РОДСТВЕННЫЕ ЗАЯВКИ
В данной заявке заявлен приоритет предварительной заявки США с серийным №61/934828, поданной 2 февраля 2014 года. Все принципы вышеупомянутой заявки включены в данное описание посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит Перечень последовательностей, который представлен в электронном ASCII-формате и тем самым включен посредством ссылки во всей своей полноте. Указанный документ в формате ASCII, созданный 29 января 2015 года, имеет название 110421-0054-WO1_SL.txt, и его размер составляет 134721 байт.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Боль является одним из наиболее распространенных симптомов, по поводу которого обращаются за медицинской помощью и который составляет основную жалобу половины всех пациентов, посещающих врача. Несмотря на наличие и широкое использование многочисленных обезболивающих средств, успех в отношении устранения боли, особенно хронической боли, не достигнут. Вследствие этого, бремя затрат, которые несет общество, остается на высоком уровне. По оценкам, полученным в различных исследованиях, боль является причиной ежегодной потери 50 миллионов рабочих дней и 61,2 миллиарда долларов в результате потери производительности. Что касается пациентов с хронической болью, то только примерно половина из них в состоянии справляться с болью путем применения предписанных имеющихся вариантов лечения. При этом общий оборот рынка отпускаемых по рецепту обезболивающих средств составляет приблизительно 25 миллиард долларов в год. Эти данные дают основание считать, что сохраняется большая потребность в безопасных и эффективных новых болеутоляющих средствах.
Терапевтические агенты, которые снижают уровни в тканях или ингибируют эффекты секретируемого фактора роста нервов (NGF или бета-NGF), могут стать как раз такими новыми болеутоляющими средствами. Хорошо известно, что NGF играет ключевую роль в развитии нервной системы; однако, NGF также представляет собой признанную мишень для устранения боли, поскольку он обуславливает боль у животных и людей. У взрослых NGF, в частности, стимулирует жизнеспособность и выживаемость субпопуляции центральных и периферических нейронов (Huang & Reichardt, Ann. Rev. Neurosci., 24: 677-736 (2001)). NGF также вносит вклад в модулирование функциональных характеристик этих нейронов и осуществляет тонический контроль чувствительности или возбудимости сенсорных болевых рецепторов, называемых ноцицепторами (Priestley et al., Can. J. Physiol. Pharmacol., 80: 495-505 (2002); Bennett, Neuroscientist, 7: 13-17 (2001)). Ноцицепторы воспринимают и передают в центральную нервную систему различные болевые стимулы, которые обуславливают восприятие боли (ноцицепции). Рецепторы NGF локализованы на ноцицепторах. Экспрессия NGF усиливается в поврежденной и воспаленной ткани и активируется при болевых состояниях у человека. Таким образом, благодаря участию NGF в ноцицепции, NGF-связывающие агенты, которые снижают уровни NGF, находят применение в качестве болеутоляющих терапевтически средств.
Подкожные инъекции самого NGF вызывают болевые ощущения у людей и животных. Инъецированный NGF приводит к развитию быстрой тепловой гипералгезии, затем замедленной тепловой гипералгезии и механической аллодинии (Petty et al., Ann. Neurol., 36: 244-46 (1994); McArthur et al., Neurology, 54: 1080-88 (2000)). Аналогичным образом, эндогенно секретированный NGF является проноцицептивным. Индуцированное повреждением ткани высвобождение NGF и его последующее действие на периферии играет важную роль в индуцировании тепловой гипералгезии благодаря процессу «периферической сенситизации» (Mendell & Arvanian, Brain Res. Rev., 40: 230-39 (2002)). Повреждение ткани стимулирует высвобождение проноцицептивных и провоспалительных цитокинов, которые в свою очередь индуцируют высвобождение NGF из кератиноцитов и фибробластов. Этот высвободившийся NGF действует непосредственно на ноцицепторы, индуцируя возникновение болевых или ноцицептивных состояний в пределах нескольких минут от начала болевого воздействия. При этом NGF также действует опосредованно, индуцируя и поддерживая ноцицептивные/болевые состояния при упреждающем высвобождении. Он запускает процесс дегрануляции тучных клеток с высвобождением проноцицептивных агентов, таких как гистамин и серотонин, и, что важно, еще большего количества NGF, и также может стимулировать окончания симпатических нервов с высвобождением проноцицептивных нейромедиаторов, таких как норадреналин (Ma & Woolf, Neuroreport, 8: 807-10 (1997)).
Уровни NGF в тканях повышены у животных, инъецированных полным адъювантом Фрейнда (CFA) и каррагинаном (Ma & Woolf, Neuroreport., 8: 807-10 (1997); Amann & Schuligoi, Neurosci. Lett., 278: 173-78 (2000)). NGF потенцирует ответ на капсаицин в дорсальных корешковых ганглиях (DRG) у крысы. Повышенные уровни NGF документально зафиксированы у пациентов, страдающих ревматоидным артритом (Aloe & Tuveri, Clin. Exp. Rheumatol. 15: 433-38 (1997)) или циститом (Lowe et al., Br. J. Urol. 79: 572-77 (1997)). Повреждение периферических нервов у грызунов усиливает экспрессию матричной РНК (мРНК) NGF в макрофагах, фибробластах и шванновских клетках (Heumann et al., J. Cell Biol. 104: 1623-31 (1987)). Сверхэкспрессия NGF у трансгенных мышей приводит к большему усилению характерного для нейропатической боли поведения после повреждения нерва, чем у мышей дикого типа (Ramer et al., Pain, Supp. 6: S111-20 (1998)). В течение промежутка времени от нескольких часов до 15 суток повышенные уровни NGF участвуют в стимулировании «центральной сенсибилизации» - усиления нейротрансмиссии в синапсах ноцицептивных путей спинного мозга. Центральная сенсибилизация приводит к развитию устойчивой и хронической гипералгезии и аллодинии. Считается, что в этот процесс вовлечена интернализация комплексов NGF и его высокоаффинного рецептора, рецепторной тирозинкиназы A (trkA). Ретроградный транспорт этих комплексов к телам ноцицептивных клеток в DRG усиливает секрецию ноцицептивных нейропептидов, например, субстанции Р или генетически родственного кальцитонину пептида (CGRP), активацию протеинкиназы С (PKC) и активацию N-метил-D-аспартатных (NMDA) рецепторов в заднем роге спинного мозга (Sah et al., Nat. Rev. Drug Disc., 2: 460-72 (2003)) - все процессы, которые стимулируют сенситизацию ноцицептивных путей. NGF также играет роль в активации и перераспределении потенциал-зависимых и лиганд-зависимых ионных каналов, включая натриевые каналы разных подтипов и рецептор капсаицина, катионный канал транзиторного рецепторного потенциала, член 1 подсемейства V (TRPV1) (Mamet et al., J. Biol. Chem., 278: 48907-13 (1999); Fjell et al., J. Neurosci. Res., 57: 39-47 (1999); Priestley et al., Can. J. Physiol. Pharmacol., 80: 495-505 (2002)). Изменение активностей и/или экспрессии трансмиттеров, рецепторов и ионных каналов лежит в основе повышения чувствительности и возбудимости ноцицепторов, ассоциированных с состояниями нейропатической боли.
NGF-индуцированная ноцицепция/боль опосредована высокоаффинным рецептором NGF, рецепторной тирозинкиназой A (trkA) (Sah, et al., Nat. Rev. Drug Disc. 2: 460-72 (2003)). Примерно 40-45% тел ноцицептивных клеток в DRG экспрессируют trkA. Они представляют собой тела клеток волокон небольшого диаметра или С-волокон, которые также экспрессируют секретируемые проноцицептивные пептиды, субстанцию Р и CGRP. Эти волокна заканчиваются в пластинках I и II заднего рога, откуда они передают в центральную нервную систему болевые стимулы, воспринимаемые периферическими ноцицепторами. Мутации или делеции в гене trkA приводят к получению фенотипа, характеризующегося потерей болевых ощущений как у людей (Indo, Clin. Auton. Res., 12 (Supp 1): I20-I32 (2002)), так и у trkA-нокаутированных мышей (de Castro et al., Eur. J. Neurosci., 10: 146-52 (1998)). Важно отметить, что экспрессия trkA активирована у животных, подвергнутых испытаниям в моделях артритической боли (Pozza et al., J. Rheumatol. 27: 1121-27 (2000)) или боли, обусловленной воспалением мочевого пузыря (Qiao & Vizzard, J. Comp. Neurol., 454: 200-11 (2002)), либо воспалительной боли, индуцированной инъекцией CFA или каррагинана в лапу (Cho et al., Brain Res., 716: 197-201 (1996)).
NGF также связывается с рецептором нейротрофинов р75 (p75NTR). Задача p75NTR зависит от его клеточного окружения и присутствия других рецепторов, вместе с которыми он, как полагают, осуществляет функцию вспомогательного рецептора или корецептора. В результате взаимодействия trkA с p75NTR образуются высокоаффинные сайты связывания с NGF. Важность взаимодействий таких рецепторов в NGF-опосредуемой передаче болевых сигналов не выяснена, однако согласно недавним исследованиям отмечено участие p75NTR в клеточных процессах, которые могут иметь к этому отношение (Zhang & Nicol, Neurosci. Lett., 366: 187-92 (2004)). Тем не менее, пока p75NTR-нокаутные мыши обнаруживают повышенные пороги к болевым стимулам, они сохраняют восприимчивость к гипералгезическим эффектам NGF, и это позволяет предположить, что для опосредования этих эффектов достаточно участия только рецепторов trkA (Bergmann et al., Neurosci. Lett., 255: 87-90 (1998)).
Блокирование NGF приводит к существенному повышению эффективности по сравнению с NSAID (нестероидные противовоспалительные лекарственные средства) при хронической ноцицептивной боли, например, при остеоартрите (OA) и при хронической боли в пояснице. Ряд терапевтических антител-кандидатов, направленно воздействующих на NGF, находятся на различных стадиях доклинической и клинической разработки. Такие антитела включают, например, танезумаб (PF-4383119; Pfizer), представляющий собой гуманизированное антитело в формате IgG2; SAR164877/REGN475 (Sanofi-Aventis/Regeneron Pharmaceuticals), представляющий собой человеческое антитело в формате IgG4; AMG 403 (Amgen/Johnson & Johnson), представляющий собой человеческое антитело в формате IgG2; PG110 (PanGenetics/Abbott), представляющий собой гуманизированное антитело в формате IgG4. Другое терапевтическое антитело-кандидат описано в заявке WO 2006/077441, касающейся антител к NGF и способов лечения заболеваний или расстройств, при которых взаимодействие NGF с описанными антителами является важным. MEDI-578 представляет собой человеческое антитело в формате IgG4. Несмотря на разработку этих кандидатов, сохраняется потребность в обеспечении аналгезирующего эффекта при болевых состояниях широкого диапазона посредством применения NGF-связывающего агента, обладающего усиленной эффективностью и улучшенным профилем безопасности.
Фактор некроза опухоли-альфа (TNFα), также называемый кахектином, представляет собой плейотропный цитокин с большим разнообразием биологических активностей, затрагивающих цитотоксичность, пролиферацию клеток иммунной системы, воспаление, онкогенез и вирусную репликацию. Kim et al., J. Mol. Biol., 374, 1374 (2007). Изначально TNFα продуцируется в виде трансмембранного белка (tmTNFα), который затем расщепляется под действием металлопротеиназы до растворимой формы (sTNFα). Wallis, Lancet Infect. Dis., 8(10): 601 (2008). TNFα (приблизительно 17 кДа) существует в виде жесткой гомотримерной молекулы, которая связывается с расположенными на клеточной поверхности рецептором 1 TNF или рецептором 2 TNF, индуцируя олигомеризацию рецепторов и передачу сигнала.
Известно, что воспалительные цитокины, и в частности TNFα, играют важную роль в возникновении гипералгезии. Leung, L., and Cahill, C.M., J. Neuroinflammation, 7: 27 (2010). С учетом некоторых предварительных данных показано, что ингибиторы TNFα могут быть полезны для контроля нейропатической боли. См., например, Sommer C. et al., J. Pehpher. Nerv. Syst., 6: 67-72 (2001); Cohen et al., A&A, Feb 2013, 116, 2, 455-462; Genevay et al., Ann. Rheum. Dis., 2004, 63, 1120-1123. Результаты клинических исследований по тестированию ингибиторов TNFα в качестве монотерапии для лечения нейропатической боли остаются неубедительными. См. Leung и Cahill (2010).
Несмотря на разработку кандидатов с направленным воздействием на NGF и направленным воздействием на TNFα для лечения боли, сохраняется потребность в обеспечении аналгезирующего эффекта при различных болевых состояниях посредством применения агентов, обладающих более высокой эффективностью, чем современный стандарт лечения. Согласно данному изобретению предложены способы комбинированного лечения, направленно воздействующего как на NGF, так и на TNFα, которые могут повысить эффективность и обладают потенциалом в отношении уменьшения как числа, так и частоты введений для испытывающих боль пациентов.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно данному изобретению предложены способы контролирования боли у субъекта, включающие введение субъекту, нуждающемуся в этом, эффективного количества антагониста фактора роста нервов (NGF) и антагониста фактора некроза опухоли-альфа (TNFα) или связывающей молекулы, содержащей домен антагониста NGF и домен антагониста TNFα. В некоторых воплощениях такое введение контролирует боль у субъекта эффективнее, чем введение эквивалентного количества антагониста NGF или антагониста TNFα по отдельности. В некоторых воплощениях способ включает совместное введение антагониста TNFα и антагониста NGF. В некоторых воплощениях антагонист TNFα и антагонист NGF вводят последовательно или одновременно.
В некоторых воплощениях данный способ является достаточным для предотвращения, ослабления, уменьшения интенсивности или устранения боли у субъекта. В некоторых воплощениях боль представляет собой острую боль, кратковременную боль, постоянную или хроническую ноцицептивную боль либо постоянную или хроническую нейропатическую боль. В некоторых воплощениях данный способ по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% более эффективен в отношении контролирования боли у субъекта, чем введение эквивалентного количества антагониста NGF или антагониста TNFα по отдельности.
В некоторых воплощениях часть связывающей молекулы, относящаяся к антагонисту TNFα, связывается с полипептидом, содержащим аминокислотную последовательность SEQ ID NO: 2. В некоторых воплощениях антагонист NGF связывается с полипептидом, содержащим аминокислотную последовательность SEQ ID NO: 1.
В некоторых воплощениях часть связывающей молекулы, относящаяся к антагонисту NGF, представляет собой антитело к NGF или его антиген-связывающий фрагмент. В некоторых воплощениях антитело к NGF или его фрагмент может ингибировать связывание NGF с TrkA, p75NTR или как с TrkA, так и с p75NTR. В некоторых воплощениях антитело к NGF предпочтительно блокирует связывание NGF с TrkA по сравнению со связыванием NGF с p75NTR. В некоторых воплощениях антитело к NGF или его фрагмент связывается с NGF человека с аффинностью примерно 0,25-0,44 нМ. В некоторых воплощениях антитело к NGF или его фрагмент связывается с тем же самым эпитопом, что и MEDI-578. В некоторых воплощениях антитело к NGF или его фрагмент конкурентно ингибирует связывание MEDI-578 с NGF человека.
В некоторых воплощениях антитело к NGF или его фрагмент содержит вариабельный домен тяжелой цепи (VH-домен) антитела, содержащий совокупность гипервариабельных участков (CDR) тяжелой цепи HCDR1, HCDR2, HCDR3, и вариабельный домен легкой цепи (VL-домен) антитела, содержащий совокупность CDR легкой цепи LCDR1, LCDR2 и LCDR3, при этом HCDR1 имеет аминокислотную последовательность SEQ ID NO: 4 или SEQ ID NO: 4 с аминокислотными заменами числом до двух включительно, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 5 с аминокислотными заменами числом до двух включительно, HCDR3 имеет аминокислотную последовательность SEQ ID NO: 6, SEQ ID NO: 6 с аминокислотными заменами числом до двух включительно, SSRIYDFNSALISYYDMDV (SEQ ID NO: 11) или SSRIYDMISSLQPYYDMDV (SEQ ID NO: 12), LCDR1 имеет аминокислотную последовательность SEQ ID NO: 8 или SEQ ID NO: 8 с аминокислотными заменами числом до двух включительно, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 9 или SEQ ID NO: 9 с аминокислотными заменами числом до двух включительно, и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 10 или SEQ ID NO: 10 с аминокислотными заменами числом до двух включительно. В некоторых воплощениях антитело к NGF или его фрагмент содержат VH-домен, имеющий аминокислотную последовательность SEQ ID NO: 3. В некоторых воплощениях антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность SEQ ID NO: 7. В некоторых воплощениях антитело к NGF или его фрагмент содержит VH-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 3. В некоторых воплощениях антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 7. В некоторых воплощениях антитело к NGF или его фрагмент содержит VH-домен, имеющий аминокислотную последовательность SEQ ID NO: 94. В некоторых воплощениях антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность SEQ ID NO: 95. В некоторых воплощениях антитело к NGF или его фрагмент содержит VH-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 94. В некоторых воплощениях антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 95.
В некоторых воплощениях антитело к NGF или его фрагмент представляет собой полноразмерное антитело в формате H2L2, Fab-фрагмент, Fab'-фрагмент, F(ab)2-фрагмент или одноцепочечный Fv (scFv)-фрагмент. В некоторых воплощениях антитело к NGF или его фрагмент является гуманизированным, химерным, приматизированным или полностью человеческим. В некоторых воплощениях антагонист NGF представляет собой scFv-фрагмент антитела к NGF. В некоторых воплощениях scFv является SS-стабилизированным. В некоторых воплощениях scFv-фрагмент антитела к NGF содержит, в направлении от N-конца к С-концу, VH-домен, имеющий аминокислотную последовательность SEQ ID NO: 3, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15), и VL-домен, имеющий аминокислотную последовательность SEQ ID NO: 7. В некоторых воплощениях scFv-фрагмент антитела к NGF содержит, в направлении от N-конца к С-концу, VH-домен, имеющий аминокислотную последовательность SEQ ID NO: 94, линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4 (SEQ ID NO: 19), и VL-домен, имеющий аминокислотную последовательность SEQ ID NO: 95.
Согласно определенным аспектам, способ включает введение антагониста TNFα, который ингибирует связывание TNFα с рецептором TNF (TNFR), блокируя тем самым активность TNFα. В некоторых воплощениях антагонист TNFα содержит антитело к TNFα или его антиген-связывающий фрагмент. В некоторых воплощениях антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий набор CDR HCDR1, HCDR2 и HCDR3, и VL-домен антитела, содержащий набор CDR LCDR1, LCDR2 и LCDR3, где указанные CDR идентичны HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 инфликсимаба или HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 адалимумаба.
В некоторых воплощениях связывающая молекула содержит полноразмерное антитело к TNFα и scFv-фрагмент антитела к NGF, слитый с С-концом тяжелой цепи антитела к TNFα. Такая связывающая молекула может содержать легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 20, и тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 22.
В некоторых воплощениях антагонист TNFα содержит растворимый TNFα-связывающий фрагмент TNFR. В некоторых воплощениях TNFR представляет собой TNFR-2 или его растворимый фрагмент. В других воплощениях TNFR представляет собой TNFR-1 или его растворимый фрагмент. В некоторых воплощениях растворимый фрагмент TNFR-1 представляет собой фрагмент с молекулярной массой 55 кДа. В других воплощениях растворимый фрагмент TNFR-2 представляет собой фрагмент с молекулярной массой 75 кДа. В некоторых воплощениях фрагмент TNFR слит с Fc-доменом иммуноглобулина. В некоторых воплощениях Fc-домен иммуноглобулина представляет собой Fc-домен IgG1 человека. В некоторых воплощениях антагонист TNFα имеет аминокислотную последовательность, приведенную в SEQ ID NO: 13, или ее функциональный фрагмент.
В некоторых воплощениях связывающая молекула представляет собой слитый белок, содержащий антагонист NGF, слитый с антагонистом TNFα через линкер. В некоторых воплощениях связывающая молекула представляет собой гомодимер слитого белка.
В некоторых воплощениях антагонист NGF представляет собой scFv-домен антитела к NGF, а антагонист TNFα представляет собой растворимый TNFα-связывающий фрагмент TNFR-2, слитый по своему карбоксиконцу с Fc-доменом иммуноглобулина. В некоторых воплощениях scFv слит с карбоксиконцом Fc-домена иммуноглобулина через линкер.
В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент (75 кДа) TNFR-2, Fc-домен IgG1 человека, линкер, состоящий из 10 аминокислот (GGGGS)2, VH-домен, содержащий аминокислотную последовательность SEQ ID NO: 3, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15), и VL-домен, содержащий аминокислотную последовательность SEQ ID NO: 7. В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность SEQ ID NO: 14. В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 14.
В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент (75 кДа) TNFR-2, Fc-домен IgG1 человека, линкер, состоящий из 10 аминокислот (GGGGS)2, VH-домен, содержащий аминокислотную последовательность SEQ ID NO: 94, линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4 (SEQ ID NO: 19), и VL-домен, содержащий аминокислотную последовательность SEQ ID NO: 95. В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность SEQ ID NO: 17. В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 17.
В некоторых воплощениях связывающая молекула содержит гомодимер слитого полипептида, содержащего, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент (75 кДа) TNFR-2, Fc-домен IgG1 человека, линкерную последовательность и scFv-домен антитела к NGF.
Согласно изобретению также предложены способы ингибирования фосфорилирования р38 в клетке, причем указанный способ включает приведение в контакт клетки с любым из полипептидов, описанных в данной заявке (например, с любой из связывающих молекул, содержащих домен антагониста NGF и домен антагониста TNFα, описанных в данной заявке). Согласно изобретению также предложены способы ингибирования фосфорилирования внеклеточной сигнал-регулируемой киназы (ERK) в клетке, при этом указанный способ включает приведение в контакт клетки с любым из полипептидов, описанных в данной заявке (например, с любой из связывающих молекул, содержащих домен антагониста NGF и домен антагониста TNFα, описанных в данной заявке). В некоторых воплощениях клетка представляет собой нейронную клетку. В других воплощениях клетка представляет собой периферическую нейронную клетку. В следующих воплощениях клетка представляет собой центральную нейронную клетку. В некоторых воплощениях клетка представляет собой клетку млекопитающего. В некоторых воплощениях млекопитающим является человек. В некоторых воплощениях клетка находится в культуре клеток.
Согласно изобретению также предложены полинуклеотидные последовательности, кодирующие связывающие молекулы, описанные в данной заявке, векторы, содержащие эти полинуклеотидные последовательности, и клетки-хозяева, содержащие эти полинуклеотиды или векторы.
Согласно изобретению также предложены способы получения связывающих молекул, описанных в данной заявке.
Согласно изобретению также предложены композиции, фармацевтические композиции и наборы, содержащие связывающие молекулы, описанные в данной заявке.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ/ФИГУР
Фиг. 1. Схематичное представление слитого белка TNFR2-Fc (панель А) и типичной мультиспецифической связывающей молекулы, TNFR2-Fc_VH#4, содержащей TNFR2-Fc-домен, слитый с scFv-доменом антитела к NGF (панель В).
На Фиг. 2А показаны результаты анализа, с использованием гель-проникающей высокоэффективной жидкостной хроматографии (SEC-HPLC), уровней агрегатов, мономеров и белковых фрагментов в партии очищенной TNFR2-Fc_VH#4.
На Фиг. 2В показаны результаты анализа посредством SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) очищенного белка TNFR2-Fc_VH#4 и очищенного белка TNFR2-Fc в восстанавливающих и невосстанавливающих условиях. Порядок нанесения на гель: 1. TNFR2-Fc_VH#4, 2. TNFR2-Fc_VL-VH (TNFR2-Fc, слитый с scFv-доменом антитела к NGF, с обратной ориентацией гена вариабельного домена), 3. TNFR2-Fc с нерелевантным scFv 1, 4. TNFR2-Fc, 5. TNFR2-Fc с нерелевантным scFv 2.
На Фиг. 3А показана чистота TNFR2-Fc_VH#4 после очистки на колонке с иммобилизованным белком А. На Фиг. 3В показана чистота TNFR2-Fc_VH#4 после второй стадии очистки на колонке с сульфопропил (SP)-сефарозой.
На Фиг. 4 показаны результаты анализа стабильности TNFR2-Fc_VH#4 с использованием дифференциальной сканирующей калориметрии.
На Фиг. 5 показано связывание TNFR2-Fc_VH#4 с TNFα и NGF, как по отдельности, так и совместно, определенное посредством иммуноферментного твердофазного анализа (ELISA). На Фиг. 5А показано связывание с NGF, на Фиг. 5В показано связывание с TNFα, а на Фиг. 5С показано связывание одновременно с TNFα и NGF.
На Фиг. 6 показана сенсограмма анализа связывания с применением поверхностного плазмонного резонанса для TNFR2-Fc_VH#4. Конкурентное связывание антигенов с мультиспецифическим антителом TNFR2-Fc_VH#4 проводили, используя BIAcore 2000. Одновременное связывание с антигенами оценивали посредством связывания сериями TNFα и NGF с TNFR2-Fc_VH#4, присоединенным к поверхности сенсорного чипа. На первой части сенсограммы показано связывание насыщающих количеств TNFα с мультиспецифическим антителом, на второй части этой сенсограммы показано связывание, когда применяли второй антиген, либо снова TNFα, что показывало полноту насыщения поверхности, либо эквимолярную смесь TNFα и NGF. Увеличение числа резонансных единиц (RU) соответствовало связыванию NGF с мультиспецифической молекулой и, следовательно, говорило о совместном участии антигенов. Анализ также проводили в условиях добавления антигенов в обратном порядке, что подтверждало эти данные.
На Фиг. 7 показано ингибирование NGF-опосредованной пролиферации клеток TF-1. А. NGF-опосредованная пролиферация в отсутствие добавленного антагониста NGF. В. Ингибирование ответа на NGF человека (hNGF) под действием TNFR2-Fc_VH#4. С. Ингибирование ответа на NGF мыши (muNGF) под действием TNFR2-Fc_VH#4. Активность NGF обычно выражают в RLU - относительных единицах люминесценции, а % NGF-опосредованной пролиферации рассчитывают как % ответа только на лиганд NGF, используя следующую формулу:
100*(RLU в лунке - RLU фона)/(общее число RLU - RLU фона),
где RLU фона обозначает среднее значение для сред, используемых в качестве контроля, а общее число RLU обозначает среднее значение для контролей, представляющих собой только лиганд. D. Ингибирование ответа на NGF человека под действием TNFR2-Fc_VarB и ндимаба VarB. Е. Ингибирование ответа на NGF мыши под действием TNFR2-Fc_VarB и ндимаба VarB.
На Фиг. 8 показано ингибирование TNFα-индуцированной активности каспазы 3 в клетках U937. А. TNFα-индуцированная активность каспазы 3 в клетках U937 в отсутствие добавленного антагониста TNFα. В. Ингибирование TNFα-индуцированной активности каспазы 3 в клетках U937, показанное в виде процента ответа в отсутствие добавленного антагониста. Активность TNF обычно выражают в RFU - относительных единицах флуоресценции, а % TNF-опосредованного высвобождения каспазы 3 рассчитывают как % ответа только на лиганд TNF, используя формулу, описанную выше для ФИГ. 7С. С. Аналогичные результаты показаны для родственной молекулы TNFR2-Fc_VarB и ндимаба VarB.
На Фиг. 9 показано влияние комбинированного лечения этанерцептом и MEDI-578 на механическую гипералгезию, индуцированную частичной перевязкой седалищного нерва. Результаты приведены в виде соотношения между ипсилатеральной/контралатеральной лапами. N=9-10 на одну группу. Данные анализировали с применением двухфакторного дисперсионного анализа (2-way ANOVA), используя в качестве зависимых факторов время и способ лечения. Далее определяли статистическую значимость, используя апостериорный критерий Бонферрони. ***р<0,001 относительно контроля Опер, (оперированные животные) + САТ-251.
На Фиг. 10А показано влияние TNFR2-Fc_VH#4 на механическую гипералгезию, индуцированную частичной перевязкой седалищного нерва. Результаты приведены в виде соотношения между ипсилатеральной/контралатеральной лапами. N=10 на одну группу. Данные анализировали с применением двухфакторного ANOVA, используя в качестве зависимых факторов время и способ лечения. Далее определяли статистическую значимость, используя апостериорный критерий Бонферрони. ***р<0,001 относительно биспецифического изотипического контроля (0,03 мг/кг, п.к.). На Фиг. 10В показаны аналогичные результаты для родственной молекулы TNFR2-Fc_VarB.
На Фиг. 11 показано влияние совместного введения MEDI-578 и этанерцепта на ослабление боли в модели гиперчувствительности к механическим раздражителям на примере суставной боли. N=9-10 на одну группу. Данные анализировали, используя двухфакторный ANOVA. Далее определяли статистическую значимость, используя апостериорный критерий Бонферрони. *р>0,05; ***р<0,001 относительно САТ-251 (0,03 мг/кг, в.в.) + PBS (10 мл/кг, в.б.).
На Фиг. 12 показано влияние TNFR2-Fc_VH#4 на ослабление боли в модели гиперчувствительности к механическим раздражителям. N=9-10 на одну группу. Данные анализировали, используя двухфакторный ANOVA. Далее определяли статистическую значимость, используя апостериорный критерий Бонферрони. ***р<0,001 относительно биспецифического изотипического контроля (0,01 мг/кг, п.к.).
На Фиг. 13 показано влияние пяти разных доз TNFR2-Fc_varB на CFA-индуцированную гипералгезию в крысиной модели. Статистический анализ: ANOVA с повторными измерениями, затем критерий планируемых сравнений с использованием InVivoStat. Значительное устранение гипералгезии по сравнению с наполнителем в каждый момент времени.
Фиг. 14: тепловая карта, показывающая HTRF (гомогенная флуоресценция с разрешением по времени) - соотношения для реакций фосфорилирования р38.
Фиг. 15: кривые зависимости доза-ответ, демонстрирующие влияние TNFα, NGF или комбинации TNFα и NGF на фосфорилирование р38.
Фиг. 16: тепловая карта, показывающая HTRF-соотношения для реакций фосфорилирования ERK.
Фиг. 17: кривые зависимости доза-ответ, демонстрирующие влияние TNFα, NGF или комбинации TNFα и NGF на фосфорилирование ERK.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Следует отметить, что термин «объект» относится к одному или более чем одному такому «объекту». По сути, термины «один/одна/одно или более» и «по меньшей мере один/одна/одно» в данном описании можно использовать взаимозаменяемо.
Кроме того, при использовании в данном описании выражение «и/или» следует толковать как конкретное описание каждого из двух конкретных признаков или компонентов вместе со вторым или без него. Таким образом, подразумевается, что термин «и/или», который использован в данном описании в такой фразе, как «А и/или В» включает в себя «А и В», «А или В», (только) «А» и (только) «В». Аналогичным образом, подразумевается, что термин «и/или», который использован в такой фразе, как «А, В и/или С», охватывает каждый из следующих аспектов: А, В и С; А, В или С; А или С; А или В; В или С; А и С; А и В; В и С; (только) А; (только) В; и (только) С.
Очевидно, что помимо случаев, где аспекты в данной заявке описаны с использованием слова «содержащий», также предложены другие аналогичные аспекты, описанные в терминах «состоящий из» и/или «состоящий по существу из».
Если не указано иное, все технические и научные термины, использованные в данном описании, имеют значение, обычно понимаемое средним специалистом в области техники, к которой данное изобретение относится. Например, издания Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press; и Oxford Dictionary of Biochemistry and Molecular Biology, Revised, 2000, Oxford University Press, дают возможность специалисту познакомиться с со словарем многих общеупотребительных терминов, используемых в этом описании.
Единицы измерения, префиксы и символы указаны в форме, принятой Международной системой единиц (СИ). Числовые диапазоны содержат числа, определяющие этот диапазон. Если не указано иное, аминокислотные последовательности записаны слева направо в направлении от амино-конца к карбокси-концу. Заголовки, представленные в данном описании, не ограничивают различные аспекты изобретения, которые могут быть определены посредством ссылки на описание в целом. Соответственно, термины, описанные непосредственно ниже, более полно определены посредством ссылки на описание во всей его полноте.
Использованный в данном описании термин «связывающая молекула» относится в своем самом широком смысле к молекуле, которая специфично связывается с антигенной детерминантой, например, антигеном. Неограничивающие примеры связывающей молекулы включают антитела или их фрагменты, слитые белки на основе растворимого рецептора или их фрагменты, неиммуноглобулиновые каркасы или их фрагменты, при этом у каждого из них сохраняется антиген-специфическое связывание. Типичные неиммуноглобулиновые каркасы включают Tn3 (Koide et al., J. Mol. Biol., 2012, Jan 13, 415(2): 393-405), DARPin (сконструированный белок с анкириновыми повторами) (Boersma & Pluckthun, Curr. Opin. Biotechnol., 2011, 22(6): 849-57), антикалин (Gebauer & Skerra, Methods Enzymol., 2012, 503: 157-88). Типичные слитые белки на основе растворимого рецептора, а также антитела приведены ниже. В некоторых воплощениях связывающая молекула может быть сконструирована в виде молекулы, содержащей комбинации таких антител или их фрагментов, слитых белков на основе растворимого рецептора или их фрагментов и неиммуноглобулиновых каркасов или их фрагментов.
Связывающая молекула или любая часть связывающей молекулы, которая распознает антиген, обозначается в данном описании как «связывающий домен». Помимо специального указания на полноразмерные связывающие молекулы, такие как природные антитела, термин «связывающая молекула» охватывает, без ограничения, полноразмерные антитела или другие не являющиеся антителами связывающие молекулы, а также антиген-связывающие фрагменты, варианты, аналоги или производные таких связывающих молекул, например, молекулы природных антител или иммуноглобулинов либо сконструированные связывающие молекулы или фрагменты, которые связываются с антигеном аналогично полноразмерной связывающей молекуле.
В некоторых воплощениях согласно изобретению предложены некоторые мультиспецифические связывающие молекулы, например, биспецифические, триспецифические, тетраспецифические и т.д. связывающие молекулы или их антиген-связывающие фрагменты, варианты или производные. Как использовано в данном описании, мультиспецифическая связывающая молекула может включать один или более чем один связывающий домен антитела, один или более чем один не относящийся к антителу связывающий домен или их комбинацию.
Использованный в данном описании термин «фактор роста нервов» («NGF»), также обозначаемый в литературе как бета-фактор роста нервов, относится к секретируемому белку, который принимает участие в процессах роста и выживаемости различных нейронов. NGF человека (humNGF) представлен номером доступа в Genbank NP_002497.2 и в данном описании приведен как SEQ ID NO: 1. Термин «NGF», использованный в данном описании, не ограничивается NGF человека и включает в себя все видовые ортологи NGF человека. Термин «NGF» охватывает проформу NGF, pro-NGF, полноразмерный NGF, а также любую форму NGF, которая образуется в результате процессинга в клетке. Термин также охватывает природные варианты NGF, например, сплайс-варианты, аллельные варианты и изоформы. NGF может связываться с двумя рецепторами: рецептором нейротрофинов р75 (p75(NTR)) и TrkA, трансмембранной тирозинкиназой. NGF представляет собой хорошо обоснованную мишень для устранения боли, при этом известно, что он опосредует сенситизацию ноцицепторов.
В настоящее время ряд агентов проходит тестирование в качестве антагонистов активности NGF. Одним из таких анти-NGF агентов является trkA-Fc, который действует как ловушка или скэвенджер, связываясь с эндогенным NGF и тем самым инактивируя его. TrkA-Fc представляет собой слитый белок, состоящий из NGF-связывающего участка trkA, присоединенного к фрагменту константного домена (Fc) IgG антитела. TrkA-Fc приводит к развитию гипоалгезии у не использовавшихся ранее в опытах животных, ослабляет ноцицептивные ответы и уменьшает разрастание немиелинизированных, воспринимающих боль нейронов (Bennett, D.L. et al. (1998) Eur. J. Neurosci., 10. 1282-91).
Связывающие молекулы, описанные в данной заявке, особенно хорошо подходят для безопасного и эффективного лечения NGF-опосредуемой боли, поскольку уровни NGF повышаются на периферии в ответ на болевые стимулы, а антитела обладают низкой проходимостью через гематоэнцефалический барьер. Ряд антител к NGF и их антиген-связывающих фрагментов, которые можно использовать в терапевтических схемах и композициях, описанных в данном изобретении, можно найти в литературе; см., например, публикации РСТ с №№ WO 02/096458 и WO 04/032870.
Термин «MEDI-578» относится к антителу, специфично связывающемуся с NGF, которое представляет собой предмет международной заявки № PCT/GB 2006/000238 и публикации заявки на патент США №2008/0107658 А1; обе они включены в данное описание посредством ссылки во всей своей полноте. Последовательности тяжелой и легкой цепи MEDI-578 показаны в SEQ ID NO: 3 и 7, соответственно.
Термин NGF-NG относится к антителу, которое специфично связывается с NGF. Последовательности тяжелой и легкой цепи NGF-NG показаны в SEQ ID NO: 24 и 26, соответственно.
Термин «фактор некроза опухоли альфа» («TNFα»), также называемый в литературе как кахектин, белок АРС1; фактор некроза опухоли; TNF; или член 2 суперсемейства лигандов фактора некроза опухоли, как использовано в данном описании, относится к конкретному белку TNFα, а не суперсемейству лигандов TNF. TNFα человека представлен в Genbank с номером доступа NP_000585.2 и представлен как SEQ ID NO: 2. Термин «TNFα», использованный в данном описании, не ограничивается TNF человека, а включает все видовые ортологи TNFα человека. Термин «TNFα» охватывает проформу TNFα, pro-TNFα, полноразмерный TNFα, а также любую форму TNFα, которая образуется в результате процессинга в клетке. Термин также охватывает природные и неприродные варианты TNFα, например, сплайс-варианты, аллельные варианты и изоформы. TNFα может связываться с двумя рецепторами, TNFR1 (рецептор TNF, тип 1; CD120a; р55/60) и TNFR2 (рецептор TNF, тип 2; CD120b; р75/80). TNFα действует в качестве провоспалительного цитокина, например, функционируя при нейровоспалении. Например, считается, что TNFα вовлечен функционально в возникновение нейропатической боли (Leung, L. and Cahill, СМ., J. Neuroinflammation, 7: 27 (2010)).
В данной области техники известно большое количество антагонистов TNFα, и многие из них продаются в качестве терапевтических средств. Примеры имеющихся в продаже антагонистов TNF-альфа, которые можно использовать в терапевтических схемах и композициях, предложенных в данном изобретении, включают этанерцепт (ENBREL®, Amgen/Pfizer), инфликсимаб (например, REMICADE®, Centocor), цертолизумаба пегол (например, CIMZIA®, UCB), голимумаб (например, SIMPONI™, Centocor) и адалимумаб (например, HUMIRA®/TRUDEXA®, Abbott).
Термин «выделенная/ый/ое» связывающая молекула, полипептид, антитело, полинуклеотид, вектор, клетка-хозяин или соединение относится к связывающей молекуле, полипептиду, антителу, полинуклеотиду, вектору, клетке хозяина или соединению, которые не находятся в форме, встречающейся в природе. Выделенные связывающие молекулы, полипептиды, антитела, полинуклеотиды, векторы, клетки хозяина или соединения включают таковые, которые изменены, адаптированы, объединены, перегруппированы, сконструированы или иным образом обработаны до такой степени, что они больше не находятся в той форме, в которой они встречаются в природе. Согласно некоторым аспектам «выделенная/ое/ый» связывающая молекула, антитело, полинуклеотид, вектор, клетка-хозяин или соединение являются «рекомбинантными».
Использованные в данном описании термины «многофункциональный полипептид» и «бифункциональный полипептид» относятся к не существующей в природе связывающей молекуле, сконструированной с целью направленного взаимодействия с двумя или более антигенами. Многофункциональный полипептид, описанный в данной заявке, обычно представляет собой генетически сконструированный слитый белок, созданный с целью объединения двух разных желаемых биологических функций в единой связывающей молекуле. Например, многофункциональный полипептид может представлять собой многофункциональную связывающую молекулу. Типичный многофункциональный полипептид, описанный в данной заявке, представляет собой многофункциональную связывающую молекулу, содержащую домен антагониста NGF, например, пептидный домен, который блокирует, ослабляет или ингибирует одну или более природных функций NGF, и домен антагониста TNFα, например, пептидный домен, который блокирует, ослабляет или ингибирует одну или более природных функций TNFα.
Одной из групп многофункциональных полипептидов, предложенных согласно данному изобретению, являются мультиспецифические связывающие молекулы, например, связывающие молекулы, которые содержат один или несколько связывающих доменов антител, например, «мультиспецифическое антитело», один или несколько не относящихся к антителам связывающих доменов, например, рецептор-ловушка, или их комбинацию. Мультиспецифические связывающие молекулы, например, содержащие один или несколько связывающих доменов антител, один или несколько не относящихся к антителам связывающих доменов или их комбинацию, представляют собой молекулы, содержащие связывающие домены, способные специфично распознавать по меньшей мере два разных эпитопа и связываться с ними. Разные эпитопы могут находиться или на одной и той же молекуле (например, на одном и том же NGF), или на разных молекулах, так что, например, мультиспецифические связывающие молекулы могут специфически распознавать и связывать NGF, а также содержащую другой эпитоп молекулу, например, TNFα, таким образом, мультиспецифическая связывающая молекула специфически распознает NGF и TNFα.
Способы получения мультиспецифических связывающих молекул, например, содержащих один или несколько связывающих доменов антител, один или несколько не относящихся к антителам связывающих доменов или их комбинацию, известны в данной области техники (Dimasi, N., et al., 2009, J. Mol. Biol., 393: 672-92; Milstein et al., 1983, Nature, 305: 537-539; Brennan et al., 1985, Science, 229:81; Suresh et al., 1986, Methods in Enzymol., 121: 120; Traunecker et al., 1991, EMBO J., 10: 3655-3659; Shalaby et al., 1992, J. Exp. Med., 175: 217-225; Kostelny et al., 1992, J. Immunol., 148: 1547-1553; Gruber et al., 1994, J. Immunol., 152. 5368; и патент США 5731168). Также охвачены антитела более чем с двумя валентностями. Например, могут быть получены триспецифические антитела (Tutt et al., J. Immunol., 147: 60 (1991)).
Термин «антитело» означает молекулу иммуноглобулина, которая распознает и специфически связывается с мишенью, такой как белок, полипептид, пептид, углевод, полинуклеотид, липид или комбинации вышеупомянутого, посредством по меньшей мере одного антиген-распознающего сайта в вариабельной области молекулы иммуноглобулина. Использованный в данном описании термин «антитело» охватывает интактные поликлональные антитела, интактные моноклональные антитела, фрагменты антител (такие как Fab, Fab', F(ab')2 и Fv фрагменты), мутантные формы одноцепочечных Fv (scFv), мультиспецифические антитела, такие как биспецифические, триспецифические, тетраспецифические и т.д. антитела, образованные по меньшей мере из двух интактных антител, химерные антитела, гуманизированные антитела, человеческие антитела, слитые белки, содержащие антиген-определяющую часть антитела, и любую другую модифицированную молекулу иммуноглобулина, которая содержит антиген-распознающий сайт, при условии, что данные антитела проявляют желаемую биологическую активность. Антитело может представлять собой любое антитело из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, или их подклассов (изотипов) (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2), на основании идентификации своих константных доменов тяжелой цепи, называемых альфа, дельта, эпсилон, гамма и мю, соответственно. Разные классы иммуноглобулинов имеют разные и хорошо известные субъединичные структуры и пространственные конфигурации.
В некоторых воплощениях «блокирующая» связывающая молекула, например, блокирующее антитело, или «антагонистическая» связывающая молекула, такая как, например, антагонистическое антитело или слитый белок, представляет собой молекулу, которая ингибирует или снижает биологическую активность антигена, с которым она связывается, такого как NGF или TNFα. Согласно определенным аспектам, блокирующие антитела или антагонистические связывающие молекулы существенно или полностью ингибируют биологическую активность данного антигена. Например, биологическая активность может быть снижена на 0,01%, 0,1%, 0,5%, 1%, 5%, 10%, 20%, 30%, 50%, 70%, 80%, 90%, 95% или даже 100%. «Антагонисты» и «домены антагонистов», как использовано в данном описании, включают полипептиды или другие молекулы, которые связываются со своей мишенью (например, TNFα или NGF), тем самым блокируя или ингибируя взаимодействие данной мишени с рецептором. Так, антагонисты NGF и/или TNFα включают молекулы, которые блокируют или ингибируют взаимодействие NGF с trkA или р75-нейротрофином либо взаимодействие TNFα с TNFR-1 или TNFR-2. Антагонисты NGF и/или TNFα также включают молекулы, которые ослабляют фосфорилирование р38 и/или фосфорилирование ERK. Типичные антагонисты включают, но не ограничиваются этим, антитела или их антиген-связывающие фрагменты и специфические к мишени пептиды растворимых несигнальных рецепторов («рецепторов-ловушек» или их лиганд-связывающих фрагментов).
Термин «фрагмент антитела» относится к части интактного антитела и относится к определяющим антигенность вариабельным областям интактного антитела. Примеры фрагментов антител включают, но не ограничиваются этим Fab, Fab', F(ab')2 и Fv фрагменты, линейные антитела, одноцепочечные антитела и мультиспецифические антитела, образованные из фрагментов антител. Антиген-связывающие фрагменты не относящихся к антителам связывающих молекул, описанные где-либо еще в данном описании, также предложены согласно данному изобретению.
«Моноклональное антитело» относится к гомогенной популяции антител, вовлеченной в высокоспецифическое распознавание и связывание одной антигенной детерминанты или одного эпитопа. Это отличает их от поликлональных антител, которые обычно включают разные антитела, направленные против разных антигенных детерминант. Термин «моноклональное антитело» охватывает как интактные и полноразмерные моноклональные антитела, так и фрагменты антител (такие как Fab, Fab', F(ab')2, Fv), мутантные формы одноцепочечных Fv (scFv), слитые белки, содержащие часть антитела, и любую другую модифицированную молекулу иммуноглобулина, содержащую антиген-распознающий сайт. Кроме того, «моноклональное антитело» относится к таким антителам, которые получают любым из ряда методов, включая, но не ограничиваясь этим, использование гибридом, отбор с применением фагов, рекомбинантную экспрессию и применение трансгенных животных.
Термин «гуманизированное антитело» относится к формам антител, не являющихся человеческими (например, являющихся мышиными) антителами, представляющими собой конкретные иммуноглобулиновые цепи, химерные иммуноглобулины или их фрагменты, которые содержат минимальное количество не являющихся человеческими (например, являющихся мышиными) последовательностей. Обычно гуманизированными антителами считаются иммуноглобулины человека, у которых остатки в гипервариабельном участке (CDR) заменены на остатки из CDR не являющегося человеком вида (например, мыши, крысы, кролика или хомяка), которые имеют желаемую специфичность, аффинность и эффективность (Jones et al., 1986, Nature, 321: 522-525; Riechmann et al., 1988, Nature, 332: 323-327; Verhoeyen et al., 1988, Science, 239: 1534-1536). В некоторых случаях остатки каркасной области Fv (FR или FW) человеческого иммуноглобулина заменены на соответствующие остатки, имеющиеся в антителе из вида не относящегося к человеку, которое имеет желаемую специфичность, аффинность и эффективность. Гуманизированное антитело может быть далее модифицировано путем замены дополнительных остатков или в каркасной области Fv, и/или среди таких замененных остатков, не относящихся к антителу человека, для улучшения и оптимизации специфичности, аффинности и/или эффективность антитела. В общем случае гуманизированное антитело будет содержать по существу все из по меньшей мере одного и обычно двух или трех вариабельных доменов, содержащих все или по существу все участки CDR, соответствующие иммуноглобулину, не являющемуся иммуноглобулином человека, в то время как все или по существу все участки FR представляют собой участки консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело также может содержать по меньшей мере часть константной области или константного домена (Fc) иммуноглобулина, обычно являющегося иммуноглобулином человека. Примеры способов, используемых для получения гуманизированных антител, описаны в патенте США 5225539 или 5639641.
«Вариабельная область» антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела, или по отдельности, или в комбинации. Вариабельные области каждой из тяжелой и легкой цепи состоят из четырех каркасных областей (FR или FW), соединенных тремя определяющими комплементарность участками (CDR), также известными как гипервариабельные участки. CDR в каждой цепи удерживаются вместе в непосредственной близости благодаря FR и, вместе с CDR из другой цепи, вносят вклад в образование антиген-связывающего сайта антител. Существуют по меньшей мере два метода определения CDR: (1) подход, основанный на межвидовой вариабельности последовательностей (т.е. Kabat et al., Sequences of Proteins of Immunological Interest (5-е изд., 1991, Национальный институт здравоохранения (National Institutes of Health), Bethesda Md.)); и (2) подход, основанный на результатах кристаллографических исследований комплексов антиген-антитело (Al-lazikani etal. (1997) J. Molec. Biol., 273: 927-948)). Помимо этого, для определения CDR в данной области техники иногда используют комбинации этих двух подходов.
Систему нумерации по Kabat обычно используют со ссылкой на остаток в вариабельном домене (приблизительно на остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, Md. (1991)).
Нумерация положений аминокислот в соответствии с подходом Kabat относится к системе нумерации, используемой для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи при компиляции антител согласно Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, Md. (1991). При использовании этой системы нумерации истинная линейная последовательность аминокислотных остатков может содержать меньшее количество аминокислот или содержать дополнительные аминокислоты в соответствии с уменьшением длины FR или CDR вариабельного домена или со вставкой в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может содержать вставку одного аминокислотного остатка (остаток 52а согласно Kabat) после остатка 52 в Н2 и встроенные остатки (например, остатки 82а, 82b и 82с и т.д. согласно Kabat) после остатка 82 в FR тяжелой цепи. Нумерация остатков по Kabat может быть проведена для заданного антитела путем выравнивания участков гомологии в последовательности этого антитела со «стандартной» пронумерованной по Kabat последовательностью. В отличие от этого, Chothia ссылается на расположение структурных петель (Chothia and Lesk, J. Mol. Biol., 196: 901-917 (1987)). Окончание петли CDR-H1 no Chothia, при определении номера с использованием правила нумерации по Kabat, варьирует между Н32 и Н34 в зависимости от длины петли (это происходит потому, что схема нумерации по Kabat допускает наличие вставок в положениях Н35А и Н35В; если ни в положении 35А, ни в положении 35В вставки нет, то петля заканчивается в положении 32; если вставка присутствует только в положении 35А, то петля заканчивается в положении 33; если присутствуют вставки в обоих положениях 35А и 35В, то петля заканчивается в положении 34). Гиперв ариабельные участки по AbM представляет собой компромисс между CDR по Kabat и структурными петлями по Chothia, и они используются программным обеспечением AbM от Oxford Molecular's для моделирования антител. Сравнение приведено ниже в Таблице 1.

