ОБЛАСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении предлагаются способы, способствующие осуществлению дифференцировки полипотентных стволовых клеток, и продукты, относящиеся к этим способам или возникающие в результате их использования. В частности, в настоящем изобретении предлагается усовершенствованный способ создания клеток, экспрессирующих гормоны поджелудочной железы, и клеток, секретирующих гормоны поджелудочной железы. Кроме того, в настоящем изобретении также предлагаются способы стимулирования дифференцировки полипотентных стволовых клеток без использования слоя питающих клеток, а также продукты, относящиеся к этим способам или возникающие в результате их использования. В настоящем изобретении также предлагаются способы, способствующие осуществлению стимулируемой глюкозой секреции инсулина в инсулинпродуцирующих клетках, полученных из полипотентных стволовых клеток.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Достижения в области клеточной заместительной терапии сахарного диабета 1 типа и нехватка трансплантируемых островков Лангерганса заставили обратить внимание на разработку источников инсулинпродуцирующих клеток, или β-клеток, подходящих для трансплантации. Одним из подходов является формирование функциональных β-клеток из полипотентных стволовых клеток, таких как, например, эмбриональные стволовые клетки.
При эмбриональном развитии позвоночных, полипотентные стволовые клетки дают начало группе клеток, формирующих три зародышевых листка (эктодерму, мезодерму и эндодерму) в ходе процесса, именуемого гаструляцией. Ткани, из которых состоят, например, щитовидная железа, тимус, поджелудочная железа, кишечник и печень, будут развиваться из эндодермы, через промежуточную стадию. Промежуточной стадией данного процесса является образование сформировавшейся эндодермы. Клетки сформировавшейся эндодермы экспрессируют ряд маркеров, таких как HNF-3beta, GATA4, Mixl1, CXCR4 и Sox-17.
Полипотентные стволовые клетки могут экспрессировать один или несколько стадийно-специфичных эмбриональных антигенов (stage-specific embryonic antigen, SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145, 1998). Дифференцировка полипотентных стволовых клеток in vitro приводит к утрате экспрессирования SSEA-4, Tra-1-60 и Tra-1-81 (если имеются) и к увеличению экспрессии SSEA-1. В недифференцированных полипотентных стволовых клетках, как правило, активна щелочная фосфатаза, которая может быть обнаружена путем фиксации клеток с помощью 4% параформальдегида, с последующим обнаружением с помощью Vector Red, применяемого в качестве субстрата, в соответствии с инструкциями производителя (Vector Laboratories, Burlingame Calif.). Недифференцированные полипотентные стволовые клетки также, как правило, экспрессируют Oct-4 и TERT, обнаруживаемые методом ОТ-ПЦР.
Полипотентные стволовые клетки, как правило, культивируются на слое питающих клеток, которые оказывают разностороннюю поддержку полипотентным клеткам. Как вариант, полипотентные стволовые клетки культивируются в культуральной системе, по существу не содержащей питающих клеток, но, тем не менее, поддерживающей пролиферацию полипотентных стволовых клеток и не допускающей существенной дифференцировки. Выращивание полипотентных стволовых клеток в культуре, не содержащей питающих клеток, без дифференцировки, осуществляется при помощи среды, кондиционированной путем предварительного культивирования в ней клеток другого типа. Как вариант, выращивание полипотентных стволовых клеток в культуре, не содержащей питающих клеток, без дифференцировки, осуществляется при помощи среды с определенным химическим составом.
Например, в публикациях Reubinoff et al. (Nature Biotechnology 18: 399-404 (2000)) и Thompson et al. (Science, 6 ноября 1998 года: Vol. 282, № 5391, pp. 1145-1147) описано культивирование линий полипотентных стволовых клеток из человеческих бластоцист с применением слоя питающих клеток из мышиных эмбриональных фибробластов.
В публикации Richards et al. (Stem Cells 21: 546-556, 2003) анализировался набор из 11 различных слоев питающих клеток, полученных от взрослых, новорожденных и эмбрионов людей, по их способности осуществлять поддержку культуры человеческих полипотентных стволовых клеток. Richards et al. сообщают: «линии человеческих эмбриональных стволовых клеток, культивируемые на питающих слоях из фибробластов кожи взрослых людей, сохраняют морфологию, характерную для эмбриональных стволовых клеток, и остаются полипотентными».
