Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые связываются с цитокиновым рецептором IL-22R, в частности, IL-22R человека. Анти-IL-22R-антитела и антигенсвязывающие фрагменты по изобретению обладают отличительными свойствами, в частности, отличительными комбинациями свойств, по сравнению с анти-IL-22R-антителами, описанными на предшествующем уровне техники.
Уровень техники
IL-22R (также известный как IL-22R1 и IL-22RA) является цитокиновым рецептором типа II, избирательно экспрессируемым в коже и эпителиальных клетках. Данный рецептор опосредует передачу сигналов через три цитокина: интерлейкин 22 (IL-22), интерлейкин 20 (IL-20) и интерлейкин 24 (IL-24). Для цитокинового сигнального пути через IL-22R требуется образования гетеродимерных комплексов на поверхности клетки. Как показано на фиг. 1, IL-22 связывается с и осуществляет передачу сигналов через комплекс, состоящий из IL-22R и IL-10Rβ (также известный как IL-10R2), тогда как IL-20 и IL-24 связываются с и осуществляют передачу сигналов через гетеродимерный комплекс, состоящий из IL-22R и IL-20Rβ (также известный как IL-20R2).
Интерлейкин-22 представляет цитокин, экспрессируемый иммунными клетками, в частности, активированными дендритными клетками и Т-клетками. После продуцирования клетками иммунной системой IL-22 проявляет свои биологические эффекты посредством связывания с IL-22R и активации IL-22R на эпителиальных клетках. Активация комплекса IL-22R-IL-10Rβ ниже связывания IL-22 приводит к провоспалительным ответным реакциям, индукции антимикробных белков, которые имеют ключевое значение для защиты хозяина от бактериальных патогенов, и защитных эффектов в некоторых органах, таких как легкие и печень. IL-22 также участвует в патологии заболеваний, в частности, в развитии воспалительных заболеваний, таких как псориаз, псориатический артрит и атопический дерматит (Ma et al., J. Clin. Invest., 118: 597-607 (2008), Van Belle et al., J. Immunol., Jan. 1, 188 (1): 462-9 (2012), Sabat et al., Nat. Rev. Drug Discov., 13 (1): 21-38 (2014)).
Была определена кристаллическая структура IL-22 в комплексе с внеклеточным доменом IL-22R, и она дала важную информацию о том, как этот лиганд связывается со своим рецептором (Jones et al., Structure, 16 (9): 1333-1344 (2008)). Внеклеточная область IL-22R включает два домена фибронектина типа III (FBNIII) (D1 и D2), ориентированные друг к другу примерно под прямыми углами. Пять петель, расположенных на границе раздела этих доменов, в первую очередь ответственны за вовлечение остатков IL-22 в комплекс лиганд-рецептор. Остатки IL-22, которые вносят вклад в связывание рецептора, кластеризуются в двух сайтах в лиганде, сайте 1а и сайте 1b. Знание ключевых остатков, обеспеченных как рецептором, так и лигандом, обеспечило способы, с помощью которых это взаимодействие может быть нарушено для отмены сигнального пути IL-22 для применения в качестве терапевтической стратегии.
Интерлейкин-20 и интерлейкин-24 экспрессируются моноцитами и кератиноцитами, и аналогично IL-22, было установлено, что эти цитокины играют роль в гомеостазе и развитии патологии кожи. С учетом этого следует, что стратегии ингибирования или подавления передачи сигналов в направлении 5'-3' IL-22R блокированием связывания лигандов, которые активируют этот рецептор, могут иметь терапевтическую применимость, в частности, в лечении кожных заболеваний, таких как псориаз и атопический дерматит.
Были разработаны антитела, которые связываются с IL-22R и блокируют взаимодействие между IL-22 и IL-22R. Например, в WO2011/061119 описывается гуманизированное анти-IL-22R-антитело, полученное из мышиного античеловеческого моноклонального антитела, первоначально описанного в WO2006/047249. Было показано, что это гуманизированное антитело, которое в настоящем описании будет относиться к «280-346-TSY», ингибирует сигнальный путь с участием IL-22 через IL-22RA в анализе пролиферации клеток и подавляет индуцированное IL-23 воспаление уха на мышиной модели псориаза.
Сущность изобретения
Настоящее изобретение улучшает уровень техники обеспечением антител или их антигенсвязывающих фрагментов, которые связываются с цитокиновым рецептором IL-22R и проявляют свойства, которые отличаются от свойств анти-IL-22R-антител, описанных на предшествующем уровне техники. Антитела или их антигенсвязывающие фрагменты, как правило, проявляют комбинации свойств, которые являются отличительными и в некоторых случаях превосходят свойства известных анти-IL-22R-антител, в частности, гуманизированного анти-IL-22R-антитела, описанного в WO 02011/01119. Свойства этих антител могут быть особенно преимущественными в отношении применения в лечении человека, в частности, для лечения таких патологических состояний, как псориаз, псориатический артрит и атопический дерматит.
В первом аспекте настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которые связываются с человеческим IL-22R, где антитело или его антигенсвязывающий фрагмент связываются с эпитопом в белке IL-22R, который не содержит Tyr60.
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты обладают одним или более дополнительными свойствами, выбранными из следующего:
(i) способности связываться с эпитопом IL-22R человека, расположенным, по меньшей мере, частично в домене D2 белка IL-22R;
(ii) способности связываться с IL-22R человека с высокой аффинностью;
(iii) способности блокировать связывание IL-22 с IL-22R человека;
(iv) способности ингибировать IL-22-зависимую активацию IL-22R;
(v) способности ингибировать IL-20-зависимую активацию IL-22R;
(vi) способности ингибировать IL-22- и IL-20-зависимую активацию IL-22R; и
(vii) отсутствия перекрестной реактивности с мышиным IL-22R.
Антитела или антигенсвязывающие фрагменты могут проявлять высокую гомологию с антителами человека, как в настоящем описании определено в других местах. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты включают вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где области VH и/или VL или один или более участков, определяющих комплементарность (CDR), происходят от животного семейства верблюдовых, то есть происходят от верблюда. Антитела или антигенсвязывающие фрагменты, обладающие высокой гомологией с человеком или имеющие, по меньшей мере, одну последовательность CDR, области VH и/или VL, полученную от верблюда, могут быть гуманизированными вариантами или вариантами зародышевого типа областей VH или VL из обычных верблюжьих антител, где термины «гуманизированный вариант» и «вариант зародышевого типа» определены в настоящем описании в других местах.
В неограничивающих вариантах осуществления изобретение обеспечивает следующие антитела или их антигенсвязывающие фрагменты, которые определяются посредством ссылки на конкретные структурные характеристики, то есть указанные аминокислотные последовательности CDR (одна или более последовательностей SEQ ID NO:2, 4, 6, 9, 11, 13, 34, 36, 41, 43 (CDR тяжелой цепи) или SEQ ID NO:16, 18, 20, 23, 25, 27, 47, 54, 57, 59 (CDR легкой цепи) или полных вариабельных областей (одна или более последовательностей SEQ ID NO:29, 31, 63, 65 (вариабельные области тяжелой цепи) или SEQ ID NO:30, 32, 62, 64, 66 (вариабельные области легкой цепи). Все эти антитела связываются с цитокиновым рецептором IL-22R человека.
В конкретных вариантах осуществления антитела, определенные следующими структурными характеристиками, могут проявлять высокую гомологию с антителами человека, как в настоящем описании определено. Антитела могут быть моноклональными антителами, полученными рекомбинантными способами. CDR следующих анти-IL-22R-антител могут быть получены от верблюда, то есть получены из обычных антител, продуцированных в ответ на иммунизацию верблюдов (в частности, ламы). Изобретение также обеспечивает гуманизированные варианты или варианты зародышевой линии человека, аффинные варианты и варианты, содержащие консервативные аминокислотные замены, как в настоящем описании определено.
Варианты осуществления анти-IL-22R-антител по изобретению теперь дополнительно описываются со ссылкой на структурные характеристики.
В одном варианте осуществления обеспечиваются антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R, где указанное антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи (VH), содержащую CDR3 тяжелой цепи, выбранный из:
SEQ ID NO:6 [VGFSGTYYSES] или ее варианта последовательности,
SEQ ID NO:13 [PPGPFKAHYNGMKY] или ее варианта последовательности,
SEQ ID NO:43 [PPGPFKAHYNGAKY] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Вариабельная область тяжелой цепи антитела или его антигенсвязывающего фрагмента может альтернативно или дополнительно содержать CDR2 тяжелой цепи, выбранный из:
SEQ ID NO:4 [SIYNDGSNTAYSDSVKG] или ее варианта последовательности,
SEQ ID NO:11 [GIHISGGITYYLDSVKG] или ее варианта последовательности,
SEQ ID NO:36 [SIYNDASNTAYSDSVKG] или ее варианта последовательности,
SEQ ID NO:41 [GIHISGGITYYTDSVKG] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Вариабельная область тяжелой цепи антитела или его антигенсвязывающего фрагмента может альтернативно или дополнительно содержать CDR1 тяжелой цепи, выбранный из:
SEQ ID NO:2 [SYDMS] или ее варианта последовательности,
SEQ ID NO:9 [SYFMS] или ее варианта последовательности,
SEQ ID NO:34 [SYDMN] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Альтернативно или дополнительно антитела или их антигенсвязывающие фрагменты, которые связываются с цитокиновым рецептором IL-22R, могут содержать вариабельную область легкой цепи (VL), содержащую CDR3 легкой цепи, выбранный из:
SEQ ID NO:20 [QSGSSSANAV] или ее варианта последовательности,
SEQ ID NO:27 [ASYRLYADYV] или ее варианта последовательности,
SEQ ID NO:54 [QSGSSSSNAV] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Вариабельная область легкой цепи антитела или его антигенсвязывающего фрагмента может альтернативно или дополнительно содержать CDR2 легкой цепи, выбранный из:
SEQ ID NO:18 [GNNNRPS] или ее варианта последовательности,
SEQ ID NO:25 [KVNTRSS] или ее варианта последовательности,
SEQ ID NO:47 [GQNNRPS] или ее варианта последовательности,
SEQ ID NO:59 [EVNKRSS] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Вариабельная область легкой цепи антитела или его антигенсвязывающего фрагмента может альтернативно или дополнительно содержать CDR1 легкой цепи, выбранный из:
SEQ ID NO:16 [QGGYYAH] или ее варианта последовательности,
SEQ ID NO:23 [TGTSRDIGDYNYVS] или ее варианта последовательности,
SEQ ID NO:57 [TGTSSDIGSYNYVS] или ее варианта последовательности,
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
В некоторых вариантах осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R, где антитело или его антигенсвязывающий фрагмент включают комбинацию CDR3 вариабельной области тяжелой цепи (HCDR3), CDR2 вариабельной области тяжелой цепи (HCDR2) и CDR1 вариабельной области тяжелой цепи (HCDR1), где комбинация выбрана из группы, включающей:
(i) HCDR3, содержащий SEQ ID NO:6; HCDR2, содержащий SEQ ID NO:36; HCDR1, содержащий SEQ ID NO:34;
(ii) HCDR3, содержащий SEQ ID NO:43; HCDR2, содержащий SEQ ID NO:41; HCDR1, содержащий SEQ ID NO:9;
(iii) HCDR3, содержащий SEQ ID NO:6; HCDR2, содержащий SEQ ID NO:4; HCDR1, содержащий SEQ ID NO:2; и
(iv) HCDR3, содержащий SEQ ID NO:13; HCDR2, содержащий SEQ ID NO:11; HCDR1, содержащий SEQ ID NO:9.
Альтернативно или дополнительно, антитела или их антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R, могут содержать комбинацию CDR3 (LCDR3) вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи (LCDR2) и CDR1 вариабельной области легкой цепи (LCDR1), выбранных из группы, включающей:
(i) LCDR3, содержащий SEQ ID NO:54; LCDR2, содержащий SEQ ID NO:47; LCDR1, содержащий SEQ ID NO:16;
(ii) LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:59; LCDR1, содержащий SEQ ID NO:57;
(iii) LCDR3, содержащий SEQ ID NO:20; LCDR2, содержащий SEQ ID NO:47; LCDR1, содержащий SEQ ID NO:16;
(iv) LCDR3, содержащий SEQ ID NO:20; LCDR2, содержащий SEQ ID NO:18; LCDR1, содержащий SEQ ID NO:16; и
(v) LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:25; LCDR1, содержащий SEQ ID NO:23.
В некоторых вариантах осуществления обеспечиваются антитела или их антигенсвязывающие фрагменты, которые связываются с цитокиновым рецептором IL-22R, где антитела или антигенсвязывающие фрагменты содержат комбинацию CDR3 (HCDR3) вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи (HCDR2) и CDR1 вариабельной области тяжелой цепи (HCDR1), CDR3 вариабельной области легкой цепи (LCDR3), CDR2 вариабельной области легкой цепи (LCDR2) и CDR1 легкой цепи легкой цепи (LCDR1) в соответствии с вариантами осуществления, описанными ниже.
В одном варианте осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R и содержат комбинацию последовательностей CDR VH и VL: HCDR3, содержащий SEQ ID NO:6; HCDR2, содержащий SEQ ID NO:36; HCDR1, содержащий SEQ ID NO:34; LCDR3, содержащий SEQ ID NO:54; LCDR2, содержащий SEQ ID NO:47; и LCDR1, содержащий SEQ ID NO:16.
В одном варианте осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R и содержат комбинацию последовательностей CDR VH и VL: HCDR3, содержащий SEQ ID NO:43; HCDR2, содержащий SEQ ID NO:41; HCDR1, содержащий SEQ ID NO:9; LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:59; и LCDR1, содержащий SEQ ID NO:57.
В одном варианте осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R и содержат комбинацию последовательностей CDR VH и VL: HCDR3, содержащий SEQ ID NO:6; HCDR2, содержащий SEQ ID NO:4; HCDR1, содержащий SEQ ID NO:2; LCDR3, содержащий SEQ ID NO:20; LCDR2, содержащий SEQ ID NO:47; и LCDR1, содержащий SEQ ID NO:16.
В одном варианте осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R и содержат комбинацию последовательностей CDR VH и VL: HCDR3, содержащий SEQ ID NO:6; HCDR2, содержащий SEQ ID NO:4; HCDR1, содержащий SEQ ID NO:2; LCDR3, содержащий SEQ ID NO:20; LCDR2, содержащий SEQ ID NO:18; и LCDR1, содержащий SEQ ID NO:16.
В одном варианте осуществления обеспечивается антитело или его антигенсвязывающий фрагмент, которые связываются с цитокиновым рецептором IL-22R и содержат комбинацию последовательностей CDR VH и VL: HCDR3, содержащий SEQ ID NO:13; HCDR2, содержащий SEQ ID NO:11; HCDR1, содержащий SEQ ID NO:9; LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:25; и LCDR1, содержащий SEQ ID NO:23.
В некоторых вариантах осуществления обеспечиваются антитела или их антигенсвязывающие фрагменты, которые связываются с цитокиновым рецептором IL-22R, где антитела или антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи (VH), выбранную из следующего:
(i) VH, содержащей или состоящей из аминокислотной последовательности SEQ ID NO:29 или 31,
(ii) аффинного варианта или варианта зародышевой линии человека VH, содержащей или состоящей из аминокислотной последовательности SEQ ID NO:29 или 31; или
(iii) VH, содержащей или состоящей из аминокислотной последовательности, обладающей, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, 99% идентичностью с аминокислотной последовательностью SEQ ID NO:29 или 31.
Альтернативно или дополнительно, антитела или антигенсвязывающие фрагменты могут содержать вариабельную область легкой цепи (VL), выбранную из следующего:
(i) VL, содержащей или состоящей из аминокислотной последовательности SEQ ID NO:30, 32 или 62,
(ii) аффинного варианта или варианта зародышевой линии человека VH, содержащей или состоящей из аминокислотной последовательности SEQ ID NO:30, 32 или 62; или
(iii) VL, содержащей или состоящей из аминокислотной последовательности, обладающей, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, 99% идентичностью с аминокислотной последовательностью SEQ ID NO:30, 32 или 62.
Для вариантов осуществления, где области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью последовательности с эталонной последовательностью, области VH и/или VL могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только в каркасных областях.
В некоторых вариантах осуществления антитела по изобретению могут включать домен CH1, шарнирную область, домен CH2 и домен CH3 человеческого антитела, в частности, человеческого IgG1, IgG2, IgG3 или IgG4.
Особенно предпочтительные антитела по настоящему изобретению описаны ниже.
230С9 и антитела, связанные с ним
В некоторых вариантах осуществления обеспечивается выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с IL-22R, где указанное антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, где:
последовательность CDR3 вариабельной области тяжелой цепи представляет SEQ ID NO:6 [VGFSGTYYSES] или ее вариант последовательности;
последовательность CDR2 вариабельной области тяжелой цепи представляет SEQ ID NO:36 [SIYNDASNTAYSDSVKG] или ее вариант последовательности; и
последовательность CDR1 вариабельной области тяжелой цепи представляет SEQ ID NO:34 [SYDMN] или ее вариант последовательности, и
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Антитело или антигенсвязывающий фрагмент может дополнительно содержать вариабельную область легкой цепи, где:
последовательность CDR3 вариабельной области легкой цепи представляет SEQ ID NO:54 [QSGSSSSNAV] или ее вариант последовательности;
последовательность CDR2 вариабельной области легкой цепи представляет SEQ ID NO:47 [GQNNRPS] или ее вариант последовательности; и
последовательность CDR1 вариабельной области легкой цепи представляет SEQ ID NO:16 [QGGYYAH] или ее вариант последовательности, и
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
В некоторых вариантах осуществления обеспечивается выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с IL-22R, где указанное антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, где:
последовательность CDR3 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:6 [VGFSGTYYSES];
последовательность CDR2 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:36 [SIYNDASNTAYSDSVKG];
последовательность CDR1 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:34 [SYDMN];
последовательность CDR3 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:54 [QSGSSSSNAV];
последовательность CDR2 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:47 [GQNNRPS]; и
последовательность CDR1 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:16 [QGGYYAH].
Антитела или их антигенсвязывающие фрагменты могут включать вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO:63 и необязательно вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO:64. В некоторых вариантах осуществления обеспечиваются моноклональные антитела или их антигенсвязывающие фрагменты, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность VH, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности, с аминокислотной последовательностью, показанной в SEQ ID NO:63, и/или вариабельная область легкой цепи содержит VL, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности, с аминокислотной последовательностью, показанной в SEQ ID NO:64. Для вариантов осуществления, где области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью с эталонной последовательностью, области VH и/или VL могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только в каркасных областях. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, содержащие вариабельные области тяжелой цепи и/или вариабельные области легкой цепи, определенные как имеющие конкретную процентную идентичность с SEQ ID NO:63 и 64 соответственно, будут иметь следующие последовательности CDR:
последовательность CDR3 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:6 [VGFSGTYYSES];
последовательность CDR2 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:36 [SIYNDASNTAYSDSVKG];
последовательность CDR1 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:34 [SYDMN];
последовательность CDR3 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:54 [QSGSSSSNAV];
последовательность CDR2 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:47 [GQNNRPS]; и
последовательность CDR1 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:16 [QGGYYAH].
Антитела, которые специфически связываются с IL-22R, могут содержать, по меньшей мере, одну полноразмерную тяжелую цепь иммуноглобулина и/или, по меньшей мере, одну полноразмерную легкую цепь лямбда или каппа. В некоторых вариантах осуществления антитела включают тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:67, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:68. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности, с аминокислотной последовательностью, показанной в SEQ ID NO:67, и/или легкую цепь, по меньшей мере, с 85% идентичностью или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности, с аминокислотной последовательностью, показанной в SEQ ID NO:68. Для вариантов осуществления, где цепи антител определяются конкретной процентной идентичностью последовательности с эталонной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только вне областей CDR.
223G5 и антитела, связанные с ним
В некоторых вариантах осуществления обеспечивается выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с IL-22R, где указанное антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, где:
последовательность CDR3 вариабельной области тяжелой цепи представляет SEQ ID NO:43 [PPGPFKAHYNGAKY] или ее вариант последовательности;
последовательность CDR2 вариабельной области тяжелой цепи представляет SEQ ID NO:41 [GIHISGGITYYTDSVKG] или ее вариант последовательности; и
последовательность CDR1 вариабельной области тяжелой цепи представляет SEQ ID NO:9 [SYFMS] или ее вариант последовательности, и
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
Антитело или антигенсвязывающий фрагмент может дополнительно содержать вариабельную область легкой цепи, где:
последовательность CDR3 вариабельной области легкой цепи представляет SEQ ID NO:27 [ASYRLYADYV] или ее вариант последовательности;
последовательность CDR2 вариабельной области легкой цепи представляет SEQ ID NO:59 [EVNKRSS] или ее вариант последовательности; и
последовательность CDR1 вариабельной области легкой цепи представляет SEQ ID NO:57 [TGTSSDIGSYNYVS] или ее вариант последовательности, и
где вариант последовательности включает одну, две или три аминокислотные замены (например, консервативные замены, гуманизирующие замены или аффинные варианты) в указанной последовательности.
В некоторых вариантах осуществления обеспечивается выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с IL-22R, где указанное антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, где:
последовательность CDR3 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:43 [PPGPFKAHYNGAKY];
последовательность CDR2 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:41 [GIHISGGITYYTDSVKG];
последовательность CDR1 вариабельной области тяжелой цепи содержит или состоит из SEQ ID NO:9 [SYFMS];
последовательность CDR3 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:27 [ASYRLYADYV];
последовательность CDR2 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:59 [EVNKRSS]; и
последовательность CDR1 вариабельной области легкой цепи содержит или состоит из SEQ ID NO:57 [TGTSSDIGSYNYVS].
Антитела или их антигенсвязывающие фрагменты могут включать вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO:65 и необязательно вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO:66. В некоторых вариантах осуществления обеспечиваются моноклональные антитела или их антигенсвязывающие фрагменты, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность VH по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:65, и/или вариабельная область легкой цепи содержит VL, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:66. Для вариантов осуществления, где области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью с эталонной последовательностью, области VH и/или VL могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только в каркасных областях. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, содержащие вариабельные области тяжелой цепи и/или вариабельные области легкой цепи, определенные как имеющие конкретную процентную идентичность с SEQ ID NO:65 и 66 соответственно, будут иметь следующие последовательности CDR:
последовательность CDR3 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:43 [PPGPFKAHYNGAKY];
последовательность CDR2 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:41 [GIHISGGITYYTDSVKG];
последовательность CDR1 вариабельной области тяжелой цепи, содержащую или состоящую из SEQ ID NO:9 [SYFMS];
последовательность CDR3 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:27 [ASYRLYADYV];
последовательность CDR2 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:59 [EVNKRSS]; и
последовательность CDR1 вариабельной области легкой цепи, содержащую или состоящую из SEQ ID NO:57 [TGTSSDIGSYNYVS].
Антитела, которые специфически связываются с IL-22R, могут содержать, по меньшей мере, одну полноразмерную тяжелую цепь иммуноглобулина и/или, по меньшей мере, одну полноразмерную легкую цепь лямбда или каппа. В некоторых вариантах осуществления антитела включают тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:69, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:70. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:69, и/или легкую цепь, по меньшей мере, с 85% идентичностью или, по меньшей мере, с 90% идентичностью последовательности или, по меньшей мере, с 95% идентичностью последовательности или, по меньшей мере, с 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:70. Для вариантов осуществления, где цепи антител определяются конкретной процентной идентичностью последовательности с эталонной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только вне областей CDR.
В тех случаях, когда конкретные антитела или антигенсвязывающие участки определены как содержащие комбинацию области VH или тяжелой цепи, определенных посредством ссылки на конкретную аминокислотную последовательность, и области VL или легкой цепи, также определенных посредством ссылки на конкретную аминокислотную последовательность, то для каждой конкретной комбинации VH/VL или тяжелой цепи/легкой цепи (если не указано иное), данное определение может быть принято с включением антител или антигенсвязывающих участков, образованных комбинацией области VH/тяжелой цепи, имеющей, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97% или, по меньшей мере, 99% идентичность последовательности с указанной аминокислотной последовательностью VH/тяжелой цепи, и области VL/легкой цепи, имеющей, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97% или, по меньшей мере, 99% идентичность последовательности с указанной аминокислотной последовательностью VL/легкой цепи. В каждом случае области/цепи, определенные процентной идентичностью последовательности с указанными аминокислотными последовательностями области/цепи, могут сохранять идентичные последовательности CDR с теми, которые присутствуют в указанных аминокислотных последовательностях области VH/VL или тяжелой/легкой цепи, одновременно проявляя вариацию аминокислотной последовательности в каркасных областях или других областях вне участков CDR.
Если не указано иное в настоящей заявке, то процентную идентичность последовательности между двумя аминокислотными последовательностями можно определить сравнением этих двух последовательностей, выравненных оптимальным образом и в которых аминокислотная последовательность, подлежащая сравнению, может содержать добавления или делеции относительно эталонной последовательности для оптимального выравнивания между этими двумя последовательностями. Процент идентичности вычисляется определением числа идентичных положений, для которых аминокислотный остаток идентичен между двумя последовательностями, делением этого числа идентичных положений на общее число положений в окне сравнения и умножением полученного результата на 100, с получением процента идентичности между этими двумя последовательностями. Например, можно использовать программу BLAST, «BLAST 2 sequences» (Tatusova and al., «Blast 2 sequences - a new tool for comparing protein and nucleotide sequences», FEMS Microbiol Lett. 174:247-250), доступной на сайте http://www.ncbi.nlm.nih.gov/ gorf/bl2.html, с параметрами по умолчанию (в частности, для параметров «штраф за создание пробела»: 5 и «штраф за удлинение пробела»: 2; где выбранной матрицей является, например, матрица «BLOSUM 62», предложенная программой), процент идентичности между двумя последовательностями, которые требуется сравнить, вычисляется непосредственно программой.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты, обеспеченные в настоящем описании, могут иметь одну или любую комбинацию следующих свойств/признаков:
антитело или антигенсвязывающий фрагмент могут связываться с эпитопом в человеческом белке IL-22R, который не содержит Tyr60;
антитело или антигенсвязывающий фрагмент могут связываться с эпитопом, расположенным, по меньшей мере, частично, в домене D2 белка IL-22R человека, где домен D2 представляет аминокислотные остатки с 125 по 228 последовательности SEQ ID NO:71;
антитело или антигенсвязывающий фрагмент могут связываться с IL-22R человека с высокой аффинностью;
антитело или антигенсвязывающий фрагмент могут блокировать связывание IL-22 с IL-22R;
антитело или антигенсвязывающий фрагмент могут ингибировать IL-22-зависимую активацию IL-22R;
антитело или антигенсвязывающий фрагмент могут ингибировать IL-20-зависимую активацию IL-22R;
антитело или антигенсвязывающий фрагмент могут ингибировать IL-22-зависимую активацию IL-22R и IL-20-зависимую активацию IL-22R;
антитело или антигенсвязывающий фрагмент не могут перекрестно реагировать с мышиным IL-22R.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты, обеспеченные в настоящем описании, предпочтительно проявляют два или более из следующих свойств/признаков:
антитело или антигенсвязывающий фрагмент связываются с эпитопом в человеческом белке IL-22R, который не содержит Tyr60;
антитело или антигенсвязывающий фрагмент связываются с IL-22R человека с высокой аффинностью;
антитело или антигенсвязывающий фрагмент ингибируют IL-22-зависимую активацию IL-22R и IL-20-зависимую активацию IL-22R.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты, обеспеченные в настоящем описании, могут быть химерными антителами. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты содержат шарнирную область, домен СН2 и/или домен СН3 человеческого IgG. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты проявляют высокую гомологию с человеческим IgG, предпочтительно человеческим IgG1, где «высокая гомология с иммуноглобулином человека» определяется в настоящем описании в других местах. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где области VH и/или VL или один или более участков, определяющих комплементарность (CDR), получены от животного семейства верблюдовых, то есть получены от верблюда, где предпочтительно верблюд является ламой.
В дополнительных аспектах изобретение также обеспечивает молекулы полинуклеотидов, которые кодируют вышеуказанные антитела и антигенсвязывающие фрагменты, в дополнение к экспрессионным векторам, содержащим полинуклеотиды, клетки-хозяева, содержащие векторы и способы рекомбинантной экспрессии/продукции антител, описанных в настоящем описании.
В еще одном аспекте изобретение обеспечивает фармацевтическую композицию, содержащую любое из анти-IL-22R-антител или антигенсвязывающих фрагментов, описанных в настоящем описании, и фармацевтически приемлемый носитель или эксципиент.
Еще один дополнительный аспект изобретения относится к способам терапевтического лечения с использованием вышеуказанных анти-IL-22R-антител или их антигенсвязывающих фрагментов, в частности, в профилактике и/или лечении таких патологических состояний, как псориаз, псориатический артрит и атопический дерматит.
Эти и другие варианты осуществления изобретения можно лучше оценить и понять при рассмотрении в сочетании со следующим описанием и прилагаемыми фигурами. Следует, однако, понимать, что нижеследующее описание, несмотря на указание различных вариантов осуществления изобретения и многочисленных их конкретных деталей, приводится в качестве иллюстрации, а не для ограничения. Многие замены, модификации, добавления и/или перегруппировки могут быть сделаны в рамках объема изобретения, не отступая от его сущности, и изобретение включает все такие замены, модификации, добавления и/или перегруппировки.
Краткое описание фигур
На фиг. 1 показаны различные рецепторные комплексы, которые опосредуют передачу сигналов через цитокины IL-22, IL-20 и IL-24. IL-22R способен образовывать гетеродимерный комплекс с двумя различными партнерами рецептора, IL-10Rβ (или IL-10R2) и IL-20Rβ (или IL-20R2), и активация этих различных комплексов связыванием лиганда запускает сигнальный путь через внутриклеточные пути в 5'-3' направлени.
На фиг.2 показана полноразмерная аминокислотная последовательность IL-22R человека (SEQ ID NO:71).
На фиг.3 показана полноразмерная нуклеотидная последовательность, кодирующая IL-22R человека (SEQ ID NO:72).
На фиг.4 показано ингибирование IL-22- и IL-20-опосредованного сигнального пути через IL-22R в анализах клеточной пролиферации. (A) Показано влияние mAb к IL-22R на пролиферацию клеточной линии BW-hIL-22R. Клетки BW-hIL-22R стабильно экспрессируют IL-22R человека, и рост ингибируется/останавливается в ответ на лиганд IL-22. Антитела, способные блокировать взаимодействие между IL-22 и hIL-22R, облегчают ингибирование роста, опосредованное связыванием лиганд-рецептор. (B) Показано влияние mAb к IL-22R на пролиферацию клеточной линии Baf3-hIL-22R/IL20Rb. Клетки Baf3-hIL-22R/IL20Rb стабильно экспрессируют компоненты рецепторного комплекса IL-22R/IL20Rb, так что клетки пролиферируют в присутствии IL-20. Антитела, способные блокировать взаимодействие между IL-20 и этим рецепторным комплексом, ингибируют пролиферацию, индуцированную связыванием лиганда.
На фиг. 5 показано выравнивание внеклеточного домена IL-22R из различных видов. (A) показаны неполные последовательности EST, доступные из Genbank; (B) показаны последовательности, определенные после клонирования IL-22R обезьяны циномолгус и макака-резус из библиотеки кДНК обезьяны циномолгус.
На фиг. 6 показаны результаты опытов с использованием конкурентного ELISA, проведенных для картирования эпитопов IL-22R mAb. Были идентифицированы различные эпитопы для антител VH-семейств 1-8, 10, 11, 19 и 22. Эпитопы были сгруппированы в зависимости от того, насколько антитела (i) блокировали связывание IL-22 in vitro и нейтрализовали сигнальный путь IL-22 в клеточном анализе (нижний левый круг); (ii) блокировали связывание IL-22 in vitro, но не обладали нейтрализующей активностью в клеточном анализе (верхний правый круг); или (iii) не блокировали связывание IL-22 in vitro, но проявляли нейтрализующую активность в клеточном анализе (нижний правый круг).
На фиг. 7 показана кристаллическая структура IL-22R в комплексе с IL-22. (A) Домены D1 и D2 IL-22R вносят остатки в поверхность раздела с лигандом IL-22. (B) Y60 в домене D1 является важным остатком IL-22R, способствующим взаимодействию с сайтом 1A лиганда IL-22.
На фиг.8 показано ингибирование IL-20-опосредованного сигнального пути через IL-22R в анализе клеточной пролиферации. Различные mAb к IL-22R тестировали на их способность ингибировать IL-20-индуцированную пролиферацию клеток Baf3-hIL-22R/IL20Rb.
На фиг.9 показано ингибирование IL-22- и IL-20-опосредованного сигнального пути через IL-22R в клеточных анализах. (A) Показано влияние mAb к IL-22R на пролиферацию клеточной линии BW-hIL-22R. Клетки BW-hIL-22R стабильно экспрессируют IL-22R человека, и рост ингибируется/останавливается в ответ на лиганд IL-22. Антитела, способные блокировать взаимодействие между IL-22 и hIL-22R, облегчают ингибирование роста, опосредованное связыванием лиганд-рецептор. (B) Показано влияние mAb к IL-22R на пролиферацию клеточной линии Baf3-hIL-22R/IL20Rb. Клетки Baf3-hIL-22R/IL20Rb стабильно экспрессируют компоненты рецепторного комплекса IL-22R/IL20Rb, так что клетки пролиферируют в присутствии IL-20. Антитела, способные блокировать взаимодействие между IL-20 и этим рецепторным комплексом, ингибируют пролиферацию, индуцированную связыванием лиганда.
