ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В данной заявке испрашивается приоритет заявки на патент США №11/104248, поданной 11 апреля 2005, которая включена в данное описание ссылкой во всей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к антителам против фактора роста нервов (NGF), таким как антагонистические антитела против NGF. Это изобретение дополнительно относится к применению антагонистов, таких как антитела, в лечении и/или предотвращении боли, включая послеоперационную боль, боль, вызванную ревматоидным артритом, и боль, вызванную остеоартритом.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Фактор роста нервов (NGF) был первым идентифицированным нейротрофином, и его роль в развитии и выживании как периферических, так и центральных нейронов хорошо охарактеризована. Показано, что при развитии NGF является критическим фактором выживания и поддержания периферических симпатических и эмбриональных сенсорных нейронов и холинергических нейронов базального переднего мозга. Smeyne et al., Nature 368:246-249 (1994), и Crowley et al., Cell 76:1001-1011 (1994). NGF повышает экспрессию нейропептидов в сенсорных нейронах (Lindsay and Harmer, Nature 337:362-364 (1989)), причем его активность опосредована двумя разными связанными с мембраной рецепторами, тирозинкиназным рецептором TrkA и общим рецептором нейротрофинов р75 (иногда называемых рецепторами NGF "высокой аффинности" и "низкой аффинности" соответственно). Chao et al., Science 232:518-521 (1986). р75-Рецептор по структуре родственен другим членам семейства рецепторов фактора некроза опухолей (Chao, et al., Science 232:518-521 (1986)). Обзор по NGF см. в Huang et al., Annu. Rev. Neurosci. 24:677-736 (2001); Bibel et al., Genes Dev. 14:2919-2937 (2000). Определена кристаллическая структура NGF и NGF в комплексе с TrkA-рецептором. См. Nature 254:411 (1991); Nature 401:184-188 (1996).
В дополнение к его эффектам в нервной системе, NGF все больше считают вовлеченным в процессы вне нервной системы. Например, показано, что NGF усиливает сосудистую проницаемость (Otten et al., Eur J Pharmacol. 106:199-201 (1984)), усиливает Т- и В-клеточные иммунные ответы (Otten, et al., Proc. Natl. Acad. Sci. USA 86:10059-10063 (1989)), индуцирует дифференцировку лимфоцитов и пролиферацию тучных клеток и вызывает высвобождение растворимых биологических сигналов из тучных клеток (Matsuda, et al., Proc. Natl. Acad. Sci. USA 85:6508-6512 (1988); Pearce, et al., J. Physiol. 372:379-393 (1986); Bischoff, et al., Blood 79:2662-2669 (1992); Horigome, et al., J. Biol. Chem. 268:14881-14887 (1993)). Хотя показано, что экзогенно добавленный NGF способен ко всем этим эффектам, важно отметить, что только в редких случаях показано, что эндогенный NGF является важным в каком-либо из этих процессов in vivo (Torcia, et al., Cell. 85(3):345-56 (1996)). Поэтому неясно, каким может быть эффект, если он вообще есть, ингибирования биологической активности эндогенного NGF.
NGF продуцируют многие типы клеток, в том числе тучные клетки (Leon, et al., Proc. Natl. Acad. Sci. USA 91:3739-3743 (1994)), В-лимфоциты (Torcia, et al., Cell 85:345-356 (1996), кератиноциты (Di Marco, et al., J. Biol. Chem. 268:22838-22846)), гладкомышечные клетки (Ueyama, et al., J. Hypertens. 11:1061-1065 (1993)), фибробласты (Lindholm, et al., Eur. J. Neurosci. 2:795-801 (1990)), бронхиальные эпителиальные клетки (Kassel, et al., Clin, Exp. Allergy. 31:1432-40 (2001)), почечные мезангиальные клетки (Steiner, et al., Am. J. Physiol. 261:F792-798 (1991)) и мышечные трубочки скелетных мышц (Schwartz, et al., J Photochem. PhotoBiol. B66:195-200 (2002)). Рецепторы NGF обнаружены на множестве разных типов клеток вне нервной системы. Например, Trka обнаружен на человеческих моноцитах, Т- и В-лимфоцитах и тучных клетках.
Взаимосвязь между повышенными уровнями NGF и множеством различнных воспалительных состояний наблюдали у больных людей, а также в некоторых животных моделях. Они включают системную красную волчанку (Bracci-Laudiero, et al., Neuroreport 4:563-565 (1993)), рассеянный склероз (Bracci-Laudiero, et al., Neurosci. Lett. 147:9-12 (1992)), псориаз (Raychaudhuri, et al., Acta Derm. l'enereol. 78:84-86 (1998)), артрит (Falcim, et al., Ann. Rheum. Dis. 55:745-748 (1996)), интерстициальный цистит (Okragly, et al., J. Urology 161:438-441 (1999)) и астму (Braun, et al., Eur. J Immunol. 28:3240-3251 (1998)).
Соответственно повышенный уровень NGF в периферических тканях связан с гипералгезией и воспалением, и его наблюдали при многих формах артрита. Синовиальная мембрана пациентов, пораженных ревматоидным артритом, экспрессирует высокие уровни NGF, в то время как сообщалось, что в невоспаленной синовиальной мембране NGF не выявлен (Aloe, et al., Arch. Rheum. 35:351-355 (1992)). Такие же результаты наблюдали у крыс с экспериментально индуцированным ревматоидным артритом (Aloe, et al., Clin. Exp.Rheumatol. 10:203-204 (1992)). Сообщалось о повышенных уровнях NGF наряду с увеличенным числом тучных клеток у трансгенных артритных мышей (Aloe, el al., Int. J. Tissue Reactions-Exp.Clin. Aspects. 15:139-143 (1993)). В публикации РСТ WO № 02/096458 раскрыто применение антител против NGF с определенными свойствами в лечении различных расстройств, связанных с NGF, таких как воспалительное состояние (например, ревматоидный артрит). Сообщалось, что очищенное антитело против NGF, инъецированное артритным трансгенным мышам, несущим ген человеческого фактора-α некроза опухолей (tumor necrosis factor, TNF-α), вызывает снижение числа тучных клеток, а также снижение уровней гистамина и вещества Р в синовиальной мембране артритных мышей (Aloe et al., Rheumatol. Int. 14: 249-252 (1995)). Показано, что экзогенное введение антитела против NGF снижает повышенные уровни TNF-α, имеющие место у артритных мышей (Manni et al., Rheumatol. Int. 18: 97-102 (1998)).
Кроме того, повышенную экспрессию NGF и высокоаффинного рецептора NGF (TrkA) наблюдали в человеческих остеоартритных хондроцитах (Iannone et al., Rheumatology. 41:1413-1418 (2002)).
Сообщалось об антагонистических антителах грызунов против NGF. См., например, Hongo et al, Hybridoma (2000), 19(3):215-227; Ruberti et al. (1993) Cell. Molec. Neurobiol. 13(5): 559-568. Однако когда антитела грызунов используют в терапевтических целях у людей, у значительного числа леченых индивидуумов развивается ответ в виде продуцирования антител человека против мыши. В дополнение, эффекторные функции мышиных антител оказались менее эффективными в контексте человека. Таким образом, имеется насущная необходимость в антагонистических антителах против NGF, в том числе в гуманизированных антагонистических антителах против NGF.
Все ссылки, публикации и заявки на патент, раскрытые здесь, включены в данное описание ссылкой во всей полноте.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Раскрытое здесь изобретение относится к антителам к фактору роста нервов.
В другом аспекте это изобретение представляет собой гуманизированное и имеющее развитую аффинность антитело, Е3, которое специфично связывается с фактором роста нервов ("NGF") человека и грызунов. Аминокислотные последовательности вариабельных областей тяжелой цепи и легкой цепи Е3 показаны на Фиг.1А (SEQ ID NO:1) и 1Б (SEQ ID NO:2) соответственно. Гипервариабельные участки (complementarity determining region, CDR) антитела Е3 (включая CDR по Chothia и по Kabat) представлены в виде диаграмм на Фиг.1А и 1Б. Аминокислотные последовательности тяжелых и легких цепей Е3 и индивидуальных расширенных CDR также показаны ниже (см. ниже "последовательности антител").
В другом аспекте это изобретение представляет собой антитело, содержащее фрагмент или область антитела Е3 (взаимозаменяемым образом обозначаемого здесь как "Е3"). В одном из воплощений этот фрагмент представляет собой легкую цепь антитела Е3, как показано на Фиг.1Б. В еще одном воплощении этот фрагмент представляет собой тяжелую цепь антитела Е3, как показано на Фиг.1А. В еще одном воплощении этот фрагмент содержит одну или более чем одну вариабельную область легкой цепи и/или тяжелой цепи антитела Е3. В еще одном воплощении этот фрагмент содержит один или более чем один гипервариабельный участок (CDR) легкой цепи и/или тяжелой цепи антитела Е3, как показано на Фиг.1А и 1Б.
В другом аспекте это изобретение представляет собой антитело, содержащее легкую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4893 или АТСС №РТА-4894. В другом аспекте это изобретение представляет собой антитело, содержащее тяжелую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895. В другом аспекте это изобретение представляет собой антитело, содержащее: (а) легкую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4894 или АТСС № РТА-4893, и (б) тяжелую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895 (для удобства, полинуклеотид(ы), продуцируемые депонированной клеткой-хозяином, обозначены здесь как имеющие номера депонирования АТСС РТА-4894, РТА-4893 и РТА-4895). В другом аспекте это изобретение представляет собой антитело, содержащее вариабельную область легкой цепи из легкой цепи, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4894 или АТСС № РТА-4893. В другом аспекте это изобретение представляет собой антитело, содержащее вариабельную область тяжелой цепи из тяжелой цепи, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895. В другом аспекте это изобретение представляет собой антитело, содержащее: (а) вариабельную область легкой цепи из легкой цепи, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4894 или АТСС № РТА-4893, и (б) вариабельную область тяжелой цепи из тяжелой цепи, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895. В еще одном аспекте это изобретение представляет собой антитело, содержащее один или более чем один CDR, кодируемый: (а) полинуклеотидом, продуцируемым клеткой-хозяином, с номером депонирования АТСС № РТА-4894, и/или (б) тяжелую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895.
В некоторых воплощениях антитело содержит константную область тяжелой цепи lgG2a человека. В некоторых воплощениях антитело содержит константную область человеческой легкой каппа-цепи. В некоторых воплощениях антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной, например не вызывает опосредованный комплементом лизис или не стимулирует антителозависимую клеточно-опосредованную цитотоксичность (ADCC). В других воплощениях константная область модифицирована, как описано в Eur. J. Immunol. (1999) 29:2613-2624; заявка РСТ № PCT/G 899/01441; и/или заявка на патент UK №9809951.8. В еще других воплощениях антитело содержит константную область тяжелой цепи lgG2a человека, содержащую следующие мутации: А330Р331 в S330S331 (нумерация аминокислот по последовательности lgG2a дикого типа). Eur. J. Immunol. (1999) 29:2613-2624.
В другом аспекте в этом изобретении предложены полипептиды (которые могут быть или могут не быть антителом), содержащие любое одно или более чем одно из следующего: а) один или более чем один CDR антитела Е3, показанный на Фиг.1А и 1Б; б) CDR-H3 из тяжелой цепи антитела Е3, показанный на Фиг.1А; в) CDR-L3 из легкой цепи антитела Е3, показанный на Фиг.1Б; г) три CDR из легкой цепи антитела Е3, показанные на Фиг.1Б; д) три CDR из тяжелой цепи антитела Е3, показанные на Фиг.1А; и е) три CDR из легкой цепи и три CDR из тяжелой цепи антитела Е3, показанные на Фиг.1А и 1Б. В этом изобретении дополнительно предложены полипептиды (которые могут быть или не быть антителом), содержащие любое одно или более чем одно из следующего: а) один или более чем один (один, два, три, четыре, пять или шесть) CDR, происходящий(е) из антитела Е3, показанный(е) на Фиг.1А и 1Б; б) CDR, происходящий из CDR-H3 из тяжелой цепи антитела Е3, показанный на Фиг.1А; и/или в) CDR, происходящий из CDR-L3 из легкой цепи антитела Е3, показанный на Фиг.1Б. В некоторых воплощениях CDR может представлять собой CDR по Kabat, CDR no Chothia или комбинацию CDR по Kabat и no Chothia (называемую здесь "расширенным" или "объединенным" CDR). В некоторых воплощениях полипептиды (такие как антитело) связываются с NGF (такими как человеческий NGF). В некоторых воплощениях полипептиды содержат любую конфигурацию CDR (включая комбинации, варианты и так далее), описанную здесь.
В одном из аспектов в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область тяжелой цепи, содержащую SEQ ID NO:9, где I34 представляет собой S, L, V А или I и N35 заменена на N, Т или S. "Заменена на" или "представляет собой" в этом контексте или как ссылка на аминокислоту для удобства относится здесь к вариантам выбора аминокислот(ы) для данного положения. Понятно, что замена или выбор может представлять собой аминокислоту, указанную в SEQ ID или на Фигуре.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые включают вариабельную область тяжелой цепи, содержащую SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L; А62 представляет собой А или S и L63 представляет собой L или V.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые включают вариабельную область тяжелой цепи, содержащую SEQ ID NO:11, где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где S105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой D, N или G и где Y110 представляет собой Y, K, S, R или Т.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область тяжелой цепи, содержащую SEQ ID NO:11, где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где S105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область тяжелой цепи, содержащую SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т; где G99 представляет собой G, S, А, С, V, N, D или Т; где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где S105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область легкой цепи, содержащую SEQ ID NO:12, где S26 представляет собой S или F; D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область легкой цепи, содержащую SEQ ID NO:13, где 151 представляет собой I, Т, V или А и S56 представляет собой S или Т.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область легкой цепи, содержащую SEQ ID NO:14, где S91 представляет собой S или Е; K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область легкой цепи, содержащую SEQ ID NO:14, где S91 представляет собой S или Е; K92 представляет собой любую аминокислоту; Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R.
В одном из аспектов в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:9, где I34 представляет собой S, L, V А или I и N35 представляет собой N, Т или S.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L; А62 представляет собой А или S и L63 представляет собой L или V.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где S105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой D, N или G и где Y110 представляет собой Y, K, S, R или Т.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где 8105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т; где G99 представляет собой G, S, А, С, V, N, D или Т; где Y100 представляет собой Y, L или R; где Y101 представляет собой Y или W; где G103 представляет собой G, А или S; где Т104 представляет собой Т или S; где S105 представляет собой S, А или Т; где Y106 представляет собой Y, R, Т или М; где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:12, где S26 представляет собой S или F; D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:13, где 151 представляет собой I, Т, V или А и S56 представляет собой S или Т.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е; K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е; K92 представляет собой любую аминокислоту; Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитела, в том числе гуманизированные антитела), которые содержат вариабельную область тяжелой цепи, содержащую CDR1-область из SEQ ID NO:9, где I34 представляет собой S, L, V A или I и N35 представляет собой N, Т или S; CDR2-область из SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L; A62 представляет собой А или S и L63 представляет собой L или V; и CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G, где Y110 представляет собой Y, K, S, R или Т. В некоторых воплощениях вариабельная область тяжелой цепи содержит CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G, где Y110 представляет собой любую аминокислоту. В других воплощениях вариабельная область тяжелой цепи содержит CDR3-область из SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т, где G99 представляет собой G, S, А, С, V, N, D или Т, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту. В некоторых воплощениях полипептид (такой как антитело) дополнительно содержит вариабельную область легкой цепи антитела.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат вариабельную область легкой цепи, содержащую CDR1-область из SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; CDR2-область из SEQ ID NO:13, где I51 представляет собой I, Т, V или А и S56 представляет собой S или Т; и CDRS-область из SEQ ID NO:14, где S91 представляет собой S или Е; K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R. В некоторых воплощениях вариабельная область легкой цепи содержит CDR3-область из SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой любую аминокислоту, Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R. В некоторых воплощениях полипептид (такой как антитело) дополнительно содержит тяжелую цепь антитела.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат (а) вариабельную область тяжелой цепи, содержащую CDR1-область из SEQ ID NO:9, где I34 представляет собой S, L, V А или I и N35 представляет собой N, Т или S; CDR2-область из SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L; A62 представляет собой А или S и L63 представляет собой L или V; и CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G; где Y110 представляет собой Y, K, S, R или Т; и (б) вариабельную область легкой цепи, содержащую CDR1-область из SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; CDR2-область из SEQ ID NO:13, где 151 представляет собой I, Т, V или А и S56 представляет собой S или Т; и CDR3-область из SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R. В некоторых воплощениях вариабельная область легкой цепи содержит CDR3-область из SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой любую аминокислоту, Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R. В некоторых воплощениях вариабельная область тяжелой цепи содержит CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где 8105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G, где Y110 представляет собой любую аминокислоту. В других воплощениях вариабельная область тяжелой цепи содержит CDRS-область из SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т, где G99 представляет собой G, S, А, С, V, N, D или Т, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F; где F108 представляет собой F или W; где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту. В некоторых воплощениях полипептид дополнительно содержит легкую цепь антитела.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело, включая гуманизированное антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:9, где I34 представляет собой S, L, V А или I и N35 представляет собой N, Т или S; аминокислотную последовательность, показанную в SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L, A62 представляет собой А или S и L63 представляет собой L или V; и аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G, где Y110 представляет собой Y, K, S, R или Т. В некоторых воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R и где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту. В других воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т, где G99 представляет собой G, S, А, С, V, N, D или Т, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту. В некоторых воплощениях полипептид (такой как антитело) дополнительно содержит вариабельную область легкой цепи антитела.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат аминокислотную последовательность, показанную в SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; аминокислотную последовательность, показанную в SEQ ID NO:13, где I51 представляет собой I, Т, V или А и S56 представляет собой S или Т; и аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R. В некоторых воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой любую аминокислоту, Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R. В некоторых воплощениях полипептид (такой как антитело) дополнительно содержит вариабельную область тяжелой цепи антитела.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), которые содержат (а) аминокислотную последовательность, показанную в SEQ ID NO:9, где I34 представляет собой S, L, V, А или I и N35 представляет собой N, Т или S; аминокислотную последовательность, показанную в SEQ ID NO:10, где М50 представляет собой М, I, G, Q, S или L, А62 представляет собой А или S и L63 представляет собой L или V; и аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G и где Y110 представляет собой Y, K, S, R или Т; и (б) аминокислотную последовательность, показанную в SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; аминокислотную последовательность, показанную в SEQ ID NO:13, где I51 представляет собой I, Т, V или А, и S56 представляет собой S или Т; и аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R. В некоторых воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой любую аминокислоту, Т93 представляет собой любую аминокислоту и где Y96 представляет собой Y или R. В некоторых воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G, где Y110 представляет собой любую аминокислоту. В других воплощениях полипептид содержит аминокислотную последовательность, показанную в SEQ ID NO:11, где G98 представляет собой G, S, А, С, V, N, D или Т, где G99 представляет собой G, S, А, С, V, N, D или Т, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой S, А, С, G, D, N, Т или G и где Y110 представляет собой любую аминокислоту. В некоторых воплощениях полипептид дополнительно содержит вариабельную область легкой цепи антитела.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий вариабельную область тяжелой цепи, содержащую: (а) CDR1-область из SEQ ID NO:9, где I34 представляет собой S, L, V А или I и N35 заменена на N, Т или S; (б) CDR2-область из SEQ ID NO:10, где М50 представляет собой I, G, Q, S или L, A62 представляет собой А или S и L63 представляет собой L или V; и (в) CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G и где Y110 представляет собой Y, K, S, R или Т; причем антитело связывается с NGF.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитела), содержащие вариабельную область легкой цепи, содержащую: (а) CDR1-область из SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; (б) CDR2-область из SEQ ID NO:13, где I51 представляет собой I, Т, V или А и S56 представляет собой S или Т; и (в) CDR3-область из SEQ ID NO:14, где K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R; причем антитело связывается с NGF.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитела), содержащие (а) вариабельную область тяжелой цепи, содержащую: (1) CDR1-область из SEQ ID NO:9, где I34 заменена на S, L, V A или I и N35 заменена на N, Т или S; (2) CDR2-область из SEQ ID NO:10, где М50 представляет собой I, G, Q, S или L, A62 представляет собой А или S и L63 представляет собой L или V; и (3) CDR3-область из SEQ ID NO:11, где Y100 представляет собой Y, L или R, где Y101 представляет собой Y или W, где G103 представляет собой G, А или S, где Т104 представляет собой Т или S, где S105 представляет собой S, А или Т, где Y106 представляет собой Y, R, Т или М, где Y107 представляет собой Y или F, где F108 представляет собой F или W, где D109 представляет собой D, N или G, где Y110 представляет собой Y, K, S, R или Т; и (б) вариабельную область легкой цепи, содержащую: (1) CDR1-область из SEQ ID NO:12, где S26 представляет собой S или F, D28 представляет собой D, S, А или Y и Н32 представляет собой Н, N или Q; (2) CDR2-область из SEQ ID NO:13, где I51 представляет собой I, Т, V или А и S56 представляет собой S или Т; и (3) CDR3-область из SEQ ID NO:14, где S91 представляет собой S или Е, K92 представляет собой K, Н, R или S и где Y96 представляет собой Y или R; причем антитело связывается с NGF.
Если не указано иначе, выбор (например замена) аминокислоты в одном положении не зависит от выбора аминокислоты в любом другом положении.
В некоторых воплощениях полинуклеотиды (такие как антитело) связываются с NGF (таким как человеческий NGF). В некоторых воплощениях полипептиды содержат любую конфигурацию CDR (включая комбинации, вариации и так далее), описанную здесь.
Как это следует из данного описания, нумерация вариабельной области, использованная здесь, представляет собой нумерацию последовательности. Специалист в данной области легко поймет то, что существует множество систем нумерации для антител (таких как нумерации по Kabat и Chothia), и то, как перевести нумерацию последовательности в другую систему нумерации, такую как нумерацию по Kabat или нумерацию по Chothia.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий аминокислотную последовательность (такую как CDR3-последовательность), выбранную из SEQ ID NO:46 или 50. В еще других воплощениях полипептид дополнительно содержит одну или более чем одну аминокислотную последовательность, показанную в SEQ ID NOS:3, 4, 5, 6, 7 и 8. В еще других воплощениях полипептид дополнительно содержит одну из дополнительных аминокислотных последовательностей, показанных в SEQ ID NOS:9, 10, 11, 12, 13, 14 и 15.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий аминокислотную последовательность (такую как CDR-область, такая как область CDR-H1 и/или CDR-H2), выбранную из (a) SEQ ID NOS:28 и/или 29; (б) SEQ ID NOS:30 и/или 31; (в) SEQ ID NOS:32 и/или 33; (г) SEQ ID NOS:34 и/или 35; (д) SEQ ID NOS:36 и/или 37; (e) SEQ ID NOS:38 и/или 39 и (ж) SEQ ID NOS:40 и 41. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-H1), выбранную из SEQ ID NOS:28, 30, 32, 34, 36, 38 и 40. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-H2), выбранную из SEQ ID NOS:29, 31, 33, 35, 37, 39 и 41. В еще других воплощениях полипептид дополнительно содержит одну или более чем одну аминокислотную последовательность, показанную в SEQ ID NOS:3, 4, 5, 6, 7 и 8. В еще других воплощениях полипептид дополнительно содержит одну из дополнительных аминокислотных последовательностей, показанных в SEQ ID NOS:9, 10, 11, 12, 13, 14 и 15.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий аминокислотную последовательность (такую как CDR-область, такая как область CDR-L1 и/или CDR-L2), выбранную из (a) SEQ ID NOS:18 и/или 19; (б) SEQ ID NOS:20 и/или 21 и (в) SEQ ID NOS:22 и/или 23. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-L1), выбранную из SEQ ID NOS: 18, 20 и 22. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-L2), выбранную из SEQ ID NOS:19, 21 и 23. В еще других воплощениях полипептид дополнительно содержит одну или более чем одну аминокислотную последовательность, показанную в SEQ ID NOS:3, 4, 5, 6, 7, 8. В еще других воплощениях полипептид дополнительно содержит одну из дополнительных аминокислотных последовательностей, показанных в SEQ ID NOS:9, 10, 11, 12, 13, 14, и 15.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий аминокислотную последовательность (такую как CDR-область, такая как область CDR-L3 и/или CDR-H3), выбранную из (a) SEQ ID NOS:51 и/или 52; (б) SEQ ID NOS:55 и/или 56; (в) SEQ ID NOS:57 и/или 58; (в) SEQ ID NOS:59 и/или 60; (г) SEQ ID NOS:61 и/или 62; (д) SEQ ID NOS:63 и/или 64. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-L3), выбранную из SEQ ID NOS:51, 55, 57, 59, 61 и 63. В некоторых воплощениях полипептид содержит аминокислотную последовательность (такую как область CDR-H3), выбранную из SEQ ID NOS:52, 56, 58, 60, 62 и 64. В еще других воплощениях полипептид дополнительно содержит аминокислотную последовательность, показанную в одном или более чем одном из SEQ ID NOS:18, 19, 30 и 31. В еще других воплощениях полипептид дополнительно содержит одну или более чем одну аминокислотную последовательность, показанную в SEQ ID NOS:3, 4, 5, 6, 7 и 8. В еще других воплощениях полипептид дополнительно содержит одну из дополнительных аминокислотных последовательностей, показанных в SEQ ID NOS:9, 10, 11, 12, 13, 14 и 15.
В другом аспекте в этом изобретении предложен полипептид (такой как антитело), содержащий одну или более чем одну аминокислотную последовательность (такую как CDR-область), показанную в SEQ ID NOS:61, 63, 18, 19, 30 и 31.
В одном из аспектов в этом изобретении предложено антитело против NGF (такое как антагонистическое антитело), которое связывается с NGF (таким как человеческий NGF) с высокой аффинностью. В некоторых воплощениях высокая аффинность представляет собой: (а) связывание с NGF при KD менее чем приблизительно 2 нМ (такой как любое из значений около 1 нМ, 800 пМ, 600 пМ, 400 пМ, 200 пМ, 100 пМ, 90 пМ, 80 пМ, 70 пМ, 60 пМ, 50 пМ или менее чем 50 пМ) и/или при koff менее чем приблизительно 6×10-5 сек-1; и/или (б) ингибирование (снижение и/или блокирование) зависимого от человеческого NGF выживания мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии около 15 пМ NGF) при любом значении около 200 пМ, 150 пМ, 100 пМ, 80 пМ, 60 пМ, 40 пМ, 20 пМ, 10 пМ или менее чем 10 пМ; и/или (в) ингибирование (снижение и/или блокирование) зависимого от человеческого NGF выживания мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 1,5 пМ NGF) при любом значении около 50 пМ, 40 пМ, 30 пМ, 10 пМ, 20 пМ, 10 пМ, 5 пМ, 2 пМ, 1 пМ или менее чем 1 пМ; и/или (г) ингибирование (снижение и/или блокирование) зависимого от крысиного NGF выживания мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 15 пМ NGF) при любом значении около 150 пМ, 125 пМ, 100 пМ, 80 пМ, 60 пМ, 40 пМ, 30 пМ, 20 пМ, 10 пМ, 5 пМ или менее чем 5 пМ; и/или (д) ингибирование (снижение и/или блокирование) зависимого от крысиного NGF выживания мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 1,5 пМ NGF) при любом значении около 30 пМ, 25 пМ, 20 пМ, 15 пМ, 10 пМ, 5 пМ, 4 пМ, 3 пМ, 2 пМ, 1 пМ или менее чем 1 пМ; и/или (е) связывание с NGF с более высокой аффинностью, чем при связывании TrkA-рецептора.
В другом аспекте в этом изобретении предложены полипептиды (такие как антитело), причем эти полипептиды: (а) связываются с NGF (таким как человеческий NGF) при KD менее чем приблизительно 2 нМ (такой как любое из значений около 1 нМ, 800 пМ, 600 пМ, 400 пМ, 200 пМ, 100 пМ, 90 пМ, 80 пМ, 70 пМ, 60 пМ, 50 пМ или менее чем 50 пМ) и/или при koff менее чем приблизительно 6×10-5 сек-1; и/или (б) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 15 пМ NGF) при любом значении около 200 пМ, 150 пМ, 100 пМ, 80 пМ, 60 пМ, 40 пМ, 20 пМ, 10 пМ или менее чем 10 пМ; и/или (в) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 1,5 пМ NGF) при любом значении около 50 пМ, 40 пМ, 30 пМ, 10 пМ, 20 пМ, 10 пМ, 5 пМ, 2 пМ, 1 пМ или меньше чем 1 пМ; и/или связываются с NGF с более высокой аффинностью, чем при связывании ТrkА-рецептора. В некоторых воплощениях полипептиды: (а) связываются с NGF при KD менее чем приблизительно 2 нМ; и/или (б) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 приблизительно 100 пМ или менее чем 100 пМ, причем IC50 измеряют в присутствии приблизительно 15 пМ NGF; и/или (в) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 приблизительно 10 пМ или менее чем 10 пМ, причем IC50 измеряют в присутствии приблизительно 1,5 пМ NGF, причем IC50 измеряют в присутствии приблизительно 15 пМ NGF. В некоторых воплощениях полипептиды: (а) связываются с NGF при KD менее чем приблизительно 100 пМ; и/или (б) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 приблизительно 20 пМ или менее чем 20 пМ, причем IC50 измеряют в присутствии приблизительно 15 пМ NGF; и/или (в) ингибируют зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 приблизительно 2 пМ или менее чем 2 пМ, причем IC50 измеряют в присутствии приблизительно 1,5 пМ NGF.
Как это следует из данного описания, из этого изобретения конкретно исключены воплощения, представляющие собой полипептиды, состоящие из аминокислотной последовательности, идентичной аминокислотной последовательности мышиного моноклонального антитела 911. Последовательности расширенных CDR моноклонального антитела (Mab) 911 показаны на Фиг.1А и 1Б и в SEQ ID NOS:9-14.
В некоторых воплощениях в этом изобретении предложены любые из указанных полипептидов или антител, причем дополнительно полипептид (такой как антитело) выделен. В некоторых воплощениях полипептид (такой как антитело) по существу очищен. В еще других воплощениях полипептид (такой как антитело) имеет развитую аффинность. В других воплощениях антитело является антагонистическим антителом. В некоторых воплощениях полипептид (такой как антитело) содержит последовательности каркасной области, не являющейся человеческой. В еще других воплощениях полипептид (такой как антитело) содержит один или более чем один остаток каркасной области, не являющейся человеческой. В некоторых воплощениях полипептид (такой как антитело) связывается с NGF (таким как человеческий NGF) при KD 2 нМ или менее чем 2 нМ. В некоторых воплощениях полипептид содержит одну или более чем одну (например, 2, 3, 4, 5, 6, 7, 8 или более) замену человеческой аминокислотой относительно аминокислотной последовательности, не являющейся человеческой (такой как последовательность вариабельной области, такой как последовательность CDR, такая как последовательность каркасной области). В некоторых воплощениях полипептид содержит по меньшей мере 1, по меньшей мере 2 или более, например по меньшей мере 3, 4, 5, 6 или более, замен аминокислот относительно аминокислотной последовательности исходного полипептида (такой как аминокислотная последовательность антитела 911, такая как любая одна или более чем одна из SEQ ID NOS:9-14). В некоторых воплощениях аффинность связывания антитела изменена (в некоторых воплощениях повышена) относительно аффинности исходного антитела (такого как моноклональное антитело 911). В еще других воплощениях аффинность связывания антитела ниже, чем аффинность связывания TrkA-рецептора для NGF (такого как человеческий NGF). В некоторых воплощениях полипептиды могут быть антителами. В некоторых воплощениях антитела являются человеческими антителами. В других воплощениях антитела являются гуманизированными антителами. В еще других воплощениях антитела являются моноклональными антителами. В некоторых воплощениях антитело представляет собой антитело с развитой аффинностью.
В этом изобретении предложены полинуклеотиды (в том числе выделенные полинуклеотиды), содержащие полинуклеотиды, кодирующие любые указанные воплощения.
В другом аспекте в этом изобретении предложен выделенный полинуклеотид, содержащий полинуклеотид, кодирующий фрагмент или область антитела Е3 (взаимозаменяемым образом обозначаемого здесь как "Е3"). В одном из воплощений этот фрагмент представляет собой легкую цепь антитела Е3, как показано на Фиг.1Б. В еще одном воплощении этот фрагмент представляет собой тяжелую цепь антитела Е3, как показано на Фиг.1А. В еще одном воплощении этот фрагмент содержит одну или более чем одну вариабельную область легкой цепи и/или тяжелой цепи антитела Е3. В еще одном воплощении этот фрагмент содержит один или более чем один гипервариабельный участок (CDR) легкой цепи и/или тяжелой цепи антитела Е3, как показано на Фиг.1А и 1Б.
В другом аспекте это изобретение представляет собой выделенный полинуклеотид, содержащий полинуклеотид, который кодирует антитело Е3. В некоторых воплощениях полинуклеотид содержит какой-либо один или оба из полинуклеотидов, показанных на Фиг.2 и 3.
В другом аспекте это изобретение представляет собой выделенный полинуклеотид, который кодирует легкую цепь Е3, с номером депонирования АТСС № РТА-4893 или АТСС № РТА-4894. В другом аспекте это изобретение представляет собой выделенный полинуклеотид, который кодирует тяжелую цепь Е3, с номером депонирования АТСС № РТА-4895. В еще одном аспекте это изобретение представляет собой выделенный полинуклеотид, содержащий (а) вариабельную область, кодируемую полинуклеотидом с номером депонирования АТСС № РТА-4893 или РТА-4894, и (б) вариабельную область, кодируемую полинуклеотидом с номером депонирования АТСС № РТА-4895. В другом аспекте это изобретение представляет собой выделенный полинуклеотид, содержащий (а) один или более чем один CDR, кодируемый полинуклеотидом с номером депонирования АТСС № РТА-4893 или РТА-4894; и/или (б) один или более чем один CDR, кодируемый полинуклеотидом с номером депонирования АТСС № РТА-4895.
В другом аспекте в этом изобретении предложены полинуклеотиды, кодирующие любые описанные здесь антитела (включая фрагменты антител) или полипептиды.
В другом аспекте в этом изобретении предложены векторы (включая векторы экспрессии и клонирования) и клетки-хозяева, содержащие любой из раскрытых здесь полинуклеотидов.
Как это следует из данного здесь описания, из этого изобретения конкретно исключены воплощения, представляющие собой полинуклеотиды, состоящие из полинуклеотидных последовательностей, идентичных полинуклеотидной последовательности мышиного моноклонального антитела 911. Последовательности расширенных CDR моноклонального антитела 911 показаны на Фиг.1А и 1Б и в SEQ ID NOS:9-14.
В другом аспекте это изобретение представляет собой клетку-хозяина, содержащую полинуклеотид, кодирующий легкую цепь Е3, и полинуклеотид, кодирующий тяжелую цепь Е3, где полинуклеотид, кодирующий легкую цепь Е3, имеет номер депонирования АТСС № РТА-4893 и/или АТСС № РТА-4894, и полинуклеотид, кодирующий тяжелую цепь Е3, имеет номер депонирования АТСС № РТА-4895. В некоторых воплощениях клетка-хозяин содержит полинуклеотид, содержащий (а) вариабельную область, кодируемую полинуклеотидом с номером депонирования АТСС № РТА-4893 или РТА-4894, и/или (б) вариабельную область, кодируемую полинуклеотидом с номером депонирования АТСС № РТА-4895. В некоторых воплощениях клетка-хозяин содержит полинуклеотид, кодирующий (а) один или более чем один CDR, кодируемый полинуклеотидом с номером депонирования АТСС № РТА-4893 или РТА-4894; и/или (б) один или более чем один CDR, кодируемый полинуклеотидом с номером депонирования АТСС № РТА-4895. В некоторых воплощениях клетка-хозяин является клеткой млекопитающего.
В другом аспекте это изобретение представляет собой комплекс NGF, связанный с антителом Е3. В другом аспекте этот комплекс выделен. В другом аспекте этот комплекс по существу очищен.
В другом аспекте это изобретение представляет собой комплекс NGF, связанный с любым описанным здесь антителом или полипептидом. В другом аспекте этот комплекс выделен. В другом аспекте этот комплекс по существу очищен.
В другом аспекте это изобретение представляет собой фармацевтическую композицию, содержащую любой описанный здесь полипептид (включая антитела, такие как антитело Е3) или полинуклеотид, такую как фармацевтические композиции, содержащие антитело Е3 или антитело, содержащее фрагмент антитела Е3, и фармацевтически приемлемый эксципиент.
В другом аспекте это изобретение представляет собой способ получения антитела Е3, включающий получение клетки-хозяина, содержащей экспрессионный вектор, который кодирует антитело Е3; культивирование клетки-хозяина или ее потомства в условиях, которые делают возможной продукцию антитела Е3; и очистку антитела Е3. В некоторых воплощениях экспрессионный вектор содержит одну или обе из полинуклеотидных последовательностей, показанных на Фиг.2 и 3.
В другом аспекте это изобретение представляет собой способ получения антитела Е3, включающий экспрессию полинуклеотида, кодирующего легкую цепь Е3, и полинуклеотида, кодирующего тяжелую цепь Е3, в подходящей клетке, где полинуклеотид, кодирующий легкую цепь Е3, имеет номер депонирования АТСС № РТА-4893 и/или АТСС № РТА-4894, и полинуклеотид, кодирующий тяжелую цепь Е3, имеет номер депонирования АТСС № РТА-4895; обычно с последующим извлечением и/или выделением антитела.
В другом аспекте в этом изобретении предложены способы получения любых описанных здесь полипептидов (таких как антитела) путем экспрессии одного или более чем одного полинуклеотида, кодирующего антитело (причем с одного вектора можно экспрессировать легкую или тяжелую цепь в отдельности или можно экспрессировать обе цепи, легкую и тяжелую) в подходящей клетке, обычно с последующим извлечением и/или выделением антитела или полипептидов, представляющих интерес.
В другом аспекте это изобретение представляет собой способ антагонизации биологической активности NGF (такого как человеческий NGF) с использованием любого раскрытого здесь полипептида (включая антитело, такое как антитело Е3). В одном из воплощений этого способа человеческий фактор роста нервов приводят в контакт с любым описанным здесь полипептидом (включая антитело Е3), тем самым активность NGF (такая как активность человеческого фактора роста нервов) антагонизируется, снижается, блокируется или подавляется.
В другом аспекте это изобретение представляет собой способ детектирования NGF с использованием любого описанного здесь полипептида (включая антитела, такие как антитело Е3). Присутствие NGF обнаруживают путем детектирования комплекса между NGF и любым описанным здесь полипептидом (таким как антитело Е3). Термин "детектирование", как он использован здесь, включает проведение качественного и/или количественного обнаружения (измерение уровней) относительно или безотносительно контроля.
В другом аспекте это изобретение представляет собой способ лечения боли путем введения эффективного количества композиции, содержащей антитело Е3 или любого описанного здесь полипептида (включая антитело) или воплощения, представляющего собой полинуклеотид. В некоторых воплощениях боль представляет собой послеоперационную боль.
В другом аспекте это изобретение представляет собой способ предотвращения или лечения боли, вызванной ревматоидным артритом, у индивида путем введения указанному индивиду эффективного количества антагонистического антитела против NGF. Было показано, что антагонистическое антитело против NGF в соответствии с этим изобретением способно ингибировать или блокировать боль, связанную с ревматоидным артритом. В некоторых воплощениях боль облегчают в пределах около 24 часов после введения антагонистического антитела против NGF. В некоторых воплощениях боль облегчают в пределах около 4 суток после введения антагонистического антитела против NGF. В некоторых воплощениях боль облегчают перед тем, как наблюдают улучшение воспалительного состояния у индивида, или в отсутствие указаний на такое улучшение.
В другом аспекте в этом изобретении предложены способы для снижения частоты случаев боли, вызванной ревматоидным артритом, для облегчения боли, вызванной ревматоидным артритом, для подавления боли, вызванной ревматоидным артритом, для облегчения проявлений боли, вызванной ревматоидным артритом, и/или для замедления начала, развития или прогрессирования боли, вызванной ревматоидным артритом, у индивида, включающие введение указанному индивиду эффективного количества антагонистического антитела против NGF.
В другом аспекте в этом изобретении предложены способы лечения воспалительной кахексии (потери массы тела), связанной с ревматоидным артритом, у индивида, включающие введение эффективного количества антагонистического антитела против NGF.
В другом аспекте это изобретение представляет собой способ предотвращения или лечения боли, вызванной остеоартритом, у индивида путем введения указанному индивиду эффективного количества антагониста фактора роста нервов (такого как антагонистическое антитело против NGF).
В другом аспекте в этом изобретении предложены способы снижения частоты случаев боли, вызванной остеоартритом, облегчения боли, вызванной остеоартритом, подавления боли, вызванной остеоартритом, облегчения проявлений боли, вызванной остеоартритом, и/или замедления начала, развития или прогрессирования боли, вызванной остеоартритом, у индивида, включающие введение указанному индивиду эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF).
В другом аспекте в этом изобретении предложены способы улучшения физической функции у индивида, имеющего остеоартрит, включающие введение указанному индивиду эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF).
В другом аспекте в этом изобретении предложены способы улучшения ригидности у индивида, имеющего остеоартрит, включающие введение указанному индивиду эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF).
В некоторых воплощениях индивидом является человек. В некоторых воплощениях для лечения боли, вызванной остеоартритом, частота введения антагонистического антитела против NGF составляет от одного раза в неделю до одного раза в 10 недель или еще реже.
В другом аспекте в этом изобретении предложены наборы и композиции, содержащие любую одну или более чем одну описанную здесь композицию. Эти наборы, обычно в подходящей упаковке и снабженные соответствующими инструкциями, являются полезными для любых описанных здесь способов. В этом изобретении также предложены фармацевтические композиции для применения в любом описанном здесь способе, причем эти композиции содержат эффективное количество антагониста NGF (такого как антитело против NGF) и фармацевтически приемлемый носитель.
В этом изобретении также предложены любые композиции и наборы, описанные для любого описанного здесь применения в контексте использования в качестве лекарственного средства и/или использования для изготовления лекарственного средства.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГ.1А показывает аминокислотную последовательность вариабельной области тяжелой цепи антитела Е3 (обозначена как "6" и "5+Н3 с развитой аффинностью"). CDR по Chothia и по Kabat представлены подчеркнутым текстом и жирным курсивом соответственно. Фиг.1А также показывает выравнивание следующих аминокислотных последовательностей вариабельных областей тяжелой цепи; (2) акцепторная последовательность человеческой зародышевой линии VH4-59 (обозначена как "VH4-59" или "2") (SEQ ID NO:69); (3) акцепторные последовательности с трансплантированным расширенным CDR мышиного антитела 911 (обозначены как "с трансплантированным CDR" или "3") (SEQ ID NO:70); (4) акцепторные последовательности с трансплантированным CDR, включая замену V71K (обозначены как "3+одна мутация каркасной области" или "4") (SEQ ID NO:71); (5) клон, содержащий Н1 и Н2 из CDR с развитой аффинностью (обозначен как "5" или "4+Н1, Н2 с развитой аффинностью") (SEQ ID NO:72); и антитело Е3 (как описано выше).
ФИГ.1Б показывает аминокислотную последовательность вариабельной области легкой цепи антитела Е3 (обозначена как "5" или "4+ L3 с развитой аффинностью). CDR Chothia и CDR по Kabat обозначены подчеркнутым текстом и жирным курсивом соответственно. Фиг.1Б также показывает выравнивание следующих аминокислотных последовательностей вариабельной области легкой цепи: (2) акцепторная последовательность человеческой зародышевой лини O8 (обозначена как "O8" или "2") (SEQ ID NO:73); (3) акцепторные последовательности с трансплантированным расширенным CDR мышиного антитела 911 (обозначены как "с трансплантированным CDR" или "3") (SEQ ID NO:74); (4) акцепторные последовательности с трансплантированным CDR (обозначены как "3+ L1, L2 с развитой аффинностью" или "4") (SEQ ID NO:75); (5) клон, содержащий L1 и L2 CDR с развитой аффинностью (обозначены как "5" или "4+L3 с развитой аффинностью"); и антитело Е3 (как описано выше).
ФИГ.2 показывает полинуклеотид, содержащий полинуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи антитела Е3 (SEQ ID NO:76).
ФИГ.3 показывает полинуклеотид, содержащий полинуклеотидную последовательность, кодирующую вариабельную область легкой цепи антитела Е3 (SEQ ID NO:77).
ФИГ.4 представляет собой график, показывающий зависимое от NGF выживание нейронов Е13.5 в присутствии вариабельной концентрации человеческого и крысиного NGF. Ось Х соответствует концентрации NGF (нг/мл), ось Y соответствует числу нейронов.
ФИГ.5 представляет собой график, сравнивающий эффект блокирования NGF различными Fab в присутствии либо 0,04 нг/мл человеческого NGF (приблизительно 1,5 пМ; показано в нижней панели), либо 0,4 нг/мл человеческого NGF (приблизительно 15 пМ; показано в верхней панели). Оценивали выживание мышиных нейронов тройничного нерва Е13.5 при различных концентрациях Fab Е3, Fab мышиных 911 и Fab H19-L129 и Fab 8L2-6D5. IC50 (в пМ) вычисляли для каждого Fab при каждой концентрации NGF и представили в Таблице 9. Fab Е3 сильно блокировал зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва при IC50 приблизительно 21 пМ в присутствии 15 пМ человеческого NGF и при IC50 приблизительно 1,2 пМ в присутствии 1,5 пМ человеческого NGF. Fab 3С и Н19-L129 также сильно блокировали зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва. На обеих панелях ось Х соответствует концентрации антитела (нМ), ось Y соответствует числу нейронов. 1,5 пМ NGF приблизительно соответствовали IC50, тогда как 15 пМ представляли собой насыщающую концентрацию NGF.
ФИГ.6 представляет собой график, сравнивающий эффект блокирования NGF различными Fab в присутствии либо 0,04 нг/мл крысиного NGF (приблизительно 1,5 пМ; показано на нижней панели), либо 0,4 нг/мл крысиного NGF (приблизительно 15 пМ; показано на верхней панели). Выживание мышиных нейронов тройничного нерва Е13.5 при различных концентрациях Fab Е3, Fab мышиного 911 и Fab H19-L129 и 8L2-6D5 оценивали, как описано выше. IC50 (в пМ) вычисляли для каждого Fab при каждой концентрации NGF и представили в Таблице 9. Fab Е3 сильно блокировал зависимое от человеческого NGF выживание нейронов тройничного нерва при IC50 приблизительно 31,6 пМ в присутствии 15 пМ крысиного NGF и при IC50 приблизительно 1,3 пМ в присутствии 1,5 пМ крысиного NGF. Fab 3С и H19-L129 также сильно блокировали зависимое от крысиного NGF выживание нейронов тройничного нерва. 1,5 пМ NGF приблизительно соответствовало IC50, тогда как 15 пМ представляло собой насыщающую концентрацию NGF. На обеих панелях ось Х соответствует концентрации антитела (нМ), ось Y соответствует числу нейронов.
ФИГ.7 представляет собой график, показывающий боль в покое, оцененную через 24 часа после хирургической операции, и показывающий, что лечение антителом Е3 против NGF при 0,02 мг/кг, 0,1 мг/кг, 0,6 мг/кг или 1 мг/кг снижало боль. Знак "*" свидетельствует о статистически достоверном отличии (p<0,5) от отрицательного контроля.
ФИГ.8 представляет собой график, показывающий боль в покое, оцененную через 24 часа после хирургической операции, и показывающий, что лечение антителом Е3 против NGF при 0,5 мг/кг достоверно (p<0,005) снижало боль в покое при инъецировании через два часа после хирургической операции.
ФИГ.9 представляет собой график, показывающий результат анализа BIAcore для аффинности связывания мышиного антитела 911 (Fab) с человеческим NGF. Мышиное антитело 911 связывалось с NGF при KD 3,7 нМ, koff 8,4×10-5 сек-1 и kon 2,2×104 мсек-1.
ФИГ.10 представляет собой график, показывающий результат анализа BIAcore для аффинности связывания антитела Е3 (Fab) (обозначен как "Fab 3Е") с человеческим NGF. Е3 связывалось с человеческим NGF при KD приблизительно 0,07 нМ (и при kon приблизительно 6,0×105 М-1 сек-1 и koff приблизительно 4,2×10-5 сек-1).