Термин «человеческое антитело» означает нативное человеческое антитело или антитело, имеющее аминокислотную последовательность, соответствующую нативному человеческому антителу, полученное с использованием любого метода, известного в данной области техники. Такое определение человеческого антитела включает в себя интактные или полноразмерные антитела, их фрагменты и/или антитела, содержащие по меньшей мере один полипептид тяжелой и/или легкой цепи иммуноглобулина человека, такие как, например, антитело, содержащее полипептиды легкой цепи иммуноглобулина мыши и тяжелой цепи иммуноглобулина человека.
Термин «химерные антитела» относится к антителам, у которых аминокислотная последовательность молекулы иммуноглобулина происходит из двух или более видов. Обычно вариабельная область и легкой, и тяжелой цепей соответствует вариабельной области антител, происходящих из одного вида млекопитающих (например, мыши, крысы, кролика и т.д.) с желаемой специфичностью, аффинностью и эффективностью, в то время как последовательности константных областей гомологичны последовательностям в антителах, происходящих из другого вида (обычно человека), чтобы избежать индуцирования иммунного ответа у этого вида. Мультиспецифические связывающие молекулы, например, содержащие один или несколько связывающих доменов антител, один или несколько не относящихся к антителам связывающих доменов или их комбинацию, например, антагонисты TNFα и/или антагонисты NGF, предложенные согласно данному изобретению, могут содержать константные области антител (например, Fc-области), в которых по меньшей мере часть одного или нескольких доменов константной области делетирована или иным образом изменена с целью обеспечения желаемых биохимических характеристик, таких как повышенная локализация в опухолях или сниженный полупериод существования в сыворотке по сравнению с антителом приблизительно той же иммуногенности, содержащим нативную или неизмененную константную область. Модифицированные константные области, приведенные в данном описании, могут содержать изменения или модификации в одном или более чем одном из трех константных доменов тяжелой цепи (СН1, СН2 или СН3) и/или в константном домене легкой цепи (CL). Согласно определенным аспектам, один или более чем один константный домен может быть частично или полностью делетирован. Согласно определенным аспектам, может быть делетирован весь домен СН2 (конструкции ΔСН2). См., например, Oganesyan V., et al., 2008, Acta Crystallogr. D Biol. Crystallogr., 64: 700-4; Oganesyan V., et al., Mol. Immunol., 46: 1750-5; Dall'Acqua, W.F., et al., 2006, J. Biol. Chem., 281: 23514-23524; и Dall'Acqua, et al., 2002, J. Immunol., 169: 5171-5180.
Термины «эпитоп» и «антигенная детерминанта» используются в данном описании взаимозаменяемо и относятся к той части антигена, которую может распознавать и с которой может специфически связываться конкретное антитело. Если антигеном является полипептид, то эпитопы могут быть образованы как из следующих одна за другой аминокислот, так и из не следующих одна за другой аминокислот, сближенных в результате формирования третичной структуры белка. Эпитопы, образованные из следующих одна за другой аминокислот, обычно сохраняются после денатурации белка, тогда как эпитопы, образованные в результате формирования третичной структуры, обычно утрачиваются после денатурации белка. В типичном случае эпитоп включает по меньшей мере 3 и обычно больше, по меньшей мере 5 или 8-10 аминокислот, в уникальной пространственной конформации. Эпитоп, описанный в данной заявке, не обязательно должен быть определен вплоть до конкретных аминокислот, которые образуют этот эпитоп. Согласно определенным аспектам, эпитоп можно идентифицировать посредством изучения связывания антитела с пептидными субъединицами полипептидного антигена или посредством изучения конкурентного связывания с данным антигеном группы антиген-специфических антител.
Под «субъектом», или «индивидуумом», или «животным», или «пациентом», или «млекопитающим» понимается любой субъект, в частности, млекопитающее, для которого желательно проведение диагностики, прогнозирования или терапии. Млекопитающие включают людей, домашних животных, сельскохозяйственных животных, участвующих в спортивных мероприятиях животных и животных зоопарков, включая, например, людей, приматов, не являющихся человеком, собак, кошек, морских свинок, кроликов, крыс, мышей, лошадей, крупный рогатый скот, медведей и так далее.
Термины «композиция» и «фармацевтическая композиция» относятся к препарату, находящемуся в такой форме, которая способствует эффективному проявлению биологической активности активного ингредиента и которая не содержит никаких дополнительных компонентов, являющихся неприемлемо токсичными для субъекта, которому данная композиция будет введена. Такие композиции могут быть стерильными.
Использованные в данном описании термины «эффективное количество» и «терапевтически эффективное количество» относятся к количеству одной или более терапевтических композиций, эффективных для контролирования боли у субъекта. Термин «контролирование боли» и его грамматические эквиваленты используются в данной заявке для описания любого полезного или желательного эффекта в отношении субъекта, нуждающегося в контролировании боли. Например, эффективное количество одной или более чем одной терапевтической композиции, описанной в данной заявке, может, например, предотвращать возникновение боли, поддерживать переносимый уровень боли, уменьшать интенсивность боли, ослаблять боль, минимизировать боль или устранять боль у субъекта.
Термин «введение», использованный в данном описании, относится к введению субъекту одной или более терапевтических композиций, описанных здесь, например, бифункционального полипептида, содержащего домен антагониста NGF и домен антагониста TNFα, терапевтической композиции, содержащей комбинацию антагониста NGF и антагониста TNFα, или отдельных терапевтических композиций, одной, содержащей антагонист NGF, и другой, содержащей антагонист TNFα. Термин «совместное введение» относится к введению субъекту двух или более терапевтических композиций, например, одной, содержащей антагонист NGF, и одной, содержащей антагонист TNFα. Как использовано в данном описании, совместное введение включает, но не требует того, чтобы две или более терапевтических композиций были введены субъекту одновременно. Такие две или более терапевтических композиций можно вводить субъекту последовательно, например, с интервалом в тридцать минут, с интервалом в один час, с интервалом в два часа, с интервалом в три часа, с интервалом в четыре часа или с интервалом в пять или более часов. Последовательность и время совместного введения, как изложено в данном описании, могут быть фиксированы или могут быть изменены на основании заключения медицинского работника.
Термины «полинуклеотид» или «нуклеиновая кислота» относятся к полимерному соединению, содержащему ковалентно связанные нуклеотидные остатки. Полинуклеотиды могут представлять собой ДНК, кДНК, РНК, одноцепочечные или двухцепочечные полинуклеотиды, векторы, плазмиды, фаговые или вирусные полинуклеотиды.
Термин «вектор» означает конструкцию, которая способна доставлять в клетку хозяина и экспрессировать в ней один ген или несколько генов либо одну последовательность или несколько последовательностей, представляющих интерес. Примеры векторов включают, но не ограничиваются этим, вирусные векторы, экспрессирующие векторы на основе «оголенной» ДНК или РНК, плазмидные, космидные или фаговые векторы, экспрессирующие векторы на основе ДНК или РНК, ассоциированные с катионными конденсирующими агентами, экспрессирующие векторы на основе ДНК или РНК, инкапсулированные в липосомы и некоторые эукариотические клетки, такие как клетки-продуценты.
Термины «полипептид», «пептид» и «белок» используются взаимозаменяемо в данном описании и относятся к полимерам аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, которые могут чередоваться с остатками, не относящимися к аминокислотам. Эти термины также охватывают состоящий из аминокислот полимер, который модифицирован естественным образом или посредством целенаправленного воздействия; например, в результате образования дисульфидных связей, гликозилирования, липидирования, ацетилирования, фосфорилирования или любой другой манипуляции или модификации, такой как конъюгирование с меткой. В это определение также включены, например, полипептиды, содержащие один или более аналогов аминокислот (включая, например, неприродные аминокислоты и т.д.), а также другие модификации, известные в данной области техники.
«Консервативная аминокислотная замена» представляет собой замену, при которой один аминокислотный остаток заменен на другой аминокислотный остаток, имеющий похожую боковую цепь. В данной области техники определены семейства аминокислотных остатков, имеющих похожие боковые цепи, в том числе основные боковые цепи (например, у лизина, аргинина, гистидина), кислотные боковые цепи (например, у аспарагиновой кислоты, глутаминовой кислоты), незаряженные полярные боковые цепи (например, у аспарагина, глутамина, серина, треонина, тирозина, цистеина), неполярные боковые цепи (например, у глицина, аланина, валина, лейцина, изолейцина, пролина, фенилаланина, метионина, триптофана), бета-разветвленные боковые цепи (например, у треонина, валина, изолейцина) и ароматические боковые цепи (например, у тирозина, фенилаланина, триптофана, гистидина). Например, замена фенилаланина на тирозин представляет собой консервативную замену. Согласно определенным аспектам, консервативные замены в последовательностях полипептидов и антител, предложенных в данном изобретении, не подавляют связывание или другую функциональную активность полипептида, содержащего такую аминокислотную последовательность. Способы идентификации нуклеотидных и аминокислотных консервативных замен, не затрагивающих функцию, хорошо известны в данной области техники (см., например, Brummell et al., Biochem., 32: 1180-1187 (1993); Kobayashi et al., Protein Eng., 12: 879-884 (1999); и Burks et al., Proc. Natl. Acad. Sci. USA, 94: 412-417 (1997)).
Связывающая молекула, содержащая домен антагониста NGF и домен антагониста TNFα
Согласно данному изобретению, предложен бифункциональный полипептид, содержащий домен антагониста NGF и домен антагониста TNFα. Согласно определенным аспектам, введение эффективного количества бифункционального полипептида, предложенного согласно изобретению, может контролировать боль у субъекта, нуждающегося в этом, эффективнее, чем введение эквивалентного количества антагониста NGF или антагониста TNFα в виде монотерапии. Бифункциональные полипептиды, предложенные согласно изобретению, могут включать домен антагониста NGF и домен антагониста TNFα в любом порядке, любой структуры или конформации. Все подходящие антагонисты NGF или антагонисты TNFα могут представлять собой часть бифункционального полипептида, предложенного согласно изобретению. Типичные антагонисты NGF и антагонисты TNFα описаны в другом месте в этой заявке.
Согласно определенным аспектам, антагонист NGF представляет собой не являющуюся антителом молекулу или ее связывающий домен, способный ингибировать активность NGF, например, растворимый NGF-связывающий фрагмент TrkA. Согласно определенным аспектам, антагонист NGF представляет собой антитело к NGF или его антиген-связывающий фрагмент. Подходящие анти-NGF антагонисты, например, антагонистические антитела, могут ингибировать связывание NGF с TrkA, p75NTR или как с TrkA, так и с p75NTR. Согласно определенным аспектам, анти-NGF антагонист, например, антагонистическое антитело или его фрагмент для применения в бифункциональной молекуле, предложенной в данной заявке, например, мультиспецифической связывающей молекуле, предпочтительно могут блокировать связывание NGF с TrkA по сравнению со связыванием NGF с P75NTR.
Типичные антитела или их фрагменты для применения в бифункциональных полипептидах, например, мультиспецифических связывающих молекулах, описанных в данной заявке, предложены в публикации заявки США №2008/0107658, которая включена в данное описание посредством ссылки во всей своей полноте. Согласно определенным аспектам, антитело к NGF или его фрагмент связывается с одним и тем же эпитопом, может конкурентно ингибировать или может связываться с NGF с более высокой аффинностью, чем антитело к NGF MEDI-578. В некоторых воплощениях антитело к NGF или его фрагмент связывается с NGF человека и/или NGF крысы с аффинностью равной или меньше 1, 0,8, 0,7, 0,6, 0,5, 0,4, 0,3 или 0,2 нМ. Например, антитело к NGF или его фрагмент может связываться с NGF человека с аффинностью примерно 0,2-0,8, 0,2-0,7, 0,2-0,6, 0,2-0,5 и/или 0,25-0,44 нМ и с NGF крысы с аффинностью примерно 0,2-0,9, 0,2-0,8 и/или 0,25-0,70 нМ.
Согласно определенным аспектам, антитело к NGF или его фрагмент представляют собой MEDI-578. Антитело MEDI-578 описано в публикации заявки США №2008/0107658 как клон 1252А5. Согласно другим аспектам, антитело к NGF или его фрагмент представляет собой танезумаб (RN-624), гуманизированное моноклональное антитело (mAb) к NGF (Pfizer; описанное в Kivitz et al. (2013) Pain, 154, 9, 1603-161), фулранумаб, полностью человеческое mAb к NGF (Amgen; описанное в Sanga et al., Pain, volume 154, issue 10, October 2013, стр. 1910-1919); REGN475/SAR164877, полностью человеческое mAb к NGF (Regeneron/Sanafi-Aventis); ABT-110 (PG110), гуманизированное mAb к NGF (Abbott Laboratories). Антитело к NGF или его фрагмент, включенное(ый) в бифункциональный полипептид, например, мультиспецифическую связывающую молекулу, предложенную согласно изобретению, могут быть, например, гуманизированным, химерным, приматизированным или полностью человеческим.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий домены HCDR1, HCDR2 и HCDR3 из MEDI-578, варианты CDR тяжелой цепи MEDI-578 с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами. Например, антитело к NGF или его фрагмент может содержать HCDR1 с аминокислотной последовательностью SEQ ID NO: 4 как таковой или с аминокислотной последовательностью SEQ ID NO: 4 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать HCDR2 с аминокислотной последовательностью SEQ ID NO: 5 как таковой или с аминокислотной последовательностью SEQ ID NO: 5 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать HCDR3 с аминокислотной последовательностью SEQ ID NO: 6 как таковой или с аминокислотной последовательностью SEQ ID NO: 6 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Согласно определенным аспектам, HCDR3 может содержать аминокислотную последовательность SSRIYDFNSALISYYDMDV (SEQ ID NO: 11) или SSRIYDMISSLQPYYDMDV (SEQ ID NO: 12).
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий домены LCDR1, LCDR2 и LCDR3 из MEDI-578, варианты CDR легкой цепи MEDI-578 с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами. Согласно определенным аспектам, антитело к NGF или его фрагмент может содержать LCDR1 с аминокислотной последовательностью SEQ ID NO: 8 как таковой или с аминокислотной последовательностью SEQ ID NO: 8 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать LCDR2 с аминокислотной последовательностью SEQ ID NO: 9 как таковой или с аминокислотной последовательностью SEQ ID NO: 9 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать LCDR3 с аминокислотной последовательностью SEQ ID NO: 10 как таковой или с аминокислотной последовательностью SEQ ID NO: 10 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 3. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH SEQ ID NO: 3.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 7. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL SEQ ID NO: 7.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 94. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH SEQ ID NO: 94.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 95. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL SEQ ID NO: 95.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий домены HCDR1, HCDR2 и HCDR3 с любой из SEQ ID NO: 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86 и 96 или их варианты с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами включительно, например, консервативными аминокислотными заменами.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий домены LCDR1, LCDR2 и LCDR3 с любой из SEQ ID NO: 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87 и 97 или их варианты с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами включительно, например, консервативными аминокислотными заменами.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности с любой из SEQ ID NO: 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86 и 96. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH с любой из SEQ ID NO: 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86 и 96.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности с любой из SEQ ID NO: 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87 и 97. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL с любой из SEQ ID NO: 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87 и 97.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий домены HCDR1, HCDR2 и HCDR3 из NGF-NG, варианты CDR тяжелой цепи NGF-NG с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами. Например, антитело к NGF или его фрагмент может содержать HCDR1 с аминокислотной последовательностью SEQ ID NO: 88 как таковой или с аминокислотной последовательностью SEQ ID NO: 88 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать HCDR2 с аминокислотной последовательностью SEQ ID NO: 89 как таковой или с аминокислотной последовательностью SEQ ID NO: 89 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать HCDR3 с аминокислотной последовательностью SEQ ID NO: 90 как таковой или с аминокислотной последовательностью SEQ ID NO: 90 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий домены LCDR1, LCDR2 и LCDR3 из NGF-NG, варианты CDR легкой цепи NGF-NG с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами. Согласно определенным аспектам, антитело к NGF или его фрагмент может содержать LCDR1 с аминокислотной последовательностью SEQ ID NO: 91 как таковой или с аминокислотной последовательностью SEQ ID NO: 91 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать LCDR2 с аминокислотной последовательностью SEQ ID NO: 92 как таковой или с аминокислотной последовательностью SEQ ID NO: 92 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами. Аналогичным образом, антитело к NGF или его фрагмент может содержать LCDR3 с аминокислотной последовательностью SEQ ID NO: 93 как таковой или с аминокислотной последовательностью SEQ ID NO: 93 с одной или более, например, одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 24. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH SEQ ID NO: 24.
Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 26. Согласно определенным аспектам, антитело к NGF или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL SEQ ID NO: 26.
Многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенная согласно изобретению, может содержать полноразмерное антитело к NGF, т.е. антитело, содержащее две полноразмерные тяжелые цепи и две полноразмерные легкие цепи в формате H2L2. Если антитело к NGF представляет собой полноразмерное антитело, то один или более чем один домен антагониста TNFα может быть соединен с N-концом или С-концом одной или более тяжелых цепей антитела к NGF или с N-концом или С-концом одной или более легких цепей антитела к NGF. Альтернативно, многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенная согласно изобретению, может содержать антиген-связывающий фрагмент антитела к NGF. Согласно определенным аспектам, фрагмент антитела к NGF может содержать любую часть константных доменов антитела или может содержать только вариабельные домены. Типичные фрагменты антитела к NGF для включения в бифункциональный полипептид, например, мультиспецифическую связывающую молекулу, включают, но не ограничиваются этим, Fab-фрагменты, Fab'-фрагменты, F(ab)2-фрагменты или одноцепочечные Fv (scFv)-фрагменты.
В некоторых типичных композициях, предложенных в данном изобретении, антитело к NGF представляет собой scFv-фрагмент, например scFv-фрагмент MEDI-578 или его NGF-связывающий вариант. В некоторых типичных композициях, предложенных в данном изобретении, антитело к NGF представляет собой scFv-фрагмент, например, scFv-фрагмент NGF-NG или его NGF-связывающий вариант. Полипептид, представляющий собой scFv-фрагмент антитела к NGF, может содержать VH- и VL-домены в любом порядке, либо N-VH-VL-C, либо N-VL-VH-C. Молекулы scFv обычно конструируют таким образом, чтобы VH- и VL-домены были соединены через гибкий линкер. Типичные структуры scFv, в том числе различные линкеры, можно найти в работе Dimasi, N., et al., J. Mol. Biol., 393: 672-92 (2009) и в публикации PCT под № WO 2013/070565, обе из которых включены в данное описание посредством ссылки во всей своей полноте. Как очевидно специалисту в данной области техники, scFv-фрагменты антитела могут иметь пониженную стабильность по сравнению с вариабельными доменами, существующими в стандартной Fab-конформации. Согласно определенным аспектам, scFv-фрагмент можно структурно стабилизировать путем введения стабилизирующих мутаций или путем введения межцепочечной(ых) дисульфидной(ых) связи(ей) (например, SS-стабилизировать). Однако необходимости в стабилизирующих мутациях и/или во введенной межцепочечной дисульфидной связи нет, и согласно определенным аспектам, они не присутствуют. Для стабилизации полипептидов scFv доступен целый ряд признанных в данной области техники способов.
Линкеры можно использовать для соединения доменов/областей бифункциональных полипептидов, предложенных согласно изобретению. Линкеры можно использовать для соединения домена антагониста NGF и домена антагониста TNFα в бифункциональной молекуле и также можно использовать для взаимного связывания вариабельных областей тяжелой и легкой цепей scFv. Типичным неограничивающим примером линкера является полипептидная цепь, содержащая по меньшей мере 4 остатка. Части таких линкеров могут быть гибкими, гидрофильными и иметь малое количество или вообще не иметь своей собственной вторичной структуры (части линкеров или части гибких линкеров). Линкеры, состоящие по меньшей мере из 4 аминокислот, можно использовать для соединения доменов и/или областей, которые расположены близко друг к другу после сборки молекулы бифункционального полипептида. Также можно использовать более длинные линкеры. Так, линкеры могут состоять из примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50 остатков. Кроме того, линкеры могут состоять, например, из приблизительно 100-175 остатков. В том случае, когда для взаимного связывания частей молекулы бифункционального полипептида используют многочисленные линкеры, такие линкеры могут быть одинаковыми или разными (например, одинаковой и разной длины и/или иметь одинаковую и разную аминокислотную последовательность).
Линкер(ы) в молекуле бифункционального полипептида способствуют образованию желаемой структуры. Линкеры могут содержать остатки (Gly-Ser)n (где n представляет собой целое число, составляющее по меньшей мере 1, 2 и до, например, 3, 4, 5, 6, 10, 20, 50, 100 или более), при этом некоторые остатки Glu или Lys распределены по всей длине для увеличения растворимости. Альтернативно, некоторые линкеры совсем не содержат остатков серина, например, в случаях, когда этот линкер является объектом О-гликозилирования. Согласно определенным аспектам, линкеры могут содержать остатки цистеина, например, если димеризацию линкеров используют для придания доменам бифункционального полипептида конфигурации правильно свернутой молекулы. Согласно определенным аспектам, бифункциональный полипептид может содержать по меньшей мере один, два, три, четыре или более полипептидных линкеров, которые соединяют домены этого полипептида.
Согласно некоторым аспектам, полипептидный линкер может содержать 1-50 остатков, 1-25 остатков, 25-50 остатков или 30-50 остатков. Согласно некоторым аспектам, полипептидный линкер может содержать часть группировки Fc. Например, согласно определенным аспектам, полипептидный линкер может содержать часть иммуноглобулинового шарнирного домена антитела IgG1, IgG2, IgG3 и/или IgG4 или его варианта.
Согласно некоторым аспектам, полипептидный линкер может представлять собой gly-ser-линкер или состоять из него. Использованный в данном описании термин «gly-ser-линкер» относится к пептиду, который состоит из остатков глицина и серина. Типичный gly-ser-линкер имеет аминокислотную последовательность формулы (Gly4Ser)n, где n представляет собой целое число, составляющее по меньшей мере 1, 2 и до, например, 3, 4, 5, 6, 10, 20, 50, 100 или более. Согласно определенным аспектам, полипептидный линкер может содержать по меньшей мере часть шарнирной области (например, происходящей из молекулы IgG1, IgG2, IgG3 или IgG4) и целый ряд остатков аминокислот gly-ser (например, gly-ser-линкер, такой как (Gly4Ser)n).
Если многофункциональный полипептид, например, мультиспецифическая связывающая молекула, содержит scFv, то гибкий линкер может соединять тяжелую и легкую цепи scFv. Такой гибкий линкер обычно не включает часть шарнирной области, а скорее представляет собой gly-ser-линкер или другой гибкий линкер. Длину и аминокислотную последовательность гибкого линкера, соединяющего друг с другом домены scFv, можно легко подобрать и оптимизировать.
Согласно определенным аспектам, многофункциональный полипептид, например, мультиспецифическая связывающая молекула, может содержать scFv-фрагмент антитела к NGF, который содержит, в направлении от N-конца к С-концу, VH, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3, и VL. В некоторых воплощениях линкер, соединяющий VH и VL scFv, представляет собой линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4. Согласно определенным аспектам, VH содержит аминокислотную последовательность SEQ ID NO: 3. Согласно определенным аспектам, VL содержит аминокислотную последовательность SEQ ID NO: 7. В некоторых воплощениях VH содержит аминокислотную последовательность с любым из SEQ ID NO: 24, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 94 и 96. В некоторых воплощениях VL содержит аминокислотную последовательность с любым из SEQ ID NO: 26, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 95 и 97. Согласно определенным аспектам, VH-домен содержит аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности с любым из SEQ ID NO: 3, 24, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 94 и 96. Согласно определенным аспектам, VL-домен содержит аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности с любым из SEQ ID NO: 7, 26, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 95 и 97.
Согласно другим аспектам, стабильность полипептида можно улучшить, добавляя межцепочечную дисульфидную связь между VH-доменом и VL-доменом путем замены некоторых остатков в VH- и VL-домене на остатки цистеина. См., например, Michaelson, J.S., et al. (2009) MAbs, 1, 128-41; Brinkmann, U., et al., (1993) Proc. Natl. Acad. Sci. USA, 90, 7538-42; Young, N.M., et al., (1995) FEBS Lett., 377, 135-9. Например, остаток глицина в положениях 100, 101 или 102 VL может быть изменен на остаток цистеина, и остаток глицина в положении 44 VH может быть изменен на остаток цистеина.
Многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенная согласно изобретению, может включать домен антагониста TNFα. Согласно определенным аспектам, домен антагониста TNFα может ингибировать связывание TNFα с рецептором TNF (TNFR) на поверхности клеток, блокируя тем самым активность TNF.
Согласно определенным аспектам, домен антагониста TNFα многофункционального полипептида, предложенного согласно изобретению, представляет собой антитело к TNFα или его антиген-связывающий фрагмент. Согласно определенным аспектам, антитело к TNFα представляет собой инфликсимаб, адалимумаб, цертолизумаба пегол, голимумаб или антиген-связывающий фрагмент любого из этих антител.
Согласно определенным аспектам, антитело к TNFα или его фрагмент связывается с одним и тем же эпитопом, может конкурентно ингибировать или может связываться с TNFα в более высокой аффинностью, чем любое из антител к TNFα инфликсимаб, адалимумаб, цертолизумаба пегол или голимумаб или антиген-связывающий фрагмент любого из этих антител. Согласно определенным аспектам, антитело к TNFα представляет собой инфликсимаб, адалимумаб, цертолизумаба пегол или голимумаб или антиген-связывающий фрагмент любого из этих антител. Информация о структуре и последовательностях таких антител к TNFα общедоступна для специалиста в данной области, и они могут быть включены в многофункциональный полипептид, например, мультиспецифическую связывающую молекулу, описанную в данной заявке, без излишнего экспериментирования. Антитело к TNFα или его фрагмент, включенное(ый) в многофункциональный полипептид, может быть, например, гуманизированным, химерным, приматизированным или полностью человеческим.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий домены HCDR1, HCDR2 и HCDR3 инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба либо вариантов CDR тяжелой цепи инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VL-домен антитела, содержащий домены LCDR1, LCDR2 и LCDR3 инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба либо варианты CDR легкой цепи инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба с одной, двумя, тремя, четырьмя, пятью или более аминокислотными заменами, например, консервативными аминокислотными заменами.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности VH инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба. Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности VL инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба. Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 28. Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VH-домен антитела, содержащий аминокислотную последовательность VH SEQ ID NO: 28.
Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 29. Согласно определенным аспектам, антитело к TNFα или его фрагмент содержит VL-домен антитела, содержащий аминокислотную последовательность VL SEQ ID NO: 29.
Многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенная согласно изобретению, может содержать полноразмерное антитело к TNFα, т.е. антитело, содержащее две полноразмерные тяжелые цепи и две полноразмерные легкие цепи в формате H2L2. Если антитело к TNFα представляет собой полноразмерное антитело, то один или более чем один домен антагониста NGF может быть соединен с N-концом или С-концом одной или более тяжелых цепей антитела к TNFα или с N-концом или С-концом одной или более легких цепей антитела к TNFα. Альтернативно, многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенная согласно изобретению, может содержать антиген-связывающий фрагмент антитела к TNFα. Согласно определенным аспектам, фрагмент антитела к TNFα может содержать любую часть константных доменов антитела или может содержать только вариабельные домены. Типичные фрагменты антитела к TNFα для включения в многофункциональный полипептид, например, мультиспецифическую связывающую молекулу, включают, но не ограничиваются этим, Fab-фрагменты, Fab'-фрагменты, F(ab)2-фрагменты или одноцепочечные Fv (scFv)-фрагменты.
Согласно определенным аспектам, многофункциональной молекулой является ндимаб varB, который представляет собой молекулу, содержащую полноразмерное антитело к TNFα, т.е. антитело, содержащее две полноразмерные тяжелые цепи и две полноразмерные легкие цепи в формате H2L2, с scFv-фрагментом MEDI-578, слитым с С-концом тяжелой цепи антитела к TNFα. Ндимаб varB содержит легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 20, и тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 22. Согласно определенным аспектам, бифункциональная молекула содержит легкую цепь, содержащую аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% последовательности SEQ ID NO: 20, и тяжелую цепь, содержащую аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% последовательности SEQ ID NO: 22.
Согласно определенным аспектам, антитело к TNFα представляет собой scFv-фрагмент, например scFv-фрагмент, происходящий из инфликсимаба, адалимумаба, цертолизумаба пегола или голимумаба, либо его TNFα-связывающий вариант. Полипептид, представляющий собой scFv-фрагмент антитела к TNFα, может содержать VH- и VL-домены в любом порядке, либо N-VH-VL-C, либо N-VL-VH-C. Молекулы scFv обычно конструируют таким образом, чтобы VH- и VL-домены были соединены через гибкий линкер и могли приобретать целый ряд разных структур, описанных выше. Полипептид, представляющий собой scFv-фрагмент антитела к TNFα, может быть стабилизирован так же, как описано выше.