На фиг. 10 схематически показаны результаты экспериментов по картированию эпитопов для вариантов зародышевого типа антител IL-22R 230C9 и 223G5. NB «Zymo» эквивалентно 280-346-TSY».
На фиг. 11 показана перекрестная реактивность вариантов зародышевого типа антител к IL-22R человека и обезьяны циномолгус, как определено анализом FACS. Антитело 230C9 перекрестно реагирует с IL-22R человека и IL-22R обезьяны циномолгус (левые панели), в то время как антитело 223G5 связывается с человеческим IL-22R, но перекрестно не реагирует с IL-22R обезьяны циномолгус (правые панели).
На фиг.12 показаны фармакокинетические данные для анти-IL-22R-антитела 230C9. Обезьянам циномолгус вводили внутривенно однократно антитело в дозе 10 мг/кг. Образцы крови отбирали на разные временные точки и определяли концентрацию антитела в плазме с помощью ELISA. Было установлено, что антитело 230C9 имеет период полураспада, равный примерно 19,4 суток.
На фиг.13 показаны фармакокинетические данные для анти-IL-22R-антитела 230C9, которое вводили в различных дозах. В более высоких дозах (≥10 мг/кг), когда мишень-опосредованное распределение препарата (TMDD) насыщается, то значения клиренса приближаются к неспецифическому клиренсу RES. B. Клиренс 230C9-N297Q у обезьяны циномолгус. Общий клиренс представляет сумму 1) TMDD, которое является нелинейной и насыщаемой, и 2) неспецифического клиренса, который является линейным и относится к RES. Период полураспада в плазме имеет обратную связь с клиренсом, что приводит к длительному периоду полураспада в высоких дозах и более короткому периоду полураспада в более низких дозах за счет мишень-опосредованного клиренса.
На фиг.14 показан фармакодинамический эффект анти-IL-22R-антитела 230C9. Обезьяну циномолгус подвергали воздействию антитела 230С9 в разных дозах и оценивали действие на обработанный IMQ участок кожи и нормальный участок кожи обезьяны. Было установлено, что повышенные дозы антитела 230C9 нормализует толщину эпидермиса (A) и уменьшает частоту Ki67-положительных ядер (B) в обработанном IMQ участке кожи.
На фиг. 15 показано влияние анти-IL-22R-антитела 230C9 (ARGX-112) на ILR-22-регулируемые уровни мРНК FLG2 в образце кожи, полученном пункционной биопсией. Обезьяне циномолгус вводили однократно внутривенно антитело 230C9 в различных дозах: 1 мг/кг, 5 мг/кг и 30 мг/кг (по 3 животных на каждую дозу). Рекомбинантный человеческий IL-22 снижал уровни общей мРНК FLG2 кожи, тогда как данный эффект отменялся антителом 230C9. На оси Y указана относительная экспрессия FLG2 по сравнению с референсным геном. Статистическое сравнение между группами обозначено линиями в верхней части графика с доверительными интервалами следующим образом: *p <0,05; **р <0,01; ***р <0,001. Показанные проценты (60%, 64%, 60% соответственно) указывают на % ингибирования сигнала.
На фиг.16 показано влияние анти-IL-22R-антитела 230C9 (ARGX-112) на IL-22-регулируемую экспрессию гена DEFB4 в эксплантах кожи человека ex vivo. Кожные экспланты с области живота обрабатывали возрастающими концентрациями антитела 230C9 перед стимуляцией 20 мл/мл rhIL-22. Антитело 230С9 было способно отменять IL-22-опосредованное повышение уровней мРНК DEFB4 дозозависимым образом. На оси Y указана относительная экспрессия DEFB4 по сравнению с референсным геном. LLOQ означает более низкий уровень количественного определения.
На фиг. 17 показано влияние анти-IL-22R-антитела 230C9 (ARGX-112) на IL-22-регулируемую экспрессию генов FLG2 и LOR в эксплантах кожи обезьяны циномолгус ex vivo. Биопсийные образцы кожи обрабатывали возрастающими концентрациями антитела 230C9 перед стимуляцией 20 мл/мл rhIL-22. Антитело 230С9 было способно отменять IL-22-опосредованное снижение уровней мРНК FLG2 (A) и мРНК LOR (B) дозозависимым образом. На оси Y указана относительная экспрессия FLG2 и LOR по сравнению с референсным геном.
На фиг. 18 показано влияние анти-IL-22R-антитела 230C9 (ARGX-112) на первичные кератиноциты человека. Кератиноциты предварительно обрабатывали антителом 230C9, и затем стимулировали смесью IL-4, IL-13, IL-22 и IFN-γ. Контрольные клетки обрабатывали смесью IL-4, IL-13 и IFN-γ. Антитело 230C9 показало дозозависимое ингибирование секреции CCL2.
Подробное описание изобретения
А. Определения
«Антитело» или «иммуноглобулин». Как в настоящем описании используется, термин «иммуноглобулин» включает полипептид, имеющий комбинацию двух тяжелых и двух легких цепей, независимо от наличия или отсутствия какой-либо соответствующей специфической иммунореактивности. Термин «антитела» относится к таким молекулам, которые имеют значительную известную специфическую иммунологическую реактивность к представляющему интерес антигену (например, цитокиновому рецептору IL-22R). Термин «анти-IL-22R-антитела» используется в настоящем описании для обозначения антител, которые проявляют иммунологическую специфичность к белку IL-22R, включая IL-22R человека, и в некоторых случаях его видовым гомологам. Антитела и иммуноглобулины содержат легкие и тяжелые цепи, с или без межцепочечной ковалентной связи между ними. Основные структуры иммуноглобулина у позвоночных относительно хорошо изучены.
Общий термин «иммуноглобулин» включает пять различных классов антител, которые можно различить биохимически. Все пять классов антител входят в объем настоящего изобретения. Последующее обсуждение, в основном, будет относиться к классу IgG молекул иммуноглобулинов. В отношение IgG, то иммуноглобулины содержат два идентичных полипептида легкой цепи с молекулярной массой примерно 23000 дальтон и два идентичных полипептида тяжелой цепи с молекулярной массой 53000-70000 дальтон. Четыре цепи соединены дисульфидными связями в конфигурации «Y», где легкие цепи обхватывают тяжелые цепи, начинающиеся у раздвоенного конца «Y» и проходящие через вариабельную область.
Легкие цепи антитела классифицируются как каппа или лямбда (κ, λ). Каждый класс тяжелой цепи может быть связан с легкой цепью либо каппа, либо лямбда. В общем, легкая и тяжелая цепи ковалентно связаны друг с другом, и «хвостовые» участки двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями, когда иммуноглобулины продуцируются гибридомами, B-клетками или клетками-хозяевами, полученными методами генной инженерии. В тяжелой цепи аминокислотные последовательности простираются от N-конца в раздвоенных концах конфигурации «Y» до С-конца в нижней части каждой цепи. Специалистам в данной области техники должно быть понятно, что тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε) с наличием некоторых подклассов среди них (например, γ1-γ4). Природа данной цепи определяет «класс» антитела как IgG, IgM, IgA, IgD или IgE соответственно. Подклассы иммуноглобулинов (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и тому подобное, хорошо охарактеризованы и, как известно, определяют функциональную специализацию. Модифицированные варианты каждого из этих классов и изотипов может легко дифференцировать квалифицированный специалист в данной области с учетом настоящего раскрытия и, соответственно, входят в объем настоящего изобретения.
Как указано выше, вариабельная область антитела обеспечивает возможность антителу избирательно распознавать и специфически связываться с эпитопами антигенов. То есть, область VL и область VH антитела объединяются с образованием вариабельной области, которая определяет трехмерный антигенсвязывающий сайт. Эта четвертичная структура антитела образует антигенсвязывающий сайт, присутствующий на конце каждого плеча Y. Более конкретно, антигенсвязывающий сайт антигена определяется тремя участками, определяющими комплементарность (CDR), в каждой из цепей VH и VL.
«IL-22R». Как в настоящем описании используется, термин «IL-22R» означает цитокиновый рецептор типа II, который опосредует сигнальный путь через лиганды IL-22, IL-20 и IL-24. IL-22R способен образовывать гетеродимерные комплексы на клеточной поверхности с IL-10R2 и IL-20R2. IL-22R также может относиться к IL22R, IL-22R1, IL22R1, IL22RA, IL-22RA, CRF2-9 и Zcytor 11. Термин IL-22R1 является достаточно широким, чтобы охватить человеческую форму рецептора и видовые гомологи. Аминокислотная последовательность полноразмерного человеческого IL-22R представлена в SEQ ID NO:71, и кодирующая нуклеотидная последовательность представлена в SEQ ID NO:72 (см. фиг. 2 и 3). Данные последовательности соответствуют последовательностям, депонированным в базе данных SwissProt, как субъединица рецептора интерлейкина-22 человека, инвентарный номер Q8N6P7.
«Сайт связывания». Как в настоящем описании используется, термин «сайт связывания» включает область полипептида, которая ответственна за избирательное связывание с антигеном-мишенью, представляющим интерес (например, IL-22R). Связующие домены содержат, по меньшей мере, один сайт связывания. Иллюстративные связывающие домены включают вариабельную область антитела. Молекулы антител по изобретению могут содержать один сайт связывания или несколько (например, два, три или четыре) сайтов связывания.
«Полученные из». Как в настоящем описании используется, термин «полученные из» обозначенного белка (например, верблюжье антитело или его антигенсвязывающий фрагмент) относится к источнику полипептида или аминокислотной последовательности. В одном варианте осуществления полипептид или аминокислотная последовательность, которые получены из конкретного исходного полипептида, представляет собой последовательность CDR или связанную с ним последовательность. В одном варианте осуществления аминокислотная последовательность, которая получена из конкретного исходного полипептида, не является смежной. Например, в одном варианте осуществления один, два, три, четыре, пять или шесть CDR получены из исходного антитела. В одном варианте осуществления полипептид или аминокислотная последовательность, которые получены из конкретного исходного полипептида или аминокислотной последовательности, имеет аминокислотную последовательность, которая по существу идентична последовательности исходной последовательности или ее фрагмента, где этот фрагмент состоит, по меньшей мере, из 3-5 аминокислот, по меньшей мере, 5-10 аминокислот, по меньшей мере, 10-20 аминокислот, по меньшей мере, 20-30 аминокислот или, по меньшей мере, 30-50 аминокислот, или которые иначе могут быть идентифицированы специалистом с обычной квалификацией в данной области как имеющее начало в исходной последовательности. В одном варианте осуществления одна или более последовательностей CDR, полученных из исходного антитела, изменены с получением вариантных последовательностей CDR, например, варианты аффинности, где вариантные последовательности CDR поддерживают активность антиген-связывающей активности.
«Полученные от верблюда». В некоторых предпочтительных вариантах осуществления антитела по изобретению содержат аминокислотные последовательности каркасной области и/или аминокислотные последовательности CDR, полученные из обычного верблюжьего антитела, продуцированного активной иммунизацией верблюда. Однако антитела по изобретению, содержащие аминокислотные последовательности, полученные от верблюда, могут быть сконструированы таким образом, чтобы содержать последовательности каркасных и/или константных областей, полученные из аминокислотной последовательности человека (то есть человеческого антитела) или других видов млекопитающих, отличных от верблюда. Например, каркасная область, участок тяжелой цепи и/или шарнирная часть человека или примата, отличного от человека, могут быть включены в конкретные анти-IL-22R-антитела. В одном варианте осуществления одна или более аминокислот, не происходящих от верблюда, могут присутствовать в каркасной области антитела, полученного от верблюда, например, аминокислотная последовательность каркасной области верблюда может содержать одну или более аминокислотных мутаций, в которых может присутствовать соответствующий аминокислотный остаток человека и примата, отличного от человека. Кроме того, области VH и VL, полученные от верблюда, или их гуманизированные варианты могут быть связаны с константными областями человеческих антител с образованием химерной молекулы, как в настоящем описании описано в других местах.
«Консервативная аминокислотная замена» - «консервативная аминокислотная замена» представляет замену, при которой аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области, включающие основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, заменимый аминокислотный остаток в полипептиде иммуноглобулина может быть заменен другим аминокислотным остатком из того же семейства боковых цепей. В еще одном варианте осуществления цепь аминокислот может быть заменена структурно сходной цепью, которая отличается по порядку и/или составу членов семейства боковых цепей.
«Часть тяжелой цепи». Как в настоящем описании используется, термин «часть тяжелой цепи» включает аминокислотные последовательности, полученные из константных областей тяжелой цепи иммуноглобулина. Полипептид, содержащий часть тяжелой цепи, содержит, по меньшей мере, один из следующих элементов: домен CH1, шарнирная область (например, верхняя, средняя и/или нижняя шарнирная область), домен CH2, домен CH3 или его вариант или его фрагмент. В одном варианте осуществления антитело или антигенсвязывающий фрагмент по изобретению могут содержать Fc-фрагмент тяжелой цепи иммуноглобулина (например, шарнирную область, домен CH2 и домен CH3). В еще одном варианте осуществления в антителе или антигенсвязывающем фрагменте по изобретению может отсутствовать, по меньшей мере, часть константной области (например, полностью или часть домена CH2). В некоторых вариантах осуществления, по меньшей мере, одна, и предпочтительно все константные области получены из тяжелой цепи иммуноглобулина человека. Например, в одном предпочтительном варианте осуществления часть тяжелой цепи содержит полностью человеческую шарнирную область. В еще одних предпочтительных вариантах осуществления часть тяжелой цепи содержит полностью человеческий Fc-фрагмент (например, последовательности шарнирной области, доменов CH2- и CH3- из иммуноглобулина человека).
В некоторых вариантах осуществления составные компоненты части тяжелой цепи константные области происходят из разных молекул иммуноглобулина. Например, часть тяжелой цепи полипептида может содержать домен CH2, полученный из молекулы IgG1, и шарнирную область, полученную из молекулы IgG3 или IgG4. В еще одних вариантах осуществления константные области являются химерными областями, содержащими участки различных молекул иммуноглобулина. Например, шарнирная область может содержать первый участок из молекулы IgG1 и второй участок из молекулы IgG3 или IgG4. Как указано выше, специалисту в данной области техники должно быть понятно, что константные области части тяжелой цепи могут быть модифицированы таким образом, что они отличаются от аминокислотной последовательности из молекулы иммуноглобулина, встречающегося в природе (дикого типа). То есть, полипептиды по изобретению, раскрытые в настоящем описании, могут содержать изменения или модификации одной или более константных областей тяжелой цепи (CH1, шарнирной области, CH2 или CH3) и/или домена константной области легкой цепи (CL). Иллюстративные модификации включают добавления, делеции или замены одной или более аминокислот в одном или нескольких доменах.
«Химерный». «Химерный» белок содержит первую аминокислотную последовательность, связанную со второй аминокислотной последовательностью, с которой она не связана в природных условиях. Аминокислотные последовательности обычно могут существовать в отдельных белках, которые объединяются вместе в слитый полипептид, или они могут обычно существовать в одном и том же белке, но помещаются в новое расположение в слитом полипептиде. Химерный белок можно получить, например, химическим синтезом или созданием и трансляцией полинуклеотида, в котором пептидные области кодированы в желаемой взаимосвязи. Типичные химерные антитела по изобретению включают слитые белки, содержащие области VH и VL, полученные от верблюда, или их гуманизированные варианты, слитые с константными областями человеческого антитела, например, человеческого IgG1, IgG2, IgG3 или IgG4.
«Вариабельная область» или «вариабельный домен». Термины «вариабельная область» и «вариабельный домен» используются в настоящем описании взаимозаменяемо и имеют эквивалентное значение. Термин «вариабельная» относится к тому факту, что некоторые участки вариабельных областей VH и VL сильно различаются по последовательности среди антител и выполняют определенную роль в связывании и определении специфичности каждого конкретного антитела для его антигена-мишени. Однако вариабельность неравномерно распределена по вариабельным областям антител. Она сконцентрирована в трех сегментах, называемых «гипервариабельными петлями» в каждой области VL и области VH, которые образуют часть антигенсвязывающего сайта. Первая, вторая и третья гипервариабельные петли области VLлямбда легкой цепи относятся в настоящем описании к L1 (λ), L2 (λ) и L3 (λ) и могут быть определены как содержащие остатки 24-33 (L1 (λ), состоящий из 9, 10 или 11 аминокислотных остатков), 49-53 (L2 (λ), состоящий из 3 остатков) и 90-96 (L3 (λ), состоящий из 5 остатков) в области VL (Morea et al., Methods, 20: 267-279 (2000)). Первая, вторая и третья гипервариабельные петли области Vкаппа легкой цепи относятся в настоящем описании к L1 (κ), L2 (κ) и L3 (κ) и могут быть определены как содержащие остатки 25-33 (L1 (κ), состоящий из 6, 7, 8, 11, 12 или 13 остатков), 49-53 (L2 (κ), состоящий из 3 остатков) и 90-97 (L3 (κ), состоящий из 6 остатков) в области VL (Мора et al., Methods, 20: 267-279 (2000)). Первая, вторая и третья гипервариабельные петли области VH относятся в настоящем описании к H1, H2 и H3 и могут быть определены как содержащие остатки 25-33 (H1, состоящий из 7, 8 или 9 остатков), 52-56 (H2, состоящий из 3 или 4 остатков) и 91-105 (H3, высоко вариабельный по длине) в области VH (Morea et al., Methods 20: 267-279 (2000)).
Если не указано иное, то термины L1, L2 и L3 соответственно относятся к первой, второй и третьей гипервариабельным петлям области VL и охватывают гипервариабельные петли, полученные как из изотипа Vкаппа, так и изотипа Vлямбда. Термины H1, H2 и H3 соответственно относятся к первой, второй и третьей гипервариабельным петлям области VH и включают гипервариабельные петли, полученные из любого известного изотипа тяжелой цепи, включая γ, ε, δ, α или μ.
Гипервариабельные петли L1, L2, L3, H1, H2 и H3 каждая может включать часть «участка, определяющего комплементарность» или «CDR», как определено ниже. Термины «гипервариабельная петля» и «участок, определяющий комплементарность», строго говоря, не являются синонимами, поскольку гипервариабельные петли (HV) определяются на основе структуры, тогда как участки, определяющие комплементарность (CDR), определяются на основе вариабельности последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD., 1983), и пределы HV и CDR могут быть разными в некоторых областях VH и VL.
CDR областей VL и VH обычно могут быть определены как содержащие следующие аминокислоты: остатки 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в вариабельной области легкой цепи и остатки 31-35 или 31-35b (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в вариабельной области тяжелой цепи (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Таким образом, HV могут находиться в соответствующих CDR, и ссылки в настоящем описании на «гипервариабельные петли» областей VH и VL следует интерпретировать как также охватывающие соответствующие CDR, и наоборот, если не указано иное.
Более высококонсервативные участки вариабельных областей называются каркасной областью (FR), как определено ниже. Вариабельные области нативных тяжелых и легких цепей содержат четыре FR (FR1, FR2, FR3 и FR4 соответственно), в основном принимая β-складчатую конфигурацию, связанную тремя гипервариабельными петлями. Гипервариабельные петли в каждой цепи удерживаются вместе в непосредственной близости с помощью FR и вместе с гипервариабельными петлями из другой цепи способствуют образованию антигенсвязывающего сайта антител. Структурный анализ антител выявил взаимосвязь между последовательностью и формой сайта связывания, образованного участками, определяющими комплементарность (Chothia et al., J. Mol. Biol., 227: 799-817 (1992)); Tramontano et al., J. Mol. Biol, 215: 175-182 (1990)). Несмотря на их высокую вариабельность последовательности, пять из шести петель принимают только небольшой репертуар конформаций основной цепи, называемый «каноническими структурами». Данные конформации в первую очередь определяются длиной петель, и во-вторых, наличием ключевых остатков в определенных положениях в петлях и в каркасных областях, которые определяют конформацию посредством их упаковки, водородных связей или способности принимать необычные конформации основной цепи.
«CDR». Как в настоящем описании используется, термин «CDR» или «участок, определяющий комплементарность» означает несмежные антигенсвязывающие сайты, находящиеся в вариабельной области полипептидов как тяжелой, так и легкой цепи. Эти конкретные области были описаны Kabat et al., J. Biol. Chem., 252, 6609-6616 (1977) and Kabat et al., Sequences of protein of immunological interest. (1991) и Chothia et al., J. Mol. Biol., 196:901-917 (1987) и MacCallum et al., J. Mol. Biol., 262:732-745 (1996), где определения включают перекрывающиеся или подгруппы аминокислотных остатков при сравнении друг с другом. Аминокислотные остатки, образующие CDR, определенные в каждой из приведенных выше ссылок, приведены для сравнения. Предпочтительно, термин «CDR» представляет CDR, как определено Kabat, на основе сравнения последовательностей.
Определения CDR
2 Нумерация остатков соответствует номенклатуре, приведенной в публикации Chothia et al., выше
3 Нумерация остатков соответствует номенклатуре, приведенной в публикации MacCallum et al., выше
«Каркасная область». Как в настоящем описании используется, термин «каркасная область» или «область FR» включает аминокислотные остатки, которые являются частью вариабельной области, но не являются частью CDR (например, с использованием системы определения CDR по Kabat). Следовательно, каркасная область вариабельной области составляет от 100 до 100 аминокислот в длину, но включает только те аминокислоты, которые находятся за пределами CDR. Для конкретного примера вариабельной области тяжелой цепи и для CDR, согласно системе нумерации по Kabat et al., каркасная область 1 соответствует области вариабельной области, охватывающей аминокислоты 1-30; каркасная область 2 соответствует области вариабельной области, охватывающей аминокислоты 36-49; каркасная область 3 соответствует области вариабельной области, охватывающей аминокислоты 66-94, и каркасная область 4 соответствует области вариабельной области от аминокислот 103 до конца вариабельной области. Каркасные области легкой цепи аналогично отделены друг от друга каждым CDR вариабельной области легкой цепи. Аналогичным образом, используя систему определения CDR по Chothia et al. или McCallum et al. границы каркасной области отделены концами соответствующих CDR, как описано выше. В предпочтительных вариантах осуществления CDR являются такими, как определено Kabat.
Во встречающихся в природе антителах шесть CDR, присутствующих в каждом мономерном антителе, представляют собой короткие, несмежные последовательности аминокислот, которые специфически располагаются с образованием антигенсвязывающего сайта, когда антитело принимает свою трехмерную конфигурацию в водной среде. Остальная часть вариабельных областей тяжелых и легких цепей проявляет меньшую межмолекулярную вариабельность в аминокислотной последовательности и называется каркасными областями. Каркасные области в значительной степени принимают β-складчатую конформацию, и CDR образуют петли, которые соединяют и в некоторых случаях составляют часть β-складчатой структуры. Таким образом, эти каркасные области функционируют с образованием каркаса, который обеспечивает расположение шести CDR в правильной ориентации за счет межцепочечных, нековалентных взаимодействий. Антигенсвязывающий сайт, образованный расположенными CDR, определяет поверхность, комплементарную эпитопу на иммунореактивном антигене. Эта комплементарная поверхность обеспечивает нековалентное связывание антитела с эпитопом иммунореактивного антигена. Положение CDR может быть легко идентифицировано специалистом в данной области техники.
«Шарнирная область». Как в настоящем описании используется, термин «шарнирная область» включает часть молекулы тяжелой цепи, которая соединяет домен CH1 с доменом CH2. Эта шарнирная область содержит примерно 25 остатков и является гибкой, что позволяет двум N-концевым антигенсвязывающим участками перемещаться независимо друг от друга. Шарнирную область можно разделить на три различных области: верхнюю, среднюю и нижнюю шарнирные области (Roux K.H. et al., J. Immunol., 161: 4083-90, 1998). Антитела по изобретению, содержащие «полностью человеческую» шарнирную область, могут содержать одну из последовательностей шарнирной области, показанных в таблице 2 ниже.
Последовательности шарнирной области человека
(SEQ ID NO:82)
(SEQ ID NO:83)
(SEQ ID NO:84)
(SEQ ID NO:85)
(SEQ ID NO:86)
(SEQ ID NO:87)
(SEQ ID NO:88)
(SEQ ID NO:89)
(SEQ ID NO:90)
(SEQ ID NO:91)
(SEQ ID NO:92)
(SEQ ID NO:93)
«Домен CH2». Как в настоящем описании используется, термин «домен CH2» включает часть молекулы тяжелой цепи, которая простирается, например, от остатка 244 до остатка 360 антитела с использованием обычных систем нумерации (остатки 244-360, система нумерации по Kabat и остатки 231-340, система нумерации ЕС, Kabat E.A. et al. Sequences of Proteins of Immunological Interest. Bethesda, US Department of Health and Human Services, NIH. 1991). Домен CH2 уникален тем, что он тесно не спарен с другим доменом. Точнее, две N-связанные разветвленные углеводные цепи находятся между двумя доменами CH2 молекулы интактного нативного IgG. Также хорошо известно, что домен CH3 простирается от домена CH2 до C-конца молекулы IgG и содержит примерно 108 остатков.
«Фрагмент». Как в настоящем описании используется, термин «фрагмент» в контексте антител по изобретению, относится к части или участку антитела или цепи антитела, содержащей меньшее число аминокислотных остатков, чем интактное или полное антитело или цепь антитела. Термин «антигенсвязывающий фрагмент» относится к полипептидному фрагменту иммуноглобулина или антитела, который связывает антиген или конкурирует с интактным антителом (то есть с интактным антителом, из которого он был получен) за связывание антигена (т.е. специфическое связывание с IL-22R). Как в настоящем описании используется, термин «фрагмент» молекулы антитела включает антигенсвязывающие участки антител, например, вариабельную область легкой цепи антитела (VL), вариабельную область тяжелой цепи антитела (VH), одноцепочечное антитело (scFv), F(ab')2-фрагмент, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент и однодоменный фрагмент антитела (DAb). Фрагменты могут быть получены, например, посредством химической или ферментативной обработки интактного или полного антитела или цепи антитела или рекомбинантными средствами.
«Валентность». Как в настоящем описании используется, термин «валентность» относится к числу потенциальных сайтов связывания мишеней в полипептиде. Каждый сайт связывания мишени специфически связывает одну молекулу-мишень или специфический сайт на молекуле-мишени. Когда полипептид содержит более одного сайта связывания мишени, то каждый сайт связывания мишени может специфически связываться с одними и теми же или разными молекулами (например, может связываться с различными лигандами или различными антигенами или различными эпитопами на одном и том же антигене).
«Специфичность». Термин «специфичность» относится к способности связываться (например, иммунологически реагировать с) с данной мишенью, например, IL-22R. Полипептид может быть моноспецифическим и содержать один или более сайтов связывания, которые специфически связываются с мишенью или полипептидом, может быть мультиспецифическим и содержать два или более сайта связывания, которые специфически связываются с одной и той же или разными мишенями.
«Синтетический». Как в настоящем описании используется, термин «синтетический» в отношении полипептидов включает полипептиды, которые содержат аминокислотную последовательность, которая не встречается в природе. Например, не встречающиеся в природе полипептиды представляют модифицированные формы встречающихся в природе полипептидов (например, включающие мутацию, такую как добавление, замена или делеция) или которые содержат первую аминокислотную последовательность (которая может быть природной или неприродной), которая связана в линейной последовательности аминокислот со второй аминокислотной последовательностью (которая может быть природной или неприродной), с которой она не связана в природе.
«Сконструированные». Как в настоящем описании используется, термин «сконструированные» включает манипулирование молекулами нуклеиновой кислоты или полипептида с помощью синтетических методов (например, с помощью рекомбинантных методов, пептидного синтеза in vitro, путем ферментативного или химического сочетания пептидов или определенной комбинации этих методов). Предпочтительно, антитела по изобретению являются сконструированными, включая, например, гуманизированные и/или химерные антитела, и антитела, которые были сконструированы для улучшения одного или более свойств, таких как связывание антигена, стабильность/ период полураспада или эффекторная функция.
«Модифицированное антитело». Как в настоящем описании используется, термин «модифицированное антитело» включает синтетические формы антител, которые изменены таким образом, что они не являются встречающимися в природе, например, антитела, которые содержат, по меньшей мере, части двух тяжелых цепей, но не две полные тяжелые цепи (например, такие как антитела с делецированным доменом или миниантитела); мультиспецифические формы антител (например, биспецифические, триспецифические и тому подобное), модифицированные с целью обеспечения связывания с двумя или более разными антигенами или с различными эпитопами на одном антигене; молекулы тяжелой цепи, присоединенные к молекулам scFv и тому подобное. scFv известны в данной области и описаны, например, в патенте США № 5892019. Кроме того, термин «модифицированное антитело» включает мультивалентные формы антител (например, трехвалентные, четырехвалентные и тому подобное, антитела, которые связываются с тремя или более копиями одного и того же антигена). В еще одном варианте осуществления модифицированное антитело по изобретению представляет слитый белок, содержащий, по меньшей мере, часть одной тяжелой цепи, без домена CH2, и содержащий связывающий домен полипептида, включающий связывающий участок одного члена пары рецептор-лиганд.
Термин «модифицированное антитело» также может быть использован в настоящем описании для обозначения вариантов аминокислотных последовательностей антител по изобретению, как это в настоящем описании определено. Специалисту в данной области должно быть понятно, что антитело может быть модифицировано с получением варианта антитела, которое изменено по аминокислотной последовательности по сравнению с антителом, из которого оно было получено. Например, могут быть сделаны нуклеотидные или аминокислотные замены, приводящие к консервативным заменам или изменениям в «заменимых» аминокислотных остатках (например, в остатках CDR и/или каркасной области). Аминокислотные замены могут включать замену одной или более аминокислот природной или неприродной аминокислотой.
«Гуманизирующие замены». Как в настоящем описании используется, термин «гуманизирующие замены» относится к аминокислотным заменам, при которых аминокислотный остаток, присутствующий в определенном положении в области VH или VL антитела (например, антитела к IL-22R, полученного из верблюжьего антитела) заменяют аминокислотным остатком, который имеет место в эквивалентном положении в эталонной человеческой области VH или VL. Эталонная область VH или VL человека может представлять область VH или VL, кодируемая зародышевой линией человека. Гуманизирующие замены могут быть сделаны в каркасных областях и/или CDR антител, описанных в настоящем описании.
«Гуманизированные варианты». Как в настоящем описании используется, термин «гуманизированный вариант» относится к варианту антитела, которое содержит одну или более «гуманизирующих замен» по сравнению с эталонным антителом, где участок эталонного антитела (например, область VH и/или VL или их участки, содержащие, по меньшей мере, один CDR) содержит аминокислоту, полученную из «нечеловеческого» вида, и «гуманизирующие замены» имеют место в аминокислотной последовательности, полученной из «нечеловеческого» вида.
«Варианты зародышевого типа». Как в настоящем описании используется, термин «вариант зародышевого типа» конкретно относится к «гуманизированным вариантам», в которых «гуманизирующие замены» приводят к замене одного или более аминокислотных остатков, присутствующих в определенном положении(ях) в области VH или VL антитела (например, анти-IL-22R-антитела, полученного от верблюда) на аминокислотный остаток, который имеет место в эквивалентном положении в эталонной человеческой области VH или VL, кодированной зародышевой линией человека. Как правило, для любого конкретного «варианта зародышевого типа» аминокислотные остатки, введенные путем замены в вариант зародышевого типа, берутся исключительно или преимущественно из одной области VH или VL, кодированной геном зародышевой линии человека. Термины «гуманизированный вариант» и «вариант зародышевого типа» часто используются в настоящем описании взаимозаменяемо. Введение одной или более «гуманизирующих замен» в полученные от верблюда (например, от ламы) области VH или VL приводит к получению «гуманизированного варианта» области VH или VL верблюда (ламы). Если аминокислотные остатки, введенные путем замены, происходят преимущественно или исключительно из последовательности одной области VH или VL, кодированной зародышевой линией человека, то результатом может быть «вариант зародышевой линии человека» области VH или VL, происходящей от верблюда (ламы).
«Аффинные варианты». Как в настоящем описании используется, термин «аффинный вариант» относится к варианту антитела, которое имеет одно или более изменений в аминокислотной последовательности по сравнению с эталонным антителом, где аффинный вариант проявляет измененную аффинность к антигену-мишени в сравнении с эталонным антителом. Например, аффинные варианты проявляют измененную аффинность к IL-22R по сравнению с эталонным анти-IL-22R-антителом. Предпочтительно аффинный вариант будет демонстрировать повышенную аффинность к антигену-мишени, например, IL-22R, по сравнению с эталонным антителом. Аффинные варианты обычно проявляют одно или более изменений в аминокислотной последовательности в CDR по сравнению с эталонным антителом. Такие замены могут приводить к замене исходной аминокислоты, присутствующей в данном положении в CDR, на другой аминокислотный остаток, который может представлять собой остаток природной аминокислоты или остаток неприродной аминокислоты. Аминокислотные замены могут быть консервативными или неконсервативными.
«Высокая гомология с человеком». Антитело, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), будет считаться обладающим высокой гомологией с антителом человека, если области VH и VL, взятые вместе, проявляют, по меньшей мере, 90% идентичность аминокислотной последовательности с ближайшими совпадающими последовательностями VH и VL зародышевой линии человека. Антитела, обладающие высокой гомологией с человеком, могут включать антитела, содержащие области VH и VL нативных «нечеловеческих» антител, которые обладают достаточно высокой степенью идентичности с последовательностями человеческой зародышевой линии, включая, например, антитела, содержащие области VH и VL обычных верблюжьих антител, а также сконструированные, в частности, гуманизированные варианты или варианты зародышевого типа таких антител, а также «полностью человеческие» антитела.
