Область изобретения
Настоящее изобретение относится к композициям и способам, применимым для диагностики и лечения связанных с иммунитетом заболеваний.
Предпосылки изобретения
Связанные с иммунитетом и воспалительные заболевания представляют собой проявление или следствие достаточно сложных, часто многократно взаимосвязанных биологических путей, которые в нормальной физиологии являются критическими для ответа на поражение или травму, инициируют восстановление после поражения или травмы и устанавливают врожденную и приобретенную защиту против чужеродных организмов. Заболевание или патология возникают, когда эти нормальные физиологические пути вызывают дополнительное поражение или травму либо напрямую связанные с интенсивностью ответа, либо как следствие аномальной регуляции или избыточной стимуляции, либо как реакцию на «свое», либо как сочетание этих механизмов.
Хотя возникновение данных заболеваний часто вовлекает многоступенчатые пути, а часто - множество различных биологических систем/путей, вмешательство в критических точках в один или несколько из этих путей может вызывать улучшение или терапевтический эффект. Терапевтическое вмешательство может происходить посредством антагонизма вредоносному процессу/пути или посредством стимуляции благоприятного процесса/пути.
Известно много связанных с иммунитетом заболеваний, и их интенсивно изучают. Такие заболевания включают в себя иммуноопосредованные воспалительные заболевания, неиммуноопосредованные воспалительные заболевания, инфекционные заболевания, иммунодефицитные заболевания, неоплазию, и т.д.
T-лимфоциты (T-клетки) являются важным компонентом иммунного ответа млекопитающих. T-клетки узнают антигены, связанные с молекулой собственного организма, кодируемой генами главного комплекса гистосовместимости (MHC). Антиген может быть представлен вместе с молекулами MHC на поверхности антигенпредставляющих клеток, инфицированных вирусом клеток, раковых клеток, трансплантатов и т.д. Система T-клеток уничтожает эти измененные клетки, которые представляют угрозу здоровью млекопитающего-хозяина. T-клетки включают в себя хелперные T-клетки и цитотоксические T-клетки. Хелперные T-клетки экстенсивно пролиферируют после узнавания комплекса антиген-MHC на антигенпредставляющих клетках. Хелперные T-клетки также секретируют множество цитокинов, т.е. лимфокинов, играющих центральную роль в активации B-клеток, цитотоксических T-клеток и множества других клеток, участвующих в иммунном ответе.
Связанные с иммунитетом заболевания можно лечить подавлением иммунного ответа. Использование нейтрализующих антител, ингибирующих молекулы, обладающие иммуностимулирующей активностью, было бы выгодно при лечении иммуноопосредованных и воспалительных заболеваний. Можно использовать молекулы, подавляющие иммунный ответ (непосредственно белки или опосредованно используя антитела-агонисты) для подавления иммунного ответа и, таким образом, облегчения иммуноопосредованного заболевания.
Известно, что CD4+ T-клетки являются важными регуляторами воспаления. Здесь активировали CD4+ T-клетки и анализировали профиль генов, дифференциально экспрессирующихся при активации. Собственно специфические для активации гены могут представлять собой потенциальные терапевтические мишени. Костимуляция in vivo необходима для продуктивного иммунопролиферативного ответа. Список костимулирующих молекул достаточно обширный, и до сих пор не ясно даже, какие костимулирующие молекулы играют критические роли при различных типах и стадиях воспаления.
Термин воспалительное заболевание кишечника («IBD») означает группу хронических воспалительных расстройств с неизвестными причинами, при которых кишечник (пищеварительный тракт) становится воспаленным, часто вызывающими спазмы или диарею. Распространение IBD в США оценивают приблизительно составляющим 200 на 100000 популяции. Пациентов с IBD можно разделить на две главные группы: пациентов с язвенным колитом («UC») и пациентов с болезнью Крона («CD»).
У пациентов с UC присутствует воспалительная реакция, первоначально вовлекающая слизистую оболочку толстой кишки. Воспаление, как правило, является однородным и непрерывным без промежуточных областей нормальной слизистой. Поверхностные клетки слизистой оболочки, так же как и эпителий крипты и подслизистой оболочки, вовлечены в воспалительную реакцию с инфильтрацией нейтрофилов. В конечном счете эта ситуация, как правило, прогрессирует до разрушения эпителия с потерей эпителиальных клеток, приводя к множественным изъязвлениям, фиброзу, дисплазии и продольной ретракции толстой кишки.
CD отличается от UC тем, что воспаление распространяется через все слои стенки кишечника и вовлекает брыжейку, так же как лимфатические узлы. CD может поражать любую часть пищеварительного тракта, от рта до ануса. Заболевание часто является периодическим, т.е. отрезки тяжелого заболевания кишечника разделены областями, кажущимися свободными от заболевания. При CD стенка кишечника также утолщается, что может привести к обструкциям. Кроме того, нередки фистулы и трещины.
Клинически IBD характеризуется разнообразными проявлениями, часто приводящими к хроническому, непредсказуемому течению. Геморрагический понос и боль в животе часто сопровождаются лихорадкой и потерей веса. Нередка анемия, так как это тяжелое истощение. Часто с IBD связаны суставные манифестации, лежащие в диапазоне от артралгии до острого артрита, та кже как и патологии функции почек. Пациенты с IBD подвержены также увеличенному риску карцином толстой кишки по сравнению с общей популяцией. Во время острых «приступов» IBD работа и другая нормальная активность обычно невозможны, и часто пациента госпитализируют.
Хотя причина IBD остается неизвестной, вовлечены несколько факторов, таких как генетическая, инфекционная и иммунологическая предрасположенность. IBD является наиболее распространенным у европеоидов, особенно у европеоидов еврейского происхождения. Хроническая воспалительная природа состояния побуждает к интенсивному поиску возможной инфекционной причины. Хотя обнаружены агенты, стимулирующие острое воспаление, ни для одного из них не показана инициация хронического воспаления, связанного с IBD. Гипотеза, что IBD представляет собой аутоиммунное заболевание, поддерживается упомянутым ранее внекишечной манифестацией IBD, например артритом суставов, и известным положительным ответом IBD на лечение такими терапевтическими средствами, как адренальные глюкокортикоиды, циклоспорин и азатиоприн, известные как подавляющие иммунный ответ. Кроме того, ЖК-тракт больше любого другого органа организма постоянно подвергается воздействию потенциальных антигенных веществ, таких как белки из пищи, побочные продукты бактерий (LPS) и т.д.
Кроме того, риск рака толстой кишки сильно повышен у пациентов с тяжелым язвенным колитом, особенно если заболевание существовало несколько лет. Приблизительно 20-25% пациентов с IBD в конечном счете нуждаются в операции по удалению толстой кишки из-за обширного кровотечения, хронического истощающего заболевания, прободения толстой кишки или риска рака. Операцию также иногда проводят, когда другие формы медицинского лечения неэффективны или когда побочные эффекты стероидов или других лекарственных средств угрожают здоровью пациента. Поскольку операция является инвазивной и существенно изменяющей жизнь, она является не очень желательной схемой лечения и, как правило, является последним средством лечения. Чтобы лучше понять это заболевание и, возможно, лечить его, экспериментально определили, регуляция какого гена повышена как при CD, так и при UC по сравнению с нормальной тканью.
Несмотря на вышеуказанный прогресс в исследовании иммунного расстройства существует большая потребность в дополнительных диагностических и терапевтических средствах, способных детектировать присутствие иммунных расстройств у млекопитающих и эффективно уменьшать эти расстройства. Соответственно целью настоящего изобретения является идентификация и характеризация полипептида, сверхэкспрессируемого в различных иммунных клетках, вовлеченных в различные иммунные расстройства, и применение указанного полипептида и кодирующих его нуклеиновых кислот для получения смеси химически связанных веществ, применимой в терапевтическом лечении и диагностической детекции иммунных расстройств у млекопитающих.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
A. Варианты осуществления
Настоящее изобретение относится к композициям и способам, применимым для диагностики и лечения связанного с иммунитетом заболевания у млекопитающих, включая человека. Настоящее изобретение основано на идентификации белков (включая антитела-агонисты и антагонисты), которые появляются в результате стимуляции иммунного ответа у млекопитающих. Связанные с иммунитетом заболевания можно лечить подавлением или усилением иммунного ответа. Молекулы, усиливающие иммунный ответ, стимулируют или усиливают иммунный ответ на антиген. Молекулы, стимулирующие иммунный ответ, можно применять терапевтически, когда усиление иммунного ответа является благоприятным. Альтернативно молекулы, подавляющие иммунный ответ, ослабляют или уменьшают иммунный ответ на антиген (например, нейтрализующие антитела), можно применять терапевтически, когда ослабление иммунного ответа является благоприятным (например, воспаление). Соответственно полипептиды PRO87299, их агонисты и антагонисты также применимы для получения лекарственных средств и медикаментов для лечения связанных с иммунитетом и воспалительных заболеваний. В конкретном аспекте такие лекарственные средства и медикаменты содержат терапевтически эффективное количество полипептида PRO87299, его агониста или антагониста с фармацевтически приемлемым носителем. Предпочтительно смесь является стерильной.
В дополнительном варианте осуществления изобретение относится к способу идентификации агонистов или антагонистов полипептида PRO87299, включающему контакт полипептида PRO87299 с молекулой-кандидатом и мониторирование биологической активности, опосредованной указанным полипептидом PRO87299. Предпочтительно полипептид PRO87299 представляет собой природную последовательность полипептида PRO87299. В конкретном аспекте агонист или антагонист PRO87299 представляет собой анти-PRO87299 антитело.
В другом варианте осуществления изобретение относится к смеси химически связанных веществ, содержащей полипептид PRO87299 или антитело-агонист или антагонист, связывающее полипептид, в смеси с носителем или наполнителем. В одном из аспектов композиция содержит терапевтически эффективное количество полипептида или антитела. В другом аспекте, где композиция содержит иммуностимулирующую молекулу, композиция является применимой для: (a) увеличения инфильтрации воспалительных клеток в ткань нуждающегося в этом млекопитающего, (b) стимуляции или усиления иммунного ответа у нуждающегося в этом млекопитающего, (c) увеличения пролиферации иммунных клеток в ответ на антиген у нуждающегося в этом млекопитающего, (d) стимуляции активности иммунных клеток или (e) увеличения проницаемости сосудов. В дополнительном аспекте, когда композиция содержит иммуноподавляющую молекулу, композиция является применимой для: (a) уменьшения инфильтрации воспалительных клеток в ткань нуждающегося в этом млекопитающего, (b) подавления или уменьшения иммунного ответа у нуждающегося в этом млекопитающего, (c) уменьшения активности иммунных клеток или (d) уменьшения пролиферации иммунных клеток в ответ на антиген у нуждающегося в этом млекопитающего. В другом аспекте композиция содержит дополнительный активный ингредиент, который может являться, например, дополнительным антителом или цитотоксическим или химиотерапевтическим средством. Предпочтительно композиция является стерильной.
В другом варианте осуществления изобретение относится к способу лечения связанного с иммунитетом расстройства у нуждающегося в этом млекопитающего, включающему введение млекопитающему эффективного количества полипептида PRO87299, его агониста или его антагониста. В предпочтительном аспекте связанное с иммунитетом расстройство выбрано из группы, состоящей из: системной красной волчанки, ревматоидного артрита, остеоартрита, ювенильного хронического артрита, спондилоартропатий, системного склероза, идиопатических воспалительных миопатий, синдрома Шегрена, системного васкулита, саркоидоза, аутоиммунной гемолитической анемии, аутоиммунной тромбоцитопении, тиреоидита, сахарного диабета, иммуноопосредованного заболевания почек, демиелинизирующих заболеваний центральной и периферической нервных систем, таких как рассеянный склероз, идиопатическая демиелинизирующая полиневропатия, или синдром Гийена-Барре, и хроническая воспалительная демиелинизирующая полиневропатия, гепатобилиарных заболеваний, таких как инфекционный, аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит и склерозирующий холангит, воспалительного заболевания кишечника, глютензависимой энтеропатии и болезни Уиппла, аутоиммунных или иммуноопосредованных заболеваний кожи, полиморфной эритемы и контактного дерматита, псориаза, аллергических заболеваний, таких как астма, аллергический ринит, атопический дерматит, гиперчувствительность к пище и уртикария, иммунологических заболеваний легкого, таких как эозинофильные пневмонии, идиопатический фиброз легких и гиперчувствительный пневмонит, связанных с трансплантацией заболеваний, включая отторжение трансплантата и реакцию «трансплантат против хозяина».
В другом варианте осуществления изобретение относится к антителу, специфически связывающемуся с любым из описанных выше или ниже полипептидов. Не обязательно, антитело представляет собой моноклональное антитело, гуманизированное антитело, фрагмент антитела или одноцепочечное антитело. В одном из аспектов настоящее изобретение относится к выделенному антителу, связывающему полипептид PRO87299. В другом аспекте антитело мимикрирует активность полипептида PRO87299 (антитело-агонист) или, напротив, антитело ингибирует или нейтрализует активность полипептида PRO87299 (антитело-антагонист). В другом аспекте антитело представляет собой моноклональное антитело, предпочтительно обладающее не относящимися к человеку остатками определяющей комплементарность области (CDR) и относящимися к человеку остатками каркасной области (FR). Антитело может являться меченым и может являться иммобилизованным на твердой подложке. В дополнительном аспекте антитело представляет собой фрагмент антитела, моноклональное антитело, одноцепочечное антитело или антиидиотипическое антитело.
В другом варианте осуществления настоящее изобретение относится к композиции, содержащей анти-PRO87299 антитело в смеси с фармацевтически приемлемым носителем. В одном из аспектов композиция содержит терапевтически эффективное количество антитела. Предпочтительно композиция является стерильной. Композицию можно вводить в форме жидкого фармацевтического состава, который можно консервировать для достижения увеличенной стабильности при хранении. Альтернативно антитело представляет собой моноклональное антитело, фрагмент антитела, гуманизированное антитело или одноцепочечное антитело.
В дополнительном варианте осуществления изобретение относится к изделию, содержащему:
(a) смесь химически связанных веществ, содержащую полипептид PRO87299 или его агонист или антагонист;
(b) контейнер, содержащий указанную композицию; и
(c) ярлык, прикрепленный к указанному контейнеру, или вкладыш в упаковку, содержащийся в указанном контейнере, относящиеся к применению указанного полипептида PRO87299 или его агониста или антагониста в лечении связанного с иммунитетом заболевания. Композиция может содержать терапевтически эффективное количество полипептида PRO87299 или его агониста или антагониста.
В другом варианте осуществления настоящее изобретение относится к способу диагностики связанного с иммунитетом заболевания у млекопитающего, включающему детекцию уровня экспрессии гена, кодирующего полипептид PRO87299, (a) в тестируемом образце клеток ткани, полученном от млекопитающего, и (b) в контрольном образце известных нормальных клеток ткани того же типа клеток, где более высокий или более низкий уровень экспрессии в тестируемом образце по сравнению с контрольным образцом указывает на присутствие связанного с иммунитетом заболевания у млекопитающего, от которого получены тестируемые клетки ткани.
В другом варианте осуществления настоящее изобретение относится к способу диагностики иммунного заболевания у млекопитающего, включающему (a) контакт анти-PRO87299 антитела с тестируемым образцом клеток ткани, полученным от млекопитающего, и (b) детекцию формирования комплекса между антителом и полипептидом PRO87299 в тестируемом образце; где формирование указанного комплекса указывает на присутствие или отсутствие указанного заболевания. Детекция может являться качественной или количественной, и ее можно проводить в сравнении с мониторингом формирования комплекса в контрольном образце известных нормальных клеток ткани того же типа клеток. Увеличенное количество комплексов в тестируемом образце указывает на присутствие или отсутствие иммунного заболевания у млекопитающего, от которого получены тестируемые клетки ткани. Антитело предпочтительно несет поддающуюся детекции метку. Формирование комплексов можно мониторировать, например, световой микроскопией, проточной цитометрией, флуорометрией или другими способами, известными в данной области. Тестируемый образец обычно получают от индивидуума, предположительно обладающего дефицитом или патологией иммунной системы.
В другом варианте осуществления изобретение относится к способу определения присутствия полипептида PRO87299 в образце, включающему воздействие на тестируемый образец клеток, предположительно содержащих полипептид PRO87299, анти-PRO87299 антителом и определения связывания указанного антитела с указанным образцом клеток. В конкретном аспекте образец содержит клетку, предположительно содержащую полипептид PRO87299, и антитело связывается с клеткой. Антитело предпочтительно является меченным поддающейся детекции меткой и/или связанным с твердой подложкой.
В другом варианте осуществления настоящее изобретение относится к диагностическому набору для связанного с иммунитетом заболевания, содержащему анти-PRO87299 антитело и носитель в соответствующей упаковке. Набор предпочтительно содержит инструкции для применения антитела для детекции присутствия полипептида PRO87299. Предпочтительно носитель является фармацевтически приемлемым.
В другом варианте осуществления настоящее изобретение относится к диагностическому набору, содержащему анти-PRO87299 антитело в соответствующей упаковке. Набор предпочтительно содержит инструкции для применения антитела для детекции присутствия полипептида PRO87299.
В другом варианте осуществления изобретение относится к способу диагностики связанного с иммунитетом заболевания у млекопитающего, включающему детекцию присутствия или отсутствия полипептида PRO87299 в тестируемом образце клеток ткани, полученном от указанного млекопитающего, где присутствие или отсутствие полипептида PRO87299 в тестируемом образце указывает на присутствие связанного с иммунитетом заболевания у указанного млекопитающего.
В другом варианте осуществления настоящее изобретение относится к способу идентификации агониста полипептида PRO87299, включающему:
(a) контакт клеток и тестируемого соединения, подлежащего скринингу, в условиях, подходящих для индукции клеточного ответа, нормально индуцируемого полипептидом PRO87299; и
(b) определение индукции указанного клеточного ответа для определения, является ли тестируемое соединение эффективным агонистом, где индукция указанного клеточного ответа указывает, что указанное тестируемое соединение является эффективным агонистом.
В другом варианте осуществления изобретение относится к способу идентификации соединения, способного ингибировать активность полипептида PRO87299, включающему контакт соединения-кандидата с полипептидом PRO87299 в адекватных условиях и в течение достаточного времени, чтобы позволить взаимодействовать данным двум компонентам, и определение, ингибирована ли активность полипептида PRO87299. В конкретном аспекте или соединение-кандидат, или полипептид PRO87299 иммобилизуют на твердой подложке. В другом аспекте неиммобилизованный компонент обладает поддающейся детекции меткой. В предпочтительном аспекте данный способ включает стадии:
(a) контакта клеток и тестируемого соединения, подлежащего скринингу, в присутствии полипептида PRO87299 в условиях, подходящих для индукции клеточного ответа, нормально индуцируемого полипептидом PRO87299; и
(b) определения индукции указанного клеточного ответа для определения, является ли тестируемое соединение эффективным антагонистом.
В другом варианте осуществления изобретение относится к способу идентификации соединения, ингибирующего экспрессию полипептида PRO87299 в клетках, нормально экспрессирующих полипептид, где способ включает приведение клеток в контакт с тестируемым соединением и определение, ингибирована ли экспрессия полипептида PRO87299. В предпочтительном аспекте данный способ включает стадии:
(a) контакта клеток и тестируемого соединения, подлежащего скринингу, в условиях, подходящих, чтобы позволять экспрессию полипептида PRO87299; и
(b) определения ингибирования экспрессии указанного полипептида.
В другом варианте осуществления настоящее изобретение относится к способу лечения связанного с иммунитетом расстройства у страдающего им млекопитающего, включающему введение млекопитающему молекулы нуклеиновой кислоты, кодирующей или (a) полипептид PRO87299, (b) агонист полипептида PRO87299, или (c) антагонист полипептида PRO87299, где указанный агонист или антагонист может представлять собой анти-PRO87299 антитело. В предпочтительном варианте осуществления млекопитающее представляет собой человека. В другом предпочтительном варианте осуществления нуклеиновую кислоту вводят посредством генной терапии ex vivo. В дополнительном предпочтительном варианте осуществления нуклеиновую кислоту содержит вектор, более предпочтительно аденовирусный, аденоассоциированный вирусный, лентивирусный или ретровирусный вектор.
В другом аспекте изобретение относится к рекомбинантной вирусной частице, содержащей вирусный вектор, по существу состоящий из промотора, нуклеиновой кислоты, кодирующей (a) полипептид PRO87299, (b) полипептид-агонист полипептида PRO87299 или (c) полипептид-антагонист полипептида PRO87299, и сигнальную последовательность для клеточной секреции полипептида, где вирусный вектор связан с вирусными структурными белками. Предпочтительно сигнальная последовательность относится к млекопитающим, такая как из природного полипептида PRO87299.
В дополнительном варианте осуществления изобретение относится к продуцирующей ex vivo клетке, содержащей конструкцию нуклеиновой кислоты, экспрессирующую ретровирусные структурные белки и содержащую также ретровирусный вектор, по существу состоящий из промотора, нуклеиновой кислоты, кодирующей (a) полипептид PRO87299, (b) полипептид-агонист полипептида PRO87299 или (c) полипептид-антагонист полипептида PRO87299, и сигнальную последовательность для клеточной секреции полипептида, где указанная продуцирующая клетка упаковывает ретровирусный вектор совместно со структурными белками для получения рекомбинантных ретровирусных частиц.
В дополнительном варианте осуществления изобретение относится к способу увеличения активности иммунных клеток у млекопитающего, включающему введение указанному млекопитающему (a) полипептида PRO87299, (b) агониста полипептида PRO87299 или (c) антагониста полипептида PRO87299, где активность иммунных клеток у млекопитающего повышается.
В дополнительном варианте осуществления изобретение относится к способу увеличения пролиферации иммунных клеток у млекопитающего, включающему введение указанному млекопитающему (a) полипептида PRO87299, (b) агониста полипептида PRO87299 или (c) антагониста полипептида PRO87299, где пролиферация иммунных клеток у млекопитающего увеличивается.
В дополнительном варианте осуществления изобретение относится к способу уменьшения пролиферации иммунных клеток у млекопитающих, включающему введение указанному млекопитающему (a) полипептида PRO87299, (b) агониста полипептида PRO87299 или (c) антагониста полипептида PRO87299, где пролиферация иммунных клеток у млекопитающего уменьшается.
B. Дополнительные варианты осуществления
В других вариантах осуществления по настоящему изобретению изобретение относится к векторам, содержащим ДНК, кодирующую любой из описанных здесь полипептидов. Предоставлена также клетка-хозяин, содержащая любой такой вектор. В качестве примера, клетки-хозяева могут представлять собой клетки CHO, E. coli или дрожжей. Дополнительно представлен способ получения любого из описанных здесь полипептидов, включающий культивирование клеток-хозяев в условиях, подходящих для экспрессии желаемого полипептида, и выделение желаемого полипептида из культуры клеток.
В других вариантах осуществления изобретение относится к химерным молекулам, содержащим любой из описанных здесь полипептидов, слитый с гетерологичной полипептидной или аминокислотной последовательностью. Примеры таких химерных молекул содержат любой из описанных здесь полипептидов, слитый с последовательностью эпитопа-метки или Fc-областью иммуноглобулина.
В другом варианте осуществления изобретение относится к антителу, специфически связывающему любой из описанных выше или ниже полипептидов. Не обязательно, антитело представляет собой моноклональное антитело, гуманизированное антитело, фрагмент антитела или одноцепочечное антитело.
В других вариантах осуществления изобретение относится к олигонуклеотидным зондам, применимым для выделения нуклеотидных последовательностей, геномных и кДНК, или в качестве антисмысловых зондов, где данные зонды можно получить из любой из описанных выше или ниже нуклеотидных последовательностей.
В других вариантах осуществления изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую полипептид PRO87299.
В одном из аспектов выделенная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, обладающую, по меньшей мере, приблизительно 80% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 81% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 82% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 83% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 84% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 85% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 86% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 87% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 88% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 89% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 90% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 91% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 92% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 93% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 94% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 95% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 96% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 97% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 98% идентичностью последовательности нуклеиновой кислоты и альтернативно, по меньшей мере, приблизительно 99% идентичностью последовательности нуклеиновой кислоты с (a) молекулой ДНК, кодирующей полипептид PRO87299, обладающий полноразмерной аминокислотной последовательностью, как описано здесь, аминокислотную последовательностью без сигнального пептида, как описано здесь, внеклеточный домен трансмембранного белка, с сигнальным пептидом или без, как описано здесь, или любой другой конкретно определенный фрагмент полноразмерной аминокислотной последовательности, как описано здесь, или (b) молекулой ДНК, комплементарной (a).
В других аспектах выделенная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, обладающую, по меньшей мере, приблизительно 80% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 81% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 82% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 83% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 84% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 85% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 86% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 87% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 88% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 89% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 90% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 91% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 92% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 93% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 94% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 95% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 96% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 97% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 98% идентичностью последовательности нуклеиновой кислоты и альтернативно, по меньшей мере, приблизительно 99% идентичностью последовательности нуклеиновой кислоты с (a) молекулой ДНК, содержащей кодирующую последовательность кДНК полноразмерного полипептида PRO87299, как описано здесь, кодирующую последовательность полипептида PRO87299 без сигнального пептида, как описано здесь, кодирующую последовательность внеклеточного домена трансмембранного полипептида PRO87299 с сигнальным пептидом или без, как описано здесь, или кодирующую последовательность любого другого конкретно определенного фрагмента полноразмерной аминокислотной последовательности как описано здесь, или (b) молекулой, комплементарной ДНК (a).
В дополнительном аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, обладающую, по меньшей мере, приблизительно 80% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 81% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 82% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 83% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 84% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 85% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере приблизительно 86% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 87% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 88% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 89% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 90% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 91% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 92% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 93% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 94% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 95% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 96% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 97% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 98% идентичностью последовательности нуклеиновой кислоты и альтернативно, по меньшей мере, приблизительно 99% идентичностью последовательности нуклеиновой кислоты с молекулой ДНК, кодирующей такой же зрелый полипептид, как показано на фиг. 2 (SEQ ID NO:2).
В другом аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую полипептид PRO87299, который обладает или делетированным трансмембранным доменом, или инактивированным трансмембранным доменом, или являющейся комплементарной такой кодирующей нуклеотидной последовательностью, где трансмембранный домен(ы) такого полипептида описаны здесь. Таким образом, рассматривают растворимые внеклеточные домены полипептидов PRO87299.
Другой вариант осуществления относится к последовательности, кодирующей фрагменты полипептида PRO87299 или комплементраной ей, которую можно применять, например, в качестве зондов для гибридизации, для кодирования фрагментов полипептида PRO87299, которые могут, не обязательно, кодировать полипептид, содержащий участок связывания для антитела анти-PRO87299 или в качестве антисмысловых олигонуклеотидных зондов. Такие фрагменты нуклеиновой кислоты обычно обладают длиной, по меньшей мере, приблизительно 20 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 30 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 40 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 50 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 60 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 70 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 80 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 90 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 100 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 110 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 120 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 130 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 140 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 150 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 160 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 170 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 180 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 190 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 200 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 250 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 300 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 350 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 400 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 450 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 500 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 600 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 700 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 800 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 900 нуклеотидов и альтернативно длиной, по меньшей мере, приблизительно 1000 нуклеотидов, где в данном контексте термин «приблизительно» означает указанную длину нуклеотидной последовательности плюс или минус 10% данной указанной длины. Отмечают, что новые фрагменты нуклеотидной последовательности, кодирующей полипептид PRO87299, можно определять общепринятым способом посредством выравнивания нуклеотидной последовательности, кодирующей полипептид PRO87299, с другими известными нуклеотидными последовательностями с использованием любой из множества хорошо известных программ выравнивания последовательностей и определения, какой фрагмент(ы) нуклеотидной последовательности, кодирующей полипептид PRO87299, является новым. Все такие нуклеотидные последовательности, кодирующие полипептид PRO87299, рассматривают здесь. Также рассматривают фрагменты полипептида PRO87299, кодируемые данными фрагментами нуклеотидной молекулы, предпочтительно те фрагменты полипептида PRO87299, котрые содержат участок связывания анти-PRO87299 антитела.
В другом варианте осуществления изобретение относится к выделенному полипептиду PRO87299, кодируемому любой из выделенных последовательностей нуклеиновой кислоты, указанных здесь выше.
В конкретном аспекте изобретение относится к выделенному полипептиду PRO87299, содержащему аминокислотную последовательность, обладающую, по меньшей мере, приблизительно 80% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 81% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 82% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 83% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере,приблизительно 84% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 85% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 86% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 87% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 88% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 89% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 90% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 91% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 92% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 93% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 94% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 95% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 96% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 97% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 98% идентичностью аминокислотной последовательности и альтернативно, по меньшей мере, приблизительно 99% идентичностью аминокислотной последовательности с полипептидом PRO87299, обладающим полноразмерной аминокислотной последовательностью, как описано здесь, аминокислотной последовательностью без сигнального пептида, как описано здесь, внеклеточным доменом трансмембранного белка с сигнальным пептидом или без, как описано здесь, или любым другим конкретно определенным фрагментом полноразмерной аминокислотной последовательности, как описано здесь.
В дополнительном аспекте изобретение относится к выделенному полипептиду PRO87299, содержащему аминокислотную последовательность, обладающую, по меньшей мере, приблизительно 80% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 81% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 82% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 83% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 84% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 85% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 86% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 87% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 88% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 89% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 90% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 91% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 92% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 93% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 94% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 95% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 96% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 97% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 98% идентичностью аминокислотной последовательности и альтернативно, по меньшей мере, приблизительно 99% идентичностью аминокислотной последовательности с аминокислотной последовательностью, показанной на фиг. 2 (SEQ ID NO:2).
В конкретном аспекте изобретение относится к выделенному полипептиду PRO87299 без N-концевой сигнальной последовательности и/или начального метионина и кодируемому нуклеотидной последовательностью, которая кодирует такую аминокислотную последовательность, как описанная здесь ранее. Описаны также способы получения полипептида, где данные способы включают культивирование клетки-хозяина, содержащей нуклеиновую кислоту, в условиях, подходящих для экспрессии полипептида PRO87299, и выделения полипептида PRO87299 из культуры клеток.
В другом аспекте изобретение относится к выделенному полипептиду PRO87299, обладающему или делетированным трансмембранным доменом, или инактивированным трансмембранным доменом. Здесь описаны также способы для получения полипептида, где данные способы включают культивирование клетки-хозяина, содержащей вектор, содержащий подходящую кодирующую молекулу нуклеиновой кислоты в условиях, подходящих для экспрессии полипептида PRO87299 и выделения полипептида PRO87299 из культуры клеток.
В другом варианте осуществления изобретение относится к агонистам и антагонистам природного полипептида PRO87299, как описано здесь. В конкретном варианте осуществления агонист или антагонист представляет собой анти-PRO87299 антитело или малую молекулу.
В другом варианте осуществления изобретение относится к способу идентификации агонистов или антагонистов полипептида PRO87299, который включает приведение в контакт полипептида PRO87299 с молекулой-кандидатом и мониторирование биологической активности, опосредованной указанным полипептидом PRO87299. Предпочтительно полипептид PRO87299 представляет собой нативный полипептид PRO87299.
В дополнительном варианте осуществления изобретение относится к смеси химически связанных веществ, содержащей полипептид PRO87299, или агонист, или антагонист полипептида PRO87299, как описано здесь, или анти-PRO87299 антитело в сочетании с носителем. Не обязательно, носитель представляет собой фармацевтически приемлемый носитель.
Другой вариант осуществления настоящего изобретения относится к применению полипептида PRO87299 или его агониста или антагониста, как описано здесь ранее, или анти-PRO87299 антитела, для получения лекарственного средства, применимого для лечения состояния, отвечающего на полипептид PRO87299, его агонист или антагонист, или на анти-PRO87299 антитело.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана нуклеотидная последовательность (SEQ ID NO:1) природной последовательности кДНК PRO87299, где SEQ ID NO:1 представляет собой клон, обозначенный здесь «ДНК332467».
На фиг.2 показана аминокислотная последовательность (SEQ ID NO:2), полученная с кодирующей последовательности SEQ ID NO:1, показанной на фиг.1.
На фиг.3 показана нуклеотидная последовательность (SEQ ID NO:3) природной последовательности кДНК HVEM (HVEM), где SEQ ID NO:3 представляет собой клон, обозначенный здесь «HVEM».
На фиг.4 показана аминокислотная последовательность (SEQ ID NO:4), полученная из кодирующей последовательности SEQ ID NO:3, показанной на фиг. 3.
На фиг.5 показана нуклеотидная последовательность (SEQ ID NO:5) природной последовательности LIGHT, где SEQ ID NO:5 представляет собой клон, обозначенный здесь «LIGHT».
На фиг.6 показана аминокислотная последовательность (SEQ ID NO:6), выведенная из кодирующей последовательности SEQ ID NO:5, показанной на фиг. 5.
На фиг.7 показана нуклеотидная последовательность (SEQ ID NO:7) варианта последовательности PRO87299, где SEQ ID NO:7 представляет собой клон, обозначенный здесь «PRO87299.short».
На фиг.8 показана аминокислотная последовательность (SEQ ID NO:8), выведенная из кодирующей последовательности SEQ ID NO:7, показанной на фиг.7.
На фиг.9 показана нуклеотидная последовательность (SEQ ID NO:9) варианта последовательности PRO87299, где SEQ ID NO:9 представляет собой клон, обозначенный здесь «PRO87299.AFTNIP».
На фиг.10 показана аминокислотная последовательность (SEQ ID NO: 10), выведенная из кодирующей последовательности SEQ ID NO:9, показанной на фиг.9.
На фиг.11 показаны варианты нуклеиновой кислоты кДНК PRO87299.
На фиг.12 показаны варианты трансляции полипептида PRO87299.
На фиг.13 показано ингибирование пролиферации CD4+ T-клеток посредством антител-агонистов.
На фиг.14 показано специфическое связывание PRO87299 с HVEM.
На фиг.15 показано связывание PRO87299 с HVEM при сравнении с другими членами семейства.
На фиг.16 показано, что связывание PRO87299 с HVEM является чувствительным к pH.
На фиг.17 показано связывание PRO87299 с HVEM в клетках, трансфицированных HVEM.
На фиг.18 показано, что антитела к PRO87299 могут блокировать взаимодействие с HVEM.
На фиг.19 показано, что PRO87299 и LIGHT могут одновременно связывать HVEM
На фиг.20 показано, что PRO87299 не блокирован взаимодействием LIGHT с HVEM.
На фиг.21 показаны пептид BP-2 и gD, блокирующие взаимодействие PRO87299/HVEM.
На фиг.22 показан ингибирующий эффект PRO87299 на CD4+ T-клетки.
На фиг.23 показано, что активация PRO87299 HVEM-Fc способствует выживанию в модели GVHR.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
I. Определения
Полипептиды PRO87299, описанные здесь, можно выделить из множества источников, таких как типы тканей человека, или из другого источника или получить рекомбинантным или синтетическим способами. Как описано в данном описании, когда ссылаются на «полипептид PRO87299», ссылаются на каждый из полипептидов отдельно, так же как совокупно. Например, описания получения, очистки, выведения, получения антител к или против введения композиций, их содержащих, лечения заболевания с помощью, и т.д., относятся к каждому полипептиду по изобретению индивидуально. Термин «полипептид PRO87299» также включает варианты полипептидов PRO87299, описанные здесь.
«Природная последовательность полипептида PRO87299» включает в себя полипептид, обладающий такой же аминокислотной последовательностью, как соответствующий полипептид PRO87299, полученный из природных источников. Такие природные последовательности полипептидов PRO87299 можно выделить из природных источников или получить рекомбинантными или синтетическими способами. Термин «природная последовательность полипептида PRO87299» конкретно охватывает существующие в природе укороченные или секретируемые формы конкретного полипептида PRO87299 (например, последовательность внеклеточного домен), варианты существующих в природе форм (например, форм альтернативного сплайсинга) и существующие в природе аллельные варианты полипептида. В различных вариантах осуществления по изобретению природные последовательности полипептидов PRO87299, описанные здесь, представляют собой зрелые или полноразмерные природные последовательности полипептидов, содержащие полноразмерные аминокислотные последовательности, показанные на сопутствующих фигурах. Старт- и стоп-кодоны на фигурах показаны жирным шрифтом и подчеркнуты. Однако в то время как показано, что полипептид PRO87299, описанный в сопутствующих фигурах, начинается с остатков метионина, обозначенного здесь на фигурах как положение аминокислоты 1, является предположительным и возможным, что другие остатки метионина, локализованные выше или ниже положения аминокислоты 1 на фигурах, могут служить начальным аминокислотным остатком для полипептидов PRO87299.
