ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области иммунологии. Конкретнее, настоящее изобретение относится к антигенсвязывающим белкам против NGF, специфично связывающимся с NGF, которые были модифицированы для устранения их иммуногенности у интересующего вида. Кроме того, изобретение относится к применению таких антигенсвязывающих белков в лечении и/или предотвращении расстройств, связанных с NGF, в частности боли.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Фактор роста нервов (NGF) был первым идентифицированным нейротрофином, и его роль в развитии и выживании как периферических, так и центральных нейронов хорошо охарактеризована. Было показано, что NGF является критически важным фактором выживания и поддерживающим фактором при развитии периферических симпатических и эмбриональных чувствительных нейронов и холинергических нейронов базальных отделов переднего мозга (Smeyne, et al., Nature 368:246-249 (1994), и Crowley, et al., Cell 76: 1001-101 I (1994)). NGF приводит к повышающей регуляции экспрессии нейропептидов в чувствительных нейронах (Lindsay, et al., Nature 337:362-364 (1989)), и его активность опосредована двумя разными мембраносвязанными рецепторами, тирозинкиназным рецептором TrkA и общим рецептором нейротрофинов р75 (иногда называемыми «высокоаффинным» и «низко аффинным» рецепторами NGF, соответственно), который является структурно родственным по отношению к другим представителям семейства рецептора фактора некроза опухоли (Chao, et al., Science 232:518-521 (1986)).
Помимо его влияния на нервную систему, все больше данных указывают на участие NGF в процессах, происходящих за пределами нервной системы. Например, было показано, что NGF повышает сосудистую проницаемость (Otten, et al., Eur J Pharmacol. 106: 199-201 (1984)), усиливает T- и B-клеточные иммунные ответы (Otten, et al., Proc. Natl. Acad. Sci. USA 86:10059-10063 (1989)), индуцирует дифференцировку лимфоцитов и пролиферацию тучных клеток и приводит к высвобождению растворимых биологических сигнальных веществ из тучных клеток (Matsuda, et al., Proc. Natl. Acad. Sci. USA 85:6508-6512 (1988); Pearce, et al., J. Physiol. 372:379-393 (1986); Bischoff, et al., Blood 79:2662-2669 (1992); Horigome, et al., J. Bioi. Chem. 268:14881-14887 (1993)).
NGF продуцируют клетки нескольких типов, включая тучные клетки (Leon, et al., Proc. Natl. Acad. Sci. USA 91:3739-3743 (1994)), В-лимфоциты (Torcia, et al., Cell 85:345-356 (1996)), кератиноциты (Di Marco, et al., J. Biol Chem. 268: 22838-22846), гладкомышечные клетки (Ueyama, et al., J. Hypertens. 11: 1061-1065 (1993)), фибробласты (Lindholm, et al., Eur. J. Neurosci. 2:795-801 (1990)), клетки бронхиального эпителия (Kassel, et al., Clin, Exp. Allergy 31:1432-40 (2001)), мезангиальные клетки почек (Steiner, et al., Am. J. Physiol. 261: F792-798 (1991)) и мышечные трубочки скелетных мышц (Schwartz, et al., J Photochem. Photobiol. B66: 195-200 (2002)). Рецепторы NGF были обнаружены на клетках различных типов за пределами нервной системы. Например, TrkA был обнаружен на человеческих моноцитах, Т- и В-лимфоцитах и тучных клетках.
У пациентов-людей, а также в нескольких моделях на животных была отмечена связь между повышенными уровнями NGF и различными воспалительными состояниями. Они включают системную красную волчанку (Bracci-Laudiero, et al., Neuroreport 4:563-565 (1993)), рассеянный склероз (Bracci-Laudiero, et al., Neurosci. Lett. 147:9-12 (1992)), псориаз (Raychaudhuri, et al., Acta Derm. l'enereol. 78:84-86 (1998)), артрит (Falcim, et al., Ann. Rheum. Dis. 55:745-748 (1996)), интерстициальный цистит (Okragly, et al., J. Urology 161: 438-441 (1999)) и астму (Braun, et al., Eur. J Immunol. 28:3240-3251 (1998)). Стабильное повышение уровня NGF в периферических тканях сопровождается гиперальгезией и воспалением, и было отмечено при нескольких формах артрита. У пациентов с ревматоидным артритом уровни экспрессии NGF в синовиальной оболочке повышены, в то время как в невоспаленной синовиальной оболочке NGF не поддавался выявлению (Aloe, et al., Arch. Rheum. 35:351-355 (1992)). Сходные результаты были получены у крыс с экспериментально индуцированным ревматоидным артритом (Aloe, et al., Clin. Exp. Rheumatol. 10:203-204 (1992)). У трансгенных мышей с артритом были отмечены повышенные уровни NGF наряду с ростом числа тучных клеток (Aloe, et al., Int. J. Tissue Reactions-Exp. Clin. Aspects 15:139-143 (1993)).
Остеоартрит (OA) является одним из наиболее распространенных хронических заболеваний костно-мышечной системы у собак, поражая 20% популяции собак в возрасте более одного года. К развитию OA приводят главным образом травмы, нестабильность суставов и заболевания, такие как дисплазия тазобедренных суставов. Остеоартрит является патологическим состоянием всего сустава, и как воспалительные, так и дегенеративные изменения всех суставных структур приводят к невозможности выполнения ими своих функций и клиническим признакам хромоты и боли. Боль представляет собой наиболее важное клиническое проявление OA у собак и является результатом сложного взаимодействия структурных изменений в суставах, биохимических и молекулярных сдвигов, а также периферических и центральных механизмов восприятия боли. В этой системе активация и повышение чувствительности периферических ноцицепторов воспалительными и гиперальгезическими медиаторами (например, цитокинами, простагландинами и нейромедиаторами) является одним из основных периферических механизмов, приводящих к боли в суставах. Лечение боли у собак нефармацевтическим лекарственным средством, которое будет обеспечивать облегчение на протяжении более продолжительных периодов времени, чем классическое лечение боли у собак, является очевидной неудовлетворенной потребностью.
Только в Соединенных Штатах Америки от OA страдают приблизительно 14,5 миллиона собак (маркетинговое исследование, 2010 г.) Чаще всего ветеринары назначают нестероидные противовоспалительные лекарственные средства (НПВС), но их эффективность и переносимость ограничены. Маркетинговое исследование показывает, что в США лечение НПВС получают приблизительно 9 миллионов собак. Кортикостероиды применяются редко, обычно на протяжении короткого периода времени и в качестве крайней меры. Очевидна неудовлетворенная потребность в удобном и безопасном средстве, которое позволяло бы эффективно лечить собак с OA.
У кошек OA представляет собой патологическое изменение диартродиального синовиального сустава, характеризующееся разрушением суставного хряща, образованием остеофитов, ремоделированием кости, изменениями в мягких тканях и слабо выраженным негнойным воспалением. Несмотря на то, что рентгенологические признаки OA у кошек описаны хорошо, клинические признаки заболевания задокументированы недостаточно и могут оставаться недиагностированными. Оценка хромоты у кошек затруднена ввиду их малых размеров и естественной подвижности, обеспечивающей им широкие компенсаторные возможности. Однако принято считать, что клинические признаки OA у кошек включают уменьшение массы тела, анорексию, депрессию, нетипичные туалетные привычки, недостаточный уход за собой, агрессивное поведение и постепенное снижение способности к прыжкам до явной хромоты. Вследствие диагностических ошибок лечение OA у кошек обычно не проводится и является неудовлетворенной потребностью в области ветеринарной медицины.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно изобретению предложены новый антигенсвязывающий белок против NGF (антитело, фрагмент антитела, антигенсвязывающий фрагмент, антигенсвязывающая часть, антитело-антагонист и так далее, как определено и взаимозаменяемо использовано здесь), и полинуклеотиды, кодирующие указанный белок. Кроме того, согласно изобретению предложены способы получения и применения указанных антигенсвязывающих белков и/или нуклеотидов в лечении и/или предотвращении расстройств, связанных с NGF, в частности боли, у субъекта. Кроме того, согласно изобретению предложены фармацевтические композиции и применения для лечения расстройств, связанных с NGF, в частности боли, у субъекта.
В одном аспекте согласно настоящему изобретению предложен рекомбинантный антигенсвязывающий белок, специфично связывающийся с фактором роста нервов (NGF), содержащий вариабельную область легкой цепи (VL), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 1 или SEQ ID NO: 21, участок 1, определяющий комплементарность (CDR2), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 2 или SEQ ID NO: 22, участок 1, определяющий комплементарность (CDR3), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 3 или SEQ ID NO: 23; и вариабельную область тяжелой цепи (VH), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 4 или SEQ ID NO: 24, участок 1, определяющий комплементарность (CDR2), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 5 или SEQ ID NO: 25, и участок 1, определяющий комплементарность (CDR3), содержащий аминокислотные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 6 или SEQ ID NO: 26; и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно настоящему изобретению предложено, что антигенсвязывающий белок содержит: вариабельную область легкой цепи (VL), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 1, участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 2, участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 3; и вариабельную область тяжелой цепи (VH), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 4, участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 5, участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 6; и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно настоящему изобретению предложено, что антигенсвязывающий белок по изобретению содержит: вариабельную область легкой цепи (VL), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 21, участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 22, участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 23; и вариабельную область тяжелой цепи (VH), содержащую участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 24, участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 25, участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность, по меньшей мере приблизительно на 90% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 26; и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
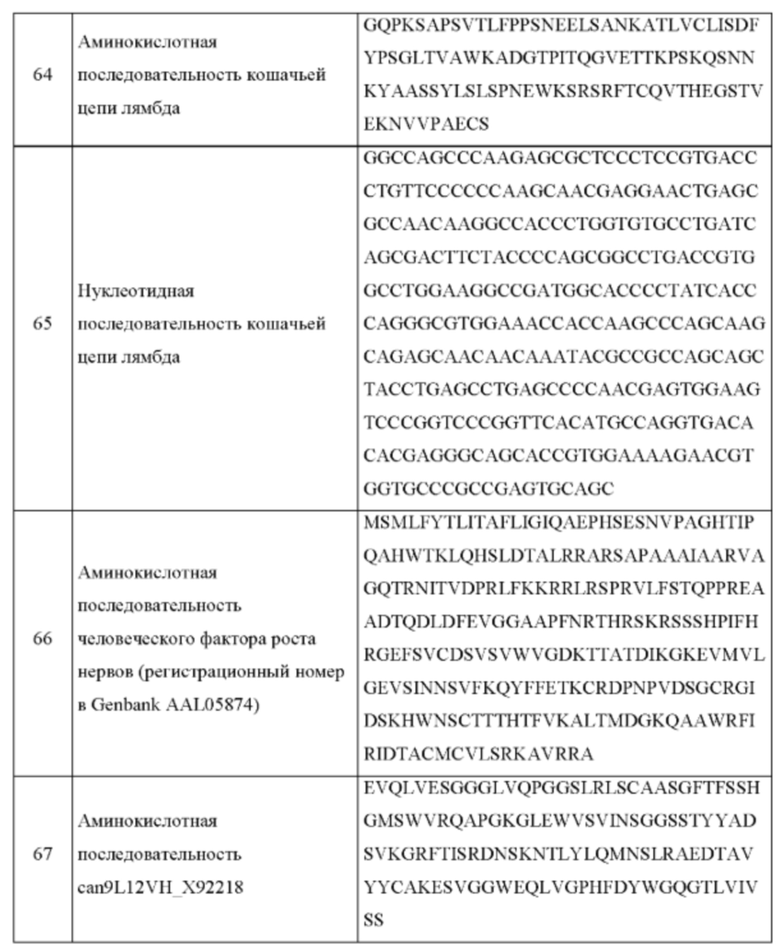
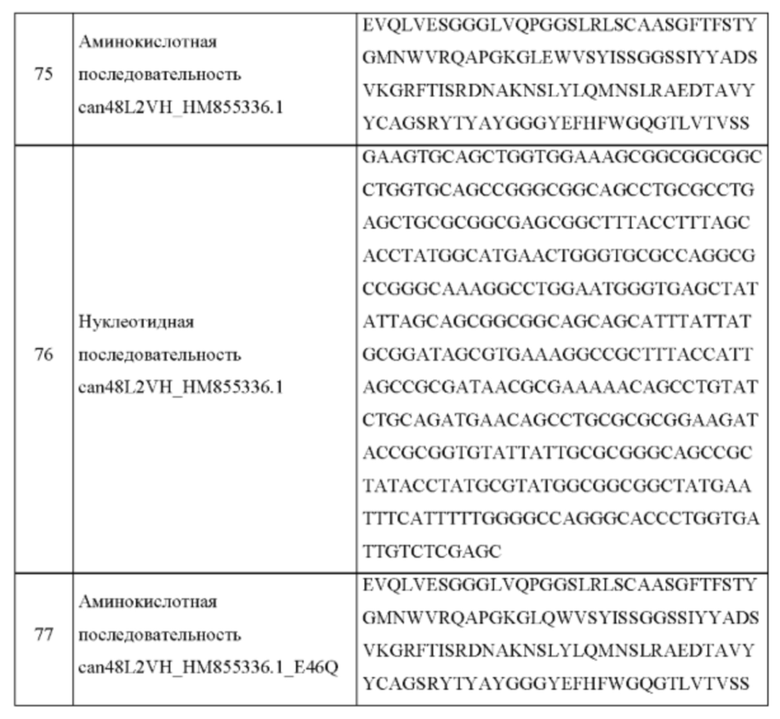
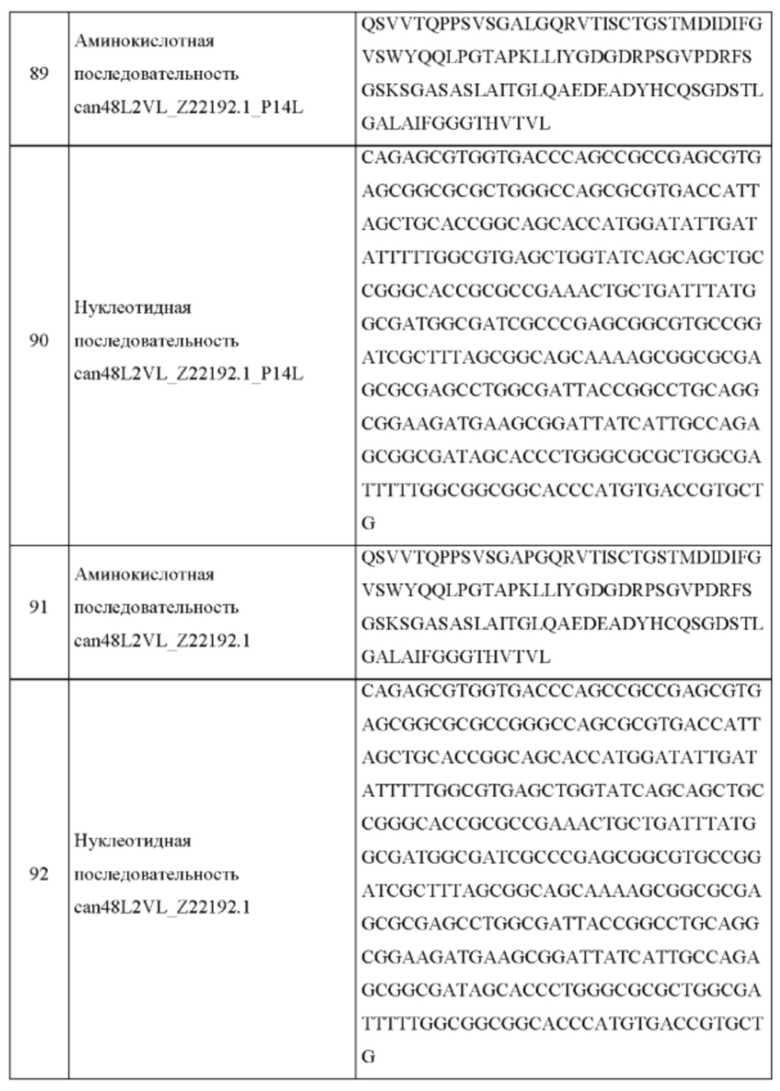
В одном аспекте согласно настоящему изобретению предложен рекомбинантный антигенсвязывающий белок, специфично связывающийся с фактором роста нервов (NGF), содержащий: вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотным последовательностям, выбранным из группы, состоящей из: SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 55, SEQ ID NO: 71, SEQ ID NO: 73, SEQ ID NO: 83, SEQ ID NO: 85, SEQ ID NO: 87, SEQ ID NO: 89 и SEQ ID NO: 91; и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотным последовательностям, выбранным из группы, состоящей из: SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 28, SEQ ID NO: 30, SEQ ID NO: 56, SEQ ID NO: 67, SEQ ID NO: 69, SEQ ID NO: 75, SEQ ID NO: 77, SEQ ID NO: 79 и SEQ ID NO: 81; и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 7, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 8, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложен антигенсвязывающий белок, где вариабельная область легкой цепи содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 27, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 28, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 9, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 10, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 29, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID N0:30, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 55, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 56, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 79, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 87, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 79, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 75, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 87, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 89, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что рекомбинантный антигенсвязывающий белок по изобретению содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 75, и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно настоящему изобретению предложен рекомбинантный антигенсвязывающий белок, специфично связывающийся с фактором роста нервов (NGF), дополнительно содержащий константную область, содержащую аминокислоты, выбранные из SEQ ID NO: 41 или 43. В одном воплощении согласно настоящему изобретению предложена нуклеотидная последовательность, кодирующая константную область, выбранная из группы, состоящей из SEQ ID NO: 42 или SEQ ID NO: 44. В одном воплощении константная область антигенсвязывающего белка по настоящему изобретению не имеет эффекторной функции. В одном воплощении изменения в константной области антигенсвязывающего белка по изобретению предотвращают деградацию антигенсвязывающего белка.
В одном воплощении согласно настоящему изобретению предложен рекомбинантный антигенсвязывающий белок, специфично связывающийся с NGF, дополнительно содержащий константную область, содержащую аминокислотную последовательность, содержащую SEQ ID NO: 62. В одном воплощении согласно настоящему изобретению предложена нуклеотидная последовательность, кодирующая константную область, содержащая SEQ ID NO: 63. В одном воплощении константная область антигенсвязывающего белка по настоящему изобретению испытывает недостаток эффекторной функции. В одном воплощении изменения в константной области антигенсвязывающего белка по изобретению предотвращают деградацию антигенсвязывающего белка.
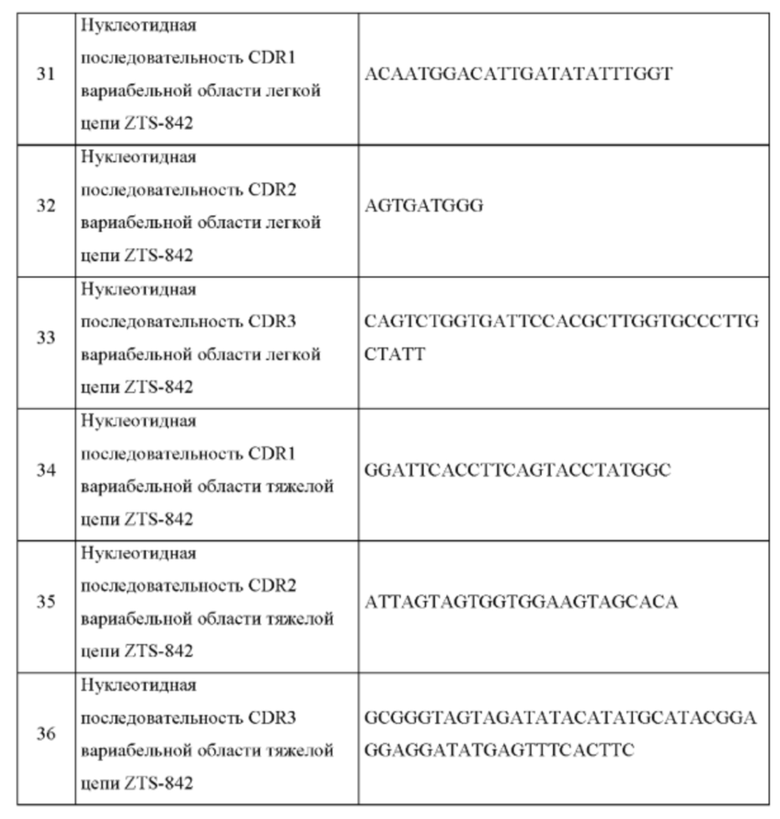
В одном аспекте согласно настоящему изобретению предложены нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, специфично связывающийся с фактором роста нервов (NGF), содержащие последовательности нуклеиновых кислот: вариабельной области легкой цепи (VL), содержащей участок 1, определяющий комплементарность (CDR1), по меньшей мере на 90% идентичный SEQ ID NO: 11 или SEQ ID NO: 31, участок 1, определяющий комплементарность (CDR2), содержащий нуклеотидные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 12 или SEQ ID NO: 32, участок 1, определяющий комплементарность (CDR3), содержащий нуклеотидные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 13 или SEQ ID NO: 33; и вариабельной области тяжелой цепи (VH), содержащей участок 1, определяющий комплементарность (CDR1), содержащий нуклеотидные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 14 или SEQ ID NO: 34, участок 1, определяющий комплементарность (CDR2), содержащий нуклеотидные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 15 или SEQ ID NO: 35, и участок 1, определяющий комплементарность (CDR3), содержащий нуклеотидные последовательности, по меньшей мере на 90% идентичные SEQ ID NO: 15 или SEQ ID NO: 36; и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно настоящему изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению, содержащая: вариабельную область легкой цепи (VL), содержащую участок 1, определяющий комплементарность (CDR1), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 11, участок 2, определяющий комплементарность (CDR2), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 12, участок 3, определяющий комплементарность (CDR3), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 13; и вариабельную область тяжелой цепи (VH), содержащую участок 1, определяющий комплементарность (CDR1), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 14, участок 2, определяющий комплементарность (CDR2), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 15, участок 3, определяющий комплементарность (CDR3), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 16; и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно настоящему изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению и содержащая: нуклеотиды, кодирующие вариабельную область легкой цепи (VL), содержащие участок 1, определяющий комплементарность (CDR1), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 31, участок 2, определяющий комплементарность (CDR2), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 32, участок 3, определяющий комплементарность (CDR3), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 33; и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи (VH), содержащие участок 1, определяющий комплементарность (CDR1), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 34, участок 2, определяющий комплементарность (CDR2), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 35, участок 3, определяющий комплементарность (CDR3), содержащий нуклеотидную последовательность, по меньшей мере приблизительно на 90% идентичную нуклеотидной последовательности, содержащей SEQ ID NO: 36; и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном аспекте согласно настоящему изобретению предложена нуклеотидная последовательность, кодирующая рекомбинантный антигенсвязывающий белок по изобретению, специфично связывающийся с фактором роста нервов (NGF), содержащая: нуклеотиды, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную нуклеотидным последовательностям, выбранным из группы, состоящей из: SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 57, SEQ ID NO: 88, SEQ ID NO: 90 и SEQ ID NO: 92; и нуклеотиды, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную нуклеотидным последовательностям, выбранным из группы, состоящей из: SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 58, SEQ ID NO: 76 и SEQ ID NO: 80; и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 17, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 18, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению, где нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 37, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 38, и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 19, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению, где нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 39, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 40, и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 57, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 58, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 92, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 80, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению, где нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 88, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 80, и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 92, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 76, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложена нуклеотидная последовательность, кодирующая антигенсвязывающий белок по изобретению, где нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 88, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержат нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 76, и любые ее варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка. В одном воплощении согласно изобретению предложено, что нуклеотидные последовательности, кодирующие рекомбинантный антигенсвязывающий белок по изобретению, содержат нуклеотидные последовательности, кодирующие вариабельную область легкой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 90, и нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи, содержащие нуклеотидную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 76, и любые их варианты, имеющие одну или более чем одну замену в нуклеиновой кислоте, основанную на вырожденности генетического кода, в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном или более чем одном воплощении антигенсвязывающий белок по изобретению ингибирует связывание NGF с рецептором TrkA. В одном или более чем одном воплощении антигенсвязывающий белок по изобретению ингибирует биологическую функцию, связанную со связыванием NGF с рецептором TrkA. В одном или более чем одном воплощении антигенсвязывающий белок по изобретению ингибирует связывание NGF с обоими рецепторами TrkA. В одном или более чем одном воплощении антигенсвязывающий белок по изобретению ингибирует биологическую функцию, связанную со связыванием NGF с рецептором TrkA с или без рецепторов р75, включая блокаду трансдукции сигналов и путей, связанных со связыванием NGF с рецептором TrkA.
В одном или более чем одном воплощении антигенсвязывающий белок по изобретению уменьшает или устраняет расстройство, связанное с NGF, посредством нарушения сигнала, связанного со связыванием NGF с рецепторами TrkA и р75. В одном или более чем одном воплощении расстройство, связанное с NGF, выбрано из группы, состоящей из: сердечно-сосудистых заболеваний, атеросклероза, ожирения, диабета 2-го типа, метаболического синдрома, боли и воспаления. В одном воплощении расстройство, связанное с NGF, представляет собой боль. В одном воплощении указанное расстройство, связанное с NGF, представляет собой болевое расстройство и выбрано из группы, состоящей из: боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли. В одном воплощении расстройство, связанное с NGF, включает боль при остеоартрите. В одном воплощении расстройство, связанное с NGF, включает операционную и послеоперационную боль. В одном воплощении расстройство, связанное с NGF, включает раковую боль.
В одном или более чем одном аспекте антигенсвязывающий белок по изобретению выбран из группы, состоящей из: моноклонального антитела, химерного антитела, одноцепочечного антитела, тетрамерного антитела, тетравалентного антитела, мультиспецифичного антитела, доменспецифичного антитела, антитела с делецией домена, слитого белка, слитого белка ScFc, Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, ScFv-фрагмента, Fd-фрагмента, однодоменного антитела, dAb-фрагмента, малого модульного иммунофармацевтического средства (SMIP), нанотела и молекулы IgNAR (иммуноглобулин с новым антигеновым рецептором, immunoglobulin new antigen receptor). В одном воплощении антигенсвязывающий белок представляет собой моноклональное антитело. В одном воплощении антигенсвязывающий белок представляет собой химерное антитело.
В одном воплощении антигенсвязывающий белок по изобретению выбран из собачьего или канинизированного моноклонального антитела, фелинизированного моноклонального антитела, эквинизированного моноклонального антитела или гуманизированного моноклонального антитела. В одном воплощении антигенсвязывающий белок представляет собой собачье или канинизированное антитело. В одном воплощении антигенсвязывающий белок по изобретению представляет собой фелинизированное антитело. В одном воплощении антигенсвязывающий белок по изобретению представляет собой эквинизированное антитело. В одном воплощении антигенсвязывающий белок по изобретению представляет собой гуманизированное антитело.
В одном или более чем одном аспекте согласно изобретению предложена фармацевтическая композиция, содержащая терапевтически эффективное количество антигенсвязывающего белка и фармацевтически приемлемый носитель. В одном воплощении согласно изобретению предложена ветеринарная композиция, содержащая терапевтически эффективное количество антигенсвязывающего белка и фармацевтически приемлемый носитель. В одном воплощении согласно изобретению предложена фармацевтическая или ветеринарная композиция, содержащая терапевтически эффективное количество антигенсвязывающего белка и фармацевтически приемлемый носитель. В одном воплощении фармацевтическую композицию по изобретению применяют в лечении расстройства, связанного с NGF. В одном воплощении расстройство, связанное с NGF, выбрано из группы, состоящей из: сердечно-сосудистых заболеваний, атеросклероза, ожирения, диабета 2-го типа, метаболического синдрома, боли и воспаления. В одном воплощении расстройство, связанное с NGF, включает боль. В одном воплощении фармацевтическую композицию применяют в лечении боли. В одном воплощении фармацевтическую композицию применяют в лечении боли и тип боли выбран из боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли. В одном воплощении боль включает боль при остеоартрите. В одном воплощении боль включает операционную и послеоперационную боль. В одном воплощении боль включает раковую боль. В одном или более чем одном воплощении фармацевтическую композицию по изобретению применяют у собаки. В одном или более чем одном воплощении фармацевтическую композицию по изобретению применяют у кошек. В одном или более чем одном воплощении фармацевтическую композицию по изобретению применяют у лошади. В одном или более чем одном воплощении фармацевтическую композицию по изобретению применяют у людей.
В одном или более чем одном воплощении фармацевтическая композиция по изобретению не оказывает существенного нежелательного эффекта на иммунную систему собаки. В одном воплощении композиция по изобретению не оказывает существенного нежелательного эффекта на иммунную систему кошки. В одном или более чем одном воплощении композиция по изобретению не оказывает существенного нежелательного эффекта на иммунную систему лошади. В одном воплощении композиция по изобретению не оказывает существенного нежелательного эффекта на иммунную систему человека. В одном воплощении фармацевтическая композиция представляет собой ветеринарную композицию.
В одном или более чем одном воплощении согласно настоящему изобретению предложена клетка-хозяин, продуцирующая любой один или более чем один из антигенсвязывающих белков по настоящему изобретению.
В одном или более чем одном воплощении согласно изобретению предложен вектор, содержащий любую одну или более чем одну из нуклеиновых кислот по настоящему изобретению.
В одном или более чем одном воплощении согласно изобретению предложена клетка-хозяин, содержащая любую одну или более чем одну из нуклеиновых кислот по настоящему изобретению.
В одном или более чем одном воплощении согласно изобретению предложена клетка-хозяин, содержащая вектор, содержащий любую одну или более чем одну из нуклеиновых кислот по настоящему изобретению.
В одном или более чем одном воплощении согласно изобретению предложена клетка-хозяин, содержащая любую одну или более чем одну из нуклеиновых кислот по настоящему изобретению.
В одном или более чем одном аспекте согласно настоящему изобретению предложен способ получения антигенсвязывающего белка по изобретению, включающий культивирование клетки-хозяина по изобретению в условиях, приводящих к продуцированию антигенсвязывающего белка, и последующее выделение антигенсвязывающего белка из клетки-хозяина или культуральной среды клетки-хозяина.
В одном или более чем одном аспекте согласно настоящему изобретению предложен способ лечения расстройства, связанного с NGF, у субъекта, включающий введение указанному субъекту терапевтически эффективного количества фармацевтической или ветеринарной композиции по настоящему изобретению. В одном воплощении согласно изобретению предложено, что расстройство, связанное с NGF, выбрано из группы, состоящей из: сердечно-сосудистых заболеваний, атеросклероза, ожирения, диабета 2-го типа, метаболического синдрома, боли и воспаления. В одном воплощении расстройство, связанное с NGF, включает боль. В одном воплощении расстройство, связанное с NGF, представляет собой болевое расстройство и выбрано из группы, состоящей из: боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли. В одном воплощении расстройство, связанное с NGF, включает боль при остеоартрите. В одном воплощении расстройство, связанное с NGF, включает операционную и послеоперационную боль. В одном воплощении расстройство, связанное с NGF, представляет собой раковую боль. В одном воплощении субъект выбран из группы, состоящей из: собак, кошек, людей и лошадей. В одном воплощении субъект включает собак. В одном воплощении субъект включает кошек. В одном воплощении субъект включает лошадей. В одном воплощении субъект включает людей.
В одном или более чем одном воплощении согласно настоящему изобретению предложен способ выявления или количественного определения уровней NGF в биологическом образце, включающий:
а) инкубацию клинического или биологического образца, содержащего NGF, в присутствии любого из антигенсвязывающих белков по настоящему изобретению; и
б) выявление антигенсвязывающего белка, связанного с NGF, в образце.
В некоторых воплощениях антигенсвязывающий белок по изобретению имеет метку, поддающуюся выявлению. В некоторых воплощениях немеченый антигенсвязывающий белок используют в комбинации со вторым антигенсвязывающим белком или его фрагментами, имеющими метку, поддающуюся выявлению. В одном воплощении изобретение включает набор, содержащий антигенсвязывающий белок по изобретению.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлено схематическое изображение общей структуры молекулы мышиного иммуноглобулина G (IgG), на котором подробно показан антигенсвязывающий сайт. Framework I, II, III каркасная область I, II, III, соответственно.
На Фиг. 2 представлено схематическое изображение общей структуры мышиного/собачьего химерного IgG.
На Фиг. 3 представлено изображение, на котором показана видовая адаптация или «канинизация» мышиного IgG, мышиные CDR перенесены на собачьи каркасные области. Данное изображение также применимо для описания фелинизации, эквинизации, гуманизации и иной видовой адаптации, как определено здесь. Canine FW1, FW2, FW3 собачья каркасная область 1, 2, 3, соответственно.
На Фиг. 4 представлено изображение «гетерохимерного» моноклонального антитела, в котором спарены химерная легкая цепь и полностью канинизированная тяжелая цепь. Canine FW1, FW2, FW3 - собачья каркасная область 1, 2, 3, соответственно - собачья каркасная область 1, 2, 3, соответственно; Mouse FW1, FW2, FW3 - мышиная каркасная область FW1, FW2, FW3, соответственно.
На Фиг. 5 представлено изображение вариабельных областей цепей антитела, на котором показаны праймеры для константных областей и вырожденные праймеры, направленные на мышиные вариабельные области.
На Фиг. 6 показаны эффекты mAb против NGF ZTS-841 и ZTS-842 на сигналы pERK 1/2, индуцированные собачьим NGF, в клетках caTrkA-CHO.
На Фиг. 7 показаны эффекты канинизированного варианта mAb aD11, отрицательного контроля и mAb 13L11 на сигналы pERK 1/2, индуцированные собачьим NGF, в клетках caTrkA-CHO.
На Фиг. 8 показаны эффекты mAb против NGF на пролиферацию клеток TF-1, индуцированную собачьим NGF, при использовании mAb ZTS-841 и ZTS-842.
На Фиг. 9 показаны эффекты mAb против NGF на пролиферацию клеток TF-1, индуцированную собачьим NGF, при использовании химерного 48L2, mAb fe148L2VH1.1 и fe148L2VH1.2.
На Фиг. 10 показаны концентрации mAb против NGF ZTS-841 при его п/к / п/к / в/в введении в дозе 2,0 мг/кг для фармакокинетических исследований.
На Фиг. 11 показаны концентрации mAb против NGF ZTS-842 при его п/к / п/к / в/в введении в дозе 2,0 мг/кг для фармакокинетических исследований.
На Фиг. 12 представлена схема MIA-анализа у крыс.
На Фиг. 13 графически представлены результаты MIA-анализа у крыс при введении mAb 841 в дозах 0,1-2 мг/кг.
На Фиг. 14 графически представлены результаты MIA-анализа у крыс при введении mAb 841 в дозах 0,01-2,0 мг/кг.
На Фиг. 15 графически представлены результаты MIA-анализа у крыс при введении mAb 842 в дозах 0,5 и 2 мг/кг.
На Фиг. 16 графически представлен показатель хромоты по VAS при использовании mAb 841 в группах введения через три и пять часов после индукции синовиита в модели LPS-синовиита.
