Эта заявка основана на предварительной заявке США № 60/741640, зарегистрированной 2 декабря 2005 г., и предварительной заявке США № 60/822597, зарегистрированной 16 августа 2006 г., раскрытие которых приведено полностью в данном документе посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к составам и способам, полезным для диагностики и лечения заболеваний и нарушений, связанных с передачей сигналов цитокинами.
УРОВЕНЬ ТЕХНИКИ
Различные заболевания и нарушения связаны с воспалением. Воспаление представляет собой процесс, связанный с рекрутированием воспалительных клеток (например, лейкоцитов) в область повреждения или инфицирования. Однако чрезмерное или неадекватное воспаление может обладать отрицательным действием. Аутоиммунные нарушения, например, часто вызывают воспаление, приводя к разрушению нормальных тканей тела. Воспаление, кроме того, связано с раком. См., например, Coussens et al. (2002) Nature 420:860-867. Например, связанное с воспалительной болезнью кишечника (IBD) хроническое воспаление строго коррелирует с кишечным канцерогенезом. Во время воспалительной реакции определенные воспалительные клетки синтезируют вещества, которые стимулируют ангиогенез, снижают антиопухолевую активность цитоксичных T-клеток и уменьшают мутации ДНК, таким образом, создавая окружение ускоренной опухолевой прогрессии Id.
IL-23 представляет собой гетеродимерный цитокин, который играет доминантную роль при аутоиммунных/воспалительных нарушениях, и в частности при хроническом воспалении. Например, в исследованиях на мышах показано, что IL-23 чрезвычайно важен для развития экспериментального аллергического энцефаломиелита (аутоиммунное воспаление мозга), который является моделью рассеянного склероза; коллаген-индуцированного артрита, который является моделью ревматоидного артрита; и гиперчувствительности замедленного типа. IL-23 также функционирует для поддержания стабилизированного колита (форма IBD). Трансгенная экспрессия IL-23 приводит к системной воспалительной реакции, а нарушение регуляции IL-23 приводит к экзематозной болезни кожи (воспалительное состояние кожи). IL-23 активирует специфичную популяцию Т-клеток (ThIL-17 клетки), которые в свою очередь вызывают продукцию ThIL-17 и провоспалительных цитокинов. Для обзора роли IL-23 при воспалении и аутоиммунной реакции см., например, Hunter (2005) Nat. Rev. Immunol. 5:521-531; и Holscher (2005) Curr. Opin. Invest. Drugs 6:489-495. Было показано, что IL-23 также ускоряет опухолевый рост путем увеличения ангиогенеза и уменьшения опухолевой инфильтрации цитотоксическими Т-клетками. Langowski et al. (2006), Nature 442:461-465.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Представлены составы и способы, полезные для диагностирования и лечения воспалительных нарушений и аутоиммунных нарушений (например, псориаз). Далее представлены составы и способы, полезные для изменения передачи сигнала IL-23 или IL-22. Эти и другие варианты осуществления изобретения представлены в данном документе. Настоящее изобретение частично основано на идентификации (определении) пути передачи сигнала, в котором IL-23 действует через IL-22, стимулируя экспрессию IL-22 в недавно открытой подгруппе Т-клеток-хелперов (Th клетки), то есть ThIL-17 линии. В одном аспекте, представлено антитело, которое специфически связывается с IL-22, в котором антитело представляет собой (a) антитело, производимое гибридомой, выбранной из 3F11.3 (ATCC каталог № PTA-7312), гибридомой 11H4.4 (ATCC каталог № PTA-7315), и гибридомой 8E11.9 (ATCC каталог № PTA-7319); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c).
В другом аспекте, представлено антитело, которое специфически связывается с IL-22R, в котором антитело представляет собой (a) антитело, производимое гибридомой, выбранной из 7E9 (ATCC каталог № PTA-7313), гибридомой 8A12 (ATCC каталог № PTA-7318), и гибридомой 8H11 (ATCC каталог № PTA-7317); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c).
В другом аспекте, представлен способ лечения аутоиммунного нарушения, в котором аутоиммунное нарушение не является артритом, способ, включающий введение млекопитающим эффективного количества фармацевтического состава, содержащего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывается с IL-22. В одном таком варианте осуществления, антитело, которое специфически связывается с IL-22, представляет собой (a) антитело, производимое гибридомой, выбранной из 3F11.3 (ATCC каталог № PTA-7312), гибридомой 11H4.4 (ATCC каталог № PTA-7315) и гибридомой 8E11.9 (ATCC каталог № PTA-7319); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В другом варианте осуществления, антагонист IL-22 представляет антитело, которое специфически связывается с IL-22R. В одном таком варианте осуществления, антитело, которое специфически связывается с IL-22R, представляет собой (a) антитело, производимое гибридомой, выбранной из (ATCC каталог № PTA-7313), гибридомой 8A12 (ATCC каталог № PTA-7318) и гибридомой 8H11 (ATCC каталог № PTA-7317); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В одном варианте осуществления, антагонист IL-22 представляет собой IL-22BP. В одном варианте осуществления, аутоиммунное заболевание представляет собой воспалительное заболевание кишечника. В одном варианте осуществления, аутоиммунное заболевание представляет собой псориаз. В одном варианте осуществления, способ в дальнейшем включает в себя введение как минимум одного вида антител, выбранного из антитела, которое специфически связывается с IL20Ra, антитела, которое специфически связывается с IL20Rb, и антитела, которое специфически связывает IL-22R. В одном варианте осуществления, способ в дальнейшем включает в себя введение как минимум одного вида антител, выбранного из антитела, которое специфически связывает IL-22, антитела, которое специфически связывает IL20Ra, и антитела, которое специфически связывает IL20Rb.
В другом аспекте, представлен способ лечения воспаления, в котором воспаление не является воспалением при артрите, способ, включающий введение млекопитающему эффективного количества фармацевтического состава, включающего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22. В одном таком варианте осуществления, антитело, которое специфически связывает IL-22, представляет собой (a) антитело, производимое гибридомой, выбранной из 3F11.3 (ATCC каталог № PTA-7312), гибридомой 11H4.4 (ATCC каталог № PTA-7315) и гибридомой 8E11.9 (ATCC каталог № PTA-7319); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22R. В одном таком варианте осуществления, антитело, которое специфически связывает IL-22R, представляет собой (a) антитело, производимое гибридомой, выбранной из 7E9 (ATCC каталог № PTA-7313), гибридомой 8A12 (ATCC каталог № PTA-7318) и гибридомой 8H11 (ATCC каталог № PTA-7317); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В одном варианте осуществления, антагонист IL-22 представляет собой IL-22BP. В одном варианте осуществления, воспаление представляет собой аутоиммунное воспаление. В одном варианте осуществления, воспаление представляет собой воспаление кожи. В одном варианте осуществления, воспаление представляет собой хроническое воспаление.
В другом аспекте, представлен способ подавления опухолевой прогрессии, способ, включающий введение млекопитающему эффективного количества фармацевтического состава, содержащего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22. В другом таком варианте осуществления, антитело, которое специфически связывает IL-22, представляет собой (a) антитело, производимое гибридомой, выбранной из 3F11.3 (ATCC каталог № PTA-7312), гибридомой 11H4.4 (ATCC каталог № PTA-7315) и гибридомой 8E11.9 (ATCC каталог № PTA-7319); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22R. В одном таком варианте осуществления, антитело, которое специфически связывает IL-22R, представляет собой (a) антитело, производимое гибридомой, выбранной из 7E9 (ATCC каталог № PTA-7313), гибридомой 8A12 (ATCC каталог № PTA-7318) и гибридомой 8H11 (ATCC каталог № PTA-7317); (b) аффинной созревшей формой антитела (a); (c) антигенсвязывающим фрагментом антитела (a) или (b); или (d) гуманизированной формой антитела (a), (b) или (c). В одном варианте осуществления, антагонист IL-22 представляет собой IL-22BP.
В другом аспекте, представлен способ стимуляции IL-23-опосредованного пути передачи сигнала в биологической системе, способ, включающий обеспечение биологической системы агонистом IL-22. В одном варианте осуществления, агонист IL-22 представляет собой IL-22. В другом аспекте, представлен способ подавления IL-23-опосредованного пути передачи сигнала в биологической системе, способ, включающий подавление биологической системы агонистом IL-22. В одном варианте осуществления, агонист IL-22 представляет собой антитело, которое специфически связывает IL-22. В одном варианте осуществления, агонист IL-22 представляет собой антитело, которое специфически связывает IL-22R.
В другом аспекте, представлен способ стимуляции функции ThIL-17 клетки, способ, включающий воздействие IL-22 агониста на ThIL-17 клетку. В одном варианте осуществления, агонист IL-22 представляет собой IL-22. В другом аспекте, представлен способ подавления функции ThIL-17 клетки, способ, включающий воздействие IL-22 антагониста на ThIL-17 клетки. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, которое специфически связывает IL-22R.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана нуклеотидная последовательность (SEQ ID NO: 1) кДНК, кодирующая нативный IL-22 человека.
На фиг.2 показана аминокислотная последовательность (SEQ ID NO:2), полученная из кодирующей последовательности SEQ ID NO: 1, показанной на фиг. 1.
На фиг.3 показана аминокислотная последовательность (SEQ ID NO:3) нативного IL-22R человека.
На фиг.4 показана аминокислотная последовательность (SEQ ID NO:4) нативного IL-22BP человека.
На фиг.5 представлен список всех полученных антител к IL-22 и их соответствующих свойств, как описано в примере 1. Внутриклеточное окрашивание сокращено как IC.
На фиг.6 показано, что анти-IL-22 антитела способны блокировать активацию STAT3, как описано в примере 2.
На фиг.7 показано, что три специфических вида анти-IL-22 антител блокируют IL-22 человека зависимым от дозы образом, как описано в примере 3.
На фиг.8 показано, что три специфических вида анти-IL-22 антител IL-22 способны блокировать мышиный IL-22 зависимым от дозы образом, как описано в примере 4.
На фиг.9 представлен расчет аффинности анти-IL-22 антител к IL-22 человека, как описано в примере 5.
На фиг.10 показано, что анти-IL-22 антитела обнаруживают внутриклеточную экспрессию IL-22, как описано в примере 6.
На фиг.11 показано внутриклеточное FACS окрашивание на IL-22 с использованием меченных анти-IL-22 антител, как описано в примере 6.
На фиг.12 показана экспрессия IL-22 в мышиных Th1 клетках, как определено 5' нуклеазным анализом, как описано в примере 7.
На фиг.13 показана экспрессия IL-22 в мышиных γδ Т-клетках, как определено 5' нуклеазным анализом, как описано в примере 8.
На фиг.14 показана экспрессия IL-22 в активированных Т-клетках человека, как определено микрочиповым анализом, как описано в примере 9.
На фиг.15 показан уровень экспрессии IL-22 в Т-клетках путем FACS, как описано в примере 10.
На фиг. 16 показано исследование анти-IL-22R антител на 293 клетках, экспрессирующих IL-22R, как описано в примере 11.
На фиг. 17 показано, что анти-IL-22R антитела могут блокировать вызванную IL-22 экспрессию STAT3 репортерного конструкта, как описано в примере 12.
На фиг. 18 показана экспрессия IL-22R и IL-10R2 на поверхности первичных кератиноцитов, как описано в примере 13.
На фиг. 19 показано, что IL-22 вызывает истончение эпидермиса человека, как описано в примере 14.
На фиг. 20 показано, что IL-22 вызывает экспрессию цитокератина 16, маркера кератиноцитарного метаболизма, как описано в примере 14.
На фиг. 21 показано, что лечение эпидермиса человека IL-22 вызывает индукцию экспрессии псориазина, высокоэкспрессируемого при псориазе гена, как описано в примере 14.
На фиг. 22 показано, что лечение кератиноцитов IL-22 увеличивает экспрессию нескольких генов, включая псориазин, как описано в примере 15.
На фиг. 23 показано, что экспрессия псориазина уменьшается при лечении анти-IL-22 и анти-IL-22R антителами, как описано в примере 14.
На фиг. 24 показано, что эпидермальное истончение уменьшается при лечении анти-IL-22 и анти-IL-22R антителами, как описано в примере 14.
На фиг. 25 показано, что эпидермальное истончение уменьшается при лечении анти-IL-22 и анти-IL-22R антителами, как описано в примере 14.
На фиг. 26 показано, что IL-23 и IL-12 вызывают эпидермальное истончение с отчетливо выраженными гистологическими признаками, как описано в примере 16.
На фиг. 27 показано, что IL-23 вызывает экспрессию IL-22, а IL-22 вызывает кожное воспаление и эпидермальное истончение in vivo, как описано в примере 17 и 18.
На фиг. 28 показано, что IL-12 и IL-23 вызывают экспрессию отдельных групп цитокинов, как описано в примере 17.
На фиг. 29 показано, что лечение анти-IL-22 моноклональными антителами значительно уменьшает IL-23-вызванный эпидермальный акантоз in vivo, как описано в примере 20.
На фиг. 30 представлена стратегия, использованная для разрушения гена IL-22 у мышей, и доказательство, подтверждающее что экспрессия IL-22 отсутствует у IL-22-/- мышей, как описано в примере 20.
На фиг. 31 показано, что IL-23-вызванный акантоз значительно уменьшается у IL-22 дефицитных мышей, как описано в примере 20.
На фиг. 32 показано, что дефицит IL-22 не имеет никакого эффекта на IL-12-вызванный акантоз, как описано в примере 20.
На фиг. 33 показано, что IL-23 вызывает продукцию IL-22 из разных IL-23-активированных лимфоцитов, как описано в примере 21.
На фиг. 34 показано, что IL-22 представляет собой новый эффекторный цитокин из ThIL-17 линии, как описано в примере 22.
На фиг. 35 показано, что IL-22 и IL-17 продуцируются той же самой Th линией (ThIL-17), как описано в примере 22.
На фиг. 36 показано, что IL-23 стимулирует продукцию IL-22 исходя из активации Т-клеток, как описано в примере 22.
На фиг. 37 показано, что IL-19, IL-20, IL-22 и IL24 вызывает эпидермальное истончение, как описано в примере 23.
На фиг. 38 показана количественная оценка эпидермального акантоза, вызванного IL-19, IL-20, IL-22 и IL24, как описано в примере 23.
На фиг.39 показано, что компоненты рецепторов для IL-19, IL-20 и IL-22 экспрессируются в кератиноцитах человека, как описано в примере 24.
На фиг. 40 показано, что блокирующие антитела к компонентам рецептора для IL-19, IL-20 и IL-22 уменьшают экспрессию псориазина, как описано в примере 24.
На фиг. 41 показано, что антитела к IL-20Ra и IL-22R, при использовании в сочетании, эффективно блокируют IL-20-вызванную экспрессию псориазина.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
I. ОПРЕДЕЛЕНИЯ
Термин "IL-22 полипептид" или "IL-22" относятся к разным интерлейкин-22 полипептидам (также обозначенный как "интерлейкин-22 лиганд" или "IL-22L" в данной области). Термин включает нативную последовательность IL-22 полипептидов и их варианты (которые определяются в дальнейшем в данном документе). IL-22 полипептиды, описанные в данном документе, могут быть выделены из различных источников, таких как ткани человека, или другого источника, или полученных рекомбинантными или синтетическими способами. Нативный IL-22 может быть любых видов, например мышиный ("mIL-22") или человеческий ("hIL-22").
Термины "IL-22R полипептид" или "IL-22R" относятся к полипептидному компоненту гетеродимера рецептора интерлейкина-22 или гетеродимера рецептора интерлейкина-20. Термин включает нативную последовательность IL-22R полипептидов и их вариантов (которые определяются в дальнейшем в данном документе). IL-22R полипептиды, описанные в данном документе, могут быть выделены из разных источников, таких как человеческая ткань, или другого источника, или получены рекомбинантными или синтетическими способами. Нативный IL-22R может быть любых видов, например мышиный ("mIL-22R") или человеческий ("hIL-22R"). Нативная последовательность IL-22R полипептидов, также называемая в данной области как "IL-22R1" и "IL22RA." "Нативная последовательность IL-22 полипептида" или "нативная последовательность IL-22R полипептида" относится к полипептиду, включающему ту же самую аминокислотную последовательность, как соответствующие IL-22 или IL-22R полипептиды, полученные в природе. Такая нативная последовательность IL-22 или IL-22R полипептидов может быть выделена природно или может быть получена рекомбинантными или синтетическими способами. Термины, в частности, включают встречающиеся в природе усеченные или секретируемые формы специфического IL-22 или IL-22R полипептида (например, IL-22, не имеющий своего связанного сигнального пептида), встречающиеся в природе различные формы (например, формы альтернативного спласинга), и встречающиеся в природе аллельные варианты полипептида. В различных вариантах осуществления изобретения, нативная последовательность IL-22 или IL-22R полипептидов, раскрытых в данном документе, представляет собой зрелые или полной длины нативные последовательности полипептидов. На фиг.2 и 3 показаны типичные человеческие полной длины IL-22 и IL-22R, соответственно. Нуклеиновая кислота, кодирующая полипептид, показанная на фиг.2, показана на фиг.1. Стартовый и стоп-кодоны показаны на этом чертеже жирным шрифтом и подчеркнуты. Хотя показано, что IL-22 и IL-22R полипептидные последовательности, раскрытые в сопроводительных чертежах, начинаются с остатков метионина, обозначенных в данном документе как аминокислотное положение 1, возможно и вероятно, что другие остатки метионина, расположенные либо в обратном направлении, либо в прямом направлении от аминокислотного положения 1 на чертежах, могут использоваться как стартовый аминокислотный остаток для IL-22 или IL-22R полипептидов.
"IL-22 вариант", "IL-22R вариант", "полипептид IL-22 варианта" или "полипептид IL-22R варианта" означает активный IL-22 или IL-22R полипептид как определено выше, имеющий как минимум примерно 80% идентичности аминокислотной последовательности с нативной последовательностью IL-22 полной длины или IL-22R полипептидной последовательностью. Обычно вариант IL-22 или IL-22R полипептида будет иметь, как минимум, примерно 80% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 81% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 82% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 83% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 84% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 85% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 86% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 87% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 88% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 89% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 90% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 91% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 92% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 93% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 94% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 95% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 96% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 97% идентичности аминокислотной последовательности, альтернативно, как минимум, примерно 98% идентичности аминокислотной последовательности и альтернативно, как минимум, примерно 99% идентичности аминокислотной последовательности к полной длины или зрелой нативной последовательности IL-22 или IL-22R полипептидной последовательности.
"Процент (%) идентичности аминокислотной последовательности" по отношению к IL-22 или IL-22R полипептидным последовательностям, идентифицированным в данном документе, определяется как процент аминокислотных остатков в подходящей последовательности, которые идентичны аминокислотным остаткам в специфической IL-22 или IL-22R полипептидной последовательности, после ориентирования последовательностей и внесения брешей, если необходимо, для получения максимального процента идентичности последовательностей, и не учитывая никаких консервативных замен в рамках идентичности последовательностей. Ориентирование для целей определения процента идентичности аминокислотной последовательности можно получить различными путями, которые известны специалисту в данной области, например с использованием общедоступного компьютерного программного обеспечения, такого как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR) программное обеспечение. Специалисты в данной области могут определять подходящие параметры измерения ориентирования, включая любые алгоритмы, необходимые для получения максимального ориентирования над полной длиной последовательностей, при сравнении. Для сравнений аминокислотных последовательностей, % идентичности аминокислотных последовательностей, данной аминокислотной последовательности A к, с, или против данной аминокислотной последовательности B (которая может альтернативно называться как данная аминокислотная последовательность A, которая имеет или включает некоторый % идентичности аминокислотной последовательности к, с или против данной аминокислотной последовательности B) рассчитывается, как указано ниже:
100 умножить на дробь X/Y,
где X представляет собой количество аминокислотных остатков, отмеченных как идентичные пары программой ориентирования последовательностей в этом программном ориентировании A и B, и где Y представляет собой общее количество аминокислотных остатков в B. Принимая во внимание, что длина аминокислотной последовательности A не эквивалентна длине аминокислотной последовательности B, % идентичности аминокислотной последовательности A к B не будет эквивалентным % идентичности аминокислотной последовательности B к A. В качестве примеров вычислений % идентичности аминокислотной последовательности с использованием этого способа, в таблицах 2 и 3 показано как вычислить % идентичности аминокислотной последовательности, обозначенной "белком сравнения" аминокислотной последовательности, к аминокислотной последовательности, обозначенной "IL-22 или IL-22R", где "IL-22 или IL-22R" представляет аминокислотную последовательность интересующего полипептида IL-22 или IL-22R, "белок сравнения" представляет аминокислотную последовательность полипептида, против которого сравнивается интересующий полипептид "IL-22 или IL-22R", и каждый "X, "Y" и "Z" представляет различные аминокислотные остатки.
% идентичности аминокислотной последовательности = (число идентичных парных аминокислотных остатков между двумя полипептидными последовательностями) деленное на (общее количество аминокислотных остатков IL-22 или IL-22R полипептида) = 5 деленное на 15=33,3%
% идентичности аминокислотной последовательности = (число идентичных парных аминокислотных остатков между двумя полипептидными последовательностями) деленное на (общее количество аминокислотных остатков IL-22 или IL-22R полипептида) = 5 деленное на 10=50%
Термин "IL-19" относится к любому нативному IL-19 из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин охватывает "полной длины", непроцессируемый IL-19, а также и любую форму IL-19, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-19, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-19, который поддерживает, как минимум, одну биологическую активность IL-19.
Термин "IL-20" относится к любому нативному IL-20 из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин включает "полной длины", непроцессируемый IL-20, а также и любую форму IL-20, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-20, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-20, который поддерживает, как минимум, одну биологическую активность IL-20.
Термин "IL-24" относится к любому нативному IL-24 из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если нет особого указания. Термин включает "полной длины", непроцессируемый IL-24, а также и любую форму IL-24, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-24, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-24, который поддерживает, как минимум, одну биологическую активность IL-24.
Термин "IL-22BP" или "IL-22 связывающий белок", как использовано в данном документе, относится к любому нативному IL-22BP из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин охватывает "полной длины", непроцессируемый IL-22BP, а также и любую форму IL-22BP, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-22BP, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-22BP, который поддерживает, как минимум, одну биологическую активность IL-22BP. Нативный IL-22BP также называется как "IL-22RA2" в данной области.
Термин IL-20Ra относится к полипептидному компоненту гетеродимера рецептора IL-19 или гетеродимера рецептора IL-20. Термин включает любой нативный IL-20Ra из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин включает "полной длины", непроцессируемый IL-20Ra, а также и любую форму IL-20Ra, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-20Ra, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-20Ra, который поддерживает, как минимум, одну биологическую активность IL-20Ra. Нативный IL-20Ra также называется как "IL-20R1" в данной области.
Термин IL-20Rb относится к полипептидному компоненту гетеродимера рецептора IL-19 или гетеродимера рецептора IL-20. Термин включает любой нативный IL-20Rb из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин включает "полной длины", непроцессируемый IL-20Rb, а также и любую форму IL-20Rb, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-20Rb, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-20Rb, который поддерживает, как минимум, одну биологическую активность IL-20Rb. Нативный IL-20Rb также называется как "IL-20R2" в данной области.
Термин "IL-10R2" относится к полипептидному компоненту гетеродимера рецептора IL-22 или гетеродимера рецептора IL-20. Термин включает любой нативный IL-10R2 из любого позвоночного животного, включая млекопитающих, таких как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо. Термин включает "полной длины", непроцессируемый IL-10R2, а также и любую форму IL-10R2, которая получается в результате процессинга в клетке. Термин также включает встречающиеся в природе варианты IL-10R2, например сплайс-варианты, аллельные варианты и другие изоформы. Термин также включает фрагменты или варианты нативного IL-10R2, который поддерживает, как минимум, одну биологическую активность IL-10R2. Нативный IL-10R2 также называется как "IL-10Rb" в данной области.
"Выделенная" биологическая молекула, такая как различные полипептиды, полинуклеотиды и описанные в данном документе антитела, относится к биологической молекуле, которая была идентифицирована и разделена и/или извлечена из, как минимум, одного компонента из естественных условий.
"Активный" или "активность", по отношению к IL-22 или IL-22R, относятся к биологической и/или иммунологической активности нативного IL-22 или IL-22R, где "биологическая" активность относится к биологической функции нативного IL-22 или IL-22R, исключая способность вызывать продукцию антител против антигенного эпитопа, обладающего нативным IL-22 или IL-22R. "Иммунологическая" активность относится к способности вызывать продукцию антител против антигенного эпитопа, обладающего нативным IL-22 или IL-22R.
Термин "антагонист" используется в широком смысле понимания и включает в себя любую молекулу, которая частично или полностью блокирует, подавляет или нейтрализует биологическую активность полипептида, такого как нативный IL-22 или IL-22R полипептид. Также включенные "антагонисты" представляют собой молекулы, которые полностью или частично подавляют транскрипцию или трансляцию мРНК, кодирующую полипептид. Подходящие молекулы-антагонисты включают в себя, например, антагонисты-антитела или фрагменты антител; фрагменты или варианты аминокислотных последовательностей нативного полипептида; пептиды; антисмысловые олигонуклеотиды; малые органические молекулы; и нуклеиновые кислоты, которые кодируют полипептидные антагонисты или антагонисты-антитела. Ссылка на "an" антагонист включает простой антагонист или комбинацию двух или более различных антагонистов. Термин "агонист" используется в широком смысле понимания и включает в себя любую молекулу, которая частично или полностью воспроизводит биологическую активность полипептида, такого как нативный IL-22 или IL-22R полипептид. Также включенный "агонист" представляет собой молекулы, которые стимулируют транскрипцию или трансляцию мРНК, кодирующую полипептид. Подходящие молекулы-агонисты включают в себя, например, агонисты-антитела или фрагменты антител; нативный полипептид; фрагменты или варианты аминокислотных последовательностей нативного полипептида; пептиды; антисмысловые олигонуклеотиды; малые органические молекулы; и нуклеиновые кислоты, которые кодируют полипептидные агонисты или антитела. Ссылка на "an" агонист включает простой агонист или комбинацию двух или более различных агонистов.
"Облегчение" относится и к терапевтическому лечению, и к профилактическим или предупредительным мерам, при которых у индивида предотвращается или снижается темп (уменьшается) нацеленного патологического состояния или нарушения. Таковые при необходимости лечения включают в себя индивидов уже с нарушением, а также индивидов, склонных к нарушению, или индивидов, у которых предупреждают нарушение.
"Хроническое" введение относится к введению веществ(а) в непрерывном режиме в отличие от острого режима, с тем чтобы поддержать начальный терапевтический эффект в течение длительного периода времени. "Прерывистое" введение представляет собой лечение, которое не обходится без последовательного прерывания, но предпочтительно является циклическим по характеру.
Термин "млекопитающие" для целей лечения относится к любому животному, классифицированному как млекопитающее, включая людей, грызунов (например, мышей и крыс) и обезьян; домашних и сельскохозяйственных животных; и животных из зоопарка, спорта, лаборатории или питомцев, таких как собаки, кошки, крупный рогатый скот, лошади, овцы, свиньи, козы, кролики и др. В некоторых вариантах осуществления, млекопитающих выбирают из человека, грызуна или обезьяны.
Введение "в комбинации с" одним или более в дальнейшем терапевтическими веществами включает в себя одновременное (параллельное) и последующее введение в любом порядке. "Носители", как использовано в данном документе, включают в себя фармацевтически приемлемые носители, наполнители или стабилизаторы, которые являются нетоксичными для клетки или млекопитающего, если подвергаются этому воздействию в дозах и концентрациях. Часто физиологически приемлемый носитель представляет собой водный pH буферный раствор. Примеры физиологически приемлемых носителей включают в себя буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; полипептиды низкого молекулярного веса (менее чем примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводороды, включая глюкозу, маннозу или декстрины; хелатирующие вещества, такие как EDTA; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEENTM, полиэтиленгликоль (PEG) и PLURONICSTM. "Антитела" (Abs) и "иммуноглобулины" (Igs) представляют собой гликопротеины, имеющие сходные структурные характеристики.
Хотя антитела подавляют специфичность связывания специфичного антигена, иммуноглобулины включают в себя и антитела, и другие подобные антителам молекулы, которые, как правило, лишены антигенной специфичности. Полипептиды последнего типа представляют собой, например, производимые лимфатической системой в низких уровнях и миеломой в высоких уровнях.
Термины "антитело" и "иммуноглобулин" используются равнозначно в широком смысле понимания и включают в себя моноклональные антитела (например, полной длины или интактные моноклональные антитела), поликлональные антитела, моновалентные антитела, мультивалентные антитела, мультиспецифичные антитела (например, биспецифические антитела, поскольку они проявляют требуемую биологическую активность) и могут также включать в себя некоторые фрагменты антител (как подробно описано в данном документе). Антитело может быть химерным, человеческим, гуманизированным и/или аффинно зрелым.
Антитело, которое специфически связывается со специфическим антигеном, относится к антителу, которое способно связывать антиген с достаточной аффиностью, так что антитело является полезным в качестве диагностического и/или терапевтического вещества в мишени-антигене. Предпочтительно, степень связывания такого антитела с ненацеленным полипептидом представляет собой менее чем примерно 10% связывания антитела с мишенью антигеном, как измерено, например, радиоиммунологическим анализом (RIA). В некоторых вариантах осуществления, антитело, которое связывает мишень антиген, имеет константу диссоциации (Kd)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ.
"Вариабельная область" или "вариабельный домен" антитела относится к амино-терминальным доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может быть назван как "VH." Вариабельный домен легкой цепи может быть назван как "VL." Эти домены, как правило, представляют собой наиболее вариабельные части антитела и содержат участки связывания антигена.
Термин "вариабельный" относится к факту, что некоторые участки вариабельных доменов значительно отличаются в последовательности антител и используются в связывании и специфичности каждого конкретного антитела для каждого конкретного антигена. Однако вариабельность неравномерно распределена по всем вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых гипервариабельными участками (CDRs) или гипервариабельными участками (HVRs) в вариабельных доменах и в легкой цепи, и в тяжелой цепи. Более высококонсервативные участки вариабельных доменов называются скелетными областями (FR). Каждый их вариабельных доменов нативных тяжелой и легкой цепей содержит четыре FR области, в основном принимая бета-видимую конфигурацию, соединяясь тремя CDRs, которые образуют петлю соединения, и в некоторых случаях образуя часть бета-видимой структуры. CDRs в каждой цепи скреплены в непосредственной близости от FR областей и, с CDRs другой цепи участвуют в образовании антигенсвязывающего участка антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены непосредственно не вовлечены в связывание антитела с антигеном, но проявляют разные эффекторные функции, такие как участие антитела в антитело-зависимой клеточной токсичности.
"Легкие цепи" антител (иммуноглобулинов) любых видов позвоночных животных можно отнести к одному или двум очевидно разным типам, названным каппа (K) и лямбда (λ), на основании аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотных последовательностей константных доменов их тяжелых цепей антитела (иммуноглобулины) можно отнести к различным классам. Существует пять больших классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них в дальнейшем могут подразделяться на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и µ, соответственно. Структуры субъединиц и пространственная конфигурация различных классов иммуноглобулинов хорошо известна и повсеместно описывается, например в Abbas et al. Cellular and Mol. Immunology, 4th ed. (2000). Антитело может быть частью большой сложной молекулы, образованной ковалентной или нековалентной связью антитела с одним или более другими белками или пептидами.
Термины "антитело полной длины", "интактное антитело" и "полное антитело" используются в данном документе равнозначно для обозначения антитела в его основной интактной форме, без фрагментов антител, как описано ниже. Термины, в частности, относятся к антителу с тяжелыми цепями, которые содержат Fc область. "Фрагменты антитела" включают только участок интактного антитела, в котором участок поддерживает как минимум одну, и столько же много или всю функцию, обычно связанную с участком при представлении в интактном антителе. В одном варианте осуществления, фрагмент антитела включает антигенсвязывающий участок интактного антитела и таким образом поддерживает способность связывать антиген. В другом варианте осуществления, фрагмент антитела, например, который включает Fc область, поддерживает как минимум одну из биологических функций, обычно связанных с Fc областью при представлении в интактном антителе, такую как FcRn связывание, регуляция времени полужизни антитела, ADCC функция и связывание комплемента. В одном варианте осуществления, фрагмент антитела представляет собой моновалентное антитело, которое имеет значительно сходное время полужизни in vivo с интактным антителом. Например, такой фрагмент антитела может лежать на антигенсвязывающем плече, связанном с Fc последовательностью, способным придавать стабильность фрагменту in vivo.
Расщепление антитела папаином приводит к образованию двух антигенсвязывающих фрагментов, названных "Fab" фрагментами, каждый с одним антигенсвязывающим участком, и остаточным "Fc" фрагментом, чье название отражает его способность быстро кристаллизоваться. Продукт обработки пепсином F(ab')2 фрагмент имеет два антигенсвязывающих участка и еще способен перекрестно связывать антиген.
"Fv" представляет собой минимальный фрагмент антитела, который содержит участок связывания полного антигена. В одном варианте осуществления, двухцепочечные Fv виды состоят из димера из вариабельного домена одной тяжелой и одной легкой цепи в устойчивой нековалентной связи. В одноцепочечных Fv (scFv) видах вариабельный домен одной тяжелой и одной легкой цепи может быть ковалентно связан свободной пептидной связью, такой что легкие и тяжелые цепи могут связываться в "димерную" структуру, аналогичную структуре двухцепочечных Fv видов. Она существует в такой конфигурации, что три CDRs каждого вариабельного домена взаимодействуют до распознавания антигенсвязывающего участка на поверхности VH-VL димера. В совокупности шесть CDRs придают антигенсвязывающую специфичность антителу. Однако даже один вариабельный домен (или половина Fv, содержащих только три CDRs, специфичных для антигена) имеет способность распознавать и связывать антиген, хотя с более низкой аффиностью, чем полный участок связывания.
Fab фрагмент содержит вариабельные домены тяжелой и легкой цепей и также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab' фрагменты отличаются от Fab фрагментов добавлением нескольких остатков на карбоксильном конце домена тяжелой цепи CH1, включая один или более цистеинов из шарнирной области антитела. Fab'-SH в данном документе представляет собой обозначение Fab', в котором цистеиновые остатки константных доменов связаны со свободной тиольной группой. F(ab')2 фрагменты антитела первоначально продуцировались как пары Fab' фрагментов, которые имеют шарнирные цистеины между ними. Также известны другие химические взаимодействия фрагментов антител.
"Одноцепочечные Fv" или "scFv" фрагменты антитела содержат VH и VL домены антитела, где эти домены представлены в одной полипептидной цепи. Как правило, scFv полипептид в дальнейшем содержит полипептидную связь между VH и VL доменами, которые позволяют scFv образовывать желательную структуру для связывания антигена. Для обзора scFv см. Pluckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Термин "димеры" относится к малым фрагментам антитела с двумя антигенсвязывающими участками, фрагменты которого содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в аналогичной полипептидной цепи (VH-VL). Используя линкер, который слишком короткий, чтобы спариваться с двумя доменами на той же самой цепи, домены должны спариваться с комплементарными доменами другой цепи и образуют два антигенсвязывающих участка. Димеры могут быть бивалентными или биспецифическими. Более подробно димеры описаны, например, в EP 404097; WO93/1161; Hudson et al. (2003) Nat. Med. 9:129-134; и Hollinger et al, Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Тримеры и тетрамеры также описаны в Hudson et al. (2003) Nat. Med. 9: 129-134.
Как в данном документе использовано, термин "моноклональное антитело" относится к антителу, полученному из популяции в значительной степени гомогенных антител, то есть отдельные антитела, входящие в состав популяции, идентичны, за исключением возможных мутаций, которые могут быть представлены в небольших количествах. Таким образом, модификатор "моноклональное" показывает характер антитела, не являющегося смесью отдельных антител. В отдельных вариантах осуществления, такое моноклональное антитело обычно включает в себя антитело, содержащее полипептидную последовательность, которая связывает мишень, где мишень-связывающая полипептидная последовательность была получена в процессе, который включает в себя селекцию одной мишень-связывающей полипептидной последовательности из множества полипептидных последовательностей. Например, процесс селекции может представлять собой селекцию уникального клона из множества клонов, такого как пул гибридомных клонов, фаговых клонов или клонов с рекомбинантной ДНК. Следует понимать, что отобранная мишень-связывающая последовательность в дальнейшем может быть изменена, например, для того чтобы улучшить аффинность к мишени, гуманизировать мишень-связывающую последовательность, улучшить ее продукцию в культуре клеток, уменьшить ее иммуногенность in vivo, получить мультиспецифичное антитело и другое, и что антитело, содержащее измененную мишень-связывающую последовательность, также представляет собой моноклональное антитело этого изобретения. В отличие от препаратов поликлональных антител, которые обычно включают в себя антитела, направленные против различных детерминант (эпитопы), каждое моноклональное антитело из препарата моноклональных антител представляет собой направленное против одной детерминанты антигена. В дополнение к их специфичности, препараты моноклональных антител эффективны тем, что они обычно не содержат другие иммуноглобулины.
Модификатор "моноклональное" показывает характер антитела, полученного в значительной степени из гомогенной популяции антител, и его невозможно считать как желательную продукцию антитела любым частным способом. Например, чтобы использоваться в соответствии с настоящим изобретением, моноклональные антитела можно получать различными способами, включающими в себя, например, гибридомный способ (например, Kohler et al, Nature, 256: 495 (1975); Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981)), способы рекомбинантной ДНК (см., например, патент США № 4816567), способы фагового дисплея (см., например, Clackson et al., Nature, 352:624-628 (1991); Marks et al., J. Mol. Biol. 222:581-597 (1992); Sidhu et al., J. Mol. Biol. 338(2):299-310 (2004); Lee et al., J. Mol. Biol. 340(5):1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34):12467-12472 (2004); и Lee et al., J. Immunol. Methods. 284(1-2):119-132(2004) и технологии получения человеческих или подобных человеческим антител у животных, которые содержат части или все локусы или гены иммуноглобулина человека, кодирующие последовательности иммуноглобулина человека (см., например, WO98/24893; WO96/34096; WO96/33735; WO91/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA 90:2551 (1993); Jakobovits et al., Nature 362:255-258 (1993); Bruggemann et al., Year in Immunol. 7:33 (1993); патенты США № 5545807; 5545806; 5569825; 5625126; 5633425; 5661016; Marks et al., Bio. Technology 10:779-783 (1992); Lonberg et al., Nature 368:856-859 (1994); Morrison, Nature 368:812-813 (1994); Fishwild et al., Nature Biotechnol. 14:845-851 (1996); Neuberger, Nature Biotechnol. 14:826 (1996) и Lonberg and Huszar, Intern. Rev. Immunol. 13:65-93 (1995).
Моноклональные антитела в данном документе специфически включают в себя "химерные" антитела, у которых участок тяжелой и/или легкой цепи аналогичен или гомологичен соответствующим последовательностям антител, полученных из отдельных видов, или принадлежащим к конкретному классу или подклассу антител, тогда как остаток цепи (цепей) аналогичен или соответствующим последовательностям антител, полученных из других видов, или принадлежащим другому классу или подклассу антител, а также и фрагменты таких антител, поскольку они проявляют желательную биологическую активность (патент США № 416567; и Morrison et al, Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
"Гуманизированные" формы нечеловеческих (например, мышиных) антител представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из нечеловеческого иммуноглобулина. В одном варианте осуществления, гуманизированное антитело представляет собой иммуноглобулин человека (антитело реципиента), в котором остатки из гипервариабельной области реципиента были замещены остатками из гипервариабельной области нечеловеческих видов (антитело донора), таких как мышь, крыса, кролик или нечеловекообразный примат, обладающих желаемой специфичностью, аффиностью и/или способностью. В отдельных случаях, остатки из скелетной области (FR) иммуноглобулина человека замещаются соответствующими нечеловеческими остатками. Кроме того, гуманизированные антитела могут включать остатки, которые не обнаруживаются у антител реципиента или донора. Эти модификации можно получить для дальнейшего улучшения продуктивности антител. Как правило, гуманизированное антитело будет содержать почти все или, по крайней мере, один, а обычно два, вариабельных домена, в которых все или почти все гипервариабельные петли соответствуют петлям нечеловеческого иммуноглобулина, и все или почти все FR представляют собой FR последовательности иммуноглобулина человека. Дополнительно, гуманизированные антитела будут также содержать по крайней мере участок константной области иммуноглобулина (Fc), обычно участок иммуноглобулина человека. Далее подробнее, см. Jones et al, Nature 321:522-525 (1986); Riechmann et al, Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). См. также приведенные в данном документе следующие обзорные статьи и ссылки: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
"Человеческое антитело" представляет собой антитело, которое содержит аминокислотную последовательность, которая соответствует последовательности антитела, полученного от человека, и/или была получена с использованием технологий для получения антител человека, как показано в данном документе. Это определение человеческого антитела специфически исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей, иммуноглобулины можно отнести к различным классам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них в дальнейшем можно разделить на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA и IgA2. "Аффинно зрелое" антитело представляет собой антитело с одним или более переходами в одном или более из HVR, которые приводят к увеличению аффинности антитела к антигену, по сравнению с родительским антителом, которое не содержит таких переходов. В одном варианте осуществления, аффинно зрелое антитело обладает наномолярными или хотя бы пикомолярными аффинностями к антигену-мишени. Аффинно зрелые антитела можно получить способами, известными в данной области. Marks et al. Bio/Technology 10:779-783 (1992) описывает созревание аффинности путем перетасовки VH и VL доменов. Неспецифический мутагенез HVR и/или скелетных остатков описывается в: Barbas et al. Proc Nat. Acad. Sci. USA 91:3809-3813 (1994); Schier et al Gene 169:147-155 (1995); Yelton et al. J. Immunol. 155:1994-2004 (1995); Jackson et al, J. Immunol. 154(7):3310-9 (1995); и Hawkins et al, J. Mol. Biol. 226:889-896 (1992).
