Настоящее изобретение относится к областям биологии и биотехнологии, в частности к молекулярной генетике, и конкретно относится к получению генетических конструкций, трансгенных клеток и новых рекомбинантных белков на основе генетической последовательности модифицированного гена GDNF для терапии ишемических инсультов, нейродегенеративных заболеваний и травматической гибели нервных клеток.
Нейротрофические факторы относятся к белкам, выполняющим функцию выживания и стимуляции фенотипической дифференцировки нервных и/или глиальных клеток (Varon, 1979; Thoenen, 1985). Нейротрофические факторы могут быть полезны при лечении нейродегенеративных заболеваний. В частности, терапия с применением GDNF признана эффективной при лечении заболеваний, связанных с дегенерацией дофаминэргических нейронов, например при болезни Паркинсона. Эффект данного применения заключается не только в увеличении нейротрансмиссии дофаминэргических синапсов в полосатом теле, но также ослаблении дегенеративных процессов нигростратиарных проекций (Aoi, 2000; Date, 1998; Connor, 2001; Jollivet, 2004; Akerud, 2001). Экспрессия мРНК GDNF обнаружена во многих областях центральной и периферической нервных систем, а также в других тканях и органах. Было найдено, что уровень мРНК GDNF быстро возрастает при повреждении нервной системы (Hoke, 2000). GDNF принадлежит к семейству трансформирующего фактора роста бета (TGFβ). Он имеет выраженное нейропротекторное действие на дофаминэргические нейроны и мотонейроны спинного мозга (Cheng, 2002), стимулирует рост аксонов (Iannotti, 2003). Исследования лекарственных препаратов на основе рекомбинантного GDNF проводились до 2009 года. Однако резкое снижение эффективности такой терапии при использовании GDNF и побочные эффекты вынудили компании отказаться от данного фактора, и проект был законсервирован. В настоящее время исследуются особенности строения гена,процессы сплайсинга и строение белка нейротрофического фактора не только человека, но и других организмов. У человека и грызунов, ген GDNF представлен двумя матричными РНК, которые образуются посредством альтернативного сплайсинга: полноразмерный транскрипт [pre-(а)pro-GDNF] и короткий транскрипт [pre-(в)pro-GDNF], у которого не хватает 78 пн в участке зашифрованного pro-домена (Suter-Crazzolara, 1994; Trupp, 1995; Matsushita, 1997; Grimm, 1998). Наличие различных сплайс вариантов может говорить об их различных функциях. В работах, выполненных в лаборатории Mart Saarma, было показано, что gdnf как с полноразмерной (α)pro областью, так и укороченной (β)pro областью секретируется в клетках нейрального происхождения. Однако было обнаружено различие транспортировки двух данных белков. Белок, кодируемый pre-(α)pro-GDNF, секретируется обычным путем, проходя через аппарат Гольджи, и транспортируется из клетки по секреторному пути. В свою очередь белок, кодируемый pre-(β) pro-GDNF, преимущественно локализуется в secretogranin II и Rab3A-позитивных секреторных везикулах и после стимуляции с помощью добавления в среду KCl движется по более быстрому секреторному пути. (Lonka-Nevalaita, 2010). Мы предполагаем, что транспортировка GDNF из клеток двумя различными путями может быть неслучайна. GDNF с «нормальной» pro-областью постоянно секретируется из клеток и, возможно, необходим для постоянного поддержания жизнеспособности нервных клеток. Другой природный сплайс-вариант с «уменьшенной pro-областью» экспрессируется в стрессовых и экстремальных условиях. Он, возможно, необходим как продукт экстренного значения в случае массовой гибели нейронов, при нейродегенеративных заболеваниях, имешических инсультах или травмах. Таким образом, используемый в предыдущих разработках рекомбинантный GDNF, полученный от «нормального» GDNF мог не обладать ожидаемыми эффектами из-за того, что исходно использовали GDNF необходимый для поддержания жизнеспособности нейронов.
Технической задачей заявленной группы изобретений является повышение эффективности нейтрофического фактора GDNF, способного при попадании в клетки или ткани млекопитающих продуцировать активный специфический GDNF, стимулирующий нейральную дифференцировку прогениторных клеток и образование нейральных отростков, используемых при терапии нейральных травм, ишемических инсультов и нейродегенеративных заболеваний.
Поставленная техническая задача и достигаемый технический результат, заключающийся в повышении активности нейтрофического фактора GDNF и расширении его функциональных возможностей при использовании в терапии достигаются заявленной группой изобретения.
Одним из изобретений заявленной группы являются генетические полинуклеотидные последовательности ДНК искусственно модифицированного GDNF с делетированной pro областью
(ttcccgctgcccgccggtaagaggcctcccgaggcgcccgccgaagaccgctccctcggccgccgccgcgcgcccttcgc gctgagtagtgactcaaatatgccagaggattatcctgatcagttcgatgatgtcatggattttattcaagccaccattaaaagactgaaaagg), которые при попадании в клетки или ткани млекопитающих в составе стандартных векторов продуцируют активный специфический GDNF стимулирующий нейральную дифференцировку прогениторных клеток и образование нейральных отростков, что пригодно для терапии нейральных травм, ишемических инсультов и нейродегенератиных заболеваний.
Другим изобретением заявленной группы является вектор, содержащий указанную выше нуклеотидную последовательность ДНК искусственно модифицированного GDNF для экспрессии в прокариотических и эукариотических клетках и тканях млекопитающих in vivo.