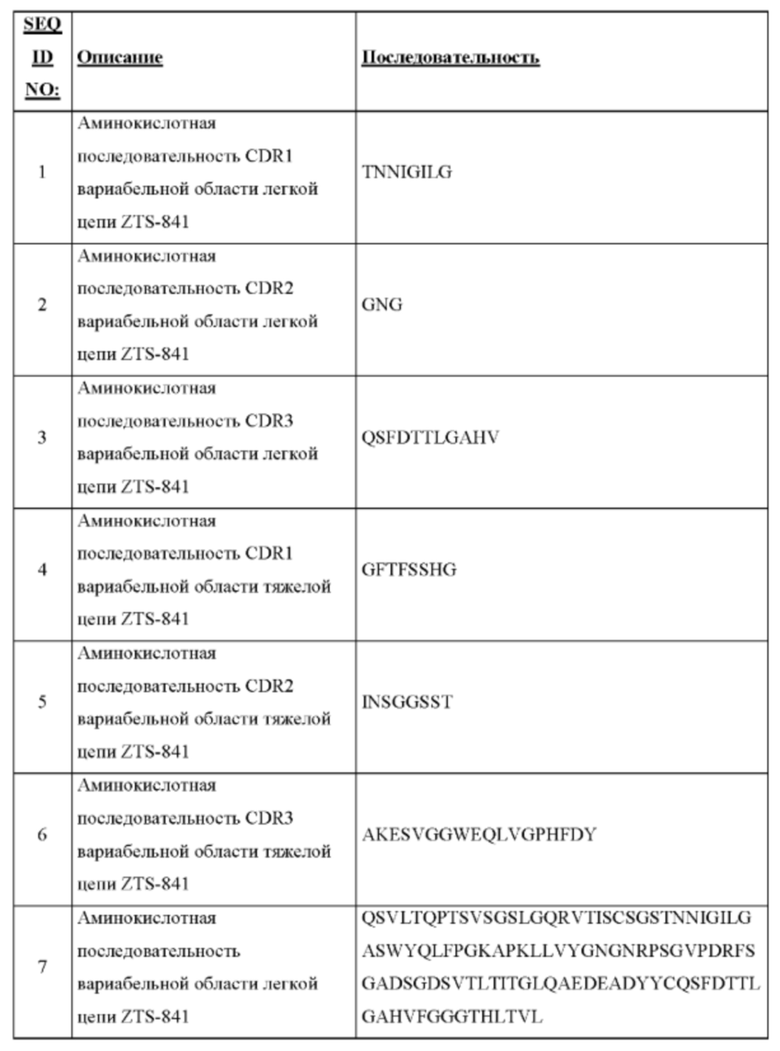
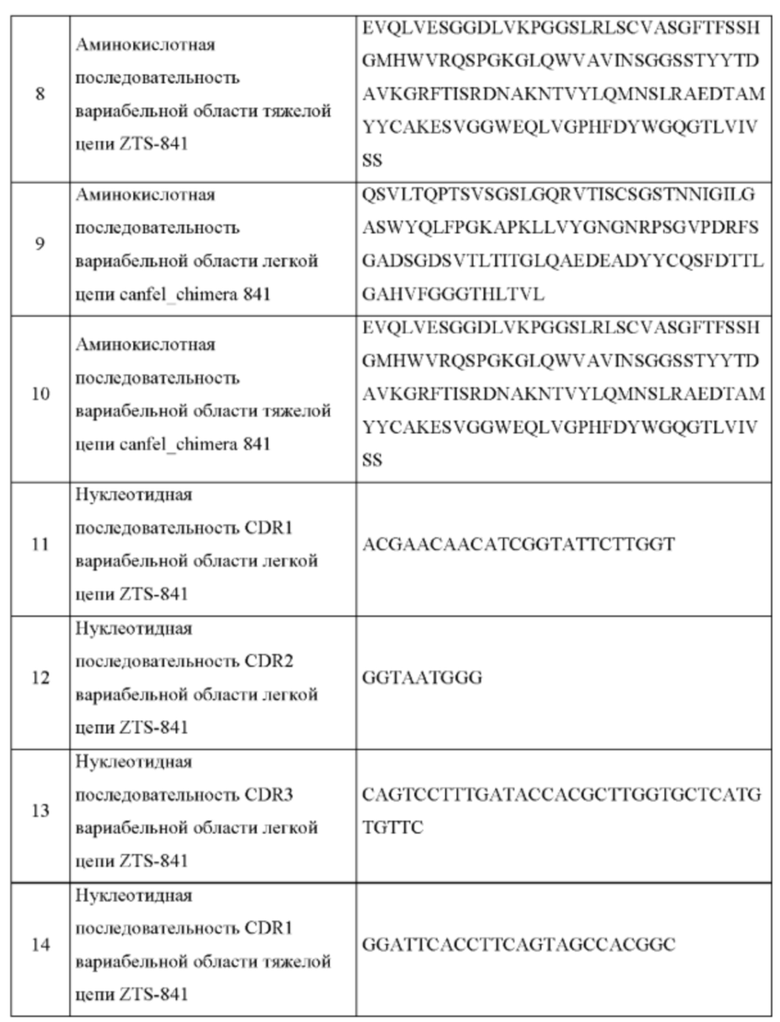
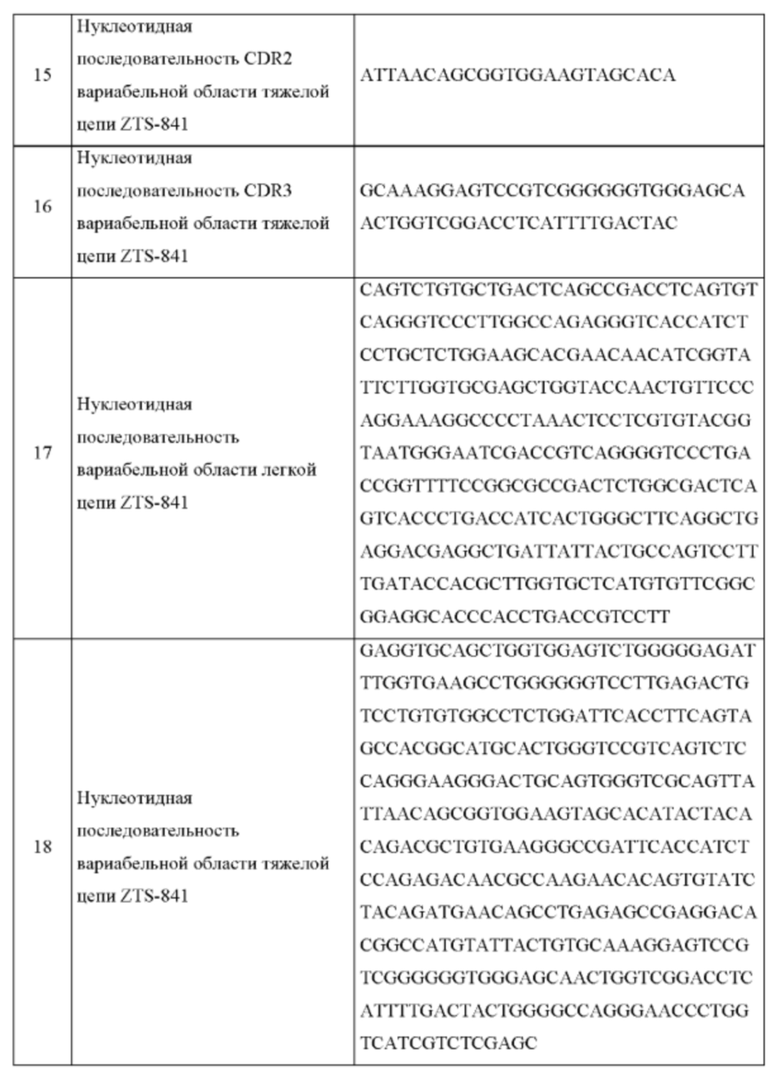

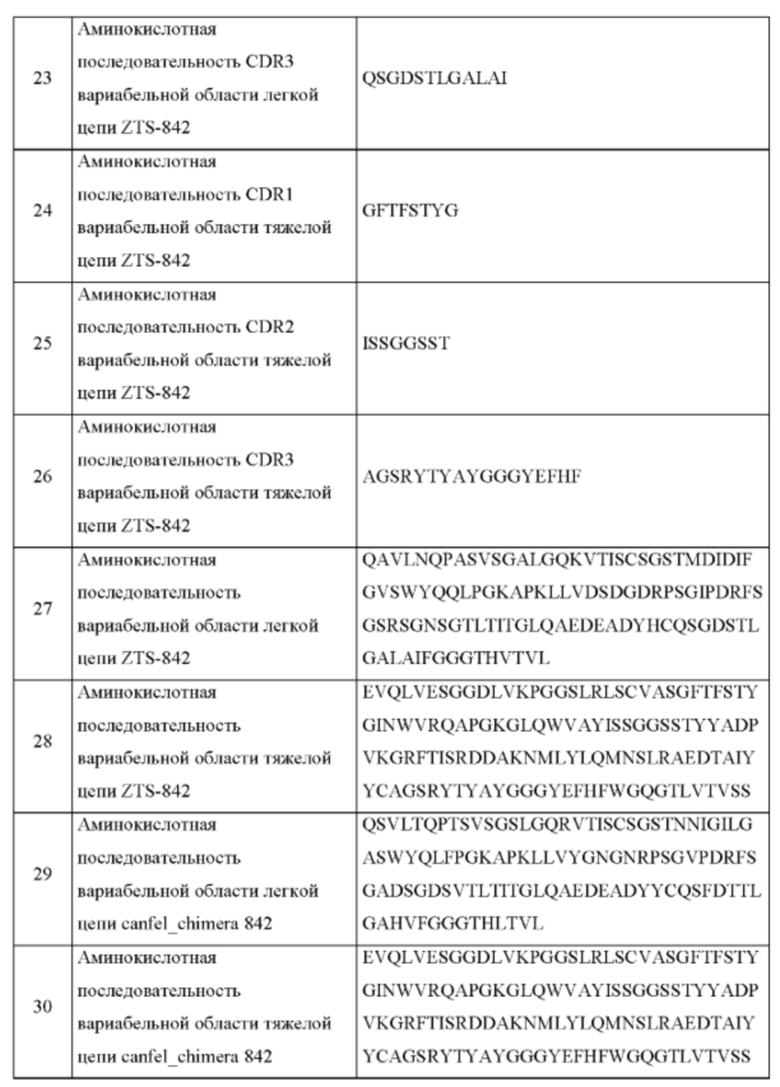

















ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно изобретению, раскрытому здесь, предложены антигенсвязывающие белки против NGF, связывающиеся с NGF с высокой аффинностью. Кроме того, согласно изобретению предложены антигенсвязывающие белки и полипептиды, которые также связываются с NGF и являются вариантами указанных антигенсвязывающих белков, а также способы получения и применения этих антигенсвязывающих белков. В некоторых воплощениях согласно изобретению также предложены полинуклеотиды, кодирующие указанные антигенсвязывающие белки и/или полипептиды. Согласно изобретению, раскрытому здесь, также предложены способы предотвращения и/или лечения боли, включающие введение терапевтически эффективного количества антигенсвязывающих белков против NGF по изобретению.
Общие методики
Следует понимать, что данное изобретение не ограничено конкретными методами, протоколами, реагентами и так далее, описанными здесь, и может, как таковое, варьировать. Термины, использованные здесь, предназначены лишь для описания конкретных воплощений и не ограничивают объем настоящего изобретения, определенный исключительно формулой изобретения.
Если не определено иное, научные и технические термины, использованные в связи с антигенсвязывающими белками, описанными здесь, имеют то значение, в котором их обычно понимают специалисты в данной области. Кроме того, если контекст не требует иного, формы единственного числа включают множественное число, и формы множественного числа включают единственное число. В целом, номенклатура, использованная в связи с культивированием клеток и тканей, молекулярной биологией, а также химией и гибридизацией белков и олиго- или полинуклеотидов, и соответствующие методики, описанные здесь, хорошо известны, широко применяются в данной области и не ограничены единственным описанием. В данной области хорошо известно, что описанные методики могут быть заменены различными другими.
Все указанные патенты и иные публикации прямо включены сюда посредством ссылки с целью описания и раскрытия, например, методов, описанных в таких публикациях, которые могут быть применены в связи с настоящим изобретением. Эти публикации приведены исключительно в силу их раскрытия до даты подачи настоящей заявки.
Для рекомбинантных ДНК, синтеза олигонуклеотидов, культивирования тканей и трансфекции применяют стандартные методики (например, электропорацию, липофекцию). Ферментативные реакции и методики очистки осуществляют в соответствии с описаниями изготовителя или так, как их обычно осуществляют в данной области или как описано здесь. Указанные выше методики обычно осуществляют, следуя общепринятым методам, хорошо известным в данной области, и как описано, без ограничения, в различных общих и более специализированных источниках, цитируемых и обсуждаемых в настоящем описании. См., например, Sambrook et al. MOLECULAR CLONING: LAB. MANUAL (3rd ed, Cold Spring Harbor Lab. Press, Cold Spring Harbor, N.Y., 2001) и Ausubel et at. Current Protocols in Molecular Biology (New York: Greene Publishing Association J Wiley biterscience), Oligonucleotide Synthesis (M.J. Gait, ed.,1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. 1. Freshney, ed. 1987); Introduction to Cell and Tissue Culture (1. P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and С.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction (Mullis et al., eds., 1994); Current Protocols in Immunology (E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane, Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (Y.T. DeVita et al., eds., J.B. Lippincott Company, 1993).
За исключением рабочих примеров или иных случаев, где это указано, все числа, выражающие количество ингредиентов или условия проведения реакций, используемые здесь, следует всегда понимать как модифицированные термином «приблизительно».
Определения
Перед подробным описанием настоящего изобретения будут определены некоторые термины, используемые в контексте настоящего изобретения. По необходимости, в дополнение к этим терминам, в других разделах заявки определены другие термины. Если здесь прямо не определено иное, термины, относящиеся к рассматриваемой области, использованы в данном описании в значениях, принятых в данной области.
Если контекстом ясно не продиктовано иное, при использовании в данном описании и формуле изобретения формы единственного числа включают множественное число. Например, ссылка на «антитело» включает множество таких антител.
При использовании здесь термин «содержащий/включающий» предназначен для обозначения того, что композиции и способы включают указанные элементы, не исключая других.
При использовании здесь термины «фактор роста нервов» и «NGF» относятся к фактору роста нервов и его вариантам, по меньшей мере частично сохраняющим биологическую активность NGF.
«Рецептор NGF» относится к полипептиду, связываемому или активируемому NGF. Рецепторы NGF включают собачий рецептор TrkA и, в меньшей степени, рецептор р75.
«Биологическая активность» NGF обычно относится к способности связываться с рецепторами NGF и/или активировать сигнальные пути рецепторов NGF. Без ограничения, биологическая активность включает любое одно или более из следующего: способность связываться с рецептором NGF (таким как TrkA и/или р75); способность стимулировать димеризацию и/или аутофосфорилирование рецепторов TrkA; способность активировать сигнальный путь рецептора NGF; способность стимулировать дифференцировку, пролиферацию, выживание, рост клеток и другие изменения в физиологии клеток, включая (в случае нейронов, включая периферические и центральные нейроны) изменение морфологии нейронов, синаптогенеза, синаптической функции, высвобождения нейромедиаторов и/или нейропептидов и регенерации после повреждения; способность стимулировать выживание мышиных тригеминальных нейронов Е13.5; и способность опосредовать боль, включая послеоперационную боль.
При использовании здесь «антигенсвязывающий белок против NGF» (взаимозаменяемо называемый «антителом против NGF» и «антителом-антагонистом против NGF», «антигенсвязывающим фрагментом», «антигенсвязывающей частью» и тому подобным) относится к антигенсвязывающему белку, способному связываться с NGF и ингибировать биологическую активность NGF и/или последующий путь (пути), опосредованный сигналами NGF. Антигенсвязывающий белок против NGF охватывает связывающие белки и антитела, которые блокируют, противодействуют, подавляют или снижают (в том числе существенно) биологическую активность NGF, включая последующие пути, опосредованные сигналами NGF, и/или ингибируют связывание NGF с его рецептором TrkA, такое как связывание с рецептором и/или стимуляция клеточного ответа на NGF. Для целей настоящего изобретения следует ясно понимать, что термин «антигенсвязывающий белок против NGF» или «антитело-антагонист против NGF» охватывает все указанные ранее термины, названия и функциональные состояния и характеристики, позволяющие существенным образом устранять, уменьшать или нейтрализовать, в любой значимой степени, сам NGF, биологическую активность NGF (включая, без ограничения, его способность опосредовать любой аспект боли при остеоартрите, воспалительной боли, послеоперационной боли, раковой боли и тому подобного) или последствия его биологической активности. В некоторых воплощениях антитело-антагонист против NGF связывается с NGF и предотвращает димеризацию NGF и/или связывание NGF с рецептором NGF (таким как TrkA и/или р75). В других воплощениях антитело-антагонист против NGF связывается с NGF и предотвращает димеризацию рецептора TrkA и/или аутофосфорилирование TrkA. В настоящей заявке приведены примеры антител-антагонистов против NGF.
При использовании здесь термины «антигенсвязывающий белок», «антитело», «антигенсвязывающий белок» и тому подобные, которые могут быть использованы взаимозаменяемо, относятся к полипептиду или его фрагменту, содержащим антигенсвязывающий сайт.В одном воплощении настоящего изобретения антигенсвязывающий белок по изобретению дополнительно подразумевает иммуноглобулин, способный специфично связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и так далее, через по меньшей мере один антигенраспознающий сайт, расположенный в одной или более чем одной вариабельной области молекулы иммуноглобулина. В некоторых воплощениях антитело имеет две легкие и две тяжелые цепи. Таким образом, выделенное итактное антитело может представлять собой антитело, выделенное из пула поликлональных антител, моноклональное антитело, синтетическое антитело, рекомбинантное антитело, химерное антитело, гетерохимерное антитело или антитело, признанное прошедшим видовую адаптацию, как определено здесь. В некоторых воплощениях термины «антигенсвязывающий белок», «антитело», «антитело-антагонист» и тому подобные предпочтительно относится к моноклональным антителам, их фрагментам и их эквивалентам по иммунологическому связыванию, которые могут связываться с белком NGF и его фрагментами. При использовании здесь данный термин охватывает не только полноразмерные (что, по стандартному определению, означает две тяжелые и две легкие цепи) поликлональные или моноклональные антитела, но также их фрагменты. Если не указано иное, для целей настоящего изобретения «антитело» и «антигенсвязывающий белок» также включают фрагменты антител. Типичные фрагменты антител включают Fab, Fab', F(ab')2, Fv, scFv, Fd, dAb, диатела, их антигенраспознающие фрагменты, малые модульные иммунофармацевтические средства (SMIP), нанотела, молекулы IgNAR и их эквиваленты, являющиеся, по мнению специалиста в данной области, антигенсвязывающими белками или фрагментами антител, и любые из указанных выше фрагментов и их аналогов, полученных в результате химических или генетических манипуляций, а также другие фрагменты антител и их мутанты, слитые белки, содержащие часть антитела, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащую антигенраспознающий сайт. Антитела и антигенсвязывающие белки могут быть получены, например, традиционными гибридомными методиками (Kohler et al., Nature 256:495-499 (1975)), методами рекомбинантных ДНК (патент США №4816567) или методиками фагового дисплея с использованием библиотек антител (Clackson et al., Nature 352:624-628 (1991); Marks et al., J. Mol. Biol. 222:581-597 (1991)). Различные другие методики получения антител описаны в Antibodies: A Laboratory Manual, eds. Harlow et al., Cold Spring Harbor Laboratory, 1988, a также других методиках, хорошо известных специалистам в данной области.
«Моноклональное антитело», как определено здесь, представляет собой антитело, продуцируемое одним клоном клеток (конкретно, одним клоном гибридомных клеток), и поэтому является чистым однородным антителом одного типа. Все моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Моноклональные антитела представляют собой однородную популяцию антител, где моноклональное антитело содержит аминокислоты (встречающиеся в природе и не встречающиеся в природе), которые вовлечены в селективное связывание с антигеном. Популяция моноклональных антител высокоспецифична и направлена против одного антигенного сайта. Термин «моноклональное антитело» охватывает не только интактные моноклональные антитела и полноразмерные моноклональные антитела, но также их фрагменты (Fab, Fab', F(ab')2, Fv, scFv, Fd, dAb, диатела, их антигенраспознающие фрагменты, малые модульные иммунофармацевтические средства (SMIP), нанотела, молекулы IgNAR и тому подобное), их мутанты, слитые белки, содержащие часть антитела, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащую антигенраспознающий сайт желаемой специфичности и способную связываться с антигеном. Он не ограничен источником антитела или способом его получения (например, гибридомой, фаговым дисплеем, рекомбинантной экспрессией, трансгенными животными и так далее).
Здесь моноклональные антитела включают, в частности, «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, имеющих происхождение от определенного вида, в то время как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям антител, имеющих происхождение от другого вида, а также фрагменты таких антител, при условии, что они демонстрируют желаемую биологическую активность. Обычно химерные антитела представляют собой антитела, гены легких и тяжелых цепей которых были сконструированы, обычно методами генной инженерии, из генов вариабельных и константных областей антител, принадлежащих к разным видам. Например, вариабельные сегменты генов мышиного моноклонального антитела могут быть объединены с собачьими константными сегментами. На Фиг. 2 представлено схематическое изображение общей структуры одного воплощения мышиного:собачьего IgG. В данном воплощении антигенсвязывающий сайт имеет происхождение от мыши, в то время как Fc-часть является собачьей.
Термин «гетерохимерный», как определено здесь, относится к антителу, в котором одна из цепей антитела (тяжелая или легкая) канинизирована, в то время как другая является химерной. На Фиг. 4 показано одно воплощение гетерохимерной молекулы. В данном воплощении канинизированная вариабельная тяжелая цепь (где все CDR мышиные, а все FR собачьи) спарена с химерной вариабельной легкой цепью (где все CDR мышиные и все FR мышиные). В данном воплощении как вариабельная тяжелая цепь, так и вариабельная легкая цепь слиты с собачьей константной областью.
Для простоты далее описаны «канинизированные» антитела, однако то же самое применимо к фелинизированному, эквинизированному, гуманизированному антигенсвязывающему белку или антигенсвязывающему белку, прошедшему любую другую «видовую адаптацию». В качестве примера, «канинизацию» определяют как метод переноса информации, обеспечивающей связывание с антигеном, не являющейся собачьей, из антитела-до нора на менее иммуногенное собачье антитело-акцептор с получением препаратов, которые можно применять в качестве терапевтических средств у собак. Канинизированные антитела представляют собой последовательности собачьих антител, в которых остатки гипервариабельных участков реципиента заменены остатками гипервариабельных участков вида, не являющегося собакой (антитело-донор), такого как мышь, крыса, кролик, кошка, собаки, коза, курица, корова, лошадь, лама, верблюд, одногорбые верблюды, акулы, приматы, не являющиеся людьми, человеческой, гуманизированной, рекомбинантной последовательностью или конструированной последовательностью, имеющей желаемые свойства, специфичность, аффинность и активность. Кроме того, канинизированные антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре. Эти модификации проводят для дополнительного улучшения свойств антитела. Модификации гипервариабельных участков и/или каркасных областей, как описано здесь, определяют для каждого отдельно конструируемого антитела, проходящего видовую адаптацию (канинизацию), исходя из экспериментов, известных специалистам в данной области, но их нельзя спрогнозировать до указанных экспериментов. Возможно, канинизированное антитело может содержать полную константную область (Fc) иммуноглобулина, обычно собачьего иммуноглобулина, или по меньшей мере ее часть. На Фиг. 3 представлено одно воплощение, демонстрирующее видовую адаптацию или канинизацию мышиного IgG. Все описания канинизации антигенсвязывающего белка и канинизированного антигенсвязывающего белка могут, в принципе, быть применимы к любому антителу, проходящему или прошедшему видовую адаптацию в форме канинизации, фелинизации, эквинизации, гуманизации и так далее.
Все фразы «рекомбинантное собачье антитело», «рекомбинантное кошачье антитело», «рекомбинантное лошадиное антитело», «рекомбинантное человеческое антитело» и тому подобные включают антитела, прошедшие видовую адаптацию, которые получены, экспрессированы, созданы или выделены рекомбинантный способом, такие как антитела, экспрессированные с использованием рекомбинантного вектора экспрессии, которым была трансфицирована клетка-хозяин, антитела, выделенные из рекомбинантной комбинаторной библиотеки собачьих (или кошачьих, человеческих и так далее) антител, антитела, выделенные из животного (например, мыши), трансгенного по генам собачьих иммуноглобулинов (см., например, Taylor, L.D., et al. (1992) Nucl. Acids Res. 20:6287-6295), или антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, включающим сплайсинг последовательностей генов собачьих (или кошачьих, человеческих и так далее) иммуноглобулинов с другими последовательностями ДНК.
При использовании здесь термины «собачье антитело», «кошачье антитело», «лошадиное антитело», «человеческое антитело» и тому подобные относятся к антителу (антигенсвязывающему белку), выработанному против мишени, и антителам, выделенным из лимфоцитов целевого вида. Эти антитела, как описано здесь, были рекомбинантный образом модифицированы in vitro для включения определенных константных областей целевого вида. Кроме того, антитела, как описано здесь, были идентифицированы, выделены, модифицированы для изменения константной области антитела с последующей экспрессией и выделением из систем культивирования клеток in vitro, известных и обычно используемых специалистами в данной области.
«Нативные антитела» и «нативные иммуноглобулины» обычно представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 Да, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как число дисульфидных связей между тяжелыми цепями иммуноглобулинов разных изотипов варьирует.В каждой тяжелой и легкой цепи также есть регулярно повторяющиеся внутрицепочечные дисульфидные связи. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следуют несколько константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на ее другом конце; константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Полагают, что определенные аминокислотные остатки формируют поверхности вариабельных доменов легкой и тяжелой цепи, обращенные друг к другу. На Фиг. 1 приведен пример общей структуры мышиного иммуноглобулина G (IgG), где подробно показан антигенсвязывающий сайт.
Здесь «исходное» антитело представляет собой антитело, кодируемое аминокислотной последовательностью, используемой для получения варианта. Предпочтительно, исходное антитело имеет собачью каркасную область и, в случае ее (их) присутствия, константную область (области) собачьего антитела. Например, исходное антитело может представлять собой канинизированое или собачье антитело.
В зависимости от аминокислотной последовательности константного домена тяжелых цепей антител, иммуноглобулины могут быть отнесены к различным классам. В настоящее время выделяют пять основных классов иммуноглобулинов, IgA, IgD, IgE, IgG, и IgM, и некоторые из них могут быть дополнительно разделены на подклассы (изотипы), например, IgGi, IgG2, IgG3, IgG4, IgA и IgA2 (как их принято обозначать у мышей и человека). Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, называют альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны у многих видов. Распространенность отдельных изотипов и функциональная активность, связанная с этими константными доменами, видоспецифичны и нуждаются в экспериментальном определении.
«Легкие цепи» антител (иммуноглобулинов) любого вида позвоночных могут быть отнесены к одному из двух четко различающихся типов, называемых каппа (κ) и лямбда (λ), исходя из аминокислотных последовательностей их константных доменов.
«Вариабельная область» антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела, самим по себе или в комбинации. Каждая вариабельная область тяжелой и легкой цепи состоит из четырех каркасных областей (FR), соединенных тремя участками, определяющими комплементарность (CDR), также известными как гипервариабельные участки. В каждой цепи FR удерживают CDR в непосредственной близости друг от друга, и они, вместе с CDR другой цепи, участвуют в образовании антигенсвязывающего сайта антитела. Существует по меньшей мере две методики определения CDR: (1) способ, основанный на межвидовой вариабельности последовательностей (то есть Kabat et al. Sequences of Proteins of Immunological Interest, (5th ed, 1991, National Institutes of Health, Bethesda Md.)); и (2) способ, основанный на кристаллографических исследованиях комплексов антиген-антитело (Chothia etal. (1989) Nature 342:877; AI-Iazikani etal. (1997) J. Molec. Bioi. 273:927-948)). При использовании здесь CDR может относиться к CDR, определенным любым из этих способов или сочетанием обоих этих способов.
При использовании здесь термин «гипервариабельный участок» относится к аминокислотным остаткам антитела, обеспечивающим связывание с антигеном. Гипервариабельный участок содержит аминокислотные остатки «участка, определяющего комплементарность», или «CDR» (Kabat et al. (1991) выше) и/или остатки «гипервариабельной петли» (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). Остатки «каркасных областей» или «FR» представляют собой остатки вариабельных доменов, не являющиеся остатками гипервариабельных участков, как определено здесь.
При использовании здесь термин «антигенсвязывающая область» относится к той части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и придающие антителу его специфичность и аффинность в отношении антигена. Связывающая область антитела содержит «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков.
«Функциональная Fc-область» обладает по меньшей мере одной эффекторной функцией Fc-области с нативной последовательностью. Типичные «эффекторные функции» включают связывание с C1q, комплемент-зависимую цитотоксичность (CDC), связывание с Fc-рецепторами, связывание с неонатальными рецепторами, антителозависимую клеточно-опосредованную цитотоксичность (ADCC), фагоцитоз, понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточных рецепторов, BCR) и так далее. Для таких эффекторных функций обычно необходимо сочетание Fc-области со связывающим доменом (например, вариабельным доменом антитела), и их можно оценить с применением различных анализов, известных в данной области, для оценки таких эффекторных функций антител.
«Fc-область с нативной последовательностью» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруженной в естественных условиях. «Вариант Fc-области» или «мутированная» или «мутантная» Fc-область содержит аминокислотную последовательность, отличающуюся от Fc-области с нативной последовательностью по меньшей мере одной аминокислотной модификацией, и может сохранять или не сохранять по меньшей мере одну эффекторную функцию Fc-области с нативной последовательностью. Предпочтительно, вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например, от приблизительно одной до приблизительно десяти аминокислотных замен и, предпочтительно, от приблизительно одной до приблизительно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. Здесь вариант Fc-области будет по меньшей мере приблизительно на 80% идентичен Fc-области с нативной последовательностью и/или Fc-области исходного полипептида, и, наиболее предпочтительно, по меньшей мере приблизительно на 90% идентичен указанной Fc-области, более предпочтительно, по меньшей мере приблизительно на 95% идентичен указанной Fc-области. Вариант Fc-области или мутированная Fc-область могут также по существу устранять функцию Fc-области антитела. Например, мутации Fc-области могут устранять эффекторную функцию антитела. В одном воплощении изобретения антитело по изобретению содержит мутированную Fc-область.
При использовании здесь «Fc-рецептор» и «FcR» описывают рецептор, связывающийся с Fc-областью антитела. Предпочтительным FcR является FcR с нативной последовательностью. Более того, предпочтительный FcR представляет собой FcR, связывающийся с антителом IgG (гамма-рецептор), и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и формы альтернативного сплайсинга этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), имеющие сходные аминокислотные последовательности и различающиеся главным образом своими цитоплазматическими доменами. Обзоры FcR приведены в Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9:457-92; Capel et at., 1994, Immunomethods, 4:25-34; и de Haas et at., 1995, J. Lab. Clin. Med., 126:330-41. «FcR» также включает неонатальный рецептор FcRn, обеспечивающий перенос материнских IgG к плоду (Guyer et at., 1976, J. Immunol., 117:587; и Kim et at., 1994, J. Immunol., 24:249).
При использовании здесь «антителозависимая клеточно-опосредованная цитотоксичность» и «ADCC» относятся к клеточно-опосредованной реакции, при которой неспецифические цитотоксические клетки, экспрессирующие Fc-рецепторы (FcR) (например, натуральные клетки-киллеры (NK), нейтрофилы и макрофаги) распознают связанное антитело на клетке-мишени и приводят впоследствии к лизису клетки-мишени. ADCC-активность интересующей молекулы может быть оценена с применением анализа ADCC in vitro, такого как описанный в патенте США №5500362 или 5821337. Эффекторные клетки, которые могут быть использованы для таких анализов, включают мононуклеарные клетки периферической крови (РВМС) и NK-клетки. Альтернативно или дополнительно, ADCC-активность интересующей молекулы может быть оценена in vivo, например, в модели на животных, такой как раскрытая в Clynes et al., 1998, PNAS (USA), 95:652-656.
«Комплемент-зависимая цитотоксичность» и «CDC» относятся к лизису мишени в присутствии комплемента. Путь активации комплемента начинается со связывания первого компонента системы комплемента (C1q) с молекулой (например, антителом), представленной в комплексе с когнатным антигеном. Для оценки активации комплемента может быть проведен анализ CDC, например, как описано в Gazzano-Santoro et at., J. Immunol. Methods, 202: 163 (1996).
Расщепление антител папаином приводит к получению двух идентичных антигенсвязывающих фрагментов, называемых «Fab-фрагментами», у каждого из которых есть один антигенсвязывающий сайт, и остаточного «Fc-фрагмента», название которого отражает его способность к быстрой кристаллизации. При обработке пепсином образуется F(ab')2-фрагмент, имеющий два антигенсвязывающих сайта и все еще способный к перекрестному связыванию антигена.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конце домена СН1 тяжелой цепи, включая один или более чем один цистеин шарнирной области антитела. Fab'-SH обозначает здесь Fab', в котором цистеиновый остаток (остатки) константного домена имеют свободную тиоловую группу. F(ab')2-фрагменты антител были исходно получены как пары Fab'-фрагментов с цистеинами шарнирной области между ними. Также известны другие химические сочетания фрагментов антител.
«Fv» представляет собой минимальный фрагмент антитела, содержащий полный антигенраспознающий и антигенсвязывающий сайт.Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочными нековалентными взаимодействиями. Именно в этой конфигурации три гипервариабельных участка каждого вариабельного домена взаимодействуют, определяя антигенсвязывающий сайт на поверхности димера VH-VL. Вместе эти шесть гипервариабельных участков обеспечивают специфичность связывания антитела с антигеном. Тем не менее, даже отдельный вариабельный домен (или половина Fv, содержащая только три гипервариабельных участка, специфичных в отношении антигена) обладает способностью к распознаванию и связыванию антигена, хотя и с меньшей аффинностью, чем целый связывающий сайт.
При использовании здесь «антиген» относится к антигенной детерминанте, распознаваемой CDR антигенсвязывающего белка или антитела, как описано здесь. Иными словами, эпитоп относится к той части любой молекулы, которая может быть распознана и связана антителом. Если не указано иное, при использовании здесь термин «эпитоп» относится к той области NGF, с которой связывается антигенсвязывающий белок / антитело / агент против NGF.
Термины «антигенсвязывающий домен», «активные фрагменты антитела» или тому подобные относится к той части антитела или антигенсвязывающего белка, которая содержит область, специфично связвающуюся со всем антигеном или его частью или комплементарную всему антигену или его части. При больших размерах антигена антитело может связываться только с определенной частью антигена. «Эпитоп», «активные фрагменты эпитопа», «антигенная детерминанта» или тому подобное представляют собой ту часть молекулы антигена, которая обеспечивает специфичное взаимодействие с антигенсвязывающий доменом антитела. Антигенсвязывающий домен может быть обеспечен одним или более чем одним вариабельным доменом антитела (например, так называемым Fd-фрагментом антитела, содержащим домен VH). Антигенсвязывающий домен может содержать вариабельный домен легкой цепи антитела (VL) и вариабельный домен тяжелой цепи антитела (VH) (патент США №5565332).
Термины «связывающая часть» антитела (или «часть антитела») или антигенсвязывающего полипептида или тому подобные включают один или более чем один полный домен, например, пару полных доменов, а также фрагменты антитела, сохраняющие способность к специфичному связыванию с антигеном, например, NGF. Показано, что связывающая функция антитела может быть выполнена фрагментами полноразмерного антитела. Связывающие фрагменты получают методиками рекомбинантных ДНК или ферментативным или химическим расщепление интактных иммуноглобулинов. Связывающие фрагменты включают Fab, Fab', F(ab')2, F(abc), Fd, dAb, Fv, одиночные цепи, одноцепочечные антитела, например, scFv, однодоменные антитела (Muyldermans et al., 2001, 26:230-5) и выделенный участок, определяющий комплементарность (CDR). Fab-фрагмент представляет собой моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1. F(ab')2-фрагмент представляет собой бивалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидной связью в шарнирной области. Fd-фрагмент состоит из доменов VH и СН1, и Fv-фрагмент состоит из доменов VL и VH одной половины антитела. dAb-фрагмент состоит из домена VH (Ward et al., (1989) Nature 341:544-546). Несмотря на то, что два домена Fv-фрагмента, VL и VH, кодированы раздельными генами, они могут быть соединены синтетическим линкером с применением рекомбинантных способов, что позволяет получать их в форме одной белковой цепи, в которой области VL и VH представлены в паре, образуя моновалентные молекулы (известные как одноцепочечные Fv (scFv) (Bird et al., 1988, Science 242:423-426)). Подразумевают, что термин «связывающая часть» антитела охватывает также такие одноцепочечные антитела. Он также охватывает другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой бивалентные биспецифичные антитела, в которых домены VH и VL экспрессированы на одной полипептидной цепи, но с использованием линкера, длина которого слишком мала для образования пары из двух доменов, расположенных на одной и той же цепи, что приводит к образованию пар из комплементарных доменов, расположенных на разных цепях, с формированием двух антигенсвязывающих сайтов (см., например, Holliger, et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-6448). Антитело или его связывающая часть могут также быть частью более крупных иммуноадгезионных молекул, образованных в результате ковалентного или не ковалентного объединения антитела или части антитела с одним или более чем одним другим белком или пептидом. Примеры таких иммуноадгезионных молекул включают использование стрептавидиновой центральной области с получением тетрамерной молекулы scFv (Kipriyanov, S. М., et al. (1995) Human Antibodies and Hybridomas 6:93-101) и использование цистеинового остатка, маркерного пептида и С-концевой полигистидиновой метки с получением бивалентных и биотинилированных молекул scFv (Kipriyanov, S. М., et al. (1994) Mol. Immunol. 31:1047-1058). Связывающие фрагменты, такие как Fab- и Е(ab')2-фрагменты, могут быть получены из целых антител с применением обычных методик, таких как расщепление целых антител папаином или пепсином, соответственно. Более того, антитела, части антител и иммуноадгезионные молекулы могут быть получены с применением стандартных методик рекомбинантных ДНК, как описано здесь и известно в данной области. Следует понимать, что связывающие сайты антитела, за исключением «биспецифичных» или «бифункциональных» антител, идентичны друг другу. «Биспецифичное» или «бифункциональное антитело» представляет собой искусственное гибридное антитело, имеющее две разные пары тяжелых/легких цепей и два разных связывающих сайта. Биспецифичное антитело может также содержать две антигенсвязывающие области, разделенные константной областью. Биспецифичные антитела могут быть получены с применением множества методов, включая слияние гибридом или соединение Fab'-фрагментов. См., например, Songsivilai et al., Clin. Exp.Immunol. 79:315-321, 1990; Kostelny et al., 1992, J. Immunol. 148, 1547-1553.
Термин «обратная мутация» относится к замене некоторых или всех аминокислот собачьего антитела, подверженных соматическим мутациям, на соответствующие остатки эмбрионального типа из гомологичной последовательности антитела эмбрионального типа. Проводят раздельное выравнивание последовательностей тяжелой и легкой цепи собачьего антитела по изобретению с последовательностями эмбрионального типа для определения последовательностей с наибольшей гомологией. Отличия собачьего антитела по изобретению от последовательности эмбрионального типа устраняют, проводя мутации в определенных нуклеотидных положениях, кодирующих такие отличающиеся аминокислоты. Роль каждой аминокислоты, идентифицированной таким образом в качестве кандидата на обратную мутацию, следует изучить на предмет прямой или косвенной роли в связывании с антигеном, и любую аминокислоту, которая, исходя из результатов мутации, влияет на любую желаемую характеристику собачьего антитела, не следует включать в окончательное собачье антитело; в качестве примера, аминокислоты, усиливающие активность, идентифицированные таким методом селективного мутагенеза, не будут подвергать обратным мутациям. Для минимизации числа аминокислот, подвергаемых обратным мутациям, аминокислотные положения, отличающиеся от ближайшей последовательности эмбрионального типа, но идентичные соответствующим аминокислотам во второй последовательности эмбрионального типа, могут быть сохранены, при условии, что вторая последовательность эмбрионального типа идентична и колинейна последовательности собачьего антитела по изобретению. Обратные мутации выбранных целевых каркасных остатков на соответствующие донорные остатки могут быть необходимы для восстановления и/или улучшения аффинности.
При использовании здесь «иммуноспецифичное» связывание антител относится к антигенспецифичному связывающему взаимодействию, происходящему между антигенсвязывающим сайтом антитела и специфическим антигеном, распознаваемым этим антителом (то есть антитело взаимодействует с белком в ELISA или другом иммунологическом анализе и не взаимодействует, поддающимся выявлению образом, с неродственными белками). Эпитоп, который «специфично связывается» или «преимущественно связывается» (использованы здесь взаимозаменяемо) с антителом или полипептидом, представляет собой термин, хорошо известный в данной области, и способы определения такого специфичного или преимущественного связывания также хорошо известны в данной области. Говорят, что молекула демонстрирует «специфичное связывание» или «преимущественное связывание», если она взаимодействует или связывается с определенной клеткой или определенным веществом чаще, быстрее, на более продолжительное время и/или с большей аффинностью, чем с альтернативными клетками или веществами. Антитело «специфично связывается» или «преимущественно связывается» с мишенью, если оно связывается с ней с большей аффинностью, авидностью, быстрее и/или на более продолжительное время, чем с другими веществами. Например, антитело, которое специфично или преимущественно связывается с эпитопом NGF, представляет собой антитело, которое связывается с этим эпитопом с большей аффинностью, авидностью, быстрее и/или на более продолжительное время, чем с другими эпитопами NGF или эпитопами, не являющимися эпитопами NGF. Читая данное определение, также следует понимать, что, например, антитело (или группировка или эпитоп), которое специфично или преимущественно связывается с первой мишенью, может специфично или преимущественно связываться со второй мишенью или не связываться с ней. Исключительное связывание не является обязательно необходимым для «специфичного связывания» или «преимущественного связывания» как таковых (но «специфичное связывание» или «преимущественное связывание» могут включать исключительное связывание). В целом, но не обязательно, ссылка на связывание обозначает преимущественное связывание.
Термин «специфично» в контексте связывания антитела относится к высокой авидности и/или высокой аффинности связывания антитела со специфическим антигеном, то есть полипептидом, или эпитопом. Специфичное связывание антитела с антигеном сильнее связывания того же антитела с другими антигенами. Антитела, специфично связывающиеся с полипептидом, могут быть способны связываться с другими полипептидами на слабом, но поддающемся выявлению уровне (например, 10% или менее от показанного связывания с интересующим полипептидом). Такое слабое связывание или фоновое связывание легко отличить от специфичного связывания антитела с соответствующим полипептидом, например, используя подходящие контроли. Обычно аффинность связывания специфичных антител с антигеном соответствует KD 10-7 М или менее, 10-8 М или менее, 10-9 М или менее, 10-10 М или менее, 10-11 М или менее, 10-12 М или менее, или 10-13 М или менее, и так далее.
При использовании здесь термин «аффинность» относится к силе связывания отдельного антигенсвязывающего сайта с антигенной детерминантой. Аффинность зависит от близости стереохимического соответствия между связывающими сайтами антитела или антигенсвязывающего белка и антигенными детерминантами, от размера площади контакта между ними, от распределения заряженных и гидрофобных групп и так далее. Аффинность антитела можно измерить равновесным анализом или методом поверхностного плазмонного резонанса («SPR», например, BIACORE™). Метод SPR основан на явлении поверхностного плазмонного резонанса (SPR), возникающего при возбуждении поверхностных плазмонных волн на границе металла и жидкости. Свет направляется на край поверхности, не контактирующей с образцом, и отражается от нее, при этом SPR приводит к снижению интенсивности отраженного света при определенном сочетании угла и длины волны. Явления бимолекулярного связывания приводят к изменению коэффициента преломления поверхности, которые выявляются как изменения SPR-сигнала.