«Внеклеточный домен», или «ECD», полипептида PRO87299 относится к форме полипептида PRO87299, являющейся в основном свободной от трансмембранного и цитоплазматического доменов. Обычно полипептид PRO87299 ECD будет обладать менее чем 1% таких трансмембранных и/или цитоплазматических доменов и предпочтительно будет обладать менее чем 0,5% таких доменов. Понятно, что любые трансмембранные домены, идентифицированные для полипептидов PRO87299 по настоящему изобретению, идентифицированы в соответствии с критериями, обычно используемыми в данной области для идентификации гидрофобного домена такого типа. Точные границы трансмембранного домена могут меняться, но наиболее вероятно, не более чем приблизительно на 5 аминокислот на любом конце домена, как первоначально идентифицировали здесь. Таким образом, не обязательно, внеклеточный домен полипептида PRO87299 может содержать от приблизительно 5 или менее аминокислот на любой стороне границы трансмембранный домен/внеклеточный домен, как указано в примерах или описании, и такие полипептиды, с присоединенным сигнальным пептидом или без и кодирующая их нуклеиновая кислота относятся к настоящему изобретению.
«Вариант полипептида PRO87299» означает активный полипептид PRO87299, как определено выше или ниже, обладающий, по меньшей мере, приблизительно 80% идентичностью аминокислотной последовательности с полноразмерной природной последовательностью полипептида PRO87299, как описано здесь, последовательностью полипептида PRO87299 без сигнального пептида, как описано здесь, внеклеточный домен полипептида PRO87299 с сигнальным пептидом или без, как описано здесь, или любой другой фрагмент полноразмерной последовательности полипептида PRO87299, как описано здесь. Такие варианты полипептида PRO87299 включают, например, полипептиды PRO87299, где добавлены или делетированы один или несколько аминокислотных остатков на N- или C-конце полноразмерной природной аминокислотной последовательности. Обычно вариант полипептида PRO87299 будет обладать, по меньшей мере, приблизительно 80% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 81% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 82% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 83% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 84% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 85% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 86% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 87% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 88% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 89% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 90% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 91% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 92% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 93% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 94% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 95% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 96% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 97% идентичностью аминокислотной последовательности, альтернативно, по меньшей мере, приблизительно 98% идентичностью аминокислотной последовательности и альтернативно, по меньшей мере, приблизительно 99% идентичностью аминокислотной последовательности с полноразмерной природной последовательностью полипептида PRO87299, как описано здесь, последовательностью полипептида PRO87299 без сигнального пептида, как описано здесь, внеклеточным доменом полипептида PRO87299 с сигнальным пептидом или без, как описано здесь, или любым другим конкретно определенным фрагментом полноразмерной последовательности полипептида PRO87299, как описано здесь. Обычно варианты полипептидов PRO87299 обладают длиной, по меньшей мере, приблизительно 10 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 20 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 30 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 40 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 50 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 60 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 70 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 80 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 90 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 100 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 150 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 200 аминокислот, альтернативно длиной, по меньшей мере, приблизительно 300 аминокислот, или более.
«Процент (%) идентичности аминокислотной последовательности» по отношению к последовательностям полипептида PRO87299, указанным в настоящем документе, определяют как процент аминокислотных остатков в последовательности-кандидате, идентичных с аминокислотными остатками конкретной последовательности полипептида PRO87299, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимального процента идентичности последовательности, и не рассматривая никаких консервативных замен как часть идентичности последовательности. Выравнивание для определения процента идентичности аминокислотной последовательности можно выполнять различными способами, известными специалистам в данной области, например с использованием общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить подходящие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по полной длине сравниваемых последовательностей. Однако для целей здесь величины % идентичности аминокислотных последовательностей получали с применением компьютерной программы сравнения последовательностей ALIGN-2, где полный исходный код программы ALIGN-2 приведен в таблице 1 ниже. Авторство компьютерной программы сравнения последовательностей ALIGN-2 принадлежит Genentech, Inc., и исходный код, представленный в таблице 1 ниже, подан с пользовательской документацией в Бюро регистрации авторских прав США, Washington D.C., 20559, где зарегистрирован под номером регистрации авторского права США No. TXU510087. Программа ALIGN-2 общедоступна из Genentech, Inc., South San Francisco, California или может быть скомпилирована из исходного кода, представленного в таблице 1 ниже. Программу ALIGN-2 следует компилировать для применения в операционной системе UNIX, предпочтительно digital UNIX V4.0D. Все параметры сравнения последовательностей установлены в программе ALIGN-2 и не меняются.
В ситуациях, где ALIGN-2 применяют для сравнений аминокислотной последовательности, % идентичности аминокислотной последовательности данной аминокислотной последовательности A к, с или против данной аминокислотной последовательности B (что альтернативно можно выразить как данную аминокислотную последовательность A, которая имеет или содержит конкретный % аминокислотной идентичности к, с или против данной аминокислотной последовательности B) вычисляют следующим образом:
100 умножить на дробь X/Y
где X представляет собой число аминокислотных остатков, отмеченных как идентичные совпадения программой сравнения последовательностей ALIGN-2 при сравнении этой программой A и B, где Y представляет собой общее число аминокислотных остатков в B. Следует учитывать, что где длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотной последовательности A с B не будет равен % идентичности аминокислотной последовательности B с A. В качестве примеров вычислений % идентичности аминокислотной последовательности с использованием данного способа в таблицах 2 и 3 показано, как вычислять % идентичности аминокислотной последовательности аминокислотной последовательности, обозначенной «Сравниваемый белок», с аминокислотной последовательностью, обозначенной «PRO87299», где «PRO87299» представляет собой аминокислотную последовательность интересующего гипотетического полипептида PRO87299, «Сравниваемый белок» представляет собой аминокислотную последовательность полипептида, против которого сравнивают интересующий полипептид «PRO87299», и каждый из «X», «Y» и «Z» представляют собой различные гипотетические аминокислотные остатки.
Пока конкретно не указано иначе, все значения % идентичности аминокислотной последовательности, применяемые здесь, получены, как описано в непосредственно предшествующем абзаце с применением компьютерной программы ALIGN-2. Однако величины % идентичности аминокислотной последовательности можно получить так же, как описано ниже, с использованием компьютерной программы WU-BLAST-2 (Altschul et al, Methods in Enzvmology 266:460-480 (1996)). Большинство параметров поиска WU-BLAST-2 установлено на значения по умолчанию. Для параметров, не установленных на значения по умолчанию, т.е. регулируемых параметров, устанавливали следующие значения: overlap span = 1, overlap fraction = 0,125, word threshold (T) = 11, и scoring matrix = BLOSUM62. Когда применяют WU-BLAST-2, значение % идентичности аминокислотной последовательности определяют делением (a) числа совпадающих идентичных аминокислотных остатков между аминокислотной последовательностью интересующего полипептида PRO87299, обладающего последовательностью, полученной из природного полипептида PRO87299, и сравниваемой интересующей аминокислотной последовательностью (т.е. последовательностью, против которой сравнивают интересующий полипептид PRO87299, который может представлять собой вариант полипептида PRO87299), как определено посредством WU-BLAST-2 на (b) общее число аминокислотных остатков интересующего полипептида PRO87299. Например, в утверждении «полипептид, содержащий аминокислотную последовательность A, которая обладает, или обладающую, по меньшей мере, 80% идентичностью аминокислотной последовательности с аминокислотной последовательностью B», аминокислотная последовательность A представляет собой сравниваемую интересующую аминокислотную последовательность, а аминокислотная последовательность B представляет собой аминокислотную последовательность интересующего полипептида PRO87299.
Процент идентичности аминокислотной последовательности можно определить также с использованием программы для сравнения последовательностей NCBI-BLAST2 (Altschul et al., Nucleic acids Res. 25:3389-3402 (1997)). Программу для сравнения последовательностей NCBI-BLAST2 можно загрузить с http://www.ncbi.nlm.nih.gov или другим способом получить из National Institute of Health, Bethesda, MD. В NCBI-BLAST2 используют несколько параметров поиска, где все эти параметры поиска устанавливают на значения по умолчанию, включая, например, unmask = yes, strand = all, expected occurrences = 10, minimum low complexity length = 15/5, multi-pass e-value = 0,01, constant for multi-pass = 25, dropoff for final gapped alignment = 25 и scoring matrix = BLOSUM62.
В ситуациях, где для сравнения аминокислотных последовательностей применяют NCBI-BLAST2, % идентичности аминокислотной последовательности данной аминокислотной последовательности A к, с, или против данной аминокислотной последовательности B (что альтернативно можно выразить как данную аминокислотную последовательность A, которая имеет или содержит конкретный % идентичности аминокислотной последовательности к, с или против данной аминокислотной последовательности B) вычисляют следующим образом:
100 умножить на дробь X/Y
где X представляет собой число аминокислотных остатков, отмеченных как идентичные совпадения программой сравнения последовательностей NCBI-BLAST2 при сравнении этой программой A и B, где Y представляет собой общее число аминокислотных остатков в B. Следует учитывать, что где длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотной последовательности A с B не будет равен % идентичности аминокислотной последовательности B с A.
«Вариант полинуклеотида RO87299» или «вариант последовательности нуклеиновой кислоты PRO87299» означает молекулу нуклеиновой кислоты, кодирующую активный полипептид PRO87299, как определено ниже, обладающую, по меньшей мере, приблизительно 80% идентичностью последовательности нуклеиновой кислоты с последовательностью нуклеиновой кислоты, кодирующей полноразмерную природную последовательность полипептида PRO87299, как описано здесь, полноразмерную природную последовательность полипептида PRO87299 без сигнального пептида, как описано здесь, внеклеточный домен полипептида PRO87299, с сигнальным пептидом или без, как описано здесь, или любой другой фрагмент последовательности полноразмерного полипептида PRO87299, как описано здесь. Обычно вариант полинуклеотида PRO87299 будет обладать, по меньшей мере, приблизительно 80% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 81% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 82% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 83% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 84% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 85% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 86% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 87% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 88% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 89% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 90% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 91% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 92% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 93% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 94% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 95% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 96% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 97% идентичностью последовательности нуклеиновой кислоты, альтернативно, по меньшей мере, приблизительно 98% идентичностью последовательности нуклеиновой кислоты и альтернативно, по меньшей мере, приблизительно 99% идентичностью последовательности нуклеиновой кислоты с последовательностью нуклеиновой кислоты, кодирующей последовательность полноразмерного природного полипептида PRO87299, как описано здесь, последовательность полноразмерного природного полипептида PRO87299 без сигнального пептида, как описано здесь, внеклеточный домен полипептида PRO87299 с сигнальной последовательностью или без, как описано здесь, или любой другой фрагмент последовательности полноразмерного полипептида PRO87299, как описано здесь. Варианты не охватывают природную нуклеотидную последовательность.
Обычно варианты полинуклеотида PRO87299 обладают длиной, по меньшей мере, приблизительно 30 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 60 нуклеотидов, альтернативно, по меньшей мере, приблизительно 90 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 120 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 150 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 180 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 210 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 240 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 270 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 300 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 450 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 600 нуклеотидов, альтернативно длиной, по меньшей мере, приблизительно 900 нуклеотидов или более.
«Процент (%) идентичности последовательности нуклеиновой кислоты» по отношению к последовательности нуклеиновой кислоты, кодирующей PRO87299, определяемый здесь, определяют как процент нуклеотидов в последовательности-кандидате, идентичных с нуклеотидами в интересующей последовательности нуклеиновой кислоты PRO87299 после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимального процента идентичности последовательности. Выравнивание для целей определения процента идентичности последовательности нуклеиновой кислоты можно выполнять различными способами, известными в данной области, например с использованием публично доступного компьютерного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Для целей здесь, однако, величины % идентичности последовательности нуклеиновой кислоты получали с использованием компьютерной программы сравнения последовательности ALIGN-2, где полный исходный код для программы ALIGN-2 представлен в таблице 1 ниже. Авторство компьютерной программы сравнения последовательностей ALIGN-2 принадлежит Genentech, Inc., и исходный код, представленный в таблице 1 ниже, подан с пользовательской документацией в Бюро регистрации авторских прав США, Washington D.C., 20559, где зарегистрирован под номером регистрации авторского права США No. TXU510087. Программа ALIGN-2 общедоступна из Genentech, Inc., South San Francisco, California или может быть скомпилирована из исходного кода, представленного в таблице 1 ниже. Программу ALIGN-2 следует компилировать для применения в операционной системе UNIX, предпочтительно digital UNIX V4.0D. Все параметры сравнения последовательностей установлены в программе ALIGN-2 и не меняются.
В ситуациях, где ALIGN-2 применяют для сравнений последовательности нуклеиновой кислоты, % идентичности последовательности нуклеиновой кислоты данной последовательности нуклеиновой кислоты C к, с или против данной последовательности нуклеиновой кислоты D (что альтернативно можно выразить как данную последовательность нуклеиновой кислоты C, которая имеет или содержит конкретный % идентичности последовательности нуклеиновой кислоты к, с или против данной последовательности нуклеиновой кислоты D) вычисляют следующим образом:
100 умножить на дробь W/Z
где W представляет собой число нуклеотидов, отмеченных как идентичные совпадения программой сравнения последовательностей ALIGN-2 при сравнении этой программой C и D, где Z представляет собой общее число нуклеотидов в D. Следует учитывать, что где длина последовательности нуклеиновой кислоты C не равна длине последовательности нуклеиновой кислоты D, % идентичности последовательности нуклеиновой кислоты C с D не будет равен % идентичности последовательности нуклеиновой кислоты D с C. В качестве примеров вычислений % идентичности последовательности нуклеиновой кислоты в таблицах 4 и 5 показано, как вычислять % идентичности последовательности нуклеиновой кислоты, обозначенной «Сравниваемая ДНК», с последовательностью нуклеиновой кислоты, обозначенной «PRO87299-ДНК», где «PRO87299-ДНК» представляет собой последовательность интересующей гипотетической нуклеиновой кислоты, кодирующей PRO87299, «Сравниваемая ДНК» представляет собой нуклеотидную последовательность молекулы нуклеиновой кислоты, против которого сравнивают интересующую молекулу нуклеиновой кислоты «PRO87299-ДНК», и каждый из «N», «L» и «V» представляют собой различные гипотетические нуклеотиды.
Если конкретно не указано иначе, все значения % идентичности последовательности нуклеиновой кислоты, применяемые здесь, получены, как описано в непосредственно предшествующем абзаце с применением компьютерной программы ALIGN-2. Однако величины % идентичности последовательности нуклеиновой кислоты можно получить так же, как описано ниже, с использованием компьютерной программы WU-BLAST-2 (Altschul et al., Methods in Enzvmology 266:460-480 (1996)). Большинство параметров поиска WU-BLAST-2 установлено на значения по умолчанию. Для параметров, не установленных на значения по умолчанию, т.е. регулируемых параметров, устанавливали следующие значения: overlap span = 1, overlap fraction = 0,125, word threshold (T) = 11, и scoring matrix = BLOSUM62. Когда применяют WU-BLAST-2, значение % идентичности последовательности нуклеиновой кислоты определяют делением (a) числа совпадающих идентичных нуклеотидов между последовательностью нуклеиновой кислоты интересующей молекулы нуклеиновой кислоты, кодирующей полипептид PRO87299, обладающей последовательностью, полученной из природной последовательности нуклеиновой кислоты, кодирующей полипептид PRO87299, и интересующей сравниваемой молекулой нуклеиновой кислоты (т.е. последовательностью, против которой сравнивают интересующую молекулу нуклеиновой кислоты, кодирующую полипептид PRO87299, которая может представлять собой вариант полинуклеотида PRO87299), как определено посредством WU-BLAST-2 на (b) общее число нуклеотидов интересующей молекулы нуклеиновой кислоты, кодирующей полипептид PRO87299. Например, в утверждении «выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты A, которая обладает, или обладающую, по меньшей мере,80% идентичностью последовательности нуклеиновой кислоты с последовательностью нуклеиновой кислоты B», последовательность нуклеиновой кислоты A представляет собой сравниваемую интересующую молекулу нуклеиновой кислоты, а последовательность нуклеиновой кислоты B представляет собой последовательность нуклеиновой кислоты интересующей молекулы нуклеиновой кислоты, кодирующей полипептид PRO87299.
Процент идентичности последовательности нуклеиновой кислоты можно определить также с использованием программы для сравнения последовательностей NCBI-BLAST2 (Altschul et al., Nucleic acids Res. 25:3389-3402 (1997)). Программу для сравнения последовательностей NCBI-BLAST2 можно загрузить с http://www.ncbi.nlm.nih.gov или другим способом получить из National Institute of Health, Bethesda, MD. В NCBI-BLAST2 спользуют несколько параметров поиска, где все эти параметры поиска устанавливают на значения по умолчанию, включая, например, unmask = yes, strand = all, expected occurrences = 10, minimum low complexity length = 15/5, multi-pass e-value = 0,01, constant for multi-pass = 25, dropoff for final gapped alignment = 25 и scoring matrix = BLOSUM62.
В ситуациях, где для сравнений последовательностей применяют NCBI-BLAST2, % идентичности последовательности нуклеиновой кислоты данной последовательности нуклеиновой кислоты C к, с, или против данной последовательности нуклеиновой кислоты D (что альтернативно можно выразить как данную последовательность нуклеиновой кислоты C, которая имеет или содержит конкретный % идентичности последовательности нуклеиновой кислоты к, с или против данной последовательности нуклеиновой кислоты D) вычисляют следующим образом:
100 умножить на дробь W/Z
где W представляет собой число нуклеотидов, отмеченных как идентичные совпадения программой сравнения последовательностей NCBI-BLAST2 при сравнении этой программой C и D, где Z представляет собой общее число нуклеотидов в D. Следует учитывать, что где длина последовательности нуклеиновой кислоты C не равна длине последовательности нуклеиновой кислоты D, % идентичности последовательности нуклеиновой кислоты C с D не будет равен % идентичности последовательности нуклеиновой кислоты D с C.
В других вариантах осуществления варианты полинуклеотида PRO87299 представляют собой молекулы нуклеиновой кислоты, кодирующей активный полипептид PRO87299 и способной гибридизоваться, предпочтительно в строгих условиях гибридизации и отмывки, с нуклеотидными последовательностями, кодирующими полноразмерный полипептид PRO87299, как описано здесь. Варианты полипептида PRO87299 могут представлять собой варианты, кодируемые вариантом полинуклеотида PRO87299.
«Выделенный» при применении здесь для описания различных полипептидов, описанных здесь, означает полипептид, который идентифицирован и отделен и/или выделен из компонента его природного окружения. Загрязняющие компоненты его натурального окружения представляют собой материалы, которые как правило, будут мешать диагностическим или терапевтическим применениям полипептида и могут включать в себя ферменты, гормоны и другие белковые или небелковые растворы. В предпочтительных вариантах осуществления полипептид будет очищен (1) до степени, достаточной для получения, по меньшей мере,15 остатков N-концевой или внутренней аминокислотной последовательности при использовании секвенатора с вращающимся стаканом, или (2) до гомогенности в SDS-PAGE в невосстанавливающих или восстанавливающих условиях с использованием окраски Кумасси синим или предпочтительно серебром. Выделенный полипептид включает в себя полипептид in situ в рекомбинантных клетках, пока не будет присутствовать, по меньшей мере, один из компонентов природного окружения полипептида PRO87299. Обычно, однако, выделенный полипептид будут получать посредством, по меньшей мере, одной стадии очистки.
«Выделенная» нуклеиновая кислота, кодирующая полипептид PRO87299 или нуклеиновая кислота, кодирующая другой полипептид, представляет собой молекулу нуклеиновой кислоты, которую идентифицируют и отделяют, по меньшей мере, от одной загрязняющей молекулы нуклеиновой кислоты, с которой она обычно связана в природном источнике нуклеиновой кислоты, кодирующей полипептид. Выделенная молекула нуклеиновой кислоты, кодирующей полипептид, является отличной от формы или окружения, в которых ее обнаружили в природе. Таким образом, выделенные молекулы нуклеиновой кислоты отличаются от молекул нуклеиновой кислоты, кодирующих конкретный полипептид, как они присутствуют в природных клетках. Однако выделенная молекула нуклеиновой кислоты, кодирующей полипептид, включает в себя молекулы нуклеиновой кислоты, кодирующие полипептид, содержащиеся в клетках, которые обычно экспрессируют полипептид, где, например, хромосомная локализация молекулы нуклеиновой кислоты отличается от локализации в природных клетках.
Термин "контрольные последовательности" относится к последовательности ДНК, необходимой для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, подходящие для прокариот, например, включают в себя последовательность промотора, необязательно, оператора и участка связывания рибосомы. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования, и энхансеры.
Нуклеиновая кислота является «функционально связанной», когда ее помещают в функциональное взаимодействие с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторный лидер является функционально связанной с ДНК для полипептида, если она экспрессируется как пребелок, который участвует в секреции полипептида; промотор или энхансер является функционально связанным с кодируюшей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосомы является функционально связанным с кодируюшей последовательностью, если он расположен так, чтобы облегчать трансляцию. Как правило, «функционально связанные» означает последовательности ДНК, связанные по соседству, и в случае секреторного лидера - по соседству и в фазе считывания. Однако энхансеры не должны находиться по соседству. Связывание осуществляют посредством лигирования в подходящие участки рестрикции. Если таких участков не существует, используют синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой.
Термин «антитело» применяют в самом широком смысле, и он конкретно охватывает, например, отдельные анти-PRO87299 моноклональные антитела (включая антитела-агонисты, антагонисты и нейтрализующие), композиции анти-PRO87299 антитела с полиэпитопной специфичностью, одноцепочечные анти-PRO87299 антитела и фрагменты анти-PRO87299 антител (см. ниже). Термин «моноклональное антитело», как применяют здесь, относится к антителу, полученному из популяции по существу однородных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах.
«Строгость» реакций гибридизации может легко определить обычный специалист в данной области, и, как правило, она представляет собой эмпирическое вычисление, зависящее от длины зонда, температуры отмывки и концентрации соли. Как правило, для более длинных зондов для правильного отжига требуются более высокие температуры, в то время как для более коротких зондов требуются более низкие температуры. Гибридизация в общем зависит от способности денатурированной ДНК к повторному отжигу с комплементарными цепями, присутствующими в окружающей среде, при температуре ниже их плавления. Чем выше желательная степень гомологии между зондом и гибридизующейся последовательностью, тем выше относительная температура, которую можно применять. В результате следует, что более высокие относительные температуры будут иметь тенденцию делать условия реакции более строгими, в то время как более низкие температуры - менее строгими. Дополнительные подробности и объяснение строгости реакций гибридизации смотри в Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995).
«Строгие условия» или «высоко строгие условия», как определяют здесь, можно идентифицировать как те, при которых: (1) применяют низкую ионную силу и высокую температуру для отмывки, например 0,015 M хлорид натрия /0,0015 M цитрат натрия /0,1% додецилсульфат натрия при 50°C; (2) во время гибридизации применяют денатурирующее средство, такое как формамид, например, 50% (об./об.) формамида с 0,1% бычьего сывороточного альбумина/0,1% фиколла/0,1% поливинилпирролидона/50 мМ натрий фосфатного буфера при pH 6,5 с 750 мМ хлорида натрия, 75 мМ цитрата натрия при 42°C; или (3) применяют 50% формамид, 5×SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5 × раствор Денхардта, обработанную ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1% SDS, и 10% декстрансульфат при 42°C, с отмывками при 42°C в 0,2 × SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C, с последующей высоко строгой отмывкой, состоящей из 0,1×SSC, содержащего EDTA, при 55°C.
«Умеренно строгие условия» можно определить, как описано в Sambrook et al., Molecular Cloning: A Laboratory Manual. New York: Cold Spring Harbor Press, 1989, и они включают в себя использование раствора для отмывки и условий гибридизации (например, температуры, ионной силы и % SDS), менее строгих, чем описанные выше. Примером умеренно строгих условий является инкубация в течение ночи при 37°C в растворе, содержащем: 20% формамид, 5×SSC (150 мМ NaCl, 15 мМ цитрат тринатрия), 50 мМ фосфат натрия (pH 7,6), 5 × раствор Денхардта, 10% декстрансульфат, и 20 мг/мл денатурированной ДНК спермы лосося, с последующей отмывкой фильтров в 1 × SSC при приблизительно 37-50°C. Опытному специалисту в данной области будет понятно, как регулировать температуру, ионную силу, и т.д., как необходимо для соответствия таким факторам, как длина зонда и т.п.
Термин «меченный эпитопом» при применении здесь относится к химерному полипептиду, содержащему полипептид PRO87299, слитый с «полипептидом-меткой». Полипептид-метка имеет достаточно остатков, чтобы обеспечивать эпитоп, против которого можно получить антитело, однако является достаточно коротким, чтобы не мешать активности полипептида, с которым он слит. Полипептид-метка предпочтительно также является достаточно уникальным, так что антитело в основном не реагирует перекрестно с другими эпитопами. Подходящие полипептиды-метки, как правило, содержат, по меньшей мере, шесть аминокислотных остатка, а обычно между приблизительно 8 и 50 аминокислотными остатками (предпочтительно между приблизительно 10 и 20 аминокислотными остатками).
Как применяют здесь, термин «иммуноадгезин» обозначает антитело-подобные молекулы, сочетающие связывающую специфичность гетерологичного белка («адгезин») с эффекторными функциями константных доменов иммуноглобулина. Структурно иммуноадгезины содержат слитые аминокислотную последовательность с желаемой связывающей специфичностью, отличную от участка узнавания и связывания антигена из антитела (т.е. являются «гетерологичными»), и последовательность константного домена иммуноглобулина. Часть адгезина молекулы иммуноадгезина, как правило, представляет собой непрерывную аминокислотную последовательность, содержащую, по меньшей мере, участок связывания рецептора или лиганда. Последовательность константного домена иммуноглобулина в иммуноадгезине можно получить из любого иммуноглобулина, такого как иммуноглобулин подтипов IgG-1, IgG-2, IgG-3, или IgG-4, IgA (включая IgA-I и IgA-2), IgE, IgD или IgM.
«Активный» или «активность» для целей здесь относится к форме (формам) полипептида PRO87299, сохраняющим биологическую и/или иммунологическую активность нативного или природного PRO87299, где «биологическая» активность относится к биологической функции (или ингибиторующей, или стимулирующей), обусловленной нативным или природным PRO87299, отличной от способности индуцировать продукцию антитела против антигенного эпитопа, которой обладает нативный или природный PRO87299, и «иммунологическая» активность относится к способности индуцировать продукцию антитела против антигенного эпитопа, которой обладает нативный или природный PRO87299.
Термин «антагонист» применен в самом широком смысле и включает в себя любую молекулу, которая частично или полностью блокирует, ингибирует или нейтрализует биологическую активность описанного здесь нативного полипептида PRO87299. Подобным образом термин «агонист» применен в самом широком смысле и включает в себя любую молекулу, которая мимикрирует биологическую активность описанного здесь нативного полипептида PRO87299. Подходящие молекулы агониста или антагониста конкретно включают в себя антитела или фрагменты антител - агонисты или антагонисты, фрагменты или варианты аминокислотной последовательности нативных полипептидов PRO87299, пептиды, антисмысловые олигонуклеотиды, малые органические молекулы, и т.д. Способы идентификации агонистов или антагонистов полипептида PRO87299 могут включать в себя привдение в контакт полипептида PRO87299 с молекулой-кандидатом агонистом или антагонистом и измерение подлежащего детекции изменения одного или нескольких видов биологической активности, нормально связанных с полипептидом PRO87299.
«Лечение» относится как к терапевтическому лечению, так и к профилактическим или предупредительным мерам, где целью является предупредить или замедлить (уменьшить) намеченное патологическое состояние или расстройство. Нуждающиеся в лечении включают в себя тех, у кого уже есть расстройство, так же как тех, кто предрасположен к расстройству, или тех, у кого нужно предотвратить расстройство.
«Хроническое» введение относится к введению средства (средств) в непрерывном режиме, в противоположность ударному режиму, так чтобы поддерживать начальный терапевтический эффект (активность) на продолжительный период времени. «Периодическое» введение представляет собой лечение, которое не является выполненным последовательно без прерывания, но является скорее циклическим по характеру.
«Млекопитающее» для целей лечения относится к любому животному, классифицируемому как млекопитающее, включая человека, домашних и сельскохозяйственных животных, и животных в зоопарке, спортивных или комнатных, таких как собаки, кошки, крупный рогатый скот, лошади, овцы, свиньи, козы, кролики и т.д. Предпочтительно млекопитающее представляет собой человека. Введение «в сочетании с» одним или несколькими дополнительнми терапевтическими средствами включает в себя одновременное (совместное) и последовательное введение в любом порядке.
«Носители», как применяют здесь, включают в себя фармацевтически приемлемые носители, наполнители, или стабилизаторы, являющиеся нетоксичными для клетки или млекопитающего, подвергающегося их воздействию в применяемых дозах и концентрациях. Часто физиологически приемлемый носитель представляет собой водный раствор с забуференным pH. Примеры физиологически приемлемых носителей включают в себя буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводороды, включая глюкозу, маннозу или декстрины; хелатобразующие агенты, такие как EDTA; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные сурфактанты, такие как TWEEN™, полиэтиленгликоль (PEG) и PLURONICS™.
«Фрагменты антител» включают в себя часть интактного антитела, предпочтительно антигенсвязывающую или вариабельную область интактного антитела. Примеры фрагментов антител включают в себя фрагменты Fab, Fab', F(ab')2, и Fv; диатела; линейные антитела (Zapata et al., Protein Eng. 8(10): 1057-1062 [1995]); молекулы одноцепочечного антитела; и мультиспецифические антитела, сформированные из фрагментов антител.
Расщеплением антител папаином получают два идентичных антигенсвязывающих фрагмента, называемых «Fab»-фрагменты, каждый с отдельным антигенсвязывающим участком, и оставшийся «Fc»-фрагмент - обозначение, отражающее способность легко кристаллизоваться. Обработкой пепсином получают F(ab')2-фрагмент, обладающий двумя соединяющимися с антигеном участками и еще способный перекрестно связывать антиген.
«Fv» представляет собой минимальный фрагмент антитела, содержащий полный участок, узнающий и связывающий антиген. Этот участок состоит из димера вариабельного домена одной тяжелой и одной легкой цепи в жесткой, нековалентной связи. В данной конфигурации это выражается в том, что три CDR каждого вариабельного домена взаимодействуют, определяя антигенсвязывающий участок на поверхности димера VH-VL. Шесть CDR совместно обеспечивают антигенсвязывающую специфичность антитела. Однако даже отдельный вариабельный домен (или половина Fv, содержащая только три CDR, специфичных к антигену) обладает способностью узнавать и связывать антиген, хотя с меньшей аффинностью, чем целый связывающий участок.
Fab-фрагмент содержит также константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab-фрагменты отличаются от Fab'-фрагментов добавлением нескольких остатков на C-конце CH1-домена тяжелой цепи, включая один или несколько цистеинов из шарнирной области антитела. F(ab')2-фрагменты антител первоначально были получены как пары Fab'-фрагментов с шарнирными цистеинами между ними. Известны также другие химические присоединения фрагментов антитела.
«Легкие цепи» антител (иммуноглобулинов) из любых видов позвоночных можно отнести к одному из двух четко различимых типов, называемых каппа и лямбда, на основании аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей антитела можно отнести к различным классам. Существует пять главных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно далее разделить на подклассы (изотипы), например, IgGl, IgG2, IgG3, IgG4, IgA и IgA2.
«Одноцепочечные Fv» или «sFv»-фрагменты антител содержат VH и VL домены антитела, где эти домены присутствуют в одной полипептидной цепи. Предпочтительно полипептид Fv дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет sFv формировать желаемую структуру для связывания антигена. Обзор sFv смотри у Pluckthun in The Pharmacology of Monoclonal antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Термин «диатела» обозначает малые фрагменты антител с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). Применением линкера, который является слишком коротким, чтобы позволять спаривание между двумя доменами в одной и той же цепи, домены вынуждают спариваться с комплементарными доменами другой цепи и образовывать два антигенсвязывающих участка. Диатела более полно описаны, например, в EP 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
«Выделенное» антитело представляет собой то, которое идентифицировали и отделили и/или извлекли из его естественного окружения. Загрязняющие компоненты его естественного окружения представляют собой вещества, которые будут мешать диагностическим или терапевтическим применениям антагониста или антитела и могут включать в себя ферменты, гормоны и другие белковые и небелковые растворы. В предпочтительных вариантах осуществления антитело будут очищать (1) до более чем 95% по массе антитела, как определяют способом Лоури, и наиболее предпочтительно до более чем 99% по массе, (2) до степени, достаточной для получения аминокислотной последовательности, по меньшей мере, 15 остатков, с N-конца или внутренних с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности в SDS-PAGE в восстанавливающих и невосстанавливающих условиях с использованием окраски Кумасси синим или, предпочтительно, серебром. Выделенное антитело включает в себя антитело in situ внутри рекомбинантных клеток, с тех пор как перестает присутствовать, по меньшей мере, один компонент естественного окружения антитела. Однако обычно выделенное антитело будут получать посредством, по меньшей мере, одной стадии очистки.
Антитело, которое «специфически связывается с» или является «специфичным к» конкретному полипептиду или эпитопу на конкретном полипептиде, представляет собой то, которое связывается с данным конкретным полипептидом или эпитопом на конкретном полипептиде без существенного связывания с любым другим полипептидом или полипептидным эпитопом.
Слово «метка» при применении здесь относится к соединению или композиции, которые можно детектировать, конъюгированным непосредственно или опосредованно с антителом, так что получают «меченое» антитело. Метка может являться поддающейся детекции сама по себе (например, радиоизотопные метки или флуоресцентные метки), или в случае ферментативной метки может катализировать химическое изменение состава субстрата или композиции, которое можно детектировать.
Под «твердой фазой» понимают неводную матрицу, к которой может присоединяться антитело по настоящему изобретению. Примеры твердых фаз, охватываемых здесь, включают в себя твердые фазы, частично или полностью сформированные из стекла (например, стекло с контролируемым размером пор), полисахариды (например, агарозу), полиакриламиды, полистирол, поливиниловый спирт и силиконы. В конкретных вариантах осуществления в зависимости от контекста твердая фаза может быть выполненной в виде лунки планшета для анализа; в других она представляет собой колонку для очистки (например, колонку для аффинной хроматографии). Данный термин включают в себя также прерывающуюся твердую фазу из дискретных частиц, таких как описанные в патенте США No 4275149.
«Липосома» представляет собой малый пузырек, состоящий из рпзличных типов липидов, фосфолипидов и/или сурфактанта, который являеется применимым для доставки млекопитающему лекарственного средства (такого как полипептид PRO87299
или антитело к нему). Компоненты липосомы обычно упорядочены в двухслойной структуре подобно упорядочиванию липидов в биологических мембранах.
«Малая молекула», как определяют здесь, обладает молекулярной массой менее приблизительно 500 Дальтон.
Термин «связанное с иммунитетом заболевание» обозначает заболевание, при котором компонент иммунной системы млекопитающего вызывает, опосредует заболеваемость млекопитающего или иначе способствует ей. Также сюда включены заболевания, при которых стимуляция иммунного ответа или вмешательство в него оказывает облегчающий эффект на прогрессирование заболевания. В данный термин включены иммуноопосредованные воспалительные заболевания, неиммуноопосредованные воспалительные заболевания, инфекционные заболевания, иммунодефицитные заболевания, неоплазия, и т.д.
Термин «опосредованное T-клетками заболевание» означает заболевание, при котором T-клетки непосредственно или опосредованно содействуют или иначе способствуют заболеваемости млекопитающих. Опосредованное T-клетками заболевание может быть связано с опосредованными клетками эффектами, опосредованными лимфокинами эффектами и т.д., и даже эффектами, связанными с B-клетками, если B-клетки являются стимулированными, например, лимфокинами, секретируемыми T-клетками.
Примеры связанных с иммунитетом и воспалительных заболеваний, некоторые из которых являются иммунными или опосредованными T-клетками, которые можно лечить по настоящему изобретению, включают в себя системную красную волчанку, ревматоидный артрит, ювенильный хронический артрит, спондилоартропатии, системный склероз (склеродермию), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (иммунную панцитопению, пароксизмальную ночную гемоглобинурию), аутоиммунную тромбоцитопению (идиопатическую тромбоцитопеническую пурпуру, иммуноопосредованную тромбоцитопению), тиреоидит (диффузный токсический зоб, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), сахарный диабет, иммуноопосредованное ренальное заболевание (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервной системы, такие как рассеянный склероз, идиопатическая демиелинизирующая полиневропатия или синдром Гийена-Барре, и хроническая воспалительная демиелинизирующая полиневропатия, гепатобилиарные заболевания, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие, не гепатотропные, вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит и склерозирующий холангит, воспалительное заболевание кишечника (язвенный колит: болезнь Крона), глютензависимую энтеропатию и заболевание Уиппла, аутоиммунные или иммуноопосредованные заболевания кожи, включая буллезные кожное заболевание, полиморфную эритему и контактный дерматит, псориаз, аллергические заболевания, такие как астма, аллергический ринит, атопический дерматит, гиперчувствительность к пище и уртикарию, иммунологические заболевания легких, такие как эозинофильные пневмонии, идиопатический фиброз легких и гиперчувствительный пневмонит, заболевания, связанные с трансплантацией, включая отторжение трансплантата и реакцию трансплантат против хозяина. Инфекционные заболевания включают в себя вирусные заболевания, такие как СПИД (инфекция HIV), гепатит A, B, C, D и E, герпес и т.д., бактериальные инфекции, грибковые инфекции, протозойные инфекции и паразитарные инфекции.