В одном варианте осуществления область VH антитела с высокой гомологией с человеком может иметь идентичность аминокислотной последовательности или гомологию последовательности на уровне 80% или более с одной или более областями VH человека в каркасных областях FR1, FR2, FR3 и FR4. В еще одних вариантах идентичность аминокислотной последовательности или гомология последовательности между областью VH полипептида по изобретению и ближайшей совпадающей последовательностью области VH зародышевой линии человека может составлять 85% или более, 90% или более, 95% или более, 97% или более или до 99% или даже 100%.
В одном варианте осуществления область VH антитела с высокой гомологией с человеком может включать одно или более (например, от 1 до 10) несовпадений аминокислотных последовательностей каркасных областей FR1, FR2, FR3 и FR4, по сравнению с ближайшей совпадающей последовательностью человеческой VH.
В еще одном варианте осуществления область VL антитела с высокой гомологией с человеком может иметь идентичность последовательности или гомологию последовательности на уровне 80% или более с одной или более областями VL человека в каркасных областях FR1, FR2, FR3 и FR4. В еще одних вариантах осуществления идентичность аминокислотной последовательности или гомология последовательности между областью VL полипептида по изобретению и ближайшей совпадающей последовательностью области VL зародышевой линии человека может составлять 85% или более, 90% или более, 95% или более, 97% или более, или до 99% или даже 100%.
В одном варианте осуществления область VL антитела с высокой гомологией с человеком может включать одно или более (например, от 1 до 10) несовпадений аминокислотных последовательностей каркасных областей FR1, FR2, FR3 и FR4, по сравнению с ближайшей совпадающей последовательностью человеческой VL.
Перед анализом процентной идентичности последовательностей антитела, обладающего высокой гомологией с человеком, и областей VH и VL зародышевой линии человека, можно определить канонические складки, позволяющие идентифицировать семейство сегментов зародышевой линии клеток человека, и идентичную комбинацию канонических складок для Н1 и Н2 или L1 и L2 (и L3). Затем для оценки гомологии последовательностей выбирают член семейства зародышевой линии человека, характеризующийся самой высокой степенью гомологии последовательности с вариабельной областью представляющего интерес антитела. Канонические классы гипервариабельных петель L1, L2, L3, H1 и H2 по классификации Chortia можно определить при помощи прикладных биоинформационных программ, которые являются общедоступными на web-странице www.bioinf.org.uk/abs/chothia.html.page. Требования, предъявляемые к ключевым остаткам, приведены в файле данных программы. В указанных файлах данных приведены положения ключевых остатков с возможными аминокислотами в каждом положении. Последовательность вариабельной области представляющего интерес антитела представлена в виде входных данных, и сначала она сравнивается с консенсусной последовательностью антитела для нумерации по системе Kabat. При выполнении анализа канонических складок используется набор матриц ключевых остатков, полученных при помощи автоматизированного метода, разработанного Martin and Thornton (Martin et al., J. Mol. Biol. 263:800-815 (1996)).
При наличии известного V-сегмента зародышевой линии человека, в котором использована такая же комбинация канонических складок для Н1 и Н2 или L1 и L2 (и L3), можно определить наиболее совпадающий член семейства с точки зрения гомологии последовательности. Прикладные биоинформационные программы позволяют определить процентную идентичность аминокислотных последовательностей доменов VH и VL каркасных областей представляющего интерес антитела и соответствующих последовательностей, кодированных зародышевой линией человека, но фактически выравнивание последовательностей также может быть выполнено вручную. Последовательности иммуноглобулинов человека можно идентифицировать в нескольких базах данных белков, таких как VBase (http://vbase.mrc-cpe.cam.ac.uk/) или база данных Pluckthun/Honegger (http://www.bioc.unizh.ch/antibody/Sequences/Germlines). Для сравнения человеческих последовательностей с V-областями доменов VH или VL в представляющем интерес антителе может быть использован алгоритм сравнения последовательностей, представленный на web-сайтах, таких как www.expasy.ch/tools/#aligh, но выравнивание с ограниченным набором последовательностей также может быть выполнено вручную. Выбирают последовательности легких и тяжелых цепей зародышевой линии человека из семейств с одинаковыми комбинациями канонических складок и с самой высокой степенью гомологии с каркасными областями 1, 2 и 3 каждой цепи и сравнивают с вариабельной областью последовательности, представляющей интерес; также сравнивают FR4 с JH- и JK- или JL-областями зародышевой линии человека.
Следует отметить, что при вычислении общей процентной гомологии последовательности остатки FR1, FR2 и FR3 оценивают, используя наиболее совпадающую последовательность из семейства зародышевой линии человека с идентичной комбинацией канонических складок. Оценивают только остатки, отличающиеся от наиболее совпадаюих или других членов семейства с такой же комбинацией канонических складок (примечание: исключая любые различия, кодированные праймером). Однако в целях гуманизации остатки в каркасных областях, идентичные членам других семейств зародышевой линии человека, не содержащие одинаковой комбинации канонических складок, могут считаться «человеческими» несмотря на «негативную» оценку в соответствии с вышеописанными строгими условиями. Данное допущение основано на методе гуманизации «mix and match», в соответствии с которым каждую FR1, FR1, FR3 и FR4 отдельно сравнивают с наиболее совпадающей последовательностью зародышевой линии человека, следовательно, гуманизированная молекула содержит комбинацию разных FR областей, как было описано в публикациях Qu и соавторов (Qu et al., Clin. Cancer Res., 5:3095-3100 (1999) и Ono и соавторов (Ono et al., Mol. Immunol. 36:387-395 (1999). Границы отдельных каркасных областей могут быть определены согласно системе нумерации IMGT, которая является адаптированной системой нумерации Chortia (Lefranc et al., NAR 27:209-212 (1999); http://imgt.cines.fr).
Антитела, обладающие высокой гомологией с человеком, могут включать гипервариабельные петли или CDR, содержащие человеческие или «человекоподобные» канонические складки, подробно рассмотренные ниже.
В одном варианте осуществления изобретения, по меньшей мере, одна гипервариабельная петля или CDR в области VH или области VL антитела с высокой гомологией с человеком, может быть получена или выделена из области VH или VL антитела, отличного от человеческого, например, обычного антитела вида семейства верблюдовых, и все ее обладает прогнозируемой или фактической канонической складчатой структурой, которая по существу идентична канонической складчатой структуре в человеческих антителах.
В данной области хорошо известно, что, хотя основные аминокислотные последовательности гипервариабельных петель в областях VH и областях VL, кодированных зародышевой линией человека, по определению являются высоковариабельными, все гипервариабельные петли, за исключением CDR Н3 области VH, образуют только несколько четко выраженных структурных конформаций, именуемых каноническими складками (Chothia et al., J. Mol. Biol. 196:901-917 (1987); Tramontano et al., Proteins, 6:382-94 (1989)), которые зависят от длины гипервариабельной петли и наличия так называемых канонических аминокислотных остатков (Chothia et al., J. Mol. Biol. 196:901-917 (1987)). Фактические канонические структуры гипервариабельных петель в интактных областях VH или VL можно определить с использованием структурного анализа (например, рентгеновской кристаллографией), но также можно прогнозировать каноническую структуру с учетом ключевых аминокислотных остатков, характерных для определенной структуры (более подробно рассмотренных ниже). По существу специфический паттерн остатков, определяющих каждую каноническую структуру, образует «сигнатуру», которая позволяет распознавать каноническую структуру в гипервариабельных петлях области VH или VL с неизвестной структурой; следовательно, канонические структуры можно прогнозировать на основе только первичной аминокислотной последовательности.
Прогнозированные канонические складчатые структуры для гипервариабельных петель любой данной последовательности VH или VL в антителе, обладающем высокой гомологией с человеком, можно анализировать, используя алгоритмы, являющиеся общедоступными на сайтах www.bioinf.org.uk/abs/chothia.html, www.biochem.ucl.ac.uk/~martin/antibodies.html и www.bioc.unizh.ch/antibody/Sequences/Germlines/Vbase_hVk.html. Указанные программы позволяют выравнивать последовательности областей VH или VL с последовательностями областей VH или VL человека, имеющими известную каноническую структуру, и прогнозировать каноническую структуру гипервариабельных петель запрашиваемой последовательности.
В случае области VH Н1 и Н2 петли могут быть оценены как имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре в человеческих антителах, если удовлетворяется, по меньшей мере, первый и предпочтительно оба следующие критерия:
1. Длина, определяемая числом остатков, идентична ближайшему совпадающему классу канонической структуры человека.
2. По меньшей мере, 33% идентичность, предпочтительно, по меньшей мере, 50% идентичность с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур петель Н1 и Н2 (следует отметить, что в соответствии с целями вышеуказанного анализа петли Н1 и Н2 анализируют отдельно и каждую петлю сравнивают с ближайшим совпадающим классом канонической структуры человека).
Вышеуказанный анализ основан на прогнозировании канонической структуры петель Н1 и Н2 представляющего интерес антитела. Если фактические структуры петель Н1 и Н2 в представляющем интерес антителе известны, например, в результате проведения рентгеновской кристаллографии, то тогда петли Н1 и Н2 в представляющем интерес антителе могут быть оценены как имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, известной в человеческих антителах, если длина петли отличается от ближайшего совпадающего класса канонической структуры человека (обычно на ±1 или ±2 аминокислоты), но фактическая структура петель Н1 и Н2 в представляющем интерес антителе соответствует канонической складчатой структуре человека.
Ключевые аминокислотные остатки, обнаруженные в классах канонических структур человека для первой и второй гипервариабельных петель областей VH человека (Н1 и Н2), описаны в публикации Chothia et al., J. Mol. Biol., 227:799-817 (1992), которая включена в настоящем описании в полном объеме посредством ссылки. В частности, в таблице 3 на странице 802 публикации Chotia et al., которая включена в настоящем описании в полном объеме посредством ссылки, представлены предпочтительные аминокислотные остатки в основных сайтах для канонических структур Н1, обнаруженные в зародышевой линии человека, и в таблице 4 на странице 803, также которая включена в настоящем описании в полном объеме посредством ссылки, представлены предпочтительные аминокислотные остатки в ключевых сайтах для канонических структур CDR H2, обнаруженные в зародышевой линии человека.
В одном варианте осуществления изобретения Н1 и Н2 в области VH антитела, обладающего высокой гомологией с человеком, имеют прогнозированную или фактическую каноническую складчатую структуру, которая по существу идентична канонической складчатой структуре в человеческих антителах.
Антитела, обладающие высокой гомологией с человеком, могут включать область VH, в которой гипервариабельные петли Н1 и Н2 образуют комбинацию канонических складчатых структур, идентичную комбинации канонических структур, известных, по меньшей мере, в одной области VH зародышевой линии человека. Установлено, что только определенные комбинации канонических складчатых структур Н1 и Н2 действительно имеют место в областях VH, кодированных зародышевой линией человека. В одном варианте осуществления изобретения петли Н1 и Н2 в области VH антитела, обладающего высокой гомологией с человеком, могут быть получены из области VH вида, отличного от человека, например, вида семейства верблюдовых, благодаря образованию комбинации прогнозируемых или фактических канонических складчатых структур, которая идентична комбинации канонических складчатых структур, известных в зародышевой линии человека или в соматически мутированной области VH. В неограничивающих вариантах осуществления изобретения петли Н1 и Н2 в области VH антитела, обладающего высокой гомологией с человеком, могут быть получены из области VH вида, отличного от человека, например, вида семейства верблюдовых, благодаря образованию одной из следующих комбинаций канонических складок: 1-1, 1-2, 1-3, 1-6, 1-4, 2-1, 3-1 и 3-5.
Антитело с высокой гомологией с антителом человека может содержать область VH, которая демонстрирует высокую идентичность/гомологию последовательности с VH человека, и которое содержит гипервариабельные петли, демонстрирующие структурную гомологию с VH человека.
Предпочтительно, чтобы канонические складки, присутствующие в петлях Н1 и Н2 области VH антитела, обладающего высокой гомологией с человеком, и их комбинация, были «правильными» для последовательности области VH в зародышевой линии человека, которая наиболее близко совпадает с областью VH антитела, обладающего высокой гомологией с человеком, с точки зрения общей идентичности первичной аминокислотной последовательности. В качестве примера можно отметить, что, если последовательность наиболее близко совпадает с областью VH3 зародышевой линии человека, то желательно, чтобы петли Н1 и Н2 образовывали комбинацию канонических складок, которая также имеет место в области VH3 человека. Это может быть особенно важным в случае антител, обладающих высокой гомологией с человеком, которые были выделены от вида, отличного от человека, например, в случае антител, содержащих области VH и VL, выделенные из обычных верблюжьих антител, в частности, антител, содержащих гуманизированные области VH и VL верблюда.
Таким образом, в одном варианте осуществления область VH анти-IL-22R-антитела с высокой гомологией с человеком может иметь идентичность последовательности или гомологию последовательности на уровне 80% или выше, 85% или выше, 90% или выше, 95% или выше, 97% или выше или до 99% или даже 100% с человеческой областью VH в каркасных областях FR1, FR2, FR3 и FR4, и в дополнении H1 и H2 в том же самом антителе получают из «нечеловеческой» области VH (например, происходят из вида семейства верблюдовых), но образуют комбинацию прогнозированных или фактических канонических складчатых структур, которая является такой же, как и комбинация канонических складок, которая, как известно, имеет место в природе в одной и той же человеческой области VH.
В еще одних вариантах осуществления изобретения петли L1 и L2 в области VL антитела, обладающего высокой гомологией с человеком, получают из области VL вида, отличного от человека (например, из области VL верблюда), и каждая имеет прогнозированную или фактическую каноническую складчатую структуру, которая по существу идентична канонической складчатой структуре в человеческих антителах.
Как и в случае областей VH, гипервариабельные петли областей VL типов Vлямбда и Vкаппа могут образовывать ограниченное число конформаций или канонических структур, частично определяемых длиной, и также наличием ключевых аминокислотных остатков в определенных положениях канонических структур.
В представляющем интерес антителе, обладающем высокой гомологией с человеком, петли L1, L2 и L3, полученные из области VL вида, отличного от человека, например, из вида семейства верблюдовых, могут быть оценены как имеющие каноническую складчатую структуру, «по существу идентичную» канонической складчатой структуре, известной в человеческих антителах, если удовлетворяется, по меньшей мере, первый и предпочтительно оба следующие критерия:
1. Длина, определяемая числом остатков, идентична ближайшему совпадающему классу канонической структуры человека.
2. По меньшей мере, 33% идентичность, предпочтительно, по меньшей мере, 50% идентичность с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур петель L1 и L2 из репертуара Vлямбда или Vкаппа (следует отметить, что в соответствии с целями вышеуказанного анализа петли L1 и L2 анализируют отдельно и каждую петлю сравнивают с ближайшим совпадающим классом канонической структуры человека).
Вышеуказанный анализ основан на прогнозировании канонической структуры петель L1, L2 и L3 области VL представляющего интерес антитела. Если фактическая структура петель L1, L2 и L3 известна, например, в результате проведения рентгеновской кристаллографии, то тогда петли L1, L2 и L3 в представляющему интерес антителе могут быть оценены как имеющие каноническую складчатую структуру «по существу идентичную» канонической складчатой структуре, известной в человеческих антителах, если длина петли отличается от ближайшего совпадающего класса канонической структуры человека (обычно на ±1 или ±2 аминокислоты), но фактическая структура петель верблюда соответствует канонической складчатой структуре человека.
Ключевые аминокислотные остатки, обнаруженные в классах канонических структур человека для CDR областей Vлямбда и Vкаппа человека, описаны в публикациях Morea et al., Methods, 20:267-279 (2000) и Martin et al., J. Mol. Biol., 263:800-815 (1996). Репертуар структур области Vкаппа человека также описан в публикации Tomlinson et al., EMBO J. 14:4628-4638 (1995), и репертуа структур области Vлямбда описан в публикации Williams et al., J. Mol. Biol., 264:220-232 (1996). Содержание всех указанных документов включено в настоящем описании посредством ссылки.
Петли L1 и L2 в области VL антитела, обладающего высокой гомологией с человеком, могут образовывать комбинацию прогнозированных или фактических канонических складчатых структур, идентичную комбинации канонических складчатых структур, известной в области VL зародышевой линии человека. В неограничивающих вариантах осуществления изобретения петли L1 и L2 в области Vлямбда антитела, обладающего высокой гомологией с человеком (например, антитело, содержащее область VL, полученную от верблюда или ее гуманизированный вариант), могут образовывать одну из следующих комбинаций канонических складок: 11-7, 13-7 (А, В, С), 14-7 (А, В), 12-11, 14-11 и 12-12 (в соответствии с определением, приведенным в публикации Williams et al., J. Mol. Biol., 264:220-32 (1996), и представленных на сайте http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVL.html). В неограничивающих вариантах осуществления изобретения петли L1 и L2 в области Vкаппа могут образовывать одну из следующих комбинаций канонических складок: 2-1, 3-1, 4-1 и 6-1 (в соответствии с определением, приведенным в публикации Tomlinson et al., EMBO J., 14:4628-38 (1995), и представленных на сайте http://www.bioc.uzh.ch/antibody/Seqiences/Germlines/VBase_hVK.html).
В еще одном варианте осуществления изобретения все три петли L1, L2 и L3 в области VL антитела, обладающего высокой гомологией с человеком, могут иметь по существу человеческую структуру. Предпочтительно, чтобы область VL антитела, обладающего высокой гомологией с человеком, имела последовательность, характеризующуюся высокой идентичностью/гомологией с областью VL человека, и чтобы также гипервариабельные петли в области VL имели структуру, гомологичную области VL человека.
В одном варианте осуществления область VL анти-IL-22R-антитела, обладающего высокой гомологией с человеком, может иметь идентичность последовательности на уровне 80% или выше, 85% или выше, 90% или выше, 95% или выше, 97% или выше или до 99% или даже 100% с областью VL человека в каркасных областях FR1, FR2, FR3 и FR4, и, кроме того, гипервариабельная петля L1 и гипервариабельная петля L2 могут образовывать комбинацию прогнозированных или фактических канонических складчатых структур, которая является такой же, как комбинация канонических складок, которая, как известно, встречается в природе в такой же области VL человека.
Конечно, предусматривается, что области VH, обладающие высокой идентичностью/гомологией последовательностей с областью VH человека, и также структурной гомологией с гипервариабельными петлями области VH человека, могут быть объединены с областями VL, обладающими высокой идентичностью/гомологией последовательности с областью VL человека, и также структурной гомологией с гипервариабельными петлями области VL человека, с образованием антител, характеризующихся высокой гомологией с человеком, которые содержат объединенные пары VH/VL (например, объединенные пары VH/VL, полученные от верблюда), обладающие максимальной гомологией последовательностей и структуры с объединенными парами VH/VL человека.
«scFv» или «scFv-фрагмент». scFv или scFv фрагмент означает одноцепочечный вариабельный фрагмент. scFv представляет слитый белок области VH и области VL антитела, соединенных через линкер.
В. Анти-IL-22R-антитела
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые специфически связываются с IL-22R человека. Анти-IL-22R-антитела и фрагменты антител, описанные в настоящем описании, обладают свойствами, в частности, комбинациями свойств, которые являются отличительными и в некоторых случаях превосходящими анти-IL-22R-антитела предшествующего уровня техники. Эти свойства и характеристики будут теперь описаны более подробно.
Эпитопы IL-22R
Анти-IL-22R-антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются с эпитопом в человеческом белке IL-22R, который не содержит аминокислотный остаток тирозина 60 (Tyr60 или Y60), где это аминокислотное положение определено со ссылкой на последовательность IL-22R, показанную на фиг. 2 (SEQ ID NO:71). Данный находящийся на поверхности остаток IL-22R ранее был идентифицирован как критически важный для связывания лиганда и, как сообщается в настоящем документе (см., в частности, пример 8) для анти-IL-22R-антитела предшествующего уровня техники 280.346.TSY (как описано в WO 02011/01119) связывается с эпитопом, включающим этот остаток. С удивлением было обнаружено, что анти-IL-22R-антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются с эпитопом, который не включает этот критический остаток, и все же в некоторых вариантах осуществления они способны блокировать связывание IL-22 и/или ингибировать IL-22- и/или IL-20-опосредованный сигнальный путь через IL-22R.
Поверхность раздела связывания внеклеточного домена IL-22R и его лиганда IL-22 охарактеризована в публикации Jones et al. (Structure 16(9): 1333-1344 (2008)), содержание которой в полном объеме включено в настоящем описании посредством ссылки. Внеклеточный домен IL-22R состоит из пяти петель (L2-L6), некоторые остатки которых являются ключевыми для опосредования взаимодействия с находящимися на поверхности остатками на лиганде IL-22. Тирозин 60 расположен в петле L2, образованной остатками в домене D1 белка IL-22R. В IL-22/растворимом комплексе IL-22R остаток Tyr60 входит в небольшую полость на IL-22, образованную в сайте 1a лиганда. Более конкретно, эта полость образуется на пересечении спирали F и петли AB, двух отдельных вторичных структур, обнаруженных в лиганде IL-22. Остатки спирали F Lys162 и Glu166 на лиганде образуют водородные связи с OH Tyr60 IL-22R и, таким образом, Tyr60 является ключевым остатком, опосредующим взаимодействие между сайтом 1a на лиганде и доменом D1 рецептора.
Антитела и антигенсвязывающие фрагменты, описанные в настоящем описании, связываются с эпитопом, который не содержит Tyr60. Как отмечено выше, Tyr60 находится в петле L2 в домене D1 рецептора. В некоторых вариантах осуществления эпитоп, с которым связываются анти-IL-22R-антитела или антигенсвязывающие фрагменты, располагается, по меньшей мере, частично в домене D2 IL-22R, т.е. между остатками 125 и 228 SEQ ID NO:71. В некоторых вариантах осуществления, эпитоп, с которым связываются анти-IL-22R-антитела или антигенсвязывающие фрагменты, содержит, по меньшей мере, один, по меньшей мере, два, по меньшей мере, три аминокислотных остатка, расположенных в домене D2 белка IL-22R.
Заявители настоящего изобретения полагают, что тот факт, что антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются с эпитопом, который не содержит Tyr60, имеет отношение к механизму их действия. Полагается, что антитела и их антигенсвязывающие фрагменты взаимодействуют с доменом D2 IL22R вместо или в дополнение к взаимодействию с доменом D1. Известно, что домен D2 IL-22R участвует во взаимодействии с корецептором. Не желая связываться с теорией, заявители изобретения полагают, что молекулы антител по настоящему изобретению могут иметь двойной механизм действия, предотвращая связывание с лигандом и предотвращая взаимодействие с корецептором.
Эпитоп, с которым связываются антитела или антигенсвязывающие фрагменты по изобретению, может быть линейным эпитопом (т.е. эпитопом, состоящим из линейной последовательности аминокислот на антигене-мишене) или конформационным эпитопом (т.е. эпитопом, состоящим из аминокислот, которые необязательно являются смежными в антигене-мишене).
Аффинность связывания
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты по изобретению связываются с человеческим IL-22R с высокой аффинностью.
Как в настоящем описании используется, термин «аффинность» или «аффинность связывания» следует понимать, основываясь на обычном значении этого термина в данной области в контексте связывания антитела и отражает прочность и/или стабильность связывания между антигеном и сайтом связывания на антителе или его антигенсвязывающем фрагменте.
Аффинность связывания антитела или его антигенсвязывающего фрагмента для его соответствующего антигена можно определить экспериментально с использованием методов, известных в данной области. Например, с помощью инструментов BIACORE измеряют аффинность, на основе иммобилизации белка-мишени или антигена на биосенсорном чипе, в то время как антитело или фрагмент антитела пропускают над иммобилизованной мишенью при определенных условиях потока. Данные эксперименты дают значения kon и koff, которые могут быть переведены в значения KD, где KD представляет константу равновесия диссоциации антигена и антитела или его фрагмента. Чем ниже значение KD, тем сильнее связывающее взаимодействие между антителом и его антигеном-мишенью. Как отмечено выше, аффинность антитела может быть определена Biacore, например, с использованием протокола, описанного в настоящем описании в других местах (см. пример 6). Аффинность антитела или антигенсвязывающего фрагмента для человеческого IL-22R, измеренная Biacore, может быть определена с использованием рекомбинантной полноразмерной конструкции IL-22R, как описано, например, в примерах 3 и 6.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты по изобретению могут демонстрировать скорость диссоциации (koff) для IL-22R ниже 2×10-3 с-1, ниже 1,5×10-3 с-1, ниже 1,2×10-3 с-1 при тестировании в виде mAb, например, когда аффинность вариабельной области тяжелой цепи в паре с вариабельной областью легкой цепи тестируется в контексте молекулы IgG1. В предпочтительных вариантах осуществления анти-IL-22R-антитела или их антигенсвязывающие фрагменты по изобретению демонстрируют скорость диссоциации (koff) для IL-22R ниже 2,5×10-4 с-1, предпочтительно ниже 1,5×10-4 с-1, более предпочтительно ниже 1×10-4 с-1 при тестировании в виде mAb, например, когда аффинность вариабельной области тяжелой цепи в паре с вариабельной областью легкой цепи тестируется в контексте молекулы IgG1.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты по изобретению могут проявлять скорость диссоциации (koff) для IL-22R в диапазоне от 1×10-6 с-1 до 2×10-3 с-1, предпочтительно в диапазоне от 1×10-6 с-1 до 2,5×10-4 с-1, более предпочтительно в диапазоне от 1×10-5 с-1 до 1×10-4 с-1.
Анти-IL-22R-антитела или их антигенсвязывающие фрагменты по изобретению могут иметь значение KD ниже 3×10-9 М, ниже 2,5×10-9 М, ниже 1×10-9 М. В предпочтительных вариантах осуществления анти-IL-22R-антитела или их антигенсвязывающие фрагменты по изобретению имеют значение KD ниже 5×10-10 М, предпочтительно ниже 2×10-10 М, предпочтительно ниже 1,5×10-10 М.
Ингибирование связывания IL-22 и IL-20 и сигнального пути в направлении 5'-3' направлении
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты по изобретению связываются с человеческим IL-22R и ингибируют связывание лиганда с этим рецептором. Это было особенно неожиданным с учетом того, что антитела, описанные в настоящем описании, связываются с эпитопом, который не содержит Tyr60. Как описано выше, Tyr60 был идентифицирован как остаток, ключевой для связывания лиганда, и поэтому анти-IL-22R-антитела и антигенсвязывающие фрагменты по настоящему изобретению вызывают удивление своей способностью ингибировать связывание лиганда и в некоторых вариантах осуществления ингибировать сигнальный путь IL-22R в направлении 5'-3'.
Антитела и антигенсвязывающие фрагменты могут связывать с IL-22R человека и ингибировать связывание IL-22 или ингибировать связывание IL-20. Предпочтительно, антитела и антигенсвязывающие фрагменты, описанные в настоящем описании, ингибируют связывание IL-22 и IL-20 с IL-22R.
Связывание лиганда с IL-22R на клетках, которые экспрессируют корецептор IL-10R2 или IL-20R2, индуцирует конформационное изменение в рецепторном комплексе, так что сигнальные пути в направлении 5'-3' от рецептора активируются. Антитела или их антигенсвязывающие фрагменты, описанные в настоящем описании, могут ингибировать IL-22-зависимую активацию IL-22R или могут ингибировать IL-20-зависимую активацию IL-22R. Как в настоящем описании используется, термин «IL-22-зависимая активация IL-22R» следует понимать как цепь сигнальных событий, которые имеют место в направлении 5'-3' от рецепторного комплекса IL-22R при связывании лиганда IL-22 с IL-22R/IL-10R2. Как в настоящем описании используется, термин «IL-20-зависимая активация IL-22R» следует понимать как цепь сигнальных событий, которые происходят в направлении 5'-3' от IL-22R при связывании лиганда IL-20 с комплексом IL-22R/IL-20R2.
IL-22-зависимая активация IL-22R может быть измерена в клетках или клеточных линиях, экспрессирующих IL-22R и ко-рецептор IL-10R2, например, клеточной линии BW-hIL-22R, описанной в публикации Dumoutier et al. J. Biol. Chem., 2009 Sep. 25; 284 (39): 26377-84, содержание которой в полном объеме включено в настоящем описании посредством ссылки. IL-22 индуцирует остановку роста в клеточной линии BW-hIL-22R, и, следовательно, IL-22-зависимую активацию IL-22R можно определить на данной клеточной линии путем измерения пролиферации клеток BW-hIL-22R, например, как описано в примере 6. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, описанные в настоящем описании, демонстрируют IC50 (концентрация, при которой достигается 50% ингибирование активации) в анализе IL-22-зависимой клеточной пролиферации составляет ниже 650 пМ, предпочтительно ниже 600 пМ. Анализ клеточной пролиферации предпочтительно представляет собой анализ, включающий клеточную линию BW-hIL-22R, как в настоящем описании описано.
IL-20-зависимая активация IL-22R может быть измерена на клетках или клеточных линиях, экспрессирующих IL-22R и ко-рецептор IL-20R2, например, клеточной линии Baf3-hIL-22R/IL20Rb, описанной в настоящем описании, и также в публикации Dumoutier et al. J Biol Chem. 2009 сент. 25; 284 (39): 26377-84, содержание которой в полном объеме включено в настоящем описании посредством ссылки. IL-20 индуцирует пролиферацию в клеточной линии Baf3-hIL-22R/IL20Rb, и, следовательно, IL-20-зависимую активацию IL-22R можно определить на этой клеточной линии путем измерения пролиферации клеток Baf3-hIL-22R/IL20Rb, например, как описано в примере 6. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, описанные в настоящем описании, демонстрируют IC50 (концентрация, при которой достигается 50% ингибирование активации) в анализе IL-20-зависимой клеточной пролиферации ниже 1 нМ, предпочтительно ниже 800 пМ. Анализ клеточной пролиферации предпочтительно представляет собой анализ, включающий клеточную линию Baf3-hIL-22R/IL20Rb, как в настоящем описании описано.
В дополнение к вышеописанным анализам пролиферации IL-22- и/или IL-20-зависимая активация IL-22R может быть измерена с использованием альтернативных клеточных анализов, например, анализа фосфорилирования STAT3, как описано в WO 02011/01119.
В предпочтительных вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению связываются с человеческим IL-22R и ингибируют как IL-22- зависимую активацию IL-22R, так и IL-20-зависимую активацию IL-22R. В таких вариантах осуществления ингибирующая или «нейтрализующая» активность антител или антигенсвязывающих фрагментов может быть эквипотентной для IL-22- и IL-20-зависимой активации. Альтернативно, ингибирующая или нейтрализующая активность антител или антигенсвязывающих фрагментов может быть выше для активации IL-22R, опосредованной одним из лигандов, по сравнению с активацией IL-22R, опосредованной другим лигандом. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по изобретению проявляют более высокую ингибирующую активность для IL-22-зависимой активации IL-22R по сравнению с IL-20-зависимой активацией IL-22R. Однако в таких вариантах ингибирующая активность антитела или антигенсвязывающего фрагмента для IL-22-зависимой активации IL-22R обычно будет в 5 раз ниже, предпочтительно в 4 раза выше, более предпочтительно в 2 раза ниже, чем ингибирующая активность антитела или антигенсвязывающего фрагмента для IL-20-зависимой активации IL-22R.
Перекрестная реактивность
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, описанные в настоящем описании, которые связываются с IL-22R человека, могут перекрестно реагировать с одним или более видовыми гомологами IL-22R, например, гомологами IL-22R приматов.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению не реагируют перекрестно с мышиным IL-22R. Альтернативно или дополнительно, антитела или антигенсвязывающие фрагменты могут связываться с одним или более гомологами IL-22R приматов, например, белками IL-22R от макака-резус и обезьяны циномолгус. Перекрестная реактивность с другими видовыми гомологами может быть особенно преимущественной при разработке и тестировании терапевтических антител. Например, доклиническое токсикологическое исследование терапевтических антител часто проводится на видах приматов, включая, не ограничиваясь ими, макак-резус и обезьян циномогус. Следовательно, перекрестная реактивность с этими видовыми гомологами может быть особенно преимущественной для разработки антител в качестве кандидатов для применения в клинике.
Антитела, полученные от верблюда
В некоторых вариантах осуществления изобретения антитела или антигенсвязывающие фрагменты, описанные в настоящем описании, могут содержать, по меньшей мере, одну гипервариабельную петлю или участок, определяющий комплементарность, полученные из области VH или области VL вида семейства верблюдовых. В частности, антитело или антигенсвязывающий фрагмент могут содержать области VH и/или VL или их CDR, полученные активной иммунизацией аутбредных верблюдов, например, ламы, антигеном IL-22R или его фрагментом.
Выражение «гипервариабельная петля или участок, определяющий комплементарность, полученные из области VH или VL вида семейства верблюдовых», означает, что эта гипервариабельная петля (HV) или CDR имеет аминокислотную последовательность, которая идентична или по существу идентична аминокислотной последовательности гипервариабельной петли или CDR, которая кодируется геном иммуноглобулина вида семейства верблюдовых. В этом контексте «ген иммуноглобулина» включает гены зародышевой линии, гены иммуноглобулина, которые подверглись перегруппировке, и также соматически мутированные гены. Таким образом, аминокислотная последовательность HV или CDR, полученная из области VH или VL вида семейства верблюдовых, может быть идентична аминокислотной последовательности HV или CDR, присутствующей в зрелом обычном антителе вида семейства верблюдовых. Выражение «полученная из» в этом контексте подразумевает структурную взаимосвязь в том смысле, что HV или CDR анти-IL-22R-антитела воплощают аминокислотную последовательность (или ее минорные варианты), которая первоначально кодировалась геном иммуноглобулина вида семейства верблюдовых. Однако это не обязательно означает конкретную взаимосвязь в отношении способа получения, используемого для продукции антитела к IL-22R.