При использовании здесь термин «KD» предназначен для обозначения константы диссоциации при взаимодействии антитела с антигеном. Константа диссоциации, KD, и константа ассоциации, Ка, являются количественными мерами аффинности. При равновесии свободный антиген (Ag) и свободное антитело (Ab) находятся в равновесии с комплексом антиген-антитело (Ag-Ab), и константы скорости, ka и kd, являются количественными показателями скоростей отдельных реакций. При равновесии ka[Ab][Ag] равна kd[Ag-Ab]. Константу диссоциации, Kd, рассчитывают следующим образом: Kd=kd/ka=[Ag][Ab]/[Ag-Ab]. KD измеряют в единицах концентрации, чаще всего в М, мМ, мкМ, нМ, пМ и так далее. При сравнении аффинности антител, выраженной как KD, большей аффинности в отношении NGF соответствуют меньшие значения. Константу ассоциации, Ka, рассчитывают следующим образом: Ka=ka/kd=[Ag-Ab]/[Ag][Ab]. Ka измеряют в единицах, обратных концентрации, чаще всего в М-1, мМ-1, мкМ-1, нМ-1, пМ-1 и так далее. При использовании здесь термин «авидность» относится к прочности связи антигена с антителом после обратимого образования комплексов. Антитела против NGF могут быть охарактеризованы по KD их связывания с белком NGF как «связывающиеся с константой диссоциации (KD) в диапазоне от приблизительно (меньшее значение KD) до приблизительно (большее значение KD)».
Термины «полипептид», «олигопептид», «пептид» и «белок» использованы здесь взаимозаменяемо для обозначения полимеров аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может прерываться не аминокислотными компонентами. Эти термины также включают аминокислотный полимер, модифицированный естественным или искусственным образом; например, образованием дисульфидных связей, гликозилированием, липидированием, ацетилированием, фосфорилированием или любой другой манипуляцией или модификацией, такой как конъюгация с компонентом, содержащим метку. В это определение также включены, например, полипептиды, содержащие один или более чем один аналог аминокислот (включая, например, искусственные аминокислоты и так далее), а также другие модификации, известные в данной области. Следует понимать, что, поскольку полипептиды по данному изобретению основаны на антителе, полипептиды могут иметь форму отдельных цепей или связанных цепей.
Термин «консервативная аминокислотная замена» указывает на любую аминокислотную замену заданного аминокислотного остатка, где заменяющий остаток химически настолько сходен с заданным остатком, что замена не приводит к существенному снижению функции (например, ферментативной активности) полипептида. Консервативные аминокислотные замены хорошо известны в данной области, и их примеры описаны, например, в патентах США №№6790639, 6774107, 6194167 или 5350576. В предпочтительном воплощении консервативная аминокислотная замена будет представлять собой любую замену, происходящую в пределах одной из следующих шести групп:
• малые алифатические практически неполярные остатки: Ala, Gly, Pro, Ser и Thr;
• большие алифатические неполярные остатки: Ile, Leu и Val; Met;
• полярные отрицательно заряженные остатки и их амиды: Asp и Glu;
• амиды полярных отрицательно заряженных остатков: Asn и Gln; His;
• полярные положительно заряженные остатки: Arg и Lys; His; и
• большие ароматические остатки: Trp и Tyr; Phe.
В предпочтительном воплощении консервативная аминокислотная замена будет представлять собой любую из следующих замен, перечисленных как пары «нативный остаток (консервативная замена)»: Ala (Ser); Arg (Lys); Asn (Gln; His); Asp (Glu); Gln (Asn); Glu (Asp); Gly (Pro); His (Asn; Gln); Ile (Leu; Val); Leu (Ile; Val); Lys (Arg; Gln; Glu); Met (Leu; Ile); Phe (Met; Leu; Tyr); Ser (Thr); Thr (Ser); Trp (Tyr); Tyr (Trp; Phe); и Val (Ile; Leu).
Термины «нуклеиновая кислота», «полинуклеотид», «молекула нуклеиновой кислоты» и тому подобные могут быть использованы здесь взаимозаменяемо для обозначения серии нуклеотидных оснований (называемых также «нуклеотидами») в ДНК и РНК. Нуклеиновая кислота может содержать дезоксирибонуклеотиды, рибонуклеотиды и/или их аналоги. Термин «нуклеиновая кислота» включает, например, одноцепочечные и двуцепочечные молекулы. Нуклеиновая кислота может представлять собой, например, ген или фрагмент гена, экзоны, интроны, молекулу ДНК (например, кДНК), молекулу РНК (например, мРНК), рекомбинантные нуклеиновые кислоты, плазмиды и другие векторы, праймеры и зонды. Включены как 5'-3'-полинуклеотиды (смысловые), так и 3'-5'-полинуклеотиды (антисмысловые). Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. В случае проведения модификаций нуклеотидная структура может быть модифицирована до или после сборки полимера. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. После полимеризации возможна дополнительная модификация полинуклеотид а, такая как конъюгация с компонентом для мечения. Другие типы модификаций включают, например, «кэпы», замену одного или более чем одного из встречающихся в природе нуклеотидов на аналог, межнуклеотидные модификации, такие как, например, модификации с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфорамидаты, карбаматы и так далее) и с заряженными связями (например, фосфоротиоаты, фосфородитиоаты и так далее), модификации, включающие боковые группировки, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и так далее), модификации с использованием интеркаляторов (например, акридина, псоралена и так далее), модификации, включающие хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и так далее), модификации, включающие алкилирующие агенты, модификации, включающие модифицированные связи (например, альфа-аномерные нуклеиновые кислоты и так далее), а также немодифицированные формы полинуклеотида (полинуклеотидов). Кроме того, любые из гидроксильных групп, обычно присутствующих в сахарах, могут быть замещены, например, фосфонатными группами, фосфатными группами, защищенными стандартными защитными группами, или активированы для образования дополнительных связей с дополнительными нуклеотидами, или могут быть конъюгированы с твердыми подложками. 5'- и 3'-концевые ОН могут быть фосфорилированы или замещены аминами или группировками органических кэпирующих групп, содержащими от 1 до 20 атомов углерода. Другие гидроксилы могут также быть дериватизированы с образованием стандартных защитных групп.Полинуклеотиды могут также содержать аналогичные формы рибозных или дезоксирибозных сахаров, которые обычно известны в данной области, включая, например, 2'-O-метил-, 2'-O-аллил-, 2'-фтор-или 2'-азидорибозу, карбоциклические аналоги Сахаров, аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и нуклеозидные аналоги без оснований, такие как метилрибозид. Одна или более чем одна фосфодиэфирная связь может быть заменена альтернативными связывающими группами. Эти альтернативные связывающие группы включают, без ограничения, воплощения, где фосфат заменен на P(O)S («тиоат»), P(S)S («дитиоат»), (O)NR2 («амидат»), P(O)R, P(O)OR', СО или СН2 («формацеталь»), где каждый R или R' независимо представляет собой Н или замещенный или незамещенный алкил (C1-20), возможно, содержащий эфирную связь (-О-), арил, алкенил, циклоалкил, циклоалкенил или аралкил. Нет необходимости в том, чтобы все связи в полинуклеотиде были идентичны. Предшествующее описание применимо ко всем полинуклеотидам, упомянутым здесь, включая РНК и ДНК.
При использовании здесь «вектор» обозначает конструкцию, которая позволяет доставлять и, предпочтительно, экспрессировать один или более чем один ген или одну или более чем одну последовательность в клетке-хозяине. Примеры векторов включают, без ограничения, вирусные векторы, векторы экспрессии на основе депротеинизированной ДНК или РНК, плазмидные, космидные или фаговые векторы, векторы экспрессии на основе ДНК или РНК, связанные с катионными конденсирующими агентами, векторы экспрессии на основе ДНК или РНК, инкапсулированные в липосомы, и определенные эукариотические клетки, такие как клетки-продуценты. Векторы, как описано здесь, имеют последовательности контроля экспрессии, под которыми понимают последовательность нуклеиновой кислоты, управляющую транскрипцией нуклеиновой кислоты. Последовательность контроля экспрессии может представлять собой промотор, такой как конститутивный или индуцируемый промотор, или энхансер. Последовательность контроля экспрессии «функционально связана» с транскрибируемой последовательностью нуклеиновой кислоты. Нуклеиновая кислота является «функционально связанной» при наличии функциональной связи между ней и другой последовательностью нуклеиновой кислоты. Например, ДНК пре-последовательности или секреторной лидерной последовательности функционально связана с ДНК полипептида в случае ее экспрессии в форме пре-белка, участвующего в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он, в силу его расположения, способствует трансляции. Обычно «функционально связанный» означает, что связанные последовательности ДНК являются смежными и, в случае секреторной лидерной последовательности, смежными в фазе считывания. Тем не менее, энхансеры не обязательно являются смежными. Связывание осуществляют лигированием в подходящих сайтах рестрикции. При отсутствии таких сайтов используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с обычной практикой.
Точно так же, как полипептид может содержать консервативную аминокислотную замену (замены), соответствующий ему полинуклеотид может содержать консервативную замену (замены) кодона. Замену кодона считают консервативной, если при экспрессии она приводит к консервативной аминокислотной замене, как описано выше. В полинуклеотид ах по настоящему изобретению может также быть применена вырожденная замена кодона, которая не приводит к аминокислотной замене. Таким образом, например, возможна мутация полинуклеотида, кодирующего выбранный полипептид, который может быть использован в одном воплощении настоящего изобретения, посредством вырожденной замены кодона для приближения частоты использования кодонов к той, которую демонстрирует экспрессионная клетка-хозяин, подлежащая трансформации указанным полинуклеотидом, или для иного улучшения его экспрессии.
«Вариант» антигенсвязывающего белка против NGF относится здесь к молекуле, отличающейся по аминокислотной последовательности от аминокислотной последовательности «исходного» антитела против NGF добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка в последовательности исходного антитела и сохраняющей по меньшей мере одну желаемую активность исходного антитела против NGF. Вариант против NGF может содержать консервативные аминокислотные замены в гипервариабельном участке антитела, как описано здесь. Желаемые активности могут включать способность к специфичному связыванию с антигеном, способность к снижению, ингибированию или нейтрализации активности NGF у животного. В одном воплощении вариант содержит одну или более чем одну аминокислотную замену в одном или более чем одном гипервариабельном участке и/или одной или более чем одной каркасной области исходного антитела. Например, вариант может содержать по меньшей мере одну, например, от приблизительно одной до приблизительно десяти и предпочтительно от приблизительно двух до приблизительно пяти замен в одном или более чем одном гипервариабельном участке и/или одной или более чем одной каркасной области исходного антитела. Обычно вариант будет иметь аминокислотную последовательность, по меньшей мере на 50% идентичную, более предпочтительно по меньшей мере приблизительно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% идентичную аминокислотным последовательностям вариабельных доменов тяжелой или легкой цепи исходного антитела. Идентичность или гомология применительно к данной последовательности определены здесь как процент аминокислотных остатков в последовательности-кандидате, идентичных остаткам исходного антитела, после выравнивания последовательностей и, по необходимости, введения разрывов для достижения максимального процента идентичности последовательностей. N-концевые, С-концевые или внутренние удлинения, делеции или вставки в последовательности антитела не следует рассматривать как влияющие на идентичность или гомологию последовательностей. Вариант сохраняет способность к связыванию с NGF и предпочтительно имеет желаемые активности, равные соответствующим активностям исходного антитела или превосходящие их. Например, вариант может иметь более высокую аффинность связывания, усиленную способность к снижению, ингибированию или нейтрализации активности NGF у животного и/или усиленную способность к ингибированию связывания NGF с TrkA и р75.
TrkA, который принято считать высокоаффинным рецептором NGF, является представителем семейства тирозинкиназных рецепторов нейротрофинов (NTKR). Эта киназа представляет собой мембраносвязанный рецептор, который, при связывании с нейротрофином, фосфорилирует самого себя (аутофосфорилирование) и представителей пути MAPK. Присутствие этой киназы приводит к дифференцировке клеток и может играть роль в определении подтипов чувствительных нейронов. Рецептор р75 принято считать низкоаффинным рецептором NGF.
«Вариант» нуклеиновой кислоты относится здесь к молекуле, последовательность которой отличается от «исходной» нуклеиновой кислоты. Расхождение полинуклеотидных последовательностей может быть результатом мутационных изменений, таких как делеции, замены или добавления одного или более чем одного нуклеотида. Каждое из этих изменений может присутствовать в заданной последовательности само по себе или в комбинации, один или более чем один раз.
Термин «выделенный» означает, что вещество (например, антигенсвязывающий белок, как описано здесь, или нуклеиновая кислота) отделено и/или выделено из компонента его естественного окружения. Примесные компоненты его естественного окружения являются веществами, которые могут препятствовать диагностическому или терапевтическому применению вещества, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. Применительно к нуклеиновой кислоте выделенная нуклеиновая кислота может включать нуклеиновую кислоту, отделенную от 5'-3'-последовательностей, с которыми она обычно связана в хромосоме. В предпочтительных воплощениях степень очистки вещества будет составлять более чем 95% по массе вещества и, наиболее предпочтительно, более чем 99% по массе. Выделенное вещество включает вещество in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент естественного окружения вещества будет отсутствовать. Однако обычно выделенное вещество будет получено посредством по меньшей мере одной стадии очистки.
При использовании здесь термины «клетка», «клеточная линия» и «клеточная культура» могут быть использованы взаимозаменяемо. Эти термины также включают их потомство, представляющее собой все последующие поколения. Следует понимать, что все потомство может не быть идентичным из-за преднамеренных или непреднамеренных мутаций. В контексте экспрессии последовательности гетерологичной нуклеиновой кислоты «клетка-хозяин» относится к прокариотической или эукариотической клетке (например, бактериальным клеткам, дрожжевым клеткам, клеткам млекопитающих и клеткам насекомых), независимо от их нахождения in vitro или in vivo. Например, клетки-хозяева могут находиться в трансгенном животном. Клетка-хозяин может быть использована в качестве реципиента векторов и может включать любой организм, поддающийся трансформации, способный реплицировать вектор и/или экспрессировать гетерологичную нуклеиновую кислоту, кодируемую вектором.
При использовании здесь слово «метка» относится к соединению или композиции, подающимся выявлению и конъюгированным прямым или непрямым образом с антителом или нуклеиновой кислотой. Сама метка может поддаваться выявлению сама по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическое изменение соединения-субстрата или композиции-субстрата, которое поддается выявлению.
«Субъект» или «пациент» относится к животному, нуждающемуся в лечении, которое может быть проведено молекулами по изобретению. Животные, которых можно лечить согласно изобретению, включают позвоночных, при этом особенно предпочтительными примерами являются млекопитающие, такие как собаки.
Термин «композиция» предназначен для обозначения комбинации активного агента, который может представлять собой любое из химической композиции, биологической композиции или биотерапевтического средства (в частности антигенсвязывающих белков, как описано здесь), и другого соединения или другой композиции, которые могут быть инертными (например, метка) или активными, такими как адъювант.
Как определено здесь, «фармацевтически приемлемые носители», подходящие для использования в изобретении, хорошо известны специалистам в данной области. Такие носители включают, без ограничения, воду, физиологический раствор, забуференный физиологический раствор, фосфатный буфер, спиртовые/водные растворы, эмульсии или суспензии. Согласно общепринятым методикам, могут быть добавлены другие часто используемые разбавители, адъюванты и эксципиенты. Такие носители могут включать этанол, полиолы и их подходящие смеси, растительные масла и органические сложные эфиры для инъекций. Могут также быть использованы буферы и агенты для коррекции рН. Буферы включают, без ограничения, соли, полученные из органической кислоты или основания. Типичные буферы включают, без ограничения, соли органических кислот, такие как соли лимонной кислоты, цитраты, соли аскорбиновой кислоты, глюконовой кислоты, гистидин-HCl, соли угольной кислоты, винной кислоты, янтарной кислоты, уксусной кислоты или фталевой кислоты, трис, гидрохлорид трометамина или фосфатные буферы. Носители для парентерального введения могут включать раствор хлорида натрия, декстрозу Рингера, декстрозу, трегалозу, сахарозу и хлорид натрия, лактат Рингера или нелетучие масла. Носители для внутривенного введения могут включать восполнители жидкости и питательных веществ, такие как восполнители на основе декстрозы Рингера и тому подобное. В фармацевтических носителях могут также присутствовать консерванты и другие добавки, такие как, например, антимикробные агенты, антиоксиданты, хелатирующие агенты (например, ЭДТА), инертные газы и тому подобное. Настоящее изобретение не ограничено выбором носителя. Изготовление этих фармацевтически приемлемых композиций из описанных выше компонентов, имеющих подходящие рН, изотоничность, стабильность и другие общепринятые характеристики, входит в компетенцию специалиста в данной области. См., например, такие публикации, как Remington: The Science and Practice of Pharmacy, 20th ed, Lippincott Williams & Wilkins, publ., 2000, и The Handbook of Pharmaceutical Excipients, 4.sup.th edit., eds. R. C. Rowe et al., APhA Publications, 2003.
«Терапевтически эффективное количество» (или «эффективное количество») относится к количеству активного ингредиента, например, агента по изобретению, достаточному для получения полезных или желаемых результатов при введении субъекту или пациенту. Эффективное количество может быть введено с применением одного или более чем одного введения, нанесения или дозы. Терапевтически эффективное количество композиции по изобретению может быть легко определено специалистом в данной области. В контексте данного изобретения «терапевтически эффективное количество» представляет собой количество, приводящее к объективно измеряемому изменению одного или более чем одного параметра, связанного с состоянием, связанным с NGF, достаточному для получения полезных или желаемых результатов, включая клинические результаты, такие как облегчение или уменьшение болевых ощущений. Эффективная доза может быть введена за одно или более чем одно введение. Для целей данного изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для лечения, облегчения, уменьшения интенсивности и/или предотвращения боли, включая послеоперационную боль, боль при ревматоидном артрите и/или боль при остеоартрите. В некоторых воплощениях «эффективное количество» может уменьшать боль в покое (боль покоя) и/или механически индуцированную боль (включая боль после движения), и оно может быть введено до, во время или после болевого стимула. Как понимают в клиническом контексте, эффективное количество лекарственного средства, соединения или фармацевтической композиции может или не может быть достигнуто в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективное количество» можно рассматривать в контексте введения одного или более чем одного терапевтического агента, и один агент можно считать вводимым в эффективном количестве, если в сочетании с одним или более чем одним другим агентом достигнут или может быть достигнут желаемый результат.Разумеется, терапевтически эффективное количество будет варьировать в зависимости от конкретного субъекта и состояния, по поводу которого проводят лечение, массы тела и возраста субъекта, тяжести состояния, конкретного выбранного соединения, применяемой схемы введения, времени введения, способа введения и тому подобного, и все это может быть легко определено специалистом в данной области.
При использовании здесь термин «терапевтический» охватывает весь спектр вариантов лечения заболевания, состояния или расстройства. «Терапевтический» агент по изобретению может действовать профилактическим или превентивным образом, в том числе включающим методики, разработанные для воздействия на животных, которые могут быть идентифицированы как подверженные риску (фармакогенетика), или облегчающим или излечивающим образом или может действовать, уменьшая скорость или степень прогрессирования по меньшей мере одного симптома заболевания или расстройства, по поводу которых проводят лечение.
В другом аспекте изобретение включает ветеринарные композиции, в которых присутствуют антитела по настоящему изобретению для терапевтического или профилактического применения. Изобретение включает способ лечения субъекта-собаки, имеющего определенный антиген, например, связанный с заболеванием или состоянием. Способ включает введение терапевтически эффективного количества рекомбинантного антитела, специфичного в отношении определенного антигена, с использованием рекомбинантного антитела, описанного здесь.
Количество антитела, которое может быть использовано для получения терапевтического эффекта, может быть определено стандартными методиками, хорошо известными специалистам в данной области. Антитела будут обычно получать по стандартной методике в фармацевтически приемлемом буфере, и они могут быть введены любым желаемым способом введения. Путь введения антитела или антигенсвязывающей группировки по изобретению может быть пероральным, парентеральным, ингаляционным или местным. В предпочтительном воплощении путь введения является парентеральным. При использовании здесь термин «парентеральный» включает внутривенное, внутримышечное, подкожное, ректальное, вагинальное или внутрибрюшинное введение.
При использовании здесь «боль» относится к боли любой этиологии, включая острую и хроническую боль, и к любой боли с воспалительным компонентом. Примеры боли включают воспалительную боль, послеоперационную инцизионную боль, нейропатическую боль, боль при переломах, боль при остеопорозных переломах, постгерпетическую невралгию, раковую боль, боль в результате ожогов, боль, связанную с ожогом или раной, боль, связанную с травмой (включая черепно-мозговую травму), нейропатическую боль, боль, связанную с расстройствами костно-мышечной системы, такими как ревматоидный артрит, остеоартрит, анкилозирующий спондилит, серонегативные (неревматоидные) артропатии, внесуставной ревматизм и периартикулярные расстройства, и боль, связанную с раком (включая «прорывную боль» и боль, связанную с терминальным раком), периферической нейропатией и постгерпетической невралгией.
При использовании здесь «лечение» представляет собой способ получения полезных или желаемых клинических результатов. Для целей данного изобретения полезные или желаемые клинические результаты включают, без ограничения, одно или более из следующего: улучшение или облегчение любого аспекта боли, включая острую, хроническую, воспалительную, нейропатическую, послеоперационную боль, боль при ревматоидном артрите или боль при остеоартрите. Для целей данного изобретения полезные или желаемые клинические результаты включают, без ограничения, одно или более из следующего: уменьшение тяжести, облегчение одного или более чем одного симптома, связанного с болью, включая любой аспект боли (такой как уменьшение продолжительности боли, снижение болевой чувствительности или болевых ощущений).
Расстройство, связанное с NGF, относится к расстройству, включающему сердечно-сосудистые заболевания, атеросклероз, ожирение, диабет 2-го типа, метаболический синдром, боль и воспаление. В некоторых воплощениях настоящего изобретения расстройство, связанное с NGF, относится к боли, в частности хронической боли, воспалительной боли, послеоперационной инцизионной боли, нейропатической боли, боли при переломах, боли при остеопорозных переломах, постгерпетической невралгии, раковой боли, боли в результате ожогов, боли, связанной с ожогом или раной, боли, связанной с травмой (включая черепно-мозговую травму), нейропатической боли, боли, связанной с расстройствами костно-мышечной системы, такими как ревматоидный артрит, остеоартрит, анкилозирующий спондилит, серонегативные (неревматоидные) артропатии, внесуставной ревматизм и периартикулярные расстройства, и боли, связанной с раком (включая «прорывную боль» и боль, связанную с терминальным раком), периферической нейропатией и постгерпетической невралгией.
«Снижение заболеваемости» болью обозначает любое снижение тяжести (что может включать уменьшение потребности в и/или количества (например, объема применения) других лекарственных средств и/или видов терапии, обычно применяемых при данных состояниях, включая, например, опиаты), продолжительности и/или частоты (включая, например, задержку или увеличение времени до послеоперационной боли у индивида). Специалистам в данной области ясно, что индивиды могут различаться по их ответу на лечение, и, как таковой, например, «способ снижения заболеваемости болью при ревматоидном артрите или болью при остеоартрите у индивида» отражает введение антитела-антагониста против NGF, исходя из обоснованного ожидания, что такое введение может, вероятно, привести к такому снижению заболеваемости у данного конкретного индивида.
«Облегчение» боли или одного или более чем одного симптома боли (такой как боль при ревматоидном артрите или боль при остеоартрите) обозначает уменьшение или улучшение одного или более чем одного симптома боли по сравнению с отсутствием введения антитела-антагониста против NGF. «Облегчение» также включает укорочение или уменьшение продолжительности симптома.
«Ослабление» боли или одного или более чем одного симптома боли (такой как боль при ревматоидном артрите или боль при остеоартрите) обозначает уменьшение степени выраженности одного или более чем одного нежелательного клинического проявления послеоперационной боли у индивида или популяции индивидов, получающих лечение антителом-антагонистом против NGF по изобретению.
При использовании здесь «задержка» развития боли обозначает приостановку, сдерживание, замедление, торможение, стабилизацию и/или отсрочку прогрессирования боли, такой как послеоперационная боль, боль при ревматоидном артрите или боль при остеоартрите. Продолжительность этой задержки может варьировать в зависимости от анамнеза заболевания и/или индивидов, проходящих лечение. Специалисту в данной области очевидно, что достаточная или существенная задержка может, в сущности, охватывать предотвращение, в том смысле, что у индивида не развивается боль. Способ, «задерживающий» развитие симптома, представляет собой способ, снижающий вероятность развития симптома в заданный период времени и/или уменьшающий степень выраженности симптомов в заданный период времени, по сравнению с отсутствием применения способа. Такие сравнения обычно основаны на клинических исследованиях, проводимых с участием статистически значимого числа субъектов.
«Послеоперационная боль» (взаимозаменяемо называемая здесь «постинцизионной» или «посттравматической болью») относится к боли, возникающей вследствие или являющейся результатом внешней травмы, такой как порез, прокол, разрез, разрыв или ранение ткани индивида (включая боль, возникающую вследствие всех хирургических процедур, независимо от их инвазивности). При использовании здесь послеоперационная боль не включает боль, развивающуюся (возникающую или появляющуюся) без внешней физической травмы. В некоторых воплощениях послеоперационная боль представляет собой внутреннюю или внешнюю (включая периферическую) боль, а ранение, порез, травма, разрыв или разрез могут быть случайными (как в случае травматического ранения) или преднамеренными (как в случае хирургического разреза). При использовании здесь «боль» включает ноцицепцию и ощущение боли, и боль может быть оценена объективно и субъективно с применением балльных оценок боли и других методов, хорошо известных в данной области. При использовании здесь послеоперационная боль включает аллодинию (то есть усиленный ответ на безболезненный в норме стимул) и гиперальгезию (то есть усиленный ответ на болезненный или неприятный в норме стимул), которая, в свою очередь, может иметь термическую или механическую (тактильную) природу. В некоторых воплощениях боль характеризуется чувствительностью к температуре, чувствительностью к механическим стимулам и/или боль покоя. В некоторых воплощениях послеоперационная боль включает механически индуцированную боль или боль покоя. В других воплощениях послеоперационная боль включает боль покоя. Боль может представлять собой первичную или вторичную боль, как хорошо известно в данной области.
Перед описанием настоящих методов следует понимать, что данное изобретение не ограничено конкретными описанными методами и условиями экспериментов, поскольку такие методы и условия могут варьировать. Также следует понимать, что термины, использованные здесь, предназначены лишь для описания конкретных воплощений и не являются ограничивающими, поскольку объем настоящего изобретения ограничен исключительно приложенной формулой изобретения.
Если не указано иное, все технические и научные термины, использованные здесь, имеют то же значение, в котором их обычно понимает специалист в области, к которой относится данное изобретение. Несмотря на то, что при практическом применении или испытании настоящего изобретения могут быть применены любые методы и материалы, сходные с описанными здесь или эквивалентные им, далее описаны предпочтительные методы и материалы. Все публикации, упомянутые здесь, полностью включены сюда посредством ссылки.
Изобретение, раскрытое здесь, относится к антигенсвязывающим белкам (использовано взаимозаменяемо с терминами «антитела», «антитела-антагонисты», «фрагменты антител» и тому подобными, как описано здесь), специфично связывающимся с фактором роста нервов (NGF), и, в частности, к антителам, которые могут представлять собой собачьи, кошачьи, лошадиные, мышиные, бычьи, человеческие антитела или антитела любого другого вида, канинизированные, фелинизированные, бовинизированные, эквинизированные, гуманизированные антитела или антитела, прошедшие любую другую видовую адаптацию, полученные с применением рекомбинантных методов, гибридомных технологий или технологии фагового дисплея, или полностью «канинизированные» (прошедшие видовую адаптацию) моноклональные антитела, которые специфично связываются с NGF и, таким образом, предотвращают связывание NGF с собачьим рецептором TrkA и, в меньшей степени, с собачьим рецептором р75, выступая, таким образом, в роли антагонистов, в том смысле, что они предотвращают активацию сигнального пути под действием NGF.
NGF был первым идентифицированным нейротрофином, и его роль в развитии и выживании как периферических, так и центральных нейронов хорошо охарактеризована. Было показано, что NGF является критически важным фактором выживания и поддерживающим фактором при развитии периферических симпатических и эмбриональных чувствительных нейронов и холинергических нейронов базальных отделов переднего мозга (Smeyne et al. (1994) Nature 368:246-249; Crowley et al. (1994) Cell 76:1001-1011). NGF приводит к повышающей регуляции экспрессии нейропептидов в чувствительных нейронах (Lindsay et al. (1989) Nature 337:362-364), и его активность опосредована двумя разными мембраносвязанными рецепторами, рецептором TrkA и общим рецептором нейротрофинов р75, который принято считать низко аффинным.
Было продемонстрировано повышение NGF при расстройствах, связанных с NGF, при которых в травмированных или болезненных тканях присутствует повышенное количество NGF. Расстройство, связанное с NGF, может быть определено как усиление боли вследствие повышения NGF в травмированной, болезненной или поврежденной ткани. При использовании здесь боль, определенная, как описано здесь, относится к расстройству, включающему хроническую боль, воспалительную боль, послеоперационную инцизионную боль, нейропатическую боль, боль при переломах, боль при остеопорозных переломах, постгерпетическую невралгию, раковую боль, боль в результате ожогов, боль, связанную с ожогом или раной, боль, связанную с травмой (включая черепно-мозговую травму), нейропатическую боль, боль, связанную с расстройствами костно-мышечной системы, такими как хроническая боль, ревматоидный артрит, остеоартрит, анкилозирующий спондилит, серонегативные (неревматоидные) артропатии, внесуставной ревматизм и периартикулярные расстройства, и боль, связанную с раком (включая «прорывную боль» и боль, связанную с терминальным раком), периферической нейропатией и постгерпетической невралгией.
В одном воплощении настоящего изобретения NGF-расстройство определено как остеоартрит у субъекта (собак, кошек, лошадей, людей и так далее). Остеоартрит (OA) представляет собой медленно прогрессирующее дегенеративное заболевание суставов, характеризующееся у собак утратой суставного хряща и обнажением субхондральной кости. В конечном счете, это приводит к самоподдерживающемуся хроническому расстройству, характеризующемуся болью в суставах. В ответ на хроническое воспаление происходят образование новой кости и местное повреждение тканей, направленные на ограничение как подвижности, так и боли. Макроскопически наблюдается утрата суставного хряща, сужение суставной щели, склероз субхондральной кости и образование суставных остеофитов (Veterinary Focus: Vol 17 No 3; 2007).
У различных видов, таких как собаки, кошки, лошади и тому подобные, развитие первичного OA зависит от породы. У собак, например, средний возраст развития заболевания составляет 3,5 года у ротвейлеров и 9,5 года у пуделей, при этом у других пород, а также у смешанных пород, возраст развития заболевания варьирует в широких пределах. Ортопедические болезни роста и связанный с ними остеоартрит являются наиболее распространенными заболеваниями суставов у собак, ими обусловлено примерно 70% обращений за медицинской помощью по поводу заболеваний суставов и связанных с ними проблем с добавочным скелетом. В двадцати двух процентах случаев это были собаки в возрасте одного года или младше. Заболеваемость OA возрастает при травмах, а также при ожирении, старении и генетических отклонениях. В частности, фактором, влияющим на заболеваемость OA, может быть возраст, при этом более 50% случаев артрита наблюдают у собак в возрасте 8-13 лет.Заболевания костно-мышечной системы очень распространены у пациентов пожилого возраста, и ортопедические расстройства есть примерно у 20% пожилых собак. Для лабрадоров-ретриверов в возрасте старше 8 лет типичен OA нескольких суставов (локтевых, плечевых, тазобедренных, коленных). Кроме того, на развитие OA также влияет размер собаки. 45% собак с артритом являются собаками крупных пород. Более 50% из них являются собаками очень крупных пород, в то время как только 28% являются собаками средних пород, а 27% - собаками мелких пород. Существует очень большая потребность в фармацевтическом вмешательстве для облегчения боли при OA у собак.
Как указано здесь, повышенные уровни NGF являются признаком, указывающим на расстройство, связанное с NGF, в частности на OA. У трансгенных мышей с артритом были отмечены повышенные уровни NGF наряду с ростом числа тучных клеток (Aloe, et al., Int. J. Tissue Reactions-Exp. Clin. Aspects 15:139-143 (1993)). В РСТ-публикации № WO 02/096458 раскрыто применение антител против NGF, обладающих определенными свойствами, в лечении различных расстройств, связанных с NGF, таких как воспалительное состояние (например, ревматоидный артрит). Сообщалось, что введение очищенного антитела против NGF трансгенным мышам с артритом, у которых был человеческий ген фактора некроза опухоли, приводило к уменьшению числа тучных клеток, а также к снижению уровней гистамина и вещества Р в синовиальной оболочке мышей с артритом (Aloe et al., Rheumatol. Int. 14: 249-252 (1995)). Было показано, что экзогенное введение антитела к NGF снижало повышенный уровень TNFα, наблюдаемый у мышей с артритом (Marmi et al., Rheumatol. Int. 18: 97-102 (1998)). Есть сообщения об антителах-антагонистах против NGF, являющихся антителами грызунов. См., например, Hongo et al., Hybridoma (2000) 19(3): 215-227; Ruberti etal. (1993) Cell. Molec. Neurobiol. 13(5): 559-568. Тем не менее, при терапевтическом применении антител грызунов у млекопитающих, не являющихся мышами, у значительного числа индивидов, получающих лечение, развивается ответ, направленный против мышиных антител. Таким образом, существует серьезная потребность в антигенсвязывающих белках-антагонистах против NGF, включая антитела-антагонисты против NGF по настоящему изобретению, для применения у собак, в частности для применения в лечении OA.
Несмотря на то, что свойства антител делают их очень привлекательными терапевтическими агентами, существует ряд ограничений. Подавляющее большинство моноклональных антител (mAb) имеют происхождение от грызунов, как отмечено ранее. При введении таких антител другому виду у пациентов возможно развитие собственного гуморального иммунного ответа на такие ксеногенные антитела. В конечном счете, такой ответ может приводить к нейтрализации и элиминации антитела. Как описано выше, мышей широко используют при получении моноклональных антител. Одна проблема при использовании антител, полученных у определенного вида, в большинстве случаев исходно у мыши, состоит в том, что субъект, не являющийся мышью, получающий лечение такими антителами, реагирует на мышиные антитела так, как если бы они представляли собой чужеродное вещество, вырабатывая, таким образом, новые антитела к мышиным антителам. Иммунная система вида, не являющегося мышью, например, собаки, «воспринимает» мышиные антитела как чужеродные, и затем у субъекта развивается иммунный ответ на эти молекулы. Специалистам в данной области будет ясно, что необходима возможность лечить субъекта антителом, специфичным в отношении антигена, но это антитело должно быть видоспецифичным. Часть реакции, вызванной введением антител другого вида, например, введением мышиного моноклонального антитела собаке, может варьировать от легкой формы, такой как сыпь, до более выраженного и жизнеугрожающего ответа, такого как почечная недостаточность. Этот иммунный ответ может также снижать эффективность лечения или приводить к реакциям в будущем при проведении у субъекта последующего лечения, включающего мышиные антитела. Соответственно, авторы изобретения описывают преодоление этого недостатка посредством «канинизации» антитела. В частности, этот способ сосредоточен на каркасных областях вариабельного домена иммуноглобулина, но может также включать участки, определяющие комплементарность (CDR), вариабельного домена. В данной заявке описаны стадии, позволяющие осуществлять этот способ, и его практическое применение.
Способ модификации моноклонального антитела (антигенсвязывающего белка, антитела-антагониста и так далее, как описано здесь, и терминов, используемых взаимозаменяемо) от животного с целью снижения его иммуногенности для терапевтического введения другому виду являлся предметом интенсивного изучения и описан в ряде публикаций (например, Antibody Engineering: A practical Guide. Carl А.K. Borrebaeck ed. W.H. Freeman and Company, 1992). Тем не менее, до недавнего времени этот способ не применяли для разработки терапевтических или диагностических средств для видов, не являющихся людьми, в частности собак. На самом деле, опубликованных данных о собачьих вариабельных доменах вообще очень немного. В Wasserman and Capra, Biochem. 6, 3160 (1977) определена аминокислотная последовательность вариабельных областей тяжелой цепи собачьего IgM и собачьего IgA. В Wasserman and Capra, Immunochem. 15, 303 (1978) определена аминокислотная последовательность легкой цепи к собачьего IgA. В McCumber and Capra, Mol. Immunol. 16, 565 (1979) раскрыта полная аминокислотная последовательность собачьей цепи мю. В Tang et al., Vet. Immunology Immunopathology 80, 259 (2001) раскрыты одна кДНК γ-цепи собачьего IgG-A и четыре белковые последовательности γ-цепи собачьего IgG-A. В ней описана ПЦР-амплификация библиотеки кДНК собачьей селезенки с вырожденными олигонуклеотидными праймерами, разработанными, исходя из консервативных областей человеческих, мышиных, свиных и бычьих IgG. Недостаток доступной информации о собачьих антителах не позволял проводить их разработку в качестве терапевтических средств для лечения заболеваний у собак.
Указанные ограничения привели к разработке технологий конструирования, известных как «видовая адаптация» и хорошо известных специалистам в данной области на примере «гуманизации» терапевтических антител. Канинизированные антитела, в качестве примера молекул, прошедших видовую адаптацию, могут быть получены как химерные антитела или их фрагменты, содержащие минимум последовательностей, имеющих происхождение от иммуноглобулина, не являющегося собачьим. Большей частью, канинизированные антитела представляют собой собачьи антитела (то есть «антитело-реципиент» или «антитело целевого вида»), в которых остатки участка, определяющего комплементарность (CDR), реципиента заменены остатками CDR вида, не являющегося собакой (то есть «антитела-донора» или «антитела исходного вида»), такого как мышь, имеющего желаемые свойства, такие как специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) собачьего иммуноглобулина заменены соответствующими остатками вида, не являющегося собакой. Эту методику канинизации называют «переносом CDR». Для восстановления и/или улучшения аффинности могут быть необходимы обратные мутации выбранных целевых каркасных остатков на соответствующие донорные остатки. Для канинизации и созревания аффинности могут также быть применены структурные методы, как описано в US 7261890.
В методиках, описанных выше, используют почти полноразмерные каркасные области одной или более чем одной из вариабельных областей тяжелой или легкой цепи антитела целевого вида, которые конструируют для присоединения CDR вида-донора. Эту методику также применяют при фелинизации антитела для снижения его антигенности при введении кошкам, таким же образом, как при канинизации. В некоторых случаях применяют обратную мутацию выбранных остатков вариабельной области для усиления представления CDR. Разработка антител, минимизирующих иммуногенную реакцию у субъекта на ненативные последовательности антитела с сохранением в то же время антигенсвязывающих областей антитела в степени, достаточной для поддержания эффективности, затруднительна.