В заявке на патент US20020072117 описываются линии клеток, продуцирующие среду, осуществляющую поддержку полипотентных стволовых клеток приматов в культуре, не содержащей питающих клеток. Использованные клеточные линии представляют собой мезенхимо- и фибробластоподобные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В заявке на патент US20020072117 также описывается использование этих клеточных линий в качестве первичного слоя питающих клеток.
В другом примере, Wang et al. (Stem Cells 23: 1221-1227, 2005) описывают способы длительного выращивания человеческих полипотентных стволовых клеток на слоях питающих клеток, полученных из человеческих эмбриональных стволовых клеток.
В другом примере, Stojkovic et al. (Stem Cells 23: 306-314, 2005) описывают систему питающих клеток, получаемую в результате спонтанной дифференцировки человеческих эмбриональных стволовых клеток.
В еще одном примере, Miyamoto et al. (Stem Cells 22: 433-440, 2004) описывают источник питающих клеток, получаемых из человеческой плаценты.
Amit et al. (Biol. Reprod. 68: 2150-2156, 2003) описывают слой питающих клеток, полученных из человеческой крайней плоти.
В другом примере, Inzunza et al. (Stem Cells 23: 544-549, 2005) описывают слой питающих клеток, полученных из человеческих постнатальных фибробластов крайней плоти.
В патенте US6642048 описывается среда, поддерживающая рост полипотентных стволовых клеток приматов (пПС) в среде, не содержащей питающих клеток, и клеточные линии, которые могут использоваться для производства такой среды. В патенте US6642048 говорится: «Данное изобретение включает мезенхимо- и фибробластоподобные клеточные линии, полученные из эмбриональной ткани или дифференцированные из эмбриональных стволовых клеток. В документе описываются и иллюстрируются способ получения таких клеточных линий, обработки среды и выращивания стволовых клеток с применением кондиционированной среды».
В другом примере, в заявке на патент WO2005014799, описывается кондиционированная среда для поддержания, пролиферации и дифференцировки клеток млекопитающих. В заявке на патент WO2005014799 говорится: «Культуральная среда, произведенная в соответствии с настоящим изобретением, кондиционируется при помощи секреторной активности клеток мыши, в частности, активности дифференцированных и иммортализованных трансгенных гепатоцитов, именуемых MMH (Met Murine Hepatocyte)».
В другом примере, Xu et al. (Stem Cells 22: 972-980, 2004) описывают кондиционированную среду, полученную из производных человеческих эмбриональных стволовых клеток, генетически модифицированных для увеличения экспрессии обратной транскриптазы человеческой теломеразы.
В другом примере, заявке на патент US20070010011, описывается культуральная среда определенного химического состава для поддержания полипотентных стволовых клеток.
В альтернативной культуральной системе используется не содержащая сыворотки среда, обогащенная факторами роста, способными стимулировать пролиферацию эмбриональных стволовых клеток. Например, Cheon et al. (BioReprod DOI:10.1095/biolreprod.105.046870, 19 октября 2005 года) описывают не содержащую питающих клеток и сыворотки культуральную систему, в которой эмбриональные стволовые клетки поддерживаются в некондиционированной, заменяющей сыворотку среде (SR), обогащенной различными факторами роста, способными запустить самообновление эмбриональных стволовых клеток.
В другом примере, Levenstein et al. (Stem Cells 24: 568-574, 2006) описывают способы длительного культивирования человеческих эмбриональных стволовых клеток в отсутствие фибробластов или кондиционированной среды, с применением среды, обогащенной основным фактором роста фибробластов (bFGF).
В другом примере, заявке на патент US20050148070, описывается способ культивирования человеческих эмбриональных стволовых клеток в среде с определенным составом без сыворотки и без питающих клеток-фибробластов, где данный способ включает: культивирование стволовых клеток в культуральной среде, содержащей альбумин, аминокислоты, витамины, минеральные вещества, по меньшей мере один трансферрин или заменитель трансферрина, по меньшей мере один инсулин или заместитель инсулина, культуральную среду, в основном не включающую эмбриональную сыворотку млекопитающих и содержащую по меньшей мере приблизительно 100 нг/мл фактора роста фибробластов, способного активировать сигнальный рецептор фактора роста фибробластов, причем фактор роста фибробластов происходит из источника, иного, чем просто слой питающих клеток-фибробластов, среду, поддерживающую пролиферацию стволовых клеток в недифференцированном состоянии без слоя питающих клеток или кондиционированной среды.