Согласно определенным аспектам, антагонист TNFα представляет собой TNFα-связывающий растворимый фрагмент рецептора TNF, например, TNFR-1 или TNFR-2, либо их варианта или их растворимого фрагмента. Согласно определенным аспектам, растворимый фрагмент TNFR-1 представляет собой фрагмент с молекулярной массой 55 кДа. В некоторых воплощениях растворимый фрагмент TNFR-2 представляет собой фрагмент с молекулярной массой 75 кДа. Согласно определенным аспектам, фрагмент рецептора TNF соединен с гетерологичным полипептидом, например, Fc-фрагментом иммуноглобулина, например, Fc-доменом IgG1. Согласно определенным аспектам, антагонист TNFα содержит аминокислотную последовательность, приведенную в SEQ ID NO: 13, или ее TNFα-связывающий фрагмент. Относящаяся к TNFR-2 часть содержит аминокислоты 1-235 из SEQ ID NO: 13. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотам 1-235 из SEQ ID NO: 13. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит аминокислоты 1-235 из SEQ ID NO: 13, за исключением, например, 1, 2, 3, 4, 5, 10, 20, 20, 40 или 50 аминокислотных вставок, замен или делеций. Fc-часть IgG1 содержит аминокислоты 236-467 из SEQ ID NO: 13. Согласно определенным аспектам, TNFα-связывающий растворимый фрагмент TNFR-2 может быть соединен с Fc-частью любого человеческого или не являющегося человеческим антитела либо с любым другим веществом белкового или небелкового происхождения, которое будет обеспечивать стабильность, например, альбумином или полиэтиленгликолем. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотам 236-467 из SEQ ID NO: 13. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит аминокислоты 236-467 из SEQ ID NO: 13, за исключением, например, 1, 2, 3, 4, 5, 10, 20, 20, 40 или 50 аминокислотных вставок, замен или делеций. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% последовательности SEQ ID NO: 13. Согласно определенным аспектам, вариант TNFα-связывающего растворимого фрагмента TNFR-2 содержит последовательность SEQ ID NO: 13, за исключением, например, 1, 2, 3, 4, 5, 10, 20, 20, 40 или 50 аминокислотных вставок, замен или делеций.
Согласно определенным аспектам, TNFα-связывающий растворимый фрагмент TNFR-2 представляет собой одноцепочечный слитый белок. Согласно определенным аспектам, TNFα-связывающий растворимый фрагмент TNFR-2 представляет собой димер двух слитых белков, образующих ассоциат, например, посредством дисульфидных связей между двумя Fc-доменами.
Многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенный в данном изобретении, может принимать целый ряд разнообразных структур и конформаций. Согласно одному из аспектов многофункциональный полипептид, предложенный в данном изобретении, содержит слитый белок, где домен антагониста NGF, описанный выше, соединен с доменом антагониста TNFα, описанным выше, через гибкий линкер. Примеры линкеров описаны в другом месте данной заявки. Согласно определенным аспектам, многофункциональный полипептид содержит гомодимер слитого белка.
Согласно типичному аспекту, предложен многофункциональный полипептид, в котором антагонист NGF представляет собой scFv-домен антитела к NGF, происходящий, например, из MEDI-578, а антагонист TNFα представляет собой растворимый TNFα-связывающий фрагмент TNFR-2, слитый по своему карбоксиконцу с Fc-доменом иммуноглобулина. scFv-домен антитела к NGF может быть, согласно определенным аспектам, соединен с карбокси концом Fc-домена иммуноглобулина через линкер. Согласно определенным аспектам, мономеры этого многофункционального полипептида образуют гомодимер, при этом каждая субъединица содержит, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент TNFR-2 с молекулярной массой 75 кДа, Fc-домен IgG1 человека, линкер, состоящий из 10 аминокислот (GGGGS)2 (SEQ ID NO: 98), VH-домен антитела к NGF, содержащий аминокислотную последовательность SEQ ID NO: 3, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15), и VL-домен антитела к NGF, содержащий аминокислотную последовательность SEQ ID NO: 7. Согласно одному из аспектов многофункциональный полипептид представляет собой TNFR2-Fc_VH#4, который содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность SEQ ID NO: 14. Согласно определенным аспектам, многофункциональный полипептид содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% последовательности SEQ ID NO: 14.
Согласно другому типичному аспекту многофункциональный полипептид содержит, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент TNFR-2 с молекулярной массой 75 кДа, Fc-домен IgG1 человека, линкер, состоящий из 10 аминокислот (GGGGS)2 (SEQ ID NO: 98), VH-домен антитела к NGF, содержащий аминокислотную последовательность SEQ ID NO: 94, линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4 (SEQ ID NO: 19), и VL-домен антитела к NGF, содержащий аминокислотную последовательность SEQ ID NO: 95. Согласно определенным аспектам, многофункциональный полипептид представляет собой TNFR2-Fc_varB, который содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность SEQ ID NO: 17. Согласно определенным аспектам, многофункциональный полипептид содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% последовательности SEQ ID NO: 17. Полипептиды
Полипептиды, предложенные в данном изобретении, например, многофункциональные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, или отдельные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, соответственно, могут представлять собой рекомбинантные полипептиды, происходящие из природных полипептидов, или синтетические полипептиды. В данной области техники считается признанным фактом, что некоторые аминокислотные последовательности могут иметь вариации без значительного влияния на структуру или функцию белка. Таким образом, согласно изобретению дополнительно предложены варианты полипептидов, предложенных согласно изобретению, которые обладают значительной активностью или которые включают домены антагониста NGF и домены антагониста TNFα. Такие мутантные формы включают делеций, вставки, инверсии, повторы и замены.
Полипептиды и аналоги могут быть дополнительно модифицированы с возможностью содержания дополнительных химических группировок, обычно не составляющих часть белка. Эти модифицированные группировки могут улучшать растворимость, период биологического полувыведения или всасывание белка. Данные группировки также могут ослаблять или устранять какие-либо нежелательные побочные эффекты белков и тому подобное. Обзор таких группировок можно найти в REMINGTON'S PHARMACEUTICAL SCIENCES, 20th ed., Mack Publishing Co., Easton, PA (2000).
Согласно определенным аспектам, многофункциональный полипептид, предложенный согласно изобретению, может включать связывающий NGF или TNFα домен, который не является антителом. В данной области техники известен ряд способов идентификации и получения не являющихся антителами полипептидов, которые связываются с высокой аффинностью с белком-мишенью. См., например, работы Skerra, Curr. Opin. Biotechnol., 18: 295-304 (2007), Hosse et al., Protein Science, 15: 14-27 (2006), Gill et al., Curr. Opin. Biotechnol., 17: 653-658 (2006), Nygren, FEBS J., 275: 2668-76 (2008) и Skerra, FEBS J., 275: 2677-83 (2008), каждая из которых включена в данное описание посредством ссылки во всей своей полноте. Согласно определенным аспектам, для идентификации/получения подходящих многофункциональных полипептидов можно использовать метод фагового дисплея. Согласно определенным аспектам, многофункциональный полипептид, например, мультиспецифическая связывающая молекула, предложенный в данном изобретении, может содержать белковый каркас типа, выбранного из группы, состоящей из белка А, липокалина, домена фибронектина, домена анкиринового консенсусного повтора и тиоредоксина.
Полинуклеотиды, векторы и клетки хозяина
Согласно данному изобретению предложены молекулы нуклеиновой кислоты, содержащие полинуклеотиды, которые кодируют многофункциональный полипептид, содержащий домен антагониста NGF и домен антагониста TNFα. Согласно данному изобретению также предложены молекулы нуклеиновой кислоты, содержащие полинуклеотиды, которые кодируют отдельные полипептиды, содержащие, соответственно, антагонист NGF и антагонист TNFα. Согласно определенным аспектам, такие полинуклеотиды кодируют пептидный домен, который специфически связывается с NGF или его фрагментом и также связывается с TNFα или его фрагментом. Например, согласно данному изобретению предложен полинуклеотид, который кодирует домен полипептида, содержащий антитело к NGF или его антиген-связывающий фрагмент, и домен полипептида, содержащий антагонист TNFα, такой как антитело к TNFα или его антиген-связывающий фрагмент, или растворимую TNFα-связывающую часть рецептора TNF, например, TNFR2. Полинуклеотиды могут быть представлены в форме РНК или в форме ДНК. ДНК включает кДНК, геномную ДНК и синтетическую ДНК; и может быть двухцепочечной или одноцепочечной, и в случае одноцепочечной может представлять собой кодирующую цепь или некодирующую (антисмысловую) цепь.
В некоторых воплощениях выделенный полинуклеотид, который кодирует многофункциональный полипептид, описанный в данной заявке, содержит нуклеотидную последовательность SEQ ID NO: 16, 18 или 99 или ее фрагменты или последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% последовательности SEQ ID NO: 16, 18 или 99 или ее фрагментам.
Выделенные полипептиды, описанные в данной заявке, могут быть получены любым подходящим способом, известным в данной области техники. Такие способы варьируют от методов прямого белкового синтеза до конструирования последовательности ДНК, кодирующей последовательности выделенных полипептидов, и экспрессии этих последовательностей в подходящем трансфицированном хозяине. Согласно определенным аспектам, последовательность ДНК конструируют с использованием рекомбинантной технологии посредством выделения или синтеза последовательности ДНК, кодирующей многофункциональный полипептид, содержащий домен антагониста NGF и домен антагониста TNFα, или отдельные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, соответственно. Соответственно, согласно данному изобретению предложен выделенный полинуклеотид, который кодирует бифункциональный полипептид, содержащий домен антагониста NGF и домен антагониста TNFα, как описано подробно выше. Также предложены выделенные полинуклеотиды, которые кодируют отдельные полипептиды, содержащие, соответственно, домен антагониста NGF и домен антагониста TNFα.
Согласно определенным аспектам, последовательность ДНК, кодирующая многофункциональный полипептид, например, представляющую интерес мультиспецифическую связывающую молекулу или отдельные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, соответственно, может быть сконструирована путем химического синтеза с использованием синтезатора олигонуклеотидов. Такие олигонуклеотиды могут быть сконструированы на основании аминокислотной последовательности желаемого многофункционального полипептида и выбора тех кодонов, которые являются предпочтительными для клетки хозяина, в которой будет продуцироваться представляющий интерес рекомбинантный полипептид. Для синтеза последовательности выделенного полинуклеотида, кодирующей представляющий интерес многофункциональный полипептид, могут быть применены стандартные методы. Например, для конструирования гена с возможной последовательностью, восстановленной по полипептиду, можно использовать полную аминокислотную последовательность. Кроме того, можно синтезировать олигомер ДНК, содержащий нуклеотидную последовательность, кодирующую конкретный многофункциональный полипептид или отдельные полипептиды. Например, можно синтезировать и затем лигировать несколько небольших олигонуклеотидов, кодирующих части желаемого полипептида. Обычно отдельные олигонуклеотиды содержат «липкие» 5'- или 3'-концы для комплементарной сборки.
Согласно определенным аспектам, полинуклеотиды, предложенные в данном изобретении, могут содержать кодирующую зрелый полипептид последовательность, слитую в одной и той же рамке считывания с маркерной последовательностью, что способствует, например, очистке кодируемого полипептида. Например, маркерная последовательность может представлять собой гекса-гистидиновую метку, доставляемую вектором pQE-9, для осуществления очистки зрелого полипептида, слитого с маркером в случае использования бактериального хозяина, или маркерная последовательность может представлять собой гемагглютининовую (НА) метку, происходящую из белка гемагглютинина вируса гриппа, в случае использования хозяина, выделенного из млекопитающего (например, клеток COS-7).
Полинуклеотиды, предложенные в настоящем изобретении, могут дополнительно содержать изменения в кодирующих областях, некодирующих областях или и там, и там. Согласно определенным аспектам, полинуклеотидные варианты содержат изменения, приводящие к молчащим заменам, вставкам или делециям, но не изменяют свойств или активностей кодируемого полипептида. Согласно определенным аспектам, нуклеотидные варианты получают благодаря присутствию молчащих замен вследствие вырожденности генетического кода. Полинуклеотидные варианты могут быть получены по разным причинам, например, с целью оптимизации кодонов для экспрессии в конкретном хозяине (чтобы изменить кодоны в мРНК человека до таких, которые будут предпочтительны для бактериального хозяина, такого как Е. coli).
Также предложены векторы и клетки, содержащие полинуклеотиды, описанные в данной заявке. После сборки (путем синтеза, сайт-специфического мутагенеза или другого метода) полинуклеотидные последовательности, кодирующие конкретный выделенный представляющий интерес полипептид, могут быть встроены в экспрессирующий вектор и функционально связаны с регулирующей экспрессию последовательностью, пригодной для экспрессии белка в желаемом хозяине. Такие векторы предложены согласно данному изобретению. Секвенирование нуклеиновой кислоты, рестрикционное картирование и экспрессия биологически активного полипептида в подходящем хозяине может подтвердить надлежащую сборку. Как хорошо известно в данной области техники, чтобы достичь высоких уровней экспрессии трансфицированного гена в хозяине, данный ген должен быть функционально связан с последовательностями, регулирующими экспрессию на уровне транскрипции и трансляции, которые функционируют в выбранном для экспрессии хозяине.
Согласно определенным аспектам, рекомбинантные экспрессирующие векторы можно использовать для амплификации и экспрессии ДНК, кодирующей многофункциональные полипептиды, например, мультиспецифические связывающие молекулы, содержащие домен антагониста NGF и домен антагониста TNFα, или отдельные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, соответственно. Рекомбинантные экспрессирующие векторы представляют собой способные реплицироваться конструкции ДНК, которые имеют синтетические или происходящие из кДНК фрагменты ДНК, кодирующие многофункциональный полипептид или отдельные полипептиды, содержащие домен антагониста NGF и домен антагониста TNFα, соответственно, функционально связанные с подходящими транскрипционными или трансляционными регуляторными элементами, происходящими из генов млекопитающих, микробов, вирусов или насекомых. Транскрипционная единица обычно содержит совокупность (1) генетического(их) элемента или элементов, играющего(их) регуляторную роль в генной экспрессии, например, транскрипционных промоторов или энхансеров, (2) структурной или кодирующей последовательности, которая транскрибируется в мРНК и транслируется в белок, и (3) соответствующих последовательностей инициации и терминации транскрипции и трансляции, как описано подробно ниже. Такие регуляторные элементы могут включать последовательность оператора для регуляции транскрипции. Помимо этого могут быть включены ориджин репликации, обычно обеспечивающий способность к репликации в хозяине, и ген, способствующий распознаванию трансформантов при селекции. Области ДНК считаются функционально связанными, когда они соотносятся друг с другом по функциям. Например, ДНК для сигнального пептида (лидерной последовательности секретируемого пептида) функционально связана с ДНК для полипептида, если он экспрессируется в виде предшественника, который принимает участие в секреции этого полипептида; промотор функционально связан с кодирующей последовательностью, если он регулирует транскрипцию данной последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен таким образом, чтобы дать возможность осуществиться трансляции. Структурные элементы, предназначенные для применения в экспрессирующих системах дрожжей, включают лидерную последовательность, обеспечивающую секрецию транслированного белка клеткой хозяина во внешнюю среду. Альтернативно, если рекомбинантный белок экспрессируется без лидерной или необходимой для транспорта последовательности, то он может включать остаток метионина на N-конце. Этот остаток впоследствии возможно может быть отщеплен от экспрессированного рекомбинантного белка с получением конечного продукта.
Выбор регулирующих экспрессию последовательностей и экспрессирующего вектора будет зависеть от выбора хозяина. Можно использовать широкий ряд комбинаций экспрессирующий хозяин/вектор. Экспрессирующие векторы, полезные для эукариотических хозяев, включают, например, векторы, содержащие регулирующие экспрессию последовательности из обезьянего вируса 40 (SV40), бычьего папилломавируса, аденовируса и цитомегаловируса. Экспрессирующие векторы, полезные для бактериальных хозяев, включают известные бактериальные плазмиды, такие как плазмиды из Е. coli, в том числе pCR 1, pBR322, рМВ9 и их производные, плазмиды для широкого диапазона хозяев, как например, плазмиды на основе М13 и содержащих одноцепочечную ДНК нитчатых бактериофагов.
Согласно данному изобретению также предложены клетки хозяина, содержащие полинуклеотиды, кодирующие полипептиды, предложенные согласно изобретению. Клетки хозяина, подходящие для экспрессии полипептидов, предложенных согласно изобретению, включают прокариот, клетки дрожжей, насекомых или высших эукариот под контролем соответствующих промоторов. Прокариоты включают грамотрицательные или грамположительные микроорганизмы, например Е. coli или бациллы. Клетки высших эукариот включают общеизвестные клеточные линии, происходящие из млекопитающих, описанные ниже. Также можно использовать бесклеточные трансляционные системы. Соответствующие клонирующие и экспрессирующие векторы для применения вместе с хозяевами - клетками бактерий, грибов, дрожжей и млекопитающих - описаны в работе Pouwels и др. (Cloning Vectors: А Laboratory Manual, Elsevier, N.Y., 1985), релевантное описание которой тем самым включено посредством ссылки. Дополнительную информацию, касающуюся способов получения белков, в том числе получения антител, можно найти, например, в публикации заявки на патент США №2008/0187954, патентах США №№6413746 и 6660501 и публикации заявки на международный патент № WO 04009823; все они тем самым включены в данное описание посредством ссылки во всей своей полноте.
Для экспрессии рекомбинантного белка также могут быть преимущественно использованы различные системы культур клеток млекопитающих или насекомых. Экспрессия рекомбинантных белков в клетках млекопитающих может быть осуществлена благодаря тому, что в них такие белки, как правило, являются правильно свернутыми, соответствующим образом модифицированными и полностью функциональными. Примеры подходящих в качестве хозяина линий клеток млекопитающих включают HEK-293 и HEK-293T, линии COS-7 клеток почки мартышки, описанные Gluzman (Cell, 23: 175, 1981) и другие клеточные линии, включая, например, L клетки, С127, 3Т3, линии клеток яичника китайского хомячка (СНО), HeLa и почки новорожденного сирийского хомячка (BHK). Векторы для экспрессии в клетках млекопитающих могут содержать нетранскрибируемые элементы, такие как ориджин репликации, подходящий промотор и энхансер, соединенные с подлежащим экспрессии геном, и другие 5'- или 3'-фланкирующие нетранскрибируемые последовательности и 5'- или 3'-нетранслируемые последовательности, такие как необходимые сайты связывания рибосомы, сайт полиаденилирования, донорный и акцепторный сайты сплайсинга и последовательности терминации транскрипции. Обзор бакуловирусных систем, используемых для получения гетерологичных белков в клетках насекомых, приведен в Luckow and Summers, Bio/Technology, 6: 47 (1988).
Согласно данному изобретению также предложен способ получения многофункционального полипептида, описанного в данной заявке, или получения отдельных полипептидов, содержащих, соответственно, антагонист NGF и антагонист TNFα. Способ включает культивирование клетки хозяина, описанной выше, в условиях, стимулирующих экспрессию многофункционального полипептида или отдельных полипептидов, и извлечение многофункционального полипептида или отдельных полипептидов.
Для долгосрочного продуцирования рекомбинантных белков с высоким выходом целесообразна стабильная экспрессия. Например, можно сконструировать клеточные линии, которые стабильно экспрессируют многофункциональный полипептид. Вместо того, чтобы использовать экспрессирующие векторы, которые содержат вирусные ориджины репликации, клетки хозяина могут быть трансфицированы ДНК под контролем соответствующих регулирующих экспрессию элементов (например, промотора, энхансера, последовательностей, терминаторов транскрипции, сайтов полиаденилирования и т.д.) и ДНК, кодирующей селектируемый маркер. После введения чужеродной ДНК сконструированные клетки могут быть оставлены расти в течение 1-2 суток в обогащенной среде и затем переведены в селективную среду. Присутствующий в рекомбинантной плазмиде селектируемый маркер придает устойчивость при селекции и способствует тому, что клетки начинают стабильно интегрировать эту плазмиду в свои хромосомы и расти с образованием бляшек, которые в свою очередь могут быть клонированы и распространены в клеточные линии. Этот метод можно использовать для конструирования клеточных линий, которые экспрессируют многофункциональный полипептид.
В некоторых воплощениях многофункциональные полипептиды, представленные в данном описании, экспрессированы в клеточной линии, характеризующейся временной экспрессией многофункционального полипептида. Временная трансфекция представляет собой процесс, при котором нуклеиновая кислота, введенная в клетку, не интегрируется в геномную или хромосомную ДНК этой клетки, а поддерживается в клетке в виде внехромосомного элемента, например в виде эписомы. Процессы транскрипции нуклеиновой кислоты этой эписомы не затрагиваются, и продуцируется белок, кодируемый нуклеиновой кислотой этой эписомы.
Клеточную линию, или стабильно, или временно трансфицированную, поддерживают в среде для культивирования клеток и в условиях, известных в данной области техники, приводящих к экспрессии и продуцированию полипептидов. В некоторых воплощениях за основу среды для культивирования клеток млекопитающих взяты составы имеющихся в продаже сред, включая, например, модифицированную Дульбекко среду Игла (DMEM) или питательную смесь Хэма F12. В некоторых воплощениях, составы среды для культивирования клеток модифицированы с целью содействия усилению и клеточного роста, и экспрессии белка в биологической системе. Использованные в данном описании термины «среда для культивирования клеток», «культуральная среда» и «состав среды» относятся к питательному раствору для поддержания, роста, размножения или распространения клеток в искусственном окружении in vitro вне многоклеточного организма или ткани. Среду для культивирования клеток можно оптимизировать для конкретного применения этой клеточной культуры, включая, например, среду для роста клеточных культур, которую готовят для стимулирования роста клеток, или среду для продуцирования из клеточных культур, которую готовят для стимулирования продуцирования рекомбинантного белка. Термины «питательное вещество», «ингредиент» и «компонент» могут быть использованы взаимозаменяемо и относятся к элементам, которые входят в состав среды для культивирования клеток.
В различных воплощениях клеточные линии поддерживают, используя метод периодического культивирования с подпиткой. Как использовано в данном описании, «метод периодического культивирования с подпиткой» относится к методу, согласно которому в подпитываемую клеточную культуру подают дополнительные питательные вещества после того, как первоначально проводят инкубацию в основной питательной среде. Например, метод периодического культивирования с подпиткой может включать добавление поддерживающей среды согласно определенному режиму подпитки в пределах заданного периода времени. Так, «подпитываемая клеточная культура» относится к клеточной культуре, где клетки, обычно клетки млекопитающих, и культуральную среду сначала загружают в сосуд для культивирования и в процессе культивирования в данную культуру подают дополнительные питательные вещества для культивирования, непрерывно или ступенчато, с периодическим отведением клеток и/или продукта, или без такого отведения, до момента окончания культивирования.
В некоторых воплощениях среда для культивирования клеток содержит основную питательную среду и по меньшей мере один гидролизат, например, гидролизат сои, гидролизат дрожжей или комбинацию этих двух типов гидролизатов, в результате чего получается модифицированная основная питательная среда. Дополнительные питательные вещества иногда могут включать только основную питательную среду, такую как концентрированная основная питательная среда, или могут включать только гидролизаты или концентрированные гидролизаты. Подходящие основные питательные среды включают, но не ограничиваются этим, модифицированную Дульбекко среду Игла (DMEM), DME/F12, минимальную поддерживающую среду (MEM), основную питательную среду Игла (ВМЕ), среду 1640 от Мемориального института Розуэлла Парка (RPMI), F-10, F-12, минимальную поддерживающую среду в альфа-модификации (альфа-MEM), минимальную поддерживающую среду Глазго (G-MEM), PF СНО (см., например, безбелковую (PF) среду для клеток СНО (Sigma) или бессывороточную среду EX-CELL™ 325 PF СНО для безбелковой среды для клеток СНО (SAFC Bioscience)) и модифицированную по методу Исков среду Дульбекко. Другие примеры основных питательных сред, которые могут быть использованы в технологическом процессе по данному изобретению, включают основную питательную среду ВМЕ (GibcoInvitrogen), модифицированную Дульбекко среду Игла (DMEM, порошок) (GibcoInvitrogen (№31600)).
В некоторых воплощениях основная питательная среда может представлять собой бессывороточную среду, и это означает, что данная среда не содержит никакой сыворотки (например, фетальной телячьей сыворотки (FBS), лошадиной сыворотки, козьей сыворотки или сыворотки, происходящей из любого другого животного, известной специалисту в данной области техники), либо представлять собой среды, не содержащие белков животного происхождения, или среды с заданным химическим составом.
Основную питательную среду можно модифицировать, чтобы исключить некоторые не являющиеся питательными компоненты, имеющиеся в стандартной основной питательной среде, такие как буферы на основе различных неорганических и органических соединений, поверхностно-активное(ые) вещество(а) и хлорид натрия. Исключение таких компонентов из основной питательной среды для клеток позволяет добиться увеличения концентрации остальных питательных компонентов и может оптимизировать клеточный рост в целом и экспрессию белков. К тому же, изъятые компоненты можно снова добавить в среду для культивирования клеток, содержащую модифицированную основную питательную среду для клеток, соответствующую требованиям условий культивирования клеток. В некоторых воплощениях среда для культивирования клеток содержит модифицированную основную питательную среду для клеток и по меньшей мере одно из следующих питательных веществ: источник железа, рекомбинантный фактор роста, буфер, поверхностно-активное вещество, регулятор осмолярности, источник энергии и гидролизаты неживотного происхождения. Помимо этого, модифицированная основная питательная среда для клеток возможно может содержать аминокислоты, витамины или комбинацию и аминокислот, и витаминов. В некоторых воплощениях модифицированная основная питательная среда дополнительно содержит глутамин, например, L-глутамин, и/или метотрексат.
В некоторых воплощениях для продуцирования белка в большом количестве используют биореакторный метод, при этом используют методы с применением биореактора периодического действия с подпиткой, биореактора периодического действия, биореактора с постоянной подпиткой или биореактора с непрерывной подачей сырья, известные в данной области техники. Емкость крупномасштабных биореакторов составляет по меньшей мере 50 литров, иногда немногим более 500 литров или от 1000 до 100000 литров. В таких биореакторах могут быть использованы лопастные мешалки для распределения кислорода и питательных веществ. Мелкомасштабные биореакторы, как правило, имеют отношение к культивированию клеток в объеме не более чем приблизительно 100 литров, и объемная емкость может изменяться в диапазоне от примерно 1 литра до примерно 100 литров. Альтернативно, как в случае крупномасштабного, так и мелкомасштабного культивирования можно применять биореакторы для однократного использования (SUB).
Температура, pH, условия перемешивания, аэрирования и плотность инокулята могут варьировать в зависимости от используемых клеток хозяина и подлежащего экспрессии рекомбинантного белка. Например, используемую для экспрессии рекомбинантного белка клеточную культуру можно поддерживать при температуре от 30 до 45 градусов по Цельсию. В процессе культивирования может быть проведен мониторинг pH культуральной среды, чтобы поддерживать значение pH на оптимальном уровне, которое для некоторых клеток хозяина может находиться в диапазоне pH 6,0-8,0. В таких методах культивирования для перемешивания можно использовать перемешивание с помощью лопастного колеса. Скорость вращения такой мешалки может составлять, в терминах окружной скорости конца лопасти, приблизительно 50-200 см/с, однако в зависимости от типа культивируемой клетки хозяина можно использовать другие системы подачи воздуха или другие системы перемешивания/аэрирования, известные в данной области техники. Для поддержания концентрации растворенного кислорода в культуре при насыщении воздухом приблизительно от 20% до 80% обеспечивается достаточное аэрирование, опять-таки в зависимости от выбранной для культивирования клетки хозяина. Альтернативно, в биореакторе может быть предусмотрено барботирование воздуха или кислорода непосредственно в культуральную среду. Существуют и другие методы подачи кислорода, включая использование систем аэрации беспузырьковым потоком, в которых применяются аэраторы с половолоконными мембранами.
Очистка белков
Белки, продуцируемые трансфицированным хозяином, описанным выше, могут быть очищены в соответствии с любым подходящим методом. Такие стандартные методы включают хроматографию (например, ионообменную, аффинную и гель-проникающую колоночную хроматографию (sizing column chromatography)), центрифугирование, использование различий в растворимости или любой другой стандартный метод очистки белка. Для облегчения очистки путем пропускания через соответствующую колонку для аффинной хроматографии к белку можно присоединить аффинные метки, такие как гексагистидин, мальтоза-связывающий домен, последовательность белка оболочки вируса гриппа и глутатион-S-трансфераза. Выделенные белки также можно охарактеризовать с использованием таких физических методов, как протеолиз, ядерный магнитный резонанс и рентгеноструктурная кристаллография.
Например, супернатанты из систем, которые секретируют рекомбинантный белок в культуральные среды, сначала можно сконцентрировать, используя имеющийся в продаже фильтр для концентрирования белков, например, ультрафильтрационную ячейку Amicon или Pellicon от Millipore. После стадии концентрирования концентрат можно нанести на подходящую матрицу для проведения очистки. Альтернативно, можно использовать анионообменную смолу, например, матрицу или носитель, имеющие боковые диэтиламиноэтильные (DEAE) группы. Такими матрицами могут быть матрицы на основе акриламида, агарозы, декстрана, целлюлозы или матрицы других типов, обычно применяемые при очистке белков. В альтернативном варианте можно использовать стадию катионного обмена. Подходящие катионообменники включают различные нерастворимые матрицы, содержащие сульфопропильные или карбоксиметильные группы. В конце, для дальнейшей очистки NGF-связывающего вещества можно выполнить одну или более чем одну стадию обращенно-фазовой высокоэффективной жидкостной хроматографии (RP-HPLC) с применением гидрофобного носителя для RP-HPLC, например, силикагеля, имеющего боковые метильные или другие алифатические группы. Для получения гомогенного рекомбинантного белка также можно использовать, в различных комбинациях, некоторые или все из вышеупомянутых стадий очистки.