Еще одним изобретением заявленной группы, обеспечивающим достижение указанного технического эффекта является модифицированный GDNF, имеющий нуклеотидную последовательность ДНК, указанную выше, встроенный в вектор экспрессии и продуцирующий в клетках и тканях активный специфический GDNF, стимулирующий нейральную дифференцировку стволовых и прогениторных клеток с образованием нейральных отростков, что пригодно для терапии нейральных травм, ишемических инсультов и нейродегенеративных заболеваний.
В заявленном изобретении синтезированы искусственные последовательности GDNF человека с делетированной pro-областью размером 174 пн. Одна последовательность размером в 459 пн, другая размером в 402 пн (отличие двух последовательностей в наличии/отсутствии сигнальной последовательности размером в 57 пн). Полученный в результате экспрессии этих последовательностей нейротрофический фактор обладает повышенной способностью индуцировать нейральную дифференцировку незрелых клеток и стимулировать образование нейральных отростков. Модификации GDNF, встроенные в любые носители (плазмиды, вирусы и т.д) возможно использовать как генетический материал при введении в зону гибели нейронов для восстановительной терапии. Также возможно получать трансгенные клеточные культуры, продуцирующие модифицированный нейротрофический фактор GDNF для клеточной терапии, и выделять белок из трансфицированных E.coli или клеток млекопитающих и использовать его для терапии.
В настоящее изобретение включено получение формы белка GDNF на основе модифицированного гена GDNF размером 459 пн, которая чрезвычайно активно стимулирует образование нейральных отростков и является сильным нейральным индуктором.
Генетические последовательности модифицированного (гена) нейтрофического фактора GDNF глиальной клеточной линии с полностью делетированной pro-областью (регионом), получают методом ПЦР реакции (полимеразной цепной реакции) с использованием прямого праймера Xhol-Pre-GDNF(F) и обратного праймера GDNF-BamHI(R) и содержащие аминокислотные последовательности одну размером в 459 пн, а другую в 402 пн в соответствии с фиг. 1.
Было обнаружено, что, действительно, уменьшение pro-области изменяет путь транспортировки нейротрофического фактора GDNF. Кроме того, GDNF транспортируемый через везикулы и в обход аппарат Гольджи, обладает более высокой активностью в качестве нейрального индуктора. На основе данного исследования мы получили модифицированные GDNF с полностью делетированной pro-областью и обнаружили значительное улучшение свойств модифицированного GDNF в качестве нейрального индуктора. Полученные нами конструкции и модифицированные генные продукты могут быть применены в терапии при необходимости стимуляции регенеративных процессов в нервной системе.
Ниже представлены конкретные примеры, иллюстрирующие изобретение, но не ограничивающие его.
Пример 1.
А. Была получена последовательность с делетированной pro-областью GDNF. На фигуре 1 представлена данная последовательность.
Серым цветом выделена N- концевая сигнальная последовательность (pre-область) и последовательность, соответствующая mature-GDNF.
Для получения с помощью метода ПЦР фрагмента ДНК размером 459 пн, содержащего необходимую часть кодирующей последовательности GDNF, были использованы праймеры. Прямой праймер содержал последовательность pre- области и часть последовательности mature-GDNF и включал сайт клонирования Xho1.
Ниже представлена последовательность праймера XhoI-Pre-Gdnf (F).
5′-CTCGAGCCACCATGAAGTTATGGGATGTCGTGGCTGTCTGCCTGGTGCTGCTC CACACCGCGTCCGCCGTCACCAGATAAACAAATGGCAGTGCTTCCTAGAA-3′
В качестве обратного праймера использовали GDNF-BamHI (R), также использованный для получения конструкции mature-GDNF/pEGFP-N1.
Используя полученные праймеры методом ПЦР была получена необходимая генетическая последовательность. Последовательность гена нейротрофического фактора человека GDNF согласно базе данных UniProtKB Р39905 первой изоформе Р39905-1 была искусственно синтезирована. Клонирование осуществлялось с помощью ПЦР реакции, точнее с амплификацией ПЦР продуктов с последующим встраиванием в коммерческий вектор pGEM-T Easy, согласно протоколу, полученная последовательность GDNF далее использовалась в качестве матрицы.
Программа амплификации с помощью ПЦР: 94°С - 1,5 мин, 94°С - 15 сек, 57-62°С - 20 сек, 72°С - 15 сек, 72°С - 10 мин, количество циклов: 30. Был получен ПЦР фрагмент Pre-(m)GDNF (459 пн). Данный фрагмент был встроены в вектор pGEM-T Easy. Конструкция была проверена с помощью рестриктного анализа и секвенирования. С использованием рестриктаз: XhoI, HindIII и BamHI, данная последовательность была встроена в вектор EGFP-N1 (Clontech), по соответствующим сайтам рестрикции. В результате переклонирования была получена конструкция Pre-GDNF/EGFP-N1.
Б. Была получена последовательность с делегированными pre- и pro-областями GDNF. На фиг. 2 представлена данная последовательность.
Серым цветом выделена используемая последовательность GDNF.
Для получения с помощью метода ПЦР фрагмента ДНК размером 402 пн, содержащего необходимую часть кодирующей последовательности GDNF, были использованы праймеры. Прямой праймер содержал последовательность pre-области и часть последовательности mature-GDNF и включал сайт клонирования XhoI.
Ниже представлена последовательность праймера XhoI-Pre-Gdnf (F).
5′-CTCGAGCCACCATGAAGTTATGGGATGTCGTGGCTGTCTGCCTGGTGCTGC TCCACACCGCGTCCGCCGTCACCAGATAAACAAATGGCAGTGCTTCCTAGAA-3′
В качестве обратного праймера использовали GDNF-BamHI.