Термин «эффективное количество» представляет собой концентрацию или количество полипептида PRO87299 и/или агониста/антагониста, которое приводит к достижению конкретной поставленной цели. «Эффективное количество» полипептида PRO87299 или его агониста или антагониста можно определить эмпирически. Кроме того, «терапевтически эффективное количество» представляет собой концентрацию или количество полипептида PRO87299 и/или агониста/антагониста, которое является эффективным для достижения конкретного заявленного терапевтического эффекта. Данное количество можно также определить эмпирически.
Термин «цитотоксическое средство», как применяют здесь, обозначает вещество, ингибирующее или предупреждающее функционирование клеток и/или вызывающее разрушение клеток. Термин предназначен, чтобы включать радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические средства, например метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин C, хлорамбуцил, даунорубицин или другие интеркалирующие средства, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины из бактерий, грибов, растений или животных, включая их фрагменты и/или варианты, и различные противоопухолевые или противораковые средства, описанные ниже. Ниже описаны другие цитотоксические средства. Уничтожающее опухолевые клетки средство вызывает разрушение опухолевых клеток. Цитотоксины можно ковалентно присоединять к антителу для нацеливания токсина на конкретную интересующую клетку, экспрессирующую антиген. Применимыми токсинами являются те, линкеры которых включают в себя, без ограничений, следующие:
ЛИНКЕРЫ:
MC=малеимидокапроил
Val Cit=валин-цитруллин, дипептидный участок в расщепляемом протеазой линкере.
Цитруллин=2-амино-5-уреидопентановая кислота
PAB=p-аминобензилкарбамоил («самоуничтожаемая» часть линкера)
Me=N-метил-валин цитруллин, где линкерная пептидная связь модифицирована для предотвращения ее расщепления катепсином B
MC(PEG)6-OH = малеимидокапроил-полиэтиленгликоль, присоединенный к цистеинам антитела.
SPP=N-сукцинимидил-4-(2-пиридилтио)пентаноат
SMCC=N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат
ЦИТОТОКСИЧЕСКИЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА:
MMAE=монометилауристатин E (MW 718)
MMAF=вариант ауристатина E (MMAE) с фенилаланином на C-конце лекарственного средства (MW 731,5)
MMAF-DMAEA=MMAF с DMAEA (диметиламиноэтиламином) в амидной связи с C-концевым фенилаланином (MW 801,5)
MMAF-TEG=MMAF с тетраэтиленгликолем, эстерифицированным с фенилаланином
MMAF-NtBu=N-t-бутил, присоединенный в форме амида к C-концу MMAF
AEVB=валерилбензилгидразон ауристатина E, кислотолабильный линкер через C-конец AE (MW 732)
AFP=фенилендиамин ауристатина F; (фенилаланиновый вариант, присоединенный к антителу через C-конец посредством фенилендиаминового спейсера) (MW 732).
«Ингибирующее рост средство» при применении здесь обозначает соединение или композицию, ингибирующие рост клетки, либо in vitro, либо in vivo. Таким образом, ингибирующее рост средство может представлять собой то, которое значительно уменьшает процент клеток в S-фазе. Примеры ингибирующих рост средств включают в себя средства, блокирующие прохождение клеточного цикла (в точке, отличной от S-фазы), такие как средства, индуцирующие Gl-арест и арест в M-фазе. Классические блокаторы M-фазы включают в себя алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Те средства, которые арестовывают Gl, распространяются также на арест S-фазы, например ДНК-алкилирующие средства, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил, и ара-C. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., часть 1, названная "Cell cycle regulation, oncogens, and antineoplastic drugs" by Murakami et al. (WB Saunders: Philadelphia, 1995), особенно p. 13. Таксаны (паклитаксел и доцетаксел) представляют собой противораковые лекарственные средства, оба полученные из both derived from the тиса. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из тиса европейского, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел активируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки посредством предотвращения деполимеризации, что приводит к ингибированию митоза в клетках.
«Химиотерапевтическое средство» представляет собой химическое соединение, используемое в лечении рака. Примеры химиотерапевтических средств включают в себя адриамицин, доксорубицин, эпирубицин, 5-фторурацил, арабинозид цитозина («Ara-C»), циклофосфамид, тиотепа, бусульфан, цитоксин, таксоиды, например, паклитаксел (Taxol, Bristol-Myers Squibb Oncology, Princeton, NJ), и доксетаксел (Taxotere, Rhone-Poulenc Rorer, Antony, France), таксотер, метотрексат, цисплатин, мелфалан, винбластин, блеомицин, этопозид, ифосфамид, митомицин C, митоксантрон, винкристин, винорелбин, карбоплатин, тенипозид, дауномицин, карминомицин, аминоптерин, дактиномицин, митомицины, эсперамицины (смотри патент США No. 4675187), мелфалан и другие родственные азотные горчичные газы. В данное определение включены также гормональные средства, которые действуют, регулируя или ингибируя гормональное действие опухолей, такие как тамоксифен и онапристон.
Термин «цитокин» представляет собой общий термин для белков, высвобождаемых одной популяцией клеток, которые действуют на другую клетку как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и общепринятые полипептидные гормоны. В число цитокинов включают гормон роста, такой как гормон роста человека, N-метионил гормон роста человека и бычий гормон роста; паратиреоидный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреостимулирующий гормон (TSH) и лютеинизирующий гормон (LH); фактор роста гепатоцитов; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухолей-α и -β; мюллерова ингибирующая субстанция; гонадотропин-ассоциированный пептид мыши; ингибин; активин, фактор роста эндотелия сосудов; интегрин; тромбопоэтин (TPO); факторы роста нервов, такие как NGF-β; фактор роста тромбоцитов; трансформирующие факторы роста (TGF), такие как TGF-α и TGF-β; инсулиноподобный фактор роста-I и -II; эритропоэтин
(EPO);остеоиндуктивные факторы; интерфероны, такие как интерферон-α, -β и -γ; колониестимулирующие факторы (CSF), такие как макрофаг-CSF (M-CSF); гранулоцит-макрофаг-CSF (GM-CSF) и гранулоцит-CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1α, IL-2, IL-3, IL- 4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12; фактор некроза опухолей, такой как TNF-α или TNF-β; и другие полипептидные факторы, включая LIF и лиганд kit (KL). Как применяют здесь, термин цитокин включает в себя белки из природных источников или из культуры рекомбинантных клеток и биологически активные эквиваленты цитокинов с природной последовательностью.
Как применяют здесь, термин «иммуноадгезин» обозначает антителоподобные молекулы, сочетающие связывающую специфичность гетерологичного белка («адгезин») с эффекторными функциями константных доменов иммуноглобулина. Структурно иммуноадгезины содержат аминокислотную последовательность с желаемой связывающей специфичностью, отличную от участка узнавания и связывания антигена из антитела (т.е. являющуюся «гетерологичной»), слитую с последовательностью константного домена иммуноглобулина. Часть адгезина молекулы иммуноадгезина, как правило, представляет собой непрерывную аминокислотную последовательность, содержащую, по меньшей мере, участок связывания рецептора или лиганда. Последовательность константного домена иммуноглобулина в иммуноадгезине можно получить из любого иммуноглобулина, такого как иммуноглобулин подтипов IgG-1, IgG-2, IgG-3, или IgG-4, IgA (включая IgA-1 и IgA-2), IgE, IgD или IgM.
Как применяют здесь, термин «воспалительные клетки» обозначает клетки, усиливающие воспалительный ответ, такие как мононуклеарные клетки, эозинофилы, макрофаги, и полиморфноядерные нейтрофилы (PMN).














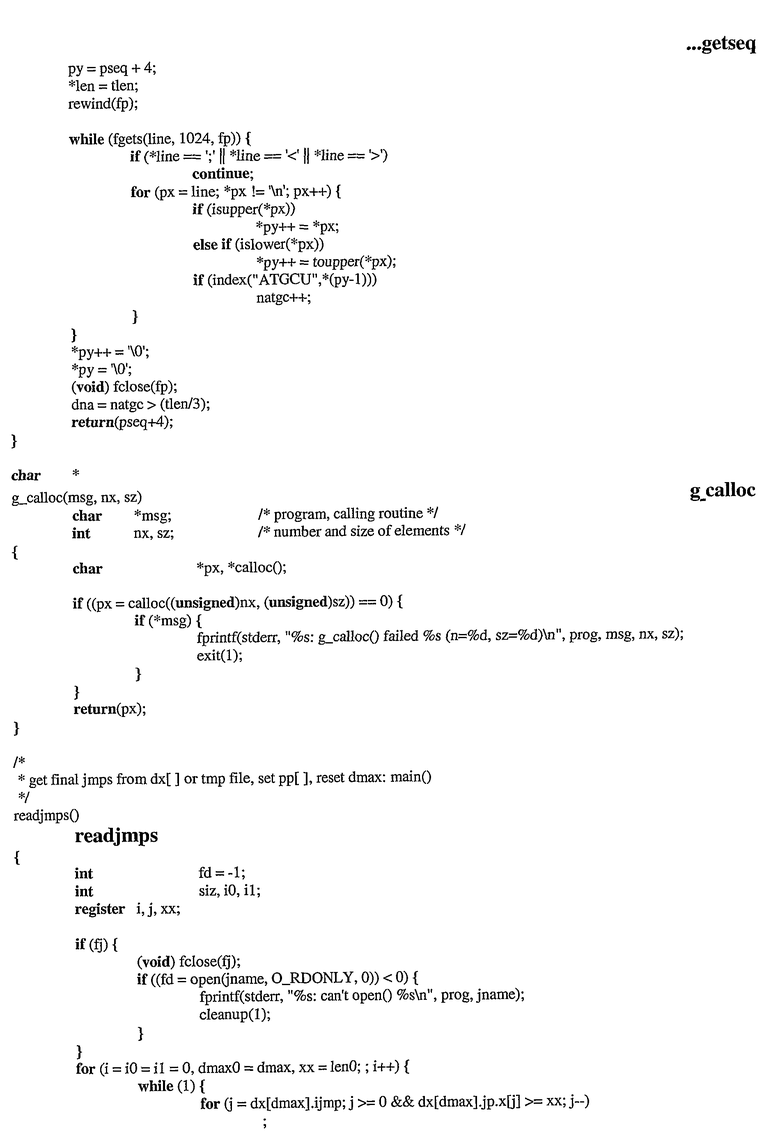


5 деленное на 15 = 33,3%
5 деленное на 10 = 50%
6 деленное на 14 = 42,9%
4 деленное на 12 = 33,3%
II. Композиции и способы по изобретению
A. Полноразмерные полипептиды PRO87299
Настоящее изобретение относится к новым идентифицированным и выделенным нуклеотидным последовательностям, кодирующим полипептиды, обозначенные в настоящей заявке как полипептиды PRO87299. В частности, идентифицированы и выделены кДНК, кодирующие различные полипептиды PRO87299, как более подробно описано в примерах ниже.
Фактические нуклеотидные последовательности данных клонов опытный специалист в данной области может легко определить секвенированием хранящегося в банке клона с использованием общепринятых в данной области способов. Предполагаемую аминокислотную последовательность можно определить из нуклеотидной последовательности с использованием общепринятого опыта. Для описанных здесь полипептидов PRO87299 и кодирующих нуклеиновых кислот авторы настоящего изобретения идентифицировали то, что, как полагают, является рамкой считывания, лучше всего идентифицируемой по информации о последовательности, доступной на тот момент.
B. Варианты полипептидов PRO87299
Кроме полноразмерной нативной последовательности полипептидов PRO87299, описанной здесь, предполагают, что можно получить варианты PRO87299. Варианты PRO87299 можно получить введением подходящих нуклеотидных замен в ДНК PRO87299 и/или синтезом желаемого полипептида PRO87299. Специалистам в данной области понятно, что аминокислотные замены могут изменять пост-трансляционные процессы PRO87299, такие как изменение числа или положения участков гликозилирования, или изменение характеристик заякоривания в мембране.
Описанные здесь изменения в нативной полноразмерной последовательности PRO87299 или в различных доменах PRO87299 можно получить, например, с использованием любого из способов и руководств для консервативных и неконсервативных мутаций, описанных, например, в патенте США No. 5364934. Изменения могут представлять собой замену, делецию или вставку одного или нескольких кодонов, кодирующих PRO87299, что приводит к изменению аминокислотной последовательности PRO87299 по сравнению с нативной последовательностью PRO87299. Необязательно изменение представляет собой замену, по меньшей мере, одной аминокислоты на другую аминокислоту в одном или нескольких доменах PRO87299. Руководство по определению, какой аминокислотный остаток можно вставить, заменить или делетировать без неблагоприятного воздействия на желаемую активность, можно найти сравнением последовательности PRO87299 с последовательностями молекул известных гомологичных белков и минимизацией количества изменений аминокислотной последовательности, выполненных в областях высокой гомологии. Аминокислотные замены могут являться результатом замены одной аминокислоты на другую аминокислоту, обладающую похожими структурными и/или химическими свойствами, такими как замена лейцина на серин, т.е. консервативными аминокислотными заменами. Вставки или делеции необязательно могут лежать в диапазоне приблизительно от 1 до 5 аминокислот. Допустимые изменения можно определить систематическим выполнением вставок, делеций или замен аминокислот в последовательности и тестированием полученных вариантов по активности, которой обладает полноразмерная или зрелая нативная последовательность.
Здесь представлены фрагменты полипептида PRO87299. Такие фрагменты могут являться укороченными на N-конце или C-конце, или в них могут отсутствовать внутренние остатки, например, по сравнению с полноразмерным нативным белком. В конкретных фрагментах отсутствуют аминокислотные остатки, не являющиеся необходимыми для желаемой биологической активности полипептида PRO87299.
Фрагменты PRO87299 можно получить любым из множества общепринятых способов. Желаемые фрагменты пептида можно синтезировать химически. Альтернативный способ включает в себя получение фрагментов PRO87299 ферментативным расщеплением, например обработкой белка ферментом, известным как расщепляющий белки в участках, определяемых конкретными аминокислотными остатками, или расщеплением ДНК подходящими рестрикционными ферментами и выделением желаемого фрагмента. Другой подходящий способ включает в себя выделение и амплификацию фрагмента ДНК, кодирующего желаемый фрагмент полипептида, посредством полимеразной цепной реакции (PCR). Олигонуклеотиды, определяющие желаемые концы фрагмента ДНК, применяют как 5'- и 3'-праймеры для PCR. Предпочтительно фрагменты полипептида PRO87299 обладают, по меньшей мере, одной общей биологической и/или иммунологической активностью с нативным полипептидом PRO87299, описанным здесь.
В конкретных вариантах осуществления интересующие консервативные замены показаны в таблице 6 под заголовком «предпочтительные замены». Если такие замены приводят к изменению биологической активности, тогда вводят более существенные изменения, обозначенные «примерные замены» в таблице 6 или описанные далее ниже по отношению к классам аминокислот, и скринируют продукты.

Существенные модификации функции или иммунологической идентичности полипептида PRO87299 выполняют выбором замен, которые значительно отличаются по их эффекту на поддержание (a) структуры полипептидного остова в области замены, например, в форме конформации листа или спирали, (b) заряда или гидрофобности молекулы в участке-мишени или (c) объема боковой цепи. Встречающиеся в природе остатки разделяют на группы на основании общих свойств боковой цепи:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gin, his, lys, arg;
(5) остатки, влияющие на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены будут вызывать замену члена одного из этих классов на другой класс. Такие замещенные остатки также можно вводить в участки консервативных замен или более предпочтительно в оставшиеся (неконсервативные) участки.
Изменения можно выполнять с использованием таких способов, известных в данной области, как олигонуклеотид-опосредованный (сайт-специфический) мутагенез, аланиновое сканирование, и PCR-мутагенез. Сайт-специфический мутагенез [Carter et al., Nucl. Acids Res., 13:4331 (1986); Zoller et al., Nucl. Acids Res., 10:6487 (1987)], кассетный мутагенез [Wells et al., Gene, 34:315 (1985)], определяемый рестрикцией мутагенез [Wells et al., Philos. Trans. R. Soc. London SerA, 317:415 (1986)] или другие известные способы можно проводить с клонированной ДНК для получения варианта ДНК PRO87299.
Сканирующий аминокислотный анализ можно также применять для идентификации одной или нескольких аминокислот в непрерывной последовательности. Среди предпочтительных сканирующих аминокислот присутствуют относительно небольшие нейтральные аминокислоты. Такие аминокислоты включают в себя аланин, глицин, серин, и цистеин. Аланин, как правило, является предпочтительной сканирующей аминокислотой среди данной группы, потому что он исключает боковую цепь после бета-углерода и менее вероятно изменяет конформацию главной цепи варианта [Cunningham and Wells, Science, 244: 1081-1085 (1989)]. Аланин также, как правило, является предпочтительным, поскольку является наиболее распространенной аминокислотой. Кроме того, его часто обнаруживают как в углубленном, так и в экспонированном положении [Creighton, The Proteins, (W.H. Freeman & Co., N.Y.); Chothia, J. MoI. Biol., 150:1 (1976)]. Если при замещении аланином не получают достаточного количества вариантов, можно использовать изостерическую аминокислоту.
C. Модификации PRO87299
Ковалентные модификации PRO87299 также включены в объем данного изобретения. Один из типов ковалентной модификации включает в себя реакцию намеченных аминокислотных остатков полипептида PRO87299 с органическим дериватизующим средством, способным реагировать с выбранными боковыми цепями или N- или C-концевыми остатками PRO87299. Дериватизацию бифункциональными средствами применяют, например, для перекрестного сшивания PRO87299 с нерастворимой в воде матрицей или поверхностью-подложкой для применения в способе очистки анти-PRO87299 антител и наоборот. Общеупотребительные сшивающие средства включают в себя, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаральдегид, сложные эфиры N-гидроксисукцинимида, например, сложные эфиры с 4-азидосалициловой кислотой, гомобифункциональные сложные имидные эфиры, включая сложные эфиры дисукцинилимидила, такие как 3,3'-дитиобис(сукцинимидилпропионат), бифункциональные малеинимиды, такие как бис-N-малеимидо-1,8-октан; и такие средства, как метил-3-[(p-азидофенил)дитио]пропиоимидат.
Другие модификации включают в себя дезамидирование остатков глутаминила и аспарагинила до соответствующих остатков глутамила и аспартила соответственно, гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серила или треонила, метилирование боковых цепей α-аминогрупп лизина, аргинина и гистидина [T.E. Creighton, Proteins: Structure и Molecular Properties, W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)], ацетилирование N-концевого амина и амидирование любой C-концевой карбоксильной группы.
Другой тип ковалентной модификации полипептида PRO87299, включенный в объем данного изобретения, включает в себя изменение характера природного гликозилирования полипептида. «Изменение характера природного гликозилирования полипептида» для целей здесь предназначено для обозначения удаления одной или нескольких углеводородных групп, обнаруженных в природной последовательности PRO87299 (или удалением лежавшего в основе участка гликозилирования, или удалением гликозилирования химическими и/или ферментативными способами), и/или добавления одного или нескольких участков гликозилирования, которые не присутствуют в природной последовательности PRO87299. Кроме того, фраза включает в себя качественное изменение гликозилирования природных белков, включающее изменение природы и соотношений различных присутствующих углеводородных групп.
Добавление участков гликозилирования к полипептиду PRO87299 может сопровождаться изменением аминокислотной последовательности. Изменение можно выполнять, например, добавлением или заменой одного или нескольких остатков серина или треонина в природной последовательности PRO87299 (для участков O-связанного гликозилирования). Аминокислотную последовательность PRO87299 можно, необязательно, изменять посредством замен на уровне ДНК, в частности, введением мутаций в ДНК, кодирующую полипептид PRO87299, по предварительно выбранным основаниям, так что получают кодоны, которые будут транслированы в желаемые аминокислоты.
Другими способами увеличения числа углеводородных групп в полипептиде PRO87299 являются химическое или ферментативное присоединение гликозидов к полипептиду. Такие способы описаны в данной области, например, в WO 87/05330, опубликованной 11 сентября 1987 г., и в Aplin and Wriston, CRC Crit. Rev. Biochem., pp. 259-306 (1981). Удаление углеводородных групп, присутствующих в полипептиде PRO87299, можно осуществлять химически или ферментативно или мутационной заменой кодонов, кодирующих аминокислотные остатки, служащие мишенями для гликозилирования. Способы химического дегликозилирования известны в данной области и описаны, например, Hakimuddin, et al., Arch. Biochem. Biophvs., 259:52 (1987) и Edge et al., Anal. Biochem., 118:131 (1981). Ферментативное расщепление углеводородных групп на полипептидах можно осуществлять с использованием множества эндо- и экзо- гликозидаз, как описано в Thotakura et al., Meth. Enzvmol., 138:350 (1987).
Другой тип ковалентной модификации PRO87299 включает в себя связывание полипептида PRO87299 с одним из множества небелковых полимеров, например, полиэтиленгликолем (PEG), полипропиленгликолем, или полиоксиалкиленами, способом, описанным в патентах США No. 4640835; 4496689; 4301144; 4670417; 4791192 или 4179337.
PRO87299 по настоящему изобретению можно модифицировать также таким способом, чтобы получить химерную молекулу, содержащую PRO87299, слитый с другим, гетерологичным полипептидом или аминокислотной последовательностью.
В одном из вариантов осуществления такая химерная молекула содержит слитый белок PRO87299 с полипептидом-меткой, который обеспечивает эпитоп, с которым может избирательно связываться антитело против метки. Эпитоп-метку, как правило, помещают на N- или C-конце PRO87299. Присутствие таких меченных эпитопом форм PRO87299 можно детектировать с использованием антитела против полипептида-метки. Снабжение эпитопом-меткой позволяет также легкую очистку PRO87299 посредством аффинной очистки с использованием антитела против метки или другого типа аффинной смолы, которая связывает эпитоп-метку. Различные полипептиды-метки и соответствующие им антитела хорошо известны в данной области. Примеры включают в себя метки полигистидин (поли-his) или полигистидин-глицин (поли-his-gly); полипептид-метку HA гриппа и антитело к нему 12CA5 [Field et al., MoI. Cell. Biol., 8:2159-2165 (1988)]; метку c-myc и антитела к нему 8F9, 3C7, 6E10, G4, B7 и 9E10 [Evan et al., Molecular и Cellular Biology, 5:3610-3616 (1985)]; и метку гликопротеин D вируса простого герпеса (gD), и антитело к нему [Paborsky et al., Pprotein Engineering, 3(6):547-553 (1990)]. Другие полипептиды-метки включают в себя Flag-пептид [Hopp et al., BioTechnology, 6:1204-1210 (1988)]; пептидный эпитоп KT3 [Martin et al., Science, 255:192-194 (1992)]; пептидный эпитоп альфа-тубулина [Skinner et al., J. Biol. Chem., 266:15163-15166 (1991)]; и пептидную метку белка гена 10 T7 [Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. USA. 87:6393-6397 (1990)].
В альтернативном варианте осуществления химерная молекула может содержать белок PRO87299, слитый с иммуноглобулином или с конкретной областью иммуноглобулина. Для бивалентной фомы химерной молекулы (также обозначаемой как «иммуноадгезин») такой слитый белок можно получить с Fc-областью молекулы IgG. Слитые белки Ig предпочтительно включают в себя замену растворимой формой полипептида PRO87299 (с делетированным или инактивированным трансмембранным доменом) вместо, по меньшей мере, одной вариабельной области в молекуле Ig. В особенно предпочтительном варианте осуществления слитый с иммуноглобулином белок содержит шарнирную CH2 и CH3 или шарнирную CH1, CH2 и CH3 области молекулы IgG1. Получение слитных с иммуноглобулином белков см. также в патенте США No. 5428130, поданном 27 июня 1995 г.
D. Получение PRO87299
Описание ниже в первую очередь относится к продукции PRO87299 культивируемыми клетками, трансформированными или трансфицированными вектором, содержащим нуклеиновую кислоту PRO87299. Это, конечно, предусматривает, что альтернативные способы, которые хорошо известны в данной области, можно применять для получения PRO87299. Например, последовательность PRO87299 или ее части можно получить прямым пептидным синтезом с использованием твердофазных способов [см., например, Stewart et al., Solid-Phase Peptide Synthesis, W.H. Freeman Co., San Francisco, CA (1969); Merrifield, J. Am. Chem. Soc, 85:2149-2154 (1963)]. Синтез белка in vitro можно проводить с использованием ручных способов или автоматически. Автоматический синтез можно выполнять, например, с использованием пептидного синтезатора Applied Biosystems (Foster City, CA) с использованием инструкций производителя. Различные части PRO87299 можно химически синтезировать отдельно и объединять с использованием химических или ферментативных способов для получения полноразмерного PRO87299.
1. Выделение ДНК, кодирующей PRO87299
ДНК, кодирующую PRO87299, можно получить из библиотеки кДНК, полученной из ткани, считающейся обладающей мРНК PRO87299 mRNA и экспрессирующей ее на поддающемся детекции уровне. Соответственно ДНК PRO87299 человека можно удобно получить из библиотеки кДНК, полученной из ткани человека, такой как описанная в примерах. Кодирующий PRO87299 ген можно получить также из геномной библиотеки или посредством известных синтетических способов (например, автоматическим синтезом нуклеиновой кислоты).
Библиотеки можно скринировать зондами (такими как антитела к PRO87299 или олигонуклеотиды из, по меньшей мере, приблизительно 20-80 оснований), сконструированными, чтобы идентифицировать интересующий ген или кодируемый им белок. Скрининг библиотеки кДНК или геномной библиотеки выбранным зондом можно проводить с использованием общепринятых способов, таких как описаны в Sambrook et al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989). Альтернативным способом выделения гена, кодирующего PRO87299, является применение способа PCR [Sambrook et al., выше; Dieffenbach et al., PCR Primer: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 1995)].
В примерах ниже описаны способы скрининга библиотеки кДНК. Олигонуклеотидные последовательности, выбранные в качестве зондов, должны обладать достаточной длиной и достаточной однозначностью, чтобы минимизировать ложноположительные сигналы. Олигонуклеотид предпочтительно является меченым, так что его можно детектировать при гибридизации с ДНК в скринируемой библиотеке. Способы мечения хорошо известны в данной области и включают в себя использование радиоактивных меток, подобных 32P-меченному ATP, биотинилирование или ферментативное мечение. Условия гибридизации, включая умеренно строгие и высоко строгие, представлены в Sambrook et al., выше.
Последовательности, идентифицированные такими способами скрининга библиотек, можно сравнивать и выравнивать с другими известными последовательностями, находящимися на хранении и доступными в публичных базах данных, таких как GenBank или другие частные базы данных последовательностей. Идентичность последовательности (или на аминокислотном, или на нуклеотидном уровне) с определенными областями молекулы или от края до края полноразмерной последовательности можно определить с использованием способов, известных в данной области, и как описано здесь.
Нуклеиновую кислоту, обладающую кодирующей белок последовательностью, можно получить посредством скрининга выбранных библиотек кДНК или геномных библиотек с использованием выведенной аминокислотной последовательности, описанной здесь, для первого раза, и, если необходимо, с использованием общепринятых способов удлинения праймеров, как описано в Sambrook et al., выше, для детекции предшественников и процессинга промежуточных соединений мРНК, которые могли не оказаться обратно транскрибированными в кДНК.
2. Выбор и трансформация клеток-хозяев
Клетки-хозяева трансфицировали или трансформировали экспрессирующими или клонирующими векторами, описанными здесь, для продукции PRO87299 и культивировали в общепринятых питательных средах, модифицированных соответствующим образом для индукции промоторов, селекции трансформантов или амплификаци генов, кодирующих желаемые последовательности. Опытный специалист в данной области может без излишних экспериментов выбрать условия культивирования, такие как среда, температура, pH и т.п. Как правило, принципы, протоколы и практические способы для максимизации продуктивности культур клеток можно найти в Mammalian Cell Biotechnology: a Practical Approach, M. Butler, ed. (IRL Press, 1991) и Sambrook et al., выше.
Способы трансфекции эукариотической клетки и трансформации прокариотической клетки известны обычному специалисту в данной области, например, c CaCl2, c CaPO4, опосредованная липосомами и электропорация. В зависимости от применяемой клетки-хозяина трансформацию проводят с использованием общепринятых способов, подходящих для таких клеток. Для прокариот, как правило, используют обработку кальцием, применяя хлорид кальция, как описано в Sambrook et al., выше, или электропорацию. Для трансформации конкретных клеток растений используют инфекцию Agrobacterium tumefaciens, как описано в Shaw et al., Gene, 23:315 (1983) и WO 89/05859, опубликованной 29 июня 1989 г. Для клеток млекопитающих без таких клеточных стенок можно использовать способ кальций-фосфатной преципитации Graham and van der Eb, Virology, 52:456-457 (1978). Общие аспекты трансфекций в системе хозяев-клеток млекопитающих описаны в патенте США No 4399216. Трансформации в дрожжах, как правило, проводят способами Van Solingen et al., J. Bact, 130:946 (1977) и Hsiao et al., Proc. Natl. Acad. Sci. (USA), 76:3829 (1979). Однако для введения ДНК в клетки можно использовать также другие способы, такие как микроинъекция в ядро, электропорация, слияние бактериальных протопластов с интактными клетками или поликатионы, например полибрен, полиорнитин. Различные способы трансформации клеток млекопитающих смотри в Keown et al., Methods in Enzymology, 185:527-537 (1990) и Mansour et al., Nature, 336:348-352 (1988).
Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах здесь включают в себя клетки прокариот, дрожжей, или высших эукариот. Подходящие прокариоты включают в себя в качестве неограничивающих примеров эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как E. coli. Публично доступны различные штаммы E. coli, такие как штамм E. coli K12 MM294 (ATCC 31446); E. coli X1776 (ATCC 31537); штамм E. coli W3110 (ATCC 27325) и K5 772 (ATCC 53635). Другие подходящие прокариотические клетки-хозяева включают в себя Enterobacteriaceae, такие как Escherichia, например E. coli, Enterohacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, так же как Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанный в DD 266710, опубликованной 12 апреля 1989 г.), Pseudomonas, такой как P. aeruginosa, и Streptomyces. Данные примеры являются скорее иллюстративными, чем ограничивающими. Штамм W3110 является одним из особенно предпочтительных хозяев или родительских хозяев, поскольку он является общеупотребительным штаммом-хозяином для ферментаций продукта рекомбинантной ДНК. Предпочтительно клетка-хозяин секретирует минимальные количества протеолитических ферментов. Например, штамм W3110 можно модифицировать для эффекта генетической мутации в генах, кодирующих белки, эндогенные для хозяина, где примеры таких хозяев включают в себя штамм E. coli W3110 1A2, который обладает полным генотипом tonA; штамм E. coli W3110 9E4, который обладает полным генотипом tonA ptr3; штамм E. coli W3110 27C7 (ATCC 55244), который обладает полным генотипом tonA ptr3 phoA E15 (argF-lac)169 degP ompT kanr; штамм E. coli W3110 37D6, который обладает полным генотипом tonA ptr3 phoA E15 (argF-lac)169 degP ompT rbs7 ilvG kanr; штамм E. coli W3110 40B4, который представляет собой штамм 37D6 с делеционной мутацией degP и неустойчивостью к канамицину; и штамм E. coli, обладающий мутантной периплазматической протеазой, описанный в патенте США No 4946783, опубликованном 7 августа 1990. Альтернативно подходят способы клонирования in vitro, например PCR или другие реакции с полимеразой нуклеиновых кислот.
Кроме прокариот, подходящими клонирующими или экспрессирующими хозяевами для кодирующих PR087299 векторов являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи. Saccharomyces cerevisiae является общеупотребительным микроорганизмом-хозяином из низших эукариот. Другие включают в себя Schizosaccharomyces pombe (Beach and Nurse, Nature, 290: 140 [1981]; EP 139383, опубликованная 2 мая 1985 г.); хозяева Kluyveromyces (патент США No 4943529; Fleer et al., Bio/Technology, 9:968-975 (1991)) такие как, например, K. lactis (MW98-8C, CBS683, CBS4574; Louvencourt et al., J. Bacteriоl, 154(2):737-742 [1983]), K. fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906; Van den Berg et al., Bio/Technology, 8:135 (1990)), K. thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070; Sreekrishna et al., J. Basic Microbiol., 28:265-278 [1988]); Candida; Trichoderma reesia (EP 244234); Neurospora crassa (Case et al., Proc. Natl. Acad. Sci. USA, 76:5259-5263 [1979]); Schwanniomyces, такие как Schwanniomyces occidentalis (EP 394538, опубликованная 31 октября 1990 г.); и мицелиальные грибы, такие как, например, Neurospora, Penicillium, Tolypocladium (WO 91/00357, опубликованная 10 января 1991 г.), и хозяева Aspergillus, такие как A. nidulans (Ballance et al., Biochem. Biophys. Res. Commun.. 112:284-289 [1983]; Tilburn et al., Gene, 26:205-221 [1983]; Yelton et al., Proc. Natl. Acad. Sci. USA. 81: 1470-1474 [1984]) и A. niger (Kelly and Hynes, EMBO J., 4:475-479 [1985]). Метилотрофные дрожжи подходят здесь и включают в себя в качестве неограничивающих примеров дрожжи, способные расти на метаноле, выбранные из родов, состоящих из Hansenula, Candida, Kloeckera, Pichia, Saccharomyces, Torulopsis и Rhodotorula. Список конкретных видов, являющихся примерами этого класса дрожжей, можно найти в C. Anthony, The Biochemistry of Methylotrophs, 269 (1982).
Подходящие клетки-хозяева для экспрессии гликозилированного PRO87299 происходят из многоклеточных организмов. Примеры клеток беспозвоночных включают в себя клетки насекомых, таких как Drosophila S2 и Spodoptera Sf9, так же как клетки растений. Примеры подходящих линий клеток-хозяев млекопитающих включают в себя клетки яичников китайского хомячка (CHO) и клетки COS. Более конкретные примеры включают в себя линию из почек обезьяны CV1, трансформированную SV40 (COS-7, ATCC CRL 1651); линию эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol., 36:59 (1977)); клетки яичников китайского хомячка/-DHFR (CHO, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA. 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod., 23:243-251 (1980)); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); и опухоли молочной железы мыши (MMT 060562, ATCC CCL51). Подразумевается, что выбор подходящей клетки-хозяина лежит в компетенции специалистов в данной области.
3. Выбор и применение реплицирующегося вектора
Нуклеиновую кислоту (например, кДНК или геномную ДНК), кодирующую PRO87299, можно вставить в реплицирующийся вектор для клонирования (амплификации ДНК) или для экспрессии. Различные векторы являются публично доступными. Вектор может, например, присутствовать в форме плазмиды, космиды, вирусной частицы или фага. Подходящую последовательность нуклеиновой кислоты можно вставить в вектор посредством множества способов. Как правило, ДНК вставляют в подходящий участок(участки) рестрикционной эндонуклеазы с использованием способов, известных в данной области. Компоненты вектора, как правило, включают в себя в качестве неограничивающих примеров одну или несколько сигнальных последовательностей, точку начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции. Для конструирования подходящих векторов, содержащих один или несколько из этих компонентов, применяют общепринятые способы лигирования, известные опытному специалисту в данной области.
Рекомбинантный PRO87299 можно получать не только напрямую, но так же, как слитый полипептид с гетерологичным полипептидом, который может представлять собой сигнальную последовательность или другой полипептид, обладающий специфическим участком расщепления на N-конце зрелого белка или полипептида. Как правило, сигнальная последовательность может являться компонентом вектора, или она может являться частью ДНК, кодирующей PRO87299, которая вставлена в вектор. Сигнальная последовательность может представлять собой прокариотическую сигнальную последовательность, выбранную, например, из группы из лидеров щелочной фосфатазы, пенициллиназы, lpp, или термостабильного энтеротоксина II. Для секреции в дрожжах сигнальная последовательность может представлять собой, например, лидер инвертазы дрожжей, лидер альфа-фактора (включая лидеры α-факторов Saccharomyces и Kluyveromyces, последний описан в патенте США No 5010182), или лидер кислой фосфатазы, лидер глюкоамилазы C. albicans (EP 362179, опубликованная 4 апреля 1990), или сигнал, описанный в WO 90/13646, опубликованной 15 ноября 1990 г. При экспрессии в клетках млекопитающих, чтобы направлять секрецию белка, можно использовать сигнальные последовательности млекопитающих, такие как сигнальные последовательности из секретируемых полипептидов того же самого или родственных видов, так же как вирусные секреторные лидеры.
