ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
1. Область изобретения
Настоящее изобретение относится к индуцированию дифференцировки стволовых клеток взрослого человека в клетки, секретирующие инсулин.
2. Предпосылки изобретения
Сахарный диабет 1 типа, который также называется инсулинозависимым сахарным диабетом, отличается утратой инсулин-продуцирующих бета-клеток, которая обычно вызвана аутоиммунной атакой островков Лангерганса поджелудочной железы и приводит к дефициту инсулина. В организме инсулин играет ключевую роль в гомеостазе уровня сахара в крови. Когда инсулин отсутствует или продуцируется в недостаточном количестве, происходит повышение уровня сахара в крови, что приводит к осложнениям в различных органах, таких как почки, нервы, сетчатка и т.д. В основном, лечение сахарного диабета 1 типа основано на использовании инсулина; так как требуется делать инъекции в течение всей жизни пациента, применение инсулина не является основным средством для лечения сахарного диабета 1 типа. С другой стороны, трансплантация экзогенной поджелудочной железы или бета-клеток позволяет лечить сахарный диабет 1 типа. Однако на эту меру налагается множество гистологических ограничений, которые связаны с хирургической трансплантацией экзогенной поджелудочной железы. Например, трансплантированные клетки могут разрушаться при помощи аутоиммунных реакций реципиента. В последнее время множество исследований направлено на разработку способов лечения диабета с использованием стволовых клеток, типичными представителями которых являются эмбриональные и взрослые стволовые клетки. Сообщалось об исследованиях, в которых эмбриональные стволовые клетки дифференцировали в инсулин-продуцирующие клетки ex vivo, с помощью которых можно лечить гипергликемию, что выяснилось при трансплантации мышам с диабетом [Fujikawa T, Oh SH, Pi L, Hatch HM, Shupe T, Petersen BE (2005) Teratoma formation leads to failure of treatment for type I diabetes using embryonic stem cell-derived insulin-producing cells. Am J Pathol166, 1781-1791; D'Amour KA, Bang AG, Eliazer S, Kelly OG, Agulnick AD, Smart NG, Moorman MA, Kroon E, Carpenter MK, Baetge ЕЕ (2006) Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol 24, 1392-1401]. Однако применение эмбриональных стволовых клеток в клинике ведет к значительным проблемам, поскольку они вызывают образование рака при трансплантации [Kroon E, Martinson LA, Kadoya K, Bang AG, Kelly OG, Eliazer S, Young H, Richardson M, Smart NG, Cunningham J, Agulnick AD, D'Amour KA, Carpenter MK, Baetge ЕЕ (2008) Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat Biotechnol. 26, 397-398].
Сообщалось о том, что эктодермальные клетки-предшественники могут дифференцироваться в инсулин-продуцирующие клетки [Hori Y, Gu X, Xie X, and Kim SK. (2005) Differentiation of insulin-producing cells from human neural progenitor cells. PLOS Med. 2, 103] и что эпителиальные клетки протока поджелудочной железы могут дифференцироваться в структуры, аналогичные структуре островков Лангерганса поджелудочной железы [Bonner-Weir S, Taneja M, Weir GC, Tatarkiewicz K, Song KH, Sharma A and O'Neil JJ (2000). In vitro cultivation of human islets from expanded ductal tissue. Proc Natl Acad Sci USA. 97, 7999-8004]. Последнее не является благоприятным, поскольку инсулин-продуцирующие клетки получали из эмбриональных клеток мозга, которые могут вызывать образование опухоли у реципиента [Amariglio N, Hirshberg A, Scheithauer BW, Cohen Y, Loewenthal R, Trakhtenbrot L, Paz N, Koren-Michowitz M, Waldman D, Leider-Trejo L, Toren A, Constantini S, Rechavi G. Donor-derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. PLoS Med. 2009 Feb 17;6(2): e1000029]. Последнее не является благоприятным, поскольку из тканей протоков можно получить только весьма ограниченное количество стволовых клеток. Более того, сообщалось о том, что когда в среде, содержащей инсулин, индуцируют дифференцировку эмбриональных стволовых клеток, инсулин, секретируемый дифференцированными клетками, фактически представляет собой инсулин, захваченный из среды, что продемонстрировано тем фактом, что не происходит секреция пропептида, который расщепляется на инсулин и C-пептид [Rajagopal J, Anderson WJ, Kume S, Martinez OI, and Melton DA (2003) Insulin staining of ES cell progeny from insulin uptake. Science. 299, 363; Hansson M, Tonning A, Frandsen U, Petri A, Rajagopal J, Englund MC, Heller RS, Hakansson J, Fleckner J, Skold HN, melton K, Semb H, and Serup P (2004) Artifactual insulin release from differentiated embryonic stem cells. Diabetes. 53, 2603-2609]. Таким образом, в последнее время было предпринято множество попыток индуцировать дифференцировку эмбриональных стволовых клеток в отсутствие инсулина. Сообщалось о дифференцировке стволовых клеток в инсулин-продуцирующие клетки в присутствие инсулиноподобных факторов роста или бетацеллюлина; эти данные объединены ниже в таблице 1. Этические проблемы и вышеуказанное образование рака затрудняют использование эмбриональных стволовых клеток при лечении сахарного диабета. Кроме того, в результате дифференцировки из плюрипотентных стволовых клеток, полученных из фибробластов кожи человека или мезенхимальных стволовых клеток, можно добиться только удивительно низких уровней инсулина и C-пептида.
Исследование дифференцировки стволовых клеток в инсулин-продуцирующие клетки ex vivo
3) Li L, Li F, Qi H, Feng G, Yuan K, Deng H, Zhou H. (2008) Coexpression of Pdx1 and betacellulin in mesenchymal stem cells could promote the differentiation of nestin-positive epithelium-like progenitors and pancreatic islet-like spheroids. Stem Cells Dev.17, 815-823.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения проводили тщательные и всесторонние исследования клеточной терапии сахарного диабета 1 типа, результатом которых стало настоящее изобретение. В этом исследовании было обнаружено, что инкубирование стволовых клеток взрослого человека, если его выполняют в среде, содержащей различные цитокины и факторы роста, о которых известно, что они эффективно способствуют дифференцировке в бета-клетки, при изменении концентрации глюкозы в среде от высокого уровня до низкого уровня, позволяет дифференцировать взрослые стволовые клетки в высокоэффективные инсулин-продуцирующие клетки.