Рекомбинантный белок, продуцируемый бактериальной культурой, можно выделить, например, проводя сначала экстракцию из осадков после центрифугирования клеток, затем выполняя одну или более стадий концентрирования, высаливания, ионообменной или гель-проникающей хроматографии в водных условиях. Для конечных стадий очистки можно использовать высокоэффективную жидкостную хроматографию (HPLC). Микробные клетки, используемые при экспрессии рекомбинантного белка, могут быть разрушены любым удобным методом, включая циклическое замораживание-оттаивание, ультразвуковую обработку, механическое разрушение или применение лизирующих клетки агентов.
Способы, известные в данной области техники для очистки рекомбинантных полипептидов, также включают, например, способы, описанные в публикациях заявок на патент США №2008/0312425, 2008/0177048 и 2009/0187005, каждая из которых тем самым включена в данное описание посредством ссылки во всей своей полноте.
Способы применения и фармацевтические композиции
Согласно данному изобретению предложены способы контролирования или лечения боли у субъекта, включающие введение терапевтически эффективного количества многофункционального полипептида-антагониста TNFα и NGF, например, мультиспецифической связывающей молекулы, предложенной в данном описании, или включающие совместное введение антагониста TNFα и антагониста NGF. Согласно определенным аспектам, таким субъектом является человек.
Согласно данному изобретению также предложены фармацевтические композиции, содержащие многофункциональный полипептид-антагонист TNFα и NGF, например, мультиспецифическую связывающую молекулу, предложенную в данном описании, или содержащие комбинацию антагониста TNFα и антагониста NGF, как предложено в данном описании. Согласно определенным аспектам, фармацевтические композиции дополнительно содержат фармацевтически приемлемый наполнитель. Эти фармацевтические композиции полезны при лечении боли, например, нейропатической и воспалительной (например, обусловленной остео- или ревматоидным артритом) боли.
Многофункциональные полипептиды и композиции, содержащие антагонист NGF и антагонист TNFα, предложенные в данном описании, могут быть полезны для целого ряда применений, включая, но не ограничиваясь этим, контроль или лечение боли, например, нейропатической боли. Способы такого применения могут представлять собой способы in vitro, ex vivo или in vivo.
Согласно определенным аспектам, заболевание, расстройство или состояние, подлежащее лечению NGF-связывающим веществом (например, антителом или полипептидом), ассоциировано с болью. Согласно определенным аспектам, данная боль ассоциирована с хронической ноцицептивной болью, хронической болью в пояснице, нейропатической болью, болью при раковом заболевании, болью при постгерпетической невралгии (PHN) или состояниями висцеральной боли.
Согласно данному изобретению предложен способ контролирования боли у субъекта, включающий введение субъекту, нуждающемуся в контролировании боли, эффективного количества антагониста фактора роста нервов (NGF) и антагониста фактора некроза опухоли (TNFα), при этом такое введение может контролировать боль у субъекта более эффективно, чем введение эквивалентного количества антагониста NGF или антагониста TNFα в виде монотерапии.
Согласно определенным аспектам, введение представляет собой совместное введение антагониста NGF и антагониста TNFα в качестве комбинированной терапии. Как уже обсуждалось в другом месте данного описания, отдельные компоненты можно вводить одновременно или последовательно. Индивидуальным антагонистом NGF или антагонистом TNFα могут быть любые антагонисты NGF или TNFα, предложенные в данном изобретении, например, растворимый NGF-связывающий фрагмент рецептора TrkA, антитело к NGF или его антиген-связывающий фрагмент, растворимый TNFα-связывающий фрагмент рецептора TNF, например, TNFR-2, или антитело к TNFα или его фрагмент, например, инфликсимаб, адалимумаб, цертолизумаба пегол, голимумаб или антиген-связывающий фрагмент любого из этих антител.
При совместном введении можно использовать различные дозы каждого из антагонистов, которые необходимы для контроля боли. Согласно определенным аспектам, при совместном введении можно использовать более низкие дозы каждого компонента или вводить дозы с меньшей частотой, чем при обычном введении в виде терапии отдельными средствами, обеспечивая тем самым дополнительно безопасность, удобство и экономичность.
Согласно определенным аспектам, способ контролирования боли, предложенный в данном изобретении, включает введение многофункционального полипептида, например, мультиспецифической связывающей молекулы, содержащей домен антагониста NGF и домен антагониста TNFα. Типичные многофункциональные полипептиды для применения в этом способе подробно изложены в данном описании. С учетом этого описания среднему специалисту в данной области техники будут очевидны дополнительные многофункциональные полипептиды, полезные для применения в этом способе.
Под «более эффективным» контролированием боли по сравнению с введением компонентов в виде монотерапии понимают, что комбинированное лечение является более эффективным в отношении контролирования боли, чем введение эквивалентных количеств или антагониста NGF, или антагониста TNFα по отдельности. Согласно определенным аспектам, и как описано более подробно ниже, способ контролирования боли, предложенный в данном изобретении, может обеспечить синергическую эффективность, например, эффект от введения и антагониста NGF, и антагониста TNFα может обеспечить более чем аддитивный эффект или такое введение может быть эффективным в тех случаях, когда ни антагонист NGF, ни антагонист TNFα не являются эффективными по отдельности. Согласно определенным аспектам, такое комбинирование может предусматривать щадящий режим дозирования, например, эффективные дозировки отдельных компонентов при совместном введении могут быть меньше эффективных доз и одного, и другого компонента по отдельности.
Согласно определенным аспектам, способ контролирования боли, предложенный в данной заявке, по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% 80%, 90% или 100% более эффективен в отношении контролирования боли у субъекта, чем введение эквивалентного количества антагониста NGF или антагониста TNFα в виде монотерапии. Согласно определенным аспектам, дозировки по отдельности антагониста NGF или антагониста TNFα при совместном введении субъекту могут быть ниже, или величина относительной дозы антагониста NGF или антагониста TNFα, предоставляемая при введении бифункционального полипептида, предложенного согласно изобретению, может быть ниже, например, на 5%, 10%, 20%, 30%, 40%, 50% 60%, 70%, 80% или 90% ниже дозировок, необходимых при введении этих компонентов в виде монотерапии.
Эффективность контролирования боли можно оценить путем опроса пациента с целью классификации характера и интенсивности испытываемой боли в соответствии с целым рядом разных шкал. В случае вербальной шкалы боли для описания используют словесные оценки в таком диапазоне: «отсутствие боли», «слабая боль», «умеренная боль» и «тяжелая боль» с баллами от 0 до 3, присваиваемыми каждому состоянию. Альтернативно, пациент может быть опрошен с целью классификации его боли в соответствии с количественной шкалой оценки боли: от 0 (отсутствие боли) до 10 (самая сильная боль). На вертикальной или горизонтальной линии визуальной аналоговой шкалы (VAS) приведены слова, описывающие боль в показателях от «отсутствия боли» до «максимально возможной боли», и пациентов просят отметить точку на этой линии, которая соответствует имеющемуся у них на данный момент уровню боли. Показатель выраженности боли по Мак-Гиллу позволяет пациентам описать как характер, так и интенсивность боли путем выбора слов, которые наилучшим образом описывают их боль, из набора коротких перечней, например, пульсирующая, жгучая, сдавливающая боль. Другие шкалы оценки боли можно применять для взрослых, которые испытывают трудности при использовании VAS или количественных шкал, например, шкалу оценки боли по выражению лица FACES, или для пациентов с невербальным поведением, например, поведенческую рейтинговую шкалу. Оценить функциональную активность в отношении того, до какой степени испытываемая пациентами боль их обременяет, можно, попросив их выполнить задание, связанное с этой болезненной областью. Улучшение балльной оценки боли, выявленное с использованием этих типов шкалы, теоретически будет указывать на повышение эффективности обезболивающего средства.
Согласно способу контролирования боли, предложенному в данном изобретении, такого введения достаточно для контролирования боли у субъекта, нуждающегося в контролировании боли, например, совместное введение антагониста NGF и антагониста TNFα или введение многофункционального полипептида, например, мультиспецифической связывающей молекулы, содержащей домен антагониста NGF и домен антагониста TNFα, может предотвращать, ослаблять, уменьшать интенсивность или устранять боль у субъекта. Согласно определенным аспектам, боль может представлять собой острую боль, кратковременную боль, постоянную или хроническую ноцицептивную боль либо постоянную или хроническую нейропатическую боль.
Согласно определенным аспектам, композиции готовят для хранения и применения путем объединения многофункционального полипептида-антагониста TNFα и NGF, например, мультиспецифической связывающей молекулы, предложенной согласно изобретению, или комбинации антагониста TNFα и антагониста NGF, предложенной согласно изобретению, с фармацевтически приемлемым наполнителем (например, носителем, эксципиентом) (Remington, The Science and Practice of Pharmacy, 20th Edition, Mack Publishing, 2000). Подходящие фармацевтически приемлемые наполнители включают, но не ограничиваются этим, нетоксичные буферы, как например, буферы на основе фосфорной, лимонной и других органических кислот; соли, такие как хлорид натрия; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (например, октадецилдиметилбензилхлорид аммония; гексаметонийхлорид; бензалконийхлорид; бензетонийхлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и мета-крезол); низкомолекулярные полипептиды (например, содержащие менее чем примерно 10 аминокислотных остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; углеводы, такие как моносахариды, дисахариды, глюкоза, манноза или декстрины; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (EDTA); сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как ион натрия; комплексы с металлами (например, комплексы Zn-белок); и неионные поверхностно-активные вещества, такие как твин или полиэтиленгликоль (ПЭГ).
На основе многофункциональных полипептидов по настоящему изобретению могут быть приготовлены композиции в жидких, полутвердых или твердых формах в зависимости от физико-химических свойств молекулы и пути доставки. Такие композиции могут включать эксципиенты или комбинации эксципиентов, например: сахара, аминокислоты и поверхностно-активные вещества. Жидкие композиции могут включать полипептиды в широком диапазоне концентраций и pH. Твердые композиции могут быть приготовлены путем лиофилизации, распылительной сушки или сушки с использованием, например, сверхкритической жидкостной технологии. В некоторых воплощениях любая из композиций, описанных в данной заявке, представляет собой лиофилизированную композицию.
В специальном воплощении на основе многофункционального полипептида по изобретению готовят композицию в 20 мМ фосфате натрия, 50 мМ L-аргинине-HCL, 150 мМ сахарозе, с 0,03% (масс./об.) полисорбата 80, pH 6,5.
Фармацевтическую композицию, предложенную в данном изобретении, можно вводить любым из ряда путей, используемых либо для местного, либо для системного варианта лечения. Введение может быть местным (например, в слизистые оболочки, включая вагинальную и ректальную доставку), например, посредством трансдермальных пластырей, мазей, лосьонов, кремов, гелей, капель, суппозиториев, спреев, жидкостей и порошков; легочным (например, посредством ингаляции или инсуффляции порошков или аэрозолей, в том числе посредством небулайзера; интратрахеальным, интраназальным, эпидермальным и трансдермальным); пероральным; или парентеральным, включая внутривенную, интраартериальную, подкожную, внутрибрюшинную или внутримышечную инъекцию или инфузию; или интракраниальным (например, интратекальным или интравентрикулярным) введением.
Многофункциональный полипептид-антагонист TNFα и NGF, предложенный согласно данному изобретению, или комбинация антагониста TNFα и антагониста NGF, предложенная согласно данному изобретению, далее могут быть объединены в фармацевтической комбинированной композиции, или в режиме введения в виде комбинированной терапии, со вторым (или третьим) соединением, обладающим антиноцицептивными свойствами.
Что касается лечения боли, то соответствующая дозировка многофункционального полипептида-антагониста TNFα и NGF, например, мультиспецифической связывающей молекулы, предложенной согласно данному изобретению, или комбинации антагониста TNFα и антагониста NGF, предложенной согласно данному изобретению, зависит от типа подлежащей лечению боли, тяжести и источника боли, восприимчивости к боли, вводят ли многофункциональный полипептид или комбинацию полипептидов для терапевтических или профилактических целей, предыдущей терапии, клинической истории пациента и так далее; все это находится на усмотрении лечащего врача. Чтобы поддерживать эффективное контролирование боли, многофункциональный полипептид или комбинацию полипептидов можно вводить один раз или в виде курса лечения продолжительностью от нескольких дней до нескольких месяцев. Оптимальные схемы введения могут быть рассчитаны на основании результатов измерения накопления лекарственного средства в организме пациента и будут варьировать в зависимости от относительной эффективности отдельного антитела или полипептида. Отвечающий за введение врач может легко определить оптимальные дозировки, методологии введения и частоту повторения.
Введение многофункционального полипептида, например, мультиспецифической связывающей молекулы, или терапия комбинацией полипептидов, как предложено в данном изобретении, может обеспечивать «синергизм» и демонстрировать «синергический характер», т.е. эффект, достигаемый при совместном использовании активных ингредиентов, превышает сумму эффектов, получаемых в результате использования этих соединений по отдельности. Синергического эффекта можно достичь, когда: (1) активные ингредиенты вводят в виде единого многофункционального слитого полипептида; (2) на основе этих активных ингредиентов готовят комбинированную стандартную лекарственную композицию, и их вводят или доставляют одновременно в ее составе; (3) активные ингредиенты доставляют путем чередования или в параллели в виде отдельных композиций; или (4) с использованием какого-либо другого режима (введения). При доставке с применением чередующейся терапии синергического эффекта можно достичь, когда соединения вводят или доставляют последовательно, например, посредством разных инъекций с использованием отдельных шприцев. В общем случае, при чередующейся терапии эффективную дозировку каждого активного ингредиента вводят последовательно, тогда как в случае комбинированной терапии эффективные дозировки двух или более активных ингредиентов вводят совместно.
Боль
В самом широком смысле термин «боль» относится к испытываемому симптому, который является весьма субъективным для испытывающего его индивидуума и находится под влиянием психического состояния индивидуума, в том числе окружающей среды и культурного фона. «Физическая» боль обычно может быть связана с раздражителем, ощущаемым извне, который является причиной истинного или возможного повреждения ткани. В этом смысле боль можно рассматривать как «сенсорное и эмоциональное переживание, связанное с истинным или возможным повреждением ткани или описываемое в терминах такого повреждения» согласно Международной ассоциации по изучению боли (IASP). Однако некоторые случаи боли не имеют никакой выявляемой причины. Например, психогенная боль, в том числе обострение ранее испытываемой физической боли под действием психогенных факторов, или синдромы иногда устойчивой, ощущаемой боли у лиц с психологическими расстройствами без каких-либо доказательств выявляемой причины боли. Типы боли
Боль включает ноцицептивную боль, нейропатическую/нейрогенную боль, «прорывную» боль, аллодинию, гипералгезию, гиперестезию, дисестезию, парестезию, гиперпатию, фантомную боль, психогенную боль, болевую анестезию, невралгию, неврит. Другие категории включают боль при злокачественных опухолях, стенокардическую боль и/или идиопатическую боль, комплексный регионарный болевой синдром I, комплексный регионарный болевой синдром II. Типы и симптомы боли не должны быть взаимоисключающими. Подразумевается, что эти термины определены в IASP.
Ноцицептивная боль инициируется в ответ на болевые стимулы специализированными сенсорными ноцицепторами в периферических нервах, преобразующими болевые стимулы в потенциалы действия. Ноцицепторы, как правило, на Aδ-волокнах и (полимодальных) С-волокнах, представляют собой свободные нервные окончания, заканчивающиеся чуть ниже кожи, в сухожилиях, суставах и в органах тела. Нейроны спинномозгового ганглия (DRG) обеспечивают место связи между периферией и спинным мозгом. Этот сигнал обрабатывается при прохождении через спинной мозг к стволу головного мозга и сайтам в таламусе и окончательно к коре головного мозга, где он обычно (но не всегда) вызывает ощущение боли. Ноцицептивная боль может быть вызвана широким спектром химических, тепловых, биологических (например, воспалительных) или механических событий, которые могут приводить к раздражению или повреждению ткани организма, как правило, превышающим определенный минимальный порог интенсивности, необходимой, чтобы вызвать ноцицептивную активность в ноцицепторах.
Нейропатическая боль, как правило, является результатом аномального функционирования в периферической или центральной нервной системе, приводя к возникновению периферической или центральной нейропатической боли, соответственно. Нейропатическая боль согласно IASP определяется как боль, инициируемая или вызываемая первичным поражением или дисфункцией в нервной системе. Нейропатическая боль часто предполагает фактическое повреждение нервной системы, в особенности в хронических случаях. Воспалительная ноцицептивная боль, как правило, является результатом повреждения ткани и возникающего в результате этого воспалительного процесса. Нейропатическая боль может сохраняться значительное время после (например, спустя месяцы или годы) очевидного выздоровления после какого-либо наблюдаемого повреждения тканей.
В случаях нейропатической боли, сенсорная обработка по пути из пораженного участка может стать аномальной, и нейтральные раздражители (например, тепловые раздражители, прикосновение/надавливание), которые обычно не вызывают боли, могут привести к ее возникновению (т.е. к аллодинии), или болевые стимулы могут вызвать ненормально усиленное восприятие боли (т.е. гипералгезию) в ответ на обычный болевой стимул. Помимо этого, обычными стимулами могут быть вызваны ощущения, аналогичные пощипыванию при прохождении электрического тока или при поражениях электрическим током либо «покалыванию в конечностях» (т.е. парестезии) и/или могут быть вызваны ощущения с характерными неприятными особенностями (т.е. дисестезии). «Прорывная» боль представляет собой обострение предсуществующей хронической боли. Гиперпатия представляет собой болевой синдром, характеризующийся аномально болезненной реакцией на раздражение. В большинстве случаев это раздражение повторяется с возрастанием болевого порога, который можно рассматривать как наименьшее восприятие боли, которое пациент может признавать как боль.
Примеры нейропатической боли включают тактильную аллодинию (например, индуцированную в результате повреждения нерва), невралгию (например, постгерпетическую невралгию (или невралгию после опоясывающего герпеса), невралгию троичного нерва), рефлекторную симпатическую дистрофию/каузалгию (травму нерва), компоненты боли при раковом заболевании (например, боли, обусловленной самим раковым заболеванием или ассоциированными с ним состояниями, такими как воспаление, или боли, обусловленной лечением, таким как химиотерапия, хирургия или лучевая терапия), фантомную боль в ампутированных конечностях, нейропатию сдавливания (например, запястный синдром) и такие нейропатии, как периферическая нейропатия (например, обусловленная диабетом, вирусом иммунодефицита человека (ВИЧ), хроническим употреблением алкоголя, воздействием других токсинов (включая многие химиотерапевтические средства), витаминной недостаточностью и широким спектром других расстройств здоровья). Нейропатическая боль включает боль, индуцированную проявлением патологического функционирования нервной системы после повреждения нерва, обусловленного разными причинами, например, хирургической операцией, ранением, опоясывающим герпесом, диабетической нейропатией, ампутацией ног или рук, раковым заболеванием и тому подобным. Расстройства здоровья, ассоциированные с нейропатической болью, включают травматическое повреждение нерва, инсульт, рассеянный склероз, сирингомиелию, травму спинного мозга и рак.
Вызывающий боль стимул часто индуцирует воспалительную реакцию, которая сама может вносить вклад в болевые ощущения. При некоторых состояниях боль, по-видимому, вызывается комплексом ноцицептивных и нейропатических факторов. Например, хроническая боль часто включает воспалительную ноцицептивную боль или нейропатическую боль или смесь их обеих. Изначальная дисфункция или повреждение нервной системы может запускать нейронное высвобождение воспалительных медиаторов и последующее нейропатическое воспаление. Например, мигренеподобные головные боли могут представлять собой смесь нейропатической и ноцицептивной боли. Кроме того, миофасциальная боль, вероятно, является вторичной по отношению к ноцицептивному сигналу из мышц, но аномальная мышечная активность может быть результатом нейропатических состояний.
Наборы, содержащие антагонисты TNFα и NGF
Согласно данному изобретению предложены наборы, содержащие многофункциональный полипептид-антагонист TNFα и NGF, например, мультиспецифическую связывающую молекулу, предложенную в данном изобретении, или комбинацию антагониста TNFα и антагониста NGF, предложенную в данном изобретении, которые можно использовать для осуществления способов, описанных в данной заявке. Согласно определенным аспектам, набор содержит по меньшей мере многофункциональный слитый полипептид, содержащий антагонист TNFα и антагонист NGF, например, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 14 или 17, в одном или нескольких контейнерах, или комбинацию антагониста NGF, например MEDI-578, и антагониста TNFα, например антитела к TNFα, такого как инфликсимаб или адалимумаб, или TNFα-связывающего растворимого фрагмента рецептора TNF, например, TNFR2-FC. Специалист в данной области техники легко поймет, что описанные антагонисты TNFα и NGF, предложенные в данном изобретении, могут быть легко включены в один из установленных форматов набора, которые хорошо известны в данной области техники.
ПРИМЕРЫ
Теперь данное изобретение в целом описано, оно будет более понятным со ссылкой на следующие далее примеры, которые включены главным образом в целях иллюстрации определенных аспектов и воплощений, изложенных в описании настоящего изобретения, и не предназначены для ограничения данного описания.
Пример 1. Конструирование мультиспецифической связывающей молекулы (scFv-фрагмент антитела к NGF)/TNFR2-Fc и определение ее характеристик
Многофункциональную молекулу, конкретно, мультиспецифическую связывающую молекулу, содержащую домен антитела к NGF и TNFR2-Fc-домен, получали так, как приведено ниже. Присоединение scFv-фрагмента анти-NGF антитела к С-концу слитого белка TNFR2-Fc (SEQ ID NO: 13) осуществляли через СН3-домен тяжелой цепи соответственно формату Bs3Ab (биспецифические антитела), описанному в Dimasi, N., et al., J. Mol. Biol., 393: 672-92 (2009) и в публикации PCT под № WO 2013/070565. Графическое изображение структуры показано на Фиг. 1. ДНК-конструкции, кодирующие полипептид TNFR2-Fc и мультиспецифическую связывающую молекулу, синтезировали посредством GeneArt (Invitrogen). Для получения такой мультиспецифической связывающей молекулы конструировали scFv-фрагмент антитела к NGF, содержащий домены VH (SEQ ID NO: 3) и VL (SEQ ID NO: 7) из MEDI-578, соединенные вместе через линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15). Присоединение N-конца scFv к С-концу последовательности SEQ ID NO: 13 осуществляли через линкерную последовательность, состоящую из 10 аминокислот (GGGGS)2. Эта мультиспецифическая связывающая молекула обозначается в данном описании как TNFR2-Fc_VH#4. ДНК-конструкцию, кодирующую такую мультиспецифическую связывающую молекулу, создавали такой, чтобы она содержала стоп-кодон и сайт рестрикции EcoRI на 3'-конце для клонирования в Bs3Ab-экспрессирующий вектор. Последовательность ДНК, кодирующая TNFR2-Fc_VH#4, представлена в виде SEQ ID NO: 16, а соответствующая ей аминокислотная последовательность - в виде SEQ ID NO: 14.
Термостабильность мультиспецифической в отношении TNF-NGF связывающей молекулы улучшали, добавляя межцепочечную дисульфидную связь между VH- и VL-доменами scFv-части MEDI-578 в мультиспецифической связывающей молекуле. Это было сделано путем введения мутации G→С по аминокислоте 44 VH-домена (SEQ ID NO: 94) и по аминокислоте 102 VL-домена (SEQ ID NO: 95). Этот клон получил название TNFR2-Fc_varB. Аминокислотная последовательность TNFR2-Fc_varB представлена в виде SEQ ID NO: 17. Последовательность ДНК, кодирующая TNFR2-Fc_varB, представлена в виде SEQ ID NO: 18. Кодон-оптимизированная последовательность ДНК, кодирующая TNFR2-Fc_varB, представлена в виде SEQ ID NO: 99. TNFR2-Fc_varB дополнительно отличается от TNFR2-Fc_VH#4 тем, что линкерная последовательность, состоящая из 15 аминокислот (GGGGS)3, соединяющая VH и VL scFv-части, заменена на состоящий из 20 аминокислот линкер (GGGGS)4 (SEQ ID NO: 19). Чтобы измерить Тпл для TNFR2-Fc_VH#4 и TNFR2-Fc_varB, использовали дифференциальную сканирующую флуориметрию (DSF). В этом способе оценивают встраивание флуоресцентного красителя Sypro Orange (Invitrogen), который связывается с гидрофобными поверхностями, открывающимися в процессе разворачивания белкового домена при воздействии повышенных температур. В анализе с применением DSF, Тпл для TNFR2-Fc_VH#4 составляла 62°С, в то время как Тпл для TNFR2-Fc_varB составляла 66°С. Таким образом, добавление межцепочечной дисульфидной связи в scFv-часть MEDI-578 мультиспецифической молекулы улучшало термостабильность молекулы на 4°С.
Белки TNFR2-Fc и TNFR2-Fc_VH#4 подвергали кратковременной экспрессии в суспензии клеток СНО с использованием полиэтиленимина (PEI) (Polysciences) в качестве реагента для трансфекции. Клетки поддерживали в среде CD-CHO (Life Technologies). Культуральные сборы после проведения мелкомасштабных трансфекций очищали, используя аффинную хроматографию на колонке HiTrap MabSelect SuRe™ (1 мл) в соответствии с протоколом производителя (GE Healthcare) и после этого буфер заменяли на буфер следующего состава: 1% сахарозы, 100 мМ NaCl, 25 мМ гидрохлорид L-аргинина и 25 мМ фосфат натрия (pH 6,3). Чистоту рекомбинантных белков анализировали, используя SDS-PAGE в восстанавливающих условиях и используя аналитическую гель-проникающую хроматографию (см. приведенный ниже метод), и концентрации определяли, измеряя поглощение при 280 нм, используя теоретически определенные коэффициенты экстинкции.
В результате проведения мелкомасштабной кратковременной экспрессии и очистки слитого белка TNFR2-Fc и мультиспецифической в отношении TNF-NGF конструкции, TNFR2-Fc_VH#4, на колонке с иммобилизованным белком А получали выходы 36,6 и 79,9 мг⋅л-1, соответственно.
Более крупную партию TNFR2-Fc_VH#4 готовили так, как приведено ниже. Неочищенный культуральный сбор после крупномасштабной трансфекции (до 6 л включительно) фильтровали, используя глубинное фильтрование, и загружали на колонку 1,6×20 см с агарозой с иммобилизованным белком A (GE Healthcare), предварительно уравновешенную буфером А (забуференным фосфатом физиологическим раствором, pH 7,2). Затем колонку промывали буфером А и продукт элюировали, используя ступенчатый градиент буфера В (50 мМ ацетата натрия, pH меньше 4,0). Продукт очищали далее, загружая на колонку 1,6×20 см Poros HS 50 (Applied Biosystems), предварительно уравновешенную буфером С (50 мМ натрий-ацетатным буфером, pH меньше 5,5), промывали буфером С и затем продукт элюировали, используя линейный градиент от 0 до 1 М NaCl в 50 мМ натрий-ацетатном буфере, pH меньше 5,5. Полученные элюаты анализировали с использованием гель-проникающей HPLC. Концентрацию белка определяли спектроскопией с измерением А280 на спектрофотометре Beckman DU520, используя рассчитанный коэффициент экстинкции 1,36.
Методы определения характеристик TNFR2-Fc_VH#4
Анализ с применением вестерн-блоттинга проводили, используя стандартные протоколы. Белки переносили на поливинилиденфторидную (PVDF) мембрану (Life Technologies), используя систему Xcell SureLock™ (Invitrogen) в соответствии с инструкциями производителя. Мембрану блокировали 3%-ным раствором (масс./об.) сухого обезжиренного молока в забуференном фосфатом физиологическом растворе (PBS) в течение 1 ч при комнатной температуре. Проявку вестерн-блотов осуществляли, используя стандартные протоколы с применением конъюгированного с пероксидазой хрена (HRP) антитела, специфического к Fc-фрагменту IgG человека (Sigma).
Гель-проникающую HPLC проводили, используя систему для HPLC от Gilson (изократический насос-307, детектор-151 для работы в ультрафиолетовой и видимой областях спектра (UVA/is), устройство подачи жидкостей-215 и модуль для введения-819) с колонкой BioSep-SEC-S3000 (300×7,8 мм) от Phenomenex и D-PBS (PBS-буфер по Дульбекко; Life Technologies) в качестве подвижной фазы при скорости потока 1 мл/мин. В колонку вводили по двадцать пять мкл раствора образцов и проводили мониторинг разделения белковых компонентов по А280 нм.
Ферментативное дегликозилирование очищенного в виде мелкой партии белка TNFR2-Fc_VH#4 проводили, используя набор EDGLY (Sigma-Aldrich) в соответствии с протоколами производителя. Белки подвергали дегликозилированию как в денатурирующих, так и в нативных условиях. Что касается денатурированных белков, то дегликозилирование 30 мкг белка проводили в течение 3 ч при 37°С, используя пептид:N-гликозидазу (PNGase; peptide:N-glycosidase) F, О-гликозидазу, α-(2→3,6,8,9)-нейраминидазу, β-N-ацетилглюкозаминидазу и β-(1→4)-галактозидазу. Используя тот же набор ферментов, которые описаны выше, проводили дегликозилирование 35 мкг белка в нативных условиях в течение 3 суток при 37°С. Дегликозилированные белки анализировали посредством SDS-PAGE с применением окрашивания с помощью кумасси и с помощью вестерн-блоттинга, используя стандартные протоколы анализа.
N-Концевое секвенирование аминокислотной последовательности TNFR2-Fc_VH#4 проводили так, как приведено ниже. Приблизительно 2 мкг TNFR2-Fc_VH#4 анализировали на геле для SDS-PAGE, используя стандартные протоколы. Белки переносили на PVDF мембрану, используя систему Xcell SureLock™ (Invitrogen) в соответствии с инструкциями производителя. Мембрану окрашивали 0,1%-ным (масс./об.) амидочерным в течение приблизительно 15 мин на орбитальном шейкере, затем промывали дистиллированной водой (dH2O) для уменьшения фонового окрашивания PVDF мембраны. Мембрану сушили на воздухе, после чего проводили N-концевое секвенирование. Представляющие интерес зоны вырезали, и определение последовательности N-конца мультиспецифической связывающей молекулы выполняли на секвенаторе 494 НТ от Applied Biosystems (Applied Biosystems, San Francisco, CA, USA), применяя фенилтиогидантоиновый метод в режиме реального времени с использованием системы микро-HPLC, модели 140А, от Applied Biosystems.
Результаты определения характеристик