ФИГ.11 представляет собой график, показывающий, что антитело Е3 блокирует взаимодействие NGF с его рецепторами, TrkA и р75, как оценено по детектированному проценту связывания между NGF и TrkA (показано черными кружками) и между NGF и р75 (показано пустыми квадратами). Ось Х соответствует концентрации антитела 3Е (Fab), ось Y соответствует связыванию NGF (максимальный процент, относительные единицы, ОЕ). Повышенные концентрации Fab Е3 блокировали взаимодействие NGF как с р75, так и с TrkA, как показано сниженным сигналом (измерен в ОЕ). Когда концентрация антитела Е3 (Fab) равна концентрации NGF, связывание NGF не наблюдали (как показано нулевым сигналом).
ФИГ.12 представляет собой график, показывающий способность полного антитела Е3 и Fab Е3 блокировать человеческий NGF. Оценивали выживание мышиных нейронов тройничного нерва Е13.5 в присутствии человеческого NGF и различных концентраций Fab Е3 и антитела Е3. Ось Х соответствует сайтам связывания NGF (нМ), ось Y соответствует нормализованному числу нейронов тройничного нерва (ТН). Полное антитело Е3 и Fab 3Е показали одинаковые уровни ингибирования зависимого от NGF выживания нейронов тройничного нерва, когда концентрации целого антитела и Fab были нормализованы относительно числа сайтов связывания NGF (Fab имеет один сайт связывания, целое антитело имеет два сайта связывания).
ФИГ.13 представляет собой график, показывающий способность различных концентраций (20, 4, 0,8, 0,16, 0,032, 0,0064, 0,00128 и 0,0 нМ) антитела Е3 (сплошные треугольники; обозначено как "3Е"), антитела 911 (сплошные кружки) и иммуноадгезина TrkA-рецептора (затемненные квадраты; обозначено как "TrkA-Fc") ингибировать зависимое от NGF выживание нейронов тройничного нерва Е13.5 в присутствии 0,4 нг/мл человеческого NGF (условия насыщения). Ось Х соответствует концентрации антитела (нМ), ось Y соответствует числу нейронов. Эти результаты продемонстрировали, что антитело Е3 блокировало NGF значительно лучше, чем мышиное моноклональное антитело против NGF 911 или TrkA иммуноадгезин.
ФИГ.14 представляет собой график, показывающий, что антагонистическое антитело Е3 против NGF (обозначено на чертеже как "3Е") или Fab 911 не ингибировало выживание нейронов, которому способствовали NT3, NT4/5 и MSP, даже при таких высоких концентрациях антитела, как 200 нМ. Для каждого эксперимента данные представляли собой средней процент выживания после 48 часов в культуре (Стандартная ошибка среднего, n=3 для каждой данной точки) относительно выживания, которое наблюдали в положительном контроле (100% выживание нейронов тройничного нерва, выращиваемых в присутствии насыщающей концентрации NGF). Различные концентрации (20 нМ, 2 нМ или 0,2 нМ) Fab Е3 (обозначен на чертеже как "3Е") и Fab мышиного антитела 911 использовали в отсутствие добавленного нейротрофина (обозначено как "контроль"), с 400 пМ NGF (обозначено как "NGF-400 пМ"), с 10 нМ NT3 (обозначено как "NT3-10HM") или с 600 пМ MSP (обозначено как "MSP-600 пМ").
ФИГ.15 представляет собой график, показывающий, что антагонистическое антитело Е3 против NGF (Fab или полное антитело) (обозначено на чертеже как "3Е") или мышиное антитело 911 (Fab или полное антитело) не ингибирует выживание нейронов, которому способствуют NT3, NT4/5 и MSP, даже при таких высоких концентрациях антитела, как 200 нМ. Различные концентрации (200 нМ и 80 нМ) Fab Е3 и полного антитела и мышиного антитела 911 (полное антитело и Fab) использовали в отсутствие добавленных нейротрофинов (обозначено как "без фактора"), с 400 пМ NGF (обозначено как "NGF-400 пМ"), с 10 нМ NT3 (обозначено как "NT3-10 нМ") или с 600 пМ MSP (обозначено как "MSP-600 пМ").
ФИГ.16 представляет собой график, показывающий, что антагонистическое антитело Е3 против NGF или Fab Е3 не ингибирует выживание нейронов нодозных ганглиев Е17, которому способствует BDNF, NT4/5 или LIF. Также тестировали мышиное антагонистическое антитело 911 против NGF и наблюдали такой же результат. Различные концентрации (200 нМ или 80 нМ) полного антитела Е3 (обозначено на чертеже как "3Е"), Fab ЕЗ, полного антитела 911 или Fab 911 тестировали в отсутствие добавленных нейротрофинов (обозначено как "без факторов"), с 400 пМ BDNF (обозначено как "BDNF-400 пМ"), с 400 пМ NT4/5 (обозначено как "NT4/5-400 пМ") или с 2,5 нМ LIF (обозначено как "LIF-2,5 нМ").
ФИГ.17 представляет собой график, показывающий, что антагонистическое антитело Е3 против NGF или Fab Е3 не ингибирует выживание нейронов нодозных ганглиев Е17, которому способствует BDNF, NT4/5 или LIF. Различные концентрации (200 нМ, 20 нМ, 2 нМ) Fab Е3 (обозначено на чертеже как "3Е") или Fab 911 тестировали в отсутствие добавленных нейротрофинов (обозначено как "контроль"), с 400 пМ BDNF (обозначено как "BDNF-400 пМ"), с 400 пМ NT4/5 (обозначено как "NT4/5-400 пМ") или с 2,5 нМ LIF (обозначено как "LIF-2,5 нМ").
ФИГ.18 представляет собой график, демонстрирующий ноцицептивный ответ у крыс с артритом (модель ревматоидного артрита) после введения антител против NGF (Е3 и 911) на сутки 14 и сутки 19. Мышам с артритом вводили Е3 (1 мг/кг внутривенно на сутки 14 и на сутки 19), 911 (10 мг/кг внутривенно на сутки 14 и на сутки 19) или индометацин (3 мг/кг перорально ежедневно в течение 10 суток). Величины интенсивности издаваемых звуков выражали в мВ как среднее ± стандартная ошибка среднего.
ФИГ.19 представляет собой график, демонстрирующий влияние антител против NGF на массу тела у крыс с артритом (модель ревматоидного артрита) после введения антител против NGF на сутки 14 и на сутки 19. Мышам с артритом вводили Е3 (1 мг/кг внутривенно на сутки 14 и на сутки 19), 911 (10 мг/кг внутривенно на сутки 14 и на сутки 19) или индометацин (3 мг/кг перорально ежедневно в течение 10 суток). Величины массы тела выражали в граммах как среднее ± стандартная ошибка среднего.
ФИГ.20 представляет собой график, демонстрирующий ноцицептивный ответ у крыс с артритом (модель ревматоидного артрита) после введения разных доз антитела Е3 против NGF (0,003 мг/кг, 0,03 мг/кг, 0,3 мг/кг и 5 мг/кг) на сутки 14 и на сутки 18. Величины интенсивности издаваемых звуков выражены в мВ как среднее ± стандартная ошибка среднего.
ФИГ.21 представляет собой график, демонстрирующий влияние антитела Е3 против NGF на процент массы на сутки 14 (нормализован относительно суток 14) у крыс с артритом (модель ревматоидного артрита) после введения разных доз антитела Е3 против NGF (0,03 мг/кг, 0,3 мг/кг и 5 мг/кг) на сутки 14 и на сутки 18.
ФИГ.22 представляет собой график, демонстрирующий влияние антитела Е3 против NGF на потерю массы тела у крыс с артритом (модель ревматоидного артрита) после введения разных доз антитела Е3 против NGF (0,03 мг/кг, 0,3 мг/кг и 5 мг/кг) на сутки 14 и на сутки 18. Величины массы тела нормализованы относительно суток 0.
ФИГ.23 показывает аминокислотную последовательность вариабельной области тяжелой цепи Е3 (Фиг.23А) и аминокислотную последовательность вариабельной области легкой цепи Е3 (Фиг.23Б) при использовании последовательной нумерации, нумерации по Kabat и нумерации по Chothia.
ФИГ.24 показывает изменения в средней ежедневной интенсивности боли после введения антитела Е3 против NGF при сравнении с исходным уровнем на сутки 0. Ось Y соответствует снижению средней ежедневной интенсивности боли (баллы по шкале VAS) при сравнении со средней ежедневной интенсивностью боли на сутки 0. Ось Х соответствует суткам после введения антитела Е3 против NGF.
ФИГ.25 показывает средние баллы по шкале VAS после введения антитела Е3 против NGF. "SE" относится к стандартной ошибке.
ФИГ.26 показывает процент максимального снижения отличия по суммарной интенсивности боли (SPID) от суток 2 по сутки 14 и от суток 2 по сутки 28 после введения антитела Е3 против NGF.
ФИГ.27 показывает среднее наименьших квадратов (LSM) ответа для WOMAC, подшкалы боли, подшкалы физической функции и подшкалы ригидности от суток 1 по сутки 28 после введения разных доз (3 мкг/кг, 10 мкг/кг, 30 мкг/кг, 100 мкг/кг и 300 мкг/кг) антитела Е3 против NGF. "SE" относится к стандартной ошибке. Оси Х соответствуют введенной дозе антитела Е3 против NGF. Знак "*" указывает на Р<0,05 по критерию Даннетта (Dunnett) при сравнении с исходным уровнем.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В раскрытом здесь изобретении предложены антагонистические антитела против NGF, которые связываются с NGF (таким как человеческий NGF) с высокой аффинностью. В этом изобретении дополнительно предложены антитела и полипептиды, происходящие из Е3, которые связываются с NGF, и способы получения и применения этих антител. В некоторых воплощениях в этом изобретении предложено гуманизированное антитело, Е3, которое связывается с фактором роста нервов ("NGF"), и способы получения и применения этого антитела. В этом изобретении также предложены полипептиды Е3 (включая антитела), которые связываются с NGF, и полинуклеотиды, кодирующие антитело и/или полипептид Е3.
В раскрытом здесь изобретении также предложены способы предотвращения и/или лечения боли, вызванной ревматоидным артритом, у индивида путем введения терапевтически эффективного количества антагонистического антитела против NGF.
В раскрытом здесь изобретении также предложены способы предотвращения и/или лечения боли, вызванной остеоартритом, у индивида путем введения терапевтически эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF).
В этом изобретении также предложены способы регуляции аффинности антитела и способы характеризации области, представляющей собой CDR.
Общие методики
В практическом применении настоящего изобретения используют, если не указано иначе, общепринятые методики молекулярной биологии (в том числе рекомбинантные методики), микробиологии, клеточной биологии, биохимии и иммунологии, которые находятся в пределах знаний специалистов в данной области техники. Такие методики подробно разъясняются в литературе, такой как Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989), Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998), Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998), Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998), J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir и С.С. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); ImmunoBiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999)); Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995) и Cancer: Principles and Practice of Oncology (V.T. DeVita et al., eds., J.B. Lippincott Company, 1993).
Определения
"Антитело" представляет собой молекулу иммуноглобулина, способную специфично связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и так далее, через по меньшей мере один сайт распознавания антигена, локализованный в вариабельной области молекулы иммуноглобулина. При использовании здесь этот термин охватывает не только интактные поликлональные или моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(ab')2, Fv), отдельные цепи (ScFv), их мутанты, слитые белки, содержащие часть антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена. Антитело включает антитело любого класса, такого как IgG, IgA или IgM (или его подкласса), и антитело не обязательно относится к какому-либо конкретному классу. В зависимости от аминокислотной последовательности константного домена тяжелых цепей антитела иммуноглобулины можно относить к разным классам. Существуют пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них можно дополнительно разделить в подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, обозначают как альфа, дельта, эпсилон, гамма и мю соответственно. Структуры субъединиц и трехмерные конфигурации разных классов иммуноглобулинов хорошо известны.
"Fv" представляет собой фрагмент антитела, который содержит полный сайт распознавания и связывания антигена. В двухцепочечном варианте Fv эта область состоит из димера одного вариабельного домена тяжелой и одного вариабельного домена легкой цепи в близкой нековалентной ассоциации. В одноцепочечном варианте Fv один вариабельный домен тяжелой и один вариабельный домен легкой цепи могут быть ковалентно связаны гибким пептидным линкером, так что легкая и тяжелая цепи могут быть ассоциированы в димерную структуру, аналогичную таковой в двухцепочечном варианте Fv. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют так, что определяют специфичность связывания антигена на поверхности димера VH-VL. Однако даже единственный вариабельный домен (или половина Fv, содержащая только 3 CDR, специфичных по отношению к антигену) имеет способность распознавать и связывать антиген, хотя обычно с более низкой аффинностью по сравнению с целым связывающим сайтом.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков по карбоксильному концу тяжелой цепи СН1-домена, в том числе одного или более чем одного цистеина из шарнирных областей антитела.
"Моноклональное антитело" относится к гомогенной популяции антител, причем моноклональное антитело состоит из аминокислот (встречающихся и не встречающихся в природе), которые участвуют в селективном связывании антигена. Популяция моноклональных антител является высокоспецифичной, будучи направленной против единственного антигенного сайта. Термин "моноклональное антитело" охватывает не только интактные моноклональные антитела и полноразмерные моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одиночную цепь (ScFv), их мутанты, слитые белки, содержащие часть антитела, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена с требуемой специфичностью и способностью связываться с антигеном. Этот термин не предназначен для ограничения в отношении источника антитела или способа получения (например, гибридома, фаговая селекция, рекомбинантная экспрессия, трансгенные животные и так далее).
При использовании здесь "человеческое антитело" означает антитело, имеющее аминокислотную последовательность, соответствующую тому, что антитело продуцировано человеком и/или было получено при использовании любой методики для получения человеческого антитела, известной в данной области или раскрытой здесь. Это определение человеческого антитела включает в себя антитела, содержащие по меньшей мере один полипептид человеческой тяжелой цепи или по меньшей мере один полипептид человеческой легкой цепи. Один такой пример представляет собой антитело, содержащее полипептиды мышиной легкой цепи и человеческой тяжелой цепи. Человеческие антитела можно продуцировать при использовании различных методик, известных в данной области. В одном из воплощений человеческое антитело выбрано из фаговой библиотеки, причем эта фаговая библиотека экспрессирует человеческие антитела (Vaughan et al., 1996, Nature Biotechnology, 14:309-314; Sheets et al., 1998, PNAS, (USA) 95:6157-6162; Hoogenboom and Winter, 1991, J. Mol. Biol., 227:381; Marks et al., 1991, J. Mol. Biol., 222:581). Человеческие антитела можно также получать введением локусов человеческих иммуноглобулинов в трансгенных животных, например мышей, в которых эндогенные гены иммуноглобулинов частично или полностью инактивированы. Этот подход описан в патентах США №№5545807, 5545806, 5569825, 5625126, 5633425 и 5661016. Или же человеческое антитело можно получать иммортализацией человеческих В-лимфоцитов, которые продуцируют антитело, направленное против целевого антигена (такие В-лимфоциты могут быть извлечены из индивида или могут быть иммунизированы in vitro). См., например, Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al., 1991, J. Immunol., 147 (1):86-95; и патент США №5750373.
"Химерные антитела" относятся к тем антителам, где одна часть каждой аминокислотной последовательности тяжелой и легкой цепи гомологична соответствующим последовательностям в антителах, происходящих из конкретных видов или принадлежащих к конкретному классу, в то время как оставшийся сегмент цепей гомологичен соответствующим последовательностям в другом. В типичных случаях в этих химерных антителах вариабельная область как легкой, так и тяжелой цепи имитирует вариабельные области антитела, происходящего из одного вида млекопитающих, в то время как константные части являются гомологичными последовательностям в антителах, происходящих из другого вида. Одним явным преимуществом таких химерных форм является, например, то, что вариабельные области может быть удобно производить из уже известных источников при использовании легкодоступных гибридом или В-клеток из организмов-хозяев, не представляющих собой человека, в комбинации с константными областями, происходящими, например, из препаратов человеческих клеток. В то время как вариабельная область имеет преимущество, состоящее в легкости получения, и их специфичность не зависит от источника, константная область, будучи человеческой, с меньшей вероятностью вызывает иммунный ответ у субъекта, представляющего собой человека, при инъецировании антител, чем если бы константная область была из источника, не представляющего собой человека. Однако это определение не ограничено этим конкретным примером.
"Функциональная Fc-область" обладает по меньшей мере одной эффекторной функцией нативной последовательности Fc-области. Примеры "эффекторных функций" включают в себя связывание C1q; комплементзависимую цитотоксичность (CDC); связывание рецептора Fc; зависимую от антител опосредованную клетками цитотоксичность (ADCC); фагоцитоз; негативную регуляцию рецепторов на поверхности клетки (например, В-клеточных рецепторов; BCR) и так далее. Такие эффекторные функции обычно требуют объединения Fc-области со связывающим доменом (например, вариабельным доменом антитела), и их можно оценивать при использовании различных анализов, известных в данной области для оценки таких эффекторных функций антитела.
"Fc-область с нативной последовательностью" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. "Вариантная Fc-область" содержит аминокислотную последовательность, которая отличается от таковой в Fc-области с нативной последовательностью из-за по меньшей мере одной модификации аминокислоты, но сохраняет по меньшей мере одну эффекторную функцию Fc-области с нативной последовательностью. Предпочтительно, вариантная Fc-область имеет по меньшей мере одну аминокислотную замену при сравнении с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например от приблизительно одной до приблизительно десяти аминокислотных замен, и предпочтительно от приблизительно одной до приблизительно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. Вариантная Fc-область здесь должна предпочтительно обладать по меньшей мере приблизительно 80%-ной идентичностью с нативной последовательностью Fc-области и/или с Fc-областью исходного полипептида и, наиболее предпочтительно, по меньшей мере приблизительно 90%-ной идентичностью последовательности с ними, более предпочтительно по меньшей мере приблизительно 95%-ной идентичностью последовательности с ними.
При использовании здесь "зависимая от антител опосредованная клетками цитотоксичность" и "ADCC" относятся к опосредованному клетками взаимодействию, в котором неспецифичные цитотоксические клетки, которые экспрессируют Fc-рецепторы (FcR) (например, натуральные киллерные (NK) клетки, нейтрофилы и макрофаги), распознают антитело, связанное на клетке-мишени, и затем вызывают лизис клетки-мишени. ADCC-активность молекулы, представляющей интерес, можно оценивать при использовании анализа ADCC in vitro, такого как описанный в патенте США №5500362 или 5821337. Полезные эффекторные клетки для таких анализов включают в себя мононуклеарные клетки периферической крови (РВМС) и NK-клетки. Альтернативно или дополнительно, ADCC-активность молекулы, представляющей интерес, можно оценивать in vivo, например, в животной модели, такой как раскрыто в Clynes et al., 1998, PNAS (USA), 95:652-656.
При использовании здесь " Fc-рецептор " и "FcR" описывает рецептор, который связывается с Fc-областью антитела. Предпочтительным FcR является человеческий FcR с нативной последовательностью. Более того, предпочтительным FcR является такой, который связывается с антителом IgG (гамма-рецептор) и включает в себя рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы типа FcγRII включают в себя FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), имеющие одинаковые аминокислотные последовательности, которые различаются, главным образом, их цитоплазматическими доменами. FcR рассмотрены в Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9:457-92; Capel et al., 1994, Immunomethods, 4:25-34; и de Haas et al., 1995, J. Lab. Clin. Med., 126:330-41. "FcR" также включает в себя неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод (Guyer et al., 1976, J. Immunol., 117:587; и Kim et al., 1994, J. Immunol., 24:249).
"Комплементзависимая цитотоксичность" и "CDC" относится к лизированию мишени в присутствии комплемента. Путь активации комплемента инициируется связыванием системы первого компонента комплемента (C1q) с молекулой (например, антителом) в комплексе с распознанным антигеном. Для оценки активации комплемента можно осуществлять анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods, 202:163 (1996).
При использовании здесь термины "Е3", "3Е" и "антитело Е3" использованы взаимозаменяемым образом так, что относятся к антителу, содержащему аминокислотную последовательность тяжелой цепи и вариабельные области легкой цепи, показанные на Фиг.1А (SEQ ID NO:1) и 1Б (SEQ ID NO:2) соответственно. Части антитела Е3, представляющие собой CDR (в том числе CDR по Chothia и по Kabat), схематически показаны на Фиг.1А и 1Б. Фиг.2 и 3 показывают полинуклеотиды, кодирующие тяжелую и легкую цепь соответственно, содержащие вариабельные области легкой цепи и тяжелой цепи, показанные на Фиг.1А и 1Б соответственно. Генерирование и характеризация Е3 описаны в примерах. С Е3 связаны разные биологические функции, в том числе, без ограничения ими, способность связываться с NGF и ингибировать биологическую активность NGF и/или дальнейшие(е) путь(и), опосредованные сигнализацией NGF, и способность ингибировать зависимое от NGF выживание мышиных нейронов тройничного нерва Е13.5. Как обсуждалось здесь, антитела по этому изобретению могут иметь любую одну или более чем одну из этих характеристик. В некоторых воплощениях термин "Е3" относится к иммуноглобулину, кодируемому (а) полинуклеотидом, кодирующим легкую цепь Е3, который имеет номер депонирования АТСС № РТА-4893 или АТСС № РТА-4894, и (б) полинуклеотидом, кодирующим тяжелую цепь Е3, который имеет номер депонирования АТСС № РТА-4895.
При использовании здесь, "иммуноспецифичное" связывание антитела относится к антигенспецифичному взаимодействию, представляющему собой связывание, которое происходит между сайтом антитела, объединяемым с антигеном, и специфическим антигеном, распознаваемым этим антителом (то есть антитело взаимодействует с белком в ELISA или в другом иммуноанализе и не взаимодействует детектируемым образом с неродственными белками).
Эпитоп, который "специфично связывается с" или "предпочтительно связывается с" (использовано здесь взаимозаменяемым образом) с антителом или полипептидом, является термином, хорошо понятным в данной области, и способы определения такого специфичного или предпочтительного связывания также хорошо известны в данной области. О молекуле говорят, что она проявляет "специфичное связывание" или "предпочтительное связывание", если она взаимодействует или ассоциируется более часто, более быстро, с большей продолжительностью и/или с большей аффинностью с конкретной клеткой или веществом, чем с альтернативными клетками или веществами. Антитело "специфично связывается с" или "предпочтительно связывается с" мишенью, если оно связывается с большей аффинностью, авидностью, более легко и/или с большей продолжительностью, чем оно связывается с другими веществами. Например, антитело, которое специфично или предпочтительно связывается с эпитопом NGF, является антителом, которое связывается с этим эпитопом с большей аффинностью, авидностью, более легко и/или с большей продолжительностью, чем оно связывается с другими эпитопами NGF или с эпитопами, не относящимися к NGF. При прочтении этого определения следует также понимать, что, например, антитело (или группировка, или эпитоп), которое специфично или предпочтительно связывается с первой мишенью, может или не может специфично или предпочтительно связываться со второй мишенью. Как таковое, "специфичное связывание" или "предпочтительное связывание" не обязательно требует (хотя оно может включать в себя) исключительное связывание. Обычно, но необязательно, ссылка на связывание означает предпочтительное связывание.
Термины "полипептид", "олигопептид", "пептид" и "белок" использованы здесь взаимозаменяемым образом так, что относятся к полимерам аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может быть прерван не аминокислотами. Эти термины также охватывают полимер аминокислот, который был модифицирован в природе или путем вмешательства, например образованием дисульфидной связи, гликозилированием, липидированием, ацетилированием, фосфорилированием или любой другой манипуляцией или модификацией, такой как конъюгация с метящим компонентом. Также в это определение включены, например, полипептиды, содержащие один или более чем один аналог аминокислот (включая, например, неприродные аминокислоты и так далее), а также другие модификации, известные в данной области. Следует понимать, что поскольку полипептиды по этому изобретению основаны на антителе, эти полипептиды могут иметь как раздельные цепи, так и связанные цепи.
"Полинуклеотид" или "нуклеиновая кислота", используемые здесь взаимозаменяемым образом, относятся к полимерам нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут быть дезоксирибонуклеотидами, рибонуклеотидами, модифицированными нуклеотидами или основаниями и/или их аналогами или любым субстратом, который можно включить в состав полимера ДНК- или РНК-полимеразой. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Модификацию структуры нуклеотида при ее наличии можно придать до или после сборки полимера. Последовательность нуклеотидов может быть прервана ненуклеотидными компонентами. Полинуклеотид можно дополнительно модифицировать после полимеризации, например, конъюгацией с метящим компонентом. Другие типы модификаций включают в себя, например, "кэпы", замену одного или более чем одного встречающегося в природе нуклеотида аналогом; интернуклеотидные модификации, такие как, например, те, которые содержат незаряженные связи (например, метилфосфонаты, трифосфорные эфиры, фосфоамидаты, карбаматы и так далее) и заряженные связи (например, фосфоротиоаты, фосфородитиоаты и так далее); те, которые содержат боковые цепи, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и так далее); те, которые содержат интеркаляторы (например, акридин, псорален и так далее); те, которые содержат хелаторы (например, металлы, радиоактивные металлы, бор, окисляющие металлы и так далее); те, которые содержат алкилирующие агенты; те, которые содержат модифицированные связи (например, альфа-аномерные нуклеиновые кислоты и так далее), а также немодифицированные формы полинуклеотида(ов). Дополнительно, любую гидроксильную группу, обычно присутствующую в сахарах, можно заменить, например, фосфонатными группами, фосфатными группами, защитить стандартными защитными группами, активировать для образования дополнительных связей с дополнительными нуклеотидами или конъюгировать с твердой подложкой. 5'- и 3'-концевую группу ОН можно фосфорилировать или замещать аминами или органическими кэп-группировками, содержащими от 1 до 20 атомов углерода. Другие гидроксилы также можно дериватизировать до стандартных защитных групп. Полинуклеотиды могут также содержать аналоги сахаров рибозы или дезоксирибозы, которые известны в данной области техники, в том числе, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидорибозу, карбоциклические аналоги сахаров, α-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и неосновные нуклеозидные аналоги, такие как метилрибозид. Одну или более чем одну фосфодиэфирную связь можно заменять альтернативными связывающими группами. Эти альтернативные связывающие группы включают в себя, без ограничения ими, такие воплощения, в которых фосфат заменен на P(O)S ("тиоат"), P(S)S ("дитиоат"), "(O)NR2 ("амидат"), P(O)R, P(O)OR', CO или СН2 ("формацеталь"), в которых каждый R или R' независимо представляет собой Н или замещенный или незамещенный алкил (C1-20), возможно, содержащий эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не все связи в полинуклеотиде должны быть идентичными. Данное выше описание применимо ко всем упомянутым здесь полинуклеотидам, в том числе РНК и ДНК.
"Вариабельная область" антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела либо в отдельности, либо в комбинации. Вариабельные области тяжелой и легкой цепи состоят каждая из четырех каркасных областей (FR), соединенных тремя определяющими комплементарность участками (CDR), также известными как гипервариабельные участки. CDR в каждой цепи удерживаются вместе в тесной близости каркасными областями и вместе с CDR из других цепей участвуют в образовании антигенсвязывающего сайта антитела. Существует по меньшей мере две методики для определения CDR: (1) подход, основанный на перекрестной вариабельности последовательностей между видами (см., Kabat et al. Sequences of Proteins of Immunological Interest (5 th ed., 1991, National Institutes of Health, Bethesda MD)); и (2) подход, основанный на кристаллографических исследованиях комплексов антиген-антитело (Chothia et al. (1989) Nature 342:877; Al-lazikani et al. (1997) J. Molec. Вiol. 273:927-948). При использовании здесь CDR может относиться к CDR, который определен при использовании любого из этих подходов или комбинации обоих подходов.
"Константная область" антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела либо в отдельности, либо в комбинации.
При использовании здесь термин "фактор роста нервов" и "NGF" относится к фактору роста нервов и его вариантам, которые сохраняют по меньшей мере часть биологической активности NGF. При использовании здесь NGF включает нативные последовательности NGF всех видов млекопитающих, в том числе человека, собак, кошек, лошадей или коров.
"Рецептор NGF" относится к полипептиду, с которым связывается или который активирует NGF. Рецепторы NGF включают в себя TrkA-рецептор и р75-рецептор любых видов млекопитающих, в том числе, без ограничения ими, человека, собак, кошек, лошадей или коров.
При использовании здесь "антагонистическое антитело против NGF" (взаимозаменяемым образом обозначаемое как "антитело против NGF") относится к антителу, которое способно связываться с NGF и ингибировать биологическую активность NGF и/или путь(и) в нисходящем направлении, опосредованный(е) сигнализацией NGF. Антагонистическое антитело против NGF охватывает антитела, которые блокируют, антагонизируют, подавляют или снижают (в том числе значительно) биологическую активность NGF, включая пути в нисходящем напрвлении, опосредованные сигнализацией NGF, такую как связывание рецептора и/или индуцирование клеточного ответа на NGF. Для целей настоящего изобретения следует ясно понимать, что термин "антагонистическое антитело против NGF" охватывает все ранее идентифицированные термины, названия и функциональные состояния и характеристики, которыми сам NGF, биологическая активность NGF (включая, без ограничения, его способность опосредовать любой аспект послеоперационной боли) или следствия этой биологической активности по существу аннулируются, снижаются или нейтрализуются в любой значимой степени. В некоторых воплощениях антагонистическое антитело против NGF связывается с NGF и предотвращает димеризацию NGF и/или связывание с рецептором NGF (таким как р75 и/или TrkA). В других воплощениях антитело против NGF связывается с NGF и предотвращает димеризацию TrkA-рецептора и/или аутофосфорилирование TrkA. Примеры антагонистических антител против NGF даны здесь.
"Биологическая активность" NGF обычно относится к способности связываться с рецепторами NGF и/или активировать сигнальные пути рецептора NGF. Биологическая активность включает в себя, без ограничения, любое одно или более чем одно из следующего: способность связываться с рецептором NGF (таким как р75 и/или TrkA); способность стимулировать димеризацию и/или аутофосфорилирование ТrkА-рецептора; способность активировать сигнальный путь рецептора NGF; способность стимулировать дифференцировку, пролиферацию, выживание, рост клеток и другие изменения в физиологии клетки, в том числе (в случае нейронов, включая периферические и центральные нейроны) изменение в нейрональной морфологии, синаптогенезе, синаптической функции, высвобождении нейротрансмиттера и/или нейропептида и регенерации после повреждения; способность стимулировать выживание мышиных нейронов тройничного нерва Е13.5 и способность опосредовать боль, включая послеоперационную боль.
При использовании здесь "по существу чистый" относится к веществу, которое является по меньшей мере на 50% чистым (то есть свободным от примесей), более предпочтительно по меньшей мере на 90% чистым, более предпочтительно по меньшей мере на 95% чистым, более предпочтительно по меньшей мере на 98% чистым, более предпочтительно по меньшей мере на 99% чистым.
"Клетка-хозяин" включает индивидуальную клетку или клеточную культуру, которая может быть или была реципиентом вектора(ов) для включения полинуклеотидных вставок. Клетки-хозяева включают в себя потомство единственной клетки-хозяина, и это потомство не обязательно может быть полностью идентичным (по морфологии или составу геномной ДНК) исходной родительской клетке ввиду естественных, случайных или намеренных мутаций. Клетка-хозяин включает клетки, трансфицированные in vivo полинуклеотидом(ами) по этому изобретению.
При использовании здесь "лечение" представляет собой подход к получению полезных или желаемых клинических результатов. Для целей этого изобретения полезные или желаемые клинические результаты включают в себя, без ограничения ими, одно или более чем одно из следующего: улучшение или облегчение любого вида боли, включая острую, хроническую, воспалительную, нейропатическую, послеоперационную боль, боль, вызванную ревматоидным артритом, или боль, вызванную остеоартритом. Для целей этого изобретения полезные или желаемые клинические результаты включают в себя, без ограничения ими, одно или более чем одно из следующего: уменьшение тяжести, облегчение одного или более чем одного симптома, связанного с болью, включающей любой вид боли (такое как уменьшение продолжительности боли, снижение чувствительности к боли или восприятия боли).
"Эффективное количество" лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для получения полезных или желаемых результатов, в том числе клинических результатов, таких как облегчение или снижение ощущения боли. Эффективное количество можно вводить одним или более чем одним введением. Для целей этого изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для лечения, облегчения, снижения интенсивности и/или предотвращения боли, включая послеоперационную боль, боль, вызванную ревматоидным артритом, и/или боль, вызванную остеоартритом. В некоторых воплощениях "эффективное количество" может снизить боль в покое (боль в состоянии покоя), или механически индуцированную боль (включая боль вследствие движения), или обе, и его можно вводить до, во время или после надреза, пореза, ссадины или повреждения и/или до, во время или после болевого стимула. Как это понимают в клиническом контексте, эффективное количество лекарственного средства, соединения или фармацевтической композиции может или не может быть достигнуто в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, "эффективное количество" можно рассматривать в контексте введения одного или более чем одного терапевтического агента, и можно считать, что единственный агент дан в эффективном количестве, если в сочетании с одним или более чем одним другим агентом можно достичь или достигнут желательный результат.
"Снижение частоты" боли означает любое из следующего: снижение тяжести (что может включать в себя снижение необходимости в количестве и/или количества (например, действия) других лекарственных средств и/или терапий, обычно используемых для этих состояний, в том числе, например, опиатов), длительности и/или частоты (включая, например, замедление или увеличение времени до послеоперационной боли у индивида). Как понятно специалистам, индивиды могут отличаться в смысле их ответа на лечение, и, например, "способ снижения частоты боли, вызванной ревматоидным артритом, или боли, вызванной остеоартритом, у индивида" как таковой соответствует введению антагонистического антитела против NGF на основании разумного ожидания, что таким введением вероятно можно вызвать такое снижение частоты у этого конкретного индивида.
"Облегчение" боли или одного или более чем одного симптома боли (такой как боль, вызванная ревматоидным артритом, или боль, вызванная остеоартритом) означает уменьшение или улучшение одного или более чем одного симптома боли по сравнению с отсутствием введения антагонистического антитела против NGF. "Облегчение" также включает в себя укорочение или снижение продолжительности проявления симптома.
"Облегчение проявлений" боли или одного или более чем одного симптома боли (такой как боль, вызванная ревматоидным артритом, или боль, вызванная остеоартритом) означает уменьшение степени одного или более чем одного нежелательного клинического проявления послеоперационной боли у индивида или популяции индивидов, которых лечили антагонистическим антителом против NGF в соответствии с этим изобретением.
При использовании здесь "замедление" развития боли означает отсрочку, торможение, сдерживание, запаздывание, стабилизацию и/или замедление прогрессирования боли, такой как послеоперационная боль, боль, вызванная ревматоидным артритом, или боль, вызванная остеоартритом. Это замедление может варьировать по длительности времени в зависимости от истории заболевания и/или индивидов, которых лечат. Как это очевидно специалисту, достаточное или значительное замедление может, в конечном счете, охватывать предотвращение в том смысле, что у индивида не развивается боль. Способ, который "замедляет" развитие симптома, представляет собой способ, который снижает вероятность развития симптома в данных временных рамках и/или снижает степень симптомов в данных временных рамках по сравнению с тем, когда данный способ не используют. Такие сравнения в типичных случаях основаны на клинических исследованиях при использовании значимого числа субъектов.
"Боль" при использовании здесь относится к боли любой этиологии, включая острую и хроническую боль и любую боль с воспалительным компонентом. Примеры боли включают в себя послеоперационную боль, боль после проведения процедур (включая зубную боль), мигрень, головную боль и невралгию тройничного нерва, боль, связанную с ожогом, раной или камнем почки, боль, связанную с травмой (включая травматическое повреждение головы), нейропатическую боль, боль, связанную с расстройствами опорно-двигательной системы, такими как ревматоидный артрит, остеоартрит, анкилозирующий спондилоартрит, серонегативные (неревматоидные) артропатии, несуставной ревматизм и околосуставные расстройства, боль, связанную с раком (включая "боль прорыва" и боль, связанную с терминальной стадией рака), периферическую нейропатию и постгерпетическую невралгию. Примеры боли с воспалительным компонентом (в дополнение к некоторым, описанным выше) включают в себя ревматическую боль, боль, связанную с воспалением слизистой оболочки, и дисменорею.
"Послеоперационная боль" (взаимозаменяемым образом обозначаемая как "боль после надреза" или "посттравматическая боль") относится к боли, возникающей или происходящей из-за внешней травмы, такой как порез, пункция, надрез, ссадина или рана в ткани индивида (включая такую, которая возникает вследствие любых хирургических процедур, как инвазивных, так и неинвазивных). При использовании здесь послеоперационная боль не включает в себя боль, которая имеет место (возникает или существует) без внешней физической травмы. В некоторых воплощениях послеоперационная боль представляет собой внутреннюю или внешнюю (включая периферическую) боль, и рана, порез, травма, ссадина или надрез могут происходить случайно (как при травматической ране) или намеренно (как при хирургическом надрезе). При использовании здесь "боль" включает в себя ноцицепцию и ощущение боли, и боль можно оценивать объективно и субъективно при использовании шкал боли и других способов, хорошо известных в данной области. Послеоперационная боль при использовании здесь включает в себя аллодинию (то есть повышенный ответ на стимул, который обычно не является вредным) и гипералгезию (то есть повышенный ответ на стимул, который обычно является вредным или неприятным), которая в свою очередь может быть температурной или механической (тактильной) по природе. В некоторых воплощениях боль характеризуется температурной чувствительностью, механической чувствительностью и/или болью в покое. В некоторых воплощениях послеоперационная боль включает в себя механически индуцированную боль или боль в покое. В других воплощениях послеоперационная боль включает в себя боль в покое. Как хорошо известно в данной области, боль может быть первичной или вторичной болью.
"Биологический образец" охватывает множество разных типов образцов, которые получены из индивида и которые можно использовать в анализе для диагноза или мониторинга. Это определение охватывает кровь и другие жидкие образцы биологического происхождения, образцы твердой ткани, такие как биоптаты или культуры ткани либо клетки, происходящие из них, и их потомство. Это определение также включает в себя образцы, с которыми после их получения манипулировали любым путем, например обрабатывали реагентами, солюбилизировали или обогащали определенными компонентами, такими как белки или полинуклеотиды, или погружали в полутвердый или твердый матрикс для получения срезов. Термин "биологический образец" охватывает клинический образец, а также включает в себя клетки в культуре, клеточные супернатанты, клеточные лизаты, сыворотку, плазму, биологическую жидкость и образцы ткани.
"Индивид" представляет собой позвоночное, предпочтительно млекопитающее, более предпочтительно человека. Млекопитающие включают в себя, без ограничения ими, сельскохозяйственных животных (таких как коровы), промысловых животных, домашних животных (таких как кошки, собаки и лошади), приматов, мышей и крыс.
При использовании здесь "вектор" означает конструкцию, которая способна доставлять и предпочтительно экспрессировать один или более чем один ген или последовательность, представляющие интерес, в клетке-хозяине. Примеры векторов включают в себя, без ограничения ими, вирусные векторы, экспрессионные векторы, представляющие собой голую ДНК или РНК, плазмидные, космидные или фаговые векторы, экспрессионные векторы на основе ДНК или РНК, связанные с катионными конденсирующими агентами, экспрессионные векторы на основе ДНК или РНК, инкапсулированные в липосомы, и некоторые эукариотические клетки, такие как клетки-продуценты.
При использовании здесь "контролирующая экспрессию последовательность" означает последовательность нуклеиновой кислоты, которая направляет транскрипцию нуклеиновой кислоты. Контролирующей экспрессию последовательностью может быть промотор, такой как конститутивный или индуцибельный промотор, или энхансер. Контролирующая экспрессию последовательность функциональным образом связана с последовательностью нуклеиновой кислоты, подлежащей транскрипции.
При использовании здесь "фармацевтически приемлемый носитель" включает в себя любое вещество, которое при объединении с активным ингредиентом позволяет этому ингредиенту сохранять биологическую активность и не взаимодействует с иммунной системой субъекта. Примеры включают в себя, без ограничения ими, любые стандартные фармацевтические носители, такие как забуференный фосфатами физиологический раствор, вода, эмульсии, такие как эмульсии масло/вода, и различные типы смачивающих агентов. Предпочтительными разбавителями для аэрозольного или парентерального введения являются забуференный фосфатами физиологический раствор или нормальный (0,9%) физиологический раствор. Композиции, содержащие такие носители, приготавливают хорошо известными общепринятыми способами (см., например, Remington's Pharmaceutical Sciences, 18th edition, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmaceutics, 20th Ed., Mack Publishing, 2000).
Термин "Koff" при использовании здесь относится к константе скорости диссоциации антитела из комплекса антитело/антиген.
Термин "Kd" при использовании здесь относится к константе диссоциации для взаимодействия антитела с антигеном.
АНТИТЕЛО Е3, Е3-ПРОИЗВОДНЫЕ АНТИТЕЛА, КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ
Композиции на основе Е3, композиции на основе производных Е3 и способы получения композиций
Это изобретение охватывает композиции, в том числе фармацевтические композиции, содержащие антитело или полипептид Е3 и полинуклеотиды, содержащие последовательности, кодирующие антитело или полипептид Е3. При использовании здесь композиции содержат одно (один) или более чем одно (один) антитело или полипептид (который может быть или не быть антителом), которое(ый) связывается с NGF, и/или один или более чем один полинуклеотид, содержащий последовательности, кодирующие одно (один) или более чем одно (один) антитело или полипептид, которое(ый) связывается с NGF. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, в том числе буферы, которые хорошо известны в данной области.
Это изобретение также охватывает воплощения, представляющие собой выделенное(ый) антитело, полипептид и полинуклеотид. Это изобретение также охватывает воплощения, представляющие собой по существу чистое(ый) антитело, полипептид и полинуклеотид.
Антитела и полипептиды по этому изобретению характеризуются любой (одной или более чем одной) следующей характеристикой: (а) способность связываться с NGF; (б) способность снижать и/или ингибировать биологическую активность NGF и/или путь(и) в нисходящем направлении, опосредованный(е) сигнализацией NGF; (в) способность снижать и/или ингибировать зависимое от NGF выживание мышиных нейронов тройничного нерва Е13.5; (г) отсутствие любого значительного перекрестного взаимодействия с NT3, NT4/5 и/или BDNF; (д) способность лечить и/или предотвращать боль (включая послеоперационную боль); (е) способность увеличивать клиренс NGF; (г) способность снижать или ингибировать активацию TrkA-рецептора, как определяют, например, при использовании анализа активации киназы рецептора (KIRA) (см. патент США №6027927).
Связывающие свойства антитела Е3, которое связывается с человеческим NGF с высокой аффинностью и медленной кинетикой диссоциации при сравнении с исходным мышиным моноклональным антителом 911 против NGF, суммированы ниже. Е3 связывается с человеческим NGF с приблизительно в 50 раз более высокой аффинностью связывания, чем исходное мышиное антитело 911.
Антитело Е3 и родственные антитела также проявляют сильную способность антагонизировать человеческий NGF, как оценивают анализами in vitro (см. Примеры 2 и 3). Например, антитело Е3 антагонизирует зависимое от NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 около 21 пМ в присутствии 15 пМ человеческого NGF и около 1,2 пМ в присутствии 1,5 пМ человеческого NGF.
Соответственно в другом аспекте дополнительно идентифицированы антитела и полипептиды по этому изобретению, которые характеризуются: (ж) высокой аффинностью связывания с человеческим NGF при низкой кинетике диссоциации (в некоторых воплощениях при KD менее чем приблизительно 2 нМ и/или koff менее чем приблизительно 6×10-5 сек-1) и/или (з) способностью ингибировать (блокировать) зависимое от NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 приблизительно 100 пМ или менее чем приблизительно 15 пМ NGF (в некоторых воплощениях человеческого NGF) и/или IC50 около 20 пМ или менее чем приблизительно 1,5 пМ NGF.
В некоторых воплощениях антитело связывается с человеческим NGF и не связывается значительно с NGF из других видов позвоночных (в некоторых воплощениях млекопитающих). В некоторых воплощениях антитело связывается с человеческим NGF, а также с одним или более чем одним NGF из других видов позвоночных (в некоторых воплощениях млекопитающих). В еще других воплощениях антитело связывается с NGF и не способно к значительному перекрестному взаимодействию с другими нейротрофинами (такими как родственные нейротрофины, NT3, NT4/5 и/или BDNF). В некоторых воплощениях антитело связывается с NGF, а также по меньшей мере с одним другим нейротрофином. В некоторых воплощениях антитело связывается с NGF таких видов млекопитающих, как лошадь или собака, но не связывается значительно с NGF из других видов млекопитающих.
В некоторых воплощениях это изобретение представляет собой антитело, содержащее легкую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4893 или АТСС № РТА-4894. В другом аспекте это изобретение представляет собой антитело, содержащее тяжелую цепь, которую кодирует полинуклеотид, продуцируемый клеткой-хозяином, с номером депонирования АТСС № РТА-4895. Настоящее изобретение также охватывает различные препараты Е3 и эквивалентных фрагментов антитела (например, Fab, Fab', F(аb')2, Fv, Fc и так далее), его единственной цепи (ScFv), его мутантов, слитых белков, содержащих часть антитела, и любую другую модифицированную конфигурацию Е3, которая содержит сайт распознавания антигена (NGF) при требуемой специфичности. Эквиваленты антитела Е3, в том числе полипептидные фрагменты антитела Е3 (которые могут быть или не быть антителом) и полипептиды, содержащие полипептидные фрагменты Е3, идентифицированы и охарактеризованы по любому (одному или более чем одному) критерию, описанному выше.
Соответственно в этом изобретении предложено или любое из следующего, или композиции (в том числе фармацевтические композиции), содержащие любое из следующего: (а) антитело Е3; (б) фрагмент или область антитела Е3; (в) легкая цепь антитела Е3, как показано на Фиг.1Б; (г) тяжелая цепь антитела Е3, как показано на Фиг.1А; (д) одна или более чем одна вариабельная область легкой цепи и/или тяжелой цепи антитела Е3; (е) один или более чем один CDR (один, два, три, четыре, пять или шесть CDR) антитела Е3, показанного на Фиг.1А и 1Б; (ж) CDR-H3 тяжелой цепи антитела Е3, показанного на Фиг.1А; (з) CDR-L3 легкой цепи антитела Е3, показанного на Фиг.1Б; (и) три CDR легкой цепи антитела Е3, показанного на Фиг.1Б; (к) три CDR тяжелой цепи антитела Е3, показанного на Фиг.1А; (л) три CDR легкой цепи и три CDR тяжелой цепи антитела Е3, показанного на Фиг.1А и 1Б; и (м) антитело, содержащее любое от (б) до (к) включительно. Как это следует из данного здесь описания, конкретно исключены из этого изобретения воплощения, представляющие собой полипептид, состоящий из аминокислотной последовательности, идентичной аминокислотной последовательности мышиного моноклонального антитела 911. Последовательности расширенных CDR моноклонального антитела 911 показаны на Фиг.1А и 1Б и в SEQ ID NOS:9-14.
Части антитела Е3, представляющие собой CDR (включая CDR по Chothia и по Kabat), схематически показаны на Фиг.1А и 1Б и состоят из следующих аминокислотных последовательностей: (a) CDR-1 тяжелой цепи ("CDR-H1") GFSLIGYDLN (SEQ ID NO:3), (б) CDR-2 тяжелой цепи ("CDR-H2") IIWGDGTTDYNSAVKS (SEQ ID NO:4), (в) CDR-3 тяжелой цепи ("CDR-H3") GGYWYATSYIFDY (SEQ ID NO:5), (г) CDR-1 легкой цепи ("CDR-L1") RASQSISNNLN (SEQ ID NO:6), (д) CDR-2 легкой цепи ("CDR-L2") YTSRFHS (SEQ ID NO:7) и (е) CDR-3 легкой цепи ("CDR-L3") QQEHTLPYT (SEQ ID NO:8). Определение CDR-областей находится в пределах знаний специалистов в данной области. Понятно, что в некоторых воплощениях CDR может быть комбинацией CDR по Kabat и по Chothia (что также обозначено как "объединенный CDR" или "расширенный CDR"). В некоторых воплощениях CDR содержат CDR по Kabat. В других воплощениях CDR представляют собой CDR по Chothia.