Другая проблема при разработке терапевтических антител, оказывающих направленное действие на белки, состоит в том, что у разных видов эпитопы гомологичных белков, а также вероятность перекрестной реактивности с другими белками часто различаются. Вследствие этого антитела необходимо получать, испытывать и разрабатывать для конкретной мишени именно того вида, у которого предполагается проводить лечение.
Антитела оказывают направленное действие на антиген посредством их связывания с определенным эпитопом антигена в результате его взаимодействия с вариабельной областью молекулы антитела. Кроме того, антитела обладают способностью опосредовать, ингибировать (как в случае антагонистических антигенсвязывающих белков против NGF по настоящему изобретению) и/или инициировать множество видов биологической активности. Терапевтические антитела могут выполнять широкий спектр функций, например, антитела могут модулировать взаимодействие рецептора с лигандом как агонисты или антагонисты. Связывание с антителом может инициировать внутриклеточную передачу сигналов, стимулируя клеточный рост, образование цитокинов или апоптоз. Антитела могут доставлять агенты, связанные с Fc-областью, в определенные места. Антитела могут также приводить к антитело-опосредованной цитотоксичности (ADCC), комплемент-опосредованной цитотоксичности (CDC) или фагоцитозу. Также существуют измененные антитела, у которых устранены функции связывания с ADCC, CDC, C1q и фагоцитоза. В одном воплощении настоящего изобретения антитело по настоящему изобретению содержит изменения в Fc-области антитела, изменяющие эффекторную функцию указанного антитела.
Канинизация и фелинизация
При использовании здесь «канинизированное антитело» обозначает антитело, имеющее аминокислотную последовательность, соответствующую аминокислотной последовательности антитела, образованного в организме собаки и/или полученного с применением любой из методик, известных в данной области или раскрытых здесь. Такой же способ применяют для фелинизации, и приведенное здесь описание применимо к фелинизации. Для простоты канинизация будет приведена в качестве примера, тем не менее, эти примеры не ограничены собакой. Те же концепции и схемы применимы к видовой адаптации других антигенсвязывающих белков (например, кошачьих, лошадиных, человеческих и тому подобных). Данное определение канинизированного антитела включает антитела, содержащие по меньшей мере один полипептид собачьей тяжелой цепи или по меньшей мере один полипептид собачьей легкой цепи. «Видовая адаптация» антител сама по себе и, в частности, гуманизация антител, является областью исследований, хорошо известной специалисту в данной области. До недавнего времени не было известно, позволяет ли видовая адаптация антител, помимо гуманизации, получить терапевтическое антитело, которое может быть эффективным у какого-либо другого вида. В настоящем изобретении приведены примеры канинизации и фелинизации антигенсвязывающего белка против NGF для терапевтического применения у собак и кошек, соответственно.
Химерные антитела содержат последовательности по меньшей мере двух разных видов. В качестве одного примера, могут быть применены рекомбинантные методики клонирования для включения вариабельных областей, содержащих антигенсвязывающие сайты, от антитела, не являющегося реципиентом (то есть антитела, полученного у вида-донора, иммунизированного антигеном) и константных областей, имеющих происхождение от иммуноглобулина-реципиента.
Антитела, прошедшие видовую адаптацию (канинизированные, фелинизированные и тому подобные), представляют собой тип химерного антитела, где остатки вариабельной области, обеспечивающие связывание с антигеном (то есть остатки участка, определяющего комплементарность, сокращенного участка, определяющего комплементарность, или любые другие остатки, участвующие в связывании с антигеном), имеют происхождение от вида, не являющегося собакой (или не являющегося кошкой), в то время как остальные остатки вариабельной области (то есть, остатки каркасных областей) и константные области имеют происхождение, по меньшей мере частично, от последовательностей собачьих (или кошачьих) антител. Часть остатков каркасной областей и остатков константных областей антитела, прошедшего видовую адаптацию, может иметь происхождение от источника, не являющегося собакой (или не являющегося кошкой). Вариабельные области антитела, прошедшего видовую адаптацию, также описывают как прошедшие видовую адаптацию (то есть как вариабельную область легкой или тяжелой цепи антитела, прошедшую видовую адаптацию). Видом, не проходящим видовую адаптацию, обычно является вид, используемый для иммунизации антигеном, такой как мышь, крыса, кролик, примат, не являющийся человеком, или млекопитающее другого вида, не являющееся собакой или не являющееся кошкой.
Участки, определяющие комплементарность (CDR), представляют собой остатки вариабельных областей антитела, участвующие в связывании с антигеном. Для определения CDR широко применяют несколько систем нумерации. Определение Kabat основано на вариабельности последовательностей, а определение Clothia основано на расположении областей структурных петель. Определение АЬМ представляет собой промежуточный вариант между методами Kabat и Chothia. Антитело, прошедшее видовую адаптацию, по изобретению может быть сконструировано так, чтобы содержать один или более чем один CDR. Кроме того, CDR могут быть использованы по отдельности или в комбинации в синтетических молекулах, таких как SMIP и малые миметики антител.
Остатки каркасных областей представляют собой остатки вариабельных областей антитела, не являющиеся гипервариабельными остатками или остатками CDR. Остатки каркасных областей могут иметь происхождение от встречающегося в природе собачьего (например, но это применимо к другим видам, таким как кошки, лошади, люди и так далее; для простоты в качестве типичного вида будет использована собака, но примеры не ограничены собакой как таковой) антитела, например, собачьей каркасной области, по существу сходной с каркасной областью антитела по изобретению. Могут также быть использованы искусственные каркасные последовательности, представляющие собой консенсус отдельных последовательностей. При выборе каркасной области для канинизации последовательности, широко представленные у собак, могут быть предпочтительнее, чем менее распространенные последовательности. Могут быть проведены дополнительные мутации акцепторных последовательностей собачьих каркасных областей для восстановления мышиных остатков, которые предположительно вовлечены в контакты с антигеном, и/или остатков, обеспечивающих структурную целостность антигенсвязывающего сайта, или для улучшения экспрессии антитела.
Перенос CDR проводят, заменяя один или более чем один CDR антитела-акцептора (например, собачьего антитела или другого антитела, содержащего желаемые остатки каркасных областей) на один или более чем один CDR антитела-донора (например, антитела, не являющегося собачьим). Антитела-акцепторы могут быть выбраны, исходя из сходства остатков каркасных областей антитела, рассматриваемого как возможное антитело-акцептор, и антител а-донор а. Например, собачьи каркасные области идентифицируют как имеющие существенную гомологию последовательностей с каждой каркасной областью соответствующего антитела, не являющегося собачьим, и CDR антитела, не являющегося собачьим, переносят на сочетание различных собачьих каркасных областей.
Анализ трехмерных структур комплексов антитела и антигена, вместе с анализом имеющихся данных по аминокислотным последовательностям, может быть применен для моделирования вариабельности последовательностей, исходя из структурных различий аминокислотных остатков, присутствующих в каждом положении CDR. CDR по настоящему изобретению могут также быть использованы в малых миметиках антител, содержащих два CDR-участка и каркасную область (Qui et al. Nature Biotechnology Vol 25;921-929; August 2007).
Акцепторные каркасные области для переноса CDR или сокращенных CDR могут быть дополнительно модифицированы для введения желаемых остатков. Например, акцепторные каркасные области могут содержать вариабельную область тяжелой цепи собачьей консенсусной последовательности, возможно с донорными остатками, не являющимися собачьими, в одном или более чем одном положении. После переноса в донорных и/или акцепторных последовательностях могут быть проведены дополнительные изменения для оптимизации связывания или функциональности антитела. См., например, международную публикацию № WO 91/09967.
Кроме того, согласно настоящему изобретению предложены клетки и клеточные линии, экспрессирующие антитела по изобретению. Типичные клетки-хозяева включают клетки бактерий, дрожжей, млекопитающих и человека, такие как клетки СНО, клетки HEK-293, клетки HeLa, клетки CV-1 и клетки COS. Методы получения стабильной клеточной линии после трансформации клетки-хозяина гетерологичной конструкцией известны в данной области. Типичные клетки-хозяева, не являющиеся клетками млекопитающих, включают клетки насекомых (Potter et al. (1993) Int. Rev. Immunol. 10(2-3): 103-112). Антитела могут также быть получены в трансгенных животных (Houdebine (2002) Curr. Opin. Biotechnol. 13(6):625-629) и трансгенных растениях (Schillberg et al. (2003) Cell Mol. Life Sci. 60(3):433-45).
Как обсуждено выше, объем изобретения также включает моноклональные, химерные, видоспецифичные антитела и антитела, прошедшие видовую адаптацию, модифицированные посредством, например, удаления, добавления или замены других частей антитела, например, константной области. Например, антитело может быть модифицировано следующим образом: (1) удалением константной области; (2) заменой константной области на другую константную область, например, константную область, предназначенную для увеличения периода полувыведения, повышения стабильности или аффинности антитела, или константную область другого вида или антитела другого класса; или (3) модификацией одной или более чем одной аминокислоты константной области для изменения, например, среди прочего, числа сайтов гликозилирования, эффекторной клеточной функции, связывания с Fc-рецепторами (FcR), фиксации комплемента. В одном воплощении настоящего изобретения антитело по изобретению содержит измененную Fc-область, изменяющую эффекторную функцию антитела. В некоторых воплощениях настоящего изобретения Fc-область антигенсвязывающего белка по изобретению заменена, модифицирована или удалена.
Методы изменения константной области антитела известны в данной области. Антитела с измененной функцией, например, измененной аффинностью в отношении эффекторного лиганда, такого как FcR на клетке, или С1-компонента комплемента, могут быть получены заменой по меньшей мере одного аминокислотного остатка константной части антитела на другой остаток (см., например, ЕР 388151 А1, патент США №5624821 и патент США №5648260, содержание всех из которых включено сюда посредством ссылки).
Например, возможно изменение аффинности Fc-области антитела в отношении FcR (например, FcγRl) или в отношении связывания с C1q посредством замены определенного остатка (остатков) на остаток (остатки), имеющий подходящую функциональность на его боковой цепи, или посредством введения заряженной функциональной группы, такой как глутамат или аспартат, или, возможно, ароматического неполярного остатка, такого как фенилаланин, тирозина, триптофана или аланина (см., например, патент США №5624821). Антитело или его связывающий фрагмент могут быть конъюгированы с цитотоксином, терапевтическим агентом или ионом радиоактивного металла. В одном воплощении конъюгированный белок представляет собой антитело или его фрагмент.Цитотоксин или цитотоксический агент включает любой агент, наносящий ущерб клеткам. Неограничивающие примеры включают калихеамицин, таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенипозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и их аналоги или гомологи. Терапевтические агенты включают, без ограничения, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин и 5-фторурацил, дакарбазин), алкилирующие агенты (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозоцин, митомицин С и цис-дихлордиамин платины (II) (DDP), цисплатин), антрациклины (например, даунорубицин и доксорубицин), антибиотики (например, дактиномицин, блеомицин, митрамицин и антрамицин) и антимитотические агенты (например, винкристин и винбластин). Методики конъюгации таких группировок с белками хорошо известны в данной области.
Композиции, производные композиции и способы изготовления композиций
Данное изобретение охватывает композиции, включая фармацевтические композиции, содержащие антигенсвязывающие белки («антитела», «фрагменты антител», «антитела-антагонисты» и тому подобное, как использовано взаимозаменяемо здесь), полипептиды и полинуклеотиды, содержащие последовательности, кодирующие антигенсвязывающие белки или полипептиды по изобретению.
При использовании здесь композиции содержат одно или более чем одно антитело, один или более чем один антигенсвязывающий белок или полипептид (который может быть или не быть антителом), связывающиеся с NGF, и/или один или более чем один полинуклеотид, содержащий последовательности, кодирующие одно или более чем одно антитело или один или более чем один полипептид, связывающиеся с NGF. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически/ветеринарно приемлемые эксципиенты, включая буферы, хорошо известные в данной области. Изобретение также охватывает воплощения с выделенными антителами, полипептидами и полинуклеотидами. Изобретение также охватывает отдельные воплощения с по существу чистыми антителами, полипептидами и полинуклеотидами.
В одном или более чем одном воплощении согласно настоящему изобретению предложены новые антигенсвязывающие белки, специфично связывающиеся с NGF. В одном или более чем одном воплощении антигенсвязывающий белок определен как антитело или фрагмент антитела. В одном или более чем одном воплощении антигенсвязывающий белок является полностью собачьим, полностью кошачьим, полностью бычьим, полностью лошадиным, полностью человеческим, канинизированным, фелинизированным, эквинизированным или гуманизированным. В одном или более чем одном воплощении антигенсвязывающий белок по настоящему изобретению связывается с собачьим, кошачьим, лошадиным или человеческим NGF. В одном воплощении антигенсвязывающий белок представляет собой моноклональное антитело. В одном воплощении моноклональное антитело по изобретению связывается с NGF и предотвращает его связывание с рецепторами TrkA и, в меньшей степени, р75 и их активацию, предотвращая, таким образом, сигнальный каскад, как описано здесь. Антигенсвязывающий белок по настоящему изобретению обозначен здесь как ZTS-841.
В одном или более чем одном воплощении согласно настоящему изобретению предложен выделенный и рекомбинантный антигенсвязывающий белок, «ZTS-841», где вариабельная область тяжелой цепи содержит аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 4 (CDR1 VH «ZTS-841»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 5 (CDR2 VH «ZTS-841»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 6 (CDR3 VH «ZTS-841»), и где вариабельная область легкой цепи содержит аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: l (CDR1 VL «ZTS-841»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 2 (CDR2 VL «ZTS-841»), и аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 3 (CDR3 VL «ZTS-841»), и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
В одном или более чем одном воплощении согласно настоящему изобретению предложен выделенный и рекомбинантный антигенсвязывающий белок, «ZTS-842», где вариабельная область тяжелой цепи содержит аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 24 (CDR1 VH «ZTS-842»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 25 (CDR2 VH «ZTS-842»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 26 (CDR3 VH «ZTS-842»), и где вариабельная область легкой цепи содержит аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 21 (CDRT VL «ZTS-842»), аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 22 (CDR2 VL «ZTS-842»), и аминокислотную последовательность, по меньшей мере приблизительно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности, содержащей SEQ ID NO: 23 (CDR3 VL «ZTS-842»), и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену по меньшей мере в одном из CDR1, CDR2 или CDR3 любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
Согласно настоящему изобретению предложены рекомбинантные антигенсвязывающие белки, в некоторых воплощениях описанные здесь, моноклональные антитела, фрагменты антител и их применение при клиническом использовании и в научных методиках, включая диагностические методики. Применение молекулярно-биологических методов и рекомбинантных технологий привело к возможности рекомбинантного получения антител и антитело подобных молекул и, таким образом, создания генных последовательностей, кодирующих определенные аминокислотные последовательности, присутствующие в полипептидной структуре антител. Такие антитела могут быть получены клонированием генных последовательностей, кодирующих полипептидные цепи указанных антител, или прямым синтезом указанных полипептидных цепей и сборкой синтезированных цепей с образованием активных тетрамерных (H2L2) структур, обладающих аффинностью в отношении определенных эпитопов и антигенных детерминант.Это позволило быстро получать антитела, имеющие последовательности, характерные для нейтрализующих антител от различных видов и источников.
Независимо от источника антител, особенностей их рекомбинантного конструирования или особенностей их синтеза, in vitro или in vivo, использования трансгенных животных, больших клеточных культур лабораторного или коммерческого размера, использования трансгенных растений или применения прямого химического синтеза без использования живых организмов на любой стадии процесса, все антитела имеют сходную общую 3-мерную структуру. Эту структуру часто обозначают как H2L2, и это указывает на тот факт, что антитела обычно содержат две легкие (L) аминокислотные цепи и 2 тяжелые (Н) аминокислотные цепи. Обе цепи имеют области, способные взаимодействовать со структурно комплементарной антигенной мишенью. Области, взаимодействующие с мишенью, называют «вариабельными» или «V»-областями, и они характеризуются различиями в аминокислотной последовательности антител с разной антигенной специфичностью. Вариабельные области Н- или L-цепей содержат аминокислотные последовательности, способные специфично связываться с антигенными мишенями.
При использовании здесь термин «антигенсвязывающая область» относится к той части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и придающие антителу его специфичность и аффинность в отношении антигена. Связывающая область антитела содержит «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. В пределах вариабельных областей Н- или L-цепей, обеспечивающих антигенсвязывающие области, есть последовательности меньших размеров, называемые «гипервариабельными» ввиду их крайней вариабельности у антител разной специфичности. Такие гипервариабельные участки также называют «участками, определяющими комплементарность», или «CDR»-участками. Эти CDR-участки обеспечивают основную специфичность антитела в отношении структуры определенной антигенной детерминанты.
CDR представляют собой несмежные участки из аминокислот в пределах вариабельных областей, но обнаружено, что, независимо от видовой принадлежности, эти критически важные аминокислотные последовательности расположены в вариабельных областях тяжелой и легкой цепи сходным образом. Вариабельные области тяжелой и легкой цепи всех антител имеют три CDR-участка, несмежных друг с другом. У всех видов млекопитающих пептиды антител содержат константные (то есть высоко консервативные) и вариабельные области, и, в пределах вариабельных областей, есть CDR и так называемые «каркасные области», образованные аминокислотными последовательностями, расположенными в пределах вариабельной области тяжелой или легкой цепи, но за пределами CDR.
Кроме того, согласно настоящему изобретению предложен вектор, содержащий по меньшей мере одну из нуклеиновых кислот, описанных выше. Ввиду вырожденности генетического кода для кодирования конкретной аминокислоты может быть использован более чем один кодон. С использованием генетического кода могут быть определены одна или несколько разных нуклеотидных последовательностей, каждая из которых будет способна кодировать указанную аминокислоту. Вероятность того, что конкретный олигонуклеотид действительно будет составлять фактическую кодирующую последовательность, может быть оценена с учетом закономерностей аномального спаривания оснований и частоты фактического использования конкретного кодона (для кодирования конкретной аминокислоты) в эукариотических или прокариотических клетках, экспрессирующих антитело против NGF или его часть. Такие «правила использования кодонов» раскрыты в Lathe, et al., 183 J. Molec. Biol. 1-12 (1985). С применением «правил использования кодонов» по Lathe может быть определена одна нуклеотидная последовательность или группа нуклеотидных последовательностей, содержащих теоретическую «наиболее вероятную» нуклеотидную последовательность, способную кодировать последовательности против NGF. Кроме того, подразумевается, что области, кодирующие антитело, для использования в настоящем изобретении могут также быть обеспечены изменением существующих генов антител с применением стандартных молекулярно-биологических методик, приводящих к получению вариантов (агонистов) антител и пептидов, описанных здесь. Такие варианты включают, без ограничения, делеции, добавления и замены в аминокислотной последовательности антител или пептидов против NGF.
Например, один класс замен представляет собой консервативные аминокислотные замены. Такие замены представляют собой замены заданной аминокислоты в пептиде антитела против NGF другой аминокислотой со сходными характеристиками. Консервативными обычно считают замены одной аминокислоты (остатка) на другую аминокислоту (остаток) в пределах следующих групп: алифатические аминокислоты Ala, Val, Leu и Ile; гидроксильные остатки Ser и Thr; кислые остатки Asp и Glu; амидные остатки Asn и Gln; основные остатки Lys и Arg; ароматические остатки Phe, Tyr; и тому подобное. Информация относительно того, какие аминокислотные изменения, вероятно, не приведут к изменениям фенотипа, представлена в Bowie et al., 247 Science 1306-10 (1990).
Вариант антигенсвязывающих белков или фрагментов антител против NGF может быть полностью функциональным или может иметь сниженную функцию по одному или более чем одному виду активности. Полностью функциональные варианты обычно содержат только консервативные изменения или изменения остатков или областей, не являющихся критически важными. Функциональные варианты могут также содержать замены сходных аминокислот, не приводящие к функциональным изменениям или приводящие к незначительным функциональным изменениям. Альтернативно, такие замены могут в некоторой степени влиять на функции положительным или отрицательным образом. Нефункциональные варианты обычно содержат одну или более чем одну неконсервативную аминокислотную замену, делецию, вставку, инверсию или усечение или замену, вставку, инверсию или делецию критически важного остатка или критически важной области.
Аминокислоты, важные для выполнения функции, могут быть определены методами, известными в данной области, такими как сайт-направленный мутагенез или мутагенез с применением аланинового сканирования. Cunningham et al., 244 Science 1081-85 (1989). Последняя методика включает введение одиночных аланиновых мутаций по каждому остатку молекулы. Полученные мутантные молекулы затем анализируют на предмет их биологической активности, такой как связывание эпитопов или ADCC-активность in vitro. Сайты, критически важные для связывания лиганда с рецептором, могут также быть определены структурным анализом, таким как кристаллография, ядерный магнитный резонанс или фотоаффинное мечение. Smith et al., 224 J. Mol. Biol. 899-904 (1992); de Vos et al., 255 Science 306-12 (1992).
Более того, полипептиды часто содержат аминокислоты, отличные от двадцати «встречающихся в природе» аминокислот.Кроме того, многие аминокислоты, включая концевые аминокислоты, могут быть модифицированы естественными процессами, такими как процессинг и другие посттрансляционные модификации, или методиками химической модификации, хорошо известными в данной области. Известные модификации включают, без ограничения, ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гемовой группировки, ковалентное присоединение нуклеотида или нуклеотидного производного, ковалентное присоединение липида или липидного производного, ковалентное присоединение фосфатидилинозита, поперечное связывание, циклизацию, образование дисульфидных связей, деметилирование, образование поперечных ковалентных связей, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, гликозилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитический процессинг, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, присоединение аминокислот к белкам, опосредованное транспортными РНК, такое как аргинилирование, и убиквтинирование. Такие модификации хорошо известны специалистам в данной области и подробно описаны в научной литературе. Некоторые особенно распространенные модификации, например, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и АДФ-рибозилирование, описаны в большинстве основополагающих публикаций, таких как Proteins-Structure and Molecular Properties (2nd ed., Т.E. Creighton, W.H. Freeman & Co., NY, 1993). По этой теме есть много подробных обзоров, такие как Wold, Posttranslational Covalent Modification of proteins, 1-12 (Johnson, ed., Academic Press, NY, 1983), Seifter et al. 182 Meth. Enzymol. 626-46 (1990) и Rattan et al. 663 Ann. NY Acad. Sci. 48-62 (1992).
Соответственно, антитела и пептиды по настоящему изобретению также охватывают производные или аналоги, в которых заменяющий аминокислотный остаток отличен от остатка, кодируемого генетическим кодом. Сходным образом, добавления и замены в аминокислотной последовательности, а также варианты и модификации, описанные выше, могут быть в равной степени применимы к аминокислотной последовательности антигена и/или эпитопа NGF или их пептидам и, таким образом, включены в настоящее изобретение. Как указано выше, гены, кодирующие моноклональное антитело по настоящему изобретению, особенно эффективны в распознавании NGF.
Производные антител
Объем данного изобретения включает производные антител. «Производное» антитела содержит дополнительные химические группировки, не являющиеся обычно частью белка. Объем данного изобретения включает ковалентные модификации белков. Такие модификации могут быть введены в молекулу посредством взаимодействия целевых аминокислотных остатков антитела с органическим дериватизирующим агентом, способным взаимодействовать с выбранными боковыми цепями или концевыми остатками. Например, дериватизация бифункциональными агентами, хорошо известная в данной области, полезна для поперечного связывания антитела или фрагмента с матрицей водонерастворимой подложки или с другими макромолекулярными носителями.
Производные также включают: радиоактивно меченные моноклональные антитела, меченные, например, радиоактивным йодом (251,1311), углеродом (4С), серой (35S), индием, тритием (Н3) или тому подобным; конъюгаты моноклональных антител с биотином или авидином, с ферментами, такими как пероксидаза хрена, щелочная фосфатаза, бета-D-галактозидаза, глюкозооксидаза, глюкоамилаза, ангидраза карбоновых кислот, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо-6-фосфатдегидрогеназа; а также конъюгаты моноклональных антител с биолюминесцентными агентами (такими как люцифераза), хемилюминесцентными агентами (такими как сложные эфиры акридина) или флуоресцентными агентами (такими как фикобилипротеины).
Другое бифункциональное производное антитела по настоящему изобретению представляет собой биспецифичное антитело, полученное объединением частей двух раздельных антител, распознающих две разные антигенные группы. Это может быть достигнуто поперечным связыванием или рекомбинантными методиками. Кроме того, в антитело или его часть могут быть добавлены группировки, увеличивающие период полувыведения in vivo (например, увеличивая время клиренса из кровотока). Такие методики включают, например, добавление ПЭГ-группировок (также называемое пегилированием), и хорошо известны в данной области. См. публикацию заявки на патент США №20030031671.
Рекомбинантная экспрессия антител
В некоторых воплощениях нуклеиновые кислоты, кодирующие соответствующее моноклональное антитело, вводят непосредственно в клетку-хозяина и клетку инкубируют в условиях, достаточных для индукции экспрессии кодируемого антитела. В типичных случаях после введения соответствующих нуклеиновых кислот в клетку ее инкубируют, обычно при 37°С, иногда проводя селекцию, на протяжении периода продолжительностью приблизительно 1-24 часа для обеспечения возможности экспрессии антитела. В одном воплощении происходит секреция антитела в супернатант среды, в которой растет клетка. Традиционно моноклональные антитела получают в форме нативных молекул в мышиных гибридомных линиях. В дополнение к этой технологии, согласно настоящему изобретению предложена рекомбинантная экспрессия ДНК моноклональных антител. Это позволяет получать канинизированные антитела, а также ряд производных антител и слитых белков у выбранного вида-хозяина.
Возможна рекомбинация последовательности нуклеиновой кислоты, кодирующей по меньшей мере одно антитело, часть или полипептид против NGF по настоящему изобретению с векторной ДНК в соответствии с общепринятыми методиками, включая тупые концы или ступенчатые концы для лигирования, расщепление рестриктазами с получением подходящих концов, соответствующее заполнение липких концов, обработку щелочной фосфатазой во избежание нежелательного соединения и лигирование подходящими лигазами. Методики для проведения таких манипуляций раскрыты, например, в Maniatis et al., MOLECULAR CLONING, LAB. MANUAL (Cold Spring Harbor Lab. Press, NY, 1982 and 1989), и Ausubel et al. 1993 supra и могут быть применены для конструирования последовательностей нуклеиновых кислот, кодирующих молекулу моноклонального антитела или его антигеневязывающую область.
Молекулу нуклеиновой кислоты, такую как ДНК, считают «способной экспрессировать» полипептид, если она содержит нуклеотидные последовательности, содержащие информацию, необходимую для регуляции транскрипции и трансляции, и такие последовательности «функционально связаны» с нуклеотидными последовательностями, кодирующими полипептид. Функциональная связь представляет собой связь, при которой регуляторные последовательности ДНК и последовательность ДНК, которую следует экспрессировать, соединены таким образом, который позволяет экспрессировать ген в виде пептидов или частей антител против NGF в количествах, поддающихся выделению. Конкретная природа регуляторных областей, необходимых для экспрессии генов, может варьировать от организма к организму, что хорошо известно в аналогичных областях. См., например, Sambrook et al., 2001 supra; Ausubel et al., 1993 supra.
Соответственно, настоящее изобретение охватывает экспрессию антитела или пептида против NGF в прокариотических или эукариотических клетках. Подходящие хозяева включают бактериальных или эукариотических хозяев, включая клетки бактерий, дрожжей, насекомых, грибов, птиц и млекопитающих in vivo или in situ или клетки-хозяева, имеющие происхождение от млекопитающих, насекомых, птиц или дрожжей. Клетка или ткань млекопитающего могут иметь происхождение от человека, примата, хомяка, кролика, грызуна, коровы, свиньи, овцы, лошади, козы, собаки или кошки, но может быть использована клетка любого другого млекопитающего.
В одном воплощении нуклеотидная последовательность по изобретению будет включена в плазмидный или вирусный вектор, способный к автономной репликации в реципиенте-хозяине. С этой целью может быть использован любой из широкого спектра векторов. См., например, Ausubel et al., 1993 supra. Факторы, важные для выбора конкретного плазмидного или вирусного вектора, включают: простоту распознавания и отбора клеток-реципиентов, содержащих вектор, от тех клеток-реципиентов, которые не содержат вектор; желаемое число копий вектора у конкретного хозяина; и желательность возможности «перехода» вектора между клетками-хозяевами разных видов.
Примеры прокариотических векторов, известных в данной области, включают плазмиды, такие как плазмиды, способные к репликации в Е. coli (такие как, без ограничения, например, pBR322, ColE1, pSC101, pACYC 184 и тому подобные). Такие плазмиды раскрыты, например, в Maniatis et al., 1989 supra; Ausubel et al., 1993 supra. Плазмиды Bacillus включают рС194, рС221, рТ127 и так далее. Такие плазмиды раскрыты Gryczan в THE MOLEC. BIO. OF THE BACILLI 307-329 (Academic Press, NY, 1982). Подходящие плазмиды Streptomyces включают pIJ101 (Kendall et al., 169 J. Bacteriol. 4177-83 (1987)) и бактериофаги Streptomyces, такие как phiC31 (Chater et al., в SIXTH INT'L SYMPOSIUM ON ACTINOMYCETALES BIO. 45-54 (Akademiai Kaido, Budapest, Hungary 1986)). Обзор плазмид Pseudomonas представлен в John et al., 8 Rev. Infect. Dis. 693-704 (1986), Izaki, 33 Jpn. J. Bacteriol. 729-42 (1978), и Ausubel et al., 1993 supra.
Альтернативно, элементы экспрессии генов, полезные для экспрессии кДНК, кодирующей антитела или пептиды против NGF, включают, без ограничения: (а) вирусные промоторы транскрипции и их энхансерные элементы, такие как ранний промотор SV40 (Okayama et al., 3 Mol. Cell. Biol. 280 (1983)), LTR вируса саркомы Рауса (Gorman et al., 79 Proc. Natl. Acad. Sci., USA 6777 (1982)) и LTR вируса мышиного лейкоза Молони (Grosschedl et al., 41 Cell 885 (1985)); (b) области сплайсинга и сайты полиаденилирования, такие как области и сайты, имеющие происхождение от поздней области SV40 (Okayarea et al., 1983); и (с) сайты полиаденилирования, такие как у SV40 (Okayama etal., 1983).
кДНК-гены иммуноглобулинов могут быть экспрессированы, как описано Weidle et al., 51 Gene 21 (1987), с использованием, в качестве экспрессионных элементов, раннего промотора SV40 и его энхансера, энхансеров промотора Н-цепи мышиных иммуноглобулинов, поздней области сплайсинга мРНК SV40, вставочной последовательности кроличьего S-глобина, сайтов полиаденилирования иммуноглобулинов и кроличьего S-глобина и элементов полиаденилирования SV40. Для генов иммуноглобулинов, представленных частично кДНК, частично геномной ДНК (Whittle et al., 1 Protein Engin. 499(1987)), транскрипционный промотор может представлять собой промотор цитомегаловируса человека, энхансеры промотора могут представлять собой энхансеры цитомегаловируса и мышиных/человеческих иммуноглобулинов, а области сплайсинга мРНК и полиаденилирования могут представлять собой нативные хромосомные последовательности иммуноглобулинов.
В одном воплощении для экспрессии кДНК-генов в клетках грызуна транскрипционный промотор представляет собой вирусную LTR-последовательность, энхансеры транскрипционного промотора представляют собой энхансер тяжелой цепи мышиного иммуноглобулина и/или вирусный LTR-энхансер, область сплайсинга содержит интрон размером более 31 п.о., а области полиаденилирования и терминации транскрипции имеют происхождение от нативной хромосомной последовательности, соответствующей синтезируемой цепи иммуноглобулина. В других воплощениях с указанными выше экспрессионными элементами комбинируют последовательности кДНК, кодирующие другие белки, для обеспечения экспрессии белков в клетках млекопитающих.
Каждый слитый ген может быть собран в векторе экспрессии или введен в вектор экспрессии. Затем клетки-реципиенты, способные экспрессировать ген химерной цепи иммуноглобулина, трансфицируют одним геном, кодирующим пептид против NGF, или химерную Н-цепь, или химерную L-цепь, или котрансфицируют генами химерной Н-цепи и химерной L-цепи. Трансфицированные клетки-реципиенты культивируют в условиях, позволяющих экспрессировать введенные гены, и экспрессированные цепи иммуноглобулинов, интактные антитела или фрагменты выделяют из культуры.
В одном воплощении слитые гены, кодирующие пептид против NGF, или химерные Н- и L-цепи, или их части, собирают в раздельных векторах экспрессии, которые затем используют для котрансфекции клетки-реципиента. Альтернативно, слитые гены, кодирующие химерные Н- и L-цепи, могут быть собраны на одном и том же векторе экспрессии. Применительно к трансфекции векторами экспрессии и получения химерного антитела линия клеток-реципиентов может представлять собой клетки миеломы. Клетки миеломы могут обеспечивать синтез, сборку и секрецию иммуноглобулинов, кодируемых генами иммуноглобулинов, использованными при трансфекции, и обладают механизмом гликозилирования иммуноглобулинов. Клетки миеломы можно выращивать в культуре или в брюшинной полости мыши, в случае чего секретированый иммуноглобулин может быть получен из асцитической жидкости. Другие подходящие клетки-реципиенты включают лимфоидные клетки, такие как В-лимфоциты, имеющие происхождение от человека или вида, не являющегося человеком, гибридомные клетки, имеющие происхождение от человека или вида, не являющегося человеком, или межвидовые гетерогибридомные клетки.
Вектор экспрессии, содержащий конструкцию химерного, канинизированного антитела или полипептид против NGF по настоящему изобретению, может быть введен в соответствующую клетку-хозяина любым из множества подходящих способов, включая такие биохимические способы, как трансформация, трансфекция, конъюгация, слияние протопластов, преципитация с фосфатом кальция и применение поликатионов, таких как диэтиламиноэтилдекстран (DEAE-декстран), и такие механические способы, как электропорация, прямая микроинъекция и бомбардировка микрочастицами. Johnston et al., 240 Science 1538 (1988).
Дрожжи могут обеспечить существенные преимущества над бактериями при получении Н- и L-цепей иммуноглобулинов. В дрожжах происходят посттрансляционные модификации пептидов, включая гликозилирование. В настоящее время существует несколько методик с рекомбинантными ДНК, в которых используют последовательности сильных промоторов и плазмиды с большим числом копий, которые могут быть применены для получения желаемых белков в дрожжах. Дрожжи распознают лидерные последовательности продуктов клонированных генов млекопитающих и секретируют пептиды, содержащие лидерные последовательности (то есть препептиды). Hitzman et al., 11th Int'l Conference on Yeast, Genetics & Molec. Biol. (Montpelier, France, 1982).
Дрожжевые системы экспрессии генов можно регулярно оценивать на предмет уровней продукции, секреции и стабильности пептидов против NGF-31, антитела и собранных мышиных и химерных, гетерохимерных, канинизированных антител, их фрагментов и областей. Можно использовать любую из ряда дрожжевых систем экспрессии генов, включающую промоторные и терминационные элементы активно экспрессируемых генов, кодирующих гликолитические ферменты, продуцируемые в больших количествах при выращивании дрожжей в средах с высоким содержанием глюкозы. Известные гликолитические гены могут также обеспечивать очень эффективные сигналы контроля транскрипции. Например, могут быть использованы промоторные и терминаторные сигналы гена фосфоглицераткиназы (ФГК). Для оценки оптимальной экспрессии плазмид для экспрессии клонированных кДНК иммуноглобулинов у дрожжей могут быть применены несколько методик. См. Vol. II DNA Cloning, 45-66, (Glover, ed.,) IRL Press, Oxford, UK (1985).
В качестве хозяев для получения молекул антител или пептидов, описанных в данном изобретении, могут также быть использованы бактериальные штаммы. Вместе с этими бактериальными хозяевами используют плазмидные векторы, содержащие репликон и контрольные последовательности, имеющие происхождение от вида, совместимого с клеткой-хозяином. Вектор содержит сайт репликации, а также определенные гены, способные обеспечивать фенотипическую селекцию трансформированных клеток. Для оценки экспрессии плазмид для получения мышиных, химерных, гетерохимерных, канинизированных антител, фрагментов и областей или цепей антител, кодируемых клонированными кДНК иммуноглобулинов, у бактерий могут быть применены несколько методик (см. Glover, 1985 supra; Ausubel, 1993 supra; Sambrook, 2001 supra; Colligan et al., eds. Current Protocols in Immunology, John Wiley & Sons, NY, NY (1994-2001); Colligan et al., eds. Current Protocols in Protein Science, John Wiley & Sons, NY, NY (1997-2001)).
Клетки-хозяева, являющиеся клетками млекопитающих, можно выращивать in vitro или in vivo. Клетки млекопитающих обеспечивают посттрансляционные модификации белковых молекул иммуноглобулинов, включая удаление лидерных пептидов, фолдинг и сборку Н- и L-цепей, гликозилирование молекул антител и секрецию функциональных белков антител. Клетки млекопитающих, которые могут быть полезны в качестве хозяев для получения белков антител, помимо клеток лимфоидного происхождения, описанных выше, включают клетки, имеющие происхождение от фибробластов, такие как клетки Vero (ATCCCRL81) или СНО-K1 (ATCCCRL61). Для экспрессии клонированных генов Н- и L-цепей пептидов против NGF в клетках млекопитающих доступно множество векторных систем (см. Glover, 1985 supra). Для получения полных антител H2L2 могут быть применены различные методики. Возможна одновременная экспрессия Н- и L-цепей в одних и тех же клетках, позволяющая достичь внутриклеточного объединения и связывания Н- и L-цепей с образованием полных тетрамерных антител H2L2 и/или пептидов против NGF. Одновременная экспрессия возможна при использовании одинаковых или разных плазмид у одного и того же хозяина. Гены Н- и L-цепей и/или пептидов против NGF могут быть помещены в одну и ту же плазмиду, которой затем трансфицируют клетки, проводя, таким образом, непосредственный отбор клеток, экспрессирующих обе цепи. Альтернативно, клетки могут сначала быть трансфицированы плазмидой, кодирующей одну цепь, например, L-цепь, с последующей трансфекцией полученной клеточной линии плазмидой с Н-цепью, содержащей второй селектируемый маркер. Клеточные линии, продуцирующие пептиды и/или молекулы H2L2 против NGF любым из указанных способов, могут быть трансфицированы плазмидами, кодирующими дополнительные копии пептидов, Н-, L-или Н- и L-цепей в сочетании с дополнительными селектируемыми маркерами, для получения клеточных линий с улучшенными свойствами, такими как повышенная продукция собранных молекул антител H2L2 или повышенная стабильность трансфицированных клеточных линий.
Для длительного получения рекомбинантных антител с высоким выходом может быть применена стабильная экспрессия. Например, могут быть получены клеточные линии, стабильно экспрессирующие молекулу антитела. Вместо использования векторов экспрессии, содержащих вирусные репликаторы, клетки-хозяева могут быть трансформированы с использованием экспрессионных кассет иммуноглобулинов и селектируемого маркера. Клетки, полученные после введения чужеродной ДНК, можно выращивать на протяжении 1-2 суток в обогащенных средах и затем переносить их на селективные среды. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к селекции и позволяет клеткам стабильно интегрировать плазмиду в хромосому и расти, образуя фокусы, которые, в свою очередь, могут быть клонированы и размножены с получением клеточных линий. Полученные таким образом клеточные линии могут быть особенно полезны при скрининге и оценке соединений/компонентов, непосредственно или косвенно взаимодействующих с молекулой антитела.