Используя полученные праймеры методом ПЦР была получена необходимая генетическая последовательность. Последовательность гена нейротрофического фактора человека GDNF согласно базе данных UniProtKB Р39905 первой изоформе Р39905-1 была искусственно синтезирована. Клонирование осуществлялось с помощью ПЦР реакции, с амплификацией ПЦР продуктов с последующим встраиванием в коммерческий вектор pGEM-T Easy, согласно протоколу. Полученная последовательность GDNF далее использовалась в качестве матрицы.
Программа амплификации с помощью ПЦР: 940С-1,5 мин, 940С - 15 сек, 57-620С- 20 сек, 720С - 15 сек, 720С - 10 мин, количество циклов: 30. Был получен ПЦР фрагмент Pre-(m)GDNF (459 пн). Данный фрагмент был встроен в вектор pGEM-T Easy. Конструкция была проверена с помощью рестриктного анализа и секвенирования. С использованием рестриктаз: XhoI, HindIII и BamHI, данная последовательность была встроена в вектор EGFP-N1 (Clontech), по соответствующим сайтам рестрикции. В результате переклонирования была получена конструкция mGDNF/EGFP-N1.
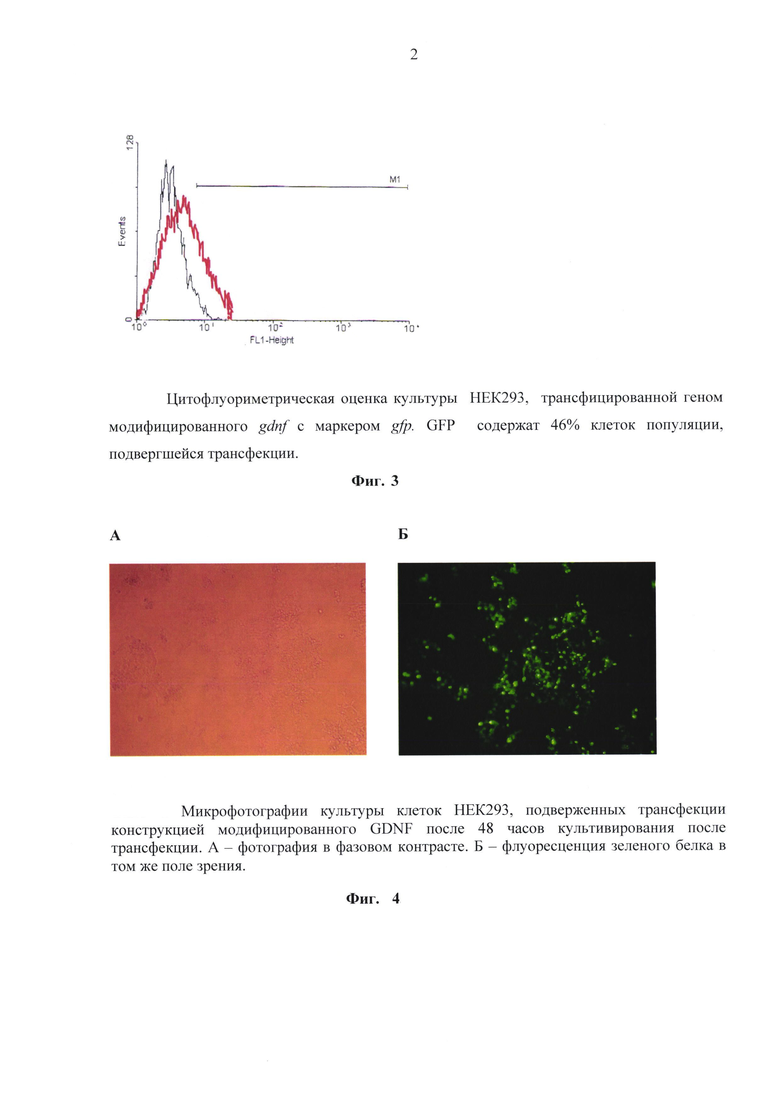
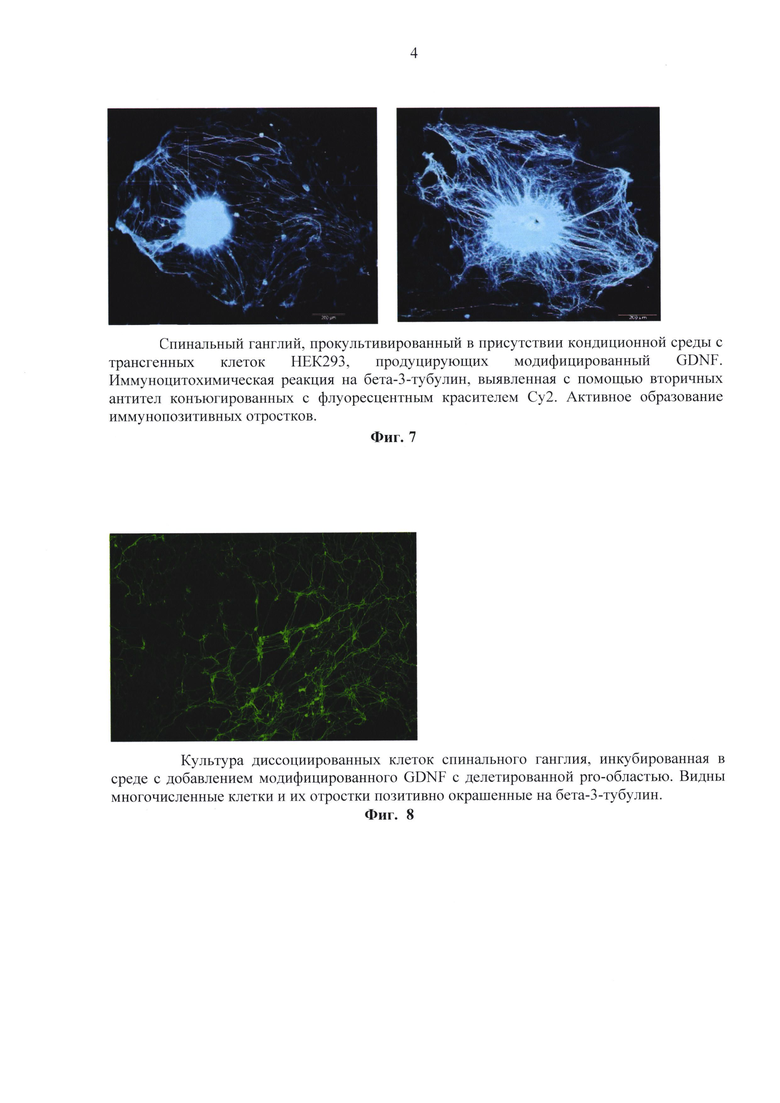
Конструкции трансфицировали в клетки линии НЕК293. Для культивирования использовали среду DMEM (ПанЭко, Россия) с добавлением 10% фетальной сыворотки (Perbio HyClone, США), L-глутамина (ПанЭко, Россия) и 4% гентамицина (ПанЭко, Россия). Культуры клеток инкубировали при 37оС в инкубаторе при уровне СО2 5%. Клетки росли на площади 25 см2, во флаконах («Costar»). После нарастания культуры клеток НЕК293 до концентрации 1.5 × 105 клеток / мл, клетки подвергали трансфекции полученными конструкциями. Трансфекцию осуществляли с помощью реагента ExGene 500(Fermentas, R0511). Селекция клонов, несущих встроенные конструкции, проводили с помощью гентамицина (G418, Invitrogen # 15750045), в течение десяти дней, после чего клоны устойчивые к G418 были исследованы на наличие встроенного гена с помощью ПЦР анализа. В ходе получения трансгенных культур было обнаружено, что трансгенные клеточные линии HEK293/(m)GDNF и HEK/Pre-GDNF обнаруживают свечение маркера gfp. С помощью цитофлуориметра была проведена оценка интенсивности свечения модифицированного GDNF. На фиг. 3 представлен результат исследования. Было обнаружено, что культура клеток подверженная трансфекции плазмидной ДНК, содержащей в своем составе модифицированный gdnf с маркерным gfp дает 46% GFP положительных клеток. На фиг. 4 представлены результаты визуализации свечения в культуре.