В некоторых воплощениях в этом изобретении предложено антитело, которое содержит по меньшей мере один CDR, который является по существу гомологичным по меньшей мере одному CDR, по меньшей мере двум, по меньшей мере трем, по меньшей мере четырем, по меньшей мере пяти CDR Е3 (или в некоторых воплощениях по существу гомологичным всем шести CDR Е3 или производных от Е3). Другие воплощения включают в себя антитела, которые имеют по меньшей мере два, три, четыре, пять или шесть CDR, которые являются по существу гомологичными по меньшей мере двум, трем, четырем, пяти или шести CDR Е3 или производных от Е3. Понятно, что для целей этого изобретения специфичность связывания и/или общая активность (которая может иметь смысл лечения и/или предотвращения боли или ингибирования зависимого от NGF выживания мышиных нейронов тройничного нерва Е13.5) обычно сохранена, хотя степень активности может варьировать по сравнению с Е3 (может быть большей или меньшей).
В этом изобретении также предложен полипептид (который может быть или не быть антителом), содержащий аминокислотную последовательность Е3 (показанную на Фиг.1А и 1Б), который имеет любое из следующего: по меньшей мере 5 последовательных аминокислот, по меньшей мере 8 последовательных аминокислот, по меньшей мере приблизительно 10 последовательных аминокислот, по меньшей мере приблизительно 15 последовательных аминокислот, по меньшей мере приблизительно 20 последовательных аминокислот, по меньшей мере приблизительно 25 последовательных аминокислот, по меньшей мере приблизительно 30 последовательных аминокислот последовательности Е3, где по меньшей мере 3 аминокислоты входят в вариабельную область Е3, при этом следует понимать, что воплощения, которые состоят из аминокислотной последовательности, которая идентична аминокислотной последовательности мышиного моноклонального антитела 911, конкретно исключены. Последовательности расширенных CDR моноклонального антитела 911 показаны на Фиг.1А и 1Б и в SEQ ID NOS:9-14. В одном из воплощений вариабельная область происходит из легкой цепи Е3. В еще одном воплощении вариабельная область происходит из тяжелой цепи Е3. В еще одном воплощении 5 (или более) последовательных аминокислот происходят из гипервариабельного участка (CDR) Е3, показанного на Фиг.1А и 1Б.
В еще одном воплощении в этом изобретении предложен полипептид, который содержит аминокислотную последовательность Е3, которая имеет любое из следующего: по меньшей мере 5 последовательных аминокислот, по меньшей мере 8 последовательных аминокислот, по меньшей мере приблизительно 10 последовательных аминокислот, по меньшей мере приблизительно 15 последовательных аминокислот, по меньшей мере приблизительно 20 последовательных аминокислот, по меньшей мере приблизительно 25 последовательных аминокислот, по меньшей мере приблизительно 30 последовательных аминокислот последовательности Е3, где последовательность Е3 содержит любое одно или более чем одно из следующего: аминокислотный остаток L29 из CDR-H1, I50 из CDR-H2, W101 из CDR-H3 и/или А103 из CDR-H3 и/или аминокислотный остаток S28 из CDR-L1, N32 из CDR-L1, Т51 из CDR-L2, 91Е из CDR-L3 и/или Н92 из CDR-L3, при этом следует понимать, что воплощения, которые состоят из аминокислотной последовательности, которая идентична аминокислотной последовательности мышиного моноклонального антитела 911, конкретно исключены.
Как это очевидно, во всем описании использована схема последовательной нумерации аминокислот, которая относится к аминокислотным остаткам в вариабельных областях (то есть аминокислотные остатки в каждой вариабельной области пронумерованы последовательно). Как хорошо известно в данной области, системы нумерации по Kabat и/или по Chothia полезны при сравнении двух антител или полипептидов, например антитела Е3 и варианта Е3 (или полипептида, предполагаемого в качестве варианта Е3). В данной области вполне понятно, как переводить последовательную нумерацию в нумерацию по Chothia и/или по Kabat, если это желательно, например, для применения при проведении сравнений между Е3 и другим полипептидом. Фиг.23 показывает вариабельные области Е3, пронумерованные при использовании последовательной нумерации, а также по Chothia и по Kabat. В дополнение, обычно понятно, что для облегчения сравнения остатки каркасной области, как правило, но не всегда, имеют приблизительно одно и то же число остатков. Однако CDR могут варьировать в размере (то есть возможно, что они имеют вставки и/или делеции одного или более чем одного аминокислотного остатка). При сравнении антитела Е3 и варианта-кандидата Е3 (например, в случае области CDR из последовательности-кандидата, которая является более длинной по последовательности в антителе Е3, с которым она выровнена) можно осуществлять следующие стадии (хотя в данной области известны другие способы). Последовательность антитела-кандидата выравнивают с вариабельными областями тяжелой цепи и легкой цепи антитела Е3. Выравнивание можно осуществлять вручную или на компьютере с использованием общепринятых компьютерных программ. Выравнивание можно облегчить при использовании некоторых аминокислотных остатков, которые являются общими для большинства последовательностей Fab. Например, легкая и тяжелая цепь в типичных случаях имеют каждая по два цистеина, которые часто находятся в консервативных положениях. Понятно, что аминокислотная последовательность варианта-кандидата антитела может быть длиннее (то есть иметь вставленные аминокислотные остатки) или короче (иметь делегированные аминокислотные остатки). К номеру остатка можно добавлять суффиксы для указания вставки дополнительных остатков, например остаток 34 abc. Для последовательностей-кандидатов, которые, например, выравнивают с последовательностью Е3 по, например, остаткам 33 и 35, но не имеют в них общего остатка для выравнивания по остатку 35, остаток 35 просто не относят к остатку. В другом подходе обычно хорошо известно, что при сравнении CDR, имеющих разную длину, сравнение можно делать между структурно эквивалентными аминокислотами (например, одно и то же положение в комплексе антиген-антитело). Например, нумерация по Chothia (см. выше Al-Lazikani et al.) обычно (но не во всех случаях) помещает вставки и делеции в структурно правильных положениях. Структурную эквивалентность можно также вывести или продемонстрировать при использовании рентгеновской кристаллографии или анализа циклов двойных мутантов (см. Pons et al. (1999) Prot. Sci. 8:958-968).
Аффинность связывания антитела против NGF с NGF (таким как человеческий NGF) может быть от приблизительно 0,10 до приблизительно 0,80 нМ, от приблизительно 0,15 до приблизительно 0,75 нМ и от приблизительно 0,18 до приблизительно 0,72 нМ. В некоторых воплощениях аффинность связывания равна приблизительно 2 пМ, приблизительно 5 пМ, приблизительно 10 пМ, приблизительно 15 пМ, приблизительно 20 пМ, приблизительно 40 пМ или более чем приблизительно 40 пМ. В одном из воплощений аффинность связывания находится между приблизительно 2 и 22 пМ. В других воплощениях аффинность связывания менее чем приблизительно 10 нМ, приблизительно 5 нМ, приблизительно 4 нМ, приблизительно 3,5 нМ, приблизительно 3 нМ, приблизительно 2,5 нМ, приблизительно 2 нМ, приблизительно 1,5 нМ, приблизительно 1 нМ, приблизительно 900 пМ, приблизительно 800 пМ, приблизительно 700 пМ, приблизительно 600 пМ, приблизительно 500 пМ, приблизительно 400 пМ, приблизительно 300 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 90 пМ, приблизительно 80 пМ, приблизительно 70 пМ, приблизительно 60 пМ, приблизительно 50 пМ, приблизительно 40 пМ, приблизительно 30 пМ, приблизительно 10 пМ. В некоторых воплощениях аффинность связывания равна приблизительно 10 нМ. В других воплощениях аффинность связывания менее чем приблизительно 10 нМ. В других воплощениях аффинность связывания представляет собой около 0,1 нМ или около 0,07 нМ. В других воплощениях аффинность связывания менее чем приблизительно 0,1 нМ или менее чем приблизительно 0,07 нМ. В других воплощениях аффинность связывания представляет собой любое значение от приблизительно 10 нМ, приблизительно 5 нМ, приблизительно 4 нМ, приблизительно 3,5 нМ, приблизительно 3 нМ, приблизительно 2,5 нМ, приблизительно 2 нМ, приблизительно 1,5 нМ, приблизительно 1 нМ, приблизительно 900 пМ, приблизительно 800 пМ, приблизительно 700 пМ, приблизительно 600 пМ, приблизительно 500 пМ, приблизительно 400 пМ, приблизительно 300 пМ, приблизительно 200 пМ, приблизительно 150 пМ, приблизительно 100 пМ, приблизительно 90 пМ, приблизительно 80 пМ, приблизительно 70 пМ, приблизительно 60 пМ, приблизительно 50 пМ, приблизительно 40 пМ, приблизительно 30 пМ, приблизительно 10 пМ до любой величины около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ или около 40 пМ. В некоторых воплощениях аффинность связывания представляет собой любое значение около 10 нМ, около 5 нМ, около 4 нМ, около 3,5 нМ, около 3 нМ, около 2,5 нМ, около 2 нМ, около 1,5 нМ, около 1 нМ, около 900 пМ, около 800 пМ, около 700 пМ, около 600 пМ, около 500 пМ, около 400 пМ, около 300 пМ, около 200 пМ, около 150 пМ, около 100 пМ, около 90 пМ, около 80 пМ, около 70 пМ, около 60 пМ, около 50 пМ, около 40 пМ, около 30 пМ, около 10 пМ. В еще других воплощениях аффинность связывания равна приблизительно 2 пМ, приблизительно 5 пМ, приблизительно 10 пМ, приблизительно 15 пМ, приблизительно 20 пМ, приблизительно 40 пМ или более чем приблизительно 40 пМ.
Аффинность связывания антитела с NGF можно определять при использовании способов, хорошо известных в данной области. Один способ определения аффинности связывания антитела с NGF представляет собой измерение аффинности монофункциональных Fab-фрагментов антитела, как описано в примерах. Для получения монофункциональных Fab-фрагментов можно расщеплять антитело (например, IgG) папаином или экспрессировать рекомбинантным способом. Аффинность Fab-фрагмента антитела против NGF можно определять поверхностным плазменным резонансом (система поверхностного плазменного резонанса (ППР) BIAcore3000™, BIAcore, INC, Piscataway NJ), как описано в примерах. Этот способ является подходящим для применения в определении аффинности связывания антител с NGF любого вида, включая NGF человека, NGF других позвоночных (в некоторых воплощениях млекопитающего) (таким как мышиный NGF, крысиный NGF, NGF примата), а также для применения с другими нейротрофинами, такими как родственные нейротрофины NT3, NT4/5 и/или BDNF.
В некоторых воплощениях антитела или пептиды по этому изобретению могут ингибировать (снижать и/или блокировать) зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 15 пМ NGF) при любом значении около 200 пМ, 150 пМ, 100 пМ, 80 пМ, 60 пМ, 40 пМ, 20 пМ, 10 пМ или менее. В некоторых воплощениях антитела или пептиды по этому изобретению могут ингибировать (снижать и/или блокировать) зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 1,5 пМ NGF) при любом значении около 50 пМ, 40 пМ, 30 пМ, 10 пМ, 20 пМ, 10 пМ, 5 пМ, 2 пМ, 1 пМ или менее. В некоторых воплощениях антитела или пептиды по этому изобретению могут ингибировать (снижать и/или блокировать) зависимое от крысиного NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 15 пМ NGF) при любом значении около 150 пМ, 125 пМ, 100 пМ, 80 пМ, 60 пМ, 40 пМ, 30 пМ, 20 пМ, 10 пМ, 5 пМ или менее. В некоторых воплощениях антитела или пептиды по этому изобретению могут ингибировать (снижать и/или блокировать) зависимое от крысиного NGF выживание мышиных нейронов тройничного нерва Е13.5 при IC50 (в присутствии приблизительно 1,5 пМ NGF) при любом значении около 30 пМ, 25 пМ, 20 пМ, 15 пМ, 10 пМ, 5 пМ, 4 пМ, 3 пМ, 2 пМ, 1 пМ или менее. Способы измерения зависимого от NGF выживания мышиных нейронов тройничного нерва Е13 известны в данной области и описаны, например, в Примере 2.
В этом изобретении также предложены способы получения любого из этих антител или полипептидов. Антитела по этому изобретению можно получать способами, известными в данной области, некоторые из которых проиллюстрированы в примерах. Полипептиды можно получать протеолитическим или другим расщеплением антител, рекомбинантными способами (а именно, одиночные или слитые полипептиды), как описано выше, или химическим синтезом. Полипептиды антител, особенно полипептиды, укороченные до приблизительно 50 аминокислот, удобно получать химическим синтезом. Способы химического синтеза известны в данной области и являются коммерчески доступными. Например, антитело Е3 можно получать на автоматическом синтезаторе полипептидов при использовании твердофазного способа. См. также патенты США №№5807715, 4816567 и 6331415. Химерные или гибридные антитела можно также получать in vitro при использовании известных способов химии синтеза белков, включая способы, в которых используют поперечно-сшивающие агенты. Например, иммунотоксины можно конструировать использованием взаимодействия, представляющего собой дисульфидный обмен, или образованием тиоэфирной связи. Примеры подходящих реагентов для этой цели включают в себя иминотиолат и метил-4-меркаптобутиримидат.
В другой альтернативе антитела можно получать рекомбинантным путем при использовании способов, хорошо известных в данной области. В одном из воплощений полинуклеотид, содержащий последовательность, которая кодирует вариабельные области антитела Е3 и представляющие собой легкую цепь области антитела Е3 (показанного на Фиг.1А и 1Б), клонируют в вектор для экспрессии или размножения в клетке-хозяине (например, в клетках СНО). В еще одном воплощении полинуклеотидные последовательности, показанные на Фиг.2 и 3, клонируют в один или более чем один вектор для экспрессии или размножения. Последовательность, кодирующая антитело, которое представляет интерес, может сохраняться в векторе в клетке-хозяине, и клетку-хозяина можно затем размножить и заморозить для будущего применения. Векторы (в том числе экспрессионные векторы) и клетки-хозяева описаны здесь далее. Раскрыты способы рекомбинантной экспрессии антител в растениях или в молоке. См., например, Peeters et al. (2001) Vaccine 19:2756; Lonberg, N. and D. Huszar. (1995) Int.Rev.Immunol 13:65; и Pollock et al. (1999) J Immunol Methods 231:147. Способы получения производных антител, например гуманизированных, одноцепочечных и так далее, известны в данной области.
Это изобретение также охватывает одноцепочечные фрагменты вариабельной области ("scFv") антител по этому изобретению, таких как Е3. Одноцепочечные фрагменты вариабельной области получают связыванием вариабельных областей легкой и/или тяжелой цепи при использовании короткого связывающего пептида. Bird et al. (1988) Science 242:423-426. Примером связывающего пептида является (GGGGS)3 (SEQ ID NO:15), который образует мостик приблизительно 3,5 нМ между карбоксильным концом одной вариабельной области и аминным концом другой вариабельной области. Созданы и используются линкеры, имеющие другие последовательности (Bird et al. (1988)). Линкеры могут в свою очередь быть модифицированными для дополнительных функций, таких как присоединение лекарственного средства или присоединение к твердой подложке. Одноцепочечные варианты можно получать либо рекомбинантными, либо синтетическими способами. Для синтетического получения scFv можно использовать автоматический синтезатор. Для рекомбинантного получения scFv подходящую плазмиду, содержащую полинуклеотид, который кодирует scFv, можно ввести в подходящую клетку-хозяина, либо эукариотическую, такую как дрожжевая, растительная, клетки насекомого или млекопитающего, либо прокариотическую, такую как Е.coli. Полинуклеотиды, кодирующие scFv, представляющие интерес, можно получать обычными манипуляциями, такими как лигирование полинуклеотидов. Получившийся scFv можно выделить с использованием стандартных методик очистки белка, известных в данной области.
Также охватываются другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой двухвалентные биспецифичные антитела, в которых VH- и VL-домены экспрессированы на единственной полипептидной цепи, но при использовании линкера, который является слишком коротким, чтобы обеспечивать спаривание между двумя доменами на одной и той же цепи, из-за чего домены вынуждены спариваться с комплементарными доменами другой цепи и создавать два сайта связывания антигена (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad Sci. USA 90:6444-6448; Poljak, R. J., et al. (1994) Structure 2:1121-1123).
Антитело может быть биспецифичным антителом, моноклональным антителом, которое имеет специфичности связывания в отношении по меньшей мере двух разных антигенов. Биспецифичное антитело можно получать при использовании раскрытых здесь антител. Способы получения биспецифичных антител известны в данной области (см., например, Suresh et al., 1986, Methods in Enzymology 121:210). Традиционно рекомбинантное получение биспецифичных антител основано на коэкспрессии двух пар тяжелой цепи и легкой цепи иммуноглобулина с двумя тяжелыми цепями, имеющими разные специфичности (Millstein and Cuello, 1983, Nature 305, 537-539).
В соответствии с одним из подходов к получению биспецифичных антител вариабельные домены антител с желаемой специфичностью связывания (сайты соединения антитела с антигеном) подвергают слиянию с последовательностями константного домена иммуноглобулина. Слияние предпочтительно происходит с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнирной области, СН2-области и СН3-области. Предпочтительным является то, что по меньшей мере в одном из слияний присутствует первая тяжелая цепь, имеющая константную область (СН1), содержащую сайт, необходимый для связывания легкой цепи. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и, если желательно, легкой цепи иммуноглобулина, встраивают в отдельные экспрессионные векторы и котрансфицируют ими подходящий организм-хозяин. Это обеспечивает более высокую гибкость в подгонке общих частей трех полипептидных фрагментов в воплощениях, когда оптимальные выходы обеспечивают неравные соотношения трех полипептидных цепей, использованных в конструировании. Однако кодирующие последовательности для двух или всех трех полипептидных цепей можно встраивать в один экспрессионный вектор, когда экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к более высоким выходам или когда эти соотношения не имеют особого значения.
В одном из подходов биспецифичные антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой специфичностью связывания в одном плече и пары, представляющей собой тяжелую цепь и легкую цепь, гибридного иммуноглобулина (придающей вторую специфичность связывания) в другом плече. Эта асимметричная структура с легкой цепью иммуноглобулина только в одной половине биспецифичной молекулы облегчает отделение желаемого биспецифичного соединения от нежелательных комбинаций цепей иммуноглобулинов. Этот подход описан в публикации РСТ WO № 94/04690, опубликована 3 марта 1994 г.
Гетероконъюгатные антитела, содержащие два ковалентно связанных антитела, также входят в объем этого изобретения. Такие антитела были использованы для нацеливания клеток иммунной системы на нежелательные клетки (патент США №4676980) и для лечения ВИЧ-инфекции (публикация заявки РСТ WO № 91/00360 и WO № 92/200373; ЕР 03089). Гетероконъюгатные антитела можно получать при использовании любых удобных способов поперечного связывания. Подходящие агенты и методики поперечного связывания хорошо известны в данной области и раскрыты в патенте США №4676980.
Антитело может быть гуманизированным антителом, например, таким, какое известно в данной области, и как описано здесь.
Антитела можно модифицировать, как описано в публикации РСТ WO № 99/58572, опубликованной 18 ноября 1999. Эти антитела содержат, в дополнение к связывающему домену, направленному на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную всему константному домену тяжелой цепи человеческого иммуноглобулина или его части. Эти антитела способны связывать молекулу-мишень, не вызывая значительный комплементзависимый лизис или опосредованное клетками разрушение мишени. Предпочтительно эффекторный домен способен специфично связывать FcRn и/или FcγRIIb. Они в типичных случаях основаны на химерных доменах, являющихся производными двух или более чем двух СН2-доменов тяжелой цепи человеческого иммуноглобулина. Антитела, модифицированные таким образом, являются предпочтительными для применения в длительном лечении антителами во избежание воспалительных и других нежелательных реакций на лечение общепринятыми антителами.
Это изобретение охватывает модификации антитела Е3, включая функционально эквивалентные антитела, что не влияет значительно на их свойства, и варианты, которые имеют повышенную или пониженную активность. Модификация полипептидов является обычной практикой в данной области и дополнительно проиллюстрирована в примерах. Примеры модифицированных полипептидов включают в себя полипептиды с заменами (в том числе консервативными заменами) аминокислотных остатков, с одной или более чем одной делецией или добавлением аминокислоты, которая не вызывает значительного изменения функциональной активности, или применение химических аналогов.
Полипептидный "вариант" при использовании здесь представляет собой полипептид, который отличается от нативного белка одной или более чем одной заменой, делецией, добавлением и/или вставкой, так что иммунореактивность полипептида по существу не уменьшается. Другими словами, способность варианта специфично связываться с антигеном может быть усилена или оставаться неизменной относительно нативного белка, или она может быть уменьшена менее чем на 50% и предпочтительно менее чем на 20% относительно нативного белка. Полипептидные варианты предпочтительно проявляют по меньшей мере приблизительно 80%, более предпочтительно по меньшей мере приблизительно 90% и наиболее предпочтительно по меньшей мере приблизительно 95% идентичность (определяемую как описано здесь) с идентифицированными полипептидами.
Варианты аминокислотной последовательности антител можно получать введением соответствующих нуклеотидных изменений в ДНК антитела или синтезом пептида. Такие варианты включают в себя, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях согласно SEQ ID NO:1 или 2, описанных здесь. Любую комбинацию делеции, вставки и замены составляют с получением конечного конструкта при условии, что конечный конструкт обладает желаемыми характеристиками. Аминокислотные изменения также могут изменять посттрансляционные процессы, происходящие с антителом, например изменять число или положение сайтов гликозилирования.
Полезный способ идентификации определенных остатков или областей антитела, которые представляют собой предпочтительные местоположения для мутагенеза или модификации, называют "аланин-сканирующим мутагенезом", он описан в Cunningham and Wells, 1989, Science, 244:1081-1085. Идентифицируют остаток или группу целевых остатков (например, заряженных остатков, таких как arg, asp, his, lys и glu) и заменяют нейтральными или отрицательно заряженными аминокислотами (наиболее предпочтительно аланином или полиаланином), чтобы повлиять на взаимодействие аминокислот с антигеном. Те местоположения аминокислот, которые демонстрируют функциональную чувствительность к заменам, затем оптимизируют введением дополнительных или других вариантов в сайтах замены или для сайтов замены. Таким образом, в то время как сайт для введения вариации аминокислотной последовательности заранее определен, природа мутации сама по себе не обязательно должна быть предопределенной. Например, для анализа свойств мутации в данном сайте осуществляют ala-сканирование или случайный мутагенез в целевом кодоне или области и подвергают экспрессирующиеся варианты антитела скринингу на желаемую активность. Сканирующий мутагенез библиотеки, как описано здесь, можно также использовать для идентификации тех местоположений в антителе, которые являются подходящими для мутагенеза или модификации.
Вставки в аминокислотную последовательность включают в себя слияния по амино- и/или карбокси-концу, варьирующие по длине от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или множества аминокислотных остатков внутрь последовательности. Примеры концевой вставки включают в себя антитело с N-концевым метионильным остатком или антитело, слитое с эпитопом tag. Другие инсерционные варианты молекулы антитела включают в себя слияние N- или С-конца антитела с ферментом или полипептидом, который увеличивает период полужизни антитела в сыворотке.
В вариантах замены удаляют по меньшей мере один аминокислотный остаток в молекуле антитела и встраивают на его место другой остаток. Сайты, представляющие наибольший интерес для замещающего мутагенеза, включают в себя гипервариабельные области, но также предусмотрены изменения каркасных областей (FR). Консервативные замены показаны в Таблице 1 под заголовком "консервативные замены". Если такие замены приводят к изменению биологической активности, то можно вводить более существенные изменения, обозначенные как "примеры замен" в Таблице 1 или дополнительно описанные ниже в ссылке на классы аминокислот, и подвергать продукты скринингу.
Существенные модификации биологических свойств антител осуществляют путем выбора замен, которые значительно отличаются по их влиянию на поддержание (а) структуры полипептидного скелета в области замены, например конформации в виде листа или спирали, (б) заряда или гидрофобности молекулы в сайте-мишени или (в) объема боковой цепи. Встречающиеся в природе остатки подразделяют на группы на основании общих свойств боковой цепи:
(1) гидрофобные: Норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr;
(3) кислые: Asp, Glu;
(4) основные: Asn, Gln, His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro; и
(6) ароматические: Тrp, Тyr, Phe.
Неконсервативные замены получают заменой члена одного из этих классов другим классом.
Любой остаток цистеина, не участвующий в поддержании правильной конформации антитела, также можно заменить, обычно серином, для улучшения устойчивости молекулы против окисления и для предотвращения аберрантного поперечного связывания. Наоборот, цистеиновую(ые) связь(и) можно добавлять к антителу для улучшения его стабильности, особенно если антитело представляет собой фрагмент антитела, такой как Fv-фрагмент.
Модификации аминокислот могут варьировать от изменения или модификации одной или более чем одной аминокислоты до полной перестройки области, такой как вариабельная область. Изменения в вариабельной области могут изменять аффинность и/или специфичность связывания. В некоторых воплощениях делают не более чем 1-5 консервативных замен аминокислот в пределах CDR-домена. В других воплощениях делают не более чем 1-3 консервативных замены аминокислот в пределах CDRS-домена. Еще в других воплощениях CDR-доменом является CDR-H3 и/или CDR-L3.
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование разными сахарами, ацетилирование и фосфорилирование. Антитела гликозилируют в консервативных положениях в их константных областях (Jefferis and Lund, 1997, Chem. Immunol. 65:111-128; Wright and Morrison, 1997, TibTECH 15:26-32). На функции белка влияют олигосахаридные боковые цепи иммуноглобулинов (Boyd et al., 1996, Mol. Immunol. 32:1311 -1318; Wittwe and Howard, 1990, Biochem. 29:4175-4180) и внутримолекулярное взаимодействие между частями гликопротеина, которое может влиять на конформацию и экспонированную трехмерную поверхность гликопротеина (Jefferis and Lund, выше; Wyss and Wagner, 1996, Current Opin. Biotech. 7:409-416). Олигосахариды могут также служить для нацеливания данного гликопротеина на определенные молекулы на основе специфичного распознавания структуры. Также сообщалось, что гликозилирование антител влияет на зависимую от антител клеточную цитотоксичность (ADCC). В частности, сообщалось, что улучшенную ADCC-активность имеют клетки СНО с регулируемой тетрациклином экспрессией β-(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), причем гликозилтрансфераза катализирует образование разделенного пополам GlcNAc (Umana et al., 1999, Nature Biotech. 17:176-180).
Гликозилирование антитела в типичных случаях является либо N-связанным, либо O-связанным. N-связанное относится к присоединению углеводной группировки к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет собой любую аминокислоту за исключением пролина, являются последовательностями распознавания для ферментативного присоединения углеводной группировки к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров, N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, наиболее обычно к серину или треонину, хотя также могут быть использованы 5-гидроксипролин или 5-гидроксилизин.
Добавление сайтов гликозилирования к антителу удобно осуществлять, изменяя аминокислотную последовательность так, чтобы она содержала одну или более чем одну вышеописанную трипептидную последовательность (для сайтов N-связанного гликозилирования). Это изменение можно также получать добавлением одного или более чем одного остатка серина или треонина к последовательности исходного антитела или ее заменой ими (для O-связанных сайтов гликозилирования).
Картину гликозилирования антител также можно изменить без изменения лежащей в основе нуклеотидной последовательности. Гликозилирование в значительной степени зависит от клетки-хозяина, использованной для экспрессии антитела. Поскольку тип клетки, использованной для экспрессии рекомбинантных гликопротеинов, например антител, в качестве потенциальных терапевтических средств, редко представляет собой нативную клетку, можно ожидать вариации в картине гликозилирования антител (см., например, Hse et al., 1997, J. Biol. Chem. 272:9062-9070).
Факторы, которые влияют на гликозилирование во время рекомбинантной продукции антител, включают в себя, в дополнение к выбору клетки-хозяина, характер роста, состав среды, плотность культуры, насыщение кислородом, рН, схемы очистки и тому подобное. Для изменения картины гликозилирования, осуществляемой в конкретном организме-хозяине, предложены различные способы, в том числе введение или сверхэкспрессию определенных ферментов, участвующих в продукции олигосахаридов (патенты США №№5047335, 5510261 и 5278299). Гликозилирование или некоторые типы гликозилирования можно удалять ферментативным путем из гликопротеина, например, с использованием эндогликозидазы Н (Endo H). В дополнение, рекомбинантную клетку-хозяина можно генетически сконструировать так, чтобы она являлась дефективной по процессингу определенных типов полисахаридов. Эти и аналогичные методики хорошо известны в данной области.
Другие способы модификации включают в себя использование методик связывания, известных в данной области, в том числе, без ограничения ими, ферментативных средств, окислительной замены и хелатирования. Модификации можно использовать, например, для присоединения меток для иммуноанализа. Модифицированные полипептиды Е3 получают при использовании процедур, признанных в данной области, и их можно подвергать скринингу при использовании стандартных анализов, известных в данной области, некоторые из которых описаны ниже и в Примерах.
Другие модификации антитела включают в себя антитела, которые были модифицированы, как описано в публикации РСТ WO № 99/58572, опубликованной 18 ноября 1999 г. Эти антитела содержат, в дополнение к связывающему домену, направленному на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную всему константному домену тяжелой цепи человеческого иммуноглобулина или части этого домена. Эти антитела способны связывать молекулу-мишень, не вызывая значительного комплементзависимого лизиса или опосредованного клетками разрушения мишени. В некоторых воплощениях эффекторный домен способен специфично связывать FcRn и/или FcγRIIb. В типичных случаях они основаны на химерных доменах, происходящих из двух или более СН2-доменов тяжелых цепей человеческих иммуноглобулинов. Антитела, модифицированные таким образом, особенно подходят для применения в длительном лечении антителами во избежание воспалительных и других нежелательных реакций на общепринятое лечение антителами.
Это изобретение также охватывает слитые белки, содержащие один или более чем один фрагмент или область антител (таких как Е3) или полипептидов по этому изобретению. В одном из воплощений предложен слитый полипептид, содержащий по меньшей мере 10 последовательных аминокислот вариабельной области легкой цепи, показанной на Фиг.1Б, и/или по меньшей мере 10 аминокислот вариабельной области тяжелой цепи, показанной на Фиг.1А. В еще одном воплощении слитый полипептид содержит вариабельную область легкой цепи и/или вариабельную область тяжелой цепи Е3, как показано на Фиг.1А и 1Б. В еще одном воплощении слитый полипептид содержит один или более чем один CDR Е3. В еще других воплощениях слитый полипептид содержит CDR-H3 и/или CDR-L3 антитела Е3. В еще одном воплощении слитый полипептид содержит любое одно или более чем одно из следующего: аминокислотный остаток L29 CDR-H1, I50 CDR-H2, W101 CDR-H3 и/или А103 CDR-H3 и/или аминокислотный остаток S28 CDR-L1, N32 CDR-L1, Т51 CDR-L2, 91Е CDR-L3 и/или Н92 CDR-L3. Для целей этого изобретения слитый белок Е3 содержит одно или более чем одно антитело Е3 и другую аминокислотную последовательность, к которой оно не присоединено в нативной молекуле, например гетерологичную последовательность или гомологичную последовательность из другой области. Примеры гетерологичных последовательностей включают в себя, без ограничения ими, "метку", такую как метка FLAG или метка 6His. Метки хорошо известны в данной области.
Слитый полипептид Е3 можно создать способами, известными в данной области, например синтетическим или рекомбинантным способом. В типичных случаях слитые белки Е3 по этому изобретению создают получением экспрессионного полинуклеотида, кодирующего их, с использованием рекомбинантных способов, описанных здесь, хотя их можно также получать другими способами, известными в данной области, в том числе, например, химическим синтезом.
В этом изобретении также предложены композиции, содержащие антитела Е3 или полипептиды, конъюгированные (например, связанные) с агентом, который облегчает связывание с твердой подложкой (такой как биотин или авидин). Для простоты, в общем, следует упомянуть Е3 или антитела, причем надо понимать, что эти способы применимы к любым связывающим NGF воплощениям, описанным здесь. Конъюгация обычно относится к связыванию этих компонентов, как описано здесь. Связывание (которое обычно представляет собой фиксацию этих компонентов в тесной ассоциации по меньшей мере для введения) можно осуществлять любым числом способов. Например, возможно прямое взаимодействие между агентом и антителом, когда каждый обладает заместителем, способным к взаимодействию с другим. Например, нуклеофильная группа, такая как группа амино или сульфгидрил, на одном может быть способна взаимодействовать с карбонилсодержащей группой, такой как ангидрид или галогенангидрид, или с алкильной группой, содержащей хорошую уходящую группу (например, галогенид), на другом.
Антитело или полипептид по этому изобретению можно связывать с метящим агентом (иначе обозначаемым как "метка"), таким как флуоресцентная молекула, радиоактивная молекула или любые другие метки, известные в данной области. Метки, известные в данной области, обычно обеспечивают сигнал (либо непосредственно, либо опосредованно). Соответственно это изобретение включает меченые антитела и полипептиды.
Способность антител и полипептидов по этому изобретению, такую как связывание NGF, снижение или ингибирование биологической активности NGF, снижение и/или блокирование NGF-индуцированного выживания мышиных нейронов тройничного нерва Е13.5, можно тестировать с использованием способов, известных в данной области, некоторые из которых описаны в примерах.
В этом изобретении также предложены композиции (в том числе фармацевтические композиции) и наборы, содержащие антитело Е3, и, как поясняется в данном описании, любое или все из антител и/или полипептидов, описанных здесь.
Полинуклеотиды, векторы и клетки-хозяева
В этом изобретении также предложены выделенные полинуклеотиды, кодирующие антитела и полипептиды по этому изобретению (в том числе антитело, содержащее полипептидные последовательности легкой цепи и вариабельные области тяжелой цепи, показанные на Фиг.1А и 1Б), и векторы и клетки-хозяева, содержащие такой полинуклеотид.
Соответственно в этом изобретении предложены полинуклеотиды (или композиции, в том числе фармацевтические композиции), содержащие полинуклеотиды, кодирующие любое из следующего: (а) антитело Е3; (б) фрагмент или область антитела Е3; (в) легкую цепь антитела Е3, как показано на Фиг.1Б; (г) тяжелую цепь антитела Е3, как показано на Фиг.1А; (д) одну или более чем одну вариабельную область легкой цепи и/или тяжелой цепи антитела Е3; (е) один или более чем один CDR (один, два, три, четыре, пять или шесть CDR) антитела Е3, показанные на Фиг.1А и 1Б; (ж) CDR-H3 тяжелой цепи антитела Е3, показанный на Фиг.1А; (з) CDR-L3 легкой цепи антитела Е3, показанный на Фиг.1Б; (и) три CDR легкой цепи антитела Е3, показанные на Фиг.1Б; (к) три CDR тяжелой цепи антитела Е3, показанные на Фиг.1А; (л) три CDR легкой цепи и три CDR тяжелой цепи антитела Е3, показанные на Фиг.1А и 1Б; или (м) антитело, содержащее любое из (б)-(л). В некоторых воплощениях полинуклеотид содержит любой или оба из полинуклеотидов, показанных на Фиг.2 и 3.
В другом аспекте это изобретение представляет собой выделенный полинуклеотид, который кодирует легкую цепь Е3, с номером депонирования АТСС № РТА-4893 или АТСС № РТА-4894. В другом аспекте это изобретение представляет собой выделенный полинуклеотид, который кодирует тяжелую цепь Е3, с номером депонирования АТСС № РТА-4895. В еще одном аспекте это изобретение представляет собой выделенный полинуклеотид, содержащий (а) вариабельную область, кодируемую в полинуклеотиде с номером депонирования АТСС № РТА-4894, и (б) вариабельную область, кодируемую в полинуклеотиде с номером депонирования АТСС № РТА-4895. В другом аспекте это изобретение представляет собой выделенный полинуклеотид, содержащий (а) один или более чем один CDR, кодируемый в полинуклеотиде с номером депонирования АТСС № РТА-4894; и/или (б) один или более чем один CDR, кодируемый в полинуклеотиде с номером депонирования АТСС № РТА-4895.
В другом аспекте в этом изобретении предложены полинуклеотиды, кодирующие любое антитело (включая фрагменты антитела) и полипептид, описанный здесь. Эти полинуклеотиды можно получать способами, известными в данной области.
В другом аспекте в этом изобретении предложены композиции (такие как фармацевтические композиции), содержащие любой полинуклеотид по этому изобретению. В некоторых воплощениях композиция содержит экспрессионный вектор, содержащий полинуклеотид, кодирующий антитело Е3, как описано здесь. В другом воплощении композиция содержит экспрессионный вектор, содержащий полинуклеотид, кодирующий любое антитело или полипептид, описанные здесь. В еще других воплощениях композиция содержит любой или оба из полинуклеотидов, показанных на Фиг.2 и 3. Экспрессионные векторы и введение полинуклеотидной композиции описаны здесь далее.
В другом аспекте в этом изобретении предложен способ получения любого полинуклеотида, описанного здесь.
Полинуклеотиды, комплементарные любым таким последовательностям, также охватываются настоящим изобретением. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными, и могут представлять собой молекулы ДНК (геномной ДНК, кДНК или синтетической ДНК) или РНК. Молекулы РНК включают в себя молекулы гетерогенной ядерной РНК, которые содержат интроны и соответствуют молекуле ДНК один к одному, и молекулы мРНК, которые не содержат интронов. В полинуклеотиде по настоящему изобретению могут присутствовать, но не обязательно, дополнительные кодирующие или некодирующие последовательности, и полинуклеотид может, но не обязательно, быть связанным с другими молекулами и/или материалами подложки.
Полинуклеотиды могут содержать нативную последовательность (то есть эндогенную последовательность, которая кодирует антитело или его часть) или могут содержать вариант такой последовательности. Варианты полинуклеотида содержат одну или более чем одну замену, добавление, делецию и/или вставку так, что иммунореактивность кодируемого полипептида не снижена относительно нативной иммунореактивной молекулы. Обычно влияние на иммунореактивность кодируемого полипептида можно оценивать, как описано здесь. Варианты предпочтительно проявляют по меньшей мере приблизительно 70% идентичность, более предпочтительно по меньшей мере приблизительно 80% идентичность и наиболее предпочтительно по меньшей мере приблизительно 90% идентичность с полинуклеотидной последовательностью, которая кодирует нативное антитело или его часть.
Две последовательности полинуклеотида или полипептида указаны как "идентичные", если последовательности нуклеотидов или аминокислот в этих двух последовательностях являются одними и теми же, когда они выровнены по максимуму соответствия, как описано ниже. Сравнения между двумя последовательностями в типичных случаях осуществляют сравнением последовательностей на протяжении окна сравнения для идентификации и сравнения локальных областей сходства последовательностей. "Окно сравнения" при использовании здесь относится к сегменту, представляющему собой по меньшей мере приблизительно 20 последовательных положений, обычно от 30 до приблизительно 75, от 40 до приблизительно 50, в котором последовательность можно сравнивать относительно последовательности, имеющей то же число последовательных положений, после оптимального выравнивания этих двух последовательностей.
Оптимальное выравнивание последовательностей для сравнения можно осуществлять при использовании программы Megalign в пакете биоинформационных программ Lasergene (DNASTAR, Inc., Madison, WI) при использовании параметров, заданных по умолчанию. Эта программа реализует несколько схем выравнивания, описанных в следующих ссылках: Dayhoff, M.O. (1978) Model of evolutionary change in proteins - Matrices for detecting distant relationships. Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington DC, Vol.5, Suppl. 3, pp.345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes, pp.626-645, Methods in Enzymology, vol.183, Academic Press, Inc., San Diego, CA; Higgins, D.G. and Sharp, P.M., 1989, CABIOS 5:151-153; Myers, E.W. and Muller W., 1988, CABIOS 4:11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R.R., 1973, Numeric Taxonomy the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80:726-730.
Предпочтительно "процент идентичности последовательностей" определяют сравнением двух оптимально выровненных последовательностей на протяжении окна сравнения, представляющего собой по меньшей мере 20 положений, причем часть последовательности полинуклеотида или полипептида в окне сравнения может содержать добавления или делеции (то есть пробелы) в количестве 20 процентов или менее чем 20 процентов, обычно от 5 до 15 процентов или от 10 до 12 процентов, при сравнении с эталонными последовательностями (которые не содержат добавления или делеции) для оптимального выравнивания двух последовательностей. Процент вычисляют определением числа положений, по которым идентичные основания нуклеиновой кислоты или аминокислотные остатки имеются в обеих последовательностях, с получением числа совпадающих положений, делением числа совпадающих положений на суммарное число положений в эталонной последовательности (то есть на размер окна) и умножением результатов на 100 с получением процента идентичности последовательности.
Варианты могут также или альтернативно быть по существу гомологичными нативному гену или его части или комплементу. Такие варианты полинуклеотидов способны к гибридизации в умеренно строгих условиях с встречающейся в природе последовательностью ДНК, кодирующей нативное антитело (или с комплементарной последовательностью).
Подходящие "умеренно строгие условия" включают в себя предварительную промывку в растворе 5×SSC (SSC представляет собой 0,15 М хлорид натрия и 0,015 М трехзамещенный цитрат натрия), 0,5%-ном додецилсульфате натрия, 1,0 мМ ЭДТА (рН 8,0); гибридизацию при 50-65°С, 5×SSC, в течение ночи и последующую промывку дважды при 65°С в течение 20 минут с каждым из 2×, 0,5× и 0,2×SSC, содержащим 0,1%-ный додецилсульфат натрия.
При использовании здесь "очень строгие условия" или "условия высокой строгости" представляют собой условия, в которых: (1) используют низкую ионную силу и высокую температуру для промывки, например 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) во время гибридизации используют денатурирующий агент, такой как формамид, например 50%-ный (объем/объем) формамид со смесью 0,1%-ный бычий сывороточный альбумин/0,1%-ный Фиколл/0,1%-ный поливинилпирролидон/50 мМ натрий-фосфатный буфер при рН 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°С; или (3) используют 50%-ный формамид, 5×SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1%-ный пирофосфат натрия, 5×раствор Денхардта (Denhardt), обработанную ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1%-ный додецилсульфат натрия и 10%-ный декстрансульфат при 42°C с промывками при 42°С в 0,2×SSC (хлорид натрия/цитрат натрия) и 50%-ном формамиде при 55°С, с последующей промывкой высокой строгости, состоящей из 0,1×SSC, содержащего ЭДТА, при 55°С. Специалист должен понимать, как подбирать температуру, ионную силу и так далее, как это необходимо для учета таких факторов, как длина зонда и тому подобные.
Обычные специалисты в данной области должны понимать, что в результате дегенерации генетического кода существует много нуклеотидных последовательностей, которые кодируют описанный здесь полипептид. Некоторые из этих полинуклеотидов имеют минимальную гомологию с нуклеотидной последовательностью любого нативного гена. Тем не менее, полинуклеотиды, которые варьируют ввиду отличий в использовании кодонов, конкретно предусмотрены настоящим изобретением. Дополнительно в объем настоящего изобретения входят аллели генов, содержащих предложенные здесь полинукпеотидные последовательности. Аллели представляют собой эндогенные гены, которые изменены в результате одной или более чем одной мутации, такой как делеция, добавление и/или замена нуклеотида. Получившиеся мРНК и белок могут, но не обязательно, иметь измененную структуру или функцию. Аллели можно идентифицировать при использовании стандартных методик (таких как гибридизация, амплификация и/или сравнение баз данных по последовательностям).
Полинуклеотиды по этому изобретению можно получать при использовании химического синтеза, рекомбинантных способов или ПЦР. Способы химического синтеза полинуклеотидов хорошо известны в данной области, и нет необходимости описывать их здесь подробно. Специалист в данной области может использовать предложенные здесь последовательности и коммерческий синтезатор ДНК с получением желаемой последовательности ДНК.
Для получения полинуклеотидов с использованием рекомбинантных способов полинуклеотид, содержащий желаемую последовательность, можно встроить в подходящий вектор, и вектор в свою очередь может быть введен в подходящую клетку-хозяина для репликации и амплификации, как обсуждается здесь далее. Полинуклеотиды можно вводить в клетки-хозяева любыми способами, известными в данной области. Клетки трансформируют введением экзогенного полинуклеотида прямым поглощением, эндоцитозом, трансфекцией, F-спариванием или электропорацией. После введения экзогенный полинуклеотид можно поддерживать в клетке в качестве неинтегрированного вектора (такого как плазмида) или вектора, интегрированного в геном клетки-хозяина. Полинуклеотид, который амплифицировали таким образом, можно выделить из клетки-хозяина способами, хорошо известными в данной области. См., например, Sambrook et al. (1989).
Альтернативно, репродукцию последовательностей ДНК делает возможной ПЦР. Технология ПЦР хорошо известна в данной области и описана в патентах США №№4683195, 4800159, 4754065 и 4683202, а также в PCR: Polymerase Chain Reaction, Mullis et al. Eds., Birkauswer Press, Boston (1994).
РНК можно получать использованием выделенной ДНК в соответствующем векторе и введением его в подходящую клетку-хозяина. Когда клетка реплицируется и ДНК транскрибируется в РНК, РНК можно затем выделить при использовании способов, хорошо известных специалистам в данной области, как указано, например, в Sambrook et al. (1989).
Подходящие векторы для клонирования можно конструировать в соответствии со стандартными методиками или можно выбирать из большого числа векторов для клонирования, имеющихся в данной области. Хотя выбранный вектор для клонирования может варьировать в соответствии с клеткой-хозяином, предназначенной для использования, полезные векторы для клонирования обычно должны иметь способность к саморепликации, могут иметь единственную мишень для конкретной рестрикционной эндонуклеазы и/или могут нести гены для маркера, который можно использовать в селекции клонов, содержащих вектор. Подходящие примеры включают в себя плазмиды и бактериальные вирусы, например pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и челночные векторы, такие как рSА3 и рАТ28. Эти и многие другие векторы для клонирования доступны от коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen.
Экспрессионные векторы обычно представляют собой реплицируемые полинуклеотидные конструкции, которые содержат полинуклеотид в соответствии с этим изобретением. Подразумевается, что экспрессионный вектор должен быть реплицируемым в клетках-хозяевах либо в виде эписом, либо в виде интегральной части хромосомной ДНК. Подходящие экспрессионные векторы включают в себя, без ограничения ими, плазмиды, вирусные векторы, в том числе аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и экспрессионный(е) вектор(ы), раскрытый(е) в публикации РСТ WO № 87/04462. Компоненты вектора обычно могут включать в себя, без ограничения, одно или более чем одно из следующего: сигнальная последовательность; точка начала репликации; один или более чем один маркерный ген; подходящие элементы транскрипционного контроля (такие как промоторы, энхансеры и терминаторы). Для экспрессии (то есть для трансляции) также обычно требуется один или более чем один элемент трансляционного контроля, такой как сайты связывания рибосом, сайты инициирования трансляции и стоп-кодоны.
Векторы, которые содержат полинуклеотиды, представляющие интерес, можно вводить в клетку-хозяина любым из многих подходящих способов, в том числе электропорацией, трансфекцией при использовании хлорида кальция, хлорида рубидия, фосфата кальция, ДЭАЭ-декстрана или других субстанций; бомбардировкой микроперфораторами; липофекцией и инфекцией (например, если вектор представляет собой инфекционный агент, такой как вирус осповакцины). Выбор введения векторов или полинуклеотидов часто должен зависеть от свойств клетки-хозяина.
В этом изобретении также предложены клетки-хозяева, содержащие любой описанный здесь полинуклеотид. Любые клетки-хозяева, способные к повышенной экспрессии гетерологичных ДНК, можно использовать для цели выделения генов, кодирующих антитело, полипептид или белок, представляющий интерес. Неограничивающие примеры клеток-хозяев из млекопитающего включают в себя, без ограничения ими, клетки COS, HeLa и СНО. См. также публикацию РСТ WO № 87/04462. Подходящие клетки-хозяева не из млекопитающего включают в себя прокариоты (такие как Е.coli или В.subtilis) и дрожжи (такие как S.cerevisiae, S.pombe или K.lactis). Предпочтительно клетки-хозяева экспрессируют кДНК на уровне, приблизительно в 5 раз более высоком, более предпочтительно в 10 раз более высоком, даже более предпочтительно в 20 раз более высоком, чем тот, который соответствует представляющему интерес эндогенному антителу или белку, при его наличии, в клетках-хозяевах. Скрининг клеток-хозяев на специфичное связывание с NGF осуществляют иммуноанализом или сортировкой клеток по активации флуоресценции (FACS). Можно идентифицировать клетку с повышенной экспрессией антитела или белка, представляющего интерес.