"Блокирующее" антитело, "нейтрализующее" антитело или антитело "антагонист" представляет собой антитело, которое подавляет или уменьшает биологическую активность связывания антигена. Такие антитела могут значительно или полностью подавлять биологическую активность антигена. Как используется в данном документе, "антитело-агонист" представляет собой антитело, которое частично или полностью воспроизводит биологическую активность интересующего полипептида.
"Эффекторные функции" антител относятся к биологическим активностям, связанным с Fc областью (Fc область нативной последовательности или вариантная Fc область аминокислотной последовательности) антитела, и изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают в себя: C1q связывание и коплемент-зависимая цитотоксичность; связывание Fc рецептора; антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC); фагоцитоз; down-регуляция рецепторов клеточной поверхности (например, рецептор B-клеток); и активация B-клеток.
"Аффинность связывания", как правило, относится к силе совокупности нековалентных взаимодействий между одним участком связывания молекулы (например, антитела) и ее связывающим партнером (например, антигеном). Если не указано особо, как использовано в данном документе, "аффинность связывания" относится к истинной связывающей аффинности, которая отражает 1:1 взаимодействие между членами пары связывания (например, антитело и антиген). Аффинность молекулы X для ее партнера Y, как правило, может быть представлена константой диссоциации (Kd). Аффинность можно измерить обычными способами, известными в данной области, включая описанные в данном документе. Антитела с низкой аффинностью, как правило, связывают антиген медленно и склонны легко диссоциировать, тогда как антитела с высокой аффинностью, как правило, более быстро связывают антиген и склонны дольше оставаться связанными. Различные способы измерения аффинности связывания известны в данной области, каждый из которых можно использовать для целей настоящего изобретения. Конкретные наглядные варианты осуществления описаны ниже. В одном варианте осуществления, "Kd" или "значение Kd" согласно этому изобретению измеряют путем анализа связывания радиоактивно меченого антигена (RIA), осуществленного с Fab вариантом интересующего антитела и его антигена, как описано следующим анализом. Растворенную аффинность связывания Fabs для антигена измеряют уравновешиванием Fab с минимальной концентрацией (125I)-меченого антигена в присутствии серий титрования немеченого антигена, далее захватывая связавшийся антиген анти-Fab покрытый антителами планшет (Chen, et al., (1999) J. Mol. Biol. 293:865-881). Для создания условий для анализа, микротитровые планшеты (Dynex) покрывают на ночь 5 мкг/мл захваченных анти-Fab антител (Cappel Labs) в 50 мМ карбонате натрия (pH 9,6), и позднее блокируют 2% (w/v) бычьего сывороточного альбумина в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В неадсорбирующем планшете (Nunc #269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями интересующего Fab (например, в соответствии с оценкой анти-VEGF антитела, Fab-12, в Presta et al., (1997) Cancer Res. 57:4593-4599). Интересующий Fab затем инкубируют в течение ночи; однако инкубацию можно продолжать в течение более длительного периода (например, примерно 65 часов), чтобы убедиться, что достигнута уравновешенность. После этого смеси переносят на захватывающий планшет для инкубации при комнатной температуре (например, в течение одного часа). Затем убирают раствор, и планшет промывают восемь раз 0,1% Tween-20 в PBS. Если планшеты были высушены, то добавляют 150 мкл/лунку сцинтиллятора (MicroScint-20; Packard), и планшеты подсчитывают на Topcount gamma counter (Packard) в течение десяти минут. Концентрации каждого Fab, который дает менее или равно 20% максимального связывания, выбирают для использования в конкурентно-связывающем анализе. Согласно другому варианту осуществления, Kd или значение Kd измеряли с помощью поверхностного плазмонного резонансного анализа, используя BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с иммобилизированными гранулами антигена CM5 ~10 единиц ответа (RU). Кратко, карбоксиметилированные декстрановые биосенсорные гранулы (CM5, BIAcore Inc.) активируются N-этил-N'-(3-диметиламинопропил)карбодиимид гидрохлорид (EDC) и N- гидроксисукцинимид (NHS) согласно инструкциям производителя. Антиген разводят 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед введением в скорость потока 5 мкл/минуту для достижения приблизительно 10 единиц ответа (RU) связанного белка. После введения антигена, вводят 1 М этаноламина, чтобы заблокировать непрореагировавшие группы. Для измерений кинетики, двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) вводят в PBS с 0,05% Tween 20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. Скорость ассоциации (kon) и скорость диссоциации (koff) рассчитывают, используя простую один к одному модель связывания Langmuir (BIAcore Evaluation Software version 3,2), одновременно выравнивая сенсограммы ассоциации и диссоциации. Равновесную константу диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen Y. et al., (1999) J. Mol. Biol. 293:865-881. Если on-скорость превышает 106 М-1с-1 с помощью вышеупомянутого поверхностного плазмонного резонансного анализа, затем on-скорость можно определять с использованием метода флуоресцентного гашения, которым измеряют увеличение или уменьшение интенсивности флуоресценции (возбуждение = 295 нм; эмиссия = 340 нм, 16 нм полоса пропускания) при 25°C 20 нМ антитело против антигена (Fab форма) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена, как измерено в спектрометре, таком как спектрофотометр, оснащенный остановленным потоком (Aviv Instruments), или 8000-серийный SLM-Aminco спектрофотометр (ThermoSpectronic) со смесительной кюветой.
"On-скорость", "скорость ассоциации", "ассоциирующая скорость" или "kon" согласно этому изобретению также можно определить как описано выше, используя BIAcoreTM-2000 или BIAcoreTM-3000 систему (BIAcore, Inc., Piscataway, NJ).
"Изолированное" антитело представляет собой антитело, которое было идентифицировано и выделено и/или извлечено из компонента его естественного окружения. Нежелательные компоненты его естественного окружения представляют собой вещества, которые возможно препятствуют диагностическому и терапевтическому применению антитела и могут включать в себя ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления, антитело будет очищено (1) до более чем 95% по весу антитела, как определено способом Лоури, и наиболее предпочтительно более чем 99% по весу, (2) в значительной степени для получения, по крайней мере, 15 остатков N-концевой или внутренней аминокислотной последовательности, используя секвенатор с вращающимся стаканом, или (3) для гомогенности путем SDS-PAGE в восстанавливающих или невосстанавливающих условиях, используя Кумасси синий или, предпочтительно, окрашивание серебром. Изолированное антитело включает в себя антитело in situ внутри рекомбинантных клеток, так как, по крайней мере, один компонент естественного окружения антител не будет представлен. Однако обычно изолированное антитело будет получено, по крайней мере, одним этапом очистки.
Слово "метка" использовано в данном документе к поддающемуся обнаружению соединению или составу, которое конъюгировано непосредственно или опосредованно к молекуле (такой как нуклеиновая кислота, полипептид или антитело), также как получение "меченой" молекулы. Метка может быть определена сама по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, может катализировать химические изменения соединения или состава субстрата, приводя к поддающемуся обнаружению продукту.
Под "твердой фазой" подразумевают неводный матрикс, к которому может прилипать молекула (такая как нуклеиновая кислота, полипептид или антитело). Примеры твердых фаз, включенные в данный документ, включают в себя фазы, образующие частично или полностью стекло (например, стекло с контролируемым размером пор), полисахариды (например, агароза), полиакриламиды, полистирол, поливиниловый спирт и силикон. В определенных вариантах осуществления, в зависимости от контекста, твердая фаза может включать в себя лунку анализируемого планшета; в других она представляет собой колонку для очистки (например, колонка для аффинной хроматографии). Этот термин также включает в себя неоднородную твердую фазу из разрозненных частиц, таких как описанные в патенте США № 4275149.
"Липосома" представляет собой маленький пузырек, состоящий из различных типов липидов, фосфолипидов и/или поверхностно-активных веществ, которые полезны для доставки препарата (такого как нуклеиновая кислота, полипептид, антитело, агонист или антагонист) млекопитающим. Компоненты липосом обычно расположены в двуслойном образовании, сходном с расположением липидов в биологических мембранах.
"Малая молекула" или "малая органическая молекула" определена в данном документе как органическая молекула, имеющая молекулярный вес ниже примерно 500 дальтон.
"Олигопептид", который связывает полипептид-мишень, представляет собой олигопептид, который способен связывать полипептид-мишень с достаточной аффинностью, так что олигопептид полезен в качестве диагностического и/или терапевтического вещества в направлении полипептида. В определенных вариантах осуществления, величина связывания олигопептида с несвязавшимся, не являющимся мишенью полипептидом является меньше, чем примерно 10% связываний олигопептида с полипептидом-мишенью, как измерено, например, поверхностным плазмонным резонансным анализом. В определенных вариантах осуществления, олигопептид связывает полипептид-мишень с константой диссоциации (Kd)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ.
"Органическая молекула", которая связывает полипептид-мишень, представляет собой органическую молекулу, за исключением олигопептида или антитела, как определено в данном документе, которая способна связывать полипептид-мишень с достаточной аффинностью, так что органическая молекула полезна в качестве диагностического и/или терапевтического вещества в направлении полипептида. В определенных вариантах осуществления, величина связывания органической молекулы с несвязавшимся, не являющимся мишенью полипептидом является меньше, чем примерно 10% связываний органической молекулы с полипептидом-мишенью, как измерено, например, поверхностным плазмонным резонансным анализом. В определенных вариантах осуществления, органическая молекула связывает полипептид-мишень с константой диссоциации (Kd)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ.
"Биологические системы" представляют собой in vitro, ex vivo или in vivo систему, включающую клетки млекопитающих, которые участвуют в обычном пути передачи сигнала.
Термином "иммунное заболевание" обозначают заболевание, при котором компонент иммунной системы млекопитающих вызывает, опосредует или иначе способствует заболеваемости млекопитающих. Также охваченными являются заболевания, при которых стимуляция или воздействие иммунного ответа благоприятно действует на прогрессию заболевания. Охваченные этим термином представляют собой иммунные воспалительные заболевания, неиммунные воспалительные заболевания, инфекционные заболевания, иммуннодефицитные заболевания и неоплазия.
Термином "заболевание, опосредованное T-клетками" обозначают заболевание, при котором T-клетки непосредственно или косвенно опосредуют или иначе способствуют заболеваемости млекопитающих. Заболевание, опосредованное T-клетками, может быть связано с опосредованными клетками эффектами, опосредованными лимфокинами эффектами, и другими, и даже эффектами, связанными с B-клетками, если, например, В-клетки стимулируются секретируемыми T-клетками лимфокинами.
Как использовано в данном документе, термин "псориаз" определен как состояние, характеризующееся высыпанием ограниченных, дискретных и конфлюентных, красноватых, чистыми-чашуйчатыми макропапулами непременно на локтях, коленях, коже головы или туловище. Как использовано в данном документе, термин "опухоль" относится ко всем видам неопластического роста и пролиферации клеток, либо доброкачественных, либо злокачественных, и всем предраковым и раковым клеткам и тканям. Термины "рак", "раковый", "нарушение клеточной пролиферации", "нарушение пролиферации" и "опухоль" не являются взаимно исключающими, как упомянуто в данном документе.
Термин "опухолевая прогрессия" относится к росту и/или пролиферации опухоли.
Термины "рак" и "раковый" относятся или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется нерегулируемым клеточным ростом/пролиферацией. Примеры рака включают в себя, но ими не ограничивается, карциному, лимфому (например, лимфома Ходжкина и не Ходжкина), бластому, саркому и лейкемию. Более частные примеры таких раков включают в себя чешуйчато-клеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, сквамозную карциному легкого, рак брюшины, печеночно-клеточный рак, желудочно-кишечный рак, панкреатический рак, глиома, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак груди, колоректальный рак, рак прямой кишки, рак желудка, эндометриальную или маточную карциному, карциному слюнных желез, рак почек, рак печени, рак простаты, рак относящийся к наружным женским половым органам, рак щитовидной железы, печеночный рак, лейкемию и другие лимфопролиферативные нарушения, и различные типы рака головы и шеи.
"Аутоиммунные нарушения" или "аутоиммунность" относится к состоянию, при котором гуморальный или опосредованный клетками иммунный ответ увеличивается, противодействуя собственной ткани тела. "Аутоиммунное нарушение, опосредованное IL-23" представляет собой любое аутоиммунное нарушение, которое вызвано непрерывной или острой активностью IL-23.
"Воспаление" относится к накоплению лейкоцитов и расширению кровеносных сосудов в области повреждения или инфекции, обычно вызывая боль, опухание и красноту.
"Хроническое воспаление" относится к воспалению, при котором причина воспаления сохраняется и ее трудно или невозможно удалить.
"Аутоиммунное воспаление" относится к воспалению, связанному с аутоиммунным нарушением.
"Артритическое воспаление" относится к воспалению, связанному с артритом.
"Воспалительная болезнь кишечника" или "IBD" относится к хроническому нарушению, характеризующемуся воспалением желудочно-кишечного пути. IBD включает в себя язвенный колит, который затрагивает толстую кишку и/или прямую кишку, и болезнь Крона, которая может затрагивать целиком желудочно-кишечную систему, но более часто затрагивает тонкую кишку (подвздошная кишка) и возможно толстую кишку.
"Артриты" относятся к воспалению суставов и включают в себя, но ими не ограничивается, остеоартрит, подагру, инфекционно-связанный артрит, артрит синдрома Рейтера и артриты, связанные с аутоиммунными нарушениями, такими как ревматоидный артрит, псориатический артрит, связанный с волчанкой артрит, артрит позвонков и склеродермия-связанный артрит.
Термин "эффективное количество" представляет собой концентрацию или количество молекулы (например, нуклеиновой кислоты, полипептида, агониста или антагониста), которое приводит к достижению той или иной обозначенной цели. "Эффективное количество" можно определить опытным путем. "Терапевтически эффективное количество" представляет собой концентрацию или количество молекул, которое эффективно для достижения обозначенного терапевтического эффекта. Это количество также можно определить опытным путем.
Как использовано в данном документе, термин "цитотоксическое вещество" относится к веществу, которое подавляет или предотвращает функцию клеток и/или вызывает разрушение клеток. Термин имеет целью включить радиоактивные изотопы (например, I131, I125, Y90 и Re186), химические терапевтические вещества и токсины, такие как ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, или их фрагменты.
"Химическое терапевтическое вещество" представляет собой химическое соединение, полезное при лечении рака. Примеры химических терапевтических веществ включают в себя адриамицин, доксорубицин, эпирубицин, 5-фторурацил, цитозин арабинозид ("Ara-C"), циклофосфамид, тиотепа, бусульфан, цитоксин, таксоиды, например паклитаксел (Taxol, Bristol-Myers Squibb Oncology, Princeton, NJ), и доксетаксел (Taxotere, Rhone-Poulenc Rorer, Antony, France), токсотер, метотрексат, цисплатин, мелфалан, винбластин, блеомицин, этопозид, ифосфамид, митомицин C, митоксантрон, винкристин, винорелбин, карбоплатин, тенипозид, дауномицин, карминомицин, аминоптерин, дактиномицин, митомицины, эсперамицины (см. патент США № 4675187), мелфалан и другие связанные азотистые иприты. Также включенные в это определение представляют собой гормональные вещества, которые влияют на регуляцию и подавление действия гормона на опухоли, такие как тамоксифен и онапристон.
"Подавляющее рост вещество" при использовании в данном документе относится к соединению или составу, которое подавляет рост клетки, в частности раковой клетки, гиперэкспрессирующей любой из генов, идентифицированный в данном документе, либо in vitro, либо in vivo. Таким образом, подавляющее рост вещество представляет собой вещество, которое значительно уменьшает процент клеток, гиперэкспрессирующих такие гены в S фазе. Примеры подавляющих рост веществ включают в себя вещества, которые блокируют продвижение клеточного цикла (в другом месте, чем S фаза), такие как вещества, которые вызывают задержку в G1 и задержку M-фазы. Классические блокаторы M-фазы включают в себя винкас (винкристин и винбластин), таксол и топо-II ингибиторы, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Те вещества, которые задерживают в G1, также перетекают в задержку S-фазы, например ДНК алкилирующие вещества, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Дальнейшую информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., глава 1, озаглавленная "Cell cycle regulation, oncogens, and antineoplastic drugs" Murakami et al. (WB Saunders: Philadelphia, 1995), в частности p. 13.
Термин "цитокин" представляет собой термин, характерный для определенного рода белков, выделяемых одной популяцией клеток, который действует на другую популяцию клеток в качестве внутриклеточных медиаторов. Примерами таких цитокинов являются лимфокины, монокины и традиционные полипептидные гормоны. Включенными в число цитокинов являются гормон роста, такой как гормон роста человека, N-метионил гормон роста человека и гормон роста крупного рогатого скота; гормон паращитовидной железы; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеины гормонов, такие как фолликуло-стимулирующий гормон (FSH), тиреотропный гормон (TSH) и лютеинизирующий гормон (LH); печеночный фактор роста; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухолей-α и -β; мюллерова ингибирующая субстанция; мышиный гонадотропин-связаный пептид; ингибин; активин; фактор роста эндотелия сосудов; интегрин; тромбопоэтин (TPO); факторы роста нервов, такие как NGF-β; фактор роста тромбоцитов; трансформирующий фактор роста (TGFs), такие как TGF-α и TGF-β; инсулиноподобный фактор роста-I и -II; эритропоэтин (EPO); остеоиндуктивные факторы; интерфероны, такие как интерферон-α, -β, и -γ; колониестимулирующие факторы (CSFs), такие как макрофаг-CSF (M-CSF); гранулоцит-макрофаг-CSF (GM-CSF); и гранулоцит-CSF (G-CSF); интерлекины (ILs), такие как IL-1, IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12; фактор некроза опухоли, такой как TNF-α или TNF-β; и другие полипептидные факторы, включая LIF и лиганд из набора (KL). Как использовано в данном документе, термин «цитокин» включает в себя белки из естественных источников или культуры рекомбинантных клеток и биологически активные аналоги нативной последовательности цитокинов.
Как использовано в данном документе, термином "клетки воспаления" именуют клетки, которые усиливают воспалительный ответ, такие как мононуклеарные клетки, эозинофилы, макрофаги и полиморфноядерные нейтрофилы (PMN).
II. СОСТАВЫ И СПОСОБЫ ИЗОБРЕТЕНИЯ
A. IL-22 или IL-22R полинуклеотиды и полипептиды
В настоящем изобретении представлены выделенные IL-22 или IL-22R полипептиды и выделенные нуклеотидные последовательности, кодирующие такие полипептиды. IL-22 или IL-22R полипептиды включают в себя нативные полной длины или зрелые IL-22 или IL-22R полипептиды, а также варианты IL-22 или IL-22R. Варианты IL-22 или IL-22R можно получить путем введения соответствующих нуклеотидных изменений в ДНК IL-22 или IL-22R, и/или путем синтеза нужного IL-22 или IL-22R полипептида. Специалисты в данной области техники будут принимать во внимание, что изменения аминокислот могут изменять посттрансляционный процессинг IL-22 или IL-22R, как например изменение количества или положения участков гликозилирования или изменения характеристик прикрепления мембраны.
Изменения нативных IL-22 или IL-22R или различных доменов IL-22 или IL-22R, как описано в данном документе, можно получить, например, используя любые способы и указания для консервативных и неконсервативных мутаций, изложенные, например, в патенте США № 5364934. Изменения могут представлять собой замену, делецию или инсерцию одного или более кодонов, кодирующих IL-22 или IL-22R, которые приводят к изменению аминокислотной последовательности IL-22 или IL- 22R по сравнению с нативной последовательностью IL-22 или IL-22R. Дополнительно, изменение является заменой по крайней мере одной аминокислоты на любую другую аминокислоту в одном или более доменах IL-22 или IL-22R. Указание по определению, какой аминокислотный остаток можно встроить, заместить или удалить без отрицательного влияния на желательную активность, можно обнаружить путем сравнения последовательности IL-22 или IL-22R с последовательностью гомологичных известных белковых молекул и минимизируя количество изменений в аминокислотной последовательности, сделанных в областях с высокой гомологией. Аминокислотные замены могут быть следствием замещения одной аминокислоты на другую аминокислоту, имеющую похожие структурные и/или химические свойства, такие как замена лейцина на серин, то есть консервативные аминокислотные замены. Инсерции и делеции дополнительно могут варьировать от примерно 1 до 5 аминокислот. Допустимые изменения можно определять системно, делая инсерции, делеции или замены аминокислот в последовательности и исследуя полученные варианты на проявляемую активность первичной или зрелой нативной последовательностью.
В конкретных вариантах осуществления, интересующие консервативные замены показаны в таблице 6 под заголовком предпочтительных замен. Если такие замены приводят к изменению биологической активности, то затем вводятся более значительные изменения, обозначенные примерные замены в таблице 6, или, как далее описано ниже в ссылке на классы аминокислот, и проводят скрининг продуктов.
Значительные модификации в функции или иммунологической идентичности IL-22 или IL-22R полипептида достигаются выбранными заменами, которые значительно отличаются по их действию на поддержание (a) структуры полипептидного остова в области замены, например, как складчатая или спиральная конформация, (b) заряда или гидрофобности молекулы в области-мишени, или (c) основной части боковой цепи. Встречающиеся в природе остатки подразделяются на группы на основе общих свойств боковой цепи:
(1) гидрофобные: norleucine, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислотные: asp, glu;
(4) основные: asn, gin, his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены будут вызывать обмен члена одного из этих классов на другой класс.
Такие замещенные остатки также можно вводить в консервативные участки замен или, более предпочтительно, в остальные (неконсервативные) участки.
Изменения можно получить, используя способы, известные в данной области, такие как олигонуклеотид-опосредованный (сайт-направленный) мутагенез, сканирование аланином, и ПЦР мутагенез. сайт-направленный мутагенез [Carter et al., Nucl. Acids Res., 12:4331 (1986); Zoller et al., Nucl. Acids Res., 10:6487 (1987)], кассетный мутагенез [Wells et al., Gene, 34:315 (1985)], рестрикционный селекционный мутагенез [Wells et al., Philos. Trans. R. Soc. London SerA, 317:415 (1986)] или другие известные способы можно проводить на клонированной ДНК для получения измененной ДНК IL-22 или IL-22R.
Полипептидные фрагменты IL-22 или IL-22R также представлены в данном документе. Такие фрагменты могут подвергаться процессингу на N-конце или C-конце или могут испытывать недостаток внутренних остатков, например, при сравнении первичного нативного белка. Некоторые фрагменты не содержат аминокислотные остатки, которые не являются существенными для желательной биологической активности IL-22 или IL-22R полипептида. Поэтому в некоторых вариантах осуществления, фрагмент IL-22 или IL-22R является биологически активным. В некоторых вариантах осуществления, фрагмент первичного IL-22 не содержит N-конечную сигнальную пептидную последовательность. В некоторых вариантах осуществления, фрагмент первичного IL-22R представляет собой растворимую форму IL-22R, которая не является мембрано-связанной, например форма IL-22R, которая не содержит трансмембранный домен. Например, растворимая форма IL-22R человека не содержит всех или значительный участок трансмембранного домена из примерно 229-251 аминокислот SEQ ID NO:3.
Ковалентные изменения IL-22 или IL-22R включены в объем этого изобретения. Один вид ковалентного изменения включает в себя реагирующие нацеленные аминокислотные остатки IL-22 или IL-22R полипептида с органическим производным веществом, который способен реагировать с выбранными боковыми цепями или N- или C-концевыми остатками IL-22 или IL-22R. Получение производных с помощью бифункциональных веществ является полезным, например, для образования поперечных связей IL-22 или IL-22R с водонерастворимой основой-носителем или поверхностью при использовании в способе очистки анти-IL-22 или IL-22R антител, и наоборот. Общеупотребительные образующие поперечные связи вещества включают в себя, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаральдегид, сложные эфиры имидов N-гидроксиянтарной кислоты, эфиры гидроксисукцинимида, например сложные эфиры с 4-азидосалициловой кислоты, гомобифункциональные имидоэфиры, включая сукцинимидилэфиры, такие как 3,3'-дитиобис(сукцинимидилпропионат), бифункциональные малеимиды, такие как бис-N-малеимидо-1,8-октан, и вещества, такие как метил-3-[(п-азидофенил)дитио]пропиоимидат.
Другие изменения включают в себя дезамидирование глутаминил- и аспарагинил-остатков до соответствующих глутамил- и аспартил-остатков, соответственно, гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп серил- или треонил-остатков, метилирование α-аминогрупп лизина, аргинина и боковых цепей гистидина [T.E. Creighton, Proteins: Structure and Molecular Properties, W. H. Freeman & Co., San Francisco, pp. 79-86 (1983)], ацетилирование N-концевого амина и амидирование любой C-концевой карбоксильной группы.
Другой вид ковалентного изменения IL-22 или IL-22R полипептида, включенный в объем этого изобретения, включает изменение характера нативного гликозилирования полипептида. "Изменение характера нативного гликозилирования полипептида" предназначено для целей данного документа, чтобы обозначить удаление одного или более углеводных компонентов, обнаруженных в нативной последовательности IL-22 или IL-22R (либо перемещением исходного участка гликозилирования, либо удалением гликозилирования химическими и/или ферментными способами), и/или добавляя один или более участков гликозилирования, которые не представлены в нативной последовательности IL-22 или IL-22R. Вдобавок фраза включает в себя качественные изменения гликозилирования нативных белков, вовлекая изменение природы и свойств различных приведенных углеводных компонентов.
IL-22 или IL-22R полипептид настоящего изобретения также можно изменить некоторым образом для образования химерной молекулы, включающей IL-22 или IL-22R, слившегося с другой гетерогенной полипептидной или аминокислотной последовательностью. В одном варианте осуществления, химерная молекула включает слияние IL-22 или IL-22R с маркерным полипептидом, который представляет эпитоп, с которым может селективно связываться антимаркер антитело. Маркер эпитопа, как правило, находится на амино- или карбокси-концах IL-22 или IL-22R. Присутствие таких эпитоп-маркерных форм IL-22 или IL-22R можно определить, используя антитела против маркерного полипептида. Также предоставление маркера эпитопа облегчает IL-22 или IL-22R быстрое очищение путем аффинного очищения с использованием антимаркерных антител или другого вида аффинной основы, которая связывает маркер эпитопа. Различные маркерные полипептиды и их соответствующие антитела хорошо известны в данной области. Примеры включают в себя полигистидин (poly-his) или полигистидин-глицин (poly-his-gly) маркеры; flu HA маркерный полипептид и его антитела 12CA5 [Field et al, Mol. Cell. Biol, 8:2159-2165 (1988)]; c-myc маркер и 8F9, 3C7, 6E10, G4, B7 и 9E10 антитела к нему [Evan et al., Molecular and Cellular Biology, 5:3610-3616 (1985)]; и маркер гликопротеина D (gD) вируса простого герпеса и его антитела [Paborsky et al., Protein Engineering, 3(6):547-553 (1990)]. Другие маркерные полипептиды включают в себя Flag-пептиды [Hopp et al., BioTechnology, 6:1204-1210 (1988)]; пептид эпитопа KT3 [Martin et al., Science, 255: 192-194 (1992)]; альфа-тубулин пептид эпитопа [Skinner et al., J Biol. Chem., 266:15163-15166 (1991)]; и пептидный маркер белка T7 гена 10 [Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. USA, 87:6393-6397 (1990)].
В другом варианте осуществления, химерная молекула может включать слияние IL-22 или IL-22R полипептида с иммуноглобулином или отдельной областью иммуноглобулина. Для бивалентной формы химерной молекулы (также называемой "иммуноадгезин") такое слияние может относиться к Fc области молекулы IgG. Слияния Ig предпочтительно включают в себя замены растворимой формы IL-22 или IL-22R полипептида на место, по крайней мере, одной вариабельной области молекулы Ig. В конкретном предпочтительном варианте осуществления, слияние иммуноглобулинов включает в себя шарнирные, CH2 и CH3, или шарнирные, CH1, CH2 и CH3 области молекулы IgG1. Получение слияний иммуноглобулинов см. также в патенте США № 5428130, опубликованном 27 июня 1995 г.
1. Получение IL-22 или IL-22R
IL-22 или IL-22R можно получить обычными рекомбинантными способами, например, путем трансформирования культивируемых клеток или вектором трансфекции, содержащим нуклеиновую кислоту, кодирующую IL-22 или IL-22R, на примере нуклеиновой кислоты, показанной на фиг. 1, которая кодирует IL-22. Также представлены клетки-хозяева, содержащие любой такой вектор. В качестве примера, клетки-хозяева могут представлять собой CHO клетки, E. coli или дрожжи. Процесс получения любых описанных в данном документе полипептидов представлен далее и включает культивирование клеток-хозяев в условиях, пригодных для экспрессии нужного полипептида, и извлечение нужного полипептида из клеточной культуры.
В других вариантах осуществления, в изобретении представлены химерные молекулы, включающие любые описанные в данном документе полипептиды, слившиеся с гетерогенной полипептидной или аминокислотной последовательностью. Пример таких химерных молекул включает любые описанные в данном документе полипептиды, слившиеся с последовательностью эпитопа маркера или Fc областью иммуноглобулина.
Конечно, можно допустить, что можно применять альтернативные способы, которые хорошо известны в данной области, для получения IL-22 или IL-22R. Например, IL-22 или IL-22R последовательность или их участки можно получить прямым пептидным синтезом, используя твердофазные способы [см., например, Stewart et al., Solid-Phase Peptide Synthesis, W.H. Freeman Co., San Francisco, CA (1969); Merrifield, J. Am. Chem. Soc, 85:2149-2154 (1963)]. Белковый синтез in vitro можно осуществить ручными или автоматическими способами. Автоматизированный синтез можно выполнять, например, с использованием Applied Biosystems Peptide Synthesizer (Foster City, CA), используя инструкции изготовителя. Различные участки IL-22 или IL-22R можно раздельно химически синтезировать и объединить, используя химические или ферментные способы получения первичного IL-22 или IL-22R.
Рекомбинантно экспрессируемый IL-22 или IL-22R можно извлечь из культуральной среды или из клеточных экстрактов клеток-хозяев. Следующие процессы являются типовыми для соответствующих процессов очистки: фракционирование на ионнообменной колонке; обращенно-фазовая HPLC; хроматография на силикатной или катионнообменной смоле, такая как DEAE; хроматофокусирование; SDS-PAGE; фракционирование сульфатом аммония; гель-фильтрация с использованием, например, сефадекса G-75; протеин A сефарозные колонки для удаления примесей, таких как IgG; и металлические хелатообразующие колонки для связывания эпитоп-маркерных форм IL-22 или IL-22R. Можно использовать различные способы очистки белков, и такие способы известны в данной области и описаны, например, в Deutscher, Methods in Enzymology, 182 (1990); Scopes, Protein Purification: Principles and Practice, Springer-Verlag, New York (1982). Выбранные этапы очистки, например, будут зависеть от природы использованного процесса получения и отдельно полученного IL-22 или IL-22R.
2. Обнаружение экспрессии генов
Экспрессию гена, кодирующего IL-22 или IL-22R, можно определить различными способами в данной области, например, определяя экспрессию мРНК, кодирующую IL-22 или IL-22R. Как использовано в данном документе, термин "определение" включает в себя количественное или качественное определение. Путем определения экспрессии генов IL-22 или IL-22R можно идентифицировать, например, те ткани, которые экспрессируют ген IL-22 или IL-22R. Экспрессию гена можно измерить, используя конкретные способы, известные специалисту в данной области, например нозерн-блоттинг (Thomas, Proc. Natl. Acad. Sci. USA, 77:5201-5205 [1980]); количественная ПЦР; или гибридизация in situ с использованием соответствующим образом меченной пробы, основанной на представленных в данном документе последовательностях. Альтернативно, экспрессию генов можно измерить иммунологическими способами, такими как иммуногистохимическое окрашивание срезов тканей и анализ клеточной культуры или жидкостей организма, чтобы количественно оценить непосредственно экспрессию генного продукта. Подходящие для иммуногистохимического окрашивания и/или анализа образца жидкости антитела включают в себя любые представленные в данном документе антитела. Удобно, антитела можно получать против нативной последовательности IL-22 или IL-22R полипептида; против синтетического пептида, содержащего фрагмент IL-22 или IL-22R полипептидной последовательности; или против экзогенной последовательности, слившейся с IL-22 или IL-22R полипептидом его фрагментом (включая синтетический пептид).
B. Антитела
Представлены антитела, которые связываются с любыми выше или ниже описанными полипептидами. В одном варианте осуществления, выделенное антитело, которое связывается с IL-19, IL-20, IL-22, IL-24, IL-20Ra, IL-20Rb, IL-10R2 или IL-22R полипептидом. Типичные антитела включают в себя поликлональные, моноклональные, гуманизированные, человеческие, биспецифические и гетероконъюгированные антитела. Антитело может представлять собой фрагмент антитела, например Fab, Fab'-SH, Fv, scFv или (Fab')2 фрагмент. В одном варианте осуществления, представлено выделенное антитело, которое связывается с IL-22 или IL-22R. В одном таком варианте осуществления, антитело частично или полностью блокирует активность IL-22 или IL-22R полипептида (то есть "блокирующее" антитело).
В данном документе приведены типичные моноклональные антитела, которые связывают IL-22 и IL-22R, и далее описаны в примерах. Такие антитела включают в себя анти-IL-22 антитела, обозначенные 3F11.3 ("3F11"), 11H4.4 ("11H4") и 8E11.9 ("8E11"), и анти-IL-22R антитела, обозначенные 7E9.10.8 ("7E9"), 8A12.32 ("8A12"), 8H11.33.28 ("8H11") и 12H5. В одном варианте осуществления, представлена гибридома, которая продуцирует любые такие антитела. В одном варианте осуществления, представлены моноклональные антитела, которые конкурируют с 3F11.3, 11H4.4 или 8E11.9 за связывание с IL-22. В другом варианте осуществления, представлены моноклональные антитела, которые связываются с таким эпитопом, как 3F11.3, 11H4.4 или 8E11.9. В другом варианте осуществления, представлены моноклональные антитела, которые конкурируют с 7E9, 8A12, 8H11 или 12H5 за связывание IL-22R. В одном варианте осуществления, представлены моноклональные антитела, которые связываются с таким эпитопом, как 7E9, 8A12, 8H11 или 12H5. Различные варианты осуществления антител представлены ниже.
1. Поликлональные антитела
Антитела могут включать в себя поликлональные антитела. Способы получения поликлональных антител известны специалисту в данной области. Поликлональные антитела можно получать из млекопитающих, например, путем одного или более введения иммунизирующего вещества и вспомогательного средства, если необходимо. Обычно иммунизирующее вещество и/или вспомогательное средство вводят млекопитающему многократными подкожными или внутрибрюшинными инъекциями. Иммунизирующее вещество может включать в себя интересующий полипептид или его гибридный белок. Может быть полезно, чтобы иммунизирующее вещество соединялось с белком, известным в качестве иммунногенного у млекопитающих при иммунизации. Примеры таких иммуногенных белков включают в себя, но ими не ограничивается, гемоцианин лимфы улитки, сывороточный альбумин, бычий тиреоглобулин и соевый ингибитор трипсина. Примеры вспомогательных средств, которые можно использовать, включают в себя полный адъювант Фрейнда и MPL-TDM вспомогательное средство (монофосфорил липид A, синтетический дикориномиколат трегалозы). Протокол иммунизации может быть выбран специалистом в данной области без лишнего проведения экспериментов.
2. Моноклональные антитела
Альтернативно, антитела могут представлять собой моноклональные антитела. Моноклональные антитела можно получить, используя гибридомные способы, такие как описанные Kohler и Milstein, Nature, 256:495 (1975). В гибридомном способе мышь, хомяк или другое подходящее животное-хозяин обычно иммунизируют иммунизирующим веществом для получения лимфоцитов, которые продуцируют или способны продуцировать антитела, которые специфически связываются с иммунизирующим веществом. Альтернативно, лимфоциты можно иммунизировать in vitro.
Иммунизирующее вещество обычно будет включать в себя интересующий полипептид или его гибридный белок. Как правило, используются либо лимфоциты периферической крови ("PBLs"), если нужны клетки из человеческого источника, либо используются спленоциты или клетки лимфатических узлов, если нужны источники нечеловеческих млекопитающих. Затем лимфоциты сливаются с иммортализированной клеточной линией, используя подходящее вещество для слияния, такое как полиэтиленгликоль, чтобы получить гибридомную клетку [Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]. Иммортализированные клеточные линии обычно представляют собой трансформированные клетки млекопитающих, в частности миеломные клетки из источника грызунов, бычьего и человеческого. Обычно используют крысиные или мышиные миеломные клеточные линии. Гибридомные клетки можно культивировать в подходящей культуральной среде, которая предпочтительно содержит одно или более веществ, которые подавляют рост или выживаемость негибридных иммортализированных клеток. Например, если родительские клетки не содержат фермент гипоксантин гуанин фосфорибозил трансферазу (HGPRT или HPRT), то культуральная среда для гибридом обычно будет включать в себя гипоксантин, аминоптерин и тимидин ("HAT среда"), вещества которой препятствуют росту HGPRT-дефицитных клеток.
Предпочтительные иммортализированные клеточные линии представляют собой линии, которые эффективно сливаются, поддерживают стабильно высокий уровень экспрессии антител выделенными антитело-продуцирующими клетками и являются чувствительными к среде, такой как HAT среда. Более предпочтительные иммортализированные клеточные линии представляют собой мышиные миеломные линии, которые можно получить, например, из Salk Institute Cell Distribution Center, San Diego, California и the American Type Culture Collection, Manassas, Virginia. Клеточные линии миеломы человека и мышино-человеческой гетеромиеломы также были описаны для получения человеческих моноклональных антител [Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63].
Культуральную среду, в которой культивируют гибридомные клетки, затем можно исследовать на присутствие моноклональных антител, которые связываются с интересующим полипептидом. Предпочтительно, специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют иммунопреципитацией или анализом связывания in vitro, таким как радиоиммуноанализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие способы и исследования известны в данной области. Аффиность связывания моноклонального антитела можно, например, определить с помощью Scatchard анализа Munson и Pollard, Anal. Biochem., 107:220 (1980).
После того как идентифицируют нужные гибридомные клетки, клоны можно субклонировать, ограничивая способы разведения и рост стандартными способами [Goding, выше]. Подходящая клеточная среда для этой цели включает в себя, например, Дульбекко модифицированную среду Игла и RPMI-1640 среду. Альтернативно, гибридомные клетки можно выращивать in vivo, внутрибрюшинно у млекопитающего.
Моноклональные антитела, выделенные из субклонов, можно разделить или очистить от культуральной среды или асцитной жидкости стандартными способами очистки иммуноглобулинов, такими как, например, белковая A-сефароза, гидроксилапатитная хроматография, гель электрофорез, диализ или аффинная хроматография.
Моноклональные антитела можно получить, используя комбинаторные библиотеки для скрининга антител с нужной активностью или активностями. Например, различные способы известны в данной области для получения библиотек фаговых дисплеев и скрининга таких библиотек для антител, обладающих нужными характеристиками связывания. Такие способы, как правило, описаны в Hoogenboom et al. (2001) в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ), и в некоторых вариантах осуществления в Lee et al. (2004) J. Mol. Biol. 340:1073-1093.
По существу, клоны синтетических антител выбирают скринингом фаговых библиотек, содержащих фаг, у которого обнаружены различные фрагменты вариабельной области антител (Fv), слившейся с белком оболочки фага. Такие фаговые библиотеки разделяются аффинной хроматографией в зависимости от нужного антигена. Клоны, экспрессирующие Fv фрагменты, способные связывать нужный антиген, абсорбируются к антигену и таким образом отделяются от не связавшихся клонов в библиотеке. Связавшиеся клоны затем элюируют от антигена и могут быть в дальнейшем обогащены дополнительными циклами адсорбции/элюции антигена. Любые антитела изобретения можно получить путем планирования подходящего способа скрининга антигенов, чтобы выбрать интересующий фаговый клон с последующим конструированием клона первичных антител, используя Fv последовательности из интересующего фагового клона и последовательностей подходящей константной области (Fc), описанной в Kabat et al, Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3.
Моноклональные антитела также можно получать способами рекомбинантной ДНК, такими как способы, описанные в патенте США № 4816567. ДНК, кодирующую моноклональные антитела изобретения, можно легко выделить и секвенировать с использованием общепринятых способов (например, используя олигонуклеотидные пробы, которые способны специфично связывать гены, кодирующие тяжелые и легкие цепи мышиных антител). Гибридомные клетки изобретения служат в качестве предпочтительного источника такой ДНК. Однажды выделенную ДНК можно помещать в экспрессирующие векторы, которыми затем трансфицируют клетки-хозяева, такие как COS клетки обезьяны, клетки яичника китайского хомячка (CHO) или клетки миеломы, которые по-другому не продуцируют белок иммуноглобулина, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Также ДНК можно модифицировать, например, путем замены кодирующей последовательности для константных доменов легкой и тяжелой цепей человека вместо гомологичных мышиных последовательностей [патент США № 4816567; Morrison et al., выше] или путем ковалентного соединения с кодирующей последовательностью иммуноглобулина, целой или частью кодирующей последовательности для неиммуноглобулинового полипептида. Такой неиммуноглобулиновый полипептид может быть замещен на константные домены антитела изобретения или может быть замещен на различные домены антигенсвязывающего участка антитела изобретения, чтобы получить химерное бивалентное антитело.
3. Моновалентные антитела
Также представлены моновалентные антитела. Способы получения моновалентных антител хорошо известны в данной области. Например, один способ включает в себя рекомбинантную экспрессию легкой цепи иммуноглобулинов и модифицированной тяжелой цепи. Как правило, тяжелую цепь усекают в любой точке Fc области, для того чтобы предотвратить поперечное связывание тяжелых цепей. Альтернативно, соответствующие остатки цистеина замещаются другим аминокислотным остатком или удаляется, для того чтобы предотвратить поперечное связывание.
Способы in vitro также подходят для получения моновалентных антител. Расщепление антител для получения их фрагментов, в частности Fab фрагментов, может производиться с использованием обычных способов, известных в данной области.