Наличие экспрессии модифицированного GDNF и его секреции из трансфецированных клеток было подтверждено с помощью Вестерн-блот гибридизации. Этим методом были исследованы лизаты трансгенных клеток НЕК293, продуцирующих модифицированные белки GDNF.с использованием антител к GDNF. Было показано, что все конструкции синтезируют модифицированные белки GDNF. Следует отметить, что во всех экспериментах pre-GDNF синтезировался значительно менее интенсивно по сравнению с mGDNF (фиг. 5).
При исследовании кондиционных сред, полученных при культивации трансгенных клеточных культур НЕК293, с помощью Вестерн-блот гибридизации с использованием антител к GDNF было обнаружено, что модифицированные белки GDNF секретируются в среду (фиг. 6).
Далее трансгенные культуры рассевали на площади 25 см2, по достижению плотности около 60% общей площади питательная среды (DMEM (ПанЭко, Россия), 10% фетальной сыворотки крови (Perbio HyClone, США), L-глутамина (ПанЭко, Россия) и 4% гентамицина (ПанЭко, Россия) была заменена на новую, и через 24 часа культивирования при 37°С кондиционированную среду собирали для последующего анализа. Кондиционированная среда с трансгенных клеточных линий была профильтрована через 0,22 нм фильтр.
Пример 2
Эмбрионы крысы Е15 были использованы для получения эмбриональных спинальных ганглиев. Эмбрионы помещали в 100 мм чашку Петри с рабочей средой (DMEM F12 (ПанЭко, Россия), при добавлении 10% фетальной сыворотки крови (Perbio HyClone, США), L-глутамина (ПанЭко, Россия) и 4% гентамицина (ПанЭко, Россия)). Спинномозговой ствол с расположенными на нем ганглиями был дважды промыт раствором Хенкса (ПанЭко, Россия) с гентамицином (100 мкг/мл). Спинальные ганглии выделяли с использованием бинокулярной лупы. Выделенные ганглии перекладывали в капли среды (100 мкл), нанесенной на дно 35-мм чашки Петри. Правильность выделения спинальных ганглиев проверяли под микроскопом. Ганглии в течение первых 3-4 часов находились в ростовой среде DMEM F12 (ПанЭко, Россия), при добавлении 10% фетальной сыворотки крови (Perbio HyClone, США), L-глутамина (ПанЭко, Россия) и 100 мкг/мл гентамицина (ПанЭко, Россия), 0,8 % глюкозы (ПанЭко, Россия) и 2 мкм HEPES(Perbio HyClone, США). После прикрепления ганглиев ко дну, ростовую среду заменяли на кондиционированные среды трансгенных клеточных культур: HEK293/(m)GDNF, HEK293/Pre-GDNF. Контролями в данном эксперименте выступали: 1) среда культивирования нетрансфицированны клеток линии НЕК293 (среда DMEM (ПанЭко, Россия), при добавлении 10% фетальной сыворотки крови (Perbio HyClone, США), L-глутамина (ПанЭко, Россия) и 4% гентамицина (ПанЭко, Россия); 2) среда с добавлением рекомбинантного GDNF (SantaCruz); 3) некондиционированная полная питательная среда. Цельные спинальные ганглии инкубировали в течение 4-10 суток (5% СО2 инкубаторе при 37оС) в 200 мкл среды. Кондиционированные и контрольные среды меняли каждые 2-3 дня.