Способы использования Е3 и антител, производных от Е3
Антитело Е3, которое связывается с NGF, можно использовать для идентификации или детектирования присутствия или отсутствия NGF. Для простоты речь, в общем, пойдет о Е3 или антителах, причем следует понимать, что эти способы применимы к любым связывающим NGF воплощениям (таким как полипептиды), описанным здесь. Детектирование обычно включает в себя приведение биологического образца в контакт с описанным здесь антителом, которое связывается с NGF и образует комплекс между NGF и антителом (например, Е3), которое специфично связывается с NGF. Образование такого комплекса возможно in vitro или in vivo. Термин "детектирование" при использовании здесь включает в себя качественное и/или количественное детектирование (измерение уровней) относительно или безотносительно контроля.
Для детектирования можно использовать любой из множества разных известных способов, в том числе, без ограничения ими, иммуноанализ при использовании антитела, которое связывается с полипептидом, например твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) и тому подобные; и функциональный анализ на кодируемый полипептид, например связывающую активность или ферментативный анализ. В некоторых воплощениях антитело детектируемым образом метят.
Диагностические применения Е3 и производных
Антитела и полипептиды по этому изобретению можно использовать в детектировании, диагностике и мониторинге заболевания, состояния или расстройства, связанного с измененной или аберрантной экспрессией NGF (повышенной или сниженной экспрессией NGF (относительно нормальной пробы в некоторых воплощениях) и/или с неприемлемой экспрессией, такой как наличие экспрессии в ткани(ях) и/или клетки(ах), где в норме нет экспрессии NGF, или отсутствие экспрессии NGF в ткани(ях) или клетке(ах), где в норме есть экспрессия NGF). Антитела и полипептиды по этому изобретению дополнительно являются полезными для детектирования экспрессии NGF, например, при заболевании, связанном с измененной или аберрантной чувствительностью к NGF или реактивностью на NGF. В некоторых воплощениях экспрессию NGF детектируют в образце от индивида, у которого предположительно имеется заболевание или расстройство, которому свойственна измененная или аберрантная чувствительность к экспрессии NGF или реактивность на экспрессию NGF или которое связано с ней (например, рак, при котором NGF стимулирует рост и/или метастазы).
Таким образом, в некоторых воплощениях в этом изобретении предложены способы, включающие приведение в контакт образца (пробы) из индивида, у которого предположительно имеется измененная или аберрантная экспрессия NGF, с антителом или полипептидом по этому изобретению и определение того, отличается ли уровень NGF от такового в контроле или образце сравнения. В некоторых воплощениях индивид имеет сердечную аритмию, болезнь Альцгеймера и/или автономную дисфункцию.
В других воплощениях в этом изобретении предложены способы, включающие приведение в контакт образца (пробы) из индивида и определение уровня экспрессии NGF. В некоторых воплощениях у индивида предположительно имеется заболевание, расстройство, которому свойственна измененная или аберрантная чувствительность к экспрессии NGF или реактивность на экспрессию NGF или которое связано с ней. В некоторых воплощениях индивид имеет мелкоклеточный рак легких, рак молочной железы, рак поджелудочной железы, рак простаты, карциному яичника, гепатоцеллюлярную карциному или меланому.
Для диагностических применений антитело в типичных случаях должно быть меченным детектируемой группировкой, включая, без ограничения ими, радиоизотопы, флуоресцентные метки и различные фермент-субстратные метки. Способы конъюгации меток с антителом известны в данной области. В других воплощениях этого изобретения антитела по этому изобретению не обязательно являются мечеными, и их присутствие можно детектировать при использовании меченого антитела, которое связывается с антителами по этому изобретению.
Антитела по настоящему изобретению можно использовать в любом известном способе анализа, таком как анализы конкурентного связывания, прямые и непрямые анализы с множественными антителами и анализы по иммунопреципитации. Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc. 1987).
Антитела можно также использовать для диагностических анализов in vivo, таких как визуализация in vivo. Обычно антитело метят радионуклидом (таким как 111In, 99Tc, 14С, 131I, 125I или 3H) так, что клетки или ткань, представляющие интерес, можно локализовать при использовании иммуносцинтиографии.
Антитело можно также использовать как окрашивающий реагент в патологии, следуя методикам, хорошо известным в данной области.
Способы применения Е3 и производных в терапевтических целях
Антитело Е3 является полезным для снижения и/или блокирования биологической активности NGF. Эту антагонистическую активность считают полезной в лечении патологических состояний, связанных с эндогенной продукцией NGF, таких как боль. Обычно в этих воплощениях индивиду вводят эффективное количество. Соответственно в одном из аспектов в этом изобретении предложен способ антагонизации биологической активности человеческого NGF при использовании любых раскрытых здесь полипептидов (в том числе антител, таких как антитело Е3). В одном из воплощений этот способ включает приведение в контакт человеческого фактора роста нервов с любым описанным здесь полипептидом (включая антитело Е3), тем самым активность человеческого фактора роста нервов антагонизируется, снижается, блокируется или подавляется. В еще одном воплощении индивид с болью (такой как послеоперационная боль или боль, вызванная ревматоидным артритом) получает лечение при использовании Е3.
Для простоты ссылка, в общем, будет дана на Е3 или антитело, причем следует понимать, что эти способы применимы к любому описанному здесь варианту антитела Е3 и полипептиду.
Для введения можно использовать различные препараты Е3 или фрагменты Е3 (например, Fab, Fab', F(ab')2, Fv, Fc и так далее), такие как одноцепочечные (ScFv), их мутанты, слитые белки, содержащие часть антитела, и любую другую модифицированную конфигурацию Е3, которая содержит сайт распознавания антигена NGF требуемой специфичности. В некоторых воплощениях можно вводить антитела Е3 или различные препараты Е3 как таковые. В других воплощениях вводят Е3 или различные препараты Е3 (в том числе любое воплощение, которое представляет собой описанную здесь композицию) и фармацевтически приемлемый эксципиент, и они могут быть в различных препаратах. Фармацевтически приемлемые эксципиенты известны в данной области и представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества. Например, эксципиент может придавать форму или консистенцию или действовать как разбавитель. Подходящие эксципиенты включают в себя, без ограничения ими, стабилизирующие агенты, смачивающие и эмульгирующие агенты, соли для варьирования осмолярности, инкапсулирующие агенты, буферы и усилители проницаемости кожи. Эксципиенты, а также препараты для парентеральной и непарентеральной доставки лекарственного средства указаны в Remington, The Science and Practice of Pharmaceutics, 20th Ed. Mack Publishing (2000).
В некоторых воплощениях эти агенты приготавливают для введения инъекцией (например, внутрибрюшинной, внутривенной, подкожной, внутримышечной и так далее), хотя также можно использовать другие формы введения (например, пероральное, на слизистые, ингаляцией, под язык и так далее). Соответственно антитело Е3 и его эквиваленты предпочтительно объединяют с фармацевтически приемлемыми носителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и тому подобные. Конкретный режим дозировки, то есть доза, время и кратность введения, должен зависеть от конкретного индивида и медицинской истории этого индивида. Обычно можно использовать любую из следующих доз: вводят дозу по меньшей мере около 50 мг/кг массы тела; по меньшей мере около 10 мг/кг массы тела; по меньшей мере около 3 мг/кг массы тела; по меньшей мере около 1 мг/кг массы тела; по меньшей мере около 750 мкг/кг массы тела; по меньшей мере около 500 мкг/кг массы тела; по меньшей мере около 250 мкг/кг массы тела; по меньшей мере около 100 мкг/кг массы тела; по меньшей мере около 50 мкг/кг массы тела; по меньшей мере около 10 мкг/кг массы тела; по меньшей мере около 1 мкг/кг массы тела или менее. Для повторных введений в течение нескольких суток или дольше в зависимости от состояния лечение продолжают, пока не наступит желаемое подавление симптомов заболевания. Пример режима введения включает введение исходной дозы около 2 мг/кг с последующими еженедельными поддерживающими дозами около 1 мг/кг антитела против NGF или с последующими поддерживающими дозами около 1 мг/кг через неделю. Однако могут быть полезными и другие режимы дозировки в зависимости от картины фармакокинетического профиля, который хочет достичь практикующий врач. Обычно в определение дозировки должны вносить вклад эмпирические соображения, такие как период полувыведения. Ход этого лечения легко контролировать общепринятыми методиками и анализами.
Для некоторых индивидов может потребоваться более чем одна доза. Частоту введения можно определять и подбирать по ходу лечения. Например, частоту введения можно определять или подбирать на основании типа и тяжести боли, подлежащей лечению, независимо от того, вводят ли агент для профилактических или терапевтических целей, предыдущего лечения, клинической истории пациента и реакции на данный агент и выбора лечащего врача. В типичных случаях врач-клиницист вводит антагонистическое антитело против NGF (такое как Е3), пока не достигнет дозировки, которая дает желаемый результат. В некоторых случаях могут быть подходящими препараты антител Е3 для непрерывного длительного высвобождения. Различные препараты и устройства для достижения непрерывного высвобождения известны в данной области.
В одном из воплощений можно определять дозировки антител Е3 (или полипептидов) эмпирически у индивидов, которые получали одно или более чем одно введение. Индивидам дают возрастающие дозировки Е3. Для оценки эффективности Е3 или другого эквивалентного антитела можно контролировать маркеры симптомов заболевания (такие как боль).
Введение антитела (такого как Е3) или полипептида в соответствии со способом по настоящему изобретению может быть непрерывным или прерывистым в зависимости, например, от физиологического состояния реципиента, от того, является ли цель введения терапевтической или профилактической, и от других факторов, известных специалисту-практику. Введение антитела может быть по существу непрерывным на протяжении предварительно выбранного периода времени или может быть серией доз, распределенных по времени, например до, во время или после развития боли, до, во время, до и после, во время и после или до, во время и после развития боли. Введение может быть до, во время и/или после получения раны, надреза, травмы, хирургической операции и любого другого случая, вероятно, вызывающего усиление послеоперационной боли.
Другие препараты включают в себя подходящие формы доставки, известные в данной области, в том числе, без ограничения ими, носители, такие как липосомы. См., например, Mahato et al. (1997) Pharm. Res. 14:853-859. Липосомальные препараты включают в себя, без ограничения ими, цитофектины, многослойные пузырьки и однослойные пузырьки.
В некоторых воплощениях может присутствовать более чем одно антитело или полипептид. Антитела могут быть моноклональными или поликлональными. Такие композиции могут содержать по меньшей мере одно, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять разных антител. Смесь антител, как их часто обозначают в данной области, может быть особенно полезной в лечении более широкого диапазона популяции индивидов.
Полинуклеотид, кодирующий любое из антител или полипептидов по этому изобретению (таких как антитело Е3), также можно использовать для доставки и экспрессии любых антител или полипептидов по этому изобретению (таких как антитело Е3) в желаемую клетку. Ясно, что экспрессионный вектор можно использовать для управления экспрессией антитела Е3 или полипептида. Экспрессионный вектор можно вводить любым способом, известным в данной области, таким как внутрибрюшинный, внутривенный, внутримышечный, подкожный, в мозговые оболочки, в желудочек мозга, пероральный, энтеральный, парентеральный, интраназальный, дермальный, сублингвальный или путем ингаляции. Например, введение экспрессионных векторов включает в себя местное или системное введение, включая инъекцию, пероральное введение, бомбардировку частицами или введение через катетер и местное введение. Специалисту известно введение экспрессионных векторов для достижения экспрессии экзогенного белка in vivo. См., например, патенты США №№6436908, 6413942 и 6376471.
Можно также использовать направленную доставку терапевтической композиции, содержащей полинуклеотид, кодирующий любое из антител или полипептидов по этому изобретению (таких как антитело Е3). Методики опосредованной рецепторами доставки ДНК раскрыты, например, в Findeis et al., Trends Biotechnol. (1993) 11:202; Chlou et al., Gene Therapeutics: Methods and Applications Of Direct Gene Transfer (J.A. Wolff, ed.) (1994); Wu et al., J. Biol. Chem. (1988) 263:621; Wu et al., J. Biol. Chem. (1994) 269:542; Zenke et al., Proc. Natl. Acad. Sci. (USA) (1990) 87:3655; Wu et al., J. Biol. Chem. (1991) 266:338. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от приблизительно 100 нг до приблизительно 200 мг ДНК для местного введения по протоколу генной терапии. Диапазоны концентрации от приблизительно 500 нг до приблизительно 50 мг, от приблизительно 1 мкг до приблизительно 2 мг, от приблизительно 5 мкг до приблизительно 500 мкг и от приблизительно 20 мкг до приблизительно 100 мкг ДНК также можно использовать в протоколе генной терапии. Терапевтические полинуклеотиды и полипептиды по настоящему изобретению можно доставлять при использовании носителей для доставки генов. Носитель для доставки генов может быть вирусного или невирусного происхождения (см., в общем, Jolly, Cancer Gene Therapy (1994) 1:51; Kimura, Human Gene Therapy (1994) 5:845; Connelly, Human Gene Therapy (1995) 1:185; и Kaplitt, Nature Genetics (1994) 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать, используя эндогенные для млекопитающего или гетерологичные промоторы. Экспрессия кодирующей последовательности может быть как конститутивной, так и регулируемой.
Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемую клетку хорошо известны в данной области. Примеры носителей на основе вирусов включают в себя, без ограничения ими, рекомбинантные ретровирусы (см., например, публикации РСТ WO 90/07936; WO 94/03622; WO 93/25698; WO 93/25234; WO 93/11230; WO 93/10218; WO 91/02805; патенты США №№5219740; 4777127; патент GB №2200651 и патент ЕР №0345242), векторы на основе альфавирусов (например, векторы, представляющие собой вирус Синдбис, вирус леса Семлики (АТСС VR-67; АТСС VR-1247), вирус реки Росс (АТСС VR-373; АТСС VR-1246) и вирус венесуэльского энцефаломиелита лошадей (АТСС VR-923; АТСС VR-1250; АТСС VR 1249; АТСС VR-532)) и векторы, представляющие собой аденоассоциированный вирус (AAV) (см., например, публикации РСТ WO 94/12649, WO 93/03769; WO 93/19191; WO 94/28938; WO 95/11984 и WO 95/00655). Можно также использовать введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, Hum. Gene Ther. (1992) 3:147.
Также можно использовать невирусные носители и способы доставки, в том числе, без ограничения ими, конденсированную с поликатионами ДНК, связанную или не связанную с убитым аденовирусом в отдельности (см., например, Curiel, Gene Ther. (1992) 3:147); лиганд-связанную ДНК (см., например, Wu, J. Biol. Chem. (1989) 264:16985); клеточные носители для доставки, представляющие собой эукариотические клетки (см., например, патент США №5814482; публикации РСТ WO 95/07994, WO 96/17072, WO 95/30763 и WO 97/42338), и нейтрализацию заряда ядра или слияние с мембранами клетки. Также можно использовать голую ДНК. Примеры способов введения голой ДНК раскрыты в публикации РСТ WO № 90/11092 и патенте США №5580859. Липосомы, которые могут действовать как носители для доставки генов, раскрыты в патенте США №5422120, в публикациях РСТ WO 95/13796, WO 94/23697, WO 91/14445 и в патенте ЕР №0524968. Дополнительные подходы раскрыты в Philip, Mol. Cell Biol. (1994) 14:2411; и в Woffendin, Proc. Natl. Acad. Sci. (1994) 91:1581.
Что касается всех описанных здесь способов, упоминание антагонистических антител против NGF также включает композиции, содержащие один или более чем один из этих агентов. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, в том числе буферы, которые хорошо известны в данной области. Настоящее изобретение можно использовать в отдельности или в комбинации с другими общепринятыми способами лечения.
СПОСОБЫ ИСПОЛЬЗОВАНИЯ АНТАГОНИСТИЧЕСКОГО АНТИТЕЛА ПРОТИВ NGF ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ БОЛИ, ВЫЗВАННОЙ РЕВМАТОИДНЫМ АРТРИТОМ
В некоторых аспектах в этом изобретении предложены способы лечения и/или предотвращения боли, вызванной ревматоидным артритом, у индивидов, в том числе млекопитающих, как людей, так и субъектов, не являющихся людьми. Соответственно в одном из аспектов в этом изобретении предложены способы лечения боли, вызванной ревматоидным артритом, у индивида, включающие введение эффективного количества антагонистического антитела против NGF. Антагонистические антитела против NGF известны в данной области и описаны здесь.
В другом аспекте в этом изобретении предложены способы снижения частоты, облегчения, подавления, облегчения проявлений и/или задержки начала, развития или прогрессирования боли, вызванной ревматоидным артритом, у индивида. Таким образом, в некоторых воплощениях антагонистическое антитело против NGF вводят до развития боли или эпизода боли у индивида, имеющего ревматоидный артрит.
В другом аспекте в этом изобретении предложены способы лечения воспалительной кахексии (потери массы тела), связанной с ревматоидным артритом, у индивида, включающие введение эффективного количества антагонистического антитела против NGF (Roubenoff et al., Arthritis Rheum. 40(3): 534-9 (1997); Roubenoff et al., J. Clin. Invest. 93(6):2379-86 (1994)).
Диагностика или оценка боли, вызванной ревматоидным артритом, установлены в данной области. Оценку можно проводить на основании измерений, известных в данной области, таких как характеризация боли пациентом при использовании различных шкал боли. См., например, Katz et al., Surg Clin North Am. (1999) 79 (2):231-52; Caraceni et al. J Pain Symptom Manage (2002) 23(3):239-55. Существуют также общепринятые шкалы для измерения болезненного состояния, такие как шкала American College of Rheumatology (ACR) (Felson, et al., Arthritis and Rheumatism (1993) 36(6):729-740), шкала Health Assesion Questionnaire (HAQ) (Fries, et al., (1982) J. Rheumatol. 9: 789-793), шкала Паулюса (Paulus, et al., Arthritis and Rheumatism (1990) 33: 477-484) и шкала Arthritis Impact Measure Scale (AIMS) (Meenam, et al., Arthritis and Rheumatology (1982) 25: 1048-1053). Антагонистическое антитело против NGF можно вводить индивиду любым подходящим путем. Здесь описаны примеры разных путей введения.
Облегчение боли можно охарактеризовать периодом облегчения. Соответственно в некоторых воплощениях облегчение боли наблюдают в пределах около 24 часов после введения антагонистического антитела против NGF. В других воплощениях облегчение боли наблюдают в пределах около 36, 48, 60, 72 часов или 4 суток после введения антагонистического антитела против NGF. В еще других воплощениях облегчение боли наблюдают перед тем, как наблюдают признаки улучшения воспалительного состояния, связанного с ревматоидным артритом. В некоторых воплощениях частота и/или интенсивность боли уменьшается, и/или качество жизни страдающего данным заболеванием повышается.
Получение и использование антагонистов NGF (в том числе антител против NGF) для этих способов раскрыты в разделах ниже ("Антагонисты NGF"; "Антагонистическое антитело против NGF"; "Другие антагонисты NGF"; "Идентификация антагонистов NGF (таких как антагонистические антитела против NGF)"; "Композиции для применения в способах по этому изобретению"; "Введение антагониста NGF (такого как антагонистическое антитело против NGF)").
СПОСОБЫ ИСПОЛЬЗОВАНИЯ АНТАГОНИСТИЧЕСКОГО АНТИТЕЛА ПРОТИВ NGF ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ
В некоторых аспектах в этом изобретении предложены способы лечения и/или предотвращения боли, вызванной остеоартритом, у индивидов, в том числе млекопитающих, как людей, так и субъектов, не являющихся людьми. Соответственно в одном из аспектов в этом изобретении предложены способы лечения боли, вызванной оетеоартритом, у индивида, включающие введение эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF). Антагонисты NGF, в том числе антагонистические антитела против NGF, известны в данной области и описаны здесь.
В другом аспекте в этом изобретении предложены способы снижения частоты, облегчения, подавления, облегчения проявлений и/или задержки начала, развития или прогрессирования боли, вызванной остеоартритом, у индивида, включающие введение эффективного количества антагониста NGF (такого как антагонистическое антитело против NGF). Таким образом, в некоторых воплощениях антагонист NGF (такой как антагонистическое антитело против NGF) вводят до развития боли или эпизода боли у индивида, имеющего остеоартрит.
Диагностика или оценка боли, вызванной остеоартритом, установлены в данной области. Оценку можно осуществлять на основании измерений, известных в данной области, таких как характеризация боли пациентом при использовании различных шкал боли. См., например, Katz et al., Surg Clin North Am. (1999) 79 (2):231-52; Caraceni et al. J Pain Symptom Manage (2002) 23(3):239-55. Например, для оценки боли и оценки ответа на лечение можно использовать шкалу WOMAC Ambulation Pain Scale (включающую в себя боль, ригидность и физическую функцию) и 100-мм визуальную аналоговую шкалу (Visual Analogue Scale, VAS).
Антагонисты NGF (такие как антагонистическое антитело против NGF) можно вводить индивиду любым подходящим путем. Здесь описаны примеры разных путей введения.
В некоторых воплощениях антагонист NGF (такой как антагонистическое антитело против NGF) вводят один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в пять недель, один раз в шесть недель, один раз в семь недель, один раз в восемь недель, один раз в девять недель, один раз в десять недель, один раз в пятнадцать недель, один раз в двадцать недель, один раз в двадцать пять недель или один раз в двадцать шесть недель. В некоторых воплощениях антагонист NGF (такой как антагонистическое антитело против NGF) вводят один раз в месяц, один раз в два месяца, один раз в три месяца, один раз в четыре месяца, один раз в пять месяцев или один раз в шесть месяцев.
Облегчение боли можно охарактеризовать периодом облегчения. Соответственно в некоторых воплощениях облегчение боли наблюдают в пределах около 24 часов после введения антагониста NGF (такого как антагонистическое антитело против NGF). В других воплощениях облегчение боли наблюдают в пределах около 36, 48, 60, 72 часов или 4 суток после введения антагониста NGF (такого как антагонистическое антитело против NGF). В некоторых воплощениях частота и/или интенсивность боли уменьшается и/или качество жизни страдающего заболеванием повышается. В некоторых воплощениях облегчение боли, вызванной остеоартритом, обеспечено в течение по меньшей мере около 7 суток, по меньшей мере около 14 суток, по меньшей мере около 21 суток, по меньшей мере около 28 суток, по меньшей мере около 35 суток, по меньшей мере около 42 суток, по меньшей мере около 49 суток, по меньшей мере около 56 суток, по меньшей мере около 63 суток, по меньшей мере около 70 суток, по меньшей мере около 77 суток, по меньшей мере около 84 суток, по меньшей мере около 180 суток или дольше после разовой дозы антагониста NGF (такого как антагонистическое антитело против NGF).
Получение и использование антагонистов NGF (в том числе антител против NGF) для этих способов раскрыты в разделах ниже ("Антагонисты NGF"; "Антагонистическое антитело против NGF"; "Другие антагонисты NGF"; "Идентификация антагонистов NGF (таких как антагонистические антитела против NGF)"; "Композиции для применения в способах по этому изобретению"; "Введение антагониста NGF (такого как антагонистическое антитело против NGF)").
Антагонисты NGF
В способах по этому изобретению (в отношении боли, вызванной ревматоидным артритом, и боли, вызванной остеоартритом) используют антагонист NGF, который относится к любой молекуле, которая блокирует, подавляет или снижает (в том числе в значительной степени) биологическую активность NGF, включая пути опосредованной NGF сигнализации в нисходящем направлении, такие как связывание рецептора и/или индуцирование клеточного ответа на NGF. Термин "антагонист" не подразумевает какого-либо конкретного механизма биологического действия и ясно предназначен включать в себя и охватывать все возможные фармакологические, физиологические и биохимические взаимодействия с NGF и их последствия, которые могут быть достигнуты множеством разных и химически разнообразных композиций. Примеры антагонистов NGF включают в себя, без ограничения ими, антитело против NGF, антисмысловую молекулу, направленную на NGF (включая антисмысловую молекулу, направленную на нуклеиновую кислоту, кодирующую NGF), антисмысловую молекулу, направленную на рецептор NGF (такой как TrkA-рецептор и/или р75-рецептор) (включая антисмысловую молекулу, направленную на нуклеиновую кислоту, кодирующую TrkA и/или р75), соединение, ингибирующее NGF, структурный аналог NGF, доминантно-негативную мутацию TrkA-рецептора, которая связывает NGF, иммуноадгезин TrkA, антитело против TrkA, доминантно-негативную мутацию р75-рецептора, которая связывает NGF, антитело против р75 и ингибитор киназы. Для цели настоящего изобретения следует ясно понимать, что термин "антагонист" охватывает все ранее идентифицированные термины, названия и функциональные состояния и характеристики, которыми сам NGF, биологическая активность NGF (включая, без ограничения, его способность опосредовать любой вид боли) или следствия биологической активности по существу аннулируются, снижаются или нейтрализуются в любой значимой степени. В некоторых воплощениях антагонист NGF (например, антитело) связывается (физически взаимодействует) с NGF, связывается с рецептором NGF (таким как TrkA-рецептор и/или р75-рецептор) и/или снижает (затрудняет и/или блокирует) сигнализацию от рецептора NGF в нисходящем направлении. Соответственно в некоторых воплощениях антагонист NGF связывается (физически взаимодействует) с NGF. В некоторых воплощениях антагонист NGF представляет собой полипептид, который связывается с NGF. В некоторых воплощениях антагонист NGF представляет собой пептид или модифицированный пептид (такой как связывающий NGF пептид, слитый с Fc-доменом), описанный в РСТ WO 2004/026329. В других воплощениях антагонист NGF связывается с рецептором NGF (таким как TrkA- или р75-рецептор). В других воплощениях антагонист NGF снижает (затрудняет и/или блокирует) сигнализацию от рецептора NGF в нисходящем направлении (например, ингибиторы киназной сигнализации). В других воплощениях антагонист NGF ингибирует (снижает) синтез и/или высвобождение NGF. В еще одном воплощении антагонист NGF является антагонистом NGF, который не представляет собой иммуноадгезин TrkA (то есть является отличным от иммуноадгезина TrkA). В еще одном воплощении антагонист NGF является отличным от антитела против NGF. В другом воплощении антагонист NGF является отличным от иммуноадгезина TrkA и отличным от антитела против NGF. В некотором воплощении антагонист NGF связывается с NGF (таким как человеческий NGF) и незначительно связывается с родственными нейротрофинами, такими как NT-3, NT4/5 и/или BDNF. В некоторых воплощениях антагонист NGF не ассоциирован с нежелательным иммунным ответом. В других воплощениях антагонист NGF представляет собой антитело против NGF. В еще других воплощениях антитело против NGF является гуманизированным (таким как описанное здесь антитело Е3). В некоторых воплощениях антитело против NGF представляет собой антитело Е3 (как описано здесь). В других воплощениях антитело против NGF содержит один или более чем один CDR антитела Е3 (например, один, два, три, четыре, пять или в некоторых воплощениях все шесть CDR Е3). В других воплощениях это антитело является человеческим. В еще других воплощениях антитело против NGF содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанной на Фиг.1А (SEQ ID NO:1), и аминокислотную последовательность вариабельной области легкой цепи, показанной на Фиг.1Б (SEQ ID NO:2). В еще других воплощениях антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной, например не вызывает опосредованный комплементом лизис или не стимулирует зависимую от антител опосредованную клетками цитотоксичность (ADCC). В других воплощениях константная область модифицирована, как описано в Eur. J. Immunol. (1999) 29:2613-2624; заявка РСТ PCT/GB99/01441 и/или заявка на патент UK №9809951.8.
Антагонистическое антитело против NGF
В способах по этому изобретению (в отношении боли, вызванной ревматоидным артритом, и боли, вызванной остеоартритом) используют антагонистическое антитело против NGF, которое относится к любой молекуле антитела, которая блокирует, подавляет или снижает (в том числе в значительной степени) биологическую активность NGF, включая пути в нисходящем направлении, опосредованные сигнализацией от NGF, такие как связывание рецептора и/или индуцирование клеточного ответа на NGF.
Антагонистическое антитело против NGF должно проявлять любую одну или более чем одну следующую характеристику: (а) связываться с NGF и ингибировать биологическую активность NGF или пути в нисходящем направлении, опосредованные сигнальной функцией NGF; (б) предотвращать, облегчать или лечить любой вид боли, вызванной ревматоидным артритом, или боли, вызванной остеоартритом; (в) блокировать или снижать активацию рецептора NGF (включая димеризацию и/или аутофосфорилирование TrkA-рецептора); (г) увеличивать клиренс NGF; (д) ингибировать (снижать) синтез, продукцию или высвобождение NGF. Антагонистические антитела против NGF известны в данной области, см., например, публикации РСТ WO 01/78698, WO 01/64247, патенты США №№5844092, 5877016 и 6153189; Hongo et al., Hybridoma, 19:215-227 (2000); Cell. Molec. Biol. 13:559-568 (1993); GenBank Accession №№ U39608, U39609, L17078 или L17077. Антагонистические антитела и полипептиды против NGF также описаны в PCT WO 2005/019266.
Для целей этого изобретения антитело взаимодействует с NGF таким образом, что ингибирует NGF и/или пути в нисходящем направлении, опосредованные сигнальной функцией NGF. В некоторых воплощениях антагонистическое антитело против NGF распознает человеческий NGF. В еще других воплощениях антагонистическое антитело против NGF специфично связывается с человеческим NGF. В некотором воплощении антагонистическое антитело против NGF незначительно связывается с родственными нейротрофинами, такими как NT-3, NT4/5 и/или BDNF. В еще других воплощениях антитело против NGF способно связывать NGF и эффективно ингибировать связывание NGF с его TrkA- и/или р75-рецептором in vivo и/или эффективно ингибировать NGF в смысле активации TrkA- и/или р75-рецептора им. В еще других воплощениях антагонистическое антитело против NGF представляет собой моноклональное антитело. В еще других воплощениях антитело против NGF является гуманизированным (таким как описанное здесь антитело Е3). В некоторых воплощениях антитело против NGF является человеческим. См., например, WO 2005/019266. В одном из воплощений антитело является человеческим антителом, которое распознает один или более чем один эпитоп на человеческом NGF. В еще одном воплощении антитело является мышиным или крысиным антителом, которое распознает один или более чем один эпитоп на человеческом NGF. В еще одном воплощении антитело распознает один или более чем один эпитоп на NGF, выбранном из группы, состоящей из антител приматов, собак, кошек, лошадей и коров. В еще дополнительных воплощениях антагонистическое антитело против NGF связывается по существу с тем же эпитопом 6 NGF, что и антитело, выбранное из любого одного или более чем одного из следующих: МАb 911, МАb 912 и МАb 938 (см. Hongo, et al., Hybridoma 19:215-227 (2000)). В других воплощениях антитело связывается с тем же эпитопом, что и моноклональное антитело 911. В еще одном воплощении антитело содержит константную область, которая является иммунологически инертной (например, не вызывает опосредованный комплементом лизис или зависимую от антител опосредованную клетками цитотоксичность (ADCC)). ADCC-активность можно оценивать при использовании способов, раскрытых в патенте США №5500362. В некоторых воплощениях константная область модифицирована, как описано в Eur. J. Immunol. (1999) 29:2613-2624; в заявке РСТ PCT/GB99/01441 и/или в заявке на патент UK №9809951.8.
В некоторых воплощениях антагонистическое антитело против NGF является гуманизированным мышиным моноклональным антителом против NGF, обозначенным как антитело "Е3", любым из антител, родственных Е3, которые описаны здесь, или любыми их фрагментами, которые являются антагонистами NGF.
Антитела, полезные в настоящем изобретении, могут охватывать моноклональные антитела, поликлональные антитела, фрагменты антитела (например, Fab, Fab', F(ab')2, Fv, Fc и так далее), химерные антитела, биспецифичные антитела, гетероконъюгатные антитела, отдельную цепь (ScFv), их мутанты, слитые белки, содержащие часть антитела, гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена требуемой специфичности, в том числе гликозилированные варианты антител, варианты аминокислотных последовательностей антител и ковалентно модифицированные антитела. Антитела могут быть мышиными, крысиными, человеческими или любого другого происхождения (включая химерные или гуманизированные антитела).
Аффинность связывания антагонистического антитела против NGF с NGF (таким как человеческий NGF) может составлять от приблизительно 0,10 до приблизительно 0,80 нМ, от приблизительно 0,15 до приблизительно 0,75 нМ и от приблизительно 0,18 до приблизительно 0,72 нМ. В одном из воплощений аффинность связывания находится между приблизительно 2 пМ и 22 пМ. В одном из воплощений аффинность связывания находится между приблизительно 23 пМ и приблизительно 100 пМ. В некотором воплощении аффинность связывания приблизительно равна 10 нМ. В других воплощениях аффинность связывания составляет менее чем приблизительно 10 нМ. В других воплощениях аффинность связывания равна приблизительно 0,1 нМ или приблизительно 0,07 нМ. В других воплощениях аффинность связывания менее чем приблизительно 0,1 нМ или менее чем приблизительно 0,07 нМ. В других воплощениях аффинность связывания представляет собой любое из значений около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ или от приблизительно 50 пМ до любого из значений около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ или около 40 пМ. В некоторых воплощениях аффинность связывания представляет собой любое из значений около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ, около 50 пМ или менее чем около 50 пМ. В некоторых воплощениях аффинность связывания менее чем любое из значений около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ или около 50 пМ. В еще других воплощениях аффинность связывания представляет собой значение около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ, около 40 пМ или более чем около 40 пМ.
Один из способов определения аффинности связывания антитела с NGF представляет собой измерение аффинности связывания монофункциональных Fab-фрагментов антитела. Для получения монофункциональных Fab-фрагментов антитело (например, IgG) можно расщеплять папаином или экспрессировать рекомбинантным способом. Аффинность Fab-фрагмента антитела против NGF можно определять поверхностным плазменным резонансом (система для поверхностного плазменного резонанса (ППР) BIAcore3000™, BIAcore, INC, Piscaway NJ). СМ5-чипы можно активировать гидрохлоридом N-этил-N+-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Человеческий NGF (или любой другой NGF) можно разбавлять в 10 мМ ацетате натрия при рН 4,0 и инъецировать на активированный чип при концентрации 0,005 мг/мл. При использовании варьирующего времени потока через индивидуальные каналы чипа можно достичь двух диапазонов плотности антигена: 100-200 единиц отклика (ЕО) для детального кинетического исследования и 500-600 ЕО для скрининговых анализов. Чип можно блокировать этаноламином. Исследования регенерации показали, что смесь пирсовского буфера для элюции (продукт №21004, Pierce Biotechnology, Rockford IL) и 4 M NaCl (2:1) эффективно удаляет связанный Fab, при этом поддерживая активность человеческого NGF на чипе для более чем 200 инъекций. В качестве рабочего буфера для анализов BIAcore используют буфер HBS-EP (0,01 M HEPES, рН 7,4, 0,15 NaCl, 3 мМ ЭДТА, 0,005% поверхностно-активного вещества Р29). Серийные разведения (0,1-10× по оценке KD) очищенных проб Fab инъецируют в течение 1 мин при 100 мкл/мин и дают до 2 ч времени для диссоциации. Концентрации Fab-белков определяют в ELISA и/или электрофорезом в полиакриламидном геле с додецилсульфатом натрия при использовании известной концентрации Fab (определенной анализом аминокислот) в качестве стандарта. Кинетические скорости ассоциации (kon) и скорости диссоциации (koff) получают одновременно, аппроксимируя данные один к одному моделью связывания по Лэнгмюру (Karlsson, R. Roos, H. Fagerstam, L. Petersson, В. (1994). Methods Enzymology 6. 99-110) при использовании программы BIAevaluation. Величины равновесной константы диссоциации (KD) вычисляют как koff/kon. Этот протокол является подходящим для применения в определении аффинности связывания антител с любыми NGF, в том числе человеческим NGF, NGF других позвоночных (в некоторых воплощениях млекопитающего) (такими как мышиный NGF, крысиный NGF, NGF приматов), а также для применения с другими нейротрофинами, такими как родственные нейротрофины NT3, NT4/5 и/или BDNF.
В некоторых воплощениях антитело связывается с человеческим NGF и незначительно связывается с NGF из других видов позвоночных (в некоторых воплощениях млекопитающего). В некоторых воплощениях антитело связывается с человеческим NGF, а также с одним или более чем одним NGF из других видов позвоночных (в некоторых воплощениях млекопитающего). В еще других воплощениях антитело связывается с NGF и не способно к значительному перекрестному взаимодействию с другими нейротрофинами (такими как родственные нейротрофины, NT3, NT4/5 и/или BDNF). В некоторых воплощениях антитело связывается с NGF, а также по меньшей мере с одним другим нейротрофином. В некоторых воплощениях антитело связывается с NGF таких видов млекопитающих, как лошадь или собака, но незначительно связывается с NGF из других видов млекопитающих.
Эпитоп(ы) может (могут) быть непрерывным(и) или прерывистым(и). В одном из воплощений антитело связывается по существу с теми же эпитопами человеческого NGF, что и антитело, выбранное из группы, состоящей из МАb 911, МАb 912 и МАb 938, как описано в Hongo et al., Hybridoma, 19:215-227 (2000). В еще одном воплощении антитело связывается по существу с тем же эпитопом человеческого NGF, что и МАb 911. В еще одном воплощении антитело связывается по существу с тем же эпитопом, что и МАb 909. Hongo et al., см. выше. Например, эпитоп может содержать одно или более чем одно из следующего: остатки K32, K34 и Е35 в пределах вариабельной области 1 (аминокислоты 23-35) человеческого NGF; остатки F79 и Т81 в пределах вариабельной области 4 (аминокислоты 81-88) человеческого NGF; остатки Н84 и K88 в пределах вариабельной области 4; остаток R103 между вариабельной областью 5 (аминокислоты 94-98) человеческого NGF и С-концом (аминокислоты 111-118) человеческого NGF; остаток Е11 в пределах до вариабельной области 1 (аминокислоты 10-23) человеческого NGF; Y52 между вариабельной областью 2 (аминокислоты 40-49) человеческого NGF и вариабельной областью 3 (аминокислоты 59-66) человеческого NGF; остатки L112 и S113 в пределах С-конца человеческого NGF; остатки R59 и R69 в пределах вариабельной области 3 человеческого NGF; или остатки V18, V20 и G23 в пределах до вариабельной области 1 человеческого NGF. В дополнение, эпитоп может содержать одну или более чем одну вариабельную область 1, вариабельную область 3, вариабельную область 4, вариабельную область 5, N-концевую область и/или С-конец человеческого NGF. В еще одном воплощении антитело значительно снижает доступность растворителя для остатка R103 человеческого NGF. Понятно, что хотя эпитопы, описанные выше, относятся к человеческому NGF, обычный специалист может выровнять структуры человеческого NGF с NGF других видов и идентифицировать вероятные соответствия этих эпитопов.
В одном из аспектов можно получать антитела, которые могут ингибировать NGF (например, человеческие, гуманизированные, мышиные, химерные), при использовании иммуногенов, которые экспрессируют полноразмерную или частичную последовательность NGF. В другом аспекте можно использовать содержащую иммуноген клетку, которая усиленно экспрессирует NGF. Другим примером иммуногена, который можно использовать, является белок NGF, который содержит полноразмерный NGF или часть белка NGF.
Антагонистические антитела против NGF можно получать любым способом, известным в данной области. Путь и режим иммунизации животного-хозяина обычно соответствует установленным и общепринятым методикам для стимуляции и продукции антител, как описано здесь далее. Общие методики получения человеческих и мышиных антител известны в данной области и раскрыты здесь.
Предусмотрено, что любым субъектом, представляющим собой млекопитающее, включая людей или продуцирующие антитело клетки из них, можно манипулировать так, чтобы он представлял собой основу для получения гибридомных клеточных линий млекопитающего, включая человека. В типичных случаях животное-хозяин инокулируют внутрибрюшинным путем, внутримышечным путем, пероральным путем, подкожным путем, в подошву и/или в кожу некоторым количеством иммуногена, включая такой, какой описан здесь.
Гибридомы можно получать из лимфоцитов и иммортализированных клеток миеломы при использовании общей методики гибридизации соматических клеток по Kohler, В. and Milstein, С. (1975) Nature 256:495-497, или как модифицировано в Buck, D. W., et al., In vitro, 18:377-381 (1982). В гибридизации можно использовать доступные линии миеломы, в том числе, без ограничения ими, Х63-Ag8.653 и из Salk Institute, Cell Distribution Center, San Diego, Calif., USA. Обычно эта методика включает в себя слияние миеломных клеток и лимфоидных клеток при использовании фузогена, такого как полиэтиленгликоль, или электрических способов, хорошо известных специалистам. После слияния отделяют клетки от среды слияния и выращивают в селективной ростовой среде, такой как среда с гипоксантином-аминоптерином-тимидином (HAT) для элиминации негибридизованных исходных клеток. Для культивирования гибридом, которые секретируют моноклональные антитела, можно использовать описанную здесь любую среду с добавлением или без добавления сыворотки. В качестве другой альтернативы этой методике слияния клеток можно использовать В-клетки, иммортализованные вирусом Эпштейна-Барр (EBV), с получением моноклональных антител против NGF, представляющих собой объект изобретения. Гибридомы размножают и субклонируют, если желательно, и супернатанты анализируют на антииммуногенную активность общепринятыми способами иммуноанализа (например, радиоиммуноанализом, иммуноферментным анализом или флуоресцентным иммуноанализом).
Гибридомы, которые можно использовать в качестве источника антител, охватывают все производные и потомство клеток исходных гибридом, которые продуцируют моноклональные антитела, специфичные по отношению к NGF, или их часть.
Гибридомы, которые продуцируют такие антитела, можно выращивать in vitro или in vivo при использовании известных способов. Моноклональные антитела можно выделять из культуральных сред или жидкостей организма общепринятыми способами очистки иммуноглобулинов, такими как осаждение сульфатом аммония, гель-электрофорез, диализ, хроматография и ультрафильтрация, если желательно. Нежелательную активность при ее наличии можно удалять, например, пропусканием препарата через сорбенты с получением иммуногена, присоединенного к твердой фазе, и элюированием или освобождением желаемого антитела от иммуногена. Популяцию антител (например, моноклональных антител) может давать иммунизация животного-хозяина человеческим NGF или фрагментом, содержащим целевую аминокислотную последовательность, который конъюгирован с белком, который является иммуногенным в видах, которые иммунизируют, например, с гемоцианином фиссуреллы (keyhole limpet), с сывороточным альбумином, с бычьим тиреоглобулином или с соевым ингибитором трипсина, при использовании бифункционального или дериватизирующего агента, например сложного эфира малеимидобензоилсульфосукцинимида (конъюгация через остатки цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 представляют собой разные алкильные группы.
Если желательно, антагонистическое антитело против NGF (моноклональное или поликлональное), представляющее интерес, можно подвергнуть определению последовательности и полинуклеотидную последовательность можно затем клонировать в векторе для экспрессии или размножения. Последовательность, кодирующую антитело, представляющее интерес, можно поддерживать в векторе в клетке-хозяине и клетку-хозяина можно затем размножить и заморозить для будущего использования. Альтернативно, полинуклеотидную последовательность можно использовать для генетической манипуляции с целью "гуманизации" антитела или улучшения аффинности или других характеристик антитела. Например, константную область можно сконструировать так, чтобы она более соответствовала человеческим константным областям во избежание иммунного ответа, если антитело используют в клинических испытаниях и средствах лечения у людей. Может быть желательно генетически манипулировать последовательностью антитела с получением большей аффинности по отношению к NGF и большей эффективности в ингибировании NGF. Специалисту в данной области должно быть очевидно, что одно или более чем одно изменение полинуклеотида можно делать в антагонистическом антителе против NGF и все еще сохранить его способность связывания с NGF.
Существует четыре общих стадии гуманизации моноклонального антитела. Они представляют собой: (1) определение нуклеотидной и предсказание аминокислотной последовательности легкого и тяжелого вариабельных доменов исходного антитела, (2) конструирование гуманизированного антитела, то есть решение того, какую каркасную область антитела использовать в процессе гуманизации; (3) реальные способы/методики гуманизации и (4) трансфекция и экспрессия гуманизированного антитела. См., например, патенты США №№4816567, 5807715, 5866692, 6331415, 5530101, 5693761, 5693762, 5585089 и 6180370.
Описано множество молекул "гуманизированных" антител, содержащих сайт связывания антигена, происходящий из иммуноглобулина, не являющегося человеческим, в том числе химерные антитела, имеющие V-области или модифицированные V-области грызунов и ассоциированные с ними гипервариабельные участки (CDR), слитые с человеческими константными доменами. См., например, Winter et al. Nature 349:293-299 (1991), Lobuglio et al. Proc. Nat. Acad. Sci. USA 86:4220-4224 (1989), Shaw et al. J Immunol. 138:4534-4538 (1987) и Brown et al. Cancer Res. 47:3577-3583 (1987). Другие ссылки описывают CDR грызунов, трансплантированные в человеческую поддерживающую каркасную область (FR) перед слиянием с соответствующим константным доменом человеческого антитела. См., например, Riechmann et al. Nature 332:323-327 (1988), Verhoeyen et al. Science 239:1534-1536 (1988) и Jones et al. Nature 321:522-525 (1986). Другая ссылка описывает CDR грызуна, поддерживаемый рекомбинантными маскированными каркасными областями грызуна. См., например, публикацию Европейского патента №0519596. Эти "гуманизированные" молекулы предназначены для минимизации нежелательного иммунологического ответа на молекулы антител грызуна против антигенов человека, который ограничивает длительность и эффективность терапевтических применений этих молекул у реципиентов, представляющих собой людей. Например, константную область антитела можно сконструировать так, что оно является иммунологически инертным (например, не вызывает комплементзависимый лизис). См., например, публикацию РСТ PCT/GB99/01441; Заявку на патент UK №9809951.8. Другие способы гуманизации антител, которые также можно использовать, раскрыты в Daugherty et al., Nucl. Acid Res. 19:2471-2476 (1991), в патентах США №№6180377; 6054297; 5997867; 5866692; 6210671 и 6350861 и в публикации РСТ WO № 01/27160.
В еще одной альтернативе полностью человеческие антитела можно получать при использовании имеющихся в продаже мышей, которые были сконструированы для экспрессии специфических белков иммуноглобулинов человека. Трансгенных животных, которые предназначены для получения более желательного (например, полностью человеческие антитела) или более устойчивого иммунного ответа, также можно использовать для получения гуманизированных или человеческих антител. Примерами такой технологии являются Xenomouse™ от Abgenix, Inc. (Fremont, СА) и HuMAb-Mouse® и ТС Mouse™ от Medarex, Inc. (Princeton, NJ).
Альтернативно, антитела можно получать рекомбинантным способом и экспрессировать при использовании любого способа, известного в данной области. В другой альтернативе антитела можно получать рекомбинантным способом технологий фагового дисплея. См., например, патенты США №№5565332, 5580717, 5733743 и 6265150 и Winter et al., Annu. Rev. Immunol. 12:433-455 (1994). Альтернативно, технологию фагового дисплея (McCafferty et al., Nature 348:552-553 (1990)) можно использовать для получения человеческих антител и фрагментов антитела in vitro из совокупности генов вариабельного (V) домена иммуноглобулина из неиммунизированных доноров. В соответствии с этой методикой гены V-доменов антител клонируют в рамке гена либо главного, либо минорного белка оболочки нитчатого бактериофага, такого как М13 или fd, и они проявляются как функциональные фрагменты антитела на поверхности фаговой частицы. Поскольку нитчатая частица содержит копию одноцепочечной ДНК фагового генома, селекция на основании функциональных свойств антитела также приводит к селекции гена, кодирующего антитело, которое проявляет эти свойства. Таким образом, фаг имитирует некоторые свойства В-клетки. Фаговый дисплей можно осуществлять во многих разных форматах; обзор см., например, в Johnson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3:564-571 (1993). Для фагового дисплея можно использовать несколько источников сегментов V-генов. Clackson et al., Nature 352:624-628 (1991) выделили широкий диапазон антител против оксазолона из небольшой случайной комбинаторной библиотеки V-генов, происходящих из селезенок иммунизированных мышей. Можно сконструировать совокупность V-генов из неиммунизированных доноров, представляющих собой людей, и выделить антитела против широкого диапазона антигенов (включая аутоантигены) в сущности по методикам, описанным в Mark et al., J. Mol. Вiоl. 222:581-597 (1991), или Griffith et al., EMBO J. 12:725-734 (1993). В естественном иммунном ответе гены антител аккумулируют мутации с высокой скоростью (соматическое гипермутирование). Некоторые внесенные изменения должны придавать более высокую аффинность, и В-клетки, проявляющие поверхностный иммуноглобулин, имеющий высокуюю аффинность, предпочтительно реплицируются и дифференцируются при последующей стимуляции антигеном. Этот естественный процесс можно имитировать использованием методики, известной как "тасовка цепей". Marks, et al., Bio/Technol. 10:779-783 (1992)). В этом способе аффинность "первичных" человеческих антител, полученных фаговым дисплеем, можно улучшить последовательной заменой генов V-областей тяжелой и легкой цепи совокупностьями встречающихся в природе вариантов (наборов) генов V-домена, полученных из неиммунизированных доноров. Это методика делает возможным получение антител и фрагментов антител с аффинностью в диапазоне от пМ до нМ. Стратегия получения очень больших популяций фаговых антител (также известно как "прототип всех библиотек") описана в Waterhouse et al., Nucl. Acid Res. 21:2265-2266 (1993). Тасовку генов также можно использовать для получения человеческих антител из антител грызунов, причем человеческое антитело имеет такую же аффинность и специфичность, как у исходного антитела грызуна. В соответствии с этим способом, который также обозначают как "импринтинг эпитопов", ген V-домена тяжелой или легкой цепи антитела грызуна, полученный методикой фагового дисплея, заменяют совокупностью человеческих генов V-доменов, что создает химеры грызун-человек. Селекция на антигене приводит к выделению человеческих вариабельных областей, способных восстанавливать функциональный сайт связывания антигена, то есть эпитоп определяет (делает импринтинг) выбор партнера. Когда этот процесс повторяют для замены оставшегося V-домена грызуна, получают человеческое антитело (см. публикацию РСТ WO № 93/06213, опубликована 1 апреля 1993). В отличие от традиционной гуманизации антител грызуна трансплантацией CDR эта методика дает полностью человеческие антитела, которые не имеют остатков каркасной области или CDR, происходящих от грызуна.