Полученные от верблюдов анти-IL-22R-антитела могут быть получены от любых видов верблюдов, включая, среди прочего, ламу, дромедара, альпака, викунью, гуанако или верблюда.
Анти-IL-22R-антитела, содержащие области VH и VL, или их CDR, полученные от верблюда, обычно представляют рекомбинантно экспрессированные полипептиды и могут представлять химерные полипептиды. Термин «химерный полипептид» относится к искусственному (не встречающемуся в природе) полипептиду, который создается сопоставлением двух или более пептидных фрагментов, которые иначе не встречаются смежными друг с другом. В это определение включены «видовые» химерные полипептиды, созданные сопоставлением пептидных фрагментов, кодированных двумя или более видами, например, верблюдом и человеком.
CDR, полученные от верблюда, могут содержать одну из последовательностей CDR, показанных в SEQ ID NO:6 и 13 (CDR3 тяжелой цепи), или SEQ ID NO:4 и 11 (CDR2 тяжелой цепи) или SEQ ID NO:2 и 9 (CDR1 тяжелой цепи) или одну из последовательностей CDR, показанных в SEQ ID NO:20 и 27 (CDR3 легкой цепи), или SEQ ID NO:18, 25 и 47 (CDR2 легкой цепи) или SEQ ID NO:16 и 23 (CDR1 легкой цепи).
В одном варианте осуществления полноразмерную область VH и/или полноразмерную область VL можно получить из вида семейства верблюдовых. В конкретных вариантах осуществления область VH, полученная от верблюда, может содержать аминокислотную последовательность, выбранную из SEQ ID NO:29 и 31, тогда как область VL, полученная от верблюда, может содержать аминокислотную последовательность, выбранную из SEQ ID NO:30, 32 и 62. Область VH, полученную от верблюда, и/или область VL, полученную от верблюда, затем можно подвергнуть генной инженерии белков, где одна или более аминокислотных замен, вставок или делеций вводятся в аминокислотную последовательность верблюда. Данные генно-инженерные изменения предпочтительно включают аминокислотные замены относительно последовательности верблюда. Такие изменения включают «гуманизацию» или «превращение в последовательность зародышевого типа», где один или более аминокислотных остатков в области VH или VL, кодированной верблюдом, заменены эквивалентными остатками из гомологичной области VH или VL, кодированной человеком. В некоторых вариантах осуществления область VH, полученная от верблюда, может проявлять, по меньшей мере, 90%, 95%, 97%, 98% или 99% идентичность с аминокислотной последовательностью, показанной в SEQ ID NO:29 или 31. Альтернативно или дополнительно область VL, полученная от верблюда, может проявлять, по меньшей мере, 90%, 95%, 97%, 98% или 99% идентичность с аминокислотной последовательностью, показанной в SEQ ID NO:30, 32 или 62.
В качестве основы для конструирования антигенсвязывающих полипептидов по изобретению можно использовать выделенные области VH и VL верблюда, полученные путем активной иммунизации верблюда (например, ламы) человеческим антигеном IL-22R. Начиная с интактных областей VH и VL верблюда, можно сконструировать одну или более аминокислотных замен, вставок или делеций, которые отходят от исходной последовательности верблюда. В некоторых вариантах осуществления такие замены, вставки или делеции могут присутствовать в каркасных доменах области VH и/или области VL. Целью таких изменений в первичной аминокислотной последовательности может быть снижение предположительно неблагоприятных свойств (например, иммуногенности у человека-хозяина (так называемая гуманизация), сайтов, вызывающих потенциальную гетерогенность продукта и/или нестабильность (гликозилирование, дезамидирование, изомеризация и тому подобное), или для усиления некоторого другого благоприятного свойства молекулы (например, растворимости, стабильности, биодоступности и тому подобное). В еще одних вариантах осуществления изменения в первичной аминокислотной последовательности могут быть проведены в одной или более гипервариабельных петлях (или CDR) областей VH и/или VL верблюда, полученных активной иммунизацией. Такие изменения могут быть введены для усиления антигенсвязывающей аффинности и/или специфичности, или для снижения предположительно неблагоприятных свойств (например, иммуногенности у человека-хозяина (так называемая гуманизация), сайтов, вызывающих потенциальную гетерогенность продукта и/или нестабильность (гликозилирование, дезамидирование, изомеризация и тому подобное), или для усиления некоторого другого благоприятного свойства молекулы, например, растворимости, стабильности, биодоступности и тому подобное.
Таким образом, в одном варианте осуществления изобретение относится к вариантному анти-IL-22R-антителу, которое содержит, по меньшей мере, одну аминокислотную замену, по меньшей мере, в одной каркасной области или CDR области VH или области VL по сравнению с областью VH или VL, полученной от верблюда, примеры которых включают, не ограничиваясь этим, области VH верблюда, содержащие аминокислотные последовательности, показанные в SEQ ID NO:29 и 31, и области VL верблюда, содержащие аминокислотные последовательности, показанные в SEQ ID NO:30, 32 или 62.
В еще одних вариантах осуществления обеспечиваются «химерные» молекулы антитела, содержащие области VH и VL, полученные от верблюда, и одну или более константных областей из антитела, отличного от верблюжьего, например, константные области человека (или их генно-инженерные варианты). В таких вариантах осуществления предпочтительно, чтобы область VH и область VL были получены от одного и того же вида семейства верблюдовых, например, как VH, так и VL могут происходить от Lama glama, или обе VH и VL могут происходить от Lama pacos (до введения изменений в аминокислотную последовательность). В таких вариантах осуществления как область VH, так и область VL могут быть получены от одного животного, в частности, одного животного, которое активно иммунизировано антигеном IL-22R.
В качестве альтернативы генно-инженерным изменениям в первичной аминокислотной последовательности верблюжьих областей VH и/или VL, могут быть выделены отдельные верблюжьи гипервариабельные петли или CDR, или их комбинации из областей VH/VL верблюда и перенесены в альтернативную (т.е. отличную от верблюжьей) каркасную область, например, каркас VH/VL человека, трансплантацией CDR. В частности, неограничивающие варианты осуществления CDR, происходящие от верблюда, могут быть выбраны из CDR, имеющих аминокислотные последовательности, показанные в SEQ ID NO:6 и 13 (CDR3 тяжелой цепи), или SEQ ID NO:4 и 11 (CDR2 тяжелой цепи) или SEQ ID NO:2 и 9 (CDR1 тяжелой цепи) или одна из последовательностей CDR, показанных в SEQ ID NO:20 и 27 (CDR3 легкой цепи), или SEQ ID NO:18, 25 и 47 (CDR2 легкой цепи) или SEQ ID NO:16 и 23 (CDR1 легкой цепи).
Анти-IL-22R-антитела, содержащие области VH и VL, или их CDR, полученные от верблюда, могут принимать разнообразные различные варианты осуществления, в которых присутствуют как область VH, так и область VL. Термин «антитело» используется в настоящем описании в самом широком смысле и охватывает, не ограничиваясь этим, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела), при условии, что они проявляют соответствующую иммунологическую специфичность для белка IL-22R. Как в настоящем описании используется, термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, то есть отдельные антитела, составляющие популяцию, идентичны, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, направленными против одного антигенного сайта. Кроме того, в отличие от препаратов обычных (поликлональных) антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов) на антигене, каждое моноклональное антитело направлено против одной детерминанты или эпитопа на антигене.
«Фрагменты антитела» включают часть полноразмерного антитела, как правило, его антигенсвязывающий участок или вариабельную область. Примеры фрагментов антител включают Fab, Fab', F(ab')2, биспецифические Fab и Fv-фрагменты, диатела, линейные антитела, молекулы одноцепочечных антител, одноцепочечный фрагмент вариабельной области (scFv) и мультиспецифические антитела, образованные из фрагментов антител (см. публикацию Holliger and Hudson, Nature Biotechnol, 23: 1126-36 (2005), содержание которой включено в настоящем описании посредством ссылки).
В неограничивающих вариантах осуществления анти-IL-22R-антитела, содержащие области VH и VL, или их CDR, полученные от верблюда, могут содержать домены CH1 и/или домены CL, аминокислотная последовательность которых является полностью или по существу человеческой. Когда антигенсвязывающий полипептид по изобретению представляет собой антитело, предназначенное для терапевтического применения человеку, то типично для всей константной области антитела или, по меньшей мере, ее части, иметь полностью или по существу человеческую аминокислотную последовательность. Следовательно, одна или более, или любая комбинация домена CH1, шарнирной области, домена CH2, домена CH3 и домена CL (и домена CH4, если он присутствует), могут быть полностью или по существу человеческими относительно его аминокислотной последовательности.
Преимущественно, домен CH1, шарнирная область, домен CH2, домен CH3 и домен CL (и домен CH4, если он присутствует), могут иметь полностью или по существу человеческую аминокислотную последовательность. В контексте константной области гуманизированного или химерного антитела или фрагмента антитела термин «по существу человеческая» относится к аминокислотной последовательности, которая, по меньшей мере, на 90% или, по меньшей мере, на 92% или, по меньшей мере, на 95% или, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична константной области человека. Термин «человеческая аминокислотная последовательность» в этом контексте относится к аминокислотной последовательности, которая кодируется геном иммуноглобулина человека, включающим гены зародышевой линии, реаранжированные и соматически мутированные гены. Изобретение также относится к полипептидам, содержащим константные области «человеческой» последовательности, которые были изменены в результате одной или более аминокислотных добавлений, делеций или замен по отношению к человеческой последовательности, за исключением тех вариантов, в которых явно необходимо присутствие «полностью человеческой» шарнирной области.
Наличие «полностью человеческой» шарнирной области в анти-IL-22R-антителах по изобретению может быть полезным как для сведения к минимуму иммуногенности, так и для оптимизации стабильности антитела. Как обсуждается в настоящем описании в других местах, предполагается, что одна или более аминокислотных замен, вставок или делеций могут быть сделаны в константной области тяжелой и/или легкой цепи, в частности, в Fc-области. Аминокислотные замены могут привести к замене замещенной аминокислоты другой природной аминокислотой или неприродной или модифицированной аминокислотой. Также допускаются другие структурные модификации, например, такие как изменения в паттерне гликозилирования (например, добавлением или делецией сайтов, связанных с N- или O-гликозилированием). В зависимости от предполагаемого применения антитела может быть желательным модифицировать антитело по изобретению в отношении его связывающих свойств с Fc-рецепторами, например, для модуляции эффекторной функции. Например, остаток(и) цистеина может быть введен в Fc-область, тем самым обеспечивая образование межцепочечной дисульфидной связи в этой области. Гомодимерное антитело, полученное таким образом, может иметь повышенную способность к интернализации и/или повышенную комплемент-опосредованную клеточную гибель и антителозависимую клеточную цитотоксичность (ADCC). См. Caron et al., J. Exp. Med., 176: 1191-1195 (1992) и Shopes B. J. Immunol., 148: 2918-2922 (1992). Изобретение также относится к иммуноконъюгатам, содержащим антитело, как в настоящем описании описано, конъюгированное с цитотоксическим агентом, таким как химиотерапевтический агент, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (например, т.е. радиоконъюгат). Fc-области также могут быть сконструированы для удлинения периода полураспада, как описано в публикации Chan and Carter, Nature Reviews: Immunology, Vol.10, pp301-316, 2010, включенной в настоящем описании посредством ссылки.
В еще одном варианте осуществления Fc-область модифицирована для увеличения способности антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или повышения аффинности антитела к рецептору Fcγ модификацией одной или более аминокислот. В альтернативных вариантах осуществления Fc-область может быть сконструирована таким образом, что эффекторная функция отсутствует. Анти-IL-22R-антитело, не имеющее эффекторной функции Fc, может быть особенно пригодно в качестве агента, блокирующего рецептор. В некоторых вариантах осуществления антитела по изобретению могут иметь Fc-область, полученную из встречающихся в природе изотипов IgG, имеющих пониженную эффекторную функцию, например IgG4. Fc-области, полученные из IgG4, могут быть дополнительно модифицированы для повышения терапевтической применимости, например, введением модификаций, которые минимизируют обмен плеч между молекулами IgG4 in vivo.
В еще одном варианте осуществления модифицировано гликозилирование антитела. Например, может быть получено негликозированное антитело (то есть, в антителе отсутствует гликозилирование). Гликозилирование может быть изменено, например, для повышения аффинности антитела к антигену-мишени. Такие модификации углеводов могут быть выполнены, например, изменением одного или более сайтов гликозилирования в последовательности антитела. Например, может быть сделана одна или более аминокислотных замен, которые приводят к элиминации одного или более сайтов гликозилирования каркаса вариабельной области, чтобы тем самым элиминировать гликозилирование в этом сайте. Такое дегликозилирование может повысить аффинность антитела к антигену.
Также настоящее изобретение относится к вариантным анти-IL-22R-антителам, имеющим измененный тип гликозилирования, таким как гипофукозилированное антитело, имеющее меньшие количества фукозильных остатков, или полностью или частично дефукозилированное антитело (как описано Natsume et al., Development Development Development and Therapy, Vol.3, pp. 7-16, 2009), или антитело, имеющее повышенное число биссекторных структур GlcNac. Было показано, что такие измененные паттерны гликозилирования повышают активность ADCC антител, обеспечивая, как правило, 10-кратное повышение ADCC относительно эквивалентного антитела, содержащего «нативную» Fc-область человека. Такие модификации углеводов могут быть выполнены, например, экспрессией антитела в клетке-хозяине с измененным ферментативным механизмом гликозилирования (как описано Yamane-Ohnuki and Satoh, mAbs 1: 3, 230-236, 2009). Примерами нефукозилированных антител с повышенной функцией ADCC являются антитела, которые получены с использованием технологии Potelligent™ BioWa Inc.
Изобретение в определенных вариантах осуществления может включать химерные верблюжьи/человеческие антитела и, в частности, химерные антитела, в которых области VH и VL имеют полностью верблюжью последовательность (например, ламы или альпако), и оставшаяся часть антитела имеет полностью человеческую последовательность. Анти-IL-22R-антитела могут включать антитела, содержащие «гуманизированные» варианты или варианты «зародышевого типа» областей VH и VL, или их CDR, полученных от верблюда, и химерные верблюжьи/человеческие антитела, в которых области VH и VL содержат одну или более аминокислотных замен в каркасных областях по сравнению с верблюжьими областями VH и VL, полученными активной иммунизацией верблюда антигеном IL-22R или его фрагментом. Такая «гуманизация» увеличивает процентную идентичность последовательности с областями VH или VL зародышевой линии человека, в результате замены несовпадений аминокислотных остатков в исходной верблюжьей области VH или VL эквивалентным остатком, найденным в области VH или VL зародышевой линии человека.
Анти-IL-22R-антитела также могут представлять CDR-привитые антитела, в которых CDR (или гипервариабельные петли), полученные из антитела верблюда, или иначе кодированные геном верблюда, пересажены на каркас VH и VL человека, и остальная часть антитела также имеет полностью человеческое происхождение. Такие CDR-привитые анти-IL-22R-антитела могут содержать CDR, имеющие аминокислотные последовательности, показанные в SEQ ID NO:6 и 13 (CDR3 тяжелой цепи), или SEQ ID NO:4 и 11 (CDR2 тяжелой цепи) или SEQ ID NO:2 и 9 (CDR1 тяжелой цепи) или одну из последовательностей CDR, показанных в SEQ ID NO:20 и 27 (CDR3 легкой цепи), или SEQ ID NO:18, 25 и 47 (CDR2 легкой цепи) или SEQ ID NO:16 и 23 (CDR1 легкой цепи).
Гуманизированные, химерные и CDR-привитые IL-22R антитела, как описано выше, в частности антитела, содержащие гипервариабельные петли или CDR, полученные активной иммунизацией верблюда, можно легко получить с использованием обычных методов рекомбинантной ДНК и методов экспрессии с использованием прокариотических и эукариотических клеток-хозяев, сконструированных для получения представляющего интерес полипептида, и включающих, не ограничиваясь этим, бактериальные клетки, дрожжевые клетки, клетки млекопитающих, клетки насекомых, растительные клетки, некоторые из которых описаны в настоящем описании и иллюстрированы в сопровождаемых примерах.
Полученные от верблюда анти-IL-22R-антитела включают варианты, в которых гипервариабельная петля(и) или CDR(и) области VH и/или области VL получены из обычного верблюжьего антитела, продуцированного против человеческого IL-22R, но где, по меньшей мере, одна из указанных (полученных от верблюда) гипервариабельных петель или CDR были сконструированы таким образом, чтобы включать одну или более аминокислотных замен, добавлений или делеций относительно последовательности, кодированной верблюдом. Такие изменения включают «гуманизацию» гипервариабельных петель/CDR. Полученные от верблюда HV/CDR, которые были сконструированы таким образом, что они могут по-прежнему иметь аминокислотную последовательность, которая «по существу идентична» аминокислотной последовательности HV/CDR, кодированных верблюдом. В этом контексте «существенная идентичность» может допускать не более одной или не более двух аминокислотных последовательностей, не совпадающих с HV/CDR, кодированных верблюдом. Конкретные варианты осуществления анти-IL-22R-антитела могут содержать гуманизированные варианты последовательностей CDR, показанных в SEQ ID NO:6 и 13 (CDR3 тяжелой цепи), или SEQ ID NO:4 и 11 (CDR2 тяжелой цепи) или SEQ ID NO:2 и 9 (CDR1 тяжелой цепи) или одну из последовательностей CDR, показанных в SEQ ID NO:20 и 27 (CDR3 легкой цепи), или SEQ ID NO:18, 25 и 47 (CDR2 легкой цепи) или SEQ ID NO:16 и 23 (CDR1 легкой цепи).
Анти-IL-22R-антитела, полученные от верблюда, могут относиться к любому изотипу. Антитела, предназначенные для терапевтического применения человеку, обычно относятся к IgG, IgD, IgE IgG, IgM типу, часто IgG типу, и в этом случае они могут относиться к любому из четырех подклассов IgG1, IgG2a и IgG2b, IgG3 или IgG4. В каждом из этих подклассов допускается выполнение одной или более аминокислотных замен, вставок или делеций в Fc-области или выполнение других структурных модификаций, например, для усиления или снижения Fc-зависимых функциональных свойств.
Предпочтительные анти-IL-22R-антитела - 230C9 и 223G5 и антитела, связанные с ними
Предпочтительными анти-IL-22R-антителами и их антигенсвязывающими фрагментами по настоящему изобретению являются гуманизированные варианты и варианты зародышевого типа антител, полученных от верблюда, описанных в настоящем описании. Гуманизированные варианты и варианты зародышевого типа демонстрируют высокую гомологию с антителом человека и предпочтительно проявляют высокую гомологию с молекулами человеческого IgG, более предпочтительно IgG1. В некоторых вариантах осуществления предпочтительные анти-IL-22R-антитела и антигенсвязывающие фрагменты по настоящему изобретению содержат гипервариабельные петли или CDR, имеющие человеческие или «человекоподобные» канонические складки, как в настоящем описании описано в других местах.
Предпочтительные анти-IL-22R-антитела по настоящему изобретению могут иметь идентичность аминокислотной последовательности на уровне 93% или выше с одной или более областями VH человека, в частности, в каркасных областях FR1, FR2, FR3 и FR4. Альтернативно или дополнительно, предпочтительные анти-IL-22R-антитела могут проявлять идентичность аминокислотной последовательности на уровне 96% или выше с одной или более областями VL человека, в частности, в каркасных областях FR1, FR2, FR3 и FR4. Предпочтительные анти-IL-22R-антитела по настоящему изобретению могут обладать идентичностью аминокислотной последовательности на уровне 93% или выше с одной или более областями VH человека, и идентичностью объединенной аминокислотной последовательности на уровне 93% или выше с одной или более областями VH человека и одной или более областями VL человека, в частности, в том случае, когда идентичность последовательности определяется каркасными областями.
За счет высокой гомологии с антителом человека предпочтительные анти-IL-22R-антитела, представленные в настоящем описании, проявляют низкую иммуногенность, что было оценено с использованием платформы ElbaseTM от lonza (оценка гена DRB-1), используя настройки «HLA class II - Caucasian v3.0». В некоторых вариантах осуществления анти-IL-22R-антитела по изобретению имеют балл DRB-1 ниже 950, предпочтительно ниже 850, более предпочтительно ниже 750, наиболее предпочтительно ниже 650.
Предпочтительными анти-IL-22R-антителами по изобретению являются моноклональные антитела, содержащие шарнирную область, домен CH2 и домен CH3 из человеческого IgG, предпочтительно IgG1. В некоторых вариантах осуществления Fc-область моноклонального антитела не имеет эффекторной функции, то есть является нулевой Fc. Это особенно выгодно для терапевтических блокирующих антител.
Предпочтительные анти-IL-22R-антитела и антигенсвязывающие фрагменты по изобретению демонстрируют комбинацию свойств, которые делают их усовершенствованными анти-IL-22R-антителами, по сравнению с описанными на предшествующем уровне техники. Предпочтительные анти-IL-22R-антитела и антигенсвязывающие фрагменты по изобретению могут проявлять следующую комбинацию свойств:
(i) связывание с эпитопом в белке IL-22R, который не содержит Tyr60;
(ii) связывание с эпитопом, который расположен, по меньшей мере, частично, в домене D2 белка IL-22R;
(iii) высокая аффинность связывания с IL-22R человека;
(iv) ингибирование IL-22-зависимой активации IL-22R и IL-20-зависимой активации IL-22R;
(v) отсутствие перекрестной реактивности с мышиным IL-22R; и
(vi) перекрестная реактивность с IL-22R макака-резус и/или обезьяны циномолгус.
Предпочтительные анти-IL-22R-антитела по изобретению могут связываться с эпитопом, который не содержит ключевой остаток Tyr60. Предпочтительные анти-IL-22R-антитела по изобретению могут связываться с эпитопом, который расположен, по меньшей мере, частично в домене D2 IL-22R, где домен D2 находится от аминокислоты 125 до аминокислоты 228 в последовательности SEQ ID NO:71. Предпочтительные анти-IL-22R-антитела по изобретению могут также связываться с IL-22R человека с высокой аффинностью, обычно проявляя скорость диссоциации (где koff измеряется Biacore) для человеческого IL-22R на уровне ниже 2,5×10-4 с-1 при измерении в виде mAb. В некоторых вариантах осуществления предпочтительные анти-IL-22R-антитела по изобретению связываются с человеческим IL-22R с высокой аффинностью, проявляя скорость диссоциации (где koff измеряется Biacore) для человеческого IL-22R в диапазоне от 1×10-6 с-1 до 2,5×10-4 с-1. В некоторых вариантах осуществления предпочтительные анти-IL-22R-антитела по изобретению связываются с человеческим IL-22R, имеющим значение KD ниже 5×10-10 М.
Предпочтительные анти-IL-22R-антитела могут также ингибировать IL-22- и IL-20-зависимую активацию IL-22R и, как правило, проявляют ингибирующую активность для IL-20-зависимой активации, которая не менее чем в четыре раза выше, чем ингибирующая активность, проявляемая для IL-22-зависимой активации. Предпочтительные анти-IL-22R-антитела могут связываться с человеческим IL-22R с высокой аффинностью и перекрестно реагировать с видовыми гомологами IL-22R от макака-резус и обезьяны циномолгус, но не реагируют перекрестно с мышиным IL-22R.
В некоторых вариантах осуществления предпочтительные антитела и антигенсвязывающие фрагменты по настоящему изобретению содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где область VH содержит:
CDR3 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [VGFSGTYYSES], SEQ ID NO:6,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [SIYNDASNTAYSDSVKG], SEQ ID NO:36,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [SYDMN], SEQ ID NO:34,
и область VL содержит:
CDR3 вариабельной области легкой цепи, содержащий или состоящий из аминокислотной последовательности [QSGSSSSNAV], SEQ ID NO:54,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [GQNNRPS], SEQ ID NO:47,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [QGGYYAH], SEQ ID NO:16.
Антитела и антигенсвязывающие фрагменты, имеющие последовательности CDR области VH и VL, как определено выше, могут содержать область VH, содержащую или состоящую из последовательности SEQ ID NO:63, и/или область VL, содержащую или состоящую из последовательности SEQ ID NO:64. В некоторых вариантах осуществления настоящего изобретения обеспечиваются антитела или их антигенсвязывающие фрагменты, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность VH, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:63, и/или вариабельная область легкой цепи содержит VL, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:64. Для вариантов осуществления, где области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью последовательности с эталонной последовательностью, области VH и/или VL могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только в каркасных областях.
Антитела и антигенсвязывающие фрагменты, имеющие последовательности CDR области VH и VL, как определено выше, могут содержать полноразмерную тяжелую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:67, и/или полноразмерную легкую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:68. В некоторых вариантах осуществления обеспечиваются антитела, содержащие тяжелую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности, или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:67, и/или легкую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:68. Для вариантов осуществления, где цепи антител определяются конкретной процентной идентичностью с эталонной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только вне участков CDR.
В некоторых вариантах осуществления предпочтительные антитела и антигенсвязывающие фрагменты по настоящему изобретению содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где область VH содержит:
CDR3 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [PPGPFKAHYNGAKY], SEQ ID NO:43,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [GIHISGGITYYTDSVKG], SEQ ID NO:41,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [SYFMS], SEQ ID NO:9,
и область VL содержит:
CDR3 вариабельной области легкой цепи, содержащий или состоящий из аминокислотной последовательности [ASYRLYADYV], SEQ ID NO:27,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [EVNKRSS], SEQ ID NO. 59,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [TGTSSDIGSYNYVS], SEQ ID NO:57.
Антитела и антигенсвязывающие фрагменты, имеющие последовательности CDR области VH и VL, как определено выше, могут содержать область VH, содержащую или состоящую из последовательности SEQ ID NO:65, и/или область VL, содержащую или состоящую из последовательности SEQ ID NO:66. В некоторых вариантах осуществления настоящего изобретения обеспечиваются антитела или их антигенсвязывающие фрагменты, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность VH, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:65, и/или вариабельная область легкой цепи содержит VL, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:66. Для вариантов осуществления, где области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью последовательности с эталонной последовательностью, области VH и/или VL могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только в каркасных областях.
Антитела и антигенсвязывающие фрагменты, имеющие последовательности CDR области VH и VL, как определено выше, могут содержать полноразмерную тяжелую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:69, и/или полноразмерную легкую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:70. В некоторых вариантах осуществления обеспечиваются антитела, содержащие тяжелую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности, или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:69, и/или легкую цепь, по меньшей мере, с 85% идентичностью последовательности или, по меньшей мере, 90% идентичностью последовательности или, по меньшей мере, 95% идентичностью последовательности или, по меньшей мере, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:70. Для вариантов осуществления, где цепи антител определяются конкретной процентной идентичностью с эталонной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять идентичные последовательности CDR с теми, которые присутствуют в эталонной последовательности, так что вариация присутствует только вне участков CDR.
Перекрестно конкурирующие антитела
Настоящее изобретение также включает (моноклональные) антитела или их антигенсвязывающие фрагменты, которые «перекрестно конкурируют» с антителами или антигенсвязывающими фрагментами, описанными в настоящем описании.
В частности, обеспечиваются антитела или их антигенсвязывающие фрагменты, которые связываются с цитокиновым рецептором IL-22R и перекрестно конкурируют с антителами или их антигенсвязывающими фрагментами, содержащими комбинацию CDR3 (HCDR3) вариабельной области тяжелой цепи, CDR2 (HCDR2) вариабельной области тяжелой цепи и CDR1 (HCDR1) вариабельной области тяжелой цепи, CDR3 (LCDR3) вариабельной области легкой цепи, CDR2 (LCDR2) вариабельной области легкой цепи и CDR1 (LCDR1) вариабельной области легкой цепи, где комбинация выбрана из группы, включающей:
(i) HCDR3, содержащий SEQ ID NO:43; HCDR2, содержащий SEQ ID NO:41; HCDR1, содержащий SEQ ID NO:9; LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:59; LCDR1, содержащий SEQ ID NO:57; а также
(ii) HCDR3, содержащий SEQ ID NO:13; HCDR2, содержащий SEQ ID NO:11; HCDR1, содержащий SEQ ID NO:9; LCDR3, содержащий SEQ ID NO:27; LCDR2, содержащий SEQ ID NO:25; LCDR1, содержащий SEQ ID NO:23.
В некоторых вариантах осуществления обеспечиваются антитела и их антигенсвязывающие фрагменты, которые связываются с цитокиновым рецептором IL-22R и перекрестно конкурируют с антителами или их антигенсвязывающими фрагментами, содержащими, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где область VH содержит:
CDR3 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [PPGPFKAHYNGAKY], SEQ ID NO:43,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [GIHISGGITYYTDSVKG], SEQ ID NO:41,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [SYFMS], SEQ ID NO:9,
и область VL содержит:
CDR3 вариабельной области легкой цепи, содержащий или состоящий из аминокислотной последовательности [ASYRLYADYV], SEQ ID NO:27,
CDR2 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [EVNKRSS], SEQ ID NO. 59,
CDR1 вариабельной области тяжелой цепи, содержащий или состоящий из аминокислотной последовательности [TGTSSDIGSYNYVS], SEQ ID NO:57.
Перекрестно конкурирующие антитела или их антигенсвязывающие фрагменты могут конкурировать с антителами или антигенсвязывающими фрагментами, имеющими область VH, содержащую или состоящую из последовательности SEQ ID NO:65, и/или область VL, содержащую или состоящую из последовательности SEQ ID NO:66. Перекрестно конкурирующие антитела или их антигенсвязывающие фрагменты могут конкурировать с антителами, имеющими полноразмерную тяжелую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:69, и/или полноразмерную легкую цепь иммуноглобулина, содержащую или состоящую из аминокислотной последовательности SEQ ID NO:70.
В контексте настоящего изобретения перекрестно конкурирующие антитела представляют антитела, которые связываются с IL-22R в сайте(сайтах), которые перекрываются или идентичны сайту (сайтам), в котором связываются анти-IL-22R-антитела по настоящему изобретению. Конкурирующие (моноклональные) антитела или их антигенсвязывающие фрагменты могут быть идентифицированы, например, с помощью конкурентного анализа антител. Например, антиген IL-22R или его фрагмент можно связать с твердой подложкой. Затем добавляют антитело или его антигенсвязывающий фрагмент по настоящему изобретению и моноклональное антитело или его антигенсвязывающий фрагмент, которые предположительно могут конкурировать с антителом по изобретению. Одна из двух молекул мечена. Если меченое соединение и немеченое антитело связываются с разными и дискретными сайтами на антигене IL-22R, то меченое соединение будет связываться с тем же уровнем независимо от присутствия или отсутствия предполагаемого конкурирующего соединения. Однако, если сайты их взаимодействия идентичны (или перекрываются), то немеченое соединение будут конкурировать, и количество меченого соединения, связанного с антигеном, будет уменьшаться. Если немеченое соединение присутствует в избытке, то очень небольшое количество, если таковое будет, меченого соединения будет связываться. Для целей настоящего изобретения конкурирующими моноклональными антителами или их антигенсвязывающими фрагментами являются те, которые уменьшают связывание антител по настоящему изобретению с IL-22R примерно на 50%, примерно 60%, примерно 70%, примерно 80%, примерно 85%, примерно 90%, примерно 95% или примерно 99%. Подробная информация о процедурах проведения таких конкурентных анализов хорошо известна в данной области и может быть найдена, например, в монографии Harlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, pages 567-569, ISBN 0-87969-314-2. Такие анализы могут быть выполнены в количественном варианте с использованием очищенных антител. Стандартная кривая строится титрованием одного антитела против самого себя, то есть одно и то же антитело используется в качестве меченого антитела и его конкурента. Титруют способность немеченого конкурентного моноклонального антитела или его антигенсвязывающего фрагмента ингибировать связывание меченой молекулы с планшетом. По результатам строят график, и сравниваются концентрации, необходимые для достижения желаемой степени ингибирования связывания.
Полинуклеотиды, кодирующие анти-IL-22R-антитела
Изобретение также обеспечивает полинуклеотидные молекулы, кодирующие анти-IL-22R-антитела по изобретению или их фрагменты, и также экспрессионные векторы, содержащие указанные нуклеотидные последовательности по изобретению, функционально связанные с регуляторными последовательностями, которые обеспечивают экспрессию антител или их фрагментов в клетке-хозяине или бесклеточной экспрессионной системы, и клетку-хозяин или бесклеточную экспрессионную систему, содержащие этот экспрессионный вектор.
В конкретных вариантах осуществления полинуклеотид, кодирующий анти-IL-22R-антитело по изобретению, может содержать одну или более полинуклеотидных последовательностей, показанных в SEQ ID NO:52, 73, 74, 75, 76, 77, 78, 79, 80 или 81, где такие последовательности кодируют области VH или VL анти-IL-22R- антител.
В некоторых вариантах осуществления полинуклеотид, кодирующий анти-IL-22R-антитело по изобретению, может содержать вариантную последовательность, которая кодирует функциональную область VH или VL анти-IL-22R-антитела, где указанная вариантная последовательность обладает, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичностью последовательности при оптимальном выравнивании с любой из SEQ ID NO:52, 73, 74, 75, 76, 77, 78, 79, 80 или 81.
В этом контексте процентную идентичность последовательности между двумя полинуклеотидными последовательностями можно определить сравнением этих двух последовательностей, выравненных оптимальным образом и в которых полинуклеотидная последовательность, подлежащая сравнению, может содержать добавления или делеции относительно эталонной последовательности для оптимального выравнивания между этими двумя последовательностями.