4. Фрагменты антител
Также представлены фрагменты антител. Фрагменты антител можно получать традиционными способами, такими как ферментное расщепление, или рекомбинантными способами. В отдельных случаях есть преимущества использования фрагментов антител, чем целых антител. Более маленький размер фрагментов способствует быстрому выведению и может приводить к улучшенному доступу к солидным опухолям. Для обзора определенных фрагментов антител см. Hudson et al. (2003) Nat. Med. 9:129-134.
Для получения фрагментов антител были разработаны различные способы. Традиционно, эти фрагменты получали путем протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24: 107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако в настоящее время эти фрагменты могут получать непосредственно из рекомбинантных клеток-хозяев. Fab, Fv и ScFv фрагменты антител все могут экспрессироваться и секретироваться в E. coli, таким образом позволяя легко получить большое количество этих фрагментов. Фрагменты антител можно выделить из библиотек фаговых антител, обсуждаемых выше. Альтернативно, Fab'-SH фрагменты можно извлекать непосредственно из E. coli и химически соединять для образования F(ab')2 фрагментов (Carter et al., Bio/Technology 10: 163-167 (1992)). Согласно другому подходу, F(ab')2 фрагменты можно выделить непосредственно из рекомбинантной культуры клеток-хозяев. Fab и F(ab')2 фрагмент с увеличенным периодом полужизни in vivo, содержащие сохраненный остаток рецепторсвязывающей антигенной детерминанты, описан в патенте США № 5869046. Другие способы получения фрагментов антител будут очевидны для квалифицированного практика. В некоторых вариантах осуществления, антитело представляет собой одноцепочечный Fv фрагмент (scFv). См. WO 93/16185; патенты США № 5571894 и 5587458. Fv и scFv представляют собой единственные виды с интактными связывающими участками, которые лишены константных областей; таким образом, они могут подходить для уменьшения неспецифического связывания при использовании in vivo. scFv гибридные белки можно конструировать до получения слияния эффекторного белка либо на амино-, либо карбокси-конце scFv. См. Antibody Engineering, ed. Borrebaeck, выше. Фрагмент антитела также может представлять собой "линейное антитело", например, как описано в патенте США № 5641870. Такие линейные антитела могут быть моноспецифическими или биспецифическими.
5. Гуманизированные антитела
Также представлены гуманизированные антитела. В данной области известны различные способы превращения нечеловеческих антител в гуманизированные. Например, гуманизированное антитело может содержать один или более аминокислотных остатка, введенных в него из нечеловеческого источника. Эти нечеловеческие аминокислотные остатки часто обозначаются как "импортируемые" остатки, которые обычно взяты из "импортируемого" вариабельного домена. Гуманизацию можно преимущественно осуществлять способом Winter и коллегами (Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-327; Verhoeyen et al. (1988) Science 239:1534-1536), замещая последовательности гипервариабельной области на соответствующие последовательности антител человека. Соответственно, такие "гуманизированные" антитела представляют собой химерные антитела (патент США № 4816567), которые значительно в меньшей степени, чем интактный человеческий вариабельный домен, замещались соответствующей последовательностью из нечеловеческих видов. На практике, гуманизированные антитела представляют собой обычные человеческие антитела, в которых несколько остатков гипервариабельной области и возможно несколько FR остатков замещаются остатками из аналогичных участков антител грызунов.
Выбор человеческих вариабельных доменов как легких, так и тяжелых для использования в получении гуманизированных антител может быть важным, чтобы уменьшить антигенность. Согласно так называемому " наилучше подобранному" способу, последовательность вариабельного домена антитела грызуна подвергают скринингу относительно целой библиотеки известных последовательностей человеческих вариабельных доменов. Человеческая последовательность, которая наиболее близка к последовательности грызунов, затем принимается в качестве человеческой основы для гуманизированного антитела (Sims et al. (1993) J. Immunol. 151:2296; Chothia et al. (1987) J. Mol. Biol. 196:901). В другом способе используют специальную основу, полученную из консенсусной последовательности всех антител человека из отдельной подгруппы легких или тяжелых цепей. Аналогичная основа может использоваться для нескольких различных гуманизированных антител (Carter et al. (1992) Proc. Natl. Acad. Sci. USA, 89:4285; Presta et al. (1993) J. Immunol., 151:2623).
В дальнейшем, как правило, желательно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и другими благоприятными биологическими свойствами. Для осуществления этой задачи, согласно одному способу, получали гуманизированные антитела с помощью способа анализа родительских последовательностей и различных воображаемых гуманизированных продуктов, используя трехмерные модели родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов обычно доступны и хорошо известны специалистам в данной области. Существуют компьютерные программы, которые изображают и показывают возможные трехмерные конформационные структуры из выбранных последовательностей кандидатов иммуноглобулинов. Изучение этих изображений дает возможность анализировать возможную роль остатков в функционировании последовательности кандидата иммуноглобулина, то есть анализ остатков, которые влияют на способность кандидата иммуноглобулина связывать его антиген. Таким образом, можно выбирать FR остатки и комбинировать с последовательностями реципиента и импортированными, так что достигается нужная характеристика антитела, такая как увеличенная аффинность для антигена-мишени. Вообще, остатки гипервариабельной области непосредственно и наиболее значительно влияют на связывание антигена.
6. Человеческие антитела
Также представлены человеческие антитела. Можно создавать человеческие антитела, соединяя Fv клонированные последовательности вариабельного домена, выделенные из производных от человека библиотек фаговых дисплеев с известными человеческими последовательностями константных доменов, как описано выше. Альтернативно, человеческие моноклональные антитела изобретения можно получить гибридомным способом. Были описаны клеточные линии миеломы человека и мышино-человеческой гетеромиеломы для получения человеческих моноклональных антител, например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147: 86 (1991).
В настоящее время возможно получение трансгенных животных (например, мышей), которые способны после иммунизации продуцировать полный набор человеческих антител в отсутствие эндогенной продукции иммуноглобулинов. Например, было описано, что гомозиготная делеция гена области связывания тяжелой цепи антитела (JH) в химерных и зародышевой линии мутантных мышах приводит к полному ингибированию продукции эндогенных антител. Перемещение группы человеческой зародышевой линии гена иммуноглобулина в такой зародышевой линии мутантных мышей будет приводить к продукции человеческих после антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255 (1993); Bruggermann et al., Year in Immunol., 7: 33 (1993).
Также может использоваться перетасовка генов для получения человеческих антител из нечеловеческих, например антитела грызунов, где человеческое антитело обладает аналогичной аффинностью к первоначальному нечеловеческому антителу. Согласно этому способу, который также называется "импринтинг эпитопа", вариабельную область либо тяжелой, либо легкой цепи фрагмента нечеловеческого антитела, полученную способами фагового дисплея, как описано в данном документе, перемещают с набором генов человеческого V домена, создавая популяцию химер с нечеловеческой цепью/человеческой цепью scFv или Fab. Разделение с антигеном приводит к выделению химерных scFv или Fab с нечеловеческой цепью/человеческой цепью, в которых человеческая цепь восстанавливает антигенсвязывающий участок, разрушенный после перемещения соответствующей нечеловеческой цепи в исходном клоне фагового дисплея, то есть эпитоп определяет (импринт) выбор партнера человеческой цепи. Если процесс повторяют, для того чтобы переместить оставшуюся нечеловеческую цепь, то получают человеческое антитело (см. PCT WO 93/06213, опубликованный 1 апреля 1993 г.). В отличие от традиционного гуманизирования нечеловеческих антител путем CDR слияния, в этом способе представлены полностью человеческие антитела, которые не имеют никаких FR или CDR остатков нечеловеческого происхождения.
7. Биспецифические антитела
Также представлены биспецифические антитела. Биспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностями связывания, по крайней мере, двух различных антигенов. В некоторых вариантах осуществления, биспецифические антитела представляют собой человеческие или гуманизированные антитела. В некоторых вариантах осуществления, одна из специфичностей связывания служит для интересующего полипептида, а другая - для любого другого антигена. В некоторых вариантах осуществления, биспецифические антитела могут связываться с двумя различными эпитопами интересующего полипептида. Биспецифические антитела также можно использовать для локализации цитотоксических веществ в клетках, которые экспрессируют интересующий полипептид, такой полипептид клеточной поверхности. Эти антитела содержат TAT226-связывающее плечо, которое связывает цитотоксическое вещество, такое как, например, сапорин, антиинтерферон-α, алкалоид барвинки(а), рициновая A цепь, метотрексат или радиоизотопный гаптен. Биспецифические антитела можно получить в виде первичных антител или фрагментов антител (например, F(ab')2 биспецифические антитела).
Способы получения биспецифических антител известны в данной области. Традиционно, рекомбинантная продукция биспецифических антител основывается на коэкспрессии двух пар тяжелая цепь - легкая цепь иммуноглобулинов, в которых две тяжелые цепи обладают различными специфичностями (Milstein и Cuello, Nature, 305: 537 (1983)). Вследствие независимого расхождения в мейозе генов тяжелых и легких цепей иммуноглобулинов, эти гибридомы (квадрогибридомы) продуцируют потенциальную смесь из 10 различных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Очистка правильной молекулы, которая обычно проводится на этапах аффинной хроматографии, достаточно трудоемка, и выход продукта низкий. Аналогичные способы раскрыты в WO 93/08829, опубликованном 13 мая 1993 г., и в Traunecker et al., EMBO J., 10: 3655 (1991).
Согласно другому подходу, вариабельные домены антител с нужными специфичностями связывания (антитело-антиген связывающие участки) сливаются с последовательностями константных доменов иммуноглобулинов. Слияние, например, происходит с константным доменом тяжелой цепи иммуноглобулина, содержащего, по крайней мере, часть шарнирных, CH2 и CH3 областей. В некоторых вариантах осуществления, первая константная область тяжелой цепи (CH1), содержащая необходимый для связывания легкой цепи участок, присутствует, по крайней мере, в одном из их слияний. ДНК, кодирующие слияния тяжелых цепей иммуноглобулинов и, если нужно, легкую цепь иммуноглобулинов, вводят внутрь выделенных экспрессирующих векторов, и проводят котрансфекцию подходящего организма хозяина. Это обеспечивает значительную гибкость в регуляции общих соотношений трех полипептидных фрагментов в вариантах осуществления, если использованные в конструкции неравноценные соотношения трех полипептидных цепей обеспечивают оптимальный выход. Однако возможно вставить кодирующие последовательности двух или всех трех полипептидных цепей в один экспрессирующий вектор, если экспрессия, по крайней мере, двух полипептидных цепей в равных соотношениях приводит к высокому выходу или если соотношения не имеет никакого особенного значения.
В одном варианте осуществления этого подхода, биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном плече и гибридной пары тяжелая-легкая цепь иммуноглобулина (обеспечивая вторую специфичность связывания) в другом плече. Было обнаружено, что эта асимметричная структура способствует разделению нужного биспецифического соединения от нежелательных сочетаний цепей иммуноглобулинов, так как присутствием легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивается легкий путь разделения. Этот подход раскрыт в WO 94/04690. Дальнейшие подробности образования биспецифических антител см., например, в Suresh et al., Methods in Enzymology, 121:210 (1986).
Согласно другому подходу, можно создать поверхность раздела между парой молекул антител, чтобы максимально увеличить процент гетеродимеров, полученных из рекомбинантной культуры клеток. Поверхность раздела содержит, по крайней мере, часть домена CH3 из константного домена антитела. В этом способе, одну или более небольшие боковые аминокислотные цепи с поверхности раздела первой молекулы антитела замещают на более большие боковые цепи (например, тирозин или триптофан). Компенсационные "полости" одного и того же или сходного размера с большими боковыми цепями создаются на поверхности раздела второй молекулы антитела путем замещения больших боковых аминокислотных цепей на более маленькие (например, аланин или треонин). Это обеспечивает механизм увеличения выхода гетеродимера относительно других нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела включают в себя поперечно сшитые или "гетероконъюгированные" антитела. Например, одно из антител в гетероконъюгате может связываться с авидином, другое - с биотином. Предполагают, что такие антитела, например, направляют клетки иммунной системы на нежелательные клетки (патент США № 4676980) и для лечения HIV инфекции (WO 91/00360, WO 92/00373 и EP 03089). Можно получить гетероконъюгированные антитела, используя любой подходящий поперечно-сшивающий способ. Подходящие поперечно-сшивающие вещества известны в данной области и раскрыты в патенте США № 4676980 вместе с рядом поперечно-сшивающих способов.
Способы образования биспецифических антител из фрагментов антител также был описан в литературе. Например, биспецифические антитела можно получать с использованием химической связи. Brennan et al., Science, 229: 81 (1985) описывают способ, в котором интактные антитела протеолитически расщепляются до образования F(ab')2 фрагментов. Присутствие дитиол комплексообразующего арсенита натрия редуцирует эти фрагменты, чтобы стабилизировать смежные дитиолы и предотвратить образование межмолекулярных дисульфидных связей. Образованные Fab' фрагменты затем преобразовывают в производные тионитробензоата (TNB). Одно из производных Fab'-TNB затем возвращается в прежнее состояние Fab'-тиол, редуцируя меркаптоэтиламином, и смешивается с эквимолярным количеством другого производного Fab'-TNB для получения биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве веществ для избирательной иммобилизации ферментов.
Последние достижения могут упростить непосредственное получение Fab'-SH фрагментов из E. coli, которые могут химически соединятся для образования биспецифических антител. Shalaby et al., J. Exp. Med., 175:217-225 (1992) описывает продукцию молекулы полностью гуманизированного биспецифического антитела F(ab')2. Каждый Fab' фрагмент был отдельно выделен из E. coli и подвергнут прямому химическому связыванию для образования биспецифического антитела. Образованные таким образом биспецифические антитела были способны связываться с клетками, гиперэкспрессирующими HER2 рецептор, и нормальными Т-клетками человека, а также как система запуска литической активности человеческих цитотоксических лимфоцитов относительно мишеней опухолей груди человека.
Также были описаны различные способы получения и разделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, были получены биспецифические антитела с использованием leucine zippers. Kostelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды leucine zipper из Fos и Jun белков были привязаны к Fab' областям двух различных антител путем генного слияния. Гомодимеры антител были разрезаны в шарнирной области для образования мономеров и затем обратно окислены для образования гетеродимеров антител. Этот способ также можно использовать для получения гомодимеров антител. В "diabody" способе, описанном Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993), был представлен альтернативный механизм получения фрагментов биспецифических антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) линкером, который слишком короткий для образования пар между двумя доменами на гомологичной цепи. Соответственно, VH и VL домены одного фрагмента вынуждены спариваться с комплементарными VL и VH доменами другого фрагмента, тем самым образуя антигенсвязывающий участок. Также сообщался другой способ получения фрагментов биспецифических антител с использованием одноцепочечных Fv (sFv) димеров. См. Gruber et al., J. Immunol., 152:5368 (1994).
Рассматриваются антитела более чем с двумя валентностями. Например, можно получить триспецифические антитела. Tutt et al., J. Immunol. 147: 60 (1991).
8. Мультивалентные антитела
Также представлены мультивалентные антитела. Мультивалентное антитело может быть интернализировано (и/или катаболизировано) быстрее, чем бивалентное антитело, клеточной экспрессией антигена, с которым связываются антитела. Антитела настоящего изобретения могут представлять собой мультивалентные антитела (которые отличаются от класса IgM) с тремя или более антигенсвязывающими участками (например, тетравалентные антитела), которые легко можно получить рекомбинантной экспрессией нуклеиновых кислот, кодирующих полипептидные цепи антитела. Мультивалентное антитело может содержать домен димеризации и три или более антигенсвязывающих участка. В некоторых вариантах осуществления, домен димеризации содержит (или состоит из) Fc область или шарнирную область. По этому сценарию, антитело будет содержать Fc область и три или более антигенсвязывающих участка на амино-конце Fc области. В некоторых вариантах осуществления, мультивалентноое антитело содержит (или состоит из) от трех до примерно восьми антигенсвязывающих участков. В одном таком варианте осуществления, мультивалентное антитело содержит (или состоит из) четырех антигенсвязывающих участков. Мультивалентное антитело содержит, по крайней мере, одну полипептидную цепь (например, две полипептидные цепи), в которой полипептидная цепь(и) содержит два или более вариабельных домена. Например, полипептидная цепь(и) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидную цепь Fc области, X1 и X2 представляют собой аминокислоту или полипептид, и n представляет собой 0 или 1. Например, полипептидная цепь(и) может содержать: VH-CH1-гибкая связь-VH-CH1-Fc цепь области; или VH-CH1-VH-CH1-Fc цепь области. Мультивалентное антитело в данном документе далее может содержать, по крайней мере, два (например, четыре) полипептида вариабельных доменов легких цепей. Мультивалентное антитело в данном документе может, например, содержать от примерно двух до примерно восьми полипептидов вариабельных доменов легких цепей. Полипептиды вариабельных доменов легких цепей, рассмотренные в данном документе, содержат вариабельный домен легких цепей и, дополнительно, далее содержат CL домен.
9. Однодоменные антитела
Также представлены однодоменные антитела. Однодоменное антитело представляет собой простую полипептидную цепь, содержащую весь или участок вариабельного домена тяжелой цепи или весь или участок вариабельного домена легкой цепи антитела. В некоторых вариантах осуществления, однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; см., например, патент США № 6248516 B1). В одном варианте осуществления, однодоменное антитело состоит из целого или участка вариабельного домена тяжелой цепи антитела.
10. Варианты антител
В некоторых вариантах осуществления рассматриваются модификации в аминокислотных последовательностях антител, описанных в данном документе. Например, может быть желательным улучшить аффинность связывания и/или других биологических свойств антитела. Варианты аминокислотных последовательностей можно получить путем введения соответствующих изменений в нуклеотидную последовательность, кодирующую антитело, или путем пептидного синтеза. Такие модификации включают в себя, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Можно производить любое сочетание делеции, инсерции и замены, чтобы получить конечный конструкт, при условии, что конечный конструкт обладает нужными характеристиками. Аминокислотные изменения можно вводить в аминокислотную последовательность антитела индивида во время получения последовательности.
Удобный способ для идентификации некоторых остатков или областей антитела, которые являются предпочтительными местами для мутагенеза, называется "сканирующий аланином мутагенез", как описано Cunningham и Wells (1989) Science, 244: 1081-1085. В данном документе идентифицирован остаток или группа остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) и замещен нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), для того чтобы действовать на взаимодействие аминокислот с антигеном. Такие аминокислотные локализации, показывающие функциональную чувствительность к заменам, затем улучшают введением далее или в, или из других вариантов в участки замены. Таким образом, предварительно определяется целый участок для введения варианта аминокислотной последовательности, природа мутации per se не нуждается в предварительном определении. Например, чтобы проанализировать выполнение мутации в данном участке, сканирование аланином или неспецифический мутагенез осуществляется в кодоне мишени или области, и проводят скрининг экспрессируемых иммуноглобулинов на нужную активность.
Вставки аминокислотных последовательностей включают в себя амино- и/или карбоксил-концевые слияния, колеблющиеся по длине от одного остатка до полипептида, состоящего из ста или более остатков, а также и вставки внутрь последовательностей из одиночных или множественных аминокислотных остатков. Примеры конечных вставок включают в себя антитело с остатком метионина на N-конце. Другие инсерционные варианты молекулы антител включают в себя слияние N- или C-концов антитела с ферментом (например, с ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
В некоторых вариантах осуществления, изменяют антитело изобретения, чтобы увеличить или уменьшить величину гликозилирования антитела. Гликозилирование полипептидов обычно является либо N-связанным или O-связанным. N-связанное относится к прикреплению углеводородного фрагмента к боковой цепи аспарагинового остатка. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту кроме пролина, представляют собой различные последовательности для прикрепления фермента углеводородного фрагмента к боковой цепи аспарагина. Таким образом, присутствие этих трипептидных последовательностей в полипептиде приводит к образованию потенциального участка гликозилирования. O-связанное гликозилирование относится к прикреплению одного из сахаров N-ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, наиболее часто к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Вставку или делецию участка гликозилирования в антитело удобно осуществлять, изменяя аминокислотную последовательность, так что получается или перемещается одна или более вышеописанные трипептидные последовательности (для N-связанных участков гликозилирования). Можно также производить изменение путем вставки, делеции или замены одного или более остатков серина или треонина в последовательности исходного антитела (для O-связанных участков гликозилирования).
Если антитело содержит Fc область, то прикрепленный к нему углеводород можно изменить. Например, антитела со зрелой углеводородной структурой, которые не содержат фукозу, прикрепленную к Fc области антитела, описаны в патенте США 2003/0157108 (Presta L.). См. также патент США 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Антитела с разделяющим N-ацетилглюкозамином (GlcNAc) в углеводороде, прикрепленном к Fc области антитела, упомянуты в WO 2003/011878, Jean-Mairet et al. и в патенте № 6602684, Umana et al. Об антителах, по крайней мере, с одним остатком галактозы в олигосахариде, прикрепленном к Fc области антитела, сообщают в WO 1997/30087, Patel et al. См. также WO 1998/58964 (Raju S.) и WO 1999/22764 (Raju S.) об антителах с измененным прикрепленным углеводородом к их Fc области. См. также в патенте США 2005/0123546 (Umana et al.) антигенсвязывающие молекулы с измененным гликозилированием.
В некоторых вариантах осуществления, вариант гликозилирования содержит Fc область, в которой углеводородная структура, прикрепленная к Fc области, не содержит фукозу. В таких вариантах улучшена ADCC функция. Дополнительно, Fc область далее содержит одну или более аминокислотных замены, которые в дальнейшем улучшают ADCC, например замены в положениях Fc области 298, 333 и/или 334 (Eu нумерация остатков). Примеры публикаций, отнесенных к "дефукозилированным" или "фукозодефицитным" антителам, включают в себя: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; Okazaki et al., J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al., Biotech. Bioeng. 87:614 (2004). Примеры клеточных линий, продуцирующих дефукозилированные антитела, включают в себя Lec13 CHO клетки, дефицитные по белковому фукозилированию (Ripka et al., Arch. Biochem. Biophys. 249:533-545 (1986); патентную заявку США № 2003/0157108 A1, Presta L.; и WO 2004/056312 A1, Adams et al., в частности пример 11), и нокаутные линии клеток, такие как по гену альфа-1,6-фукозилтрансферазы, FUT8, нокаутные CHO клетки (Yamane-Ohnuki et al., Biotech. Bioeng. 87: 614 (2004)).
Другой вариант представляет собой вариант аминокислотной замены. Эти варианты имеют, по крайней мере, один аминокислотный остаток в молекуле антитела, замещенный другим остатком. Интересные для заместительного мутагенеза участки включают в себя гипервариабельные области, но также предполагаются FR изменения. Консервативные замены показаны в таблице 6 выше под заголовком "предпочтительные замены". Если такие замены приводят к нужному изменению биологической активности, тогда можно вводить более существенные изменения, называемые "типичные замены" в таблице 6, или, как далее описано выше в отношении к классам аминокислот, и полученные антитела скринировали на желательные свойства связывания.
Один тип заместительного варианта включает в себя замещение одного или более остатков гипервариабельной области родительского антитела (например, гуманизированное или человеческое антитело). Как правило, полученные варианты, выбранные для дальнейшего развития, будут модифицированы (например, улучшены) биологические свойства по сравнению с родительским антителом, из которого они получены. Удобный способ для получения таких заместительных вариантов включает в себя созревание аффинности с использованием фагового дисплея. Кратко, несколько участков гипервариабельной области (например, 6-7 участков) мутируют для образования всех возможных аминокислотных замен в каждом участке. Полученные таким образом антитела изображаются от частиц нитчатых фагов, как слияния, по крайней мере, части белка оболочки фага (например, продукта гена III M13), упакованного внутри каждой частицы. Затем проводят скрининг вариантов фаговых дисплеев на их биологическую активность (например, аффинность связывания). Для того чтобы идентифицировать кандидатные участки гипервариабельной области для модификации, можно проводить сканирующий мутагенез (например, сканирующий аланином) для идентификации остатков гипервариабельной области, вносящих значительный вклад в связывание антигена. Альтернативно, или дополнительно, может оказаться полезным анализировать кристаллическую структуру комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактирующие остатки и соседние остатки являются кандидатами для замены в соответствии со способами, известными в данной области, включая способы, представленные в данном документе. Если создают такие варианты, то группу вариантов подвергают скринингу, используя способы, известные в данной области, включая способы, описанные в данном документе, и антитела с лучшими свойствами в одном или более соответствующих исследованиях можно отбирать для дальнейшего усовершенствования.
Молекулы нуклеиновых кислот, кодирующих варианты аминокислотных последовательностей антител, получают различными способами, известными в данной области. Эти способы включают в себя, но ими не ограничиваются, выделение из естественного источника (в случае встречающихся в природе вариантов аминокислотных последовательностей) или получение путем олигонуклеотид-опосредованного (или сайт-специфического) мутагенеза, ПЦР мутагенеза и кассетного мутагенеза полученного ранее варианта или инвариантной версии антитела.
Может быть желательно, чтобы введение одной или более аминокислотной модификации в Fc область антитела изобретения способствовало образованию варианта Fc области. Вариант Fc области может содержать человеческую последовательность Fc области (например, человеческую Fc область IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одном или более аминокислотных положениях, включаю замену шарнирного цистеина.
Согласно этому описанию и обучению уровня техники, предполагают, что в некоторых вариантах осуществления, антитело изобретения может содержать одно или более изменений по сравнению с вариантом антитела дикого типа, например в Fc области. Эти антитела, вероятно, все же в значительной степени сохраняют аналогичные характеристики, необходимые для терапевтического пользования, по сравнению с вариантом дикого типа. Например, полагают, что некоторые изменения можно получить в Fc области, которая вероятно приводит к измененному (то есть, либо улучшенному, либо уменьшенному) C1q связыванию и/или комлемент-зависимой цитотоксичности (CDC), например, как описано в WO99/51642. См. также Duncan & Winter Nature 322:738-40 (1988); патент США № 5648260; патент США № 5624821; и WO 94/29351, касающиеся других примеров вариантов Fc области. В WO 00/42072 (Presta) и WO 2004/056312 (Lowman) описывают варианты антител с улучшенным или уменьшенным связыванием с FcRs. Содержание этих патентных публикаций включено в частности, в данный документ посредством ссылки. См. также Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001). Антитела с увеличенными временем полужизни и улучшенным связыванием с неонатальным Fc рецептором (FcRn), который отвечает за перемещение материнских IgG к плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US2005/0014934A1 (Hinton et al.). Эти антитела содержат Fc область с одной или более заменами, в которых улучшено связывание Fc области с FcRn. Варианты полипептидов с измененными аминокислотными последовательностями Fc области и улучшенной или увеличенной способностью связывания C1q описаны в патенте США № 6194551B1, WO99/51642. Содержание этих патентных публикаций включено, в частности, в данный документ посредством ссылки. См. также Idusogie et al., J. Immunol. 164: 4178-4184 (2000).
В одном аспекте в изобретении представлены антитела, содержащие модификации в области контакта Fc полипептидов, содержащих Fc область, где модификации способствуют и/или вызывают гетеродимеризацию. Эти модификации включают введение выпячивания внутри первого Fc полипептида и полость внутри второго Fc полипептида, где выпячивание устанавливается в полость с тем, чтобы вызвать образование комплекса первого и второго Fc полипептидов. Способы образования антител с этими модификациями известны в данной области, например, как описанные в патенте США № 5731168.
11. Производные антител
Антитела можно далее модифицировать для включения дополнительных небелковых компонентов, которые известны в данной области и легкодоступны. Предпочтительно, подходящие компоненты для получения производных антитела представляют собой водорастворимые полимеры. Неограниченные примеры водорастворимых полимеров включают в себя, но ими не ограничиваются, полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо случайные сополимеры), и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные многоатомные спирты (например, глицерин), поливиниловый спирт и их смеси. Пропионовый альдегид полиэтиленгликоля может иметь преимущество при производстве вследствие его стабильности в воде. Полимер может иметь любой молекулярный вес, и может быть разветвленным или неразветвленным. Количество полимеров, прикрепленных к антителу может варьировать, и если прикреплено более, чем один полимер, то они могут быть одни и те же или разные молекулы. Как правило, число и/или тип полимеров, использованных для получения производных, может определяться в зависимости от факторов, включающих в себя, но ими не ограничиваясь, особые свойства или функции антитела для улучшения, будет ли производное антитело использоваться в терапии в определенных состояниях, и так далее.
В другом варианте осуществления, представлены конъюгаты антитела и небелковый фрагмент, которые выборочно можно нагреть радиоактивным облучением. В одном варианте осуществления, небелковый фрагмент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. 102: 11600-11605 (2005)). Облучение может быть любых длин волн и включает в себя, но ими не ограничивается, длины волн, которые не вредят обычным клеткам, но которые повреждают небелковый фрагмент при температуре, при которой убиваются клетки, близкие к антитело-небелковому фрагменту.
В некоторых вариантах осуществления, антитело может быть меченым и/или может быть иммобилизировано на твердой основе. В дальнейшем аспекте антитело представляет собой антиидиотипическое антитело.
12. Гетероконъюгированные антитела
Также представлены гетероконъюгированные антитела. Гетероконъюгированные антитела содержат два ковалентно связанных антитела. Такие антитела, например, направлены на нацеливание клеток иммунной системы на нежелательные клетки [патент США № 4676980] и для лечения HIV инфекции [WO 91/00360; WO 92/200373; EP 03089]. Предполагают, что антитела можно получить in vitro, используя способы химии синтетических белков, включая способы с использованием сшивающих веществ. Например, иммунотоксины можно сконструировать, используя дисульфидную реакцию замещения или образуя тиоэфирную связь. Примеры подходящих для этой цели реагентов включают в себя иминотиолят и метил-4-меркаптобутиримидат, показанные, например, в патенте США № 4676980.
13. Цитотоксические антитела
Также представлены цитотоксические антитела. В некоторых вариантах осуществления, цитотоксическое антитело представляет собой анти-IL22 антитело, такое как представленное ниже антитело, которое действует на эффекторную функцию и/или вызывает гибель клеток. В некоторых вариантах осуществления, цитотоксическое анти-IL-22R антитело связывается с внеклеточным доменом IL-22R.
14. Конструирование эффекторной функции
Может быть желательно модифицировать антитело с соответствующей эффекторной функцией с тем, чтобы увеличить, например, эффективность антитела при лечении болезни, такой как рак. Например, цистеиновые остатки можно вводить в Fc область, таким образом, позволяя образовывать дисульфидные связи внутри цепи в этой области. Полученные таким образом гомодимерные антитела могут обладать улучшенной способностью к интернализации и/или увеличенной комплемент-опосредованной клеточной гибелью и антитело-зависимой клеточной цитотоксичностью (ADCC). См. Caron et al., J. Exp Med., 176: 1191-1195 (1992) и Shopes J. Immunol., 148: 2918-2922 (1992). Гомодимерные антитела с увеличенной анти-опухолевой активностью также можно получить, используя гетеробифункциональные перекрестносшивающие вещества, как описано в Wolff et al., Cancer Research, 53: 2560-2565 (1993). Альтернативно, антитела можно сконструировать, чтобы они имели двойные Fc области и, таким образом, могли иметь увеличенные способности лизиса комплемента и ADCC. См. Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989).
15. Векторы, клетки-хозяева и рекомбинантные способы
Для рекомбинантного получения антитела, в одном варианте осуществления, выделяют кодирующие их нуклеиновые кислоты и вставляют в реплицируемый вектор для дальнейшего клонирования (амплификация ДНК) или для экспрессии. Кодирующую антитело ДНК легко выделяют и секвенируют с использованием обычных способов (например, используя олигонуклеотидные пробы, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Существует много векторов. Выбор вектора частично зависит от используемой клетки-хозяина. Как правило, клетки-хозяева представляют собой клетки либо прокариотического, либо эукариотического (как правило, млекопитающих) источника. Будет оценено, что константные области любого изотипа можно использовать для этой цели, включая IgG, IgM, IgA, IgD и IgE константные области и что такие константные области можно получить от любого человека или видов животных.
a) Получение антител с использованием прокариотических клеток-хозяев
(1) Векторная конструкция
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела, можно получить с использованием стандартных рекомбинантных способов. Нужные полинуклеотидные последовательности можно выделить и секвенировать из продуцирующих антитела клеток, таких как клетки гибридомы. Альтернативно, можно синтезировать полинуклеотиды, используя нуклеотидные синтезаторы или способы ПЦР. Однажды полученные, кодирующие полипептиды последовательности вставляют в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерогенные полинуклеотиды в прокариотических хозяевах. Многие векторы, которые существуют и известны в данной области, можно использовать для цели настоящего изобретения. Выбор подходящего вектора будет зависеть, главным образом, от размера нуклеиновых кислот, вставленных в вектор, и от конкретной клетки-хозяина, трансформированной вектором. Каждый вектор содержит различные компоненты, в зависимости от их функции (амплификация или экспрессия гетерогенного полинуклеотида, или и то и другое) и их совместимость с конкретной клеткой-хозяином, в которой он находится. Как правило, компоненты вектора включают в себя, но ими не ограничиваются, точку начала репликации, маркерный ген селекции, промотор, рибосомный связывающий сайт (RBS), сигнальную последовательность, гетерогенную вставка нуклеиновых кислот и последовательность окончания транскрипции.
Как правило, плазмидные векторы, содержащие репликон и контрольные последовательности, которые получены из видов соответствующей клетки-хозяина, можно использовать применительно к этим хозяевам. Обычно вектор несет сайт репликации, а также и маркирующие последовательности, которые способны обеспечить селекцию фенотипа в трансформированных клетках. Например, E. coli обычно трансформируют с использованием pBR322, плазмиды, полученной из видов E. coli. pBR322 содержит гены, кодирующие резистентность к ампициллину (Amp) и тетрациклину (Tet) и таким образом обеспечивает легкие способы идентификации трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаги также могут содержать или быть модифицированы, чтобы содержали, промоторы, которые можно использовать микробным организмом для экспрессии эндогенных белков. Примеры производных pBR322, использованных для экспрессии конкретных антител, описаны подробно в Carter et al., патенте США № 5648237.
Кроме того, фаговые векторы, содержащие репликон и контрольные последовательности, которые совместимы с хозяйским микроорганизмом, можно использовать в качестве трансформирующих векторов применительно к этим хозяевам. Например, бактериофаг, такой как λGEM.TM.-11 можно использовать для получения рекомбинантного вектора, который можно использовать для транформации компетентных клеток-хозяев, таких как E. coli LE392.
Вектор экспрессии изобретения может содержать две или более промотор-цистронных пары, кодирующих каждый из полипептидных компонентов. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5') к цистрону, который изменяет ее экспрессию. Обычно прокариотические промоторы разделяют на два класса, индуцибельные и конститутивные. Индуцибельный промотор представляет собой промотор, который вызывает увеличение уровня транскрипции цистрона под его контролем в ответ на изменения условий культивирования, например присутствие или отсутствие питательного вещества или изменения температуры.
Хорошо известно большое количество промоторов, выявленных в различных потенциальных клетках хозяевах. Выбранный промотор может быть функционально связан с ДНК цистрона, кодирующей легкую или тяжелую цепь с помощью перемещения промотора из источника ДНК посредством расщепления рестриктазами и вставки выделенной последовательности промотора в вектор из изобретения. Как нативная последовательность промотора, так и многие гетерогенные промоторы можно использовать для прямой амплификации и/или экспрессии генов мишеней. В некоторых вариантах осуществления, гетерогенные промоторы используют, так как они, как правило, позволяют более высокую транскрипцию и более высокий выход экспрессируемого гена мишени по сравнению с нативным полипептидным промотором мишени.
Подходящие для использования в прокариотических хозяевах промоторы включают в себя PhoA промотор, промоторные системы β-галактомазы и лактозы, промоторную систему триптофана (trp) и гибридные промоторы, такие как tac или trc промотор. Однако другие промоторы, которые функционируют у бактерий (такие как другие известные бактериальные или фаговые промоторы), также подходят. Их нуклеотидные последовательности были опубликованы, таким образом давая возможность квалифицированному специалисту оперативно связывать их с цистронами, кодирующими легкие и тяжелые цепи мишени (Siebenlist et al. (1980) Cell 20:269), используя линкеры или адапторы для получения любых нужных сайтов рестрикции.
В одном аспекте изобретения, каждый цистрон в рекомбинантном векторе содержит секретируемый компонент сигнальной последовательности, который определяет перемещение экспрессируемых полипептидов через мембрану. Как правило, сигнальная последовательность может являться компонентом вектора или может быть частью полипептидной ДНК мишени, которая вставлена в вектор. Сигнальная последовательность, выбранная для цели этого изобретения, должна быть выбранной и процессированной (то есть расщепленная сигнальной пептидазой) клетками хозяевами. Для прокариотических клеток-хозяев, для которых не выбрана и не процессируется сигнальная последовательность, нативная к гетерогенным полипептидам, сигнальная последовательность замещается прокариотической сигнальной последовательностью, выбранной, например, из группы состоящей из щелочной фосфотазы, пенициллиназы, Ipp или лидерных последовательностей термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP. В одном варианте осуществления изобретения, сигнальные последовательности, использованные в обоих цистронах экспрессирующей системы, представляют собой STII сигнальные последовательности или их варианты.
В другом аспекте, продукция иммуноглобулинов согласно изобретению может наблюдаться в цитоплазме клетки хозяев и поэтому нежелательно присутствие секретируемых сигнальных последовательностей в каждом цистроне. При этом легкие и тяжелые цепи иммуноглобулинов экспрессируются, сворачиваются и собираются для образования функционирующих иммуноглобулинов в цитоплазме. Некоторые штаммы-хозяева (например, E. coli trxB-штаммы) обеспечивают состояние цитоплазмы, которое способствует образованию дисульфидных связей, таким образом, обеспечивает правильное сворачивание и сборку экспрессируемых белковых субъединиц. Proba and Pluckthun Gene, 159:203 (1995).
Антитела изобретения также можно получить путем использования экспрессионной системы, в которой количественное соотношение экспрессируемых полипептидных компонентов можно изменять, чтобы максимизировать выход секретируемых и правильно собранных антител изобретения. Такие изменения выполняются, по крайней мере, частично одновременным изменением эффективности трансляции полипептидных компонентов.
Один способ изменения трансляционной эффективности описан в Simmons et al., патент США № 5840523. В нем используют варианты области инициации трансляции (TIR) в цистроне. Для данной TIR, ряд вариантов аминокислотных или нуклеиновых кислотных последовательностей можно получать с различными трансляционными эффективностями, поэтому обеспечивая подходящие значения, которыми регулируют этот фактор для желательного уровня экспрессии специфической цепи. Варианты TIR можно получать обычным способом мутагенеза, который приводит к изменениям в кодонах, которые могут изменять аминокислотную последовательность. В некоторых вариантах осуществления, изменения в нуклеотидной последовательности молчащие. Изменения в TIR могут включать в себя, например, изменения в количестве или расстоянии между последовательностями Шайн-Дальгарно наряду с изменениями сигнальной последовательности. Один способ получения мутантных сигнальных последовательностей представлен образованием "банка кодонов" в начале кодирующей последовательности, которая не изменяет аминокислотную последовательность сигнальной последовательности (то есть изменения молчащие). Это можно выполнить, изменяя третье нуклеотидное положение каждого кодона; дополнительно, несколько аминокислот, такие как лейцин, серин, и аргинин, имеют множество первых и вторых положений, которые могут добавлять сложность в формировании банка. Этот способ мутагенеза описан подробно в Yansura et al. (1992) METHODS: A Companion to Methods in Enzymol. 4: 151-158.
В одном варианте осуществления, получают множество векторов с разными эффективностями TIR для каждого цистрона. Это ограниченное множество обеспечивает сравнение уровня экспрессии каждой цепи, а также выход продуктов нужных антител при различных сочетаниях эффективности TIR. Эффективности TIR можно определить, количественно измеряя уровень экспрессии репортерного гена, как описано подробно в Simmons et al. патенте США № 5840523. Основываясь на сравнении трансляционной эффективности, выбирают нужные отдельные TIR, чтобы сочетать экспрессирующие векторные конструкты изобретения.
Прокариотические клетки-хозяева, подходящие для экспрессии антител изобретения, включают в себя Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры нужных бактерий включают в себя Escherichia (например, E. coli), Bacilli (например, B. subtilis), Enterobacteria, Pseudomonas виды (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla, или Paracoccus. В одном варианте осуществления, используют грамотрицательные клетки. В одном варианте осуществления, используют клетки E. coli в качестве хозяев для изобретения. Примеры штаммов E. coli включают в себя штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219; ATCC Deposit № 27325) и производные его, включая штамм 33D3, имеющий генотип W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTΔ(nmpc-fepE) degP41 kanR (патент США № 5639635). Другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coliλ 1776 (ATCC 31537) и E. coli RV308(ATCC 31608) также подходят. Эти примеры иллюстрируют, а не ограничивают. Способы конструирования производных любых вышеупомянутых бактерий, имеющих определенные генотипы, известны в данной области и описаны, например, в Bass et al., Proteins, 8:309-314 (1990). Как правило, необходимо выбрать подходящую бактерию, имеющую оценку реплицируемости репликонов в клетках бактерий. Например, виды E. coli, Serratia, или Salmonella можно подходящим образом использовать как хозяев, когда хорошо известны плазмиды, такие как pBR322, pBR325, pACYC177, или pKN410 используются для получения репликона. Обычно клетки хозяев будут секретировать минимальные количества протеолитических ферментов, и дополнительные ингибиторы протеаз можно по необходимости вводить в клеточную культуру.
(2) Получение антител
Клетки-хозяева трансформируют вышеописанными экспрессирующими векторами и культивируют в обычной питательной среде, измененной соответствующим образом для индукции промоторов, отбора трансформантов или амплифицирования генов, кодирующих нужные последовательности.