Очищенные белки TNFR2-Fc_VH#4 и TNFR2-Fc анализировали посредством SEC-HPLC в отношении уровней агрегатов, мономеров и белковых фрагментов (Фиг. 2А и 2В). Основной пик, содержащий мономер, составлял приблизительно 90% от общего присутствующего белка, при этом остальные приблизительно 10% соответствовали белковой массе с более низким временем удерживания в колонке, что указывало на присутствие структур более высокого порядка или агрегатов. Однако, согласно SEC-HPLC пик мономера имел два отчетливых плеча, указывая на то, что белок в этом пике не является гомогенным. Анализ с применением SDS-PAGE с окрашиванием с помощью кумасси показал две четко выраженные зоны для TNFR2-Fc_VH#4 (при приблизительно 100 и 75 кДа) и, аналогичным образом, также две четко выраженные зоны для слитого белка TNFR2-Fc (при приблизительно 70 и 45 кДа) в восстанавливающих условиях (Фиг. 2В). В случае невосстанавливающих условий обнаружены три основные зоны для TNFR2-Fc_VH#4 (от 150 до 250 кДа) и одна основная и одна минорная зона для слитого белка TNFR2-Fc при приблизительно 150 и 120 кДа, соответственно. Поскольку разница в молекулярных массах для этих двух зон в восстанавливающих условиях была приблизительно эквивалентна размеру scFv-фрагмента (примерно 26,5 кДа), выполняли дальнейший анализ, чтобы понять, в каких формах была получена мультиспецифическая связывающая молекула. Масс-спектроскопический анализ в нативных условиях подтверждал данные SDS-PAGE о том, что в случае двух отдельных препаратов очищенного белка в очищенном препарате TNFR2-Fc_VH#4 присутствовали три белка с молекулярными массами, приблизительно соответствующими 125, 152 и 176 кДа (Фиг. 2С).
Если картина зон, наблюдаемая в геле после SDS-PAGE, была обусловлена разным гликозилированием TNFR2-Fc_VH#4, то после дегликозилирования должен быть возврат обратно к одной зоне. Однако, такая картина зон сохранялась как в восстанавливающих, так и в невосстанавливающих условиях, когда TNFR2-Fc_VH#4 подвергали дегликозилированию либо в виде нативного белка, либо в виде денатурированного белка (данные не показаны). Окрашивание на вестерн-блотах как гликозилированного, так и дегликозилированного белка TNFR2-Fc_VH#4 с использованием поликлонального антитела, специфического к Fc-фрагменту IgG человека показало, что как ожидаемая зона полноразмерного белка, так и зона с более низкой молекулярной массой были реакционноспособными в отношении специфического к Fc-фрагменту антитела (данные не показаны).
Окончательную идентификацию укороченного продукта проводили с использованием N-концевого аминокислотного секвенирования белка. При этом было выявлено, что первыми 8 аминокислотами с N-конца укороченного белка являются SMAPGAVH, что соответствует аминокислотам 176-183 в последовательности TNFR2-Fc_VH#4 (SEQ ID NO: 14). Это соответствовало укорачиванию на 175 аминокислот с N-конца TNFR2-Fc_VH#4, при котором оставались только 42 аминокислоты домена TNFR2. Это позволяет авторам изобретения точно интерпретировать касающиеся масс данные по результатам SDS-PAGE, масс-спектроскопии и анализа посредством SEC-HPLC. Возможны три комбинации димеров TNFR2-Fc_VH#4, и все они присутствовали в препаратах очищенных белков: (1) полноразмерный гомодимер, (2) гетеродимер полноразмерной и укороченной структур и (3) гомодимер укороченных структур. Чтобы точно измерить биологическую активность как in vitro, так и in vivo, готовили препарат полноразмерного гомодимера, используя двухстадийный процесс колоночной хроматографии. На первой стадии, после очистки на колонке с иммобилизованным белком А, продукт содержал 80,5% мономера (Фиг. 3А), а после второй стадии очистки на колонке (SP сефароза) процентное содержание мономера составляло 97,8% (Фиг. 3В). Выход после проведения всего процесса составлял 7,3%.
Пример 2. Анализ термостабильности с использованием дифференциальной сканирующей калориметрии (DSC)
Для калориметрических измерений использовали автоматизированную систему для DSC MicroCal VP-Capillary (GE Healthcare, USA). Образцы белков тестировали в концентрации 1 мг/мл в 25 мМ буфере на основе гистидина/гистидина-HCl, pH 6,0. Образцы белка и буфер подвергали воздействию линейного изменения температуры от 25°С до 100°С со скоростью 95°С за один час. Данные для буфера как референсного образца вычитали из данных для образца белка, используя программное обеспечение Origin 7, и определяли области температурных переходов.
На термограмме для TNFR2-Fc_VH#4 (Фиг. 4) показаны три четко выраженных перехода, соответствующих разворачиванию, с температурами денатурации (Тпл) 64, 67 и 84°С. Авторы изобретения сделали вывод, что Тпл 64°С соответствовала денатурации как домена TNFR2, так и scFv домена антитела к NGF, при этом Тпл 67°С и 84°С представляли собой типичные Тпл, соответствующие денатурации для СН2- и СН3-доменов IgG1, соответственно (например, Dimasi, N., et al., J. Mol. Biol., 393: 672-92 (2009) и публикация PCT № WO 2013/070565). Без связи с теорией отметим, что scFv обычно имеют более низкие температуры денатурации по сравнению с другими доменами антитела, и их разворачивание характеризуется одиночным переходом (Roberge и др., 2006, Jung и др., 1999, Tischenko и др., 1998).
Пример 3. Подтверждение связывания антигенов с TNFR2-Fc_VH#4
А. Связывание одиночного и двойного антигена по данным ELISA
На поверхность лунок планшетов Nunc Maxisorp при 4°С в течение ночи наносили покрытие, используя 50 мкл раствора TNFα (R&D Systems), разбавленного до концентрации 5 мкг мл-1 в PBS (pH 7,4). На следующий день раствор для нанесения покрытия удаляли и лунки блокировали 150 мкл блокирующего буфера (3% обезжиренного молока в PBS) в течение 1 ч при комнатной температуре. Лунки промывали три раза в PBS, после чего добавляли по 50 мкл растворов из серии разведений белка TNFR2-Fc_VH#4, приготовленного в блокирующем буфере. Через 1 ч инкубации при комнатной температуре лунки трижды промывали в смеси PBS-твин 20 (0,1% об./об.; PBS-Т). Затем в лунки добавляли по пятьдесят микролитров раствора биотинилированного NGF и инкубировали в течение еще одного часа при комнатной температуре, после чего промывали, как указано выше, и добавляли по 50 мкл раствора стрептавидин-HRP (1:100). Через 1 час инкубации при комнатной температуре лунки промывали PBS-T, добавляли по 50 мкл раствора субстрата 3,3',5,5'-тетраметилбензидина и отставляли для развития окраски. Реакцию останавливали добавлением 1 М раствора H2SO4 и измеряли поглощение при 450 нм, используя титрационный микропланшетный ридер. Полученные данные анализировали, используя программное обеспечение Prism 5 (GraphPad, San Diego, CA). Чтобы провести ELISA связывания одиночного антигена, лунки покрывали либо TNFα, либо NGF-биотином, как описано выше, и детекцию связывания с антителом выполняли, используя конъюгированное с HRP антитело, специфичное к Fc-фрагменту IgG человека (1:5000), а окрашивание осуществляли так, как описано выше.
Результаты ELISA показаны на Фиг. 5. Белок TNFR2-Fc_VH#4 предназначался для связывания с обоими антигенами TNFα и NGF. Связывание одиночного антигена осуществляли, проводя сначала иммобилизацию одного антигена на 96-луночном титрационном микропланшете, затем добавляя TNFR2-Fc_VH#4 в серийных разведениях. Детекцию специфического связывания проводили, используя конъюгированное с пероксидазой хрена (HRP) антитело, специфичное к Fc-фрагменту IgG. Чтобы провести ELISA связывания двойного антигена, первый антиген, TNFα, иммобилизовали на планшете для ELISA и затем добавляли TNFR2-Fc_VH#4 в серийных разведениях, после чего добавляли второй биотинилированный антиген, NGF, в фиксированной концентрации. Затем проводили детекцию специфического связывания, используя HRP-конъюгированный стрептавидин. Белок TNFR2-Fc_VH#4 связывался с TNFα и NGF в ELISA в режиме связывания одиночного антигена (Фиг. 5А и В). В ELISA в режиме связывания двойного антигена, TNFR2-Fc_VH#4 связывался одновременно как с TNFα, так и NGF (Фиг. 5С).
Б. Одновременное связывание антигенов с применением поверхностного плазмонного резонанса
Эксперименты по одновременному связыванию с антигенами проводили по существу так, как описано в Dimasi, N., et al., J. Mol. Biol., 393: 672-92 (2009), используя прибор BIAcore 2000 (GE Healthcare). Кратко, для иммобилизации приблизительно 1500 резонансных единиц TNFR2-Fc_VH#4 в концентрации 100 нМ использовали сенсорный чип СМ5. Поверхности сенсорного чипа затем использовали для изучения конкурентного связывания с TNFα и NGF. Образцы антигенов готовили в HBS-EP буфере (10 мМ HEPES (4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота) (pH 7,4), 150 мМ NaCl, 3 мМ этилендиаминтетрауксусная кислота (EDTA), 0,005% полисорбата Р20). Для всех измерений связывания использовали скорость потока 30 мкл/мин. Чтобы оценить одновременное связывание мультиспецифического антитела с TNFα и NGF, над поверхностью сенсорного чипа вводили 1 мкМ раствор TNFα (молекулярная масса 17,5 кДа) и по завершении введения далее вводили смесь TNFα и NGF (молекулярная масса 13,5 кДа), оба в концентрации 1 мкМ. TNFα включали в смесь с NGF для предупреждения потери сигнала, обусловленной диссоциацией TNFα во время фазы связывания с NGF. В качестве контроля осуществляли аналогичную процедуру связывания и при последнем введении добавляли только TNFα; отсутствие какого-либо увеличения числа резонансных единиц для этого введения указывало на то, что связывание с TNFα происходило на уровнях насыщения. Проводили аналогичные эксперименты по связыванию и контрольные эксперименты, в которых порядок введения TNFα и NGF был обратным.
Одновременное связывание TNFR2-Fc_VH#4 с антигенами характеризовали с использованием поверхностного плазмонного резонанса. Качественный анализ относящихся к связыванию событий проводили последовательно друг за другом. TNFR2-Fc_VH#4 ковалентно иммобилизовали на поверхности сенсорного чипа, применяя химические реакции аминосочетания. После этого вводили первый антиген, получая насыщающие уровни связывания с TNFR2-Fc_VH#4, затем вводили второй антиген в виде эквимолярной смеси с антигеном 1. Сенсограмма связывания отчетливо показала, что белок TNFR2-Fc_VH#4 связывался одновременно с TNFα и NGF (Фиг. 6). Одновременное связывание с двумя данными антигенами происходило независимо от порядка введения антигенов.
Пример 4. Ингибирование пролиферации клеток TF-1, индуцированной NGF
Клетки TF-1 (ЕСАСС, № по каталогу 93022307) рассевали в количестве 1,5×104 клеток/лунка в 50 мкл бессывороточных культуральных сред в 96-луночный планшет для тканевых культур (Corning Costar) и инкубировали в течение 18 ч при 37°С в атмосфере с 5% СО2. Рекомбинантный NGF человека (Sigma) или мыши (R&D Systems) предварительно инкубировали с разведениями TNFR2-Fc_VH#4, MEDI-578 в виде IgG1 ТМ YTE, не связывающегося изотипического контроля для MEDI-578 IgG1 ТМ YTE или не связывающегося биспецифического изотипического контроля R347 Bs3Ab в течение 30 мин при 37°С в 96-луночном круглодонном планшете (Greiner). Затем в планшет с клетками добавляли по пятьдесят микролитров каждого образца и инкубировали в течение 48 ч при 37°С.По окончании периода инкубации добавляли по 100 мкл буфера для анализа клеток CellTiter-GLO® (Promega) и планшет инкубировали в течение 10 мин при 37°С в атмосфере с 5% СО2. Затем измеряли люминесценцию, используя стандартный протокол для определения люминесценции. Стандартная NGF-индуцированная пролиферация клеток TF-1 в отсутствие антитела показана на Фиг. 7А.
Функциональную активность TNFR2-Fc_VH#4 определяли, используя NGF-индуцированную пролиферацию клеток TF-1. Белок TNFR2-Fc_VH#4 способен полностью ингибировать пролиферацию, индуцированную NGF как человека, так и мыши (Фиг. 7B и 7С, соответственно). Фиг. 7В: клетки TF-1 стимулировали рекомбинантным NGF человека в концентрации, соответствующей ЕС80. Клетки инкубировали с лигандом в присутствии серийных разведений антитела в течение 48 ч, после чего проводили количественную оценку пролиферации клеток путем культивирования в течение 10 мин, используя буфер для анализа клеток CellTiter-GLO® (Promega). Фиг. 7С: клетки TF-1 стимулировали рекомбинантным мышиным NGF в концентрации, соответствующей ЕС80. Клетки инкубировали с лигандом в присутствии серийных разведений антитела в течение 48 ч, после чего проводили количественную оценку пролиферации клеток путем культивирования в течение 10 мин, используя буфер для анализа клеток CellTiter-GLO® (Promega). Эти данные демонстрируют, что NGF-ингибирующая часть TNFR2-Fc_VH#4 является биологически активной и ингибирует NGF-индуцированную пролиферацию со схожей эффективностью, проявляемой MEDI-578 в виде IgG1TM. Аналогичные данные также наблюдали для TNFR2-Fc_varB и другой мультиспецифической в отношении TNF-NGF связывающей молекулы - ндимаба varB (ФИГ. 7D и 7Е). Ндимаб varB содержит полноразмерное антитело к TNFα, т.е. антитело, содержащее две полные тяжелые цепи и две полные легкие цепи в формате H2L2, с scFv-фрагментом MEDI-578, слитым с С-концом тяжелой цепи антитела к TNFα. Легкая цепь ндимаба varB представлена в SEQ ID NO: 20, а тяжелая цепь ндимаба varB представлена в SEQ ID NO: 22.
Пример 5. Ингибирование апоптоза клеток U937, индуцированного TNFα
Клетки U937 (ЕСАСС, № по каталогу 85011440) помещали в 96-луночный планшет для тканевых культур с черными стенками (Coming Costar) в количестве 8×105 клеток/лунка в 50 мкл культуральных сред. Клетки U937 стимулировали рекомбинантным TNFα человека в концентрации, соответствующей ЕС80. Клетки инкубировали с лигандом в присутствии серийных разведений антитела в течение 2 ч, после чего выполняли количественное определение активности каспазы 3 путем культивирования в течение 2 часов, используя буферный раствор для измерения активности каспазы 3. TNFR2-Fc_VH#4, не связывающийся биспецифический изотипический контроль R347 Bs3Ab и этанерцепт предварительно инкубировали с клетками в течение 30 мин при 37°С. После этого добавляли 50 мкл рекомбинантного TNFα человека (R&D Systems), получая конечную концентрацию в анализе 20 нг/мл, и далее инкубировали в течение 2 ч при 37°С. По окончании периода инкубации добавляли 50 мкл буферного раствора для измерения активности каспазы 3 (0,2% (масс./об.) CHAPS (3-[(3-холамидопропил)диметил-аммонио]-1-пропансульфонат); 0,5% (об./об.) игепала СА-630; 200 мМ NaCl; 50 мМ HEPES; 20 мкМ субстрат DEVD-R110 (Invitrogen)) и клетки инкубировали в течение 2,5 ч при 37°С. Флуоресценцию измеряли при возбуждении при 475 нм и испускании при 512 нм. Каспазная активность в отсутствие антагониста TNFα показана на Фиг. 8А.
Функциональную активность TNFR2-Fc_VH#4 определяли, используя анализ TNFα-индуцированной активности каспазы 3 в клетках U937. Белок TNFR2-Fc_VH#4 полностью ингибировал TNFα-индуцированную активность каспазы 3 так же, как этанерцепт (Фиг. 8В). Это ясно показывает, что TNFα-ингибирующая часть TNFR2-Fc_VH#4 является биологически активной и обладает эффективностью, аналогичной этанерцепту. Аналогичные данные также наблюдали для TNFR2-Fc_varB и ндимаба varB (см. Фиг. 8С).
Пример 6. Анализы in vivo
Все процедуры in vivo проводили в соответствии с Актом Министерства внутренних дел Великобритании о научных процедурах с животными (UK Home Office Animals (Scientific Procedures) Act) (1986), одобренным местным комитетом по этике. На всех этапах использовали самок мышей C57BI/6 (Charles River, UK). Мышей размещали группами по 5-6 штук на одну клетку в отдельных вентилируемых клетках (IVC) со свободным доступом к корму и воде в условиях 12-часового цикла свет/темнота (свет: 07:00-19:00). Температуру в виварии и комнате для проведения экспериментов поддерживали при 24°С, а постоянный фоновый шум обеспечивали посредством обычного радиоузла. В целях идентификации всем мышам по меньшей мере за 5 суток до начала каждого исследования под анестезией (3%-ным изофлураном в кислороде) вставляли транспондеры.
А. Модель нейропатической боли Зельтцера
Механическую гипералгезию оценивали с использованием аналгезиметра (Randall L.O., Selitto J.J., Arch. Int. Pharmacodyn. Ther., 111: 409-19 (1957)) (Ugo Basile). К дорсальной поверхности каждой задней лапы по очереди прикладывали возрастающее усилие до тех пор, пока не наблюдали реакцию в виде отдергивания. В этот момент приложение усилия останавливали и регистрировали нагрузку в граммах. Данные выражали в виде порога отдергивания в граммах для ипсилатеральной и контралатеральной лап. После установления исходных показателей мышей разделяли на 2 группы с приблизительно одинаковыми соотношениями между ипсилатеральной/контралатеральной лапами и проводили хирургическую операцию. Мышей подвергали анестезии, используя 3%-ный изофлуран. После этого обнажали приблизительно 1 см левого седалищного нерва методом тупой диссекции через разрез на уровне середины бедра. Затем через дорсальную треть нерва пропускали шовный материал (10/0 натуральный шелк : этикон) и туго перевязывали. Разрез закрывали с помощью клея, и мышей оставляли восстанавливаться в течение по меньшей мере семи суток перед началом исследований. Ложнооперированных (sham operated) мышей обрабатывали согласно такому же протоколу, но после обнажения нерва рану заклеивали и мышам давали возможность восстановиться. Мышей тестировали в отношении гипералгезии на 7-е и 10-е сутки после хирургического вмешательства. На 10-е сутки тестирования прооперированных мышей далее подразделяли на группы, которые получали изотипический контроль САТ251 IgG1 (0,03 мг/кг, подкожно (п.к.)), этанерцепт (0,01 мг/кг, п.к.), MEDI-578 (0,03 мг/кг, п.к.) или комбинацию этанерцепта (0,01 мг/кг, п.к.) и MEDI-578 (0,03 мг/кг, п.к.). Все ложнооперированные мыши получали САТ251 (0,03 мг/кг, п.к.). Механическую гипералгезию измеряли в моменты времени 4 ч, 1-е, 2-е, 3-й, 4-е и 7-е сутки после введения дозы.
Совместное введение этанерцепта и MEDI-578 в модели механической гипералгезии проявляется как значительное уменьшение соотношения между ипсилатеральной/контралатеральной лапами на 10-е сутки после хирургического вмешательства по сравнению с ложнооперированными контролями (Фиг. 9). При введении разовой дозы или этанерцепта (0,01 мг/кг, п.к.), или MEDI-578 (0,03 мг/кг, п.к.) не удавалось устранить в значительной степени эту гипералгезию. Совместное введение этанерцепта (0,01 мг/кг, п.к.) вместе с MEDI-578 (0,03 мг/кг, п.к.) способствовало значительному устранению механической гипералгезии через 4 ч после введения дозы, и этот эффект сохранялся на протяжении 7 суток после введения дозы.
Во втором исследовании оценивали действие TNFR2-Fc_VH#4. После установления механической гипералгезии мышам на 13-е сутки после хирургического вмешательства вводили изотипический контроль R347 Bs3Ab (0,03 мг/кг, п.к.), этанерцепт (0,01 мг/кг, п.к.), MEDI-578 (0,03 мг/кг, п.к.) или TNFR2-Fc_VH#4 (0,01 мг/кг или 0,03 мг/кг, п.к.). Ложнооперированные животные получали изотипический контроль R347 Bs3Ab (0,03 мг/кг, п.к.). Мышей тестировали в отношении механической гипералгезии через 4 ч после введения и на 1-е, 2-е, 4-е и 7-е сутки после введения, как описано выше.
Введение TNFR2-Fc_VH#4 приводило к значительному уменьшению соотношения между ипсилатеральной/контралатеральной лапами на 10-е сутки после хирургического вмешательства по сравнению с ложнооперированными контролями (Фиг.10А). При введении или этанерцепта (0,01 мг/кг, п.к.), или MEDI-578 (0,03 мг/кг, п.к.) не удавалось устранить в значительной степени механическую гипералгезию. Между тем, введение TNFR2-Fc_VH#4 (0,01 и 0,03 мг/кг, п.к.) приводило к значительному устранению механической гипералгезии через 4 ч после введения дозы, эффекту, который сохранялся на протяжении 6 суток после введения. Никакого эффекта не наблюдали после введения контроля R347 Bs3Ab. Аналогичные результаты получали при введении TNFR2-Fc_varB (см. Фиг. 10В). Эти данные подтверждают, что TNFR2-Fc_VH#4 в очень низких дозах может способствовать значительному устранению боли, в то же время показано, что эквивалентные дозы одного лишь MEDI-578 или этанерцепта оказались неэффективными или незначительно эффективными.
Б. Модель хронической суставной боли
Гиперчувствительность к механическим раздражителям определяли, используя контрольно-измерительный прибор для фиксации состояния временного выведения из строя (incapacitance tester) для мышей (Linton Instrumentation). Мышей помещали в это устройство, располагая их задние лапы на отдельных датчиках, и распределение массы тела рассчитывали в течение 4 с. Данные выражали в виде соотношения весовой нагрузки между ипсилатеральной и контралатеральной лапами в граммах.
После установления исходных показателей мышей разделяли на 2 группы с приблизительно одинаковыми соотношениями между ипсилатеральной/контралатеральной лапами. Внутрисуставные инъекции осуществляли, применяя следующий метод: животных подвергали анестезии, используя 3%-ный изофлуран в кислороде, и левое колено выбривали и протирали. В коленный сустав каждой мыши вводили инъекцией или 10 мкл полного адъюванта Фрейнда (FCA) (10 мг/мл), или наполнитель (легкое минеральное масло), используя иглу 25-го калибра, установленную на шприц фирмы Гамильтон емкостью 100 мкл. Инъекции выполняли непосредственно в синовиальное пространство коленного сустава. Мышей оставляли восстанавливаться и повторно тестировали в отношении изменений гиперчувствительности к механическим раздражителям на 7-е и 10-е сутки после инъекции, как описано выше. На 10-е сутки тестирования обработанных FCA мышей далее рандомизировали в группы и на 13-е сутки мышам вводили этанерцепт (0,01 мг/кг, внутрибрюшинно (в.б.)) или наполнитель, после чего они получали дозу MEDI-578 (0,03 мг/кг, внутривенно (в.в.)) или изотипического контроля САТ251 (0,03 мг/кг, в.в.). Мышей тестировали в отношении гиперчувствительности к механическим раздражителям через 4 ч после введения и на 1-е, 2-е, 4-е и 7-е сутки после введения, как описано выше.
Эффект совместного введения этанерцепта и MEDI-578 оценивали, используя модель воспалительной боли, индуцированной введением FCA внутрь сустава. Внутрисуставное введение FCA вызывало гиперчувствительность к механическим раздражителям, которая проявлялась в виде значительного уменьшения соотношения между ипсилатеральной/контралатеральной лапами на 7-е и 10-е сутки по сравнению с введением наполнителя в качестве контроля (Фиг. 11). Никакого уменьшения соотношения между ипсилатеральной/контралатеральной лапами не наблюдали в ложнообработанных группах по сравнению с исходными уровнями до проведения обработки. Введение этанерцепта (0,01 мг/кг, в.б.) + САТ251 (0,03 мг/кг, в.в.) или PBS (10 мл/кг, в.б.) + MEDI-578 (0,03 мг/кг, в.в.) вызывало незначительное устранение FCA-индуцированной гиперчувствительности к механическим раздражителям через 4 ч и на 1-е, 2-е, 4-е и 7-е сутки после введения дозы, но при этом не удавалось достичь статистической значимости. Между тем, введение этанерцепта (0,01 мг/кг, в.б.) + MEDI-578 (0,03 мг/кг, в.в.) способствовало значительному устранению FCA-индуцированной гиперчувствительности к механическим раздражителям во все моменты времени тестирования после введения.
Во втором исследовании оценивали действие TNFR2-Fc_VH#4. После установления FCA-индуцированной гиперчувствительности к механическим раздражителям мышам на 13-е сутки после введения FCA вводили: изотипический контроль R347 Bs3Ab (0,01 мг/кг, п.к.), этанерцепт (0,01 мг/кг, п.к.), MEDI-578 (0,01 мг/кг, п.к.) или TNFR2-Fc_VH#4 (0,003 мг/кг или 0,01 мг/кг, п.к.). Мышей опять тестировали в отношении гиперчувствительности к механическим раздражителям через 4 ч после введения и на 1-е, 2-е, 4-е и 7-е сутки после введения, как описано выше.
Действие TNFR2-Fc_VH#4 («биспецифический») по сравнению с действиями по отдельности этанерцепта и MEDI-578 показано на Фиг. 12. Ни этанерцепт (0,01 мг/кг, п.к.), ни MEDI-578 (0,01 мг/кг, п.к.) не вызывали значительного устранения FCA-индуцированной гиперчувствительности к механическим раздражителям в любой момент времени после введения. В то же время, введение TNFR2-Fc_VH#4 способствовало значительному устранению FCA-индуцированной гиперчувствительности к механическим раздражителям. Для более высокой дозы TNFR2-Fc_VH#4 (0,01 мг/кг, п.к.) продемонстрирована значительная активность на протяжении всего исследования, тогда как для более низкой дозы (0,003 мг/кг, п.к.) уровня значимости достигали в 1-е сутки после введения, который сохранялся аналогичным уровню для более высокой дозы на протяжении всего исследования.
В. Утвержденная модель FCA-индуцированной гиперчувствительности к механическим раздражителям у крыс
Внутриподошвенная инъекция полного адъюванта Фрейнда (FCA) вызывает воспалительную реакцию, что индуцирует гиперчувствительность и отек и имитирует некоторые аспекты клинического проявления воспалительной боли. Эти эффекты можно исследовать с использованием оборудования для измерения весовой нагрузки. Для оценки возможных антигипералгезических свойств TNFR2-Fc_VH#4 при FCA-индуцированной гиперчувствительности использовали метод весовой нагрузки. Не подвергавшиеся ранее воздействию (наивные) крысы распределяют вес своего тела одинаково между двумя задними лапами. Однако, когда после инъекции задняя (левая) лапа воспаляется и/или болит, вес перераспределяется таким образом, что на пораженную лапу приходится меньший вес (уменьшение весовой нагрузки на поврежденную лапу). Весовую нагрузку, приходящуюся на каждую заднюю конечность, измеряют, используя контрольно-измерительный прибор для фиксации состояния временного выведения из строя (далее «контрольно-измерительный прибор») (Linton Instruments, UK) для крыс. Крыс помещают в этот контрольно-измерительный прибор, располагая задние лапы на отдельных датчиках, и среднее усилие, прилагаемое к обеим задним конечностям, регистрируют в течение 4 секунд.
Для этого исследования проводили акклиматизацию не подвергавшихся ранее воздействию крыс (самцов крыс Sprague Dawley (Harlan, UK), 198-258 г) в процедурной комнате в их собственных домашних клетках со свободным доступом к пище и воде. Привыкание к контрольно-измерительному прибору проводили в течение нескольких дней. Перед внесением повреждающего фактора осуществляли регистрацию исходной весовой нагрузки. Гиперчувствительность к воспалению индуцировали путем внутриподошвенной инъекции FCA (поставляемого Sigma, 100 мкл раствора в концентрации 1 мг/мл) в левую заднюю лапу. Через 23 часа после введения FCA выполняли измерение весовой нагрузки до лечения для оценки гиперчувствительности.
Затем животных ранжировали и рандомизировали в подвергаемые обработке группы в соответствии с FCA-диапазоном весовой нагрузки в опыте по схеме латинского квадрата. Через 24 часа после инъекции FCA животных обрабатывали или TNFR2-Fc_VH#4 («биспецифический»), вводимым в.в. в дозе 0,003, 0,01, 0,03, 0,3 и 3 мг/кг, или антителом NIP228 в качестве отрицательного контроля (антителом, выработанным для связывания с гаптеном нитрофенолом), вводимым в.в. в дозе 3 мг/кг, или наполнителем (1%-ным раствором метилцеллюлозы), вводимым перорально (п.о.) в дозе 2 мл/кг, или индометацином, вводимым п.о. в дозе 10 мг/кг.
Весовую нагрузку оценивали через 4 и 24 часа после обработки антителом/лекарственными средствами. Данные для каждого момента времени анализировали, сравнивая подвергаемые обработке группы с контрольной получавшей наполнитель группой. Статистический анализ включал ANOVA с повторными измерениями, затем критерий планируемых сравнений (planned comparison test) с использованием InVivoStat (invivostat.co.uk) (p<0,05 говорит о статистической значимости). Результаты показаны на Фиг. 13. Наблюдали существенное устранение гиперчувствительности на моменты времени 4 и 24 часа при использовании индометацина (10 мг/кг). Для TNFR2-Fc_VH#4 в дозе 0,3 и 3 мг/кг показано значительное устранение гиперчувствительности как для 4, так и 24 часов, для TNFR2-Fc_VH#4 в дозе 0,003 и 0,03 мг/кг также показано значительное устранение гиперчувствительности, но только для 24 часов. Изотипический контроль NIP228 не оказывал никакого значительного эффекта на FCA-ответ в любой момент времени.
Пример 7. Фосфорилирование р38 под действием TNFα и NGF
В научной литературе высказано предположение, что фосфорилирование р38 играет важную роль в возникновении нейропатической боли. Например, показано, что лечение ингибиторами р38 предотвращало возникновение симптомов нейропатической боли в модели частичного повреждения нерва (Wen Y.R. et al., Anesthesiology, 2007, 107: 312-321) и в модели воспалительной нейропатии седалищного нерва (Milligan E.D. et al., J. Neurosci., 2003, 23: 1026-1040). В эксперименте, представленном в настоящей заявке, исследовали влияние TNFα, NGF и комбинации TNFα и NGF на фосфорилирование р38 в анализе с использованием клеточных культур. Кратко, клетки Neuroscreen-1 (субклон нейроэндокринных крысиных клеток PC-12) инкубировали с TNFα, NGF или комбинацией TNFα и NGF в возрастающих количествах. По окончании 20-минутного периода инкубации проводили количественное определение фосфорилированной р38 (фосфо-р38), используя анализ с применением гомогенной флуоресценции с разрешением по времени (HTRF) (Cisbio).
HTRF-анализ: после стимуляции TNFα, NGF или комбинацией TNFα и NGF, клеточные супернатанты быстро удаляли и клетки подвергали лизису в лизирующем буфере. Детекцию фосфорилированной митоген-активируемой протеинкиназы фосфо-р38 MAPK (Thr180/Tyr182) выполняли в лизатах в формате сэндвич-анализа, используя два антитела с разной специфичностью: антитело к фосфо-р38, конъюгированное с криптатом европия (донорным флуорофором), и антитело к р38 (общее), конъюгированное с d2 (акцепторым флуорофором). Антитела инкубировали с клеточными лизатами и HTRF-соотношения рассчитывали, исходя из данных измерений флуоресценции при 665 нм и 620 нм, полученных с использованием планшетного ридера EnVision Multilabel (Perkin Elmer).
Данные представлены в виде HTRF-соотношений, которые рассчитывают как соотношение между излучением при 665 нм и излучением при 620 нм. Тепловая карта, показывающая HTRF-соотношения для реакций фосфорилирования р38, показана на ФИГ. 14. Кривые зависимости доза-ответ, демонстрирующие эффект TNFα, NGF или комбинации TNFα и NGF, показаны на ФИГ. 15. Как можно видеть из ФИГ. 15, комбинированное влияние более высоких концентраций TNFα и NGF на процесс фосфорилирования р38 превышает предсказанную сумму сигналов от фосфо-р38, индуцированных и тем и другим фактором по отдельности. Эти данные позволяют предположить, что TNFα и NGF могут действовать совместно в отношении индуцирования фосфорилирования р38 и что эти два пути могут быть вовлечены в молекулярный механизм передачи сигнала, вызывающего боль.
Пример 8. Фосфорилирование ERK под действием TNFα и NGF
Подобно р38, ERK также активируется в процессе развития нейропатической боли. В эксперименте, представленном в настоящей заявке, исследовали влияние TNFα, NGF и комбинации TNFα и NGF на фосфорилирование ERK в анализе с использованием клеточных культур. Кратко, клетки Neuroscreen-1 (субклон нейроэндокринных крысиных клеток PC-12) инкубировали с TNFα, NGF или комбинацией TNFα и NGF в возрастающих количествах. По окончании 20-минутного периода инкубации проводили количественное определение фосфо-ERK, используя HTRF-анализ (Cisbio).
HTRF-анализ: после стимуляции клеточные супернатанты быстро удаляли и клетки подвергали лизису в лизирующем буфере. Детекцию фосфо-ERK MAPK (Thr202/Tyr204) выполняли в лизатах в формате сэндвич-анализа, используя два антитела с разной специфичностью: антитело к фосфо-ERK, конъюгированное с криптатом европия (донорным флуорофором), и антитело к ERK (общее), конъюгированное с d2 (акцепторным флуорофором). Антитела инкубировали с клеточными лизатами и HTRF-соотношения рассчитывали, исходя из данных измерений флуоресценции при 665 нм и 620 нм, полученных с использованием планшетного ридера EnVision Multilabel (Perkin Elmer).
Данные представлены в виде HTRF-соотношений, которые рассчитывают как соотношение между излучением при 665 нм и излучением при 620 нм. Тепловая карта, показывающая HTRF-соотношения для реакций фосфорилирования ERK, показана на ФИГ. 16. Кривые зависимости доза-ответ, демонстрирующие эффект TNFα, NGF или комбинации TNFα и NGF, показаны на ФИГ. 17. Как можно видеть из ФИГ. 17, небольшие количества одного TNFα не индуцировали фосфорилирование ERK, однако более высокие количества усиливали NGF-индуцированное фосфорилирование ERK. Эти данные позволяют предположить, что TNFα и NGF могут действовать совместно в отношении фосфорилирования ERK и что эти два пути могут быть вовлечены в молекулярный механизм передачи сигнала, вызывающего боль.
Перечень последовательностей
SEQ ID NO: 1 - NP_002497.2⎢ предшественник бета-фактора роста нервов (Homo sapiens)