Очевидно, что хотя вышерассмотренное относится к гуманизированным антителам, общие рассмотренные принципы применимы к приспособлению антител для применения, например, у собак, кошек, приматов, лошадей и коров. Далее очевидно, что один или более чем один описанный здесь аспект гуманизации антитела можно объединять, например трансплантацию CDR, мутацию каркасной области и мутацию CDR.
Антитела можно получать рекомбинантным способом, сначала выделением антител и клеток, продуцирующих антитело, из животных-хозяев, получением последовательности гена и использованием последовательности гена для рекомбинантной экспрессии антитела в клетках-хозяевах (например, в клетках СНО). Другой способ, который можно использовать, представляет собой экспрессию последовательности антитела в растениях (например, в табаке) или в трансгенном молоке. Способы рекомбинантной экспрессии антител в растениях или молоке были раскрыты. См., например, Peeters, et al. Vacine 19:2756 (2001); Lonberg, N. and D. Huszar Int. Rev. Immunol 13:65 (1995); и Pollock, et al., J Immunol Methods 231:147(1999). Способы получения производных антител, например гуманизированных, одноцепочечных и так далее, известны в данной области.
Для выделения антител, которые являются специфичными по отношению к NGF, также можно использовать методики сортировки иммуноанализами и проточной цитометрией, такие как сортировка клеток при активации флуоресценцией (FACS).
Антитела можно связывать со многими разными носителями. Носители могут быть активными и/или инертными. Примеры хорошо известных носителей включают в себя полипропилен, полистирол, полиэтилен, декстран, нейлон, амилазы, стекло, натуральные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Для цели этого изобретения носитель может быть по природе либо растворимым, либо нерастворимым. Специалисты должны знать о других подходящих носителях для связывания антител или должны быть способны подтвердить, что они являются таковыми, используя обычное экспериментирование. В некоторых воплощениях носитель содержит группировку, которая нацелена на миокард.
ДНК, кодирующие моноклональные антитела, легко выделить и подвергнуть определению последовательности при использовании общепринятых способов (например, при использовании олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи моноклонального антитела). В качестве предпочтительного источника таких ДНК служат гибридомные клетки. ДНК после выделения можно помещать в экспрессионные векторы (такие как экспрессионные векторы, раскрытые в публикации РСТ WO № 87/04462), которыми затем трансфицируют клетки-хозяева, такие как клетки Е.coli, клетки COS мартышек, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые в противном случае не продуцируют белок иммуноглобулин, с достижением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. См., например, публикацию РСТ WO № 87/04462. ДНК также можно модифицировать, например, последовательностями, кодирующими константные домены человеческой тяжелой и легкой цепи вместо гомологичных мышиных последовательностей, Morrison et al., Proc. Nat. Acad. Sci. 81:6851 (1984), или ковалентным присоединением всей или части последовательности, кодирующей неиммуноглобулиновый полипептид, к последовательности, кодирующей иммуноглобулин. Таким образом получают "химерные" или "гибридные" антитела, которые имеют специфичность связывания, свойственную данному моноклональному антителу против NGF.
Антагонистические антитела против NGF можно охарактеризовать при использовании способов, хорошо известных в данной области. Например, один из способов представляет собой идентификацию эпитопа, с которым оно связывается, или "эпитопное картирование". Существует много способов, известных в данной области, для картирования и характеризации местоположения эпитопов на белках, в том числе нахождение кристаллической структуры комплекса антитела с антигеном, конкурентные анализы, анализы экспрессии фрагментов гена и анализы на основе синтетических пептидов, как описано, например, в Chapter 11, Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В дополнительном примере картирование эпитопа можно использовать для определения последовательности, с которой связывается антагонистическое антитело против NGF. Картирование эпитопов доступно из различных источников, например Pepscan Systems (Edelhertweg 15, 8219 PH Leiystad, Нидерланды). Эпитоп может быть линейным эпитопом, то есть содержаться в единой последовательности аминокислот, или конформационным эпитопом, образованным трехмерным взаимодействием аминокислот, которые не обязательно могут содержаться в единой последовательности. Пептиды вариабельных длин (например, длиной по меньшей мере 4-6 аминокислот) можно выделить или синтезировать (например, рекомбинантным способом) и использовать для анализов связывания с антагонистическим антителом против NGF. В другом примере эпитоп, с которым связывается антагонистическое антитело против NGF, можно определять в систематическом скрининге использованием перекрывающихся пептидов, производных от последовательности NGF, и определением связывания антагонистического антитела против NGF. В соответствии с анализами экспрессии фрагментов гена фрагментируют открытую рамку считывания, кодирующую NGF, либо случайным образом, либо специфическими генными конструктами и определяют взаимодействие экспрессирующихся фрагментов NGF с тестируемым антителом. Фрагменты гена можно, например, получать при использовании ПЦР и затем транскрибировать и транслировать в белок in vitro в присутствии радиоактивных аминокислот. Связывание антитела с меченными радиоактивными изотопами фрагментами NGF затем определяют иммунопреципитацией и гель-электрофорезом. Определенные эпитопы можно также идентифицировать при использовании больших библиотек случайных пептидных последовательностей, проявляющихся на поверхности фаговых частиц (фаговые библиотеки). Альтернативно, определенную библиотеку перекрывающихся пептидных фрагментов можно тестировать по связыванию с тестируемым антителом в простых анализах связывания. В дополнительном примере для идентификации остатков, требуемых, достаточных и/или необходимых для связывания с эпитопом, можно осуществлять мутагенез домена, связывающего антиген, эксперименты по перестановке доменов и мутагенез со сканированием по аланину. Например, эксперименты по перестановке доменов можно осуществлять при использовании мутантного NGF, в котором различные фрагменты полипептида NGF были заменены (переставлены с) последовательностями из близкородственного, но отличающегося по антигенности белка (такого как другой член белкового семейства нейротрофинов). По оценке связывания антитела с мутантным NGF можно оценить важность конкретного фрагмента NGF для связывания с антителом.
Еще один способ, который можно использовать для характеризации антагонистического антитела против NGF, представляет собой использование анализов конкуренции с другими антителами, известными связыванием с тем же антигеном, то есть с различными фрагментами на NGF, для определения того, связывается ли антагонистическое антитело против NGF с тем же эпитопом, что и другие антитела. Конкурентные анализы хорошо известны специалистам в данной области. Пример антител, которые можно использовать в конкурентных анализах для настоящего изобретения, включает в себя МАb 911, 912, 938, как описано в Hongo, et al., Hybridoma 19:215-227 (2000).
Для управления экспрессией антагонистического антитела против NGF можно использовать экспрессионный вектор. Специалисту в данной области известно введение экспрессионных векторов для достижения экспрессии экзогенного белка in vivo. См., например, патенты США №№6436908, 6413942 и 6376471. Введение экспрессионных векторов включает в себя локальное или системное введение, включая инъекцию, пероральное введение, бомбардировку частиц или введение катетером и местное введение. В еще одном воплощении экспрессионный вектор вводят непосредственно в симпатический ствол или ганглий или в коронарную артерию, предсердие, желудочек или перикард.
Также можно использовать направленную доставку терапевтической композиции, содержащей экспрессионный вектор или субгеномные полинуклеотиды. Методики опосредованной рецепторами доставки ДНК раскрыты, например, в Findeis et al., Trends Biotechnol. (1993) 11:202; Chlou et al., Gene Therapeutics: Methods and Applications of Direct Gene Transfer (J.A. Wolff, ed.) (1994); Wu et al., J. Biol. Chem. (1988) 263:621; Wu et al., J. Biol. Chem. (1994) 269:542; Zenke et al., Proc. Natl. Acad. Sci. USA (1990) 87:3655; Wu et al., J. Biol. Chem. (1991) 266:338. В протоколе локального введения при генной терапии терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от приблизительно 100 нг до приблизительно 200 мг ДНК. В протоколе генной терапии также можно использовать диапазоны концентраций от приблизительно 500 нг до приблизительно 50 мг, от приблизительно 1 мкг до приблизительно 2 мг, от приблизительно 5 мкг до приблизительно 500 мкг и от приблизительно 20 мкг до приблизительно 100 мкг ДНК. Терапевтические полинуклеотиды и полипептиды можно доставлять при использовании носителей для доставки генов. Носители для доставки генов могут быть вирусного или невирусного происхождения (см., в общем, Jolly, Cancer Gene Therapy (1994) 1:51; Kimura, Human Gene Therapy (1994) 5:845; Connelly, Human Gene Therapy (1995) 1:185, и Kaplitt, Nature Genetics (1994) 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать при использовании эндогенных промоторов млекопитающего или гетерологичных промоторов. Экспрессия кодирующей последовательности может быть либо конститутивной, либо регулируемой.
Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в данной области. Примеры носителей на основе вирусов включают в себя, без ограничения ими, рекомбинантные ретровирусы (см., например, публикации РСТ WO 90/07936, WO 94/03622, WO 93/25698, WO 93/25234, WO 93/11230, WO 93/10218, WO 91/02805; патенты США №№5219740 и 4777127; патент GB №2200651 и патент ЕР №0345242), векторы на основе альфавирусов (например, векторы, представляющие собой вирус Синдбис, вирус леса Семлики (АТСС VR-67; АТСС VR-1247), вирус реки Росс (АТСС VR-373; АТСС VR-1246) и вирус венесуэльского энцефаломиелита лошадей (АТСС VR-923; АТСС VR-1250; АТСС VR 1249; АТСС VR-532)), и векторы, представляющие собой аденоассоциированный вирус (AAV) (см., например, публикации РСТ WO 94/12649, WO 93/03769, WO 93/19191, WO 94/28938, WO 95/11984 и WO 95/00655). Также можно использовать введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, Hum. Gene Ther. (1992) 3:147.
Также можно использовать невирусные носители и способы для доставки, в том числе, без ограничения ими, только конденсированную поликатионами ДНК, связанную или несвязанную с убитым аденовирусом (см., например, Curiel, Hum. Gene Ther. (1992) 3:147); связанную с лигандом ДНК (см., например, Wu, J. Biol. Chem. (1989) 264:16985); носители, представляющие собой эукариотическую клетку-носитель (см., например, патент США №5814482; публикации РСТ WO 95/07994, WO 96/17072, WO 95/30763 и WO 97/42338), и нейтрализацию заряда ядра или слияние с клеточными мембранами. Также можно использовать голую ДНК. Способы ведения голой ДНК раскрыты в публикации РСТ WO № 90/11092 и патенте США №5580859. Липосомы, которые могут действовать как носители для доставки генов, раскрыты в патенте США №5422120; в публикациях РСТ WO 95/13796, WO 94/23697, WO 91/14445 и в ЕР 0524968. Дополнительные подходы раскрыты в Philip, Mol. Cell Вiоl. (1994) 14:2411, и в Woffendin, Proc. Natl. Acad. Sci. (1994) 91:1581.
Другие антагонисты NGF
Можно использовать антагонисты NGF, отличные от антител против NGF (в отношении боли, вызванной ревматоидным артритом, и боли, вызванной остеоартритом). В некоторых воплощениях этого изобретения антагонист NGF содержит по меньшей мере одну антисмысловую молекулу, способную блокировать или снижать экспрессию функционального NGF. Нуклеотидные последовательности NGF известны и легко доступны из публично доступных баз данных. См., например, Borsani et al., Nuc. Acid Res. 1990, 18, 4020; номер доступа NM 002506; Ullrich et al. Nature 303:821-825 (1983). Обычным является получение молекулы антисмыслового олигонуклеотида, которая должна специфично связываться с мРНК NGF без перекрестного взаимодействия с другими полинуклеотидами. Примеры сайтов-мишеней включают в себя, без ограничения ими, кодон инициирования, 5'-регуляторные области, кодирующую последовательность и 3'-нетранслируемую область. В некоторых воплощениях олигонуклеотиды имеют длину приблизительно от 10 до 100 нуклеотидов, длину приблизительно от 15 до 50 нуклеотидов, длину приблизительно от 18 до 25 нуклеотидов или длину более чем 25 нуклеотидов. Олигонуклеотиды могут содержать модификации каркаса, такие как, например, фосфоротиоатные связи и модификации сахара по 2'-O, хорошо известные в данной области. Примеры антисмысловых молекул включают в себя молекулы, являющиеся антисмысловыми по отношению к NGF, описанные в публикации заявки США №20010046959; см. также http://www.rna-tec.com/repair.htm.
В других воплощениях антагонист NGF содержит по меньшей мере одну антисмысловую молекулу, способную блокировать или снижать экспрессию функционального рецептора NGF (такого как TrkА и/или р75). Woolf et al., J. Neuroscii. (2001) 21(3): 1047-55; Taglialetela et al., J Neurochem (1996) 66(5): 1826-35. Нуклеотидные последовательности TrkA и р75 известны и легко доступны из публично доступных баз данных.
Альтернативно, экспрессию и/или высвобождение NGF и/или экспрессию рецепторов NGF можно снижать при использовании способов, представляющих собой нокдаун генов, морфолиноолигонуклеотиды, РНК-интерференцию или рибозимы, хорошо известных в данной области. См.
http://www.macalester.edu/~montgomery/RNAi.html;
http://pub32.ezboard.com/fmorpholinosfrm19.showMessage?topicID=6.topic;
http://www.highveld.com/ribozyme.html.
В других воплощениях антагонист NGF содержит по меньшей мере одно соединение, ингибирующее NGF. "Соединение, ингибирующее NGF" при использовании здесь относится к соединению, которое является иным, чем антитело против NGF, и которое прямо или опосредованно снижает, ингибирует, нейтрализует или отменяет биологическую активность NGF. Соединение, ингибирующее NGF, должно показывать любую одну или более чем одну из следующих характеристик: (а) связывание с NGF и ингибирование биологической активности NGF и/или путей в нисходящем направлении, опосредованных сигнальной функцией NGF; (б) предотвращение, облегчение или лечение любого вида боли (такой как боль, вызванная остеоартритом); (в) блокирование или снижение активации рецептора NGF (включая димеризацию и/или аутофосфорилирование TrkA-рецептора); (г) увеличение клиренса NGF; (д) ингибирование (снижение) синтеза, продукции или высвобождения NGF. Примеры соединений, ингибирующих NGF, включают в себя низкомолекулярные ингибиторы NGF, описанные в публикации заявки США №20010046959; соединения, которые ингибируют связывание NGF с р75, как описано в публикации РСТ WO № 00/69829, и PD90780 [7-(бензолиламино)-4,9-дигидро-4-метил-9-оксо-пиразоло[5,1-b]хиназолин-2-карбоновую кислоту], как описано в Colquhoun et al., J. Pharmacol. Exp. Ther. 310(2):505-11 (2004); соединения, которые ингибируют связывание NGF с TrkA и/или р75, как описано в публикации РСТ WO № 98/17278. Дополнительные примеры соединений, ингибирующих NGF, включают в себя соединения, описанные в публикациях РСТ WO 02/17914 и WO 02/20479 и в патентах США №№5342942, 6127401 и 6359130. Дополнительные примеры соединений, ингибирующих NGF, представляют собой соединения, которые являются конкурентными ингибиторами NGF. См. патент США №6291247. Более того, специалист может получить другие низкомолекулярные соединения, ингибирующие NGF.
В некоторых воплощениях соединение, ингибирующее NGF, связывается с NGF. Примеры целевых (связываемых) сайтов включают в себя, без ограничения ими, часть NGF, которая связывается с TrkA-рецептором и/или р75-рецептором, и те части NGF, которые примыкают к участку, связывающему рецептор, и определяют частично правильную трехмерную форму участка, связывающего рецептор. В еще одном воплощении соединение, ингибирующее NGF, связывается с рецептором NGF (таким как TrkA и/или р75) и ингибирует биологическую активность NGF. Примеры сайтов для нацеливания включают в себя те части TrkA и/или р75, которые связываются с NGF.
В воплощениях, содержащих малые молекулы, малая молекула может иметь любую молекулярную массу приблизительно от 100 до 20000 дальтон, от 500 до 15000 дальтон или от 1000 до 10000 дальтон. Библиотеки малых молекул являются доступными. Малые молекулы можно вводить при использовании любого способа, известного в данной области, включая ингаляционный, внутрибрюшинный, внутривенный, внутримышечный, подкожный, в мозговую оболочку, в желудочек мозга, пероральным путем, энтеральным путем, парентеральным путем, интраназальным путем или в кожу. В общем, когда антагонист NGF в соответствии с этим изобретением представляет собой малую молекулу, его следует вводить в режиме от 0,1 до 300 мг/кг массы пациента, разделенных на 1-3 или более доз. Для взрослого пациента нормальной массы можно вводить дозы в диапазоне от 1 мг до 5 г на дозу.
В других воплощениях антагонист NGF содержит по меньшей мере один структурный аналог NGF. "Структурные аналоги NGF" в настоящем изобретении относятся к соединениям, которые имеют такую же 3-мерную структуру, как часть таковой у NGF, и которые связываются с рецептором NGF в физиологических условиях in vitro или in vivo, причем это связывание по меньшей мере частично ингибирует биологическую активность NGF. В одном из воплощений структурный аналог NGF связывается с TrkA- и/или р75-рецептором. Примеры структурных аналогов NGF включают в себя, без ограничения ими, бициклические пептиды, описанные в публикации РСТ WO № 97/15593; бициклические пептиды, описанные в патенте США №6291247; циклические соединения, описанные в патенте США №6017878; и пептиды, являющиеся производными NGF, которые описаны в публикации РСТ WO № 89/09225. Подходящие структурные аналоги NGF можно также конструировать и синтезировать при использовании молекулярного моделирования связывания с рецептором NGF, например, способом, описанным в публикации РСТ WO № 98/06048. Структурные аналоги NGF могут быть мономерами или димерами/олигомерами в любой желаемой комбинации одних и тех же или разных структур с получением улучшенной аффинности и улучшенных биологических эффектов.
В других воплощениях в этом изобретении предложен антагонист NGF, содержащий по меньшей мере один доминантно-негативный мутант TrkA-рецептора и/или р75-рецептора. Специалист может получать доминантно-негативные мутанты, например, TrkA-рецептора так, что рецептор должен связываться с NGF и, таким образом, действовать в качестве "ловушки" для захвата NGF. Однако доминантно-негативные мутанты не должны иметь нормальной биоактивности TrkA-рецептора при связывании с NGF. Примеры доминантно-негативных мутантов включают в себя, без ограничения ими, мутанты, описанные в следующих ссылках: Li et al., Proc. Natl. Acad. Sci. USA 1998, 95, 10884; Eide et al., J. Neurosci. 1996, 16, 3123; Liu et al., J. Neurosci 1997, 17, 8749; Klein et al., Cell 1990, 61, 647; Valenzuela et al., Neuron 1993, 10, 963; Tsoulfas et al., Neuron 1993, 10, 975; и Lamballe et al., EMBO J. 1993, 12, 3083, каждая из которых включена сюда ссылкой во всей полноте. Доминантно-негативные мутанты можно вводить в форме белка или в форме экспрессионного вектора так, что доминантно-негативный мутант, например мутант TrkA-рецептора, экспрессируется in vivo. Белок или экспрессионный вектор можно вводить при использовании любого способа, известного в данной области, например внутрибрюшинным путем, внутривенным путем, внутримышечным путем, подкожным путем, в мозговую оболочку, в желудочек мозга, пероральным путем, энтеральным путем, парентеральным путем, интраназальным путем, в кожу или путем ингаляции. Например, введение экспрессионных векторов включает в себя локальное или системное введение, включая инъекцию, пероральное введение, бомбардировку частиц или введение катетером и местное введение. Специалист должен быть знаком с введением экспрессионных векторов для достижения экспрессии экзогенного белка in vivo. См., например, патенты США №№6436908, 6413942 и 6376471.
Также можно использовать направленную доставку терапевтической композиции, содержащей антисмысловой полинуклеотид, экспрессионный вектор или субгеномные полинуклеотиды. Методики опосредованной рецепторами доставки ДНК раскрыты, например, в Findeis et al., Trends Biotechnol. (1993) 11:202; Chlou et al., Gene Therapeutics: Methods and Applications Of Direct Gene Transfer (J.A. Wolff, ed.) (1994); Wu et al., J. Biol. Chem. (1988) 263:621; Wu et al., J. Biol. Chem. (1994) 269:542; Zenke et al., Proc. Natl. Acad. Sci. USA (1990) 87:3655; Wu et al., J. Biol. Chem. (1991) 266:338. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от приблизительно 100 нг до приблизительно 200 мг ДНК для локального введения в протоколе генной терапии. В некоторых воплощениях в протоколе генной терапии можно также использовать диапазоны концентрации от приблизительно 500 нг до приблизительно 50 мг, от приблизительно 1 мкг до приблизительно 2 мг, от приблизительно 5 мкг до приблизительно 500 мкг и от приблизительно 20 мкг до приблизительно 100 мкг или более чем 100 мкг ДНК или терапевтических полинуклеотидов и полипептидов по настоящему изобретению при использовании носителей для доставки гена. Носитель для доставки гена может быть вирусного или невирусного происхождение (см., в общем, Jolly, Cancer Gene Therapy (1994) 1:51; Kimura, Human Gene Therapy (1994) 5:845; Connelly, Human Gene Therapy (1995) 1:185; и Kaplitt, Nature Genetics (1994) 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать при использовании эндогенных для млекопитающего или гетерологичных промоторов и/или энхансеров. Экспрессия кодирующей последовательности может быть либо конститутивной, либо регулируемой.
Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в данной области. Примеры носителей на основе вирусов включают в себя, без ограничения ими, рекомбинантные ретровирусы (см., например, публикации РСТ WO 90/07936, WO 94/03622, WO 93/25698, WO 93/25234, WO 93/11230, WO 93/10218, WO 91/02805; патенты США №№5219740 и 4777127; патент GB №2200651 и патент ЕР №0345242), векторы на основе альфа-вируса (например, векторы, представляющие собой вирус Синдбис, вирус леса Семлики (АТСС VR-67; АТСС VR-1247), вирус реки Росс (АТСС VR-373; АТСС VR-1246) и вирус венесуэльского энцефаломиелита лошадей (АТСС VR-923; АТСС VR-1250; АТСС VR 1249; АТСС VR-532), и векторы, представляющие собой аденоассоциированный вирус (AAV) (см., например, публикации РСТ WO 94/12649, WO 93/03769, WO 93/19191, WO 94/28938, WO 95/11984 и WO 95/00655). Также можно использовать введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, Hum. Gene Ther. (1992) 3:147.
Также можно использовать невирусные носители и способы для доставки, в том числе, без ограничения ими, поликатионную конденсированную ДНК, связанную или несвязанную с убитым аденовирусом в отдельности (см., например, Curiel, Hum. Gene Ther. (1992) 3:147); ДНК, связанную с лигандом (см., например, Wu, J. Biol. Chem. (1989) 264:16985); клетки-носители, представляющие собой эукариотическую клетку (см., например, патент США №5814482; публикации РСТ WO 95/07994, WO 96/17072, WO 95/30763 и WO 97/42338) и нейтрализацию заряда ядра или слияние с клеточными мембранами. Также можно использовать голую ДНК. Примеры способов введения голой ДНК раскрыты в публикации РСТ WO № 90/11092 и патенте США №5580859. Липосомы, которые могут действовать как носители для доставки гена, раскрыты в патенте США №5422120; в публикациях РСТ WO 95/13796, WO 94/23697, WO 91/14445 и в патенте ЕР №0524968. Дополнительные подходы раскрыты в Philip, Mol. Cell Biol. (1994) 14:2411, и в Woffendin, Proc. Natl. Acad. Sci. (1994) 91:1581.
Также ясно, что экспрессионный вектор можно использовать для прямой экспрессии любых описанных здесь антагонистов NGF на основе белка (например, антитела против NGF, иммуноадгезин TrkA и так далее). В данной области известны, например, другие фрагменты TrkA-рецептора, которые способны блокировать (от частичного до полного блокирования) NGF и/или биологическую активность NGF.
В еще одном воплощении антагонист NGF содержит по меньшей мере один иммуноадгезин TrkA. Иммуноадгезины TrkA при использовании здесь относятся к растворимым химерным молекулам, содержащим внеклеточный домен TrkA-рецептора и последовательность иммуноглобулина, которая сохраняет специфичность связывания, свойственную TrkA-рецептору (по существу сохраняет специфичность связывания, свойственную TrkA-рецептору) и способна связываться с NGF.
Иммуноадгезины TrkA известны в данной области, и было обнаружено, что они блокируют связывание NGF с ТrkА-рецептором. См., например, патент США №6153189. Brennan et al. сообщают о введении иммуноадгезина TrkA крысам, представляющим собой модель послеоперационной боли. См. Society for Neuroscience Abstracts 24 (1-2) 880 (1998). В одном из воплощений иммуноадгезин TrkA включает слияние аминокислотной последовательности TrkA-рецептора (или ее части) из внеклеточного домена TrkA, способного связываться с NGF (в некоторых воплощениях аминокислотную последовательность, которая по существу сохраняет специфичность связывания, свойственную TrkA-рецептору), и последовательности иммуноглобулина. В некоторых воплощениях TrkA-рецептор представляет собой последовательность человеческого TrkA-рецептора, и слияние происходит с последовательностью константного домена иммуноглобулина. В других воплощениях последовательность константного домена иммуноглобулина представляет собой последовательность константного домена тяжелой цепи иммуноглобулина. В других воплощениях ассоциация двух продуктов слияния тяжелой цепи иммуноглобулина с TrkA-рецептором (например, через ковалентную(ые) дисульфидную(ые) связь(и)) приводит к гомодимерной структуре, подобной иммуноглобулину. Легкая цепь иммуноглобулина может быть дополнительно связана с одним или обоими из химер TrkA-рецептор-иммуноглобулин в связанном дисульфидной связью димере с получением гомотримерной или гомотетрамерной структуры. Примеры подходящих иммуноадгезинов TrkA включают в себя описанные в патенте США №6153189.
В еще одном воплощении антагонист NGF содержит по меньшей мере одно антитело против TrkA, способное блокировать, подавлять, изменять и/или снижать физическое взаимодействие NGF с TrkA-рецептором и/или сигнализацию в нисходящем направлении, что снижает и/или блокирует биологическую активность NGF. Антитела против TrkA известны в данной области. Примеры антител против TrkA включают в себя описанные в публикациях РСТ WO 97/21732, WO 00/73344, WO 02/15924 и в публикации США №20010046959.
В еще одном воплощении антагонист NGF содержит по меньшей мере одно антитело против р75, способное блокировать, подавлять и/или снижать физическое взаимодействие NGF с р75-рецептором и/или сигнализацию в нисходящем направлении, что снижает и/или блокирует биологическую активность NGF.
В еще одном воплощении антагонист NGF содержит по меньшей мере один ингибитор киназы, способный ингибировать киназную сигнализацию в нисходящем направлении, связанную с активностью TrkA- и/или р75-рецептора. Примерами ингибитора киназ являются K252а или K252b, которые известны в данной области и описаны в Knusel et al., J. Neurochem. 59:715-722 (1992); в Knusel et al., J. Neurochemistry 57:955-962 (1991); в Koizumi et al., J. Neuroscience 8:715-721 (1988); в Hirata et al., Chemical Abstracts 111:728, XP00204135, см. abstract и 12th Collective Chemical Substance Index, p.34237, с.3 (5-7), 55-60, 66-69), р.34238, с.1 (41-44), с.2 (25-27, 32-33), р.3423, с.3 (48-50, 52-53); и патент США №6306849.
Предполагается, что множество других категорий антагонистов NGF будут идентифицированы при поиске клиницистом.
Идентификация антагонистов NGF (таких как антагонистические антитела против NGF)
Антагонисты NGF, в том числе антагонистические антитела против NGF, можно идентифицировать или охарактеризовать при использовании способов, известных в данной области, которыми детектируют и/или измеряют снижение, облегчение или нейтрализацию биологической активности NGF. Можно использовать способы, описанные в РСТ WO 04/065560. Для идентификации агентов против NGF можно использовать другой способ, например анализ активации рецептора киназы (KIRA), описанный в патентах США №№5766863 и 5891650. Этот анализ типа ELISA подходит для качественного или количественного определения активации киназы путем измерения аутофосфорилирования киназного домена рецептора протеин-тирозинкиназы (здесь ниже "rРТK") например TrkA-рецептора, а также для идентификации и характеризации потенциальных антагонистов выбранных rРТK, например TrkA. Первая стадия анализа включает в себя фосфорилирование киназного домена рецептора киназы, например TrkA-рецептора, причем рецептор присутствует в клеточной мембране эукариотической клетки. Рецептор может быть эндогенным рецептором, или клетку можно трансформировать нуклеиновой кислотой, кодирующей рецептор или рецепторную конструкцию. В типичных случаях первую твердую фазу (например, лунку первого планшета для анализа) покрывают по существу гомогенной популяцией таких клеток (обычно клеточной линии млекопитающего) так, что клетки прилипают к твердой фазе. Часто клетки являются адгезивными и поэтому естественно прилипают к первой твердой фазе. Если используют "рецепторную конструкцию", она обычно содержит продукт слияния рецептора киназы и меченого (flag) полипептида. Опознавательный полипептид распознается захватывающим агентом, часто захватывающим антителом, в части анализа, представляющей собой ELISA. Затем анализаруемый компонент, например антагонист NGF-кандидат (включая антагонистическое антитело против NGF), добавляют вместе с NGF в лунки, содержащие прилипшие клетки, так что тирозинкиназный рецептор (например, TrkA-рецептор) подвергается воздействию (или контакту с) NGF и анализируемым компонентом. Это анализ делает возможным идентификацию антагонистов (в том числе антител), которые ингибируют активацию TrkA его лигандом NGF. После воздействия с NGF и анализируемым компонентом прилипающие клетки солюбилизируют при использовании лизисного буфера (в котором имеется солюбилизирующий детергент) и мягкого перемешивания, что высвобождает клеточный лизат, который можно подвергать анализу ELISA непосредственно, без необходимости в концентрировании или осветлении клеточного лизата.
Клеточный лизат, который получают таким образом, затем готов к стадии анализа, представляющей собой ELISA. В качестве первой стадии в стадии ELISA вторую твердую фазу (обычно лунку микротитрационного планшета для ELISA) покрывают захватывающим агентом (часто - захватывающим антителом), который специфично связывается с тирозинкиназным рецептором или, в случае рецепторной конструкции, с меченым полипептидом. Покрытие второй твердой фазы осуществляют так, что захватывающий агент прилипает ко второй твердой фазе. Захватывающий агент обычно представляет собой моноклональное антитело, но, как описано здесь в примерах, также можно использовать поликлональные антитела. Полученный клеточный лизат затем подвергают воздействию или приводят в контакт с прилипшим захватывающим агентом так, что рецептор или рецепторная конструкция прилипает ко второй твердой фазе (или захватывается ею). Затем осуществляют стадию промывки так, что удаляют несвязанный клеточный лизат, оставляя захваченный рецептор или рецепторную конструкцию. Затем прилипший или захваченный рецептор или рецепторную конструкцию экспонируют или приводят в контакт с антителом против фосфотирозина, которое распознает фосфорилированные остатки тирозина в тирозинкиназном рецепторе. В одном из воплощений антитело против фосфотирозина конъюгируют (прямо или опосредованно) с ферментом, который катализирует изменение цвета нерадиоактивного цветного реагента. Соответственно фосфорилирование рецептора можно измерять по последующему изменению цвета реагента. Фермент можно связать с антителом против фосфотирозина прямо, или антитело против фосфотирозина можно конъюгировать с молекулой конъюгации (например биотином), и фермент можно затем связать с антителом против фосфотирозина через молекулу конъюгации. Наконец, связывание антитела против фосфотирозина с захваченным рецептором или рецепторной конструкцией измеряют, например, по изменению цвета цветного реагента.
Антагонисты NGF (такие как антагонистическое антитело против NGF) можно также идентифицировать инкубированием агента-кандидата с NGF и мониторингом любой одной или более чем одной следующей характеристики: (а) связывание с NGF и ингибирование биологической активности NGF или путей в нисходящем направлении, опосредованных сигнальной функцией NGF; (б) ингибирование, блокирование или снижение активации рецептора NGF (включая димеризацию и/или аутофосфорилирование TrkA); (в) увеличение клиренса NGF; (г) лечение или предотвращение любого вида боли, вызванной ревматоидным артритом, или боли, вызванной остеоартритом; (д) ингибирование (снижение) синтеза, продукции или высвобождения NGF. В некоторых воплощениях антагонист NGF (например, антагонистическое антитело против NGF) идентифицируют инкубированием агента-кандидата с NGF и мониторингом связывания и/или сопутствующего снижения или нейтрализации биологической активности NGF. Анализ связывания можно осуществлять с очищенным(и) полипептидом(ами) NGF или с клетками, естественно экспрессирующими полипептид(ы) NGF или трансфицированными для такой экспрессии. В одном из воплощений анализ связывания представляет собой анализ конкурентного связывания, где оценивают способность агента-кандидата (такого как антитело) конкурировать с известным антагонистическим антителом против NGF за связывание NGF. Анализ можно осуществлять в различных форматах, в том числе в формате ELISA. В других воплощениях антагонист NGF (такой как антагонистическое антитело против NGF) идентифицируют инкубированием агента-кандидата с NGF и мониторингом связывания и сопутствующего ингибирования димеризации и/или аутофосфорилирования TrkA-рецептора.
После исходной идентификации активность антагониста NGF-кандидата (такого как антагонистическое антитело против NGF) можно дополнительно подтвердить и уточнить биоанализами, известными тем, что они тестируют целевые биологические активности. Альтернативно, биоанализы можно использовать для прямого скрининга агентов-кандидатов. Например, NGF стимулирует ряд морфологически распознаваемых изменений в чувствительных клетках. Это включают в себя, без ограничения, стимулирование дифференцировки клеток РС12 и усиление роста нейритов этих клеток (Greene et al., Proc Natl Acad Sci USA. 73(7):2424-8, 1976), стимулирование отрастания нейритов из эксплантатов чувствительных сенсорных и симпатических ганглиев (Levi-Montalcini, R. and Angeletti, P. Nerve growth factor. Physiol. Rev. 48:534-569, 1968) и стимулирование выживания нейронов, зависимых от NGF, таких как нейроны эмбриональных спинномозговых корешков, тройничного ганглия или симпатического ганглия (например, Chun & Patterson, Dev. Biol. 75:705-711, (1977); Buchman & Davies, Developmenti 18:989-1001 (1993). Таким образом, анализ ингибирования биологической активности NGF включает в себя культивирование клеток, чувствительных к NGF, с NGF вместе с анализируемым компонентом, например, антагонистом NGF-кандидатом (включая антагонистическое антитело против NGF). После соответствующего периода времени анализируют клеточный ответ (клеточную дифференцировку, отрастание нейрита или выживание клетки).
Способность антагониста NGF-кандидата (включая антагонистическое антитело против NGF) блокировать или нейтрализовать биологическую активность NGF можно также оценивать мониторингом способности агента-кандидата ингибировать опосредованное NGF выживание в эмбриональных ганглиях спинномозговых корешков крысы биоанализом, как описано в Hongo et al., Hybridoma 19:215-227 (2000).
Композиции для применения в способах по этому изобретению
Композиции, используемые в способах по этому изобретению (в отношении боли, вызванной ревматоидным артритом, и боли, вызванной остеоартритом) содержат эффективное количество антагониста NGF (такого как антитело против NGF) и в некоторых воплощениях дополнительно содержат фармацевтически приемлемый эксципиент. В некоторых воплощениях композиция предназначена для применения в любом описанном здесь способе. Примеры таких композиций, а также того, как их приготавливать, также описаны в предыдущем разделе и ниже. В одном из воплощений композиция содержит антагонист NGF. В еще одном воплощении композиция содержит один или более чем один антагонист NGF. В еще одном воплощении композиция содержит один или более чем один антагонист NGF, выбранный из любого одного или более чем одного из следующего: антагонист (например, антитело), который связывается (физически взаимодействует) с NGF, антагонист, который связывается с рецептором NGF (таким как TrkA- и/или р75-рецептор), и антагонист, который снижает (затрудняет и/или блокирует) сигнализацию от рецептора NGF в нисходящем направлении. В еще других воплощениях композиция содержит любой антагонист NGF, который не представляет собой иммуноадгезин TrkA (то есть является отличным от иммуноадгезина TrkA). В других воплощениях композиция содержит любой антагонист NGF, который является отличным от антитела против NGF. В еще других воплощениях композиция содержит любой антагонист NGF, который является отличным от иммуноадгезина TrkA и отличным от антитела против NGF. В других воплощениях антагонист NGF ингибирует (снижает) синтез, продукцию или высвобождение NGF. В некоторых воплощениях антагонист NGF связывается с NGF и в незначительной степени перекрестно взаимодействует с родственными нейротрофинами (такими как NT3, NT4/5 и/или BDNF). В некоторых воплощениях антагонист NGF не связан с нежелательным иммунным ответом. В некоторых воплощениях антагонист NGF выбран из группы, состоящей из антитела против NGF, антисмысловой молекулы, направленной на NGF (включая антисмысловую молекулу, направленную на нуклеиновую кислоту, кодирующую NGF), антисмысловой молекулы, направленной на рецептор NGF (такой как TrkA и/или р75), соединения, ингибирующего NGF, структурного аналога NGF, доминантно-негативной мутации TrkA-рецептора, которая связывается с NGF, иммуноадгезина TrkA, антитела против TrkA, антитела против р75 и ингибитора киназы. В еще одном воплощении антагонист NGF представляет собой антитело против NGF. В других воплощениях антитело против NGF распознает человеческий NGF. В некоторых воплощениях антитело против NGF является человеческим. В еще других воплощениях антитело против NGF является гуманизированным (таким как описанное здесь антитело Е3). В еще другом воплощении антитело против NGF содержит константную область, которая не вызывает нежелательный или неподходящий иммунный ответ, такой как опосредованный антителом лизис или ADCC. В других воплощениях антитело против NGF содержит один или более чем один CDR антитела Е3 (например, один, два, три, четыре, пять или в некоторых воплощениях все шесть CDR Е3).
Понятно, что композиции могут содержать более чем один из антагонистов NGF. Например, композиция может содержать более чем один из членов класса антагонистов NGF (например, смесь антител против NGF, которые распознают разные эпитопы NGF), а также из членов разных классов антагонистов NGF (например, антитело против NGF и соединение, ингибирующее NGF). Другие примеры композиции содержат более чем одно антитело против NGF, которые распознают один и тот (одни и те же) эпитоп(ы), разные виды антител против NGF, которые связываются с разными эпитопами NGF, или разные соединения, ингибирующие NGF.
Композиция, используемая в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and Practice of Pharmaceutics, 20th Ed. (2000) Lippincott Williams and Wilkins, Ed. K.Е.Hoover.) в форме лиофилизированных препаратов или водных растворов. Фармацевтически приемлемые эксципиенты описаны здесь далее.
Антагонист NGF и его композиции можно также использовать в сочетании с другими агентами, которые служат для усиления и/или дополнения эффективности этих агентов. При боли, вызванной остеоартритом, антагонист NGF можно вводить в сочетании с одним или более чем одним другим анальгетиком, нестероидным противовоспалительным средством (НПВС) или стероидами. Анальгетики включают в себя, без ограничения ими, ацетаминофен, трамадол, капсаицин (местного применения). Примерами НПВС являются ацетилированные салицилаты, в том числе аспирин; неацетилированные салицилаты, в том числе сальсалат, дифлунисал; уксусные кислоты, в том числе этодолак, диклофенак, индометацин, кеторолак, набумектон; пропионовые кислоты, в том числе фенопрофен, флурбипрофен, ибупрофен, кетопрофен, напроксен, напроксен в натриевой форме, оксапрозин; фенаматы, в том числе меклофенамат, мефенамовая кислота; фенилбутазон, пироксикам; ингибиторы циклоокигеназы-2 (ЦОГ-2), в том числе целекоксиб, эторикоксиб, валдекоксиб, рофенкоксиб, лумиракоксиб. Примерами стероидов являются внутрисуставные кортикостероиды (ВСК).
Для лечения боли, вызванной ревматоидным артритом, антагонист NGF можно вводить в сочетании с одним или более чем одним другим анальгетиком, НПВС, кортикостероидом (например, преднизоном) или другим модифицирующим заболевание противоревматическим лекарственным средством. Примерами модифицирующих заболевание противоревматических лекарственных средств являются метотрексат, гидроксихлорохин, сульфасалазин, лефлуномид, ингибиторы фактора некроза опухолей (TNF), растворимый рецептор интерлейкина-1, конъюгаты золота, цитотоксические агенты (азатиприн, циклофосфамид, циклоспорин А).
Введение антагониста NGF (такого как антагонистическое антитело против NGF)
Антагонист NGF (такой как антагонистическое антитело против NGF) можно вводить индивиду (при ревматоидном артрите и остеоартрите) любым подходящим путем. Специалисту в данной области должно быть ясно, что примеры, описанные здесь, предназначены не ограничивать, а иллюстрировать доступные методики. Соответственно в некоторых воплощениях антагонист NGF (такой как антагонистическое антитело против NGF) вводят индивиду в соответствии с известными способами, такими как внутривенное введение, например, в виде болюсной инъекции или непрерывной инфузии в течение некоторого периода времени, внутримышечным путем, внутрибрюшинным путем, в спинномозговую жидкость, подкожно, внутрисуставно, подъязычно, в синовиальную жидкость, путем инсуффляции, в мозговую оболочку, перорально, ингаляцией или местным путем. Введение может быть системным, например внутривенным введением, или локализованным. Для введения полезны имеющиеся в продаже распылители для жидких препаратов, в том числе струйные распылители и ультразвуковые распылители. Жидкие препараты можно распылять непосредственно, лиофилизированный порошок можно распылять после его разведения. Альтернативно, антагонист NGF (такой как антагонистическое антитело против NGF) можно распылять при использовании фторуглеродного препарата и ингалятора отмеренной дозы или ингалировать в виде лиофилизированного и измельченного порошка.
В одном из воплощений антагонист NGF (такой как антагонистическое антитело против NGF) вводят методиками сайт-специфичной или направленной локальной доставки. Примеры методик сайт-специфичной или направленной локальной доставки включают в себя различные имплантируемые деп-препараты, являющиеся источниками антагониста NGF (такого как антагонистическое антитело против NGF), или катетеры для локальной доставки, такие как инфузионные катетеры, постоянный катетер или игольный катетер, синтетические трансплантаты, наружные повязки, шунты и стенты или другие имплантируемые устройства, сайт-специфичные носители, прямая инъекция или прямое нанесение. См., например, публикацию РСТ WO № 00/53211 и патент США №5981568.
Для введения можно использовать различные препараты антагониста NGF (такого как антагонистическое антитело против NGF). В некоторых воплощениях антагонист NGF (например, антагонистическое антитело против NGF) можно вводить как таковой. В некоторых воплощениях антагонист NGF (например, антагонистическое антитело против NGF) и фармацевтически приемлемый эксципиент могут быть в различных препаратах. Фармацевтически приемлемые эксципиенты известны в данной области и представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества. Например, эксципиент может придавать форму или консистенцию или действовать в качестве разбавителя. Подходящие эксципиенты включают в себя, без ограничения ими, стабилизирующие агенты, смачивающие и эмульгирующие агенты, соли для варьирования осмолярности, инкапсулирующие агенты, буферы и усилители проницаемости кожи. Эксципиенты, а также препараты для парентеральной и непарентеральной доставки лекарственного средства указаны в Remington, The Science and Practice of Pharmaceutics, 20th Ed. Mack Publishing (2000).
В некоторых воплощениях эти агенты включают в состав перпарата для введения инъекцией (например, внутрибрюшинной, внутривенной, подкожной, внутримышечной и так далее). Соответственно эти агенты можно объединять с фармацевтически приемлемыми носителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и тому подобные. Конкретный режим дозировки, то есть доза, время и повторность, должны зависеть от конкретного индивида и его медицинской истории.
Антитело против NGF можно вводить при использовании любого подходящего способа, включающего в себя инъекцию (например, внутрибрюшинного, внутривенного, подкожного, внутримышечного и так далее). Антитела против NGF можно также вводить ингаляцией, как описано здесь. Обычно для введения антител против NGF исходная вероятно подходящая дозировка может быть от приблизительно 2 мг/кг. Для цели настоящего изобретения типичная ежедневная дозировка может быть в любом диапазоне от приблизительно 0,1 мкг/кг до 1 мкг/кг, до 3 мкг/кг, до 30 мкг/кг до 300 мкг/кг, до 3 мг/кг, до 30 мг/кг, до 100 мг/кг или более чем 100 мг/кг в зависимости от факторов, упомянутых выше. Например, антитело против NGF можно вводить при приблизительно 1 мкг/кг, приблизительно 10 мкг/кг, приблизительно 20 мкг/кг, приблизительно 50 мкг/кг, приблизительно 100 мкг/кг, приблизительно 200 мкг/кг, приблизительно 500 мкг/кг, приблизительно 1 мг/кг или приблизительно 2 мг/кг. Для повторных введений в течение нескольких суток или дольше в зависимости от состояния лечение продолжают, пока не происходит желаемое подавление симптомов или пока не достигают достаточных терапевтических уровней для снижения боли. Пример режима введения включает в себя введение исходной дозы около 2 мг/кг с последующей еженедельной поддерживающей дозой около 1 мг/кг антител против NGF или с последующей поддерживающей дозой около 1 мг/кг через неделю. Однако могут быть полезными другие режимы дозировки в зависимости от картины фармакокинетического профиля, которого хочет достичь практикующий врач. Например, в некоторых воплощениях предусмотрено введение от одного до четырех раз в неделю. Можно использовать даже менее частое введение. В некоторых воплощениях антитело против NGF вводят один раз в 2 недели, в 3 недели, в 4 недели, в 5 недель, в 6 недель, в 7 недель, в 8 недель, в 9 недель, в 10 недель, в 15 недель, в 20 недель, в 25 недель или реже. В некоторых воплощениях антитело против NGF вводят один раз в 1 месяц, в 2 месяца, в 3 месяца, в 4 месяца, в 5 месяцев, в 6 месяцев или реже. Ход этого лечения легко подвергать мониторингу общепринятыми методиками и анализами. Режим введения (включающий в себя использование антагониста(ов) NGF) можно варьировать в течение времени.
Исследования, выполненные на пациентах с болью, вызванной остеоартритом колена, от умеренной до сильной (суммированы в Примере 9), продемонстрировали, что дозировки в диапазоне от 3 до 300 мкг/кг обеспечивают облегчение боли при вариабельной продолжительности. Все тестированные дозировки (3, 10, 30, 100 и 300 мкг/кг) вызывали снижение боли в течение по меньшей мере 7 суток; более высокие дозировки приводили к продолжительному облегчению боли в течение по меньшей мере 28 суток (Фиг.24). Доза 100 мкг/кг вызывала облегчение боли в течение по меньшей мере 80 суток (Фиг.25).