Трансформация означает введение ДНК в прокариотического хозяина, так что ДНК реплицируется либо как нехромосомный элемент, либо путем хромосомной интеграции. В зависимости от использованной клетки хозяина, трансформацию проводят, используя стандартные способы, соответствующие таким клеткам. Обработку кальцием, используя хлорид кальция, как правило используют для бактериальных клеток, которые содержат устойчивые барьеры клеточной стенки. В другом способе трансформации используют полиэтиленгликоль/ДМСО. В еще одном способе используют электропарацию.
Прокариотические клетки, использованные для получения полипептидов изобретения, выращивают в среде известной в данной области и подходящей для культуры выбранных клеток-хозяев. Примеры подходящей среды включают в себя luria broth (LB) плюс необходимые питательные добавки. В некоторых вариантах осуществления, среда также содержит селектирующее вещество, выбранное на основании конструкции экспрессирующего вектора, чтобы обеспечить селективный рост прокариотических клеток, содержащих экспрессирующий вектор. Например, добавляют ампицилин в среду для роста клеток, экспрессирующих ген устойчивости к ампицилину.
Любые необходимые добавки, кроме источников углерода, азота и неорганического фосфата, также могут быть включены в соответствующих концентрациях, вводимые по отдельности или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Дополнительно культуральная среда может содержать одно или более восстанавливающие вещества, выбранные из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при подходящих температурах. В некоторых вариантах осуществления, для роста E. coli, температуры роста варьируют от примерно 20°C до примерно 39°C; от примерно 25°C до примерно 37°C; или примерно 30°C. pH среды может быть любым pH, варьирующим от примерно 5 до примерно 9, в зависимости в основном от организма хозяина. В некоторых вариантах осуществления, для E. coli, pH составляет от примерно от 6,8 до примерно 7,4, или примерно 7,0.
Если в экспрессируемом векторе изобретения используют индуцибельный промотор, то экспрессия белков происходит в условиях, подходящих для активации промотора. В одном аспекте изобретения, PhoA промоторы используют для контроля транскрипции полипептидов. Следовательно, трансформированные клетки-хозяева культивируют в фосфатно-лимитирующей среде для индукции. В некоторых вариантах осуществления, фосфатно-лимитирующая среда представляет собой C.R.A.P. среду (см., например, Simmons et al., J. Immunol. Methods (2002), 263: 133-147). Множество других индукторов можно использовать в соответствии с используемым векторным конструктом, как известно в данной области.
В одном варианте осуществления, экспрессированные полипептиды настоящего изобретения секретируются и выходят из периплазмы клеток-хозяев. Выделение белка обычно включает в себя разрушение микроорганизма, как правило, такими способами как осмотический шок, разрушение ультразвуком или лизис. Как только клетки разрушают, клеточный детрит или целые клетки удаляли центрифугированием или фильтрацией. В дальнейшем белки можно очистить, например, путем хроматографии на аффинной смоле. Альтернативно, белки могут транспортироваться в клеточную среду и выделяться там. Клетки можно удалять из культуры и культурального супернатанта, фильтрованных и концентрированных, для дальнейшей очистки полученных белков. Экспрессированные полипептиды далее можно выделить и идентифицировать, используя общие известные способы, такие как электрофорез в полиакриламидном геле (PAGE) и вестерн-блот анализ.
В одном аспекте изобретения, проводится получение антител в большом количестве посредством ферментации. Различные массовые ферментационные способы с периодической подпиткой доступны для получения рекомбинантных белков. Массовые ферментации содержат, по крайней мере, 1000 литров объема, а в некоторых вариантах осуществления, примерно от 1000 до 100000 литров объема. В этих ферментерах используют лопастные мешалки, чтобы распределять кислород и питательные вещества, особенно глюкозу (предпочтительно углеродно-энергетический источник). Ферментации небольшого количества обычно относят к ферментации в ферментере, который не содержит более чем приблизительно 100 литров объемной вместимости и может варьировать от примерно 1 литра до примерно 100 литров.
В способе ферментации, индукция экспрессии белка обычно вызывается после того, как клетки были выращены в подходящих условиях до нужной плотности, например OD550 примерно 180-220, в которой клетки находятся в ранней стационарной фазе. Можно использовать ряд индукторов в соответствии с использованным векторным конструктом, как известно в данной области и описано выше. Клетки можно выращивать в течение более коротких периодов до индукции. Обычно клетки индуцируются примерно за 12-50 часов, хотя можно использовать более продолжительное или более короткое время индукции.
Чтобы улучшить выход продукции и качество полипептидов изобретения, можно изменять различные ферментативные условия. Например, чтобы улучшить правильную сборку и сворачивание секретируемых антителами полипептидов, добавленные векторы, гиперэкспрессирующие белки-шапероны, такие как Dsb белки (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил цис,транс-изомераза с шаперонной активностью), можно использовать для ко-трансформации прокариотических клеток-хозяев. Было показано, что белки-шапероны способствуют правильному сворачиванию и растворимости гетерогенных белков, полученных из бактериальных клеток-хозяев. Chen et al. (1999) J. Biol. Chem. 274: 19601-19605; Georgiou et al., патент США № 6083715; Georgiou et al., патент США № 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275: 17100-17105; Ramm and Pluckthun (2000) J. Biol. Chem. 275: 17106-17113; Arie et al. (2001) Mol. Microbiol. 39:199-210.
Чтобы минимизировать протеолиз экспрессированных гетерогенных белков (особенно тех, которые являются протеолитически чувствительными), в настоящем изобретении можно использовать некоторые штаммы хозяев, дефицитных протеолитическими ферментами. Например, штаммы клеток-хозяев можно изменить, чтобы вызвать генетические мутации в генах, кодирующих известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их сочетание. Некоторые протеазодефицитные штаммы E. coli доступны и описаны в, например, Joly et al. (1998), выше; Georgiou et al., патент США № 5264365; Georgiou et al., патент США № 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном варианте осуществления, штаммы E. coli, дефицитные протеолитическими ферментами и трансформированные плазмидами, гиперэкспрессирующие один или более белков-шаперонов, используют в качестве клеток-хозяев в экспрессирующих системах изобретения.
(3) Очистка антител
В одном варианте осуществления, полученные в данном документе антитела далее очищают для получения препаратов, которые являются в значительной степени гомогенными для дальнейших способов и применений. Можно использовать стандартные способы очистки белков, известные в данной области. Следующие способы представляют собой типичные подходящие способы очистки: фракционирование на иммуно-аффинных или ионно-обменных колонках, преципитация этанолом, обращенно-фазовая HPLC, хроматография на силикагеле или на катионно-обменной смоле, такая как DEAE, хроматофокусирование, SDS-PAGE, фракционирование сульфатом аммония и гель-фильтрация с использованием, например, сефадекса G-75.
В одном аспекте, белок A, иммобилизированный на твердой фазе, используют для иммунно-аффинной очистки продуктов антител изобретения. Белок A представляет собой белок клеточной стенки 41 кД из Staphylococcus aureas, который связывается с высокой аффинностью к Fc области антител. Lindmark et al (1983) J. Immunol. Meth. 62: 1-13. Твердая фаза, к которой иммобилизован белок A, может представлять собой колонку, включающую поверхность стеклянную или кремнеземную, или стеклянную колонку с контролируемым размером пор, или колонку с кремниевой кислотой. В некоторых заявках, колонки покрыты реагентом, таким как глицерин, для возможного предотвращения неспецифического прилипания примесей.
В качестве первого этапа очистки, полученный из клеточной культуры препарат, как описано выше, можно использовать по отношению к белку A, иммобилизованному на твердой фазе, чтобы обеспечить специфическое связывание интересующего антитела к белку A. Затем отмывают твердую фазу, чтобы удалить примеси, неспецифически связанные с твердой фазой. Наконец, интересующее антитело удаляют из твердой фазы элюцией.
b) Получение антител с использованием эукариотических клеток-хозяев
Обычно вектор для использования в эукариотических клетках-хозяевах включает в себя один или более следующих неограничивающих компонентов: сигнальную последовательность, точку начала репликации, один или более маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
(1) Компонент сигнальной последовательности
Вектор для использования в эукариотической клетке-хозяине также может содержать сигнальную последовательность или другой полипептид, имеющий специфический сайт расщепления на N-конце зрелого белка или интересующего полипептида. Выбранная гетерогенная сигнальная последовательность может быть узнанной и процессированной (то есть расщепленная сигнальной пептидазой) клеткой-хозяином. Экспрессия в клетках млекопитающих, сигнальные последовательности млекопитающих, а также и вирусные секреторные лидерные последовательности, например сигнал простого герпеса gD, являются доступными. ДНК для такой области предшественника замыкается в рамке считывания ДНК, кодирующей антитело.
(2) Точка начала репликации
Как правило, компонент точки начала репликации не является необходимым для экспрессирующих векторов млекопитающих. Например, обычно можно использовать только точку начала SV40, так как она содержит начальный промотор.
(3) Селекционный генный компонент
Экспрессирующие и клонирующие векторы могут содержать селекционный ген, также обозначенный селектируемый маркер. Обычные селекционные гены кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампицилину, неомицину, метотрексату или тетрациклину, (b) ауксотрофный дефицит комплемента, где необходимо, или (c) обеспечение необходимыми питательными веществами, недостаточными из сложной среды.
В одном примере схемы селекции используется препарат, останавливающий рост клетки-хозяина. Такие клетки, которые успешно трансформируются гетерогенным геном, продуцируют белок, придающий лекарственную устойчивость и таким образом сохраняющий схему селекции. В примерах такой доминантной селекции используют препараты неомицин, микофиноликовую кислоту и гидромицин.
Другой пример подходящих селектируемых маркеров для клеток млекопитающих представляет собой пример, который позволяет идентифицировать компетентные клетки, чтобы выбрать нуклеиновую кислоту антитела, такую как DHFR, тимидинкиназу, металлотионеин-I и -II, предпочтительно гены металлотионеинов приматов, аденозиндезаминазу, орнитиндекарбоксилазу и другие.
Например, в некоторых вариантах осуществления, клетки, трансформированные DHFR селектирующим геном, были впервые идентифицированы посредством культивирования всех трансформантов в культуральной среде, содержащей метотрексат (Mtx), антагонист конкурентный DHFR. В некоторых вариантах осуществления, подходящие клетки-хозяева при использовании DHFR дикого типа представляют собой клеточную линию яичника китайского хомячка (CHO), дефицитную по DHFR активности (например, ATCC CRL-9096).
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, которые содержат индогенный DHFR), трансформированные или ко-трансформированные ДНК последовательностями, кодирующими антитело, белок дикого типа DHFR и другой селектируемый маркер, такой как аминогликозид 3'-фосфотрансфераза (APH), можно выбрать посредством роста клеток в среде, содержащей селектирующее вещество для селектируемого маркера, такое как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США № 4965199.
(4) Промоторный компонент
Экспрессирующие и клонирующие векторы обычно содержат промотор, который узнается организмом хозяина и функционально связан с нуклеиновой кислотой, кодирующей интересующий полипептид (например, антитело). Известны промоторные последовательности для эукариот. Например, фактически все эукариотические гены имеют AT-богатую область, локализованную приблизительно на 25-30 оснований выше участка инициации транскрипции. Другая последовательность, обнаруженная на 70-80 оснований выше от начала транскрипции многих генов, представляет собой CNCAAT область, где N может быть любым нуклеотидом. На 3' конце большинства эукариотических генов есть AATAAA последовательность, которая может быть сигнальной для добавления поли A хвоста к 3' концу кодирующей последовательности. В некоторых вариантах осуществления, любые или все эти последовательности можно подходящим образом вставить в эукариотические экспрессирующие векторы.
Транскрипция с векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами полученными из геномов вирусов, таких как вирус полиомы, fowlpox вирус, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и вирус обезьяны 40 (SV40), из геторогенных промоторов млекопитающих, например промотор актина или промотор иммуноглобулина, из промоторов белков теплового шока предоставлены такие промоторы, которые находятся в соответствии с системами клеток-хозяев. Начальные и конечные промоторы SV40 вируса удобно получать в виде рестрикционного фрагмента SV40, который также содержит SV40 вирусную точку начала репликации. Самый первый промотор человеческого цитомегаловируса удобно получен в виде HindIII E рестрикционного фрагмента. Система для экспрессии ДНК в хозяевах млекопитающих с использованием вируса папилломы крупного рогатого скота представлена в качестве вектора, описанного в Патенте США № 4419446. Модификация этой системы описана в Патенте США № 4601978. См. также Reyes et al, Nature 297:598-601 (1982), описывающий экспрессию кДНК β-интерферона человека в клетках мышей под контролем тимидинкиназного промотора из вируса простого герпеса. Альтернативно, длинный терминальный повтор вируса саркомы Рауса можно использовать в качестве промотора.
(5) Компонент энхансерного элемента
Транскрипцию ДНК, кодирующую антитело этого изобретения, высшими эукариотами часто усиливают посредством вставки энхансерной последовательности в вектор. В настоящее время известны многие энхансерные последовательности из генов млекопитающих (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Обычно, однако, будет использоваться энхансер из вируса эукариотических клеток. Примеры включают в себя SV40 энхансер на последнем участке точки начала репликации (bp 100-270), начального промоторного энхансера цитомегаловируса, энхансер полиомы на последнем участке точки начала репликации и энхансеры аденовирусов. См. также Yaniv, Nature 297:17-18 (1982), описывающий энхансерные элементы активации эукариотических промоторов. Энхансер может вставляться в вектор в положении 5' или 3' к последовательности, кодирующей полипептид антитела, но, как правило, локализован в области 5' промотора.
(6) Компонент терминации транскрипции
Использованные в эукариотических клетках-хозяевах экспрессирующие векторы также могут содержать последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Обычно такие последовательности доступны с 5' и изредка 3' нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслируемом участке мРНК, кодирующей антитело. Один удобный компонент терминации транскрипции представляет собой полиаденилированную область гормона роста крупного рогатого скота. См. WO94/11026 и описанный там экспрессирующий вектор.
(7) Селекция и трансформация клеток-хозяев
Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах в данном документе включают в себя клетки высших эукариот, описанные в данном документе, включая клетки-хозяева позвоночных. Воспроизводство клеток позвоночных в культуре (культура тканей) стало обычной процедурой. Примеры полезных линий клеток-хозяев млекопитающих представляют собой линию CV1 почек обезьян, трансформированную SV40 (COS-7, ATCC CRL 1651); линию почек эмбриона человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре, Graham et al, J. Gen Virol. 36:59 (1977)); клетки почек карликовых хомячков (BHK, ATCC CCL 10); клетки яичника китайского хомячка-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки сертоли мышей (TM4, Mather, Biol. Reprod 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почек африканских зеленых обезьян (VERO-76, ATCC CRL-1587); клетки карциномы шеи человека (HELA, ATCC CCL 2); клетки почек собак (MDCK, ATCC CCL 34); клетки печени буфало крыс (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N. Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; FS4 клетки; и линия гепатомы человека(Hep G2).
Клетки-хозяева трансформируют вышеописанными экспрессирующими или клонирующими векторами для получения антител и культивирования в подходящей питательной среде, измененной как нужно для активации промоторов, селекции трансформантов или амплификации генов, кодирующих нужные последовательности.
(8) Культивирование клеток-хозяев
Используемые клетки-хозяева для получения антител этого изобретения можно культивировать в различных средах. Коммерчески доступная среда, такая как Ham's F10 (Sigma), Minimal Essential Medium ((MEM), (Sigma), RPMI-1640 (Sigma), и Дульбекко модифицированная среда Игла ((DMEM), Sigma) подходят для культивирования клеток-хозяев. Кроме того, можно использовать в качестве культуральной среды для клеток-хозяев любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), патенты США № 4767704; 4657866; 4927762; 4560655; или 5122469; WO 90/03430; WO87/00195; или Патент США Re. 30985. Любую из этих сред можно дополнить необходимыми гормонами и/или другими ростовыми факторами (такими как инсулин, трансферрин или эпидермальными факторами роста), солями (такими как хлорид и фосфат натрия, кальция, магния), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимедин), антибиотиками (такими как GENTAMYCINTM препарат), микроэлементами (определяемыми как неорганические соединения, присутствующие обычно в конечных концентрациях на микромолярном уровне) и глюкозой или эквивалентным источником энергии. Любые другие дополнения также могут быть включены в подходящих концентрациях, которые должны быть известны специалистам в данной области. Условия культивирования, такие как температура, pH и другие, представляют собой условия, ранее использованные с выбранными для экспрессии клеток-хозяев, и будут очевидны обычному квалифицированному специалисту.
(9) Очистка антител
При использовании рекомбинантных способов, антитела можно получать внутриклеточно или непосредственно секретированные в среду. Если антитело получают внутриклеточно, как первый этап, дисперсный детрит, либо клетки-хозяева, либо лизированные фрагменты, можно удалить, например, посредством центрифугирования или ультрацентрифугирования. Если антитела секретируются в среду, супернатант из таких экспрессирующих систем можно исходно концентрировать, используя коммерчески доступный фильтр для концентрирования белка, например Amicon и Millipore Pellicon ultrafiltration unit. Ингибитор протеаз, такой как PMSF, может включать в себя любые вышеупомянутые этапы для подавления протеолиза, и антибиотики могут включать в себя этапы предотвращения роста случайных примесей.
Полученные из клеток составы антител можно очистить, используя, например, гидроксиапатитовую хроматографию, гель-электрофорез, диализ и аффинную хроматографию, с аффинной хроматографией, что является удобным способом. Подходящий белок A в качестве аффинного лиганда зависит от видов и изотипа любого Fc домена иммуноглобулина, который представлен в антителе. Белок A может использоваться для очистки антител, в основе которых находятся человеческие тяжелые цепи γ1, γ2, или γ4 (Lindmark et al., J. Immunol. Methods 62:1-13 (1983)). Белок G рекомендуется для всех изотипов мышей и для человеческого γ3 (Guss et al., EMBO J. 5:1567-1575 (1986)). Матриксом, к которому прикрепляется аффинный лиганд, может быть агароза, но допустимы другие матриксы. Механически прочные матриксы, такие как стекло с контролируемым размером пор или поли(стириндивинил)бензол, обеспечивают более быструю скорость потока и более короткое время процесса, чем это обеспечит агароза. Если антитело содержит CH3 домен, Bakerbond ABXTMresin (J. T. Baker, Phillipsburg, NJ) удобна для очистки. Другие способы очистки белка, такие как фракционирование на ионно-обменной колонке, преципитация этанолом, обращенно-фазовая ВЭЖХ, хроматография на силикагеле, хроматография на гепарине, SEPHAROSETM, хроматография на анионной или катионной обменной смоле (такие как колонка с полиаспартиковой кислотой), хроматофокусирование, SDS-PAGE, и преципитация аммонием сульфатом также доступны в зависимости от выделяемого антитела.
После любых предварительных этапов очистки смесь, содержащая интересующее антитело и примеси, можно подвергнуть дальнейшей очистке, например, посредством хроматографии гидрофобного взаимодействия при низком pH, используя элюирующий буфер при pH в промежутке примерно 2,5-4,5, предпочтительно произведенный при низких концентрациях солей (например, от примерно 0-0,25 М соли).
Как правило, различные способы получения антител для использования в исследовании, тестирования и клинического использования точно установлены в данной области, в соответствии с вышеописанными способами и/или, как предполагается целесообразным специалистами в данной области, для отдельного интересующего антитела.
C. Иммуноконъюгаты
Иммуноконъюгаты, или "нагруженное лекарственным средством антитело", полезны для доставки на место цитотоксических веществ при лечении рака. См., например, Syrigos et al. (1999) Anticancer Research 19:605-614; Niculescu-Duvaz et al. (1997) Adv. Drug Deliv. Rev. 26:151-172; Патент США № 4975278. Иммуноконъюгаты обеспечивают направленную доставку фрагмента лекарств к опухоли, поскольку системное введение неконъюгированных цитотоксических веществ может приводить к недопустимому уровню цитотоксичности в нормальных клетках, а также опухолевые клетки стремятся к подавлению. См. Baldwin et al. (Mar. 15, 1986) Lancet pp. 603-05; Thorpe (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review," в Monoclonal Antibodies '84: Biological and Clinical Applications (A. Pinchera et al., eds.) pp. 475-506.
В одном аспекте, иммуноконъюгат содержит антитело, которое связывает IL-19, IL-20, IL-22, IL-24, IL22R, IL-20Ra, IL-20Rb, или IL-10R2, такие как антитела, представленные в данном документе, и цитотоксическое вещество, такое как химиотерапевтическое вещество, вещество, подавляющее рост, токсин (например, ферментативно активный токсин из бактериального, грибного, растительного или животного источника, или их фрагменты), или радиоактивный изотоп (то есть радиоконъюгат).
Химиотерапевтические вещества, полезные для получения таких иммуноконъюгатов, были описаны выше. Ферментативно активные токсины и их фрагменты, которые можно использовать, включают в себя цепь А дифтерии, несвязавшиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модецина, альфа-сарцин, Aleurites fordii белки, белки диантина, Phytolaca americana белки (PAPI, PAPII, и PAP-S), momordica charantia ингибитор, цурцин, кротин, sapaonaria officinalis ингибитор, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Ряд радионуклеидов доступен для получения радиоконъюгированных антител. Примеры включают в себя 212Bi, 131I, 131In, 90Y и 186Re.
Конъюгаты антитела и цитотоксического вещества можно получить, используя ряд бифункциональных белок-связывающих веществ, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметил адипимидат HCL), активные эфиры (такие как дисукцинимидил суберат), альдегиды (такие как глютарелдегид), биз-азидные соединения (такие как бис-(п-азидобензоил)гександиамин), бис-диазоний производные (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные фтористые соединения (такие как 1,5-дифтор-2,4-динитробензен). Например, иммунотоксин рицина можно получить, как описано в Vitetta et al, Science, 238: 1098 (1987). Углерод-14-меченная 1-изотиоцианатбензил-3-метилдиэтилен триаминпентауксусная кислота (MX-DTPA) представляет собой типичное хелатообразующее вещество для конъюгации радионуклеотида к антителу. См. WO94/11026.
Конъюгаты антитела и одна или более маленьких молекул токсина, такие как калихеамицина, майтансиноиды, трихотин и CC1065, и производные этих токсинов, которые обладают активностью токсинов, также предложены в данном документе.
1. Майтансин и майтансиноиды
В одном варианте осуществления, иммуноконъюгаты содержат антитело, конъюгированное с одной или более молекулами майтансиноидов. Майтансиноиды представляют собой митотоксические ингибиторы, которые действуют, ингибируя полимеризацию тубулина. Майтансин был впервые выделен из восточного африканского кустарника Maytenus serrata (патент США № 3896111). Впоследствии было обнаружено, что некоторые микробы также вырабатывают майтансиноиды, такие как майтансинол и C-3 майтансинол эфиры (патент США № 4151042). Синтетический майтансинол и производные и их аналоги раскрыты, например, в патентах США № 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533, раскрытие которых прямо включено посредством ссылки.
При попытке улучшения их терапевтического индекса, майтансин и майтансиноиды были конъюгированы с антителами, которые связывают антигены на поверхности опухолевых клеток. Иммуноконъюгаты, содержащие майтансиноиды и их терапевтическое применение, раскрыты, например, в патентах США № 5208020, 5416064 и европейском патенте 0 425 235 B1, раскрытие которых прямо включено посредством ссылки. У Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описаны иммуноконъюгаты, содержащие майтансиноид, обозначенный DM1, связанный с моноклональным антителом C242, направленным против человеческого колоректального рака. Было обнаружено, что конъюгат является высокотоксичным по отношению к культивируемым клеткам колоректального рака, и показана антиопухолевая активность при анализе роста опухоли in vivo. В Chari et al., Cancer Research 52:127-131 (1992) описаны иммуноконъюгаты, в которых майтансиноид был конъюгирован через дисульфидную связь к мышиному антителу A7, связывающему антиген на клеточных линиях человеческого колоректального рака, или к другому мышиному моноклональному антителу TA.1, которое связывает HER-2/neu онкоген. Цитотоксичность TA.1-майтансиноидного конъюгата была испытана in vitro на клеточной линии человеческого рака груди SK-BR-3, которая экспрессирует 3×105 HER-2 поверхностных антигена на клетку. Лекарственный конъюгат достигал уровня токсичности сходного со свободным препаратом майтансиноида, который мог увеличиваться путем увеличения числа молекул майтансиноидов на молекулу антитела. A7-майтансиноидный конъюгат показал низкую системную токсичность у мышей.
Антитело-майтансиноидные конъюгаты получают посредством химического связывания антитела с молекулой майтансиноида без значительного снижения биологической активности либо антитела, либо молекулы майтансиноида. Средняя величина 3-4 конъюгированных молекул майтансиноидов на антитело показывает эффективность увеличения цитотоксичности клеток-мишеней без отрицательно влияющей функции или растворимости антитела, хотя ожидается, что даже одна молекула токсина на антитело увеличит цитотоксичность более, чем при использовании чистого антитела. Майтансиноиды хорошо известны в данной области и могут синтезироваться с использованием известных способов или быть выделены из природных источников. Подходящие майтансиноиды раскрыты, например, в патенте США № 5208020 и в других патентах и непатентных публикациях, отнесенных к данному документу выше. Предпочтительные майтансиноиды представляют собой майтансинол и модифицированные аналоги майтансинола в ароматическом кольце или в других положениях молекулы майтансинола, такие как различные эфиры майтансинола.
Существует много связывающих групп, известных в данной области, для получения антитело-майтансиноид конъюгатов, включая, например, описанные в патенте США № 5208020 или европейском патенте 0 425 235 B1, и Chari et al, Cancer Research 52:127-131 (1992). Связывающие группы включают в себя дисульфидные группы, тиоэфирные группы, кислотолабильные группы, фотолабильные группы, пептидазолабильные группы или эстеразолабильные группы, как раскрыто в вышеупомянутых патентах, предпочтительнее являются дисульфидные и тиоэфирные группы.
Конъюгаты антитела и майтансиноида можно получить, используя ряд бифункциональных белок-связывающих веществ, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметил адипимидат HCL), активные эфиры (такие как дисукцинимидил суберат), альдегиды (такие как глютарелдегид), бис-азидные соединения (такие как бис-(п-азидобензоил)гександиамин), бис-диазоний производные (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные фтористые соединения (такие как 1,5-дифтор-2,4-динитробензен). Некоторые связывающие вещества, включая N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP), обеспечивают дисульфидную связь.
Связующее вещество может быть прикреплено к молекуле майтансиноида в различных положениях, зависящих от типа связи. Например, эфирная связь может быть образована посредством реакции с гидроксильной группой при использовании обычных способов связывания. Реакция может происходить в C-3 положении, имеющем гидроксильную группу, C-14 положении, измененном гидроксиметильной группой, C-15 положении, измененном гидроксильной группой, и C-20 положении, имеющем гидроксильную группу. В предпочтительном варианте осуществления, связь образуется в C-3 положении майтансинола или аналога майтансинола.
2. Ауристатины и доластатины
В некоторых вариантах осуществления, иммуноконъюгат содержит антитело, конъюгированное к доластатину или пептидному аналогу или производному долостатина, например ауристатину (патенты США № 5635483; 5780588). Было показано, что доластатины и ауристатины препятствуют изменению микротрубочек, GTP гидролизу и ядерному и клеточному делению (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают антираковой (патент США № 5663149) и антигрибковой активностью (Pettit et al. (1998) Antimicrob. Agents Chemother. 42:2961-2965). Фрагмент лекарства с доластатином или ауристатином может быть прикреплен к антителу через N (амино) конец или C (карбоксил) конец пептидного лекарственного фрагмента (WO 02/088172).
Типичные варианты осуществления с ауристатином включают в себя N-терминальные связанные монометилауристатиновые лекарственные компоненты DE и DF, раскрытые в "Monomethylvaline Compounds Capable of Conjugation to Ligands," патентная заявка США, публикация № US 2005-0238649 A1, раскрытие которых прямо включено посредством ссылки полностью.
Обычно пептидные лекарственные компоненты можно получить посредством образования пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получить, например, согласно жидкофазному способу синтеза (см. E. Schroder and K. Lubke, "The Peptides", volume 1, pp 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Ауристатин/доластатин лекарственные компоненты можно получить согласно способам из: US 5635483; US 5780588; Pettit et al (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al (1998) Anti-Cancer Drug Design 13:243-277; Pettit, G.R. et al. Synthesis, 1996, 719-725; и Pettit et al (1996) J. Chem. Soc. Perkin Trans. 15:859-863. См. также Doronina (2003) Nat. Biotechnol. 21(7):778-784; патентная заявка США, публикация № 2005-0238649 A1, раскрытие которых включено посредством ссылки полностью (раскрывая, например, связывающие вещества и способы получения соединений монометилвалина, таких как MMAE и MMAF, конъюгированных со связывающими веществами).
3. Калихеамицин
Другой интересующий иммуноконъюгат содержит антитело, конъюгированное к одной или более молекулам калихеамицина. Семейство антибиотиков калихеамицина способно образовывать двунитевые разрывы ДНК в субпикомолярных концентрациях. Для получения конъюгатов семейства калихеамицина, см. патенты США 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все от компании American Cyanamid). Структурные аналоги калихеамицина, которые можно использовать, включают в себя, но ими не ограничиваются, γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θ1 I (Hinman et al., Cancer Research 53:3336-3342 (1993), Lode et al., Cancer Research 58:2925-2928 (1998) и вышеупомянутые патенты США от American Cyanamid). Другие антиопухолевые препараты, с которым антитело может образовывать конъюгаты, представляют собой QFA, являющееся антифолатом. И калихеамицин, и QFA обладают внутриклеточными участками действия и с трудом проникают через плазматическую мембрану. Поэтому клеточное накопление этих веществ через антитело-опосредованную интернализацию в значительной степени усиливает их цитотоксические эффекты.
4. Другие цитотоксические вещества
Другие антиопухолевые вещества, которые могут конъюгировать с антителом, включают в себя BCNU, стрептозоицин, винкристин и 5-фторурацил, семейство веществ, известных в собирательном значении как LL-E33288 комплекс, описанный в патентах США 5053394, 5770710, а также и эсперамицины (патент США 5877296).
Ферментативно активные токсины и их фрагменты, которые могут использоваться, включают в себя цепь А дифтерии, несвязавшиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модецина, альфа-сарцин, Aleurites fordii белки, белки диантина, Phytolaca americana белки (PAPI, PAPII, и PAP-S), momordica charantia ингибитор, цурцин, кротин, sapaonaria officinalis ингибитор, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, WO 93/21232, опубликованный 28 октября 1993 г.
В другом аспекте, иммуноконъюгаты могут содержать антитело и соединение с нуклеолитической активностью (например, рибонуклеазу или ДНК эндонуклеазу, такую как дезоксирибонуклеазу; ДНКазу).
Для селективного разрушения опухоли, иммуноконъюгат может содержать анти-FGFR2 антитело и высокорадиоактивный атом. Ряд радиоактивных изотопов доступен для получения радиоконъюгированных анти-FGFR2 антител. Примеры включают в себя At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если конъюгат используют для диагностики, то он может содержать радиоактивный атом для сцинтиграфических исследований, например Tc99m, или I123, или спиновую метку для ядерной магнитной резонансной (NMR) визуализации (также известной как магнитно-резонансная визуализация, mri), такую как снова йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радио- или другие метки можно встраивать в конъюгат известными способами. Например, можно биосинтезировать пептид или можно синтезировать его химическим аминокислотным синтезом с использованием подходящих аминокислотных предшественников, включая, например, фтор-19 на место водорода. Такие метки, как Tc99m или I123, Re186, Re188 и In111 можно прикреплять через остаток цистеина в пептиде. Иттрий-90 можно прикреплять через остаток лизина. IODOGEN способ (Fraker et al (1978) Biochem. Biophys. Res. Commun. 80: 49-57 можно использовать для встраивания йода-123. В "Monoclonal Antibodies in Immunoscintigraphy" (Chatal, CRC Press 1989) описаны другие способы подробно.
D. Антагонисты и агонисты
Представлены антагонисты IL-22. Такие антагонисты включают в себя антагонисты, которые прямо действуют на IL-22 (например, анти-IL-22 антитело) и антагонисты, которые опосредованно действуют на IL-22 активность (например, анти-IL-22R антитело). Такие антагонисты нужны для, например, 1) лечения воспалительных нарушений и аутоиммунных нарушений, и 2) изменения передачи сигнала IL-23 или IL-22. В одном конкретном варианте осуществления, состав, содержащий антагонист IL-22 или IL-22R, нужен для уменьшения количества псориазной ткани у млекопитающего. В другом конкретном варианте осуществления, состав, содержащий антагонист IL-22 или IL-22R, нужен для частичного или полного подавления пролиферации опухолевых клеток.
В одном аспекте, антагонист IL-22 представляет собой анти-IL-22 антитело или анти-IL-22R антитело. В некоторых вариантах осуществления, анти-IL-22 антитело представляет собой блокирующее антитело, которое частично или полностью блокирует взаимодействие IL-22 с его рецептором. В некоторых вариантах осуществления, анти-IL-22R антитело представляет собой блокирующее антитело, которое частично или полностью блокирует взаимодействие IL-22R с IL-22. В некоторых вариантах осуществления, анти-IL-22R антитело связывается с внеклеточным лигандсвязывающим доменом IL-22R. Например, анти-IL-22R антитело может связываться с внеклеточным лигандсвязывающим доменом человеческого IL-22R, который обнаружен в SEQ ID NO: 3 из примерно 18-228 аминоксилот.
В другом аспекте, антагонист IL-22 представляет собой олигопептид, который связывается с IL-22 или IL-22R. В одном варианте осуществления, олигопептид связывается с внеклеточным лигандсвязывающим доменом IL-22R. Олигопептиды могут быть химически синтезированными с использованием известного способа синтеза олигопептидов или могут быть получены и очищены с использованием рекомбинантного способа. Такие олигопептиды обычно представляют собой, по крайней мере, примерно 5 аминокислот в длину, альтернативно, по крайней мере, примерно 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100 аминокислот в длину. Такие олигопептиды можно идентифицировать без лишнего экспериментирования с использованием хорошо известных способов. При этом указано, что способы скрининга олигопептидных библиотек для олигопептидов, которые способны специфически связываться с полипептидной мишенью, представляют собой хорошо известные в данной области (см., например, патенты США № 5556762, 5750373, 4708871, 4833092, 5223409, 5403484, 5571689, 5663143; PCT публикации № WO 84/03506 и WO84/03564; Geysen et al., Proc. Natl. Acad. Sci. U.S.A., 81:3998-4002 (1984); Geysen et al. Proc. Natl. Acad. Sci. USA, 82: 178-182 (1985); Geysen et al, in Synthetic Peptides as Antigens, 130-149 (1986); Geysen et al., J. Immunol. Meth., 102:259-274 (1987); Schoofs et al., J. Immunol, 140:611-616 (1988), Cwirla, S. E. et al. (1990) Proc. Natl. Acad. Sci. USA, 87:6378; Lowman, H.B. et al. (1991) Biochemistry, 30: 10832; Clackson T. et al. (1991) Nature, 352: 624; Marks, J. D. et al. (1991), J. Mol. Biol, 222:581; Kang A.S. et al. (1991) Proc. Natl. Acad. Sci. USA, 88:8363, и Smith G. P. (1991) Current Opin. Biotechnol, 2:668). В некоторых вариантах осуществления, олигопептид может конъюгироваться с цитотоксическим веществом.
В еще одном аспекте, антагонист IL-22 представляет собой органическую молекулу, которая связывается с IL-22 или IL-22R, кроме олигопептида или антитела, как описано в данном документе. Органической молекулой может быть, например, маленькая молекула. В одном варианте осуществления, органическая молекула связывается с внеклеточным доменом IL-22R. Органическая молекула, которая связывается с IL-22 или IL-22R, может быть идентифицирована и химически синтезирована с использованием известного способа (см., например, PCT публикации № WO00/00823 и WO00/39585). Такие органические молекулы обычно меньше размером, чем примерно 2000 дальтон, альтернативно меньше размером, чем примерно 1500, 750, 500, 250 или 200 дальтон, где такие органические молекулы, которые способны связываться с IL-22 или IL-22R, можно идентифицировать без лишнего экспериментирования с использованием хорошо известных способов. При этом указано, что способы скрининга библиотек органических молекул для молекул, которые способны связываться с полипептидной мишенью, представляют собой хорошо известные в данной области (см., например, PCT публикации № WO00/00823 и WO00/39585). В некоторых вариантах осуществления, органические молекулы могут конъюгировать с цитотоксическим веществом.
В еще одном аспекте, IL-22 антагонист представляет собой растворимый IL-22 рецептор, например, форму IL-22R, которая не является мембраносвязанной. Такие растворимые формы IL-22R могут конкурировать с мембраносвязанным IL-22R за связывание с IL-22. В некоторых вариантах осуществления, растворимая форма IL-22R может содержать весь или лиганд-связывающий участок внеклеточного домена IL-22R, например весь или лиганд-связывающий участок полипептида, содержащего 18-228 аминокислот из SEQ ID NO: 3. В некоторых вариантах осуществления, растворимая форма IL-22R не имеет трансмембранного домена. Например, растворимая форма человеческого IL-22R может не иметь всего или значительного участка трансмембранного домена из примерно 229-251 аминокислот из SEQ ID NO:3.
Сообщали о встречающемся в природе растворимом рецепторе для IL-22. См. Dumoutier L. et al., "Cloning и characterization of IL-22 binding protein, a natural antagonist of IL-10-related T cell-derived inducible factor/IL-22," J. Immunol. 166:7090-7095 (2001); и Xu W. et al., "A soluble class II cytokine receptor, IL-22RA2, is a naturally occurring IL-22 antagonist," Proc. Natl. Acad. Sci. U.S.A. 98:9511-9516 (2001). Этот рецептор различно обозначают "IL-22BP" или "IL-22RA2" в данной области. Последовательность человеческого IL-22BP показана на фиг. 4. Термин "IL-22BP" или "IL-22 связывающий белок", как использовано в данном документе, относится к любому нативному IL-22BP из любого источника позвоночных, включая таких млекопитающих, как приматы (например, люди и обезьяны) и грызуны (например, мыши и крысы), если не указано особо.
В еще одном аспекте, антагонист IL-22 представляет собой антисмысловую нуклеиновую кислоту, которая уменьшает экспрессию гена IL-22 или IL-22R (то есть, которая уменьшает транскрипцию гена IL-22 или IL-22R и/или трансляцию мРНК IL-22 или IL-22R). В некоторых вариантах осуществления, антисмысловая нуклеиновая кислота связывается с нуклеиновой кислотой (ДНК или РНК), кодирующей IL-22 или IL-22R. В некоторых вариантах осуществления, антисмысловая нуклеиновая кислота представляет собой олигонуклеотид из примерно 10-30 нуклеотидов в длину (включая все точки между такими конечными точками). В некоторых вариантах осуществления, антисмысловой олигонуклеотид содержит измененные сахар-фосфодиэфирные остовы (или другие связи сахаров, включая фосфотиоатные связи, как описано в WO 91/06629), где такие измененные сахар-фосфодиэфирные остовы устойчивы к эндогенным нуклеазам. В одном варианте осуществления, антисмысловая нуклеиновая кислота представляет собой олигодезоксирибонуклеотид, который вызывает разрушение и/или уменьшение транскрипции или трансляции мРНК, кодирующей IL-22 или IL-22R. В некоторых вариантах осуществления, антисмысловая нуклеиновая кислота представляет собой РНК, которая уменьшает экспрессию нуклеиновой кислоты мишени посредством "РНК интерференции" ("RNAi"). Для обзора RNAi, см., например, Novina et al. (2004) Nature 430: 161-164. Такие РНК получают из, например, коротких интерферирующих РНК (siRNAs) и микроРНК. siRNAs, например, можно синтезировать в виде двунитевых олигорибонуклеотидов из примерно 18-26 нуклеотидов в длину.
В еще одном аспекте, представлены агонисты IL-22. Типичные агонисты включают в себя, но ими не ограничиваются, нативный IL-22 или IL-22R; фрагменты, варианты или измененные формы IL-22 или IL-22R, которые сохраняют, по крайней мере, одну активность нативного полипептида; вещества, которые способны связываться и активировать IL-22R; и вещества, которые вызывают гиперэкспрессию IL-22, или IL-22R, или нуклеиновых кислот, кодирующих IL-22 или IL-22R.
E. Фармацевтические составы
В изобретении представлены фармацевтические составы. В одном варианте осуществления, фармацевтический состав содержит 1) активное вещество, например любой из вышеописанных полипептидов, антитела, агонисты или антагонисты; и 2) фармацевтически приемлемый носитель. В дальнейшем варианте осуществления, фармацевтический состав далее содержит, по крайней мере, одно дополнительное терапевтическое вещество.
Фармацевтические составы представляют собой приготовленные для хранения посредством смешивания вещества, имеющего определенную степень очистки, с дополнительными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизированных составов или водных растворов. Приемлемые носители, наполнители или стабилизаторы нетоксичны для реципиентов в применяемых дозах и концентрациях и включают в себя буферы, такие как фосфатные, цитратные и других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консервирующие вещества (такие как октадецилдиметилбензил хлорида аммония; гексаметоний хлорид; бензалконий хлорид, бензетоний хлорид; фенол, бутанол или бензиловый спирт; алкил парааминобензойные кислоты, такие как метил или пропил парааминобензойная кислота; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); полипептиды низкого молекулярного веса (меньше чем примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводороды, включая глюкозу, маннозу или декстрины; хелатообразующие вещества, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие контр-ионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (PEG).