Как экспрессионные, так и клонирующие векторы содержат последовательность нуклеиновой кислоты, позволяющую вектору реплицироваться в одной или нескольких выбранных клетках-хозяевах. Такие последовательности хорошо известны для множества бактерий, дрожжжей и вирусов. Точка начала репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий, точка начала репликации плазмиды 2μ подходит для дрожжей, и различные вирусные точки начала репликации (SV40, полиомы, аденовируса, VSV или BPV) применимы для клонирующих векторов в клетках млекопитающих. Экспрессирующие и клонирующие векторы, как правило, будут содержать ген для селекции, называемый также селективным маркером. Типичные гены для селекции кодируют белки, которые (a) обеспечивают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату, или тетрациклину, (b) комплементируют ауксотрофную недостаточность, или (c) снабжают критическими питательными веществами, не доступными из комплексной среды, например, ген, кодирующий D-аланин-рацемазу для Bacilli.
Примером пригодных селективных маркеров для клеток млекопитающих являются маркеры, обеспечивающие идентификацию клеток, компетентных для поглощения нуклеиновой кислоты, кодирующей PRO87299, такие как DHFR или тимидинкиназа. При применении DHFR дикого типа подходящей клеткой-хозяином является линия клеток CHO, дефектных по активности DHFR, полученная и размноженная, как описано у Urlaub et al., Proc. Natl. Acad. Sci. USA, 77:4216 (1980). Пригодным геном для селекции для использования в дрожжах является ген trp1, присутствующий в плазмиде дрожжевой YRp7 [Stinchcomb et al., Nature, 282:39 (1979); Kingsman et al., Gene. 7:141 (1979); Tschemper et al., Gene, 10:157 (1980)]. Ген trp1 обеспечивает селективный маркер для мутантного штамма дрожжей, лишенных способности расти на триптофане, например, ATCC No 44076 или PEP4-1 [Jones, Genetics, 85:12 (1977)].
Экспрессирующие и клонирующие векторы обычно содержат промотор, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей PRO87299 для непосредственного синтеза мРНК. Промоторы, узнаваемые множеством потенциальных клеток-хозяев, хорошо известны. Промоторы, пригодные для использования в прокариотических хозяевах, включают в себя промоторные системы β-лактамазы и лактозы [Chang et al., Nature, 275:615 (1978); Goeddel et al., Nature, 281:544 (1979)], щелочной фосфатазы, промоторную систему триптофана (trp) [Goeddel, Nucleic acids Res., 8:4057 (1980); EP 36,776], и гибридные промоторы, такие как промотор tac [deBoer et al., Proc Natl. Acad. Sci. USA, 80: 21-25 (1983)]. Промоторы для использования в бактериальных системах будут содержать также последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей PRO87299.
Примеры пригодных промоторных последовательностей для использования в дрожжевых хозяевах включают в себя промоторы для 3-фосфоглицераткиназы [Hitzeman et al., J. Biol. Chem.. 255:2073 (1980)] или других гликолитических ферментов [Hess et al., J. Adv. Enzyme Reg., 7:149 (1968); Holland, Biochemistry, 17:4900 (1978)], таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируват декарбоксилаза, фосфофруктокиназа, глюкоза-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, изомераза фосфоглюкозы и глюкокиназа.
Другими промоторами дрожжей, представляюшими собой индуцируемые промоторы, обладающие дополнительным преимуществом транскрипции, контролируемой условиями роста, являются промоторные области алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, ферментов деградации, связанных с метаболизмом азота, металлотеонеина, глицеральдегид-3-фосфатдегидрогеназы, и ферментов, ответственных за утилизацию мальтозы и галактозы. Пригодные векторы и промоторы для использования для экспрессии в дрожжах дополнительно описаны в EP 73657.
Транскрипцию PRO87299 с векторов в клетках-хозяевах из млекопитающих контролируют, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус оспы-дифтерита птиц (UK 2211504, опубликованная 5 июля 1989 г.), аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B и вирус обезьян 40 (SV40), гетерологичными промоторами млекопитающих, например промоторами актина или иммуноглобулина, и промоторами теплового шока, при условии что такие промоторы совместимы с системами клетки-хозяина.
Транскрипцию ДНК, кодирующую PRO87299, у высших эукариот, можно увеличить вставкой в вектор энхансерной последовательности. Энхансеры представляют собой цис-действующие элементы ДНК, обычно приблизительно от 10 до 300 п.н., которые действуют на промотор для увеличения его транскрипции. В настоящее время известно много энхансерных последовательностей из генов млекопитающих (глобина, эластазы, альбумина, α- фетопротеина и инсулина). Как правило, однако, будут использовать энхансер из вируса эукариотической клетки. Примеры включают в себя энхансер SV40 на поздней стороне от точки начала репликации (п.н. 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне от точки начала репликации, и энхансеры аденовируса. Энхансер можно вставить в вектор в положении 5' или 3' к PRO87299 кодирующей последовательности, но предпочтительно его локализуют в участке 5' от промотора.
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (клетках дрожжей, грибов, насекомых, растений, животных, человека, или содержащих ядро клетках из других многоклеточных организмов), будут содержать также последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности из 5' и, иногда 3'-нетранслируемых областей эукариотических или вирусных ДНК или кДНК, являются общедоступными. Эти области содержат нуклеотидные фрагменты, транскрибируемые как полиаденилированные фрагменты в нетранслируемой части мРНК, кодирующей PRO87299.
Другие способы, векторы и клетки-хозяева, подходящие для адаптации для синтеза PRO87299 в культуре рекомбинантных клеток позвоночных, описаны в Gething et al., Nature. 293:620-625 (1981); Mantei et al., Nature, 281:40-46 (1979); EP 117060; и EP 117058.
4. Детекция амплификации/экспрессии гена
Амплификацию и/или экспрессию гена можно измерять в образце непосредственно, например, посредством общепринятого Саузерн-блоттинга, Нозерн-блоттинга для количественной оценки транскрипции мРНК [Thomas, Proc. Natl. Acad. Sci. USA, 77:5201-5205 (1980)], дот-блоттинга (анализ ДНК), или гибридизации in situ с использованием соответствующим образом меченного зонда на основе представленной здесь последовательности. Альтернативно можно применять антитела, которые могут узнавать специфические дуплексы, включая дуплексы ДНК, дуплексы РНК и гибридные дуплексы ДНК-РНК или дуплексы ДНК-белок. Антитела, в свою очередь, можно метить и можно проводить анализ, где дуплекс связан с подложкой, так что при формировании дуплекса на подложке можно детектировать антитело, связанное с дуплексом.
Альтернативно экспрессию гена можно измерять иммунологическими способами, такими как иммуногистохимическое окрашивание клеток или срезов ткани и анализ культуры клеток или жидкостей организма для непосредственной количественной оценки экспрессии продукта гена. Антитела, применимые для иммуногистохимического окрашивания и/или анализа жидкостей организма, могут являться либо моноклональными, либо поликлональными, и их можно получить в любом млекопитающем. Можно легко получить антитела против нативной последовательности полипептида PRO87299 или против синтетического пептида на основе последовательности ДНК, представленной здесь, или против экзогенной последовательности, слитой с ДНК PRO87299 и кодирующей специфический эпитоп антитела.
5. Очистка полипептида
Формы PRO87299 можно выделить из культуральной среды или из лизатов клетки-хозяина. Если они связаны с мембраной, их можно высвобождать из мембраны с использованием раствора подходящего детергента (например, Тритона-X 100) или ферментативного расщепления. Клетки, применяемые для экспрессии PRO87299, можно разрушить различными физическими или химическими способами, такими как циклы замораживания и оттаивания, обработка ультразвуком, механическое разрушение или лизирующими клетки средствами.
Может являться желательным очистить PRO87299 от рекомбинантных клеточных белков или полипептидов. Следующие способы являются примерами подходящих способов очистки: фракционирование на ионообменной колонке; осаждение этанолом; обращенно-фазовая HPLC; хроматография на оксиде кремния или на катионообменной смоле, такой как DEAE; хроматофокусирование; SDS-PAGE; осаждение сульфатом аммония; гель-фильтрация с использованием, например, Sephadex G-75; колонки с белок A сефарозой для удаления примесей, таких как IgG; и металл-хелатные колонки для связывания меченных эпитопом форм PRO87299. Можно применять различные способы очистки белка, и такие способы известны в данной области и описаны, например, в Deutscher, Methods in Enzymology, 182 (1990); Scopes, Protein Purification: Principles and Practice, Springer-Verlag, New York (1982). Выбранная стадия(стадии) очистки будет зависеть, например, от природы применяемого способа получения и конкретного полученного PRO87299.
E. Распределение в тканях
Локализацию тканей, экспрессирующих PRO87299, можно идентифицировать определением экспрессии мРНК в различных тканях человека. По локализации таких генов получают информацию о том, на какие ткани наиболее вероятно влияет стимулирующая и ингибирующая активность полипептидов PRO87299. По локализации гена в конкретной ткани получают также образец ткани для анализов блокирования активности, обсуждаемых ниже.
Как указано ранее, экспрессию генов в различных тканях можно измерять общепринятыми Саузерн-блоттингом, Нозерн-блоттингом для количественной оценки транскрипции мРНК (Thomas, Proc. Natl. Acad. ScL USA, 77:5201-5205 [1980]), дот-блоттингом (анализом ДНК) или гибридизацией in situ с использованием соответствующим образом меченного зонда на основе представленной здесь последовательности. Альтернативно можно применять антитела, которые могут узнавать специфические дуплексы, включая дуплексы ДНК, дуплексы РНК и гибридные дуплексы ДНК-РНК или дуплексы ДНК-белок.
Альтернативно экспрессию гена в различных тканях можно измерять иммунологическими способами, такими как иммуногистохимическое окрашивание срезов тканей и анализ культуры клеток или жидкостей организма для прямой количественной оценки экспрессии продукта гена. Антитела, применимые для иммуногистохимического окрашивания и/или анализа жидкостей организма, могут являться либо моноклональными, либо поликлональными, и их можно получить в любом млекопитающем. Можно легко получить антитела против нативной последовательности полипептида PRO87299 или против синтетического пептида на основе последовательности ДНК, кодирующей полипептид PRO87299, или против экзогенной последовательности, слитой с ДНК PRO87299 и кодирующей специфический эпитоп антитела. Общие способы получения антител и конкретные протоколы Нозерн-блоттинга и гибридизации in situ приведены ниже.
F. Исследования связывания антитела
Активность полипептидов PRO87299 можно далее верифицировать исследованиями связывания антитела, в которых тестируют способность анти-PRO87299 антител ингибировать эффект полипептидов PRO87299 соответственно в клетках ткани. Примерные антитела включают в себя поликлональные, моноклональные, гуманизированные, биспецифические, и гетероконъюгированные антитела, получение которых будет описано здесь ниже. Исследования связывания антитела можно проводить любым известным способом анализа, таким как анализы конкурентного связывания, прямые и непрямые сендвич-анализы, и анализы иммунопреципитации. Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc., 1987).
Анализы конкурентного связывания основаны на способности меченого стандарта конкурировать с аналитом тестового образца за связывание с ограниченным количеством антитела. Количество белка-мишени в тестовом образце обратно пропорционально количеству стандарта, который становится связанным с антителами. Для облегчения определения количества стандарта, становящегося связанным, предпочтительно понижают растворимость антител до или после конкуренции, так что стандарт и аналит, связанные с антителами, можно удобно отделить от стандрта и аналита, которые остаются несвязанными.
Сэндвич-анализы включают в себя использование двух антител, каждое из которых способно связываться с различной иммуногенной частью, или эпитопом подлежащего детекции белка. В сэндвич-анализе аналит в тестовом образце связан с первичным антителом, иммобилизованном на твердой подложке, и затем вторичное антитело связывется с аналитом, таким образом формируя нерастворимый комплекс из трех частей. См., например, патент США No 4376110. Вторичное антитело само по себе может являться меченным детектируемой группой (прямые сэндвич-анализы), или его можно измерять с использованием антииммуноглобулинового антитела, меченного детектируемой группой (непрямой сэндвич-анализ). Например, одним из типов сэндвич-анализа является анализ ELISA, в этом случае детектируемая группа представляет собой фермент.
Образец ткани для иммунохимии может являться свежим или замороженным или его можно погружать в парафин и фиксировать консервантом, таким как, например, формалин.
G. Анализы на основе клеток
Анализы на основе клеток и модели на животных для связанных с иммунитетом заболеваний можно использовать для дальнейшего понимания связи между генами и полипептидами, указанными в настоящем документе, и развитием и патогенезом связанного с иммунитетом заболевания.
В другом способе клетки из типа клеток, как известно, вовлеченных в конкретное связанное с иммунитетом заболевание, трансфицировали кДНК, описанными здесь, и анализировали способность этих кДНК стимулировать или ингибировать иммунную функцию. Подходящие клетки можно трансфицировать желаемым геном и контролировать активность иммунной функции. Такие трансфицированные линии клеток затем можно использовать для тестирования способности композиций поли- или моноклональных антител или антитела ингибировать или стимулировать иммунную функцию, например, чтобы регулировать пролиферацию T-клетки или инфильтрацию воспалительной клетки. Клетки, трансфицированные кодирующими последовательностями генов, указанными в настоящем документе, можно далее использовать для идентификации лекарственных средств-кандидатов для лечения связанных с иммунитетом заболеваний.
Кроме того, первичные культуры, полученные из трансгенных животных (как описано ниже) можно использовать в анализах на основе клеток здесь, хотя предпочтительными являются стабильные линии клеток. Способы получения непрерывных линий клеток хорошо известны в данной области (см., например, Small et al, MoI. Cell. Biol. 5: 642-648 [1985]).
Одним из подходящих анализов на основе клеток является реакция смешанных лимфоцитов (MLR). Current Protocols in Immunology, unit 3.12; edited by J E Coligan, A M Kruisbeek, D H Marglies, E M Shevach, W Strober, National Institutes of Health, Published by John Wiley & Sons, Inc. В данном анилизе исследуют способность тестируемого соединения стимулировать или ингибировать пролиферацию активированных T-клеток. Суспензию отвечающих T-клеток культивируют с аллогенными стимулирующими клетками и пролиферацию T-клеток измеряют по поглощению меченного тритием тимидина. Этот анализ представляет собой общее измерение реактивности T-клеток. Поскольку большинство T-клеток отвечают и при активации продуцируют IL-2, различия в отвечаемости в данном анализе частично отражают различия в продукции IL-2 отвечающими клетками. Результаты MLR можно верифицировать общепринятым анализом детекции лимфокина (IL-2). Current Protocols in Immunology, выше, 3.15, 6.3.
Пролиферативный ответ T-клеток в анализе MLR может являться обусловленным непосредственными митогенными свойствами анализируемых молекул или активацией, индуцируемой внешним антигеном. Активация T-клеток требует антигенспецифического сигнала, опосредованного через T-клеточный рецептор (TCR), и костимулирующего сигнала, опосредованного через связывающее взаимодействие со вторым лигандом, например, связывающее взаимодействие с B7 (CD80, CD86)/CD28. Перекрестное связывание с CD28 увеличивает секрецию лимфокина активированными T-клетками. Активация T-клеток обладает как отрицательным, так и положительным контролями через связывание лигандов, обладающих отрицательным или положительным эффектом. CD28 и CTLA-4 являются родственными гликопротеинами суперсемейства Ig, которые связываются с B7. Связывание CD28 с B7 обладает положительным костимулирующим эффектом на активацию T-клетки; связывание CTLA-4 с B7 напротив, напротив, обладает дезактивирующим эффектом на T-клетку. Chambers, C. A. and Allison, J. P., Curr. Opin. Immunol. (1997) 9:396. Schwartz, R. H., Cell (1992) 71; 1065; Linsey, P. S. и Ledbetter, J. A., Annu. Rev. Immunol. (1993) JUL: 191; June, C. H. et al, Immunol. Today (1994) 15:321; Jenkins, M. K., Immunity (1994) 1:405. При исследовании костимуляции анализировали костимулирующую или ингибирующую активность полипептидов PRO87299 для T-клеток.
Непосредственное применение стимулирующего соединения, как по изобретению, подтверждали в экспериментах с гликопротеином 4-IBB, членом семейства рецепторов фактора некроза опухоли, который связывается с лигандом (4-IBBL), экспрессированным на примированных T-клеток, и передает сигнал активации и роста T-клеток. Alderson, M. E. et al., J. Immunol. (1994) 24:2219.
Использование стимулирующего соединения-агониста также подтверждено экспериментально. Активация 4-1BB обработкой анти-4-1BB антителом-агонистом усиливает эрадикацию опухолей. Hellstrom, I. and Hellstrom, K. E., Crit. Rev. Immunol. (1998) 18:l. Терапия иммуноадъювантом для лечения опухолей, более подробно описанная ниже, является другим примером использования стимулирующего соединения по изобретению.
Альтернативно иммуностимулирующего или усиливающего эффекта можно достичь также введением PRO87299, который обладает свойствами, усиливающими свойства проницаемости сосудов. Усиленная проницаемость сосудов может являться благоприятной при расстройствах, которые можно уменьшить локальной инфильтрацией иммунных клеток (например, моноцитов, эозинофилов, PMN) и воспалением.
С другой стороны, полипептиды PRO87299, так же как другие соединения по изобретению, являющиеся прямыми ингибиторами пролиферации/активации T-клеток, секреции лимфокинов и/или проницаемости сосудов, можно непосредственно использовать для супрессии иммунного ответа. Данные соединения являются применимыми для уменьшения степени иммунного ответа и для лечения связанных с иммунитетом заболеваний, характеризующихся гиперактивным, сверхоптимальным или аутоиммунным ответом. Такое применение соединений по изобретению подтвердили экспериментами, описанными выше, в которых связывание CTLA-4 с рецептором B7 дезактивирует T-клетки. Непосредственно ингибирующие соединения по изобретению функционируют аналогичным образом. Ожидают, что использование соединений, подавляющих проницаемость сосудов, будет уменьшать воспаление. Такие применения будут благоприятными для условий лечения, связанных с избыточным воспалением.
Альтернативно соединения, например антитела, связывающиеся со стимулирующими полипептидами PRO87299 и блокирующие стимулирующий эффект этих молекул, оказывают сетевой ингибирующий эффект, и их можно использовать для подавления опосредованного T-клетками иммунного ответа посредством ингибирования пролиферации/активации T-клеток и/или посредством ингибирования секреции лимфокинов. Блокирование стимулирующего эффекта полипептидов подавляет иммунный ответ млекопитающего. Это применение подтвердили в экспериментах с использованием анти-IL2 антитела. В этих экспериментах антитело связывается с IL2 и блокирует связывание IL2 с его рецептором, таким образом обеспечивая ингибирующий T-клетку эффект.
H. Модели на животных
Результаты анализов на основе клеток in vitro можно далее верифицировать с использованием моделей на животных in vivo и анализов функционирования T-клеток. Множество хорошо известных моделей на животных можно использовать для дальнейшего понимания роли генов, указанных в настоящем документе, в развитии и патогенeзе связанного с иммунитетом заболевания, и для тестирования эффективности терапевтических средств, включая антитела, и другие антагонисты природных полипептидов, включая низкомолекулярные антагонисты. Сущность таких моделей in vivo делает их предсказательными для ответа у пациентов-людей. Модели связанных с иммунитетом заболеваний на животных включают в себя как нерекомбинантных, так и рекомбинантных (трансгенных) животных. Нерекомбинантные модели на животных включают в себя, например, модели на грызунах, например мышиные модели. Такие модели можно получить введением клеток в сингенную мышь с использованием общепринятых способов, например подкожной инъекции, инъекции в хвостовую вену, имплантации селезенки, интраперитонеальной имплантации, имплантации под капсулу почки и т.д.
Реакция трансплантат против хозяина возникает, когда иммунокомпетентные клетки трансплантируют пациентам с ослабленным иммунитетом или толерантным пациентам. Клетки донора распознают антигены хозяина и отвечают на них. Ответ может варьировать от угрожающего жизни тяжелого воспаления до умеренных случаев диареи и потери веса. Модели реакции трансплантат против хозяина предоставляют способы оценки реактивности T-клетки против антигенов MHC и минорных антигенов трансплантата. Подходящие способы подробно описаны в Current Protocols in Immunology, выше, часть 4.3.
Модели на животных для отторжения кожного аллотрансплантата являются средствами тестирования способности T-клеток опосредовать разрушение ткани in vivo и измерения их роли в отторжении трансплантата. В наиболее распространенных и принятых моделях используют мышиные трансплантаты хвост-кожа. В повторных экспериментах показали, что отторжение кожных аллотрансплантатов опосредовано T-клетками, T-клетками-хелперами и T-клетками килерами-эффекторами, а не антителами. Auchincloss, H. Jr. and Sachs, D. H., Fundamental Immunology, 2nd ed., W. E. Paul ed., Raven Press, NY, 1989, 889-992. Подходящие способы подробно описаны в Current Protocols in Immunology, выше, часть 4.4. Другими моделями отторжения трансплантата, которые можно использовать для тестирования соединений по изобретению, являются модели аллогенного сердечного трансплантата, описанные Tanabe, M. et al, Transplantation (1994) 58:23 и Tinubu, S. A. et al, J. Immunol. (1994) 4330-4338.
Модели на животных для гиперчувствительности замедленного типа обеспечивают также анализ опосредованной клетками иммунной функции. Реакции гиперчувствительности замедленного типа представляют собой иммунный ответ, опосредованный T-клетками in vivo, характеризующийся воспалением, который не достигает пика, пока не пройдет период времени после антигенного стимула. Эти реакции возникают также при тканеспецифических аутоиммунных заболеваниях, таких как рассеянный склероз (MS) и экспериментальный аутоиммунный энцефаломиелит (EAE, модель MS). Подходящий способ подробно описан в Current Protocols in Immunology, выше, часть 4.5.
EAE представляет собой аутоиммунное заболевание, опосредованное T-клетками, характеризующееся воспалением с участием T-клеток и мононуклеарных клеток и последующей демиелинизацией аксонов в центральной нервной системе. EAE, в основном считают подходящей моделью на животных для MS у человека. Bolton, C, Multiple sclerosis (1995) 1:143. Разработаны как модели с обострением, так и модели с обострением-ремиссией. Соединения по изобретению можно тестировать по стимулирующей или ингибирующей T-клетки активности против иммуноопосредованного демиелинизирующего заболевания с использованием способа, описанного в Current Protocols in Immunology, выше, части 15.1 и 15.2. Смотри также модели заболевания миелина, при котором олигодендроциты или Шванновские клетки трансплантируют в центральную нервную систему, как описано в Duncan, I. D. et al, Molec. Med. Today (1997) 554-561.
Контактная гиперчувствительность представляет собой простой анализ in vivo гиперчувствительности замедленного типа для опосредованной клетками иммунной функции. В данном способе кожу подвергают воздействию экзогенных гаптенов, что вызывает реакцию гиперчувствительности замедленного типа, которую измеряют и оценивают количественно. Контактная чувствительность включает в себя фазу начальной сенсибилизации с последующей фазой активации. Фаза активации происходит, когда T лимфоциты сталкиваются с антигеном, с которым они контактировали ранее. Происходит отек и воспаление, что делает ее отличной моделью аллергического контактного дерматита человека. Подходящий способ подробно описан в Current Protocols in Immunology, Eds. J. E. Cologan, A. M. Kruisbeek, D. H. Margulies, E. M. Shevach и W. Strober, John Wiley & Sons, Inc., 1994, unit 4.2. Смотри также Grabbe, S. and Schwarz, T, Immun. Today 19 (1): 37-44 (1998).
Моделью артрита на животных является индуцированный коллагеном артрит. Эта модель обладает общими клиническими, гистологическими и иммунологическими характеристиками с аутоиммунным ревматоидным артритом человека и является приемлемой моделью аутоиммунного артрита человека. Модели на мыши и крысе характеризуются синовитом, эрозией хряща и субхондральной кости. Соединения по изобретению можно тестировать по активности против аутоиммунного артрита с использованием способов, описанных в Current Protocols in Immunology, выше, части 15.5. См. также модель с использованием моноклонального антитела к CD18 и интегринов VLA-4, описанную в Issekutz, A.C. et al, Immunology (1996) 88:569.
Описана модель астмы, в которой индуцированную антигеном гиперреактивность, легочную эозинофилию и воспаление индуцировали сенсибилизацией животного овальбумином и затем стимуляцией животного тем же белком, введенным в аэрозоли. Для некоторых моделей на животных (морской свинке, крысе, не относящемся к человеку примате) показаны симптомы, сходные с симптомами атопической бронхиальной астмы при стимуляции аэрозольными антигенами. Модели на мышах обладают многими свойствами астмы человека. Подходящие способы тестирования соединений по изобретению по активности и эффективности для лечения астмы описаны в Wolyniec, W. W. et al, Am. J. Respir. Cell MoI. Biol. (1998) 18:777 и процитированных там ссылках.
Кроме того, соединения по изобретению можно тестировать на моделях на животных для заболеваний, подобных псориазу. Факты позволяют предполагать для псориаза T-клеточный патогенeз. Соединения по изобретению можно тестировать в модели на мышах scid/scid, описанной Schon, M. P. et al, Nat. Med. (1997) 3:183, в которой для мышей показаны гистопатологические повреждения кожи, сходные с псориазом. Другой подходящей моделью является химера кожа человека/мышь scid, полученная, как описано Nickoloff, B. J. et al, Am. J. Path. (1995) 146:580.
Рекомбинантные (трансгенные) модели на животных можно конструировать введением кодирующей части генов, указанных в настоящем документе, в геном интересующих животных с использованием общепринятых способов получения трансгенных животных. Животные, которых можно применять в качестве мишени для трансгенной манипуляции, включают в себя в качестве неограничивающих примеров мышей, крыс, кроликов, морских свинок, овец, коз, свиней, и не относящихся к человеку приматов, например павианов, шимпанзе и мартышек. В данной области известны способы введения трансгена в таких животных, включая микроинъекцию в пронуклеус (Hoppe and Wanger, патент США No 4873191); опосредованный ретровирусом перенос в зародышевые линии (например, Van der Putten et al., Proc. Natl. Acad. Sci USA 82, 6148-615 [1985]); направленное воздействие на ген в эмбриональных стволовых клетках (Thompson et al., Cell 56, 313-321 [1989]); электропорация эмбрионов (Lo, Mol. Cel. Biol. 3, 1803-1814 [1983]); опосредованный спермиями перенос генов (Lavitrano et al., Cell 57, 717-73 [1989]). Обзор см., например, в патенте США No 4736866.
Для целей настоящего изобретения трансгенные животные включают в себя тех, которые несут трансген только в части их клеток («мозаичные животные»). Трансген можно вводить или как отдельный трансген, или в конкатемерах, например тандемах голова-к-голове или голова-к-хвосту. Избирательное введение трансгена в конкретный тип клеток возможно также, например, следующим способом Lasko et al., Proc. Natl. Acad. ScL USA 89, 6232-636 (1992).
Экспрессию трансгена в трансгенных животных можно контролировать общепринятыми способами. Например, анализы Саузерн-блоттингом или амплификацией PCR можно применять для проверки интеграции трансгена. Затем можно анализировать уровень экспрессии мРНК с использованием таких способов, как гибридизация in situ, анализы Нозерн-блоттингом, PCR или иммуноцитохимию. У животных можно далее исследовать патологичесикие признаки иммунного заболевания, например, гистологическим исследованием для определения инфильтрации иммунных клеток в конкретные ткани. Можно проводить также эксперименты по блокированию, в которых трансгенных животных обрабатывают соединениями по изобретению для определения степени стимуляции или подавления соединениями пролиферации T-клеток. В этих экспериментах животному вводят блокирующие антитела, которые связываются с полипептидом PRO87299, полученные, как описано выше, и определяют эффект на иммунную функцию.
Альтернативно можно сконструировать животных с «нокаутом», обладающих дефектным или измененным геном, кодирующим полипептид, идентифицированный здесь, в результате гомологичной рекомбинации между эндогенным геном, кодирующим полипептид, и измененной геномной ДНК, кодирующей тот же полипептид, введенной в эмбриональную клетку животного. Например, кДНК, кодирующую конкретный полипептид, можно использовать для клонирования геномной ДНК, кодирующей этот полипептид, в соответствии с общепринятыми способами. Часть геномной ДНК, кодирующей конкретный полипептид, можно делетировать или заменить другим геном, таким как ген, кодирующий селективный маркер, который можно использовать для контроля интеграции. Как правило, в вектор включают несколько тысяч пар нуклеотидов неизмененной фланкирующей ДНК (и на 5'-, и на 3'-концах) [см., например, описание векторов для гомологичной рекомбинации в Thomas and Capecchi, Cell, 51:503 (1987)]. Вектор вводят в линию эмбриональных стволовых клеток (например, электропорацией) и отбирают клетки, в которых произошла гомологичная рекомбинация введенной ДНК с эндогенной ДНК [см., например, Li et al., Cell, 69:915 (1992)]. Затем отобранные клетки инъецируют в бластоцисту животного (например, мыши или крысы) для получения агрегированных химер [см., например, Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp. 113-152]. Затем химерный эмбрион можно имплантировать в подходящую ложнобеременную патронатную самку животного, которая вынашивает эмбриона до срока для получения животного «с нокаутом». Потомство, несущее гомологично рекомбинировавшую ДНК в половых клетках, можно идентифицировать общепринятыми способами и использовать для выведения животных, у которых все клетки содержат гомологично рекомбинировавшую ДНК. Животных с нокаутом можно характеризовать, например, по их способности к защите против конкретных патологических состояний и по развитию у них патологических состояний из-за отсутствия полипептида.
I. Иммуноадъювантная терапия
В одном из вариантов осуществления иммуностимулирующие соединения по изобретению можно применять в иммуноадъювантной терапии для лечения опухолей (рака). В настоящее время является общепризнанным, что T-клетки узнают опухолеспецифические антигены человека. Одна группа опухолевых антигенов, кодируемых семействами генов MAGE, BAGE и GAGE, является молчащей во всех нормальных тканях взрослых, но в значительных количествах экспрессируется в опухолях, таких как меланомы, опухоли легкого, опухоли головы и шеи, и карциномы мочевого пузыря. DeSmet, C. et ah, (1996) Proc. Natl. Acad. Sci. USA, 93:7149. Показано, что костимуляция T-клеток индуцирует регрессию опухоли и противоопухолевый ответ как in vitro, так и in vivo. Melero, I. et al, Nature Medicine (1997) 3:682; Kwon, E. D. et al, Proc. Natl. Acad. Sci. USA (1997) 94: 8099; Lynch, D. H. et al, Nature Medicine (1997) 3:625; Finn, O. J. and Lotze, M. T., J. Immunol. (1998) 21:114. Стимулирующие соединения по изобретению можно вводить в качестве адъювантов, отдельно или вместе с регулирующим рост средством, цитотоксическим средством или химиотерапевтическим средством для стимуляции пролиферации/активации T-клетки и противоопухолевого ответа на опухолевые антигены. Регулирующее рост, цитотоксическое или химиотерапевтическое средство можно вводить в общепринятых количествах с использованием известных режимов введения. Иммуностимулирующая активность соединений по изобретению позволяет снижать количества регулирующих рост, цитотоксических, или химиотерапевтических средств, таким образом потенциально снижая токсичность для пациента.
J. Анализы скрининга кандидатов на лекарственное средство
Анализы скрининга кандидатов на лекарственное средство разработаны для идентификации соединений, которые связываются или образуют комплекс с полипептидами, кодируемыми указанными в настоящем документе генами, или их биологически активными фрагментами, или иным образом мешают взаимодействию кодируемых полипептидов с другими клеточными белками. Такие анализы скрининга будут включать в себя анализы, пригодные для высокопроизводительного скрининга химических библиотек, что делает их особенно подходящими для идентификации низкомолекулярных кандидатов на лекарственное средство. Рассматриваемые малые молекулы включают в себя синтетические органические или неорганические соединения, включая пептиды, предпочтительно растворимые пептиды, слитые белки (поли)пептид-иммуноглобулин, и, в частности, антитела, включая, без ограничения, поли- и моноклональные антитела и фрагменты антитела, одноцепочечные антитела, антиидиотипические антитела и химерные или гуманизированные варианты таких антител или фрагментов, так же как антитела и фрагменты антител человека. Анализы можно проводить во множестве форматов, включая анализы белок-белкового связывания, анализы биохимического скрининга, иммуноанализы и анализы на основе клеток, хорошо охарактеризованные в данной области. Общим для всех анализов является то, что они включают в себя контакт кандидата на лекарственное средство с полипептидом, кодируемым указанной в настоящем документе нуклеиновой кислотой при условиях и в течение времени, позволяющих взаимодействие этих двух компонентов.
В анализах связывания взаимодействие представляет собой связывание, и сформированный комплекс можно выделить или детектировать в реакционной смеси. В конкретном варианте осуществления полипептид, кодируемый указанным в настоящем документе геном, или кандидат на лекарственное средство иммобилизуют на твердой фазе, например на планшете для микротитрования, посредством ковалентных или нековалентных присоединений. Нековалентное присоединение, как правило, осуществляют покрытием твердой поверхности раствором полипептида и высушиванием. Альтернативно иммобилизованное антитело, например моноклональное антитело, специфическое для подлежащего иммобилизации полипептида, можно использовать для его заякоривания на твердой поверхности. Анализ осуществляют добавлением неиммобилизованного компонента, который можно метить поддающейся детекции меткой, к иммобилизованному компоненту, например покрытой поверхности, содержащей заякоренный компонент. После завершения реакции непрореагировавшие компоненты удаляют, например, отмывкой, и комплексы, заякоренные на твердой поверхности, детектируют. Когда первоначально не иммобилизованный компонент несет детектируемую метку, детекция метки, иммобилизованной на поверхности, указывает на образование комплексов. Когда первоначально не иммобилизованный компонент не несет детектируемой метки, образование комплекса можно детектировать, например, с использованием меченого антитела, специфически связывающего иммобилизованный комплекс.
Если соединение-кандидат взаимодействует, но не связывается с конкретным белком, кодируемым указанным в настоящем документе геном, его взаимодействие с этим белком можно анализировать способами, хорошо известными для детекции белок-белковых взаимодействий. Такие анализы включают в себя общепринятые способы, такие как перекрестное сшивание, ко-иммунопреципитация и совместная очистка на градиентах или хроматографических колонках. Кроме того, белок-белковые взаимодействия можно контролировать с использованием генетической системы на основе дрожжей, описанной Fields и соавторами [Fields and Song, Nature (London) 340, 245-246 (1989); Chien et al, Proc. Natl. Acad. ScL USA 88, 9578-9582 (1991)], как описано Chevray and Nathans, Proc. Natl. Acad. Sci USA 89, 5789-5793 (1991). Многие транскрипционные активаторы, такие как GAL4 дрожжей, состоят из двух физически раздельных модулярных доменов, один из которых действует как ДНК-связывающий домен, в то время как другой функционирует как активирующий транскрипцию домен. В системе экспрессии в дрожжах, описанной в вышеупомянутых публикациях (обычно обозначаемой «двугибридная система»), пользуются преимуществом этого свойства и применяют два гибридных белка: один, в котором белок-мишень слит с ДНК-связывающим доменом GAL4, и другой, в котором кандидаты на активирующие белки слиты с активирующим доменом. Экспрессия репортерного гена GAL1-lacZ под контролем активируемого GAL4 промотора зависит от восстановления активности GAL4 посредством белок-белкового взаимодействия. Колонии, содержащие взаимодействующие полипептиды, детектируют хромогенным субстратом для β-галактозидазы. Полный набор (MATCHMAKER™) для идентификации белок-белковых взаимодействий между двумя конкретными белками с использованием двугибридного способа коммерчески доступен от Clontech. Эту систему можно также распространить на картирование доменов белка, вовлеченных в специфические белковые взаимодействия, так же как на точное определения аминокислотных остатков, критических для этих взаимодействий.
Чтобы найти и тестировать соединения, которые мешают взаимодействию указанного в настоящем документе гена и других внутри- или внеклеточных компонентов, обычно получают реакционную смесь, содержащую продукт гена и внутри- или внеклеточный компонент при условиях и в течение времени, позволяющих взаимодействие и связывание двух продуктов. Для проверки способности тестируемого соединения ингибировать связывание, реакцию проводят в отсутствие и в присутствии тестируемого соединения. Кроме того, в третью реакционную смесь можно добавлять плацебо, что служит положительным контролем. Связывание (формирование комплекса) между тестируемым соединением и внутри- или внеклеточным компонентом, присутствующим в смеси, контролируют, как указано выше. Формирование комплекса в контрольной реакционной смеси (смесях), но не в реакционной смеси, содержащей тестируемое соединение, указывает на то, что тестируемое соединение мешает взаимодействию тестируемого соединения и его реакционного партнера.
K. Композиции и способы для лечения связанных с иммунитетом заболеваний
Композиции, применимые для лечения связанных с иммунитетом заболеваний, включают в себя без ограничения белки, антитела, малые органические молекулы, пептиды, фосфопептиды, антисмысловые и рибозимные молекулы, молекулы с тройной спиралью, и т.д., которые подавляют или стимулируют иммунную функцию, например пролиферацию/активацию T-клетки, высвобождение лимфокинов или инфильтрацию иммунной клетки.