Следовательно, цель настоящего изобретения состоит в разработке способа дифференцировки взрослых стволовых клеток в клетки, секретирующие инсулин.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Вышеупомянутые и другие цели, отличительные признаки и другие преимущества настоящего изобретения станут более понятны из следующего подробного описания в сочетании с прилагаемыми чертежами, на которых:
Фиг.1 представляет собой световые микрофотографии, на которых показана морфология клеток после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин;
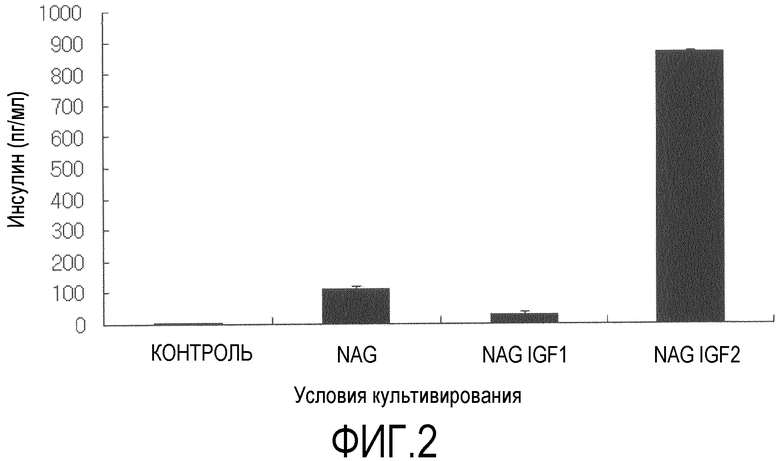
Фиг.2 представляет собой гистограмму, на которой показаны измеренные с помощью твердофазного иммуноферментного анализа количества секретированного инсулина после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин, и в течение двух часов подвергали их воздействию 25 мМ глюкозы;
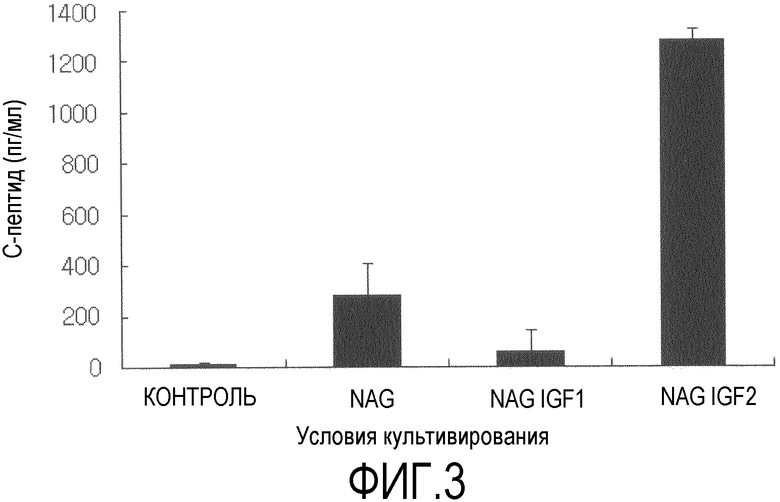
Фиг.3 представляет собой гистограмму, на которой показаны измеренные с помощью твердофазного иммуноферментного анализа количества C-пептида, секретированного после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин, и в течение двух часов подвергали их воздействию 25 мМ глюкозы;
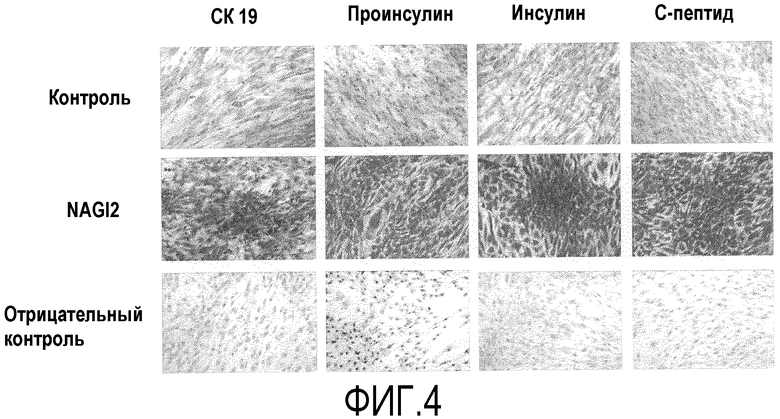
Фиг.4 представляет собой световые микрофотографии, на которых показана измеренная с помощью иммуноцитохимического анализа экспрессия инсулина, проинсулина, CK19 и C-пептида после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин;
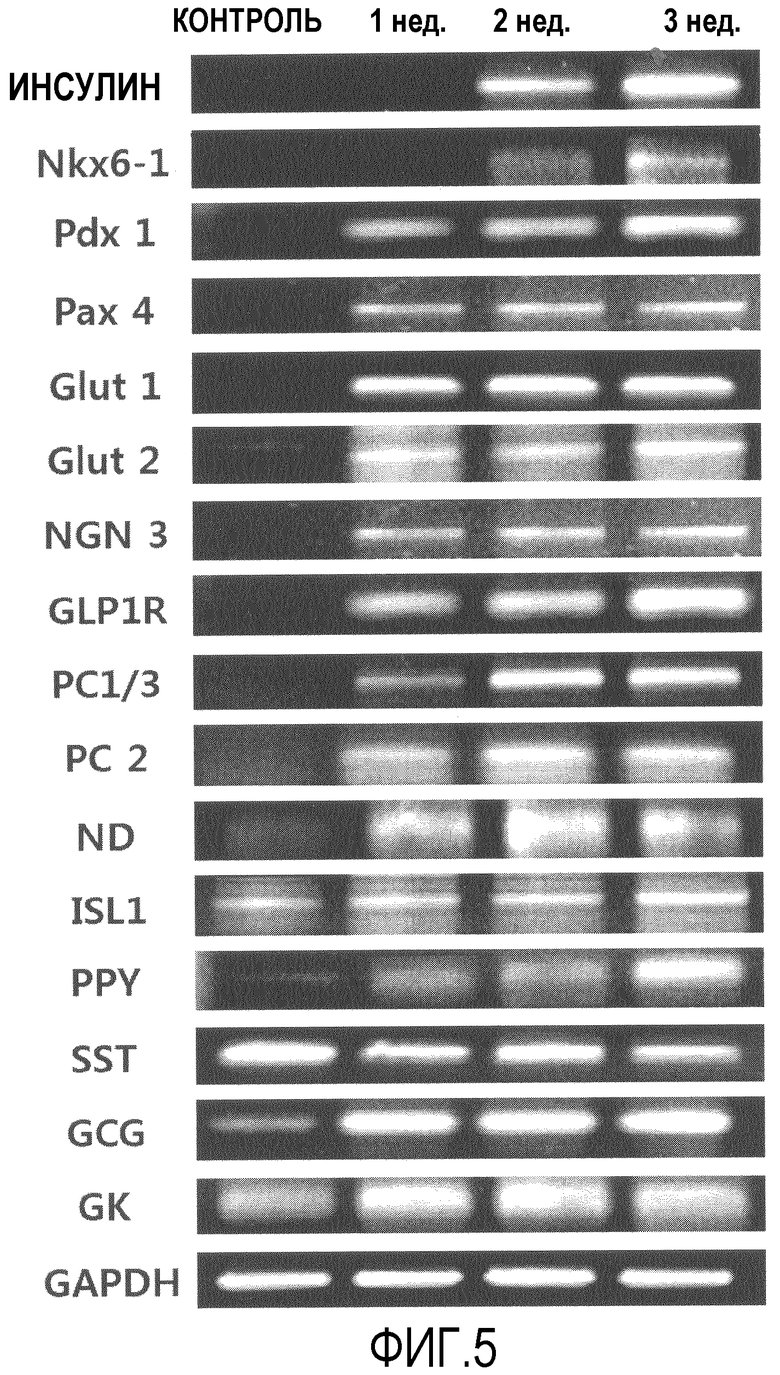
Фиг.5 представляет собой фотографию агарозного геля, на которой показана экспрессия генов, характеризующих бета-клетки, выделенных из клеток, полученных после того, как в течение трех недель стволовые клетки, полученные из подкожного слоя вокруг глаза, дифференцировались в клетки, секретирующие инсулин;
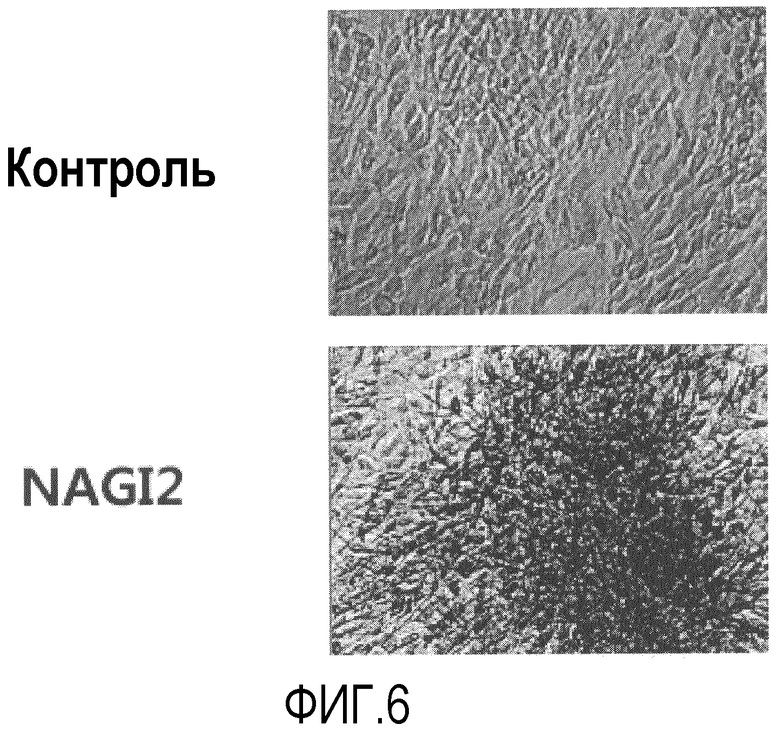
Фиг.6 представляет собой микрофотографии, на которых показано окрашивание клеток, полученных после того, как в течение трех недель клетки, полученные из подкожного слоя вокруг глаза, дифференцировались в клетки, секретирующие инсулин, с использованием дитизона, который специфичен к инсулин-продуцирующим бета-клеткам;