Также можно использовать липофекции или липосомы для доставки вещества внутрь клетки. Если вещество представляет собой фрагмент антитела, то наименьший ингибиторный фрагмент, который специфически связывается с белком мишенью, является предпочтительным. Например, на основании последовательностей вариабельной области антитела можно сконструировать пептидные молекулы, которые сохраняют способность связывать белковую последовательность. Такие пептиды можно синтезировать химически и/или получить рекомбинантным ДНК способом (см., например, Marasco et al., Proc. Natl. Acad. Sci. USA 90, 7889-7893 (1993)). Антитела, раскрытые в данном документе, также можно сформулировать как иммунолипосомы. Липосомы, содержащие антитело, получают способами, известными в данной области, так как описано в Epstein et al, Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al, Proc. Natl Acad. Sci. USA, 77: 4030 (1980); и патентах США № 4485045 и 4544545. Липосомы с увеличенным временем циркуляции раскрыты в патенте США № 5013556. Особенно полезные липосомы можно получить посредством способа обращенно-фазового выпаривания липидного состава, содержащего фосфатидилхолин, холестерин и PEG-производный фосфатидилэтаноламин (PEG-PE). Липосомы пропускают через фильтры с определенным размером пор для получения липосом нужного диаметра. Fab' фрагменты антитела настоящего изобретения можно конъюгировать с липосомами, как описано в Martin et al., J. Biol. Chem., 257: 286-288 (1982) посредством реакции дисульфидного обмена. Химиотерапевтическое вещество (такое как доксорибуцин) дополнительно содержится в липосоме. См. Gabizon et al, J. National Cancer Inst., 81(19): 1484 (1989).
Вещество также может быть заключено в микрокапсулы, полученные, например, способами коацервации или межфазной полимеризацией, например, гидроксиэтилцеллюлозных или желатиновых микрокапсул и микрокапсулы полиметилметацилата, соответственно, в коллоидные системы доставки лекарственного вещества к участку действия (например, липосомы, альбуминовые микросферы, микроэмульсия, наночастицы и нанокапсулы) или в макроэмульсиях. Такие способы раскрыты в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получить препараты с замедленным высвобождением вещества. Подходящие примеры препаратов с замедленным высвобождением включают в себя полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих вещество, матрицы которого представлены в форме профилированных изделий, например, пленки или микрокапсул. Примеры матриц с замедленным высвобождением включают в себя полиэфиры, гидрогели, например, поли(2-гидроксиэтилметакрилат), или поливиниловый спирт, полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глютамата, неразлагающийся этиленвинилацетат, разлагающиеся сополимеры молочной кислоты и гликолиевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолиевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксибутановую кислоту. Кроме того, полимеры, такие как этиленвинилацетат и молочная кислота - гликолиевая кислота, способствующие высвобождению молекул в течение более 100 дней, некоторые гидрогели высвобождают белки в течение более короткого периода времени. Если инкапсулированные антитела сохраняются в теле в течение долгого времени, то они могут денатурировать или агрегировать в результате воздействия воды при 37°C, приводя к уменьшению биологической активности и возможных изменений в иммуногенности. Могут быть разработаны рациональные стратегии для стабилизации в зависимости от включенного механизма. Например, если обнаружено, что механизм агрегации является внутримолекулярным образованием S-S связей путем тиодисульфидного обмена, то стабилизации можно достигнуть посредством модифицирования сульфгидрильных остатков, лиофилизированных кислыми растворами, регулируя влагосодержание, используя подходящие добавки и разрабатывая специфические составы полимерных матриксов.
Фармацевтический состав в данном документе также может содержать более чем одно активное соединение в случае необходимости указания лечения. Например, в одном аспекте, фармацевтический состав, содержащий более чем одно активное соединение, содержит 1) по крайней мере, один антагонист IL-22, например антитело, которое связывается с IL-22, и/или антитело, которое связывается с IL-22R; и 2) по крайней мере, одно антитело, которое связывается с IL-19, IL-20, IL-24, IL20Ra, IL-20Rb или IL-10R2 (где любое количество антител, перечисленных в 2), можно выбрать в любом сочетании). В другом аспекте, фармацевтический состав содержит два или более активных соединения, обладающих взаимодополняющими активностями. Например, в одном варианте осуществления, фармацевтический состав может содержать 1) по крайней мере, один антагонист IL-22, например антитело, которое связывается с IL-22, и/или антитело, которое связывается с IL-22R; и 2) антагонист TNF-α или IL-12. В еще одном аспекте, фармацевтический состав, содержащий более чем одно активное соединение, может содержать цитотоксическое вещество или ингибитор роста.
F. Способы лечения
Представлены терапевтические способы с использованием вышеупомянутых составов или фармацевтических составов. Такие способы включают в себя in vitro, ex vivo и in vivo терапевтические способы, если не указано особо. В различных аспектах, представлены способы стимуляции или ингибирования IL-23-опосредованного пути передачи сигнала. Представлены способы стимуляции или ингибирования функции ThIL-17 клеток. Также представлены способы лечения воспалительных и/или аутоиммунных нарушений. Далее представлены способы лечения нарушений, связанных с IL-23 или IL-22 передачей сигнала. Также представлены способы лечения ThIL-17-апосредованных нарушений. Ниже представлены эти и другие аспекты изобретения.
В одном аспекте, представлен способ стимуляции IL-23 опосредованного пути передачи сигнала в биологической системе, способ, включающий обеспечение биологических систем IL-22 агонистом. Биологические системы включают в себя, например, клетки млекопитающих в in vitro системы клеточной культуры или в организме in vivo. Типичные биологические системы, которые моделируют псориаз, представлены в примерах и включают в себя восстановленный человеческий эпидермис (RHE) (пример 14) или животные модели (пример 16). В одном варианте осуществления, IL-22 агонист представляет собой IL-22. В другом аспекте, представлен способ ингибирования IL-23-опосредованного пути передачи сигнала в биологической системе, способ, включающий обеспечение биологических систем IL-22 антагонистом. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело.
В другом аспекте, представлен способ стимуляции функции ThIL-17 клеток, способ, включающий воздействие IL-22 агониста на ThIL-17 клетки. В одном варианте осуществления, IL-22 агонист представляет собой IL-22. В другом аспекте, представлен способ ингибирования функции ThIL-17 клеток, способ, включающий воздействие IL-22 антагонист на ThIL-17 клетки. В одном варианте осуществления, IL-22 антагонист представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело. Типичные функции ThIL-17 клеток включают в себя, но ими не ограничиваются, стимуляцию клеточно-опосредованного иммунитета (гиперчувствительность замедленного типа); рекрутирование характерных иммунных клеток, таких как миелоидные клетки (например, моноциты и нейтрофилы), к участкам воспаления; и стимуляцию инфильтрации воспалительных клеток в ткани. В одном варианте осуществления, функция ThIL-17 клеток опосредуется IL-23.
В еще одном аспекте, представлен способ лечения воспаления, способ, включающий введение млекопитающему при необходимости такого лечения эффективного количества фармацевтического состава, включающего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело. Воспаление включает в себя, но ими не ограничивается, аутоиммунное воспаление (воспаление, связанное с аутоиммунным нарушением), хроническое воспаление, кожное воспаление, артритическое воспаление (включая воспаление, связанное с ревматоидным артритом) и системную воспалительную реакцию. В одном варианте осуществления, воспаление опосредуется IL-23.
В еще одном аспекте, представлен способ лечения аутоиммунного нарушения, способ, включающий введение млекопитающему при необходимости такого лечения эффективного количества фармацевтического состава, включающего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело. Аутоиммунное нарушение включает в себя, но ими не ограничивается, болезни соединительной ткани, множественный склероз, системную красную волчанку, воспалительный артрит (например, ревматоидный артрит), аутоиммунное воспаление легких, синдром Гийена-Барре, аутоиммунный тиреоидит, инсулинозависимый сахарный диабет, увеит, миастению gravis, реакцию "трансплантант против хозяина", аутоиммунное воспалительное заболевание глаз, псориаз, артрит, связанный с аутоиммунной реакцией (например, ревматоидный артрит), аутоиммунное воспаление мозга и воспалительную болезнь кишечника. В одном варианте осуществления, аутоиммунное нарушение представляет собой IL-23-опосредованное аутоиммунное нарушение.
В отдельном аспекте, представлены способы лечения псориаза и/или нарушений, характеризующиеся псориатическими симптомами. Псориаз рассматривается как аутоиммунное заболевание, при котором T-клетки иммунной системы узнают белок в коже и атакуют область, в которой обнаружен этот белок, вызывая очень быстрый рост новых клеток кожи и болезненность, припухание, поверхностные повреждения. Эти повреждения характеризуются гиперпролиферацией кератиноцитов и накоплением активированных T-клеток в эпидермисе псориатических повреждений. Хотя начальная молекулярная причина заболевания не известна, были установлены генетические связи к, по крайней мере, 7 локусу предрасположенности к псориазу (Psor1 в 6p21,3, Psor2 в 17q, Psor3 в 4q, Psor4 в 1 cent-q21, Psor5 в 3q21, Psor6 в 19pl3, и Psor7 в 1p). Некоторые из этих локусов связаны с другими аутоиммунными/воспалительными заболеваниями, включающими ревматоидный артрит, атопический дерматит и воспалительную болезнь кишечника (IBD). Общепринятые подходы к лечению псориаза включают в себя введение IL-12 или TNF-α антагонистов. См., например, Nickoloff et al. (2004) J.Clin. Invest. 113:1664-1675; Bowcock et al. (2005) Nat. Rev. Immunol. 5:699-711; Kauffman et al. (2004) J. Invest. Dermatol. 123:1037-1044. Данные, представленные в данном документе, однако, включают в себя отдельный IL-23/IL-22 путь передачи сигнала в патогенезе псориаза. Соответственно, лечение, которое модулирует этот путь передачи сигнала, может представлять альтернативу или может дополнять другие подходы к лечению псориаза.
В одном варианте осуществления, способ лечения псориаза включает введение пациенту эффективного количества фармацевтического состава, содержащего IL-22 антагониста. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело. В различных вариантах осуществления, способ далее включает введение (либо в одном и том же фармацевтическом составе, либо в раздельном фармацевтическом составе), по крайней мере, одного дополнительного терапевтического вещества. В одном таком варианте осуществления, дополнительное терапевтическое вещество представляет собой по крайней мере один антагонист цитокина, выбранного из IL-19, IL-20 и IL-24. Такие антагонисты включают в себя, но ими неограничиваются, антитело, которое связывает IL-19, IL-20, IL-24, IL-20Ra, IL-20Rb или IL-10R2. Любое количество таких антител можно выбирать в любом сочетании. В другом варианте осуществления, дополнительное терапевтическое вещество представляет собой вещество, известное эффективностью при лечении псориаза. Некоторые из таких терапевтических веществ описаны, например, в Nickoloff et al. (2004) J. Clin. Invest. 113:1664-1675; Bowcock et al. (2005) Nat. Rev. Immunol. 5:699-711; и Kauffman et al. (2004) J. Invest. Dermatol. 123:1037-1044. Такие вещества включают в себя, но ими не ограничиваются, терапевтическое вещество, которое нацеливает Т-клетки, например эфализумаб и/или алефацепт; антагонист IL-12, например, блокирующий антитело, которое связывает IL-12 или его рецептор; и антагонист TNF-α, например блокирующее антитело, которое связывает TNF-α или его рецептор.
В еще одном аспекте, представлен способ ингибирования опухолевой прогрессии, способ, включающий введение млекопитающему эффективного количества фармацевтического состава, содержащего антагонист IL-22. В одном варианте осуществления, антагонист IL-22 представляет собой антитело, например нейтрализующее анти-IL-22 антитело и/или нейтрализующее анти-IL-22R антитело. В одном варианте осуществления, опухолевая прогрессия является IL-23 опосредованной.
Составы настоящего изобретения (например, полипептиды, антитела, антагонисты, агонисты и фармацевтические составы, содержащие любые из вышеупомянутых) вводят млекопитающему, предпочтительно человеку, в соответствии с известными способами, такими как внутривенное введение в виде болюса или непрерывным вливанием на протяжении времени, внутримышечными, внутрибрюшинными, внутрицереброспинальными, подкожными, внутрисуставными, внутриоболочечными, пероральными, местными или ингаляционными (интраназальными, внутрилегочными) курсами. Внутривенное или ингаляционное введение полипептидов и антител предпочтительны.
В некоторых вариантах осуществления, введение антиракового вещества можно сочетать с введением состава этого изобретения. Например, чтобы лечиться составом изобретения, пациент также может получать антираковое вещество (химиотерапевтическое вещество) или радиационную терапию. Графики приема и дозирование таких химиотерапевтических веществ можно использовать согласно инструкциям изготовителя или как имперически определено квалифицированным специалистом. Графики приема и дозирование такой химиотерапии также описаны в Chemotherapy Service Ed., M. C. Perry, Williams & Wilkins, Baltimore, MD (1992). Химиотерапевтическое вещество может предшествовать или следовать за введением состава или может быть дан одновременно с тем. Дополнительно, соединение антиэстрогена, такое как тамоксифен, или антипрогестерон, такой как онапристон (см., EP 616812) могут быть даны в дозах, известных для таких молекул.
Желательным может быть также введение антител против антигенов, связанных с другим иммунным заболеванием или опухолью, таких как антитела, которые связываются с CD20, CD11a, CD18, ErbB2, EGFR, ErbB3, ErbB4 или фактором роста эндотелия сосудов (VEGF). Альтернативно, или в дополнение, два или более антитела, связывающие те же самые или два или более различных антигена, описанных в данном документе, можно вводить одновременно пациенту. В некоторых вариантах осуществления, благоприятным может быть также введение одного или более цитокинов пациенту. В некоторых вариантах осуществления, составы изобретения вводятся одновременно с ингибитором роста. Например, ингибитор роста можно вводить до, после или одновременно с введением состава. Подходящие дозы ингибитора роста представляют собой используемые в настоящее время дозы и могут быть снижены вследствие комбинированного действия (синергизм) ингибитора роста и состава.
Для лечения или уменьшения тяжести иммунного заболевания подходящая доза состава изобретения будет зависеть от типа заболевания, которое лечат, как определено выше, тяжести и причины болезни, вводится ли вещество для предупреждения или с терапевтическими целями, проведенной ранее терапии, клинической истории болезни пациента и ответа на соединение, и усмотрения лечащего врача. Соединения удобным способом вводят пациенту единовременно или в течение всего периода лечения.
Например, в зависимости от типа и тяжести заболевания, примерно 1 мкг/кг-15 мг/кг (например, 0,1-20 мг/кг) полипептида или антитела является начальной дозой для введения пациенту, либо, например, одним или более раздельными введениями, либо продолжительным вливанием. Обычно дневная доза может варьировать от примерно 1 мкг/кг до 100 мг/кг или более, в зависимости от вышеупомянутых факторов. Для повторного введения через несколько дней или больше в зависимости от состояния лечение поддерживают до желательного подавления наблюдаемых симптомов заболевания. Однако могут использоваться другие режимы доз. Прогресс этой терапии легко контролировать обычными способами и исследованиями.
G. Диагностические способы и способы определения
В одном аспекте, представлен способ диагностирования псориаза у млекопитающих, способ, включающий определение уровня экспрессии гена, кодирующего IL-22 или IL-22R полипептид, в исследуемом образце клеток ткани, полученных из млекопитающего, где более высокий уровень экспрессии в исследуемом образце по сравнению с контрольным образцом (например, образец известных нормальных клеток ткани того же типа клеток) показывает наличие псориаза у млекопитающего, из которого был получен исследуемый образец. Определение может быть количественным или качественным. В одном варианте осуществления, исследуемый образец содержит кровь или сыворотку. В одном варианте осуществления, определение уровня экспрессии гена, кодирующего IL-22 или IL-22R полипептид, содержит (a) взаимодействие анти-IL-22 или анти-IL-22R антитела с исследуемым образцом, полученного из млекопитающего, и (b) определение образования комплекса между антителом и полипептидом IL-22 или IL-22R в исследуемом образце. Антитело может быть связано с определяемой меткой. Образование комплекса можно наблюдать, например, с помощью световой микроскопии, проточной цитометрии, флуорометрии или другими способами, известными в данной области. Исследуемый образец можно получать от индивидов, подозреваемых в наличии псориаза.
В одном варианте осуществления, определение уровня экспрессии гена, кодирующего IL-22 или IL-22R полипептид, включает определение уровня транскрипции мРНК с гена. Уровни транскрипции мРНК можно определить либо количественно, либо качественно, различными способами, известными специалисту в данной области. Уровни транскрипции мРНК можно также определить прямо или косвенно путем определения уровней кДНК, полученной из мРНК. Обычные способы определения уровней транскрипции мРНК включают в себя, но ими не ограничивается, количественную в реальном времени RT-ПЦР и способ, основанный на гибридизации, включая микрочиповый анализ и анализы, основанные на фильтрах, такие как Нозерн-блотты.
К другому варианту осуществления настоящего изобретения относится диагностический набор, содержащий анти-IL-22 или анти-IL-22R антитело в удобной упаковке. Предпочтительно набор содержит инструкцию для использования антитела, для определения IL-22 или IL-22R полипептида. В одном аспекте, диагностический набор представляет собой диагностический набор для псориаза.
H. Исследования
1. Исследования, основанные на клетках и животной модели
Исследования, основанные на клетках и животной модели для иммунных заболеваний, используют в практике некоторых вариантов осуществления изобретения. Некоторые исследования, основанные на клетках, представленные в примерах ниже, используют, например, для исследования эффективности IL-22 антагонистов или агонистов.
Также используют животные модели in vivo в практике некоторых вариантов осуществления изобретения. Типичные животные модели также описаны в примерах ниже. Природа in vivo таких моделей позволяет предсказывать ответы у человеческих пациентов. Животные модели иммуносвязанных заболеваний включает в себя и нерекомбинантных, и рекомбинантных (трансгенных) животных. Нерекомбинантные животные модели включают в себя, например, грызунов, например, мышиные модели. Такие модели можно получать путем введения клеток в сингенных мышей с использованием стандартных способов, например подкожного введения, введения в хвостовую вену, имплантации селезенки, внутрибрюшинной имплантации, имплантации в капсулу почки и другие.
Модели реакции "трансплантант против хозяина" представляют способы оценки T-клеточной реактивности против MHC антигенов и минорные трансплантируемые антигены. Реакция "трансплантант против хозяина" наблюдается, когда иммунокомпетентные клетки трансплантируют пациентам с подавленной иммунной системой или устойчивым. Донорные клетки узнают и реагируют на антигены. Ответ может варьировать в пределах от угрожающего жизни тяжелого воспаления до легкой степени заболевания диареей и потери веса. Подходящий способ оценки реакции "трансплантант против хозяина" описан подробно в Current Protocols in Immunology, выше, часть 4.3.
Животная модель отторжения кожного аллотрансплантанта представляет собой способ исследования способности Т-клеток вызывать in vivo разрушение тканей и оценку их роли в отторжении трансплантанта. Самые распространенные и признанные модели используют кожный трансплантат мышиного хвоста. Повторные эксперименты показали, что отторжение кожного аллотрансплантанта опосредуется Т-клетками, хелперными Т-клетками и киллерными Т-клетками, а не антителами. Auchincloss, H. Jr. и Sachs, D. H., Fundamental Immunology, 2nd ed., W. E. Paul ed., Raven Press, NY, 1989, 889-992. Подходящий способ описан подробно в Current Protocols in Immunology, выше, часть 4.4. Другие модели отторжения транслантанта, которые можно использовать для исследования соединений изобретения, представляют собой модели аллогенной пересадки сердца, описанные Tanabe, M. et al, Transplantation (1994) 58:23 и Tinubu, S. A. et al, J. Immunol. (1994) 4330-4338.
Контактная гиперчувствительность является простым исследованием in vivo для клеточно-опосредованной иммунной функции (гиперчувствительность замедленного типа). В этом способе, взаимодействие кожи с экзогенными гаптенами приводит к реакции гиперчувствительности замедленного типа, которая оценивается и измеряется количественно. Контактная гиперчувствительность включает в себя начальную фазу сенсибилизации, за которой следует фаза получения вывода. Фазу получения вывода наблюдают, если T-лимфоциты встречают антиген, с которым они прежде имели контакт. Опухание и воспаление наблюдают, производя это на прекрасной модели аллергического контактного дерматита человека. Подходящий способ описан подробно в Current Protocols in Immunology, Eds. J. E. Cologan, A. M. Kruisbeek, D. H. Margulies, E. M. Shevach and W. Strober, John Wiley & Sons, Inc., 1994, unit 4.2. См. также Grabbe S. and Schwarz T., Immun. Today 19 (1): 37-44 (1998).
Дополнительно, составы изобретения можно исследовать на животных моделях для псориазподобных заболеваний. Например, составы изобретения можно исследовать на scid/scid мышиной модели, описанной Schon, M. P. et al, Nat. Med. (1997) 3: 183, в которой мыши показывают гистопатологические кожные повреждения, сходные с псориазом. Другая подходящая модель представляет собой химеру человеческая кожа/scid мыши, полученные, как описано в Nickoloff B. J. et al, Am. J. Path. (1995) 146:580. Другая подходящая модель описана в Boyman et al., J. Exp. Med. (2004) 199(5):731-6, в котором кожа человека до развития псориаза трансплантируется в AGR129 мышей, приводя к развитию псориатических кожных повреждений.
Можно сконструировать нокаутных животных, которые содержат дефектный или измененный ген, кодирующий полипептид, идентифицированный в данном документе, как результат гомологичной рекомбинации между эндогенным геном, кодирующим полипептид, и молекулой ДНК, в которой этот ген был изменен. Например, кДНК, кодирующую отдельный полипептид, можно использовать для клонирования геномной ДНК, кодирующей этот полипептид в соответствии с установленными способами. Участок геномной ДНК, кодирующей отдельный полипептид, можно удалить или переместить с другим геном, таким как ген, кодирующий селектируемый маркер, который можно использовать для контролируемой интеграции. Обычно несколько тысяч пар оснований неизмененной фланкирующей ДНК (и на 5', и 3' концах) включаются в вектор (см., например, Thomas and Capecchi, Cell, 51:503 (1987) для описания гомологичных рекомбинантных векторов). Вектор вводят в линию эмбриональных стволовых клеток (например, посредством электропорации) и отбираются клетки, в которых введенная ДНК гомологично рекомбинировала с эндогенной ДНК (см., например, Li et al., Cell, 69:915 (1992)). Выбранные клетки затем вводили в бластоцисту животного (например, мыши или крысы) для получения агрегации химер (см., например, Bradley, в Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp. 113-152). Затем химерный эмбрион можно имплантировать в подходящую псевдобеременную самку приемного животного и эмбриону присваивают термин "нокаутного" животного. Потомка, содежащего гомологично рекомбинированную ДНК в своих половых клетках, можно идентифицировать стандартными способами и использовать для выведения животных, у которых все клетки животного содержат гомологично рекомбинированную ДНК. Нокаутных животных можно характеризовать, например, по их способности сопротивляться некоторым патологическим условиям и по их развитию в патологических условиях вследствие отсутствия полипептида.
2. Скрининговые исследования лекарственных кандидатов
Скрининговые исследования лекарственных кандидатов направлены на идентификацию соединений, которые связываются или образуют комплекс с полипептидами, идентифицированными в данном документе, или их биологически активными компонентами, или по-другому препятствуют взаимодействию полипептида с другими клеточными белками. Такие скрининговые исследования будут включать в себя исследования, ответственные за высокопроизводительный скрининг из химических библиотек, делая их более удобными для идентификации малых молекул лекарственных кандидатов. Предполагаемые малые молекулы включают в себя синтетические органические или неорганические соединения, включая пептиды, предпочтительно растворимые пептиды, полипептидные - иммуноглобулиновые слияния и, в частности, антитела, включающие, без ограничений, поли- и моноклональные антитела и фрагменты антител, одноцепочечные антитела, анти-идиотипические антитела и химерные или гуманизированные версии таких антител или фрагментов, а также и человеческие антитела и фрагменты антител. Исследования можно осуществлять в различных форматах, включая исследования белок-белкового связывания, исследования биохимического скрининга, иммунологический анализ и исследования, основанные на клетках, которые хорошо охарактеризованы в данной области. Все исследования являются общепринятыми в том, что они требуют контактирования исследуемого соединения с полипептидом, идентифицированным в данном документе при условиях и в течение времени, достаточного, чтобы позволить полипептиду взаимодействовать с исследуемым веществом.
При анализе связывания взаимодействие представляет собой связывание, и образованный комплекс можно выделить или определить в реакционной смеси. В отдельном варианте осуществления, полипептид или исследуемое вещество иммобилизованы на твердой фазе, например на титрационном микропланшете, путем ковалентного или нековалентного связывания. Нековалентное связывание, как правило, осуществляется покрытием твердой поверхности раствором полипептида или исследуемого соединения и высушивания. Альтернативно, иммобилизированное антитело, например моноклональное антитело, специфическое для полипептида, для того чтобы иммобилизовать, можно использовать фиксированный полипептид к твердой поверхности. Исследование осуществляют добавлением неиммобилизованного компонента, который можно метить определяемой меткой, к иммобилизованному компоненту, например поверхности с покрытием, содержащей фиксированный компонент. Когда заканчивается реакция, непрореагировавшие компоненты удаляют, например, отмыванием и определяют фиксированные комплексы на твердой поверхности. Когда первоначально неиммобилизированный компонент несет определяемую метку, определение метки, иммобилизированной на поверхности, показывает, что произошло образование комплекса. Где первоначально неиммобилизированный компонент не несет метку, образование комплекса можно обнаружить, например, используя меченые антитела, специфически связывающие иммобилизованный комплекс.
Если исследуемое соединение взаимодействует, но не связывается с отдельным полипептидом, идентифицированным в данном документе, это взаимодействие с этим белком можно исследовать хорошо известными способами определения белок-белковых взаимодействий. Такие исследования включают в себя традиционные способы, такие как образование поперечных связей, ко-иммуннопреципитация и ко-очистка по градиентам или на хроматографических колонках. Кроме того, белок-белковые взаимодействия можно наблюдать, используя генетические системы дрожжей, описанные Fields и соавторами (Fields and Song, Nature (London) 340, 245-246 (1989); Chien et al, Proc. Natl. Acad. Sci. USA 88, 9578-9582 (1991)), как раскрыто Chevray and Nathans, Proc. Natl. Acad. Sci. USA 89, 5789-5793 (1991). Многие активаторы транскрипции, такой как GAL4 дрожжей, состоят из двух физически отдельных модулярных доменов, один - действующий в качестве ДНК-связывающего домена, хотя другой - функционирующий в качестве домена активации транскрипции. Экспрессирующая система дрожжей, описанная в вышеупомянутых публикациях (как правило, называемые как "двугибридная система"), использует преимущество этого свойства и использует двугибридные белки, один, в котором белок-мишень сливается с ДНК-связывающим доменом GAL4, и другой, в котором активирующие кандидата белки сливаются с активационным доменом. Экспрессия GAL1-lacZ репортерного гена под контролем GAL4-активированного промотора зависит от восстановления GAL4 активности посредством белок-белкового взаимодействия. Колонии, содержащие взаимодействующие полипептиды, определяются хромогенным субстратом для β-галактозидазы. Полный набор (MATCHMAKERTM) для идентификации белок-белковых взаимодействий между специфическими белками с использованием двухгибридного способа является коммерчески доступным от Clontech. Эта система также может распространяться на картированные белковые домены, включенные в специфические взаимодействия белков, а также и для точек определения аминокислотных остатков, которые являются значимыми для этих взаимодействий.
Для идентификации соединений, которые препятствуют взаимодействию полипептида, идентифицированного в данном документе, и другим внутри- или внеклеточным компонентам, можно приготовить реакционную смесь, содержащую полипептид и компонент в условиях, подходящих для взаимодействия полипептида с компонентом. Для исследования способности исследуемого соединения ингибировать взаимодействие готовят реакционную смесь в отсутствие и в присутствии исследуемого соединения. Если есть уменьшение взаимодействия полипептида с компонентом в присутствии исследуемого соединения, то затем говорят, что исследуемое соединение ингибирует взаимодействие полипептида с компонентом.
В некоторых вариантах осуществления, способы идентификации агонистов или антагонистов IL-22 или IL-22R полипептида включают контактирование IL-22 или IL-22R полипептида с молекулой кандидатом агониста или антагониста и оценку определяемого изменения в одной или более биологических активностях, обычно связанных с IL-22 или IL-22R полипептидом. Такие активности включают в себя, но ими не ограничиваются, описанные в примерах ниже.
3. Анализ связывания антител
Изучение связывания антител можно производить любым известным способом исследования, таким как конкурентно-связывающий анализ, прямые и непрямые "сэндвич" исследования и исследование иммунопреципитацией. Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc., 1987).
Конкурентно-связывающий анализ основан на способности меченого стандарта конкурировать с аналитом исследуемого образца за связывание с ограниченным количеством антитела. Количество белка-мишени в исследуемом образце обратно пропорционально количеству стандарта, которое связывается с антителами. Чтобы способствовать определению количества стандарта, который связывается, антитела предпочтительно переводят в нерастворимую форму до или после конкурирования, так что стандарт и аналит, связанные с антителами, можно удобным образом отделить от стандарта и аналита, которые остались несвязанными.
"Сэндвич" исследования включают использования двух антител, каждое из которых способно связываться с различным иммуногенным участком или эпитопом белка для обнаружения. В "сэндвич" исследовании, аналит исследуемого образца связан первым антителом, которое иммобилизировано на твердом носителе, и после этого второе антитело связывается с аналитом, образуя таким образом нерастворимый трехчленный комплекс. См., например, патент США № 4376110. Второе антитело само может быть мечено определяемым фрагментом (прямые "сэндвич" исследования) или может быть оценено с использованием антииммуноглобулинового антитела, которое мечено определяемым фрагментом (непрямые "сэндвич" исследования). Например, один тип "сэндвич" исследования представляет собой ELISA исследование, в случае которого определяемым фрагментом является фермент.
Иммуногистохимия также может использоваться для определения клеточной локализации антигена, с которым связывается антитело. Для иммуногистохимии образец тканей может быть свежим или замороженным или может быть помещен в парафин и фиксирован консервирующим веществом, таким как формалин, например.
Изделия
В другом аспекте, представлены изделия, содержащие составы, используемые для диагностики и лечения нарушений, описанные выше. Изделия содержат контейнер и инструкцию. Подходящие контейнеры включают в себя, например, колбы, ампулы, пипетки и пробирки. Контейнеры могут быть сделаны из различных материалов, таких как стекло или пластик. Контейнер содержит в себе составы, которые эффективны для диагностики или лечения состояния, и может иметь стерильное выпускное отверстие (например, контейнер может представлять собой внутривенный мешок с раствором или ампулу, имеющую крышку, поддающуюся прокалыванию иглой для подкожных инъекций). Активное вещество в составе обычно представляет собой полипептид, антитело, агонист или антагонист изобретения. Инструкция или наклейки на, или связанные с контейнером показывают, что состав используют для диагностики или лечения состояния из вариантов. Изделия далее могут содержать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как фосфатно-буферный солевой раствор, раствор Рингера и раствор декстрозы. Далее он может включать в себя другие материалы, нужные с коммерческой и потребительской точки зрения, включая другие буферы, растворители, фильтры, иглы, шприцы и информацию о лекарстве с инструкциями использования.
В одном варианте осуществления изобретения представлено изделие, включающее:
a) химический состав, содержащий агонист или антагонист IL-22 или IL-22R;
b) контейнер, содержащий указанный состав; и
c) метку, прикрепленную к указанному контейнеру, или информацию о лекарстве, включаущую в себя указанный контейнер, отнесенный к использованию указанного антагониста при лечении иммунного заболевания или рака. Состав может содержать эффективное количество антагониста.
Следующие примеры представлены только для иллюстративных целей и не предназначены ограничивать объем настоящего изобретения никаким образом.
Раскрытие всех патентных и литературных ссылок, упомянутых в настоящем описании, включено посредством ссылки полностью.
III. ПРИМЕРЫ
Коммерчески доступные реактивы, упомянутые в примерах, использовали согласно инструкциям изготовителя, если не указано особо. Источник таких клеток, идентифицированных в следующих примерах и во всем описании с помощью ATCC инвентарных номеров, представляет собой American Type Culture Collection, Manassas, VA.
ПРИМЕР 1: Получение анти-IL-22 и анти-IL-22R антител
В этом примере показано получение моноклональных антител, которые специфически связывают IL-22 или IL-22R. Использованные для получения способы основывались на способах, известных в данной области, и описаны, например, в Goding, выше. Использованные иммуногены были полноразмерными очищенными человеческими IL-22 (hIL-22) или полноразмерными очищенными человеческими IL-22R (hIL-22R). Кратко, мышей иммунизировали примерно 1-100 мкг hIL-22 или hIL-22R иммуногена, эмульгированного во вспомогательном веществе. Иммунизированных мышей затем повторно подвергают стимуляции спустя 10-12 дней дополнительным иммуногеном, эмульгированным во вспомогательном веществе. Периодически от мышей получали образцы сыворотки для исследования способами ELISA, чтобы определить анти-IL-22 или IL-22R антитела.
После определения нужного титра антител "положительных" по антителам животных умерщвляли и получали клетки селезенки. Клетки селезенки затем сливались (используя 35% полиэтиленгликоль) с мышиной линией миеломных клеток. Слиянием были получены гибридомные клетки, которые были клонированы и культивировались в среде, содержащей HAT (гипоксантин, аминоптерин и тимидин). Гибридомные клетки скринировали ELISA на реактивность против IL-22 или IL-22R. (см. фиг.5). Список антител, полученных из таких гибридом, и их соответствующие свойства находятся на фиг. 5.
ПРИМЕР 2: передача сигнала IL-22 блокируется анти-IL-22 антителами
Активация STAT3 представляет собой отличительный признак активации рецептора IL-22 и внутриклеточной передачи сигнала. Полученные против человеческого IL-22 антитела были исследованы на их способность блокировать IL-22-индуцированную активацию STAT3. 293 Т-клетки, экспрессирующие гетеродимер рецептора IL-22 человека (hIL-22R/hIL-10R2), были рассажены в 0,2×106/лунку 24-луночного планшета. Клетки трансфицировали STAT3 люциферазным репортером (TK-SIE-SRE-S) с использованием Lipofectamine 2000TM (Invitrogen). Следовательно, при активации STAT3 клетки будут продуцировать люциферазу, ферментную активность которой можно определить добавлением люциферина. Уменьшение люциферазной активности означает, что блокируется STAT3. На следующий день добавляли 0,5 нМ hIL-22 (R&D Systems) в каждую лунку вместе с 20 мкг/мл антитела. Спустя шестнадцать часов клетки лизировали и определяли показания образцов на люминометре. Показанные на фиг. 6 данные представляют собой люциферазную активность относительно Renilla внутреннего контроля, которая является мерой относительной активации STAT3. Как показано на фиг. 6, антитела 3F11.3, 11H4.4 и 8E11.9 обладали значительной блокирующей способностью.
ПРИМЕР 3: Дозозависимый ответ анти-IL-22 антител
Ряд доз антител, полученных против человеческого IL-22, были исследованы на их способность блокировать человеческий IL-22 в исследовании активации STAT3. 293 клетки, экспрессирующие hIL-22R/hIL-10R2, были рассажены в 0,2×106/лунку 24-луночного планшета. Клетки трансфицировали STAT3 люциферазным репортером (TK-SIE-SRE-S) с использованием Lipofectamine 2000TM (Invitrogen). На следующий день добавляли 0,5 нМ hIL-22 (R&D Systems) в каждую лунку вместе с различными концентрациями анти-IL-22 антителами 3F11, 8E11 или 11H4. Разброс концентраций антитела начинался с 40 мкг/мл с 2-кратными разведениями до конечной концентрации 0,012 мкг/мл. Спустя шестнадцать часов клетки лизировали, и определяли показания образцов на люминометре. Три антитела показали сходную кривую доза/ответ блокирования активации STAT3, как показано на фиг. 7.
ПРИМЕР 4: Дозозависимый ответ анти-IL-22 антител
Ряд доз антител, полученных против человеческого IL-22, были исследованы на их способность блокировать мышиный IL-22 (mIL-22) в исследовании активации STAT3. 293 клетки, экспрессирующие mIL-22R/mIL-10Rb, были рассажены в 0,2×106/лунку 24-луночного планшета. Клетки трансфицировали STAT3 люциферазным репортером (TK-SIE-SRE-S) с использованием Lipofectamine 2000TM (Invitrogen). На следующий день добавляли 0,5 нМ mIL-22 (полигистидин меченый) в каждую лунку вместе с различными концентрациями антител 3F11, 8E11 или 11H4. Разброс концентраций антитела начинался с 40 мкг/мл с 2-кратными разведениями до конечной концентрации 0,012 мкг/мл. Спустя шестнадцать часов клетки лизировали и определяли показания образцов на люминометре. На фиг. 8 показано, что анти-IL-22 антитела перекрестно реагируют с мышиным IL-22, и показана сходная, но не жестко, кривая доза/ответ. Это показывает, что анти-IL-22 антитела можно использовать в мышиных экспериментах.
ПРИМЕР 5: Аффинность анти-IL-22 к человеческому IL-22
На фиг. 9 показана аффинность анти-IL-22 к человеческому IL-22. Аффинность оценивали BIACore анализом. Различные количества анти-IL-22 IgG были иммобилизованы на CM 5 чип (845 RU (ссылочные единицы) для 11H4 IgG, 1933 RU для 8E11 IgG, & 7914 RU для 3F11 IgG) посредством гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS) химического соединения. Двукратные серийные разведения IL-22 были приготовлены в пределах 0,5-250 нМ. Образцы антигена были нанесены поверх IgG-иммобилизованной поверхности при скорости потока 20 мкл/мин в течении 6 минут, и связанные комплексы были подвергнуты диссоциации на 10 минут. Поверхности IgG восстанавливали 10 мМ Gly, pH 1,5 после каждого периода введения антигена. В качестве отрицательного контроля клеток, посторонний IgG (3A5 RF трансплантант) был иммобилизирован для вычитания уровня фона. Разделяющий буфер, PBS, содержащий 0,05% Tween 20 с 0,01% NaN3, использовали для всех разведений образцов и эксперимент связывания проводили при 25°C. Данные анализировали универсально в соответствии с 1:1 моделью связывания. Эти результаты показывают, что анти-IL-22 антитела обладают очень хорошей аффинностью к человеческому IL-22.
ПРИМЕР 6: Анти-IL-22 антитела определяют IL-22 в клетке
Были исследованы антитела против IL-22 на способность определять внутриклеточный IL-22. Для внутриклеточного FACS окрашивания IL-22, были использованы следующие линии 293 клеток: клетки, экспрессирующие hIL-22-GFP, mIL-22-GFP, mIL20-GFP и GFP только. Исследуемые антитела представляли собой античеловеческие IL-22 антитела 3F11, 8E11 и 17F6. Мышиные анти-gp120 использовали в качестве контрольного изотипа. Вторичное использованное антитело представляло собой антимышиный IgG-PE из Jackson labs. Клетки инкубировали с Brefeldin A в течение 2 часов, отмывали в PBS и затем фиксировали 2% параформальдегидом в течение ночи при 4°C. Затем клетки отмывали в PBS и инкубировали в 5 мл 0,2% Tween-20 в течение 30 минут при 37°C. Окрашивание антителами проводили в течение 30 минут при 4°C, затем отмывали раствором Tween-20. Клетки ресуспендировали в FACS буфере и анализировали в FACScan. На фиг. 10 показаны FACS результаты. FACS результаты показывают, что антитела 3F11 и 8E11 вызывают изменение паттерна окрашивания клеток, показывая, что эти антитела связывают и мышиный, и человеческий внутриклеточный IL-22.
Анти-IL-22 антитело 3F11 использовали в дополнительных экспериментах окрашивания клеток. 3F11 антитело конъгировали с Alexa 647, флуорофором фикоэритрином. Мышиный IgG2a, конъюгированный к Alexa 647, был использован в качестве контроля изотипа (Caltag). 293 клеточные линии, экспрессирующие hIL-22-GFP и GFP только были исследованы на связывание антитела 3F11. 293 клетки фиксировали 2% параформальдегидом в течение 30 минут, затем отмывали дважды PBS/2% FCS. Клетки ресуспендировали в 0,5% сапонине 15 минут. Нормальную мышиную сыворотку добавляли в другие 15 минут, затем добавляли антитела 0,5 мкг/миллион клеток на 30 минут. Клетки отмывали и ресуспендировали в FACS буфере и анализировали в FACScan. На фиг. 11 показано в нижней левой панели изменение в клетках внутри верхнего правого квадрата. Этот результат показывает, что конъюгированное 3F11 антитело связано с внутриклеточным IL-22.
ПРИМЕР 7: Экспрессия IL-22 в Th1 T-клетках
Когда CD4+ T-клетки созревают в тимусе и выходят в периферическую лимфатическую систему, они, как правило, сохраняют свой начальный фенотип до контактирования с антигенами, специфическими к их T-клеточному рецептору (TCR) (Sprent et al., Annu. Rev. Immunol. (2002); 20:551-79). Связывание TCR со специфическими антигенами, представленными антиген-представляющими клетками (APC), вызывает активацию T-клеток. В зависимости от окружения и цитокиновой стимуляции, CD4+ Т-клетки могут дифференцироваться на Th1 или Th2 фенотипы и стать эффекторными или клетками памяти (Sprent et al., Annu Rev Immunol. (2002); 20:551-79 и Murphy et al., Nat Rev Immunol. (2002) Dec;2(12):933-44). Этот процесс известен как первичная активация. Испытав первичную активацию, CD4+ T-клетки становятся эффекторными или клетками, и они сохраняют свой фенотип как Th1 или Th2. После того как эти клетки встретили вновь антиген, они подверглись вторичной активации, но это время ответа на антиген будет более быстрым, чем первичная активация, и приводит к продукции эффекторных цитокинов, как определено первичной активацией (Sprent et al., Annu Rev Immunol. (2002); 20:551-79 и Murphy et al., Annu Rev Immunol. 2000; 18:451-94). Было обнаружено в исследованиях во время первичной и вторичной активации CD4+ T-клеток, что экспрессия некоторых генов различна (Rogge et al., Nature Genetics. 25, 96-101 (2000) и Ouyang et al., Proc Natl Acad Sci USA. (1999) Mar 30;96(7):3888-93).