В другом примере, заявке на патент US20050233446, описывается среда с определенным составом, которая может использоваться при культивировании стволовых клеток, включая недифференцированные зародышевые стволовые клетки приматов. В растворе, среда, по существу, является изотонической относительно культивируемых стволовых клеток. В данной культуре указанная среда содержит основную среду и количество bFGF, инсулина и аскорбиновой кислоты, достаточное для поддержки роста зародышевых стволовых клеток без существенной дифференцировки.
В другом примере, патенте US6800480, говорится: «В одном варианте осуществления предлагается культуральная среда для выращивания клеток зародышевых стволовых клеток приматов, в значительной степени в недифференцированном состоянии, включающая основную среду с низким содержанием эндотоксина и низким осмотическим давлением, которая эффективно поддерживает рост зародышевых стволовых клеток приматов. Основная среда объединяется с питательной сывороткой, способной поддерживать рост зародышевых стволовых клеток приматов, и субстратом, выбранным из группы, состоящей из питающей клетки и экстраклеточного матрикса, полученного из питающих клеток. Среда также содержит аминокислоты, не относящиеся к незаменимым, антиоксидант и первый фактор роста, выбираемый из группы, состоящей из нуклеозидов и соль-пирувата».
В другом примере, заявке на патент US20050244962, говорится: «В одном аспекте, в изобретении предлагается способ культивирования эмбриональных стволовых клеток приматов. Стволовые клетки культивируются в культуре, в основном, не содержащей эмбриональной сыворотки млекопитающих (предпочтительно, также, в основном, не содержащей эмбриональной сыворотки любых животных) и в присутствии фактора роста фибробластов, полученного из источника, иного, чем просто питающие клетки-фибробласты. В предпочтительной форме, слой питающих фибробластов, ранее необходимый для поддержания культуры стволовых клеток, становится необязательным вследствие добавления достаточного количества фактора роста фибробластов».
В другом примере, заявке на патент WO2005065354, описывается изотоническая культуральная среда определенного состава, в основном не содержащая питающих клеток и сыворотки, включающая: a) базовую среду; b) количество bFGF, достаточное для поддержания роста, в основном недифференцированных стволовых клеток млекопитающих; c) количество инсулина, достаточное для поддержания роста, в основном недифференцированных стволовых клеток млекопитающих; и d) количество аскорбиновой кислоты, достаточное для поддержки роста, в основном недифференцированных стволовых клеток млекопитающих.
В другом примере, WO2005086845, описывается способ поддержания недифференцированных стволовых клеток, где упомянутый способ включает воздействие на стволовые клетки одним членом семейства белков трансформирующего ростового фактора-бета (TGFβ), одним членом семейства белков фактора роста фибробластов (FGF) или никотинамидом (NIC) в количестве, достаточном для поддержания клеток в недифференцированном состоянии в течение периода времени, достаточного для получения желаемого результата.
Полипотентные стволовые клетки могут культивироваться и дифференцироваться на субстрате тканевой культуры, покрытом экстраклеточным матриксом. Экстраклеточный матрикс может быть растворен перед покрытием им субстрата тканевой культуры. Примеры подходящих способов растворения экстраклеточного матрикса и покрытия субстрата тканевой культуры можно найти в публикации Kleinman, H.K., et al., Biochemistry 25: 312 (1986), и Hadley, M.A., et al., J. Cell. Biol. 101: 1511 (1985).
Формирование поджелудочной железы происходит при дифференцировке сформировавшейся эндодермы в панкреатическую эндодерму. Клетки панкреатической эндодермы экспрессируют ген панкреатическо-дуоденального гомеобокса, Pdx1. При отсутствии Pdx1 развитие поджелудочной железы не идет дальше формирования вентрального и дорзального зачатков. Следовательно, экспрессия Pdx1 является важным этапом онтогенеза поджелудочной железы. Зрелая поджелудочная железа содержит, среди других типов клеток, экзокринную ткань и эндокринную ткань. Экзокринная и эндокринная ткани образуются при дифференцировке панкреатической эндодермы.