SEQ ID NO: 2 - NP_000585.2⎢ фактор некроза опухоли (Homo sapiens)

SEQ ID NO: 3 - VH MEDI-578 (VH 1256A5)
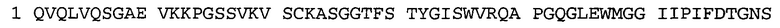

SEQ ID NO: 4 - VHCDR1 MEDI-578
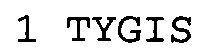
SEQ ID NO: 5 - VHCDR2 MEDI-578
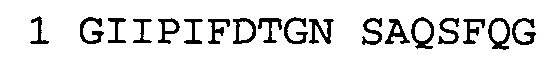
SEQ ID NO: 6 - VHCDR3 MEDI-578
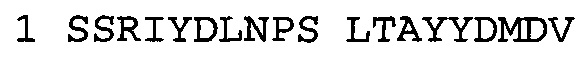
SEQ ID NO: 7 - VL MEDI-578 (VL 1256A5)

SEQ ID NO: 8 - VLCDR1 MEDI-578
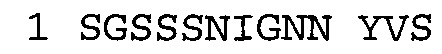
SEQ ID NO: 9 - VLCDR2 MEDI-578
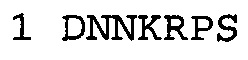
SEQ ID NO: 10 - VLCDR3 MEDI-578

SEQ ID NO: 11

SEQ ID NO: 12

SEQ ID NO: 13 - аминокислотная последовательность растворимого TNFR2

SEQ ID NO: 14 - TNFR2-Fc_VH#4 - аминокислотная последовательность

SEQ ID NO: 15 - состоящая из 15 аминокислот (а.к.) линкерная последовательность (Gly4Ser)3

SEQ ID NO: 16 - TNFR2-Fc_VH#4 - нуклеотидная последовательность
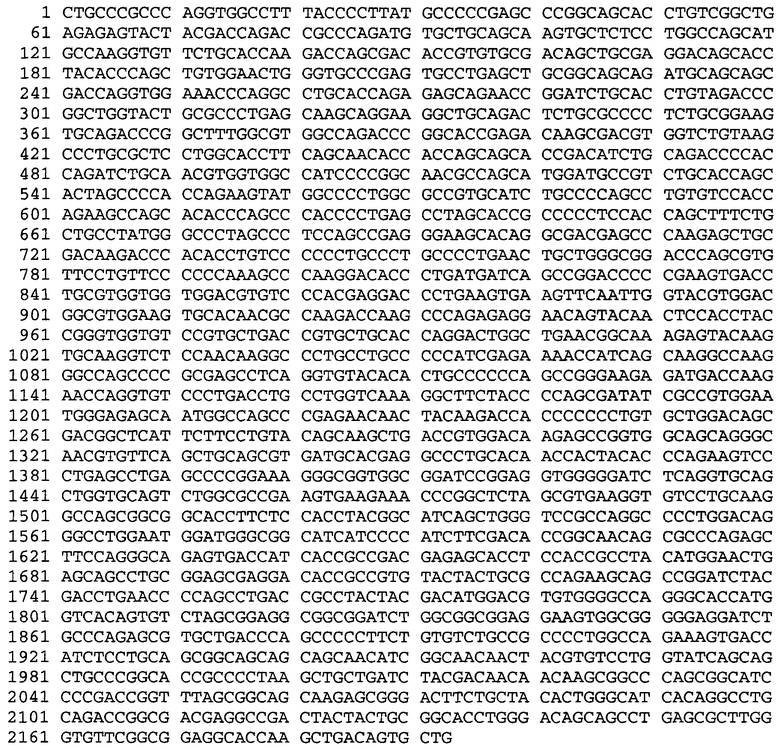
SEQ ID NO: 17 - TNFR2-Fc_varB - аминокислотная последовательность

SEQ ID NO: 18 - TNFR2-Fc_varB - нуклеотидная последовательность
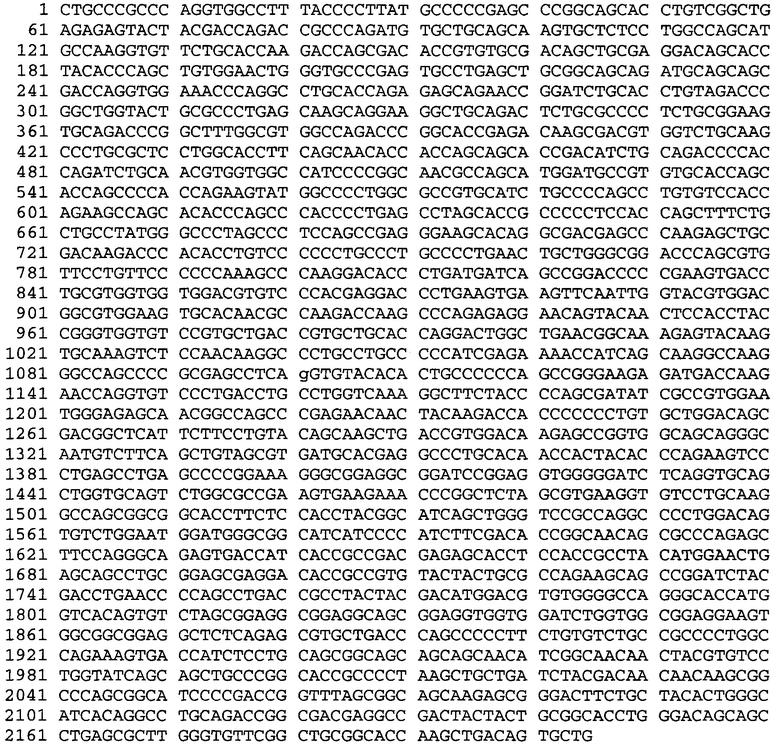
SEQ ID NO: 19 - состоящая из 20 а.к. линкерная последовательность (Gly4Ser)4

SEQ ID NO: 20 - ндимаб varB - аминокислотная последовательность L-цепи

SEQ ID NO: 21 - ндимаб varB - нуклеотидная последовательность L-цепи

SEQ ID NO: 22 - ндимаб varB - аминокислотная последовательность Н-цепи

SEQ ID NO: 23 - ндимаб varB - нуклеотидная последовательность Н-цепи
SEQ ID NO: 24 - аминокислотная последовательность VH NGF-NG

SEQ ID NO: 25 - нуклеотидная последовательность VH NGF-NG

SEQ ID NO: 26 - аминокислотная последовательность VL NGF-NG
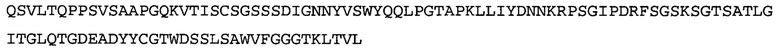
SEQ ID NO: 27 - нуклеотидная последовательность VL NGF-NG

SEQ ID NO: 28 - аминокислотная последовательность VH ндимаба

SEQ ID NO: 29 - аминокислотная последовательность VL ндимаба
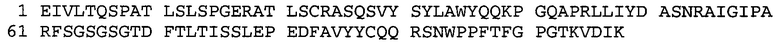
SEQ ID NO: 30 - аминокислотная последовательность VH 1126F1
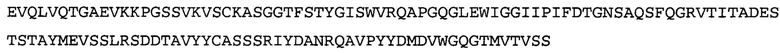
SEQ ID NO: 31 - аминокислотная последовательность VL 1126F1
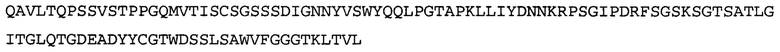
SEQ ID NO: 32 - аминокислотная последовательность VH 1126G5
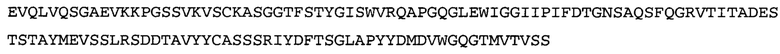
SEQ ID NO: 33 - аминокислотная последовательность VL 1126G5
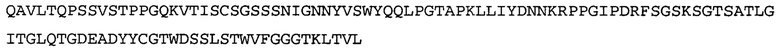
SEQ ID NO: 34 - аминокислотная последовательность VH 1126H5
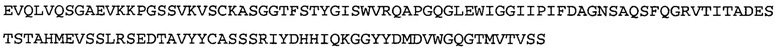
SEQ ID NO: 35 - аминокислотная последовательность VL 1126H5
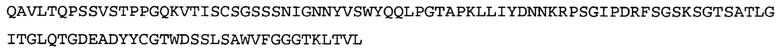
SEQ ID NO: 36 - аминокислотная последовательность VH 1127D9

SEQ ID NO: 37 - аминокислотная последовательность VL 1127D9

SEQ ID NO: 38 - аминокислотная последовательность VH 1127F9

SEQ ID NO: 39 - аминокислотная последовательность VL 1127F9
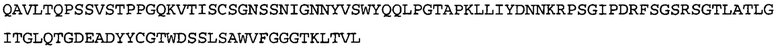
SEQ ID NO: 40 - аминокислотная последовательность VH 1131D7

SEQ ID NO: 41 - аминокислотная последовательность VL 1131D7
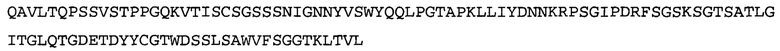
SEQ ID NO: 42 - аминокислотная последовательность VH 1131H2
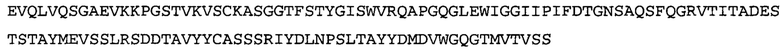
SEQ ID NO: 43 - аминокислотная последовательность VL 1131H2
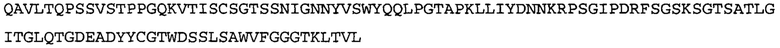
SEQ ID NO: 44 - аминокислотная последовательность VH 132A9
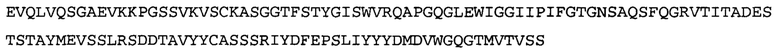
SEQ ID NO: 45 - аминокислотная последовательность VL 132A9
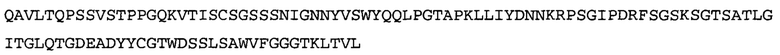
SEQ ID NO: 46 - аминокислотная последовательность VH 1132H9

SEQ ID NO: 47 - аминокислотная последовательность VL 1132H9
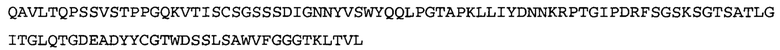
SEQ ID NO: 48 - аминокислотная последовательность VH 1133C11
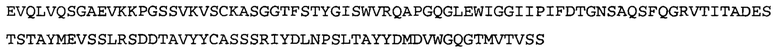
SEQ ID NO: 49 - аминокислотная последовательность VL 1133C11
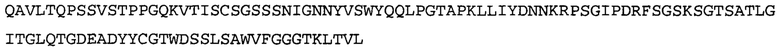
SEQ ID NO: 50 - аминокислотная последовательность VH 1134D9

SEQ ID NO: 51 - аминокислотная последовательность VL 1134D9
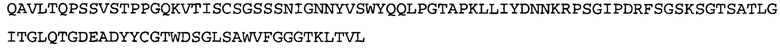
SEQ ID NO: 52 - аминокислотная последовательность VH 1145D1
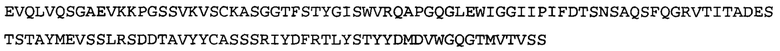
SEQ ID NO: 53 - аминокислотная последовательность VL 1145D1
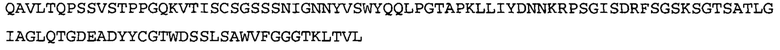
SEQ ID NO: 54 - аминокислотная последовательность VH 1146D7

SEQ ID NO: 55 - аминокислотная последовательность VL 1146D7
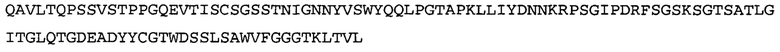
SEQ ID NO: 56 - аминокислотная последовательность VH 1147D2

SEQ ID NO: 57 - аминокислотная последовательность VL 147D2
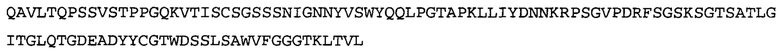
SEQ ID NO: 58 - аминокислотная последовательность VH 1147G9

SEQ ID NO: 59 - аминокислотная последовательность VL 1147G9
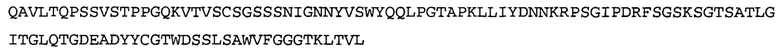
SEQ ID NO: 60 - аминокислотная последовательность VH 1150F1

SEQ ID NO: 61 - аминокислотная последовательность VL 1150F1
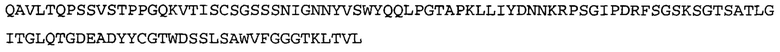
SEQ ID NO: 62 - аминокислотная последовательность VH 1152H5
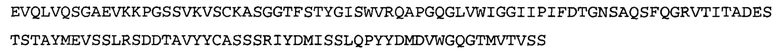
SEQ ID NO: 63 - аминокислотная последовательность VL 1152H5
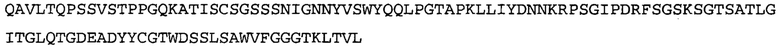
SEQ ID NO: 64 - аминокислотная последовательность VH 1155H1
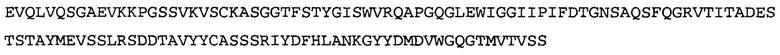
SEQ ID NO: 65 - аминокислотная последовательность VL 1155H1
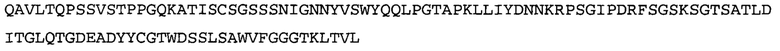
SEQ ID NO: 66 - аминокислотная последовательность VH 1158A1
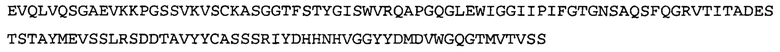
SEQ ID NO: 67 - аминокислотная последовательность VL 1158A1
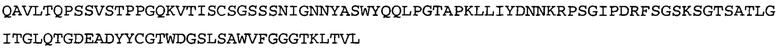
SEQ ID NO: 68 - аминокислотная последовательность VH 1160E3

SEQ ID NO: 69 - аминокислотная последовательность VL 1160E3
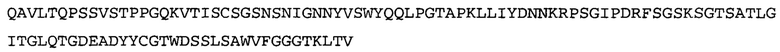
SEQ ID NO: 70 - аминокислотная последовательность VH 1165D4

SEQ ID NO: 71 - аминокислотная последовательность VL 1165D4
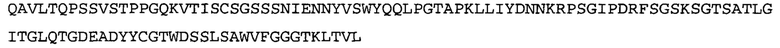
SEQ ID NO: 72 - аминокислотная последовательность VH 1175H8

SEQ ID NO: 73 - аминокислотная последовательность VL 1175H8
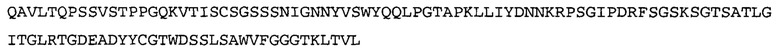
SEQ ID NO: 74 - аминокислотная последовательность VH 1211G10

SEQ ID NO: 75 - аминокислотная последовательность VL 1211G10

SEQ ID NO: 76 - аминокислотная последовательность VH 1214A1
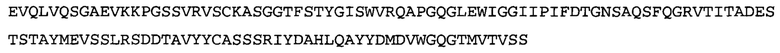
SEQ ID NO: 77 аминокислотная последовательность VL 1214A1

SEQ ID NO: 78 - аминокислотная последовательность VH 1214D10
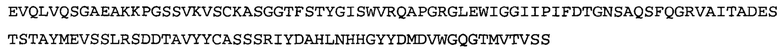
SEQ ID NO: 79 - аминокислотная последовательность VL 1214D10

SEQ ID NO: 80 - аминокислотная последовательность VH 1218H5

SEQ ID NO: 81 - аминокислотная последовательность VL 1218H5
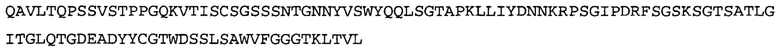
SEQ ID NO: 82 - аминокислотная последовательность VH 1230H7
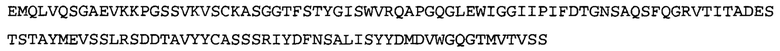
SEQ ID NO: 83 - аминокислотная последовательность VL 1230H7
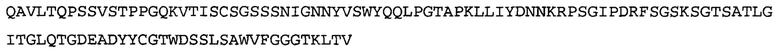
SEQ ID NO: 84 - аминокислотная последовательность VH 1083H4
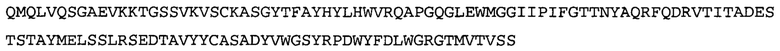
SEQ ID NO: 85 - аминокислотная последовательность VL 1083H4
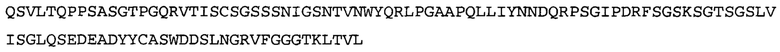
SEQ ID NO: 86 - аминокислотная последовательность VH 1227H8