В общем, антагонист NGF, когда он не представляет собой антитело, можно (в некоторых воплощениях) вводить при скорости приблизительно от 0,1 до 300 мг/кг массы пациента, разделив на 1-3 дозы или как раскрыто здесь. В некоторых воплощениях для взрослого пациента нормальной массы можно вводить дозы в диапазоне приблизительно от 0,3 до 5,00 мг/кг. Конкретный режим дозировки, то есть время и кратность введения дозы, должен зависеть от конкретного индивида и его истории болезни, а также от свойств индивидуальных агентов (таких как период полувыведения агента и другие соображения, хорошо известные в данной области).
Для цели настоящего изобретения подходящая дозировка антагониста NGF (включая антагонистическое антитело против NGF) должна зависеть от используемого антагониста NGF (или его композиции), от типа и тяжести боли, подлежащей лечению, от того, вводят ли агент в профилактических или терапевтических целях, от предыдущего лечения, от клинической истории пациента и реакции на агент и от решения лечащего врача. В типичных случаях врач-клиницист должен вводить антагонист NGF (например, антагонистическое антитело против NGF), пока не достигнет дозировки, которая дает желаемый результат. Дозу и/или частоту можно варьировать по ходу лечения.
Обычно вклад в определение дозировки должны вносить эмпирические соображения, такие как период полувыведения. Например, антитела, которые являются совместимыми с человеческой иммунной системой, такие как гуманизированные антитела или полностью человеческие антитела, можно использовать для увеличения периода полувыведения антител и для предотвращения атаки антитела со стороны иммунной системы хозяина. Частоту введения можно определять и корректировать по ходу лечения и обычно, но не обязательно, основывать на излечении, и/или подавлении, и/или облегчении, и/или задержке боли. Альтернативно, могут быть подходящими препараты для постоянного непрерывного высвобождения антагонистов NGF (например, антагонистических антител против NGF). Различные препараты и устройства для достижения непрерывного высвобождения известны в данной области.
В одном из воплощений дозировки антагониста NGF (например, антагонистического антитела против NGF) можно определять эмпирически у индивидов, которые получали одно или более чем одно введение антагониста NGF. Индивидам дают возрастающие дозировки антагониста NGF (например, антагонистического антитела против NGF). Для оценки эффективности антагониста NGF (например, антагонистического антитела против NGF) можно отслеживать показатель боли.
Введение антагониста NGF (например, антагонистического антитела против NGF) в соответствии со способом по настоящему изобретению может быть непрерывным или прерывистым, например, в зависимости от физиологического состояния реципиента, от того, является ли введение терапевтическим или профилактическим, и от других факторов, известных специалистам. Введение антагониста NGF (например, антагонистического антитела против NGF) может быть по существу непрерывным в течение предопределенного периода времени и может быть серией разделенных доз, например либо до, либо во время, либо после развития боли; до; во время; до и после; во время и после; до и во время или до, во время и после развития боли.
В некоторых воплощениях может присутствовать более чем один антагонист NGF, такой как антагонистическое антитело против NGF. Возможно присутствие по меньшей мере одного, по меньшей мере двух, по меньшей мере трех, по меньшей мере четырех, по меньшей мере пяти или более чем пяти разных антагонистов NGF (например, антагонистических антител против NGF). Обычно эти антагонисты NGF (такие как антагонистические антитела против NGF) имеют комплементарные активности, которые не влияют друг на друга нежелательным образом. Антагонисты NGF можно также использовать в сочетании с другими агентами, которые служат для усиления и/или дополнения эффективности агентов.
Терапевтические препараты антагониста NGF (например, антагонистического антитела против NGF), используемые в соответствии с настоящим изобретением, приготавливают для хранения путем смешивания антагониста NGF (например, антитела), имеющего желаемую степень чистоты, с возможными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington, The Science and Practice of Pharmaceutics, 20th Ed. Mack Publishing (2000)) в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов при используемых дозировках и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводороды, в том числе глюкозу, маннозу, или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металла (например комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ).
Липосомы, содержащие антагонист NGF (такой как антагонистическое антитело против NGF), получают способами, известными в данной области, такими как описано в Epstein, et al., Proc. Natl. Acad. Sci. USA 82:3688 (1985); Hwang, et al., Proc. Natl Acad. Sci. USA 77:4030 (1980); и патентах США №№4485045 и 4544545. Липосомы с улучшенным временем циркуляции раскрыты в патенте США №5013556. Особенно полезные липосомы можно получать способом обратнофазного упаривания с липидной композицией, содержащей фосфатидилхолин, холестерин и дериватизированный полиэтиленгликолем фосфатидилэтаноламин (ПЭГ-ФЭ). Липосомы экструдируют через фильтры с определенным размером пор, получая липосомы с желаемым диаметром.
Активные ингредиенты можно также заключать в микрокапсулы, получаемые, например, методиками коацервации или межфазной полимеризации, например гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметацилатные) микрокапсулы соответственно, в коллоидных системах доставки лекарственного средства (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методики раскрыты в Remington, The Science and Practice of Pharmaceutics, 20th Ed. Mack Publishing (2000).
Можно получать препараты с непрерывным высвобождением. Подходящие примеры препаратов с непрерывным высвобождением включают в себя полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антагонист (такой как антитело), где эти матрицы представляют собой частицы определенной формы, например пленки или микрокапсулы. Примеры матриц для непрерывного высвобождения включают в себя полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT ™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида), ацетат-изобутират сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
Препараты для использования для введения in vivo должны быть стерильными. Это легко осуществить, например, фильтрованием через стерильные фильтровальные мембраны. Композиции терапевтического антагониста NGF (например, антагонистического антитела против NGF) обычно помещают в контейнер, имеющий порт для стерильного доступа, например в мешок для внутривенного раствора или во флакон, имеющий пробку, прокалываемую гиподермальной инъекционной иглой.
Композиции в соответствии с настоящим изобретением могут быть в стандартных лекарственных формах, таких как таблетки, пилюли, капсулы, порошки, гранулы, растворы или суспензии или суппозитории для перорального, парентерального или ректального введения или введения ингаляцией или инсуффляцией.
Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим носителем, например общепринятыми таблеточными ингредиентами, такими как кукурузный крахмал, лактоза, сахароза, сорбит, тальк, стеариновая кислота, стеарат магния, двукальциевая соль фосфорной кислоты или смолы, и другими фармацевтическими разбавителями, например водой, с образованием твердой предварительной композиции, содержащей гомогенную смесь соединения по настоящему изобретению или его нетоксичной фармацевтически приемлемой соли. Упоминание этих предварительных композиций как гомогенных означает, что активный ингредиент равномерно диспергирован по композиции так, что композицию можно легко разделить на равные по эффективности стандартные лекарственные формы, такие как таблетки, пилюли и капсулы. Эту твердую предварительную композицию затем делят на стандартные лекарственные формы вышеописанного типа, содержащие от 0,1 до приблизительно 500 мг активного ингредиента по настоящему изобретению. Таблетки или пилюли новой композиции можно покрывать оболочкой или приготавливать иным образом с получением лекарственной формы, дающей преимущество пролонгированного действия. Например, таблетка или пилюля может содержать внутренний дозовый и внешний дозовый компонент, причем последний из них находится в форме оболочки над первым. Два компонента можно разделить энтеросолюбильным слоем, который служит для предотвращения разложения в желудке и позволяет внутреннему компоненту проходить интактным в поджелудочную железу или замедлять его высвобождение. Для таких энтеросолюбильных слоев и оболочек можно использовать множество разных веществ, причем такие вещества включают в себя множество полимерных кислот и смесей полимерных кислот с такими веществами, как шеллак, цетиловый спирт и ацетат целлюлозы.
Подходящие поверхностно-активные агенты включают в себя, в частности, неионные агенты, такие как полиоксиэтиленсорбитаны (например, Tween™ 20, 40, 60, 80 или 85) и другие сорбитаны (например Span™ 20, 40, 60, 80 или 85). Удобным является то, что композиции с поверхностно-активным агентом содержат от 0,05 до 5% поверхностно-активного агента и могут содержать от 0,1 до 2,5%. Следует понимать, что если необходимо, можно добавлять другие ингредиенты, например маннит или другие фармацевтически приемлемые носители.
Подходящие эмульсии можно получать при использовании имеющихся в продаже жировых эмульсий, таких как Intralipid™, Liposyn™, Infonutrol™, Lipofundin™ и Lipiphysan™. Активный ингредиент можно растворить либо в предварительно размешанной композиции эмульсии, либо в масле (например, соевом масле, подсолнечном масле, хлопковом масле, кунжутном масле, кукурузном масле или миндальном масле) и образовать эмульсию перемешиванием с фосфолипидом (например, яичными фосфолипидами, соевыми фосфолипидами или соевым лецитином) и водой. Следует понимать, что для регуляции тоничности эмульсии можно добавлять другие ингредиенты, например глицерин или глюкозу. Подходящие эмульсии должны в типичных случаях содержать до 20% масла, например от 5 до 20%. Жировая эмульсия может содержать жировые капельки размером от 0,1 до 1,0 мкм, в частности 0,1 и 0,5 мкм, и иметь рН в диапазоне от 5,5 до 8,0.
Композиции, представляющие собой эмульсию, можно получать перемешиванием антагониста NGF (такого как антитело против фактора роста нервов) с Intralipid™ или его компонентами (соевым маслом, яичными фосфолипидами, глицерином и водой).
Композиции для ингаляции или инсуффляции включают в себя растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях и порошки. Жидкие или твердые композиции могут содержать подходящие фармацевтически приемлемые эксципиенты, как указано выше. В некоторых воплощениях композиции вводят пероральным или назальным респираторным путем для местного или системного эффекта. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно распылять с использованием газов. Распыленные растворы можно вдыхать прямо из распылительного устройства, или распылительное устройство можно прикрепить к маске для лица, тенту или дыхательному аппарату пульсирующего положительного давления. Композиции, представляющие собой раствор, суспензию или порошок, можно вводить, предпочтительно перорально или назально, из устройств, которые доставляют препарат подходящим образом.
Эффективность лечения можно оценивать способами, хорошо известными в данной области.
НАБОРЫ, СОДЕРЖАЩИЕ АНТИТЕЛА И ПОЛИНУКЛЕОТИДЫ ПО ДАННОМУ ИЗОБРЕТЕНИЮ
В этом изобретении также предложены наборы, содержащие антитела или полипептиды для применения в детектировании и/или лечении. Соответственно в некоторых воплощениях наборы содержат антитело Е3. В некоторых воплощениях набор содержит любое антитело или полипептид, описанные здесь.
В других аспектах наборы можно использовать для любых описанных здесь способов, в том числе, например, для лечения индивида с болью (включая послеоперационную боль, боль, вызванную ревматоидным артритом, и боль, вызванную остеоартритом). Наборы по этому изобретению находятся в подходящей упаковке и, возможно, могут предоставлять дополнительные компоненты, такие как буферы, и инструкции для применения антител в любом описанном здесь способе. В некоторых воплощениях наборы включают в себя инструкции по лечению боли. В некоторых воплощениях набор содержит описанное здесь антагонистическое антитело против NGF и инструкции по лечению и/или предотвращению боли, вызванной ревматоидным артритом, у индивида. В других воплощениях набор содержит описанный здесь антагонист NGF (такой как антагонистическое антитело против NGF) и инструкции по лечению и/или предотвращению боли, вызванной остеоартритом, у индивида. В некоторых воплощениях антагонистическое антитело против NGF представляет собой антитело Е3.
В другом аспекте в этом изобретении предложены наборы, содержащие полинуклеотид, кодирующий полипептид Е3, как описано здесь. В некоторых воплощениях наборы дополнительно содержат инструкции для применения полинуклеотида в любом описанном здесь способе.
СПОСОБЫ КОРРЕКЦИИ АФФИННОСТИ АНТИТЕЛА И СПОСОБЫ ХАРАКТЕРИЗАЦИИ CDR
Авторы разработали новый способ характеризации CDR антитела и/или изменения (такого как улучшение) аффинности связывания, свойственной полипептиду, такому как антитело, названный "сканирующий библиотечный мутагенез" ("library scanning mutagenesis"). Обычно сканирующий библиотечный мутагенез работает следующим образом. Одно или более чем одно аминокислотное положение в CDR заменяют двумя или более чем двумя (например 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислотами при использовании способов, признанных в данной области. Это генерирует малые библиотеки клонов (в некоторых воплощениях один для каждого анализируемого аминокислотного положения), причем каждая укомплектована двумя или более чем двумя членами (если замены в каждом положении включают в себя две или более чем две аминокислоты). Обычно библиотека также включает в себя клон, содержащий нативную (незамененную) аминокислоту. Малое число клонов, например около 20-80 клонов (в зависимости от комплектности библиотеки), из каждой библиотеки подвергают скринингу на аффинность связывания с целевым полипептидом и идентифицируют вероятно подходящие варианты с повышенным, таким же, сниженным или отсутствующим связыванием. Способы определения аффинности связывания хорошо известны в данной области. В некоторых воплощениях аффинность связывания определяют при использовании анализа поверхностного плазменного резонанса BIAcore, который детектирует приблизительно 2-кратные или более чем 2-кратные различия в аффинности связывания. BIAcore является особенно полезным, когда исходное антитело уже связывается при относительно высокой аффинности, например при KD около 10 нМ или ниже чем 10 нМ. Скрининг при использовании поверхностного плазменного резонанса BIAcore описан здесь в примерах.
В других воплощениях аффинность связывания определяют при использовании Kinexa Biosensor, анализов сцинтилляционной близости, ELISA, иммуноанализа ORIGEN (IGEN), гашения флуоресценции, переноса флуоресценции и/или дрожжевого дисплея. В других воплощениях аффинность связывания подвергают скринингу при использовании подходящего биоанализа.
В некоторых воплощениях каждое аминокислотное положение в CDR (в некоторых воплощениях одну за раз) заменяют всеми 20 природными аминокислотами при использовании способов мутагенеза, признанных в данной области (некоторые из которых описаны здесь). Это генерирует малые библиотеки клонов (в некоторых воплощениях одну для каждого анализируемого аминокислотного положения), каждая с комплектностью 20 членов (если в каждом положении замены включают в себя все 20 аминокислот).
В некоторых воплощениях библиотека, подлежащая скринингу, содержит замены в двух или более чем двух положениях, которые могут быть в одном и том же CDR или в двух или более чем двух CDR. Таким образом, в некоторых воплощениях библиотека содержит замены в двух или более чем двух положениях в одном CDR. В других воплощениях библиотека содержит замену в двух или более чем двух положениях в двух или более чем двух CDR. В еще других воплощениях библиотека содержит замену в 3, 4, 5 или более чем 5 положениях, причем указанные положения находятся в двух, трех, четырех, пяти или шести CDR. В некоторых воплощениях замену осуществляют при использовании кодонов с низкой избыточностью. См., например, Таблицу 2 в Balint et al., (1993) Gene 137(1): 109-18.
В некоторых воплощениях CDR представляет собой CDR-Н3 и/или CDR-L3. В других воплощениях CDR представляет собой один или более чем один из CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и/или CDR-H3. В некоторых воплощениях CDR представляет собой CDR no Kabat, CDR по Chothia или расширенный CDR.
Вероятно подходящие варианты с улучшенным связыванием можно подвергнуть определению последовательности и тем самым идентифицировать мутант, представляющий собой замену в CDR, которая приводит к улучшенной аффинности (также обозначаемый как "улучшенная" замена). Например, как продемонстрировано в Примере 1, применение этого способа позволяет идентифицировать единственную замену, которая улучшает связывание, даже когда оценивают 18 других замен в том же аминокислотном положении, приводящих к отсутствию связывания (то есть к потере функции антитела). Вероятно подходящие варианты, которые связываются, также можно подвергнуть определению последовательности и тем самым идентифицировать замену в CDR, которая сохраняет связывание.
В некоторых воплощениях осуществляют множественные раунды скрининга. Например, вероятно подходящие варианты (каждый содержит аминокислотную замену в одном или более чем одном положении одного или более чем одного CDR) с улучшенным связыванием также являются полезными для конструирования второй библиотеки, содержащей по меньшей мере исходные и замененные аминокислоты в каждом улучшенном положении CDR (то есть аминокислотном положении в CDR, при котором мутант, представляющий собой замену, показал улучшенное связывание). Получение и скрининг или селекция этой библиотеки обсуждены дополнительно ниже.
Сканирующий библиотечный мутагенез также обеспечивает способ характеризации CDR, поскольку частота клонов с улучшенным связыванием, таким же связыванием, сниженным связыванием или отсутствием связывания также дает информацию, относящуюся к важности каждого аминокислотного положения для стабильности комплекса антитела с антигеном. Например, если некоторое положение в CDR сохраняет связывание при замене на все 20 аминокислот, это положение идентифицируют как положение, которое едва ли требуется для связывания антигена. Наоборот, если положение в CDR сохраняет связывание в только малом проценте замен, это положение идентифицируют как положение, которое является важным для функции CDR. Таким образом, способы сканирующего библиотечного мутагенеза генерируют информацию о положениях в CDR, которые можно заменять многими разными аминокислотами (в том числе всеми 20 аминокислотами), и о положениях в CDR, которые нельзя заменить или которые можно заменить только несколькими аминокислотами. Этот аспект обсужден и проиллюстрирован в Примере 1.
В некоторых воплощениях вероятно подходящие варианты с улучшенной аффинностью объединяют во второй библиотеке, которая включает в себя улучшенную аминокислоту и исходную аминокислоту в этом положении, и могут дополнительно включать в себя дополнительные замены в этом положении в зависимости от комплектности библиотеки, которая является желаемой или допустимой при использовании желаемого способа скрининга или селекции. В дополнение, если желательно, соседние аминокислотные положения можно рандомизировать по меньшей мере до двух или более чем двух аминокислот. Рандомизация соседних аминокислот может придать дополнительную конформационную гибкость в мутанте CDR, что может в свою очередь сделать возможным или облегчить введение большего числа улучшающих мутаций. В некоторых воплощениях библиотека также содержит замену в положениях, которые не показывают улучшенную аффинность в первом раунде скрининга.
Вторую библиотеку подвергают скринингу или селекции на те члены библиотеки, которые имеют улучшенную и/или измененную аффинность связывания, при использовании любых способов, известных в данной области, в том числе скрининга при использовании анализа поверхностным плазменным резонансом BIAcore и селекции при использовании любых способов, известных в данной области для селекции, включая фаговый дисплей, дрожжевой дисплей и рибосомный дисплей.
Преимущества способов коррекции аффинности антител и характеризации CDR
Эти способы полезны для предварительного скрининга аминокислотных положений CDR для идентификации аминокислотных замен, которые улучшают связывание или сохраняют связывание. Предварительная идентификация важных остатков и замен, которые улучшают связывание и/или замены, которые сохраняют функцию антитела, делает возможным эффективное конструирование и скрининг библиотеки для созревания аффинности.
Настоящий способ также полезен для характеризации CDR и дает обширную информацию относительно важности каждого аминокислотного положения в CDR для связывания антигена. Настоящий способ можно также использовать для идентификации замен, которые улучшают связывание.
Применение малых библиотек, в которых каждое положение можно рандомизировать (в некоторых воплощениях, одно за один раз), делает возможным скрининг мутантов, представляющих собой замену, при использовании чувствительных способов, таких как BIAcore, который дает детальную кинетическую информацию. Способы скрининга обычно являются непрактичными, когда скринингу подвергают большую библиотеку. Вместо этого для идентификации клонов, которые сохраняют связывание, обычно используют способы селекции, такие как фаговый дисплей, дрожжевой дисплей и рибосомный дисплей. Фаговый дисплей и анализы ELISA могут сильно зависеть от концентрации белковой пробы, полученной из клона, и, таким образом, имеют тенденцию к сильному предпочтению клонов, которые имеют повышенную экспрессию, повышенную стабильность или сниженную токсичность, а не к идентификации клонов с повышенной аффинностью связывания. В дополнение, отличия в уровне экспрессии клонов могут маскировать небольшие улучшения в аффинности связывания. Эти недостатки являются особенно острыми, когда в качестве исходного материала используют антитело с высокой аффинностью связывания, потому что для того, чтобы сделать скрининг достаточно строгим, следует использовать очень низкие уровни антигена.
Напротив, способы по этому изобретению, такие как рандомизация в каждом положении (в некоторых воплощениях одно положение за раз), делает возможным введение и характеризацию эффекта замены, например, всеми 20 аминокислотами в данном положении. Этот анализ дает информацию о том, как много замен в данном положении допустимо (то есть сохраняют связывание антитела), что в свою очередь дает информацию относительно важности каждой аминокислоты для функции антитела. Дополнительно, замены, которые приводят к улучшенному связыванию, можно идентифицировать даже в обстоятельствах, при которых многие или большинство из замен в данном положении дают нефункциональные (несвязывающие) антитела. Напротив, мутагенез со сканированием по аланину, который обычно используют для идентификации важных положений в CDR, дает информацию относительно того, делает ли возможным или предотвращает связывание замена аланином. Обычно положения, в которых замена аланином предотвращает связывание, удалены из библиотеки с развитой аффинностью. Однако во многих случаях аланин может быть плохой заменой в этом положении CDR.
Настоящие способы также делают возможной идентификацию и характеризацию эффекта отдельных мутаций CDR. Напротив, такие способы, как фаговый дисплей, вводят и подвергают селекции много мутаций одновременно и, таким образом, потенциально увеличивают риск того, что положительные мутации будут маскированы наличием разрушительной мутации, присутствующей в конкретном клоне.
Настоящие способы также являются полезными для улучшения аффинности при сохранении специфичности связывания, свойственной изначальному (исходному) антителу, поскольку настоящие способы делают возможной идентификацию малых чисел мутаций (например, 1, 2, 3, 4 или 5 мутаций в единственном CDR), которые приводят к улучшенной аффинности связывания. Напротив, такие способы, как фаговый дисплей, в типичных случаях улучшают аффинность связывания при использовании множественных мутаций за один раз, что может приводить к сдвигу специфичности антитела и/или увеличению нежелательных перекрестных взаимодействий.
Следующее примеры даны для иллюстрации, но не для ограничения этого изобретения.
ПРИМЕРЫ
Пример 1. Гуманизация и развитие аффинности мышиного антагонистического антитела 911 против NGF
А. Общие способы
В этом примере использовали следующие общие способы.
Генерирование библиотеки
Библиотеки генерировали кассетным ПЦР-мутагенезом с вырожденными олигонуклеотидами, как описано в Kау et al. (1996), Phage display of peptides and proteins: a laboratory manual, San-Diego, Academic Press (см. стр.277-291). Допинговый кодон NNK использовали для рандомизации одного аминокислотного положения, так чтобы она включала в себя 20 возможных аминокислот. Для рандомизации одного аминокислотного положения, так чтобы она включала в себя только подмножество аминокислот со специфическими свойствами, использовали допинговые кодоны, как описано в Balint et al. (1993) Gene 137(1): 109-18. Направленный мутагенез осуществляли при использовании рекомбинантной ПЦР, как описано в Innis et al. (1990) PCR protocols: A guide to methods and applications (см. стр.177-183).
Получение Fab в малом масштабе
Экспрессию в малом масштабе в 96-луночных планшетах оптимизировали для скрининга библиотеки Fab. Начиная с Е.coli, трансформированных библиотекой Fab, брали колонии для инокуляции главного планшета (агар LB с ампициллином (50 мкг/мл) + 2% глюкозы) и рабочего планшета (2 мл/лунка, 96 лунок/планшет, содержат 1,5 мл LB с ампициллином (50 мкг/мл) + 2% глюкозы). Оба планшета инкубировали при 30°С в течение 8-12 часов. Главный планшет хранили при 4°С, клетки из рабочего планшета осаждали при 5000 об/мин и снова суспендировали с 1 мл LB с ампициллином (50 мкг/мл) + 1 мМ изопропил-β-тиогалактопиранозид (IPTG) для индукции экспрессии Fab. Клетки собирали центрифугированием после 5-ч периода экспрессии при 30°С, затем снова суспендировали в 500 мкл буфера HBS-EP (100 мМ буфер HEPES при рН 7,4, 150 мМ NaCl, 0,005% Р20, 3 мМ ЭДТА). Лизис клеток, ресуспендированных в HBS-EP, осуществляли одним циклом замораживания (-80°С) и оттаивания при 37°С. Клеточный лизат центрифугировали при 5000 об/мин в течение 30 мин для отделения клеточных остатков от супернатантов, содержащих Fab. Супернатанты затем инъецировали в аппарат BIAcore для плазмонного резонанса с получением информации об аффинности для каждого Fab. Клоны, экспрессирующие Fab, собирали из главного планшета для определения последовательности ДНК и для крупномасштабного получения и подробного определения характеристик Fab, как описано ниже.
Крупномасштабное получение Fab
Для получения подробных кинетических параметров экспрессировали Fab и очищали их из больших культур. В колбы Эрленмейера, содержащие 200 мл LB с ампициллином (50 мкг/мл) + 2% глюкозы, вносили по 5 мл ночной культуры из выбранного клона E.coli, экспрессирующего Fab. Клоны инкубировали при 30°С до достижения величины оптической плотности 1,0 при 550 нм (OD550nm) и затем индуцировали заменой среды на 200 мл LB с ампициллином (50 мкг/ мл) + 1 мМ IPTG. После 5-ч периода экспрессии при 30°С клетки осаждали центрифугированием, затем снова суспендировали в 10 мл забуференного фосфатами физиологического раствора (PBS) при рН 8. Лизис клеток осуществляли двумя циклами замораживания/оттаивания (при -80°С и 37°С соответственно). Супернатант клеточного лизата наносили на колонки Superflow с сефарозой Ni-NTA (Qiagen, Valencia. CA), уравновешенные с PBS, рН 8, затем промывали при использовании PBS (5 объемов колонки, рН 8). Индивидуальные Fab элюировали в разных фракциях при использовании PBS (рН 8) + 300 мМ имидазол. Фракции, содержащие Fab, объединяли в пул и диализировали в PBS, затем перед характеризацией аффинности количественно анализировали при использовании ELISA.
Получение полных антител
Для экспрессии полных антител вариабельные области тяжелой и легкой цепи клонировали в 2 экспрессионных вектора млекопитающего (Eb.911.E3 или Eb.pur.911.3E для легкой цепи и Db.911.3E для тяжелой цепи; описаны здесь) и трансфицировали, используя липофектемин, клетки НЕK 293 для временной экспрессии. Антитела очищали при использовании протеина А стандартными способами.
Анализ Biacore
Аффинности Fab и моноклональных антител против NGF определяли при использовании системы BIAcore 3000™ для поверхностного плазменного резонанса (ППР) (BIAcore, INC, Piscaway NJ). Чипы СМ5 активировали гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Человеческий NGF разбавляли в 10 мМ ацетате натрия при рН 4,0 и инъецировали на активированный чип при концентрации 0,005 мг/мл. При использовании варьирующего времени потока через индивидуальные каналы чипа достигали два диапазона плотности антигена: 100-200 единиц отклика (ЕО) для детального кинетического исследования и 500-600 ЕО для скрининговых анализов. Чип блокировали этаноламином. Исследования регенерации показали, что смесь пирсовского буфера для элюции (продукт №21004, Pierce Biotechnology, Rockford, IL) и 4 M NaCl (2:1) эффективно удаляла связанный Fab, сохраняя активность человеческого NGF на чипе для более чем 200 инъекций. В качестве рабочего буфера для всех анализов BIAcore использовали буфер HBS-EP (0,01 M HEPES, рН 7,4, 0,15 NaCl, 3 мМ ЭДТА, 0,005% поверхностно-активного вещества Р29).
Скрининговый анализ
Скрининговый анализ BIAcore оптимизировали для определения аффинности клонов Fab из библиотеки. Супернатанты лизата малой культуры инъецировали при 50 мкл/мин в течение 2 мин. Для определения единственной экспоненциальной скорости диссоциации (koff) при использовании программного обеспечения BIAevaluation использовали длительности диссоциации от 10 до 15 минут. Для подтверждения инъецировали пробы, которые показали оценки koff в том же диапазоне, что и матрица, использованная для создания библиотеки (клон 8L2-6D5, koff 1×10-3 сек-1), и для диссоциации обеспечивали время до 45 мин для получения более точных величин koff. Клоны, показывающие улучшенные (сниженные) величины koff, экспрессировали в крупном масштабе и полные кинетические параметры, kon и koff, определяли на очищенном белке. Анализ оказался способным обнаруживать отличия в аффинности, которые были приблизительно 2-кратными или более чем 2-кратными.
Анализ для определения аффинности
Инъецировали серийные разведения (0,1-10× оценки KD) проб очищенного Fab в течение 1 мин при 100 мкл/мин и для диссоциации обеспечивали время до 2 ч. Концентрации белков Fab определяли при использовании ELISA и/или электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), используя в качестве стандарта Fab известной концентрации (определена анализом аминокислот). Кинетические скорости ассоциации (kon) и диссоциации (koff) получали одновременной аппроксимацией данных 1:1 на модели связывания Лэнгмюра (Karlsson, R. Roos, H. Fagerstam, L. Petersson, В. (1994). Methods Enzymology 6. 99-110) при использовании программы BIAevaluation. Величины равновесной константы диссоциации (KD) вычисляли как koff/kon.
Б. Гуманизация и созревание аффинности мышиного антагонистического антитела 911 против NGF
Для гуманизации и созревания аффинности было выбрано мышиное антагонистическое антитело 911 против NGF (см. Hongo et al., (2000) Hybridoma 19(3):215-227). Моноклональное антитело 911 связывается с человеческим и крысиным NGF с высокой аффинностью и не проявляет значительного перекрестного взаимодействия с нейротрофинами NT3, NT4/5 или BDNF. См. Hongo, id. Аффинность отщепленного папаином Fab-фрагмента мышиного моноклонального антитела 911 определяли при использовании анализа BIAcore, как описано выше. Отщепленный папаином Fab-фрагмент мышиного моноклонального антитела 911 связывал человеческий NGF при KD приблизительно 10 нМ.
Гуманизацию и созревание аффинности осуществляли в несколько стадий следующим образом.
(1) Получение матрицы с трансплантированным CDR. Расширенные CDR легкой цепи антитела 911 (то есть включающие в себя области CDR по Kabat и по Chothia) трансплантировали в акцепторные последовательности человеческой зародышевой линии O8 при использовании JK2, расширенные CDR тяжелой цепи антитела 911 трансплантировали в акцепторную последовательность человеческой зародышевой линии VH4-59 при использовании JH4. Аминокислотные последовательности акцепторных последовательностей человеческой зародышевой линии показаны на Фиг.1А и 1Б. Нумерация аминокислот является последовательной. Используя указанные выше каркасные области белка, конструировали последовательности ДНК для синтетических генов, кодирующих человеческие каркасные области с мышиными CDR. Эти гуманизированные тяжелые и легкие вариабельные домены были обозначены как hVH и hVL соответственно. Оптимизировали кодоны для использования в Е.coli и в хомячках. Несколько перекрывающихся олигонуклеотидов (69-90 оснований в длину), увеличивающих полную длину hVL и hVH на два коротких фланкирующих праймера для каждой цепи, использовали для раздельного синтеза этих двух генов рекурсивной ПЦР, по существу как описано в Prodromou et al., (1992) Protein Eng 5(8); 827-9. Получившиеся фрагменты ДНК правильной длины очищали в геле и затем клонировали в бицистронную экспрессионную плазмиду для Е.coli (устойчивую к ампициллину). Экспрессия антител была под контролем IPTG-индуцируемого промотора lacZ, подобного описанному в Barbas (2001) Phage display: a laboratory manual, Cold Spring Harbor, NY, Cold Spring Harbor Laboratory Press (см. Вектор pComb3X, при pg 2.10), однако модификации включали в себя добавление и экспрессию следующих дополнительных доменов: константный домен человеческой легкой каппа-цепи (см. GenBank Accession № САА09181) и константный домен CHI человеческого иммуноглобулина IgG2 (GenBank Accession № Р01859).
Аминокислотные последовательности вариабельных областей антитела с трансплантированным CDR (также называемого "матрицей"), обозначенные как 8L2-4D5, также показаны на Фиг.1А и 1Б. Аффинность 8L2-4D5 определяли при использовании анализа BIAcore, как описано выше. 8L2-4D5 связывали человеческий NGF при KD приблизительно 38 нМ.
(2) Введение точечной мутации в последовательность каркасной области. Замену V71K вводили в тяжелую цепь с трансплантированным CDR при использовании направленного мутагенеза рекомбинантной ПЦР, как описано в Innis et al., (1995) PCR strategies, San Diego, Academic Press. Эта замена заменяла остаток человеческой каркасной области соответствующим остатком мышиной каркасной области. Получившееся антитело было обозначено как 8L2-6D5, и аминокислотная последовательность вариабельной области тяжелой цепи 8L2-6D5 показана на Фиг.1А. Аффинность 8L2-6D5 определяли при использовании анализа BIAcore, как описано выше. Fab-фрагмент 8L2-6D5 связывал человеческий NGF при KD приблизительно 15 нМ. 8L2-6D5 выбрали в качестве матрицы для созревания аффинности.
(3) Гуманизация и созревание аффинности CDRs L1, L2, H1 и Н2. CDRs L1, L2, Н1 и Н2 подвергали гуманизации и созреванию аффинности. В CDRs L1, L2, Н1 и Н2 идентифицировали аминокислотные положения, которые не являются существенными для структуры CDR на основании канонической структуры по Chothia (см. Al-Lazikani et al. (1997) J. Mol. Biol. 273(4):927-48), и подвергали их рандомизации следующим образом. Получали две библиотеки, содержащие мутации легкой цепи или мутации тяжелой цепи, показанные в Таблице 2, и трансплантированные (мышиные) CDR-L3 или CDR-H3 соответственно при использовании кассетного ПЦР-мутагенеза с вырожденными олигонуклеотидами, как описано в Kаy et al. (1996), Phage display of peptides and proteins: a laboratory manual. San Diego, Academic Press, с использованием допинговых кодонов, как описано в Balint et al., (1993) Gene 137(1):109-18). В общем, заменяли аминокислотные остатки остатками, которые являются более распространенными в человеческих антителах, на основании выравнивания аминокислотных последовательностей легкой цепи и тяжелой цепи антитела 911 с последовательностями антитела человеческой зародышевой линии. Аминокислотный остаток дикого типа (не замененный) также представлен в библиотеке, за исключением остатка 50 CDR-H2, метионина, в таком случае метионин дикого типа не представлен в библиотеке. Метиониновые остатки подвержены окислению; таким образом, предполагается, что замена этого остатка улучшает стабильность полученного антитела. Библиотеки Fab клонировали в вектор рСоmb3Х с человеческими СН1- и Сκ-областями, описанными выше.
Таблица 2
1. Библиотека тяжелых цепей Н1/Н2:
CDR-H1
I34 заменен на F, L, V, S, Р, Т, А или I
N35 заменен на N, Т, S или Y
CDR-H2
М50 заменен на все 20 природных аминокислот
А62 заменен на А или S
L63 заменен на L или V
2. Библиотека легких цепей L1/L2
CDR-L1
S26 заменен на S, А, V или F
D28 заменен на D, А, S или Y
Н32 заменен на Н, N, K, D, Е, Q или Y
CDR-L2
Y50 заменен на Y, D, А или S
I51 заменен на I, Т, А или V
F54 заменен на F или L
S56 заменен на S и Т
Для экспериментов по скринингу аффинности каждую библиотеку дополнительно объединяли с соответствующей легкой или тяжелой цепью, имеющей трансплантированные CDR (например, библиотеку Н1/Н2 объединяли с легкой цепью, имеющей трансплантированные CDR), антитело экспрессировали и аффинность индивидуальных клонов по отношению к человеческому NGF подвергали скринингу при использовании системы BIAcore для поверхностного плазмонного резонанса (ППР) (BIAcore, Inc. Piscataway, NJ) в соответствии с инструкциями изготовителя и как описано выше. Определяли koff, kon и KD. Клоны антитела ранжировали на основании величин koff, поскольку обычно наибольшее варьирование аффинности заметно в величинах koff и, дополнительно, потому что величины koff не зависят от концентрации антител.
Определяли последовательность связываемых клонов, и последовательность клонов, которые связывались, показана в Таблице 3.
Связывание сохранили CDR, содержащие следующие замены:
CDR-H1
I34: связывание при наличии S, L, V, I и А.
N35: связывание при наличии N, Т и S.
CDR-H2
М50: связывание при наличии М, I, G, Q, S, L.
А62: связывание при наличии А и S.
L63: связывание при наличии L и V.
CDR-L1
S26: связывание при наличии S и F.
D28: связывание при наличии D, S, A, Y.
Н32: связывание при наличии Н, N, Q.
CDR-L2
Y50: связывание при наличии Y.
I51: связывание при наличии I, Т, V, А.
F54: связывание при наличии F.
S56: связывание при наличии S и Т.
CDR, содержащие следующие замены, были выбраны, в общем, на основании аффинности связывания и объединены в единый клон, обозначенный как H19-L129:
CDR-H1: I34L; N35N (нет изменений)
CDR-H2: M50I; А62А (нет изменений); L63V
CDR-L1: S26S (нет изменений); D28S; H32N
CDR-L2: Y50Y (нет изменений); I51Т; F54F (нет изменений); S56S (нет изменений).
Эти мутации объединяли (амплификацией Н- и L-цепей в ПЦР, разрезанием продуктов ПЦР и вектора (pRN8) рестрикционным ферментом и осуществлением 3-фрагментного лигирования) в единый клон, обозначенный как H19-L129, который также включал в себя трансплантированные CDR-H3 и L3. Последовательности вариабельных областей тяжелой цепи и легкой цепи H19-L129 показаны на Фиг.1А и 1Б, и Таблица 4 показывает аминокислотную последовательность CDR-L1, -L2, -Н1 и -Н2. H19-L129 связывали NGF при KD приблизительно 1 нМ, определенной при использовании анализа BIAcore, как описано здесь.
(4) Созревание аффинности CDR-H3 и L3. Созревание аффинности CDR-H3 и L3 осуществляли в две стадии. Сначала в процессе, называемом "сканирующий библиотечный мутагенез", каждый аминокислотный остаток в Н3 и L3 индивидуально подвергали предварительному скринингу для идентификации аминокислотных положений, в которых мутация приводит к повышенной аффинности связывания с человеческим NGF. На основании результатов сканирующего библиотечного мутагенеза (также называемого "анализ рандомизации малой библиотеки") выбирали подмножество аминокислотных положений в Н3 и L3 для получения библиотеки созревания аффинности и подвергали библиотеку созревания аффинности скринингу по аффинности к человеческому NGF при использовании анализа BIAcore, как описано здесь. Следует понимать, что эти методики могут быть общеупотребительными.
(а) Сканирующий библиотечный мутагенез
Каждое аминокислотное положение в CDR-H3 и -L3 индивидуально подвергали предварительному скринингу на замены, которые приводят к повышенной аффинности связывания с человеческим NGF. Частота аминокислотных замен в любом данном положении, которые приводят к улучшению связывания, к такому же связыванию, к худшему связыванию или к отсутствию связывания, дает информацию о положениях в CDR, которые можно заменять многими различными аминокислотами (в том числе всеми 20 аминокислотами), и о положениях в CDR, которые нельзя заменять или которые можно заменять лишь несколькими аминокислотами. Также идентифицировали аминокислотные замены, приводящие к повышенной аффинности связывания. На основании результатов этого скрининга выбирали подмножество аминокислотных положений в CDR-H3 и -L3 для получения библиотеки созревания аффинности.
Получали индивидуальные библиотеки Fab, в которых каждая аминокислота CDR-L3 и Н3 была рандомизирована во все 20 аминокислот, по одной за один раз, что приводило к нескольким малым библиотекам (5 библиотек для легкой цепи и 13 библиотек для тяжелой цепи), каждая с разнообразием возможностей для 20 аминокислот в каждом аминокислотном положении. Во всех случаях в библиотеке представлена нативная (то есть не измененная) аминокислота. Библиотеки получали кассетным ПЦР-мутагенезом с вырожденными олигонуклеотидами, как описано в Kay et al. (1996), Phage display of Peptides and Proteins: a laboratory manual, San Diego, Academic Press, при использовании допингового кодона NNK, чтобы рандомизировать одно аминокислотное положение так, чтобы оно включало 20 возможных аминокислот. Матрицей для конструирования библиотеки служило 8L2-6D5 (антитело с трансплантированными CDR, имеющее мутацию в каркасной области V71K), так как более низкая аффинность антитела с трансплантированными CDR делает возможным более легкое детектирование отличий по аффинности у мутантов по Н3 и L3 при скрининге. Таким образом, каждый член библиотеки содержал CDR3 (либо Н3, либо L3) с одной аминокислотной заменой и 5 трансплантированных CDR.
По 20-80 клонов из каждой малой библиотеки подвергали скринингу при использовании анализа BIAcore, как описано здесь. Пробы одновременно анализировали в BIAcore по аффинности связывания с NGF в одном канале чипа BIAcore и по присутствию Fab связыванием с антителом, имеющим пентагистидиновую метку, в другом канале сенсорного чипа для детектирования гистидиновой метки на С-конце тяжелой цепи. Клоны, которые экспрессировали белок, классифицировали как имеющие такую же аффинность, худшую аффинность, лучшую аффинность или отсутствие связывания при использовании koff для классификации. Результаты этого анализа показаны в Таблице 5.
Определяли последовательность всех клонов с улучшенной аффинностью, выявляя частоту и идентичность аминокислотных замен, которые приводят к повышенной аффинности. В дополнение, из каждой библиотеки выбирали несколько клонов, которые сохранили такую же аффинность, как клон 8I2-6D5, для того, чтобы уточнить замены в аминокислотной последовательности, которые допустимы в данном положении, даже если замена не обязательно увеличивает аффинность связывания. Результаты этого анализа суммированы в Таблице 6.
Несколько мутаций привели к повышенной аффинности связывания. По меньшей мере следующие мутации привели к значительно повышенной аффинности связывания при сравнении с матрицей 8L2-6D5: (H_Y101W (последовательность CDR GGYWYGTSYYFDY (SEQ ID NO:46)); H_S105A (последовательность CDR GGYYYGTAYYFDY (SEQ ID NO:47)); H_S105T (последовательность CDR GGYYYGTTYYFDY (SEQ ID NO:48)); H_G103A (последовательность CDR GGYYYATSYYFDY (SEQ ID NO:49) и L_S91E (последовательность CDR QQEKTLPYT (SEQ ID NO:50)).
Результаты этого эксперимента использовали, чтобы направлять селекцию аминокислотных положений для генерирования библиотеки созревания аффинности.
Этот эксперимент также дал информацию о частоте аминокислотных замен в любом данном положении, которые приводят к лучшему связыванию, такому же связыванию, худшему связыванию или отсутствию связывания, как показано в Таблице 5. Эта информация сделала возможной идентификацию аминокислотных положений в CDR, которые можно заменить многими различными аминокислотами (в том числе всеми 20 аминокислотами), и положений в CDR, которые можно заменить несколькими аминокислотами или очень малым числом аминокислот (в некоторых воплощениях таких аминокислот нет). Эти результаты также показали аминокислотные замены, которые повышают аффинность связывания.
(б) Созревание аффинности
Затем результаты анализа рандомизации малой библиотеки (см. выше) использовали для селекции остатков с целью получения библиотек H3 и L3 для созревания аффинности CDR-H3 и -L3. Остатки Y101 и G103 из CDR-H3 и остатки S91 и K92 из CDR-L3 были отобраны для получения библиотек Н3 и L3 для созревания аффинности Н3 и L3 CDR.
Эта библиотека объединяла мутации одновременно в Н3 и в L3 в клоне 8L2-6D5 с трансплантированными CDR и отдельно на фоне H19-L129 и имела разнообразие в 80 различных клонах. Таблица 7 показывает аминокислотные остатки, выбранные для замены, и аминокислоты, которые были заменены в каждом положении.
Таблица 7. Аминокислотные остатки в Н3 и L3, выбранные для замены, и аминокислоты, которые были заменены в каждом положении
CDR-H3:
Y101 заменен на Y и W, С. (Следует иметь в виду, что С был включен, так как применение кодона TRS в одном из вырожденных олигонуклеотидов также генерировало кодон для С.)
G103 заменен на А, Р, S.
CDR-L3:
S91 заменен на Е.
K92 заменен на все двадцать аминокислот. Связывание имело место при A, R, K и Н.
Каждый полипептид экспрессировали в виде Fab и аффинность по отношению к человеческому NGF у 96 индивидуальных клонов подвергали скринингу для каждой библиотеки при использовании анализа BIACORE в соответствии с инструкциями изготовителя и как описано выше. Результаты этого анализа показаны в Таблице 8.
На основании аффинности связывания наилучшие клоны, Е3 (взаимозаменяемым образом обозначаемый как "3Е") и 3С, выбрали для дополнительной характеризации. Е3 содержит в CDR следующие замены: CDR-H3: Y101W, G103A; и CDR-L3: S91E, K92Н, которые были объединены в единый клон, который также включал в себя следующие мутации: L1, L2, Н1 и Н2:
CDR-H1: I34L;
CDR-H2: M50I; L63V:
CDR-L1: D28S; H32N;
CDR-L2: I51Т.
Последовательности вариабельных областей тяжелой цепи и легкой цепи Е3 показаны на Фиг.1А и 1Б. 3С содержал в CDR следующие замены: CDR-L3: S91E; K92R; CDR-H3: Y101W; G103A, которые были объединены в единый клон, который также включал в себя мутации L1, L2, Н1 и Н2, описанные для клона 3Е.
Последовательности 3Е и 3С клонировали в экспрессионные векторы для млекопитающего для продукции Fab и полного антитела, экспрессировали в клетках НЕK293 и очищали при использовании хроматографии с Ni-NTA или протеином А. Чистый белок точно определяли количественным анализом аминокислот.
Аффинности связывания Fab Е3 и 3С с человеческим NGF измеряли при использовании анализа BIAcore в соответствии с инструкциями изготовителя и как описано выше за исключением того, что для предотвращения эффекта повторного связывания использовали 100 ЕО NGF на чипе. Вкратце, несколько концентраций антител (Fab) инъецировали в течение 2 минут на чип СМ5 с 100 RU иммобилизованного человеческого NGF на нем и давали для диссоциации 1800 секунд. Мышиное антитело 911 (Fab) анализировали в качестве контроля. Данные анализировали при использовании программного обеспечения BIAevaluation, следуя инструкциям изготовителя. Результаты анализа антител Е3 и 911 показаны на Фиг.9 и 10. Е3 связывало человеческий NGF при KD приблизительно 0,07 нМ (и при kon около 6,0е5 М-1 сек-1 и koff около 4,2е-5 сек-1). 3С связывало человеческий NGF при KD приблизительно 0,35 нМ (при koff около 1,4Е-5). Напротив, мышиное антитело 911 связывало NGF при KD 3,7 нМ, koff 8,4×10-5 сек-1 и kon 2,2×104 M1- сек-1.
Антитело Е3 (взаимозаменяемым образом обозначаемое как 3Е) выбрали для дальнейшего анализа на основании высокой аффинности связывания. Для тестирования способности Е3 предотвращать взаимодействие NGF с рецепторами NGF TrkA и р75 предварительно примешивали 2,5 нМ человеческого NGF и инкубировали в течение одного часа с 0-50 нМ антитела Е3 (Fab). После инкубации пробы инъецировали при 10 мкл/минута на чип СМ5 BIAcore, содержащий 260 RU р75 (канал 2) и 600 RU TrkA (канал 3) и определяли процент связывания. Результаты этого анализа показаны на Фиг.11. Повышенные концентрации Fab Е3 блокировали взаимодействие NGF с р75 и TrkA, как показано снижением сигнала (измеренного в RU), что указывает на то, что Fab Е3 блокирует взаимодействие человеческого NGF с TrkA и р75. Когда концентрация антитела Е3 (Fab) равна концентрации NGF (при концентрации NGF около 2,5 нМ), связывание NGF не наблюдали (как показано по нулевому сигналу). Из того факта, что нулевой процент связывания NGF с рецептором имел место, когда концентрация NGF была равна концентрации антитела 3Е, следует то, что 2,5 нМ NGF по меньшей мере в 10 раз выше, чем KD Е3 для NGF при равновесии.