Процент идентичности вычисляется определением числа идентичных положений, для которых нуклеотидный остаток идентичен между двумя последовательностями, делением этого числа идентичных положений на общее число положений в окне сравнения и умножением полученного результата на 100, с получением процента идентичности между этими двумя последовательностями. Например, можно использовать программу BLAST, «BLAST 2 sequences» (Tatusova and al., «Blast 2 sequences - a new tool for comparing protein and nucleotide sequences», FEMS Microbiol Lett. 174:247-250), доступной на сайте http://www.ncbi.nlm.nih.gov/ gorf/bl2.html, с параметрами по умолчанию (в частности, для параметров «штраф за создание пробела»: 5 и «штраф за удлинение пробела»: 2; где выбранной матрицей является, например, матрица «BLOSUM 62», предложенная программой), процент идентичности между двумя последовательностями, которые требуется сравнить, вычисляется непосредственно программой.
В некоторых вариантах осуществления вариабельная область тяжелой цепи и вариабельная область легкой цепи анти-IL-22R-антитела или антигенсвязывающих фрагментов по настоящему изобретению кодируются комбинацией первой и второй полинуклеотидных последовательностей, где первая и вторая полинуклеотидные последовательности выбраны из следующих пар:
(i) первый полинуклеотид, кодирующий вариабельную область тяжелой цепи, содержащий SEQ ID NO:52, и второй полинуклеотид, кодирующий вариабельную область легкой цепи, содержащий SEQ ID NO:73;
(ii) первый полинуклеотид, кодирующий вариабельную область тяжелой цепи, содержащий SEQ ID NO:74, и второй полинуклеотид, кодирующий вариабельную область легкой цепи, содержащий SEQ ID NO:75;
(iii) первый полинуклеотид, кодирующий вариабельную область тяжелой цепи, содержащий SEQ ID NO:76, и второй полинуклеотид, кодирующий вариабельную область легкой цепи, содержащий SEQ ID NO:77;
(iv) первый полинуклеотид, кодирующий вариабельную область тяжелой цепи, содержащий SEQ ID NO:78, и второй полинуклеотид, кодирующий вариабельную область легкой цепи, содержащий SEQ ID NO:79; или
(v) первый полинуклеотид, кодирующий вариабельную область тяжелой цепи, содержащий SEQ ID NO:80, и второй полинуклеотид, кодирующий вариабельную область легкой цепи, содержащий SEQ ID NO:81.
Полинуклеотидные молекулы, кодирующие антитела по изобретению, включают, например, молекулы рекомбинантной ДНК. Термины «нуклеиновая кислота», «полинуклеотид» или «полинуклеотидная молекула», используются в настоящем описании взаимозаменяемо и относятся к любой молекуле ДНК или РНК, одно- или двухцепочечной и в случае одноцепочечной молекулы ее комплементарной последовательности. При рассмотрении молекул нуклеиновой кислоты последовательность или структура конкретной молекулы нуклеиновой кислоты могут быть описаны в настоящем описании в соответствии с общепринятыми правилами представления последовательности в направлении от 5' до 3'. В некоторых вариантах осуществления изобретения нуклеиновые кислоты или полинуклеотиды являются «выделенными». Данный термин, применяемый к молекуле нуклеиновой кислоты, относится к молекуле нуклеиновой кислоты, которая отделена от последовательностей, с которыми она непосредственно является смежной в природном геноме организма, в котором она находится. Например, «выделенная нуклеиновая кислота» может включать молекулу ДНК, вставленную в вектор, такой как плазмида или вирусный вектор, или встроенную в геномную ДНК прокариотической или эукариотической клетки или организма-хозяина, отличного от человека. Применительно к РНК термин «выделенный полинуклеотид» означает главным образом молекулу РНК, кодированную выделенной молекулой ДНК. Альтернативно, данный термин может означать молекулу РНК, которая была очищена/отделена от других нуклеиновых кислот, с которыми данная РНК ассоциирована в природном состоянии (то есть в клетках или тканях). Выделенный полинуклеотид (ДНК или РНК) может также представлять молекулу, полученную биологическими или синтетическими методами и отделенную от других компонентов, присутствующих в процессе получения.
Для рекомбинантной продукции антитела по изобретению рекомбинантный полинуклеотид, кодирующий его, может быть получен (с использованием стандартных методов молекулярной биологии) и вставлен в реплицируемый вектор для экспрессии в выбранной клетке-хозяине или бесклеточной экспрессионной системе. Подходящими клетками-хозяевами могут быть прокариотические, дрожжевые клетки или высшие эукариотические клетки, в частности, клетки млекопитающих. Примерами пригодных клеточных линий млекопитающих являются линия CV1 клеток почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol., 36: 59 (1977)); клетки почки новорожденного сирийского хомячка (BHK, ATCC CCL 10); клетки/-DHFR яичника китайского хомячка (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA, 77: 4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod., 23: 243-251 (1980)); клетки мышиной миеломы SP2/0-AG14 (ATCC CRL 1581; ATCC CRL 8287) или NS0 (номер в коллекциях культур HPA 85110503); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы линии buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и линия клеток гепатомы человека (Hep G2), а также DSM линия клеток PERC-6. Экспрессионные векторы, подходящие для применения в каждой из этих клеток-хозяев, также хорошо известны в данной области.
Следует отметить, что термин «клетка-хозяин» обычно относится к культивируемой клеточной линии. Весь организм человека, в который был введен экспрессионный вектор, кодирующий антигенсвязывающий полипептид по изобретению, однозначно исключается из определения «клетка-хозяин».
Продукция антител
В важном аспекте изобретение также относится к способу получения антител по изобретению, который включает культивирование клетки-хозяина (или бесклеточной системы экспрессии), содержащей полинуклеотид (например, экспрессионный вектор), кодирующий антитело, в условиях, обеспечивающих экспрессию антитела, и выделение экспрессированного антитела. Данный процесс рекомбинантной экспрессии может быть использован для крупномасштабного производства антител, включая анти-IL-22R-антитела по изобретению, включая моноклональные антитела, предназначенные для терапевтического применения человеку. Подходящие векторы, клеточные линии и способы получения для крупномасштабного производства рекомбинантных антител, подходящих для терапевтического применения in vivo, обычно доступны в данной области и хорошо известны специалистам в данной области.
Терапевтическая применимость анти-IL-22R-антител
Анти-IL-22R-антитела, обеспеченные в настоящем описании, можно использовать в качестве лекарственных средств, в частности для применения в лечении или профилактике расстройств, где патология обусловлена нарушенной передачей сигналов через комплексы IL-22R клеточной поверхности. Такая нарушенная передача сигналов может быть связана со сверхэкспрессией или избыточной продукцией любого из цитокинов IL-22, IL-20 и/или IL-24.
Термин «лечение» или «проводить лечение» означает замедление, прерывание, прекращение, контролирование, остановку, снижение тяжести симптома, расстройства, патологического состояния или заболевания, но не обязательно включает полное устранение всех связанных с заболеванием симптомов, патологических состояний или расстройств. Термин «профилактика» означает предупреждение развития расстройства, патологического состояния или заболевания или предупреждение начала развития симптомов, связанных с расстройством, патологическим состоянием или заболеванием.
Лиганды, которые передают сигналы через IL-22R, вовлечены в развитие ряда заболеваний, и поскольку IL-22R избирательно экспрессируется в коже и эпителиальных клетках, основными заболеваниями являются те, которые поражают кожу и эпителий, включая, не ограничиваясь этим, псориаз, псориатический артрит и атопический дерматит. Высокие уровни IL-22 были обнаружены в псориатических бляшках человека (Boniface et al., Clin. Exp. Immunol., 150: 407-415 (2007)), и вовлечение этого цитокина в патогенез псориаза было экспериментально продемонстрировано на мышиных моделях воспаления кожи (Ma et al., J. Clin. Invest., 118: 597-607 (2008), Van Belle et al., J. Immunol. Jan 1, 188 (1): 462-9 (2012)).
В некоторых вариантах осуществления обеспечиваются способы лечения воспалительных заболеваний кожи. В некоторых вариантах осуществления обеспечиваются способы лечения воспалительных заболеваний кожи, выбранных из псориаза, псориатического артрита, контактного дерматита или атопического дерматита у человека. Способы включают введение пациенту, нуждающемуся в этом, терапевтически эффективного количества любого из анти-IL-22R-антител или антигенсвязывающих фрагментов, как определено в настоящем описании в других местах. Все варианты осуществления анти-IL-22R-антител или антигенсвязывающих фрагментов, как в настоящем описании описано, одинаково применимы к способам лечения по настоящему изобретению.
В некоторых вариантах осуществления обеспечиваются способы лечения синдрома Шергена или рака, выбранного из гепатокарциномы, липосаркомы, плоскоклеточной карциномы ротовой полости, рака ободочной кишки и колоректального рака, рака поджелудочной железы, мелкоклеточного и крупноклеточного рака легких, рака молочной железы, глиобластомы, T-клеточной лимфомы кожи, анапластической крупноклеточной лимфомы, лимфомы из клеток мантийной зоны у человека, которые включают введение пациенту, нуждающемуся в этом, терапевтически эффективного количества любого из анти-IL-22R-антител или антигенсвязывающих фрагментов, как в настоящем описании определено в других местах. Все варианты осуществления анти-IL-22R-антител или антигенсвязывающих фрагментов, как в настоящем описании описано, одинаково применимы к способам лечения по настоящему изобретению.
Для терапевтического применения человеку анти-IL-22R-антитела, описанные в настоящем описании, можно вводить человеку, нуждающемуся в лечении, в «эффективном количестве». Термин «эффективное количество» относится к количеству или дозе анти-IL-22R-антитела, которое при однократном или многократном введении пациенту обеспечивает терапевтическую эффективность в лечении заболевания. Терапевтически эффективные количества анти-IL-22R-антитела могут включать количество в диапазоне от примерно 0,1 мг/кг до примерно 20 мг/кг на одну дозу. Количество антитела, вводимого в любую заданную временную точку, может варьироваться таким образом, чтобы оптимальное количество анти-IL-22R-антитела, независимо от того, используется оно в отдельности или в комбинации с любым другим терапевтическим агентом, вводилось во время курса лечения.
Также предусматривается введение анти-IL-22R-антител, описанных в настоящем описании, или фармацевтических композиций, содержащих такие антитела, в комбинации с любым другим подходящим лечением для вышеуказанных заболеваний, в виде комбинированной терапии.
Фармацевтические композиции
В объем изобретения входят фармацевтические композиции, содержащие одно или комбинацию анти-IL-22R-антител по изобретению или их антигенсвязывающие фрагменты, формулированные с одним или более фармацевтически приемлемыми носителями или эксципиентами. Такие композиции могут включать одно или комбинацию (например, двух или более разных) анти-IL-22R-антител. Способы формуляции моноклональных антител для терапевтического применения человеку хорошо известны в данной области, и их обзор имеется, например, в публикации Wang et al., Journal of Pharmaceutical Sciences, Vol.96, pp1-26, 2007, содержание которой в полном объеме включено в настоящем описании посредством ссылки.
В некоторых вариантах осуществления фармацевтические композиции формулированы для введения субъекту любым подходящим путем введения, включая, не ограничиваясь этим, внутримышечную, внутривенную, внутрикожную, внутрибрюшинную инъекцию, подкожное, эпидуральное, интраназальное, пероральное, ректальное, местное, ингаляционное, буккальное (например, сублингвальное) и чрескожное введение. В предпочтительных вариантах осуществления композиция формулирована для подкожного введения.
Включение посредством ссылки
В вышеприведенном описании и в следующих примерах упоминаются различные публикации, каждая из которых включена в настоящее описание посредством ссылки в полном объеме.
Примеры
Изобретение будет дополнительно пояснено со ссылкой на следующие неограничивающие примеры.
Пример 1. Иммунизация ламы
Двух лам иммунизировали смесью рекомбинантного белка IL22R человека (R & D systems) и человеческого Fn14-Fc согласно схеме иммунизации, показанной в таблице 5 ниже. После шести инъекций рекомбинантных белков раз в неделю отбирали образцы крови и сыворотки иммунизированных лам использовали для оценки гуморального иммунного ответа против IL22R по детектированию присутствия антител против иммунизирующего антигена до иммунизации и после иммунизации. Обе ламы генерировали значительный и специфический иммунный ответ против IL22R.
Схема иммунизации для каждой ламы
+Fn14-Fc (80мкг)
+Fn14-Fc (40мкг)
+Fn14-Fc (40мкг)
+Fn14-Fc (40мкг)
+Fn14-Fc (40мкг)
+Fn14-Fc (40мкг)
10 мл иммунной крови (плазмы)
Примеры 2. Конструирование и селекция библиотек
После иммунизации собирали PBMC и экстрагировали РНК. Проводили синтез случайной праймированной кДНК и с использованием ПЦР амплифицировали сегменты генов VHCH1, VλCλ и VκCκ ламы. Для ПЦР-амплификации легких цепей использовали два подхода. В первом подходе использовали первичную ПЦР-амплификацию с праймерами без линкеров с сайтами рестрикции с последующей ПЦР-амплификацией с праймерами с сайтами рестрикции (ApaLI и AscI). Для второго подхода использовали меченые праймеры для амплификации кДНК непосредственно. Библиотеки VHCH1 конструировали с использованием двухступенчатой ПЦР, в которой проводили 25 циклов с немечеными праймерами, и затем 10 циклов с использованием меченых вариантов этих праймеров (содержащих сайты рестрикции SfiI и NotI) (см. WO2010/001251).
Амплифицированные ПЦР легкие цепи расщепляли ApaLI и AscI, в то время как амплифицированные ПЦР тяжелые цепи расщепляли SfiI и NotI и объединяли в библиотеку Fab клонированием вставки библиотеки легкой цепи в вектор pCB3 библиотеки тяжелой цепи с использованием сайтов рестрикции ApaLI и AscI. Было установлено, что конечные библиотеки Fab имеют требуемое разнообразие > 109 различных клонов.
После конструирования библиотеки продуцировали фаги, и селекцию с использованием фагового дисплея выполняли на IL22R человека (Biotechne, 2770-LR). Для селекции IL22R-специфических клонов человеческий IL22R (i) наносили непосредственно на планшет MaxiSorp™ (Nunc); или (ii) захватывали неконкурентным антителом против человеческого IL22R (MAB2770, R & D systems); или (iii) захватывали нейтравидином после биотинилирования. Покрытие и захват IL22R обычно проводили в двух разных концентрациях; например, 5 мкг/мл и 0,1 мкг/мл.
После первого раунда селекции не было видно четкого обогащения, но после второго и третьего раундов наблюдали дозозависимые обогащения во всех библиотеках; до 100 раз после второго раунда и в 10000 раз после третьего раунда. Было обнаружено, что обогащения на захваченном антителом IL22R выше по сравнению с обогащениями с прямым покрытием IL22R. Это может быть за счет того, что IL22R составляет всего 25 кДа, и прямое покрытие влияет на его конформацию или ограничивает доступный эпитоп. Поэтому проводили третий раунд селекции с использованием биотинилированного hIL22R.
В дополнение к вышеописанным селекциям выполняли два дополнительных раунда селекции на биотинилированном рекомбинантном мышином IL22R после первого раунда селекции на MAB2770-захваченном человеческом IL-22R. Данный отбор проводили для определения перекрестной реактивности Fab с мышиным IL22R. В качестве положительного контроля селекции параллельно проводили отбор на человеческом IL-22R-биотине, захваченном нейтравидином. В целом процесс селекции был очень успешным со значительным обогащением, наблюдаемым для IL22R-связывающих Fab.
Пример 3. Скрининг на IL22R-специфические Fab
После успешной селекции на основе фагового дисплея Fab, присутствующие в периплазме, тестировали на их способность блокировать связывание IL-22 с IL22R. Кроме того, проводили скрининг для определения связывания клона с мышиным IL22R1. Был разработан новый подход, основанный на поверхностном плазмонном резонансе (SPR), который позволил тестировать в одном скрининге конкуренцию за лиганд, аффинность к человеческому IL22R и перекрестную реактивность к мышиному IL22R.
Различные каналы чипа Biacore (CM5) покрывали:
1. Ничем (холостой контроль)
2. IL-22 (3000 RU) для тестирования на конкурентную активность Fab (когда IL22R вводится совместно)
3. Человеческим IL22R (3000 RU) для тестирования на связывание с мишенью
4. Мышиным IL22R (2500RU) для тестирования на перекрестную реактивность.
Для тестирования конкуренции за лиганд перед инжектированием предварительно смешивали периплазматические экстракты с низкой концентрацией растворимого IL22R1 (0,2 мкг/мл). Таким образом, также можно было измерить способность Fab блокировать взаимодействие лиганд-рецептор в канале 2. Полученные периплазматические экстракты подвергали скринингу на связывание с человеческим IL22R (канал 3) и мышиным IL22R (канал 4).
Преимущество данного метода по сравнению с обычным ELISA (связывающим или конкурентным) заключается в том, что одновременно тестируются несколько характеристик Fab: 1) связывание с целевым человеческим IL-22R; 2) конкурентная активность; и 3) перекрестная реактивность. Кроме того, показатель скорости диссоциации дает четкое указание на аффинность тестированных Fab.
Используя данный альтернативный метод скрининга, Fab со всеми возможными характеристиками были идентифицированы и распределены по категориям на основе значения их скорости диссоциации:
• конкурирующие или не конкурирующие
• перекрестно реагирующие и не реагирующие перекрестно
• конкурирующие и перекрестно реагирующие
• неконкурирующие и перекрестно реагирующие
Затем различные клоны подвергали секвенированию.
Пример 4. Анализ последовательности IL-22R Fab
Клоны, идентифицированные анализом Biacore (например, конкуренция за лиганд, максимальное связывание, низкая скорость диссоциации или перекрестная реактивность с мышиным белком), были секвенированы. Затем клоны были сгруппированы на основе идентичности их CDR3 с образованием семейств VH.
В целом было идентифицировано 67 разных VH, относящихся к 13 разным семействам VH. Было идентифицировано значительно больше семейств легких цепей (более 80 последовательностей Vлямбда и Vкаппа). Большое разнообразие в последовательности и функции Fab показывает, что иммунизация и селекция были очень успешными.
Аминокислотные последовательности областей VH и VL клонов, представляющих особый интерес, показаны ниже в таблицах 6-8, и полинуклеотидные последовательности этих клонов показаны в таблице 9.
Последовательности каркасных областей и CDR VH-областей IL-22R Fab
Последовательности каркасных областей и CDR VL-областей IL-22R Fab
Последовательности вариабельной области IL-22R Fab
Полинуклеотидные последовательности IL-22R Fab
ACCGTCCTG
Пример 5. Характеристика Fab
Клоны с наилучшими значениями скорости диссоциации реклонировали в pCB4, экспрессировали и очищали с помощью IMAC (Talon от Clonentech). Данные клоны снова тестировали с использованием анализа Biacore. Результаты дальнейшей характеристики Fab приведены в таблице 10 ниже.
Характеристика связывания IL-22R Fab
VH
IL-22
В зависимости от разнообразия в семействе VH, конкурирующей активности и аффинности (измеренных с использованием значения koff, полученного в Biacore), отбирали один или более клонов из каждого семейства и переформатировали в моноклональные антитела (mAb) для дальнейшей характеристики.
Пример 6. Характеристика IL-22R-связывающих mAb
Как отмечено выше, отобранные клоны каждого семейства VH получали в виде IgG1. После реклонирования вариабельных областей HC и LC в отдельные экспрессионные векторы pUPE, содержащие константные области человека, клетки HEK293E транзиентно трансфицировали двумя плазмидами, одна из которых кодирует полную тяжелую цепь, и вторая кодирует легкую цепь. Трансфицированным клеткам давали возможность экспрессировать антитела в течение 6-суточного периода культивирования клеток. После выделения антител из клеточного культурального супернатанта с использованием магнитных шариков с иммобилизованным белком А очищенные антитела тестировали на их способность нейтрализовать сигнальный путь человеческого и мышиного IL-22R с использованием различных клеточных линий, как описано ниже.
Клеточная линия BW-hIL22R: получена из клеточной линии BW и имеет стабильную экспрессию человеческого IL22R, что обеспечивает IL22-зависимую задержку роста. Следовательно, клетки будут пролиферировать только тогда, когда блокируется взаимодействие IL22-IL22R. Активное антитело будет способствовать пролиферации в низкой концентрации антитела. Пролиферация измеряется по включению тимидина.
Клеточная линия Baf3-mIL22R: получена из клеточной линии Baf3 и имеет стабильную экспрессию мышиного IL22R, что обеспечивает IL22-зависимую пролиферацию. Антитело, нейтрализующее mIL22R, будет блокировать пролиферацию зависимым от концентрации антител образом.
Клеточная линия Baf3-hIL22R/IL20Rb: получена из клеточной линии Baf3, но коэкспрессирует hIL22R с hIL20Rbeta, позволяя клеткам пролиферировать в присутствии IL20. Антитело, нейтрализующее hIL22R, будет блокировать пролиферацию клеток, стимулированных IL20.
Результаты тестирования различных mAb к IL22R против клеточной линии BW-hIL22R и клеточной линии Baf3-hIL22R/IL20Rb приведены на фиг.4 и в таблице 11. На фиг.4А показано влияние нейтрализующих анти-IL-22R-антител на восстановление пролиферации клеточной линии BW-hIL22R. На фиг.4В можно видеть влияние нейтрализующих анти-IL-22R-антител на ингибирование IL-20 пролиферации клеточной линии, экспрессирующей IL-22R. Антитело к IL22R, описанное в WO 2011/061119 (280.346.TSY), было включено в эксперименты в качестве стандартного или эталонного антитела к IL-22R.
В дополнение к оценке активности на клетках измеряли кинетику связывания in vitro и блокирующую активность очищенных антител в анализе Biacore (3000). Для измерения аффинности рекомбинантный IL22R наносили на чип Biacore CM5 при 200 RU с использованием стандартного протокола покрытия (IL22R инжектируют в концентрации 10 мкг/мл в ацетатном буфере, рН 4,5). Антитела, испытанные в диапазоне концентраций и приготовленные в буфере HBSEP при pH 7,4 (Biacore), инжектировали при 30 мкл/мин в течение 60 с и промывали буфером HBSEP с pH 7,4 (Biacore) в течение 10-20 мин. Полученные сенсограммы анализировали с помощью программного обеспечения для оценки BIA, и кинетику определяли с использованием стандартного подбора кривой (уравнение Ленгмюра 1:1 с переносом масс).
Для оценки блокирующей активности с использованием анализа Biacore 3000 рекомбинантный IL22 (R& D System) наносили при 2000 RU на чип CM5. Антигенсвязывающие участки (очищенный IgG, очищенные Fab или периплазматический экстракт, содержащий Fab) предварительно инкубировали с рекомбинантным человеческим IL22R (концентрация от 0,2 до 1 мкг) перед инжектированием в канал, покрытый hIL22. Детектирование связывания (IL22R:IL22) указывало, что антигенсвязывающий участок не конкурировал, когда связывание не детектировали (IL22R:IL22), то это указывало на то, что антигенсвязывающий участок блокировал взаимодействие IL22:IL22R in vitro. Результаты анализа Biacore и пролиферации обобщены в таблице 11 ниже.
Обобщенные данные по характеристике панели анти-IL-22R-антител
VH
(константа диссоциации)
Антитела к IL22R 157A2 и 166G8 проявляют активность на пикомолярном уровне.
Пример 7. Видовая перекрестная реактивность mAb к IL-22R
mAb к IL22R тестировали на видовую перекрестную реактивность против мышиного IL-22R и IL-22R примата, отличного от человека.
кДНК для IL22R обезьяны циномолгус была недоступна, и ее необходимо было извлечь из библиотек кДНК обезьяны циномолгус. Праймеры были сконструированы на основе общедоступной базы данных NCBI. Несколько последовательностей для ECD cynoIL22R были доступны из общедоступных баз данных (Genbank, NCBI) и содержали инсерцию, делецию или и то и другое (см. фиг. 5). Неопределенность в отношении истинной последовательности IL-22R обезьяны циномолгус подверждали клонированием из кДНК обезьяны циномолгус.
ECD IL-22R обезьяны циномолгус амплифицировали с помощью ПЦР из библиотеки кДНК и клонировали в рамке считывания в направлении 5'-3' сигнального пептида IgGкаппа и апстрим человеческого Fc в вектор pUPE. Несмотря на то, что используемая кДНК была получена от обезьяны циномолгус, секвенирование нескольких клонов показало, что IL22R макака-резус также клонировался. IL22R-ECD макака-резус характеризовался отсутствием делеции и вставки, обнаруженных у обезьяны циномолгус (в рамке на фиг. 5), и двумя аминокислотными различиями между IL22R макака-резус и обезьяны циномолгус (жирным шрифтом на фиг. 5).
После получения и очистки cyIL22R-Fc и rhIL22R-Fc тестировали специфичность связывания с IL-22R ECD обезьяны циномолгус, макака-резус и мыши всех антител из первичной селекции в ELISA и SPR (Biacore). Мономерный мышиный IL22R получали от R & D Systems (номер по каталогу 4248-MR). Результаты обобщены в таблице 12.
Видовая перекрестная реактивность mAb к IL-22R
Пример 8. Картирование эпитопа
Для идентификации и сравнения эпитопов, связанных с mAb IL22R, использовали два метода: конкурентный ELISA и FACS.
8.1. Картирование эпитопа с использованием конкурентного ELISA
Эпитопы, распознаваемые антителами, сравнивали друг с другом с помощью конкурентного ELISA. По меньшей мере, одно репрезентативное mAb из каждого семейства VH наносили на планшет Maxisorp. Затем добавляли биотинилированный человеческий IL22R в присутствии большого избытка mAb, предназначенных для тестирования. Связывание биотинилированного IL22R с покрытым антителом детектировали с использованием HRP-конъюгированного стрептавидина. Когда биотинилированный IL22R не детектировался, означая, что растворимое mAb предотвращало связывание с покрытым mAb, то было определено, что оба mAb связываются с одним и тем же эпитопом или имеют перекрывающийся эпитоп. Картирование эпитопов дополнительно уточняли с использованием Fab вместо mAb. Меньший по размеру Fab-фрагмент давал возможность различить очень близкие эпитопы.
Идентифицировали различные эпитопы для антител VH-семейств 1-8, 10, 11, 19 и 22. Картирование эпитопов показывает очень широкое покрытие эпитопов для такого небольшого белка (25 кДа). Как показано на фиг.6, эпитопы были сгруппированы в соответствии с тем, насколько ли антитела: (i) блокировали связывание IL-22 in vitro и нейтрализовали сигнальный путь IL-22 в клеточном анализе (нижний левый круг); (ii) блокировали связывание IL-22 in vitro, но не обладали нейтрализующей активностью (верхний правый круг); или (iii) не блокировали связывание IL-22 in vitro, но обладали нейтрализующей активностью в клеточном анализе (нижний правый круг). Было установлено, что шесть перекрывающихся, но различимых эпитопов блокируются и нейтрализуются (см. нижний левый круг), и из этих шести эпитопов два (mAb из семейства 4 и 22) имели перекрывающийся эпитоп со стандартным антителом (280.346.TSY, см. заявку на патент WO2011/061119). Эпитоп не был идентичен такому для стандарта, поскольку эти mAb (из семейства 4 и 22) были способны конкурировать также со всеми mAb из семейств 1, 5, 6 и 7, даже если они не могли конкурировать со стандартным антителом. В совокупности, эти данные позволяют предположить о наличии двух разных групп эпитопов, одной, сформированной стандартом, и одной, образованной mAb из семейств 1,5,6 и 7, и третьей группы (mAb из семейств 4 и 22), которая перекрывается с обеими группами (фиг. 6).
Как показано в верхнем правом круге на фиг. 6, было идентифицировано 3 антитела, которые связываются с IL22R, но не блокируют связывание IL22 или нейтрализуют активность in vivo. Эти антитела, вероятно, связываются с областями IL22R, не участвующими в связывании лиганда или активации передачи сигналов, и могут быть использованы просто для детектирования. 3 антитела, показанные в правом нижнем круге на фиг.6, необычны тем, что они нейтрализуют активность IL22R in vivo, но не блокируют IL22 in vitro, что предполагает наличие нового функционального эпитопа и неожиданного механизма действия.
8.2. Картирование эпитопа с использованием анализа FACS
Jones et al. (Structure 16 (9): 1333-1344 (2008)) сообщают о структуре IL-22, связанного с внеклеточным доменом IL22R1. В этой статье авторы показывают, что два домена IL22R, называемые D1 и D2, взаимодействуют с лигандом IL-22 в сайте 1A и сайте 1B, соответственно (см. фиг. 7). Два ключевых остатка в IL22R-D1, вовлеченных в прямое взаимодействие с сайтом 1A IL22, представляют собой лизин 58 (K58) и тирозин 60 (Y60). Триптофан 208 (W208) IL22R-D2 непосредственно участвует во взаимодействии с сайтом 1B IL22 (см. фиг. 7).
Тестировали связывание анти-IL-22R-антител и стандартного антитела (280.346.TSY) с клетками BW, сверхэкспрессирующими различные мутанты IL-22R. Мутанты IL22R содержат мутации аминокислот, о которых известно, что они участвуют во взаимодействии IL-22/IL22R. В данном аналитическом режиме антитела биотинилировали, добавляли к клеткам, и детектировали связывание антитела с IL22R меченым стрептавидином проточной цитометрией (FACS). В таблице 13 обобщены полученные результаты.
Связывание антител с клетками, экспрессирующими мутанты IL-22R
Результаты FACS четко показывают, что стандартное антитело 280.346.TSY взаимодействует с остатками домена D1 (K58 и Y60). То же самое имеет место для второго контрольного антитела к IL-22R, AMR22, хотя эпитоп не является полностью тем же самым, поскольку мутация R112A влияет на связывание AMR22. Как и предполагалось по результатам картирования эпитопов с использованием ELISA, показано, что некоторые антитела ламы не конкурируют с антителом 280.346.TSY, другие тестированные антитела не были затронуты мутациями, влияющими на связывание 280,346.TSY с IL22R. Это подтвердило, что антитела нацелены на другой эпитоп, возможно, включая остатки в домене D2. Фактически, мутация A209D в домене D2 IL22R элиминировала связывание 198A1, подтверждая, что 198A1 и антитело из семейства 5 связываются с доменом D2 IL22R (см. фиг. 7B).
Удивительно, но на антитело 166G8, которое имеет точно такую же VH, что и 198A1, но имеет другую VL, мутация A209D не оказывала влияния, что позволяет предположить, что VL участвует во взаимодействии. Возможно, VL 166G8 допускает наличие более крупного остатка в положении 209, в то время как VL 198A1 - нет. Эта гипотеза подтверждается их специфичностью связывания с IL22R макака-резус. В IL22R макака-резус имеет место серин в положении 209. Поскольку 166G8 допускает наличие более крупной аминокислоты в этом положении, то оно может связывать IL22R макака-резус, в то время как 198A1 не способно связывать rhIL22R, поскольку оно не допускает крупных остатков.
Пример 9. Нейтрализация сигнального пути IL20 и IL24 в клеточном анализе
Анти-IL22R-антитела тестировали на их способность блокировать IL-20- и IL-24-зависимую передачу сигналов в клетках, коэкспрессирующих IL22R и IL20Rb. Существует механистическое различие между IL-22-, IL-20- и IL-24-зависимой активацией рецептора. Полагается, что IL-22 индуцирует передачу сигналов, сначала связываясь с IL22R, запуская рекрутинг корецептора IL10R2 и активацию тримерного комплекса. Предполагается, что для IL-20 этот цитокин сначала связывается с корецептором (IL20R2) перед рекрутингом IL22R и активацией тримерного комплекса. Наконец, для IL-24 остается неясным, как рецепторный комплекс подвергается рекрутингу и активации, а также неясно, где связывается IL-24.
9.1. Сигнальный путь IL-24
Несколько клеточных линий использовали для тестирования IL-24-зависимой передачи сигналов, Baf3-hIL22R/20R2 или Baf3-mIL22R/20R2. IL-24 индуцировал пролиферацию этих клеточных линий со значительным окном, что определяли по уровню гексозаминидазы (данный показатель используется для подсчета клеток) в диапазоне OD от 0,17 до 1,5 для клеток Baf3-hIL22R/20Rb и в диапазоне от 0,15 до 0,8 для клеток Baf3-mIL22R/20Rb. Ни одно из тестированных антител не оказывало влияния на пролиферацию, индуцированную IL-24. Эти результаты предполагают, что либо IL-24 имеет совершенно другой механизм активации IL22R путем связывания с сайтом, который резко отличается от IL22 и IL-20, либо что IL22R не участвует в передаче сигналов IL-24 в тестированной клеточной линии (и анализ не прогнозирует нейтрализующую активность).
9.2. Сигнальный путь IL-20
Клеточная линия Baf3-hIL22R/IL20Rb происходит от клеточной линии Baf3, но коэкспрессирует hIL22R с hIL20Rβ, позволяя клеткам пролиферировать в присутствии IL-20. Антитело, нейтрализующее hIL22R, будет блокировать пролиферацию клеток, стимулированных IL-20. Результаты тестирования антител ламы вместе со стандартными антителами 280.346.TSY и AMR22 показаны на фиг. 8.
IL-20-зависимую пролиферацию в Baf3-hIL22R/IL20Rb тестировали параллельно с анализом IL-22-зависимой пролиферации в BW-hIL22R, что позволило провести адекватное сравнение между двумя цитокинами. Результаты трех независимых экспериментов приведены в таблице 14.