Фиг.7 представляет собой гистограмму, на которой показаны измеренные с помощью твердофазного иммуноферментного анализа количества секретированного инсулина после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин, и в течение 2 часов подвергали их воздействию 25 мМ глюкозы; и
Фиг.8 представляет собой гистограмму, на которой показаны измеренные с помощью твердофазного иммуноферментного анализа количества секретированного C-пептида после того, как при различных условиях в течение трех недель индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, в клетки, секретирующие инсулин, и в течение 2 часов подвергали их воздействию 25 мМ глюкозы.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
В значительной степени настоящее изобретение было выполнено с помощью двух экспериментов: в одном из них проверяли влияние инсулиноподобных факторов роста на дифференцировку, и в другом дифференцировку выполняли в присутствии бетацеллюлина вместо активина A.
В соответствии с одним из их аспектов, настоящее изобретение относится к способу дифференцировки взрослых стволовых клеток ex vivo в клетки, секретирующие инсулин, который содержит: культивирование взрослых стволовых клеток в течение 4-10 дней в среде, содержащей 20-30 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина A, 5-20 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2, и затем в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 5-20 мМ никотинамида, 1-10 нМ активина A, 5-20 нМ глюкагоноподобного пептида-1 и 10-100 нг/мл инсулиноподобного фактора роста.
Более конкретно, индуцировали дифференцировку стволовых клеток взрослого человека, выделенных из подкожных жировых тканей вокруг глаз, в клетки, секретирующие инсулин, для лечения сахарного диабета 1 типа, в среде, содержащей цитокины и факторы роста, включая фактор роста фибробластов-2, никотинамид, глюкагоноподобный пептид-1, активин A, инсулиноподобный фактор роста и т.д., при этом высокое содержание глюкозы в первой культуре сменяли на низкое содержание глюкозы во второй культуре.