Например, антисмысловые молекулы РНК и ДНК действуют, чтобы напрямую блокировать трансляцию мРНК посредством гибридизации с намеченной мРНК и предупреждения трансляции белка. Когда используют антисмысловую ДНК, предпочтительными являются олигодезоксирибонуклеотиды, полученные из участка инициации трансляции, например, между положениями приблизительно -10 и +10 в нуклеотидной последовательности гена-мишени.
Рибозимы представляют собой ферментативные молекулы РНК, способные катализировать специфическое расщепление РНК. Рибозимы действуют посредством специфической для последовательности гибридизации с комплементарной РНК-мишенью с последующим эндонуклеолитическим расщеплением. Конкретные участки расщепления рибозимом в потенциальной РНК-мишени можно идентифицировать известными способами. Дополнительные подробности см., например, в Rossi, Current Biology 4, 469-471 (1994), и публикации PCT No WO 97/33551 (опубликована 18 сентября 1997).
Молекулы нуклеиновой кислоты молекулы для формирования тройной спирали, используемые для подавления транскрипции, должны являться одноцепочечными и состоящими из дезоксинуклеотидов. Состав оснований этих олигонуклеотидов рассчитывают так, чтобы он способствовал формированию тройной спирали по правилам спаривания оснований Хугстэна, что в основном требует протяженных участков пуринов или пиримидинов на одной цепи дуплекса. Дополнительные подробности см., например, в публикации PCT No WO 97/33551, выше.
Эти молекулы можно идентифицировать посредством любых анализов скрининга или любого сочетания любых анализов скрининга, обсуждаемых выше, и/или посредством любых способов скрининга, хорошо известных специалистам в данной области.
L. Анти-PRO87299 антитела
Настоящее изобретение далее относится к анти-PRO87299 антителам. Примеры антител включают в себя поликлональные, моноклональные, гуманизированные, биспецифические и гетероконъюгированные антитела.
1. Поликлональные антитела
Анти-PRO87299 антитела могут включать в себя поликлональные антитела. Способы получения поликлональных антител известны опытному специалисту в данной области. Образование поликлональных антител можно индуцировать у млекопитающих, например, одной или несколькими инъекциями иммунизирующего вещества и, если желательно, адъюванта. Как правило, иммунизирующее вещество и/или адъювант можно инъецировать млекопитающему посредством множественных подкожных или внутрибрюшинных инъекций. Иммунизирующее вещество может включать в себя полипептид PRO87299 или слитый с ним белок. Может являться полезным конъюгировать иммунизирующее вещество с белком, известным как иммуногенное для иммунизируемого млекопитающего. Примеры таких иммуногенных белков включают в себя в качестве неограничивающих примеров гемоцианин морского блюдечка, сывороточный альбумин, бычий тиреоглобулин и соевый ингибитор трипсина. Примеры адъювантов, которые можно применять, включают в себя полный адъювант Фрейнда и адъювант MPL-TDM (монофосфорил липид A, синтетический дикориномиколат трегалозы). Специалист в данной области может выбрать способ иммунизации без излишних экспериментов.
2. Моноклональные антитела
Анти-PRO87299 антитела альтернативно могут представлять собой моноклональные антитела. Моноклональные антитела можно получить с использованием способов гибридомы, таких как описанные в Kohler and Milstein, Nature, 256:495 (1975). В способе гибридомы мышь, хомяка или другого подходящего животного-хозяина, как правило, иммунизируют иммунизирующим веществом для активации лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с иммунизирующим веществом. Альтернативно можно иммунизировать лимфоциты in vitro.
Иммунизирующее вещество, как правило, будет включать в себя полипептид PRO87299 или слитый с ним белок. Как правило, используют или лимфоциты периферической крови («PBL»), если желательны клетки человеческого происхождения, или клетки селезенки или клетки лимфоузлов, если желательны источники из не относящихся к человеку млекопитающих. Затем лимфоциты сливают с иммортализованной линией клеток с использованием подходящего вызывающего слияние клеток вещества, такого как полиэтиленгликоль, для получения клетки гибридомы [Goding, Monoclonal antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]. Иммортализованные линии клеток обычно представляют собой трансформированные клетки млекопитающих, в частности клетки миеломы, относящиеся к грызунам, быку и человеку. Обычно применяют линии клеток миеломы крысы или мыши. Клетки гибридомы можно культивировать в подходящей культуральной среде, предпочтительно содержащей одно или несколько веществ, подавляющих рост или выживание не слитых иммортализованных клеток. Например, если в родительских клетках отсутствует фермент гипоксантин-гуанин-фосфорибозилтрансфераза (HGPRT или HPRT), культуральная среда для гибридомы, как правило, будет содержать гипоксантин, аминоптерин и тимидин («среда HAT») - вещества, предотвращающие рост HGPRT-дефектных клеток.
Предпочтительными иммортализованными линиями клеток являются клетки, которые эффективно сливаются, поддерживают высокий уровень экспрессии антитела выбранными продуцирующими антитело клетками и являются чувствительными к среде, такой как среда HAT. Более предпочтительными иммортализованными линиями клеток являются линии миеломы мышей, которые можно получить, например, из Salk Institute Cell Distribution Center, San Diego, California и American Type Culture Collection, Manassas, Virginia. Описана также продукция моноклональных антител в линиях клеток миеломы человека и гетеромиеломы мыши-человека [Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal antibody Production Techniques и Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63].
Затем в культуральной среде, в которой культивировали клетки гибридомы, можно анализировать присутствие моноклональных антител, направленных против PRO87299. Предпочтительно специфичность связывания моноклонального антитела, продуцируемого клетками гибридомы, определяют иммунопреципитацией или анализом связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие способы и анализы известны в данной области. Аффинность связывания моноклонального антитела можно, например, определять анализом Скэтчарда по Munson and Pollard, Anal. Biochem., 107:220 (1980).
После идентификации желательных клеток гибридомы клоны можно субклонировать способами лимитирующих разведений и выращивать общепринятыми способами [Goding, выше]. Подходящая для данной цели культуральная среда включает в себя, например, среду Игла, модифицированную Дульбекко и среду RPMI-1640. Альтернативно клетки гибридомы можно выращивать in vivo в форме асцитов у млекопитающего.
Моноклональные антитела, секретируемые субклонами, можно выделять или очищать из культуральной среды или асцитной жидкости общепринятыми способами очистки иммуноглобулина, такими как, например, очистка на белок A-сефарозе, хроматография на гидроксилапатите, гель-электрофорез, диализ, или аффинная хроматография.
Моноклональные антитела можно получить также способами рекомбинантных ДНК, такими, как описаны в патенте США No. 4816567. ДНК, кодирующие моноклональные антитела по изобретению, можно легко выделить и секвенировать с применением общепринятых способов (например, с применением олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Клетки гибридомы по изобретению являются подходящим источником такой ДНК. После выделения ДНК можно поместить в экспрессирующие векторы, которыми затем трансфицируют клетки-хозяева, такие как обезьяньи клетки COS, клетки яичников китайского хомячка (CHO) или клетки миеломы, которые иначе не продуцируют белок иммуноглобулин, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК также можно модифицировать, например, заменой кодирующими последовательностями константных доменов тяжелых и легких цепей человека гомологичных мышиных последовательностей [патент США No. 4816567; Morrison et al., выше] или ковалентным присоединением к кодирующей последовательности иммуноглобулина всей или части кодирующей последовательности не относящегося к иммуноглобулинам полипептида. Таким не относящимся к иммуноглобулинам полипептидом можно заменить константные домены антитела по изобретению или заменить их вариабельными доменами совмещающегося с антигеном участка антитела по изобретению для получения химерного бивалентного антитела.
Антитела могут представлять собой моновалентные антитела. Способы получения моновалентных антител хорошо известны в данной области. Например, один из способов включает в себя рекомбинантную экспрессию легкой цепи иммуноглобулина и модифицированной тяжелой цепи. Как правило, тяжелую цепь укорачивают в какой-либо точке Fc-области, чтобы предотвратить перекрестное сшивание тяжелых цепей. Альтернативно существенные остатки цистеина заменяют другим аминокислотным остатком или делетируют для предотвращения перекрестного сшивания.
Способы in vitro также пригодны для получения моновалентных антител. Расщепление антител для получения их фрагментов, особенно Fab-фрагментов, можно осуществлять с применением общепринятых способов, известных в данной области.
3. Человеческие и гуманизированные антитела
Анти-PRO87299 антитела по изобретению могут дополнительно относиться к гуманизированным антителам или человеческим антителам. Гуманизированные формы не относящихся к человеку (например, мышиных) антител представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие связывающие антиген последовательности антител), содержащие минимальную последовательность, полученную из не относящегося к человеку иммуноглобулина. Гуманизированные антитела включают в себя человеческие иммуноглобулины (антитело-реципиент), в которых остатки из определяющей комплементарность области (CDR) рецепиента заменяют остатками из CDR не относящихся к человеку видов (антитело-донор), таких как мышь, крыса или кролик, обладающими желаемой специфичностью, аффинностью и емкостью. В некоторых случаях каркасные остатки Fv человеческого иммуноглобулина заменяют соответствующими не относящимися к человеку остатками. Гуманизированные антитела могут также содержать остатки, не обнаруженные ни в антителе-реципиенте, ни в импортируемых последовательностях CDR или каркаса. Как правило, гуманизированное антитело содержит в основном все из, по меньшей мере, одного, а обычно двух вариабельных доменов, в которых все или в основном все области CDR соответствуют областям не относящегося к человеку иммуноглобулина и все или в основном все области FR представляют собой области из консенсусной последовательности человеческого иммуноглобулина. Оптимально, гуманизированное антитело будет содержать также, по меньшей мере, часть константной области иммуноглобулина (Fc), обычно из человеческого иммуноглобулина [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-329 (1988); and Presta, Curr. Op. Struct. Biol., 2:593-596 (1992)].
Способы гуманизации не относящихся к человеку антител хорошо известны в данной области. Как правило, гуманизированное антитело обладает одним или несколькими аминокислотными остатками, введенными в него из не относящегося к человеку источника. Эти не относящиеся к человеку аминокислотные остатки часто обозначают как «импортируемые» остатки, которые, как правило, берут из «импортируемого» вариабельного домена. Гуманизацию по существу можно провести по способу Winter и соавторов [Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)] посредством замены заменой CDR или последовательностями CDR грызунов соответствующих последовательностей антитела человека. Следовательно, такие «гуманизированные» антитела представляют собой химерные антитела (патент США No 4816567), где по существу менее чем один интактный человеческий вариабельный домен заменен соответствующей последовательностью из не относящихся к человеку видов. На практике гуманизированные антитела, как правило, представляют собой человеческие антитела, в которых некоторые остатки CDR и, возможно, некоторые остатки FR заменены остатками из аналогичных участков антител грызунов.
Человеческие антитела можно получать также с применением различных способов, известных в данной области, включая библиотеки фагового дисплея [Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991)]. Способы Cole et al. и Boerner et al. также доступны для получения человеческих моноклональных антител (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985) и Boerner et al., J. Immunol., 147(1):86-95 (1991)]. Подобным образом человеческие антитела можно получить введением локусов человеческих иммуноглобулинов в трансгенных животных, например мышей, у которых эндогенные гены иммуноглобулинов частично или полностью инактивированы. При стимуляции обнаружена продукция человеческих антител, близко сходная с обнаруженной у человека продукцией во всех отношениях, включая реаранжировку генов, сборку и репертуар антител. Данный способ описан, например, в патентах США No 5545807, 5545806, 5569825, 5625126, 5633425, 5661016 и в следующих научных публикациях: Marks et al., Bio/Technology 10, 779-783 (1992); Lonberg et al., Nature 368 856-859 (1994); Morrison, Nature 368, 812-13 (1994); Fishwild et al., Nature Biotechnology 14, 845-51 (1996); Neuberger, Nature Biotechnology 14, 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13 65-93 (1995).
Можно осуществлять также аффинное созревание антител с применением известных способов отбора и/или мутагенеза, как описано выше. Предпочтительно аффинно зрелые антитела обладают аффинностью в пять раз, более предпочтительно в 10 раз, даже более предпочтительно в 20 или 30 раз большей, чем у исходного антитела (как правило, мышиного, гуманизированного или человеческого), из которого получают зрелое антитело.
4. Биспецифические антитела
Биспецифические антитела представляют собой моноклональные, предпочтительно человеческие или гуманизированные, антитела, обладающие специфичностями связывания, по меньшей мере, для двух различных антигенов. В настоящем случае присутствует одна из связывающих специфичностей для домена PRO87299, другая для любого другого антигена, и предпочтительно для белка клеточной поверхности или рецептора или субъединицы рецептора.
Способы получения биспецифических антител известны в данной области. Обычно рекомбинантная продукция биспецифических антител основана на коэкспрессии двух пар тяжелая цепь/легкая цепь иммуноглобулина, где две тяжелые цепи обладают различными специфичностями. [Milstein and Cuello, Nature, 305:537-539 (1983)]. Из-за случайного ассортимента тяжелых и легких цепей иммуноглобулинов данные гибридомы (квадромы) продуцируют потенциальную смесь из десяти различных молекул антител, из которых только одно обладает корректной биспецифической структурой. Выделение корректной молекулы обычно включает в себя стадии аффинной хроматографии. Подобные процедуры описаны в WO 93/08829, опубликованном 13 мая 1993, и у Traunecker et al., EMBO J., 10:3655-3659 (1991).
Вариабельные домены антител с желаемыми связывающими специфичностями (участками совмещения антитело-антиген) можно присоединять к последовательностям константных доменов иммуноглобулинов. Слитый белок предпочтительно содержит константный домен тяжелой цепи иммуноглобулина, содержащий, по меньшей мере, часть шарнирных областей CH2 и CH3. Предпочтительно, чтобы первая константная область тяжелой цепи (CH1), содержащая участок, необходимый для связывания легкой цепи, присутствовала, по меньшей мере, в одном из слитых белков. ДНК, кодирующие слитые белки с тяжелыми цепями иммуноглобулинов и, если желательно, с легкими цепями иммуноглобулинов, вставляют в различные экспрессирующие векторы и котрансфицируют в подходящий организм-хозяин. Дополнительные детали получения биспецифических антител см., например, в Suresh et al., Methods in Enzymology, 121:210 (1986).
Согласно другому способу, описанному в WO 96/27011, границу раздела между парой молекул антител можно модифицировать для максимизации процента гетеродимеров, выделенных из культуры рекомбинантных клеток. Предпочтительная граница раздела содержит, по меньшей мере, часть CH3-области константного домена антитела. По данному способу одну или несколько малых боковых цепей аминокислот из границы раздела первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозина или триптофана). Компенсаторные «полости» такого же размера, как большая боковая цепь (цепи), или похожего размера создают на границе раздела второй молекулы антитела посредством замены больших боковых цепей аминокислот на меньшие (например, аланин или треонин). Это предоставляет механизм для увеличения выхода гетеродимера выше, чем у других нежелательных конечных продуктов, таких как гомодимеров.
Биспецифические антитела можно получить как полноразмерные антитела или фрагменты антител (например, F(ab')2 биспецифические антитела). Способы получения биспецифических антител из фрагментов антител описаны в литературе. Например, биспецифические антитела можно получать с применением химического связывания. Brennan et al., Science 229:81 (1985) описывают способ, где интактные антитела протеолитически расщепляют для получения F(ab')2-фрагментов. Эти фрагменты восстанавливают в присутствии комплексообразователя с дитиолом арсенита натрия для стабилизации соседних дитиолов и предотвращения формирования междумолекулярных дисульфидов. Полученные Fab'-фрагменты затем переводят в производные тионитробензоата (TNB). Одно из производных Fab'-TNB затем переводят обратно в Fab'-тиол посредством восстановления с меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB для формирования биспецифического антитела. Полученные биспецифические антитела можно применять в качестве средств избирательной иммобилизации ферментов.
Fab'-фрагменты можно выделять непосредственно из E. coli и соединять химически для получения биспецифических антител. Shalaby et al., J. Exp. Med. 175:217-225 (1992) описали получение молекулы полностью гуманизированного биспецифического антитела F(ab')2. Каждый Fab'-фрагмент отдельно секретировали из E. coli и подвергали прямому химическому связыванию in vitro для получения биспецифического антитела. Биспецифическое антитело, полученное таким образом, способно связываться с клетками, сверхэкспрессирующими рецептор ErbB2, и нормальными T-клетками человека, так же как запускать литическую активность человеческих цитотоксических лимфоцитов против мишеней рака молочной железы человека.
Описаны также различные способы для получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получали с применением лейциновых молний. Kostelny et al., J. Immunol. 148(5):1547-1553 (1992). Пептиды с лейциновой молнией из белков Fos и Jun слиянием генов присоединяли к Fab'-участкам двух различных антител. Гомодимеры антител восстанавливали в шарнирной области для получения мономеров и затем вновь окисляли для формирования гетеродимеров антител. Данный способ можно применять также для получения гомодимеров антител. Способ «диател», описанный Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993), представляет собой альтернативный механизм получения фрагментов биспецифических антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) линкером, слишком коротким, чтобы позволить спаривание двух доменов одной и той же цепи. Соответственно VH- и VL-домены одного фрагмента вынуждены спариваться с комплементарными VH- и VL-доменами другого фрагмента, таким образом формируя два антигенсвязывающих участка. Опубликована также другая стратегия получения фрагментов биспецифического антитела посредством применения одноцепочечных димеров Fv (sFv). Смотри Gruber et al., J. Immunol. 152:5368 (1994). Рассматривают антитела с более чем двумя валентностями. Например, можно получить триспецифические антитела. Tutt et al., J. Immunol. 147:60 (1991).
Примерные биспецифические антитела могут связывать два различных эпитопа на данном полипептиде PRO87299 здесь. Альтернативно плечо против полипептида PRO87299 можно объединять с плечом, которое связывается с запускающей молекулой на лейкоците, такой как молекула T-клеточного рецептора (например, CD2, CD3, CD28, или B7), или Fc-рецепторов для IgG (FcγR), таких как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16), чтобы фокусировать механизмы клеточной защиты на клетке, экспрессирующей конкретный полипептид PRO87299. Биспецифические антитела можно использовать также для локализации цитотоксических средств на клетках, экспрессирующих конкретный полипептид PRO87299. Эти антитела обладают плечом, связывающим PRO87299, и плечом, связывающим цитотоксическое средство или радиоизотопное хелатирующее средство, такое как EOTUBE, DPTA, DOTA, или TETA. Другое интересующее биспецифическое антитело связывает полипептид PRO87299 и дополнительно связывает тканевой фактор (TF).
5. Гетероконъюгированные антитела
Настоящее изобретение относится также к гетероконъюгированным антителам. Гетероконъюгированные антитела составлены из двух ковалентно соединенных антител. Такие антитела предназначены, например, для нацеливания клеток иммунной системы на нежелательные клетки [патент США No 4676980] и для лечения инфекции HIV [WO 91/00360; WO 92/200373; EP 03089]. Предполагают, что антитела можно получить in vitro с применением известных способов синтетической химии белков, включая способы, включающие в себя средства для перекрестного сшивания. Например, можно конструировать иммунотоксины с применением реакции дисульфидного обмена или формированием тиоэфирной связи. Примеры подходящих для данной цели реагентов включают в себя иминотиолат, метил-4-меркаптобутиримидат и реагенты, описанные, например, в патенте США No 4676980.
6. Модификация эффекторной функции
Может являться желательным модифицировать антитело по изобретению по отношению к эффекторной функции, чтобы, например, улучшить эффективность антитела для лечения рака. Например, в Fc-область можно ввести остаток (остатки) цистеина, таким образом позволяя формирование межцепьевой дисульфидной связи в данной области. Полученное таким образом гомодимерное антитело может обладать улучшенной способностью к интернализации и/или увеличенным комплемент-зависимым цитолизом и антитело-зависимой клеточной цитотоксичностью (ADCC). См. Caron et al., J. Exp Med., 176: 1191-1195 (1992) и Shopes, J. Immunol., 148: 2918-2922 (1992). Гомодимерные антитела с улучшенной противоопухолевой активностью можно также получить с применением гетеробифункциональных перекрестносшивающих средств, как описано в Wolff et al. Cancer Research, 53: 2560-2565 (1993). Альтернативно можно сконструировать антитело, обладающее двойными Fc-областями и, таким образом, увеличить лизис комплементом и способность к ADCC. Смотри Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989).
7. Иммуноконъюгаты
Изобретение относится также к иммуноконъюгатам, содержащим антитело, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, токсин (например, ферментативно активный токсин из бактерий, грибов, растений или животных или его фрагмент), или радиоактивный изотоп (т.е. радиоконъюгат).
Химиотерапевтические средства, применимые для получения таких иммуноконъюгатов, описаны выше. Ферментативно активные токсины и их фрагменты, которые можно применять, включают в себя цепь A дифтерии, несвязывающиеся активные фрагменты дифтерийного токсина, цепь A экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модецина, альфа-сарцин, белки Aleurites fordii, белки с диантином, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из momordica charantia, курцин, кротин, ингибитор из sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. Множество радиоактивных изотопов доступно для производства радиоконъюгированных антител. Примеры включают в себя 212Bi, 131I, 131In, 90Y и 186Re.
Конъюгаты антитела и цитотоксического средства получают с применением множества бифункциональных связывающих белки средств, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (таких как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидил суберат), альдегиды (такие как глутаральдегид), бис-азидо соединения (такие как бис-(p-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(p-диазоний-бензоил)-этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат), и бис-активные соединения фтора (такие как 1,5-дифторо-2,4-динитробензол). Например, рициновый иммунотоксин можно получить, как описано в Vitetta et al., Science, 238: 1098 (1987). Меченная углеродом-14 1- изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего вещества для конъюгации радиоактивных изотопов с антителом. См. WO94/11026.
В другом варианте осуществления антитело можно конъюгировать с «рецептором» (таким как стрептавидин) для применения в предварительном нацеливании на опухоль, где пациенту вводят конъюгат антитело-рецептор с последующим выведением несвязавшегося конъюгата из циркуляции с применением очищающего средства и затем введением «лиганда» (например, авидина), конъюгированного с цитотоксическим средством (например, радиоактивным изотопом).
8. Иммунолипосомы
Описанные здесь антитела можно также составлять как иммунолипосомы. Липосомы, содержащие антитело, получают способами, известными в данной области, такими, как описаны в Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl Acad. Sci. USA, 77: 4030 (1980); и патентах США No 4485045 и 4544545. Липосомы с увеличенным временем циркуляции описаны в патенте США No. 5013556.
Особенно полезные липосомы можно получить способом обращенно-фазового испарения со смесью липидов, содержащей фосфатидилхолин, холестерин и PEG-дериватизированный фосфатидилэтаноламин (PEG-PE). Липосомы выдавливают через фильтры с определенным размером пор для получения липосом желаемого диаметра. Fab'-фрагменты антитела по настоящему изобретению можно конъюгировать с липосомами, как описано в Martin et al., J. Biol. Chem., 257: 286-288 (1982), посредством дисульфиднообменной реакции. Внутри липосомы, не обязательно, заключено химиотерапевтическое средство (такое как доксорубицин). Смотри Gabizon et al., J. National Cancer Inst., 81(19): 1484 (1989).
M. Фармацевтические композиции
Активные молекулы PRO87299 по изобретению (например, полипептиды PRO87299, анти-PRO87299 антитела, и/или варианты каждого из них), так же как другие молекулы, идентифицируемые анализами скрининга, описанными здесь ранее, можно вводить для лечения связанных с иммунитетом заболеваний в форме фармацевтических композиций.
Терапевтические составы с активной молекулой PRO87299, предпочтительно полипептидом или антителом по изобретению, подготавливают для хранения смешиванием активной молекулы, обладающей желаемой степенью чистоты, с необязательными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. [1980]), в форме in the form of лиофилизированных составов или водных ратворов. Приемлемые носители, наполнители или стабилизаторы являются нетоксичными для реципиентов в применяемых дозах и концентрациях и включают в себя буферы, такие как фосфат, цитрат, и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензил аммоний хлорид; хлорид гексония; хлорид бензалкония, хлорид бензетония; фенольный, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и m-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводороды, включая глюкозу, маннозу, или декстрины; хелатобразующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные сурфактанты, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Соединения, идентифицированные в анализах скрининга, описанных здесь, можно составлять аналогичным способом с использованием общепринятых способов, хорошо известных в данной области. Для доставки молекулы PRO87299 в клетки можно использовать также липофекции или липосомы. При использовании фрагментов антител предпочтительным является наименьший подавляющий фрагмент, специфически связывающийся со связывающим доменом белка-мишени. Например, на основе последовательностей вариабельной области антитела можно сконструировать пептидные молекулы, сохраняющие способность связывать намеченную последовательность белка. Такие пептиды можно синтезировать химически и/или получить способом рекомбинантной ДНК (см., например, Marasco et al., Proc. Natl. Acad. ScL USA 90, 7889-7893 [1993]).
Составы здесь могут содержать также более одного активного компонента, как необходимо для конкретного симптома, подлежащего лечению, предпочтительно компоненты с взаимодополняющим действием, не оказывающие отрицательного влияния друг на друга. Альтернативно или дополнительно композиция может содержать цитотоксическое средство, цитокин, химиотерапевтическое средство или средство, подавляющее рост. Такие молекулы соответственно присутствуют в сочетании в количествах, эффективных для предназначенной цели.
Активные молекулы PRO87299 можно захватывать в микрокапсулы, полученные, например, способами коацервации или межфазной полимеризацией, например гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметакрилатные) микрокапсулы соответственно в коллоидных системах доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие способы описаны в Remington's Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980).
Составы, применяемые для введения in vivo, должны быть стерильными. Этого легко достичь фильтрованием через стерилизующие фильтрационные мембраны.
Можно получить препараты молекул PRO87299 с пролонгированным действием. Подходящие примеры препаратов с пролонгированным действием включают в себя полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело, где матрицы присутствуют в форме профилированных изделий, например пленок или микрокапсул. Примеры матриц с пролонгированным действием включают в себя полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли(виниловый спирт)), полилактиды (патент США No 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты - гликолевой кислоты, такие как LUPRON DEPOT™ (пригодные для инъекции микросферы, состоящие из сополимера молочной кислоты - гликолевой кислоты и ацетата лейпролида), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как этиленвинилацетат и молочная кислота - гликолевая кислота способны высвобождать молекулы более 100 суток, конкретные гидрогели высвобождают белки за более короткие периоды времени. Когда инкапсулированные антитела остаются в организме на долгое время, они могут денатурировать или агрегировать в результате воздействия влажности при 37°C, что приводит к потере биологической активности и возможным изменениям иммуногенности. Можно разрабатывать разумные стратегии стабилизации в зависимости от вовлеченного механизма. Например, если обнаружено, что механизмом агрегации является образование междумолекулярной S-S связи посредством тиол-дисульфидного обмена, можно достичь стабилизации модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролированием содержания влаги, применением соответствующих добавок и разработкой специальных композиций полимерной матрицы.
N. Способы лечения
Предполагают, что полипептиды, антитела и другие активные соединения по настоящему изобретению можно применять для лечения различных связанных с иммунитетом заболеваний и состояний, таких как опосредованные T-клетками заболевания, включая заболевания, характеризующиеся инфильтрацией воспалительных клеток в ткань, стимуляцией пролиферации T-клеток, подавлением пролиферации T-клеток, увеличением или уменьшением проницаемости сосудов или ее замедлением.
Состояния или расстройства, подлежащие лечению полипептидами, антителами и другими соединениями по изобретению, включают в себя в качестве неограничивающих примеров системную красную волчанку, ревматоидный артрит, ювенильный хронический артрит, остеоартрит, спондилоартропатии, системный склероз (склеродермию), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (иммунную панцитопению, пароксизмальную ночную гемоглобинурию), аутоиммунную тромбоцитопению (идиопатическую тромбоцитопеническую пурпуру, иммуноопосредованную тромбоцитопению), тиреоидит (базедову болезнь, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), сахарный диабет, иммуноопосредованное заболевание почек (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервных систем, такие как рассеянный склероз, идиопатическая демиелинизирующая полиневропатия или синдром Гийена - Барре, и хроническая воспалительная демиелинизирующая полиневропатия, гепатобилиарные заболевания, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит и склерозирующий холангит, воспалительное заболевание кишечника (язвенный колит: болезнь Крона), глютензависимую энтеропатию и болезнь Уиппла, аутоиммунные или иммуноопосредованные заболевания кожи, включая буллезные заболевания кожи, полиморфную эритему и контактный дерматит, псориаз, аллергические заболевания, такие как астма, аллергический ринит, атопический дерматит, гиперчувствительность к пище и уртикарию, иммунологические заболевания лекого, такие как эозинофильные пневмонии, идиопатический фиброз легких и гиперчувствительный пневмонит, связанные с трансплантацией заболевания, включая отторжение трансплантата и реакцию «трансплантат против хозяина».
При системной красной волчанке центральным медиатором заболевания является продукция аутореактивных антител к собственным белкам/тканям и последующее возникновение иммуноопосредованного воспаления. Антитела напрямую или опосредованно опосредуют повреждение ткани. Хотя не было показано, что T-лимфоциты непосредственно вовлечены в разрушение тканей, T-лимфоциты необходимы для формирования аутореактивных антител. Таким образом, генез заболевания является зависимым от T-лимфоцитов. Множество органов и систем являются клинически пораженными, включая почки, легкое, скелетно-мышечную систему, кожу и слизистые оболочки, глаза, центральную нервную систему, сердечно-сосудистую систему, желудочно-кишечный тракт, костный мозг и кровь.
Ревматоидный артрит (RA) представляет собой хроническое системное аутоиммунное воспалительное заболевание, в основном затрагивающее синовиальную мембрану множества суставов, что приводит к повреждению суставного хряща. Патогенез зависит от T-лимфоцитов и связан с продукцией ревматоидных факторов, аутоантител, направленных против собственного IgG, с формированием в результате иммунных комплексов, достигающих высокого уровня в синовиальной жидкости и крови. Эти комплексы в суставе могут индуцировать заметную инфильтрацию лимфоцитов и моноцитов в синовиальную мембрану и последующие заметные изменения синовии; суставная щель/жидкость инфильтрована такими же клетками с добавлением многочисленных нейтрофилов. Пораженными тканями в первую очередь являются суставы, часто симметричным образом. Однако возникает также внесуставное заболевание двух основных форм. Одной формой является развитие внесуставных очагов при продолжающем прогрессировать заболевании суставов и типичные очаги фиброза легких, васкулита и кожных язв. Вторая форма внесуставного заболевания представляет собой так называемый синдром Фелти, возникающий поздно в ходе заболевания, иногда после того как суставное заболевание становится неактивным, и включающий присутствие нейтропении, тромбоцитопении и спленомегалии. Это может сопровождаться васкулитами во множестве органов с возникновением инфарктов, кожных язв и гангрены. У пациентов часто развиваются ревматоидные узелки в подкожном слое, лежащем выше поврежденных суставов; на поздней стадии узелки обладают некротическими центрами, окруженными смешанным инфильтратом воспалительных клеток. Другие манифестации, которые могут возникать при RA, включают в себя: перикардит, плеврит, коронарный артериит, интерстициальный пневмонит с фиброзом легких, cухой конъюнктивит и ревматоидные узелки.
Ювенильный хронический артрит представляет собой хроническое идиопатическое воспалительное заболевание, которое часто начинается в возрасте менее 16 лет. Его фенотип обладает некоторым сходством с RA; некоторых пациентов, положительных по ревматоидному фактору, классифицируют как страдающих ювенильным хроническим артритом. Заболевание подразделяют на три главных категории: олигоартикулярный, полиартикулярный и системный. Артрит может являться тяжелым и, как правило, является деструктивным и приводит к анкилозу суставов и задержке роста. Другие проявления могут включать в себя хронический передний увеит и системный амилоидоз.
Спондилоартропатии представляют собой группу расстройств с некоторыми общими клиническими признаками и общей связью с экспрессией продукта гена HLA-B27. Расстройства включают в себя: анкилозирующий спондилит, синдром Рейтера (реактивный артрит), артрит, связанный с воспалительным заболеванием кишечника, спондилит, связанный с псориазом, спондилоартропатию с ранним дебютом и недифференцированную спондилоартропатию. Отличительные признаки включают в себя сакроилеит со спондилитом или без; воспалительный асимметрический артрит; связь с HLA-B27 (серологически определяемым аллелем в локусе HLA-B MHC I класса); воспаление глаз и отсутствие аутоантител, связанных с другим ревматоидным заболеванием. Клеткой, наиболее вовлеченной как ключевая для индукции заболевания, является CD8+ T-лимфоцит, клетка, нацеленная на антиген, презентируемый молекулами MHC I класса. CD8+ T-клетки могут реагировать против аллеля HLA-B27 MHC I класса, как если бы он являлся чужеродным пептидом, экспрессированным молекулой MHC I класса. Выдвинули гипотезу, что эпитоп HLA-B27 может мимикрировать бактериальный или другой микробный антигенный эпитоп, и таким образом индуцировать ответ CD8+T-клеток.
Системный склероз (склеродермия) обладает неизвестной этиологией. Критерием заболевания является уплотнение кожи; вероятно, оно индуцировано активным воспалительным процессом. Склеродермия может являться местной или системной; распространенными являются очаги в сосудах, и повреждение эндотелиальных клеток в микроциркуляторном русле является ранним и важным событием в развитии системного склероза; повреждение сосудов может являться иммуноопосредованным. Иммунологическая основа выражается в присутствии инфильтратов мононуклеарных клеток в кожных очагах и в присутствии у многих пациентов антинуклеарных антител. Регуляция ICAM-1 на поверхности клеток фибробластов часто повышена в очагах на коже, что позволяет предполагать, что взаимодействие T-клетки с этими клетками может играть роль в патогенезе заболевания. Другие затрагиваемые органы включают в себя: желудочно-кишечный тракт: атрофия гладкой мускулатуры и фиброз, приводящие к ненормальной перистальтике/сократительной способности; почки: концентрическая субэндотелиальная пролиферация интимы, поражающая малые интерлобулярные и дуговые артерии с получающимся в результате пониженным ренальным кортикальным кровотоком, приводящая к протеинурии, азотемии и гипертензии; скелетная мышца: атрофия, интерстициальный фиброз; воспаление; легкое: интерстициальный пневмонит и интерстициальный фиброз; и сердце: некроз полосы сокращения, рубцевание/фиброз.
Идиопатические воспалительные миопатии, включая дерматомиозит, полимиозит и другие, представляют собой расстройства из хронического мышечного воспаления неизвестной этиологии, приводящие к мышечной слабости. Повреждение/воспаление мышц часто является симметричным и прогрессирующим. С большинством форм связаны аутоантитела. Эти миозит-специфические аутоантитела направлены против белковых и РНК-компонентов, вовлеченных в синтез белка, и подавляют их функцию.
Синдром Шегрена обусловлен иммуноопосредованным воспалением и последующим функциональным повреждением слезных желез и слюнных желез. Заболевание может являться связанным с воспалительными заболеваниями соединительной ткани или сопровождаться ими. Заболевание связано с продукцией аутоантител против антигенов Ro и La, которые оба представляют собой малые комплексы РНК-белок. Поражения приводят к cухому конъюнктивиту, ксеростомии, с другими манифестациями или связанными поражениями, включая билиарный цирроз, периферическую или сенсорную невропатию и пальпируемую пурпуру.
Системные васкулиты представляют собой заболевания, при которых первичным поражением является воспаление и последующее разрушение кровеносных сосудов, приводящее к ишемии/некрозу/дегенерации тканей, снабжаемых пораженными сосудами, и возможной в некоторых случаях дисфункции окончания двигательного нерва. Васкулиты могут возникать также как вторичное повреждение или остаточное явление других обусловленных иммунным воспалением заболеваний, таких как ревматоидный артрит, системный склероз и т.д., в частности заболеваний, также связанных с образованием иммунных комплексов. Заболевания группы первичного системного васкулита включают в себя: системный некротический васкулит: нодозный полиартериит, аллергический васкулит и гранулематоз, поливаскулит; гранулематоз Вегенера; лимфоидный гранулематоз; и гигантоклеточный артериит. Смешанный васкулит включает в себя: слизисто-кожный лимфоузелковый синдром (MLNS или болезнь Кавасаки), изолированный васкулит ЦНС, болезнь Бехчета, облитерирующий тромбангиит (болезнь Бюргера) и кожный некротизирующий венулит. Считают, что патогенный механизм большинства перечисленных типов васкулита в первую очередь обусловлен депонированием иммуноглобулиновых комплексов в стенке сосуда и последующей индукцией воспалительного ответа или посредством ADCC, или посредством активации комплемента, или и тем, и другим.