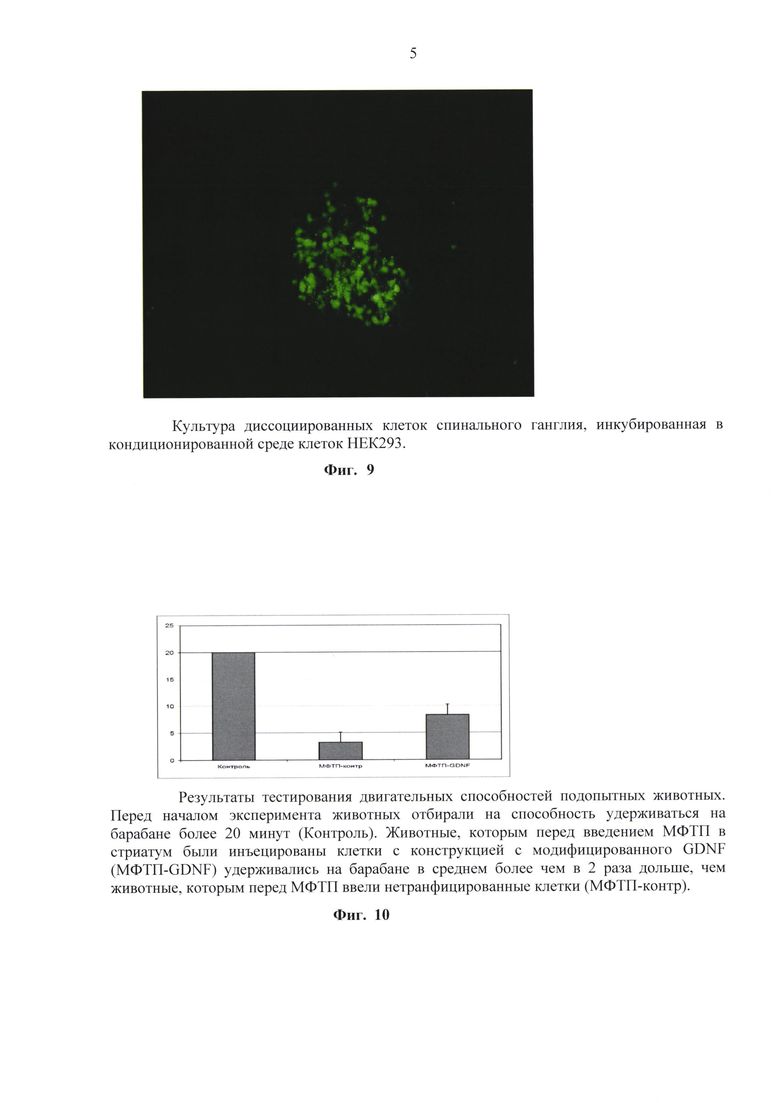
Наблюдали появление отдельных отростков уже через 12 часов после помещения в кондиционированную среду с mGDNF (фиг. 7). Через сутки наблюдались отростки, образовавшиеся у спинальных ганглиев, культивированных с добавлением pre-GDNF. Причем следует отметить, что при добавлении кондиционной среды с mGDNF отростки образовывались на самых ранних этапах и через сутки и далее имели большую интенсивность и разветвленность. Большинство отростков росли по дну культуральной чашки, но наблюдались и выходящие из ганглия на небольшом расстоянии от дна и идущие напрямую к более или менее отдаленной точке на дне чашки, находясь на весу. Чтобы проанализировать нейрональные свойства данных отростков, был проведен иммуногистохимический анализ с использованием антител на нейроспецифический фактор β3-тубулин. При окраске на β3тубулин использовали кроличьи поликлональные антитела, разведенные в соотношении 1:100 раствором PBS содержащим 0,3% детергента Triton X100 и 2% нормальной сыворотки осла для блокирования неспецифического связывания.