После получения антитела по изобретению оно может быть очищено любым методом очистки молекул иммуноглобулинов, известным в данной области, например, хроматографией (например, ионообменной, аффинной, особенно аффинной хроматографией с использованием специфического антигена после белка А, и гель-фильтрацией на колонках), центрифугированием, по дифференциальной растворимости или любой другой стандартной методикой очистки белков. Многие воплощения включают секрецию антител из клетки в культуральную среду и их выделение из культуральной среды.
Фармацевтические и ветеринарные применения
Антигенсвязывающий белок или фрагменты антител против NGF, как описано здесь, по настоящему изобретению могут быть использованы, например, в лечении расстройств, связанных с NGF, у собак и кошек. Конкретнее, согласно изобретению также предложена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель или разбавитель и, в качестве активного ингредиента, антитело или фрагмент антитела по изобретению. Антитело может быть химерным, гетерохимерным, канинизированным, фелинизированным, эквинизированным, гуманизированным или прошедшим видовую адаптацию для его применения у другого вида. Также предусмотрены интактные иммуноглобулины или их связывающие фрагменты, такие как Fab. Антитело и его фармацевтические композиции по данному изобретению могут быть введены парентерально, например, подкожно, внутримышечно или внутривенно.
Антитела и/или пептиды против NGF, по настоящему изобретению могут быть введены как отдельные терапевтические агенты или в комбинации с другими терапевтическими агентами. Они могут быть введены сами по себе, но обычно их вводят с фармацевтическим носителем, выбранным, исходя из выбранного пути введения и стандартной фармацевтической практики. Введение антител, раскрытых здесь, может быть осуществлено любым подходящим способом, включая парентеральную инъекцию (такую как внутрибрюшинная, подкожная или внутримышечная инъекция), пероральное или местное введение антител (обычно в составе фармацевтической композиции) на поверхность дыхательных путей. Местное введение на поверхность дыхательных путей может быть осуществлено посредством интраназального введения (например с использованием капельницы, тампона или ингалятора). Местное введение антител на поверхность дыхательных путей может также быть осуществлено посредством ингаляционного введения, такого как получение пригодных для вдыхания частиц фармацевтической композиции (включая как твердые, так и жидкие частицы), содержащей антитела, в форме аэрозольной суспензии с последующим вдыханием этих пригодных для вдыхания частиц субъектом. Способы и устройства для введения пригодных для вдыхания частиц фармацевтических композиций хорошо известны, и может быть применена любая общепринятая методика.
В некоторых желаемых воплощениях антитела вводят парентеральной инъекцией. Для парентерального введения антитела или пептиды против NGF могут быть изготовлены в форме раствора, суспензии, эмульсии или лиофилизированного порошка вместе с фармацевтически приемлемым наполнителем для парентерального введения. Например, наполнитель может представлять собой раствор антитела или их смесь, растворенные в приемлемом носителе, таком как водный носитель; такими наполнителями являются вода, физиологический раствор, раствор Рингера, раствор декстрозы, раствор трегалозы или сахарозы или 5%-й сывороточный альбумин, 0,4%-й физиологический раствор, 0,3%-й глицин и тому подобное. Также могут быть использованы липосомы и неводные наполнители, такие как нелетучие масла. Эти растворы стерильны и обычно не содержат твердых частиц. Эти композиции можно стерилизовать общепринятыми и хорошо известными методиками стерилизации. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для создания условий, близких к физиологическим, такие как агенты для коррекции рН и буферные агенты, агенты для коррекции тоничности и тому подобное, например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и так далее. Концентрация антитела в этих композициях может варьировать в широких пределах, например от менее чем приблизительно 0,5%, обычно от 1% или по меньшей мере приблизительно 1% до 15% или 20% по массе, и ее будут выбирать, исходя в первую очередь из объемов жидкостей, значений вязкости и так далее, в соответствии с конкретным выбранным способом введения. Наполнитель или лиофилизированный порошок может содержать добавки, которые поддерживают изотоничность (например хлорид натрия, маннит) и химическую стабильность (например буферы и консерванты). Композицию стерилизуют широко применяемыми методиками. Существующие способы изготовления парентерально вводимых композиций будут известны или очевидны специалистам в данной области и описаны более подробно, например, в REMINGTON'S Р HARMA. SCI. (15th ed., Mack Pub. Co., Easton, Pa., 1980).
Антитела по данному изобретению могут быть лиофилизированы для хранения и восстановлены в подходящем носителе перед применением. Было показано, что эта методика эффективна при использовании обычных иммунноглобулинов. Могут быть применены любые подходящие методики лиофилизации и восстановления. Специалистам в данной области будет ясно, что лиофилизация и восстановление могут привести к той или иной степени потери активности антител и что это может привести к необходимости коррекции применяемых уровней. Композиции, содержащие настоящие антитела или их смесь, могут быть введены для предотвращения рецидива и/или терапевтического лечения существующего заболевания. Подходящие фармацевтические носители описаны в последнем издании REMINGTON'S PHARMACEUTICAL SCIENCES, стандартном руководстве в данной области. При терапевтическом применении композиции вводят субъекту, уже страдающему от заболевания, в количестве, достаточном для излечения или по меньшей мере частичной остановки развития или облегчения заболевания и его осложнений. Количество, адекватное для достижения этого, определяют как «терапевтически эффективную дозу» или «терапевтически эффективное количество». Количества, эффективные для такого применения, будут зависеть от тяжести заболевания и общего состояния собственной иммунной системы субъекта, но обычно входят в диапазон от приблизительно 0,1 мг антитела на кг массы тела до приблизительно 10 мг антитела на кг массы тела, предпочтительно от приблизительно 0,3 мг антитела на кг массы тела до приблизительно 5 мг антитела на кг массы тела. Минимальное содержание посторонних веществ и снижение вероятности отторжения «чужеродного вещества», достигаемые настоящими антителами, подобными собачьим антителам, по данному изобретению, могут обеспечить возможность введения этих антител в существенном избытке.
Разумеется, вводимая доза будет варьировать в зависимости от известных факторов, таких как фармакодинамические характеристики конкретного агента, а также способ и путь его введения, возраст, состояние здоровья и масса тела реципиента, характер и выраженность симптомов, тип сопутствующего лечения, частота введения и желаемый эффект.
В качестве неограничивающего примера, лечение патологии, связанной с NGF, у собак или кошек может быть проведено в форме введения антител против NGF по настоящему изобретению один раз в две недели или один раз в месяц в необходимом диапазоне доз. Примерами антител для терапевтического применения у собак являются высокоаффиные (они могут также быть высокоавидными) антитела и их фрагменты, области и производные, имеющие высокую активность против NGF in vivo, по настоящему изобретению. Однократное или многократное введение композиций может быть осуществлено при уровнях и типе дозы, выбранных лечащим ветеринаром. В любом случае, фармацевтические композиции должны обеспечивать количество антитела (антител) по данному изобретению, достаточное для эффективного лечения субъекта.
Диагностические применения
Согласно настоящему изобретению также предложены указанные выше антитела и пептиды против NGF для использования в диагностических методах для выявления NGF у вида, в частности у собак и кошек, с подтвержденным расстройством, связанным с NGF, или подозрением на наличие расстройства, связанного с NGF. В одном воплощении изобретения расстройство, связанное с NGF, представляет собой боль. В другом воплощении расстройство, связанное с NGF, представляет собой остеоартрит. Антитела и/или пептиды против NGF по настоящему изобретению полезны для иммунологических анализов, позволяющих выявлять или определять количество NGF или антител против NGF в образце. Иммунологический анализ на предмет NGF обычно включает инкубацию клинического или биологического образца в присутствии высокоаффинного (или высокоавидного) антитела или полипептида против NGF по настоящему изобретению, имеющего метку, поддающуюся выявлению, и способного селективно связываться с NGF, и выявление связанного меченого пептида или антитела в образце. Различные методики клинических анализов хорошо известны в данной области. См., например, IMMUNOASSAYS FOR THE 80'S (Voller et al., eds., Univ. Park, 1981). Такие образцы включают образцы тканевых биоптатов, крови, сыворотки и кала или жидкости, полученные у субъектов-животных и анализируемые посредством ELISA, как описано ниже. Таким образом, антитело или полипептид против NGF могут быть зафиксированы на нитроцеллюлозе или другой твердой подложке, позволяющей иммобилизовать клетки, клеточные частицы или растворимые белки. Подложку можно затем промыть подходящими буферами с последующей обработкой пептидом, антителом или антигенсвязывающим белком, имеющими метку, поддающуюся выявлению, и специфичными в отношении NGF. После этого твердую подложку можно промыть буфером во второй раз для удаления несвязанных пептида или антитела. Затем, на известных стадиях метода, можно определить количество связанной метки на твердой подложке.
«Твердая подложка» или «носитель» относится к любой подложке, способной связывать пептид, антиген или антитело. Хорошо известные подложки или носители включают стекло, полистирол, полипропилен, полиэтилен, поливинилиденфторид (PVDF), декстран, нейлон, амилозы, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Для целей настоящего изобретения носитель должен быть частично растворимым или нерастворимым. Вещество подложки может иметь практически любую возможную структурную конфигурацию при условии, что связанная с ним молекула сохраняет способность к связыванию с NGF или антителом против NGF. Таким образом, конфигурация подложки может быть сферической, как у гранулы, или цилиндрической, как у внутренней поверхности пробирки или внешней поверхности стержня. Альтернативно, поверхность может быть плоской, такой как лист, чашка для культивирования, тест-полоска и так далее. Например, подложки могут включать полистироловые гранулы. Специалистам в данной области будут известны многие другие подходящие носители для связывания антитела, пептида или антигена, или они смогут определить такие носители посредством рутинных экспериментов. На хорошо известных стадиях метода может быть определена связывающая активность заданной партии пептида, и/или антитела, или антигенсвязывающего белка против NGF. Специалисты в данной области могут определить оптимальные рабочие условия для проведения анализа посредством рутинных экспериментов.
Мечение пептида и/или антитела, специфичных в отношении NGF, меткой, поддающейся выявлению, может быть осуществлено посредством их связывания с ферментом для использования в ферментном иммунологическом анализе (EIA) или твердофазном иммуноферментном анализе (ELISA). Связанный фермент взаимодействует с доступным субстратом, приводя к образованию химической группировки, которая может быть выявлена, например, спектрофотометрическими, флуориметрическими или визуальными методами. Ферменты, которые могут быть использованы для мечения антител, специфичных в отношении NGF, по настоящему изобретению меткой, поддающейся выявлению, включают, без ограничения, малатдегидрогеназу, стафилококковую нуклеазу, дельта-5-стероидизомеразу, дрожжевую алкогольдегидрогеназу, альфа-глицерофосфатдегидрогеназу, триозофосфатизомеразу, пероксидазу хрена, щелочную фосфатазу, аспарагиназу, глюкозооксидазу, бета-галактозидазу, рибонуклеазу, уреазу, каталазу, глюкозо-6-фосфатдегидрогеназу, глюкоамилазу и ацетилхолинэстеразу. Радиоактивное мечение антител, специфичных в отношении NGF, позволяет выявлять NGF с применением радиоиммунного анализа (RIA). См. Work et al., LAB. TECHNIQUES & BIOCHEM. IN MOLEC. BIO (No. Holland Pub. Co., NY, 1978). Радиоактивный изотоп может быть выявлен такими методами, как применение гамма-счетчика или сцинтилляционного счетчика, или авторадиографией. Изотопы, особенно полезные для целей настоящего изобретения, включают 3H, 125I, 131I, 35S и 14С.
Также возможно мечение антител, специфичных в отношении NGF, флуоресцентным соединением. При воздействии света с соответствующей длиной волны на флуоресцентно меченое антитело его присутствие можно выявить по флуоресценции. Соединениями, чаще всего используемыми для флуоресцентного мечения, являются изотиоцианат флуоресцеина, родамин, фикоэритрин, фикоцианин, аллофикоцианин, o-фталевый альдегид и флуорескамин. Мечение антител или антигенсвязывающих белков, специфичных в отношении NGF, меткой, поддающейся выявлению, может также быть осуществлено с использованием флуоресцентных металлов, таких как 125Eu или другие металлы группы лантаноидов. Эти металлы могут быть присоединены к антителу, специфичному в отношении NGF, с использованием таких металлохелатирующих групп, как диэтилентриаминпентауксусная кислота (ДТПА) или этилендиаминтетрауксусная кислота (ЭДТА).
Кроме того, мечение антител, специфичных в отношении NGF, меткой, поддающейся выявлению, может быть осуществлено сочетанием с хемилюминесцентным соединением. Присутствие хемилюминесцентно меченого антитела затем определяют, выявляя люминесценцию, возникающую при химическом взаимодействии. Примерами соединений, полезных для хемилюминесцентного мечения, являются люминол, изолюминол, тероматический (theromatic) сложный эфир акридиния, имидазол, соль акридиния и сложный эфир щавелевой кислоты.
Сходным образом, для мечения антитела, его части, фрагмента, полипептида или производного, специфичных в отношении NGF, по настоящему изобретению может быть использовано биолюминесцентное соединение. Биолюминесценция представляет собой тип хемилюминесценции в биологических системах, где эффективность хемилюминесцентного взаимодействия повышена каталитическим белком. Присутствие биолюминесцентного белка определяют, выявляя люминесценцию. Важными биолюминесцентными соединениями, используемыми для мечения, являются люциферин, люцифераза и экворин.
Выявление антитела, его части, фрагмента, полипептида или производного, специфичных в отношении NGF, может быть осуществлено сцинтилляционным счетчиком, например, если метка, поддающаяся выявлению, представляет собой радиоактивный гамма-излучатель, или флуориметром, например, если метка представляет собой флуоресцентное вещество. В случае ферментной метки выявление может быть осуществлено колориметрическими методами с использованием субстрата соответствующего фермента. Выявление может также быть осуществлено визуальным сравнением степени ферментативного взаимодействия субстрата со стандартными образцами, полученными сходным образом.
Для целей настоящего изобретения NGF, выявляемый указанными выше анализами, может присутствовать в биологическом образце. Может быть использован любой образец, содержащий NGF. Например, образец представляет собой биологическую жидкость, такую как, например, кровь, сыворотка, лимфа, моча, кал, воспалительный экссудат, спинномозговая жидкость, амниотическая жидкость, экстракт или гомогенат ткани и тому подобное. Изобретение не ограничено анализами с использованием только этих образцов, однако в свете настоящего описания специалист в данной области сможет определить подходящие условия, позволяющие использовать другие образцы.
Выявление in situ может быть осуществлено посредством получения гистологического образца от субъекта-животного и обработки такого образца комбинацией меченых антител по настоящему изобретению. Антитело (или его часть) может быть доставлено нанесением или наложением меченого антитела (или его части) на биологический образец. Применяя такую методику, можно определять не только присутствие NGF, но и распределение NGF в анализируемой ткани. При применении настоящего изобретения специалисту в данной области будет ясно, что любой из широкого спектра гистологических методов (таких как методики окрашивания) может быть модифицирован для проведения такого выявления in situ.
Антитело, фрагмент или производное по настоящему изобретению могут быть адаптированы для использования в иммунометрическом анализе, также известном как «двухсайтовый» или «сэндвич»-анализ. Типичный иммунометрический анализ включает связывание некоторого количества немеченого антитела (или фрагмента антитела) с твердой подложкой, нерастворимой в анализируемой жидкости, добавление некоторого количества растворимого антитела, имеющего метку, поддающуюся выявлению, и/или определение количества тройного комплекса, образованного антителом, фиксированным на твердой фазе, антигеном и меченым антителом.
Антитела могут быть использованы для количественного или качественного выявления NGF в образце или для выявления присутствия клеток, экспрессирующих NGF. Это может быть осуществлено иммунофлуоресцентными методиками с использованием флуоресцентно меченого антитела (см. ниже) в сочетании с флуоресцентной микроскопией, проточно-цитометрическим или флуориметрическим выявлением. Для диагностических целей антитела могут быть мечеными или немечеными. Немеченые антитела могут быть использованы в комбинации с другими мечеными антителами (вторичными антителами), способными взаимодействовать с антителом, такими как антитела, специфичные в отношении константных областей собачьих иммуноглобулинов. Альтернативно, возможно прямое мечение антител. Может быть использован широкий спектр меток, таких как радионуклиды, флуоресцентные метки, ферменты, субстраты ферментов, кофакторы ферментов, ингибиторы ферментов, лиганды (в частности гаптены) и так далее. Доступно множество типов иммунологических анализов, таких как анализы, обсужденные ранее, и они хорошо известны специалистам в данной области. Важно, что антитела по настоящему изобретению могут быть полезны в диагностике расстройства, связанного с NGF, у собак. Конкретнее, антитело по настоящему изобретению позволяет определять сверхэкспрессию NGF у домашних животных. Таким образом, антитело по настоящему изобретению может стать важным инструментом для иммуногистохимии. Антитела по настоящему изобретению могут быть использованы в микроматрицах с антителами, особенно подходящих для оценки профилей экспрессии генов.
Наборы
Объем настоящего изобретения также включает наборы для практического применения соответствующих способов. Наборы содержат по меньшей мере одно или более чем одно из антител по настоящему изобретению, нуклеиновую кислоту, кодирующую указанное антитело/антитела, или клетку, содержащую указанную нуклеиновую кислоту. Антитело по настоящему изобретению может быть представлено в контейнере, обычно в лиофилизированной форме. Антитела, которые могут быть конъюгированы с меткой или токсином или быть неконъюгированными, обычно включены в наборы с буферами, такими как трис, фосфатный, карбонатный и так далее, стабилизаторами, биоцидами, инертными белками, например, сывороточным альбумином, или тому подобным. В большинстве случаев эти вещества будут присутствовать в количестве менее 5 масс. %, исходя из количества активного антитела, и обычно присутствуют в общем количестве по меньшей мере приблизительно 0,001 масс. %, снова исходя из концентрации антитела. Во многих случаях будет желательным включить инертный наполнитель или эксципиент для разбавления активных ингредиентов, где эксципиент может присутствовать в количестве приблизительно от 1% до 99 масс. % всей композиции. Если в анализе используют вторичное антитело, способное связываться с первичным антителом, оно будет обычно представлено в отдельном флаконе. Вторичное антитело обычно конъюгировано с меткой и включено в композицию, аналогично композициям антител, описанным выше. В большинстве случаев набор будет также содержать инструкции по применению.
Теперь изобретение будет описано далее посредством неограничивающих примеров, приведенных ниже.
ПРИМЕРЫ
Далее изобретение пояснено и обосновано посредством следующих примеров. Тем не менее, эти примеры никоим образом не следует рассматривать как ограничивающие объем изобретения. Наоборот, специалисту в данной области будет очевидно, что существуют другие воплощения, модификации и эквиваленты настоящего изобретения, не выходящие за рамки сущности настоящего изобретения и/или объема приложенной формулы изобретения.
Пример 1:
Синтез и очистка собачьего NGF (cNGF)
Разрабатывали ПЦР-праймеры с подходящими сайтами рестрикции для амплификации собачьего пре-про-β-NGF (SEQ ID NO: 59). Ген β-NGF клонировали в плазмиду pCTV927 (направленная плазмида Chromos) через сайты EcoRV/KpnI. Проводили котрансфекцию клеток CHOK1SV плазмидой pCTV927/β-NGF вместе с плазмидой pSIO343, кодирующей интегразу системы Chromos, с использованием трансфекционного реагента Lipofectamine 2000. Отдельные стабильные клоны анализировали на предмет экспрессии, и клон с высокой экспрессией был выбран для размножения и экспрессии для последующей очистки. Собачий β-NGF (cNGF), полученный в результате этих трансфекции, очищали с применением ионообменной хроматографии. Первичную очистку проводили в проточном режиме с использованием Q Sepharose FF (GE Healthcare #17-0510-01). Осветленный супернатант разводили водой 1:1 и рН корректировали до 8,5 с использованием 1 М трис. Разведенный образец смешивали с Q Sepharose FF в отношении 150:1 на протяжении более чем 1,5 часа. Смоле давали осесть и собирали несвязанный белок. Проводили дальнейшую очистку cNGF катионообменной хроматографией; его снова разводили водой 1:1 и загружали на SP-Sepharose FF (GE Healthcare #17-0729-01), предварительно уравновешенную с использованием 20 мМ трис, рН 8,5. После загрузки колонку промывали и затем проводили элюирование с линейным градиентом от 0 до 210 мМ NaCl (каждый в 20 мМ трис, рН 8,5) за 20 объемов колонки. Фракции анализировали посредством SDS-PAGE, объединяли и проводили диализ (3.5K mwco) против PBS при 4°С. Диализат собирали, стерилизовали фильтрацией и концентрацию измеряли по оптической плотности при 280 нм (1 мг/мл равен 1,48 А280).
Пример 2:
Иммунизация собак
Иммунизация собак может быть проведена методами, известными в данной области, и не ограничена каким-либо одним методом. В одном примере собачий NGF (как описано в Примере 1) вводят непосредственно собакам с адъювантом для стимуляции иммунного ответа. Для получения оптимальных ответов против антигена собакам проводили повторные инъекции и регулярно получали образцы сыворотки. Проводили мониторинг и определение гуморального иммунного ответа у иммунизированных собак с применением стандартных методов твердофазного иммуноферментного анализа (ELISA) с прямым связыванием антигена, как хорошо известно специалисту в данной области и описано ниже.
Пример 3:
Связывание с основным антигеном и активация В-клеток
Для оценки титров собачьих антител против NGF 100 мкл рекомбинантного собачьего NGF (10 мкг/мл) сорбировали в течение ночи в планшетах Immunolon 2Hb при 4°С. Лунки промывали с использованием PBS-T (PBS с 0,1% Tween) три раза и неспецифическое связывание блокировали, используя 200 мкл PBS и 5% нежирного снятого молока с инкубацией на протяжении 1 часа при комнатной температуре. После трехкратной промывки с использованием 300 мкл PBD-T проводили инкубацию разведений собачьей сыворотки на протяжении одного часа. Связывание с собачьими IgG против NGF выявляли с использованием смеси 100 мкл антител Bethyl против собачьего IgG1 (А40-120Р) и против собачьего IgG2 (А40-121Р), 0,2 мкг/мл. После добавления хромогенного субстрата (SureBlue Reserve ТМВ 1-Component Microwell Peroxidase Substrate, KPL 53-00-01) и десяти минут инкубации при комнатной температуре (к.т.) реакцию останавливали добавлением 100 мкл 0,1 Н HCl. Поглощение каждой лунки определяли как оптическую плотность (OD) при 450 нм.
Протокол активации собачьих В-клеток памяти
Мононуклеарные клетки периферической крови (РВМС) выделяли с применением разделения центрифугированием в градиенте Ficoll™. После выделения РВМС из образца проводили специфическую селекцию клеток, секретировавших антитела, по экспрессии специфических поверхностных маркеров антителообразующих клеток, хорошо известных специалисту в данной области и описанных в US 2014/0287402 и Callard and Kotowicz "Cytokine Cell Biology: A practical approach" Oxford University Press, 2000, pg 17-31, включенных сюда посредством ссылки. Перед фиксацией В-клеток на сортировочных чипах проводили активацию клеток in vitro. После выделения и заморозки клетки (РВМС, приблизительно 107 клеток на флакон) извлекали из жидкого азота и быстро размораживали на водяной бане. Клетки переносили в центрифужные пробирки объемом 15 мл и по каплям добавляли 12 мл полной среды. После центрифугирования клеток при 1000 об/мин на протяжении 10 мин осадок ресуспендировали в 10 мл полной среды и центрифугировали еще раз при 1000 об/мин на протяжении 10 мин. В завершение, клетки ресуспендировали в 4 мл полной среды.
Пример 4:
Последовательности ДНК, кодирующие антитела 9L12 (ZTS-841), 48L2 (ZTS-842) и 13L11
Отдельные интересующие клетки отделяли от микроматриц посредством микроманипуляции и помещали в микропробирки, содержавшие лизирующий буфер и магнитные гранулы, для захвата мРНК. кДНК получали из общей РНК с использованием смеси геноспецифичных праймеров, гибридизовавшихся в начале конкретных доменов НС гамма, LC каппа и LC лямбда. Фермент терминальную дезоксинуклеотидилтрансферазу (TdT) использовали для присоединения к 3'-концу продукта первой цепи кДНК. Для последующей первой ПЦР использовали смесь геноспецифичных обратных праймеров и универсальный прямой праймер. Затем проводили вложенную ПЦР по отдельности для каждой VH и VL, амплифицируя вариабельные области антител. Для этого использовали обратные праймеры, располагавшиеся в константных доменах НС или LC, вместе с универсальным прямым праймером. Фрагменты, амплифицированные при ПЦР, разделяли электрофорезом в агарозном геле. Ампликоны полноразмерных VH и VL, выделенные из отдельных клеток, клонировали в векторы экспрессии, содержавшие константные части соответствующих НС или LC. Последовательности собачьих вариабельных доменов представляли собой следующее: вариабельная область легкой цепи 9L12 (841) (SEQ ID NO: 7), которой соответствует нуклеотидная последовательность SEQ ID NO: 17; вариабельная область тяжелой цепи 9L12 (841) (SEQ ID NO: 8), которой соответствует нуклеотидная последовательность SEQ ID NO: 18; вариабельная область легкой цепи 48L2 (842) (SEQ ID NO: 27), которой соответствует нуклеотидная последовательность SEQ ID NO: 36; вариабельная область тяжелой цепи 48L2 (842) (SEQ ID NO: 28), которой соответствует нуклеотидная последовательность SEQ ID NO: 38; вариабельная область легкой цепи 13L11 (SEQ ID N0:51), которой соответствует нуклеотидная последовательность SEQ ID NO: 53; вариабельная область тяжелой цепи 13L11 (SEQ ID NO: 52), которой соответствует нуклеотидная последовательность SEQ ID NO: 54.
Константные области выделенных антител, которые были выделены, как описано выше, не использовали при последующем конструировании антител по изобретению. Fc-область рекомбинантных антител по изобретению содержит модифицированный вариант собачьего IgGB (Bergeron et al., Vet Immunol knmunopathol 2014 Jan 15:157 (1-2): 31-41) и была выбрана в силу ее периода полувыведения, биофизических свойств и сниженных эффекторных функций. Согласно Bergeron et al., собачий IgGB имеет хорошую аффинность в отношении собачьего FcRn и биофизические свойства, подходящие для его дальнейшей обработки. По результатам дифференциальной сканирующей калориметрии (DSC), проведенной с использованием собачьей Fc-области самой по себе, температурная стабильность константных областей составляла 70°С и 83°С. Эти температуры плавления сходны с температурами плавления коммерческих гуманизированных mAb или превышают их.
В домене СН2 собачьего IgGB были проведены три точковые мутации для устранения ADCC- и CDC-активности. Эта мутированная Fc названа здесь IgGB(e-) (SEQ ID NO: 43). Несмотря на то, что NGF является растворимой мишенью, эффекторные функции антитела против NGF были устранены для защиты от любых возможных нежелательных эффектов, связанных с действием на неспецифические мишени или эффекторной функцией. Эти мутации не оказывали влияния на иммуногенность данного mAb. Кроме того, мутации, проведенные для устранения эффекторных функций, не повлияли на связывание с FcRn или белком А. Было отмечено снижение связывания с собачьими FcγRI и FcγRIII, а также снижение ADCC-активности. Белок C1q является первым белком в каскаде комплемента и необходим для комплемент-зависимой цитотоксичности (CDC). Было показано снижение связывания IgGB(e-) с белком C1q. Аминокислотная последовательность описанной собачьей константной области НС-65 представлена как SEQ ID NO: 41, а соответствующая ей нуклеотидная последовательность представлена как SEQ ID NO: 42. Аминокислотная последовательность собачьей константной области лямбда представлена как SEQ ID NO: 60, а соответствующая ей нуклеотидная последовательность представлена как SEQ ID NO: 61.
Пример 5:
Определение аффинности связывания с антигеном
Аффинность связывания антител против собачьего NGF определяли с применением поверхностного плазмонного резонанса (SPR) и системы Biacore (Biacore Life Sciences (GE Healthcare), Уппсала, Швеция). Иммобилизацию собачьего и крысиного NGF проводили аминным сочетанием 5 мкг/мл NGF с применением N-гидроксисукцинимидной (NHS)/1-этил-3-(3-диметиламинопропил)карбодиимидной (EDC) химии. Чипы гасили этаноламином и оценивали аффинность, с которой все mAb-кандидаты связывались с иммобилизованным NGF. Различные концентрации собачьих и фелинизированных антител против NGF пропускали над поверхностями с NGF при мониторинге ассоциации антитела с антигеном и диссоциации образованного комплекса в реальном времени. Проводили кинетический анализ, получая равновесную константу диссоциации (KD). Результаты показаны в Таблице 1 ниже.
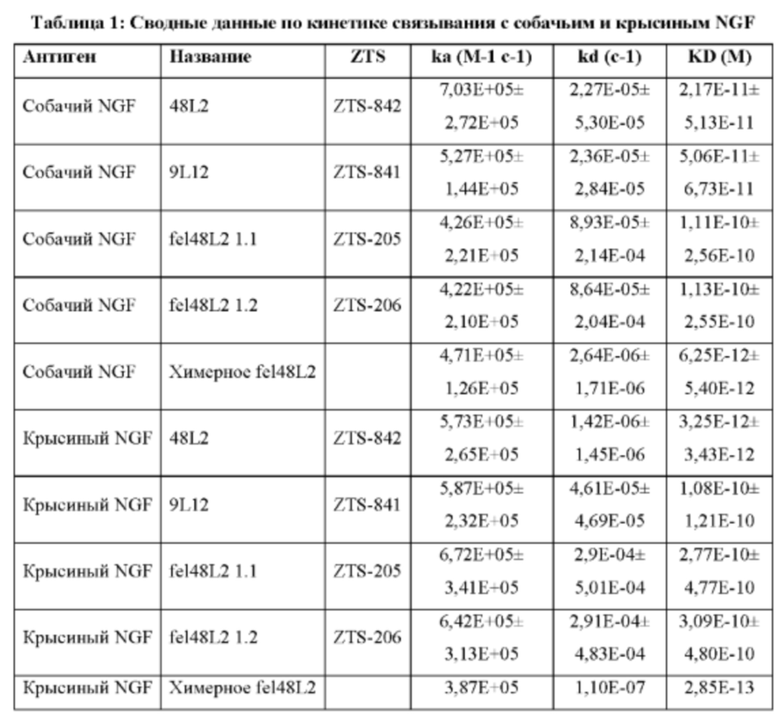
Пример 6:
Конструирование химерных антител 9L12 (841) и 48L2 (842)
Вариабельные домены антитела обеспечивают его связывание с антигеном. Антитела состоят из гомодимера, образованного парой двух гетеродимерных белков. Каждая белковая цепь (одна тяжелая и одна легкая) гетеродимера состоит из вариабельного домена и константного домена. Каждый вариабельный домен содержит три участка, определяющих комплементарность (CDR), которые участвуют в связывании с антигеном. CDR разделены в вариабельном домене каркасными областями, обеспечивающими остов для правильного пространственного представления связывающих сайтов антитела. Вместе взятые, CDR и каркасные области придают антителам способность к связыванию с их когнатными антигенами. Предполагается, что перенос полноразмерного вариабельного домена на соответствующую константную область окажет незначительное влияние или не повлияет на способность антитела связываться с NGF. Для одновременного подтверждения правильности определения последовательности вариабельных областей тяжелых и легких цепей и получения однородного материала были разработаны векторы экспрессии для получения рекомбинантных химерных или собачьих антител в системах экспрессии с клетками млекопитающих. Химерные антитела, описанные здесь, состоят из вариабельной последовательности (как CDR, так и каркасных областей) антитела вида-хозяина, перенесенной на соответствующие константные области тяжелой и легкой цепи молекулы собачьего IgG. Химерное антитело, описанное здесь, состоит из вариабельного сегмента (как CDR, так и каркасных областей) собачьей молекулы, перенесенного на соответствующие константные области тяжелой и легкой цепи молекулы кошачьего IgG. Поскольку связывание с антигеном обеспечивает вариабельный домен, предполагается, что перенос полностью собачьего вариабельного домена на кошачий константный домен окажет незначительное влияние или не повлияет на способность антитела связываться с NGF. Последовательности химерных вариабельных доменов представляли собой следующее: вариабельная область легкой цепи canfel_chimera 9L12 (841) (SEQ ID NO: 9), которой соответствует нуклеотидная последовательность SEQ ID NO: 19; вариабельная область тяжелой цепи canfel_chimera 9L12 (841) (SEQ ID NO: 10), которой соответствует нуклеотидная последовательность SEQ ID NO: 21; вариабельная область легкой цепи canfel_chimera 48L2 (842) (SEQ ID NO: 29), которой соответствует нуклеотидная последовательность SEQ ID NO: 39; тяжелая цепь canfel_chimera 48L2 (842) (SEQ ID NO: 30), которой соответствует нуклеотидная последовательность SEQ ID NO: 40. Каждый вариабельный сегмент клонировали в плазмиду для экспрессии в клетках млекопитающих, содержавшую тяжелую или легкую цепь кошачьего IgG. Аминокислотная последовательность кошачьей константной области тяжелой цепи представлена как SEQ ID NO: 62, а соответствующая ей нуклеотидная последовательность представлена как SEQ ID NO: 63. Аминокислотная последовательность кошачьей константной области легкой цепи представлена как SEQ ID NO: 64, а соответствующая ей нуклеотидная последовательность представлена как SEQ ID NO: 65.
Пример 7:
Фелинизация антител 48L2 и 9L12
Выработка противолекарственных антител (ADA) может приводить к снижению эффективности любых биотерапевтических белков, включая моноклональные антитела. Всесторонний анализ литературы показал, что видовая адаптация моноклональных антител может уменьшать склонность mAb к иммуногенности. Для снижения рисков, связанных с выработкой ADA, при применении собачьих моноклональных антител против NGF, предложенных здесь, была применена методика фелинизации для того, чтобы, в конечном счете, использовать эти антитела у кошек Эта методика фелинизации основана на идентификации кошачьей последовательности антитела эмбрионального типа, наиболее подходящей для использования при переносе CDR. После тщательного анализа всех кошачьих последовательностей эмбрионального типа, как тяжелой цепи, так и легкой цепи, кандидаты эмбрионального типа были отобраны, исходя из их гомологии собачьим mAb, и CDR собачьих сегментов-предшественников были использованы для замены нативных кошачьих CDR. Проводили экспрессию фелинизированных mAb и определение их способности связываться с NGF. Задача состояла в сохранении высокой аффинности и активности в отношении клеток при использовании каркасных областей кошачьих антител для минимизации иммуногенного потенциала in vivo. Были получены синтетические конструкции, в которых были представлены фелинизированные вариабельные области тяжелой и легкой цепи mAb 48L2 (ZTS-842), в котором использованы SEQ ID NO: 21-26, и 9L12 (ZTS-841), в котором использованы SEQ ID NO: 1-6. После субклонирования вариабельных областей каждой цепи в плазмиды, содержавшие кошачью константную область тяжелой цепи (SEQ ID NO: 62) и кошачью константную область легкой цепи (SEQ ID NO: 64), проводили котрансфекцию клеток HEK293 этими плазмидами для экспрессии антител. Проводили экспрессию химерных, гетерохимерных и фелинизированных вариантов mAb 48L2 и 9L12 и определяли их способность связываться с NGF посредством SPR.
Пример 8:
Получение антител с использованием глутаминсинтетазных (GS) плазмид
Гены, кодирующие собачьи и фелинизированные 9L12 и 48L2 (ZTS-841 и ZTS-842, соответственно), как описано здесь, и фелинизированные тяжелые и легкие цепи 9L12, клонировали в GS-плазмиды рЕЕ6.4 и рЕЕ12.4 (Lonza, Базель, Швейцария) с применением стандартных молекулярно-биологических методик, хорошо известных специалистам в данной области. Полученные отдельные плазмиды расщепляли по протоколу изготовителя и лигировали друг с другом, получая единую плазмиду для экспрессии в клетках млекопитающих. Для демонстрации транзиторной продукции каждого антитела каждую плазмиду использовали для трансфекции клеток HEK293 и проводили экспрессию в культурах разного размера. Белок выделяли из кондиционированной среды HEK с применением хроматографии с белом А в соответствии со стандартными методами очистки белков. Среду загружали на хроматографическую смолу и проводили элюирование со сдвигом рН. Перед использованием элюированного белка его подвергали коррекции рН, диализу и стерилизующей фильтрации. Анализировали аффинность и активность полученных антител.
Для получения стабильной клеточной линии, продуцирующей антитела-кандидаты, GS-плазмиду линеаризовали перед трансфекцией, используя рестриктазу PvuI, расщепляющую остов плазмиды в единственном сайте. Клетки GS-CHOK1SV (клон 144Е12) трансфицировали линеаризованной плазмидной ДНК посредством электропорации. После трансфекции клетки высевали в 48-луночные планшеты (48WP) для получения стабильных пулов. При по меньшей мере 50% смыкания монослоя в 48-луночных планшетах 100 мкл супернатанта анализировали на предмет экспрессии IgG с применением ForteBio Octet и биосенсоров с белком A (Pall ForteBio, Фримонт, штат Калифорния). Клоны с наилучшей экспрессией масштабировали в 6-луночных планшетах (6WP), а затем во встряхиваемых колбах (SF) объемом 125 мл. После адаптации клеток к суспензионной культуре в колбах объемом 125 мл по 2 колбы пула каждой клеточной линии сохраняли для длительного хранения. Поскольку промышленные клеточные линии должны быть клональными, 3 пула с наиболее высокой экспрессией были субклонированы серийным разведением в 96-луночных культуральных планшетах. Для подтверждения клональности и во избежание второго раунда серийного разведения 96-луночные планшеты визуализировали с использованием устройства Molecular Devices Clone-Select Imager (CSI) (Molecular Devices LLC, Сан-Хосе, штат Калифорния), позволяющего получить изображения отдельных клеток и их последующего роста. Отбор клонов проводили, исходя из успешных CSI-изображений, роста и продукции антител в 96-луночных планшетах.
Для оценки роста и продуктивности клеточных культур проводили дальнейшую оценку пулов с наибольшей экспрессией в подпитываемой культуре во встряхиваемых колбах объемом 125 мл на протяжении 14 суток. Посев клеток проводили с использованием сред-платформ и подпиток, состоявших из CD СНО от Life Technologies с 4 аминокислотами, собственной подпитки CDF v6.2 и 10%-й глюкозы. После 14 суток в подпитываемой культуре пулы центрифугировали и полученное mAb в CD СНО выделяли фильтрацией супернатанта через 0,20 мкм полиэфирсульфоновую мембрану (PES) перед очисткой.
Типичная очистка состоит из двух литров кондиционированной среды (из культуры клеток СНО, профильтрованной через 0,2 мкм фильтр), загружаемых на колонку MabSelect (GE healthcare, № по каталогу 17-5199-02) объемом 235 мл. Колонку предварительно уравновешивали с использованием PBS. Образец загружали при времени удержания более 2,5 минуты. После загрузки колонку промывали еще раз с использованием PBS и затем 25 мМ ацетатом натрия с рН, близким к нейтральному. Проводили элюирование с использованием 25 мМ уксусной кислоты, рН 3,6, и затем десорбцию с использованием 250 мМ уксусной кислоты, 250 мМ хлорида натрия, рН приблизительно 2,2. Фракции (50 мл) собирали на стадиях элюирования и десорбции. Проводили постоянный мониторинг поглощения УФ при А280. Пиковые фракции объединяли, рН корректировали приблизительно до 5,5 добавлением 20 мМ ацетата натрия и затем проводили диализ против трех смен буфера. Диализат собирали, стерилизовали фильтрацией и хранили при 4°С.
