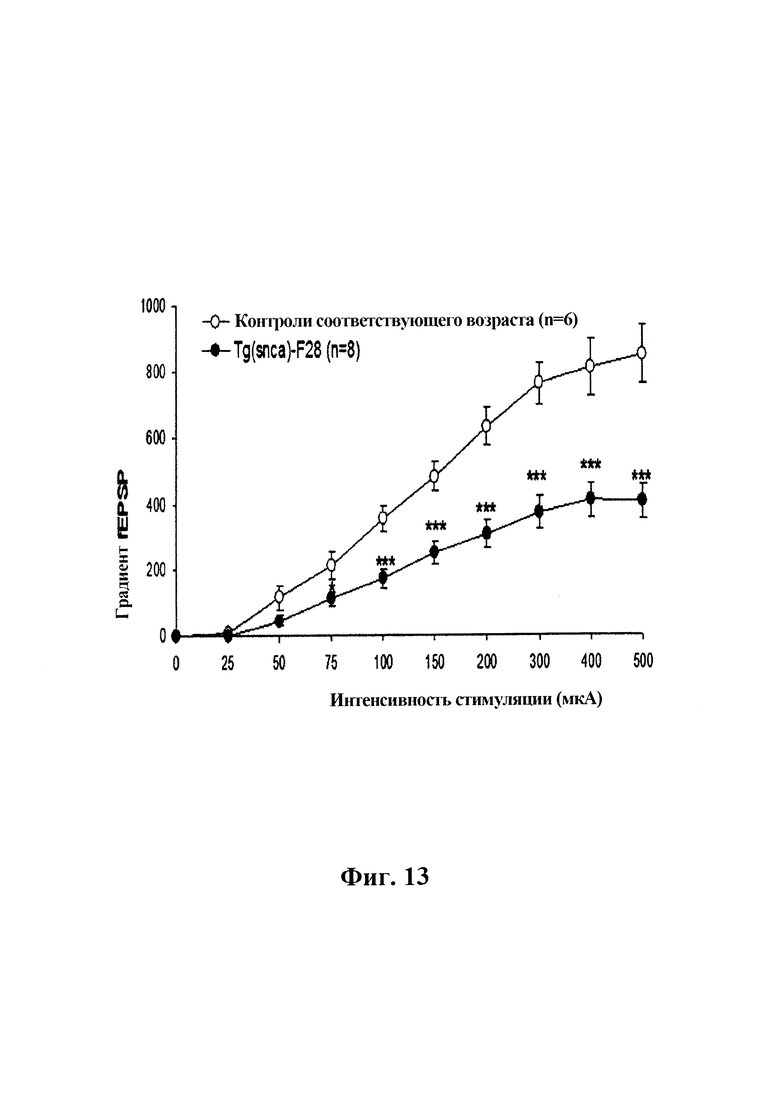
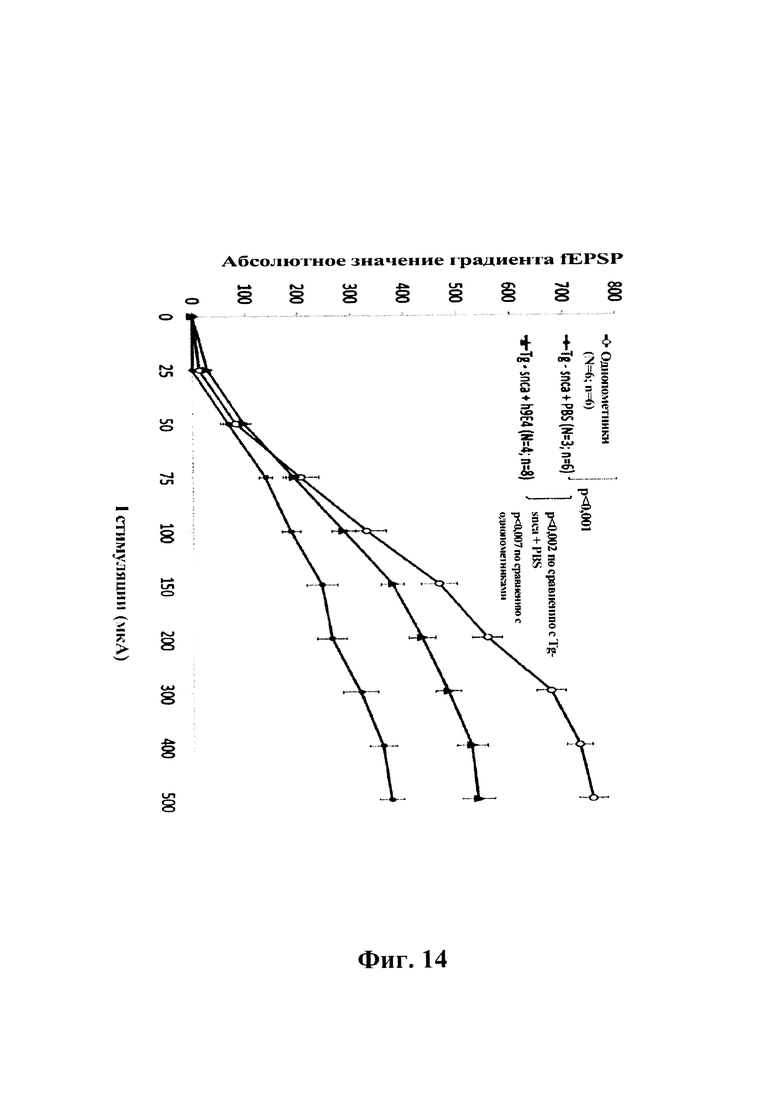
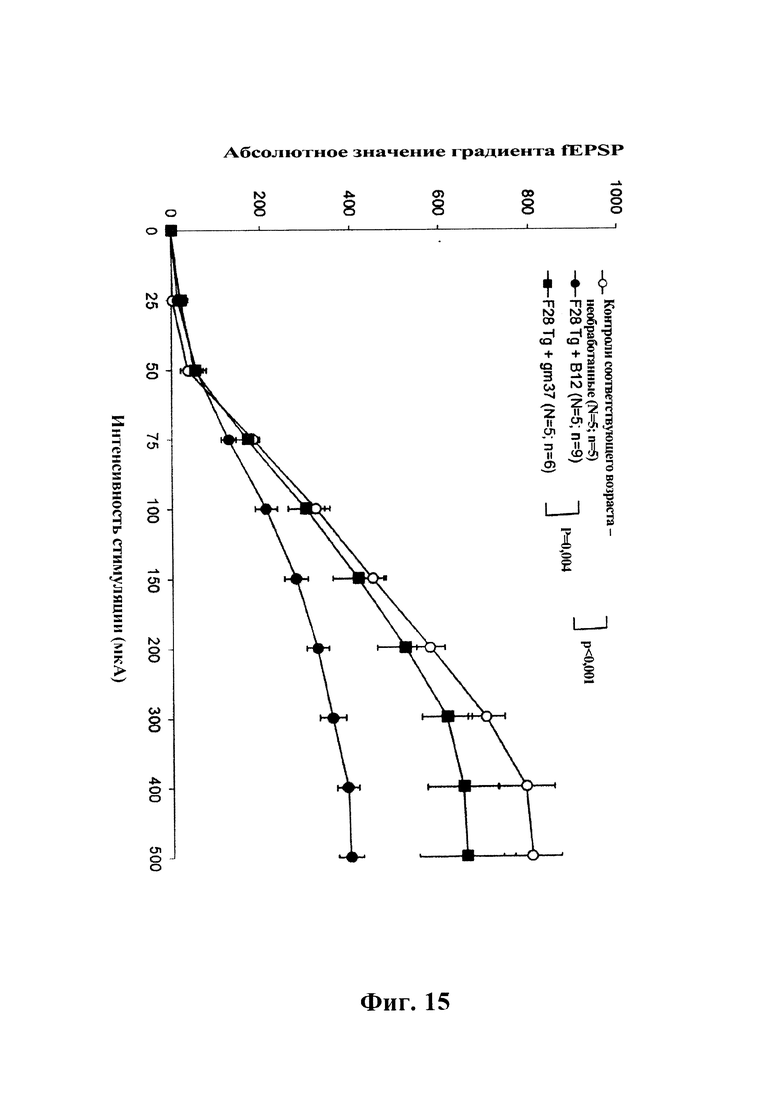
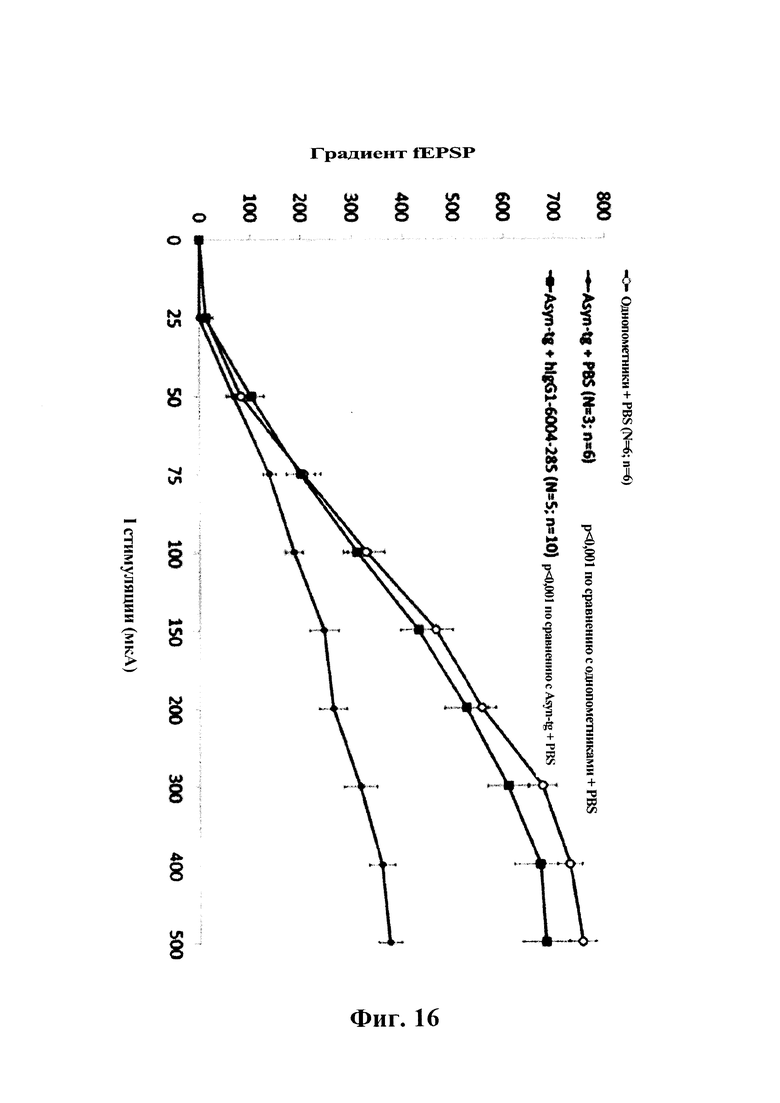
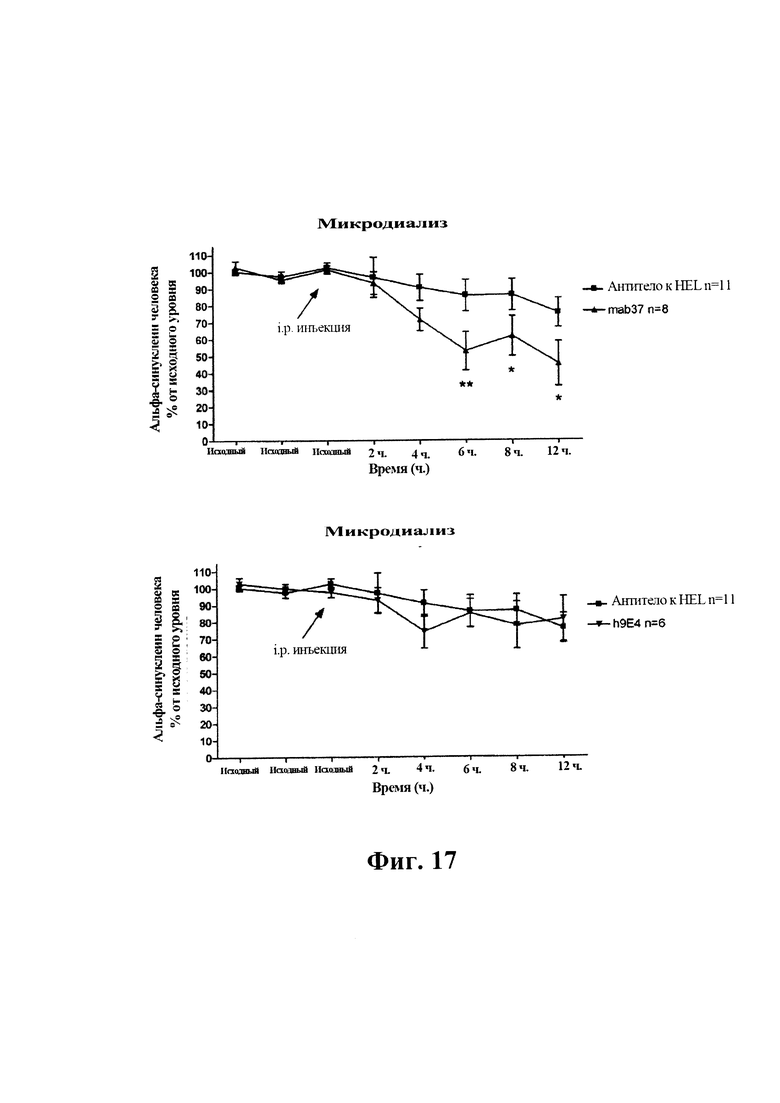



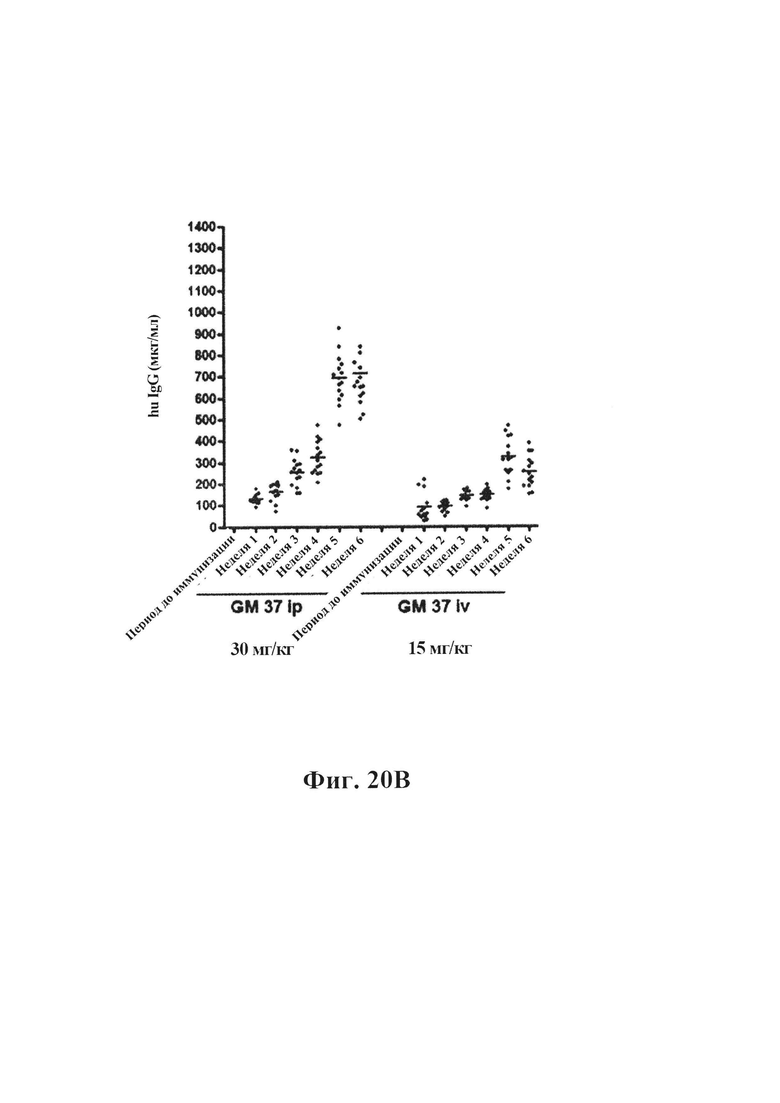
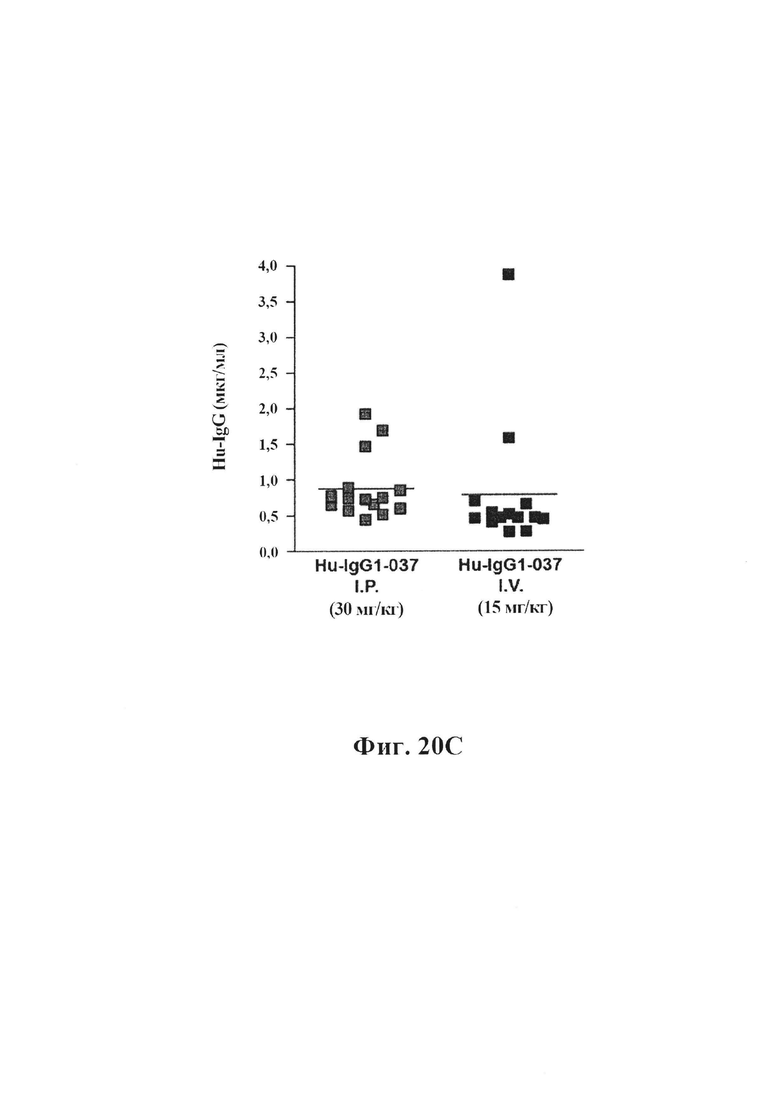
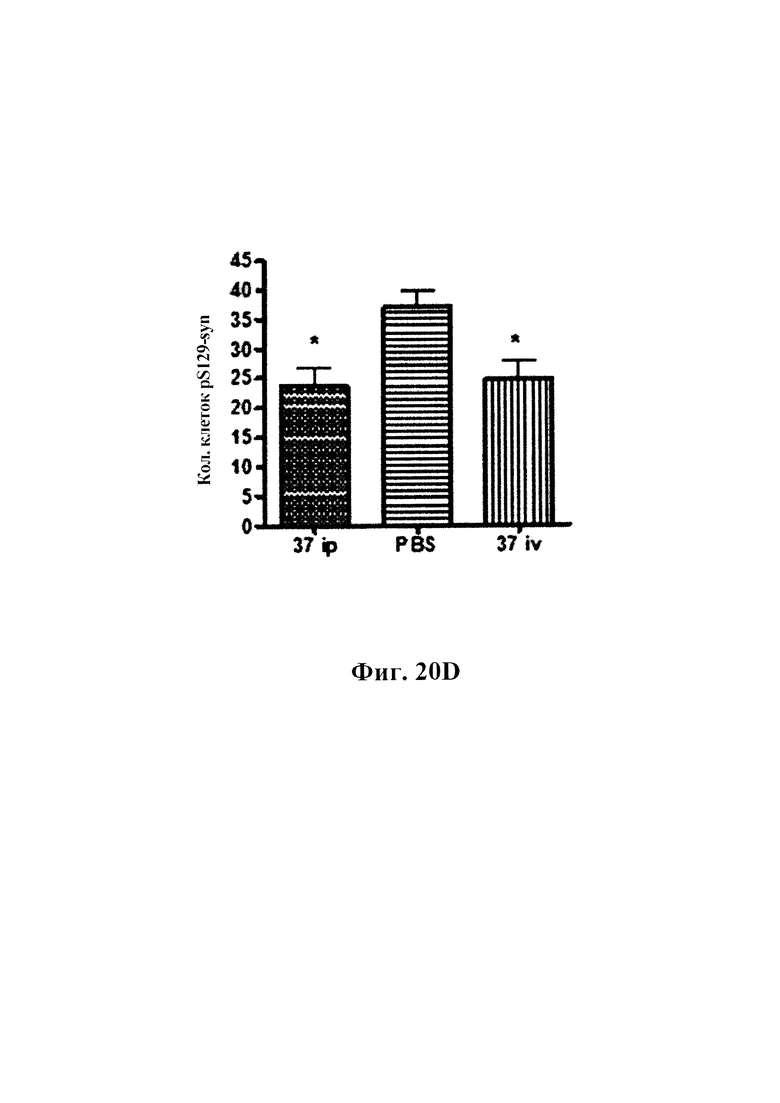
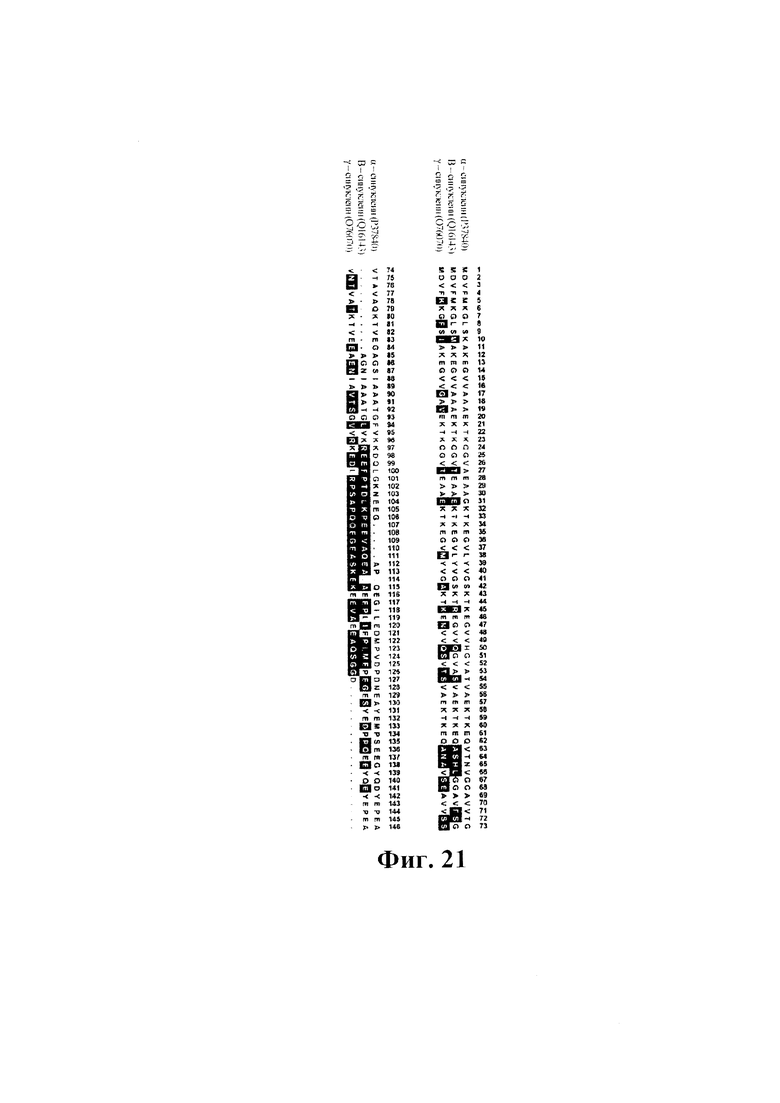
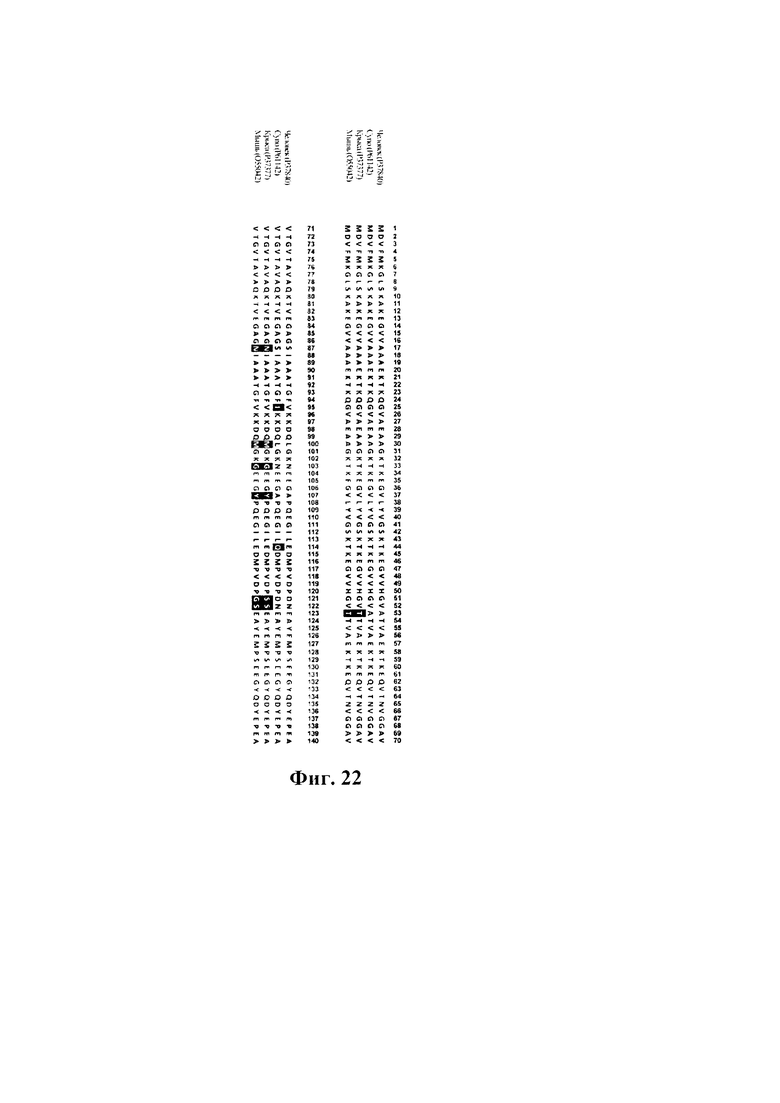

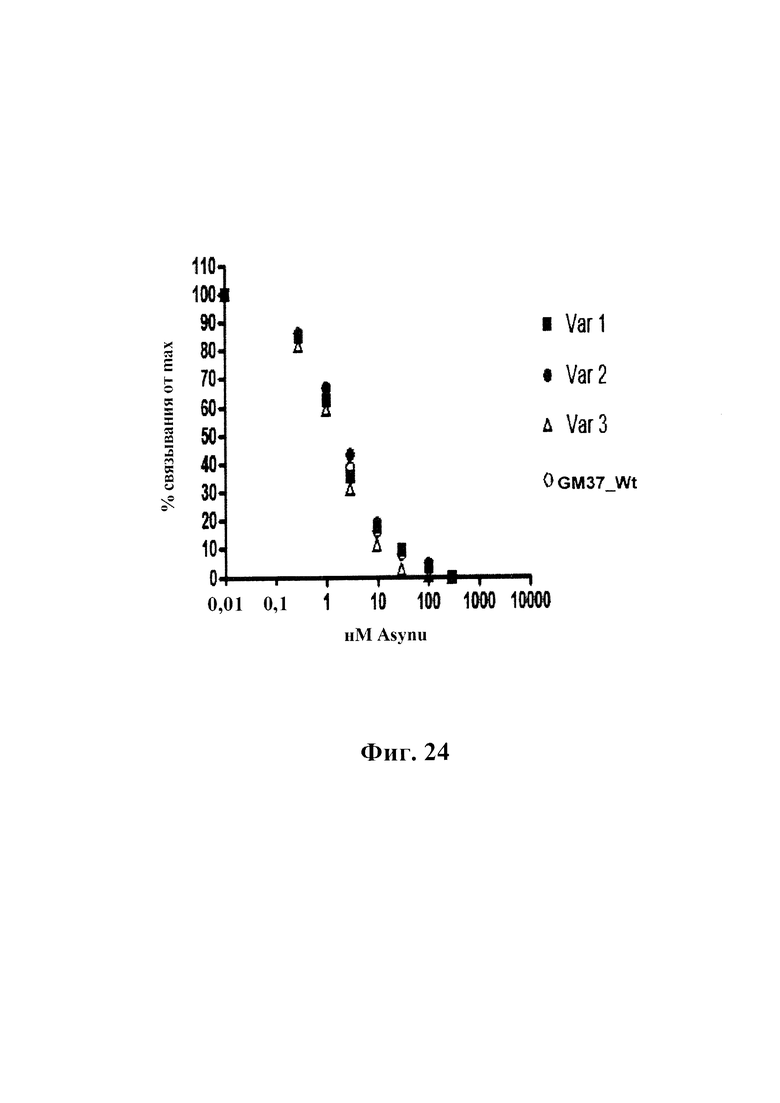
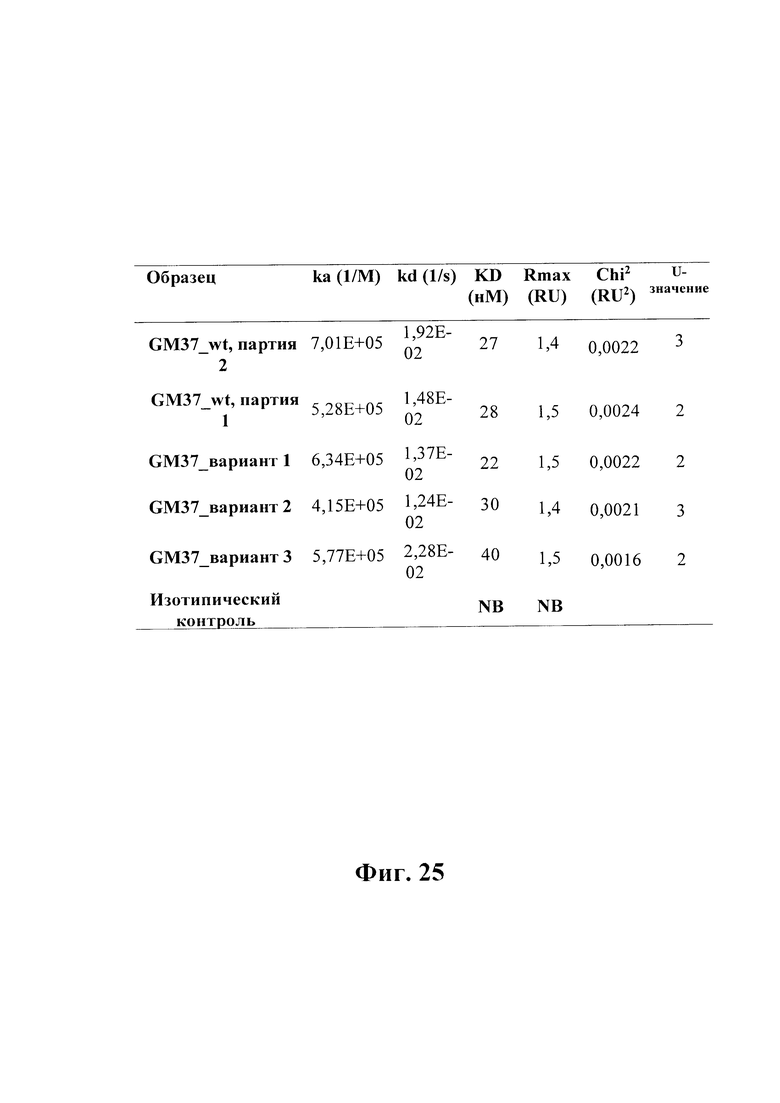
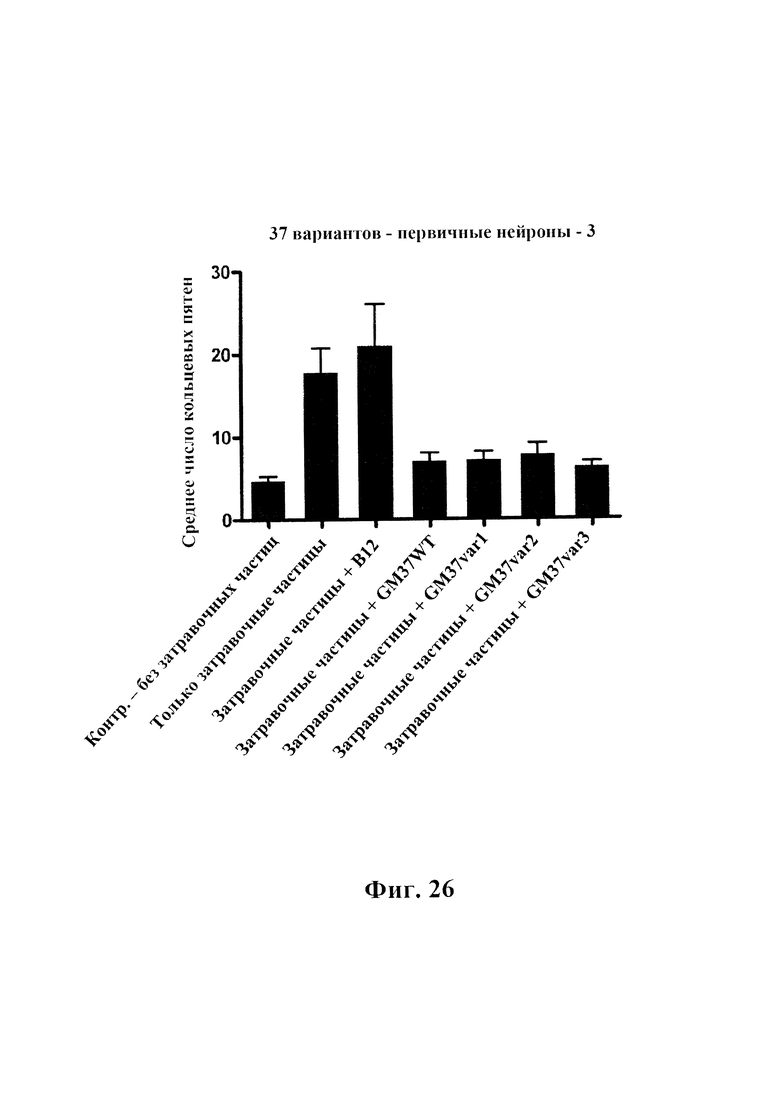
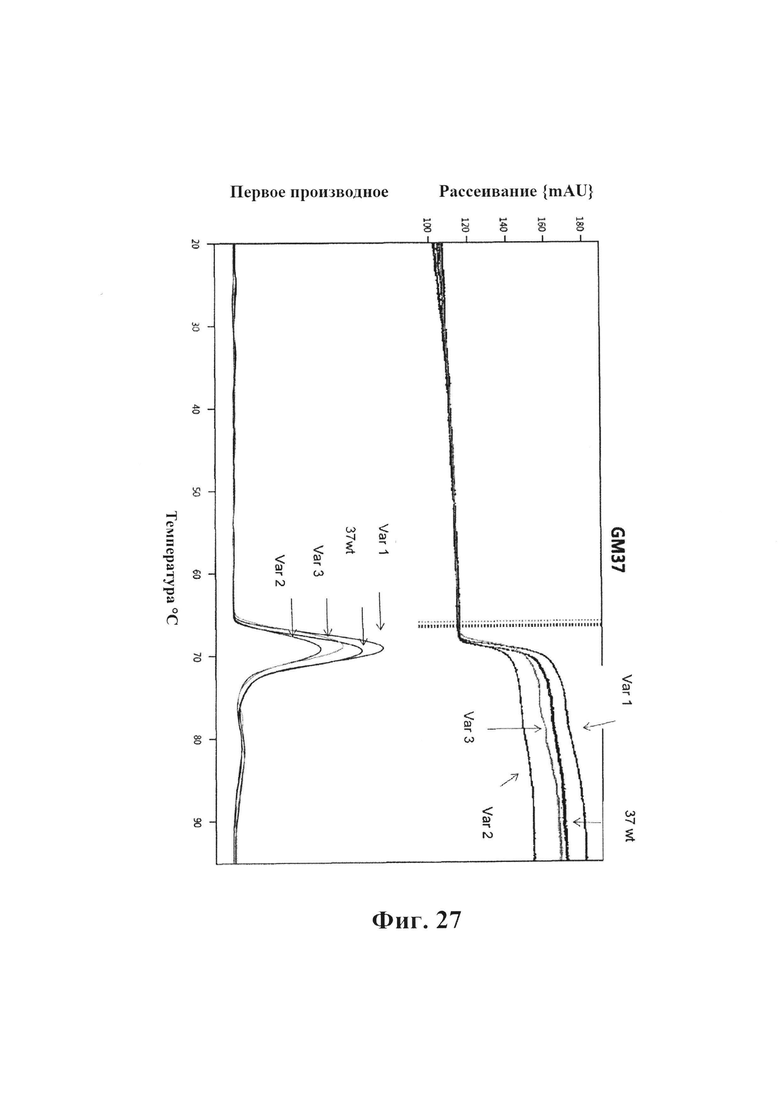
Изобретение относится к области биотехнологии, в частности к моноклональному антителу или его антигенсвязывающему фрагменту, которое способно специфически связываться с альфа-синуклеином человека, а также к содержащему его препарату, композиции и конъюгату. Также раскрыта нуклеиновая кислота, кодирующая указанное