Пример 3
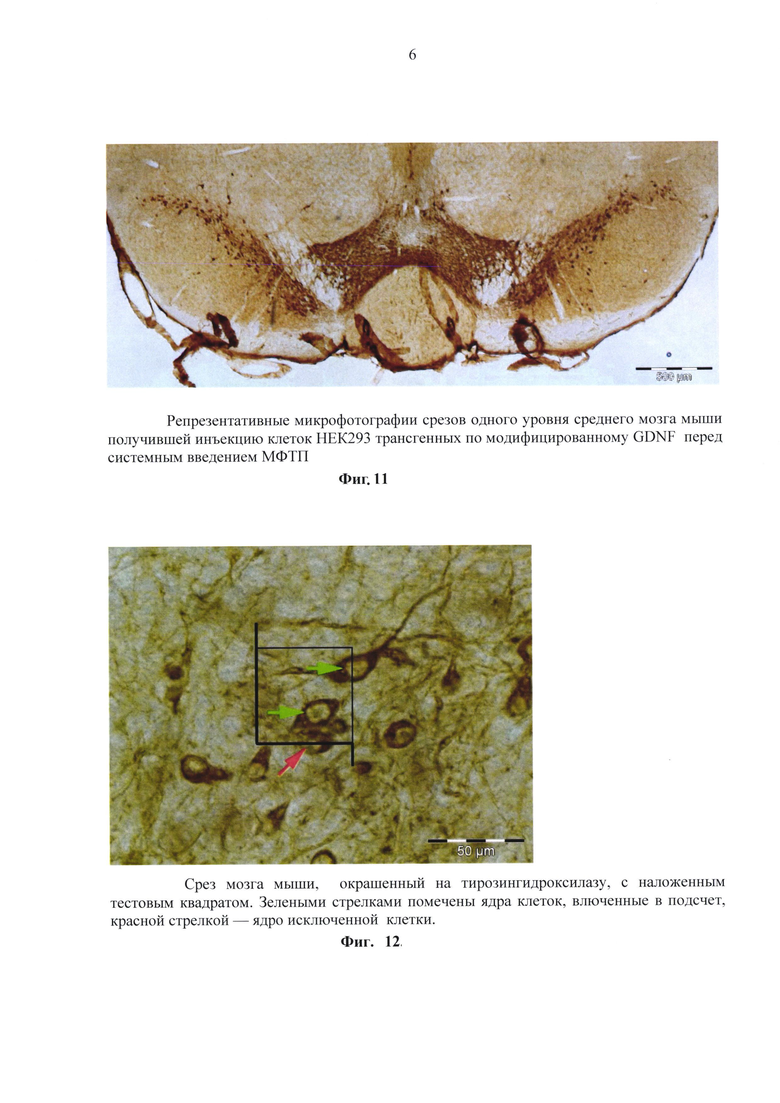
В другой серии экспериментов ганглии разделяли на 3-5 фрагментов и помещали в лунку 4-луночой плашки фрагменты одного ганглия. Эксперимент, в котором цельные спинальные ганглии подвергались механически диссоциации дали сходные результаты. Клетки спинальных ганглиев помещались в 4-луночные плашки по 50 000 клеток на лунку. Далее проводилась инкубация с кондиционированной средой с трансгенных клеток продуцирующих модифицированные GDNF. Клетки диссоциированных спинальных ганглиев располагались на дне культуральных чашек поодиночке или группами. Часть клеток позитивно окрашивались на бета-3-тубулин. В культурах, инкубированных со средой, кондиционированной трансгенными клетками содержащими модифицированный GDNF (фиг. 8) клетки, позитивно окрашенные на бета-3-тубулин, присутствовали в большом количестве и активно образовывали отростки. Все отростки интенсивно ветвились. Ветвления начинались в большинстве случаев в 150-200 микрометрах от границы ганглия.
Было показано, что модифицированный GDNF обладает чрезвычайно высокими способностями стимулировать образование нейральных отростков у эмбрионального спинального ганглия, что не только сопоставимо со свойствами рекомбинантного GDNF (recGDNF), но и превосходит их.
Для оценки влияния модифицированного GDNF на способность стимулировать образование нейральных отростков был произведен подсчет аксонов с использованием рамки с двумя радиусами Мерца (Jack W. Lipton et al., Neuropharmacology. 2008). Данные этих измерений показывают, что модифицированные GDNF с делетированной pro-областью позитивно воздействуют на длину отростков. При этом интенсивность воздействия модифицированных-GDNF значительно превышает таковую рекомбинантного GDNF.
Пример 4
Эффективность модифицированного GDNF на модели Паркинсона.
Исследовали влияние введения клеток линии НЕК293 в каудатум-путамен на нарушение двигательной активности мышей после системного введения нейротоксина МФТП (1-метил-4-фенил-1,2,3,6-тетрагидропиридин - нейротоксин, вызывающий массовую гибель дофамин-эргических нейронов, обычно применяется для моделирования болезни Паркинсона). Двигательную активность исследовали с помощью теста RotaRod - (Удерживание на вращающемся барабане). Удерживание животных на вращающемся стержне небольшого диаметра один из распространенных способов оценки нарушений движений. Измеряют время в течение, которого животное удерживается на барабане прибора RotaRod Advanced фирмы TSE systems (США) при скорости его вращения 15 оборотов в минуту. Для опытов отбирали животных способных удерживаться на барабане непрерывно в течение 20 минут. Проводили первое тестирование непосредственно перед введением клеток. Животные подвергались поведенческим тестированиям спустя 14 суток после воздействия Результаты тестирования представлены на фиг 10. Животные, которым перед введением МФТП в стриатум были инъецированы клетки, продуцирующие модифицированный GDNF с делетированной pro-областью удерживались на барабане в среднем более чем в 2 раза дольше, чем животные, которым перед МФТП ввели нетранфицированные клетки (МФТП-контр). Таким образом, введение в стриатум животных перед инъекцией МФТП клеток, продуцирующих новую форму модифицированного GDNF предотвращает развитие двигательных нарушений, обусловленных гибелью дофаминэргических нейронов. Исследуемая форма нейротрофического фактора обладает нейропротективными свойствами превышающая ранее исследованные свойства рекомбинантного GDNF.
Перед началом эксперимента животных отбирали на способность удерживаться на барабане более 20 минут (Контроль). Животные, которым перед введением МФТП в стриатум были инъецированы клетки с конструкцией с модифицированного GDNF (МФТП-GDNF), удерживались на барабане в среднем более чем в 2 раза дольше, чем животные, которым перед МФТП ввели нетранфицированные клетки (МФТП-контр).
Пример 5
Исследование влияния модифицированного GDNF на жизнеспособность дофаминэргических нейронов черной субстанции.
Для исследования влияния введения трансгенных клеток с модифицированным GDNF в каудатум-путамен подопытных мышей на сохранность дофаминэргических нейронов черной субстанции проводили подсчет тирозингидроксилаза иммунопозитивных клеток в компактной части этого ядра на срезах среднего мозга животных спустя 14 суток после введения нейротоксина МФТП. Опытным животным за 48 часов перед введением токсина инъецировали трансгенные клетки HEK293 с модифицированным GDNF. Контрольным животным инъецировали клетки HEK293 трансфицированные геном зеленого флуоресцентного белка.