Пример 2. Оценка способности антител против NGF блокировать NGF при использовании анализа выживания мышиных нейронов тройничного нерва Е13.5
Способность Fab Е3 или полного антитела Е3 блокировать активность NGF оценивали по измерению способности антител ингибировать зависимое от NGF выживание мышиных нейронов тройничного нерва Е13.5 in vitro. Тройничный ганглий состоит из кожных сенсорных нейронов, которые иннервируют лицевую область. Выживание мышиных нейронов тройничного нерва Е13.5 представляет собой чувствительный анализ для оценки блокирующей NGF активности антагонистических антител против NGF, так как NGF требуется для поддержания выживания этих нейронов. Например, при насыщающих концентрациях NGF выживание к 48 часам в культуре близко к 100%. Напротив, менее чем 5% нейронов выживают к 48 часам в отсутствие NGF.
Анализ выживания осуществляли следующим образом. Спаренных в определенное время беременных самок мышей Swiss Webster подвергали эвтаназии ингаляцией CO2. Рог матки удаляли и эмбрионов на эмбриональной стадии Е13.5 извлекали и декапитировали. Тройничные ганглии вырезали при использовании электролитически заостренных вольфрамовых игл. Эти ганглии затем трипсинизировали, механически диссоциировали и высевали при плотности 200-300 клеток на лунку в бессыворочной среде определенного состава в 96-луночных планшетах, покрытых поли-L-орнитином и ламинином.
Блокирующую активность Fab или антител против NGF оценивали, добавляя к нейронам тройничного нерва вариабельные дозы антител против NGF: моноклонального антитела 911 против NGF (Fab), 8L2-6D5, H19-L129, Е3 и 3С; и человеческий или крысиный NGF при следующих концентрациях: 0,4 нг/мл (~15 пМ; эта концентрация является насыщающей концентрацией NGF для выживания) и 0,04 нг/мл (~1,5 пМ; эта концентрация приблизительно соответствует IC50). После 48 часов в культуре клетки подвергали автоматической иммуноцитохимической процедуре, осуществляемой на аппарате для работы с жидкостями Biomek FX (Beckman Coulter) следующим образом: фиксация при использовании 4%-ного формальдегида, 5%-ной сахарозы и PBS; придание проницаемости при использовании 0,3%-ного Triton Х-100 в PBS; блокирование сайтов неспецифичного связывания при использовании 5%-ной нормальной козьей сыворотки, 0,11%-ного BSA в PBS и последовательная инкубация с первичными и вторичными антителами для детектирования нейронов. Первичным антителом было кроличье поликлональное антитело против белкового продукта гена 89.5 (PGP9.5, Chemicon), признанного маркера нейронального фенотипа. Вторичным антителом было козье Alexa Fluor 488 против кролика (Molecular Probes) вместе с красителем ядер Hoechst 33342 (Molecular Probes) для мечения ядер всех клеток, присутствующих в культуре. Получение изображения и анализ изображения осуществляли на визуализаторе Discovery-I/GenII (Universal Imaging Corporation). Изображения получали автоматически при двух длинах волн для Alexa Fluor 488 и Hoechst 33342, причем окрашивание ядер использовали как референтный показатель для основанной на изображении системы автофокусировки визуализатора, поскольку окрашивание ядер присутствует во всех лунках. Подходящие объективы и число визуализированных сайтов на лунку выбирали для охвата всей поверхности каждой лунки. Автоматический анализ изображений настраивали для подсчета числа нейронов, присутствующих в каждой лунке после 48 часов в культуре, на основании их специфичного окрашивания антителом против PGP9.5. Внимательное определение пороговых показателей изображения и применение селективного фильтра на основе морфологии и интенсивности флуоресценции приводили к точному подсчету нейронов в лунке.
Результаты этого эксперимента продемонстрировали, что Fab Е3 блокировал активность NGF с высокой аффинностью. Результаты показаны на Фиг.4-6 и в Таблице 9.
Фиг.4 представляет собой график, показывающий зависимое от NGF выживание нейронов Е13.5 в присутствии варьирующей концентрации человеческого и крысиного NGF.
Фиг.5 представляет собой график, сравнивающий эффект блокирования NGF различными Fab в присутствии либо человеческого NGF при 0,04 нг/мл (приблизительно 1,5 пМ; показано на нижней панели), либо человеческого NGF при 0,4 нг/мл (приблизительно 15 пМ; показано на верхней панели). 1,5 пМ NGF приблизительно соответствовало ЕС50 для NGF-стимулирующего выживания, в то время как 15 пМ представляло собой насыщающую концентрацию NGF. Выживание мышиных нейронов тройничного нерва Е13.5 при различных концентрациях Fab Е3, Fab мышиного 911 и Fab H19-L129 и Fab 8L2-6D5 оценивали, как описано выше. IC50 (в пМ) вычисляли для каждого Fab при каждой концентрации NGF и представили в Таблице 9. Fab Е3 сильно блокировал зависимое от человеческого NGF выживание нейронов тройничного нерва при IC50 приблизительно 21 пМ в присутствии 15 пМ человеческого NGF и IC50 приблизительно 1,2 пМ в присутствии 1,5 пМ человеческого NGF. Fab 3C и H19-L129 также сильно блокировали зависимое от человеческого NGF выживание нейронов тройничного нерва.
Фиг.6 представляет собой график, сравнивающий эффект блокирования NGF различными Fab в присутствии либо крысиного NGF при 0,04 нг/мл (приблизительно 1,5 пМ; показано на нижней панели), либо крысиного NGF при 0,4 нг/мл (приблизительно 15 пМ; показано на верхней панели). 1,5 пМ NGF приблизительно соответствовало EC50, в то время как 15 пМ представляло собой насыщающую концентрацию NGF. Выживание мышиных нейронов тройничного нерва Е13.5 при различных концентрациях Fab E3; мышиного Fab 911 и Fab H19-L129 и 8L2-6D5 оценивали, как описано выше. EC50 (в пМ) вычисляли для каждого Fab при каждой концентрации NGF и представили в Таблице 9. Fab E3 сильно блокировал зависимое от человеческого NGF выживание нейронов тройничного нерва: при IC50 приблизительно 31,6 пМ в присутствии 15 пМ крысиного NGF и при IC50 приблизительно 1,3 пМ в присутствии 1,5 пМ крысиного NGF. Fab 3C и H19-L129 также сильно блокировали зависимое от крысиного NGF выживание нейронов тройничного нерва.
В другом эксперименте авторы сравнили способность полного антитела Е3 и Fab 3Е ингибировать зависимое от NGF выживание нейронов Е13.5 в присутствии человеческого NGF при 0,4 нг/мл (насыщающая концентрация). Результаты этого анализа показаны на Фиг.12. Полное антитело Е3 и Fab 3Е показали одинаковые уровни ингибирования зависимого от NGF выживания, когда концентрации целого антитела и Fab были нормализованы относительно числа сайтов связывания NGF (Fab имеет один сайт связывания, а целое антитело имеет два сайта связывания). Эти результаты продемонстрировали отсутствие эффекта авидности из-за связывания полного антитела с димером NGF.
В других экспериментах авторы сравнили способность различных концентраций (20, 4, 0,8, 0,16, 0,032, 0,0064, 0,00128 и 0,0 нМ) антитела Е3, антитела 911 и иммуноадгезина TrkA-рецептора (состоящего из внеклеточного домена ТrkА-рецептора NGF, слитого с Fc-доменом иммуноглобулина, СН2-СН3) ингибировать зависимое от NGF выживание нейронов Е13.5 в присутствии 0,4 нг/мл (насыщающие условия). Эти результаты показаны на Фиг.13. Эти результаты продемонстрировали, что антитело Е3 блокировало NGF лучше, чем антитело 911 или иммуноадгезин TrkA.
Пример 3. Оценка специфичности антитела Е3 против NGF с использованием анализов выживания мышиных нейронов тройничного нерва и нодозных ганглиев
Способность антитела Е3 специфично блокировать активность NGF оценивали измерением способности этого антитела ингибировать выживание мышиных нейронов тройничного нерва Е17/18 in vitro в присутствии насыщающих концентраций NGF, родственного NGF нейротрофина NT3 или неродственного NGF нейротрофического фактора: белка, стимулирующего макрофаги (MSP). Выживание мышиных нейронов тройничного нерва Е17/18 представляет собой чувствительный анализ для оценки активности, блокирующей NGF, антагонистических антител против NGF, так как NGF требуется для поддержания выживания этих нейронов в более высоких концентрациях, чем уровень NGF, требуемый для поддержания выживания нейронов тройничного нерва Е13.5. Выживание этих нейронов также поддерживают NT3 или MSP; поэтому выживание этих нейронов также представляет собой чувствительный анализ для оценки того, блокирует ли антагонистическое антитело против NGF также и NT3 или MSP.
Способность антитела Е3 специфично блокировать активность NGF также оценивали измерением способности этого антитела ингибировать выживание мышиных нейронов нодозных ганглиев Е17 в присутствии насыщающих концентраций BDNF или NT4/5. Выживание нейронов нодозных ганглиев поддерживают BDNF или NT4/5, поэтому выживание этих нейронов представляет собой чувствительный анализ для оценки способности антагонистического антитела против NGF блокировать BDNF или NT4/5.
Анализ выживания осуществляли следующим образом. Спаренных в определенное время беременных самок мышей Swiss Webster подвергали эвтаназии ингаляцией CO2. Удаляли рог матки, экстрагировали эмбрионы (в день 17 или 18 эмбрионального развития) и декапитировали их. Вырезали и очищали тройничные и нодозные ганглии. Ганглии затем трипсинизировали, подвергали механической диссоциации и высевали при плотности 100-300 клеток на лунку в бессывороточной среде определенного состава в 4-луночные планшеты (Greiner), покрытые поли-L-орнитином и ламинином.
Нейроны тройничного нерва Е17/18 выращивали либо без добавления нейротрофических факторов (отрицательный контроль), либо в присутствии насыщающих концентраций человеческого NGF (400 пМ и 15 пМ) (положительный контроль), NT3 (400 пМ) или MSP (600 пМ). Делали по две параллельные культуры, которые включали в себя вариабельные концентрации Fab Е3 и 911 и полных антител. Концентрации Fab и полных антител указывали в расчете на сайт связывания (например, полное антитело содержит два сайта связывания, в то время как Fab содержит один сайт связывания).
Нейроны нодозных ганглиев Е17 выращивали либо без добавления нейротрофических факторов (отрицательный контроль), либо с насыщающими концентрациями BDNF (400 пМ) (положительный контроль), или NT4/5 (400 пМ), или неродственного NGF ростового фактора ILF (фактор ингибирования интерлейкинов). Использовали высокие концентрации нейротрофинов, так как цель этого эксперимента заключалась в том, чтобы протестировать специфичность антител. Делали по две параллельных культуры, которые включали в себя варьирование опять же с добавлением и без добавления антител Е3 и 911. После 48 часов в культуре суммарное число нейронов, выживающих в каждой лунке в каждых условиях, определяли подсчетом вручную, используя фазово-контрастный микроскоп.
Результаты этих экспериментов продемонстрировали, что антитела Е3 и 911 полностью блокировали эффекты NGF по стимулированию выживания нейронов тройничного нерва Е18. Напротив, антитела Е3 и 911 не влияли на выживание нейронов тройничного нерва, стимулированных NT3 или MSP, или на выживание нейронов нодозных ганглиев, стимулированных BDNF, или NT4/5, или LIF. Эти результаты продемонстрировали, что антитело Е3 обладало селективной специфичностью по отношению к NGF, так как не детектировалось взаимодействие между этими антителами и другими родственными NGF нейротрофинами (NT3, NT4/5, BDNF) при концентрациях, которые в 1000-10000 раз более высокие, чем эффективная концентрация для блокирования NGF. Дополнительно, эти результаты продемонстрировали, что гибель нейронов, наблюдаемая в культурах с добавленным NGF у нейронов, зависимых от NGF, при добавлении антител или Fab Е3, происходила из-за специфичного взаимодействия между этими антителами и NGF, а не из-за генерализованного токсичного эффекта. Также тестировали мышиное антагонистическое антитело 911 против NGF и наблюдали такие же результаты. Следует иметь в виду, что из-за высоких концентраций использованных нейротрофинов оба антитела, Е3 и 911, очень близки к условиям их титрования, и предполагалось, что они связываются с NGF при одинаковых уровнях, так как отличия в аффинности связывания этих антител с NGF в таких условиях должны быть менее заметными.
Результаты этих экспериментов показаны на Фиг.14, 15, 16 и 17. Эти данные показали средний процент выживания после 48 часов в культуре (Стандартная ошибка среднего, n=3 для каждой точки данных) относительно выживания, наблюдаемого в положительном контроле в каждом эксперименте (например, 100%-ное выживание нейронов тройничного нерва, выращиваемых в присутствии насыщающей концентрации NGF, и 100%-ное выживание нейронов нодозных ганглиев, выращиваемых в присутствии насыщающей концентрации BDNF соответственно). Фиг.14-15 представляют собой графики, показывающие, что антагонистическое антитело против NGF Е3 или Fab Е3 не ингибирует выживание, стимулированное NT3 и MSP, даже при столь высоких концентрациях антитела, как 200 нМ. Напротив, 20 нМ антитела Е3 или Fab 3Е и Fab 911 полностью блокировали выживание, которое вызывает NGF. Также тестировали мышиное антагонистическое антитело 911 против NGF и наблюдали такие же результаты. Конкретно, Фиг.14 представляет собой график, показывающий сравнение эффектов различных концентраций (20 нМ, 2 нМ или 0,2 нМ) Fab Е3 (на чертеже обозначено как "3Е") и Fab мышиного антитела 911 на выживание нейронов тройничного нерва Е18 в отсутствие добавленного нейротрофина (обозначено как "контроль"), 400 пМ NGF (обозначено как "NGF-400 пМ"), 10 нМ NT3 (обозначено как "NT3-10 нМ") или 600 пМ MSP (обозначено как "MSP-600 пМ"). Фиг.15 представляет собой график, показывающий сравнение влияния различных концентраций (200 нМ и 80 нМ) Fab и полного антитела Е3 и полного антитела и Fab мышиного антитела 911 на выживание нейронов тройничного нерва Е17 в отсутствие добавленных нейротрофинов (обозначено как "без фактора"), 400 пМ NGF (обозначено как "NGF-400 пМ"), 10 нМ NT3 (обозначено как "NT3-10 нМ") или 600 пМ MSP (обозначено как "MSP-600 пМ").
Фиг.16-17 представляют собой графики, показывающие, что антагонистическое антитело против NGF Е3 или Fab Е3 не ингибировал(о) выживание нейронов нодозных ганглиев Е17, стимулированное BDNF, NT4/5 или LIF. Также тестировали мышиное антагонистическое антитело 911 против NGF и наблюдали такие же результаты. Конкретно, Фиг.16 представляет собой график, показывающий сравнение влияний различных концентраций (200 нМ или 80 нМ) полного антитела Е3 (на чертеже обозначено как "3Е"), Fab Е3, полного антитела 911 или Fab 911 на выживание нейронов нодозных ганглиев Е17 в отсутствие добавленных нейротрофинов (обозначено как "без факторов"), 400 пМ BDNF (обозначено как "BDNF-400 пМ"), 400 пМ NT4/5 (обозначено как "NT4/5-400 пМ") или 2,5 нМ LIF (обозначено как "LIF-2,5 нМ"). Фиг.17 представляет собой график, показывающий сравнение влияний различных концентраций (200 нМ, 20 нМ, 2 нМ) Fab Е3 (на чертеже обозначено как "3Е") или Fab 911 на выживание нейронов нодозных ганглиев Е17 в отсутствие добавленных нейротрофинов (обозначено как "контроль"), 400 пМ BDNF (обозначено как "BDNF-400 пМ"), 400 пМ NT4/5 (обозначено как "NT4/5-400 пМ") или 2,5 нМ LIF (обозначено как "LIF-2,5 нМ").
Пример 4. Получение экспрессионных векторов для млекопитающего и экспрессия антитела Е3 в клетках млекопитающего
Для применения в экспрессии антитела Е3 в клетках млекопитающего разработали и сконструировали три экспрессионных вектора для млекопитающего.
Вектор Db.911.3E представляет собой экспрессионный вектор, содержащий вариабельную область тяжелой цепи антитела Е3 и константную область человеческого IgG2a, и подходит для временной или стабильной экспрессии тяжелой цепи. Db.911.3E состоит из нуклеотидных последовательностей, соответствующих следующим участкам: промоторный участок мышиного цитомегаловируса (нуклеотиды 1-612); синтетический интрон (нуклеотиды 619-1507); участок, кодирующий дигидрофолатредуктазу (DHFR, нуклеотиды 707-1267); сигнальный пептид человеческого гормона роста (нуклеотиды 1525-1602); вариабельная область тяжелой цепи антитела 3Е (нуклеотиды 1603-1965); константная область тяжелой цепи IgG2a человека, содержащая следующие мутации: А330Р331 в S330S331 (нумерация аминокислот в соответствии с последовательностью IgG2 дикого типа; см. Eur. J. Immunol. (1999) 29:2613-2624); поздний сигнал полиаденилирования SV40 (нуклеотиды 2974-3217); энхансерный участок SV40 (нуклеотиды 3218-3463); участок, представляющий собой фаг f1 (нуклеотиды 3551-4006), и участок, кодирующий бета-лактамазу (AmpR, нуклеотиды 4443-5300). Db.911.3E депонирован в АТСС 8 января 2003, и ему был присвоен номер АТСС №РТА-4895.
Вектор Eb.911.3E представляет собой экспрессионный вектор, содержащий вариабельную область легкой цепи антитела Е3 и константную область человеческой каппа-цепи, и подходит для временной экспрессии легкой цепи. Eb.911.3E состоит из нуклеотидных последовательностей, соответствующих следующим участкам: промоторный участок мышиного цитомегаловируса (нуклеотиды 1-612); человеческий интрон EF-1 (нуклеотиды 619-1142); сигнальный пептид человеческого гормона роста (нуклеотиды 1173-1150); вариабельная область легкой цепи антитела Е3 (нуклеотиды 1251-1571); константная область человеческой каппа-цепи (нуклеотиды 1572-1892); поздний сигнал полиаденилирования SV40 (нуклеотиды 1910-2153); энхансерный участок SV40 (нуклеотиды 2154-2399); участок, представляющий собой фаг f1 (нуклеотиды 2487-2942), и участок, кодирующий бета-лактамазу (AmpR, нуклеотиды 3379-4236). Eb.911.3E депонирован в АТСС 8 января 2003, и ему был присвоен номер АТСС № РТА-4893.
Вектор Eb.pur.911.3E представляет собой экспрессионный вектор, содержащий вариабельную область легкой цепи антитела Е3 и человеческую константную область-каппа, и подходит для стабильной экспрессии легкой цепи. Eb.pur.911.3E состоит из нуклеотидных последовательностей, соответствующих следующим участкам: промоторный участок мышиного цитомегаловируса (нуклеотиды 1-612); человеческий интрон EF-1 (нуклеотиды 619-1758); участок, кодирующий ген рас (puromycinR, нуклеотиды 739-1235); участок, представляющий собой 5'-нетранслируемый конец гена человеческого белка-70 теплового шока (hsp70 5'UTR, нуклеотиды 1771-1973); сигнальный пептид человеческого гормона роста (нуклеотиды 1985-2062); вариабельная область легкой цепи антитела Е3 (нуклеотиды 2063-2383); константная область человеческой каппа-цепи (нуклеотиды 2384-2704); поздний сигнал полиаденилирования SV40 (нуклеотиды 2722-2965); энхансерный участок SV40 (нуклеотиды 2966-3211); участок, представляющий собой фаг f1 (нуклеотиды 3299-3654), и участок, кодирующий бета-лактамазу (AmpR, нуклеотиды 4191-5048). Eb.pur.911.E3 депонирован в АТСС 8 января 2003, и ему был присвоен номер АТСС № РТА-4894.
Временную экспрессию в клетках осуществляли следующим образом. Клетки СНО и НЕK293Т в 150-мм чашках подвергали временной ко-трансфекции с 25 нг каждой плазмиды (то есть одной плазмиды, содержащей тяжелую цепь, и одной плазмиды, содержащей легкую цепь). ДНК смешивали с 100 мкл липофектамина-2000 (Invitrogen) в соответствии с инструкциями изготовителя. Комплексам ДНК-липид давали контактировать с клетками в среде DMEM/F12 без сыворотки или антибиотиков в течение 5 часов. После этой инкубации заменяли среды для экспрессии на Opti-MEM (Invitrogen) без каких-либо добавок в течение двух суток. Клеточные супернатанты, содержащие антитело, собирали последовательно до четырех раз с последующими заменами среды. Супернатанты очищали аффинной хроматографией при использовании смолы MapSelect с протеином А (Amersham biosciences 17-5199-02). Антитело связывалось с протеином А смолы в буфере, представляющем собой 0,3 М глицин, 0,6М NaCl при рН 8, затем его элюировали 0,1 М цитратным буфером при рН 3. Фракции, содержащие антитело, немедленно нейтрализовали 1 М трис-буфером при рН 8,0. Фракции антитела затем диализировали и концентрировали в PBS.
Пример 5. Антитело Е3 против NGF эффективно в лечении послеоперационной боли
Для оценки эффективности лечения антителом Е3 авторы использовали модель боли, которая имитирует послеоперационную боль. Антитело Е3 содержало константную область тяжелой цепи IgG2a человека, содержащую следующие мутации: А330Р331 в S330S331 (нумерация аминокислот соответствует последовательности IgG2 дикого типа; см. Eur. J. Immunol. (1999) 29:2613-2624); константную область человеческой легкой каппа-цепи и вариабельные области тяжелой и легкой цепи, показанные на фиг.1А и 1Б.
Животные. Самцов крыс Sprague Dawley с массой тела 220-240 г приобретали у Harlan (Wisconsin) и акклиматизировали к условиям содержания в течение одной недели перед хирургической операцией.
Хирургическая операция. Хирургическая операция была основана на процедуре, описанной в Brennan, et al. Pain 64:493-501 (1996). Животных анестезировали 2%-ным изофлураном в смеси с воздухом, которую подавали во время проведения хирургической операции через носовой конус. Подготавливали подошвенную поверхность правой задней лапы при использовании подушечки из йод-повидона и делали 1-см центральный продольный надрез через кожу и связки начиная с 0,5 см от края пятки и продолжая к кончикам пальцев. Измерения проводили линейкой, удерживая лапу в согнутом положении. Подошвенную мышцу приподнимали, используя изогнутый пинцет, и надрезали вдоль. Мышцу надрезали на полную глубину между началом и местом прикрепления. Кровотечение контролировали по ходу хирургической операции приложением давления через марлевый компресс. Рану зашивали двумя матрацными швами (черная хирургическая мононить, этилон 5-0). Эти швы завязывали 5-6 раз, первый узел делали слабым. Место раны промазывали раствором бацитрацина. Животным давали поправляться в покое в чистых клетках в течение двух часов или более до начала тестирования их поведения.
Оценка боли в покое. Для оценки боли, связанной с весовой нагрузкой, использовали кумулятивную балльную оценку боли. Животных помещали на пластиковую сетку (калибр: 8 мм2) в прозрачных пластиковых клетках, которые приподнимали на платформе (высота: 18 дюймов), что делало возможной проверку нижней стороны лап. После 20-минутного периода привыкания оценивали весовую нагрузку по шкале от 0 до 2 баллов. Балл 0 давали, если лапа была побелевшей или прижатой к сетке, что указывает на полную весовую нагрузку. Балл 1 давали, если кожа лапы предпочтительно только слегка касалась сетки без побеления или вдавления кожи. Балл 2 давали, если лапа была полностью вне контакта с сеткой. Подергивание лапой расценивали как 2, если крыса все еще была спокойной. Каждое животное наблюдали в течение 1 минуты через каждые 5 минут на протяжении 30 минут. Для оценки боли в порезанной ноге использовали сумму 6 баллов (0-12), полученных за время 1/2 часа. Также вычисляли частоту балла 2 и использовали для оценки частоты сильной боли или полного оберегания лапы животным. Каждое животное тестировали за 24 часа до хирургической операции (исходный уровень) и через 2 ч, 24 ч, 48 ч и 72 ч после операции. Результаты этого эксперимента показаны на Фиг.1, которая отражает кумулятивный балл боли в покое, который наблюдали у животных, которых лечили мышиным антителом 911 против NGF (35 мг/кг). Эти результаты продемонстрировали, что лечение антителом против NGF значительно снижает послеоперационную боль в покое. Весовая нагрузка была хорошим коррелятом того, как животное стремится использовать конечность, и поэтому было эффективным показателем облегчения боли.
Антитело Е3 инъецировали внутрибрюшинным путем (i.p.) при различных концентрациях антитела (0,004, 0,01, 0,02, 0,1, 0,6 и 1 мг на килограмм массы животного) за 15 часов до надреза. Группа отрицательного контроля получала не антитело, а инъекцию физиологического раствора внутрибрюшинно (i.p.). Фентанил при 0,01 мг/кг инъецировали i.p. в качестве положительного контроля за 30 минут до тестирования на момент времени 24 часа после хирургической операции. Каждый эксперимент включал по 8 животных (n=8 на группу) для каждого состояния, и в контрольной группе было 56 животных. Осуществляли хирургическую операцию и определяли кумулятивный балл боли, как описано выше. Боль в покое оценивали через двадцать четыре часа после хирургической операции.
Как показано на Фиг.7, гуманизированное антитело Е3 против NGF значительно снижало боль в покое (p<0,05) после хирургической операции при введении в дозе от 0,02 мг/кг до 1 мг/кг. Знак "*" обозначает достоверно значимое отличие от контроля (p<0,05). Лечение при использовании 0,02 мг/кг ослабляло болевое поведение по меньшей мере так же эффективно, как лечение при использовании фентанила (0,01 мг/кг). Эта доза фентанила представляет собой 10-кратную нормальную для человека дозу этого сильнодействующего опиоида.
В другом эксперименте тестировали эффективность антитела Е3 в снижении послеоперационной боли при введении после операции. Антитело Е3 (0,5 мг/кг) инъецировали внутривенно (i.v.) через два часа после хирургической операции. Контрольная группа получала не антитело, а внутривенную инъекцию физиологического раствора. Осуществляли хирургическую операцию и выражали боль в покое как кумулятивный балл боли, определенный через 24 часа после хирургической операции. Как показано на Фиг.8, лечение антителом против NGF значительно (p<0,05) снижало боль в покое на момент времени двадцать четыре часа после надреза, когда антитело вводили через 2 часа после надреза. Эти результаты продемонстрировали, что антитело Е3 эффективно облегчает послеоперационную боль при введении после хирургической операции.
Пример 6. Оценка анальгетических эффектов антагонистического антитела против NGF 911 в модели ревматоидного артрита на крысах
Анальгетические эффекты антитела 911 против NGF (см. Hongo et al., Hybridoma 19(3):215-227 (2000)) в сравнении с индометацином, использованным в качестве эталонного вещества, исследовали при использовании тестирования издаваемых звуков у крыс с хроническим артритом, индуцированным полным адъювантом Фрейнда (CFA).
В это исследование включили пятьдесят (50) самцов крыс Lewis (LEWIS LEW/Crl Ico) (Charles River, Бельгия) массой от 150 г до 220 г в начале экспериментальной фазы. Всех животных держали по меньшей мере 5 суток до эксперимента в помещении с контролируемой температурой (19,5-24,5°С), относительной влажностью (45-65%) и 12-часовым циклом света/темноты при неограниченном доступе к фильтрованной водопроводной воде и стандартному гранулированному лабораторному корму (U.A.R., Франция) на протяжении всего исследования. Животным делали индивидуальную метку на хвосте.
На сутки 0 (D0) у крыс индуцировали артрит инъекцией 0,05 мл суспензии Mycobacterium butyricum (Difco, США) в минеральном масле (10 мг/ мл) в кожу хвоста. На сутки 14 (D14) крыс с артритом включали в исследование в соответствии с их способностью издавать звуки при мягком сгибании задней лапы и по их индексу артрита, оцениваемому с использованием балльной шкалы воспаления для каждой задней и передней лапы (см. Kuzuna et al., Chem. Pharm. Bull. (Tokyo) 23:1184-1191 (1975); Pearson et al., Arthritis Rheum. 2:440-459 (1959)). Животным давали балльную оценку на основании следующих критериев: Балл 0: нормальный вид; Балл 1: эритема; Балл 2: эритема с легким отеком; Балл 3: сильное воспаление без анкилоза; Балл 4: анкилоз. В исследование включали только животных, способных издавать звуки при мягком сгибании и дающих балл 2 или 3.
В исследование включили четыре группы по 10 крыс в каждой. Для группы 1 (носитель) на сутки 14 (D14) после отбора крысам внутривенно вводили носитель (физиологический раствор). На сутки 18 (D18) интенсивность ноцицепции оценивали мягким сгибанием задней лапы и интенсивность уровня издаваемых звуков записывали для каждого животного. Для группы 2 (4 суток) в D14 после отбора крысам внутривенно вводили 911 (10 мг/кг). На сутки 18 (D18) интенсивность ноцицепции оценивали мягким сгибанием задней лапы и интенсивность уровня издаваемых звуков записывали для каждого животного. Для группы 3 (24 часа) на сутки 17 после инъекции CFA крысам внутривенно вводили 911 (10 мг/кг). Интенсивность ноцицепции оценивали мягким сгибанием задней лапы 24 часа спустя и интенсивность уровня издаваемых звуков записывали для каждого животного. Для группы 4 (индометацин) на сутки 18 (D18) интенсивность ноцицепции оценивали мягким сгибанием задней лапы через один час после перорального введения индометацина (10 мг/кг). Интенсивность уровня издаваемых звуков также записывали для каждого животного. Тестируемые вещества вводили слепым случайным образом внутривенным путем в объеме 5 мл/кг, тогда как индометацин вводили пероральным путем в объеме 10 мл/кг.
Анальгетические эффекты антитела 911 против NGF показаны в Таблице 10. Результаты выражали для каждой группы как интенсивность ноцицепции, оцениваемой по интенсивности уровня издаваемых звуков, записанной для каждого животного в мВ (среднее ± стандартная ошибка среднего), процент отличия интенсивности ноцицепции вычисляли относительно средней величины в группе, которой давали носитель. Статистическую достоверность между группами, получившими лечение, и группой, получившей носитель, определяли с помощью теста Даннетта (Dunnett) при использовании остаточной дисперсии в соответствии с однофакторным дисперсионным анализом (Р<0,05).
Как показано в Таблице 10, антитело 911 против NGF значительно снижало боль в модели ревматоидного артрита у крыс на момент времени 24 часа или 4 суток после разового введения антител.
Пример 7. Фармакологические эффекты антагонистического антитела Е3 и 911 против NGF в модели ревматоидного артрита у крыс
Фармакологические эффекты (противовоспалительные и анальгетические эффекты) антагонистического антитела против NGF Е3 и 911 в сравнении с индометацином, используемым в качестве вещества для внутреннего положительного контроля, исследовали в модели хронического артрита, индуцированного полным адъювантом Фрейнда (CFA), у крыс. Анальгетические эффекты Е3 и 911 оценивали измерением ноцицептивного ответа. Противовоспалительные эффекты оценивали по объему лапы, артритному индексу (балл воспаления) и массе тела и задних лап. Уровни цитокинов (IL-6, IL-1β, TNF-α и TGF-β1) в лапе, циркулирующего TGF-R1 в сыворотке, концентрации Е3 и 911 в плазме, биологические параметры и рентгенограммы получали в конце эксперимента.
Экспериментальный протокол
1. Схема исследования
В это исследование включили 80 самцов крыс Lewis (LEWIS Lew/Ico) (Charles River Laboratories - Бельгия) в возрасте 5 недель. Их держали в помещении с контролируемой температурой (19,5-24,5°С) и относительной влажностью (45-65%) с 12-часовым циклом свет/темнота и при неограниченном доступе к фильтрованной водопроводной воде и стандартному гранулированному лабораторному корму (SAFE, Франция) на протяжении всего исследования. По получении в виварии их держали по 5 в клетке и перед любым тестированием давали 10-дневный период для акклиматизации. Животным делали индивидуальные метки на хвосте.
В это исследование включили пять групп по 10 животных в каждой (5-недельные самцы крыс Lewis - LEWIS Lew/Ico из Charles River Laboratories - Бельгия): Группа 1: крысы без артрита / физиологический раствор (носитель), болюсная инъекция внутривенно, n=10; Группа 2: крысы с артритом / физиологический раствор (носитель), болюсная инъекция внутривенно, n=10; Группа 3: крысы с артритом / индометацин 3 мг/кг, перорально ежедневно в течение 10 суток, n=10; Группа 4: крысы с артритом / Е3, 1 мг/кг, болюсная инъекция внутривенно, n=10; Группа 5: крысы с артритом / 911, 10 мг/кг, болюсная инъекция внутривенно, n=10. Дозы выражали по свободному активному веществу (мг/кг). Е3 и 911 приготавливали экстемпорально в физиологическом растворе из маточного раствора до желаемой концентрации. Е3, 1 мг/кг: 3,41 мл маточного раствора (0,88 мг/мл) с физиологическим раствором в достаточном количестве до 15 мл. 911, 10 мг/кг: 12 мл маточного раствора (2,5 мг/мл) с физиологическим раствором в достаточном количестве до 15 мл. Все разбавленные растворы (до внутривенной инъекции) стерилизовали при использовании установки со стерильным 0,20-мкм фильтром. Величины рН и осмолярности разбавленных растворов измеряли перед каждой внутривенной инъекцией. Перед первой внутривенной инъекцией осмолярности (мосм/л) для физиологического раствора, Е3 и 911 были 278, 269 и 308 соответственно; рН для физиологического раствора, Е3 и 911 были 5,93, 6,76 и 6,71 соответственно. Перед второй внутривенной инъекцией осмолярности (мосм/л) для физиологического раствора, Е3 и 911 были 280, 270 и 309, соответственно; рН для физиологического раствора, Е3 и 911 были 5,86, 6,72 и 6,59 соответственно.
Е3 или 911 либо физиологический раствор вводили болюсной инъекцией внутривенно в объеме 5 мл/кг на сутки 14 и сутки 19 после индукции артрита в закодированном случайном порядке. В группе, не имеющей артрит, давали физиологический раствор внутривенной болюсной инъекцией в объеме 5 мл/кг на сутки 14 и сутки 19. Индометацин приготавливали экстемпорально в 1%-ной метилцеллюлозе. Индометацин вводили в объеме 10 мл/кг пероральным путем (перорально) один раз в день в течение 10 суток начиная с суток 14 по сутки 23 после индукции артрита в закодированном случайном порядке.
2. Индукция артрита
На сутки 0 (D0) артрит индуцировали у 70 крыс инъекцией 0,05 мл суспензии Mycobacterium butyricum в кожу хвоста. Группа из 10 крыс не получала никакой инъекции в кожу (крысы без артрита). На сутки 14 (D14) крыс с артритом включали в исследование при использовании следующих критериев: все включенные крысы проявляли увеличение среднего объема лап (среднего объема левой и правой лапы) по меньшей мере на 0,30 мл при сравнении со средним объемом лап (средний объем левой и правой лапы) в группе без артрита (объем лап измеряли, как описано ниже); все включенные крысы издавали звуки при мягком сгибании (ноцицептивный ответ измеряли, как описано ниже); и все включенные крысы проявляли балл индекса артрита 2-3 по каждой задней лапе (индекс артрита измеряли, как описано ниже) (животных с баллом 0, 1 или 4 отбрасывали).
3. Масса тела
Животных взвешивали один раз в день начиная от суток 0 по сутки 24 (за исключением выходных дней перед лечением: D1, D2, D8, D9, D10). Все измерения осуществляли между 9:00 и 12:00 за исключением D14 (7:30-9:00) и D24 (7:30-8:00 утра).
3. Измерение объема лапы
Объем правой и левой задней лапы каждой крысы (крысы с артритом и крысы без артрита) измеряли при использовании плетизмометра. Измерения осуществляли в следующие периоды времени (после индукции артрита): сутки 14 (до внутривенной болюсной инъекции или перорального введения) и сутки 24 (5 суток после последней внутривенной болюсной инъекции или через 24 часа после последнего перорального введения). Все измерения осуществляли между 9:00 и 12:00. Все данные собирали и хранили при использовании программного обеспечения WinDas.
4. Индекс артрита
Индекс артрита оценивали при использовании балльной оценки воспаления для каждой задней и передней лапы (крысы с артритом): Балл 0: нормальный вид; Балл 1: эритема; Балл 2: эритема и легкий отек; Балл 3: сильное воспаление без анкилоза; Балл 4: анкилоз. Эти оценки делали в следующие периоды времени (после индукции артрита): сутки 14 (до внутривенной болюсной инъекции или перорального введения) и сутки 24 (5 суток после последней внутривенной болюсной инъекции или через 24 часа после последнего перорального введения). Все измерения делали между 2:00 и 3:00 дня (D14), 8:00 и 9:00 утра (D24). Все данные собирали и хранили при использовании программного обеспечения WinDas.
5. Измерение ноцицептивного ответа (тестирование издаваемых звуков)
Ноцицептивный ответ оценивали мягким сгибанием правой и левой задней лапы повторно 2 раза с интервалами от 4 до 5 сек пальцем оператора (крысы с артритом). Интенсивность уровня издаваемых звуков записывали для каждого животного для каждой задней лапы (2 раза: на правой задней лапе: s1 и s3; 2 раза: на левой задней лапе: s2 и s4). Эту оценку делали в следующие периоды времени (после индукции артрита): сутки 14 (до внутривенной болюсной инъекции или перорального введения); сутки 18 (до второй внутривенной болюсной инъекции или через 1 час после перорального введения) и сутки 24 (5 суток после последней внутривенной болюсной инъекции или через 24 часа после последнего перорального введения). Все измерения делали между 9:00 и 12:00 за исключением D14 (7:30-9:00) и D24 (7:30-9:00).
6. Сбор крови для измерения концентрации Е3 или 911 и циркулирующего TGF-β1 и гематологических параметров
На сутки 24 (после измерений объема лапы и индекса артрита и тестирования издаваемых звуков) под общей анестезией при использовании изофлурана (в смеси кислорода и закиси азота) пробы крови (около 800-1000 мкл) собирали из ретроорбитального синуса капиллярным действием микропипетки.
Измерение концентрации Е3 или 911 (группы 2, 4 и 5): часть образца крови собирали в пробирки, содержащие Li-гепарин (держали на льду) и центрифугировали при 2500-3000g в течение 10 мин. Получали образцы плазмы (по меньшей мере 100 мкл), замораживали их в жидком азоте и хранили при -80°С. Один образец был слегка гемолизированным (крыса с артритом #36, получившая носитель).
Измерение циркулирующего TGF-β1 (группы 1-2-3-4-5): часть образца крови собирали в микропробирки для получения сыворотки при температуре окружающей среды. После получения образца кровь перемешивали и давали ей свертываться в течение 30 минут перед центрифугированием. Пробирки центрифугировали при приблизительно 6000g в течение 3 минут. Каждый образец сыворотки (по меньшей мере 100 мкл, за исключением крыс #52 и #53) делии на аликвоты и хранили при -20°С до активирования образца для анализа TGF-β1. Эти аликвоты (50 флаконов) держали в течение 6 месяцев начиная с конца исследования. Некоторые образцы были слегка гемолизированными (крысы без артрита #2, #5, #9 и #10, получившие носитель; крысы с артритом #53 и #63, получившие носитель; крысы с артритом #31 и #51, получившие Е3; крысы с артритом #52, #62 и #64, получившие 911). Уровни TGF-β1 измеряли при использовании набора для ELISA на человеческий TGF-β1 (номер по каталогу DB100, партия 212258 и 213610, R&D Systems - Франция).
Сбор крови для гематологических параметров (группы 1-2-3-4-5: 50 флаконов): часть образца крови собирали в пробирки, содержащие K3-ЭДТА (по меньшей мере 100 мкл). Определение параметров осуществляли в день сбора и образцы не хранили. Гематологические параметры включали в себя эритроциты, лейкоциты, тромбоциты, гемоглобин, гематокрит, определяемые гематологическим счетчиком клеток (D24). Некоторые гематологические параметры не измеряли из-за свернувшихся проб (крыса без артрита #10, получившая носитель; крысы с артритом #59 и #67, получившие Е3; крыса с артритом #16, получившая 911).
7. Уровни цитокинов в лапе
На сутки 24 (5 суток после последней внутривенной болюсной инъекции или через 24 часа после последнего перорального введения) (после рентгеновской радиографии) каждую заднюю лапу животного (крысы с артритом и без артрита) взвешивали и помещали в помеченный полиэтиленовый флакон. Образцы ткани замораживали в жидком азоте и хранили при -80°С.
Получение гомогенатов суставов. Замороженные задние лапы распыляли при использовании Bio-Pulverizer. Порошки задних лап затем помещали в 50-мл коническую центрифужную пробирку, содержащую 3 мл PBS с добавленными 50 мкл антипротеазной смеси и гомогенизировали на льду при использовании гомогенизатора Ultra-Turrax (50% максимальной скорости). Гомогенат затем центрифугировали при 2000g в течение 15 минут при 4°С и супернатанты фильтровали через 0,2-мкм фильтры Sartorius, делили на аликвоты и хранили при -80°С до использования.
Измерение уровней цитокинов. Уровни цитокинов TNF-α (набор для ELISA на крысиный TNF-α, номер по каталогу RTA00, партия 213718, R&D Systems, Франция), IL-β (набор для ELISA на крысиный IL-1β, номер по каталогу RLB00, партия 212435, R&D Systems, Франция), IL-6 (набор для ELISA на крысиный IL-6, номер по каталогу R6000, партия 211773, 214008 и 214362, R&D Systems, Франция) и TGF-R1 (набор для ELISA на человеческий TGF-β1, номер по каталогу DB100, партия 212258 и 213610, R&D Systems, Франция) определяли в двух параллельных опытах в соответствии с процедурой изготовителей. Аликвоты гомогенатов задней лапы хранили при -80°С.
8. Рентгеновский анализ
На сутки 24 после сбора крови животных забивали и получали рентгенограммы (задних лап) для оценки патологических изменений сустава. Рентгеновский анализ фокусировали на суставной эрозии, суставном пространстве, аномалии надкостницы в обеих задних лапах. Все рентгенограммы анализировали наблюдением семи разных аспектов: повреждение мягкой ткани, деформация, деминерализация, суставное пространство, эрозии, остеогенез и реакция надкостницы. Для каждого животного первые шесть аспектов анализировали независимо наблюдением наихудшей задней ноги. Реакцию надкостницы анализировали наблюдением хвоста. Для каждого аспекта балл составляет от 0 (норма) до 4 (максимальное повреждение). Поэтому суммарный балл составляет от 0 до 28. Рентгенограммы интерпретировал один и тот же наблюдатель без знания чего-либо о животных (леченых или нелеченых).
10. Наблюдения
Одно животное (#65) умерло в D23 после введения индометацина (до введения в D23) по неизвестной причине.
11. Анализ и представление результатов
Все результаты были представлены в виде среднего значения ± стандартная ошибка среднего (S.E.M.) для 10 крыс в каждой группе в каждой точке времени. Объем лапы выражали в мл, вычисленных из средней величины по объему правой и левой лапы. Индекс артрита вычисляли из суммы баллов, полученных для каждой из 4 лап. Ноцицептивный ответ оценивали по интенсивности уровня издаваемых звуков, записанного для каждого животного (среднее по 4 величинам: 2 момента времени/лапы) в мВ. Процент ингибирования ноцицептивного ответа вычисляли из средней величины для группы с артритом, получившей носитель [(средняя величина для группы с артритом, получившей носитель - средняя величина для группы с артритом, получившей лечение / средняя величина для группы с артритом, получившей носитель)×100]. Массу тела выражали в граммах. Массу задних лап (левой и правой) выражали в граммах. Уровни цитокинов (IL-6, IL-1β, TNF-α и TGF-α1) в каждой задней лапе выражали в пг/мл. Циркулирующие уровни TGF-β1 выражали в пг/мл. Рентгенологический индекс для каждого параметра (деминерализация, эрозии, реакция надкостницы, повреждение мягких тканей, суставное пространство, деформация, остеогенез) и суммарный рентгенологический индекс (суммарный балл) вычисляли из суммы баллов, полученных для каждого параметра. Межгрупповые достоверности различий между величинами для группы, получившей носитель (крысы с артритом), и группы, получившей носитель (крысы без артрита), оценивали по t-критерию Стьюдента или критерию Манна-Уитни для суммы рангов, когда критерий равной дисперсии или нормальности не соблюдался. Межгрупповые достоверности различий между величинами для группы, получившей носитель (крысы с артритом), и групп, получивших Е3, 911 и индометацин, оценивали при использовании однофакторного дисперсионного анализа ANOVA с последующим неспаренным t-критерием Даннетта. Вероятность Р≤0,05 считали признаком достоверности. Весь статистический анализ осуществляли при использовании программного обеспечения Sigmastat™.
Результаты
1. Ноцицептивный ответ (тестирование издаваемых звуков)
Как показано в Таблице 11 и на Фиг.18, в D14 ноцицептивный ответ составлял 4147±331, 4386±235, 4644±367 и 4468±143 в группах с артритом, получивших носитель, индометацин, Е3 и 911 соответственно. Индометацин сильно и достоверно снижал ноцицептивный ответ после введения 3 мг/кг/сутки перорально (в течение 10 суток) приблизительно на -3768 мВ (% ингибирования: 71%) и -4353 мВ (% ингибирования: 74%) в D18 и D24 соответственно при сравнении с группой с артритом, получившей носитель (D18: 1511±398 против 5279±326 мВ; D24: 1552±508 против 5905±345 мВ). Е3 (1 мг/кг внутривенно в D14 и D19) сильно и достоверно снижало ноцицептивный ответ приблизительно на -4167 мВ (% ингибирования: 79%) и -5905 мВ (% ингибирования: 100%) в D18 и D24 соответственно при сравнении с группой с артритом, получившей носитель (D18: 1112±401 против 5279±326 мВ; D24: 0±0 против 5905±345 мВ). 911 (10 мг/кг внутривенно 2 суток в D14 и D19) сильно и достоверно снижало ноцицептивный ответ приблизительно на -3932 (% ингибирования: 74%) и -5358 мВ (% ингибирования: 91%) в D18 и D24 соответственно при сравнении с группой с артритом, получившей носитель (D18: 1347±492 против 5279±326 мВ; D24: 547±307 против 5905±345 мВ).
2. Масса тела
Как показано в Таблице 12 и на Фиг.19, заметное снижение прибавки массы тела наблюдали у крыс с артритом в сравнении с крысами без артрита с D0 по D14 из-за развития артрита. В D14 (день отбора) крысы с артритом проявляли значительное снижение массы при сравнении с крысами без артрита (289±2 против 217±4 г) (t-критерий Стьюдента Р<0,05). Однако отсутствие значимого отличия по массе (D14) детектировали во всех группах с артритом (t-критерий Даннетта Р>0,05). Масса тела умеренно и достоверно повышалась в группе, получившей индометацин (3 мг/кг/сутки в течение 10 дней), с D17 по D24 с максимумом около 43 г в D24 при сравнении с группой с артритом, получившей носитель (261±5 против 218±3 г). После лечения при использовании ЕЗ (1 мг/кг внутривенно в D14 и D19) масса тела умеренно и достоверно повышалась с D17 по D24 с максимумом около 46 г в D24 при сравнении с группой с артритом, получившей носитель (264±5 г против 218±3 г). После лечения при использовании 911 (10 мг/кг внутривенно в D14 и D19) масса тела умеренно и достоверно повышалась с D18 по D24 с максимумом около 47 г в D24 при сравнении с группой с артритом, получившей носитель (265±7 против 218±3 г).
3. Объем лап
В D14 осуществляли рандомизацию с целью получения групп, гомогенных по объему лап. Как показано в Таблице 13, в D14 объем задних лап (средний объем правой и левой лапы) был значительно больше в группе с артритом, чем в группе без артрита (2,10±0,05 против 1,44±0,02 мл (t-критерий Стьюдента Р≤0,05)). Индометацин (3 мг/кг/сутки перорально в течение 10 суток) достоверно снижал объем лап приблизительно на -0,75 мл (D24) при сравнении с группой, получившей носитель (1,59±0,03 мл против 2,34±0,08 мл). Е3 (1 мг/кг внутривенно в D14 и D19) слегка и достоверно повышало объем лап приблизительно на 0,37 мл при сравнении с группой с артритом, получившей носитель (2,71±0,09 мл против 2,34±0,08 мл). 911 (10 мг/кг внутривенно в D14 и D19) слегка и достоверно повышало объем лап приблизительно на 0,36 мл при сравнении с группой с артритом, получившей носитель (2,70±0,11 мл против 2,34±0,08 мл).