Для состояний первичной активации, нативные Т-клетки могут активироваться Ova и APC. Выделенная из клеток в этом состоянии РНК может предоставить информацию о том, что гены дифференциально регулируются во время первичной активации и что цитокины воздействуют на экспрессию генов во время Th1 и Th2 развития. После первичной активации, CD4+ Т-клетки могут сохраняться в культуре. Так как предыдущая активация и обработка цитокинами были зафиксированы в этих клетках, то они становятся либо эффекторными, либо клетками памяти. Во время этого периода, потому что нет никаких APC или антигенов, CD4+ Т-клетки входят в стадию отдыха. Эта стадия отдыха предоставляет информацию о различиях между нативными и клетками памяти, и отдыхающими клетками памяти Th1 и отдыхающими клетками памяти Th2. Отдыхающие клетки памяти Th1 и Th2 затем подвергаются вторичной активации анти-CD3/CD28 антителами или стимуляцией IL 12/IL 18 цитокинами. Эти состояния предоставляют информацию о различиях между активированными нативными и активированными Т-клетками памяти и различиях между активированными Th1 памяти и активированными Th2 памяти клетками.
Для эксперимента, показанного на фиг. F12, были выделены спленоциты DO11.10 мышей и активированы OVA в тех или иных Th1 состояниях: [IL-12 (1 нг/мл), IFN-γ, и IL-4 (1 мкг/мл)]; Th0 состояниях: [(анти-IL12, анти-IFN-γ, и анти-IL4)]; или Th2 состояниях: [(анти-IL-12 (0,5 мкг/мл), анти-IFN-γ, и IL-4 (5 нг/мл]). РНК выделяли спустя 48 часов (первичная стимуляция). Отдых клеток сохранялся в культуре до 7 дней, а затем они повторно стимулировались (вторичная стимуляция) OVA и облученными Balb/c спленоцитами. Субпопуляция клеток из Th1 состояния также стимулировалась лишь IL-12 и IL-18. Спустя 48 часов выделяли РНК. Экспрессию IL-22, IFN-γ и IL-4 в этих образцах РНК анализировали 5'-нуклеазным анализом (TaqManTM). Экспрессия была первично нормализована для образцов HPRT аутогенов, затем записанная в виде кратного увеличения по сравнению с уровнем экспрессии спленоцитов. Результат показан на фиг. 12, и данные показывают, что IL-22 высокоэкспрессируются в Th1 клетках после повторной стимуляции. Следовательно лечение анти-IL-22 возможно полезно для нацеливания этих клеток либо для лечения Th1 опосредованных нарушений, когда желательно очистить Th1 клетки из крови, или как диагностики Th1 опосредованных нарушений, когда подозревают важную роль IL-22.
ПРИМЕР 8: IL-22 продуцируется γδ Т-клетками
Для того чтобы анализировать экспрессию IL-22 в γδ Т-клетках, клетки были выделены из селезенки мыши, и γδ Т-клетки разделяли MACS сортером. GL4 представляет собой анти-γδ TCR антитело, которое специфически активирует γδ Т-клетки (Becton-Dickenson). Был использован набор для выделения РНК Qiagen MINI, чтобы выделить РНК из клеток для 5' нуклеазного анализа (TaqManTM). Master Mix one-step RT-ПЦР Master Mix Reagent (Applied Biosystems; 4309169) был использован, и аутогены RPL10 и SPF31 были использованы для нормализации. Были использованы целые спленоциты для определения относительного уровня экспрессии IL-22. На фиг. 13 показано, что IL-22 высокоэкспрессируется в γδ Т-клетках, стимулированных GL4 антителом.
ПРИМЕР 9: IL-22 продуцируется активированными человеческими Т-клетками
Микрочипы для нуклеиновых кислот используются для идентифицирования различно экспрессируемых генов в больных тканях по сравнению с их нормальными аналогами. Используя микрочипы для нуклеиновых кислот, исследуемые и контрольные образцы мРНК из исследуемых и контрольных образцов тканей подвергают обратной транскрипции и метят для получения проб кДНК. Пробы кДНК затем гибридизуют к чипам нуклеиновых кислот, иммобилизованных на твердом носителе. Чипы изменяют конформацию, так что известна последовательность и положение каждого члена чипа. Например, выбранные гены, для которых известно, что они экспрессируются при некотором состоянии заболевания, могут располагаться на твердом носителе. Гибридизация меченой пробы с конкретным членом группы показывает, что образец, из которого была получена проба, экспрессирует этот ген. Если сигнал гибридизации пробы из исследуемого образца (в этом примере, активированных CD4+ Т-клеток) больше сигнала гибридизации пробы из контрольного образца (в этом примере, нестимулированных CD4+ Т-клеток), идентифицируются ген или гены, гиперэкспрессированные в исследуемой ткани. Этот результат означает, что гиперэкспресссированный белок в исследуемой ткани используется не только как диагностический маркер для определения состояния болезни, но также как терапевтическая мишень для лечения состояния болезни.
Способ гибридизации нуклеиновых кислот и способ микрочипов хорошо известны в данной области. Например, специфический препарат нуклеиновых кислот для гибридизации и пробы, препараты и условия гибридизации подробно описаны в PCT патентной заявке серия № PCT/USO 1/10482, зарегистрированной 30 марта 2001 г., и которая включена в данное описание посредством ссылки.
В этом эксперименте, CD4+ Т-клетки были очищены от единичного донора, используя RossetteSepTM протокол из Stem Cell Technologies (Vancouver BC), в котором используют анти-CD8, анти-CD16, анти-CD19, анти-CD36 и анти-CD56 антитела, использованные для выделения CD4+ Т-клеток. Выделенные CD4+ Т-клетки активировали анти-CD3 антителом (использованное в концентрации, которая не стимулировала пролиферацию) вместе с либо ICAM-I, либо анти-CD28 антителом. Через 24 или 72 часа клетки собирали, выделяли РНК и запускали анализ на чипах микрочипов Affimax (Affymetrix Inc., Santa Clara, CA). Нестимулированные (отдыхающие) клетки собирали сразу после очистки и подвергали тому же самому анализу. Сравнивали гены, чья экспрессия увеличивалась в либо в одной, либо другой из двух временных точек в активированных по сравнению с отдыхающими клетками.
Результаты этого эксперимента показаны на фиг. 14. Результаты микрочипов подтверждают и одобряют данные в примере 7. Th1 Т-клетки продуцируют большое количество IL-22 при стимуляции в противоположность Th2 клеткам, которые продуцируют IL-4 или IL-5. Этот результат, вероятно, позволит разделить Th1 и Th2 связанные иммунные нарушения, основанные на цитокиновом профиле. Th1 клетки, экспрессирующие IL-22 и IFN-γ, можно лечить терапией, направленной на эти цитокины, без влияния на популяцию клеток Th2.
ПРИМЕР 10: Th1 клетки экспрессируют внутриклеточный IL-22
Чтобы определить уровень экспрессии IL-22 в Т-клетках посредством FACS, проводили внутриклеточное окрашивание мышиных Th1/Th2 клеток. Первичные спленоциты были разделены на Th1 или Th2. Для FACS окрашивания, 1 миллион клеток был посеян на лунку 96-луночного планшета, и были обработаны PMA/Иономицином в течение 2 часов, затем для других Brefeldin A на 2 часа. Использованные антитела представляли собой античеловеческие IL-22 (антитело 3F11.1) и анти-gp120 в качестве контроля. Антимышиный IFN-γ-FITC и антимышиный IL-4-PE были получены из BD Bioscience (San Diego CA). PE-конъюгированные козлиные антимышиные IgG (также из BD Bioscience) были использованы в качестве вторичного антитела. Клетки фиксировали 2% параформальдегидом в течение 30 минут, затем отмывали дважды PBS/2% FCS. Клетки ресуспендировали в 0,5% сапонине в течение 15 минут, затем добавляли антитела 0,5 мкг/миллион клеток на 30 минут. Клетки затем отмывали дважды и добавляли вторичные антитела в 0,5% сапонине на 15 минут. Наконец, клетки отмывали и ресуспендировали в FACS буфере и анализировали на FACScan. На верхних панелях фиг. 15 показано, что Th1 клетки можно отличить от Th2 клеток. Th1 клетки являются положительными для IFN-γ, отрицательными для IL4 и положительными для IL-22. Th2 клетки в большей степени негативны для IFN-γ, положительны для IL4 и негативны для IL-22.
ПРИМЕР 11: Получение анти-IL-22 рецептора (IL-22R)
Чтобы исследовать связывание анти-IL-22R антител, используют 293 клетки, экспрессирующие ML-22R, и клетки, экспрессирующие GFP. Один миллион клеток окрашивали различными анти-hIL-22R антителами в концентрации 0,3 мкг/миллион клеток. Исследуемые антитела представляли собой 7E9, 8A12, 8H11 и 12H5. Вторичное антитело было козлиным антимышиным PE конъюгированным (Jackson Labs), использованный коэффициент разбавления 1:200. Клетки отмывали и окрашивали в FACS буфере (0,5% BSA/PBS). Окрашивание исследуемыми антителами проводили в течение 15 минут при 4°C, затем клетки отмывали и добавили вторичное антитело на 15 минут при 4°C. Клетки отмывали дважды перед анализом в FACScan. Результаты представлены на фиг. 16. Для каждого графика, в котором пики не перекрываются, пик слева соответствуют контролю, а пики справа соответствуют исследуемому антителу. На фиг. 16 показано, что все четыре исследуемые анти-IL-22R антитела были положительны для связывания IL-22R в транфицированных 293 клетках. Антитела 7E9, 8A12, 8H11 и 12H5 хорошо связываются с очень небольшим фоном.
ПРИМЕР 12: IL-22R блокирующие антитела
Для исследования блокирующей активности анти-IL-22R антител, был использован люциферазный репортерный конструкт (как описано в примере 2). Если антитело блокирует активность, STAT3 не будет активироваться, и люциферазный сигнал будет низким. Клетки, экспрессирующие hIL-22R/hIL10Rb, были рассажены в 0,2×106/лунку 24-луночного планшета, и люциферазные репортеры TK-SIE-SRE-S (0,8 мкг/лунку) и RL-TK-Luc (0,16 мкг/лунку) были трансфицированы в клетки. На следующий день, был добавлен hIL-22 0,5 нМ в лунки, и каждое антитело было добавлено 20 мкг/мл. Исследованные анти-IL-22R антитела были 7E9, 8A12, 8H11 и 12H5. Использованные контрольные антитела были GP120 и 11H4, анти-hIL-22 антитело показало блокирующую активность в примере 2. Шестнадцатью часами позже клетки лизировали и образцы оценивали на люминометре для определения люциферазной активности. На фиг. 17 показано, что все четыре исследованных анти-IL-22R антитела блокируют IL-22R-IL-22 взаимодействие.
ПРИМЕР 13: IL-22R экспрессируется в первичных кератиноцитах
Кератиноциты представляют собой клеточную популяцию, которая гиперпролиферирует во время псориаза. Терапия мишеней кератиноцитов используется для ослабления псориаза. Экспрессию IL-22R на первичных человеческих кератиноцитах определяли FACS анализом. Нормальные человеческие эпидермальные кератиноциты (NHEK) донора 0526 были получены из Cascade Biologies, пассаж #2, выращенные до 80% плотности, и были окрашены 300-600K клеток на образец. Анти-IL-22R сыворотку использовали в разведении 1:50 и предвыделенную сыворотку использовали в разведении 1:50 как контроль. Для IL10R2 окрашивания, антитело из R&D (клон #90220, мышиный IgG1) использовали 0,3 мкг на образец с мышиным IgG1-PE контрольным изотипом (BD Pharmingen #33815X). Вторичное антитело для анти-IL-22R сыворотки было крысиным анти-мышиным IgG1-PE (BD Pharmingen #550083), использованным 0,1 мкг на образец. На фиг. 18 показано, что IL-22R и IL10R2 экспрессируется на NHEK. Следовательно, блокирование IL-22R или IL-22 может быть полезным для ослабления нарушений, связанных с гиперпролиферацией кератиноцитов, таких как псориазы.
ПРИМЕР 14: Эффект IL-22 на эпидермальные культуры
Восстановленный человеческий эпидермис (RHE) можно использовать в качестве модели для действия цитокинов на кожу. RHE и культуральная среда были получены из MatTek Corporation (Ashland, MA). RHE уравновешивали всю ночь (20-22 часа) с 0,9 мл среды при 37°C, 5% CO2, для удаления нагрузки до начала эксперимента и затем культивировали в области контакта воздух/жидкость в 5 мл среды при 37°C, 5% CO2. Эффект IL-22 на RHE исследовали с использованием трех различных состояний. IL-22 (1,2 нМ) или эпидермальный фактор роста (EGF-R&D системы) (1 нМ) добавили в среду. Контроль состоит из необработанной среды. RHE культивировали 4 дня, со смены среды каждые два дня, добавляя свежий EGF или IL-22. RHE снимали, фиксировали в 10% нейтральном буферном формалине (NBF) всю ночь, разделяли и окрашивали с гематоксилином и эозином (H&E). На фиг. 19 показано, что обработка IL-22 вызывает истончение эпидермиса. Это показывает, что IL-22 вызывает гиперплазию, или пролиферацию клеток, которые формирую эпидермис.
Если эти части были окрашены на кератин 16 (K16), маркера пролиферации кератиноцитов, RHE обработанные IL-22 показали более яркое окрашивание к K16. K16 экспрессируются только в пролиферирующих клетках кожи, таких как псориаз и заживление ран (рассмотренные в Freedberg et al, Soc. Invest. Derm. 116:633-640 (2001)). На фиг. 20 показано K16 окрашивание в обработанных IL-22 RHE относительно необработанных RHE и обработанных EGF. IL-22 отработанные RHE показали K16 по всей ткани, поскольку окрашивание локализуется в необработанных и EGF обработанных частях.
Обработка RHE IL-22 также стимулирует псориазин, гиперэкспрессируемый ген при псориазе. Псориазин (S100A7) первоначально открыли как белок, экспрессируемый при псориазе, но не в нормальной коже (Madsen P., et al., J. Invest. Derm. 97: 701-712 (1991)). Псориазин экспрессируется в активированных культивированных и злокачественных кератиноцитах и в злокачественных эпителиальных клетках рака груди (Watson et al., Int. J. of Biochem. and Cell Bio. 30:567-571 (1998)). Текущие данные подтверждают роль псориазина в воспалительной болезни кожи, хемотаксисе и прогрессии опухоли груди. Корреляция псориазина с псориазоподобной гиперплазией кожи предполагает роли дифференцировки в кератиноцитах. Псориазин может также быть хемотаксическим, стимулирующим инфильтрацию нейтрофилами и CD4+ T-лимфоцитами эпидермиса, который является отличительным признаком псориазина. На фиг. 21 показано, что обработка RHE IL-22 вызывает высокие уровни экспрессии псориазина. Этот результат подтвердил, что IL-22 и IL-22R играют роль при псориазе.
Индуцирующий эффект IL-22 пути на псориазин может блокироваться антителами, направленными IL-22 или IL-22R. Анти-IL-22 антитело 8E11, введенное в концентрации 20 мкг/мл, уменьшало экспрессию псориазина до неопределяемых уровней (см., фиг. 23). Когда используется в концентрации 20 мкг/мл, анти-IL-22R антитело (7E9) также значительно уменьшало экспрессию псориазина, как показано на фиг. 23.
Анти-IL-22 и анти-IL-22R антитела были исследованы для определения, могли ли они уменьшать эпидермальное истончение, наблюдаемое при обработке RHE IL-22. Анти-IL-22 антитело (8E11), введенное при концентрации 20 мкг/мл, показало значительное уменьшение эпидермального истончения (см. фиг.24). RHE, обработанные IL-22, достигают толщины 80-90 мкм, и обработка анти-IL-22(8E11) антителами уменьшает RHE толщину до 50-60 мкм (фиг. 25). Анти-IL-22R антитело (7E9) также уменьшает толщину кожи. При использовании концентрации 20 мкг/мл, анти-IL-22R антитела уменьшают RHE толщину от 80-90 мкм до 55-60 мкм (фиг. 25). Эти данные показывают, что анти-IL-22 или анти-IL-22R антитела могут облегчать симптомы, связанные с псориазом, такие как эпидермальная пролиферация и толщина.
ПРИМЕР 15: Применение микрочипов генов, индуцированных IL-22
Чтобы определить, что гены стимулировались IL-22, нормальные человеческие эпидермальные кератиноциты (NHEK) получали из единичного донора, рассеивали и обрабатывали при 70% плотности в течение 24 часов 20 нг/мл IL-22. Среду и добавки (EpiLifeR+HKGS) получали из Cascade BiologicsTM (Portland, OR). Клетки отмывали и лизировали. Общую РНК очищали из NHEK клеток с использованием Qiagen RNeasy Mini Kit. РНК подвергали микрочипному анализу и определяли количественно экспрессию генов (см. вышеописанный пример 9 для описания микрочипового анализа).
Псориазин стимулируется 81 кратно индукцией IL-22. SPR-2G увеличивается 11 кратно (см. фиг. 22.). Эти результаты показывают, что IL-22 путь участвует в псориазе. Следовательно, антагонисты и антагонисты антитела, направленные против IL-22 или IL-22R, используют для облегчения псориаза.
ПРИМЕР 16: IL-23 вызывает отличительные признаки псориаза in vivo
Мышиная модель была использована для сравнения способности IL-12 и IL-23 вызывать псориазные кожные признаки. C57B1/6 мышам вводили подкожно в ухо 500 нг либо рекомбинантный IL-12, либо рекомбинантный IL-23 в общем объеме 20 мкл PBS. Контрольным мышам вводили 20 мкл PBS только. Мышам вводили однократно каждые 2 дня в течение 16 дней. Каждая экспериментальная группа состояла из пяти мышей. Толщину уха измеряли до и на множестве временных точек после введения штангенциркулем (Mitutoyo America Corporation) и сообщали в виде среднего +/- стандартного отклонения. Для этого эксперимента и последующих экспериментов, статистическую значимость считали одним способом или двумя способами ANOVA, используя Prism software (GraphPad). Все p значения <0,05 были оценены как значимые. Уши мышей собирали стандартным гистологическим анализом с использованием гематоксилин-и-эозин (H&E) окрашиванием.
Как показано на фиг. 26A, и IL-12, и IL-23 введение вызывает значительное увеличение толщины ушей в течение одной недели после первого введения. Для мышей, сохранивших IL-12, p был <0,001 (дни 12, 14 и 16 против PBS контроля, соответственно). Для мышей, сохранивших IL-23, p был <0,001 (дни 8, 12, 14 и 16 против PBS контроля, соответственно). Гистологический анализ показал, что и у IL-12 и у IL-23 введенных на ушах развиваются заметная воспалительная клеточная инфильтрация и эпидермальное истончение (акантоз) по сравнению с обработанной PBS контрольной группой; однако существовало несколько точных гистологических отличий между этими двумя группами. Первая, IL-12 вызывал умеренное изменение акантоза с заметной, предоминантно мононуклеарной дермальной воспалительной клеточной инфильтрацией (фиг. 26 D, E) по сравнению с PBS контрольной группой (фиг. 26B, C), поскольку IL-23 вызывал заметный акантоз со смешанной дермальной воспалительной клеточной инфильтрацией многими полиморфонуклеарными лейкоцитами (фиг. 26F, G), включая и нейтрофилы (arrows) и эозинофилы. Эпидермальная гиперплазия и присутствие полиморфонуклеарных лейкоцитов являются гистологическими отличительными признаками псориаза у человека, а также самые простые гистологические обнаружения на мышиной модели псориаза. См. P. C. van de Kerkhof et al, Dermatologica 174: 224 (1987) и P. R. Mangan et al., Nature (2006) 441:235.
ПРИМЕР 17: IL-22 действует после IL-23 in vivo
Чтобы идентифицировать цитокины, которые потенциально подавляют IL-12 или IL-23, использовали ПЦР в реальном времени для определения экспрессии панели цитокинов из образцов кожи каждого уха, в которые вводили IL-12 или IL-23. Инъекции в кожу ушей и гистологический анализ проводили, как описано в предыдущем примере. На 8 день эксперимента, РНК выделяли из каждого уха мыши и проводили ПЦР в реальном времени для количественного определения уровней мРНК, кодирующей IFN-γ, IL-17 и IL-22. Специфически, РНК выделили RNeasy Mini Kit (Qiagen, Valencia, CA) согласно инструкциям производителя. RT-ПЦР в реальном времени проводили с использованием ABI 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA) с праймерами и пробами с использованием TaqManTM One-Step RT-PCR Master Mix reagents (Applied Biosystems). Реакцию проводили дважды и образцы были нормализованы по контрольному гену домашнего хозяйства RPL-19 и сообщали согласно ∆∆Ct способу.
Как показано на фиг. 27A, IL-12 вызывал значительное увеличение IFN-γ экспрессии в ухе спустя восемь дней после первой инъекции. IL-23 вызывали продукцию IL-17 и ингибировали IFN-γ продукцию относительно PBS-обработанной контрольной группы (фиг. 27A). Интересно, IL-22 также был значительно увеличен после инъекции IL-23, но не после инъекции IL-12 (фиг. 27A). Эти данные предположили связь между IL-23 и IL-22.
Чтобы подтвердить, что цитокины продуцировались лимфоцитами, которые инфильтрировали уши, лимфоциты были элюированы из обработанных ушей, и продукция цитокинов была измерена после активации посредством ELISA. В соответствии с данными RT-ПЦР в реальном времени, клетки из IL-23 инъецированных ушей предпочтительно продуцировали IL-22 и IL-17, хотя клетки из IL-12 инъецированных ушей секретировали большое количество IFN-γ (фиг. 28).
ПРИМЕР 18: IL-22 вызывает дермальное воспаление и эпидермальную гиперплазию in vivo
Чтобы определить, является ли IL-22, подобно IL-23, способным вызывать псориатические кожные признаки in vivo, мышей инъецировали подкожно в уши IL-22 или PBS только, как описано выше в примере 16. Как показано на фиг. 27B, IL-22 вызывал значительное увеличение толщины ушей относительно PBS обработанной группы. IL-20, другой цитокин из IL-10 семейства, вызывает только очень умеренное и локализованное увеличение толщины ушей. Это открытие противоречило предыдущему сообщению, где эпидермальная трансгенная гиперэкспрессия IL-20 вызывала заметную эпидермальную гиперплазию, результат которой предположил, что IL-20 может потенциально играть роль в эпидермальной функции и при псориазе. См. Blumberg et al, Cell 104:9 (2001). Гистологический анализ показал, что обработанные IL-22 уши мышей имеют сходный гистологический вид с ушами обработанной IL-23 группы, показанной на фиг. 26F и G, проявляя заметный акантоз и смешанную дермальную воспалительную клеточную инфильтрацию (фиг. 27G, H), включая многие нейтрофилы (arrows) и некоторые эозинофилы. В отличие от этого, обработанные IL-20 уши имели только умеренный очаговый акантоз с умеренным и сильно очаговое смешанное воспаление (фиг. 27D, E) относительно PBS обработанной группы (фиг. 27C, F). В этих данные предположили, что IL-22 имеет важное значение для IL-23-индуцированного кожного воспаления и акантоза.
ПРИМЕР 19: Анти-IL-22 блокирующее антитело значительно уменьшало IL-23-индуцированный акантоз
Чтобы подтвердить, что IL-23 действует через IL-22, чтобы вызвать псориатические кожные признаки, было исследовано действие анти-IL-22 моноклонального антитела 8E11 на IL-23 индуцированное дермальное воспаление и акантоз. Мышам вводили подкожно в уши IL-23 или PBS, как описано выше (пример 16), за исключением того, что инъекции производили через 14 дней. Мыши также были инъецированы внутрибрюшинно 8E11 или контрольным моноклональным антителом IgG1 изотипа в концентрации 200 мкг на мышь и частоте раз в каждые два дня в течении 14 дней. На 14 день, собирали уши мышей для гистологического анализа, используя H&E окрашивание.
Как показано на фиг. 29A, 8E11 ("анти-IL-22 mAb") значительно уменьшало IL-23-индуцированный эпидермальный акантоз (*p<0,001) относительно обработки контрольным IgG1 антителом. (Сравни также фигуры 29D и E (анти-IL-2 mAb) с B и C (контрольный IgG1)). Далее, мыши, обработанные анти-IL-22 mAb, также показали умеренное снижение дермального воспаления. Однако мыши, обработанные анти-IL-22 mAb, по-прежнему показали умеренную воспалительную клеточную инфильтрацию,по сравнению с кожей ушей, обработанной PBS. (Ср. фиг. 29D и E (анти-IL-22 mAb) с F и G (PBS)).
ПРИМЕР 20: IL-23-индуцированный акантоз значительно уменьшался у IL-22 дефицитных мышей
Чтобы далее подтвердить, что IL-23 действует через IL-22, вызывая псориатические кожные признаки, действие IL-23 и на дикого типа, и на IL-22 дефицитных мышей было исследовано. IL-22 дефицитные мыши (например, гомозиготные IL-22 нокаутные мыши, называемые "IL-22'' мыши") были получены разрушением гена-мишени согласно стратегии, изображенной на фиг. 30A. Экзоны 1-4 (закрытые боксы) IL-22 кодирующей последовательности были заменены кассетой резистентностью к неомицину, фланкированной loxP сайтами. Гетерозиготные мыши, несущие зависящий аллель, были скрещены с трансгенной линией, в которой протамин 1 (Prm) промотор запускал Cre рекомбиназу. Зависимый аллель был вырезан во время сперматогенеза в смеси гетерозиготных самцов (то есть гетерозиготы для зависимого аллеля и PrmCre трангена). Смеси гетерозиготных самцов были скрещены с самками дикого типа, и полученное в результате потомство было скринировано по скрещиваемому аллелю и снижению PrmCre трансгена. Потомки были обратно скрещены с C57B1/6 фоном для, по крайней мере, шести генераций. Мышиные генотипы были подтверждены с помощью ПЦР, использующей праймеры, показанные на фиг. 30B.
IL-22 экспрессия была исследована на уровнях мРНК и белка в Th клетках дикого типа и IL-22-/- мышей. IL-22 мРНК экспрессия была исследована в Th1, Th2 и ThIL-17 клетках дикого типа ("+/+") и IL-22-/- ("-/-") мышах (фиг. 30C), используя RT-ПЦР, подтверждающую, что IL-22 мРНК не обнаруживается у IL-22-/- мышей. Экспрессия IL-22, IL-17, IFN-γ и IL-4 была исследована в Th1, Th2 и ThIL-17 клетки из дикого типа ("WT") и IL-22-/- ("KO") мыши, используя ELISA. Результаты показаны на фиг. 30D для каждого IL-22, IL-17, IFN-γ и IL-4, как показано на вершине каждого графика, закрашенными полосами и открытыми полосами, показывающими уровни экспрессии у WT и KO мышей, соответственно. Дополнительно, CD4 Т-клетки из IL-22-/- мыши были способны активироваться и дифференцироваться во всех субпопуляциях Т-хелперов и способны продуцировать нормальный уровень IL-17, IFN-γ и IL-4 относительно дикого типа CD4 Т-клеток. Как предполагалось, однако, IL-22 отсутствовал в IL-22-/- CD4 Т-клетках. Были получены IL-22-/- мыши, чтобы развивались нормально и имели аналогичные составы лимфоцитов во всех главных лимфоидных органах, рассмотренные по отношению к мышам дикого типа (данные не представлены).
IL-22-/- мышам и детенышам одного помета дикого типа вводили подкожные инъекции в уши IL-23 или PBS, как описано выше (пример 16). На 16 день, уши мышей анализировали стандартным гистологическим анализом. Как показано на фиг. 31A и B, IL-23 вызывал значительно меньшее ушное истончение и эпидермальное истончение у IL-22-/- мышей по сравнению с контрольными группами. (IL-22-/- мыши названы на этой фиг. и фиг. 32 "KO" или "IL-22 KO"; мыши дикого типа названы на этой фиг. и фиг. 32 "WT" или "IL-22 WT.") При гистологическом окрашивании, и эпидермальный акантоз, и дермальное воспаление были значительно уменьшены у IL-22-/- мышей (фиг. 31E и F, соответственно) по сравнению с IL-23-обработанными дикого типа детенышами одного помета (фиг. 31C и D, соответственно). В отличие от этих результатов, IL-22 дефицит не влияет на IL-12 индуцированное кожное ушное воспаление во всех (фиг. 32). Следовательно, данные показывают, что IL-22 играет значительную роль в дермальном воспалении, и эпидермальный акантоз вызывается IL-23, а не IL-12.
ПРИМЕР 21: IL-23 вызывает IL-22 продукцию в различных IL-23-активированных лимфоцитах
Затем исследована способность IL-23 индуцировать IL-22, различные популяции лимфоцитов были выделены и стимулированы in vitro в условиях, показанных на фиг. 33. Проводили ELISA для определения IL-22 в культуральных супернатантах и сообщали о результатах на фиг. 33 A в виде среднее +/- стандартное отклонение. Способность IL-23 индуцировать IL-10 семейство цитокинов, а не IL-22, также была исследована. Спленоциты из DO 11.10 TCR трансгенной мыши стимулировали 0,3 мкМ OVA пептидом в обозначенных условиях поляризации T-хелперных клеток в течение 4 дней, затем делали перерыв на два дня и повторно стимулировали связанный с пластиной анти-CD3 (10 мкг/мл) и растворимый анти-CD28 (5 мкг/мл) в течение других 2 дней. Проводили RT-ПЦР в реальном времени на РНК, выделенной из клеток в указанных условиях для количественного определения экспрессии мышиных IL-19, IL-20 и IL-24. РНК из нормальных мышиных спленоцитов также включалась как контроль. Как показано на фиг. 33B, IL-23 не вызывал экспрессию любого другого исследуемого IL-10 семейства цитокинов.
ПРИМЕР 22: IL-22 представляет собой новый эффекторный цитокин из линии ThIL-17
Недавно, IL-23 был связан с развитием новой IL-17 продуцирующей эффектор CD4+ Т-клеточной линии (ThIL-17). L. E. Harrington., Nat. Immunol 6:1123 (2005); H. Park., Nat. Immunol. 6:1133 (2005). IL-23 способен индуцировать ThIL-17 клетки из нативных CD4+ Т-клеток в присутствии APC и антигена, но не способен вызывать IL-17 продукцию, когда использовали нативно очищенные Т-клетки, активированные анти-CD3/анти-CD28. L. E. Harrington et al., Nat. Immunol 6: 1123 (2005); M. Veldhoen et al, Immunity 24: 179 (2006). Более того, предполагали, что TGF-β и IL-6 представляют de novo факторы для дифференцировки ThIL-17 субпопуляций. M. Veldhoen et al., Immunity 24:179 (2006).
Эксперименты можно проводить для исследования, может ли IL-22 быть дополнительным эффектором Т-клеток цитокина, вызванного IL-23, при дополнительной стимуляции TCR. CD4+ Т-клетки из DO 11.10 TCR трансгенной мыши активировали 0.3 мкМ OVA пептидом в течение четырех дней при условиях Th1-поляризации (IL-12 и анти-IL-4), Th2-поляризации (IL-4, анти-IL-12 и анти-IFN-γ), ThIL-17-поляризации (IL-23, анти-IFN-γ и анти-IL-4) или Th0 (анти-IL 12/23 p40, анти-IFN-γ и анти-IL-4), как ранее описано. L. E. Harrington et al, Nat Immunol 6:1123 (2005). РНК выделяли из клеток и проводили ПЦР в реальном времени для определения экспрессии мРНК, кодирующей различные мышиные цитокины (показанные выше графики на фиг. 34A). Дополнительно, проводили ELISA на культуральных супернатантах для определения экспрессии различных цитокинов на уровне белка. Как показано на фиг. 34A, IL-17 индуцировался IL-23, тогда как IFN-γ и IL-4 продуцировались Th1 и Th2 клетками соответственно. IL-22 был получен и на уровне мРНК, и на уровне белка из IL-17 продуцирующих ThIL-17 клеток.
Чтобы определить, является ли IL-22 новым эфекторным цитокином из полностью коммитированной ThIL-17 линии, поляризованные Т-клетки, как описано выше, отдыхали два дня и затем повторно стимулировались в течение двух дней связанным с пластиной анти-CD3 (10 мкг/мл) и растворимым анти-CD28 (5 мкг/мл) в отсутствие или присутствии IL-23. Проводили ELISA для определения экспрессии мышиных цитокинов, показанные выше графики на фиг. 34B. Результаты показывают, что IL-17 специфически продуцируется ThIL-17 субпопуляцией, даже в отсутствие IL-23, и IL-23 увеличивал IL-17 продукцию. IL-23 не активирует IL-17 продукцию коммитированными Th1 и Th2 клетками. IL-22 показал идентичную модель экспрессии как IL-17, показывающий, что IL-22 действительно представлял собой эффекторный цитокин, экспрессированный такой новой ThIL-17 субпопуляцией.
Предварительно сообщали, что IL-23 рецептор может экспрессироваться на активированных Т-клетках памяти. C. Parham et al, J Immunol 168:5699 (2002). Вышеописанные эксперименты не исключают возможность, что IL-23 действовали на Т-клетки памяти для продукции IL-22. Чтобы отнестись к этому более критично, вышеописанные эксперименты были повторены с использованием нативных CD4+ Т-клеток, выделенных из DO 11.10 TCR трансгенных мышей. Специфически, CD4+ Т-клетки из Rag2-/- DO 11.10. TCR-трансгенных мышей стимулировали OVA пептид-импульсные BALB/c клетки каналов селезенки (облученные, обедненные Т-клетки) в течение 72 часов в ThI-поляризующих состояниях (IL-12 и анти-IL-4), Th2-поляризующих состояниях (IL-4, анти-IL-12 и анти-IFN-γ), ThIL-17-поляризующих (IL-23, анти-IFN-γ и анти-IL-4) или других состояниях, как показано на фиг. 35A. Как показано на этом чертеже, ThIL-17 клетки продуцировали самые высокие уровни IL-22, хотя Th1 также секретировал определенные уровни IL-22. Далее, добавление либо IFN-γ, либо IL-4 полностью прекращало продукцию IL-17; однако эти два цитокина только умеренно подавляли продукцию IL-22 (фиг. 35A). Эти данные предполагают потенциально различные пути индукции экспрессии IL-17 против IL-22. Однако более полно показанные ThIL-17 клетки продуцировали и IL-17, и IL-22 после повторной стимуляции в течение 48 часов в указанных вторичных состояниях (фиг. 35B). IL-23 далее увеличивал уровни этих цитокинов в способе, который не может блокироваться ни IFN-γ, ни IL-4 (фиг. 35B). Эти данные подтверждают стабильность этой ThIL-17 линии.
Чтобы далее исследовать продуцируются ли IL-17 и IL-22 теми же клетками во время активации, ThIL-17 клетки были стимулированы PMA и иономицином, и антитела к IL-22 или IL-17 были использованы для внутриклеточного окрашивания. Как показано на фиг. 35C, IL-17-продуцирующие клетки главным образом получались на ThIL-17 оси (левая панель). IL-22-продуцирующие клетки также предпочтительно обнаружили из линии ThIL-17 (правая панель). Содержание и IL-22, и IL-17 выявили, что значительная часть клеток из ThIL-17 линии продуцирует и IL-22, и IL-17 одновременно, показывая, что IL-22 и IL-17 продуцируются теми же самыми клетками.
Как обсуждалось выше, недавние исследования также показали, что другие факторы из APC могут быть первичной движущей силой вслед за дифференцировкой IL-17-продуцирующих Т-клеток из нативных CD4+ Т-клеток, поскольку IL-23 не могут индуцировать de novo IL-17 продукцию очищенных нативных CD4 Т-клеток. M. Veldhoen et al., Immunity 24: 179 (2006). Два из таких факторов, критических для продукции IL-17, из нативных CD4 Т-клеток были идентифицированы как TGF-β и IL-6. Id. Чтобы определить, являлись ли эти факторы также критическими для продукции IL-22 у мышей, очищенные нативные CD4 Т-клетки (>98%) стимулировали связанными с пластинами анти-CD3 (10 мкг/мл) и растворимыми анти-CD28 (5 мкг/мл). В соответствии с публикованными данными, TGF-β и IL-6, а не IL-23 вызывали продукцию IL-17 (фиг. 36A, правая панель). Удивительно, в отличие от индукции IL-17, IL-22 мог индуцироваться только в присутствии IL-23 и не может индуцироваться TGF-β и IL-6 (фиг. 36A, левая панель). Эти данные предполагают, что транскрипция IL-17 и IL-22 может регулироваться по-разному. Однако, как ранее сообщалось, TGF-β и IL-6 не могут сохранять долговременно IL-17 продуцирующих линией Т-клеток без IL-23 (фиг. 36B). Таким образом данные показывают, что IL-23 может быть одним из главных факторов, стимулирующих продукцию IL-22 сублинией Т-клеток.
Далее мы исследовали, какая сходная IL-22 продуцирующая линия Т-клеток может сохраняться из человеческих CD4 Т-клеток. Мы обнаружили, что IL-23 может вызывать секрецию IL-22 из очищенных нативных человеческих CD4+ Т-клеток, стимулированных анти-CD3/анти-CD28 при ThIL-17-поляризующих состояниях (фиг. 36C, левая панель). Эти клетки могут продуцировать IL-22 вновь при повторной стимуляции без добавления экзогенного IL-23 (фиг. 36C, правая панель), показывая образование стабильной Т-клеточной линии. Хотя эти клетки культивировали в сходных условиях, таких как вышеописанные исследования на мышах, мы не смогли определить продукцию IL-17 в связи с ограниченностью (данные не показаны).
В заключение, получены данные для первого времени, что IL-23 может индуцировать IL-22-продуцирующая Т-клеточная сублиния из мышиных и человеческих нативных CD4 Т-клеток. Продукция IL-17 этой линии зависит от других факторов окружения. Между тем аутентичный антиген и APC стимулирующие условия, IL-23 заводит Т-клеточную сублинию, продуцирующую и IL-22, и IL-17. IL-23 также стимулируется IL-22 продукцией, если нативные Т-клетки стимулировались анти-CD3 и анти-CD28. TGF-β и IL-6 могут индуцировать временную продукцию IL-17 из нативных Т-клеток, но недолговременно коммитируют, не заводя продукцию IL-22.
ПРИМЕР 23: IL-19, IL-20 и IL-24 также вызывают эпидермальное истончение
IL-22 принадлежит к семейству цитокинов, которые включают в себя IL-19, IL-20 и IL-24, все из которых показывают увеличенную экспрессию в псориатической коже. Такие цитокины также были исследованы, чтобы определить, являются ли они, подобные IL-22, способными индуцировать эпидермальную гиперплазию и акантоз. RHE культивировали в течение четырех дней и обрабатывали 20 нг/мл IL-19, IL20, IL-22 или IL-24 или 6 нг/мл EGF. Обработанные RHE окрашивали H&E. Результаты показаны на фиг. 37A. Все цитокины вызывали аконтоз жизнеспособного содержащего ядра эпидермиса, обозначенного увеличенной длиной двойных стрелок. В соответствии с предыдущими наблюдениями (выше), IL-22 вызывает гипогранулез или уменьшение в гранулярном клеточном слое (стрелка-указатель), а также гиалинизацию ниже основания (звездочки). IL-22 также вызывал парокиратоз в RHE, культивированном в течение 7 дней (данные не приведены). Гипогранулез и паракиратоз представляют собой часто встречающиеся гистологические признаки псориаза. IL-19, IL-22 и IL-24 вызывали только эпидермальный акантоз с небольшим или незначительным действием на либо гранулярный клеточный слой, или на основание. EGF вызывал эпидермальный акантоз с гипергранулезом и уплотнением кератиноцитов в stratum granulosum (стрелка). Эпидермальное истончение, вызванное IL-19, IL-20, IL-22 или IL-24, было определено количественно в независимом эксперименте и представлено графически на фиг. 38. IL-22 обладал наибольшим действием. Воспалительные цитокины TNF-α, IFN-γ и IL-1β, которые, как полагают, играют роль при псориазе, не стимулируют пролиферацию кератиноцитов в этой RHE системе (данные не приведены). Таким образом, такие цитокины могут играть вторичную роль при псориазе или могут играть роль через зависимый путь IL-19, IL-20, IL-22 и/или IL-24.
Использовали иммуногистохимию для определения цитокератина 16 (CK16), маркера эпидермальной гиперплазии. IL-24, IL-22 и EGF индуцировали CK16 экспрессию во всем неороговевающем эпидермисе, хотя только IL-19 и IL-20 индуцировали CK16 экспрессию в базальной зоне (фиг. 37B).
Иммуногистохимию также использовали для определения псориазина (S100A7), одного из нескольких белков семейства S100, увеличенного при некоторых гиперпролиферативных и воспалительных кожных состояниях, включая псориаз. IL-19, IL-20, IL-22 и IL-24 все индуцировали S100A7 экспрессию в супрабазальном эпидермисе, IL-22 и IL-24 обладающие наибольшим действием (фиг. 37C). S100A7 окрашивание получили в ядре и цитоплазме кератиноцитов, по-видимому, также с некоторым белком, являющимся внеклеточным. Результаты, показанные на фиг. 37B и C, были количественно определены и изображены графически на фиг. 37E и F.
Иммуногистохимию также использовали для определения pY(705)-STAT3, трансактивирующей формы STAT3. Было показано, что активированный STAT3 увеличен в коже, поврежденной псориазом. IL-19, IL-20, IL-22 и IL-24 все индуцировали постоянную активацию STAT3 в RHE кератиноцитах, обнаруженных во всех слоях живых клеток, показанных их ядерной локализацией (фиг. 37D).