Фиксация материала. Животных усыпляли внутрибрюшинным введением летальной дозы уретана. Кровеносную систему промывали транскардиально сначала раствором фосфатного буфера (рН 7,4) в физиологическом растворе (PBS) а затем фиксирующим раствором: 4%-ным раствором параформальдегида в PBS. Фиксированный таким образом мозг извлекали из черепной коробки и погружали в упомянутый выше фиксирующий раствор и держали в холодильнике при +4оС 12 часов, после чего мозг пропитывали в 30%-ном растворе сахарозы в PBS 24 часа при 4оС. В дальнейшем мозг замораживали и резали на замораживающем микротоме. Фронтальные срезы толщиной 40 микрометров собирали в PBS. Было собрано по 4 серии срезов каждого мозга. Таким образом, в каждую серию срезов попадал каждый 4-й срез. Промежуток между соседними срезами в серии равнялся 160 микрометров. Срезы помещали в антифриз и хранили при -20оС до процедуры окрашивания.
Иммуногистохимическое окрашивание на тирозингидроксилазу. Для окрашивания срезы после отмывки от антифриза в 3х сменах PBS (по 5 минут) помещали в раствор первичных антител: моноклональных мышиных антител против тирозингидроксилазы, разведенных в соотношении 1:200 в растворе PBS с добавлением 2% нормальной сыворотки лошади, 0.3% детергента Triton X100 и 0.01% азида натрия.
В растворе первичных антител срезы содержали с перемешиванием при 4-8 оС в течение 12 часов. Далее после троекратной промывки в PBS срезы на 1 час погружали в раствор биотинилированных антител лошади против иммуноглобулина мыши, разведенных PBS в соотношении 1:100 с добавлением 0,3% Triton X-100 при комнатной температуре. После этого срезы троекратно промывали в PBS и помещали в раствор ABC комплекса в PBS при разведении 1:200 также на 1 час. После троекратной промывки проводили стандартную реакцию на пероксидазу с помощью 0,03% раствора диаминобензидина в PBS с добавлением 0,01% перекиси водорода. Окрашенные срезы помещали на предметные стекла, покрывали 50% глицерином и покровным стеклом.
Подсчет тирозингидроксилаза-позитивных клеток. Количественный анализ тирозингидроксилаза-позитивных клеток на иммуногистохимически окрашенных срезах проводили с помощью микроскопа Olympus IX81 снабженного моторизованным предметным столиком Märzhäuser motorised stage управляемым с компьютера. и цифровой фотокамерой Olympus DP72. Подсчет клеток осуществляли с монитора компьютера с использованием программы “Cell*”. Сначала при малом увеличении (объектив 10х) получали обзорное изображение участка среза, с комплексом ядер среднего мозга, содержащих тирозингидроксилаза-позитивные клетки (фиг. 11): компактную часть черной субстанции (SNC substantia nigra, compact part), вентральную область покрышки (VTA - ventral tegmental area).
Затем подсчитывали количество тирозингидроксилаза-позитивных клеток в тестовом квадрате, который перемещали, отслеживая его перемещение на обзорном изображении. Положение тестового квадрата со стороной 50 микрометров меняли с шагом 150 микрометров по оси Х и 100 микрометров по оси Y в пределах вентральной части среднего мозга. Оператор, неинформированный о нумерации групп животных, подсчитывал количество ядер тирозингидроксилаза-иммунопозитивных клеток в тестовом квадрате в соответствии со стандартными критериями подсчета. Клетку, окрашенную на тирозингидроксилазу, включали в подсчет, если ее ядро (округлая не окрашенная структура, окруженная окрашенной цитоплазмой) лежало внутри тестового квадрата или касалось двух его смежных сторон (фиг. 12) и не касалось двух других сторон отмеченных на фиг. 10 более толстым контуром. Так как просчитываемая фракция объема исследованной области мозга составляла 1/16 исследуемого объёма, для определения общего количества тирозингидроксилаза-позитивных клеток подсчитанное количество умножали на 16. Подсчитывали количество тирозингидроксилаза-иммунопозитивных клеток, лежащих в пределах компактной части черной субстанции.
Исследование показало, увеличение количества тирозингидроксилаза-позитивных нейронов в компактной части черной субстанции животных, в куадатум-путамен которых были инъецированы клетки трансгенные по модифицированному GDNF. Оно было в 2 раза больше, чем у животных, получивших инъекцию клеток НЕК293, не содержащих модифицированный GDNF (фиг. 13).
Таким образом, можно утверждать, что введение модифицированного GDNF в зону гибели нейронов, уменьшает гибель дофамин-эргических нейронов черной субстанции и улучшает восстановительные процессы в зоне повреждения.
Значимой проблемой для жизнеспособности и трудоспособности человека является массовая гибель нервных клеток вследствие нейродегенеративных заболеваний, травм, а также ишемических инсультов. Эти явления во многих случаях ведут к инвалидности и гибели пациента. К сожалению известные на данный момент методы терапии в большинстве случаев не влияют на течение болезни и не предотвращают гибель нейронов и не способствуют их восстановлению. GDNF предотвращает гибель дофаминергических нейронов (а значит и может быть полезен для лечения болезни Паркинсона), а также участвует в поддержании мотонейронов (Airaksinen et al., 2002). GDNF (глия-производный нейротрофический фактор) играет ключевую роль в развитии и поддержании жизнеспособности клеток центральной и периферической нервных систем. Полученная модификация, GDNF транспортируется из клетки везикулярным способом в обход аппарата Гольджи и обладает лучшими нейропротективными и нейроиндуктивными характеристиками, чем известный рекомбинантный белок GDNF.