SEQ ID NO: 87 - аминокислотная последовательность VL 1227H8
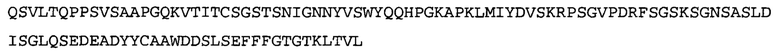
SEQ ID NO: 88 - HCDR1 NGF-NG
SEQ ID NO: 89 - HCDR2 NGF-NG
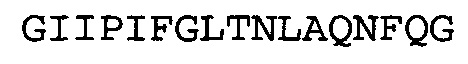
SEQ ID NO: 90 - HCDR3 NGF-NG
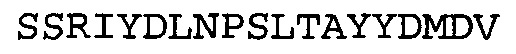
SEQ ID NO: 91 - LCDR1 NGF-NG
SEQ ID NO: 92 - LCDR2 NGF-NG
SEQ ID NO: 93 - LCDR3 NGF-NG
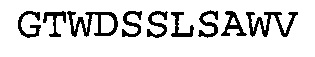
SEQ ID NO: 94 - аминокислотная последовательность VH MEDI-578 с заменой G→C

SEQ ID NO: 95 - аминокислотная последовательность VL MEDI-578 с заменой G→C

SEQ ID NO: 96 - аминокислотная последовательность VH 1230D8

SEQ ID NO: 97 - аминокислотная последовательность VL1230D8

SEQ ID NO: 98

SEQ ID NO: 99 - TNFR2-Fc_varB - кодон-оптимизированная нуклеотидная последовательность


Объем данного изобретения не следует ограничивать конкретными описанными аспектами, которые предназначены просто в качестве иллюстраций отдельных аспектов изобретения, и любые композиции или способы, которые являются функционально эквивалентными, включены в объем данного описания. В действительности, различные модификации изобретения, помимо показанных и изложенных в данном описании, станут очевидны специалистам в данной области техники из приведенного выше описания и прилагаемых фигур. Подразумевается, что такие модификации попадают в объем прилагаемой формулы изобретения.
Все публикации и патентные заявки, упомянутые в данном описании, включены в данное описание посредством ссылки в той же степени, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно указана как включенная посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| Химерный белок, составленный из домена антагониста NGF и домена антагониста TNFα | 2015 |
|
RU2788122C2 |
| АНТАГОНИСТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2005 |
|
RU2401842C2 |
| ПАРТНЕРЫ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С NGF | 2006 |
|
RU2406728C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ hTNFα С ИХ ПОСРЕДСТВОМ (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, РЕКОМБИНАНТНОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА (ВАРИАНТЫ) | 2005 |
|
RU2458704C9 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2644235C2 |
| АНТАГОНИСТЫ ИЛ-6 И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2670943C9 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА НЕРВОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2640252C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И СОДЕРЖАЩИХ ЕГО КОМПОЗИЦИЙ | 2006 |
|
RU2429013C2 |











