Двухстадийный способ культивирования применяли к трем группам. Контроль подвергали воздействию среды, содержащей 20-30 мМ глюкозы, в присутствии никотинамида, активина A и глюкагоноподобного пептида-1 (NAG) в течение 4-10 дней, с последующим инкубированием в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, в присутствии тех же цитокинов [группа NAG], как описано в корейской патентной заявке № 2007-44663, которая была подана авторами настоящего изобретения ранее, в мае 2007. Вторую группу сначала в течение 4-10 дней подвергали воздействию среды, содержащей 20-30 мМ глюкозы, в сочетании с эмбриональной телячьей сывороткой, активином A, фактором роста фибробластов-2 и глюкагоноподобным пептидом-1, за которым следовало второе инкубирование в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, в сочетании с эмбриональной телячьей сывороткой, никотинамидом, активином A, глюкагоноподобным пептидом-1 и инсулиноподобным фактором роста-1 [группа NAGI1]. Дифференцировку третьей группы индуцировали тем же способом, который применяли ко второй группе, за исключением того, что на второй стадии вместо инсулиноподобного фактора роста-1 использовали инсулиноподобный фактор роста-2.
В соответствии с другим аспектом, настоящее изобретение относится к способу дифференцировки взрослых стволовых клеток ex vivo в клетки, секретирующие инсулин, который отличается использованием бетацеллюлина (известно, что он является эффективным средством, которое способствует дифференцировке) и добавлением B27 (известен в качестве сывороточной добавки), чтобы обеспечить условия для дифференцировки и высокоэффективной секреции инсулина (пример 7).
Более конкретно, чтобы дифференцировать взрослые стволовые клетки, инкубирование осуществляли в течение 1-7 дней в среде, содержащей 20-30 мМ глюкозы (среда с высоким содержанием глюкозы), с добавлением добавки B27, фактора роста фибробластов-2 и эпидермального фактора роста, и затем в течение 1-7 дней в среде с такой же концентрацией глюкозы, с добавлением эмбриональной телячьей сыворотки, активина A, фактора роста фибробластов-2 и глюкагоноподобного пептида-1.
После этого клетки культивировали в течение 1-7 дней в среде, содержащей 4-7 мМ глюкозы (среда с низким содержанием глюкозы), в присутствии цитокинов и факторов роста, включая эмбриональную телячью сыворотку, активин A, фактор роста фибробластов-2, глюкагоноподобный пептид-1, и, наконец, в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, с добавлением эмбриональной телячьей сыворотки, бетацеллюлина, никотинамида и глюкагоноподобного пептида-1.
В настоящем изобретении можно использовать DMEM (модифицированная по Дульбекко среда Игла) или DMEM/F12 (1:1 модифицированная по Дульбекко среда Игла: Смесь питательных веществ F-12).
Подробное описание индукции дифференцировки взрослых стволовых клеток в клетки, секретирующие инсулин, приведено ниже.
Жировую ткань, полученную хирургическим способом из подкожных слоев вокруг глаза или в щеке, обрабатывали коллагеназой I типа при 37°C в течение 30 мин и затем центрифугировали. Полученный таким образом осадок клеток культивировали в среде с добавлением эмбриональной телячьей сыворотки, чтобы получить взрослые стволовые клетки.
Чтобы дифференцировать стволовые клетки в клетки, секретирующие инсулин, их культивировали в течение 15-30 дней в среде, индуцирующей дифференцировку. Сначала их инкубировали в течение 4-10 дней в культуральных чашках, покрытых коллагеном, в среде с высоким содержанием глюкозы и с добавлением 5-20% эмбриональной телячьей сыворотки, 5-20 нМ глюкагоноподобного пептида-1, 1-10 нМ активина A и 10-30 нг/мл фактора роста фибробластов-2. После этого их переносили в среду с низким содержанием глюкозы и с добавлением 5-20% эмбриональной телячьей сыворотки, 5-20 мМ никотинамида, 5-20 нМ глюкагоноподобного пептида-1, 1-10 нМ активина A и 10-100 нг/мл инсулиноподобного фактора роста, с последующим инкубированием в течение 10-20 дней.
По завершению инкубации в течение 15-30 дней, перед тем как подвергнуть клетки воздействию среды с высоким содержанием глюкозы в течение 2 часов, в течение 10-15 часов их обрабатывали бычьим сывороточным альбумином (BSA) в среде с низким содержанием глюкозы. Наконец в полученной таким образом культуре проводили количественный анализ образования инсулина и C-пептида с использованием твердофазного иммуноферментного анализа. Было обнаружено, что условия дифференцировки, установленные в настоящем изобретении, индуцируют секрецию увеличенного количества инсулина и C-пептида по сравнению с недифференцированным состоянием (отрицательный контроль). Тогда как между группой NAGI1 (с добавлением инсулиноподобного пептида-1) и группой NAG отсутствовали различия в секреции инсулина и C-пептида, использование инсулиноподобного пептида-2 (группа NAGI2) давало возможность увеличить секрецию инсулина в восемь раз и C-пептида в шесть раз по сравнению с использованием инсулиноподобного пептида-1 (группа NAGI1). Генетический анализ клеток, дифференцированных в присутствии инсулиноподобного пептида-2, показал, что ND (neuro D1), PC (конвертаза прогормона)1/3, PC2, NGN3, Glut (транспортер глюкозы) 1, Glut 2, pax4, pdx1 - известно, что все эти гены специфичны для бета-клеток поджелудочной железы - начинали экспрессироваться, когда их дифференцировку индуцировали в среде для культивирования на первой стадии, и что ген Nkx6.1 и ген инсулина экспрессировались в процессе инкубирования в среде для культивирования со второй стадии. Также у клеток наблюдали экспрессию CK19, проинсулина, инсулина и C-пептида, которую измеряли с помощью иммунохимического окрашивания, и сильное окрашивание дитизоном, краской, которая специфична к бета-клеткам поджелудочной железы.