Оригинальность предложенного изобретения заключается в получении модифицированной последовательности GDNF, продукт которой обладает повышенной способностью в качестве нейрального индуктора и стимулятора образования нейральных отростков. Транспортировка модифицированного GDNF через везикулы (минуя аппарат Гольджи) не подвергает GDNF фосфолированию и гликозилированию, что улучшает его индукционные свойства.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ, СОДЕРЖАЩАЯ hGDNF ПОД КОНТРОЛЕМ ТЕМПЕРАТУРОЧУВСТВИТЕЛЬНОГО ПРОМОТОРА ДЛЯ РЕГУЛИРУЕМОЙ ЭКСПРЕССИИ НЕЙРОТРОФИЧЕСКОГО ФАКТОРА КАК В КЛЕТКАХ, ТАК И НЕПОСРЕДСТВЕННО В ОРГАНИЗМЕ МЛЕКОПИТАЮЩИХ | 2012 |
|
RU2527169C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА В ДОФАМИНЭРГИЧЕСКИЕ НЕЙРОНЫ | 2023 |
|
RU2823290C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК МЛЕКОПИТАЮЩИХ, МОДИФИЦИРОВАННЫХ ГЕНОМ ФАКТОРА РОСТА НЕРВОВ ЧЕЛОВЕКА | 2011 |
|
RU2458984C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПОПУЛЯЦИИ СТРОМАЛЬНЫХ КЛЕТОК ЖИРОВОЙ ТКАНИ, ИНДУЦИРОВАННЫХ К НЕЙРАЛЬНОЙ ДИФФЕРЕНЦИРОВКЕ | 2008 |
|
RU2396345C1 |
| ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ ДЛЯ СТИМУЛЯЦИИ ПОСТТРАВМАТИЧЕСКОЙ РЕГЕНЕРАЦИИ ПЕРИФЕРИЧЕСКИХ НЕРВОВ | 2019 |
|
RU2719013C1 |
| СПОСОБ СТИМУЛИРОВАНИЯ РЕГЕНЕРАЦИИ СПИННОГО МОЗГА С ПОМОЩЬЮ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ КЛЕТОК КРОВИ ПУПОВИНЫ ЧЕЛОВЕКА | 2013 |
|
RU2521225C2 |
| Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией NGF для терапии нейродегенеративных заболеваний | 2023 |
|
RU2829363C1 |
| Способ получения глиальных производных индуцированных плюрипотентных стволовых клеток с повышенной экспрессией BDNF для терапии ишемического инсульта | 2023 |
|
RU2829361C1 |
| 2-((1H-1,2,4-Триазо-3-ил)тио)-3-метил-6-(проп-1-ен-2-ил)циклогекс-3-ен-1-ол в качестве противопаркинсонического средства | 2022 |
|
RU2796729C1 |
| ПОЛУЧЕНИЕ ТЕРМИНАЛЬНО ДИФФЕРЕНЦИРОВАННЫХ ДОФАМИНЕРГИЧЕСКИХ НЕЙРОНОВ ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА | 2004 |
|
RU2345133C2 |



Изобретение относится к области биохимии. Заявлена кДНК природного гена GDNF, искусственно модифицированная удалением pro-области, которая при попадании в клетки или ткани млекопитающих в составе стандартных векторов продуцирует активный специфический GDNF. Кроме того, изобретение относится к вектору экспрессии, содержащему указанную кДНК, а также к активному специфическому GDNF, кодированному указанной кДНК. При трансляции в клетках и тканях указанный GDNF стимулирует нейральную дифференцировку стволовых и прогениторных клеток с образованием нейральных отростков. Изобретение позволяет повысить активность GDNF в качестве нейрального индуктора и стимулятора образования нейральных отростков и может применяться при терапии нейродегенеративных заболеваний, травматических нарушений иннервации, а также ишемических инсультов головного мозга млекопитающих, в том числе человека. 3 н.п. ф-лы, 13 ил., 5 пр.
1. кДНК природного гена GDNF, искусственно модифицированная удалением pro-области
(ttcccgctgcccgccggtaagaggcctcccgaggcgcccgccgaagaccgctccctcggccgccgccgcgcgcccttcgc gctgagtagtgactcaaatatgccagaggattatcctgatcagttcgatgatgtcatggattttattcaagccaccattaaaagactgaaaagg), которая при попадании в клетки или ткани млекопитающих в составе стандартных векторов продуцирует активный специфический GDNF, стимулирующий нейральную дифференцировку прогениторных клеток и образование нейральных отростков, что пригодно для терапии нейральных травм, ишемических инсультов и нейродегенеративных заболеваний.
2. Вектор, содержащий кДНК по п.1, для экспрессии в прокариотических и эукариотических клетках и в тканях млекопитающих in vivo.
3. Активный специфический GDNF, кодированный кДНК по п.1, при трансляции в клетках и тканях стимулирующий нейральную дифференцировку стволовых и прогениторных клеток с образованием нейральных отростков, что пригодно для терапии нейральных травм, ишемических инсультов и нейродегенеративных заболеваний.
| PICCININI E | |||
| et al., Glial cell line-derived neurotrophic factor: Characterization of mammalian posttranslational modifications, Annals of Medicine, 2013, vol.45, pp.66-73 | |||
| YAN CHEN, Effects of glial line-derived neurotrophic factor (GDNF) on stem/progenitor cell proliferation and differentiation, 2005, University of Kentucky Doctoral |