антитело или его фрагмент. Изобретение эффективно для лечения синуклеопатии, для выявления наличия альфа-синуклеина в головном мозге субъекта, а также для измерения количества альфа-синуклеина в головном мозге субъекта. 19 н. и 37 з.п. ф-лы, 27 ил., 4 табл., 11 пр.
1. Моноклональное антитело или его антигенсвязывающий фрагмент, которое способно специфически связываться с альфа-синуклеином человека, где указанное антитело связывает эпитоп в пределах аминокислот 112-117, имеющих аминокислотную последовательность SEQ ID NO:9 альфа-синуклеина человека, имеющего аминокислотную последовательность SEQ ID NO:10, причем антитело или его антигенсвязывающий фрагмент включает:
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 1;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 2;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 3;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6; или
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 1;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 33;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 3;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6; или
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 1;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 34;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 3;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6; или
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 1;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 35;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 3;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 4;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 5; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 6; или
(a) CDR1 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 20;
(b) CDR2 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 21;
(c) CDR3 тяжелой цепи с аминокислотной последовательностью с SEQ ID NO: 22;
(d) CDR1 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 23;
(e) CDR2 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 24; и
(f) CDR3 легкой цепи с аминокислотной последовательностью с SEQ ID NO: 25.
2. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, которое способно специфически связываться с эпитопом в пределах аминокислот 112-115, имеющих аминокислотную последовательность SEQ ID NO: 19 альфа-синуклеина человека, имеющего аминокислотную последовательность SEQ ID NO: 10.
3. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-2, причем моноклональное антитело представляет собой интактное антитело.
4. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, где моноклональное антитело выбрано из группы, состоящей из антител подтипа IgG1, IgG2, IgG3 или IgG4.
5. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-4, причем антигенсвязывающий фрагмент выбран из группы, состоящей из Fv-фрагментов, Fab-подобных фрагментов и доменных антител.
6. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-5, где антитело или антигенсвязывающий фрагмент характеризуются одним или несколькими из следующих свойств:
a) аффинностью связывания к альфа-синуклеину в диапазоне 0,5-10 нМ, например 1-5 нМ или 1-2 нМ;
b) способностью подавлять усечение протеазами фибрилл альфа-синуклеина;
c) способностью обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
d) способностью снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo;
e) способностью, при длительном введении, восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
f) способностью предотвращать затравочное действие альфа-синуклеина; и/или
g) способностью связывать усеченный альфа-синуклеин в головном мозге человека.
7. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, причем моноклональное антитело представляет собой человеческое, гуманизированное, рекомбинантное или химерное антитело.
8. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельный домен тяжелой цепи с SEQ ID NO: 7 или вариабельный домен легкой цепи с SEQ ID NO: 8.
9. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 8, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 7, и легкую цепь, состоящую из вариабельного домена с SEQ ID NO: 8.
10. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельный домен тяжелой цепи с SEQ ID NO: 30 или вариабельный домен легкой цепи с SEQ ID NO: 8.
11. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 10, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 30, и легкую цепь, состоящую из вариабельного домена с SEQ ID NO: 8.
12. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельный домен тяжелой цепи с SEQ ID NO: 31 или вариабельный домен легкой цепи с SEQ ID NO: 8.
13. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 12, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 31 и вариабельного домена с SEQ ID NO: 8.
14. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельный домен тяжелой цепи с SEQ ID NO: 32 или вариабельный домен легкой цепи с SEQ ID NO: 8.
15. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 14, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 32 и вариабельного домена с SEQ ID NO: 8.
16. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельный домен тяжелой цепи с SEQ ID NO: 26 или вариабельный домен легкой цепи с SEQ ID NO: 27.
17. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 16, содержащее тяжелую цепь, состоящую из вариабельного домена с SEQ ID NO: 26 и вариабельного домена с SEQ ID NO: 27.