Активность антител в блокировании IL-22- и IL-20-зависимого сигнального пути через IL-22R
IL-22 (пM)
IL-20 (пM)
*Диапазон, наблюдаемый в трех экспериментах (2250, 2292, 2301 для IL-20 и 2253, 2292, 2301, 2428 для IL-22); nc: не конкурирует; nt: не тестировали; cs: переставленная цепь.
Можно сделать вывод, что все антитела из семейств 1, 4 и 5 были способны блокировать как IL-20, так и IL-22. Трудно сравнивать активность между IL-22- и IL-20-стимулированной пролиферацией за счет разницы в чувствительности, которая зависит от концентрации цитокинов, числа рецепторов и внутриклеточной биологической вариабельности. Однако было установлено, что большинство антител более эффективно блокируют IL-22-зависимую передачу сигналов по сравнению с блокированием IL-20-зависимой передачи сигналов.
Пример 10. Селекция примерных антител и превращение в антитела зародышевого типа
Аффинность антител, представляющих наибольший интерес (нейтрализация сигнального пути IL22), дополнительно повышали путем перестановки легких цепей. Во время перестановки легкой цепи VHCH1 отобранного антитела объединяют с библиотекой легкой цепи от той же ламы, из которой был получен VHCH. Затем проводили несколько раундов селекции с использованием фагового дисплея по скорости диссоциации для идентификации пар VHCH:VLCL, которые имеют наиболее высокую аффинность. В случае антитела 166G8 было обнаружено несколько клонов с улучшенной кинетикой связывания, в том числе 198A1, но, к сожалению, данный клон потерял большую часть своей перекрестной реактивности для макака-резуса, и поэтому антитело 166G8 осталось наиболее привлекательным кандидатом семейства 5 для дальнейшей разработки. В случае 157A2 (из семейства 4) было выявлено несколько Fab (например, 218A7) с улучшенной аффинностью по сравнению с 157A2. Следующие антитела были взяты для дальнейшей разработки. Данные антитела были отобраны на основе активности, видовой перекрестной реактивности и эпитопа или механизма действия.
218A7 (улучшенный вариант 157A2, семейство 4) - было установлено, что это антитело обладает очень хорошей активностью в отношении IL-22 и IL-20 (на субнаномолярном уровне), перекрестно реагирует с IL22R примата, отличного от человека, но не реагирует перекрестно с мышиным IL22R. Было обнаружено, что 157A2 имеет на IL22R эпитоп, отличный, но перекрывающийся со стандартным антителом 280.346.TSY.
166G8 (семейство 5) - было установлено, что это антитело обладает высокой активностью (на пикомолярном уровне) и обладает перекрестной реактивностью для некоторых видов, поскольку оно связывается с IL22R макака-резуса (но не с IL22R обезьяны циномолгус и не с мышиным IL22R).
Характеристики отобранных анти-IL-22R-антител приведены в таблице 15 ниже.
Характеристики анти-IL-22R-антител ламы, отобранных для превращения в антитела зародышевого типа
(пM)
(пM)
Пример 11. Превращение антител 218A7 и 166G8 в антитела зародышевого типа
Превращение антител в зародышевый тип проводили, как описано в международной патентной заявке WO2011/080350, содержание которой включено в настоящем описании в полном объеме посредством ссылки. CDR были мутированы для удаления потенциальных посттрансляционных модификаций вместе с некоторыми другими положениями, которые, как было установлено, различаются в антителах из одинаковых семейств VH и VL с идентичным спариванием. Области VH и VL антител зародышевого типа, полученных из антител ламы 218A7 и 166G8, оценивали на их идентичность с человеческими последовательностями. Также было проведено сравнение со стандартным антителом 280.346.TSY. Результаты приведены в таблице 16 ниже.
Идентичность анти-IL-22R-антитела зародышевого типа, полученных из антител 218A7 и 166G8, с антителом человека
Антитело 230C9 было отобрано в качестве варианта зародышевого типа антитела 218A7, и антитело 223G5 было отобрано в качестве варианта зародышевого типа антитела 166G8. Последовательности областей VH и VL антител 218A7, 230C9 и 223G5 показаны в таблицах 17-21 ниже (последовательности 166G8 показаны в таблицах 6-9 выше).
Последовательности каркасных областей и CDR для VH-областей антител 218A7, 230C9 и 223G5
Последовательности каркасных областей и CDR для VL-областей антител 218A7, 230C9 и 223G5
Последовательности вариабельных областей антител 218A7, 230C9 и 223G5
Последовательности тяжелой цепи и легкой цепи антител 218A7 и 223G5
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYQSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
GQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Затемненные участки последовательности представляют константные области тяжелой и легкой цепей. Остаток «Q», выделенный жирным шрифтом в константной области тяжелой цепи, представляет мутацию из «N».
Полинуклеотидные последовательности, кодирующие последовательности вариабельной области/последовательности тяжелой цепи и легкой цепи антител 218A7, 230C9 и 223G5
Пример 12. Характеристика вариантов зародышевого типа антител 230C9 и 223G5
12. 1. Аффинность связывания вариантов mAb зародышевого типа
Аффинность антител 230C9 и 223G5 измеряли с использованием SPR (Biacore 300). Вкратце, 250 RU человеческого IL22R (Biotechne, 2770-LR) наносили на чип CM5 с использованием стандартных методов (EDC-сочетание IL22R в концентрации 2 мкг/мл в ацетатном буфере, pH 4,5). Затем антитело в различных концентрациях инжектировали в HBSEP+ буфере (pH 7,4) в течение 2 мин. Связывание контролировали также во время 10-мин промывания (HBSEP+).
Аффинность, рассчитанная с использованием программного обеспечения BIAevaluation с использованием подбора кривой уравнением Ленгмюра 1:1, показана в таблице 22 ниже.
Аффинность связывания вариантов зародышевого типа антител
Антитела 223G5 и 224C4 (оба варианта зародышевого типа антитела 166G8) показали очень высокую аффинность к IL-22R (10-20 пМ), тогда как антитело 230C9 имело аффинность, равную 0,13 нМ. Все варианты зародышевого типа антитела показали более высокую аффинность, чем стандартное антитело 280.346.TSY (0,69 нМ).
За счет потенциального эффекта авидности, наблюдаемого при использовании бивалентной молекулы в качестве лиганда во время измерения аффинности, проводили тестирование в обратном экспериментальном режиме. В этом аналитическом режиме антитела покрывали до 250 RU с использованием стандартных методов. Мономерный человеческий IL22R добавляли в различных концентрациях, и значения аффинности, рассчитанные с использованием программного обеспечения BIAevaluation с использованием подбора кривой уравнением Ленгмюра 1:1, приведены в таблице 23 ниже.
Аффинность связывания антитела 230С9 с человеческим IL-22R
К сожалению, антитело 223G5 инактивировалось в процессе нанесения, и с помощью этого аналитического режима не удалось определить аффинность.
Аффинность антитела 230C9 является постоянной независимо от режима измерения аффинности, показывая, что низкое покрытие, используемое в обоих экспериментальных условиях, является достаточным для снижения любого эффекта авидности.
12.2. Ингибирование IL-22- и IL-20-зависимой активации и сигнального пути IL-22R1
Антитела 218A7 (варианты не зародышевого типа) и 230C9 (эквивалентный вариант зародышевого типа) тестировали вместе с другими вариантами зародышевого типа антител на их способность нейтрализовать IL-22- и IL-20-опосредованную передачу сигналов через IL-22R1. Тестирование проводили с использованием анализа клеточной пролиферации, описанного в примерах 6 и 9.
На фиг. 9A показана способность 218A7 и 230C9 нейтрализовать IL-22-зависимую активацию IL-22R1. На фиг. 9В показана активность различных серий 230C9 нейтрализовать IL-20-зависимую активацию IL-22R1. Относительные активности тестированных антител показаны в таблицах 24 и 25 ниже.
Активность антител 218A7 и 230C9 в блокировании IL-22-зависимого сигнального пути через IL22R (клеточная линия BWhIL22R)
IL-22 (нМ)
Активность антитела 230C9 в блокировании IL-20-зависимого сигнального пути через IL22R (клеточная линия Baf3hIL22R/IL20Rb)
IL-20 (нМ)
12.3. Подтверждающее картирование эпитопа
Подтверждающее картирование эпитопа для вариантов зародышевого типа антител 230C9 и 223G5 проводили с использованием анализа FACS, как описано в примере 8.2. Было установлено, что антитело 230C9 конкурирует за связывание с IL-22R1 со стандартным антителом 280.346.TSY, а также с антителом 223G5. Антитело 223G5 не конкурировало за связывание с IL-22R1 со стандартным антителом 280.346.TSY.
Результаты анализа FACS объединяли с результатами анализа пролиферации, описанными выше, и выводы, касающиеся эпитопов, приведены на фиг.10. Как указано, можно сделать вывод о том, что: (i) антитело 230C9 связывается с отличающимся, но перекрывающимся эпитопом по сравнению с эпитопами, которые связываются 280.346.TSY; и (ii) антитело 223G5 связывается с эпитопом, отличным от эпитопа для 280.346.TSY.
Анализ FACS также проводили, как описано в примере 8.2, для исследования связывания вариантов зародышевого типа антител с различными мутантами IL-22R. Результаты приведены в таблице 26 ниже.
Связывание антител с клетками, экспрессирующими мутанты IL-22R
Полученные результаты подтверждают, что антитела 230C9 и 223G5 связываются с эпитопом, который не содержит Y60. Было установлено, что мутация A209D оказывает влияние на связывание 223G5, тогда как на связывание антитела ламы 166G8 это изменение не оказывало влияния. Вероятно, это связано с трудностями определения чувствительности 166G8 к мутации A209D (166G8 тестировали только один раз) и, возможно, связано с незначительными изменениями в последовательностях CDR во время превращения в вариант зародышевого типа 166G8. Полученные данные подтверждают, что антитело 280.346.TSY связывается с D1 IL22R, что антитело 223G5 связывается с D2 IL22R и что 230C9 имеет перекрывающийся, но отличный эпитоп.
12.4. Видовая перекрестная реактивность вариантов зародышевого типа антител
Антитело 230C9 связывается с человеческим IL-22R, но не проявляет перекрестной реактивности для IL-22R мыши, крысы и кролика. Это согласуется с тем, что 230C9 имеет отличный эпитоп по сравнению со стандартным антителом 280.346.TSY - см. п. 12.3 выше.
Способность антител связываться с IL22R обезьяны циномолгус, экспрессированным на клетках, тестировали созданием химерных конструкций внеклеточного домена (ECD) IL22R из различных видов, слитого с человеческим трансмембранным (TM) и человеческим внутриклеточным доменами (ID). Несмотря на то, что эффективность транзиентной трансфекции была невысокой, результаты анализа FACS показали, что антитело 230C9-N297Q, но не 223G5-N297Q, связывается с cynoIL22R_ECD-человеческийTM_человеческийICD, о чем судили по наличию более широкого пика на графике FACS (см. фиг. 11 со стрелками, указывающими на более широкие пики).
Кроме того, когда химерные конструкции также котрансфицировали с репортерным геном STAT3 в клетки HEK, то активность IL22R после добавления человеческого IL22 можно было измерять в присутствии или в отсутствие нейтрализующего антитела. Результаты, представленные в таблице 27 ниже, показывают, что все тестированные антитела были способны нейтрализовать человеческий IL22R и что только перекрестно реагирующие с приматами (230C9 и 280.346.TSY) были способны связываться и блокировать активность химер обезьяна циномолгус-макак-резус, что свидетельствует о том, что эти антитела будут функциональными в исследованиях на приматах.
Перекрестная реактивность анти-IL-22R-антител с IL-22R человека, макака-резус и обезьяны циномолгус
ECD человека
ECD макака-резус
ECD обезьяны циномолгус
12.5. Анализ иммуногенности
Иммуногенность вариантов зародышевого типа антител оценивали с использованием инструментов прогнозирования. В частности, наличие потенциальных иммуногенных пептидов в вариабельных областях оценивали с использованием платформы Epibase™ (LRZ-1) от lonza с использованием настроек «HLA class II - Caucasian v3.0». Данная платформа анализирует специфичность связывания HLA всех 10-мерных пептидов, полученных из последовательностей VH и VL. Профилирование выполняли на уровне аллотипа для 15 DRB1, 6 DRB3/4/5, 12 DQ и 7 DP, т.е. всего 40 рецепторов HLA класса II. Были идентифицированы сильные и средние связывающие антитела для DRB1, DRB3/4/5, а также сильные связывающие антитела для эпитопов DQ и DP. Подсчет эпитопов проводили отдельно для высокоаффинных и среднеаффинных DRB1-связывающих антител. Пептиды, связывающиеся с несколькими аллотипами одной и той же группы, определялись как один. Приблизительный балл, выражающий наихудший иммуногенный риск, рассчитывали следующим образом: балл = ∑ (число эпитопов × частота аллотипов). Другими словами, число эпитопов, влияющих на конкретный аллотип HLA, умножается на частоту аллеля затронутого аллотипа. Для данной последовательности продукты суммировали для всех аллотипов DRB1, использованных в исследовании, которые присутствуют в 2% или более популяции субъектов европейской национальности.
Результаты приведены в таблице 28 ниже.
Баллы DRB1 для вариантов зародышевого типа анти-IL-22R-антител по сравнению с различными другими коммерчески доступными антителами
Как следует из таблицы 28, антитело 230C9 имеет очень низкий балл DRB1, значительно ниже чем у других коммерчески доступных антител, тогда как антитело 223G5 имеет низкий балл DRB1, сравнимый с другими коммерчески доступными человеческими или гуманизированными антителами. Эти данные предполагают, что антитело 230C9 (и 223G5) будут иметь предпочтительный профиль иммуногенности.
Пример 13. Фармакокинетическое исследование (PK) антитела 230C9-N297Q на обезьянах циномолгус
Проводили фармакокинетический анализ клона антител 230C9. Двум обезьянам циномолгус внутривенно вводили однократную дозу 10 мг/кг антитела в течение 2 ч. Образцы крови отбирали на различные временные точки и определяли концентрацию mAb в плазме с помощью ELISA. В частности, микротитрационный планшет (Maxisorb Nunc) покрывали рекомбинантным hIL22R 2 мкг/мл (Biotechne; номер по каталогу 2770-LR) в PBS в течение ночи при 4°C. Планшет промывали 3 раза смесью PBS-твин и блокировали в течение 2 ч 250 мкл PBS-1% казеина. После 3 промываний смесью PBS-твин наносили образцы. Все разведения готовили на 1% пулированной плазме (это пул от 3 нативных обезьян циномолгус). Образцы оставляли для связывания на 2 ч при комнатной температуре. Затем планшеты 5 раз промывали смесью PBS-твин и поликлональные антитела с адсорбированными тяжелой и легкой цепями козьего биотинилированного античеловеческого IgG, связанные с HRP, наносили в 50000-кратном разведении (Bethyl, номер по каталогу: A80-319P) и оставляли для связывания на 1 ч при комнатной температуре. Затем планшеты 5 раз промывали смесью PBS-твин и добавляли реагент s(HS)ослабитель TMB (SDT, # sTMB-W). Окрашивание проводили в течение 10 мин и затем останавливали добавлением 1 Н серной кислоты, после чего измеряли оптическую плотность при 450 нм. Образцы анализировали 3 раза, и 230C9-N297Q (из той же партии, которую вводили животным) использовали для построения стандартной кривой.
Соответствующие фармакокинетические параметры для анализа двухкомпартментной модели при в/в инфузии приведены в таблице 29. Фармакокинетические профили антитела 230C9-N297Q показаны графически на фиг. 12 (приведенный результат является средним значением для двух обезьян). Эти данные четко показывают, что антитело имеет длительное среднее время удерживания (MRT). Интересно, что хотя антитело содержит негликозилированную Fc-область IgG1 без каких-либо модификаций, обеспечивающих повышение периода полураспада, период полураспада 230C9-N297Q неожиданно был высоким. В частности, 230C9-N297Q имеет период полураспада, равный примерно 19,4 суток. Таким образом, по-видимому, увеличенный период полураспада антител по изобретению обусловлен свойствами Fab-областей.
Фармакокинетические параметры
Дальнейшее фармакокинетическое исследование проводили на обезьянах циномолгус при однократном внутривенном введении 230C9-N297Q в дозах (0,1; 1; 3; 10 и 30 мг/кг) в целом 10 обезьянам. Кинетика у обезьян циномолгус демонстрирует двухкомпартментную нелинейную кинетику. В дозах ниже 10 мг/кг клиренс (CL) увеличивался с уменьшением дозы, что приводило к пропорциональному снижению биодоступности (AUC) с уменьшением дозы. Это хорошо известное явление для моноклональных антител (см. таблицу 30 и фиг. 13).
Фармакокинетические параметры у 2 обезьян/дозу и в 5 группах с введением разных доз
Масштабирование этих параметров для прогнозирования фармакокинетики у человека дает профиль у человека, сходный с тем, который можно было ожидать на основании эмпирических данных, полученных с другими молекулами IgG1 человека, включая точечную оценку периода полураспада у человека (T), равного примерно 18 суткам, который согласуется с результатами предыдущего исследования, приведенного выше.
Пример 14. Фармакодинамические эффекты (PD) антитела 230C9-N297Q на обезьянах циномолгус
Фармакодинамические эффекты антитела 230C9 N297Q (ARGX-112) исследовали на обезьяне циномолгус при внутривенном введении обезьяне антитела 230C9 в различных дозах (0,3; 1; 3; 10; 30 мг/кг). Срез кожи обезьяны также обрабатывали имиквимодом (IMQ) для оценки воздействия 230C9 N297Q на кожное воспаление. Сообщалось, что IMQ вызывает воспаление кожи у мышей (Van Belle et al. 2012, J. Immunol. Jan 1; 188 (1): 462-9), и в одном сообщении также были описаны аналогичные эффекты у приматов, отличных от человека (Poirier et al., 2016, Exp. Dermatol. Mar, 25 (3): 233-4; Poirier et al., 2016, J. Immunol. Jan 1, 196 (1): 274-83) и у человека (Vinter et al., 2015, Br. J. Dermatol. Feb, 172 (2): 345-53).
Через 5 суток после обработки IMQ отбирали биопсийный образец из участка кожи, не обработанного IMQ, и участка кожи, обработанного IMQ, для оценки эффектов 230C9 N297Q на действие, вызванное IMQ. Эффекты оценивали сравнением толщины эпидермиса и частоты пролиферирующих ядер в эпидермисе (частота Ki67-положительных ядер), и результаты показаны на фиг. 14. Возрастающие дозы антитела 230C9 N297Q были способны нормализовать толщину эпидермиса (фиг. 14A) и уменьшали частоту Ki67-положительных ядер (фиг. 14В). Значение ЕС50 составляло примерно 3 мг/кг.
Пример 15. Взаимосвязь PK в сыворотке крови и PD ответной реакции кожного экспланта после однократной инфузии 230C9 N297Q у обезьян циномолгус
Контактирование кожи с мишенью исследовали на обезьянах циномолгус, которым внутривенно однократно в течение 15 мин вводили антитело 230C9 N297Q (в дозе 5 мл/кг). Самок обезьян оставляли без обработки или подвергали воздействию антитела 230C9 в дозах 1; 5 или 30 мг/кг в день проведения испытания 1 (по три животных на группу). Кровь отбирали у всех обработанных животных (3×150 мкл) на разные временные точки, при этом последний образец отбирали на сутки 7. Образцы сыворотки (1% об/об) анализировали на уровни экспозиции 230C9 N297Q с использованием ELISA.
Образцы кожи, полученные пункционной биопсией (3 мм), отбирали на сутки -1 (до введения) и сутки 7 от каждого животного и инкубировали в присутствии (сутки -1, сутки 7) или в отсутствии (сутки 7) rhIL-22 в течение 24 ч при 37°C во влажной атмосфере воздух/CO2 (95%/5%) перед измерением уровней мРНК FLG2 с помощью кПЦР. Результаты показаны на фиг. 15.
Стимуляция rhIL22 приводила к 5-кратному снижению общего уровня мРНК FLG2 в коже. IL-22-опосредованная репрессия транскрипта FLG2 достоверно восстанавливалась у животных, обработанных 230C9 N297Q, что указывает на значительное воздействие на кожу во всех группах с введением разных доз.
Пример 16. Влияние 230C9 N297Q на жизнеспособные кожные экспланты человека и обезьяны циномолгус ex vivo
Для оценки способности антитела 230C9 N297Q ингибировать уровни мРНК DEFB4, индуцированные rhIL-22, использовали свежеотобранную кожу с области живота у здоровых доноров.
Образцы кожи, полученные пункционной биопсией (3 мм), помещали в вертикальном положении в 96-луночные планшеты для культивирования тканей и культивировали на границе раздела воздух-жидкость в 100 мкл среды EpiLife, с добавлением добавки для роста кератиноцитов человека (HKGS) без гидрокортизона (Invitrogen).
Образцы инкубировали с возрастающими концентрациями антитела 230C9 N297Q в течение 24 ч при 37°C во влажной атмосфере воздух/CO2 (95%/5%) перед стимуляцией 20 нг/мл rhIL-22 (R & D Systems) в течение еще 24 ч при 37°C. Относительные уровни экспрессии гена DEFB4 в лизатах кожи определяли количественной ПЦР в режиме реального времени (кПЦР) с использованием валидированных анализов экспрессии генов (Applied Biosystems) и системы определения последовательности ABI PRISM® 7900HT.
На фиг.16 показано дозозависимое снижение уровней мРНК DEFB4, индуцированных rhIL-22, по данным одного репрезентативного эксперимента (EC50 [CI95%]=4 нМ [0,5-29 нМ]; 4 донора). Аналогичную эффективность получали при увеличении времени предварительной инкубации 230C9 N297Q до 48 ч, что дает основание предположить о достижении равновесия связывания IL-22RA. Контрольное антитело того же изотипа не оказывало влияния на опосредованные rhIL-22 повышение уровней мРНК DEFB4.
В свежеотобранных биопсийных образцах кожи (3 мм) от обезьян циномолгус (Macaca Fascicularis) ген DEFB4 не регулировался rhIL-22. В противоположность, как FLG2, так и LOR подвергались понижающей регуляции после 24-ч инкубации с rhIL-22 с использованием того же протокола, который описан для опыта с кожным эксплантом человека. Данные реакции полностью блокировались 24-ч предварительной инкубацией с антителом 230C9 N297Q (EC50=11 нМ; оба гена), как показано на фиг. 17.
Пример 17. Эффективность антитела 230C9 N297Q на кератиноцитах человека
Дизайн был разработан для тестирования активности анти-IL-22R-антител в функциональном анализе на первичных кератиноцитах человека. Клетки стимулировали смесью цитокинов IL-4, IL-13, IL-22 (все 10 нг/мл) и IFN-γ, 1 нг/мл в течение 48 ч, и затем в культуральном супернатанте измеряли уровень CCL2 с использованием платформы MSD.
Кератиноциты суспендировали в среде EpiLife (Life Technologies) со следующими добавками для роста: EGF; BPE; инсулин; трансферрин и гентамицин/амфотерицин. Клетки высевали в 384-луночные белые прокси планшеты (Perkin Elmer) и инкубировали в течение 2 ч при 37°C, в атмосфере 5% CO2/95% воздуха. Затем клетки обрабатывали 80 нл антитела (в различных концентрациях, как показано на фиг. 18) или носителем. Затем добавляли 40 мкл стимулирующей смеси (IL-4, IL-13, IL-22 (R & D systems, все в конечной концентрации 10 нг/мл) и IFN-γ (R & D systems, конечная концентрация 1 нг/мл)) и планшеты инкубировали при 37°С, в атмосфере 5% СО2/95% воздуха в течение 2 суток. Контрольные лунки обрабатывали смесью IL-4 и IL-13 (оба 10 нг/мл) и IFN-γ, 1 нг/мл, и определяли 100% ингибирование сигнального пути IL-22. Концентрацию CCL2 в культуральном супернатанте измеряли с помощью набора MSD CCL2 (Mesoscale Cat # K151AYB-2). Жизнеспособность клеток оценивали с использованием реагента PrestoBlue®.
Активность 230C9 N297Q тестировали в 5 экспериментах, и было показано дозозависимое ингибирование уровней CCL2 со значениями EC50 0,10 нМ [0,042-0,24 нМ]. Репрезентативные экспериментальные результаты одного опыта показаны на фиг. 18.
Настоящее изобретение не должно ограничиваться в объеме конкретными вариантами осуществления, описанными в настоящем описании. Фактически, различные модификации изобретения в дополнение к тем, которые описаны в настоящем описании, станут очевидными для специалистов в данной области техники из предшествующего описания и сопровождающих фигур. Предполагается, что такие модификации попадают в объем прилагаемой формулы изобретения. Кроме того, все варианты осуществления, описанные в данном документе, считаются широко применимыми и могут комбинироваться с любыми и всеми другими совместимыми вариантами осуществления, в зависимости от ситуации.