Саркоидоз представляет собой состояние неизвестной этиологии, характеризующееся присутствием эпителиоидных гранулем почти в любой ткани организма; наиболее распространенным является вовлечение легкого. Патогенез включает в себя персистенцию активированных макрофагов и лимфоцитов в участках заболевания с последующими хроническими остаточными явлениями, происходящими от высвобождения местно и системно активных продуктов, высвобождаемыми этими типами клеток.
Аутоиммунная гемолитическая анемия, включая аутоиммунную гемолитическую анемию, иммунную панцитопению и пароксизмальную ночную гемоглобинурию, является результатом продукции антител, реагирующих с антигенами, экспрессированными на поверхности красных кровяных клеток (и в некоторых случаях, других клеток крови, включая также тромбоциты), и является отражением удаления этих покрытых антителами клеток посредством опосредованного комплементом лизиса и/или ADCC/Fc-рецептор-опосредованных механизмов.
При аутоиммунной тромбоцитопении, включая тромбоцитопеническую пурпуру и иммуноопосредованную тромбоцитопению, разрушение/удаление тромбоцитов происходит в результате присоединения к тромбоцитам или антитела, или комплемента и последующего удаления посредством механизмов лизиса комплементом, ADCC или опосредованного FC-рецептором механизма.
Тиреоидит, включая базедову болезнь, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит и атрофический тиреоидит, является результатом аутоиммунного ответа против антигенов щитовидной железы с продукцией антител, реагирующих с белками, присутствующими в щитовидной железе, и часто специфических для нее. Существуют экспериментальные модели, включая спонтанные модели: крыс (крыс BUF и BB) и кур (линия кур с ожирением); индуцируемые модели: иммунизация животных или тиреоглобулином, или антигеном микросомальной фракции щитовидной железы (тироидная пероксидаза).
Сахарный диабет I типа или инсулинзависимый диабет представляет собой аутоиммунное разрушение панкреатических островков β-клеток; это разрушение опосредовано аутоантителами и аутореактивными T-клетками. Антитела к инсулину или рецептору инсулина могут также вызывать фенотип нечувствительности к инсулину.
Иммуноопосредованные ренальные заболевания, включая гломерулонефрит и тубулоинтерстициальный нефрит, являются результатом опосредованного антителами или T-лимфоцитами повреждения ренальной ткани или непосредственно в результате продукции аутореактивных антител или T-клеток против ренальных антигенов, или опосредованно в результате депонирования в почке антител и/или иммунных комплексов, реактивных против других не относящихся к почке антигенов. Таким образом, другие иммуноопосредованные заболевания, приводящие к формированию иммунокомплексов, могут также индуцировать иммуноопосредованное ренальное заболевание как опосредованное остаточное явление. Как прямые, так и опосредованные иммунные механизмы приводят к воспалительному ответу, приводящему к развитию/индуцирующему развитие повреждений в ренальной ткани, что приводит к нарушению функции органа и в некоторых случаях прогрессированию до почечной недостаточности. Как гуморальный, так и клеточный иммунные механизмы могут являться вовлеченными в патогенез повреждений.
Считают, что демиелинизирующие заболевания центральной и периферической нервных систем, включая рассеянный склероз; идиопатическую демиелинизирующую полиневропатию или синдром Гийена-Барре; и хроническую воспалительную демиелинизирующую полиневропатию, имеют аутоиммунную природу и приводят к демиелинизации нерва в результате разрушения олигодендроцитов или непосредственно миелина. При РС существуют основания предполагать, что индукция и прогрессирование заболевания зависит от T-лимфоцитов. Рассеянный склероз представляет собой демиелинизирующее заболевание, являющееся T-лимфоцит-зависимым и имеющее или течение с обострением-ремиссией, или хронически прогрессирующее течение. Этиология является неизвестной; однако вирусные инфекции, генетическая предрасположенность, внешняя среда и аутоиммунность - все вносят свой вклад. Бляшки содержат инфильтрат, преимущественно опосредованный T-лимфоцитами, микроглиальными клетками и инфильтрированными макрофагами; CD4+T-лимфоциты являются преимущественным типом клеток в бляшках. Механизм клеточной смерти олигодендроцитов и последующей демиелинизации является неизвестным, однако, вероятно, его запускают T-лимфоциты.
Воспалительное и фиброзное заболевание легких, включая эозинофильную пневмонию; идиопатический фиброз легких и гиперчувствительный пневмонит, может включать в себя дисрегулированный иммунно-воспалительный ответ. Подавление этого ответа может являться терапевтически выгодным.
Аутоиммунные или иммуноопосредованные заболевания кожи, включая буллезные заболевания кожи, полиморфную эритему и контактный дерматит, опосредованы аутоантителами, образование которых зависит от T-лимфоцитов.
Псориаз является опосредованным T-лимфоцитами воспалительным заболеванием. Очаги содержат инфильтраты T-лимфоцитов, макрофагов и антиген-процессирующих клеток и некоторые нейтрофилы.
Аллергические заболевания, включая астму, аллергический ринит, атопический дерматит, гиперчувствительность к пище, и уртикарию, являются зависимыми от T-лимфоцитов. Эти заболевания преимущественно опосредованы индуцированным T-лимфоцитами воспалением, IgE-опосредованным воспалением или сочетанием того и другого.
Заболевания, связанные с трансплантацией, включая отторжение трансплантата и реакцию трансплантат против хозяина (GVHD), являются зависимыми от T-лимфоцитов; подавление функции T-лимфоцитов является улучшающим.
Другие заболевания, при которых вмешательство в иммунный и/или воспалительный ответ являются полезными, включают в себя в качестве неограничивающих примеров вирусную инфекцию (включая в качестве неограничивающих примеров СПИД, гепатит A, B, C, D, E и герпес) бактериальную инфекцию, грибковые инфекции, и протозойные инфекции и паразитарные инфекции (молекулы (или производные/агонисты), стимулирующие MLR, можно использовать терапевтически для усиления иммунного ответа на инфекционные средства), иммунодефицитные заболевания (молекулы/производные/агонисты), стимулирующие MLR, можно использовать терапевтически для усиления иммунного ответа при состояниях врожденного, приобретенного, индуцируемого инфекцией (как при инфекции HIV), или ятрогенного (т.е. как от химиотерапии) иммунодефицита и неоплазии.
Показано, что у некоторых пациентов с раком человека развивается ответ антител и/или T-лимфоцитов на антигены неопластических клеток. На моделях неоплазии на животных показано также, что усиление иммунного ответа может приводить к отторжению или регрессии данной конкретной неоплазмы. Молекулы, усиливающие ответ T-лимфоцитов в MLR, имеют применение in vivo для усиления иммунного ответа против неоплазии. Молекулы, усиливающие пролиферативный ответ T-лимфоцитов в MLR (или низкомолекулярные агонисты или антитела, влияющие на тот же рецептор как агонист), можно использовать терапевтически для лечения рака. Молекулы, подавляющие ответ лимфоцитов в MLR, функционируют также in vivo в ходе неоплазии для подавления иммунного ответа на неоплазму; или такие молекулы могут являться экспрессированными собственно неопластическими клетками, или их экспрессия в других клетках может являться индуцированной неоплазмой. Антагонизм таких подавляющих молекул (или антителом, низкомолекулярными антагонистами или другими способами) усиливает иммуноопосредованное отторжение опухоли.
Кроме того, подавление молекул с провоспалительными свойствами может обладать терапевтическим преимуществом при реперфузионном повреждении, инсульте, инфаркте миокарда, атеросклерозе, остром повреждении легких, геморрагическом шоке, ожоге, сепсисе/септическом шоке, остром тубулярном некрозе, эндометриозе, дегенеративное заболевание суставов и панкреатите. Соединения по настоящему изобретению, например полипептиды или антитела, вводят млекопитающему, предпочтительно человеку, согласно известным способам, таким как внутривенное введение в виде болюса или длительным вливанием в течение периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, интраартикулярным, внутрисуставным, интратекальным, пероральным, способами или способами ингаляции (интраназальным, внутрилегочным). Предпочтительным является внутривенное или ингаляционное введение полипептидов и антител.
В иммуноадъювантной терапии другие терапевтические режимы, подобные введению противораковых средств, можно сочетать с введением белков, антител или соединений по настоящему изобретению. Например, пациенту, подлежащему лечению иммуноадъювантом по изобретению, можно вводить также противораковое средство (химиотерапевтическое средство) или радиотерапию. Для таких химиотерапевтических средств можно применять препараты и режимы дозирования согласно инструкциям производителя или по эмпирическому определению опытного специалиста-практика в данной области. Препараты и режимы дозирования для такой химиотерапии описаны также в Chemotherapy Seiyice Ed., M.C. Perry, Williams & Wilkins, Baltimore, MD (1992). Химиотерапевтическое средство может вводить до или после введения иммуноадъюванта или можно вводить одновременно с ним. Кроме того, можно вводить антиэстрогенные соединения, такие как тамоксифен или антипрогестерон, такой как онапристон (см., EP 616812), в дозах, известных для таких молекул.
Может являться желательным также вводить антитела против других иммунных заболеваний, связанных с ассоциированными с опухолью антигенами, такими как антитела, связывающиеся с CD20, CD11a, CD18, ErbB2, EGFR, ErbB3, ErbB4, или фактор роста эндотелия сосудов (VEGF). Альтернативно или дополнительно, два или несколько антител, связывающих один и тот же антиген или два или несколько различных антигенов, описанных здесь, можно совместно вводить пациенту. Иногда может являться благоприятным также вводить пациенту один или несколько цитокинов. В одном из вариантов осуществления полипептиды PRO87299 вводят совместно с подавляющим рост средством. Например, подавляющее рост средство можно вводить первым, затем полипептид PRO87299. Однако предусматривают также его одновременное введение или введение первым. Подходящими дозами для подавляющего рост средства являются дозы, применяемые в настоящее время, и их можно понизить из-за совместного действия (синергизма) подавляющего рост средства и полипептида PRO87299.
Для лечения или уменьшения тяжести связанного с иммунитетом заболевания подходящая дозировка соединения по изобретению будет зависеть от типа заболевания, подлежащего лечению, как определено выше, тяжести и течения заболевания, введения средства для профилактических или терапевтических целей, предшествующей терапии, истории болезни пациента и чувствительности к соединению и усмотрения лечащего врача. Соединение является применимым для введения пациенту за один раз или в течение серии обработок.
Например, в зависимости от типа и тяжести заболевания приблизительно 1 мкг/кг - 15 мг/кг (например, 0,1-20 мг/кг) полипептида или антитела является начальной возможной дозой для введения пациенту, например, посредством одного или нескольких отдельных введений или посредством длительного вливания. Обычная ежесуточная доза может лежать в диапазоне от приблизительно 1 мкг/кг до 100 мг/кг или более в зависимости от факторов, упомянутых выше. При повторных введениях в течение нескольких дней или дольше в зависимости от состояния, лечение продолжают, пока не происходит желаемого подавления симптомов заболевания. Однако можно применять другие режимы дозирования. Прогресс этой терапии легко мониторировать общепринятыми способами и анализами.
O. Изделия
В другом варианте осуществления изобретение относится к изделию, содержащему вещества (например, содержащему молекулу PRO87299), пригодные для диагностики или лечения описанных выше расстройств. Изделие содержит контейнер и инструкцию. Подходящие контейнеры включают в себя, например, флаконы, ампулы, шприцы и пробирки. Контейнеры можно изготавливать из множества материалов, таких как стекло или пластик. Контейнер содержит композицию, эффективную для диагностики или лечения состояния, и может иметь стерильное отверстие для доступа (например, контейнер может представлять собой пакет с раствором для внутривенного введения или флакон с пробкой, прокалываемой стерильной иглой для подкожных инъекций). Активное вещество в композиции, как правило, представляет собой полипептид или антитело по изобретению. На ярлыке или в листовке-вкладыше указано, что композицию применяют для лечения выбранного состояния, такого как рак. В инструкции или ярлыке на контейнере или приложенным к контейнеру, указано, что композицию применяют для диагностики или лечения выбранного состояния. Изделие может, кроме того, содержать второй контейнер, содержащий фармацевтически пригодный буфер, такой как фосфатно-солевой буфер, раствор Рингера и раствор глюкозы. Он может дополнительно содержать другие вещества, желательные с коммерческой или пользовательской точки зрения, включая другие буферы, растворители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями по применению.
P. Диагностика и прогноз связанного с иммунитетом заболевания
Белки клеточной поверхности, такие как белки, сверхэкспрессированные при конкретных связанных с иммунитетом заболеваниях, являются отличными мишенями для лекарственных средств-кандидатов или для лечения заболевания. Те же самые белки вместе с секретируемыми белками, кодируемыми генами, амплифицированными при связанных с иммунитетом болезненных состояниях, находят дополнительное применение для диагностики и прогноза этих заболеваний. Например, антитела, направленные против белковых продуктов генов, амплифицированных при рассеянном склерозе, ревматоидном артрите или другом связанным с иммунитетом заболевании, можно применять как диагностические или прогностические.
Например, антитела, включая фрагменты антител, можно использовать для количественной или качественной детекции экспрессии белков, кодируемых амплифицированными или сверхэкспрессированными генами («продукты маркерного гена»). Антитело предпочтительно снабжают поддающейся детекции, например, флуоресцентной меткой и связывание можно мониторировать световой микроскопией, проточной цитометрией, флуорометрией, или другими способами, известными в данной области. Данные способы особенно подходят, если сверхэкспрессированный ген кодирует белок клеточной поверхности. Такие анализы связывания проводят в основном, как описано выше.
Детекцию in situ антитела, связанного с продуктами маркерного гена, можно осуществлять, например, посредством иммунофлуоресцентной или иммуноэлектронной микроскопии. Для этой цели у пациента отбирают гистологический препарат и на него наносят меченое антитело, предпочтительно покрытием биологического образца антителом. Этот способ позволяет также определять распределение продукта маркерного гена в исследуемой ткани. Специалистам в данной области будет очевидно, что широкое множество гистологических способов легко доступны для детекции in situ.
Следующие примеры представлены только для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения каким-либо образом.
Полное содержание всех патентов и ссылок на литературу, процитированных в настоящей заявке, приведено здесь в качестве ссылки.
ПРИМЕРЫ
Коммерчески доступные реагенты, указанные в примерах, использовали согласно инструкциям производителя, если не указано иначе. Источником клеток, указанных в следующих примерах и на всем протяжении описания под инвентарными номерами ATCC, является Американская коллекция типовых культур, Manassas, VA.
ПРИМЕР 1: Клонирование PRO87299
Провели поиск в базе данных последовательности нуклеотидов транскрибируемых участков ДНК (EST) (Merck/Washington University) и идентифицировали EST, содержащую интересующие домены, конкретно, домен(ы) иммуноглобулина (Ig) и иммунорецепторный тирозин-связывающий ингибиторный мотив(ы) (ITIM). Поиск проводили с использованием компьютерной программы BLAST или BLAST2 [Altschul et al., Methods in Enzvmology. 266:460-480 (1996)] с использованием в качестве сравнения интересующих доменов в 6 рамках трансляции последовательности. Сравнения, приводящие к баллу BLAST 70 (или в некоторых случаях 90) или выше, не кодирующие известные белки, кластеризовали и, если необходимо, объединяли в консенсусные последовательности ДНК программой «phrap» (Phil Green, University of Washington, Seattle, Washington).
На основе последовательности, как описано выше, синтезировали олигонуклеотиды: 1) для идентификации посредством PCR библиотеки кДНК, содержащие интересующую последовательность, и 2) для использования в качестве зондов для выделения клона полноразмерной кодирующей последовательности для PRO87299. Прямой и обратный праймеры для PCR, как правило, лежали в диапазоне от 20 до 30 нуклеотидов, и часто их конструировали для получения продукта PCR длиной приблизительно 100-1000 п.н. Последовательности зондов, как правило, обладают длиной 40-55 п.н. В некоторых случаях синтезировали дополнительные олигонуклеотиды, когда консенсусная последовательность превышала приблизительно l-1,5 т.п.н. Чтобы скринировать несколько библиотек по полноразмерному клону, ДНК из библиотек скринировали амплификацией PCR, как в Ausubel et al., Current Protocols in Molecular Biology, выше, с парой праймеров для PCR. Затем положительную библиотеку использовали для выделения клонов, кодирующих интересующий ген с использованием олигонуклеотиданого зонда и одной из пар праймеров.
Применяли следующие олигонуклеотидные зонды:
Прямой праймер:
hBTig.EcoRI.F2 5' TTGAATTCATGAAGACATTGCCTGCCATGC 3'
(SEQ ID NO 11)
Обратный праймер:
hBTig.BamHI.R2 5' TTGGATCCTTAACTCCTCACACATATGGATGCATATTC 3'
(SEQ ID NO 12)
Для клонирования использовали библиотеку кДНК из крови человека. Библиотеку кДНК, используемую для выделения клонов кДНК, конструировали общепринятыми способами с использованием коммерчески доступных реагентов, таких как реагенты из Invitrogen, San Diego, CA. кДНК праймировали олиго-dT, содержащим участок NotI, соединяли с затуплением с SalI-адапторами, наполовину обработанными киназой, разрезали NotI, соответствующим образом сортировали по размеру гель-электрофорезом и клонировали в определенной ориентации в подходящем клонирующем векторе (таком как pRKB или pRKD; pRK5B является предшественником pRK5D, не содержащим участок SfiI; смотри, Holmes et al., Science, 253:1278-1280 (1991)) по уникальным участкам XhoI и NotI.
Полная нуклеотидная последовательность клона, обозначенного здесь как ДНК332467, показана на фиг.1 (SEQ ID NO: 1). Клон ДНК332467 содержит одну открытую рамку считывания с очевидным участком инициации трансляции в положениях нуклеотидов 24-26 и стоп-сигналом в положениях нуклеотидов positions 891-893 (фиг.1, SEQ ID NO: 1). Предполагаемый полипептидный предшественник обладает длиной 289 аминокислот, обладает рассчитанной молекулярной массой приблизительно 32781 дальтон и рассчитанной pI приблизительно 6,27. Анализ полноразмерной последовательности PRO87299, показанной на фиг.2 (SEQ ID NO: 2) доказывает присутствие множества важных полипептидных доменов, как показано на фиг. 2, где положения, приведенные для этих полипептидных доменов, являются приблизительно такими, как описаны.
Анализом существующей базы данных белков с использованием анализа выравнивания последовательности ALIGN-2 для полноразмерной последовательности, показанной на фиг.2 (SEQ ID NO: 2), показали идентичность последовательности между аминокислотной последовательностью PRO87299 и неизвестной белковой последовательностью.
ПРИМЕР 2: Клонирование вариантов PRO87299
Скринировали варианты последовательности PRO87299 в РНК B-клеток от 16 различных доноров. С этой РНК проводили RT-PCR для получения полноразмерного PRO87299. Продукты PCR клонировали в векторы, позволяющие высокопроизводительное секвенирование, и анализировали секвенированием с двойным проходом. Для нескольких вариантов PRO87299 показали минимальное изменение (фиг.11 A-F). Однако обнаружили усеченный вариант, обладающий экзоном 3 с делетированным трансмембранным доменом (фиг.7, SEQ ID NO: 7 ). В усеченном варианте делетированы нуклеиновые кислоты 403-547 из нативного белка, что приводит к варианту полипептида PRO87299 (фиг.8, SEQ ID NO: 8) длиной только 241 аминокислоты, в то время как нативный PRO87299 обладает длиной 289 аминокислот. Отсутствие трансмембранного домена может означать, что этот вариант PRO87299 представляет собой секретируемую форму.
Обнаружили дополнительный вариант PRO87299, содержащий вставку 18 пар нуклеотидных оснований на 5'-конце экзона 3 (фиг.9, SEQ ID NO: 9). Эти вставленные 18 пар оснований кодируют дополнительные 6 аминокислот (AFTNIP) и вставлены в кодирующую PRO87299 нуклеиновую кислоту в рамке считывания, приводя к варианту полипептида PRO87299 (фиг. 10, SEQ ID NO: 10), имеющему длину 295 аминокислот. И короткая форма, и форма AFTNIP показаны вместе с другими вариантами на фиг.12A-B. Домен IgG, обнаруженный в аминокислотах 51-117 всех полипептидов PRO87299, может являться важным для функционирования полипептида PRO87299.
ПРИМЕР 3: Анализ стимулированных T-клеток на микрочипах
Микрочипы нуклеиновых кислот, часто содержащие тысячи последовательностей генов, являются применимыми для идентификации дифференциально экспрессирующихся генов в пораженных заболеваниям тканях по сравнению с их нормальными аналогами. С использованием микрочипов нуклеиновых кислот тестируемые и контрольные образцы мРНК из тестируемых и контрольных образцов ткани подвергали обратной транскрипции и метили для получения зондов кДНК. Затем зонды кДНК гибридизовали с массивом нуклеиновых кислот, иммобилизованных на твердой подложке. Массив конструировали так, что последовательность и положение каждого члена массива является известной. Например, выборку генов, известных как экспрессирующиеся в конкретных стадиях заболевания, можно упорядочивать на твердой подложке. Гибридизациия меченого зонда с конкретным членом массива указывает на экспрессию данного гена в образце, из которого получен зонд. Если сигнал гибридизации с зондом из тестируемого образца (в данном случае, активированных CD4+ T-клеток) сильнее, чем сигнал гибридизации зонда из контрольного образца (в данном случае, не стимулированных CD4+ T-клетках), идентифицированы ген или гены, сверхэкспрессированные в тестируемой ткани. Следствием данного результата является то, что сверхэкспрессированный белок в тестируемом образце не только является применимым в качестве диагностического маркера для присутствия состояния заболевания, но также является терапевтической мишенью для лечения состояния заболевания.
Способы гибридизации нуклеиновых кислот и спсособ микрочипов хорошо известны в данной области. В одном примере конкретное получение нуклеиновых кислот для гибридизации и зондов, стекол, и условий гибридизации, все, подробно описаны в заявке на патент PCT серийный No PCT/USOl/10482, поданной 30 марта 2001 и приведенной здесь в качестве ссылки. В этом эксперименте с использованием способа RossetteSep™ от (Stem Cell Technologies, Vancouver BC) очищали CD4+ T-клетки от одного донора, содержащие анти-CD8, анти-CD16, анти-CD19, анти-CD36 и анти-CD56 антитела, применяя для получения выделенных CD4 + T-клеток. Выделенные CD4+ T-клетки активировали анти-CD3 антителом (использованным в концентрации, не стимулирующей пролиферацию) вместе или с ICAM-I, или с анти-CD28 антителом. После 24 или 72 часов клетки собирали, выделяли РНК и проводили анализ на микрочипах Affimax™ (Affymetrix Inc. Santa Clara, CA). Нестимулированные (покоящиеся) клетки собирали немедленно после очистки и подвергали такому же анализу. Сравнивали гены, экспрессия которых имела повышенную регуляцию в любой из двух временных точек в активированных клетках по сравнению с покоящимися.
Результатом данных экспериментов является то, что полипептиды PRO87299 по настоящему изобретению значительно сверхэкспрессированы в выделенных CD4 + T-клетках, активированных анти-CD3/ICAM-1 и анти-CD3/анти-CD28, по сравнению с выделенными покоящимися CD4+ T-клетками. Как описано выше, этими данными показывают, что полипептиды PRO87299 по настоящему изобретению не только являются применимым в качестве диагностических маркеров присутствия одного или нескольких иммунных расстройств, но также служат терапевтическими мишенями для лечения этих иммунных расстройств.
ПРИМЕР 4: PRO87299 при лимфоме
Лимфома является 6-й из наиболее распространенных злокачественный новообразований в Соединенных Штатах. В 1990 г. в Соединенных Штатах было 43000 предположительных новых случаев лимфомы. Неходжскинская лимфома составляет большинство случаев, а лимфома Ходжкина занимает отдаленное второе место. Заболеваемость неходжскинской лимфомой постепенно увеличивается с возрастом. Однако для болезни Ходжкина существует высокая заболеваемость у пациентов в возрасте 20-30, плато между 30-55 и другой подъем после возраста 55. Мужчины более подвержены риску как болезни Ходжкина, так и неходжскинской лимфомы, чем женщины. Основным клиническим проявлением злокачественной лимфомы является разбухание лимфатического узла и симптомов, включая лихорадку, дисфорию и потерю массы. Обычными первичными очагами лимфомы являются надключичные, подмышечные, медиастинальные, периаортальные, цервикальные и паховые лимфоузлы. Лимфома обладает также способностью метастазировать в другие органы.
Болезнь Ходжкина впервые описана Томасом Ходжкином в 1832. Болезнь Ходжкина представляет собой неограниченную пролиферацию лимфоцитов, которые становятся больше, с обильной бледной цитоплазмой и двумя или более овальными мелкодольчатыми ядрами, содержащими большие ядрышки. Клетки такого вида известны как клетки Рида-Штернберга. Клетки Рида-Штернберга являются важными для диагностики болезни Ходжкина, однако их присутствие само по себе не является достаточным для диагноза. Болезнь Ходжкина отличается от неходжскинской лимфомы типом клеток, гистологией лимфоузлов и симптоматологией, такой как лихорадка. Болезнь Ходжкина обычно проявляется как увеличение отдельной группы периферических лимфоузлов и может затрагивать соседние лимфоузлы, однако нечасто является внеузловой. Причина болезни Ходжкина является неизвестной, однако предшествующая инфекция вирусом Эпштейна-Барра и транслокации bcl-2 связаны с развитием болезни Ходжкина.
Неходжскинские лимфомы представляют собой неоплазии иммунной системы, возникающие в лимфоузлах, однако они отличаются от болезни Ходжкина такими факторами, как тип клеток и симптоматология, проявляющаяся у пациента. Большинство неходжскинских лимфом обладают B-клеточным фенотипом и являются положительными по маркерам CD19 и CD20. Меньшее число являются T-клеточными лимфомами и являются положительными по маркерам CD2 и CD3.
Патентованную базу данных, содержащую информацию по экспрессии генов (GeneExpress®, Gene Logic Inc., Gaithersburg, MD), анализировали, чтобы удостовериться, присутствует ли повышенная регуляция полипептида PRO87299 (и кодирующей его нуклеиновых кислот) в лимфоме по сравнению с нормальными лимфатическими тканями. Конкретно анализ базы данных GeneExpress® проводили с использованием или программного обеспечения, доступного в Gene Logic Inc., Gaithersburg, MD для использования с базой данных GeneExpress® или патентованного программного обеспечения, написанного и разработаанного в Genentech, Inc. для использования с базой данных GeneExpress®. Рейтинг положительных ответов в анализе основан на нескольких критериях, включая, например, тканевую специфичность, опухолевую специфичность и уровень экспрессии в нормальных эссенциальных и/или нормально пролиферирующих тканях. В результате показали высокую экспрессию PRO87299 в лимфоме по сравнению с другими опухолями и нормальными тканями.
ПРИМЕР 5: PRO87299 при воспалительном заболевании кишечника
В этом эксперименте анализ на микрочипах использовали для поиска генов, сверхэкспрессированных при IBD по сравнению с нормальной тканью кишечника. Получали образцы биопсии от пациентов с IBD. Для каждого пациента с IBD получали образцы из ткани, пораженной заболеванием (или UC, или болезнью Крона) и из здорового кишечника, так чтобы можно было лучше сравнивать картины экспрессии. Все образцы хранили при -70°C до подготовки к выделению РНК. Образцы биопсии гомогенизировали в 600 мкл буфера RLT (+ BME) и выделяли РНК с использованием колонок Rneasy Mini Qiagen™ (Qiagen) с обработкой ДНКазой на колонке согласно рекомендациям производителя. После выделения РНК количество РНК оценивали с использованием RiboGreen™ (Molecular Probes), следуя рекомендациям производителя, и проверяли ее сохранность в агарозных гелях. Соответствующие количества РНК метили для анализа на микрочипах и образцы анализировали на патентованном микрочипе Genentech и микрочипах Affymetrics™. Сравнивали гены, регуляция экспрессии которых повышена в ткани, пораженной IBD, по сравнению с нормальным кишечником, подбирая пары образцов биопсии из нормального кишечника и пораженной IBD ткани от одного и того же пациента. Результаты этого эксперимента показали, что идентифицирована значительная сверхэкспрессия PRO87299 в образцах при болезни Крона по сравнению с нормальной тканью кишечника.
ПРИМЕР 6: Экспрессия PRO87299 в NK-клетках
Клетки натуральные киллеры (NK) являются важными эффекторными клетками врожденной иммунной системы. Они специализируются на эффекте цитолиза против клеток хозяина, которые или инфицированы вирусом, паразитами, или стали злокачественными. Фенотипически NK-клетки представляют собой большие гранулярные лимфоциты, составляющие приблизтельно 2 % циркулирующей популяции лимфоцитов. Их обычно идентифицируют по экспрессии на клеточной поверхности CD56 и CD16. Они созревают в костном мозге из клеток-предшественников CD34+, которые у них являются общими с T-клетками. Зрелые NK-клетки разделяют с T-клетками экспрессию CD8, цитолитический механизм и некоторые KIR, но остаются отличающимися от T-клеток отсутствием CD3 и T-клеточных рецепторов. Подобно цитотоксическим T-клеткам они содержат гранулы, заполненные порообразующим белком, цитотоксинами, серин эстеразы и протеогликанами, которые опосредуют лизис клеток-мишеней. И цитотоксические T-клетки и NK-клетки уничтожают при контакте посредством связывания со своими мишенями и доставкой их летального выброса химикатов, что приводит к отверстиям в мембране клеток-мишеней. В отличие от цитотоксических T-клеток NK-клетки не нуждаются в узнавании специфического антигена перед инициированием лизиса. Скорее активация NK-клетки может являться опосредованным факторами роста и цитокинами (в частности, показано, что IL-2, IL-12 и IL-15 опосредуют пролиферативные и цитотоксические активности) или неустойчивым равновесием между двумя классами рецепторов NK-клеток, один из которых активирует клетки, а другой ингибирует. Ig-подобные рецепторы килеров (KIR) представляют собой рецепторы NK-клеток, переносящие ингибирyющий сигнал при встрече с молекулами MHC I класса на клеточной поверхности. Это является важным для уничтожения как злокачественных клеток, так и инфицированных вирусом клеток. Поскольку вирусы часто подавляют экспрессию MHC I класса в инфицированных ими клетками, инфицированные вирусом клетки становятся чувствительными к уничтожению NK-клетками. Подобным образом в злокачественных клетках уменьшена или отсутствует экспрессия MHC I класса, и они также становятся чувствительными к уничтожению NK-клетками. Натуральные цитотоксические рецепторы (NCR) составляют семейство активирующих рецепторов на NK-клетках. В некоторых эффекторных системах поверхностная плотность NCR коррелирует цитолитической активностью NK-клеток, в то время как в других системах уничтожение требует взаимодействия между NCR, другим активирующим рецептором NKG2D и его адаптером полипептидом DAP10. Кроме того, на силу сигнала может влиять привлечение корецепторов, таких как 2B4 и NTB-A. Лиганды для NCR и NKG2D, гемагглютинин и MICA, MICB соотвественно, не экспрессируются большинством нормальных клеток, однако индуцированы в большинстве линий опухолевых клеток. Экспрессия лигандов опухолевыми клетками запускает сильный иммунный ответ, приводящий к отторжению опухолевых клеток. Показано, что активация NK-клеток IL-15 или IL-12 индуцирует как цитотоксические, так и пролиферативные эффекты. Показано что молекула контактной адгезии 2 (JAM2) связывается с NK-клетками, и выдвинули гипотезу о ее роли в экстравазации лимфоцитов к участкам воспаления.
Таким образом, эксперимент с микрочипами ДНК, сравнивающий дифференциальную экспрессию генов с тремя этими способами активации по сравнению с покоящимися NK-клетками, позволяет выявлять новые гены или новые связи генов с активностью NK-клетки. Можно разрабатывать антитела, пептиды или малые молекулы, нацеленные на конкретные гены, выявленные этими микрочипами, для лечения иммуноопосредованных воспалительных заболеваний и злокачественных новообразований. NK-клетки периферической крови выделяли из лейкаферезной массы посредством негативного отбора с использованием набора для выделения NK-клеток с магнитной системой сортировки клеток MACS™ (Miltenyi Biotec). Очистку клеток подтверждали окрашиванием PE анти-CD56 для анализа FACS. Чистота препаратов клеток лежала в диапазоне от 89% до 96%. Культура клеток: начинали культуры in-vitro в 6-луночных планшетам по 5 мл культуры/лунку. Среда: RPMI 1640, 10% инактивированной нагреванием FBS, 100 ед./мл пенициллина, 100 мг/мл стрептомицина, 2 мМ L-глутамин, и 5,5×10-5 бета-меркаптоэтанол. Экспериментальные обработки: время 0 час, необработанные CD56(+) клетки. Время 16 час. Необработанные, стимулированные IL2 (10 нМ), IL15(10 нМ), JAM-IT(10 нМ). Активацию NK-клеток контролировали посредством FACS по экспрессии на клеточной поверхности CD56 и CD69. В этой серии экспериментов определили, что CD56+ NK-клетки экспрессируют PRO87299 по сравнению с нормальными покоящимися NK-клетками.
ПРИМЕР 7: Применение PRO87299 в качестве зонда для гибридизации
В следующем способе описано применение нуклеотидной последовательности, кодирующей PRO87299, в качестве зонда для гибридизации.
ДНК, содержащую кодирующую последовательность полноразмерного или зрелого PRO87299 (такую как ДНК, кодирующую встречающиеся в природе варианты PRO87299), как описано здесь, применяли в качестве зонда для скрининга гомологичных ДНК в библиотеках кДНК из тканей человека или геномных библиотеках из тканей человека.
Гибридизацию и отмывку фильтров, содержащих любую из библиотек ДНК, проводили в следующих высокострогих условиях. Гибридизацию радиоактивно меченного полученного из PRO87299 зонда с фильтрами проводили в растворе 50% формамида, 5×SSC, 0,1% SDS, 0,1% пирофосфата натрия, 50 мМ фосфата натрия, pH 6,8, 2× раствора Денхардта и 10% декстрансульфата при at 42°C в течение 20 часов. Отмывку фильтров проводили в водном растворе 0,1×SSC и 0,1% SDS при 42°C.
ДНК, обладающие желаемой идентичностью последовательностью с ДНК, кодирующей полноразмерную нативную последовательность PRO87299, можно затем идентифицировать с использованием общепринятых способов, известных в данной области.
ПРИМЕР 8: Экспрессия PRO87299 в E. coli
В данном примере проиллюстрировано получение негликозилированной формы PRO87299 рекомбинантной экспрессией в E. coli.
Сначала последовательность ДНК, кодирующую PRO87299, амплифицировали с использованием выбранных праймеров для PCR. Праймеры должны содержать участки для ферментов рестрикции из выбранного экспрессирующего вектора. Можно применять множество экспрессирующих векторов. Примером подходящего вектора является pBR322 (полученный из E. coli; смотри Bolivar et al., Gene, 2:95 (1977)), содержащий гены устойчивости к ампициллину и тетрациклину. Вектор расщепляли ферментом рестрикции и дефосфорилировали. Затем амплифицированные PCR последовательности лигировали в вектор. Вектор будет предпочтительно содержать последовательности, кодирующие ген устойчивости к антибиотику, промотор trp, a поли-his лидер (включая первые шесть кодонов STII, последовательность поли-his, и участок расщепления энтерокиназой), кодирующая область PRO87299, терминатор транскрипции лямбда и ген argU.
Затем смесь для лигирования использовали для трансформации выбранного штамма E. coli с использованием способов, описанных в Sambrook et al., выше. Трансформанты идентифицировали по их способности расти на чашках с LB и затем отбирали колонии, устойчивые к антибиотику. Плазмидную ДНК можно выделить и подтвердить рестрикционным анализом и секвенированием ДНК.
Отобранные клоны можно выращивать в течение ночи в жидкой культуральной среде, такой как LB-среда, дополненная антибиотиками. Ночную культуру можно затем использовать для инокуляции культуры большего объема. Затем клетки растили до желаемой оптической плотности, во время чего промотор для экспрессии является включенным.
После культивирования клеток в течение еще нескольких часов клетки можно собрать центрифугированием. Осадок клеток, полученный центрифугированием, можно солюбилизировать с использованием различных средств, известных в данной области, и затем солюбилизированный белок PRO87299 можно очистить с использованием a металл-хелатной колонки в условиях, позволяющих жесткое связывание белка.