Пример 9:
Нейтрализация биологической активности собачьего NGF in vitro
Аффинность каждого собачьего и фелинизированного антитела против NGF в отношении собачьего NGF измеряли с применением SPR (поверхностного плазмонного резонанса) на системе Biacore system (Biacore Life Sciences (GE Healthcare), Уппсала, Швеция), как описано в Примере 5. Кроме того, был разработан функциональный in vitro-анализ для измерения констант ингибирования применительно к способности mAb ингибировать связывание NGF с TrkA. Для определения того, блокировали ли mAb против NGF по изобретению последующие клеточные сигналы в результате ингибирования связывания NGF с TrkA, очищенные антитела оценивали в анализе, позволявшем измерять индуцированное собачьим NGF фосфорилирование внутриклеточной сигнально регулируемой киназы 1 и 2 (pERK 1/2). Клетки, использованные в анализе представляли собой СНО-K1, экспрессировавшие собачий TrkA (Life Technologies), которые выращивали в среде DMEM/F12 с GlutaMAX™-1 (Life Technologies), дополненной 10%-й диализованной FBS, 20 мМ HEPES, 500 мкг/мл генетицина и 1х антибиотиком-антимикотиком мг/мл (Life Technologies) при 37°С в инкубаторе с 5% СО2, 95% воздуха с увлажением. Для анализа pERK 1/2 клетки высевали в количестве 5,0×104 клеток на лунку в 96-луночные планшеты для тканевых культур (Costar) и инкубировали в течение ночи при 37°С, позволяя клеткам закрепиться. Клетки подвергали сывороточному голоданию на протяжении 2 часов в HBSS, содержавшем хлориды кальция и магния (Life Technologies). Перед добавлением клеток проводили серийное разведение антител против NGF в HBSS и предварительную инкубацию с рекомбинантным собачьим NGF, разведенным в HBSS/0,1% BSA, при комнатной температуре на протяжении 1 часа. Конечные концентрации собачьего NGF и BSA при анализе составляли 15 нг/мл (ЕС90) и 0,025%, соответственно. Клетки стимулировали на протяжении 10 минут при 37°С перед удалением анализируемых смесей и добавлением 100 мкл лизирующего буфера, поставляемого с набором pERK 1/2 AlphaLISA® SureFire® Ultra Assay Kit (PerkinElmer). Затем лизаты обрабатывали, следуя инструкциям изготовителя, и планшеты считывали с использованием устройства для считывания планшетов EnSpire® (PerkinElmer). Максимальный ответ в анализе определяют как фосфорилирование ERK 1/2, измеренное в присутствии собачьего NGF самого по себе (без mAb). Минимальный ответ определяют как базальные уровни фосфорилирования ERK 1/2 (без стимуляции). Расчетные показатели ингибирования для антител против NGF выражены как процент от минимального и максимального ответа. Полученные данные по проценту ингибирования наносили на график с использованием GraphPad Prism 5 для определения IC50 (подбор кривых по 4 параметрам). См. Фиг. 6-7.
Анализ пролиферации клеток TF-1
Клетки TF-1 (АТСС) выращивали обычным образом в АТСС-модифицированной среде RPMI 1640 (Life Technologies), дополненной 10% FBS и 2 нг/мл рекомбинантного человеческого GM-CSF (R&D Systems Inc.). Среда для анализа пролиферации TF-1 представляла собой RPMI 1640, дополненную 10% BIT 9500 (Stemcell Technologies) и 10 мкг/мл гентамицина. Пролиферацию TF-1 проводили в 96-луночных микропланшетах (Costar), инкубируя 15000 клеток на лунку с собачьими и фелинизированными антителами против NGF в указанных концентрациях и 2 нг/мл рекомбинантного собачьего NGF. После 65-часового периода культивирования использовали набор CellTiter-GLO Luminescent Assay Kit (Promega) для оценки эффектов антител против NGF на пролиферацию клеток, индуцированную собачьим NGF. Максимальный ответ в анализе определяют как пролиферацию в присутствии собачьего NGF самого по себе (без антител). Минимальный ответ определяют как пролиферацию, измеренную без собачьего NGF. Расчетные показатели ингибирования (нейтрализация NGF) для антител против NGF выражены как процент от минимального и максимального ответа. Полученные данные по проценту ингибирования наносили на график с использованием GraphPad Prism 5 для определения IC50 (подбор кривых по 4 параметрам). См. Фиг. 8-9.
Пример 10:
Фармакокинетика
Фармакокинетика (ФК) обоих собачьих mAb против NGF 48L2 (ZTS-842) и 9L12 (ZTS-841). Исследования у собак проводили после двух подкожных (п/к) и одного внутривенного (в/в) введения в дозе 2,0 мг/кг, проводимых с интервалом 28 суток. Данные, полученные после в/в введения, продемонстрировали, что период полувыведения составлял 13,3 плюс/минус 3,4 суток (среднее плюс/минут стандартное отклонение), а клиренс был медленным (3,9 плюс/минус 0,2 мл/сутки/кг). При п/к введении пиковые концентрации в сыворотке наблюдались через 1-7 суток после введения. Абсолютная биодоступность при п/к введении составляла, в среднем, 88% плюс/минус 41%. Связывание с NGF in vivo было подтверждено с применением высоко чувствительного анализа общего NGF (свободного NGF и комплекса NGF-mAb). До введения 48L2 (ZTS-842) концентрации NGF были меньше нижнего предела количественного определения, 10 пг/мл. После введения 48L2 (ZTS-842) концентрации общего NGF возрастали у всех животных, составляя, в среднем, 1300 плюс/минус 500 пг/мл на 84 сутки исследования. Концентрации 48L2 (ZTS-842) были в большом избытке на всем протяжении исследования, составляя, в среднем, 7,8 плюс/минус 1,3 мкг/мл на 84 сутки, через 28 суток после последнего введения, указывая на то, что даже меньшие дозы будут адекватны для связывания эндогенного NGF на протяжении по меньшей мере одного месяца после введения. Несмотря на отсутствие прямой оценки иммуногенности, данных по динамике концентрации 48L2 (ZTS-842), которые указывали бы на индукцию противолекарственных антител, у четырех собак за время исследования продолжительностью 84 суток с тремя введениями, не было. См. Фиг. 11. Кроме того, ZTS-841 было также изучено с применением тех же параметров, что описаны выше; при п/к / п/к/ в/в введении с интервалом 28 суток в дозе 20 мг/кг период полувыведения составлял 11,8 плюс 4,1 суток. Биодоступность при п/к введении 94% плюс 12%. См. Фиг. 10-11.
ФК фелинизированного mAb против NGF fel48L21.1 (ZTS-205) изучали у 3 самцов и 3 самок кошек после двух подкожных (п/к) и одного внутривенного (в/в) введения в дозе 1,5 мг/кг с интервалом 28 суток. Данные, полученные после в/в введения, продемонстрировали, что период полувыведения составлял 10,8 плюс/минус 2,5 суток (среднее плюс/минут стандартное отклонение), а клиренс был медленным (3,0 плюс/минус 1,0 мл/сутки/кг). При п/к введении пиковые концентрации в сыворотке наблюдались через 2-7 суток после введения. Абсолютная биодоступность при п/к введении составляла, в среднем, 88% плюс/минус 17%. Концентрации fel48L21.1 (ZTS-205) были в высокими на всем протяжении исследования, составляя, в среднем, 7,2 плюс/минус 4,0 мкг/мл на 84 сутки, через 28 суток после последнего введения, указывая на то, что даже меньшие дозы будут адекватны для связывания эндогенного NGF на протяжении по меньшей мере одного месяца после введения. Несмотря на отсутствие прямой оценки иммуногенности, данных по динамике концентрации ZTS-842, которые указывали бы на индукцию противолекарственных антител, у шести кошек за время исследования продолжительностью 84 суток с тремя введениями, не было.
Методология биоаналитического анализа
Анализ связывания лиганда для определения свободного 48L2 (ZTS-842) был разработан на основе захвата свободного mAb биотинилированным собачьим NGF на стрептавидиновом диске Gyrolab™ и флуоресцентном выявлении после добавления мышиного моноклонального антитела против собачьего IgG, меченного AlexaFluor™. Анализ связывания лиганда для определения свободного fe148L21.1 (ZTS-205) был разработан на основе захвата свободного mAb биотинилированным собачьим NGF на стрептавидиновом диске Gyrolab™ и флуоресцентном выявлении после добавления козьего поликлонального антитела против кошачьего IgG, меченного AlexaFluor™.
Пример 11:
Изучение собачьего и фелинизированного антитела против NGF в MIA-модели у крыс
Остеоартрит (OA) представляет собой дегенеративное збаолевание суставов, характеризующееся прогрессирующим разрушением суставного хряща. Внутрисуставная инъекция MIA приводит к разрушению суставного хряща и развитием в субхондральной кости очагов, имитирующих OA. Данная модель является быстрым и минимально инвазивным методом воспроизведения очагов, сходных с OA, у грызунов.
Болеутоляющий эффект антител против NGF, прошедших видовую адаптацию (например, канинизированных, фелинизированных и тому подобных), после однократного введения MIA в MIA-модели остеоартрита у крыс был продемонстрирован раздельным введением моноклональных антител ZTS-841, ZTS-842 во время исследования на 7 сутки исследования и на 14 сутки исследования. Боль оценивали с применением нагрузочного теста для оценки стойкой боли и теста компрессии сустава (теста Рэндалла-Селитто) для оценки механической гиперальгезии с использованием анальгезимерта (Ugo Basile). Тест проводили, оказывая давление на заднюю лапу. При нажатии на педаль, активировавшую мотор, сила возрастала с постоянной скоростью по линейной шкале. При возникновении боли отмечают отдергивание лапы или вокализацию, нажатие на педаль немедленно прекращали и определяли болевой порог по шкале. Во избежание возможных травм максимальная нагрузка составляла 400 г. Тест Рэндалла-Селитто проводили на -1 (исходно), 20 и 28 сутки исследования. Схема MIA-анализа у крыс представлена на Фиг. 12.
Разрушение хряща индуцировали введением метаболического ингибитора монойодацетата (MIA). Крыс анестезировали изофлураном (3-5% в 100% O2). По достижении у животных полной анестезии проводят инъекцию 50 мкл MIA, 40 мг на один миллилитр физиологического раствора, в суставную щель левого коленного сустава с использованием шприца 1 сс с иглой 27G. Животному переставали давать изофлуран, позволяли полностью прийти в сознание и затем помещали обратно в его клетку.
Для оценки эффекта mAb против NGF по изобретению у этих животных оценивали их способность переносить нагрузку с использованием устройства Incapacitance Tester. Животных помещали в акриловую стетовую камеру при ее прямом положении и проводили оценку нагрузки. В каждой временной точке оценку проводили три раза. При каждой оценке рассчитывают процентный показатель нагрузки (WBS) с применением следующей формулы:
Среднее 3 WBS принимали как WBS для данной временной точки. На -21 сутки рассчитывали WBS до MIA-индукции, MIA вводили в левое колено в дозе 2 мг/0,05 мл. На -1 сутки WBS измеряли для рандомизации. На 0 сутки вводили mAb против NGF или плацебо и затем нагрузку оценивали на 7, 14, 21 и 28 сутки. Массу тела отмечали один раз в неделю в день оценки нагрузки.
Образцы сыворотки получали на 28 сутки после введения посредством терминальной кардиальной пункции. После эвтаназии посредством асфиксии в С02 проводили забор цельной крови посредством кардиальной пункции, помещали ее в пробирки для отделения сыворотки, давали ей свернуться при комнатной температуре, затем центрифугировали (3500 об/мин, 15 мин) и переносили в 96-луночные планшеты в двух аликвотах по 300 мкл каждая, как указано в таблице ниже. Образцы замораживали при температуре -10°С или ниже до анализа. Графическое представление оценки ZTS-841 и ZTS-842 в MIA-анализе у крыс приведено на Фиг. 13-15.
Пример 12:
Эффекты на хромоту: оценка канинизированного антитела в модели синовиита у собак
Хорошо известно, что воспалительные процессы в мягких тканях являются важным компонентом остеоартрита. В модели боли при синовиите транзиторное воспаление синовиальной оболочки одного колена индуцируют внутрисуставной инъекцией бактериального липополисахарида (LPS). Хромота, поддающаяся количественной оценке, появляется в течение 2 ч после индукции синовиита, достигает пика через 3-4 ч, ослабевает к 6 ч и полностью исчезает после 24 ч. Эту модель широко применяют в исследованиях мишеней для контроля боли.
Однократная внутривенная инъекция ZTS-841 в дозе 5 мг/кг интактным самцам биглей уменьшала хромоту, по сравнению с плацебо в виде физиологического раствора, в модели LPS-синовиита у собак. Как видно в Таблице 2 ниже, ZTS-841 продемонстрировало эффективность через 3 и часа после индукции LPS-синовиита.
В Таблице 2 и на Фиг. 16 приведены средние значения, полученные методом наименьших квадратов (со стандартной ошибкой), для хромоты по VAS в группах лечения через три и пять часов после индукции синовиита. Различия между 5 мг/кг ZTS-841 и плацебо были статистически значимыми.
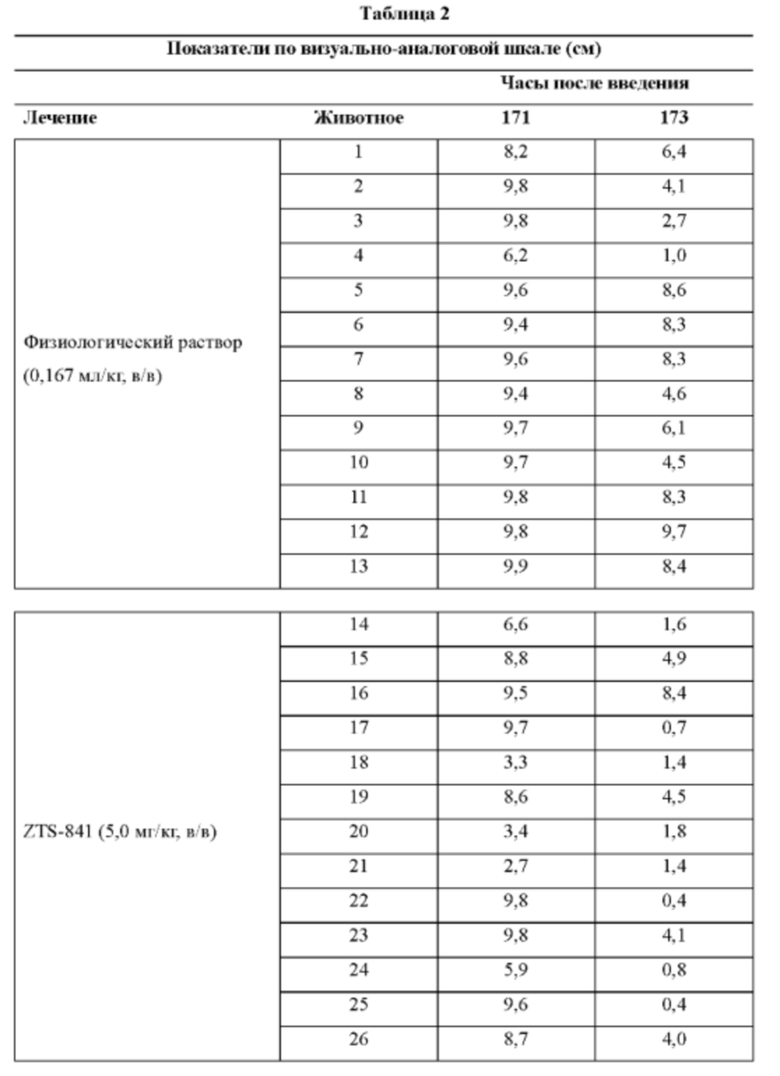
Пример 13:
Гуманизация антител 48L2 (ZTS-842) и 9L12 rZTS-841)
Аналогично описанной методике фелинизации, и как хорошо известно специалисту в данной области, из всех доступных человеческих последовательностей были идентифицированы последовательности антител эмбрионального типа, подходящие для переноса CDR с mAb 48L2 и 9L12. Вариабельные области легкой цепи и вариабельные области тяжелой цепи были выбраны, исходя из наибольшей гомологии соответствующим собачьим каркасным областям. CDR нативных человеческих сегментов удаляли и заменяли на исходные собачьи CDR. Рекомбинантные гуманизированные 48L2 и 9L12 получали с использованием выбранных вариабельных областей, соединенных с последовательностями соответствующих им константных областей тяжелой цепи собачьего IgG. Антитела получали из клеток HEK, очищали, как описано ранее, и затем оценивали на предмет их способности связываться с человеческим NGF, как показано в Таблице 3 ниже. Были сконструированы синтетические конструкции, в которых были представлены гуманизированные вариабельные области тяжелой и легкой цепи mAb 48L2 (ZTS-842) и 9L12 (ZTS-841). Были синтезированы различные комбинации вариабельных областей тяжелой и легкой цепи в рамках обеих групп, и они были проанализированы на предмет связывания (см. ниже). Последовательности CDR во время конструирования не изменяли, изменяли только последовательности каркасных областей.
Аффинность связывания антител против человеческого NGF (SEQ ID NO: 66) определяли с применением поверхностного плазмонного резонанса (SPR). Человеческий NGF иммобилизовали на поверхности чипа BIACORE прямым аминным сочетанием. Различные концентрации описанных гуманизированных антител против NGF пропускали над поверхностями с человеческим NGF при мониторинге ассоциации антитела с антигеном и диссоциации образованного комплекса в реальном времени. Проводили кинетический анализ, получая равновесную константу диссоциации (KD). Результаты показаны в Таблице 3 ниже.
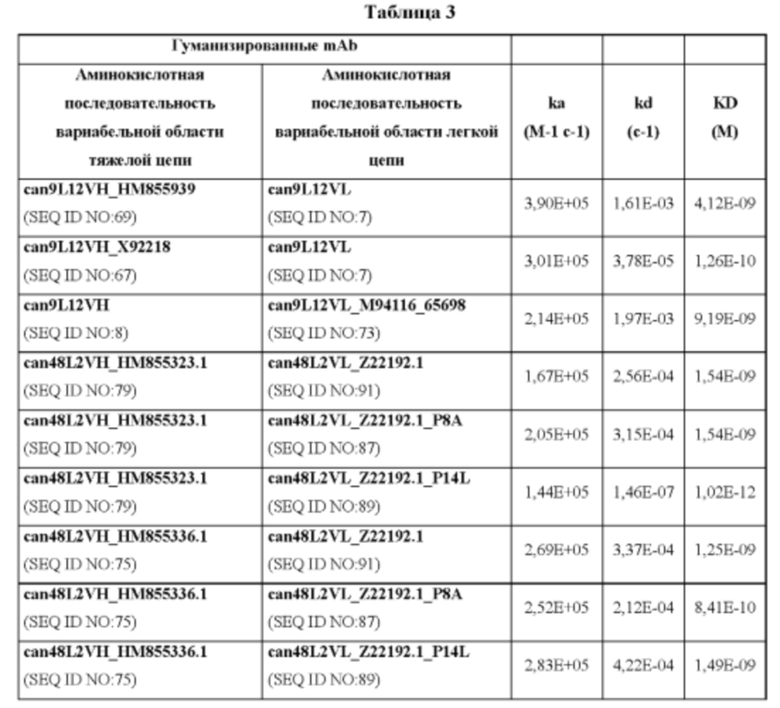
Пример 14:
Сканирующий мутагенез паратопов антител 48L2 9L12 (ZTS-841) и 48L2 (ZTS-842)
Область антитела, обеспечивающая распознавание антигена, представляет собой паратоп.Паратоп образован комбинацией аминокислот гипервариабельных участков (CDR) вариабельных областей тяжелой и легкой цепи. Связывание антитела с антигеном часто опосредовано боковыми цепями остатков CDR и боковыми цепями или углеводными группировками антигена. Для содействия определению критически важных боковых цепей, вовлеченных в распознавание антигена антителом, проводили мутагенез с аланиновым сканированием по каждому остатку CDR как тяжелой, так и легкой цепи; методика описана в Cunningham and Wells (1989) Science, Vol. 244, Issue 4908, pp. 1081-1085. Полученные мутанты затем по отдельности анализировали на предмет их способности связываться с NGF с применением Biacore. Измеряли аффинность связывания с человеческим NGF (hN ниже), собачьим NGF (cN ниже) и крысиным NGF (rN ниже) и значения KD получали по тем же протоколам, что описаны в Примерах 5 и 9 выше. Затем полученные значения сравнивали с антителами дикого типа, и они представлены в таблицах как процент связывания дикого типа. Данные, представленные в Таблицах 4 и 5 ниже, показаны как «процентный показатель сходства» в сравнении с диким типом.
Для определения относительной аффинности мутантных mAb, полученных при аланиновом сканировании, в сравнении с исходным mAb, определяли профили связывания с чипами, покрытыми NGF, при 100 нМ с использованием Biacore Т200. Среднее число единиц ответа после четырех повторов, полученное для исходного mAb, плюс/минус 3 стандартных отклонения использовали для расчета параметров, определяющих пороговое число единиц ответа, включая скорости ассоциации и диссоциации при связывании антитела. Процент точек данных для каждого мутанта, соответствовавших этому пороговому значению, затем использовали для определения «% показателя сходства». Показатель сходства после замены каждого положения в CDR тяжелой и легкой цепи ZTS-841 и ZTS-842 на аланин показан как «процент ингибирования в сравнении с исходным антителом» Таблицах 4 и 5, соответственно. Результаты замены каждого положения в CDR на аланин.
Последовательности, представленные в Таблице 4, относятся к аланиновому мутагенезу аминокислотных последовательностей CDR вариабельной области тяжелой цепи и вариабельной области легкой цепи ZTS-841 (9L12). Таблица 5 относится к аланиновым заменам в аминокислотных последовательностях CDR вариабельных областях тяжелой и легкой цепи ZTS-842 (48L2). Мутированные аминокислоты указаны в таблице в соответствии с нумерацией дикого типа как для последовательностей вариабельной области тяжелой цепи, так и для последовательностей вариабельной области легкой цепи, как описано ранее и включено ниже. Ниже выделены аминокислотные положения 1, 25, 50, 75 и 100. В столбце «название образца» последовательности вариабельных областей тяжелой или легкой цепи указаны с номером аминокислотного положения, в котором проведена заменена на аланин.
ZTS-841 VH: SEP ID NO: 8:
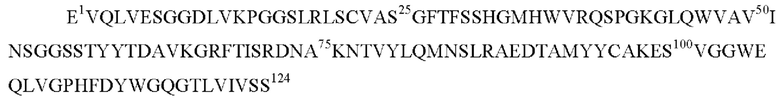
ZTS-841 VL: SEP ID NO: 7:
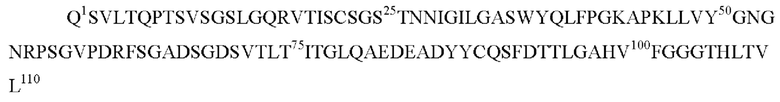
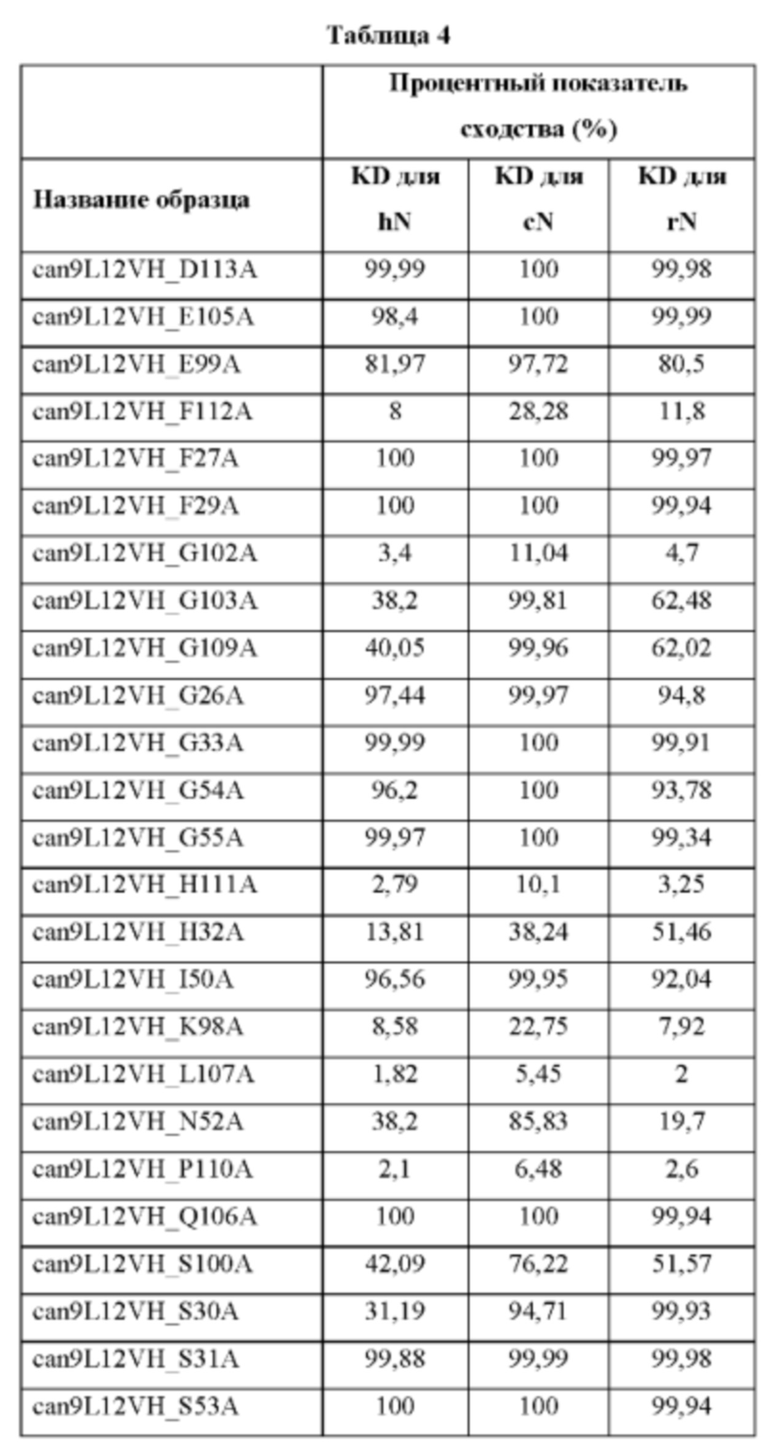
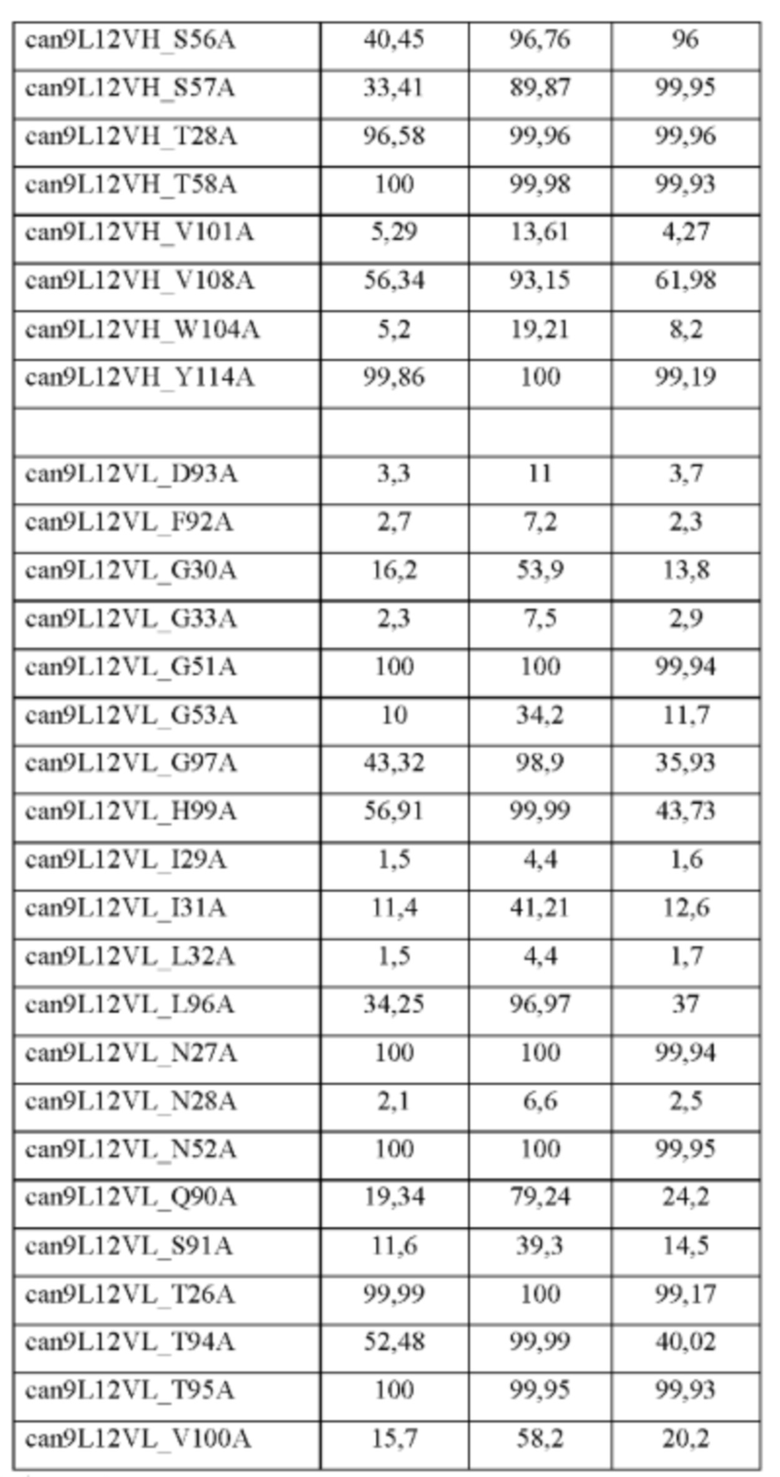
Для Таблицы 5:
ZTS-842 VH: SEP ID NO: 28:
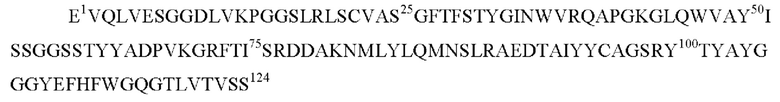
ZTS-842 VL: SEP ID NO: 27:
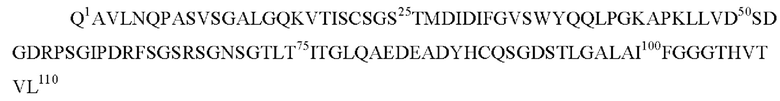
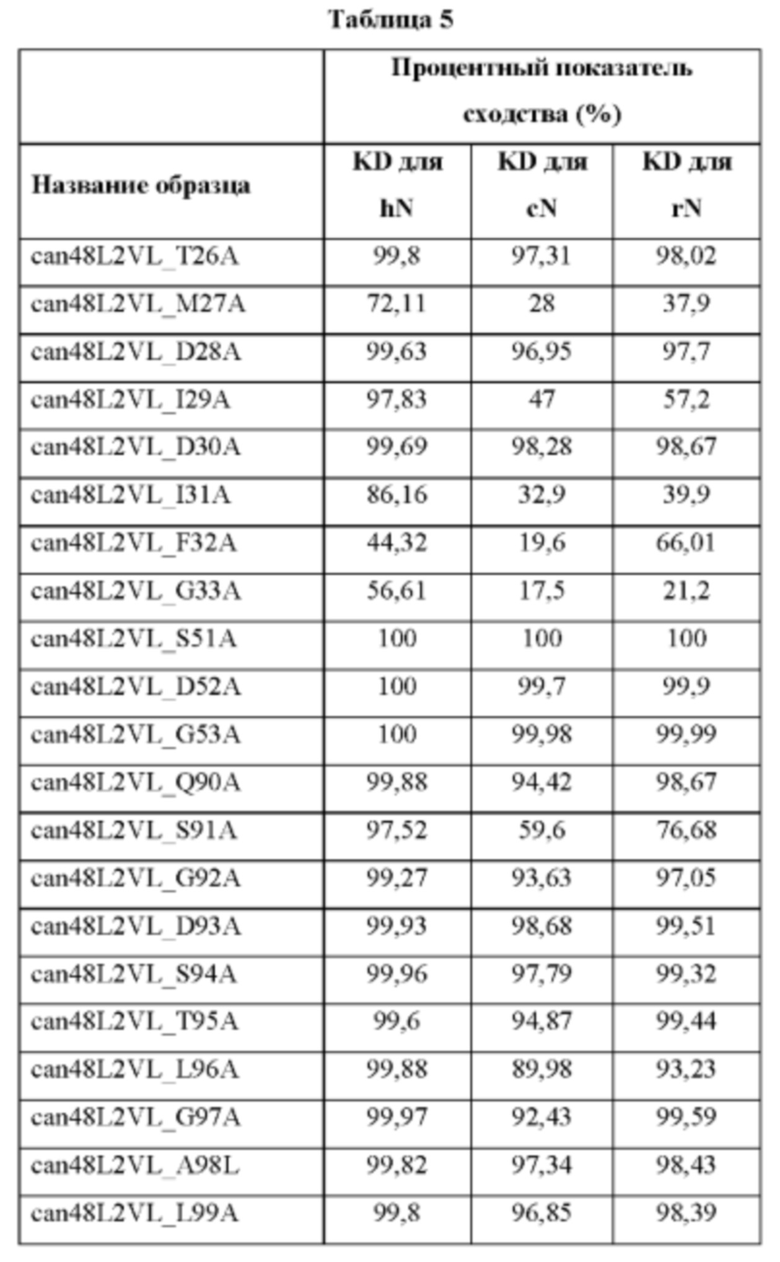
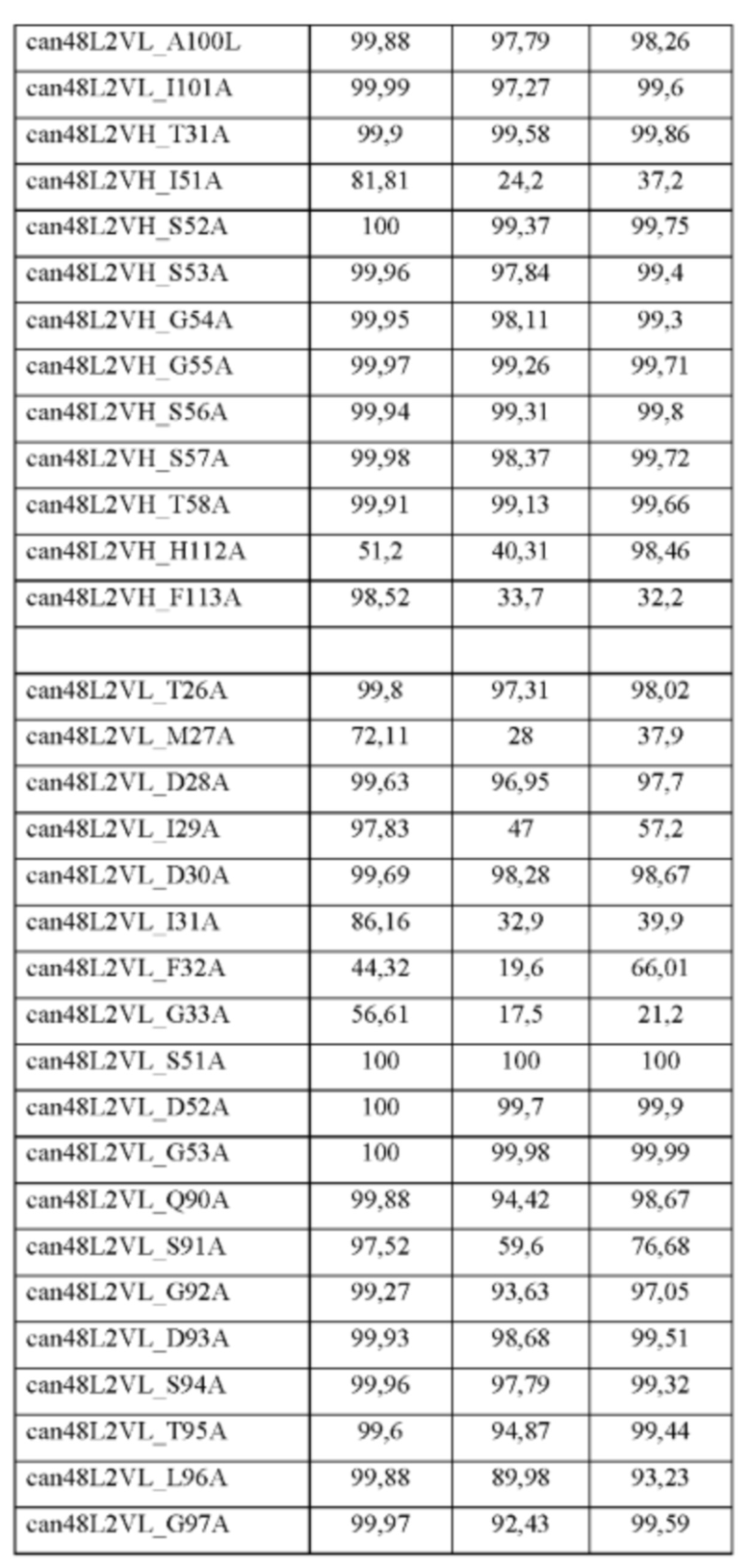
Значения, приведенные в Таблицах 4 и 5, имеющие процент сходства менее 50%, указывают на то, что аминокислотные положения необходимы для связывания паратопа антитела с NGF. Мутации аминокислот дикого типа в указанных положениях с их заменой на аланин, приводящие к снижению или полному отсутствию связывания с NGF, показывают, какие аминокислоты необходимы для связывания, а какие аминокислоты могут быть заменены, по меньшей мере посредством консервативных аминокислотных замен.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Zoetis Services LLC
Steiniger, Sebastian C.J.
Dunkle, William
Rugg, Catherine
Dunham, Steven A.