Таким образом, способ дифференцировки взрослых стволовых клеток в клетки, секретирующие инсулин в соответствии с настоящим изобретением, по существу состоял из двухстадийного процесса культивирования, в котором индуцировали дифференцировку взрослых стволовых клеток сначала в среде с высоким содержанием глюкозы в присутствии фактора роста фибробластов-2, активина A и глюкагоноподобного пептида-1, и затем в среде с низким содержанием глюкозы в присутствии никотинамида, активина A, глюкагоноподобного пептида-1 и инсулиноподобного фактора роста-2, который обладает способностью вызывать увеличенную секрецию инсулина по отношению к средству, которое применяли в группе NAG, как описано в корейской патентной заявке № 2007-44663, которая подана в мае 2007 года.
Альтернативно, взрослые стволовые клетки можно дифференцировать в клетки, эффективно секретирующие инсулин, в соответствии с настоящим изобретением следующим образом.
Взрослые стволовые клетки культивировали в течение 1-7 дней в среде с высоким содержанием глюкозы, которая содержала 20-30 мМ глюкозы, 0,5×-4× добавку B27, 10-30 нг/мл фактора роста фибробластов-2 и 10-30 нг/мл эпидермального фактора роста, затем в течение 1-7 дней в среде с высоким содержанием глюкозы, которая содержала 20-30 мМ глюкозы, 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина A, 5-20 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2, и затем в течение 1-7 дней в среде с низким содержанием глюкозы, которая содержала 4-7 мМ глюкозы, 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина A, 5-15 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2, и, наконец, в течение 10-20 дней в среде с низким содержанием глюкозы, которая содержала 5-20% эмбриональной телячьей сыворотки, 5-20 мМ никотинамида, 1-10 нг/мл бетацеллюлина и 5-20 нМ глюкагоноподобного пептида-1.
Было обнаружено, что после дифференцировки в присутствии бетацеллюлина, дифференцированные клетки (BTC) продуцируют как инсулин, так и C-пептид, каждый в шестикратном количестве по сравнению с группой NAG.
Следовательно, клетки, дифференцированные в присутствии добавки B27 и бетацеллюлина вместо эмбриональной телячьей сыворотки и активина A, соответственно, могут секретировать инсулин более эффективно, чем клетки из группы NAG.
Лучшее понимание настоящего изобретения может быть достигнуто с помощью следующих примеров, которые приведены для того, чтобы проиллюстрировать настоящее изобретение, и не должны рассматриваться как его ограничение.
Пример 1: Выделение стволовых клеток
Жировую ткань, с разрешения пациентов удаленную хирургическим способом из подкожного слоя вокруг глаза в клинике пластической хирургии, обрабатывали 0,075% раствором коллагеназы 1 типа при 37°C в течение 30 мин и затем таким же объемом среды с добавлением эмбриональной телячьей сыворотки, что и раствор коллагеназы, чтобы подавить активность фермента. После центрифугирования, образованный таким образом клеточный осадок дважды промывали средой для культивирования и инкубировали в DMEM-LG с добавлением 10% эмбриональной телячьей сыворотки.
Пример 2: Культура стволовых клеток
Полученные стволовые клетки культивировали в течение 21 для в среде, индуцирующей дифференцировку. Для этого клетки сначала высевали при плотности 5×103 клеток/лунка на 48-луночные планшеты, покрытые коллагеном, в которые перед инкубированием в течение 7 дней добавляли среду, содержащую 25 мМ глюкозы (DMEM-HG), с добавлением 10% эмбриональной телячьей сыворотки, 4 нМ активина A, 10 нМ глюкагоноподобного пептида-1 и 20 нг/мл фактора роста фибробластов-2. После этого клетки дополнительно культивировали в течение еще 14 дней в среде, содержащей 5,5 мМ глюкозы (DMEM-LG), с добавлением 10% эмбриональной телячьей сыворотки, 10 мМ никотинамида, 4 нМ активина A, 10 нМ глюкагоноподобного пептида-1 и 50 нг/мл инсулиноподобного фактора роста-1 или инсулиноподобного фактора роста-2.
В ходе инкубации старую среду регулярно заменяли на свежую с интервалом в 3-4 дня. После инкубации в целом в течение 3 недель, дифференцировку клеток исследовали следующим образом.
Пример 3: Морфологические изменения клеток после дифференцировки
Стволовые клетки выделяли из подкожного жира вокруг глаза, культивировали и наблюдали их морфологические изменения. Во время инкубирования в среде, не содержащей факторы роста и цитокины, морфология клеток не менялась, тогда как под воздействием факторов роста и/или цитокинов, таких как никотинамид, активин A и глюкагоноподобный пептид-1 или инсулиноподобный фактор роста-1 или -2, наблюдали, что стволовые клетки подвергались морфологическим изменениям и обретали округлую форму (фиг.1). На 3 неделе после индукции дифференцировки, клетки приобретали округлую форму и образовывали кластеры клеток.
Пример 4: Исследование секреции инсулина и C-пептида
После завершения культивирования в течение 3 недель, клетки в течение 12 часов подвергали воздействию среды с низким содержанием глюкозы (DMEM-LG) с добавлением 0,5% бычьего сывороточного альбумина. Затем клетки в течение 2 часов подвергали воздействию DMEM-HG, и проводили количественный анализ секретированного инсулина с использованием твердофазного иммуноферментного анализа (фиг.2). Также количество C-пептида, соединенного с инсулином в проинсулине, также измеряли с использованием твердофазного иммуноферментного анализа (фиг.3). Для твердофазного иммуноферментного анализа инсулина и C-пептида использовали коммерчески доступные наборы из Mercodia, Sweden. Стандартные растворы с заранее известными концентрациями и ранее полученные культуры в количестве 25 мкл на лунку высевали на 96-луночные микропланшеты, покрытые инсулином или C-пептидом человека, после чего в планшеты добавляли антитела к инсулину или C-пептиду и инкубировали в течение 1 часа. Затем проводили инкубирование в течение 30 мин в растворе субстрата TMB и измеряли оптическую плотность. Ни инсулин, ни C-пептид не секретировался клетками, у которых не индуцировали дифференцировку, тогда как было обнаружено, что группы, которые культивировали с использованием двухстадийного способа по настоящему изобретению, секретировали инсулин и C-пептид. В особенности, при измерении инсулина и C-пептида, группа, которую культивировали в присутствии инсулиноподобного пептида-2 (группа NAGI2), показала восьмикратный уровень секреции инсулина и шестикратный уровень секреции C-пептида по сравнению с соответствующими уровнями у группы NAG.