--->
Список последовательностей
<110> ардженкс БВБА
<120> Анти-IL-22R-антитела
<130> P144494WO00
<150> GB1612337.4
<151> 2016-07-15
<160> 101
<170> PatentIn version 3.5
<210> 1
<211> 30
<212> БЕЛОК
<213> Лама
<400> 1
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 2
<211> 5
<212> БЕЛОК
<213> Лама
<400> 2
Ser Tyr Asp Met Ser
1 5
<210> 3
<211> 14
<212> БЕЛОК
<213> Лама
<400> 3
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 4
<211> 17
<212> БЕЛОК
<213> Лама
<400> 4
Ser Ile Tyr Asn Asp Gly Ser Asn Thr Ala Tyr Ser Asp Ser Val Lys
1 5 10 15
Gly
<210> 5
<211> 32
<212> БЕЛОК
<213> Лама
<400> 5
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 6
<211> 11
<212> БЕЛОК
<213> Лама
<400> 6
Val Gly Phe Ser Gly Thr Tyr Tyr Ser Glu Ser
1 5 10
<210> 7
<211> 11
<212> БЕЛОК
<213> Лама
<400> 7
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 8
<211> 30
<212> БЕЛОК
<213> Лама
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asp
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly
20 25 30
<210> 9
<211> 5
<212> БЕЛОК
<213> Лама
<400> 9
Ser Tyr Phe Met Ser
1 5
<210> 10
<211> 14
<212> БЕЛОК
<213> Лама
<400> 10
Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val Ser
1 5 10
<210> 11
<211> 17
<212> БЕЛОК
<213> Лама
<400> 11
Gly Ile His Ile Ser Gly Gly Ile Thr Tyr Tyr Leu Asp Ser Val Lys
1 5 10 15
Gly
<210> 12
<211> 32
<212> БЕЛОК
<213> Лама
<400> 12
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Val Thr
20 25 30
<210> 13
<211> 14
<212> БЕЛОК
<213> Лама
<400> 13
Pro Pro Gly Pro Phe Lys Ala His Tyr Asn Gly Met Lys Tyr
1 5 10
<210> 14
<211> 11
<212> БЕЛОК
<213> Лама
<400> 14
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 15
<211> 22
<212> БЕЛОК
<213> Лама
<400> 15
Asn Phe Met Leu Thr Gln Pro Ser Ala Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys
20
<210> 16
<211> 7
<212> БЕЛОК
<213> Лама
<400> 16
Gln Gly Gly Tyr Tyr Ala His
1 5
<210> 17
<211> 15
<212> БЕЛОК
<213> Лама
<400> 17
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 18
<211> 7
<212> БЕЛОК
<213> Лама
<400> 18
Gly Asn Asn Asn Arg Pro Ser
1 5
<210> 19
<211> 20
<212> БЕЛОК
<213> Лама
<400> 19
Asn Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Ala
1 5 10 15
Glu Tyr Tyr Cys
20
<210> 20
<211> 10
<212> БЕЛОК
<213> Лама
<400> 20
Gln Ser Gly Ser Ser Ser Ala Asn Ala Val
1 5 10
<210> 21
<211> 10
<212> БЕЛОК
<213> Лама
<400> 21
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 22
<211> 22
<212> БЕЛОК
<213> Лама
<400> 22
Asn Phe Met Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Leu Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys
20
<210> 23
<211> 14
<212> БЕЛОК
<213> Лама
<400> 23
Thr Gly Thr Ser Arg Asp Ile Gly Asp Tyr Asn Tyr Val Ser
1 5 10
<210> 24
<211> 15
<212> БЕЛОК
<213> Лама
<400> 24
Trp Tyr Gln Gln Leu Pro Gly Leu Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 25
<211> 7
<212> БЕЛОК
<213> Лама
<400> 25
Lys Val Asn Thr Arg Ser Ser
1 5
<210> 26
<211> 20
<212> БЕЛОК
<213> Лама
<400> 26
Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala
1 5 10 15
Asp Tyr Tyr Cys
20
<210> 27
<211> 10
<212> БЕЛОК
<213> Лама
<400> 27
Ala Ser Tyr Arg Leu Tyr Ala Asp Tyr Val
1 5 10
<210> 28
<211> 10
<212> БЕЛОК
<213> Лама
<400> 28
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 29
<211> 120
<212> БЕЛОК
<213> Лама
<400> 29
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Asn Asp Gly Ser Asn Thr Ala Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Gly Phe Ser Gly Thr Tyr Tyr Ser Glu Ser Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 30
<211> 103
<212> БЕЛОК
<213> Лама
<400> 30
Asn Phe Met Leu Thr Gln Pro Ser Ala Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Tyr Tyr Ala His Trp Tyr Gln
20 25 30
Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gly Asn Asn Asn
35 40 45
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Asn
50 55 60
Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Ala Glu
65 70 75 80
Tyr Tyr Cys Gln Ser Gly Ser Ser Ser Ala Asn Ala Val Phe Gly Gly
85 90 95
Gly Thr His Leu Thr Val Leu
100
<210> 31
<211> 123
<212> БЕЛОК
<213> Лама
<400> 31
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asp
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Ser Tyr
20 25 30
Phe Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile His Ile Ser Gly Gly Ile Thr Tyr Tyr Leu Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Thr Pro Pro Gly Pro Phe Lys Ala His Tyr Asn Gly Met Lys Tyr
100 105 110
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 32
<211> 110
<212> БЕЛОК
<213> Лама
<400> 32
Asn Phe Met Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Leu Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Arg Asp Ile Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Leu Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Lys Val Asn Thr Arg Ser Ser Gly Thr Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Tyr Arg Leu Tyr
85 90 95
Ala Asp Tyr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 33
<211> 30
<212> БЕЛОК
<213> Лама
<400> 33
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 34
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Фрагмент варианта зародышевого типа антитела
<400> 34
Ser Tyr Asp Met Asn
1 5
<210> 35
<211> 14
<212> БЕЛОК
<213> Лама
<400> 35
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 36
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 36
Ser Ile Tyr Asn Asp Ala Ser Asn Thr Ala Tyr Ser Asp Ser Val Lys
1 5 10 15
Gly
<210> 37
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 37
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 38
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 38
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 39
<211> 30
<212> БЕЛОК
<213> Лама
<400> 39
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 40
<211> 14
<212> БЕЛОК
<213> Лама
<400> 40
Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val Ser
1 5 10
<210> 41
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 41
Gly Ile His Ile Ser Gly Gly Ile Thr Tyr Tyr Thr Asp Ser Val Lys
1 5 10 15
Gly
<210> 42
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 42
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Val Thr
20 25 30
<210> 43
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 43
Pro Pro Gly Pro Phe Lys Ala His Tyr Asn Gly Ala Lys Tyr
1 5 10
<210> 44
<211> 11
<212> БЕЛОК
<213> Лама
<400> 44
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 45
<211> 22
<212> БЕЛОК
<213> Лама
<400> 45
Leu Pro Val Leu Thr Gln Pro Ser Ala Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys
20
<210> 46
<211> 15
<212> БЕЛОК
<213> Лама
<400> 46
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 47
<211> 7
<212> БЕЛОК
<213> Лама
<400> 47
Gly Gln Asn Asn Arg Pro Ser
1 5
<210> 48
<211> 20
<212> БЕЛОК
<213> Лама
<400> 48
Asn Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Ala
1 5 10 15
Glu Tyr Tyr Cys
20
<210> 49
<211> 10
<212> БЕЛОК
<213> Лама
<400> 49
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 50
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 50
Ser Tyr Glu Leu Thr Gln Pro Ser Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys
20
<210> 51
<211> 15
<212> БЕЛОК
<213> Лама
<400> 51
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 52
<211> 360
<212> БЕЛОК
<213> Лама
<400> 52
Cys Ala Gly Gly Thr Gly Cys Ala Gly Cys Thr Cys Gly Thr Gly Gly
1 5 10 15
Ala Gly Thr Cys Thr Gly Gly Gly Gly Gly Ala Gly Gly Cys Thr Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Gly Gly Gly Thr
35 40 45
Thr Cys Thr Cys Thr Gly Ala Gly Ala Cys Thr Cys Thr Cys Cys Thr
50 55 60
Gly Thr Gly Cys Ala Gly Cys Cys Thr Cys Thr Gly Gly Ala Thr Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Ala Gly Thr Ala Gly Cys Thr Ala Cys
85 90 95
Gly Ala Cys Ala Thr Gly Ala Gly Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Cys Cys Ala Gly Gly Cys Thr Cys Cys Ala Gly Gly Gly Ala Ala
115 120 125
Gly Gly Gly Gly Cys Thr Gly Gly Ala Gly Thr Gly Gly Gly Thr Gly
130 135 140
Thr Cys Cys Ala Gly Thr Ala Thr Thr Thr Ala Thr Ala Ala Thr Gly
145 150 155 160
Ala Cys Gly Gly Thr Ala Gly Thr Ala Ala Cys Ala Cys Ala Gly Cys
165 170 175
Cys Thr Ala Thr Thr Cys Ala Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Cys Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Cys Ala Gly Ala Gly Ala Cys Ala Ala Cys Gly Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Gly Thr Thr Gly Thr Ala Thr
225 230 235 240
Cys Thr Gly Cys Ala Ala Ala Thr Gly Ala Ala Cys Ala Gly Cys Thr
245 250 255
Thr Gly Ala Ala Ala Thr Cys Thr Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Gly Gly Cys Cys Gly Thr Gly Thr Ala Thr Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Ala Ala Ala Ala Gly Thr Thr Gly Gly Cys Thr Thr Thr Ala
290 295 300
Gly Thr Gly Gly Thr Ala Cys Thr Thr Ala Cys Thr Ala Cys Ala Gly
305 310 315 320
Thr Gly Ala Ala Thr Cys Ala Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Gly Ala Cys Cys Cys Ala Gly Gly Thr Cys Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Ala
355 360
<210> 53
<211> 20
<212> БЕЛОК
<213> Artificial Seqence
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 53
Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Glu Asp Glu Ala
1 5 10 15
Asp Tyr Tyr Cys
20
<210> 54
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 54
Gln Ser Gly Ser Ser Ser Ser Asn Ala Val
1 5 10
<210> 55
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 55
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 56
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 56
Gln Ser Ala Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys
20
<210> 57
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 57
Thr Gly Thr Ser Ser Asp Ile Gly Ser Tyr Asn Tyr Val Ser
1 5 10
<210> 58
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 58
Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 59
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 59
Glu Val Asn Lys Arg Ser Ser
1 5
<210> 60
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 60
Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu Ala
1 5 10 15
Asp Tyr Tyr Cys
20
<210> 61
<211> 10
<212> БЕЛОК
<213> Лама
<400> 61
Phe Gly Gly Gly Thr Gln Leu Thr Val Leu
1 5 10
<210> 62
<211> 103
<212> БЕЛОК
<213> Лама
<400> 62
Leu Pro Val Leu Thr Gln Pro Ser Ala Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Tyr Tyr Ala His Trp Tyr Gln
20 25 30
Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gly Gln Asn Asn
35 40 45
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Gly Ala Gly Asn
50 55 60
Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Ala Glu
65 70 75 80
Tyr Tyr Cys Gln Ser Gly Ser Ser Ser Ala Asn Ala Val Phe Gly Gly
85 90 95
Gly Thr Lys Leu Thr Val Leu
100
<210> 63
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела а
<400> 63
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Asn Asp Ala Ser Asn Thr Ala Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Gly Phe Ser Gly Thr Tyr Tyr Ser Glu Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 64
<211> 103
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 64
Ser Tyr Glu Leu Thr Gln Pro Ser Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Tyr Tyr Ala His Trp Tyr Gln
20 25 30
Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gly Gln Asn Asn
35 40 45
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Gly Ala Gly Asn
50 55 60
Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Glu Asp Glu Ala Asp
65 70 75 80
Tyr Tyr Cys Gln Ser Gly Ser Ser Ser Ser Asn Ala Val Phe Gly Gly
85 90 95
Gly Thr Lys Leu Thr Val Leu
100
<210> 65
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 65
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Phe Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile His Ile Ser Gly Gly Ile Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Thr Pro Pro Gly Pro Phe Lys Ala His Tyr Asn Gly Ala Lys Tyr
100 105 110
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 66
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 66
Gln Ser Ala Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Ser Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Glu Val Asn Lys Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Tyr Arg Leu Tyr
85 90 95
Ala Asp Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu
100 105 110
<210> 67
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 67
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Asn Asp Ala Ser Asn Thr Ala Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Gly Phe Ser Gly Thr Tyr Tyr Ser Glu Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 68
<211> 209
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 68
Ser Tyr Glu Leu Thr Gln Pro Ser Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Tyr Tyr Ala His Trp Tyr Gln
20 25 30
Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gly Gln Asn Asn
35 40 45
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Gly Ala Gly Asn
50 55 60
Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Glu Asp Glu Ala Asp
65 70 75 80
Tyr Tyr Cys Gln Ser Gly Ser Ser Ser Ser Asn Ala Val Phe Gly Gly
85 90 95
Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala Pro Ser Val
100 105 110
Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn Lys Ala Thr
115 120 125
Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val Thr Val Ala
130 135 140
Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu Thr Thr Thr
145 150 155 160
Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser Tyr Leu Ser
165 170 175
Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser Cys Gln Val
180 185 190
Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro Thr Glu Cys
195 200 205
Ser
<210> 69
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 69
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Phe Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile His Ile Ser Gly Gly Ile Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Thr Pro Pro Gly Pro Phe Lys Ala His Tyr Asn Gly Ala Lys Tyr
100 105 110
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 70
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 70
Gln Ser Ala Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Ser Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Glu Val Asn Lys Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Tyr Arg Leu Tyr
85 90 95
Ala Asp Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 71
<211> 574
<212> БЕЛОК
<213> homo sapiens
<400> 71
Met Arg Thr Leu Leu Thr Ile Leu Thr Val Gly Ser Leu Ala Ala His
1 5 10 15
Ala Pro Glu Asp Pro Ser Asp Leu Leu Gln His Val Lys Phe Gln Ser
20 25 30
Ser Asn Phe Glu Asn Ile Leu Thr Trp Asp Ser Gly Pro Glu Gly Thr
35 40 45
Pro Asp Thr Val Tyr Ser Ile Glu Tyr Lys Thr Tyr Gly Glu Arg Asp
50 55 60
Trp Val Ala Lys Lys Gly Cys Gln Arg Ile Thr Arg Lys Ser Cys Asn
65 70 75 80
Leu Thr Val Glu Thr Gly Asn Leu Thr Glu Leu Tyr Tyr Ala Arg Val
85 90 95
Thr Ala Val Ser Ala Gly Gly Arg Ser Ala Thr Lys Met Thr Asp Arg
100 105 110
Phe Ser Ser Leu Gln His Thr Thr Leu Lys Pro Pro Asp Val Thr Cys
115 120 125
Ile Ser Lys Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr
130 135 140
Pro Ile Arg Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Phe
145 150 155 160
His Asp Leu Phe Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln
165 170 175
Met His Leu Gly Gly Lys Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr
180 185 190
Pro Asp Thr Glu Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp
195 200 205
Ala Lys Glu Ser Ala Pro Tyr Met Cys Arg Val Lys Thr Leu Pro Asp
210 215 220
Arg Thr Trp Thr Tyr Ser Phe Ser Gly Ala Phe Leu Phe Ser Met Gly
225 230 235 240
Phe Leu Val Ala Val Leu Cys Tyr Leu Ser Tyr Arg Tyr Val Thr Lys
245 250 255
Pro Pro Ala Pro Pro Asn Ser Leu Asn Val Gln Arg Val Leu Thr Phe
260 265 270
Gln Pro Leu Arg Phe Ile Gln Glu His Val Leu Ile Pro Val Phe Asp
275 280 285
Leu Ser Gly Pro Ser Ser Leu Ala Gln Pro Val Gln Tyr Ser Gln Ile
290 295 300
Arg Val Ser Gly Pro Arg Glu Pro Ala Gly Ala Pro Gln Arg His Ser
305 310 315 320
Leu Ser Glu Ile Thr Tyr Leu Gly Gln Pro Asp Ile Ser Ile Leu Gln
325 330 335
Pro Ser Asn Val Pro Pro Pro Gln Ile Leu Ser Pro Leu Ser Tyr Ala
340 345 350
Pro Asn Ala Ala Pro Glu Val Gly Pro Pro Ser Tyr Ala Pro Gln Val
355 360 365
Thr Pro Glu Ala Gln Phe Pro Phe Tyr Ala Pro Gln Ala Ile Ser Lys
370 375 380
Val Gln Pro Ser Ser Tyr Ala Pro Gln Ala Thr Pro Asp Ser Trp Pro
385 390 395 400
Pro Ser Tyr Gly Val Cys Met Glu Gly Ser Gly Lys Asp Ser Pro Thr
405 410 415
Gly Thr Leu Ser Ser Pro Lys His Leu Arg Pro Lys Gly Gln Leu Gln
420 425 430
Lys Glu Pro Pro Ala Gly Ser Cys Met Leu Gly Gly Leu Ser Leu Gln
435 440 445
Glu Val Thr Ser Leu Ala Met Glu Glu Ser Gln Glu Ala Lys Ser Leu
450 455 460
His Gln Pro Leu Gly Ile Cys Thr Asp Arg Thr Ser Asp Pro Asn Val
465 470 475 480
Leu His Ser Gly Glu Glu Gly Thr Pro Gln Tyr Leu Lys Gly Gln Leu
485 490 495
Pro Leu Leu Ser Ser Val Gln Ile Glu Gly His Pro Met Ser Leu Pro
500 505 510
Leu Gln Pro Pro Ser Arg Pro Cys Ser Pro Ser Asp Gln Gly Pro Ser
515 520 525
Pro Trp Gly Leu Leu Glu Ser Leu Val Cys Pro Lys Asp Glu Ala Lys
530 535 540
Ser Pro Ala Pro Glu Thr Ser Asp Leu Glu Gln Pro Thr Glu Leu Asp
545 550 555 560
Ser Leu Phe Arg Gly Leu Ala Leu Thr Val Gln Trp Glu Ser
565 570
<210> 72
<211> 1725
<212> ДНК
<213> homo sapiens
<400> 72
atgaggacgc tgctgaccat cttgactgtg ggatccctgg ctgctcacgc ccctgaggac 60
ccctcggatc tgctccagca cgtgaaattc cagtccagca actttgaaaa catcctgacg 120
tgggacagcg ggccggaggg caccccagac acggtctaca gcatcgagta taagacgtac 180
ggagagaggg actgggtggc aaagaagggc tgtcagcgga tcacccggaa gtcctgcaac 240
ctgacggtgg agacgggcaa cctcacggag ctctactatg ccagggtcac cgctgtcagt 300
gcgggaggcc ggtcagccac caagatgact gacaggttca gctctctgca gcacactacc 360
ctcaagccac ctgatgtgac ctgtatctcc aaagtgagat cgattcagat gattgttcat 420
cctaccccca cgcccatccg tgcaggcgat ggccaccggc taaccctgga agacatcttc 480
catgacctgt tctaccactt agagctccag gtcaaccgca cctaccaaat gcaccttgga 540
gggaagcaga gagaatatga gttcttcggc ctgacccctg acacagagtt ccttggcacc 600
atcatgattt gcgttcccac ctgggccaag gagagtgccc cctacatgtg ccgagtgaag 660
acactgccag accggacatg gacctactcc ttctccggag ccttcctgtt ctccatgggc 720
ttcctcgtcg cagtactctg ctacctgagc tacagatatg tcaccaagcc gcctgcacct 780
cccaactccc tgaacgtcca gcgagtcctg actttccagc cgctgcgctt catccaggag 840
cacgtcctga tccctgtctt tgacctcagc ggccccagca gtctggccca gcctgtccag 900
tactcccaga tcagggtgtc tggacccagg gagcctgcag gagctccaca gcggcatagc 960
ctgtccgaga tcacctactt agggcagcca gacatctcca tcctccagcc ctccaacgtg 1020
ccacctcccc agatcctctc cccactgtcc tatgccccaa acgctgcccc tgaggtcggg 1080
cccccatcct atgcacctca ggtgaccccc gaagctcaat tcccattcta cgccccacag 1140
gccatctcta aggtccagcc ttcctcctat gcccctcaag ccactccgga cagctggcct 1200
ccctcctatg gggtatgcat ggaaggttct ggcaaagact cccccactgg gacactttct 1260
agtcctaaac accttaggcc taaaggtcag cttcagaaag agccaccagc tggaagctgc 1320
atgttaggtg gcctttctct gcaggaggtg acctccttgg ctatggagga atcccaagaa 1380
gcaaaatcat tgcaccagcc cctggggatt tgcacagaca gaacatctga cccaaatgtg 1440
ctacacagtg gggaggaagg gacaccacag tacctaaagg gccagctccc cctcctctcc 1500
tcagtccaga tcgagggcca ccccatgtcc ctccctttgc aacctccttc ccgtccatgt 1560
tccccctcgg accaaggtcc aagtccctgg ggcctgctgg agtcccttgt gtgtcccaag 1620
gatgaagcca agagcccagc ccctgagacc tcagacctgg agcagcccac agaactggat 1680
tctcttttca gaggcctggc cctgactgtg cagtgggagt cctga 1725
<210> 73
<211> 309
<212> ДНК
<213> Лама
<400> 73
aattttatgc tgactcagcc ctccgcggtg tccgtgtctt tgggacagac ggccaagatc 60
acctgccaag ggggctatta tgctcactgg taccagcaga agccaggcca ggcccctgtg 120