4. Индекс артрита
Как показано в Таблице 14, индекс артрита в D 14 составлял 10,1±0,8, 8,7±0,6, 10,2±0,4 и 9,4±0,7 в группах с артритом, получивших носитель, индометацин, Е3 и 911 соответственно. Индометацин сильно и достоверно снижал индекс артрита после введения 3 мг/кг/сутки перорально (в течение 10 суток) максимум приблизительно на -8,0 при сравнении с группой с артритом, получившей носитель (2,7±0,7 против 10,7±0,6). Е3 (1 мг/кг внутривенно в D14 и D19) не влияло на индекс артрита при сравнении с группой с артритом, получившей носитель (11,4±0,4 против 10,7±0,6). 911 (10 мг/кг внутривенно в D14 и D19) не влияло на индекс артрита при сравнении с группой с артритом, получившей носитель (10,9±0,7 против 10,7±0,6).
5. Уровни цитокинов в лапе
Как показано в Таблице 15, в D24 уровни цитокинов в левой и правой лапе повышены в группе с артритом, получившей носитель, максимум приблизительно в 3,5 раза (IL-1β), в 4 раза (TNF-α) и в 1,8 раза (TGF-β1) при сравнении с группой без артрита, получившей носитель. Достоверных различий между этими двумя группами не наблюдали по уровням IL-6 в правой и левой лапе. Уровни цитокинов в группе с артритом были одинаковыми в левой и правой лапе: 259,7±38,5 против 219,2±32,4, 4802,8±365,5 против 4007,1±380,4, 17,8±1,6 против 18,6±1,9 и 9735,0±1219,8 против 9161,4±846,1 пг/мл для IL-6, IL-1β, TNF-α и TGF-β1 соответственно. Индометацин слегка, но достоверно снижал уровень TGF-β1 в правой лапе после введения 3 мг/кг/сутки перорально (в течение 10 суток) примерно в 1,3 раза при сравнении с группой с артритом, получившей носитель (7057,4±335,6 против 9161,4±846,1), и при этом он не модифицировал уровни IL-6, TNF-α или IL-1β. Такой же, но не достоверный эффект наблюдали в левой лапе. Е3 (1 мг/кг внутривенно в D14 и D19) не влияло на уровни IL-6, IL-1β, TNF-α или TGF-β1 в обеих лапах при сравнении с группой с артритом, получившей носитель. 911 (10 мг/кг внутривенно в D14 и D19) повышало уровень IL-1β в правой лапе при сравнении с группой с артритом, получившей носитель (6215,3±666,7 против 4007,1±380,4). Оно не влияло на уровни других цитокинов в обеих лапах.
6. Измерение циркулирующего TGF-β1
Как показано в Таблице 16, в D24 уровни TGF-β1 в сыворотке были повышены в группе с артритом, получившей носитель, при сравнении с группой без артрита, получившей носитель (81715,7±1984,1 против 60269,9±2142,8). Индометацин достоверно снижал уровень TGF-β1 в сыворотке после введения 3 мг/кг/сутки перорально (в течение 10 суток) примерно в 1,5 раза при сравнении с группой с артритом, получившей носитель (57222,2±3194,1 против 81715,7±1984,1). Е3 (1 мг/кг внутривенно в D14 и D19) и 911 (10 мг/кг внутривенно в D14 и D19) значительно снижали уровень TGF-β1 в сыворотке, так что уровни цитокина в группах, получивших Е3 и 911, были сравнимы с теми, которые наблюдались в группе без артрита, получившей носитель (69408,8±3926,7 и 67214,5±3649,4 соответственно против 60269,9±2142,8).
7. Гематологические параметры
Как показано в Таблице 17, гематологические параметры, такие как лейкоциты и тромбоциты, были больше у крыс с артритом, получивших носитель, в сравнении с крысами без артрита, получившими носитель (t-критерий Стьюдента Р<0,05), тогда как эритроциты, гемоглобин и гематокрит (t-критерий Стьюдента Р>0,05) не изменились. Индометацин не влиял на параметры крови после введения 3 мг/кг/сутки перорально (в течение 10 суток) при сравнении с группой с артритом, получившей носитель. Е3 (1 мг/кг внутривенно в D14 и D19) не влияло на параметры крови при сравнении с группой с артритом, получившей носитель. 911 (10 мг/кг внутривенно в D14 и D19) не влияло на параметры крови при сравнении с группой с артритом, получившей носитель.

7. Масса задних лап
Как показано в Таблице 18, масса левой и правой задней лапы была больше у крыс с артритом, получивших носитель, чем у крыс без артрита, получивших носитель (3,43±0,11 против 1,98±0,01 и 3,32±0,12 против 1,99±0,02 г соответственно) (t-критерий Стьюдента или Манна-Уитни Р<0,05). Индометацин достоверно снижал массу задних лап после введения 3 мг/кг/сутки перорально (в течение 10 суток) при сравнении с группой с артритом, получившей носитель (левая задняя лапа: 2,23±0,04 против 3,43±0,11 г; правая задняя лапа: 2,20±0,05 против 3,32±0,12 г). Е3 (1 мг/кг внутривенно в D14 и D19) достоверно повышало массу только левой задней лапы при сравнении с группой с артритом, получившей носитель (левая задняя лапа: 3,86±0,14 против 3,43±0,11 г; правая задняя лапа: 3,72±0,13 против 3,32±0,12 г). 911 (10 мг/кг внутривенно в D14 и D19) достоверно повышало массу только правой задней лапы при сравнении с группой с артритом, получившей носитель (левая задняя лапа: 3,73±0,12 против 3,43±0,11 г; правая задняя лапа: 3,83±0,15 против 3,32±0,12 г).
8. Рентгеновский анализ
Как показано в Таблице 19, у крыс без артрита, получивших носитель, наблюдали суммарный балл 0,0±0,0. Крысы с артритом, получившие носитель, имели суммарный балл 15,1±1,3 с высокими баллами по деминерализации (2,4±0,3), эрозии (2,7±0,3), повреждению мягких тканей (3,1±0,2) и суставному пространству (3,3±0,2) и умеренным баллом по реакции надкостницы (1,0±0,3), остеогенезу (0,8±0,2) и деформации (1,8±0,2). Индометацин (3 мг/кг/сутки перорально в течение 10 суток) сильно и достоверно снижал суммарный балл приблизительно на 10,7 при сравнении с крысами с артритом, получившими носитель (4,4±0,9 против 15,1±1,3). Е3 (1 мг/кг внутривенно в D14 и D19) не влияло на суммарный балл при сравнении с группой с артритом, получившей носитель (14,2±1,3 против 15,1±1,3). 911 (10 мг/кг внутривенно в D14 и D19) не влияло на суммарный балл при сравнении с группой с артритом, получившей носитель (15,4±1,0 против 15,1±1,3).
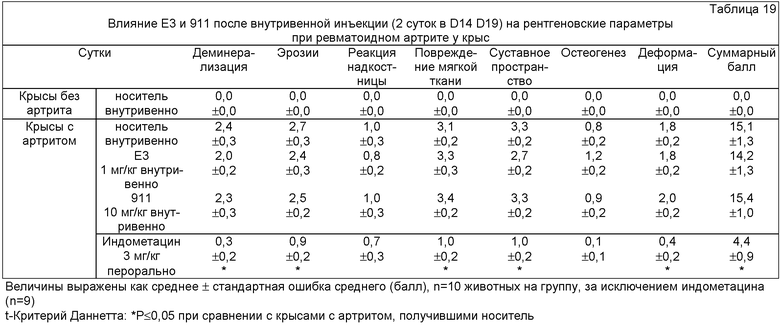
Заключение
В экспериментальных условиях, описанных выше, Е3 (1 мг/кг внутривенно 2 суток: D14-D19) и 911 (10 мг/кг внутривенно 2 суток: D14-D19) в этой модели артрита показали сильные анальгетические эффекты, но не показали значительных противовоспалительных эффектов.
Пример 8. Влияние разных доз антитела Е3 против NGF в модели ревматоидного артрита у крыс
Способность Е3 вызывать снижение боли у крыс с артритом дополнительно испытывали путем исследования зависимости доза-ответ между введением Е3 и снижением боли. Крыс обрабатывали адъювантом для индукции артрита, как описано выше. Десять крыс, которым не инъецировали адъювант, использовали в качестве контролей без артрита. Через четырнадцать суток после инъекции адъюванта животных отбирали в исследование на основании критериев, указанных выше, рандомизировали в восемь групп по десять крыс и тестировали на интенсивность их ответа в виде издаваемых звуков. Затем им вводили на сутки 14 физиологический раствор или 0,003 мг/кг, 0,01 мг/кг, 0,03 мг/кг, 0,1 мг/кг, 0,3 мг/кг, 1 мг/кг или 5 мг/кг антитела Е3, как описано выше. Животных тестировали на ответ в виде издаваемых звуков на сутки 16, 18, 20 и 24. Животным снова вводили физиологический раствор или такую же дозу Е3 на сутки 18 после тестирования на издаваемые звуки. Животных также взвешивали каждые сутки начиная с суток 14. Таким образом, животным дважды вводили данную дозу антитела или физиологический раствор на сутки 14 и 18 и оценивали на боль пять раз на сутки 14, 16, 18, 20 и 24. Данные показаны в Таблицах 20-22 и на Фиг.20-22.

Влияние лечения животных различными дозами антитела Е3 против NGF на индуцированные болью издаваемые звуки (данные показаны в Таблице 20) статистически анализировали при использовании двухстороннего вариационного анализа для сравнения результатов, полученных попарно между животными с артритом, которым давали носитель, и теми, которым давали указанную дозу антитела Е3. При всех тестированных уровнях Е3 имел место сильный достоверный эффект (p<0,0001). Даже при самой низкой тестированной дозе (0,003 мг/кг Е3) различия в издаваемых звуках были достоверными (р<0,0001).
Как показано в Таблице 20 и на Фиг.20, в соответствии с указанными экспериментами лечение антителом Е3 при 1 мг/кг показало быстрое и устойчивое облегчение боли. В пределах двух суток (самая ранняя временная точка тестирования) интенсивность издаваемых звуков упала на 90%. Лечение более низкими концентрациями Е3 также давало устойчивое облегчение боли, хотя при самых низких дозах облегчение боли требовало несколько больше времени для проявления. Вероятно, кажущееся снижение эффективности на сутки 24 у всех, кроме самых высоких тестированных доз, происходит из-за уменьшения реального уровня Е3 в плазме, вторичного по отношению к иммунному ответу у субъекта крысы. Очевидно, что столь низкие дозы, как 0,003 мг/кг, дают в этой модели по меньшей мере частичное облегчение боли.
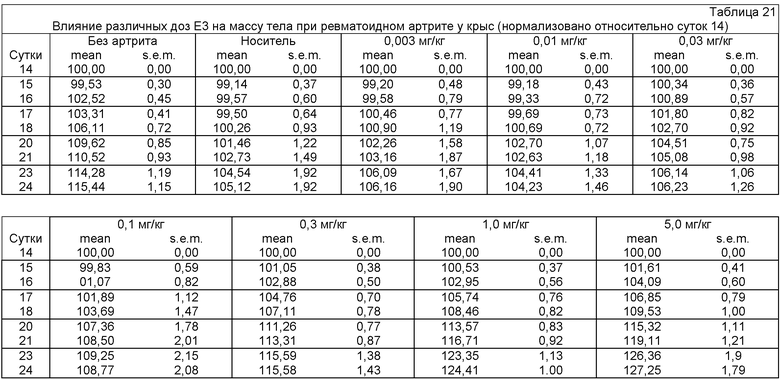

Влияние лечения животных различными дозами антитела Е3 против NGF на массу тела статистически анализировали при использовании двухстороннего дисперсионного анализа ANOVA для сравнения результатов, полученных попарно, между животными с артритом, которым давали носитель, и теми, которых лечили указанной дозой антитела Е3. При использовании данных, нормализованных относительно массы на сутки 14 (Таблица 21), дозы 0,03 мг/кг Е3 приводили к достоверному изменению массы тела (p<0,005). При всех высоких дозах Е3 различие между лечеными и нелечеными животными с артритом было достоверным (p= или <0,0001). При использовании данных, нормализованных относительно массы на сутки 0 (Таблица 22), доза 0,03 мг/кг Е3 приводила к значительному изменению массы тела (p<0,002). При всех более высоких дозах Е3 различие между лечеными и нелечеными животными с артритом было достоверным (p<0,0001).
Опять же в соответствии с более ранними исследованиями крысы, которым давали Е3, показали менее заметную потерю массы тела, чем крысы с артритом, которым давали физиологический раствор (Таблица 22 и Фиг.22). Фактически, крысы, которым давали высокие дозы антитела Е3, восстанавливали произошедшую ранее потерю массы тела и на самом деле прибавляли в массе быстрее, чем группы без артрита (Таблица 21 и Фиг.21).
Пример 9. Анальгетические эффекты антитела Е3 против NGF у пациентов с болью от умеренной до сильной, вызванной остеоартритом колена
В этом рандомизированном двойном слепом исследовании с повышением дозы и с плацебо в качестве контроля анальгетические эффекты разовых внутривенных доз (3 мкг/кг, 10 мкг/кг, 30 мкг/кг, 100 мкг/кг или 300 мкг/кг) антитела Е3 против NGF сравнивали с плацебо у пациентов с болью от умеренной до сильной, вызванной остеоартритом колена. В этом исследовании участвовали взрослые мужчины и женщины (возраст между 35 и 65), которые испытывали боль от умеренной до сильной, вызванную остеоартритом колена, но в других отношениях имели, в общем, хорошее состояние здоровья. Во время скринингового периода пациентам требовалось прекратить прием лекарственных средств от артрита, таких как нестероидные противовоспалительные средства (НПВС), ингибиторы циклооксигеназы типа-2 (ЦОГ-2) и опиаты, по меньшей мере за 14 суток до введения антитела Е3 против NGF. Пациентам разрешалось использовать лечение ацетаминофеном или ибупрофеном по неотложным показаниям с некоторыми ограничениями на основании лабораторных данных или прошлой истории болезни. Некоторые пациенты принимали лекарства по неотложным показаниям во время исследования, но лекарства по неотложным показаниям не принимались в течение 2 суток до и после введения антитела Е3 против NGF.
Тридцать четыре пациента были допущены к исследованию, и их оценивали в госпитальных условиях на сутки исследования -1, 1 и 2. Введение антитела Е3 против NGF осуществляли утром в первые сутки. Для записи индекса боли в колене и приема лекарств по неотложным показаниям в клинике и дома использовали электронные дневники. Десяти пациентам давали плацебо. Двадцать четыре пациента получали антитело Е3 против NGF: по четыре пациента при уровне дозы 3 мкг/кг (группа 1), 10 мкг/кг (группа 2) и 30 мкг/кг (группа 3) и по шесть пациентов при уровне дозы 100 мкг/кг (группа 4) и 300 мкг/кг (группа 5). В качестве плацебо использовали стерильный 0,9%-ный хлорид натрия для инъекций в соответствии с Фармакопеей США (USP) (нормальный физиологический раствор). Антитело Е3 против NGF представляло собой замороженный жидкий препарат, состоящий из 10 мг/мл антитела в водном растворе 10 мМ гистидина, 275 мМ сахарозы, 0,01% полисорбата 20, рН 6,0. За один час до внутривенного введения замороженное антитело подвергали оттаиванию и разбавляли в хлориде натрия для инъекций, USP. Объем для каждого пациента вычисляли на основании массы субъекта на сутки -1 и относили к уровню дозы в диапазоне 3-6 мл для группы 1, 10-20 мл для группы 2, 30-60 мл для группы 3 и 100 мл для группы 4 и группы 5. Для групп 1 и 2 антитело Е3 вводили медленной внутривенной болюсной инъекцией в течение 3-5 минут с последующим промыванием 5 мл хлорида натрия для инъекций, USP, внутривенно. Для групп 3-5 антитело Е3 вводили при 100 мл/ч инфузионным насосом с последующим промыванием 5 мл хлорида натрия для инъекций, USP, внутривенно и прекращением внутривенного введения. В каждой группе плацебо (хлорид натрия для инъекций, USP) вводили медленной болюсной инъекцией или непрерывной инфузией так же, как и антитело Е3.
После двух суток в клинике пациентов выписывали домой. В зависимости от группы по уровню дозы, их продолжали оценивать по уровням боли в течение 28 суток после введения антитела Е3 (группы 1, 2 и 3) или вплоть до 181 суток после введения антитела Е3 (группы 4 и 5). Периодические оценки безопасности осуществляли в течение 181 суток для всех пациентов.
Анальгетические эффекты оценивали до введения и многократно после введения антитела Е3. Для индексации текущей боли в колене использовали утвержденную электронную визуальную аналоговую шкалу (VAS) с диапазоном от 0-100 и разрешением 101 точка. Пациентов стимулировали 4 раза в сутки и инструктировали указывать уровень боли, вызванной артритом, в индексируемом колене на данный момент. Утвержденную электронную VAS в диапазоне от 0-100 с разрешением 101 точка также использовали для индексации боли в колене во время ходьбы. VAS для индексации боли в колене во время ходьбы заполняли ежедневно одновременно с VAS для текущей четырехкратной индексации боли в колене.
Электронный дневник для сбора данных выдавали на сутки -8 и осуществляли 7-дневную скрининговую оценку заполнения электронного дневника начиная от суток -8 по сутки -2 для определения успешности использования электронных дневников данным субъектом. Субъектов инструктировали делать 4 записи в сутки. В качестве исходного уровня для оценки эффективности использовали среднюю оценку по VAS, записанную во время скринингового периода.
Для оценки боли, вызванной артритом, также использовали шкалу WOMAC 3.1 (Артритная шкала университетов Западного Онтарио и МакМастера) и Индекс остеоартрита, Версия VA.1®. WOMAC 3.1™ состоит из 24 вопросов, разделенных по 3 группам: боль (5 вопросов), ригидность (2 вопроса) и физическая функция (17 вопросов). Bellamy et al., J. Rheumatol 15:1833-40 (1988); и Bellamy, Semin. Arthritis Rheum. 18:14-7 (1989). Группа физической функции дает информацию о способности к повседневной жизненной активности. Пациенты выполняли тест непосредственно на электронном дневнике во время назначенных посещений клиники при использовании утвержденной электронной VAS в диапазоне 0-100 при разрешении 101 точка. Каждая группа получала балльную оценку определением среднего от баллов по VAS для компонентных вопросов. Суммарную WOMAC 3.1™ определяли из среднего по баллам для каждой из 3 групп. От пациентов также требовали сообщать в электронном дневнике об использованном лекарстве для неотложных целей.
Результат оценивали как изменение от исходного уровня (уровень боли во время скринингового периода), выражали как отличие интенсивности боли (PID) или отличие суммарной интенсивности боли (SPID) для разных уровней дозы. Изменения от исходного уровня в показателях активности анализировали при использовании модели, представляющей собой анализ ковариации (ANCOVA), с лечением в качестве основного фактора и исходным уровнем боли в качестве ковариантного параметра. Для средних и изменений среднего делали сравнения антитела Е3 с плацебо при использовании критерия Даннетта. Для оценки зависимости доза-ответ использовали критерий тренда по Тьюки-Циминере-Хейсу (Tukey-Ciminera-Heyse).
Как показано на Фиг.24, снижение средней интенсивности ежедневной боли после разового введения антитела Е3 против NGF наблюдали при каждой тестированной дозе. Обычно эффект снижения интенсивности боли длился дольше в группе, которую лечили более высокой дозой (30 мкг/кг, 100 мкг/кг и 300 мкг/кг), чем в группе, которую лечили более низкой дозой (3 мкг/кг и 10 мкг/кг). Как показано на Фиг.25, снижение интенсивности боли продолжалось по меньшей мере 80 суток после введения Е3 при 100 мкг/кг.
Таблица 23 ниже показывает отличия суммарной интенсивности боли (SPID) для каждого введения Е3 на сутки 2-8, сутки 2-14, сутки 2-21 и сутки 2-28 при сравнении с исходным уровнем. Для статистического анализа использовали ANCOVA. Таблица 23 свидетельствует о том, что после разового введения антитела Е3 против NGF снижение боли было статистически достоверным (p<0,05).
Статистический анализ осуществляли без поправок и с поправками на множественные сравнения. Первый ряд величин p в скобках для каждого интервала времени дан без поправок; второй ряд величин p в скобках дан с поправкой Даннетта.
Как показано на Фиг.26, процент максимального снижения SPID разовым введением антитела Е3 против NGF достигал величины приблизительно от 45% до 65% от суток 2 до суток 14 для каждой тестированной дозы Е3 и достигал величины приблизительно от 45% до 60% от суток 2 до суток 28 для дозы Е3 10 мкг/кг, 30 мкг/кг, 100 мкг/кг и 300 мкг/кг.
Как показано на Фиг.27, введение антитела Е3 против NGF также снижало балл WOMAC от суток 1 до суток 28. Баллы боли, физической функции и ригидности были достоверно снижены при дозе Е3 100 мкг/кг. Эти данные показывают, что разовое введение антитела против NGF не только снижает боль, но также улучшает физическую функцию и улучшает ригидность у пациентов, имеющих остеоартрит.
Депонирование биологического материала
Следующие материалы были депонированы в Американской коллекции типовых культур (АТСС: American Type Culture Collection, 10801 University Boulevard, Manassas, Virginia, USA):
Вектор Eb.911.3E представляет собой полинуклеотид, кодирующий вариабельную область легкой цепи Е3; вектор Eb.pur.911.3E представляет собой полинуклеотид, кодирующий вариабельную область легкой цепи Е3, и вектор Db.911.3E представляет собой полинуклеотид, кодирующий вариабельную область тяжелой цепи Е3.
Это депонирование было сделано в соответствии с Будапештским договором о международном признании депонирования микроорганизмов для целей патентной процедуры и Инструкций к ним (Будапештский договор). Это гарантирует поддержание депонированной жизнеспособной культуры в течение 30 лет со дня депонирования. АТСС должна сделать депозит доступным в соответствии с условиями Будапештского договора и соглашения между Rinat Neuroscience Corp. и АТСС, которое гарантирует постоянную и неограниченную доступность потомства депонированной культуры при публикации соответствующего патента США или публичном раскрытии любой заявки на патент США или иностранный патент в зависимости от того, какое событие произойдет первым, и гарантирует доступность этого потомства для любого, кто определен Комиссаром по патентам и товарным знакам США быть уполномоченным на это в соответствии со Сводом законов США 35 USC, раздел 122, и соответствующими правилами для Комиссара (включая Свод федеральных постановлений США 37 CFR, Секция 1.14, особенно 886 OG 638).
Агенты по настоящей заявке согласны в том, что если культура депонированного материала погибнет или будет потеряна или разрушена при культивировании в подходящих условиях, эти материалы должны быть немедленно заменены по уведомлению другого об этом. Доступность депонированного материала не должна расцениваться как лицензия на практическое применение этого изобретения в нарушение прав, даваемых любым государством в соответствии с его патентным законодательством.
Последовательности антитела
Вариабельная область тяжелой цепи (CDR по Kabat подчеркнуты; CDR по Chothia обозначены жирным курсивом)
QVQLQESGPGLVKPSETLSLTCTVSGFSLI G YDLN WIRQPPGKGLEWIG IIWGPGTTDYNSAVKS RVTISKDTSKNQFSLKLSSVTAADTAVYYCAR GGYWYATSYYFDAY WGQGTLVTVS (SEQ ID NO:1)
Вариабельные области легкой цепи (CDR по Kabat подчеркнуты; CDR по Chothia обозначены жирным курсивом)
DIQMTQSPSSLSASVGDRVTITC RASQSISNNLN WYQQKPGKAPKLLIY YTSRFHS GVPSRFSGSGSGTDFTFTISSLQPEDIATYYC QQEHTLPYT FGQGTKLEIKRT (SEQ ID NO:2)
Расширенные CDR тяжелой цепи Е3
CDR-H1: GFSLIGYDLN (SEQ ID NO:3)
CDR-H2: IIWGDGTTDYNSAVKS (SEQ ID NO:4)
CDR-H3: GGYWYATSYYFDY (SEQ ID NO:5)
Расширенные CDR легкой цепи Е3
CDR-L1: RASQSISNNLN (SEQ ID NO:6)
CDR-L2: YTSRFHS (SEQ ID NO:7)
CDR-L3: QQEHTLPYT (SEQ ID NO:8)
Расширенные CDR мышиного моноклонального антитела 911
Расширенные CDR тяжелой цепи 911
CDR-H1: GFSLIGYDIN (SEQ ID NO:9)
CDR-H2: MIWGDGTTDYNSALKS (SEQ ID NO:10)
CDR-H3: GGYYYGTSYYFDY (SEQ ID NO:11)
Расширенные CDR легкой цепи 911
CDR-L1: RASQDISNHLN (SEQ ID NO:12)
CDR-L2: YISRFHS (SEQ ID NO:13)
CDR-L3: QQSKTLPYT (SEQ ID NO:14)
Аминокислотная последовательность (полная) тяжелой цепи Е3 QVQLQESGPGLVKPSETLSLTCTVSGFSLIGYDLNWIRQPPGKGLEWIGIIWGDGTTDYNSAVKSRVTISKDTSKNQFSLKLSSVTAADTAVYYCARGGYWYATSYYFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYLPEHYTVSWNSGALTSGVHTFPAYLQSSGLYSLSSWTVPSSNFGTQTYTCNVDHKPSNTKVDKTVERKCCVECPPCPAPHYAGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRWSYLTWHQDWLNGKEYKCKVSNKGLPSSIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO:16)
Аминокислотная последовательность легкой цепи 3Е (полное антитело) DIQMTQSPSSLSASVGDRVTITCRASQSISNNLNWYQQKPGKAPKLLIYYTSRFHSGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQEHTLPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVYCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHXVYACEVTHQGLSSHYTKSFNRGEC (SEQ ID NO:17)
Нуклеотидная последовательность тяжелой цепи 3Е (полное антитело)
CAGGTGCAGCTGCAGGAGTCTGGCCCAGGACTGGTGAAGCCTTCCGAGACCCTGTCCCTCACCTGCACTGTCTCTGGGTTCTCACTTATCGGCTATGATCTTAACTGGATCCGACAGCCTCCAGGGAAGGGACTGGAGTGGATTGGGATTATCTGGGGTGATGGAACCACAGACTATAATTCAGCTGTCAAATCCCGCGTCACCATCTCAAAAGACACCTCCAAGAACCAGTTCTCCCTGAAGCTGAGCTCTGTGACCGCCGCGGACACGGCCGTGTATTACTGTGCGAGAGGAGGTTATTGGTACGCCACTAGCTACTACTTTGACTACTGGGGCCAGGGCACCCTGGTCACCGTCTCCTCAGCCTCCACCAAGGGCCCATCTGTCTTCCCACTGGCCCCATGCTCCCGCAGCACCTCCGAGAGCACAGCCGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCAGAACCTGTGACCGTGTCCTGGAACTCTGGCGCTCTGACCAGCGGCGTGCACACCTTCCCAGCTGTCCTGCAGTCCTCAGGTCTCTACTCCCTCAGCAGCGTGGTGACCGTGCCATCCAGCAACTTCGGCACCCAGACCTACACCTGCAACGTAGATCACAAGCCAAGCAACACCAAGGTCGACAAGACCGTGGAGAGAAAGTGTTGTGTGGAGTGTCCACCTTGTCCAGCCCCTCCAGTGGCCGGACCATCCGTGTTCCTGTTCCCTCCAAAGCCAAAGGACACCCTGATGATCTCCAGAACCCCAGAGGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCAGAGGTGCAGTTCAACTGGTATGTGGACGGAGTGGAGGTGCACAACGCCAAGACCAAGCCAAGAGAGGAGCAGTTCAACTCCACCTTCAGAGTGGTGAGCGTGCTGACCGTGGTGCACCAGGACTGGCTGAACGGAAAGGAGTATAAGTGTAAGGTGTCCAACAAGGGACTGCCATCCAGCATCGAGAAGACCATCTCCAAGACCAAGGGACAGCCAAGAGAGCCACAGGTGTATACCCTGCCACCATCCAGAGAGGAGATGACCAAGAACCAGGTGTCCCTGACCTGTCTGGTGAAGGGATTCTATCCATCCGACATCGCCGTGGAGTGGGAGTCCAACGGACAGCCAGAGAACAACTATAAGACCACCCCTCCAATGCTGGACTCCGACGGATCCTTCTTCCTGTATTCCAAGCTGACCGTGGACAAGTCCAGATGGCAGCAGGGAAACGTGTTCTCTTGTTCCGTGATGCACGAGGCCCTGCACAACCACTATACCCAGAAGAGCCTGTCCCTGTCTCCAGGAAAGTAA (SEQ ID NO:65)
Нуклеотидная последовательность вариабельного домена тяжелой цепи 3Е
CAGGTGCAGCTGCAGGAGTCTGGCCCAGGACTGGTGAAGCCTTCCGAGACCCTGTCCCTCACCTGCACTGTCTCTGGGTTCTCACTTATCGGCTATGATCTTAACTGGATCCGACAGCCTCCAGGGAAGGGACTGGAGTGGATTGGGATTATCTGGGGTGATGGAACCACAGACTATAATTCAGCTGTCAAATCCCGCGTCACCATCTCAAAAGACACCTCCAAGAACCAGTTCTCCCTGAAGCTGAGCTCTGTGACCGCCGCGGACACGGCCGTGTATTACTGTGCGAGAGGAGGTTATTGGTACGCCACTAGCTACTACTTTGACTACTGGGGCCAGGGCACCCTGGTCACCGTCTCCTCA (SEQ ID NO:66)
Нуклеотидная последовательность легкой цепи 3Е (полное антитело)
GATATCCAGATGACACAGTCCCCATCCTCCCTGTCTGCCTCTGTGGGTGACCGCGTCACCATCACCTGCCGCGCATCTCAGTCCATTAGCAATAATCTGAACTGGTATCAGCAGAAGCCAGGCAAAGCCCCAAAACTCCTGATCTACTACACCTCACGCTTCCACTCAGGTGTCCCATCACGCTTCAGTGGCAGTGGCTCTGGTACAGATTTCACCTTCACCATTAGCAGCCTGCAACCAGAAGATATTGCCACTTATTACTGCCAACAGGAGCATACCCTTCCATATACCTTCGGTCAAGGCACCAAGCTGGAGATCAAACGCACTGTGGCTGCACCATCTGTCTTCATCTTTCCTCCATCTGATGAGCAGTTGAAATCCGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCACGCGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCCGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACCCTGAGCAAAGCAGACTACGAGAAACACMAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGTTCTCCAGTCACAAAGAGCTTCAACCGCGGTGAGTGCTAA (SEQ ID NO:67)
Нуклеотидная последовательность вариабельного домена легкой цепи 3Е
GATATCCAGATGACACAGTCCCCATCCTCCCTGTCTGCCTCTGTGGGTGACCGCGTCACCATCACCTGCCGCGCATCTCAGTCCATTAGCAATAATCTGAACTGGTATCAGCAGAAGCCAGGCAAAGCCCCAAAACTCCTGATCTACTACACCTCACGCTTCCACTCAGGTGTCCCATCACGCTTCAGTGGCAGTGGCTCTGGTACAGATTTCACCTTCACCATTAGCAGCCTGCAACCAGAAGATATTGCCACTTATTACTGCCAACAGGAGCATACCCTTCCATATACCTTCGGTCAAGGCACCAAGCTGGAGATCAAACGC (SEQ ID NO:68)
Понятно, что описанные здесь примеры и воплощения предназначены только для иллюстративных целей и что в свете их различные модификации или изменения будут предложены специалистам в данной области и должны быть включены в пределы сущности и объема этого изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ ИНТЕРСТИЦИАЛЬНОГО ЦИСТИТА | 2008 |
|
RU2570559C2 |
| СТАБИЛЬНЫЙ ЖИДКИЙ ПРЕПАРАТ АНТИТЕЛА | 2009 |
|
RU2518278C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ПОСЛЕОПЕРАЦИОННОЙ БОЛИ ВВЕДЕНИЕМ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ФАКТОР РОСТА НЕРВОВ | 2003 |
|
RU2338555C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2644235C2 |
| ХИМЕРНЫЙ БЕЛОК, СОСТАВЛЕННЫЙ ИЗ ДОМЕНА АНТАГОНИСТА NGF И ДОМЕНА АНТАГОНИСТА TNFα | 2015 |
|
RU2678810C2 |
| АНТИТЕЛА ПРОТИВ NGF И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2019 |
|
RU2838175C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640252C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640254C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ БОЛИ | 2009 |
|
RU2522493C2 |
| АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА IL-7 И СПОСОБЫ | 2011 |
|
RU2533809C9 |



























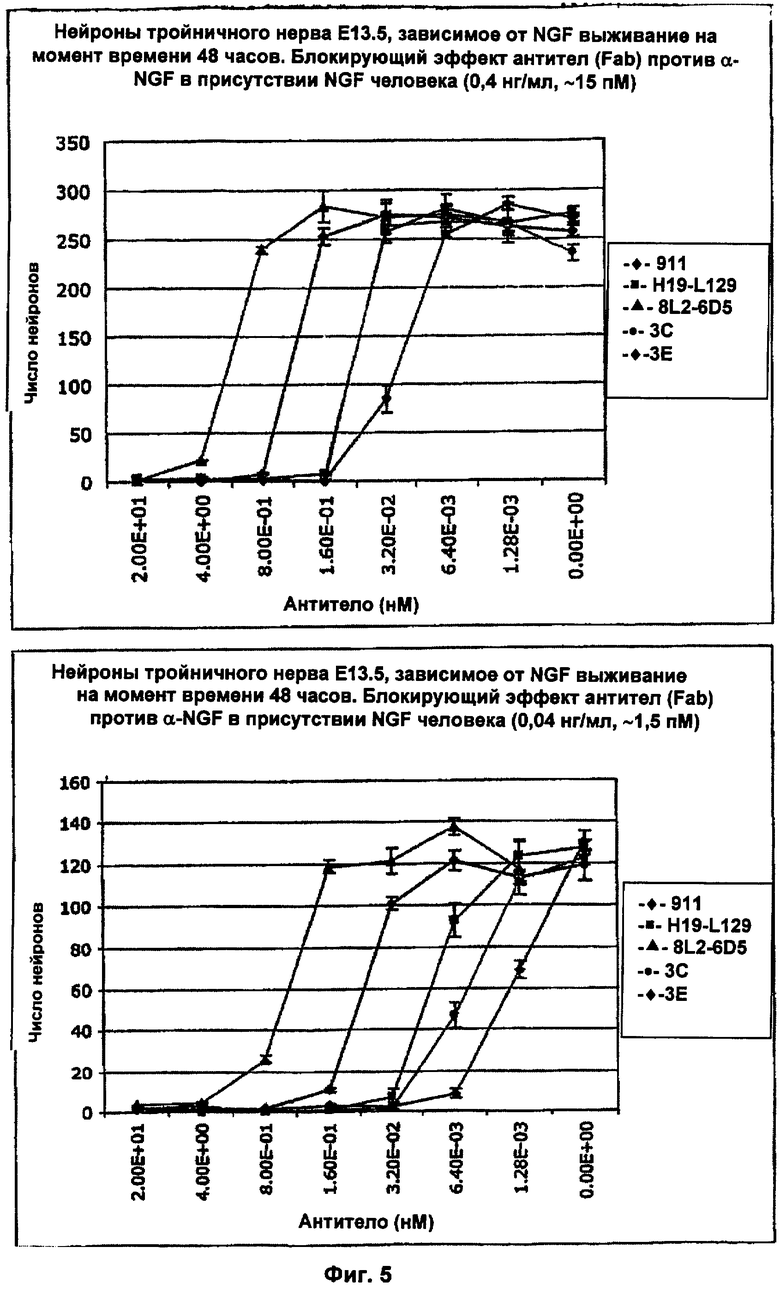




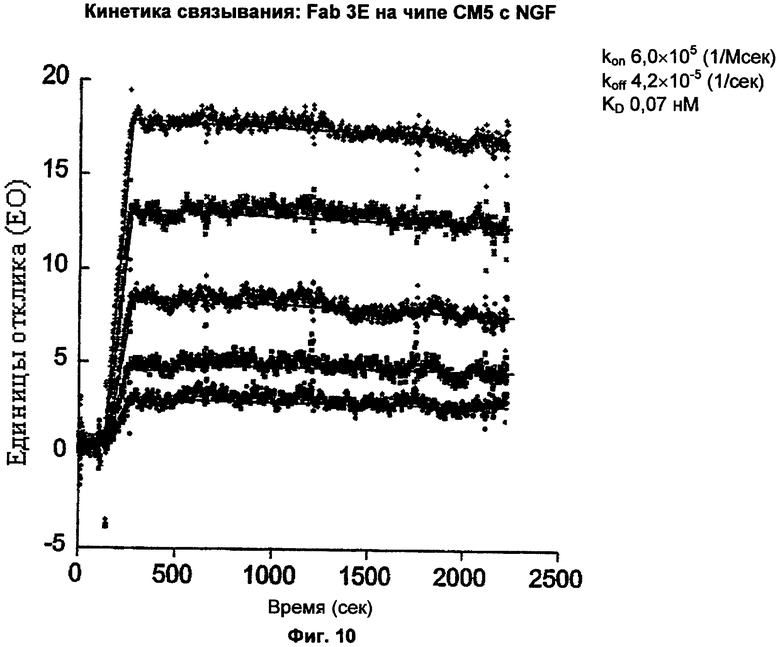

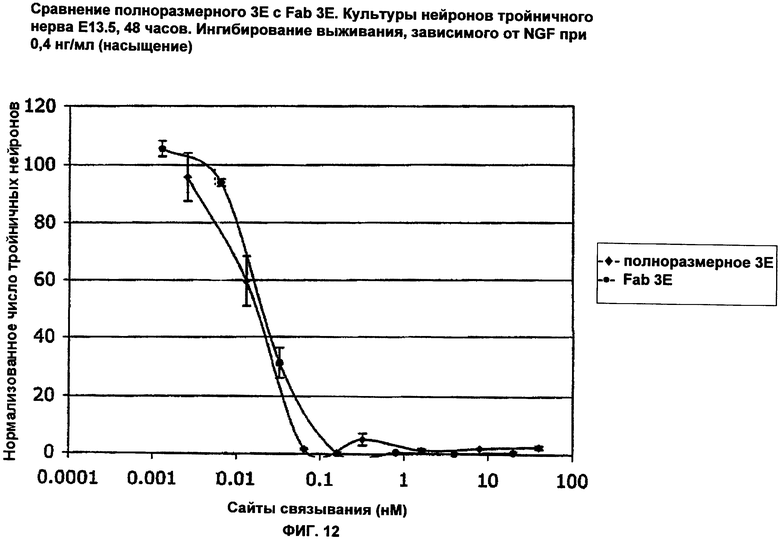






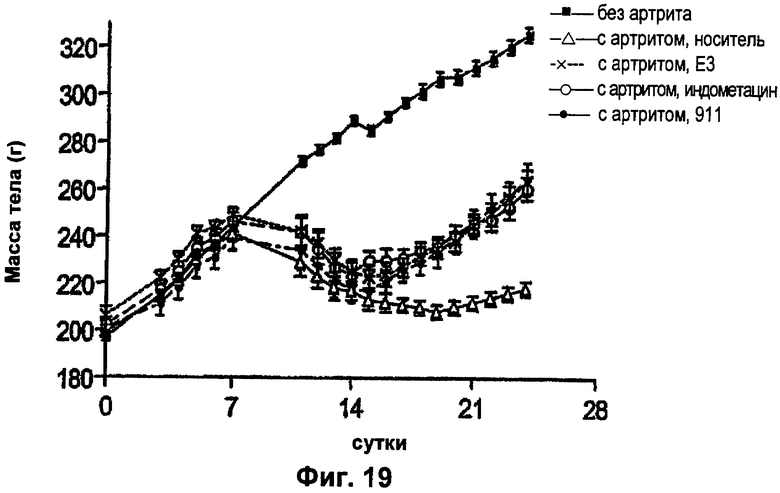
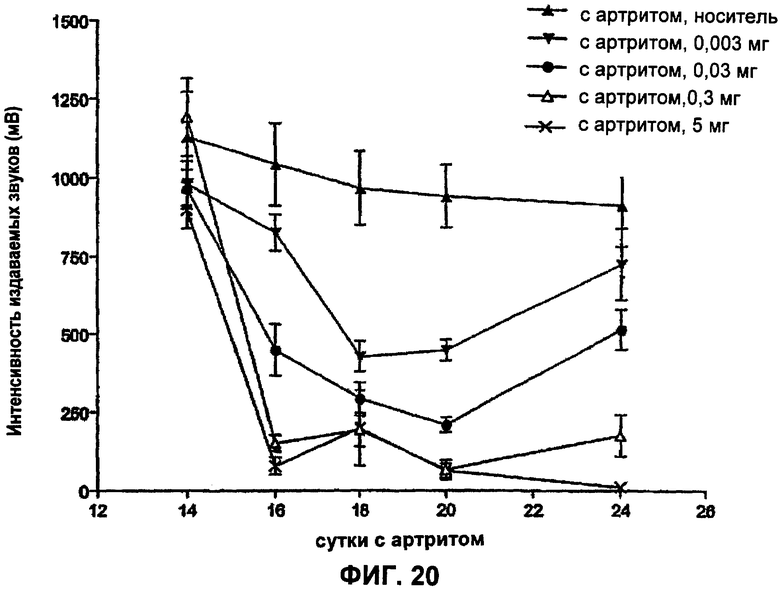

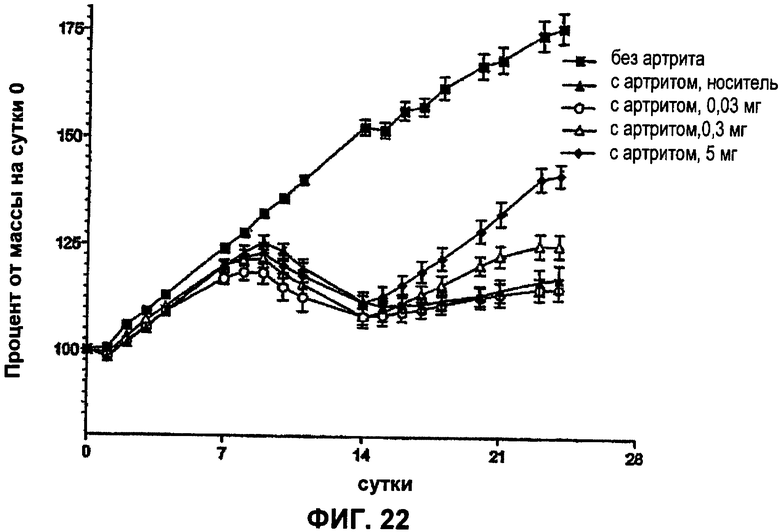








Группа изобретений относится к применению антител против фактора роста нервов (NGF) (анти-NGF антитела) для лечения боли, улучшения физической функции и уменьшения ригидности у индивида, имеющего остеоартрит. Представлены: способ лечения боли, улучшения физической функции и уменьшения ригидности у индивида, имеющего остеоартрит, включает введение указанному индивиду эффективного количества анти-NGF антитела, а также набор и фармацевтическая композиция для осуществления такого способа лечения. Показано, что разовое введение анти-NGF антитела не только снижает боль, но также улучшает физическую функцию и уменьшает ригидность у пациентов, имеющих остеоартрит. 3 н. 21 з.п. ф-лы, 23 табл., 27 ил.
1. Способ лечения боли, улучшения физической функции и уменьшения ригидности у индивида, имеющего остеоартрит, включающий введение указанному индивиду эффективного количества анти-NGF антитела.
2. Способ по п.1, где индивид представляет собой человека.
3. Способ по п.1, где анти-NGF антитело вводят с частотой введения в диапазоне от одного раза в неделю до одного раза в двенадцать недель.
4. Способ по п.1, где анти-NGF антитело вводят один раз в месяц, один раз в два месяца, один раз в три месяца, один раз в четыре месяца, один раз в пять месяцев или один раз в шесть месяцев.
5. Способ по п.1, где боль облегчают на период по меньшей мере приблизительно семь суток, по меньшей мере приблизительно четырнадцать суток, по меньшей мере приблизительно четыре недели или по меньшей мере приблизительно двенадцать недель после введения разовой дозы анти-NGF антитела.
6. Способ по п.1, где анти-NGF антитело вводят в дозе в диапазоне от приблизительно 3 мкг/кг до приблизительно 1 мг/кг.
7. Способ по п.1, где анти-NGF антитело вводят в дозе приблизительно 100 мкг/кг или приблизительно 300 мкг/кг.
8. Способ по п.1, где анти-NGF антитело вводят внутривенно или подкожно.
9. Способ по п.1, где анти-NGF антитело представляет собой моноклональное антитело.
10. Способ по п.1, где анти-NGF антитело представляет собой гуманизированное антитело.
11. Способ по п.1, где анти-NGF антитело связывается с человеческим NGF.
12. Способ по п.11, где анти-NGF антитело дополнительно связывается с NGF грызуна.
13. Способ по п.1, где анти-NGF антитело:
(а) связывается с NGF при KD менее чем приблизительно 2 нМ;
(б) ингибирует зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при значении IC50 приблизительно 100 пМ или менее, где IC50 измеряют в присутствии приблизительно 15 пМ человеческого NGF; и
(в) ингибирует зависимое от человеческого NGF выживание мышиных нейронов тройничного нерва Е13.5 при значении IС50 приблизительно 10 пМ или менее, где IC50 измеряют в присутствии приблизительно 1,5 пМ NGF.
14. Способ по любому из пп.1-13, где анти-NGF антитело содержит вариабельную область тяжелой цепи, содержащую:
(а) область CDR1 (гипервариабельную область), показанную в SEQ ID NO:3;
(б) область CDR2, показанную в SEQ ID NO:4; и
(в) область CDR3, показанную в SEQ ID NO:5.
15. Способ по любому из пп.1-13, где анти-NGF антитело содержит вариабельную область легкой цепи, содержащую:
(а) область CDR1, показанную в SEQ ID NO:6;
(б) область CDR2, показанную в SEQ ID NO:7; и
(в) область CDR3, показанную в SEQ ID NO:8.
16. Способ по п.15, где анти-NGF антитело дополнительно содержит вариабельную область тяжелой цепи, содержащую:
(а) область CDR1, показанную в SEQ ID NO:3;
(б) область CDR2, показанную в SEQ ID NO:4; и
(в) область CDR3, показанную в SEQ ID NO:5.
17. Способ по п.16, где анти-NGF антитело представляет собой антитело, содержащее аминокислотные последовательности, показанные в SEQ ID NO:1 и 2.
18. Способ по п.17, где анти-NGF антитело представляет собой антитело, содержащее аминокислотные последовательности, показанные в SEQ ID NO:16 и 17.
19. Набор, содержащий эффективное количество анти-NGF антитела и инструкции по введению эффективного количества анти-NGF антитела индивиду, имеющему остеоартрит, для лечения боли, улучшения физической функции, вызванной остеоартритом, и уменьшения ригидности у данного индивида.
20. Набор по п.19, где индивид представляет собой человека.
21. Набор по п.19, где анти-NGF антитело вводят с частотой введения в диапазоне от одного раза в неделю до одного раза в двенадцать недель.
22. Набор по п.19, где боль облегчают на период по меньшей мере приблизительно семь суток после введения разовой дозы анти-NGF антитела.
23. Набор по любому из пп.19-22, где анти-NGF антитело вводят в дозе в диапазоне от приблизительно 3 мкг/кг до приблизительно 1 мг/кг.
24. Фармацевтическая композиция для лечения боли, улучшения физической функции и уменьшения ригидности у индивида, имеющего остеоартрит, содержащая анти-NGF антитело и фармацевтически приемлемый носитель.
| WO 2004058184 А2, 15.07.2004 | |||
| НАСОНОВА В.А | |||
| и др | |||
| Клиническая ревматология | |||
| - М.: Медицина, 1989, с.432,466-471 | |||
| МАЗУРОВ В.И | |||
| Клиническая ревматология | |||
| - С.-Пб.: Фолиант, 2001, с.364-365 | |||
| US 2004237124 A1, 25.11.2004 | |||
| WO 2005019266 A2, 03.03.2005 | |||
| HALLIDAY DALE A | |||
| et al | |||
| Elevated nerve growth factor levels in the synovial fluid of patients whith |