Пример 5: Экспрессия белков, специфичных для клеток, секретирующих инсулин
После того как стволовые клетки, полученные из подкожного жира вокруг глаза, индуцировали к дифференцировке в присутствии инсулиноподобного пептида-2, для которого с помощью твердофазного иммуноферментного анализа была доказана наибольшая эффективность, у полученных клеток, секретирующих инсулин, анализировали экспрессию CK19, инсулина, C-пептида и проинсулина (который не расщеплялся перед внеклеточной секрецией) с использованием иммуноцитохимического анализа (фиг.4). После дифференцировки, продолжавшейся в течение 3 недель, в течение 2 часов клетки подвергали воздействию высокой концентрации глюкозы, фиксировали при 4°C в течение 90 мин в 4% параформальдегиде и промывали три раза в фосфатно-солевом буфере. Для удаления эндогенной пероксидазы клетки обрабатывали в течение 10 мин в 3% пероксиде водорода, а затем инкубировали при комнатной температуре в течение 60 мин в PBS с добавлением 2% бычьего сывороточного альбумина, чтобы блокировать фоновые сигналы. При 4°C в течение 12 часов проводили реакции клеточных культур с разведенными в PBS первичными человеческими антителами против CK19 (1:400), C-пептида (1:500), инсулина (1:500) и проинсулина (1:500). В качестве отрицательного контроля использовали PBS, который не содержал первичные антитела. В течение 20 мин их обрабатывали каждым вторичным антителом, конъюгированным с биотином, и стрептавидином, конъюгированным с перекисью водорода, а затем окрашивали 3,3'-диаминобензидином (DAB). По сравнению с контролем, в котором не индуцировали дифференцировку, у групп, которые обрабатывали сочетанием никотинамида, активина A и глюкагоноподобного пептида, наблюдали значительно увеличенные уровни экспрессии CK19, C-пептида, инсулина и проинсулина (фиг.4).
Пример 6: Экспрессия генов, участвующих в секреции инсулина
После того как индуцировали дифференцировку стволовых клеток, полученных из подкожного слоя вокруг глаза, клетки брали на 7, 14 и 21 день и с помощью полимеразной цепной реакции с обратной транскриптазой (RT-ПЦР) исследовали экспрессию генов, характерных для клеток, секретирующих инсулин.
Сначала из дифференцированных клеток выделяли общую РНК. Один мл Tri Reagent добавляли в пробирку с клетками и тщательно перемешивали с 200 мкл хлороформа, а затем инкубировали при комнатной температуре в течение 15 мин. После центрифугирования при 14000 об/мин в течение 15 мин, полученный таким образом супернатант переносили в новую пробирку и смешивали с изопропанолом. После центрифугирования при 14000 об/мин в течение 10 мин образовывался осадок, к которому затем добавляли 75% этанол. Эту суспензию снова центрифугировали при 14000 об/мин в течение 10 мин. Супернатант удаляли, а оставшийся 75% этанол выпаривали, после чего остаток растворяли в стерилизованной, деионизированной воде и инкубировали при 65°C в течение 5 мин, чтобы элюировать РНК. Проводили количественный анализ этой РНК посредством определения оптической плотности при 260 нм в УФ спектрофотометре. В целом 7,5 мкг РНК смешивали с 1× реакционным буфером, 1 мМ dNTP, 0,5 мг/мл олиго-dT, 20 ЕД ингибитора РНКазы и 20 ЕД обратной транскриптазы M-MuLV и добавляли стерильную деионизированную воду до конечного объема 50 мкл перед инкубированием при 42°C в течение 60 мин.
ПЦР осуществляли в перемешанном растворе, содержащем 1× буфер Taq, 2 мМ MgCl2, 0,25 ЕД Taq-полимеразы и 10 пмоль праймеров, каждая кДНК последовательность которых комплементарна каждому гену, приведенному в таблице 2.
ПЦР начиналась с предварительной денатурации при 94°С в течение 5 мин, за которой следовало 35 циклов денатурации при 94° С в течение 30 сек, отжига при соответствующей температуре в течение 30 сек и элонгации при 72° С в течение 30 сек, и финальное удлинение фрагментов при 72°C в течение 5 мин. По 15 мкл каждого полученного таким образом продукта ПЦР загружали вместе с 6× буфером для нанесения (0,25% бромфенол синий, 0,25% ксилолцианол, 40% сахароза) на 2% агарозный гель и проводили электрофорез при электрическом напряжении 100 В в течение 30 мин. После завершения электрофореза гели окрашивали бромистым этидием и исследовали в УФ трансиллюминаторе (Vilber loumat, FRANCE). Уровень экспрессии каждого гена определяли и сравнивали с уровнем глицеральдегид-3-фосфатдегидрогеназы (GAPDH) в качестве контроля. Данные для сравнительного анализа получали на основе трех экспериментов.
Последовательности праймеров, использованных в RT-ПЦР для амплификации соответствующих генов, которые характерны для клеток, секретирующих инсулин, их названия и размеры продуктов ПЦР перечислены ниже в таблице 2.