ttggtcatct atggaaataa taataggccc tcagggatcc ctgagcgctt ctctggctcc 180
agttctggga acacagccac cctgaccatc agcggggccc aggctgagga cgaggccgag 240
tattactgtc agtcaggaag cagtagtgct aatgctgtgt tcggcggagg gacccatctg 300
accgtcctg 309
<210> 74
<211> 369
<212> ДНК
<213> Лама
<400> 74
caggtgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggattc tctgagactc 60
tcctgtgcag cctctggatt caccttcgga agctatttca tgagctgggt ccgccaggct 120
ccaggaaagg ggcccgagtg ggtctcaggt attcatatta gtggtggtat tacatactac 180
ttagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctgcaaatga acaacctgaa acctgaggac acggccgtgt attattgtgt aacacccccg 300
ggccccttta aggcccatta caatggcatg aagtactggg gcaaagggac cctggtcacc 360
gtctcctca 369
<210> 75
<211> 330
<212> ДНК
<213> Лама
<400> 75
aattttatgc tgactcagcc tccctccgtg tctggaactc tgggaaagac ggtcaccatc 60
tcctgcactg gaaccagtcg tgacattggg gactataact atgtctcctg gtatcaacag 120
ctcccaggat tggcccccaa actcctgatc tataaagtca acactcgatc ctcagggacc 180
cctgatcgct tctctggctc caagtcaggc aacacggcct ccctgaccat ctctgggctc 240
cagtctgagg acgaggctga ttattactgt gcctcatata gactgtacgc cgattatgtg 300
ttcggcggag ggacccatct gaccgtcctg 330
<210> 76
<211> 360
<212> ДНК
<213> Лама
<400> 76
caggtgcagc tcgtggagtc tgggggaggc ttggtgcagc ctgggggttc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctacgaca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtccagt atttataatg acggtagtaa cacagcctat 180
tcagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgttgtat 240
ctgcaaatga acagcttgaa atctgaggac acggccgtgt attactgtgc aaaagttggc 300
tttagtggta cttactacag tgaatcatgg ggccagggga cccaggtcac cgtgtcctca 360
<210> 77
<211> 309
<212> ДНК
<213> Лама
<400> 77
ctgcctgtgc tgactcagcc ctccgcggtg tccgtgtctt tgggacagac ggccaggatc 60
acctgccaag ggggctatta tgctcactgg taccagcaga agccaggcca ggcccctgtg 120
ctggtcatct atggacagaa taataggccc tcagggatcc ctgagcgctt ctctggctcc 180
ggcgctggga acacagccac cctgaccatc agcggggccc aggctgagga cgaggctgag 240
tattactgtc agtcaggaag cagtagtgct aatgctgtgt tcggcggagg gaccaagctg 300
accgtcctc 309
<210> 78
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 78
caggtgcagc tcgtggagtc tgggggaggc ttggtgcagc ctgggggttc tctgagactc 60
tcctgtgctg cctctggatt caccttcagt agctacgaca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtgtccagc atttataacg acgccagtaa cacagcctat 180
tcagactccg tgaagggccg attcaccatc tccagagaca actcaaagaa cacgttgtat 240
ctgcaaatga acagcctgag agctgaggac acggccgtgt attactgtgc gaaagttggc 300
tttagtggta cttactacag tgaatcatgg ggccagggga ccctcgtcac tgtctcctca 360
<210> 79
<211> 309
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 79
tcctacgaac tgactcagcc ctcctcggtg tccgtggcgt tgggacagac ggccaggatc 60
acctgccaag gaggctatta tgcacactgg taccagcaga agccaggcca ggcccctgtg 120
ctggtcatct atggacagaa taataggccc tcagggatcc ctgagcgctt ctctggctcc 180
ggcgctggga acacagccac cctgacaatc agccgcgccc aggctgagga cgaggctgac 240
tattactgtc agtcaggaag cagtagttct aatgctgtgt tcggcggagg gaccaagctg 300
accgtcctc 309
<210> 80
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 80
caggtgcagc tcgtggagtc tgggggcggc ttggtgcagc ctggggattc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatttca tgagctgggt ccgccaggct 120
ccaggaaagg ggcccgagtg ggtctcaggt attcatatta gtggtggtat tacatactac 180
acggactccg tgaagggccg attcaccatc tccagagaca acgcaaagaa cacgctgtat 240
ctgcaaatga acagcctgag agctgaggac acggccgtgt attattgtgt aacacccccg 300
ggccccttta aggcccatta caatggcgcg aagtactggg gcaaagggac cctggtcact 360
gtctcctca 369
<210> 81
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность варианта зародышевого типа антитела
<400> 81
cagtctgccc tgactcagcc tccctccgtg tctggatctc ctggacagtc tgtcaccatc 60
tcctgcactg gaaccagtag tgacattggg tcctataact atgtctcctg gtatcaacag 120
ctcccaggaa aggcccccaa actcctgatc tatgaggtca acaagcgatc ctcaggggtc 180
cctgatcgct tctctggctc caagtcaggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgt gcctcatata gactgtacgc cgattatgtg 300
ttcggcggag ggacccaact gaccgtcctc 330
<210> 82
<211> 10
<212> БЕЛОК
<213> homo sapiens
<400> 82
Glu Pro Lys Ser Cys Asp Lys Thr His Thr
1 5 10
<210> 83
<211> 5
<212> БЕЛОК
<213> homo sapiens
<400> 83
Cys Pro Pro Cys Pro
1 5
<210> 84
<211> 8
<212> БЕЛОК
<213> homo sapiens
<400> 84
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 85
<211> 12
<212> БЕЛОК
<213> homo sapiens
<400> 85
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr
1 5 10
<210> 86
<211> 50
<212> БЕЛОК
<213> homo sapiens
<400> 86
Cys Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys
1 5 10 15
Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro
20 25 30
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
35 40 45
Cys Pro
50
<210> 87
<211> 8
<212> БЕЛОК
<213> homo sapiens
<400> 87
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 88
<211> 7
<212> БЕЛОК
<213> homo sapiens
<400> 88
Glu Ser Lys Tyr Gly Pro Pro
1 5
<210> 89
<211> 5
<212> БЕЛОК
<213> homo sapiens
<400> 89
Cys Pro Ser Cys Pro
1 5
<210> 90
<211> 8
<212> БЕЛОК
<213> homo sapiens
<400> 90
Ala Pro Glu Phe Leu Gly Gly Pro
1 5
<210> 91
<211> 3
<212> БЕЛОК
<213> homo sapiens
<400> 91
Glu Arg Lys
1
<210> 92
<211> 10
<212> БЕЛОК
<213> homo sapiens
<400> 92
Cys Cys Val Glu Cys Pro Pro Pro Cys Pro
1 5 10
<210> 93
<211> 7
<212> БЕЛОК
<213> homo sapiens
<400> 93
Ala Pro Pro Val Ala Gly Pro
1 5
<210> 94
<211> 78
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность EST
<400> 94
Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr Pro Ile Arg
1 5 10 15
Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Phe His Asp Leu
20 25 30
Phe Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln Met His Leu
35 40 45
Gly Gly Lys Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr Pro Asp Thr
50 55 60
Glu Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp Ala
65 70 75
<210> 95
<211> 78
<212> БЕЛОК
<213> Макак-резус
<400> 95
Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr Pro Ile Arg
1 5 10 15
Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Phe His Asp Leu
20 25 30
Phe Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln Met His Leu
35 40 45
Gly Gly Glu Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr Pro Asp Thr
50 55 60
Glu Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp Ser
65 70 75
<210> 96
<211> 79
<212> БЕЛОК
<213> Обезьяна циномолгус
<400> 96
Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr Pro Ile Arg
1 5 10 15
Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Tyr Asp Leu Ser
20 25 30
Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln Met Val Asn His
35 40 45
Leu Gly Gly Lys Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr Pro Asp
50 55 60
Thr Glu Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp Ser
65 70 75
<210> 97
<211> 79
<212> БЕЛОК
<213> Обезьяна циномолгус
<400> 97
Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr Pro Ile Arg
1 5 10 15
Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Tyr Asp Leu Ser
20 25 30
Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln Met Val Asn His
35 40 45
Leu Gly Gly Lys Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr Pro Asp
50 55 60
Thr Glu Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp Ser
65 70 75
<210> 98
<211> 77
<212> БЕЛОК
<213> Обезьяна циномолгус
<400> 98
Val Arg Ser Ile Gln Met Ile Val His Pro Thr Pro Thr Pro Ile Arg
1 5 10 15
Ala Gly Asp Gly His Arg Leu Thr Leu Glu Asp Ile Tyr Asp Leu Ser
20 25 30
Tyr His Leu Glu Leu Gln Val Asn Arg Thr Tyr Gln Met His Leu Gly
35 40 45
Gly Lys Gln Arg Glu Tyr Glu Phe Phe Gly Leu Thr Pro Asp Thr Glu
50 55 60
Phe Leu Gly Thr Ile Met Ile Cys Val Pro Thr Trp Ser
65 70 75
<210> 99
<211> 293
<212> БЕЛОК
<213> homo sapiens
<400> 99
Leu Arg Leu Glu Tyr Leu Ile Arg Leu Thr Ile Gly Tyr Arg Leu Asn
1 5 10 15
Gly Thr Ala Ala Thr Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val
20 25 30
Ala Thr Ala Thr Gly Val His Ser Pro Glu Asp Pro Ser Asp Leu Leu
35 40 45
Gln His Val Lys Phe Gln Ser Ser Asn Phe Glu Asn Ile Leu Thr Trp
50 55 60
Asp Ser Gly Pro Glu Gly Thr Pro Asp Thr Val Tyr Ser Ile Glu Tyr
65 70 75 80
Lys Thr Tyr Gly Glu Arg Asp Trp Val Ala Lys Lys Gly Cys Gln Arg
85 90 95
Ile Thr Arg Lys Ser Cys Asn Leu Thr Val Glu Thr Gly Asn Leu Thr
100 105 110
Glu Leu Tyr Tyr Ala Arg Val Thr Ala Val Ser Ala Gly Gly Arg Ser
115 120 125
Ala Thr Lys Met Thr Asp Arg Phe Ser Ser Leu Gln His Thr Thr Leu
130 135 140
Lys Pro Pro Asp Val Thr Cys Ile Ser Lys Val Arg Ser Ile Gln Met
145 150 155 160
Ile Val His Pro Thr Pro Thr Pro Ile Arg Ala Gly Asp Gly His Arg
165 170 175
Leu Thr Leu Glu Asp Ile Phe His Asp Leu Phe Tyr His Leu Glu Leu
180 185 190
Gln Val Asn Arg Thr Tyr Gln Met His Leu Gly Gly Lys Gln Arg Glu
195 200 205
Tyr Glu Phe Phe Gly Leu Thr Pro Asp Thr Glu Phe Leu Gly Thr Ile
210 215 220
Met Ile Cys Val Pro Thr Trp Ala Lys Glu Ser Ala Pro Tyr Met Cys
225 230 235 240
Arg Val Lys Thr Leu Pro Asp Arg Thr Trp Thr Ile Glu Gly Arg Asp
245 250 255
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
260 265 270
Ala Pro Glu Leu Leu Gly Arg Pro Ser Val Phe Leu Phe Pro Pro Lys
275 280 285
Pro Lys Asp Thr Leu
290
<210> 100
<211> 293
<212> БЕЛОК
<213> Макак-резус
<400> 100
Leu Arg Leu Glu Tyr Leu Ile Arg Leu Thr Ile Gly Tyr Arg Leu Asn
1 5 10 15
Gly Thr Ala Ala Thr Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val
20 25 30
Ala Thr Ala Thr Gly Val His Ser Pro Glu Asp Pro Ser Asp Leu Leu
35 40 45
Gln His Val Lys Phe Gln Ser Asn Asn Phe Glu Asn Ile Leu Thr Trp
50 55 60
Asp Ser Gly Pro Glu Gly Thr Pro Asp Thr Val Tyr Ser Ile Glu Tyr
65 70 75 80
Lys Thr Tyr Gly Glu Arg Asp Trp Val Ala Lys Lys Gly Cys Gln Arg
85 90 95
Ile Thr Arg Lys Ser Cys Asn Leu Thr Val Glu Thr Gly Asn His Thr
100 105 110
Glu Leu Tyr Tyr Ala Arg Val Thr Ala Val Ser Ala Gly Gly Arg Ser
115 120 125
Ala Thr Lys Met Thr Asp Arg Phe Asn Ser Leu Gln His Thr Ala Leu
130 135 140
Lys Pro Pro Asp Val Thr Cys Ile Pro Lys Val Arg Ser Ile Gln Met
145 150 155 160
Ile Val His Pro Thr Pro Thr Pro Ile Arg Ala Gly Asp Gly His Arg
165 170 175
Leu Thr Leu Glu Asp Ile Phe His Asp Leu Phe Tyr His Leu Glu Leu
180 185 190
Gln Val Asn Arg Thr Tyr Gln Met His Leu Gly Gly Glu Gln Arg Glu
195 200 205
Tyr Glu Phe Phe Gly Leu Thr Pro Asp Thr Glu Phe Leu Gly Thr Ile
210 215 220
Met Ile Cys Val Pro Thr Trp Ser Lys Lys Ser Ala Pro Tyr Met Cys
225 230 235 240
Arg Val Arg Thr Leu Pro Asp Arg Thr Trp Thr Ile Glu Gly Arg Asp
245 250 255
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
260 265 270
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
275 280 285
Pro Lys Asp Thr Leu
290
<210> 101
<211> 292
<212> БЕЛОК
<213> Обезьяна циномолгус
<400> 101
Leu Arg Leu Glu Tyr Leu Ile Arg Leu Thr Ile Gly Tyr Arg Leu Asn
1 5 10 15
Gly Thr Ala Ala Thr Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val
20 25 30
Ala Thr Ala Thr Gly Val His Ser Pro Glu Asp Pro Ser Asp Leu Leu
35 40 45
Gln His Val Lys Phe Gln Ser Asn Asn Phe Glu Asn Ile Leu Thr Trp
50 55 60
Asp Ser Gly Pro Glu Gly Thr Pro Asp Thr Val Tyr Ser Ile Glu Tyr
65 70 75 80
Lys Thr Tyr Gly Glu Arg Asp Trp Val Ala Lys Lys Gly Cys Gln Arg
85 90 95
Ile Thr Arg Lys Ser Cys Asn Leu Thr Val Glu Thr Gly Asn His Thr
100 105 110
Glu Leu Tyr Tyr Ala Arg Val Thr Ala Val Ser Ala Gly Gly Arg Ser
115 120 125
Ala Thr Lys Met Thr Asp Arg Phe Asn Ser Leu Gln His Thr Ala Leu
130 135 140
Lys Pro Pro Asp Val Thr Cys Ile Pro Lys Val Arg Ser Ile Gln Met
145 150 155 160
Ile Val His Pro Thr Pro Thr Pro Ile Arg Ala Gly Asp Gly His Arg
165 170 175
Leu Thr Leu Glu Asp Ile Tyr Asp Leu Ser Tyr His Leu Glu Leu Gln
180 185 190
Val Asn Arg Thr Tyr Gln Met His Leu Gly Gly Lys Gln Arg Glu Tyr
195 200 205
Glu Phe Phe Gly Leu Thr Pro Asp Thr Glu Phe Leu Gly Thr Ile Met
210 215 220
Ile Cys Val Pro Thr Trp Ser Lys Lys Ser Ala Pro Tyr Met Cys Arg
225 230 235 240
Val Arg Thr Leu Pro Asp Arg Thr Trp Thr Ile Glu Gly Arg Asp Met
245 250 255
Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
260 265 270
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
275 280 285
Lys Asp Thr Leu
290
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| АНТИ-ROR1/АНТИ-CD3 БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2831608C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИТЕЛО, СВЯЗЫВАЮЩЕЕ ЧЕЛОВЕЧЕСКИЙ IL-4R, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2779649C1 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| ПОЛИПЕПТИД, СОДЕРЖАЩИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН И ТРАНСПОРТИРУЮЩИЙ СЕГМЕНТ | 2017 |
|
RU2827545C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ИНТЕРЛЕЙКИНУ-31 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2019 |
|
RU2795411C2 |
| АНТИТЕЛА К ENTPD2, ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ И СПОСОБЫ ПРИМЕНЕНИЯ АНТИТЕЛ И ВИДОВ КОМБИНИРОВАННОЙ ТЕРАПИИ | 2019 |
|
RU2790991C2 |
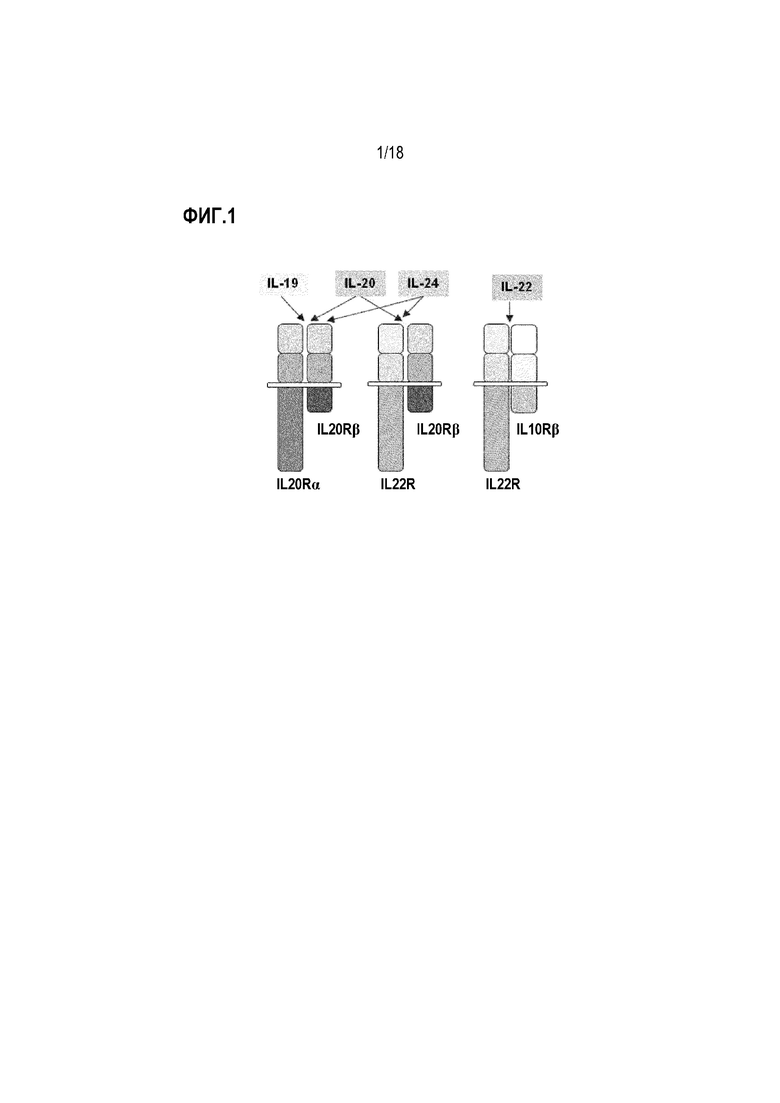




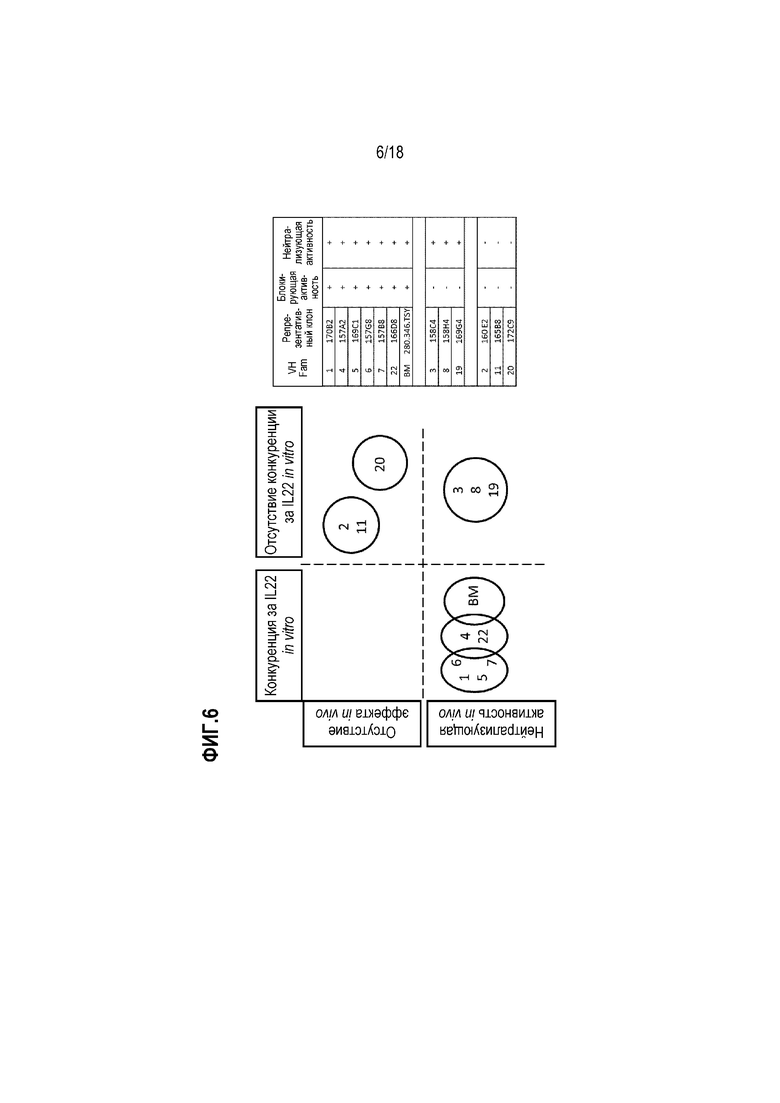
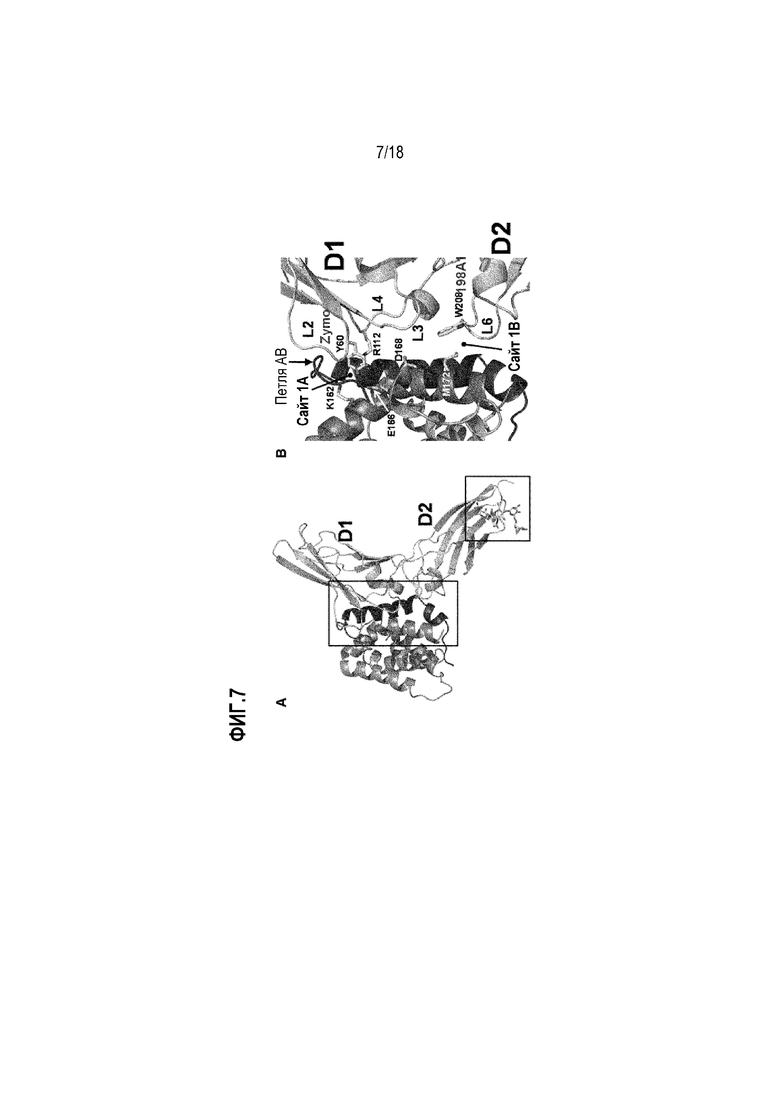
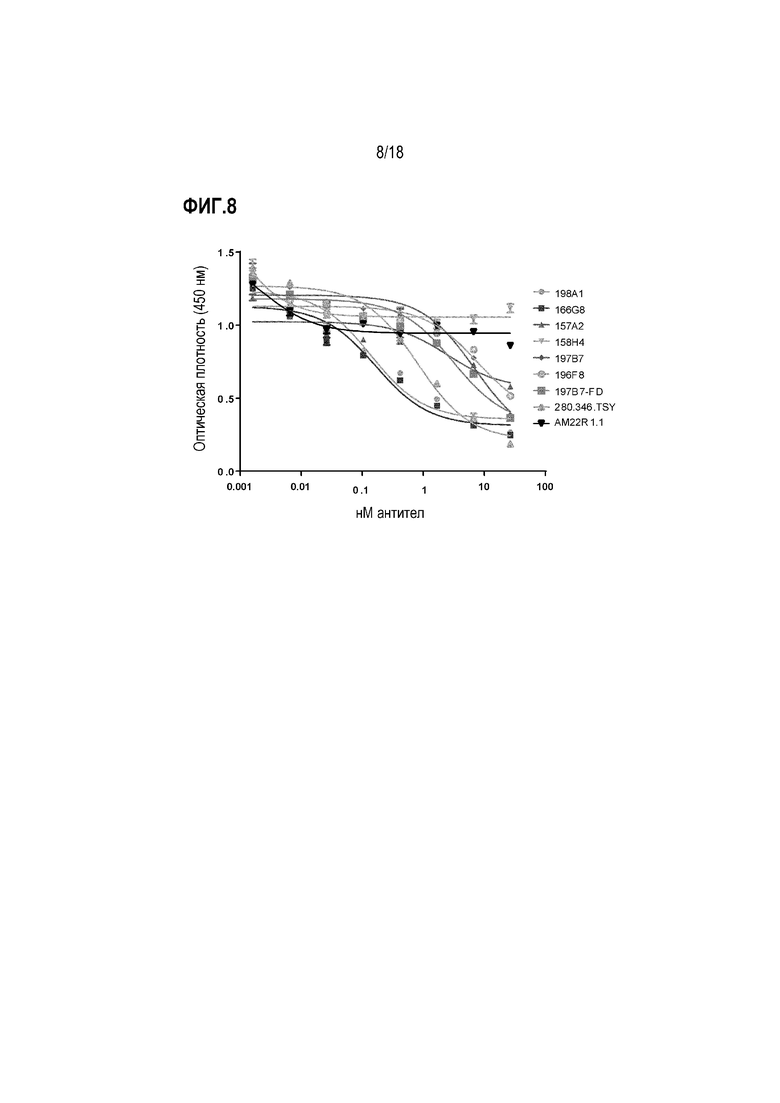
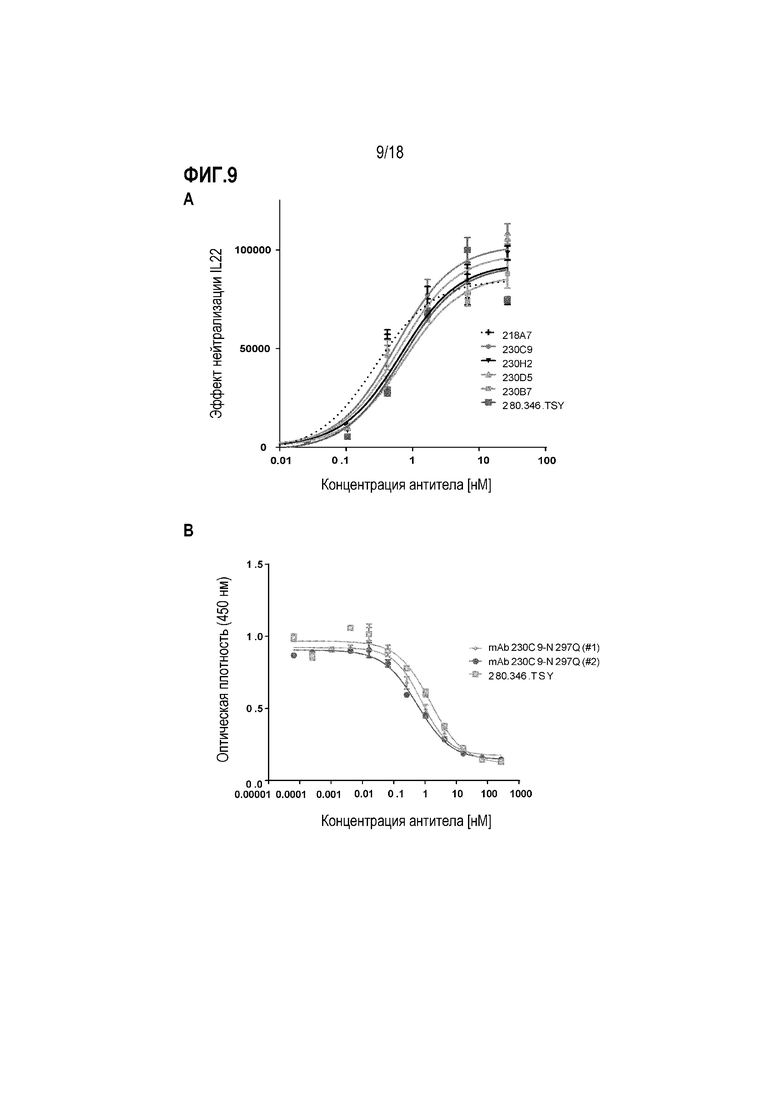
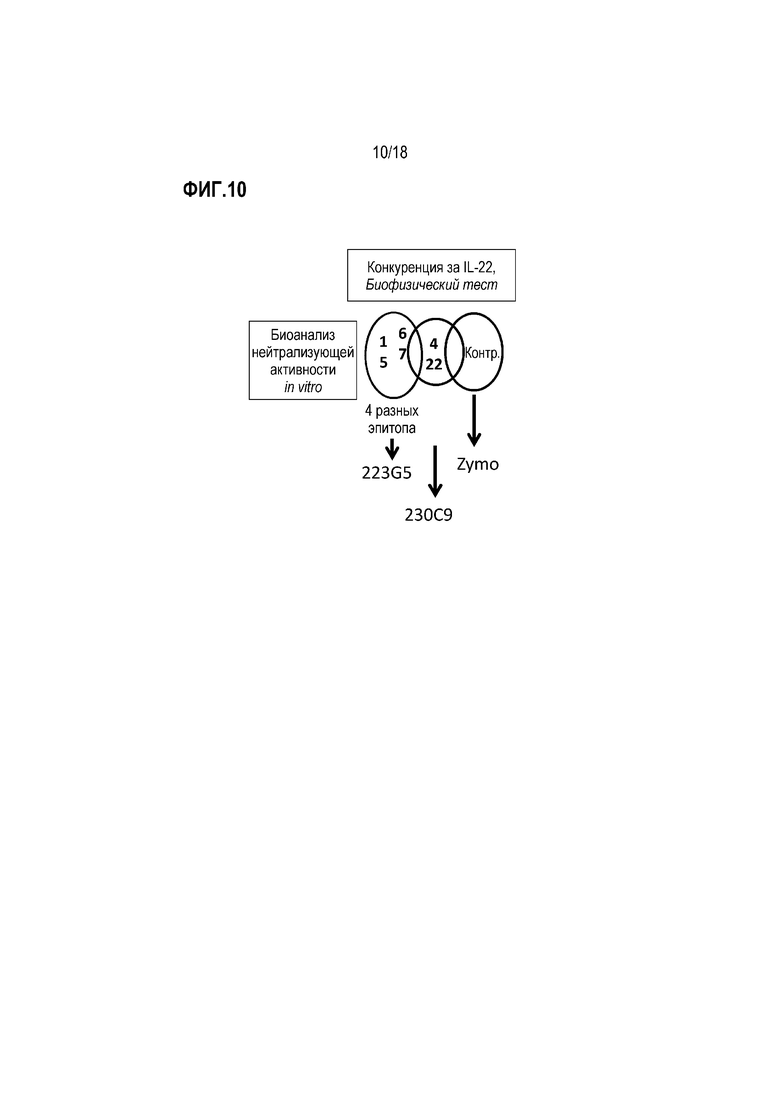

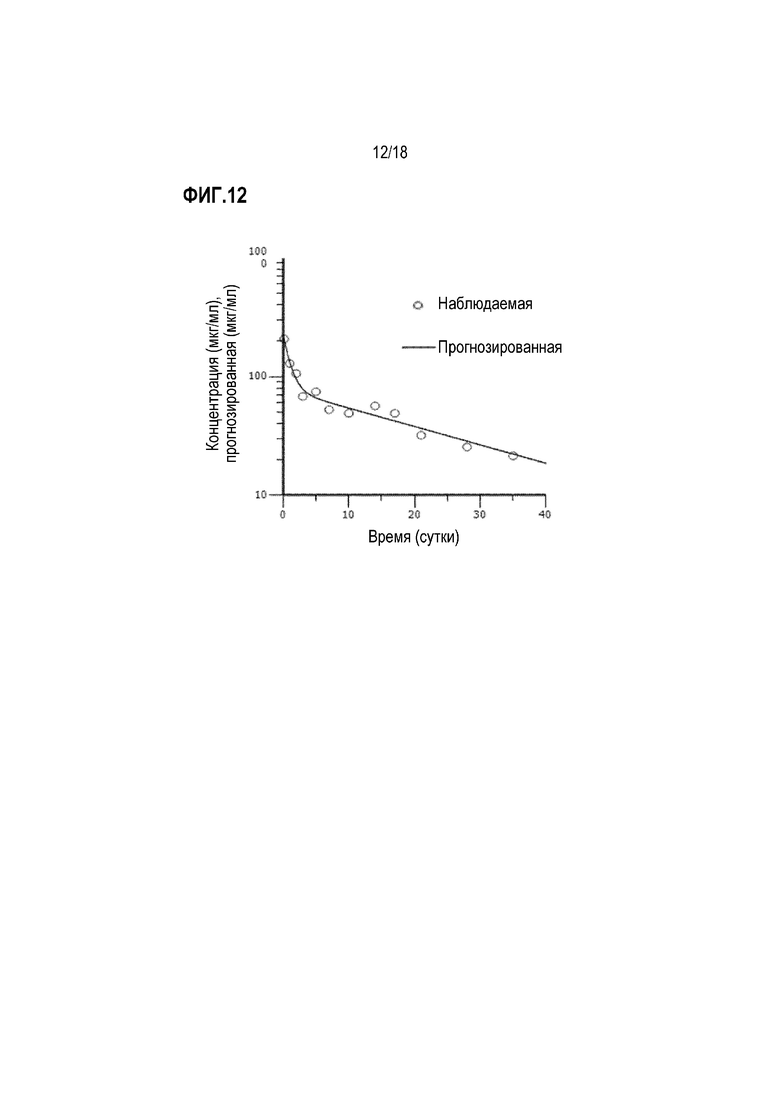
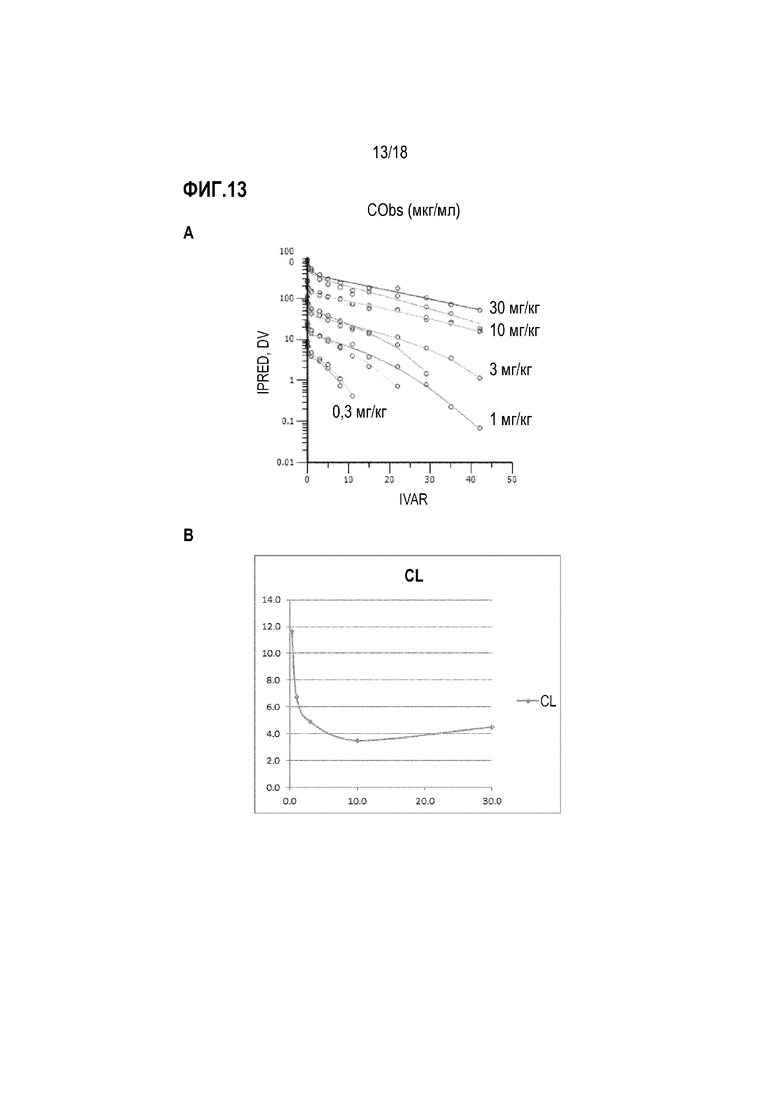
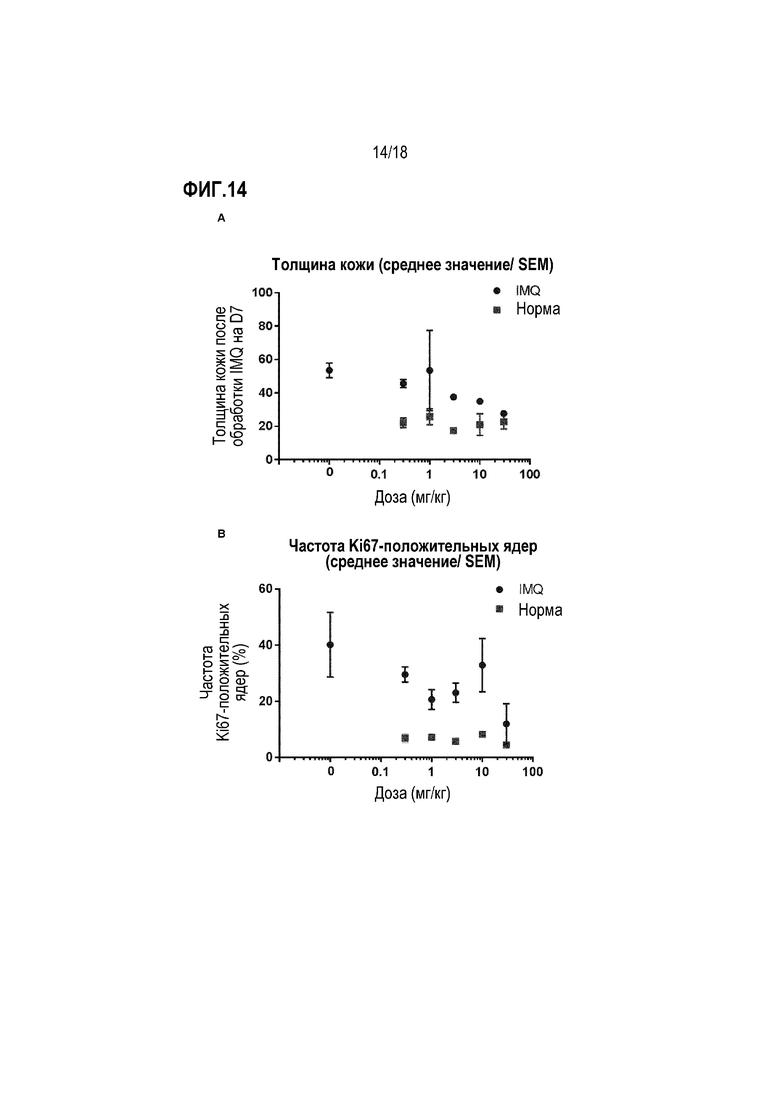

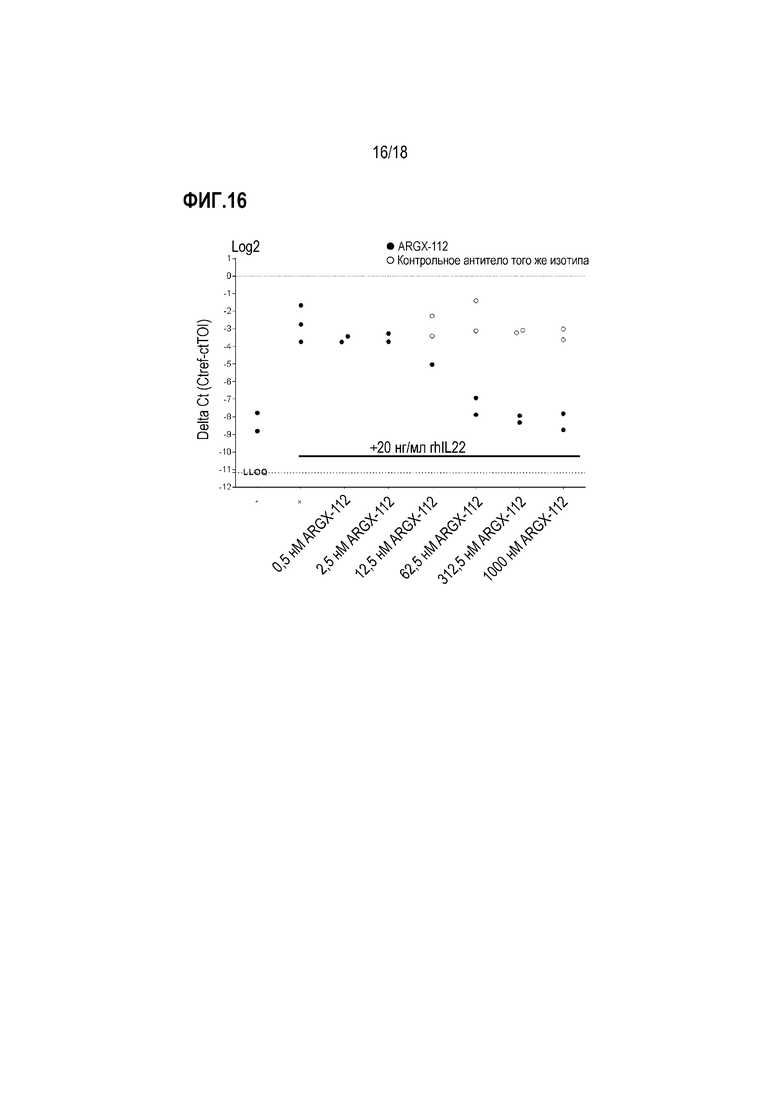
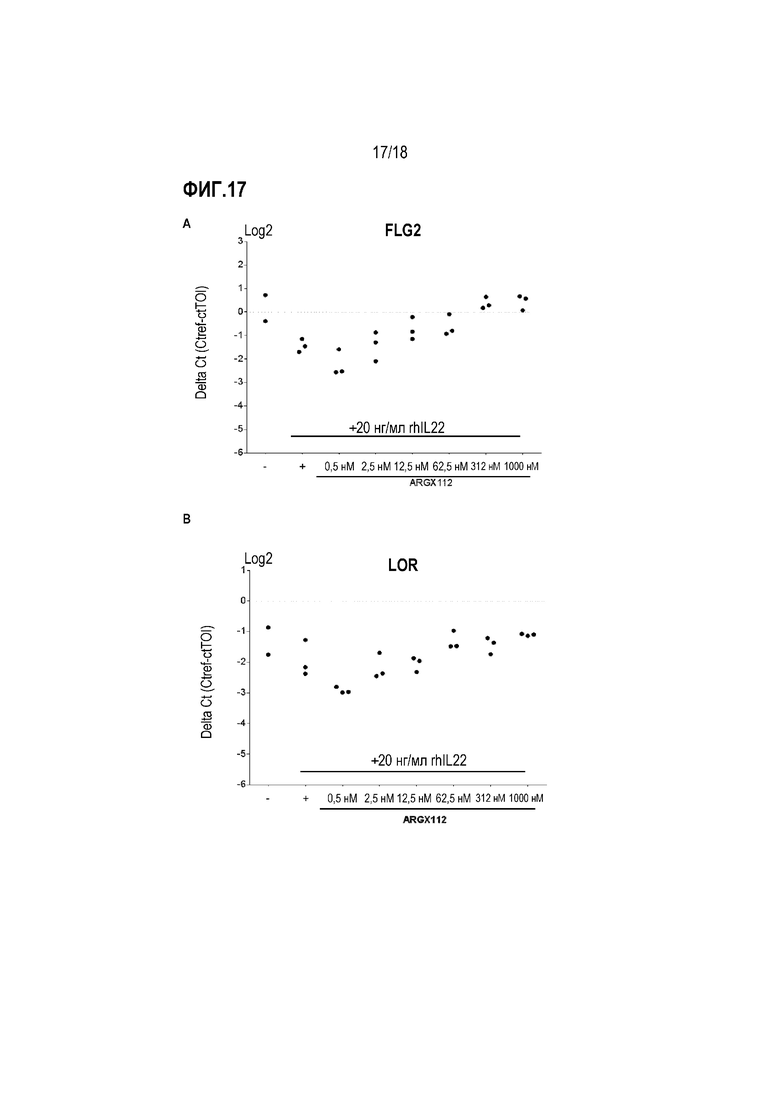
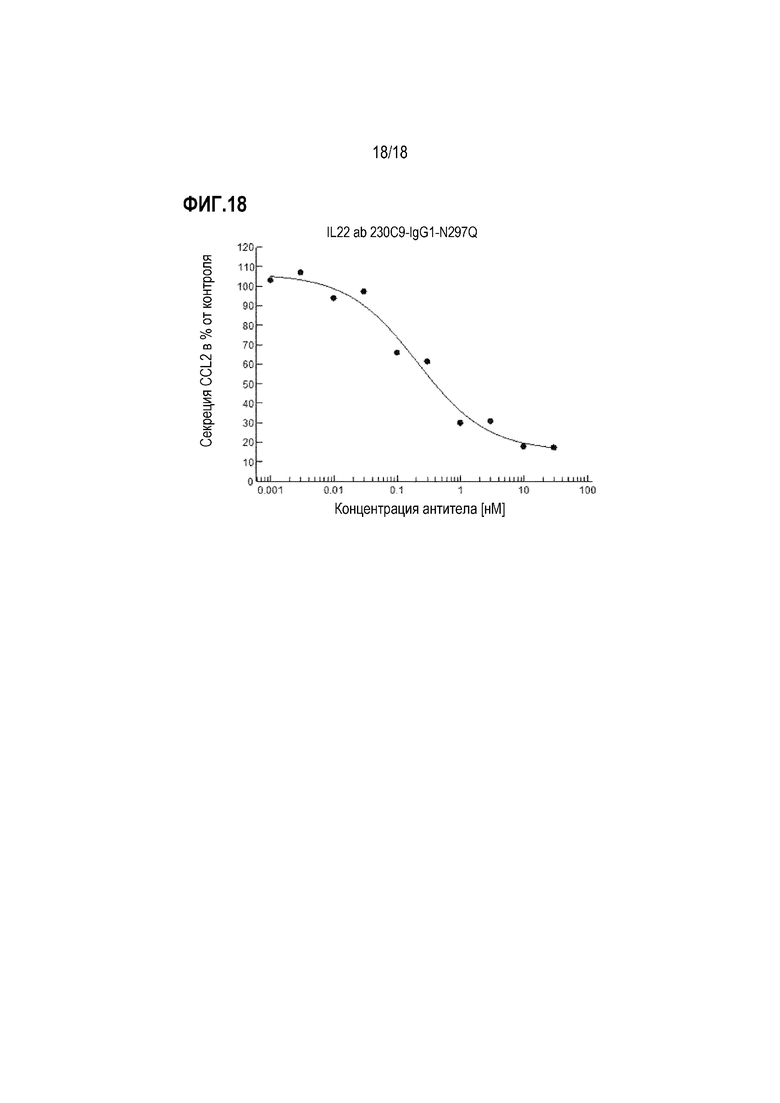
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые связываются с цитокиновым рецептором IL-22R, в частности IL-22R человека. Изобретение также относится к выделенному полинуклеотиду, вектору экспрессии, клетке-хозяину, бесклеточной экспрессионной системе, способу получения рекомбинантного антитела или его антигенсвязывающего фрагмента, фармацевтической композиции, способу лечения атопического дерматита, псориатического артрита или псориаза, применению в качестве лекарственного средства, применению в лечении или профилактике псориаза, псориатического артрита или атопического дерматита. 14 н. и 7 з.п. ф-лы, 18 ил., 30 табл., 17 пр.
1. Антитело или его антигенсвязывающий фрагмент, которое связывается с IL-22R человека,
где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую комбинацию последовательностей CDR вариабельной области тяжелой цепи: HCDR3, содержащий SEQ ID NO: 6; HCDR2, содержащий SEQ ID NO: 36; и HCDR1, содержащий SEQ ID NO: 34;
и вариабельную область легкой цепи, содержащую комбинацию последовательностей CDR вариабельной области легкой цепи: LCDR3, содержащий SEQ ID NO: 54; LCDR2, содержащий SEQ ID NO: 47; и LCDR1, содержащий SEQ ID NO: 16.
2. Антитело или антигенсвязывающий фрагмент по п.1, где антитело или его антигенсвязывающий фрагмент содержит комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), выбранных из следующего: VH, содержащей аминокислотную последовательность SEQ ID NO: 63 или аминокислотную последовательность, которая по меньшей мере на 90%, 95%, 97%, 98% или 99% идентична ей, и VL, содержащей аминокислотную последовательность SEQ ID NO: 64 или аминокислотную последовательность, которая по меньшей мере на 90%, 95%, 97%, 98% или 99% идентична ей.
3. Антитело по п.1, содержащее по меньшей мере одну тяжелую цепь и по меньшей мере одну легкую цепь, выбранные из следующего: тяжелой цепи, содержащей аминокислотную последовательность SEQ ID NO: 67 или аминокислотную последовательность, которая по меньшей мере на 90%, 95%, 97%, 98% или 99% идентична ей, и легкой цепи, содержащей аминокислотную последовательность SEQ ID NO: 68 или аминокислотную последовательность, которая по меньшей мере на 90%, 95%, 97%, 98% или 99% идентична ей.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-3, где антитело или его антигенсвязывающий фрагмент проявляет по меньшей мере одно или более из следующих свойств:
(i) связывается с эпитопом в пределах белка IL-22R, который не содержит Tyr60;
(ii) связывается с IL-22R человека с высокой аффинностью;
(iii) блокирует связывание IL-22 с IL-22R;
(iv) ингибирует IL-22-зависимую активацию IL-22R и/или IL-20-зависимую активацию IL-22R;
(v) не реагирует перекрестным образом с IL-22R мыши.
5. Антитело или антигенсвязывающий фрагмент по любому из пп.1-4, которое содержит шарнирную область, домен CH2 и/или домен CH3 IgG человека.
6. Антитело по любому из пп.1-5, которое проявляет высокую гомологию с IgG человека, предпочтительно с IgG1.
7. Антитело или антигенсвязывающий фрагмент по любому из пп.1-6, где области VH и/или VL, или один или более CDR, получены от животного семейства верблюдовых.
8. Антитело или антигенсвязывающий фрагмент по п.7, где животное представляет ламу.
9. Выделенный полинуклеотид, который кодирует антитело или антигенсвязывающий фрагмент по любому из пп.1-8.
10. Вектор экспрессии, содержащий полинуклеотид по п.9, функционально связанный с регуляторными последовательностями, которые обеспечивают экспрессию антитела, антигенсвязывающего фрагмента, вариабельной области тяжелой цепи или вариабельной области легкой цепи в клетке-хозяине или бесклеточной экспрессионной системе.
11. Клетка-хозяин для экспрессии антитела или его антигенсвязывающего фрагмента, которое связывается с белком IL-22R человека по любому из пп.1-8, содержащая вектор экспрессии по п.10.
12. Бесклеточная экспрессионная система для экспрессии антитела или его антигенсвязывающего фрагмента, которое связывается с белком IL-22R человека по любому из пп.1-8, содержащая вектор экспрессии по п.10.
13. Способ получения рекомбинантного антитела или его антигенсвязывающего фрагмента, который включает культивирование клетки-хозяина по п.11 или бесклеточной экспрессионной системы по п.12 в условиях, которые обеспечивают экспрессию антитела или антигенсвязывающего фрагмента, и выделение экспрессированного антитела или антигенсвязывающего фрагмента.
14. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент, которое связывается с белком IL-22R человека, по любому из пп.1-8 и по меньшей мере один фармацевтически приемлемый носитель или эксципиент,
где фармацевтическая композиция предназначена для связывания белка IL-22R человека.
15. Способ лечения псориаза у человека, который включает введение пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-8.
16. Способ лечения псориатического артрита у человека, который включает введение пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-8.
17. Способ лечения атопического дерматита у человека, который включает введение пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-8.
18. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-8 или фармацевтической композиции по п.14 в качестве лекарственного средства для связывания белка IL-22R человека, где антитело или антигенсвязывающий фрагмент связывается с эпитопом в пределах белка IL-22R, который не содержит Tyr60.
19. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-8 или фармацевтической композиции по п.14 в лечении или профилактике псориаза.
20. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-8 или фармацевтической композиции по п.14 в лечении или профилактике псориатического артрита.
21. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-8 или фармацевтической композиции по п.14 в лечении или профилактике атопического дерматита.
| WO 2006047249 A1, 22.10.2004 | |||
| JING LI et al., Temporal associations between interleukin 22 and the extracellular domains of IL-22R and IL-10R2, International Immunopharmacology, 2004, Vol.4, N5, pp.693-708 | |||
| DUDAKOV J.A | |||
| et al., Interleukin-22: immunobiology and pathology, Annu Rev Immunol, 2015, N33, pp.747-785 | |||
| СИНГЕР М | |||
| и др., Гены и геномы, |