18. Препарат, содержащий моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17, для применения в лечении синуклеопатии, где указанный препарат практически не содержит антител, образующихся в естественных условиях, которые либо не способны связываться с альфа-синуклеином, либо существенным образом не изменяют функциональные характеристики антитела к альфа-синуклеину в препарате, при этом указанные функциональные характеристики выбраны из группы, состоящей из:
(i) аффинности связывания антитела к альфа-синуклеину в отношении альфа-синуклеина;
(ii) способности антитела к альфа-синуклеину подавлять усечение протеазами фибрилл альфа-синуклеина;
(iii) способности антитела к альфа-синуклеину обращать нарушение базальной синаптической передачи у трансгенных мышей F28-snca;
(iv) способности антитела к альфа-синуклеину снижать уровни альфа-синуклеина в гиппокампе мыши, по результатам измерения с помощью микродиализа in vivo;
(v) способности, при длительном введении, антитела к альфа-синуклеину восстанавливать двигательную функцию в крысиной модели болезни Паркинсона;
(vi) способности предотвращать затравочное действие альфа-синуклеина; или
(vii) способности связывать усеченный альфа-синуклеин в головном мозге человека.
19. Фармацевтическая композиция, содержащая эффективное количество моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-17 и фармацевтически приемлемый носитель для применения в лечении синуклеопатии.
20. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17.
21. Применение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-17 для лечения синуклеопатии.
22. Применение по п. 21, причем синуклеопатия выбрана из болезни Паркинсона, идиопатической и наследственной форм болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
23. Применение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-17 для изготовления лекарственного препарата для лечения синуклеопатии.
24. Применение по п. 23, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
25. Способ лечения синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии, при этом указанный способ предусматривает введение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-17, или фармацевтической композиции по п. 19 указанному субъекту в эффективном количестве.
26. Способ по п. 25, где лечение является длительным.
27. Способ по п. 26, где длительное лечение длится по меньшей мере 2 недели.
28. Способ по п. 27, где субъектом является человек.
29. Конъюгат для лечения синуклеопатии, содержащий моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17, который помечен с помощью выявляемой метки.
30. Конъюгат по п. 29, где указанная выявляемая метка представляет собой флуоресцентную метку, хемилюминесцентную метку, парамагнитную метку, радиоизотопную метку или ферментную метку.
31. Применение конъюгата по любому из пп. 29-30 для выявления наличия указанного альфа-синуклеина в головном мозге субъекта.
32. Применение по п. 31, где указанное выявление предусматривает визуализацию in vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
33. Применение по любому из пп. 31-32, где указанное выявление предусматривает визуализацию ex vivo указанного антитела к синуклеину или указанного его антигенсвязывающего фрагмента, связанных с указанным альфа-синуклеином.
34. Применение конъюгата по любому из пп. 29-30 для измерения количества указанного альфа-синуклеина в головном мозге субъекта.
35. Применение по п. 34, где указанное измерение предусматривает визуализацию in vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
36. Применение по любому из пп. 34-35, где указанное измерение предусматривает визуализацию ex vivo указанного антитела к синуклеину или указанного его антигенсвязывающего фрагмента, связанных с указанным альфа-синуклеином.
37. Применение фармацевтической композиции по п. 19 для изготовления лекарственного препарата для лечения синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
38. Применение фармацевтической композиции по п. 19 для изготовления лекарственного препарата для диагностики синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
39. Применение фармацевтической композиции по п. 19 для изготовления лекарственного препарата для визуализации синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
40. Применение конъюгата по любому из пп. 29-30 для изготовления лекарственного препарата для лечения синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
41. Применение конъюгата по любому из пп. 29-30 для изготовления лекарственного препарата для диагностики синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
42. Применение конъюгата по любому из пп. 29-30 для изготовления лекарственного препарата для визуализации синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии.
43. Способ лечения синуклеопатии, причем синуклеопатия выбрана из болезни Паркинсона, болезни Гоше, болезни диффузных телец Леви, варианта болезни Альцгеймера с тельцами Леви, комбинированной болезни Альцгеймера и Паркинсона, истинной вегетативной недостаточности или множественной системной атрофии, при этом указанный способ предусматривает введение конъюгата по любому из пп. 29-30 указанному субъекту в эффективном количестве.
44. Способ по п. 43, где лечение является длительным.
45. Способ по п. 44, где длительное лечение длится по меньшей мере 2 недели.
46. Способ по п. 45, где субъектом является человек.
47. Применение конъюгата по любому из пп. 29-30 для выявления наличия указанного альфа-синуклеина в головном мозге или жидкости организма субъекта.
48. Применение по п. 47, где указанное выявление предусматривает визуализацию in vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
49. Применение по п. 48, где указанное выявление предусматривает визуализацию ex vivo указанного антитела к синуклеину или указанного его антигенсвязывающего фрагмента, связанных с указанным альфа-синуклеином.
50. Применение конъюгата по любому из пп. 29-30 для измерения количества указанного альфа-синуклеина в головном мозге или жидкости организма субъекта.
51. Применение по п. 50, где указанное измерение предусматривает визуализацию in vivo указанного антитела к синуклеину, связанного с указанным альфа-синуклеином.
52. Применение по п. 51, где указанное измерение предусматривает визуализацию ex vivo указанного антитела к синуклеину или указанного его антигенсвязывающего фрагмента, связанных с указанным альфа-синуклеином.
53. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-17, которые были получены или произведены в линии клеток, такой как линия клеток человека, линия клеток млекопитающего, отличного от человека, линия клеток насекомого, дрожжей или бактерии.
54. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-15, полученные в линии клеток СНО, линии клеток HEK, линии клеток BHK-21, линии клеток мыши, линии клеток фибросаркомы, линии клеток PER.C6, линии клеток HKB-11, линии клеток САР и линии клеток НиН-7 человека.
55. Моноклональное антитело, его антигенсвязывающий фрагмент по любому из пп. 1-15, содержащие константный домен, определенный в SEQ ID NO: 18, и константный каппа-домен, определенный в SEQ ID NO: 17.
56. Моноклональное антитело, его антигенсвязывающий фрагмент по любому из пп. 16-17, содержащие константный домен, определенный в SEQ ID NO: 28, и константный каппа-домен, определенный в SEQ ID NO: 29.
| WO2007011907 A2, 25.01.2007 | |||
| WO2015075011 A1, 21.05.2015 | |||
| ROSS JAKES et al., Epitope mapping of LB509, a monoclonal antibodydirected against α-synuclein, Neuroscience Letters, 1999, Vol.269, pp.13-16 | |||
| US2009208487 A1, 20.08.2009 | |||
| ДВУХТАКТНЫЙ ДВИГАТЕЛЬ ВНУТРЕННЕГО ГОРЕНИЯ | 1929 |
|
SU13752A1 |