<120> ANTI-NGF ANTIBODIES AND METHODS THEREOF
<130> ZP000167
<140> 62/641,538
<141> 2018-03-12
<160> 92
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> PRT
<213> Canis familiaris
<400> 1
Thr Asn Asn Ile Gly Ile Leu Gly
1 5
<210> 2
<211> 3
<212> PRT
<213> Canis familiaris
<400> 2
Gly Asn Gly
1
<210> 3
<211> 11
<212> PRT
<213> Canis familiaris
<400> 3
Gln Ser Phe Asp Thr Thr Leu Gly Ala His Val
1 5 10
<210> 4
<211> 8
<212> PRT
<213> Canis familiaris
<400> 4
Gly Phe Thr Phe Ser Ser His Gly
1 5
<210> 5
<211> 8
<212> PRT
<213> Canis familiaris
<400> 5
Ile Asn Ser Gly Gly Ser Ser Thr
1 5
<210> 6
<211> 18
<212> PRT
<213> Canis familiaris
<400> 6
Ala Lys Glu Ser Val Gly Gly Trp Glu Gln Leu Val Gly Pro His Phe
1 5 10 15
Asp Tyr
<210> 7
<211> 110
<212> PRT
<213> Canis familiaris
<400> 7
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Gly Ala Ser Trp Tyr Gln Leu Phe Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Val Tyr Gly Asn Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ala Asp Ser Gly Asp Ser Val Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Gly Ala His Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 8
<211> 125
<212> PRT
<213> Canis familiaris
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Val Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Thr Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Lys Glu Ser Val Gly Gly Trp Glu Gln Leu Val Gly Pro His Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Ile Val Ser Ser
115 120 125
<210> 9
<211> 110
<212> PRT
<213> Canis familiaris
<400> 9
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Gly Ala Ser Trp Tyr Gln Leu Phe Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Val Tyr Gly Asn Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ala Asp Ser Gly Asp Ser Val Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Gly Ala His Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 10
<211> 125
<212> PRT
<213> Canis familiaris
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Val Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Thr Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Lys Glu Ser Val Gly Gly Trp Glu Gln Leu Val Gly Pro His Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Ile Val Ser Ser
115 120 125
<210> 11
<211> 24
<212> DNA
<213> Canis familiaris
<400> 11
acgaacaaca tcggtattct tggt 24
<210> 12
<211> 9
<212> DNA
<213> Canis familiaris
<400> 12
ggtaatggg 9
<210> 13
<211> 36
<212> DNA
<213> Canis familiaris
<400> 13
cagtcctttg ataccacgct tggtgctcat gtgttc 36
<210> 14
<211> 24
<212> DNA
<213> Canis familiaris
<400> 14
ggattcacct tcagtagcca cggc 24
<210> 15
<211> 24
<212> DNA
<213> Canis familiaris
<400> 15
attaacagcg gtggaagtag caca 24
<210> 16
<211> 54
<212> DNA
<213> Canis familiaris
<400> 16
gcaaaggagt ccgtcggggg gtgggagcaa ctggtcggac ctcattttga ctac 54
<210> 17
<211> 330
<212> DNA
<213> Canis familiaris
<400> 17
cagtctgtgc tgactcagcc gacctcagtg tcagggtccc ttggccagag ggtcaccatc 60
tcctgctctg gaagcacgaa caacatcggt attcttggtg cgagctggta ccaactgttc 120
ccaggaaagg cccctaaact cctcgtgtac ggtaatggga atcgaccgtc aggggtccct 180
gaccggtttt ccggcgccga ctctggcgac tcagtcaccc tgaccatcac tgggcttcag 240
gctgaggacg aggctgatta ttactgccag tcctttgata ccacgcttgg tgctcatgtg 300
ttcggcggag gcacccacct gaccgtcctt 330
<210> 18
<211> 375
<212> DNA
<213> Canis familiaris
<400> 18
gaggtgcagc tggtggagtc tgggggagat ttggtgaagc ctggggggtc cttgagactg 60
tcctgtgtgg cctctggatt caccttcagt agccacggca tgcactgggt ccgtcagtct 120
ccagggaagg gactgcagtg ggtcgcagtt attaacagcg gtggaagtag cacatactac 180
acagacgctg tgaagggccg attcaccatc tccagagaca acgccaagaa cacagtgtat 240
ctacagatga acagcctgag agccgaggac acggccatgt attactgtgc aaaggagtcc 300
gtcggggggt gggagcaact ggtcggacct cattttgact actggggcca gggaaccctg 360
gtcatcgtct cgagc 375
<210> 19
<211> 333
<212> DNA
<213> Canis familiaris
<400> 19
caggcggtgc tgaaccagcc ggcgagcgtg agcggcgcgc tgggccagaa agtgaccatt 60
agctgcagcg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagctg 120
ccgggcaaag cgccgaaact gctggtggat agcgatggcg atcgcccgag cggcattccg 180
gatcgcttta gcggcagccg cagcggcaac agcggcaccc tgaccattac cggcctgcag 240
gcggaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 20
<211> 375
<212> DNA
<213> Canis familiaris
<400> 20
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggctt tacctttagc agccatggca tgcattgggt gcgccagagc 120
ccgggcaaag gcctgcagtg ggtggcggtg attaacagcg gcggcagcag cacctattat 180
accgatgcgg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccgtgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcgatgt attattgcgc gaaagaaagc 300
gtgggcggct gggaacagct ggtgggcccg cattttgatt attggggcca gggcaccctg 360
gtgattgtct cgagc 375
<210> 21
<211> 8
<212> PRT
<213> Canis familiaris
<400> 21
Thr Met Asp Ile Asp Ile Phe Gly
1 5
<210> 22
<211> 3
<212> PRT
<213> Canis familiaris
<400> 22
Ser Asp Gly
1
<210> 23
<211> 12
<212> PRT
<213> Canis familiaris
<400> 23
Gln Ser Gly Asp Ser Thr Leu Gly Ala Leu Ala Ile
1 5 10
<210> 24
<211> 8
<212> PRT
<213> Canis familiaris
<400> 24
Gly Phe Thr Phe Ser Thr Tyr Gly
1 5
<210> 25
<211> 8
<212> PRT
<213> Canis familiaris
<400> 25
Ile Ser Ser Gly Gly Ser Ser Thr
1 5
<210> 26
<211> 17
<212> PRT
<213> Canis familiaris
<400> 26
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
1 5 10 15
Phe
<210> 27
<211> 111
<212> PRT
<213> Canis familiaris
<400> 27
Gln Ala Val Leu Asn Gln Pro Ala Ser Val Ser Gly Ala Leu Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Ser Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Val Asp Ser Asp Gly Asp Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Arg Ser Gly Asn Ser Gly Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 28
<211> 124
<212> PRT
<213> Canis familiaris
<400> 28
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Pro Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Met Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 29
<211> 110
<212> PRT
<213> Canis familiaris
<400> 29
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Gly Ala Ser Trp Tyr Gln Leu Phe Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Val Tyr Gly Asn Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ala Asp Ser Gly Asp Ser Val Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Gly Ala His Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 30
<211> 124
<212> PRT
<213> Canis familiaris
<400> 30
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Pro Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Met Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 31
<211> 24
<212> DNA
<213> Canis familiaris
<400> 31
acaatggaca ttgatatatt tggt 24
<210> 32
<211> 9
<212> DNA
<213> Canis familiaris
<400> 32
agtgatggg 9
<210> 33
<211> 36
<212> DNA
<213> Canis familiaris
<400> 33
cagtctggtg attccacgct tggtgccctt gctatt 36
<210> 34
<211> 24
<212> DNA
<213> Canis familiaris
<400> 34
ggattcacct tcagtaccta tggc 24
<210> 35
<211> 24
<212> DNA
<213> Canis familiaris
<400> 35
attagtagtg gtggaagtag caca 24
<210> 36
<211> 51
<212> DNA
<213> Canis familiaris
<400> 36
gcgggtagta gatatacata tgcatacgga ggaggatatg agtttcactt c 51
<210> 37
<211> 333
<212> DNA
<213> Canis familiaris
<400> 37
caggctgtgc tgaatcagcc ggcctcagtg tctggggccc tgggccagaa ggtcaccatc 60
tcctgctctg gaagcacaat ggacattgat atatttggtg tgagctggta ccaacagctc 120
ccaggaaagg cccctaaact cctcgtggac agtgatgggg atcgaccctc agggatccct 180
gacagatttt ctggctccag gtctggcaac tcaggcaccc tgaccatcac tgggctccag 240
gctgaggacg aggctgatta tcactgtcag tctggtgatt ccacgcttgg tgcccttgct 300
attttcggcg gaggcaccca tgtgaccgtc ctt 333
<210> 38
<211> 372
<212> DNA
<213> Canis familiaris
<400> 38
gaggtacaac tggtggaatc tgggggagac ctggtgaagc ctgggggatc cctgagactc 60
tcctgtgtgg cctctggatt caccttcagt acctatggca tcaactgggt ccgccaggct 120
ccagggaagg ggctgcagtg ggtcgcatac attagtagtg gtggaagtag cacatactat 180
gcagatcctg tgaagggccg gttcaccatc tccagagacg acgccaagaa catgctgtat 240
cttcagatga acagcctgag agccgaggac acggccatat attactgtgc gggtagtaga 300
tatacatatg catacggagg aggatatgag tttcacttct ggggccaggg aaccctggtc 360
accgtctcga gc 372
<210> 39
<211> 333
<212> DNA
<213> Canis familiaris
<400> 39
caggcggtgc tgaaccagcc ggcgagcgtg agcggcgcgc tgggccagaa agtgaccatt 60
agctgcagcg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagctg 120
ccgggcaaag cgccgaaact gctggtggat agcgatggcg atcgcccgag cggcattccg 180
gatcgcttta gcggcagccg cagcggcaac agcggcaccc tgaccattac cggcctgcag 240
gcggaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 40
<211> 372
<212> DNA
<213> Canis familiaris
<400> 40
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggctt tacctttagc acctatggca ttaactgggt gcgccaggcg 120
ccgggcaaag gcctgcagtg ggtggcgtat attagcagcg gcggcagcag cacctattat 180
gcggatccgg tgaaaggccg ctttaccatt agccgcgatg atgcgaaaaa catgctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcgattt attattgcgc gggcagccgc 300
tatacctatg cgtatggcgg cggctatgaa tttcattttt ggggccaggg caccctggtg 360
accgtctcga gc 372
<210> 41
<211> 335
<212> PRT
<213> Canis familiaris
<400> 41
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp Lys
85 90 95
Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys
100 105 110
Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg Thr Pro Glu
130 135 140
Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln
145 150 155 160
Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln
165 170 175
Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys
195 200 205
Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser
225 230 235 240
Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys
245 250 255
Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln
260 265 270
Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu
275 280 285
Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg
290 295 300
Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu
305 310 315 320
His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 42
<211> 1005
<212> DNA
<213> Canis familiaris
<400> 42
gcctcaacaa ctgctcctag cgtgtttccc ctggccccta gctgcggaag tacctcaggc 60
agcacagtgg ccctggcttg tctggtgtct ggatatttcc ctgagccagt gaccgtgagt 120
tggaacagcg gctctctgac ctccggggtg cacacatttc catctgtgct gcagtctagt 180
ggcctgtact ccctgtcaag catggtgact gtgccttcct ctaggtggcc atcagaaact 240
ttcacctgca acgtggccca tcccgccagc aagaccaaag tggacaagcc cgtgcctaaa 300
agggagaatg gaagggtgcc aagaccacct gattgcccta agtgtccagc tccagaaatg 360
ctgggaggac caagcgtgtt catctttcca cccaagccca aagacacact gctgattgct 420
agaactcccg aggtgacctg cgtggtggtg gacctggatc cagaggaccc cgaagtgcag 480
atctcctggt tcgtggatgg gaagcagatg cagacagcca aaactcagcc tcgggaggaa 540
cagtttaacg gaacctatag agtggtgtct gtgctgccaa ttggacacca ggactggctg 600
aagggcaaac agtttacatg caaggtgaac aacaaggccc tgcctagtcc aatcgagagg 660
actatttcaa aagctagggg acaggctcat cagccttccg tgtatgtgct gcctccatcc 720
cgggaggaac tgtctaagaa cacagtgagt ctgacttgtc tgatcaaaga tttctttccc 780
cctgacattg atgtggagtg gcagagcaat gggcagcagg agccagaatc caagtacaga 840
accacaccac cccagctgga cgaagatggc tcctatttcc tgtacagtaa gctgtcagtg 900
gacaaatcta ggtggcagcg cggggatacc tttatctgcg ccgtgatgca cgaggctctg 960
cacaatcatt acacacaaga aagtctgtca catagccccg gcaag 1005
<210> 43
<211> 335
<212> PRT
<213> Canis familiaris
<400> 43
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp Lys
85 90 95
Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys
100 105 110
Pro Lys Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg Thr Pro Glu
130 135 140
Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln
145 150 155 160
Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln
165 170 175
Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys
195 200 205
Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser
225 230 235 240
Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys
245 250 255
Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln
260 265 270
Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu
275 280 285
Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg
290 295 300
Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu
305 310 315 320
His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 44
<211> 1005
<212> DNA
<213> Canis familiaris
<400> 44
gcctcaacaa ctgctcctag cgtgtttccc ctggccccta gctgcggaag tacctcaggc 60
agcacagtgg ccctggcttg tctggtgtct ggatatttcc ctgagccagt gaccgtgagt 120
tggaacagcg gctctctgac ctccggggtg cacacatttc catctgtgct gcagtctagt 180
ggcctgtact ccctgtcaag catggtgact gtgccttcct ctaggtggcc atcagaaact 240
ttcacctgca acgtggccca tcccgccagc aagaccaaag tggacaagcc cgtgcctaaa 300
agggagaatg gaagggtgcc aagaccacct gattgcccta agtgtccagc tccagaagcg 360
gcgggagcac caagcgtgtt catctttcca cccaagccca aagacacact gctgattgct 420
agaactcccg aggtgacctg cgtggtggtg gacctggatc cagaggaccc cgaagtgcag 480
atctcctggt tcgtggatgg gaagcagatg cagacagcca aaactcagcc tcgggaggaa 540
cagtttaacg gaacctatag agtggtgtct gtgctgccaa ttggacacca ggactggctg 600
aagggcaaac agtttacatg caaggtgaac aacaaggccc tgcctagtcc aatcgagagg 660
actatttcaa aagctagggg acaggctcat cagccttccg tgtatgtgct gcctccatcc 720
cgggaggaac tgtctaagaa cacagtgagt ctgacttgtc tgatcaaaga tttctttccc 780
cctgacattg atgtggagtg gcagagcaat gggcagcagg agccagaatc caagtacaga 840
accacaccac cccagctgga cgaagatggc tcctatttcc tgtacagtaa gctgtcagtg 900
gacaaatcta ggtggcagcg cggggatacc tttatctgcg ccgtgatgca cgaggctctg 960
cacaatcatt acacacaaga aagtctgtca catagccccg gcaag 1005
<210> 45
<211> 6
<212> PRT
<213> Canis familiaris
<400> 45
Asn Ile Gly Ser Lys Asp
1 5
<210> 46
<211> 3
<212> PRT
<213> Canis familiaris
<400> 46
Ser Asp Ser
1
<210> 47
<211> 11
<212> PRT
<213> Canis familiaris
<400> 47
Gln Val Trp Asp Ile Ser Ala Asp Ala Ile Val
1 5 10
<210> 48
<211> 8
<212> PRT
<213> Canis familiaris
<400> 48
Gly Tyr Thr Phe Thr Asp Tyr Tyr
1 5
<210> 49
<211> 8
<212> PRT
<213> Canis familiaris
<400> 49
Ile Asp Pro Gly Asn Gly Ala Thr
1 5
<210> 50
<211> 13
<212> PRT
<213> Canis familiaris
<400> 50
Ala Pro Leu Gly Tyr Val Pro Ala Ser Thr Ser Glu Tyr
1 5 10
<210> 51
<211> 108
<212> PRT
<213> Canis familiaris
<400> 51
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Thr Val Thr Leu Arg Gln
1 5 10 15
Thr Ala His Ile Thr Cys Gly Gly Asp Asn Ile Gly Ser Lys Asp Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Ile Ile Tyr
35 40 45
Ser Asp Ser Lys Arg Pro Thr Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Met Ala Thr Leu Thr Ile Ser Gly Ala Leu Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ile Ser Ala Asp Ala
85 90 95
Ile Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 52
<211> 120
<212> PRT
<213> Canis familiaris
<400> 52
Glu Val Gln Leu Val Gln Ser Ala Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Ala Gly Leu Asn Trp Met
35 40 45
Gly Arg Ile Asp Pro Gly Asn Gly Ala Thr Arg Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Gly Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Leu Gly Tyr Val Pro Ala Ser Thr Ser Glu Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Ser Val Ser Ser
115 120
<210> 53
<211> 324
<212> DNA
<213> Canis familiaris
<400> 53
tcctatgtgc tgacccagcc accatcagtg actgtgaccc tgaggcagac ggcccacatc 60
acctgtgggg gagacaacat tggaagtaaa gatgtttatt ggtaccagca gaagccgggc 120
caggcccccg tgttgattat ctatagtgat agcaagaggc cgacagggat ccctgagcga 180
ttctccggct ccaactcggg gaacatggcc accctgacca tcagtggggc cttggcggag 240
gatgaggctg actattactg ccaggtatgg gacatcagtg ctgatgctat tgtgttcggc 300
ggaggcaccc atctgaccgt cctt 324
<210> 54
<211> 360
<212> DNA
<213> Canis familiaris
<400> 54
gaggtccagc tggtgcagtc tgcagctgag gttaagaagc caggggcatc tgtaaaggtc 60
tcctgcaaga cctctggata caccttcact gactactata tgcactgggt acaacaggct 120
ccaggagcag ggctcaattg gatgggacgg attgatcctg gaaatggtgc cacaaggtat 180
gcacagaagt tccagggcag actcaccctg acggcagaca catccacaag cacagcctac 240
atggagctga gcggtctgag agctgaggac acagctgtgt actactgtgc gcccctaggg 300
tacgtgcctg catcaacatc tgagtactgg ggccagggca ccctggtcag cgtctcgagc 360
<210> 55
<211> 111
<212> PRT
<213> Felis catus
<400> 55
Gln Ala Val Leu Asn Gln Pro Ser Ser Val Ser Gly Ala Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Ser Trp Tyr Gln Gln Ile Pro Gly Met Ala Pro Lys Thr Ile
35 40 45
Ile Asp Ser Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ser Thr Gly Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 56
<211> 124
<212> PRT
<213> Felis catus
<400> 56
Asp Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Thr Cys Val Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Pro Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met Leu Tyr
65 70 75 80
Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 57
<211> 333
<212> DNA
<213> Felis catus
<400> 57
caggcggtgc tgaaccagcc gagcagcgtg agcggcgcgc tgggccagcg cgtgaccatt 60
agctgcagcg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagatt 120
ccgggcatgg cgccgaaaac cattattgat agcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcagc accggcaccc tgaccattac cggcctgcag 240
gcggaagatg aagcggatta ttattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 58
<211> 372
<212> DNA
<213> Felis catus
<400> 58
gaggtacaac tggtggaatc tgggggagac ctggtgaagc ctgggggatc cctgagactc 60
tcctgtgtgg cctctggatt caccttcagt acctatggca tcaactgggt ccgccaggct 120
ccagggaagg ggctgcagtg ggtcgcatac attagtagtg gtggaagtag cacatactat 180
gcagatcctg tgaagggccg gttcaccatc tccagagacg acgccaagaa catgctgtat 240
cttcagatga acagcctgag agccgaggac acggccatat attactgtgc gggtagtaga 300
tatacatatg catacggagg aggatatgag tttcacttct ggggccaggg aaccctggtc 360
accgtctcga gc 372
<210> 59
<211> 201
<212> PRT
<213> Canis familiaris
<400> 59
Gln His Ser Leu Asp Thr Ala Leu Arg Arg Ala Arg Ser Ala Pro Ala
1 5 10 15
Gly Ala Ile Ala Ala Arg Val Thr Gly Gln Thr Arg Asn Ile Thr Val
20 25 30
Asp Pro Lys Leu Phe Lys Lys Arg Arg Leu Arg Ser Pro Arg Val Leu
35 40 45
Phe Ser Thr His Pro Pro Pro Val Ala Ala Asp Ala Gln Asp Leu Asp
50 55 60
Leu Glu Ala Gly Ser Thr Ala Ser Val Asn Arg Thr His Arg Ser Lys
65 70 75 80
Arg Ser Ser Pro His Pro Val Phe His Arg Gly Glu Phe Ser Val Cys
85 90 95
Asp Ser Val Ser Val Trp Val Gly Asp Lys Thr Thr Ala Thr Asp Ile
100 105 110
Lys Gly Lys Glu Val Met Val Leu Gly Glu Val Asn Ile Asn Asn Ser
115 120 125
Val Phe Lys Gln Tyr Phe Phe Glu Thr Lys Cys Arg Asp Pro Thr Pro
130 135 140
Val Asp Ser Gly Cys Arg Gly Ile Asp Ser Lys His Trp Asn Ser Tyr
145 150 155 160
Cys Thr Thr Thr His Thr Phe Val Lys Ala Leu Thr Met Asp Gly Lys
165 170 175
Gln Ala Ala Trp Arg Phe Ile Arg Ile Asp Thr Ala Cys Val Cys Val
180 185 190
Leu Ser Arg Lys Ala Gly Arg Arg Ala
195 200
<210> 60
<211> 106
<212> PRT
<213> Canis familiaris
<400> 60
Gly Gln Pro Lys Ala Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gly Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Ser Gly Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Thr Gln Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Asp Lys Trp Lys
65 70 75 80
Ser His Ser Ser Phe Ser Cys Leu Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Lys Val Ala Pro Ala Glu Cys Ser
100 105
<210> 61
<211> 318
<212> DNA
<213> Canis familiaris
<400> 61
ggacaaccga aggcctcccc ctcggtcaca ctcttcccgc cctcctctga ggagctcggc 60
gccaacaagg ccaccctggt gtgcctcatc agcgacttct accccagcgg cgtgacggtg 120
gcctggaagg cagacggcag ccccgtcacc cagggcgtgg agaccaccaa gccctccaag 180
cagagcaaca acaagtacgc ggccagcagc tacctgagcc tgacgcctga caagtggaaa 240
tctcacagca gcttcagctg cctggtcacg cacgagggga gcaccgtgga gaagaaggtg 300
gcccccgcag agtgctct 318
<210> 62
<211> 335
<212> PRT
<213> Felis catus
<400> 62
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Thr Thr Ser Gly Ala Thr Val Ala Leu Ala Cys Leu Val Leu Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ala Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Leu Ser Asp Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Arg Lys Thr Asp His Pro Pro Gly Pro Lys Pro Cys Asp Cys
100 105 110
Pro Lys Cys Pro Pro Pro Glu Met Leu Gly Gly Pro Ser Ile Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Ser Ile Ser Arg Thr Pro Glu
130 135 140
Val Thr Cys Leu Val Val Asp Leu Gly Pro Asp Asp Ser Asp Val Gln
145 150 155 160
Ile Thr Trp Phe Val Asp Asn Thr Gln Val Tyr Thr Ala Lys Thr Ser
165 170 175
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Leu His Gln Asp Trp Leu Lys Gly Lys Glu Phe Lys Cys Lys
195 200 205
Val Asn Ser Lys Ser Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Lys Gly Gln Pro His Glu Pro Gln Val Tyr Val Leu Pro Pro Ala
225 230 235 240
Gln Glu Glu Leu Ser Arg Asn Lys Val Ser Val Thr Cys Leu Ile Lys
245 250 255
Ser Phe His Pro Pro Asp Ile Ala Val Glu Trp Glu Ile Thr Gly Gln
260 265 270
Pro Glu Pro Glu Asn Asn Tyr Arg Thr Thr Pro Pro Gln Leu Asp Ser
275 280 285
Asp Gly Thr Tyr Phe Val Tyr Ser Lys Leu Ser Val Asp Arg Ser His
290 295 300
Trp Gln Arg Gly Asn Thr Tyr Thr Cys Ser Val Ser His Glu Ala Leu
305 310 315 320
His Ser His His Thr Gln Lys Ser Leu Thr Gln Ser Pro Gly Lys
325 330 335
<210> 63
<211> 1005
<212> DNA
<213> Felis catus
<400> 63
gcctccacca cggccccatc ggtgttccca ctggccccca gctgcgggac cacatctggc 60
gccaccgtgg ccctggcctg cctggtgtta ggctacttcc ctgagccggt gaccgtgtcc 120
tggaactccg gcgccctgac cagcggtgtg cacaccttcc cggccgtcct gcaggcctcg 180
gggctgtact ctctcagcag catggtgaca gtgccctcca gcaggtggct cagtgacacc 240
ttcacctgca acgtggccca cccgcccagc aacaccaagg tggacaagac cgtgcgcaaa 300