PRO87299 можно экспрессировать в E. coli в меченной поли-His форме с использованием следующего способа. Сначала ДНК, кодирующую PRO87299, амплифицируют с использованием выбранных праймеров для PCR. Праймеры будут содержать участки для ферментов рестрикции, соответствующие участкам для ферментов рестрикции в выбранном экспрессирующем векторе, и другие полезные последовательности, обеспечивающие эффективную и надежную инициацию трансляции, быструю очистку на металл-хелатной колонке и протеолитическое удаление энтерокиназой. Затем амплифицированные PCR, меченные поли-His последовательности лигировали в экспрессирующий вектор, который использовали для трансформации хозяина - E. coli на основе штамма 52 (W3110 fuhA(tonA) lon galE rpoHts(htpRts) clpP(lacIq). Сначала трансформанты выращивали в LB, содержащей 50 мг/мл карбенициллина, при 30°C с покачиванием до достижения O.D.600 3-5. Затем культуры разводили в 50-100 раз средой CRAP (полученной смешиванием 3,57 г (NH4)2SO4, 0,71 г цитрата натрия·2H2O, 1,07 г KCl, 5,36 г дрожжевого экстракта Difco, 5,36 г Sheffield hycase SF в 500 мл воды, так же как 110 нМ MPOS, pH 7,3, 0,55% (мас./об.) глюкозы и 7 мМ MgSO4) и выращивали в течение приблизительно 20-30 часов при 30°C с покачиванием. Отбирали образцы для подтверждения экспрессии анализом в SDS-PAGE и смешанную культуру центрифугировали для осаждения клеток. Осадок клеток замораживали до очистки и повторного сворачивания.
Массу E. coli после ферментаций в от 0,5 до 1 л (осадки 6-10 г) ресуспендировали в 10 объемах (мас./об.) буфера 7 M гуанидин, 20 мМ Трис, pH 8. Добавляли твердый сульфит натрия и тетратионат натрия для получения конечных концентраций 0,1 M и 0,02 M соответственно и перемешивали раствор в течение ночи при 4°C. В результате данной стадии получают денатурированный белок со всеми остатками цистеина, блокированными сульфитизацией. Раствор центрифугируют при 40000 об/мин в ультацентрифуге Beckman в течение 30 мин. Супернатант разводят 3-5 объемами буфера для металл-хелатной колонки (6 M гуанидин, 20 мМ Трис, pH 7,4) и фильтруют через фильтры 0,22 микрон для осветления. Осветленный экстракт наносят на 5 мл металл-хелатную колонку Qiagen Ni-NTA, уравновешенную буфером для металл-хелатной колонки. Колонку промывали дополнительным буфером, содержащими 50 мМ имидазол (Calbiochem, Utrol grade), pH 7,4. Белок элюировали буфером, содержащим 250 мМ имидазол. Фракции, содержащие желаемый белок, пулировали и хранили при 4°C. Концентрацию белка оценивали по его оптической плотности при 280 нм с использованием коэффициента поглощения, рассчитанного на основе его аминокислотной последовательности.
Белки повторно сворачивают посредством медленного растворения образца в свежеприготовленном буфере для повторного сворачивания, состоящего из: 20 мМ Триса, pH 8,6, 0,3 M NaCl, 2,5 M мочевины, 5 мМ цистеина, 20 мМ глицина и 1 мМ EDTA. Объемы для повторного сворачивания выбирали так, что конечная концентрация белка составляет между 50 и 100 микрограмм/мл. Раствор для повторного сворачивания осторожно перемешивали при 4°C в течение 12-36 часов. Реакцию повторного сворачивания останавливали добавлением TFA до конечной концентрации 0,4% (pH приблизительно 3). Перед дальнейшей очисткой белка раствор фильтровали через фильтр 0,22 микрон и добавляли ацетонитрил до конечной концентрации 2-10%. Повторно свернутый белок подвергали хроматографии на обращенно-фазовой колонке Poros Rl/H с использованием подвижной фазы - буфера 0,1% TFA с элюцией градиентом ацетонитрила от 10 до 80%. Аликвоты фракций с оптическим поглощением при A280 анализировали в SDS-полиакриламидных гелях и пулировали фракции, содержашие гомогенный повторно свернутый белок. Как правило, правильно свернутые формы большинства белков является элюированными при наименьшей концентрации ацетонитрила, так как эти формы являются наиболее компактными с их гидрофобными внутренними частями, огражденными от взаимодействия с обращенно-фазовой смолой. Агрегированные формы обычно элюируют при более высокой концентрации ацетонитрила. Кроме отделения неправильно свернутых форм белков от желаемой формы на обращенно-фазовой стадии удаляют также эндотоксин из образцов.
Фракции, содержащие формы полипептида PRO87299 с желаемым сворачиванием, объединяют и ацетонитрил удаляют с использованием мягкого потока азота, направленного на раствор. Белки составляли в 20 мМ Hepes, pH 6,8 с 0,14 M хлоридом натрия и 4% маннитом посредством диализа или гель-фильтрации с использованием смол G25 Superfine (Pharmacia), уравновешенных в буфере для состава, и стерилизовали фильтрованием.
Полипептиды PRO87299, описанные здесь, успешно экспрессировали, как описано выше.
ПРИМЕР 9: Экспрессия PRO87299 в клетках млекопитающих
В данном примере проиллюстрированно получение потенциально гликозилированной формы PRO87299 посредством рекомбинантной экспрессия в клетках млекопитающих.
Вектор pRK5 (см. EP 307247, опубликованную 15 марта 1989 г.) применяли в качестве экспрессирующего вектора. Необязательно ДНК PRO87299 лигировали в pRK5 с использованием выбранных рестрикционных ферментов, чтобы обеспечить вставку ДНК PRO87299 с использованием способов лигирования, таких как описаны в Sambrook et al., выше. Полученный вектор назвали pRK5-PRO87299.
В одном из вариантов осуществления выбранными клетками-хозяевами могут являться клетки 293. Клетки 293 человека (ATCC CCL 1573) выращивали до конфлюентности в чашках для культивирования клеток в среде, такой как DMEM, дополненной фетальной телячьей сывороткой и, необязательно, питательными компонентами и/или антибиотиками. Приблизительно 10 мкг ДНК pRK5-PRO87299 смешивают с приблизительно 1 мкг ДНК, кодирующей ген VA РНК [Thimmappaya et al., Cell, 31:543 (1982)] и растворяют в 500 мкл 1 мМ Трис-HCl, 0,1 мМ EDTA, 0,227 M CaCl2. К этой смеси по каплям добавляют 500 мкл 50 мМ HEPES (pH 7,35), 280 мМ NaCl, 1,5 мМ NaPO4 и позволяют формирование преципитата в течение 10 минут при 25°C. Преципитат суспендируют, добавляют к клеткам 293 и позволяют осесть в течение приблизительно четырех часов при 37°C. Культуральную среду отбирали и добавляли 2 мл 20% глицерина в PBS на 30 секунд. Затем клетки 293 промывали бессывороточной средой, добавляли свежую среду и инкубировали клетки в течение приблизительно 5 суток.
Приблизительно через 24 часа после трансфекции культуральную среду удаляли и заменяли культуральной средой (в чистом виде) или культуральной средой, содержащей 200 мкКи/мл 35S-цистеина и 200 мкКи/мл 35S-метионина. После 12 часов инкубации кондиционированную среду собирали, концентрировали на центрифугируемом фильтре и наносили на 15% SDS-гель. Обработанный гель можно высушить и экспонировать на пленку в течение выбранного периода времени для выявления присутствия полипептида PRO87299. Культуры, содержащие трансфицированные клетки, можно подвергать дальнейшей инкубации (в бессывороточной среде) и тестировать среду в выбранных биологических анализах.
Альтернативным способом PRO87299 можно временно ввести в клетки 293 с использованием способа с декстрансульфатом, описанного Somparyrac et al., Proc. Natl. Acad. Sci 12:7575 (1981). 293 выращивают до максимальной плотности во вращающейся колбе и добавляют 700 мкг ДНК pRK5-PRO87299. Сначала клетки концентрируют из вращающейся колбы центрифугированием и промывают PBS. Преципитат ДНК-декстран инкубируют на осадке клеток в течение четырех часов. Клетки обрабатывают 20% глицерином в течение 90 секунд, промывают средой для культивирования клеток и вводят обратно во вращающуюся колбу, содержащую среду для культивирования клеток, 5 мкг/мл бычьего инсулина и 0,1 мкг/мл бычьего трансферрина. После приблизительно четырех суток кондиционированную среду центрифугируют и фильтруют для удаления клеток и осколков. Образец, содержащий экспрессированный PRO87299, можно затем концентрировать и очищать любым выбранным способом, таким как диализ и/или хроматография на колонке.
В другом варианте осуществления можно экспрессировать PRO87299 в клетках CHO. pRK5-PRO87299 можно трансфицировать в клетки CHO с использованием известных реагентов, таких как CaPO4 или DEAE-декстран. Как описано выше, культуры клеток можно инкубировать и заменять среду культуральной средой (в чистом виде) или культуральной средой, содержащей радиоактивную метку, такую как 35S-метионин. После определения присутствия полипептида PRO87299 культуральную среду можно заменить бессывороточной средой. Предпочтительно культуры инкубируют приблизительно 6 суток и затем кондиционированную среду собирают. Среду, содержащую экспрессированный PRO87299, можно затем концентрировать и очищать любым выбранным способом.
Меченный эпитопом PRO87299 также можно экспрессировать в клетках-хозяевах CHO. PRO87299 можно субклонировать из вектора pRK5. Вставку субклона можно подвергать PCR для слияния в рамке с выбранным эпитопом-меткой, таким как метка поли-his, в экспрессирующем векторе Baculovirus. Меченную поли-his вставку PRO87299 можно затем субклонировать в вектор, содержащий промотор/энхансер SV40, содержащий селективный маркер, такой как DHFR для отбора стабильных клонов. Наконец, клетки CHO можно трансфицировать (как описано выше) вектором, содержащим промотор/энхансер SV40. Для подтверждения экспрессии можно выполнять мечение, как описано выше. Затем культуральную среду, содержащую экспрессированный PRO87299, меченный поли-His, можно концентрировать и очищать любым выбранным способом, таким как Ni2+-хелатную аффинную хроматографию.
PRO87299 можно также экспрессировать в клетках CHO и/или COS способом временной экспрессии или в клетках CHO другим способом стабильной экспрессии.
Стабильную экспрессию в клетках CHO осуществляли с использованием следующего способа. Белки экспрессировали в форме IgG-конструкции (иммуноадгезина), в которой кодирующие последовательности для растворимых форм (например, внеклеточных доменов) соответствующих белков слиты с последовательностью константной области IgG1, содержащей шарнирный, CH2 и CH2 домены и/или представляющей собой меченную поли-His форму.
После амплификации PCR соответствующие ДНК субклонировали в экспрессирующий вектор для CHO с использованием общепринятых способов, как описано в Ausubel et al., Current Protocols of Molecular Biology, Unit 3,16, John Wiley и Sons (1997). Экспрессирующие векторы для CHO конструировали с совместимыми участками рестрикции 5' и 3' от интересующей ДНК, чтобы обеспечить подходящую перезарядку челнока кДНК. Вектор, используемый для экспрессии в клетках CHO, является таким, как описано у Lucas et al., Nucl. Acids Res. 24:9 (1774-1779 (1996), с использованием раннего промотора/энхансера SV40, чтобы управлять экспрессией интересующей кДНК и дигидрофолатредуктазы (DHFR). Экспрессия DHFR позволяет отбор по стабильному поддержанию плазмиды после трансфекции.
Двенадцать микрограмм желаемой плазмидной ДНК вводили в приблизительно 10 миллионов клеток CHO с использованием коммерчески доступных реагентов для трансфекции Superfect® (Quiagen), Dosper® или Fugene® (Boehringer Mannheim). Клетки выращивали, как описано в Lucas et al., выше. Приблизительно 3×107 клеток замораживали в ампуле для дальнейшего роста и продукции, как описано ниже.
Ампулы, содержащие плазмидную ДНК, оттаивали помещением в водяную баню и перемешивали с встряхиванием. Содержимое переливали пипеткой в центрифужную пробирку, содержащую 10 мл среды, и центрифугировали при 1000 об/мин в течение 5 минут. Спернатант отбирали и клетки ресуспендировали в 10 мл селективной среды (PS20, фильтрованной через 0,2 мкм с 5% диафильтрованной через 0,2 мкм). Затем клетки аликвотировали во вращающуюся емкость 100 мл, содержащую 90 мл селективной среды. После 1-2 суток клетки переносили во вращающуюся емкость 250 мл, заполненную 150 мл селективной ростовой среды, и инкубировали при 37°C. После других 2-3 суток вращающиеся емкости 250 мл, 500 мл и 2000 мл засевали 3×105 клеток/мл. Среду для клеток заменяли свежей средой центрифугированием и ресуспендированием в среде для продукции. Хотя можно применять любую подходящую среду для CHO, в настоящее время можно применять среду для продукции, описанную в патенте США No 5122469, опубликованном 16 июня 1992 г. Вращающуюся емкость для продукции 3л засевали 1,2×106 клеток/мл. На сутки 0 определяли pH. На сутки 1 из вращающейся емкости отбирали образец и начинали разбрызгивание фильтрованным воздухом. На сутки 2 из вращающейся емкости отбирали образец, температуру изменяли на 33°C и добавляли 30 мл 500 г/л глюкозы и 0,6 мл 10% пеногасителя (например, 35% эмульсию полидиметилсилоксана, эмульсия медицинского качества Dow Corning 365). На всем протяжении продукции pH регулировали по мере необходимости для поддержания около 7,2. После 10 суток или когда жизнеспособность падала ниже 70%, культуру клеток собирали центрифугированием и фильтровали через фильтр 0,22 мкм. Фильтрат или хранили при 4°C, или немедленно наносили на колонки для очистки.
В случае конструкций, меченных поли-His, белки очищали с использованием колонки Ni-NTA (Qiagen). Перед очисткой к кондиционированной среде добавляли имидазол до концентрации 5 мМ. Кондиционированную среду подавали насосом на 6 мл колонку Ni-NTA, уравновешенную 20 мМ Hepes, pH 7,4, буфером, содержащим 0,3 M NaCl и 5 мМ имидазол, при скорости потока 4-5 мл/мин при 4°C. После нанесения колонку промывали дополнительным буфером для уравновешивания и элюировали белок буфером для уравновешивания, содержащим 0,25 M имидазол. Затем высокоочищенный белок обессоливали буфером для хранения, содержащим 10 мМ Hepes, 0,14 M NaCl и 4% маннит, pH 6,8, на 25 мл колонке G25 Superfine (Pharmacia) и хранили при -80°C.
Конструкции с иммуноадгезином (Fc-содержащие) очищали из кондиционированной среды следующим образом. Кондиционированную среду подавали насосом на 5 мл колонку с белком A (Pharmacia), уравновешенную в 20 мМ Na фосфатном буфере, pH 6,8. После нанесения, колонку интенсивно промывали буфером для уравновешивания перед элюцией 100 мМ лимонной кислотой, pH 3,5. Элюированный белок немедленно нейтрализовали сбором фракций по 1 мл в пробирки, содержащие 275 мкл 1 M Трис-буфера, pH 9. Затем высокоочищенный белок обессоливали буфером для хранения, как описано выше для белков, меченных поли-His. Гомогенность оценивали в SDS полиакриламидных гелях и N-концевым аминокислотным секвенированием деградацией по Эдману.
Многие из полипептидов PRO87299, описанных здесь, успешно экспрессировали, как описано выше.
ПРИМЕР 10: Экспрессия PRO87299 в дрожжах
В следующем способе описана рекомбинантная экспрессия PRO87299 в дрожжах.
Сначала конструировали экспрессирующие векторы для дрожжей для внутриклеточной продукции или секреции PRO87299 с промотора ADH2/GAPDH. ДНК, кодирующую PRO87299 и промотор вставляли в подходящие участки рестрикционных ферментов в выбранной плазмиде для управления внутриклеточной экспрессией PRO87299. Для секреции ДНК, кодирующую PRO87299, можно клонировать в выбранную плазмиду вместе с ДНК, кодирующей промотор ADH2/GAPDH, сигнальный пептид нативного PRO87299 или другой сигнальный пептид млекопитающих или, например, секреторный сигнал/лидерную последовательность альфа-фактора или инвертазы дрожжей, и линкерные последовательности (если неоюходимо) для экспрессии PRO87299.
Затем клетки дрожжей, такие как штамм дрожжей AB110, можно трансформировать экспрессирующими плазмидами, описанными выше, и культивировать в выбранной среде для ферментации. Супернатанты трансформированных дрожжей можно анализировать осаждением 10% трихлоруксусной кислотой и разделением SDS-PAGE с последующим окрашиванием гелей Кумасси синим.
Рекомбинантный PRO87299 можно затем выделить и очистить удалением дрожжевых клеток из среды для ферментации центрифугированием и затем концентрированием среды с использованием выбранных патронных фильтров. Концентрат, содержащий PRO87299, можно очищать далее с использованием выбранных смол для хроматографии на колонках.
Множество полипептидов PRO87299, описанных здесь, успешно экспрессировали, как описано выше.
ПРИМЕР 11: Экспрессия PRO87299 в клетках насекомых, инфицированных бакуловирусом
В следующем способе описана рекомбинантная экспрессия PRO87299 в клетках насекомых, инфицированных бакуловирусом.
Последовательность, кодирующую PRO87299, вставляли выше эпитопа-метки, содержащегося в бакуловирусном экспрессирующем векторе. Такие эпитопы-метки включают в себя метки поли-his и иммуноглобулиновые метки (подобные Fc-областям IgG). Можно применять множество плазмид, включая плазмиды, полученные из коммерчески доступных плазмид, таких как pVL1393 (Novagen). Коротко, последовательность, кодирующую PRO87299 или желаемую часть кодирующей последовательности PRO87299, такую как последовательность, кодирующую внеклеточный домен трансмембранного белка, или последовательность, кодирующую зрелый белок, если белок является внеклеточным, амплифицируют PCR с праймерами, комплементарными 5'- и 3'-областям. 5'-праймер может содержать фланкирующие (выбранные) участки для ферментов рестрикции. Затем продукт расщепляют этими выбранными ферментами рестрикции и субклонируют в экспрессирующий вектор.
Рекомбинантный бакуловирус получают котрансфекцией вышеуказанной плазмиды и ДНК вируса BaculoGold™ (Pharmingen) в клетки Spodoptera frugiperda ("Sf9") (ATCC CRL 1711) с использованием липофектина (коммерчески доступного из GIBCO-BRL). После 4-5 суток инкубации при 28°C высвобожденные вирусы собирают и используют для дальнейших амплификаций. Вирусную инфекцию и экспрессию белка осуществляют, как описано в O'Reilley et al., Baculovirus expression vectors: A Laboratory Manual. Oxford: Oxford University Press (1994).
Экспрессированные меченные поли-his PRO87299 можно затем очистить, например, Ni2+-хелатной аффинной хроматографией следующим образом. Из инфицированных рекомбинантным вирусом клеток Sf9 получали экстракты, как описано в Rupert et al., Nature, 362:175-179 (1993). Коротко, клетки Sf9 промывали, ресуспендировали в буфере для обработки ультразвуком (25 мл Hepes, pH 7,9; 12,5 мМ MgCl2; 0,1 мМ EDTA; 10% глицерин; 0,1% NP-40; 0,4 M KCl) и обрабатывали дважды в течение 20 секунд на льду. Полученные воздействием ультразвука гомогенаты осветляли центрифугированием и супернатант разводили в 50 раз в буфере для нанесения (50 мМ фосфат, 300 мМ NaCl, 10% глицерин, pH 7,8) и фильтровали через 0,45 мкм фильтр. Получали колонку с Ni2+-NTA агарозой (коммерчески доступной из Qiagen) с объемом слоя 5 мл, промывали 25 мл воды и уравновешивали 25 мл буфера для нанесения. Фильтрованный экстракт клеток наносили на колонку при 0,5 мл в минуту. Колонку промывали до фонового A280 буфером для нанесения и в этой точки начинали сбор фракций. Затем колонку промывали вторым буфером для промывки (50 мМ фосфат; 300 мМ NaCl, 10% глицерин, pH 6,0), элюирующим неспецифически связавшийся белок. После повторного достижения фонового A280, колонку обрабатывали градиентом имидазола 0-500 мМ во втором буфере для промывки. Собирали фракции по одному мл и анализировали SDS- PAGE и окрашиванием серебром или вестерн-блоттингом с Ni2+-NTA, конъюгированным со щелочной фосфатазой (Qiagen). Фракции, содержащие элюированный элюированный меченный His10 PRO87299, объединяли и диализовали против буфера для нанесения.
Альтернативно очистку меченного IgG (или меченого Fc) PRO87299 можно проводить с использованием известных способов хроматографии, включая, например, хроматографию на колонке с белком A или белком G.
Множество полипептидов PRO87299, описанных здесь, успешно экспрессировали, как описано выше.
ПРИМЕР 12: Получение антител, связывающих PRO87299
Антитела, специфически связывающие PRO87299, получали иммунизацией мыши конструкцией PRO87299-Fc. Эту конструкцию получили лигированием области, кодирующей внеклеточный домен PRO87299 (аминокислоты 1-155), в плазмиду, содержащую Fc-домен человека, таким образом получая химеру PRO87299(ECD)-Fc. Этот белок продуцировали, очищали и инъецировали в подушечку лапы мыши.
Способы получения моноклональных антител известны в данной области и описаны, например, в Goding, выше. Мышей, таких как Balb/c, иммунизировали химерой PRO87299(ECD)-Fc, эмульгированной в полном адъюванте Фрейнда и инъецированной подкожно или внутрибрюшинно в количестве от 1-100 микрограмм. Альтернативно иммуноген эмульгируют в адъюванте MPL-TDM (Ribi Immunochemical Research, Hamilton, MT) и инъецируют в подушечку задней лапы животного. Иммунизированных мышей стимулировали 10-12 суток спустя дополнительной химерой PRO87299(ECD)-Fc, эмульгированной в выбранном адъюванте. Затем в течение нескольких недель мышей можно также стимулировать дополнительными иммунизирующими инъекциями. Образцы сыворотки периодически получали от мышей посредством ретроорбитального забора крови для тестирования в анализах ELISA для детекции анти-PRO87299 антител.
После детекции подходящего титра антитела животным, «положительным» по антителам, инъецировали последнюю внутривенную инъекцию химеры PRO87299(ECD)-Fc. От трех до четырех суток спустя мышей умерщвляли и собирали клетки селезенки. Затем клетки селезенки сливали (с использованием 35% полиэтиленгликоля) с выбранной линией клеток миеломы мыши, такой как P3X63AgU.l, доступной из ATCC, No. CRL 1597. Слиянием получали клетки гибридомы, которые затем рассевали на 96-луночные культуральные планшеты, содержащие среду HAT (гипоксантин, аминоптерин и тимидин) для подавления пролиферации неслитых клеток, гибридов миеломы и гибридов клеток селезенки.
Клетки гибридомы скринировали в ELISA по реактивности против PRO87299. Получили девять антител, специфически связывающихся с PRO87299. Положительные клетки гибридомы инъецировали внутрибрюшинно сингенным мышам Balb/c для получения асцитов, содержащих анти-PRO87299 моноклональные антитела. Альтернативно клетки гибридомы можно выращивать во флаконах для культуры клеток или роллерных флаконах. Очистку моноклональных антител, продуцированных в асцитах, осуществляли с использованием осаждения сульфатом аммония с последующей эксклюзионной хроматографией. Альтернативно можно применять аффинную хроматографию, основанную на связывании антитела с белком A или белком G.
Как указано ранее, получили девять (9) антител, специфически связывающихся с PRO87299. Определили, что из этих девяти антитело, обозначенное 5F5.1, является антителом-агонистом. Активность агониста определяли по подавлению пролиферации CD4+ T-клеток. CD 4+ T-клетки выделяли из крови человека, как описано ранее в примере 2, и культивировали с перекрестно-сшитыми антителами, связанными с плашетом. 96-луночные планшеты подготавливали инкубацией в течение ночи при 37 °C с антителами козы против IgG мыши в концентрации 10 мкг/мл и промывали PBS для удаления избытка. Анти-CD3/анти-CD28 антитела добавляли к покрытым IgG планшетам с анти-PRO87229 антителом или без него. Сочетание анти-CD3(0,1 мкг/мл) и анти-CD28 (0,25 мкг/мл) стимулирует пролиферацию CD4+ T-клеток, и добавление анти-PRO87299 антитела уменьшает пролиферацию в 5 раз, как измеряют по поглощению тимидина (фиг.13). Для одного анти-CD3 антитела не показали пролиферации и для контрольного антитела также не показали пролиферации. Активность агониста анти-PRO87299 антитела 5F5.1 можно прекратить тепловой инактивацией. Линию гибридомы 5F5.1 депонировали в ATCC согласно Будапештскому соглашению (см. пример 19).
Второй эксперимент проводили с использованием растворимых антител. Для анти-CD3 (10 мкг/мл) и анти-CD28 (5 мкг/мл) при добавлении к кльтуральной среде также показали увеличение пролиферации CD 4+ T-клеток. Для анти-PRO87299 антитела, добавленного в культуральную среду с двумя стимулирующими антителами, показали подавление пролиферации CD4+ T-клеток вплоть до 50%. Анти-PRO87299 антитело использовали в диапазоне 5-200 мкг/мл, и подавление пролиферации CD4+ клеток являлось зависимым от дозы в этом диапазоне.
Антитело 5E10 является антителом, специфически связывающимся с PRO87299, но не обладающим агонистическим эффектом. Показано, что это антитело распознает PRO87299 на первичных B-клетках и CD4+ T-клетках. С ним можно также распознавать трансфицированные PRO87299 клетки с использованием анализа FACS. Антитело 5E10 воспроизводимо не обладает агонистическим эффектом.
Таким образом, получены антитела, специфически связывающие PRO87299. Анти-PRO87299 антитела-антагонисты являются полезными для стимуляции иммунного ответа, что может являться полезным для лечения иммунодефицитов и отражения инфекции патогенами. Анти-PRO87299 антитела-агонисты являются полезными для уменьшения пролиферации CD4+ T-клеток, таким образом, уменьшая иммунный ответ, и могут являться полезными для лечения аутоиммунных заболеваний, лимфомы и воспалительного заболевания кишечника.
ПРИМЕР 13: Очистка полипептидов PRO87299 с использованием специфических антител
Природные или рекомбинантные полипептиды PRO87299 можно очищать множеством общепринятых в данной области способов очистки белка. Например, прополипептид PRO87299, зрелый полипептид PRO87299, или преполипептид PRO87299 очищают иммуноаффинной хроматографией с использованием антител, специфических для интересующего полипептида PRO87299. Как правило, иммуноаффинную колонку получают ковалентным присоединением антитела против полипептида PRO87299 к активированной хроматографической смоле.
Поликлональные иммуноглобулины получают из иммунной сыворотки или осаждением сульфатом аммония или очисткой на иммобилизованном белке A (Pharmacia LKB Biotechnology, Piscataway, N.J.). Подобным образом моноклональные антитела получают из асцитной жидкости мыши осаждением сульфатом аммония или хроматографией на иммобилизованном белке A. Частично очищенный иммуноглобулин ковалентно присоединяют к хроматографической смоле, такой как CnBr-активированной SEPHAROSE™ (Pharmacia LKB Biotechnology). Антитело соединяли со смолой, смолу блокировали и производное смолы промывали согласно инструкциям производителя.
Такую иммуноаффинную колонку использовали для очистки полипептида PRO87299 посредством получения фракции из клеток, содержащих полипептид PRO87299 в растворимой форме. Этот препарат получали солюбилизацией целой клетки или внутриклеточной фракции, полученной дифференциальным центрифугированием, добавлением детергента или другими способами, хорошо известными в данной области. Альтернативно растворимый полипептид PRO87299, содержащий сигнальную последовательность, может являться секретированным в приемлемом количестве в среду, в которой выращивали клетки.
Препарат, содержащий растворимый полипептид PRO87299, пропускали через иммуноаффинную колонку и промывали колонку в условиях, позволяющих предпочтительное поглощение полипептида PRO87299 (например, буферами высокой ионной силы в присутствии детергента). Затем проводили элюцию с колонки в условиях, нарушающих связывание антитело/PR087299 полипептид (например, буфером с низким pH, таким как приблизительно pH 2-3, или с высокой концентрацией хаотропного средства, такого как мочевина или ион тиоцианата), и собирали полипептид PRO87299.
ПРИМЕР 14: Скрининг лекарственных средств
Настоящее изобретение является особенно применимым для скрининга соединений с использованием полипептидов PRO87299 или их связывающих фрагментов в любом из множества способов скрининга лекарственных средств. Полипептид PRO87299 или фрагмент, применяемые в таком тесте, могут присутствовать или свободными в растворе, прикрепленными к твердой подложке, переносимыми на клеточной поверхности, или локализованными внутри клетки. В одном способе скрининга лекарственных средств используют эукариотические или прокариотические клетки-хозяева, стабильно трансформированные рекомбинантными нуклеиновыми кислотами, экспрессирующими полипептид или фрагмент PRO87299. Проводят скрининг лекарственных средств против таких трансформированных клеток в анализах конкурентного связывания. Такие клетки в жизнеспособной или фиксированной форме можно использовать для общепринятых анализов связывания. Можно измерять, например, формирование комплексов между полипептидом фрагментом PRO87299 или и тестируемым средством. Альтернативно можно исследовать уменьшение формирования комплекса между полипептидом PRO87299 и его клеткой-мишенью или рецепторами-мишенями, вызванное тестируемым средством.
Таким образом, настоящее изобретение относится к способам скрининга лекарственных средств или любых других средств, которые могут влиять на связанное с полипептидом PRO87299 заболевание или расстройство. Эти способы включают в себя контактирование такого средства с полипептидом или фрагментом PRO87299 и анализ (i) присутствия комплекса между средством и полипептидом или фрагментом PRO87299, или (ii) присутствия комплекса между полипептидом или фрагментом PRO87299 и клеткой, способами, хорошо известными в данной области. В таких анализах конкурентного связывания полипептид или фрагмент PRO87299, как правило, является меченым. После соответствующей инкубации свободный полипептид или фрагмент PRO87299 отделяют от полипептида или фрагмента PRO87299, присутствующего в связанной форме, и количество свободной или не входящей в комплекс метки является показателем способности конкретного средства связывать полипептид PRO87299 или мешать комплексу полипептид PRO87299/клетка.
Другой способ скрининга лекарственного средства включает в себя высокопроизводительный скрининг соединений, обладающих подходящей аффинностью связывания с полипептидом и подробно описан в WO84/03564, опубликованном 13 сентября 1984 г. В кратком изложении, большое число различных малых пептидных тестируемых соединений синтезируют на твердом субстрате, таком как пластиковые иглы или какая-либо другая поверхность. Применительно к полипептиду PRO87299 проводят реакцию тестируемых пептидных соединений с полипептидом PRO87299 и промывку. Связанный полипептид PRO87299 детектируют способами, хорошо известными в данной области. Очищенным полипептидом PRO87299 можно также непосредственно покрывать планшеты для использования в вышеупомянутых способах скрининга лекарственных средств. Кроме того, можно использовать не нейтрализующие антитела для захвата пептида и иммобилизации его на твердой подложке.
Настоящее изобретение относится также к применению конкурентных анализов скрининга лекарственных средств, в которых нейтрализующие антитела, способные связывать полипептид PRO87299, специфически конкурируют с тестируемым соединением за связывание с полипептидом PRO87299 или его фрагментами. Таким способом антитела можно использовать для детекции любого пептида, разделяющего одну или несколько антигенных детерминант с полипептидом PRO87299.
ПРИМЕР 15: Рациональный дизайн лекарственных средств
Целью рационального дизайна лекарственных средств является получение структурных аналогов интересующего биологически активного полипептида (т.е. полипептида PRO87299) или малых молекул, с которыми они взаимодействуют, например агонистов, антагонистов или ингибиторов. Любой из данных примеров можно использовать для моделирования лекарственных средств, которые являются более активными или стабильными формами полипептида PRO87299 или которые усиливают или препятствуют функционированию полипептида PRO87299 in vivo (c.f., Hodgson, Bio/Technology, 9: 19-21 (1991)).
По одному споособу трехмерную структуру полипептида PRO87299 или комплекса полипептид PRO87299 - ингибитор определяют рентгенокристаллографией, компьютерным моделированием или, более обычно, сочетанием двух способов. Следует установить и форму, и заряды полипептида PRO87299 для выяснения структуры и определения активного сайта(сайтов) молекулы. Менее часто полезную информацию относительно структуры полипептида PRO87299 можно получать посредством моделирования на основе структуры гомологичных белков. В обоих случаях важную информацию о структуре используют для дизайна аналогичных молекул, подобных полипептиду PRO87299, или для идентификации эффективных ингибиторов. Полезные примеры рационального дизайна лекарственных средств могут включать в себя молекулы, обладающие улучшенной активностью или стабильностью, как показано Braxton and Wells, Biochemistry. 3_l:7796-7801 (1992), или действующие как ингибиторы, агонисты или антагонисты природных пептидов, как показано Athauda et at, J. Biochem., 113:742-746 (1993).
Возможно также выделить специфическое для мишени антитело, отобранное функциональным анализом, как описано выше, и затем решить его кристаллическую структуру. Этим способом, в принципе, получают фармакор, на котором можно основывать последующий дизайн лекарственного средства. Возможно полностью избежать кристаллографии белка, получая антиидиотипические антитела (анти-id) к функциональному фармакологически активному антителу. Ожидают, что как зеркальное отражение зеркального отражения участок связывания анти-id будет аналогом исходного рецептора. Анти-id можно затем использовать для идентификации и выделения пептидов из банков химически или биологически полученных пептидов. Затем выделенные пептиды будут играть роль фармакора.
Благодаря настоящему изобретению можно сделать доступными значительные количества полипептида PRO87299 для проведения таких аналитических исследований, как рентгенокристаллография. Кроме того, знание аминокислотной последовательности полипептида PRO87299, представленной здесь, будет предоставлять руководство для использования способов компьютерного моделирования вместо или в дополнение к рентгенокристаллографии.
ПРИМЕР 16 - PRO87299 специфически связывает HVEM
Скринингом библиотеки белков определили, что PRO87299 специфически связывается с HVEM. Внеклеточный домен PRO87299 сливали с Fc человека для получения PRO87299(ECD)-Fc. Этот слитый белок присоединяли по аминогруппе к сенсорному чипу CM5 Biacore™ (BIAcore, Inc., Piscataway, NJ) на приблизительно 9000 единиц ответа, как описано в общих чертах в Chen, Y. et al., J. MoI Biol 293:865-881 (1999). Коротко, карбоксиметилированные декстрановые биосенсорные чипы (CM5, BIAcore™ Inc.) активировали гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) согласно инструкциям поставщика. PRO87299(ECD)-Fc растворяли и впрыскивали при скорости потока для достижения приблизительно 9000 единиц ответа (RU) присоединенного белка. Во второй проточной ячейке на тот же чип присоединяли по аминогруппе контрольный белок, PRO4346-Fc человека (инвентарный номер в Genbank AK057097), на приблизительно 18500 единиц ответа. Выполняли впрыскивание 1М этаноламина для блокирования непрореагировавших групп. Затем отдельные белки из библиотеки белков SPDI, состоящей из приблизительно 2000 белков, инъецировали в концентрации 2мкг/мл и оценивали связывание по изменению единиц ответа как функции от времени. Обнаружили, что HVEM связывает PRO87299(ECD)-Fc, но не контрольный PRO4346-Fc. Как показано на фиг.14, связывание PRO87299 и HVEM приводит к высокодостоверному увеличению 1200 единиц ответа по сравнению с контролем. Скорости связывания (kon) и скорости диссоциации (koff) рассчитывали с использованием простой модели связывания Ленгмюра один к одному (программное обеспечение BIAcore™ Evaluation версии 3.2) посредством одновременного подбора кривой по точкам для сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) рассчитывали как отношение koff/kon.
PRO87299 избирательно связывается с HVEM, как показано на фиг.15. В этом эксперименте PRO87299(ECD)-Fc и члены семейства CD28 hCTLA4-Fc, hPD-l-Fc, hICOS-Fc или hCD28-Fc присоединяли по аминогруппе к чипу Biacore™ CM5 на приблизительно 8000 единиц ответа. Затем каждый анализировали по способности связывать HVEM. Белок HVEM-Fc получали лигированием нуклеиновых кислот, кодирующих аминокислоты 1-199 HVEM, с Fc, получая слитый белок HVEM-Fc. Этот HVEM-Fc клонировали в экспрессирующий вектор, продуцирующий слитый белок при стабильной трансфекции в клетки CHO. Все меченные Fc белки очищали до более чем 90% чистоты аффинной хроматографией с использованием белок A Sepharose™ (Amersham). В результате получили, что 2мкг/мл HVEM-Fc, инъецированный при 5мкл/мин, избирательно связывается с PRO87299, но не с другими тестированными членами семейства CD28. Положительную активность каждого члена семейства CD28 подтверждали тестированием ответа связывания с их известными лигандами (данные не представлены) (члены семейства CD28 и лиганды закупали в R&D systems).
Связывание PRO87299 с HVEM является зависимым от pH, но независимым от концентрации NaCl. При использовании анализа Biacore™, как описано выше, два независимых препарата HVEM-Fc связывались с PRO87299, присоединенным по аминогруппе к чипу Biacore™ CM5. Как показано на фиг.16, связывание не нарушалось при обработке 2,5M NaCl; однако комплекс PRO87299/HVEM можно было диссоциировать 10мМ глицином при pH 3,0.