(SEQ ID NO: 1)
5'-aaattcgttgtcataccagg-3'
(SEQ ID NO: 2)
(SEQ ID NO: 3)
5'-aagggctttattccatctctctcg-3'
(SEQ ID NO: 4)
(SEQ ID NO: 5)
5'-tggcactcctgggacgagtttc-3'
(SEQ ID NO: 6)
(SEQ ID NO: 7)
5'-gtcctcctcctttttccac-3'
(SEQ ID NO: 8)
(SEQ ID NO: 9)
5'-ggtttcttcctgagccagcg-3'
(SEQ ID NO: 10)
(SEQ ID NO: 11)
5'-tgtttggcaaggcaatgacc-3'
(SEQ ID NO: 12)
(SEQ ID NO: 13)
5'-tgctggacttgtgcttcttcaac-3'
(SEQ ID NO: 14)
(SEQ ID NO: 15)
5'-aataagaatgcccgtgacga-3'
(SEQ ID NO: 16)
(SEQ ID NO: 17)
5'-atcaaaggaagggctggtg-3'
(SEQ ID NO: 18)
(SEQ ID NO: 19)
5'-gtaacgaaccgaccactt-3'
(SEQ ID NO: 20)
(SEQ ID NO: 21)
5'-ttgcgttctcggggtgccatagc-3'
(SEQ ID NO: 22)
(SEQ ID NO: 23)
5'-agtcgtaggagacagaaggt-3'
(SEQ ID NO: 24)
(SEQ ID NO: 25)
5'-acttctttggtgattgctttggcggtg-3'
(SEQ ID NO: 26)
(SEQ ID NO: 27)
5'-gagacacaaccacccttcatccttc-3'
(SEQ ID NO: 28)
(SEQ ID NO: 29)
5'-ctgggtaacagggatcaaacag-3'
(SEQ ID NO: 30)
(SEQ ID NO: 31)
5'-cttggcaagtctgcatttga-3'
(SEQ ID NO: 32)
(SEQ ID NO: 33)
5'-ctgcctcattccaagccatacagtagtg-3'
(SEQ ID NO: 34)
Как следует из данных на фиг.5, в клетках, в которых не индуцировали дифференцировку, экспрессировались neuro D и islet 1, о которых известно, что они играют важные роли в развитии поджелудочной железы, и глюкагон, SST (соматостатин) и PP (панкреатический полипептид), секретируемые клетками, отличающимися от бета-клеток эндокринных клеток поджелудочной железы, но когда индуцировали дифференцировку, клетки начинали экспрессировать PC2, PC1/3, Glp1R, нейрогенин 3, Glut2, Glut1, pax4 и pdx1 на 7 день и Nkx6.1 и инсулин на 14 день, а их уровни экспрессии значительно увеличивались на 21 день.
Пример 7: Окрашивание дитизоном
Известно, что инсулин образует комплекс с ионом цинка (Zn++) с образованием гексамеров. Известно, что дитизон представляет собой цинк-хелатирующее средство, которое избирательно окрашивает бета-клетки поджелудочной железы в связи с высоким содержанием цинка в этих клетках. Для этого клетки, которые дифференцировались в присутствии инсулиноподобного пептида-2, окрашивали дитизоном. Для этого 50 мг порошка дитизона растворяли в 5 мл диметилсульфоксида (DMSO) и 10 мкл раствора дитизола разводили в 1 мл среды для культивирования. После фильтрации через 0,2 мкм нейлоновый фильтр, окрашивающий раствор дитизона капали на культуральную чашку и инкубировали ее при 37°С в течение 15 мин. Клетки промывали три раза в PBS и осматривали под оптическим микроскопом. Как видно на фиг.6, дифференцированные клетки четко окрашивались, при этом кластеры клеток имели насыщенную окраску.
Вкратце, как описано выше, когда стволовые клетки, полученные из подкожного жира вокруг глаза, сначала подвергали воздействию глюкозы в высокой концентрации в сочетании с активином A, фактором роста фибробластов-2 и глюкагоноподобным пептидом-1, а затем подвергали воздействию глюкозы в низкой концентрации в присутствии никотинамида, активина A, глюкагоноподобного пептида-1 и инсулиноподобного фактора роста-2, они дифференцировались в секретирующие инсулин клетки, у которых наблюдали образование инсулина, C-пептида и проинсулина в больших количествах. Кроме того, они экспрессировали гены, характерные для бета-клеток поджелудочной железы, а также четко окрашивались дитизоном. Таким образом, установленный способ по настоящему изобретению дает возможность эффективной дифференцировки взрослых стволовых клеток в инсулин-секретирующие бета-клетки.
Пример 8: Культура стволовых клеток
Полученные стволовые клетки культивировали в течение 21 для в среде, индуцирующей дифференцировку. Для этого клетки сначала высевали при плотности 5×103 клеток/лунка на 48-луночные планшеты с коллагеновым покрытием, в которые перед инкубацией в течение 3 дней добавляли среду, содержащую 25 мМ глюкозы (DMEM-HG), с добавлением 1× добавки B27, 20 нг/мл фактора роста фибробластов-2 и 20 нг/мл эпидермального фактора роста. После этого клетки дополнительно инкубировали еще в течение 4 дней в среде, содержащей 25 мМ глюкозы, с добавлением 10% эмбриональной телячьей сыворотки, 4 нМ активина A, 20 нг/мл фактора роста фибробластов-2 и 10 нМ глюкагоноподобного пептида-1. Впоследствии клетки культивировали в течение 3 дней в среде, содержащей 5,5 мМ глюкозы (DMEM-LG), с добавлением 10% эмбриональной телячьей сыворотки, 4 нМ активина A, 20 нг/мл фактора роста фибробластов-2 и 10 нМ глюкагоноподобного пептида-1, и затем, перед инкубацией в течение дополнительных 10 дней, переносили в новую среду, содержащую 5,5 мМ глюкозы (DMEM-LG), с добавлением 10 мМ никотинамида, 10 нМ глюкагоноподобного пептида и 10 нг/мл бетацеллюлина.
В процессе инкубации старую среду периодически заменяли на свежую с интервалами в 3-4 дня. После инкубации, в целом, в течение 3 недель, дифференцировку клеток исследовали следующим образом.
Пример 9: Исследование секреции инсулина и C-пептида
После завершения 3-недельного культивирования, клетки в течение 12 часов подвергали воздействию среды с низким содержанием глюкозы (DMEM-LG), с добавлением 0,5% бычьего сывороточного альбумина. Затем клетки в течение 2 часов подвергали воздействию DMEM-HG и проводили количественный анализ секретированного инсулина с использованием твердофазного иммуноферментного анализа (фиг.7). Также с использованием иммуноцитохимического анализа измеряли количество C-пептида, соединенного с инсулином в проинсулине (фиг.8). Для твердофазного иммуноферментного анализа инсулина и C-пептида использовали коммерчески доступные наборы из Mercodia, Sweden. Стандартные растворы с уже известными концентрациями и полученные ранее культуры высевали в количестве 25 мкл на лунку в 96-луночные планшеты, покрытые инсулином или C-пептидом человека, после чего в лунки добавляли антитела к инсулину или C-пептиду и инкубировали в течение 1 часа. После этого в лунки добавляли по 200 мкл раствора субстрата TMB. После инкубации в течение 30 мин, измеряли их оптические плотности. Клетки, у которых не индуцировали дифференцировку, не секретировали ни инсулин, ни C-пептид, и при этом было обнаружено, что группы, которые культивировали с использованием двухстадийного способа по настоящему изобретению, образуют инсулин и C-пептид. В особенности, измерения показали, что группа, которую культивировали тем же способом, что в примере 8, секретирует приблизительно в 6 раз больше инсулина и приблизительно в 6 раз больше C-пептида, чем группа NAG.