acagaccacc caccgggacc caaaccctgc gactgtccca aatgcccacc ccctgagatg 360
cttggaggac cgtccatctt catcttcccc ccaaaaccca aggacaccct ctcgatttcc 420
cggacgcccg aggtcacatg cttggtggtg gacttgggcc cagatgactc cgatgtccag 480
atcacatggt ttgtggataa cacccaggtg tacacagcca agacgagtcc gcgtgaggag 540
cagttcaaca gcacctaccg tgtggtcagt gtcctcccca tcctacacca ggactggctc 600
aaggggaagg agttcaagtg caaggtcaac agcaaatccc tcccctcccc catcgagagg 660
accatctcca aggccaaagg acagccccac gagccccagg tgtacgtcct gcctccagcc 720
caggaggagc tcagcaggaa caaagtcagt gtgacctgcc tgatcaaatc cttccacccg 780
cctgacattg ccgtcgagtg ggagatcacc ggacagccgg agccagagaa caactaccgg 840
acgaccccgc cccagctgga cagcgacggg acctacttcg tgtacagcaa gctctcggtg 900
gacaggtccc actggcagag gggaaacacc tacacctgct cggtgtcaca cgaagctctg 960
cacagccacc acacacagaa atccctcacc cagtctccgg gtaaa 1005
<210> 64
<211> 106
<212> PRT
<213> Felis catus
<400> 64
Gly Gln Pro Lys Ser Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Asn
1 5 10 15
Glu Glu Leu Ser Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Ser Gly Leu Thr Val Ala Trp Lys Ala Asp Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Ser Pro Asn Glu Trp Lys
65 70 75 80
Ser Arg Ser Arg Phe Thr Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Asn Val Val Pro Ala Glu Cys Ser
100 105
<210> 65
<211> 318
<212> DNA
<213> Felis catus
<400> 65
ggccagccca agagcgctcc ctccgtgacc ctgttccccc caagcaacga ggaactgagc 60
gccaacaagg ccaccctggt gtgcctgatc agcgacttct accccagcgg cctgaccgtg 120
gcctggaagg ccgatggcac ccctatcacc cagggcgtgg aaaccaccaa gcccagcaag 180
cagagcaaca acaaatacgc cgccagcagc tacctgagcc tgagccccaa cgagtggaag 240
tcccggtccc ggttcacatg ccaggtgaca cacgagggca gcaccgtgga aaagaacgtg 300
gtgcccgccg agtgcagc 318
<210> 66
<211> 240
<212> PRT
<213> Homo sapiens
<400> 66
Met Ser Met Leu Phe Tyr Thr Leu Ile Thr Ala Phe Leu Ile Gly Ile
1 5 10 15
Gln Ala Glu Pro His Ser Glu Ser Asn Val Pro Ala Gly His Thr Ile
20 25 30
Pro Gln Ala His Trp Thr Lys Leu Gln His Ser Leu Asp Thr Ala Leu
35 40 45
Arg Arg Ala Arg Ser Ala Pro Ala Ala Ala Ile Ala Ala Arg Val Ala
50 55 60
Gly Gln Thr Arg Asn Ile Thr Val Asp Pro Arg Leu Phe Lys Lys Arg
65 70 75 80
Arg Leu Arg Ser Pro Arg Val Leu Phe Ser Thr Gln Pro Pro Arg Glu
85 90 95
Ala Ala Asp Thr Gln Asp Leu Asp Phe Glu Val Gly Gly Ala Ala Pro
100 105 110
Phe Asn Arg Thr His Arg Ser Lys Arg Ser Ser Ser His Pro Ile Phe
115 120 125
His Arg Gly Glu Phe Ser Val Cys Asp Ser Val Ser Val Trp Val Gly
130 135 140
Asp Lys Thr Thr Ala Thr Asp Ile Lys Gly Lys Glu Val Met Val Leu
145 150 155 160
Gly Glu Val Ser Ile Asn Asn Ser Val Phe Lys Gln Tyr Phe Phe Glu
165 170 175
Thr Lys Cys Arg Asp Pro Asn Pro Val Asp Ser Gly Cys Arg Gly Ile
180 185 190
Asp Ser Lys His Trp Asn Ser Cys Thr Thr Thr His Thr Phe Val Lys
195 200 205
Ala Leu Thr Met Asp Gly Lys Gln Ala Ala Trp Arg Phe Ile Arg Ile
210 215 220
Asp Thr Ala Cys Met Cys Val Leu Ser Arg Lys Ala Val Arg Arg Ala
225 230 235 240
<210> 67
<211> 125
<212> PRT
<213> Homo sapiens
<400> 67
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Ser Val Gly Gly Trp Glu Gln Leu Val Gly Pro His Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Ile Val Ser Ser
115 120 125
<210> 68
<211> 375
<212> DNA
<213> Homo sapiens
<400> 68
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc agccatggca tgagctgggt gcgccaggcg 120
ccgggcaaag gcctggaatg ggtgagcgtg attaacagcg gcggcagcag cacctattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acagcaaaaa caccctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gaaagaaagc 300
gtgggcggct gggaacagct ggtgggcccg cattttgatt attggggcca gggcaccctg 360
gtgattgtct cgagc 375
<210> 69
<211> 125
<212> PRT
<213> Homo sapiens
<400> 69
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Ser Val Gly Gly Trp Glu Gln Leu Val Gly Pro His Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Ile Val Ser Ser
115 120 125
<210> 70
<211> 375
<212> DNA
<213> Homo sapiens
<400> 70
caggtgcagc tggtggaaag cggcggcggc gtggtgcagc cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc agccatggca tgcattgggt gcgccaggcg 120
ccgggcaaag gcctggaatg ggtgagcgtg attaacagcg gcggcagcag cacctattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acagcaaaaa caccctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gaaagaaagc 300
gtgggcggct gggaacagct ggtgggcccg cattttgatt attggggcca gggcaccctg 360
gtgattgtct cgagc 375
<210> 71
<211> 110
<212> PRT
<213> Homo sapiens
<400> 71
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Gly Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Gly Ala His Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 72
<211> 330
<212> DNA
<213> Homo sapiens
<400> 72
cagagcgtgc tgacccagcc gccgagcgtg agcggcgcgc cgggccagcg cgtgaccatt 60
agctgcaccg gcagcaccaa caacattggc attctgggcg tgcattggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat ggcaacggca accgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcacc agcgcgagcc tggcgattac cggcctgcag 240
gcggaagatg aagcggatta ttattgccag agctttgata ccaccctggg cgcgcatgtg 300
tttggcggcg gcacccatct gaccgtgctg 330
<210> 73
<211> 110
<212> PRT
<213> Homo sapiens
<400> 73
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Gly Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ala Asp Ser Gly Asp Ser Val Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Gly Ala His Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 74
<211> 330
<212> DNA
<213> Homo sapiens
<400> 74
cagagcgtgc tgacccagcc gaccagcgtg agcggcgcgc cgggccagcg cgtgaccatt 60
agctgcaccg gcagcaccaa caacattggc attctgggcg tgcattggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat ggcaacggca accgcccgag cggcgtgccg 180
gatcgcttta gcggcgcgga tagcggcgat agcgtgagcc tggcgattac cggcctgcag 240
gcggaagatg aagcggatta ttattgccag agctttgata ccaccctggg cgcgcatgtg 300
tttggcggcg gcacccatct gaccgtgctg 330
<210> 75
<211> 124
<212> PRT
<213> Homo sapiens
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Ser Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 76
<211> 372
<212> DNA
<213> Homo sapiens
<400> 76
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc acctatggca tgaactgggt gcgccaggcg 120
ccgggcaaag gcctggaatg ggtgagctat attagcagcg gcggcagcag catttattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gggcagccgc 300
tatacctatg cgtatggcgg cggctatgaa tttcattttt ggggccaggg caccctggtg 360
attgtctcga gc 372
<210> 77
<211> 124
<212> PRT
<213> Homo sapiens
<400> 77
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Ser Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 78
<211> 372
<212> DNA
<213> Homo sapiens
<400> 78
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc acctatggca tgaactgggt gcgccaggcg 120
ccgggcaaag gcctgcagtg ggtgagctat attagcagcg gcggcagcag catttattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gggcagccgc 300
tatacctatg cgtatggcgg cggctatgaa tttcattttt ggggccaggg caccctggtg 360
attgtctcga gc 372
<210> 79
<211> 124
<212> PRT
<213> Homo sapiens
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 80
<211> 372
<212> DNA
<213> Homo sapiens
<400> 80
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc acctatggca ttaactgggt gcgccaggcg 120
ccgggcaaag gcctggaatg ggtgagctat attagcagcg gcggcagcag cacctattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gggcagccgc 300
tatacctatg cgtatggcgg cggctatgaa tttcattttt ggggccaggg caccctggtg 360
attgtctcga gc 372
<210> 81
<211> 124
<212> PRT
<213> Homo sapiens
<400> 81
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Tyr Thr Tyr Ala Tyr Gly Gly Gly Tyr Glu Phe His
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 82
<211> 372
<212> DNA
<213> Homo sapiens
<400> 82
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcgg cgagcggctt tacctttagc acctatggca ttaactgggt gcgccaggcg 120
ccgggcaaag gcctgcagtg ggtgagctat attagcagcg gcggcagcag cacctattat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat 240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gggcagccgc 300
tatacctatg cgtatggcgg cggctatgaa tttcattttt ggggccaggg caccctggtg 360
attgtctcga gc 372
<210> 83
<211> 110
<212> PRT
<213> Homo sapiens
<400> 83
Gln Ser Val Leu Thr Gln Pro Ala Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val
100 105 110
<210> 84
<211> 333
<212> DNA
<213> Homo sapiens
<400> 84
cagagcgtgc tgacccagcc ggcgagcgcg agcggcaccc cgggccagcg cgtgaccatt 60
agctgcagcg gcagcaccat ggatattgat atttttggcg tgaactggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat agcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcacc agcgcgagcc tggcgattag cggcctgcag 240
agcgaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 85
<211> 111
<212> PRT
<213> Homo sapiens
<400> 85
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 86
<211> 333
<212> DNA
<213> Homo sapiens
<400> 86
cagagcgtgc tgacccagcc gccgagcgcg agcggcaccc tgggccagcg cgtgaccatt 60
agctgcagcg gcagcaccat ggatattgat atttttggcg tgaactggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat agcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcacc agcgcgagcc tggcgattag cggcctgcag 240
agcgaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 87
<211> 111
<212> PRT
<213> Homo sapiens
<400> 87
Gln Ser Val Val Thr Gln Pro Ala Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ala Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 88
<211> 333
<212> DNA
<213> Homo sapiens
<400> 88
cagagcgtgg tgacccagcc ggcgagcgtg agcggcgcgc cgggccagcg cgtgaccatt 60
agctgcaccg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat ggcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcgcg agcgcgagcc tggcgattac cggcctgcag 240
gcggaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 89
<211> 111
<212> PRT
<213> Homo sapiens
<400> 89
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Gly Ala Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ala Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 90
<211> 333
<212> DNA
<213> Homo sapiens
<400> 90
cagagcgtgg tgacccagcc gccgagcgtg agcggcgcgc tgggccagcg cgtgaccatt 60
agctgcaccg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat ggcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcgcg agcgcgagcc tggcgattac cggcctgcag 240
gcggaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<210> 91
<211> 111
<212> PRT
<213> Canis familiaris
<400> 91
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Thr Met Asp Ile Asp Ile Phe
20 25 30
Gly Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asp Gly Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ala Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr His Cys Gln Ser Gly Asp Ser Thr Leu
85 90 95
Gly Ala Leu Ala Ile Phe Gly Gly Gly Thr His Val Thr Val Leu
100 105 110
<210> 92
<211> 333
<212> DNA
<213> Canis familiaris
<400> 92
cagagcgtgg tgacccagcc gccgagcgtg agcggcgcgc cgggccagcg cgtgaccatt 60
agctgcaccg gcagcaccat ggatattgat atttttggcg tgagctggta tcagcagctg 120
ccgggcaccg cgccgaaact gctgatttat ggcgatggcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcgcg agcgcgagcc tggcgattac cggcctgcag 240
gcggaagatg aagcggatta tcattgccag agcggcgata gcaccctggg cgcgctggcg 300
atttttggcg gcggcaccca tgtgaccgtg ctg 333
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ИНТЕРЛЕЙКИНУ-31 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2019 |
|
RU2795411C2 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
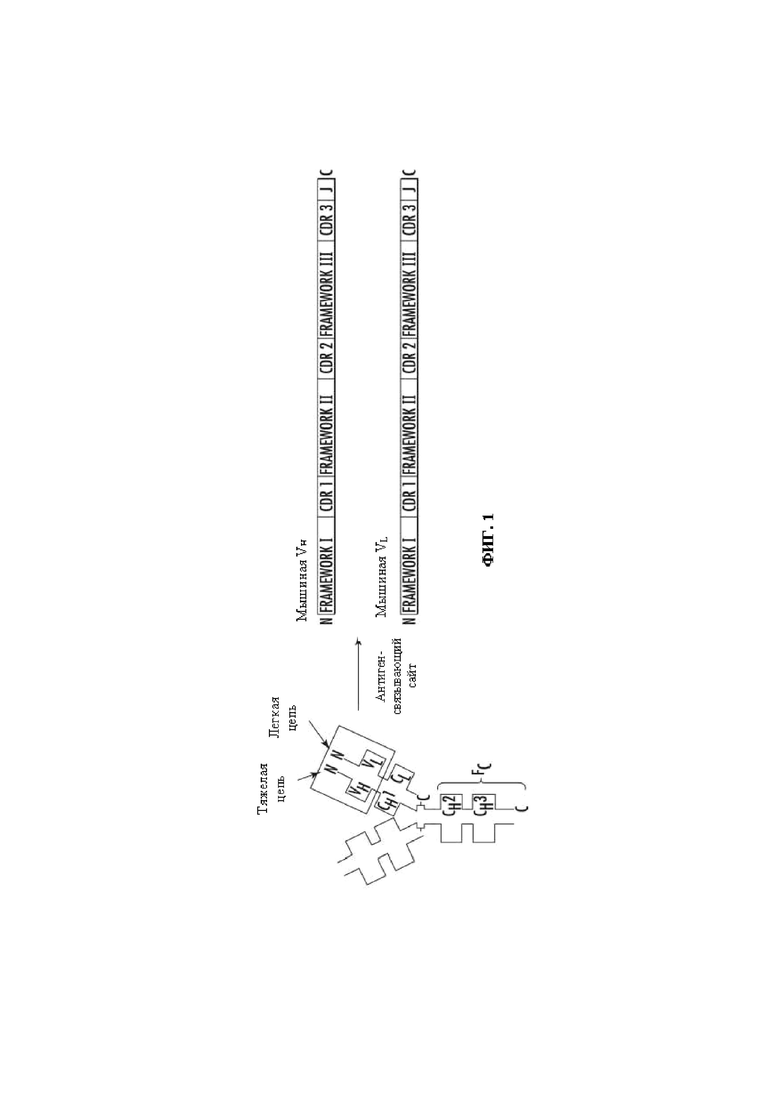
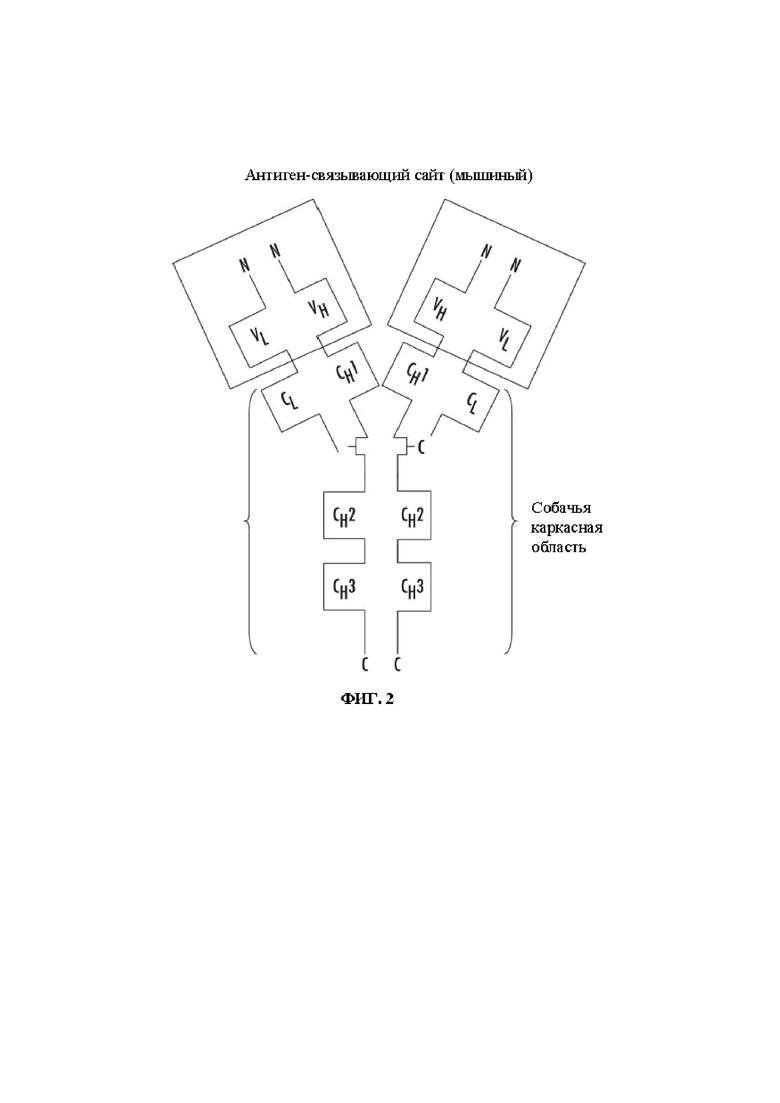


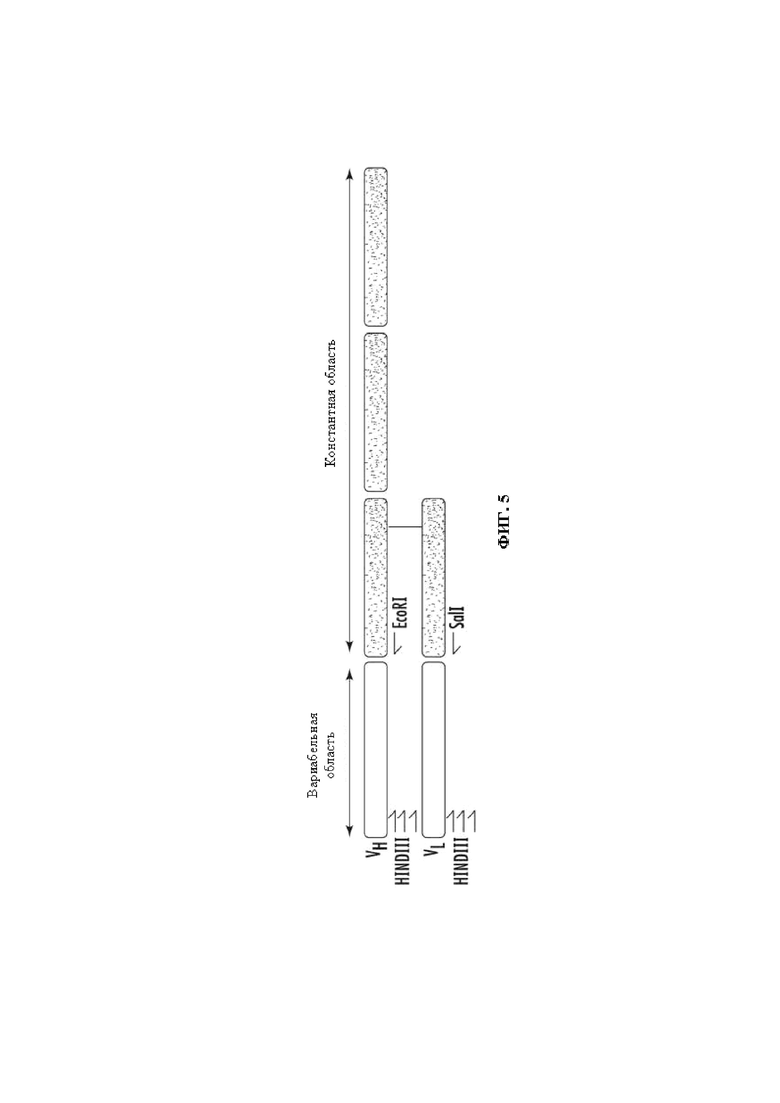
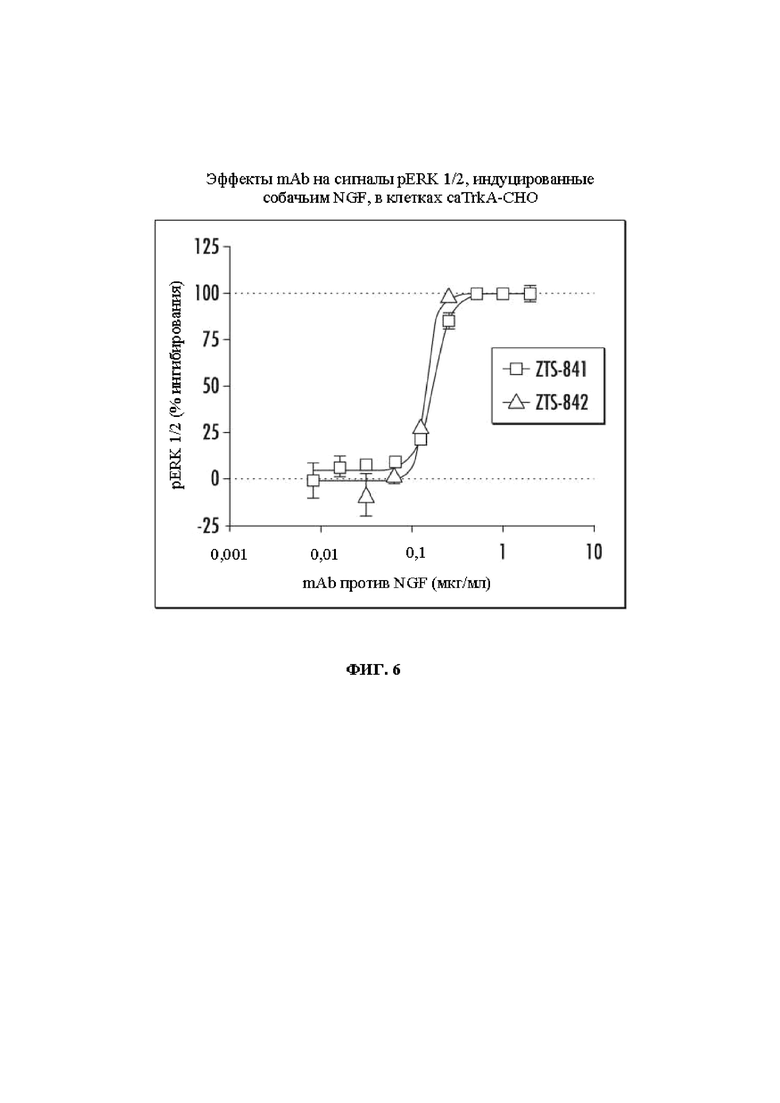
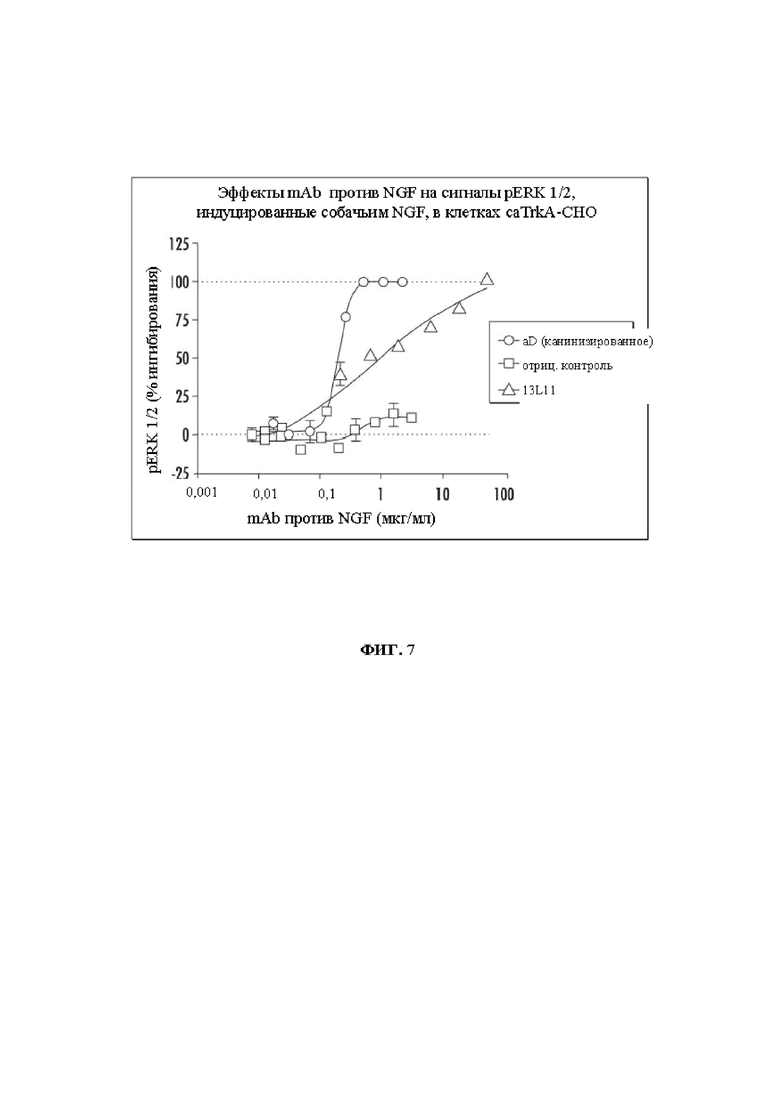
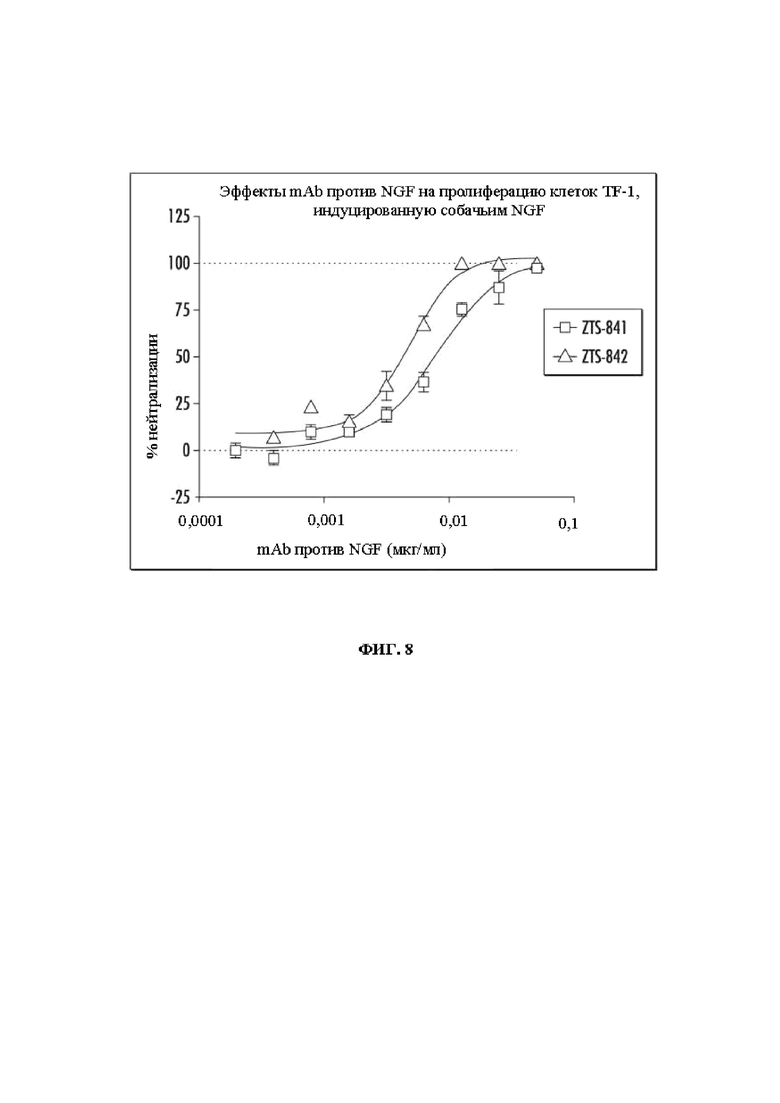
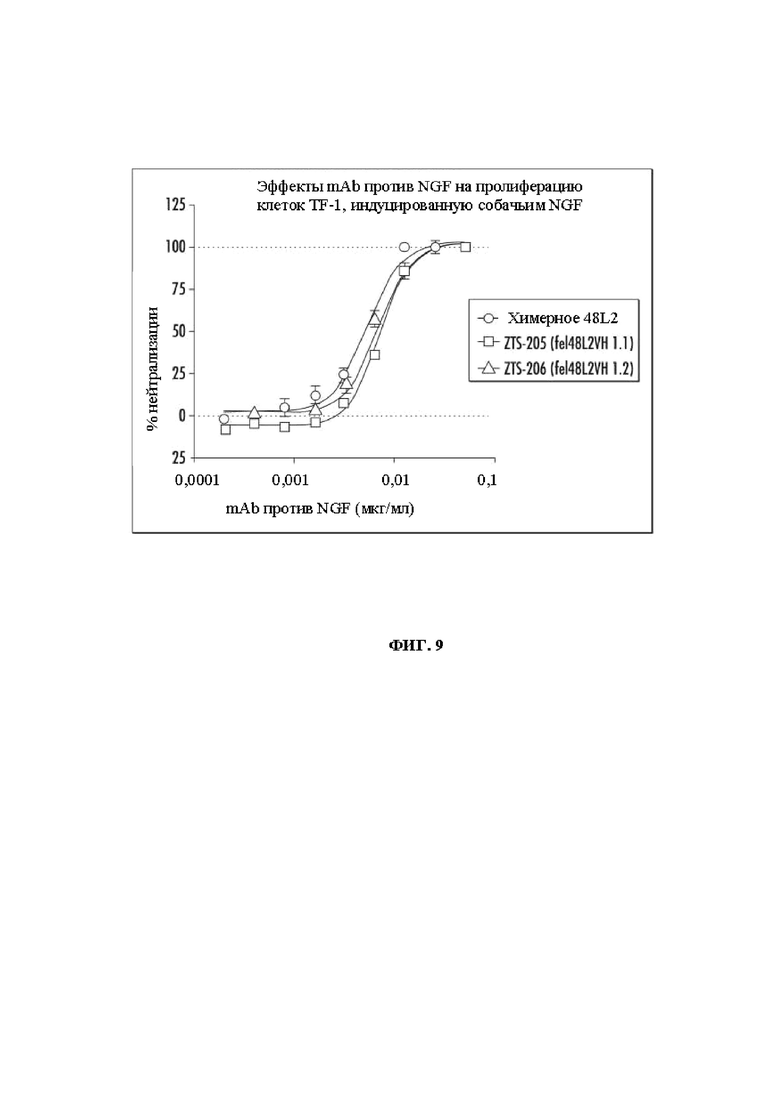
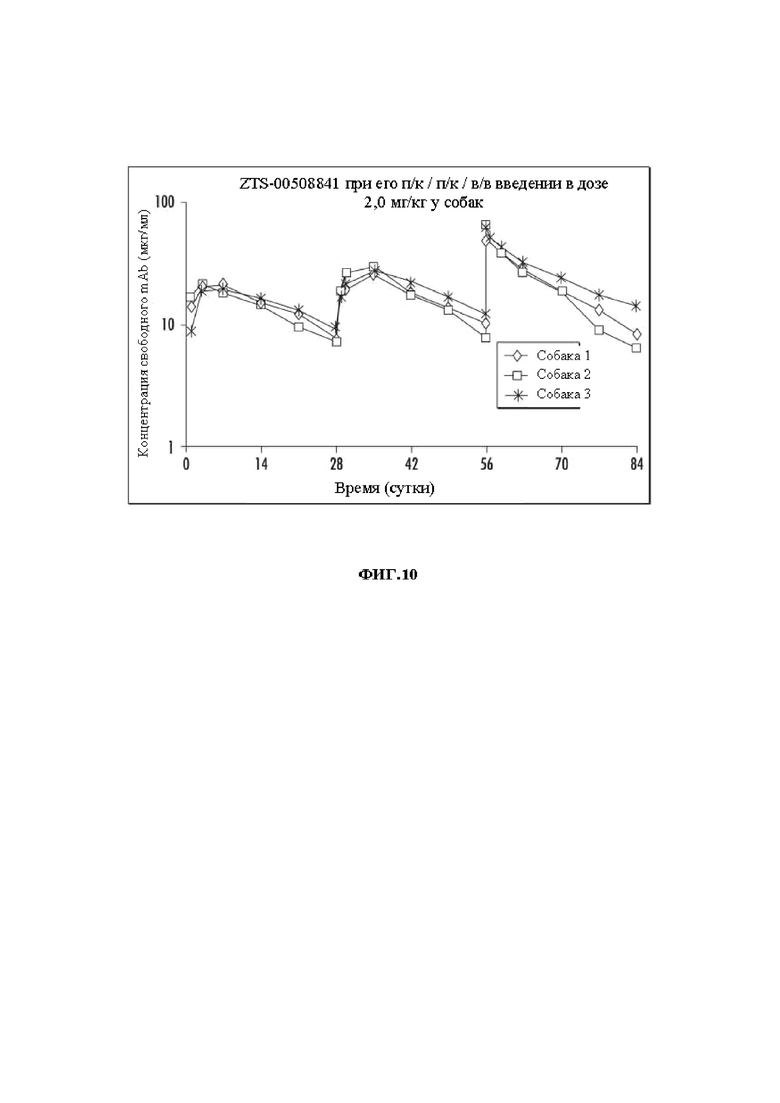
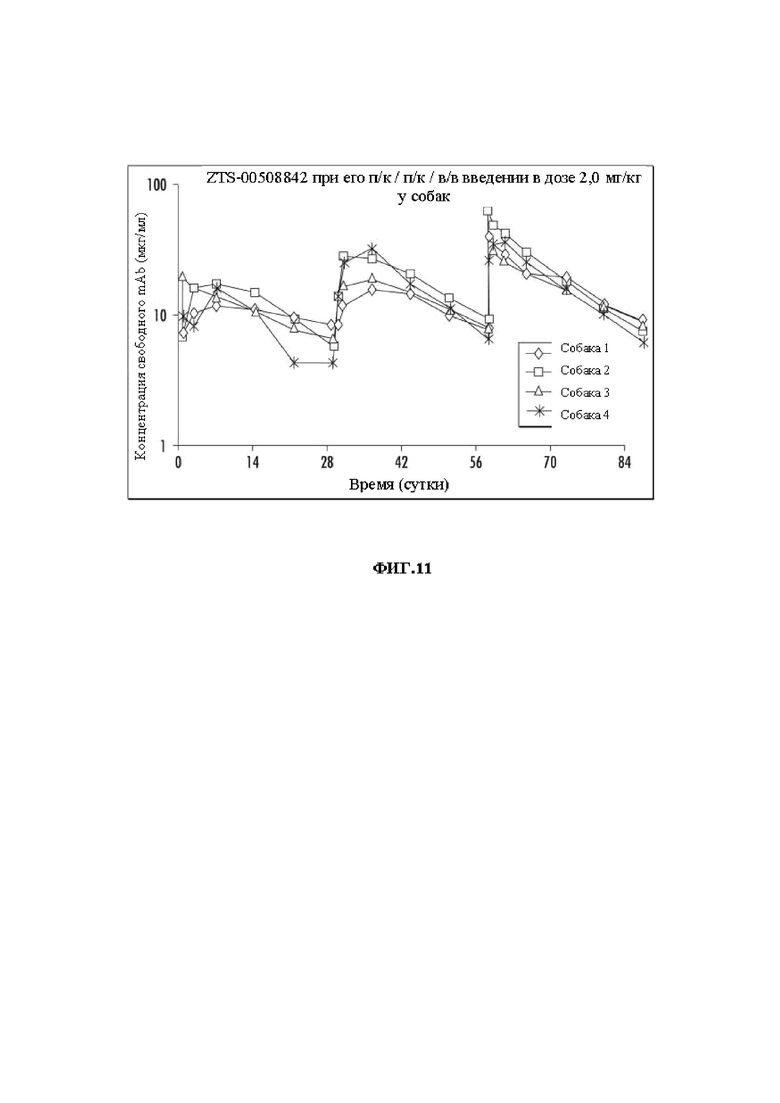

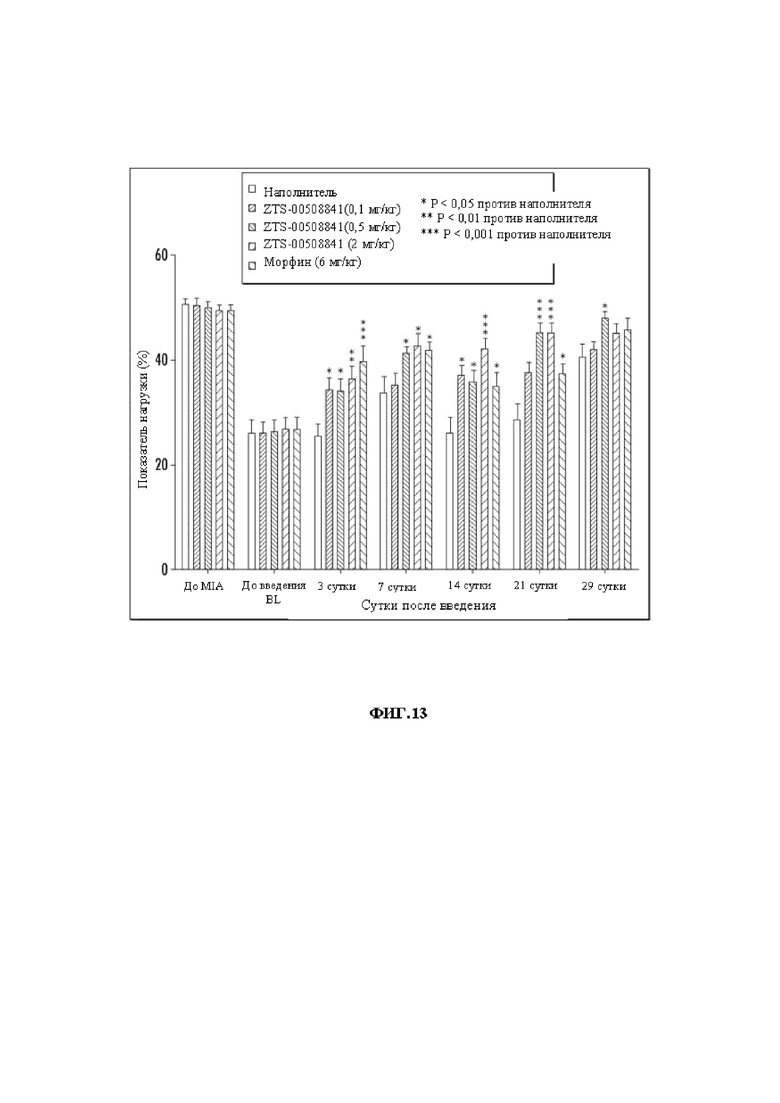
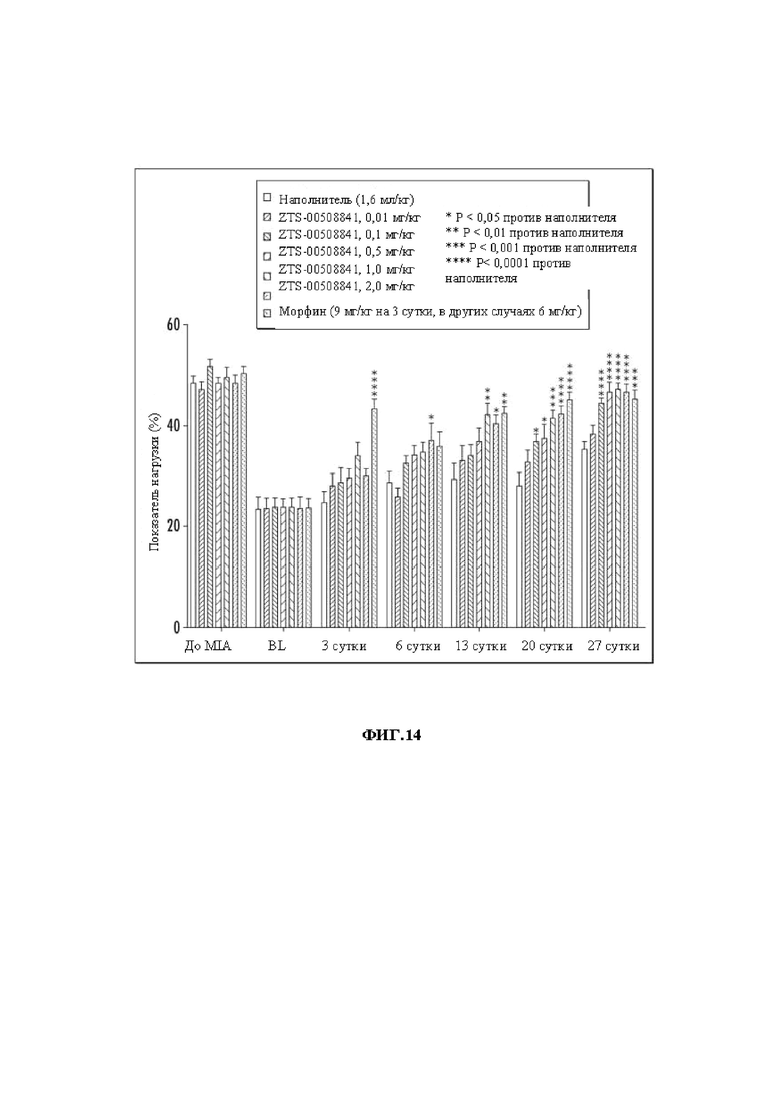
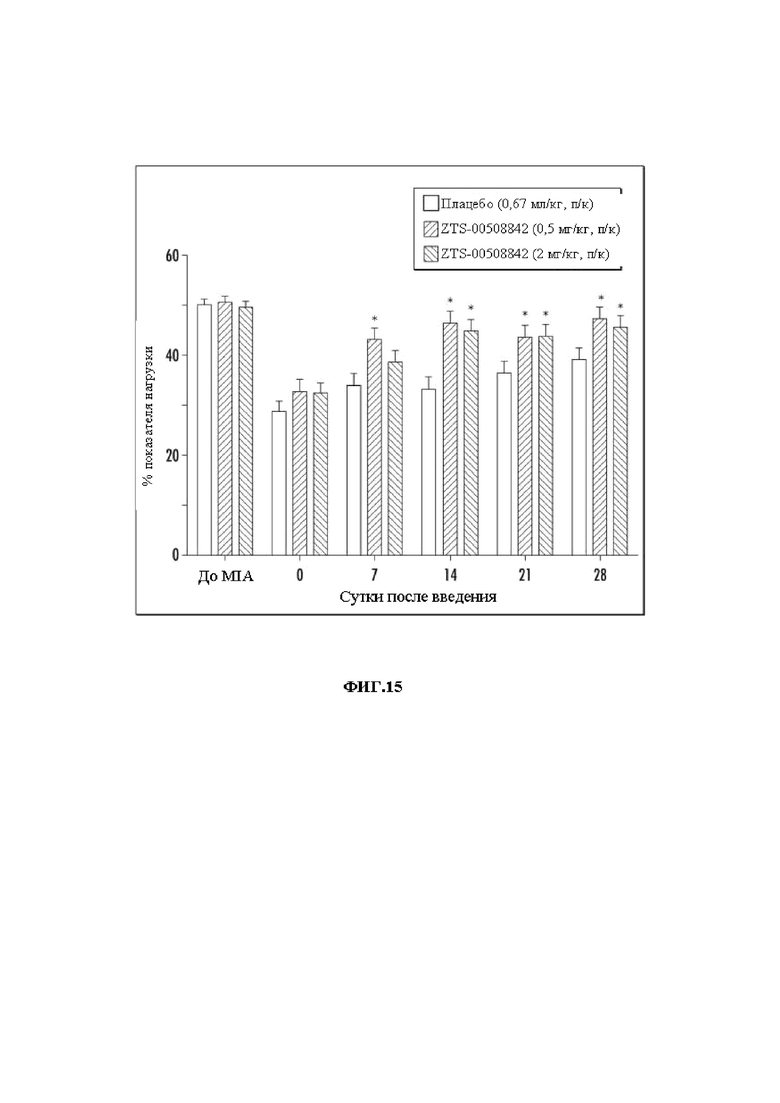
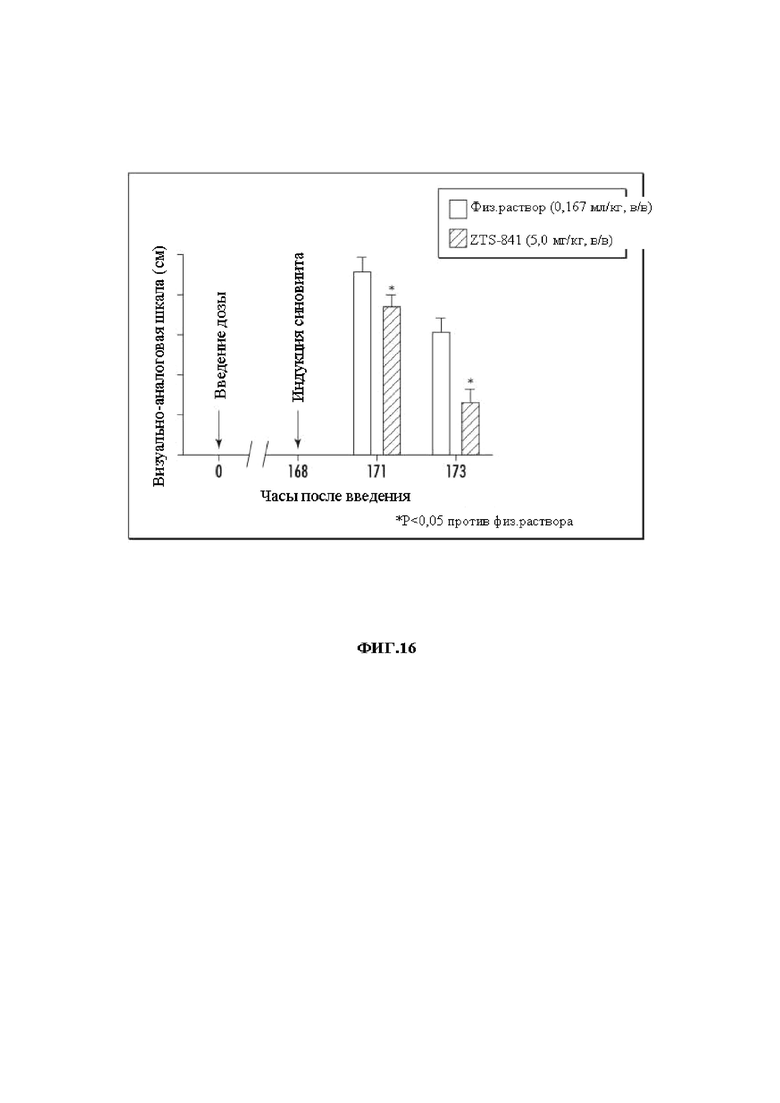
Группа изобретений относится к биотехнологии. Представлены: рекомбинантные антигенсвязывающие белки, специфично связывающиеся с фактором роста нервов (NGF), способ их получения, нуклеиновая кислота, кодирующая указанные белки, ветеринарная композиция для лечения заболеваний, связанных с повышенными уровнями NGF, вектор экспрессии, содержащий выделенную нуклеиновую кислоту. Также раскрыты клетка-хозяин, содержащая вектор, для продуцирования антигенсвязывающего белка и клетка-хозяин, содержащая нуклеиновую кислоту, для продуцирования антигенсвязывающего белка, способ лечения у субъекта расстройства, связанного с повышенными уровнями NGF, и способ ингибирования активности NGF у субъекта, включающий введение указанному субъекту фармацевтической композиции. Изобретение может использоваться для лечения расстройства, связанного с повышенными уровнями NGF. 10 н. и 27 з.п. ф-лы, 16 ил., 5 табл., 14 пр.
1. Рекомбинантный антигенсвязывающий белок, специфично связывающийся с фактором роста нервов (NGF), содержащий:
a) вариабельную область легкой цепи (VL), содержащую:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 1;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 2;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 3; и
б) вариабельную область тяжелой цепи (VH), содержащую:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 4;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 5; и
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 6; или
a) вариабельную область легкой цепи (VL), содержащую:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 21;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 22;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 23; и
б) вариабельную область тяжелой цепи (VH), содержащую:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 24;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 25; и
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 26;
и любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
2. Антигенсвязывающий белок по п. 1, где:
a) вариабельная область легкой цепи (VL) содержит:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 1;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 2;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 3; и
б) вариабельная область тяжелой цепи (VH) содержит:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 4;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 5;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 6; и
любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
3. Антигенсвязывающий белок по п. 1, где:
в) вариабельная область легкой цепи (VL) содержит:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 21;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 22;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 23; и
г) вариабельная область тяжелой цепи (VH) содержит:
1) участок 1, определяющий комплементарность (CDR1), содержащий аминокислотную последовательность SEQ ID NO: 24;
2) участок 2, определяющий комплементарность (CDR2), содержащий аминокислотную последовательность SEQ ID NO: 25;
3) участок 3, определяющий комплементарность (CDR3), содержащий аминокислотную последовательность SEQ ID NO: 26; и
любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
4. Рекомбинантный антигенсвязывающий белок, специфично связывающийся с фактором роста нервов (NGF), содержащий:
a) вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 7, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 8;
б) вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 9, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 10;
в) вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 27, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 28; или
г) вариабельную область легкой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 29, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO: 30; и
любые его варианты, имеющие одну или более чем одну консервативную аминокислотную замену в любой из вариабельных областей легкой или тяжелой цепи указанного антигенсвязывающего белка.
5. Антигенсвязывающий белок по любому из пп. 1-4, ингибирующий связывание NGF с рецептором TrkA.
6. Антигенсвязывающий белок по любому из пп. 1-5, уменьшающий или устраняющий расстройство, связанное с повышенными уровнями NGF.
7. Антигенсвязывающий белок по п. 6, где расстройство, связанное с повышенными уровнями NGF, выбрано из группы, состоящей из сердечно-сосудистых заболеваний, атеросклероза, ожирения, диабета 2-го типа, метаболического синдрома, боли и воспаления.
8. Антигенсвязывающий белок по п. 7, где расстройство, связанное с повышенными уровнями NGF, представляет собой боль.
9. Антигенсвязывающий белок по п. 8, где указанное расстройство, связанное с повышенными уровнями NGF, представляет собой болевое расстройство и выбрано из группы, состоящей из боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли.
10. Антигенсвязывающий белок по п. 9, где расстройство, связанное с повышенными уровнями NGF, включает боль при остеоартрите.
11. Антигенсвязывающий белок по любому из пп. 1-10, содержащий моноклональное антитело.
12. Антигенсвязывающий белок по п. 11, где указанное моноклональное антитело представляет собой собачье моноклональное антитело, канинизированное моноклональное антитело, кошачье моноклональное антитело, фелинизированное моноклональное антитело, человеческое моноклональное антитело, гуманизированное моноклональное антитело.
13. Ветеринарная композиция для лечения заболеваний, связанных с повышенными уровнями NGF, содержащая терапевтически эффективное количество антигенсвязывающего белка по любому из пп. 1-12 и фармацевтически приемлемый носитель.
14. Ветеринарная композиция по п. 13 для применения в лечении боли у субъекта.
15. Ветеринарная композиция по п. 14, где боль выбрана из группы, состоящей из боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли.
16. Ветеринарная композиция по п. 15, где боль включает боль при остеоартрите.
17. Ветеринарная композиция по п. 15, где боль включает операционную и послеоперационную боль.
18. Ветеринарная композиция по п. 15, где боль включает раковую боль.
19. Ветеринарная композиция по любому из пп. 13-18 для применения у собак или кошек.
20. Ветеринарная композиция по п. 19 для применения у собак.
21. Ветеринарная композиция по п. 19 для применения у кошек.
22. Выделенная нуклеиновая кислота, кодирующая антигенсвязывающий белок по любому из пп. 1-12, для лечения заболеваний, связанных с повышенными уровнями NGF.
23. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 22.
24. Клетка-хозяин, содержащая вектор по п. 23, для продуцирования антигенсвязывающего белка по любому из пп. 1-12.
25. Клетка-хозяин, содержащая нуклеиновую кислоту по п. 22, для продуцирования антигенсвязывающего белка по любому из пп. 1-12.
26. Способ получения антигенсвязывающего белка по любому из пп. 1-12, включающий культивирование клетки-хозяина по п. 24 или 25 в условиях, приводящих к продуцированию антигенсвязывающего белка, и выделение антигенсвязывающего белка из клетки-хозяина или культуральной среды клетки-хозяина.
27. Способ лечения у субъекта расстройства, связанного с повышенными уровнями NGF, включающий введение указанному субъекту терапевтически эффективного количества фармацевтической композиции по п. 13.
28. Способ по п. 27, где расстройство, связанное с повышенными уровнями NGF, выбрано из группы, состоящей из сердечно-сосудистых заболеваний, атеросклероза, ожирения, диабета 2-го типа, метаболического синдрома, боли и воспаления.
29. Способ по п. 28, где расстройство, связанное с повышенными уровнями NGF, включает боль.
30. Способ по п. 29, где указанное расстройство, связанное с повышенными уровнями NGF, представляет собой болевое расстройство и выбрано из группы, состоящей из боли при остеоартрите, боли при ревматоидном артрите, операционной и послеоперационной боли, инцизионной боли, общевоспалительной боли, раковой боли, боли в результате травмы, нейропатической боли, невралгии, боли при диабетической нейропатии, боли, связанной с ревматическими заболеваниями, боли, связанной с заболеваниями костно-мышечной системы, висцеральной боли и желудочно-кишечной боли.
31. Способ по п. 30, где расстройство, связанное с повышенными уровнями NGF, включает боль при остеоартрите.
32. Способ по п. 30, где расстройство, связанное с повышенными уровнями NGF, включает операционную и послеоперационную боль.
33. Способ по п. 30, где расстройство, связанное с повышенными уровнями NGF, включает раковую боль.
34. Способ ингибирования активности NGF у субъекта, включающий введение указанному субъекту фармацевтической композиции по п. 13.
35. Способ по любому из пп. 27-34, где субъект выбран из группы, состоящей из собак или кошек.
36. Способ по п. 35, где субъект включает собак.
37. Способ по п. 35, где субъект включает кошек.
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640254C2 |
| SKAPER S.D | |||
| Nerve growth factor: a neuroimmune crosstalk mediator for all seasons | |||
| Immunology | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Двухколейная подвесная дорога | 1919 |
|
SU151A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| См | |||
| реферат, стр | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И СОДЕРЖАЩИХ ЕГО КОМПОЗИЦИЙ | 2006 |
|
RU2429013C2 |