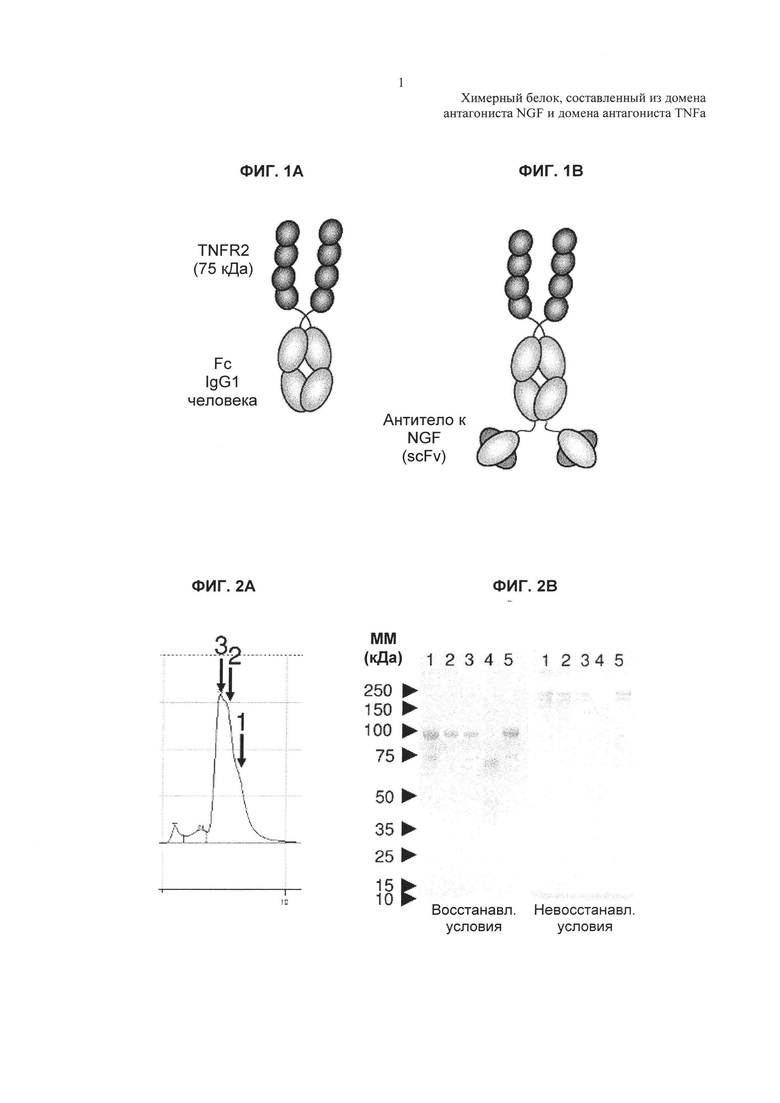
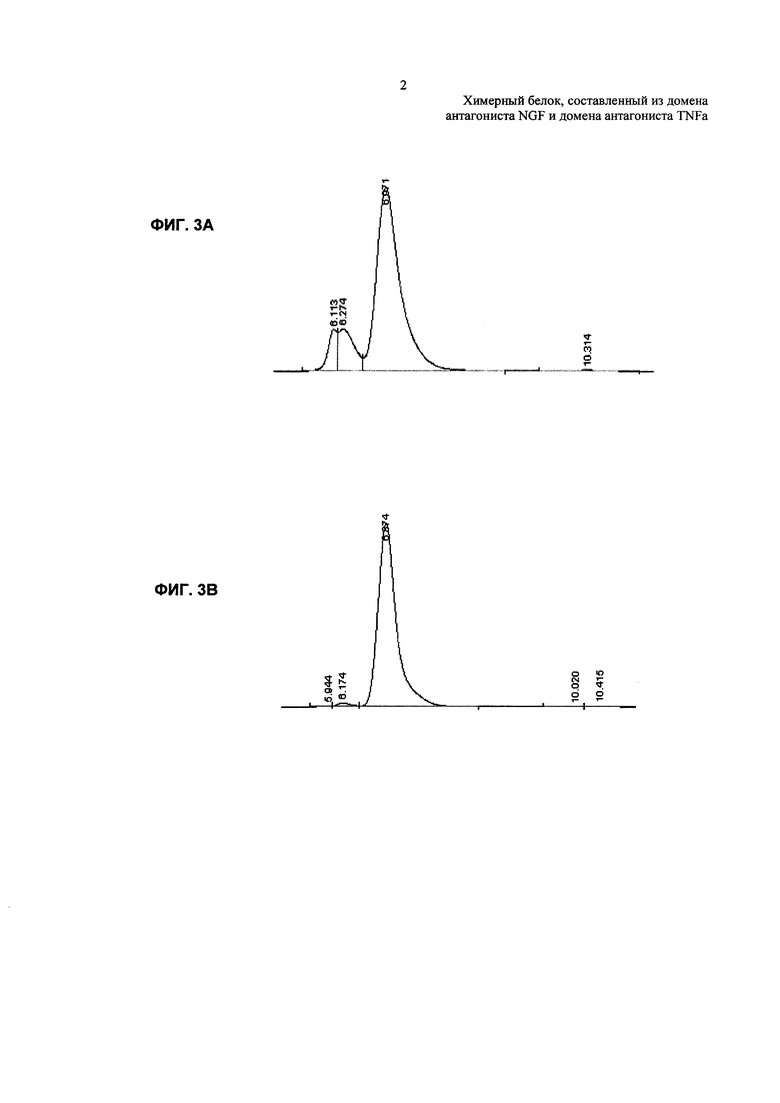
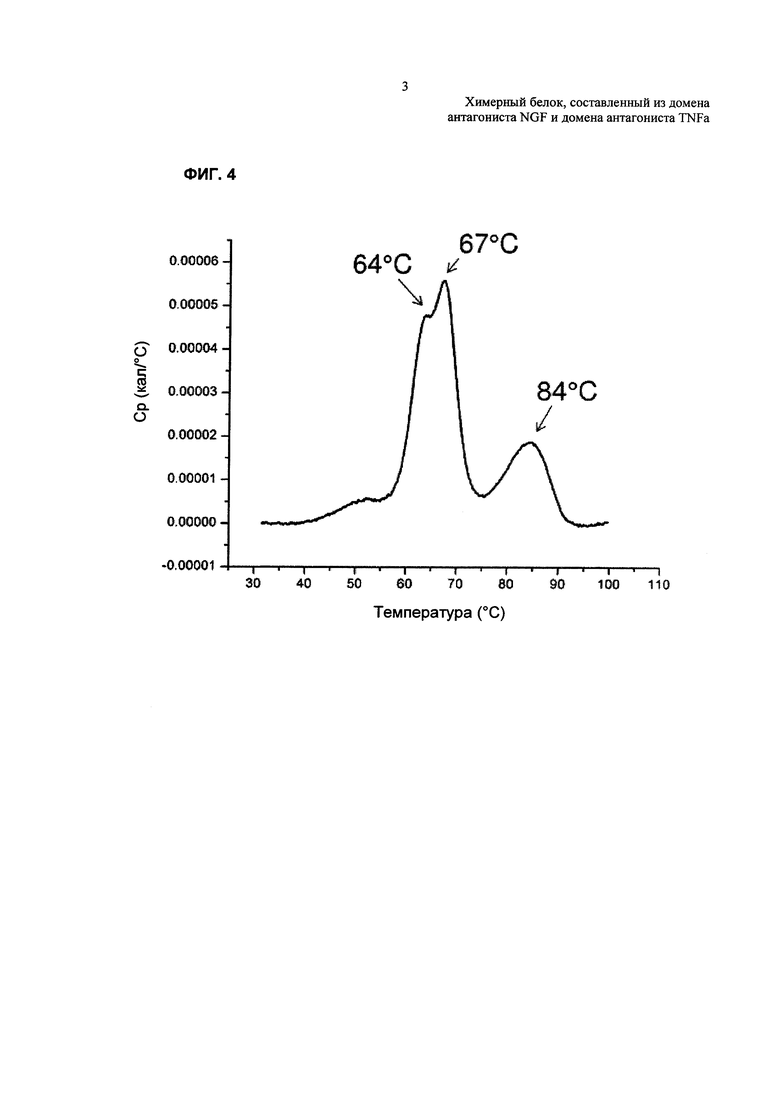
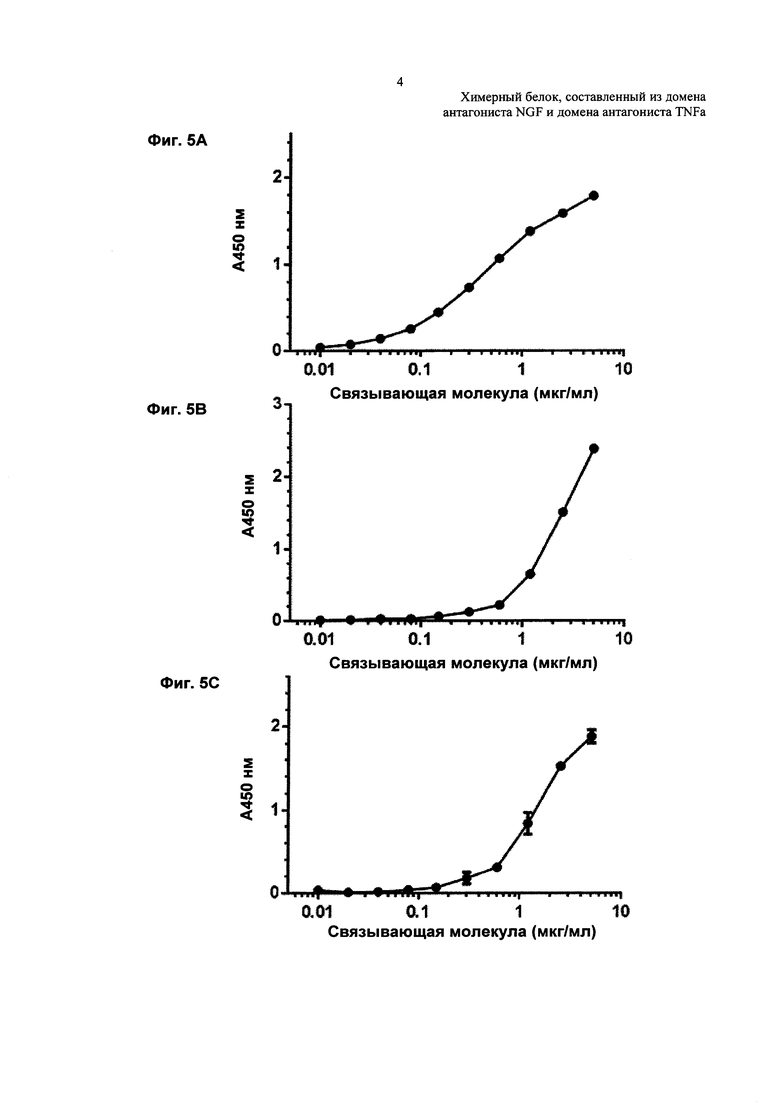
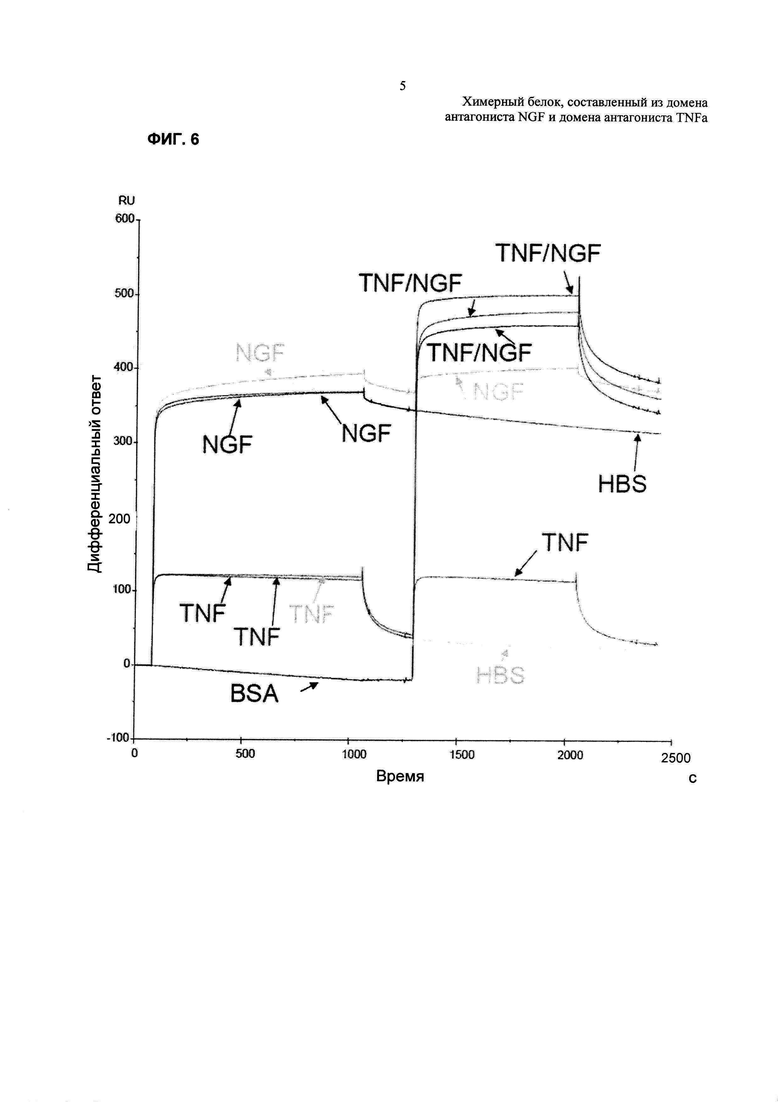
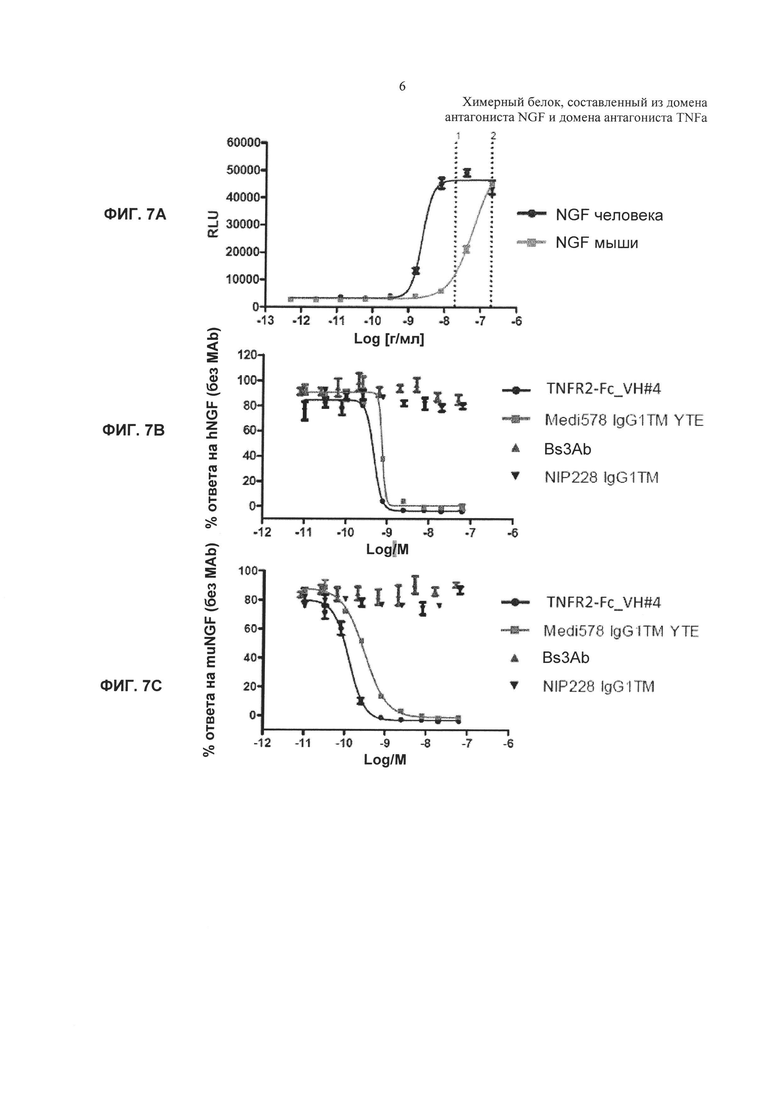
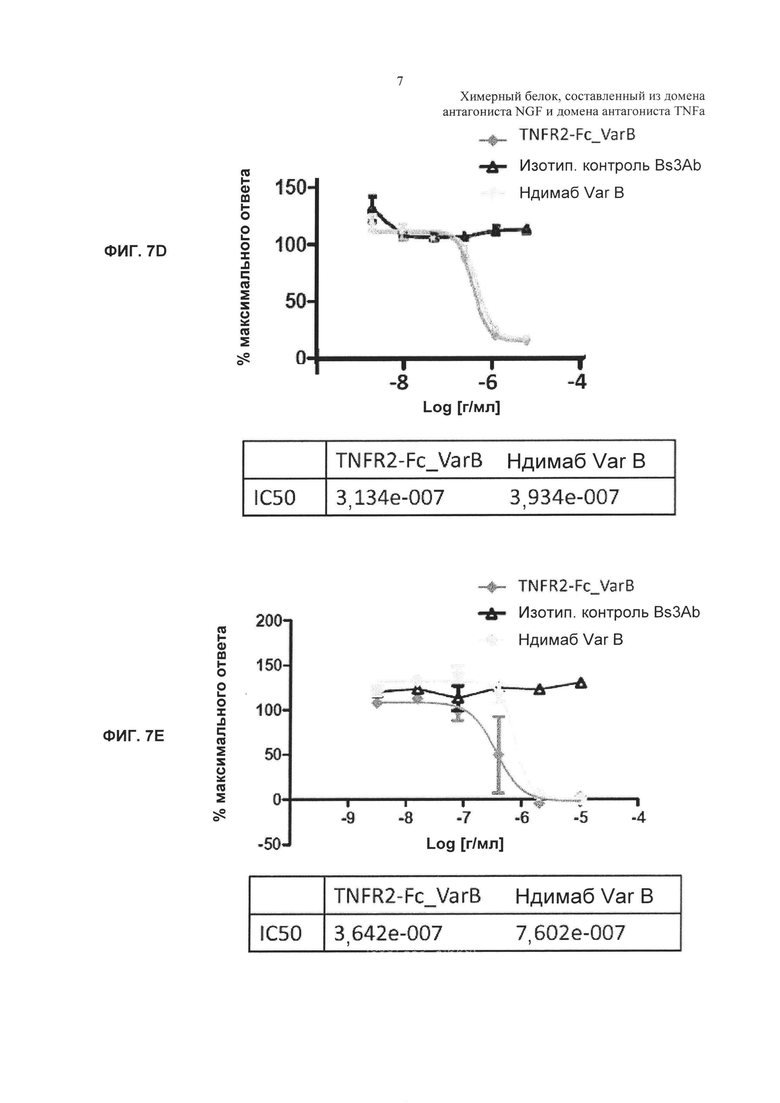
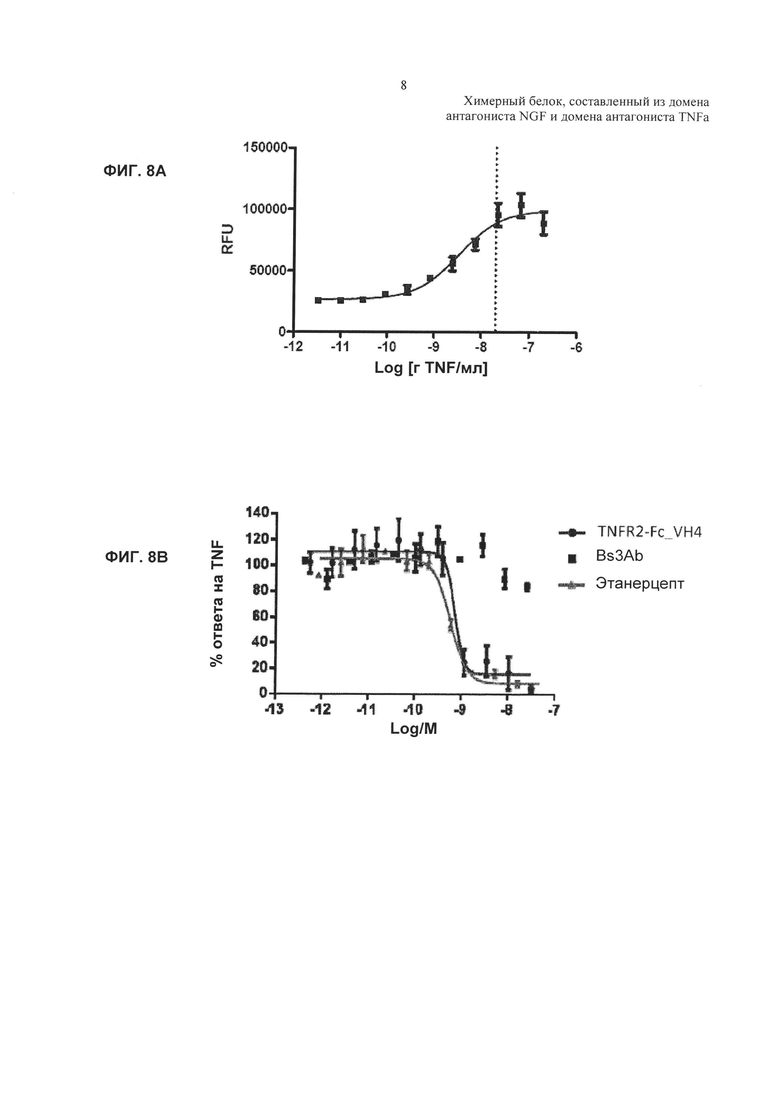
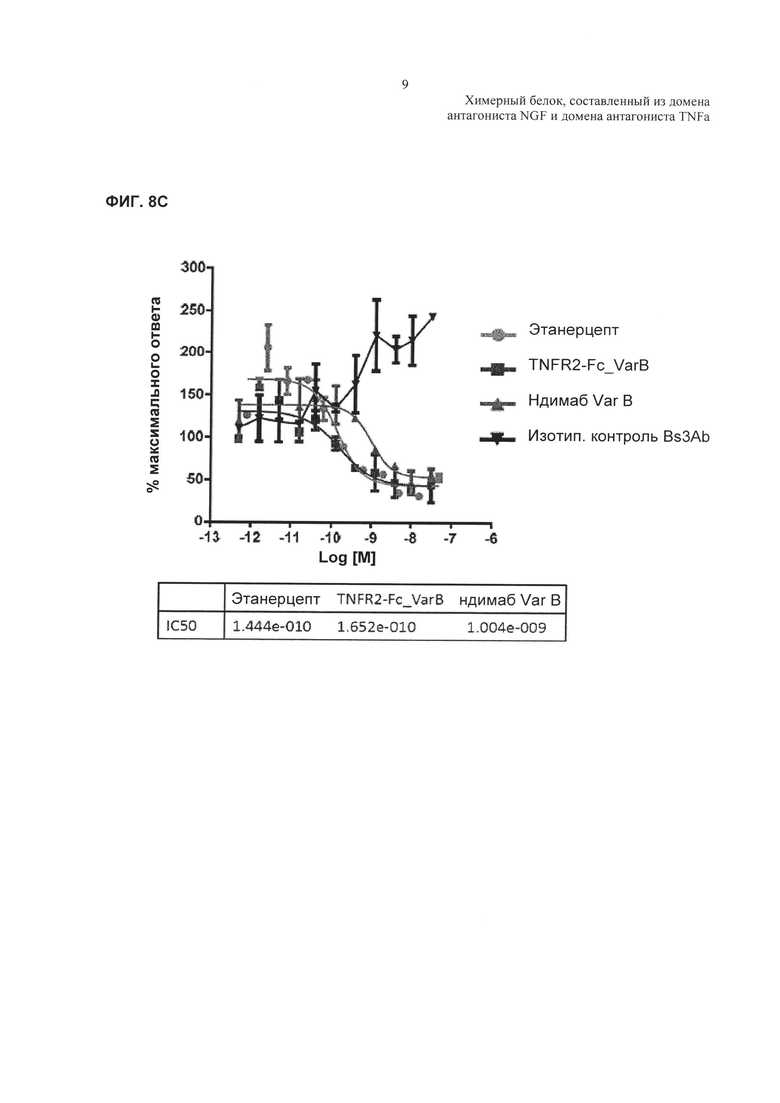
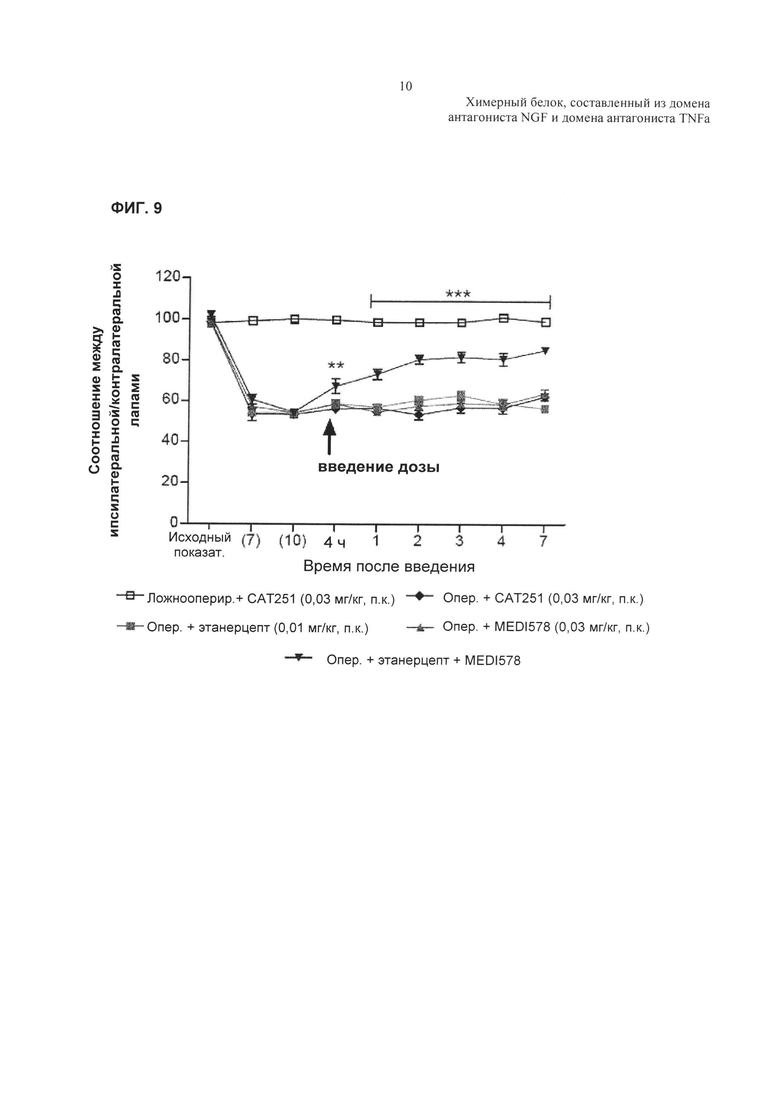
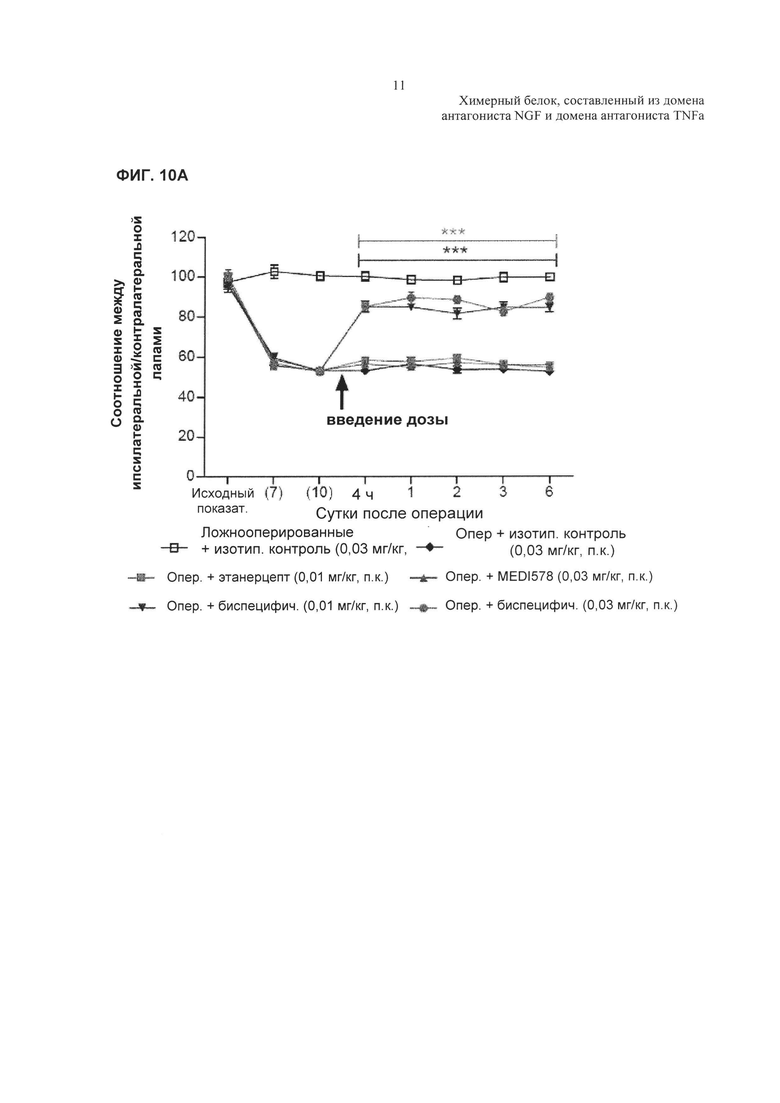
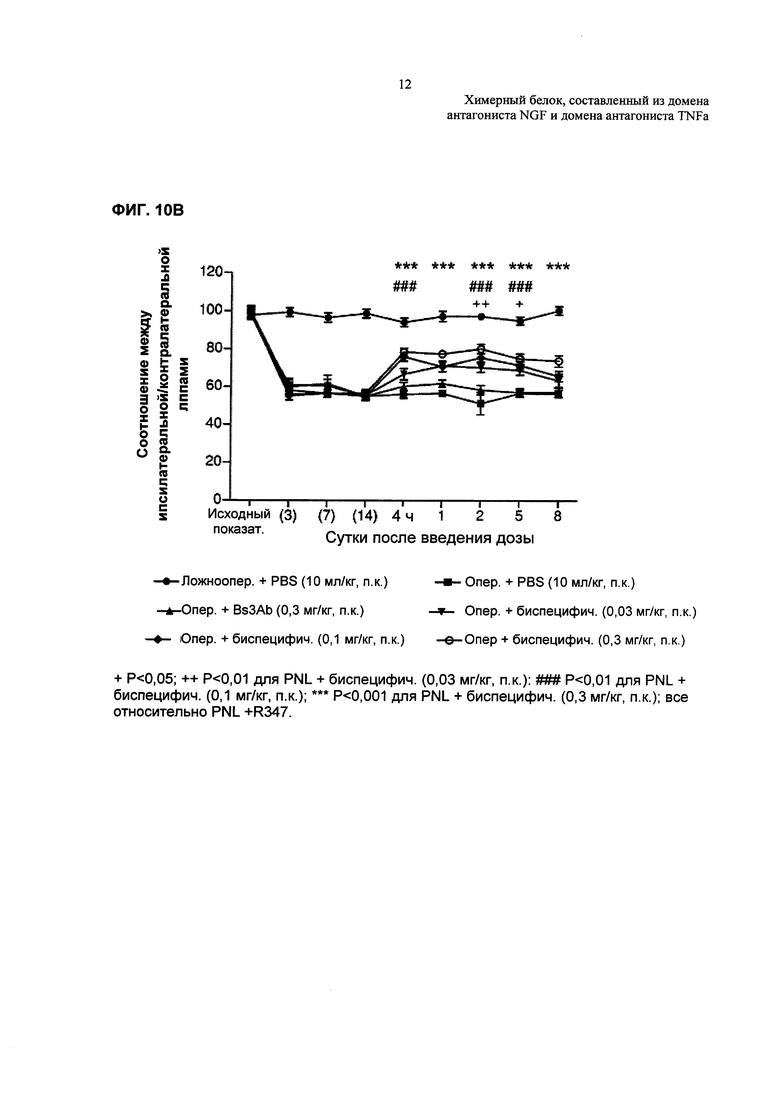
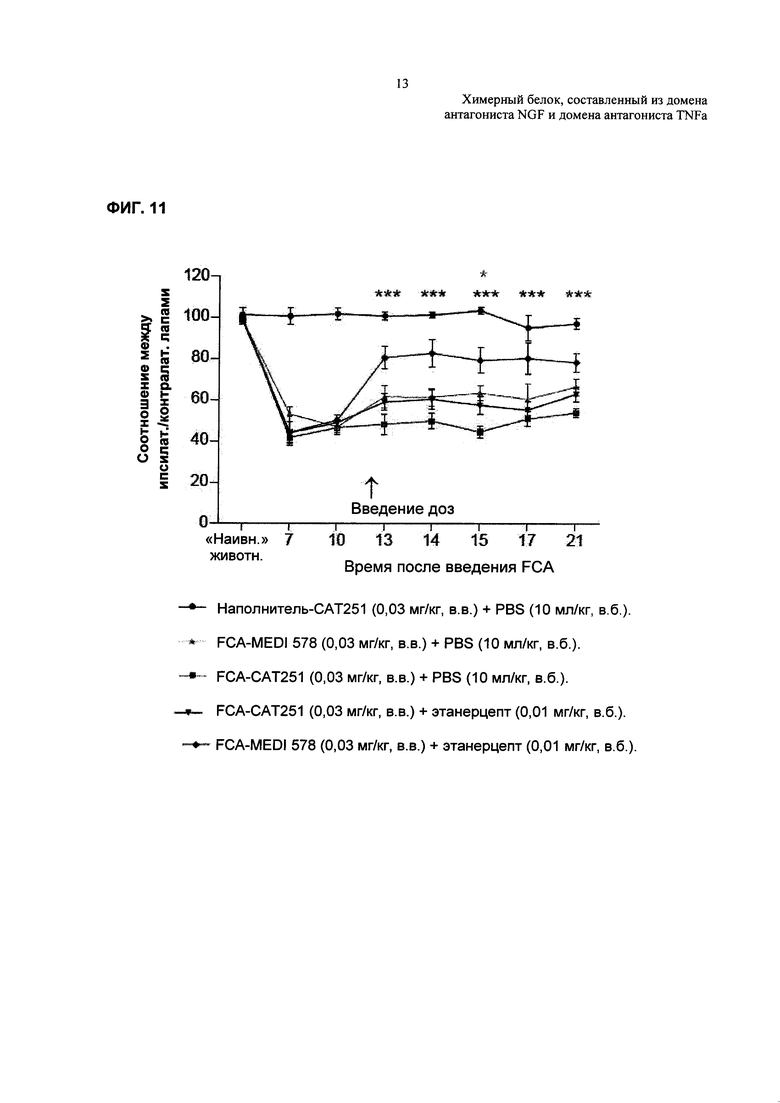
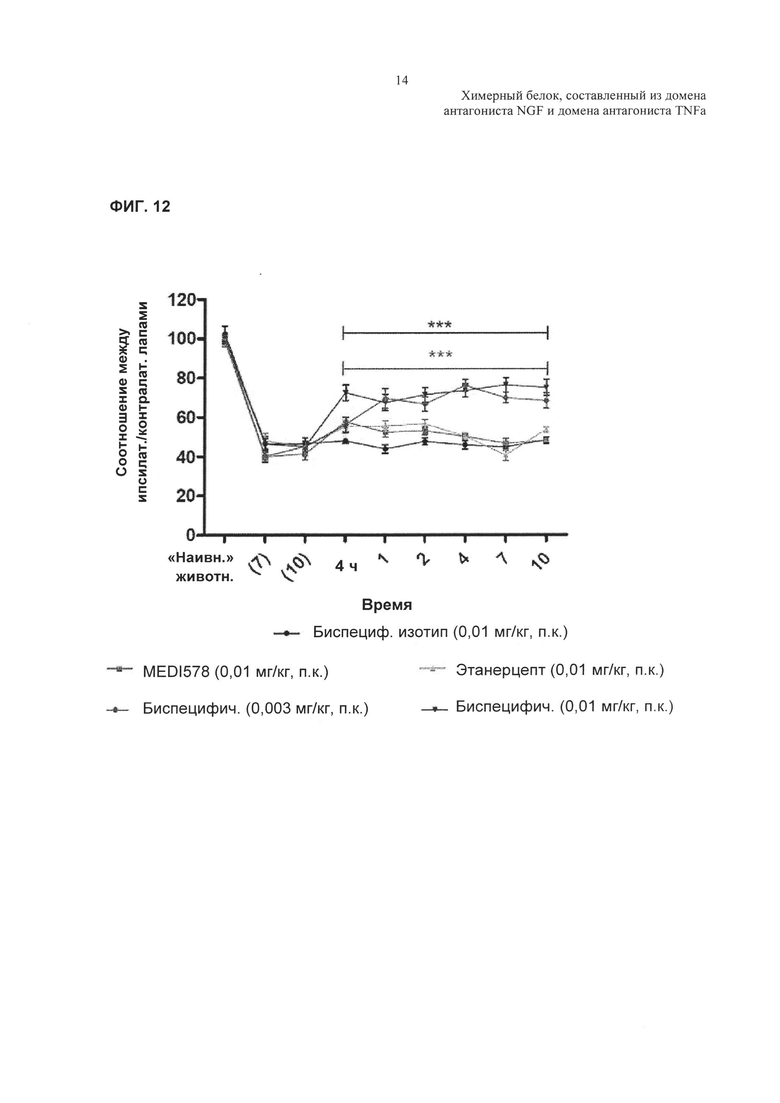
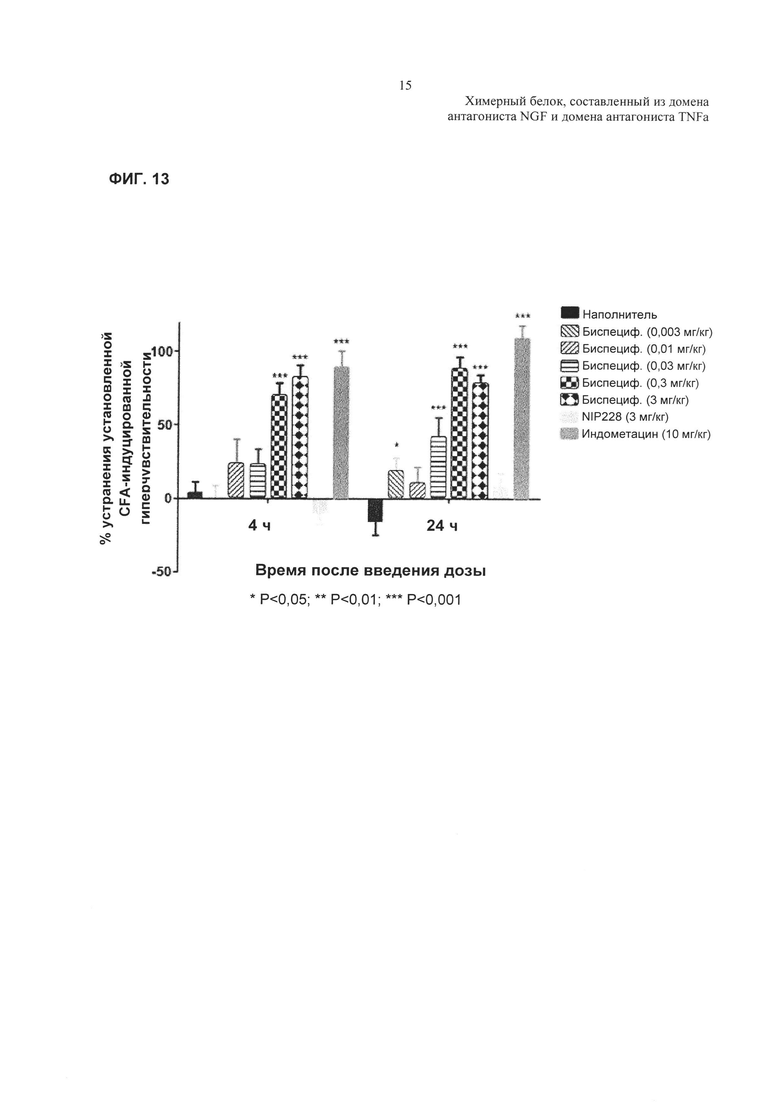
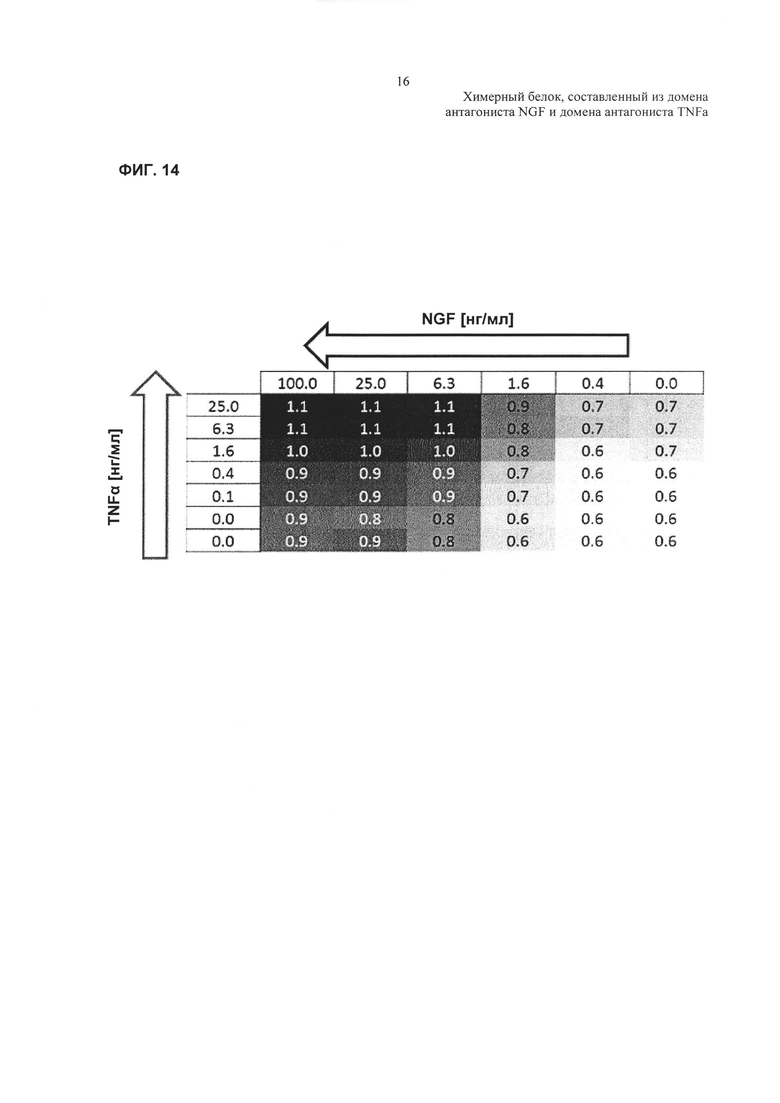
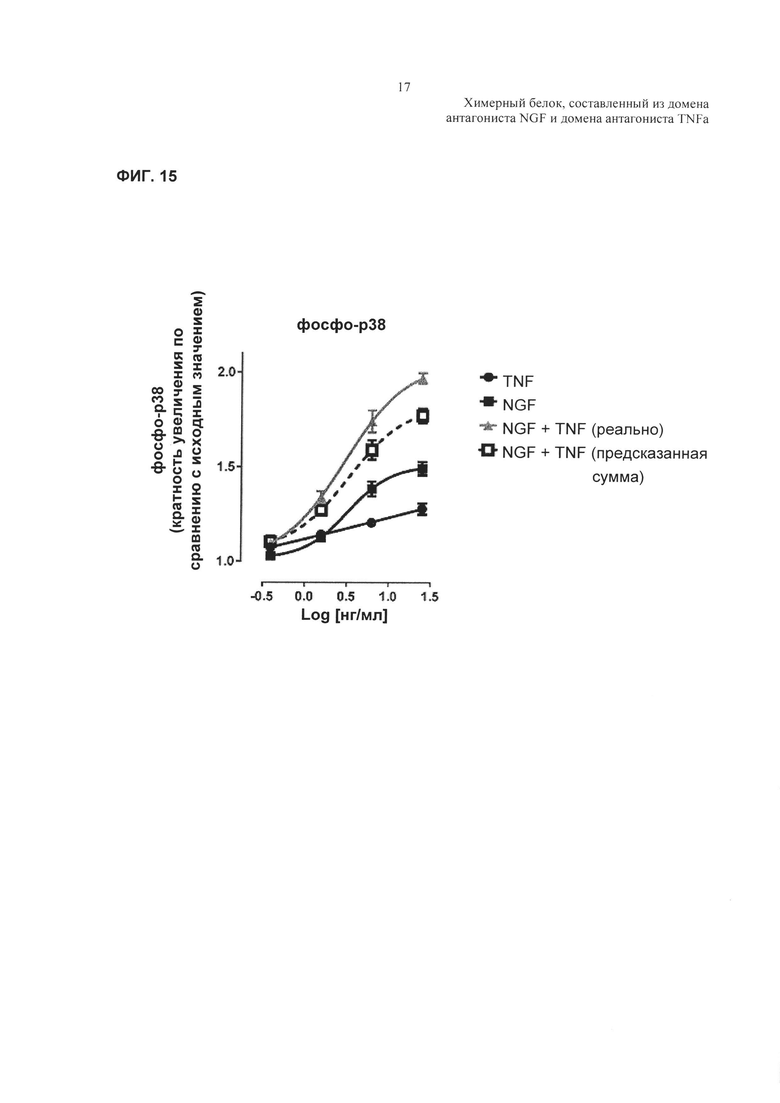
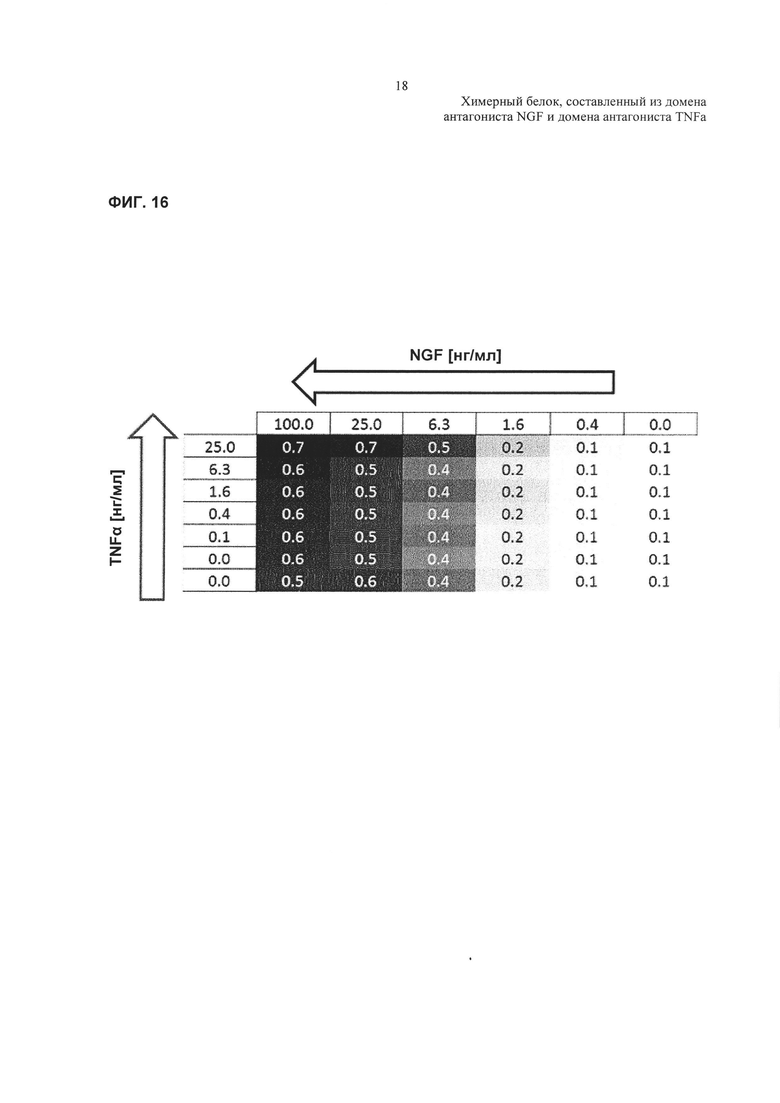
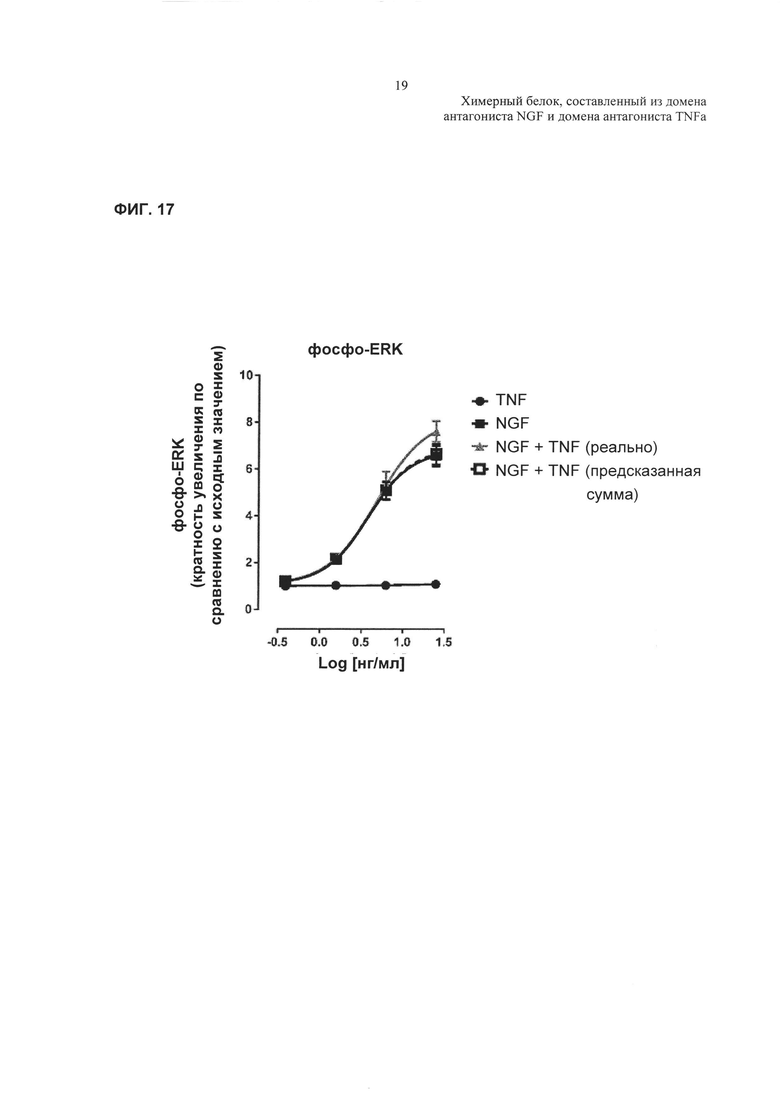
Изобретение относится к биотехнологии и представляет собой связывающую молекулу, которая содержит домен антагониста NGF (фактор роста нервов) и домен антагониста TNFα (α-фактор некроза опухоли), где антагонист NGF представляет собой антитело к NGF или его антиген-связывающий фрагмент, и где домен антагониста TNFα содержит растворимый TNFα-связывающий фрагмент TNFR; где антитело к NGF или его фрагмент содержит вариабельный домен тяжелой цепи (VH-домен), содержащий совокупность гипервариабельных участков (CDR) HCDR1, HCDR2, HCDR3, и вариабельный домен легкой цепи (VL-домен), содержащий совокупность CDR LCDR1, LCDR2 и LCDR3, причем HCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 5, HCDR3 имеет аминокислотную последовательность любой из SEQ ID NO: 6, 11 или 12, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 8, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 9, и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 10; где домен антагониста TNFα содержит аминокислотную последовательность, идентичную по меньшей мере на 80% последовательности SEQ ID NO: 13, или ее функциональный фрагмент; и где указанная связывающая молекула способна к связыванию NGF и TNFα. Изобретение относится также к аналгезирующей композиции, содержащей связывающую молекулу и носитель. Введение антагониста NGF и антагониста TNFα, предложенных в данном изобретении, может контролировать боль у субъекта более эффективно, чем введение эквивалентного количества антагониста NGF или антагониста TNFα в виде монотерапии. 7 н. и 25 з.п. ф-лы, 17 ил., 1 табл., 8 пр.
1. Связывающая молекула, содержащая домен антагониста NGF (фактор роста нервов) и домен антагониста TNFα (α-фактор некроза опухоли), где антагонист NGF представляет собой антитело к NGF или его антиген-связывающий фрагмент, и где домен антагониста TNFα содержит растворимый TNFα-связывающий фрагмент TNFR; где антитело к NGF или его фрагмент содержит вариабельный домен тяжелой цепи (VH-домен), содержащий совокупность гипервариабельных участков (CDR) HCDR1, HCDR2, HCDR3, и вариабельный домен легкой цепи (VL-домен), содержащий совокупность CDR LCDR1, LCDR2 и LCDR3, причем HCDR1 имеет аминокислотную последовательность SEQ ID NO: 4, HCDR2 имеет аминокислотную последовательность SEQ ID NO: 5, HCDR3 имеет аминокислотную последовательность любой из SEQ ID NO: 6, 11 или 12, LCDR1 имеет аминокислотную последовательность SEQ ID NO: 8, LCDR2 имеет аминокислотную последовательность SEQ ID NO: 9, и LCDR3 имеет аминокислотную последовательность SEQ ID NO: 10; где домен антагониста TNFα содержит аминокислотную последовательность, идентичную по меньшей мере на 80% последовательности SEQ ID NO: 13, или ее функциональный фрагмент; и
где указанная связывающая молекула способна к связыванию NGF и TNFα.
2. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент может ингибировать связывание NGF с TrkA (тирозинкиназа А), с p75NRT (рецептор нейротрофинов р75) или как с TrkA, так и с p75NRT.
3. Связывающая молекула по п. 1, которая предпочтительно блокирует связывание NGF с TrkA по сравнению со связыванием NGF с p75NRT.
4. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент связывается с NGF человека с аффинностью примерно 0,25-0,44 нМ.
5. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент содержит VH-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 3 или 94.
6. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 7 или 95.
7. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент содержит VH-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 3 или 94, и где антитело к NGF или его фрагмент содержит VL-домен, имеющий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 7 или 95.
8. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент представляет собой полноразмерное антитело в формате H2L2, Fab-фрагмент, Fab'-фрагмент, F(ab)2-фрагмент или одноцепочечный Fv (scFv)-фрагмент.
9. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент является гуманизированным, химерным, приматизированным или полностью человеческим.
10. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент представляет собой scFv-фрагмент антитела к NGF, который содержит, в направлении от N-конца к С-концу, VH-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 3, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15), и VL-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 7.
11. Связывающая молекула по п. 1, где антитело к NGF или его фрагмент представляет собой scFv-фрагмент антитела к NGF, который содержит, в направлении от N-конца к С-концу, VH-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 94, линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4 (SEQ ID NO: 19), и VL-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 95.
12. Связывающая молекула по любому из пп. 1-11, где TNFR представляет собой TNFR-2.
13. Связывающая молекула по п. 12, где TNFR-2 фрагмент слит с Fc-доменом иммуноглобулина.
14. Связывающая молекула по п. 13, где Fc-домен иммуноглобулина представляет собой Fc-домен IgG1 человека.
15. Связывающая молекула по п. 12, где антагонист TNFα содержит аминокислотную последовательность, идентичную по меньшей мере на 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности, приведенной в SEQ ID NO: 13, или ее функциональный фрагмент.
16. Связывающая молекула по любому из пп. 1-11, которая содержит слитый белок, содержащий антагонист NGF, слитый с антагонистом TNFα через линкер.
17. Связывающая молекула по п. 16, содержащая гомодимер слитого белка.
18. Связывающая молекула по п. 1, которая содержит гомодимер слитого полипептида, содержащего, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент TNFR-2, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% последовательности, соответствующей аминокислотам 1-235 SEQ ID NO: 13, Fc-домен IgG1 человека, линкерную последовательность, состоящую из 10 аминокислот (GGGGS)2 (SEQ ID NO: 98), VH-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 3 или 94, линкерную последовательность, состоящую из 15 аминокислот (GGGGS)3 (SEQ ID NO: 15), и VL-домен, содержащий аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 7 или 95.
19. Связывающая молекула по п. 1, которая содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95%, 97%, 99% или 100% аминокислотной последовательности SEQ ID NO: 14.
20. Связывающая молекула по п. 1, которая содержит гомодимер слитого полипептида, содержащего, в направлении от N-конца к С-концу, TNFα-связывающий фрагмент TNFR-2 с молекулярной массой 75 кДа, содержащий аминокислотную последовательность SEQ ID NO: 13, линкерную последовательность, состоящую из 10 аминокислот (GGGGS)2 (SEQ ID NO: 98), VH-домен, содержащий аминокислотную последовательность SEQ ID NO: 94, линкерную последовательность, состоящую из 20 аминокислот (GGGGS)4 (SEQ ID NO: 19), и VL-домен, содержащий аминокислотную последовательность SEQ ID NO: 95.
21. Связывающая молекула по любому из пп. 1-11, где остаток глицина в положении аминокислоты, соответствующем положению 102, 103 или 104 SEQ ID NO: 7, заменен на остаток цистеина.
22. Связывающая молекула по любому из пп. 1-11, где остаток глицина в положении аминокислоты, соответствующем положению 44 SEQ ID NO: 3, заменен на остаток цистеина.
23. Связывающая молекула по любому из пп. 1-11, где остаток глицина в положении аминокислоты, соответствующем положению 102, 103 или 104 SEQ ID NO: 7, заменен на остаток цистеина, и где остаток глицина в положении аминокислоты, соответствующем положению 44 SEQ ID NO: 3, заменен на остаток цистеина.
24. Связывающая молекула по любому из пп. 1-11, которая содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность SEQ ID NO: 17.
25. Связывающая молекула по любому из пп. 1-11, которая содержит гомодимер слитого полипептида, содержащего аминокислотную последовательность, идентичную по меньшей мере на 80%, 85%, 90%, 95% или 99% аминокислотной последовательности SEQ ID NO: 17.
26. Выделенный полинуклеотид, кодирующий связывающую молекулу по любому из пп. 1-25.
27. Экспрессирующий вектор, содержащий полинуклеотид по п. 26, функционально связанный с промотором.
28. Клетка-хозяин, содержащая полинуклеотид по п. 26 или вектор по п. 27, которая продуцирует связывающую молекулу, способную к связыванию NGF и TNFα.
29. Аналгезирующая композиция, содержащая связывающую молекулу по любому из пп. 1-25 и носитель.
30. Набор, содержащий связывающую молекулу по любому из пп. 1-25, полинуклеотид по п. 26, вектор по п. 27 и/или композицию по п. 29, где указанный набор дополнительно содержит один или несколько контейнеров, и где связывающая молекула, присутствующая в наборе или кодируемая полинуклеотидом или вектором, предназначена для использования в качестве аналгезирующего средства.
31. Применение связывающей молекулы в изготовлении лекарственного средства для предупреждения, ослабления, уменьшения интенсивности или устранения боли у субъекта, где связывающая молекула представляет собой связывающую молекулу по любому из пп. 1-25.
32. Применение по п. 31, где боль представляет собой острую боль, кратковременную боль, постоянную ноцицептивную боль либо постоянную или хроническую нейропатическую боль.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛИ, ВЫЗВАННОЙ ОСТЕОАРТРИТОМ, ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА ФАКТОРА РОСТА НЕРВОВ И СОДЕРЖАЩИХ ЕГО КОМПОЗИЦИЙ | 2006 |
|
RU2429013C2 |