ПРИМЕР 24: Блокирование антителами рецепторов для IL-20 и IL-22 уменьшают экспрессию псориазина
И IL-19, и IL-20 передают сигнал посредством гетеродимера рецептора IL20Ra и IL20Rb. IL-20 также передает сигнал посредством гетеродимера рецептора IL-22R и IL-20Rb. IL-22 передает сигнал посредством гетеродимера IL-22R и IL10R2. Экспрессия этих компонентов рецептора на поверхности кератиноцитов, выделенных из RHE или первичной культуры нормальных человеческих эпидермальных кератиноцитов (NHEK, из донорской неонатальной крайней плоти), была исследована проточной цитометрией. Для проточной цитометрии были использованы следующие мноклональные антитела: анти-IL20Ra (полученные из мышей для целей этого исследования); анти-IL20Rb (полученные из мышей для целей этого исследования); анти-IL-22R антитело 7E9 (описанные выше); и анти-IL-10R2 FAB874P (PE-конъюгированные) (R&D Systems, Minneapolis, MN). Результаты показаны на фиг. 39. Компонент рецептора, с которым связывается антитело, показан в верхнем правом углу каждого графика (IL-22R обозначается как "IL-22R1"). IL-20Rb и IL10R2 постоянно экспрессировались на поверхности NHEK, несмотря на вырождение, количества пассажей или уровней кальция в среде (фиг. 39A.). В отличие от этого, экспрессия и IL-20Ra, и IL-22R1 на поверхности NHEK варьировала от донора к донору и была постоянно низкого, но определяемого уровня (фиг. 39A и данные не приведены). По сравнению с уровнями экспрессии в монослое NHEK, IL-20Ra и IL-22R экспрессировались в намного более высоких уровнях в кератиноцитах, выделенных из RHE (фиг. 39B). Причины такого различия неизвестны. Однако все же очевидно, что все анализированные рецепторные компоненты экспрессируются в человеческих кератиноцитах. Экспрессия этих рецепторных компонентов на иммунных клетках (T клетки, B клетки, естественные киллеры и моноциты) не определялась (данные не приведены). Таким образом, видимо, лиганды для этих рецепторных компонентов обеспечивают связь между иммунной системой и нарушениями в кератиноцитах.
Чтобы исследовать, могут ли вышеописанные антитела блокировать эффекты обработки IL-19, IL-20 и IL-22, как описано в предыдущем примере, к RHE культуральной среде добавляли 20 микроорганизмов/мл анти-IL20Ra, анти-IL20Rb или анти-IL-22R за один час до добавления 20 нг/мл IL-19, IL-20 или IL-22. Затем в течение четырех дней клетки RHE культивировали в среде, которую меняли на второй день (4,5 мл свежей среды, включая цитокин и антитело). Затем RHE иммуногистохимически окрашивали на псориазин (S100A7). Результаты показаны на фиг. 40. IL-19, IL-20 и IL-22-обработанные RHE показаны на первой, второй и третьей строках, соответственно. Предобработанные анти-IL20Ra (αIL-20Ra), анти-IL20Rb (αIL-20Rb) или анти-IL-22R (αIL-22R1) клетки RHE показаны в третьем, четвертом и пятом столбцах, соответственно. Контроли без антител и контрольные антитела изотипа показаны в первом и втором столбцах.
В результатах показано, что либо анти-IL20Ra, либо анти-IL20Rb эффективно блокировали IL-19-индуцированную экспрессию псориазина. Аналогично, анти-IL-22R эффективно блокировали IL-22-индуцированную экспрессию псориазина. Анти-IL20Rb эффективно блокировали IL-20-индуцированную экспрессию псориазина, а анти-IL20Ra - нет. Аналогично, анти-IL-22R не могли блокировать IL-20-индуцированную экспрессию псориазина.
Чтобы далее исследовать эффекты анти-IL-22R и анти-IL20Ra на IL-20-индуцированную экспрессию псориазина, проводили предобработку RHE этими антителами либо по отдельности, либо в сочетании до обработки IL-20. Результаты показаны на фиг. 41. Как описано выше, ни анти-IL-22R, ни анти-IL20Ra сами по себе не могли блокировать IL-20-индуцированную экспрессию псориазина (второй столбец, обе панели). Однако сочетание и анти-IL20Ra, и анти-IL-22R эффективно блокировали IL-20-индуцированную экспрессию псориазина, указывая, что IL-20Ra и IL-22R играют дополнительную роль при передаче сигнала IL-20 в человеческих кератиноцитах (левая панель ниже).
ПРИМЕР 25: IL-19, IL-20, IL-22 и IL-24 вызывают аналогичные характеры генной экспрессии
Чтобы идентифицировать гены, индуцируемые IL-19, IL-20, IL-22 и IL-24, обрабатывали RHE 20 нг/мл IL-19, IL-20, IL-22 или IL-24 в течение четырех дней. Выделяли РНК, и кДНК гибридизовали к Affymetrix U133 Plus Gene Chips (Affymetrix, Santa Clara, CA), которые содержат наборы на 54675 проб. Данные анализировали на гены, экспрессия которых увеличивалась, по крайней мере, 2-кратно. IL-20, IL-22 и IL-24 показали аналогичные профили экспрессии генов. Ранее сообщалось, что из 20 лидерных генов, обычно индуцированных IL-20, IL-22 и IL-24, семь генов связаны с псориазом. Это гены псориазина (S100A7), S100A12, SCCA2, SERPINB4, CCL20, CD36 и Stat3.
Чтобы исследовать, увеличивают ли псориазин гены, индуцированные IL-20, IL-22 и IL-24, вышеописанные микрочиповые анализы сравнили с предыдущим микрочиповым исследованием псориатических клеток (Zhou et al. (2003) Physiol. Genomics 13:69-78). Так как это исследование проводили, используя различные микрочиповые чипы, то сравнили только в общем refseqs между этим исследованием и настоящим исследованием. Из 468 refseqs, которые увеличивались в псориатической коже, 356 индуцировались IL-20, IL-22 и IL-24, и 188 из них были значительными (p<0,05). Взятые вместе, вышеописанные исследования показывают значительное перекрывание между генами, которые индуцируются IL-20, IL-22 и IL-24, и генами, которые увеличиваются в псориатической коже.
ПРИМЕР 26: Депонирование материалов
Следующие гибридомные клеточные линии сданы на хранение в American Type Culture Collection, 10801 University Blvd., Manassas, VA 20110-2209 USA (ATCC).
Этот депозит был сделан в соответствии с положениями Будапештского договора о Международном признании депонирования микроорганизмов для целей патентной процедуры (Будапештский договор). Это гарантирует сохранение жизнеспособной культуры в течение 30 лет после даты депозита. Клеточная линия будет предоставлена ATCC в рамках Будапештского договора, и по соглашению между Genentech, Inc. и ATCC, которое гарантирует (a) что доступ к культуре будет предоставлен во время рассмотрения патентной заявки до определенного комиссаром, являющимся уполномоченным согласно 37 CFR §1.14 и 35 USC §122, и (b) что все ограничения на предоставление неограниченному кругу лиц таким образом хранящейся культуры будут окончательно удалены после выдачи патента.
Правопреемник настоящей заявки согласился, что в случае гибели хранящейся культуры на депозите или утери, или разрушения при культивировании в соответствующих условиях, она будет немедленно замещена по уведомлению на жизнеспособный образец той же самой культуры. Предоставление хранящейся клеточной линии нельзя объяснить лицензией на практику изобретения в нарушение предоставленных прав под уполномочиванием любого правительства в соответствии с его патентными законами.