Ожидалось, что синтезируя инсулин и C-пептид в количествах, в 7-8 раз превышающих количества, синтезируемые группой NAG, согласно раскрытию корейской патентной заявки № 2007-44663, поданной в мае 2007 года, инсулин-секретирующие клетки, дифференцированные из взрослых стволовых клеток в средах для культивирования по настоящему изобретению, как описано ранее, являются превосходным средством для лечения сахарного диабета 1 типа.
Хотя предпочтительные варианты осуществления настоящего изобретения раскрыты с иллюстративными целями, специалисты в данной области признают, что возможны различные модификации, дополнения и замены без отклонения от объема и сущности изобретения, раскрытых в пунктах приложенной формулы изобретения.









Изобретение относится к области биотехнологии, конкретно к дифференцировке стволовых клеток взрослого человека в клетки, секретирующие инсулин, и может быть использовано в медицине для лечения сахарного диабета 1 типа. Стволовые клетки взрослого человека, выделенные из подкожных жировых тканей вокруг глаз, индуцируют к дифференцировке в клетки, секретирующие инсулин, в среде, в которой присутствуют цитокины и факторы роста, включая добавку В27, фактор роста фибробластов-2, эпидермальный фактор роста, никотинамид, глюкагоноподобный пептид-1, активин А, инсулиноподобный фактор роста и бетацеллюлин. При культивировании глюкозу в высокой концентрации заменяют глюкозой в низкой концентрации. Изобретение позволяет получить клетки, синтезирующие большие количества инсулина и С-пептида. 2 н. и 1 з.п. ф-лы, 8 ил., 2 табл.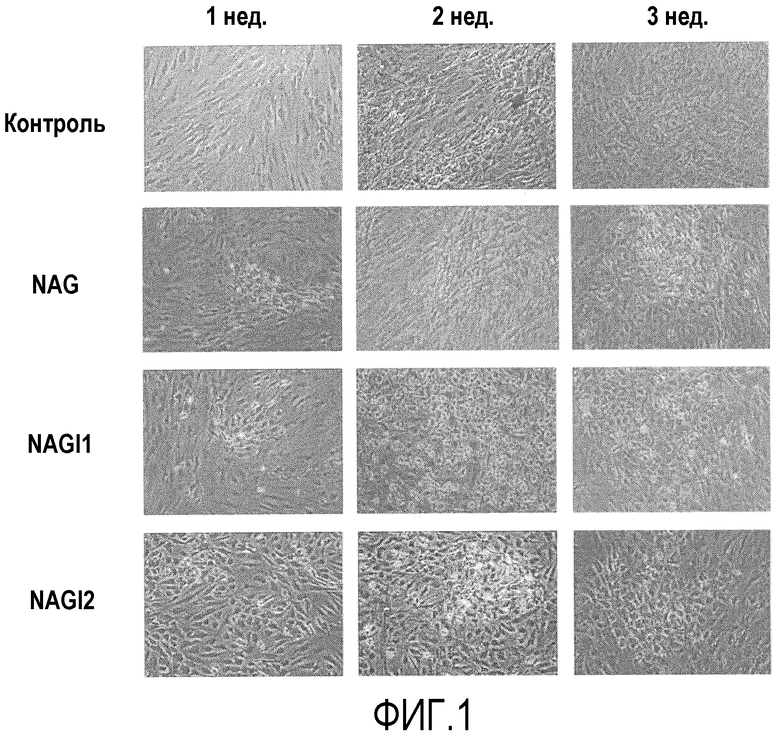
1. Способ дифференцировки стволовых клеток взрослого человека ex vivo в клетки, секретирующие инсулин, который содержит:
культивирование взрослых стволовых клеток в течение 4-10 дней в среде, содержащей 20-30 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина А, 5-20 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2; и
культивирование культивированных стволовых клеток в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 5-20 мМ никотинамида, 1-10 нМ активина А, 5-20 нМ глюкагоноподобного пептида-1 и 10-100 нг/мл инсулиноподобного фактора роста.
2. Способ по п.1, где инсулиноподобный фактор роста представляет собой инсулиноподобный фактор роста-2.
3. Способ дифференцировки стволовых клеток взрослого человека ex vivo в клетки, секретирующие инсулин, который содержит последовательное культивирование взрослых стволовых клеток:
в течение 1-7 дней в среде, содержащей 20-30 мМ глюкозы, с добавлением добавки В27, 10-30 нг/мл фактора роста фибробластов-2 и 10-30 нг/мл эпидермального фактора роста;
в течение 1-7 дней в среде, содержащей 20-30 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина А, 5-20 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2;
в течение 1-7 дней в среде, содержащей 4-7 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 1-10 нМ активина А, 5-20 нМ глюкагоноподобного пептида-1 и 10-30 нг/мл фактора роста фибробластов-2; и
в течение 10-20 дней в среде, содержащей 4-7 мМ глюкозы, с добавлением 5-20% эмбриональной телячьей сыворотки, 5-20 мМ никотинамида, 1-10 нг/мл бетацеллюлина и 5-15 нМ глюкагоноподобного пептида.
| KR 2008099052 А, 12.11.2008 | |||
| ДИФФЕРЕНЦИРОВКА СТРОМАЛЬНЫХ КЛЕТОК, ПОЛУЧЕННЫХ ИЗ ЖИРОВОЙ ТКАНИ, В ЭНДОКРИННЫЕ КЛЕТКИ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2002 |
|
RU2351648C2 |
| SEGEV Н | |||
| et al | |||
| Differentiation of human embryonic stem cells into insulin-producing clusters, Stem Cells, 2004, v.22, n.3, p.265-274 | |||
| SEABERG R.M | |||
| et al | |||
| Clonal identification of multipotent precursors from adult mouse pancreas that generate neural and pancreatic lineages, Nat | |||
| Biotechnol., | |||