Взаимодействие PRO87299/HVEM можно блокировать антителами к PRO87299. В данном эксперименте HVEM-Fc присоединяли по аминогруппе к сенсорному чипу Biacore™ CM5 на приблизительно 7500 единиц ответа. Впрыскивания PRO87299(ECD)-Fc проводили две минуты при скорости потока 5мкл/мин. Единицы ответа (RU) считывали через 105 секунд после впрыскивания. Для PRO87299(ECD)-Fc (8 нМ) получили ответ 100 RU. Каждое антитело в увеличивающейся концентрации предварительно смешивали с PRO87299(ECD)-Fc (8 нМ) и инкубировали в течение 1 час. Связывание каждого образца измеряли в случайном порядке и в двух повторах и 10 мМ глицина pH 2,5 использовали для регенерации связывающей поверхности после каждого впрыскивания. Как показано на фиг.18, анти-PRO87299 антитела 5E10 и 3Bl.9 блокируют связывание PRO87299 с HVEM зависимым от концентрации образом, в то время как антитело-агонист 5F5.1 не обладает блокирующим эффектом. Для впрыскиваемых антител самих по себе не показали связывания с иммобилизованным HVEM (данные не представлены). Концентрации рассчитывали на основании кажущейся уменьшенной молекулярной массы of PRO87299(ECD)-Fc (55 кДа) и массы антитела 150 кДа.
PRO87299 связывается с HVEM в анализе на основе клеток. В этом эксперименте клетки 293 человека (ATCC CCL 1573) выращивают до конфлюентности в планшетах для культивирования клеток в среде, такой как DMEM, дополненный фетальной телячьей сывороткой и, необязательно, питательными компонентами и/или антибиотиками. Приблизительно 10 мкг ДНК pRK5-HVEM смешивают с приблизительно 1 мкг ДНК, кодирующей ген VA РНК [Thimmappaya et al., Cell, 3_1:543 (1982)], и растворяли в 500 мкл 1 мМ Трис-HCl, 0,1 мМ EDTA, 0,227 M CaCl2. К этой смеси по каплям добавляют 500 мкл 50 мМ HEPES (pH 7,35), 280 мМ NaCl, 1,5 мМ NaPO4 и осадку позволяют формироваться в течение 10 минут при 25°C. Осадок суспендируют и добавляют к клеткам 293 и позволяют осесть в течение приблизительно четырех часов при 37°C. Культуральную среду отбирают и 2 мл 20% глицерина в PBS добавляют в течение 30 секунд. Затем клетки 293 промывают в бессывороточной среде, добавляют свежую среду и клетки инкубируют в течение приблизительно 5 суток. Альтернативно приблизительно 10 мкг ДНК pRK5-HVEM смешивают с реагентом Lipofectamine™ (Gibco/BRL, Gaithersburg MD) и проводят трансфекцию, следуя инструкциям производителя. После трансфекции клеток 293 HVEM их инкубировали с радиоактивно меченным I125 PRO87299(ECD)-Fc и позволяли происходить связыванию. Затем радиоактивно меченный PRO87299 конкурировал с немеченым PRO87299(ECD)-Fc и ожидаемое число общих участков связывания и константу диссоциации (Kd) определяли анализом Скэтчарда. Фиг.17(A) представляет собой график Скэтчарда, и фиг. 17(B) представляет собой график смещения, показывающий связывание PRO87299(ECD)-Fc с клетками, временно трансфицированными HVEM. Эти данные показывают, что Kd для PRO87299 составляет приблизительно 25 нМ. Общее связывание радиоактивно меченного PRO87299(ECD)-Fc с клетками 293 HEK с пустой трансфекцией составляло 2,5% от связывания с трансфицированными HVEM клетками (данные не представлены). С использованием этого способа определяли аффинность взаимодействий HVEM/LIGHT/PRO87299. Она указана в таблице 7 ниже.
Анти-PRO87299 антитело (3B1.9) дозозависимым образом конкурирует с HVEM за связывание с PRO87299(ECD)-Fc, как обсуждают ранее в этом примере.
Вместе взятые, эти данные показывают, что PRO87299 специфически связывается с HVEM, как определили посредством белок-белкового взаимодействия, блокирования белок-антитело и анализом in vivo.
ПРИМЕР 17 - PRO87299 и LIGHT могут связывать HVEM одновременно
Опубликовано, что LIGHT (GenBank Accession No: NM_172014, SEQ ID NO:5, SEQ ID NO:6) может связываться с HVEM (Marsters, S.A. et al., Curr. Biol. 8 (9), 525-528 (1998). Mauri D.N. et al, Immunity (8), 21-30, (1998)). Чтобы подтвердить, что LIGHT и PRO87299 могут связывать HVEM одновременно, авторы настоящего изобретения провели эксперимент по совместному связыванию. HVEM-Fc присоединяли по аминогруппе к сенсорному чипу Biacore™ CM5 на приблизительно 150 единиц ответа. Меньшее количество иммобилизованного HVEM-Fc являлось важным для достижения почти насыщенного связывания PRO87299-Fc. На фиг.19 показано, что LIGHT (25 нМ) и PRO87299-Fc (17 нМ) впрыскивали независимо при 5мкл/мин в течение 1200 секунд (красные и синие кривые соответственно). Затем таким же образом впрыскивали смесь LIGHT и PRO87299-Fc в тех же самых конечных концентрациях (зеленая кривая). Рассчитанная сумма сенсограмм отдельно впрыснутых LIGHT и PRO87299-Fc (серая кривая) близко совпадает с экспериментальными данными, подтверждая, что LIGHT и PRO87299 способны связывать HVEM одновременно. Концентрации основаны на молекулярной массе мономера LIGHT 25 кДа и сниженной молекулярной массе PRO87299-Fc (55 кДа).
LIGHT не блокирует взаимодействие PRO87299 с HVEM. PRO87299-Fc присоединяли по аминогруппе к сенсорному чипу Biacore™ CM5 на приблизительно 9800 единиц ответа. LIGHT (закупленный в Alexis™, Cat.#552-018- C010) инкубировали в увеличивающейся концентрации с 4 нМ HVEM в течение 1 часа. Как показано на фиг.20, увеличивающиеся концентрации LIGHT до конечной концентрации 300 нМ не блокируют связывание PRO87299-Fc/HVEM. Активность LIGHT подтверждали связыванием с иммобилизованным HVEM-Fc на сенсорном чипе CM5 (данные не представлены). LIGHT один не связывается с иммобилизованным PRO87299-Fc (данные не представлены). Все сенсограммы связывания выполняли в случайном порядке и в двух повторах. Единицы ответа записывали как разницу между фоновым ответом и ответом за 15 секунд перед окончанием 2 мин впрыскивания при 5 мкл/мин. Концентрации основаны на молекулярной массе 25 кДа для мономера LIGHT и 55 кДа для мономера HVEM. Эти данные показывают, что LIGHT не блокирует связывание HVEM с PRO87299.
HVEM первоначально идентифицировали как клеточный рецептор для входа в клетку вируса простого герпеса - 1 (HSV-1), и показали, что он связывает гликопротеин D (gD) HSV-I (Mongomery et al., Cell 87:427-436 (1996)). HVEM содержит три богатых цистеином домена (CRD), которые являются общим структурным свойством всех членов семейства TNFR. Решение кристаллической структуры белка gD в комплексе с HVEM показало, что первая CRD HVEM связывает белок gD, тогда как вторая CRD обеспечивает структурную поддержку первой CRD (Carfi et al., MoI. Cell 8: 169-179 (2001)). Тот результат, что PRO87299 взаимодействует с LIGHT и не конкурирует за связывание с HVEM, приводит к гипотезе, что PRO87299 взаимодействует с внешней поверхностью комплекса HVEM/LIGHT. Для проверки этой гипотезы сконструировали полученный из фага пептид (BP-2), способный блокировать связывание HVEM с гликопротеином D (gD) герпеса, но не с LIGHT (Sarrias et al., Mol. Immuno. 37:665-673: (2000)). В концентрациях, ингибирующих связывание gD, BP-2 ингибирует связывание PRO87299 с HVEM (фиг.21A). При прямом сравнении рекомбинантный белок gD (форма Δ290-299, Milne et al., J Virology. 77:8962-8972 (2003)) также ингибирует связывание HVEM с PRO87299 (фиг.21B). Эти результаты повторяли в анализах связывания клеток, показывая, что рекомбинантный gD (Δ 290-299) ингибирует связывание растворимого PRO87299-Fc и HVEM-Fc с клетками 293, экспрессирующими HVEM или PRO87299 (фиг.21C и D соответственно). Эти результаты показывают, что PRO87299 взаимодействует с первой CRD HVEM в участке, отдаленном от участка связывания LIGHT. Обнаружение, что PRO87299 взаимодействует и с LIGHT и с HVEM, будет позволять получение антител или малых молекул против HVEM, которые могут избирательно ингибировать взаимодействие PRO87299/HVEM без нарушения взаимодействия PRO87299/LIGHT. Другим типом антитела или малой молекулы, рассматриваемой при этом обнаружении, является тот, который будет ингибировать и взаимодействие PRO87299/HVEM, и взаимодействие PRO87299/LIGHT. Эти два различных класса молекул могут обладать различными терапевтическими эффектами.
ПРИМЕР 18: PRO87299 подавляет активацию T-клетки
Как упомянуто в примере 1, PRO87299 клонировали для дальнейших исследований белков, содержащих домен ITIM. Внутриклеточный домен PRO87299 содержит два домена ITIM, которые являются индуцибельно фосфорилированными, что позволяет привлечение и связывание SHP-I и SHP-2, что указывает на то, что функция PRO87299 является подавляющей (Watanabe N., et al., Nature Immuno. 1-10 (2003)). В этом эксперименте первичные CD4+ T-клетки стимулируют различными концентрациями иммобилизованного анти-CD3 антителом. Fc-меченный контрольный белок, HVEM-Fc или DcR3-Fc, также перекрестно сшивали с планшетом с использованием предварительного покрытия анти-Fc антителом. Пролиферацию CD4+ T-клеток измеряли включением H3 тимидина после 72-часовой инкубации. Этот эксперимент проводили в трех повторах лунок, и подавление указано как среднее (фиг.22A). Выдвинули гипотезу, что HVEM является подавляющим для пролиферации T-клеток из-за его блокирования LIGHT. Этот эксперимент показал, что неподавление HVEM/LIGHT, активация HVEM PRO87299 вызывает уменьшение пролиферации T-клеток.
Чтобы дополнительно показать, что PRO87299 является подавляющим для пролиферации T-клеток, проводили эксперимент, как описано выше, но включая подавляющее анти-PRO87299 антитело 3Bl.9. Показано, что антитело 3B1.9 блокирует связывание PRO87299 с HVEM (см. пример 16), когда CD4+ T-клетки стимулировали иммобилизованными на планшетах анти-CD3 и +/- HVEM. Затем к среде добавляли антитело 3B1.9 и контрольное антитело. Данные показывают, что антитело 3B1.9 может мешать взаимодействию PRO87299/HVEM и, таким образом, освобождает клетку от подавляющего сигнала. В результате обработанные 3B1.9 пролиферируют с той же самой скоростью, как и необработанные клетки (фиг.22B). Только при использовании больших концентраций HVEM антитело 3B1.9 являлось неэффективным. Эти данные показывают, что PRO87299 или антитела-агонисты можно применять для подавления связанных с T-клетками аутоиммунных заболеваний или, напротив, антитела-антагонисты, такие как 3B1.9, будут применимыми для стимуляции T-клеток для отражения инфекции патогенами.
ПРИМЕР 19: Реакция транплантат против хозяина
Реакция транплантат против хозяина возникает, когда иммунокомпетентные клетки трансплантируют пациентам с ослабленным иммунитетом. T-клетки донора узнают антигены хозяина и становятся активированными, секретируют цитокины, пролиферируют и дифференцируются в эффекторные клетки. Этот ответ известен как реакция транплантат против хозяина (GVHR). Ответ GVHR включают в себя синдром полиорганной недостаточности, и эффекты могут варьировать от угрожающего жизни тяжелого воспаления до умеренных случаев диареи и потери веса. Модели реакции трансплантат против хозяина на мышах используют для моделирования клинических расстройств острой и хронической GVHR, которые могут возникать после трансплантации костного мозга и аутоиммунных заболеваний. Общий способ подробно описан в Current Protocols in Immunology, выше, часть 4.3. В данном случае PBMC человека очищали из лейкаферезной массы нормального донора градиентом фикола. CD8 и NK истощали с использованием наборов для истощения MACS CD8 NK-клеток. 40×106 клеток инъецировали самке мыши SCID Beige в возрасте 8-10 недель на сутки 0. 100 мкг HVEM-Fc или контрольного белка инъецировали внутривенно на сутки 0, 2, 4, 6, 8, 10. Как показано на фиг.23, активация PRO87299 посредством HVEM-Fc в модели GVHR значительно продлевала выживание. Мыши, не обработанные HVEM-Fc, обладали 100% смертностью на 13 сутки после восстановительной терапии. Мыши, обработанные HVEM-Fc, оставались живыми на 19 сутки после восстановительной терапии в одной процедуре и до 30 суток после восстановительной терапии в другой. Этот результат показывает, что активация PRO87299 агонистом будет являться применимой в трансплантации ткани, где введение агониста PRO87299 будет предупреждать или уменьшать отторжение трансплантированной ткани хозяином.
ПРИМЕР 20: Депонирование материалов
Следующая линия клеток гибридомы депонирована в Американскую коллекцию типовых культур, 10801 University Blvd., Manassas, VA 20110-2209 USA (ATCC):
Данное депонирование выполнено согласно условиям Будапештского Соглашения о международном признании депонирования микроорганизмов для целей патентной процедуры и инструкциям в силу данного соглашения (Будапештское Соглашение). Это гарантирует поддержание жизнеспособной культуры в течение 30 лет от даты депонирования. Линия клеток будет сделана доступной в ATCC по положениям Будапештского Соглашения и подчиняется соглашению между Genentech, Inc и ATCC, которое гарантирует, что (a) доступ к культуре будет обеспечен в течение рассмотрения патентной заявки для кого-либо определенного членом комиссии как указанного для данной цели согласно 37 CFR § 1.14 и 35 USC § 122, и (b) все данные ограничения публичную доступность культуры, депонированной таким образом, будут окончательно сняты после выдачи патента. Патентообладатель настоящей заявки согласен с тем, что если культура в депозите погибнет или будет потеряна или уничтожена при культивировании при соответствующих условиях, ее незамедлительно заменят по уведомлении жизнеспособным образцом такой же культуры. Доступность депонированной линии клеток не следует рассматривать как лицензию для осуществления изобретения на практике в нарушение прав, предоставленных под властью любого правительства в соответствии с его патентным законодательством.
Приведенное выше письменное описание полагают достаточным для обеспечения осуществления изобретения на практике специалистом в данной области. Объем настоящего изобретения не является ограниченным депонированным материалом, поскольку депонированный вариант осуществления предназначен в качестве отдельной иллюстрации конкретных аспектов по изобретению и любые конструкции, являющиеся функционально эквивалентными, входят в объем настоящего изобретения. Депонирование материала здесь не составляет признания, что письменное описание, содержащееся здесь, не является адекватным, чтобы позволить практическое применение любого аспекта по изобретению, включая его лучший вариант, и его не следует истолковывать как ограничивающее объем формулы изобретения до конкретных представленных в нем иллюстраций. Действительно, различные модификации по изобретению в дополнение к показанным и описанным здесь, становятся очевидными специалистам в данной области из приведенного выше описания и попадают в объем прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕТЕРОДИМЕРНЫЕ ПОЛИПЕПТИДЫ IL-17 A/F И ВОЗМОЖНОСТИ ИХ ЛЕЧЕБНОГО ПРИМЕНЕНИЯ | 2007 |
|
RU2440134C2 |
| ИММУНОСУПРЕССОРНЫЕ ПОЛИПЕПТИДЫ И НУКЛЕИНОВЫЕ КИСЛОТЫ | 2008 |
|
RU2506275C2 |
| АГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ NOTCH3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ NOTCH3-АССОЦИИРОВАННЫХ ЗАБОЛЕВАНИЙ | 2007 |
|
RU2461569C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТИЖЕНИЯ ИММУННОЙ СУПРЕССИИ | 2001 |
|
RU2270691C2 |
| АНТИТЕЛА К ОХ-2/СD200 И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2520088C2 |
| СПОСОБЫ ВЫЯВЛЕНИЯ ВОСПАЛИТЕЛЬНОГО ЗАБОЛЕВАНИЯ КИШЕЧНИКА | 2008 |
|
RU2502806C2 |
| АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ Notch3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С Notch3 ЗАБОЛЕВАНИЙ | 2007 |
|
RU2493167C2 |
| ПОЛИПЕПТИДЫ И ПОЛИНУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ИММУНОЛОГИЧЕСКИХ НАРУШЕНИЙ И РАКА | 2012 |
|
RU2623161C2 |
| КОНСТРУКЦИИ СЛИЯНИЯ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ АНТИТЕЛ С ПОВЫШЕННЫМИ АФФИННОСТЬЮ СВЯЗЫВАНИЯ Fc-РЕЦЕПТОРА И ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2010 |
|
RU2623167C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДУЛЯЦИИ ГЕПСИНОМ СТИМУЛИРУЮЩЕГО МАКРОФАГИ БЕЛКА | 2010 |
|
RU2539772C2 |
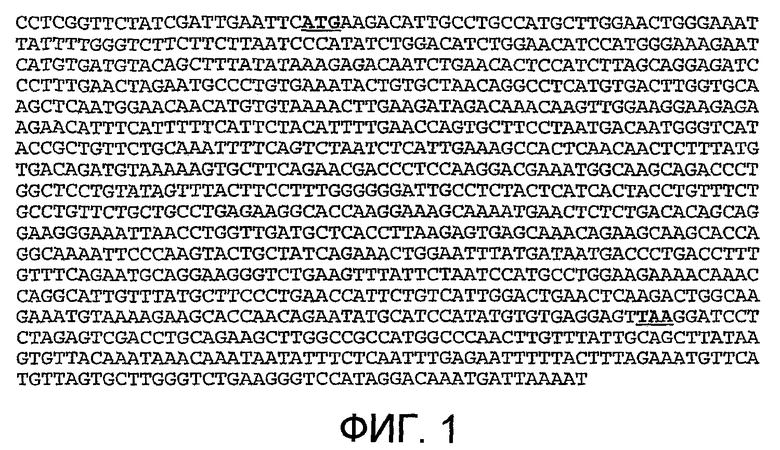



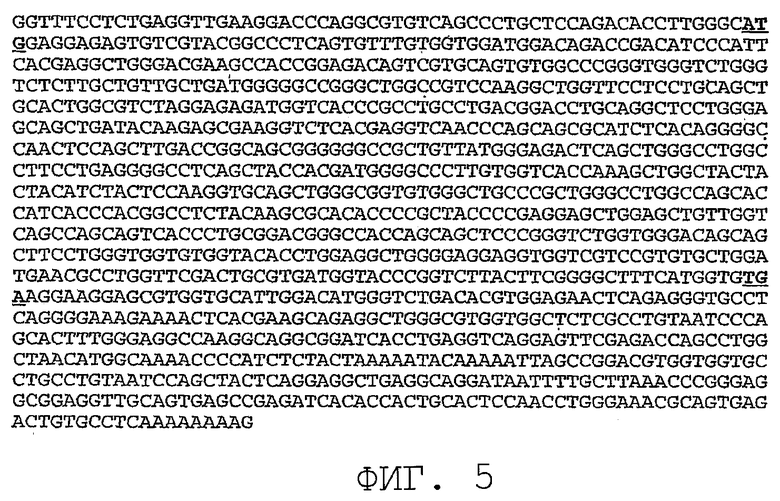

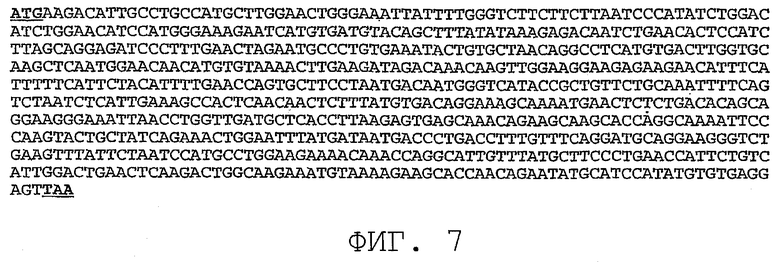











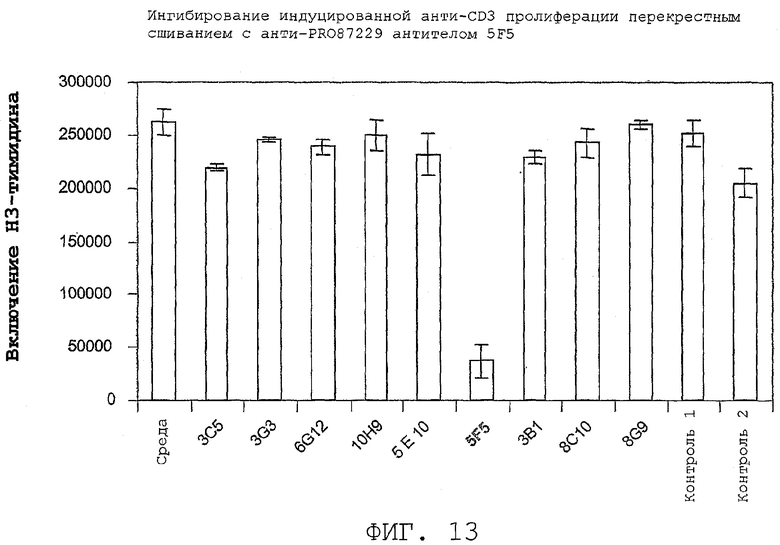
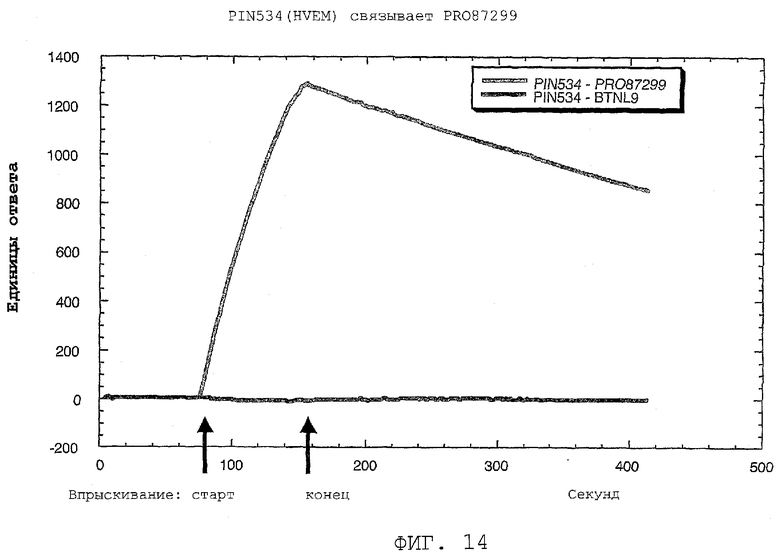
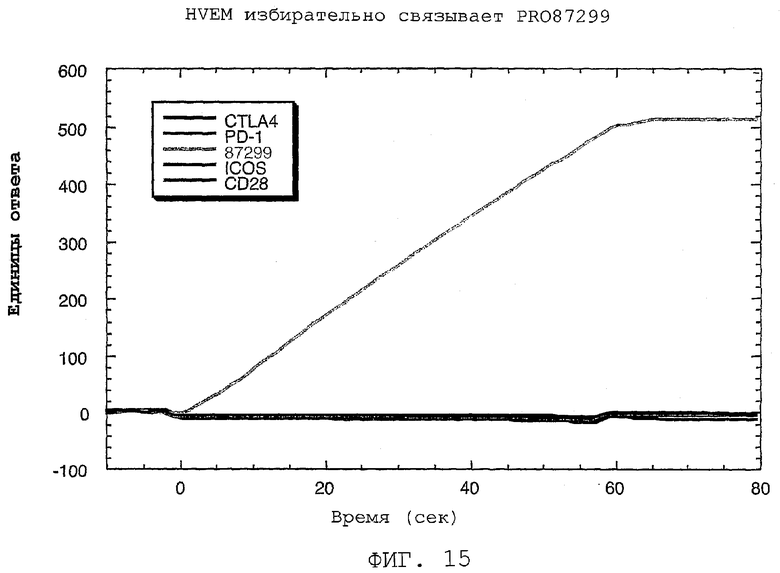

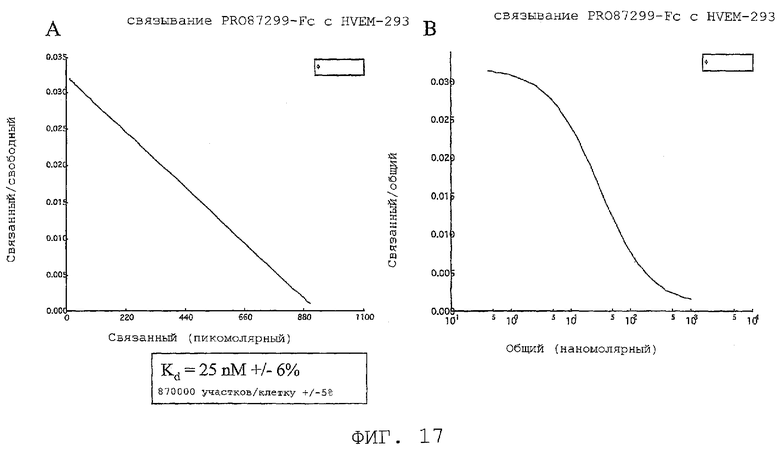
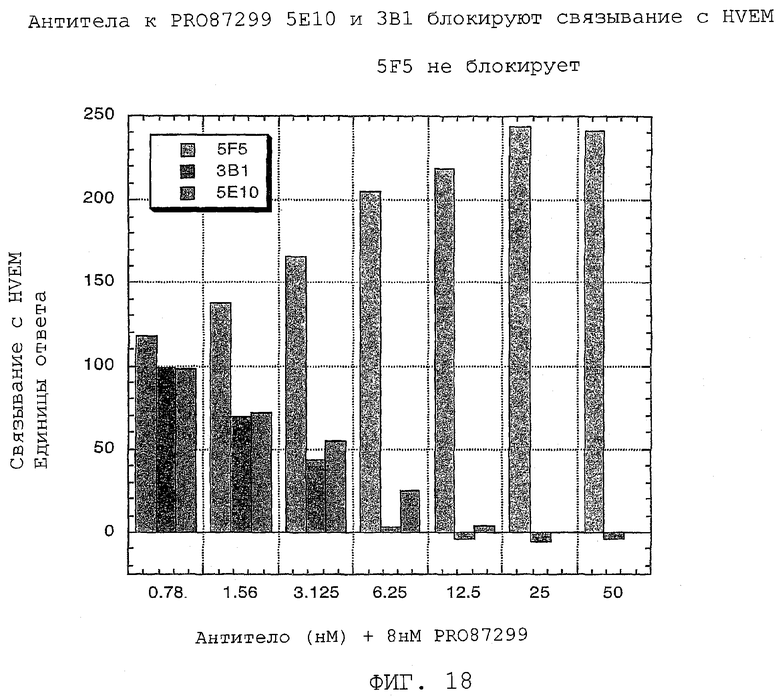
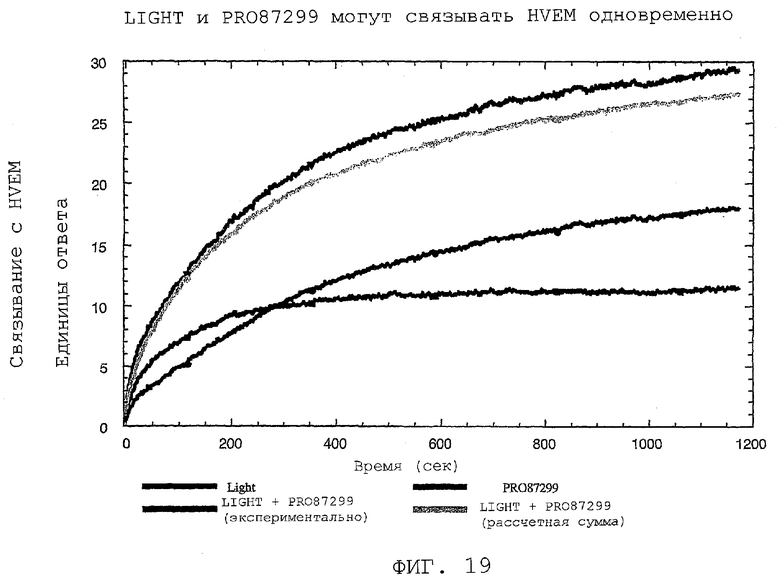

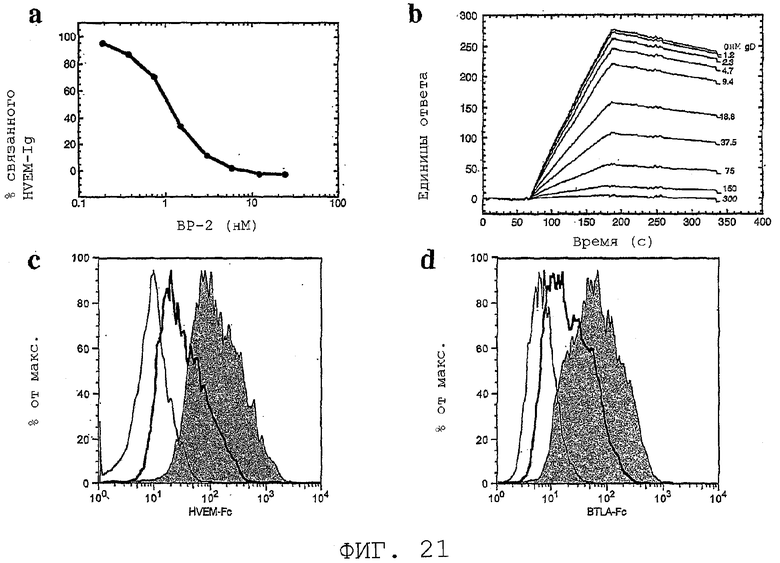

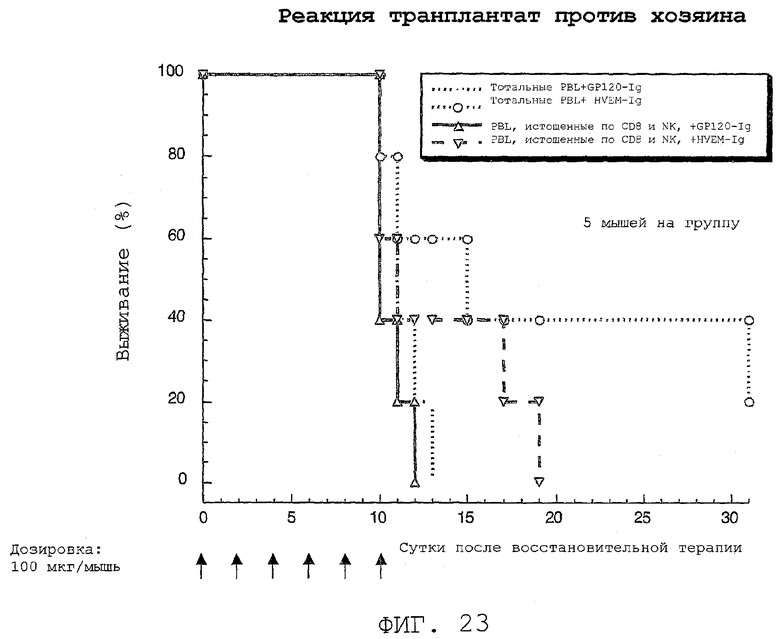
Изобретение относится к антителу, специфически связывающемуся с вариантом PRO87299. Антитело по изобретению дополнительно обладает свойством блокировать взаимодействие HVEM с PRO87299 и выполнять функцию агониста PRO87299. Антитело, проявляющее свойство агониста, продуцируется гибридомой Btig5F5.1 или Btig3B1.9. Для антитела установлена аминокислотная последовательность, приведенная в описании. Изобретение раскрывает способы использования антител для стимуляции или понижения иммунного ответа при заболеваниях, связанных с иммунитетом, облегчения лимфомы и воспалительного заболевания у нуждающегося в этом млекопитающего, определения присутствия полипептида PRO87299 в образце, ослабления отторжения трансплантированных клеток. Полученное по изобретению антитело является иммуномодулятором, что позволяет применять терапевтически одно и то же лекарственное средство как для усиления иммунного ответа, так и для его уменьшения. 9 н. и 7 з.п. ф-лы, 23 ил., 7 табл.
1. Антитело, специфически связывающееся с вариантом PRO87299, показанным на фиг.10 (SEQ ID NO:10).
2. Антитело, специфически связывающееся с PRO87299 и блокирующее взаимодействие HVEM с PRO87299.
3. Антитело, специфически связывающееся с PRO87299, где указанное антитело представляет собой агонист PRO87299.
4. Антитело по п.3, где указанное антитело подавляет пролиферацию СD4+Т-клетки.
5. Антитело по п.3, где указанное антитело подавляет пролиферацию клетки лимфомы.
6. Антитело по п.3, где указанное антитело подавляет пролиферацию NK-клетки.
7. Антитело по любому из пп.1-6, где указанное антитело представляет собой моноклональное антитело, гуманизированное антитело, химерное антитело, фрагмент антитела или одноцепочечное антитело.
8. Выделенное антитело по п.3, продуцируемое гибридомой Btig5F5.1 (инвентарный No в АТСС 6302).
9. Выделенное антитело по п.3, обладающее биологической активностью антитела, продуцируемого гибридомой Btig5F5.1 (инвентарный No в АТСС 6302), где биологическая активность представляет собой подавление пролиферации СD4+Т-клетки.
10. Способ стимуляции иммунного ответа у нуждающегося в этом млекопитающего, включающий в себя введение указанному млекопитающему терапевтически эффективного количества антитела по п.2.
11. Способ понижения иммунного ответа у нуждающегося в этом млекопитающего, включающий в себя введение указанному млекопитающему терапевтически эффективного количества антитела по п.3.
12. Способ по п.11, где животное страдает расстройством, выбранным из системной красной волчанки, ревматоидного артрита, остеоартрита, ювенильного хронического артрита, спондилоартропатии, системного склероза, идиопатической воспалительной миопатии, синдрома Шегрена, системного васкулита, саркоидоза, аутоиммунной гемолитической анемии, аутоиммунной тромбоцитопении, тиреоидита, сахарного диабета, иммуноопосредованного заболевания почек, демиелинизирующего заболевания центральной или периферической нервной системы, идиопатической демиелинизирующей полиневропатии, синдрома Гийена-Барре, хронической воспалительной демиелинизирующей полиневропатии, гепатобилиарного заболевания, инфекционного или аутоиммунного хронического активного гепатита, первичного биллиарного цирроза, гранулематозного гепатита, склерозирующего холангита, воспалительного заболевания кишечника, глютензависимой энтеропатии, болезни Уиппла, аутоиммунного или иммуноопосредованного заболевания кожи, буллезного заболевания кожи, полиморфной эритемы, контактного дерматита, псориаза, аллергического заболевания, астмы, аллергического ринита, атопического дерматита, гиперчувствительности к пище, уртикарии, иммунологического заболевания легкого, эозинофильных пневмоний, идиопатического фиброза легких, гиперчувствительного пневмонита, связанного с трансплантацией заболевания, отторжения трансплантата и реакции трансплантат против хозяина.
13. Способ определения присутствия полипептида PRO87299 в образце, предположительно содержащем указанный полипептид, где указанный способ включает в себя воздействие на указанный образец анти-РRO87299 антителом по п.1 и по наличию связывания указанного антитела с PRO87299 определяют присутствие в образце полипептида.
14. Способ облегчения лимфомы у нуждающегося в этом млекопитающего, включающий в себя введение указанному млекопитающему терапевтически эффективного количества анти-РRO87299-антитела по п.3, которое специфически связывается с полипептидом PRO87299.
15. Способ облегчения воспалительного заболевания кишечника у нуждающегося в этом млекопитающего, включающий в себя введение указанному млекопитающему терапевтически эффективного количества анти-РRO87299-антитела по п.3, которое специфически связывается с полипептидом PRO87299.
16. Способ ослабления отторжения трансплантированных клеток у млекопитающего, включающий в себя введение эффективного количества анти-РRO87299-антитела по п.3.
| US 2003215442 A1, 20.11.2003 | |||
| EP 1336619 A2, 20.08.2003 | |||
| AU 1566602 A1, 11.04.2002 | |||
| РОЙТ А | |||
| и др | |||
| Иммунология | |||
| - Мир, 2000, с.153-155. |