Считается, что изложенное выше описание достаточно, чтобы позволить квалифицированному специалисту в данной области применять изобретение. Настоящее изобретение не ограничивается областью хранящихся материалов, так как хранящиеся варианты осуществления предполагались в качестве простой иллюстрации некоторых аспектов изобретения и любые конструкты, которые функционально эквиваленты, лежат в области этого изобретения. Депозит материалов здесь не представляет собой признание того, что написанное описание, заключенное в данный документ, является недостаточным для разрешения применения любого аспекта изобретения, включая лучший вариант его, а также нельзя объяснить ограничением области формул изобретения для специальных примеров, в которых это представлено. Действительно, различные модификации изобретения в дополнение к представленным и описанным в данном описании станут очевидными квалифицированным специалистам в данной области из изложенного выше описания и лежат в области прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2702108C2 |
| ИНГИБИТОРЫ АНГИОПОЭТИНПОДОБНОГО БЕЛКА 4, ИХ КОМБИНАЦИИ И ПРИМЕНЕНИЕ | 2005 |
|
RU2392966C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2825390C2 |
| НОВАЯ КОМПОЗИЦИЯ И СПОСОБЫ ЛЕЧЕНИЯ СВЯЗАННЫХ С ИММУНИТЕТОМ ЗАБОЛЕВАНИЙ | 2004 |
|
RU2375078C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| ИНГИБИРОВАНИЕ МЕТАСТАЗИРОВАНИЯ ОПУХОЛИ ПРИ ПОМОЩИ АНТАГОНИСТОВ Bv8 ИЛИ G-CSF | 2010 |
|
RU2567803C2 |
| ГЕТЕРОДИМЕРНЫЕ ПОЛИПЕПТИДЫ IL-17 A/F И ВОЗМОЖНОСТИ ИХ ЛЕЧЕБНОГО ПРИМЕНЕНИЯ | 2007 |
|
RU2440134C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| ТРИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IL-17A, IL-17F И ДРУГОЙ ПРОВОСПАЛИТЕЛЬНОЙ МОЛЕКУЛЫ | 2016 |
|
RU2680011C2 |





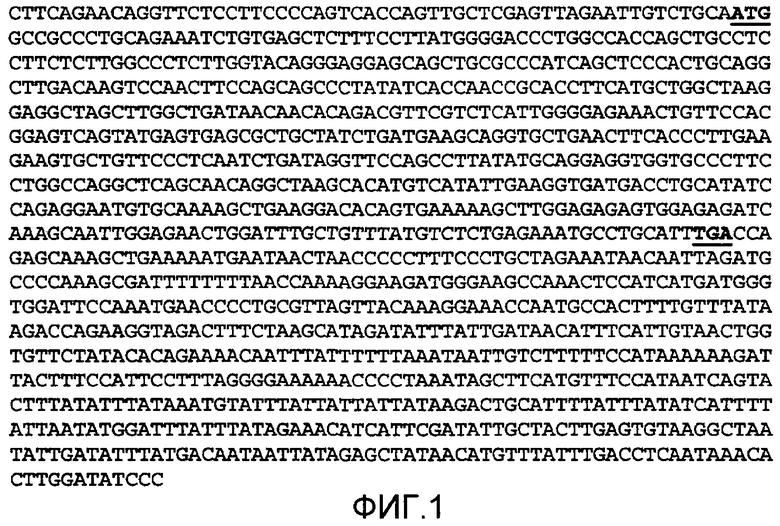



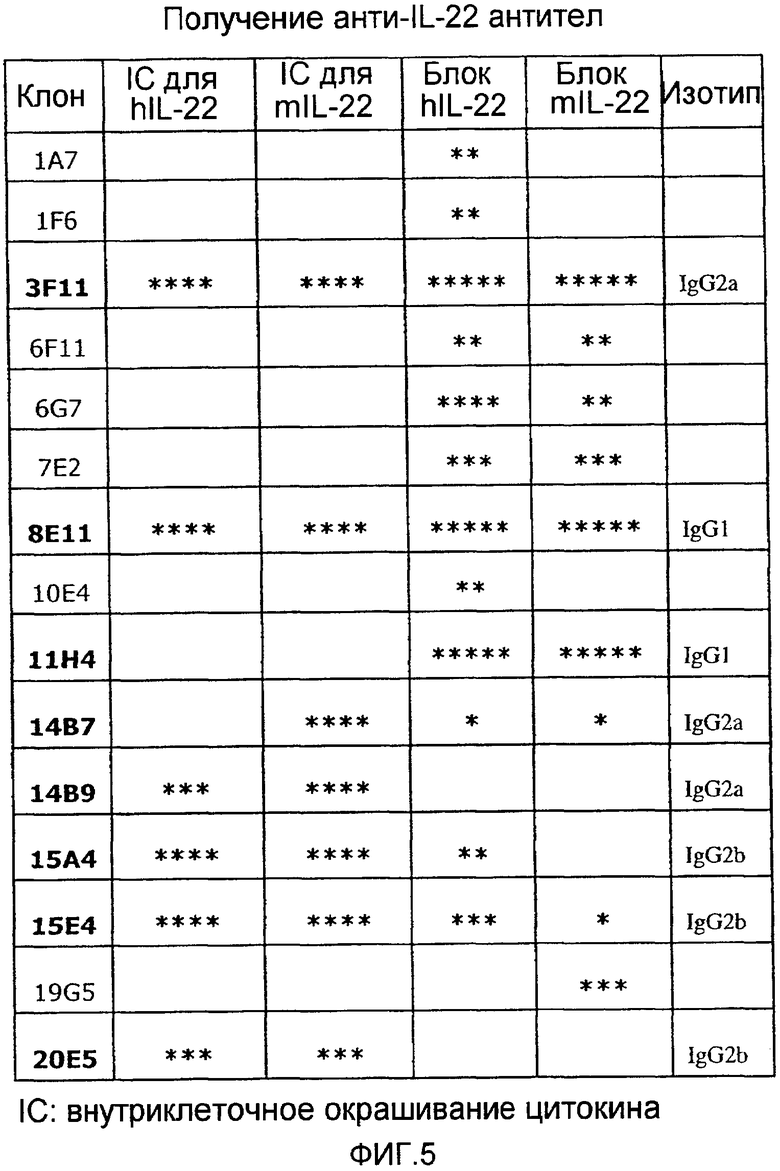
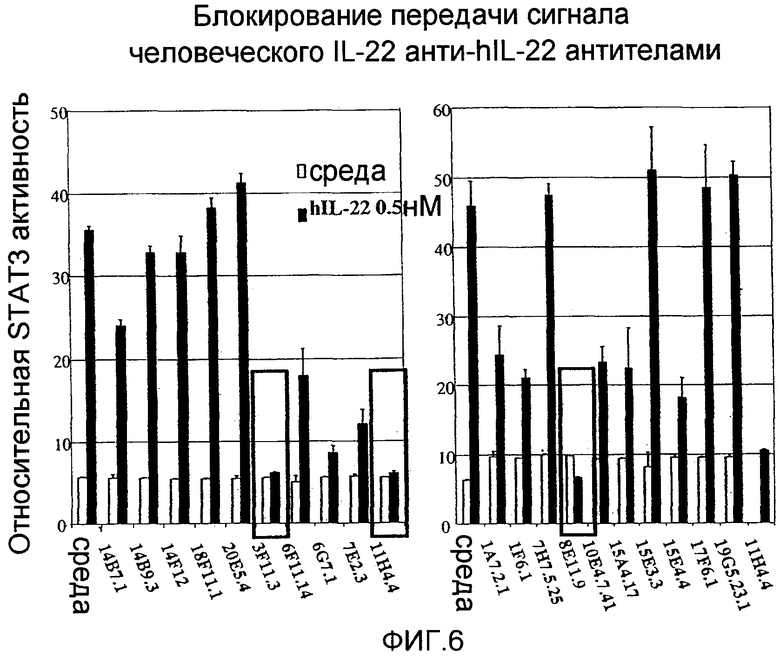
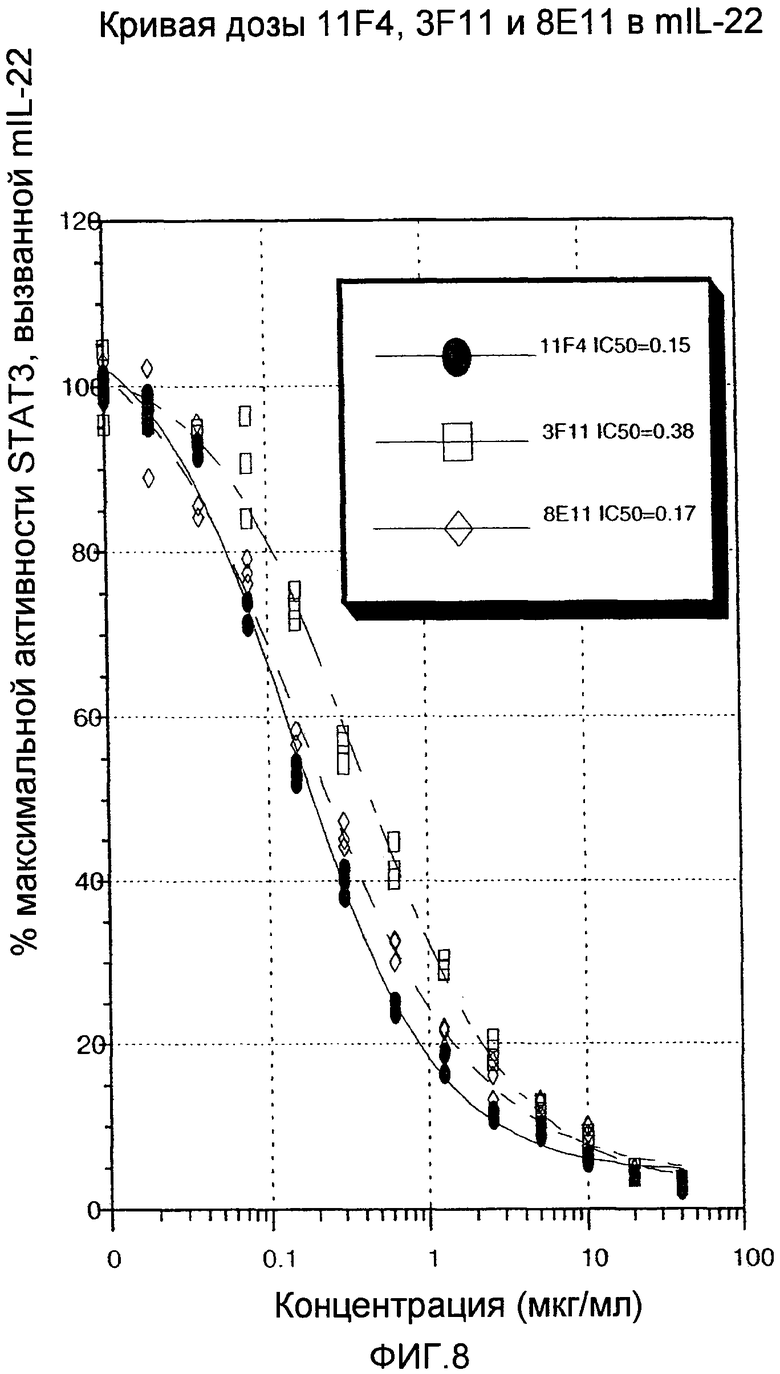


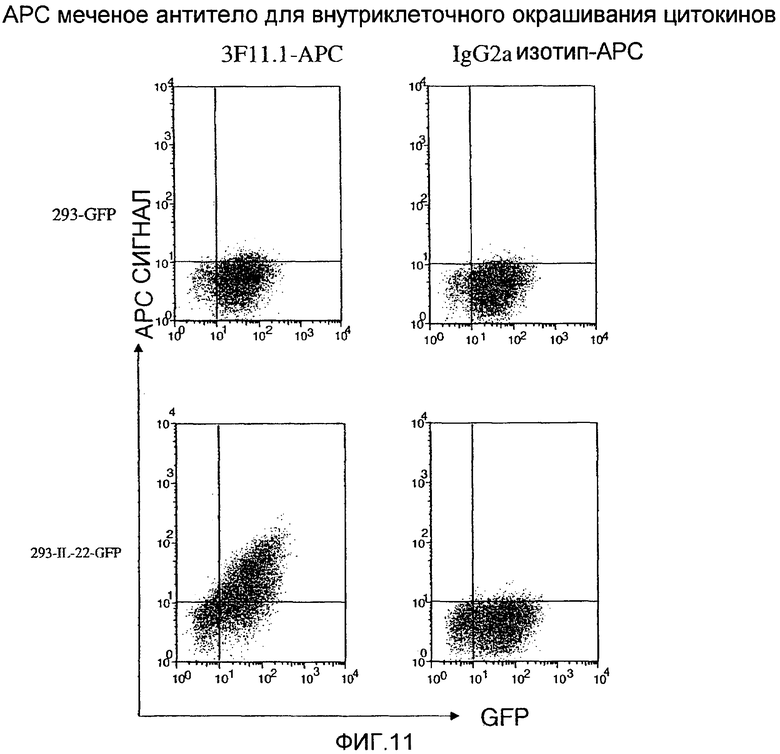
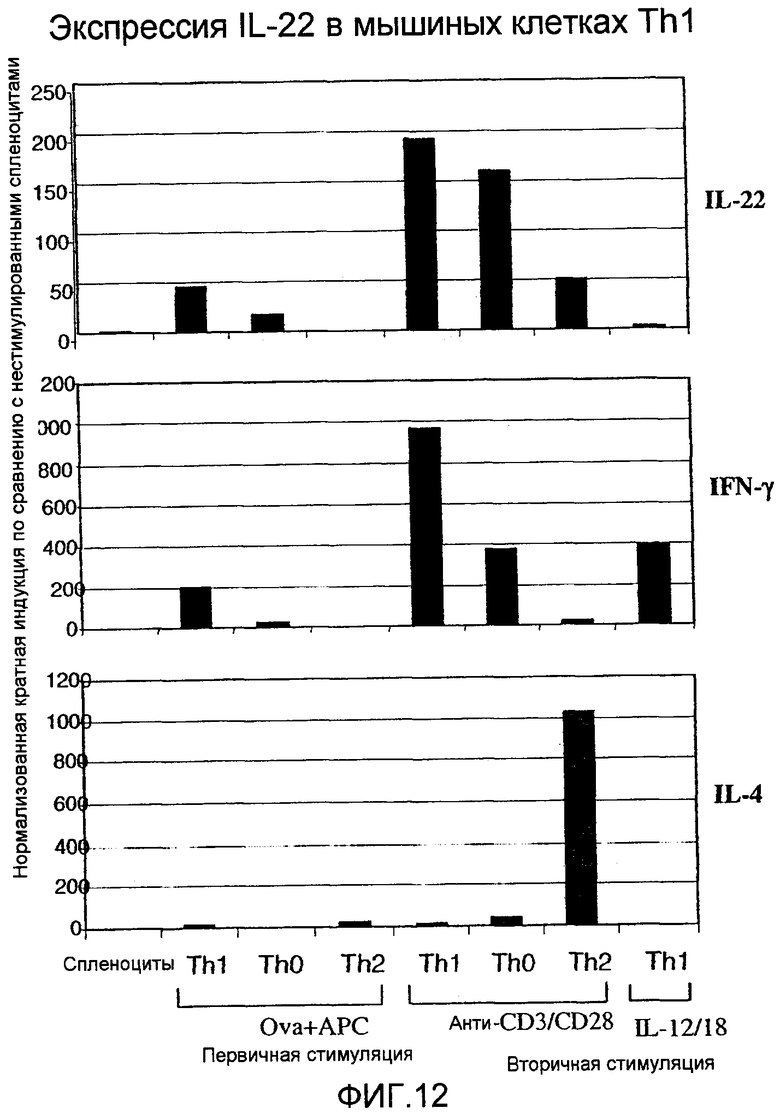
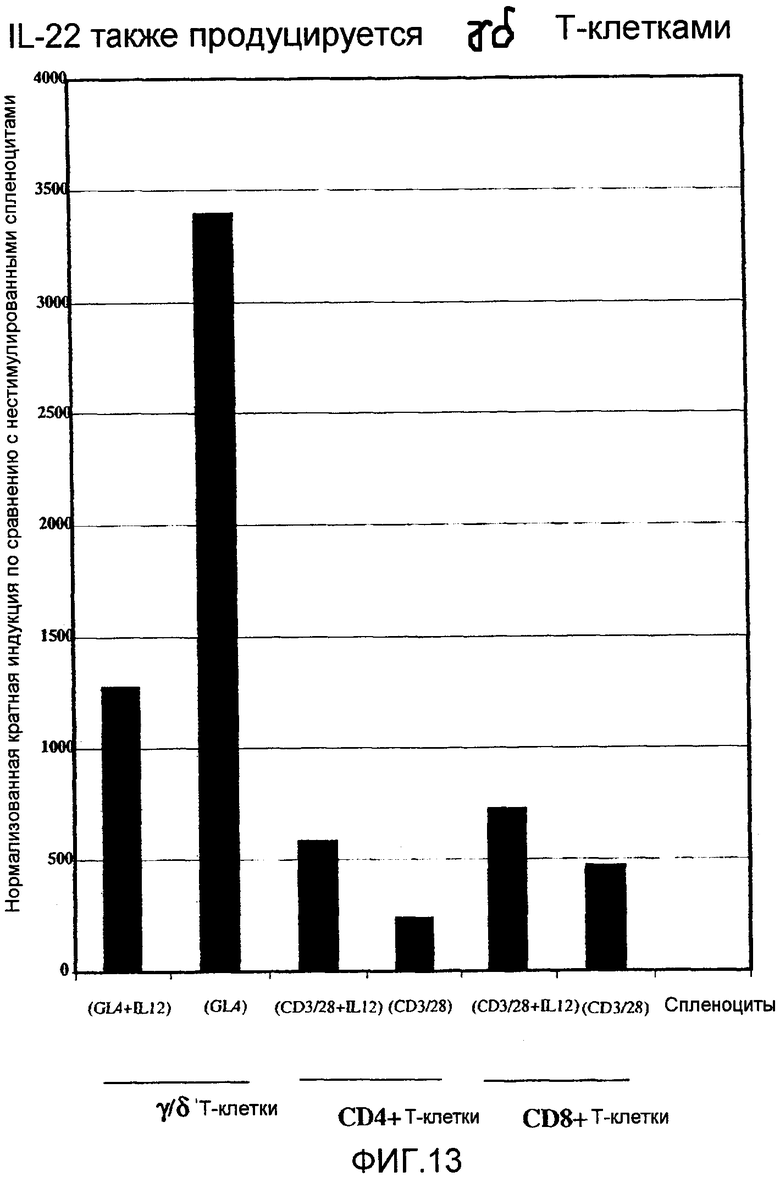
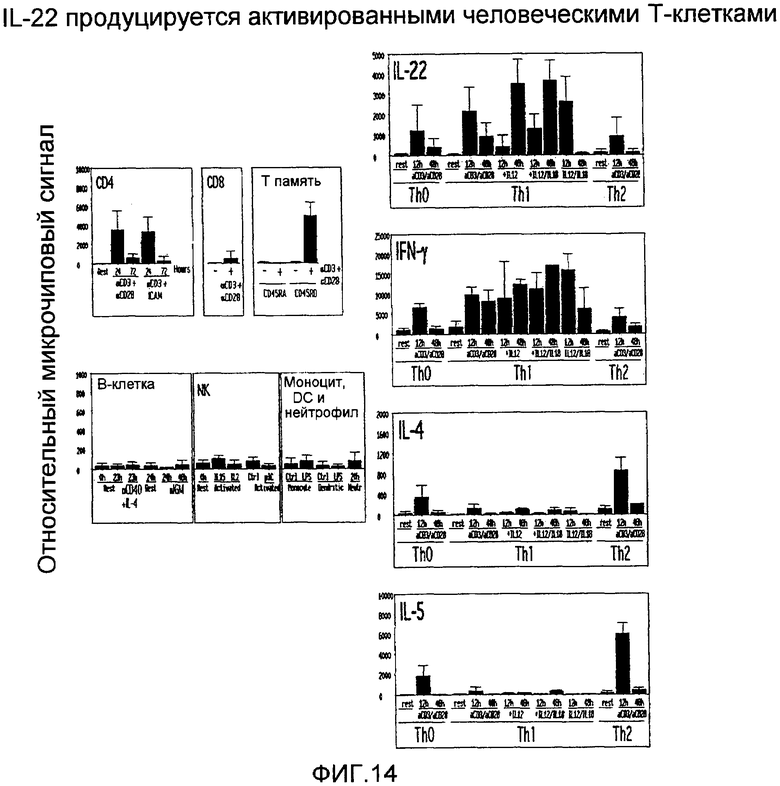
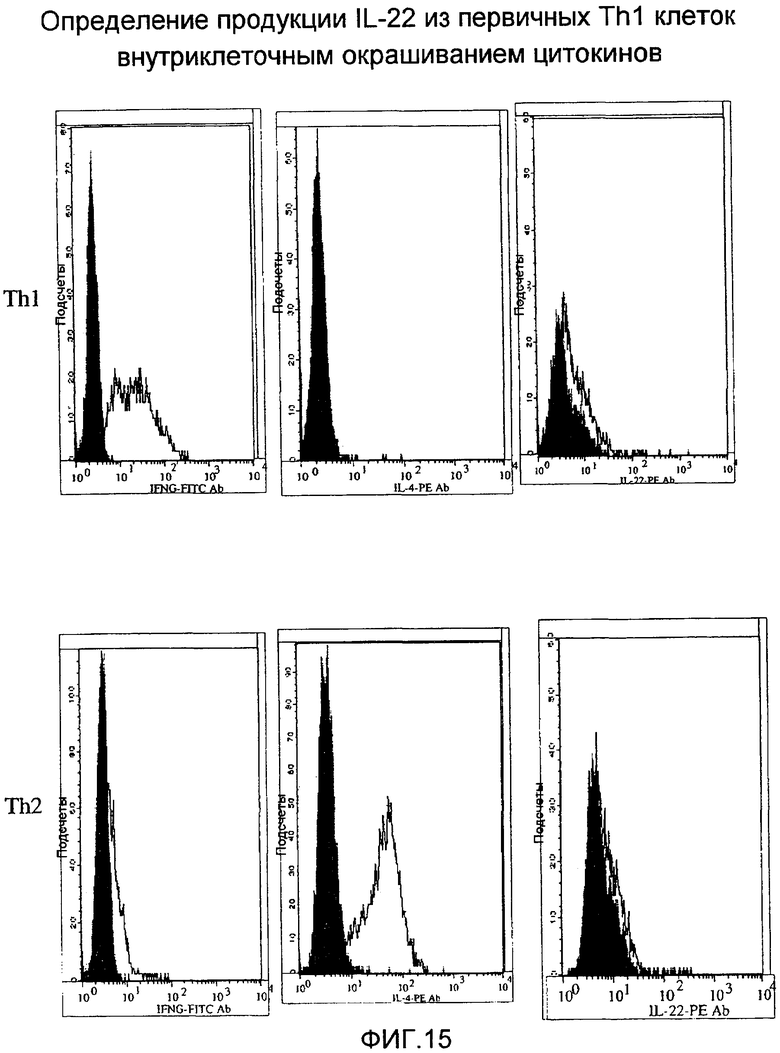
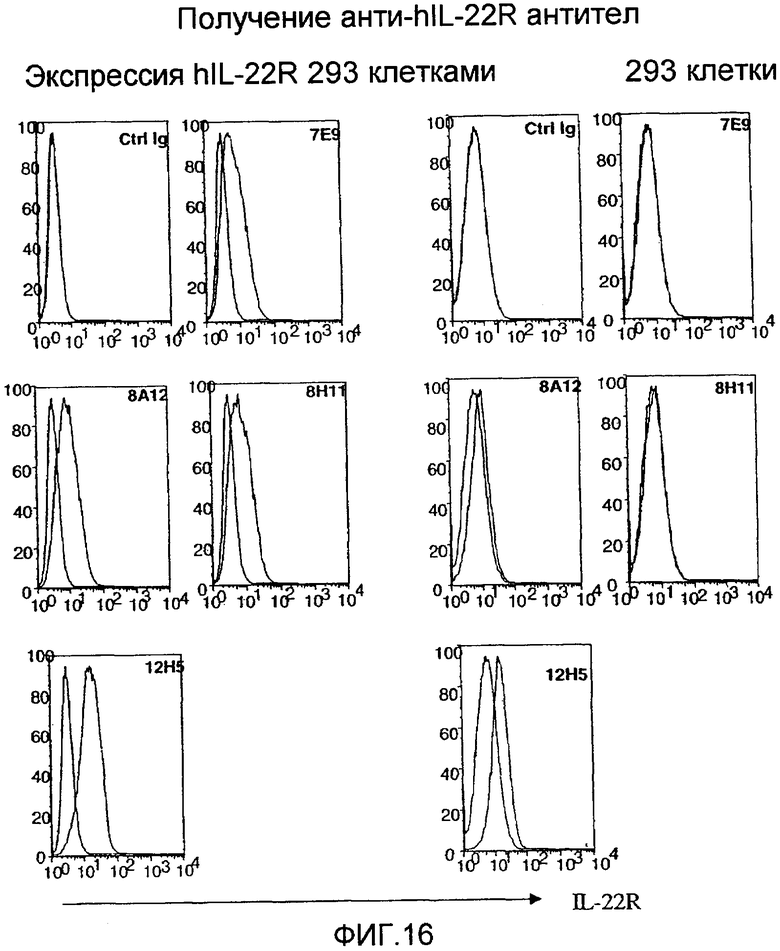
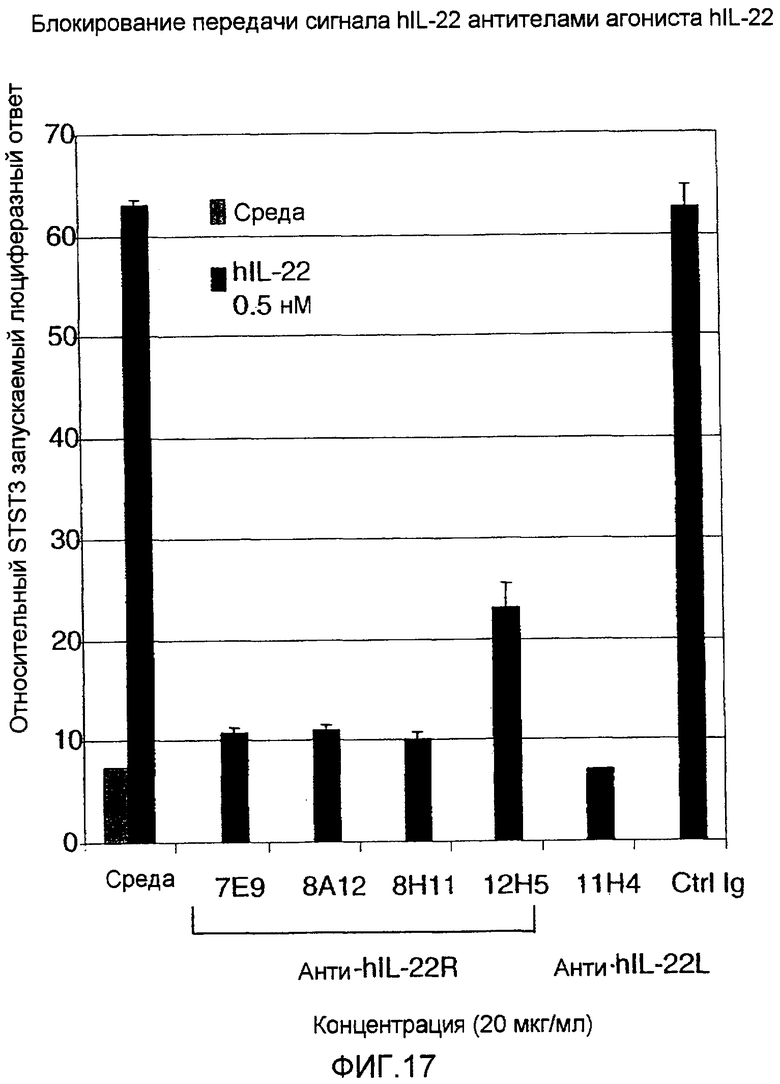
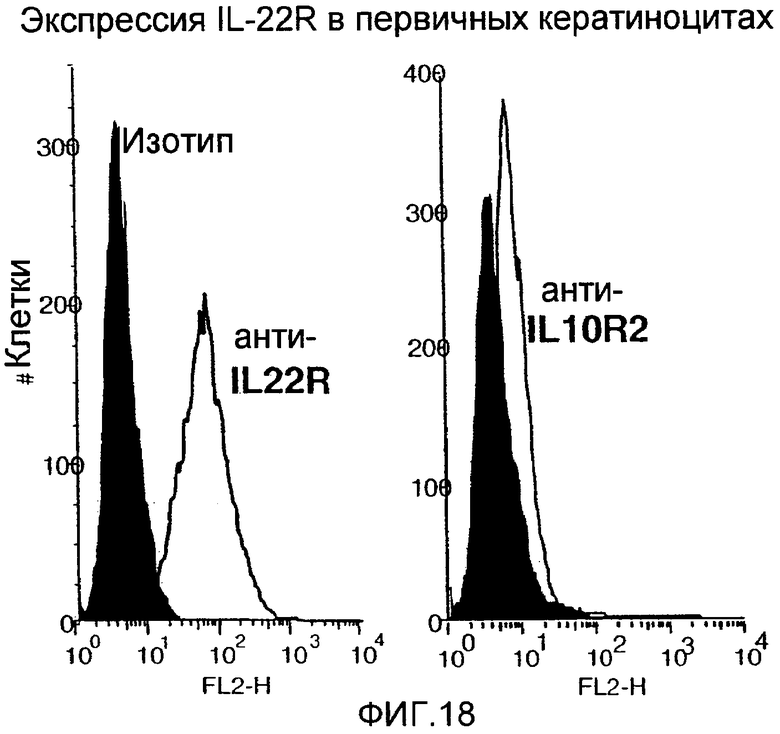
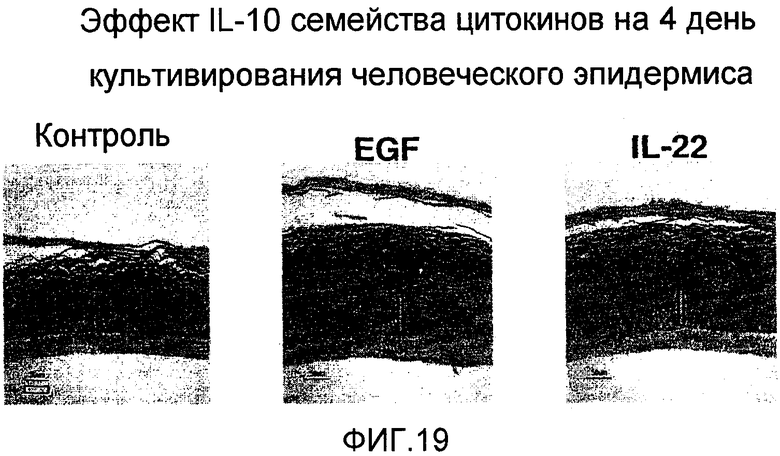


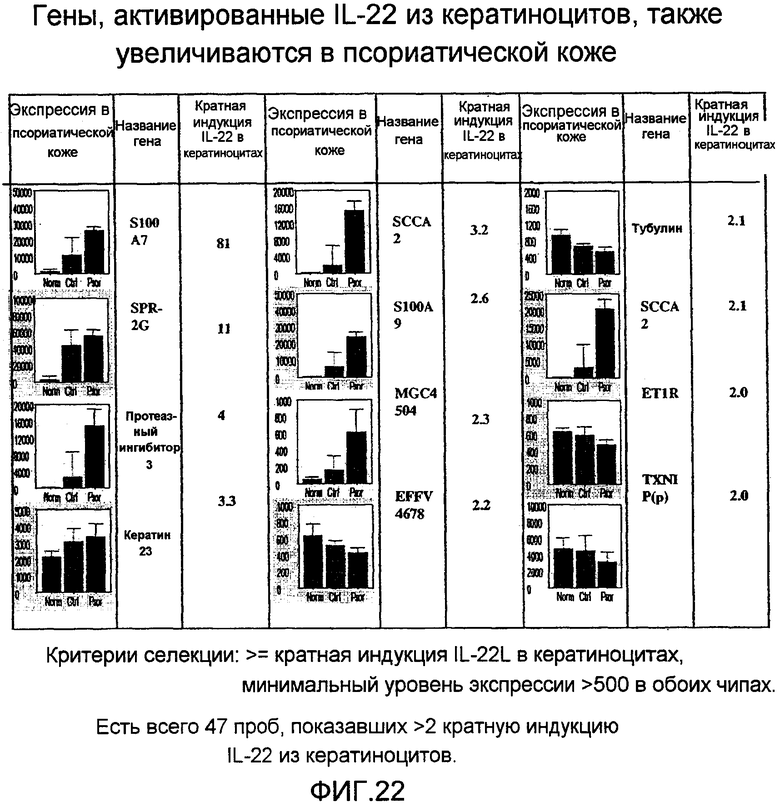
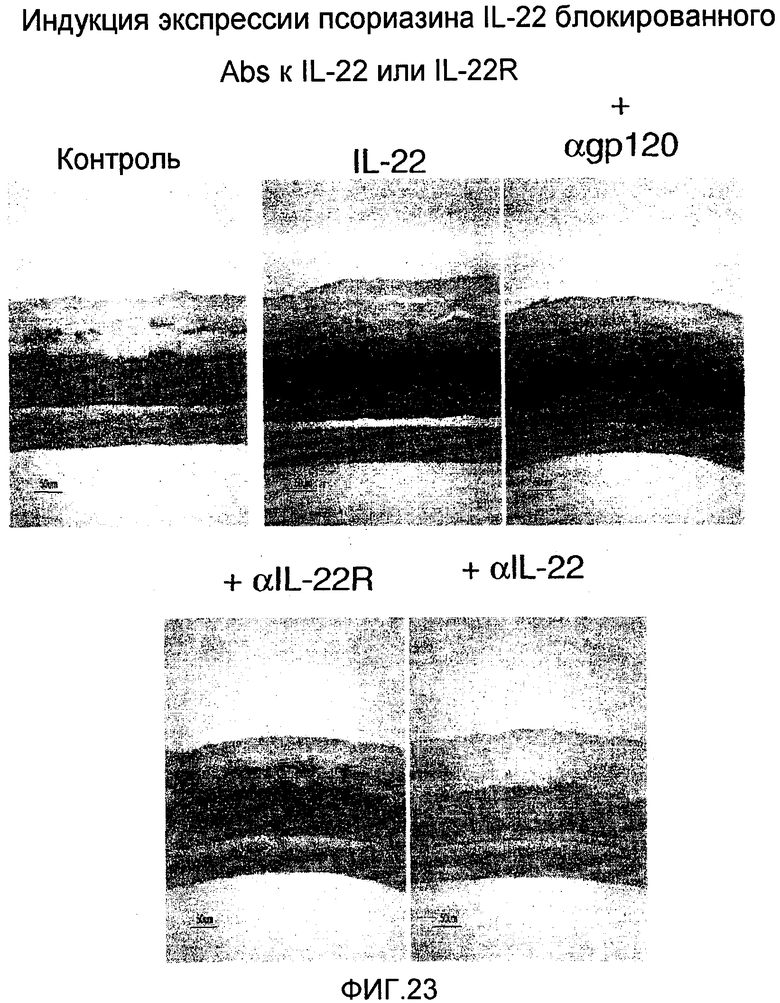
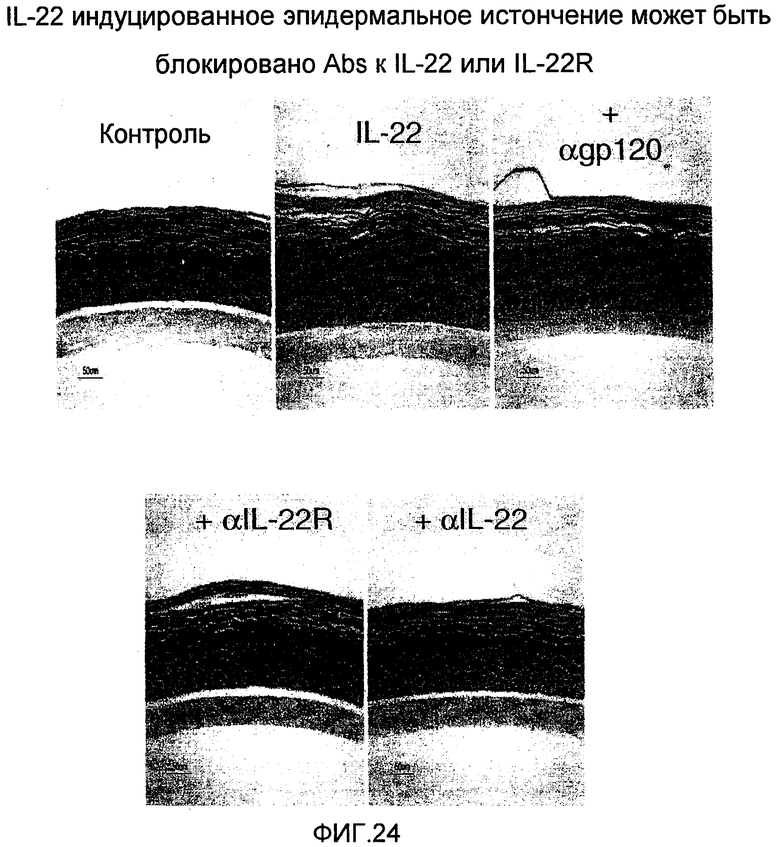
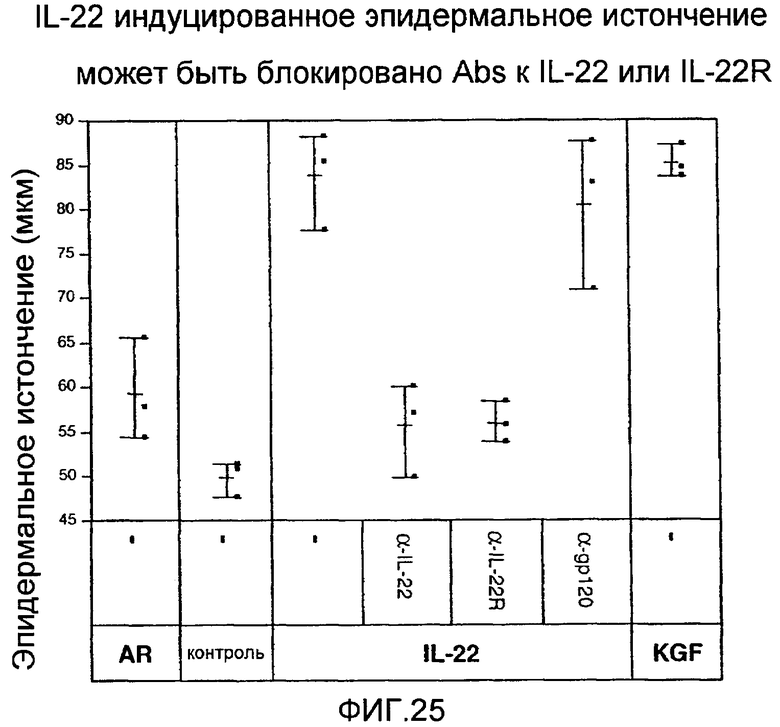
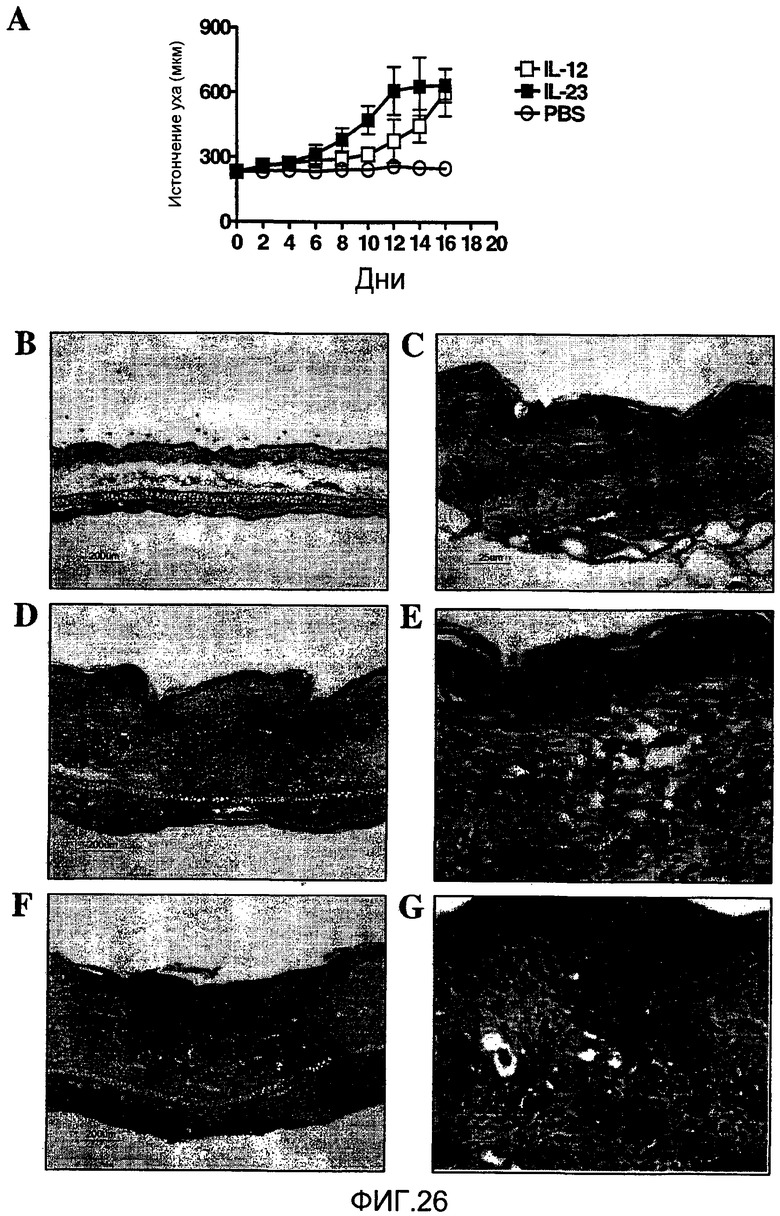
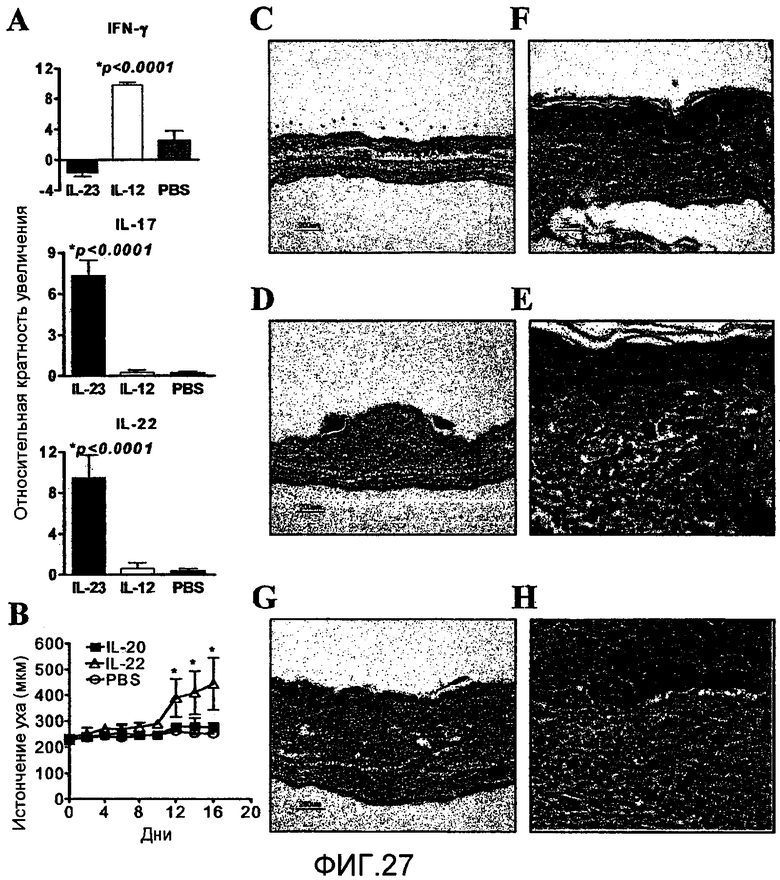
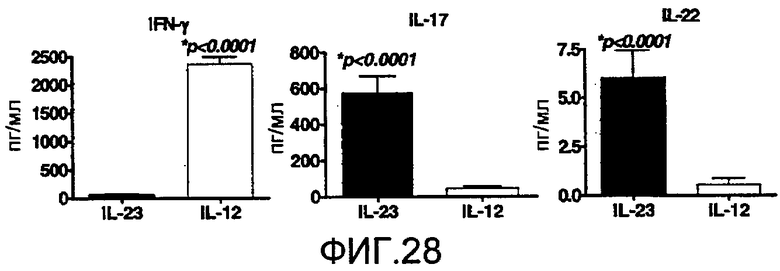
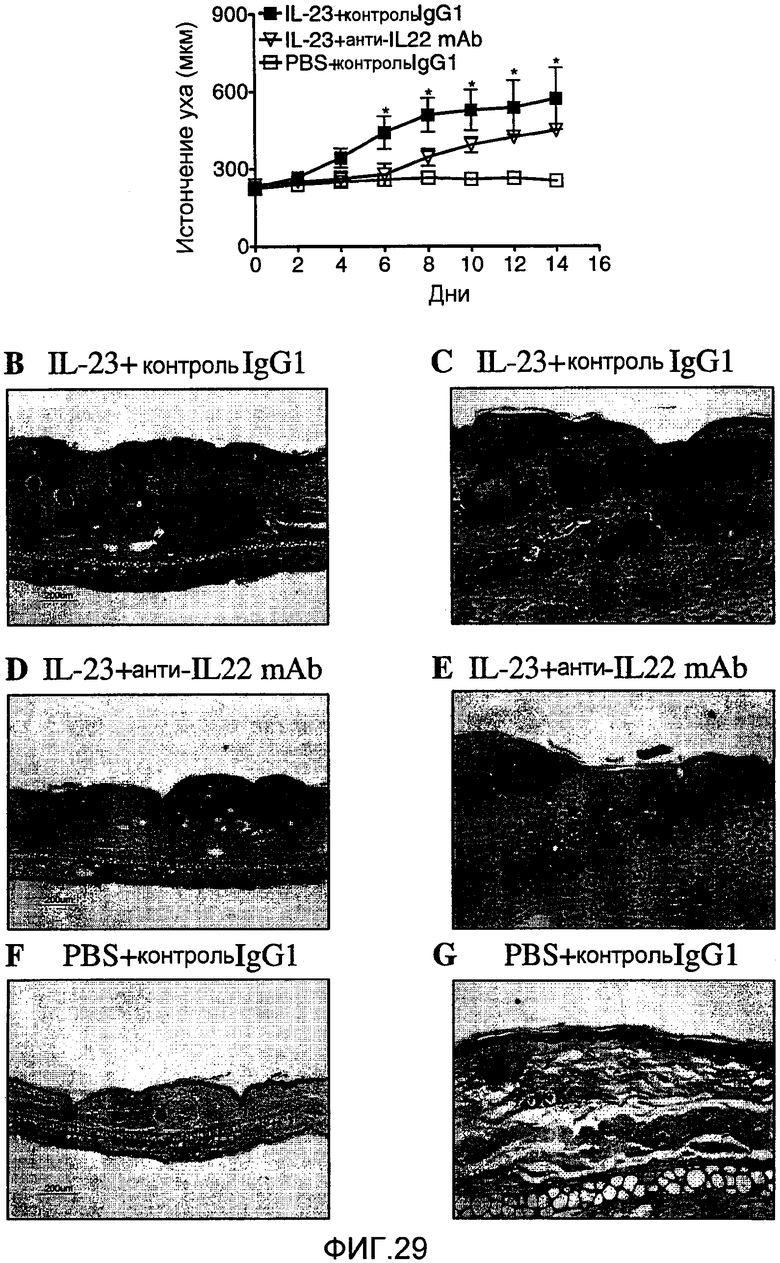
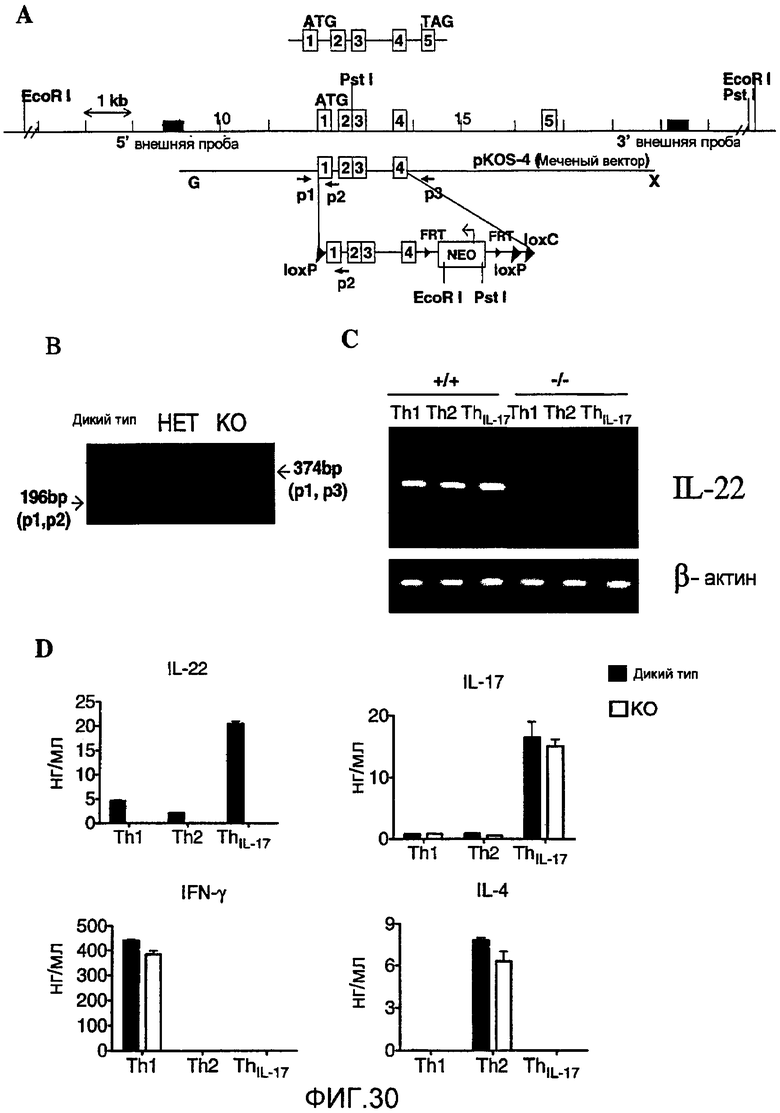
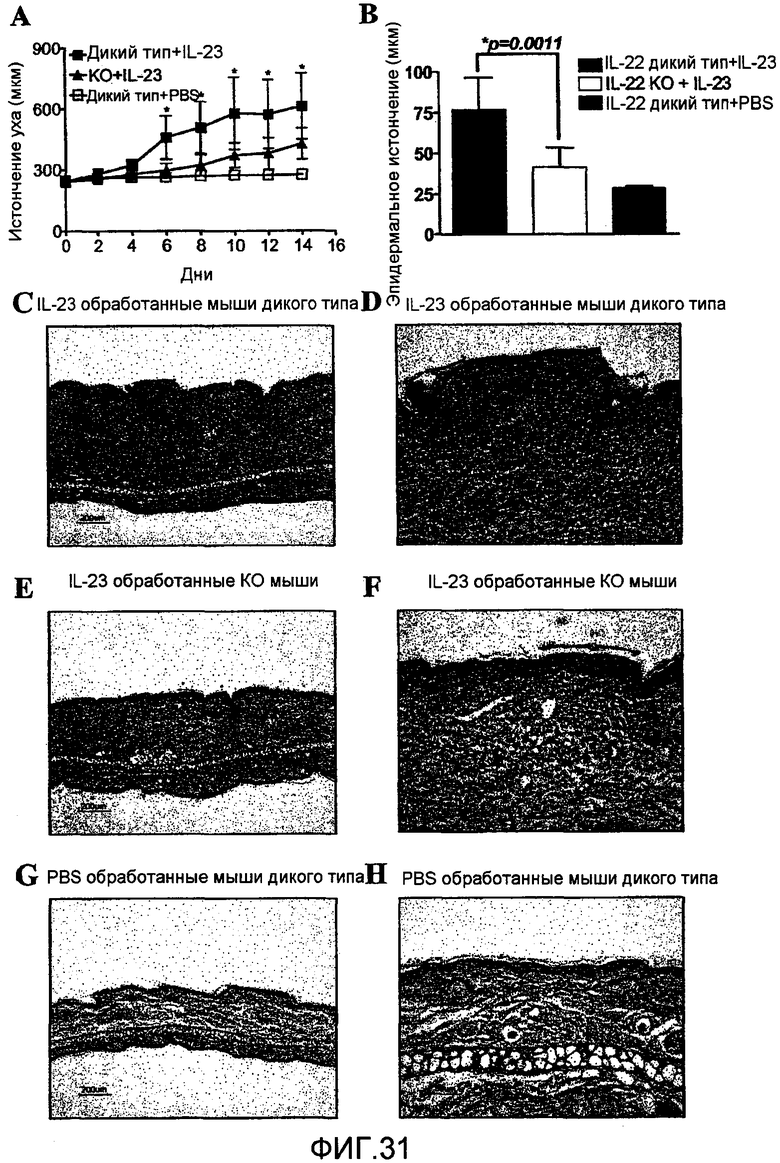
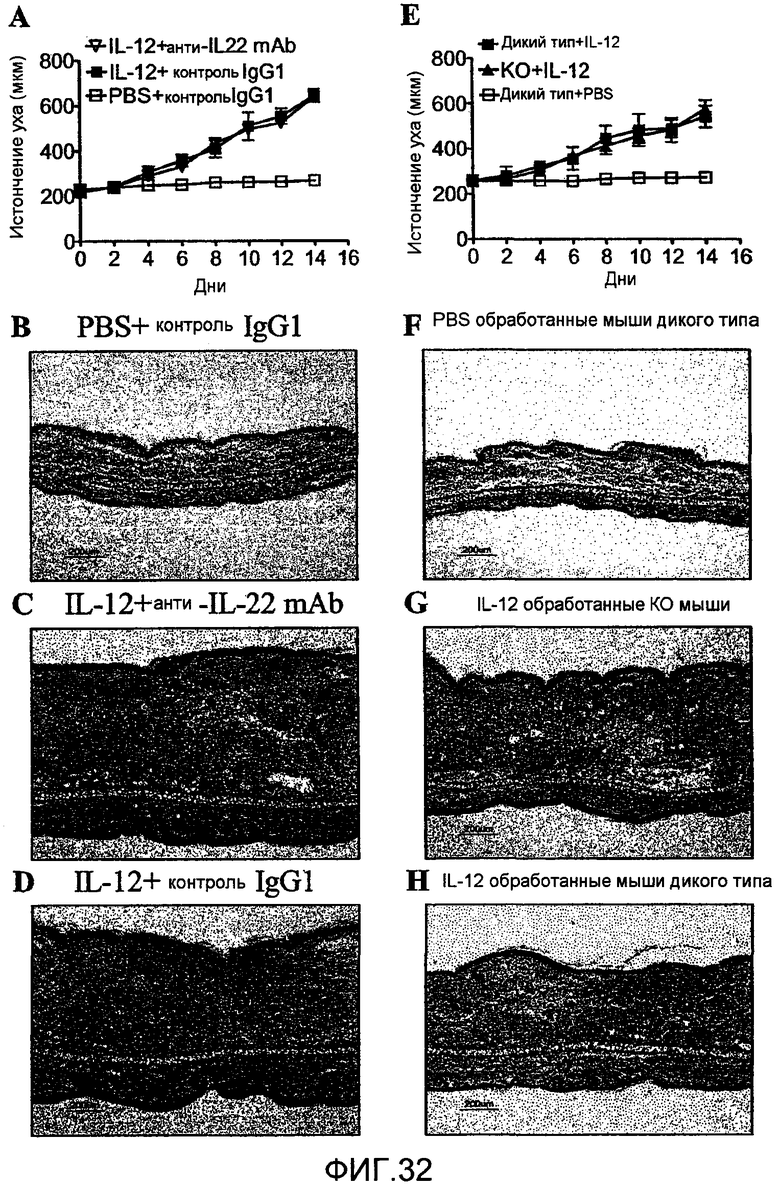
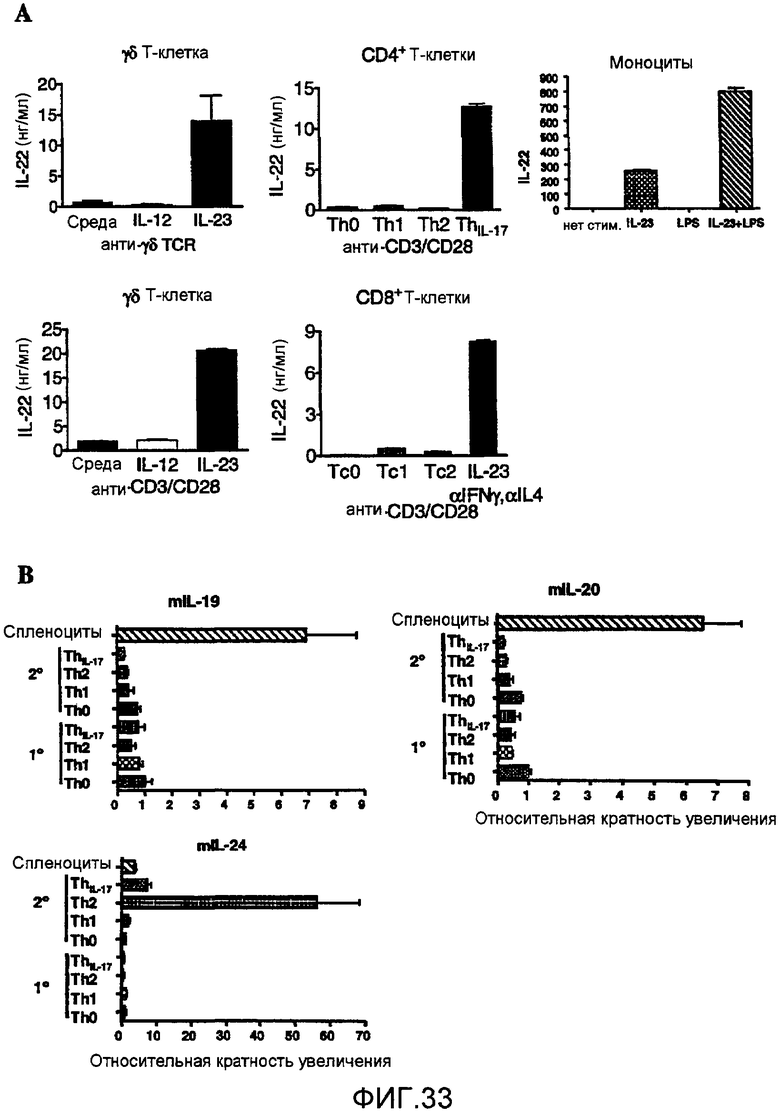

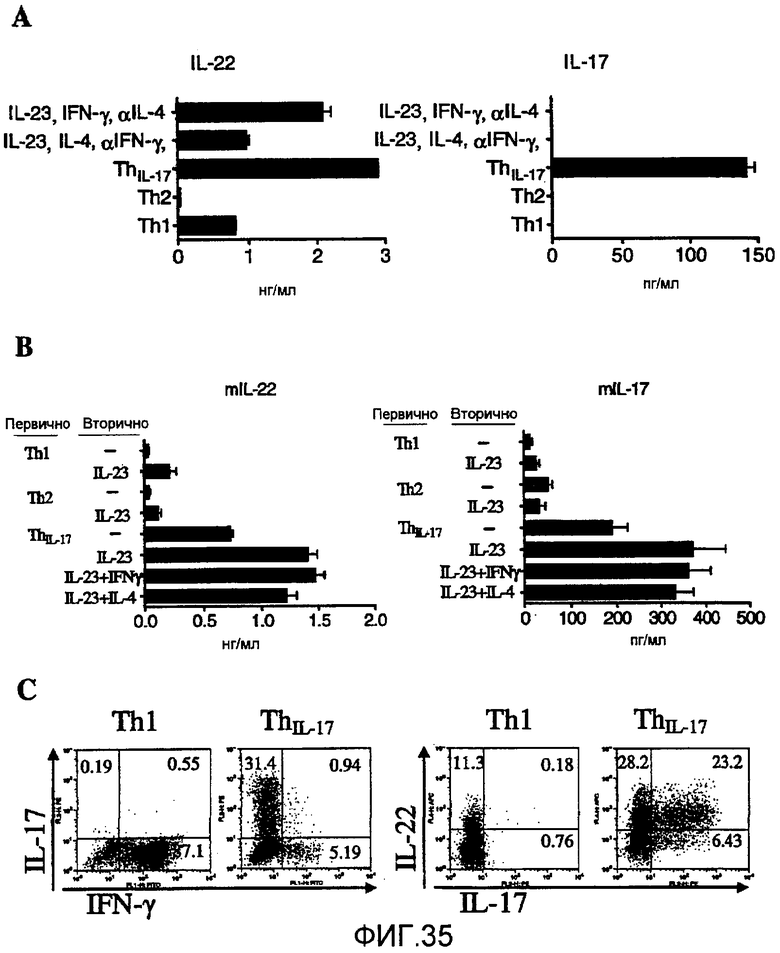
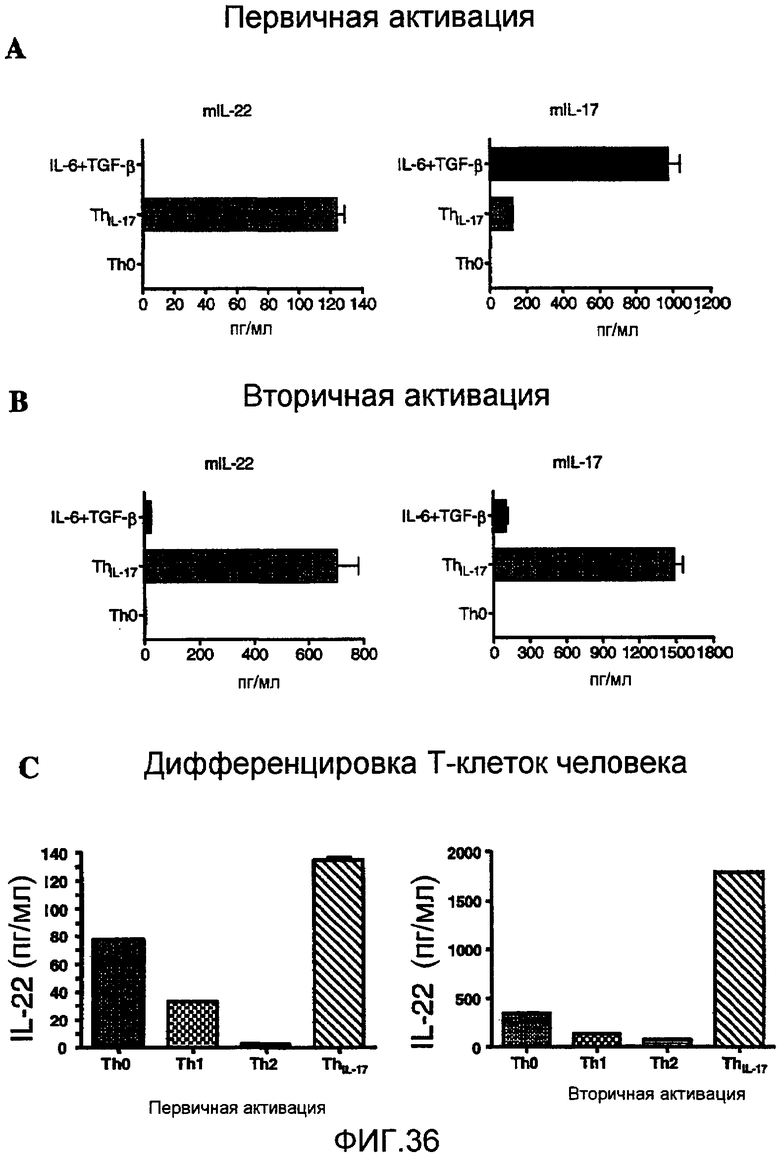
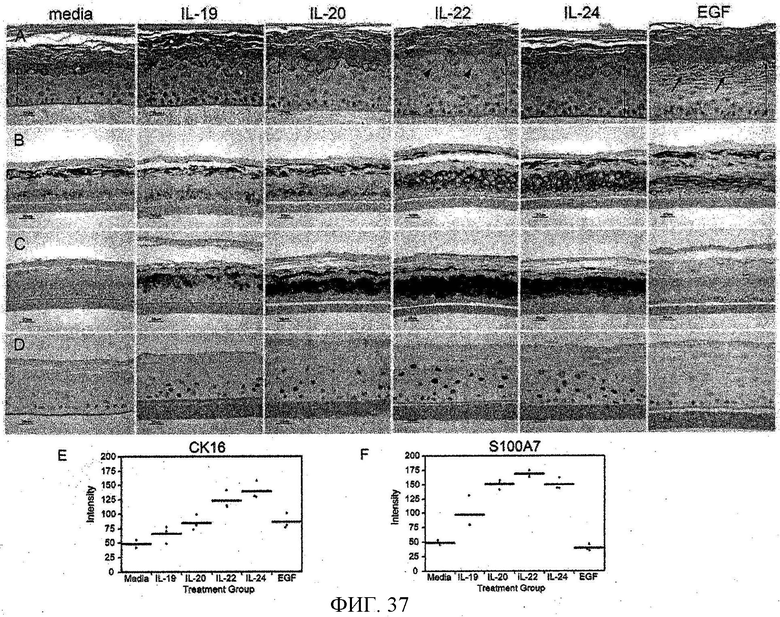
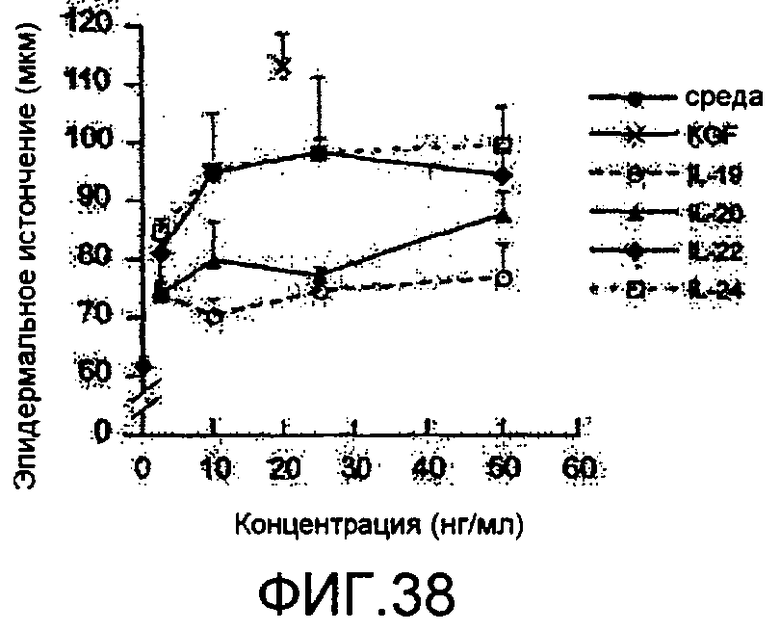
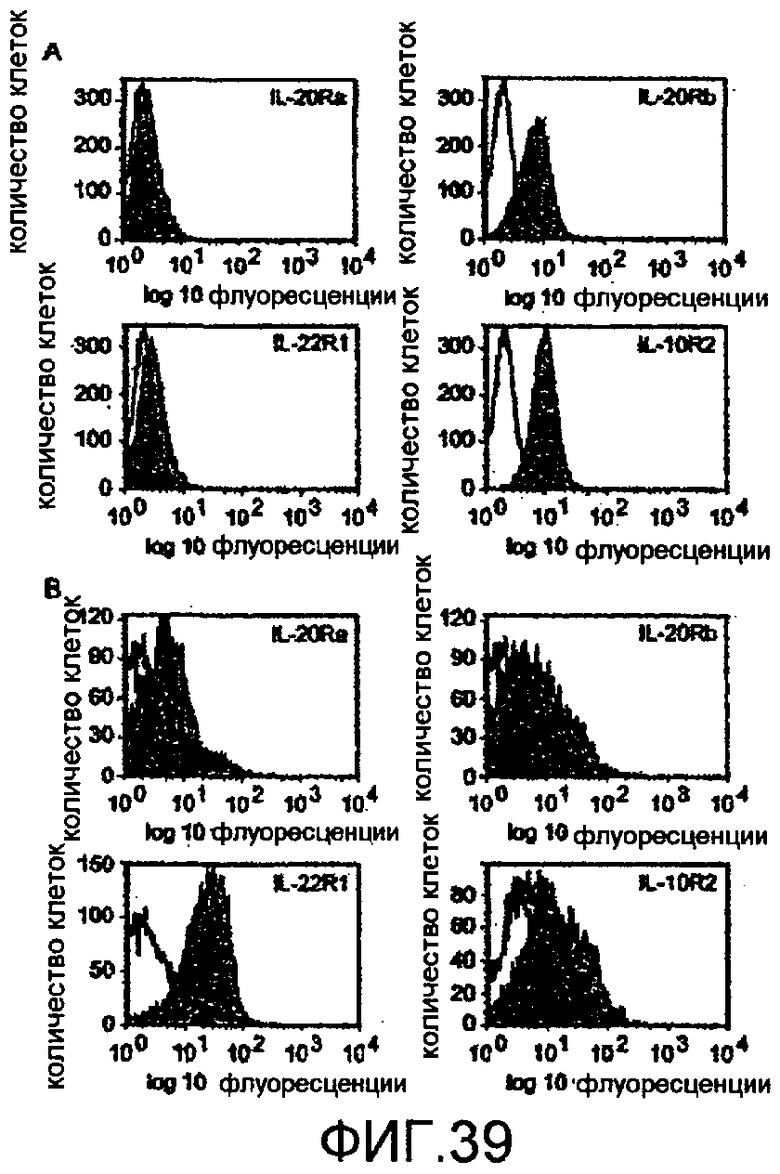
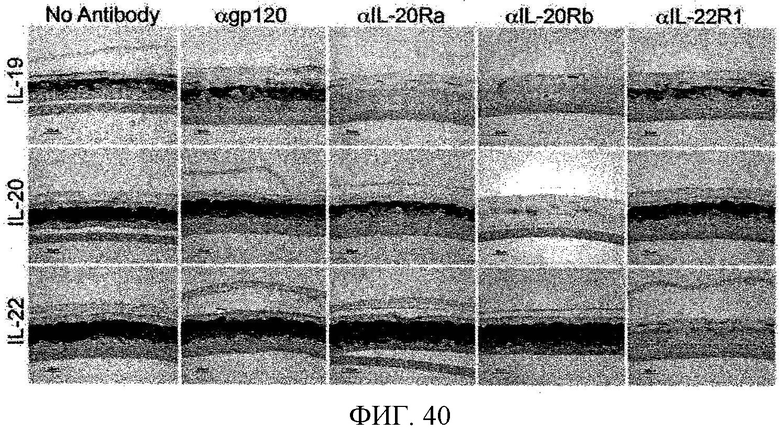
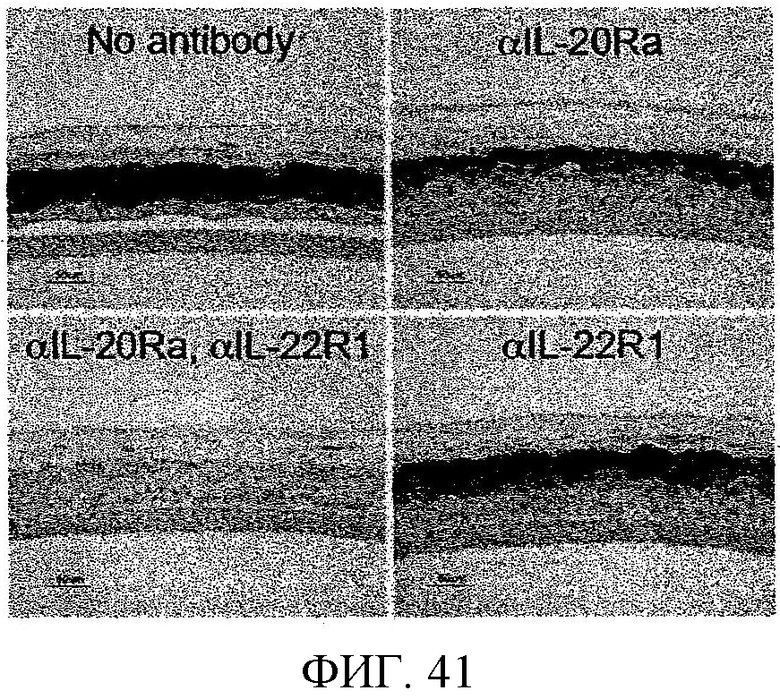
Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты антител, специфичных к IL22 человека. Антитела отличаются тем, что их получают из разных гибридом РТА-7312, РТА-7315, РТА-7319. Антитело из РТА-7312 характеризуется тем, что ингибирует активацию STAT3, достигая IC50 при концентрации 0,14 мкг/мл, тогда как антитела из РТА-7315, РТА-7319 характеризуются Kd менее 1 нМ. Использование изобретения может найти дальнейшее применение в терапии иммунных расстройств, опосредованных IL22. 3 н. и 21 з.п. ф-лы, 1 табл., 41 ил.
1. Антитело, которое специфически связывается с IL-22 человека, содержащее последовательности вариабельных областей, специфичные к IL-22, где антитело продуцируется гибридомой 3F11.3 (АТСС Accession № РТА-7312), где антитело ингибирует активацию STAT3, достигая IC50 при концентрации 0,14 мкг/мл.
2. Антитело по п.1, которое представляет собой аффинно зрелую форму антитела.
3. Антитело по п.2, которое представляет собой антиген-связывающий фрагмент антитела.
4. Антитело по п.3, которое представляет собой гуманизированную форму антитела.
5. Антитело по п.2, которое представляет собой гуманизированную форму антитела.
6. Антитело по п.1, которое представляет собой антиген-связывающий фрагмент антитела.
7. Антитело по п.6, которое представляет собой гуманизированную форму антитела.
8. Антитело по п.1, которое представляет собой гуманизированную форму антитела.
9. Антитело, которое специфически связывается с IL-22 человека с аффинностью, составляющей менее 1 нм, содержащее последовательности вариабельных областей, специфичные к IL-22, где антитело продуцируется гибридомой 11Н4.4 (АТСС Accession № РТА-7315).
10. Антитело по п.9, которое представляет собой аффинно зрелую форму антитела.
11. Антитело по п.10, которое представляет собой антиген-связывающий фрагмент антитела.
12. Антитело по п.11, которое представляет собой гуманизированную форму антитела.
13. Антитело по п.10, которое представляет собой гуманизированную форму антитела.
14. Антитело по п.9, которое представляет собой антиген-связывающий фрагмент антитела.
15. Антитело по п.14, которое представляет собой гуманизированную форму антитела.
16. Антитело по п.9, которое представляет собой гуманизированную форму антитела.
17. Антитело, которое специфически связывается с IL-22 человека с аффинностью, составляющей менее 1 нм, содержащее последовательности вариабельных областей, специфичные к IL-22, где антитело продуцируется гибридомой 8Е11.9 (АТСС Accession № РТА-7319).
18. Антитело по п.17, которое представляет собой аффинно зрелую форму антитела.
19. Антитело по п.18, которое представляет собой антиген-связывающий фрагмент антитела.
20. Антитело по п.19, которое представляет собой гуманизированную форму антитела.
21. Антитело по п.18, которое представляет собой гуманизированную форму антитела.
22. Антитело по п.17, которое представляет собой антиген-связывающий фрагмент антитела.
23. Антитело по п.22, которое представляет собой гуманизированную форму антитела.
24. Антитело по п.17, которое представляет собой гуманизированную форму антитела.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Nagalakshmi et al., "Interieukin-22 activates STATS and induces IL-10 by colon epithelial cells", Int | |||
| Immunopharmacol, 2004; 4(5), реферат | |||
| RU 2005131852 A, 20.04.2006. | |||