Клетки, обладающие свойствами островковых клеток, как сообщалось, были получены из эмбриональных клеток мыши. Например, в публикации Lumelsky et al. (Science 292: 1389, 2001) сообщается о дифференцировке мышиных эмбриональных стволовых клеток в инсулинсекретирующие структуры, сходные с островками поджелудочной железы. Soria et al. (Diabetes 49: 157, 2000) сообщают, что инсулинсекретирующие клетки, полученные из мышиных эмбриональных стволовых клеток, нормализовали гликемию у мышей с диабетом, вызванным стрептозотоцином.
В одном примере, в публикации Hori et al. (PNAS 99: 16105, 2002) описывается, что обработка мышиных эмбриональных стволовых клеток ингибиторами фосфоинозитид 3-киназы (LY294002) приводила к получению клеток, сходных с β-клетками.
В другом примере, Blyszczuk et al. (PNAS 100: 998, 2003) сообщается о получении инсулинпродуцирующих клеток из мышиных эмбриональных стволовых клеток с конститутивной экспрессией Pax4.
Micallef et al. сообщают, что ретиноевая кислота может регулировать способность эмбриональных стволовых клеток формировать Pdx1-положительную панкреатическую эндодерму. Ретиноевая кислота с наибольшей эффективностью индуцирует экспрессию Pdx1 при добавлении в культуру на 4 день дифференцировки эмбриональных стволовых клеток в течение периода, соответствующего концу гаструляции эмбриона (Diabetes 54: 301, 2005).
В публикации Miyazaki et al. сообщается о линии мышиных эмбриональных стволовых клеток со сверхэкспрессией Pdx1. Эти результаты показывают, что экспрессия экзогенного Pdx1 очевидно повышает экспрессию генов инсулина, соматостатина, глюкокиназы, нейрогенина 3, P48, Pax6 и HNF6 в образующихся дифференцированных клетках (Diabetes 53: 1030, 2004).
В публикации Skoudy et al. сообщается, что активин A (входящий в суперсемейство TGF-β) повышает экспрессию экзокринных панкреатических генов (p48 и амилаза) и эндокринных генов (Pdx1, инсулин и глюкагон) в эмбриональных стволовых клетках мыши. Максимальный эффект наблюдался при использовании активина A в концентрации 1 нМ. Также эти авторы наблюдали, что на уровень экспрессии мРНК инсулина и Pdx1 не влияла ретиноевая кислота; однако обработка раствором FGF7 с концентрацией 3 нМ приводила к повышению уровня транскрипта Pdx1 (Biochem. J. 379: 749, 2004).
В работе Shiraki et al. изучались эффекты факторов роста, специфически увеличивающих дифференцировку эмбриональных стволовых клеток в Pdx1-положительные клетки. Эти авторы наблюдали, что TGF-β2 приводил к воспроизводимому увеличению доли Pdx1-положительных клеток (Genes Cells, июнь 2005 г.; 10(6): 503-16).
В публикации Gordon et al. продемонстрирована индукция образования эндодермальных клеток brachyury+/HNF-3beta+ из эмбриональных стволовых клеток мыши в отсутствие сыворотки и в присутствии активина в сочетании с ингибитором сигнального пути Wnt (US 2006/0003446A1).
В публикации Gordon et al. (PNAS, Vol. 103, страница 16806, 2006) говорится: «Для образования передней первичной полоски требовались, одновременно, сигнальные пути Wnt и TGF-бета/Nodal/активин».
Однако модель развития эмбриональных стволовых клеток на мышах может не имитировать в точности программу развития у высших млекопитающих, как, например, у человека.
Thomson et al. выделяли эмбриональные стволовые клетки из человеческих бластоцист (Science 282: 114, 1998). Параллельно, Gearhart и соавторы получили клеточные линии человеческих эмбриональных зародышевых клеток (чЭЗ) из ткани половых желез эмбриона (Shamblott et al., Proc. Natl. Acad. Sci. USA 95: 13726, 1998). В отличие от эмбриональных стволовых клеток мыши, воспрепятствовать дифференцировке которых можно путем простого культивирования с фактором торможения лейкемиии (Leukemia Inhibitory Factor, LIF), человеческие эмбриональные стволовые клетки должны культивироваться в очень специфических условиях (США, патенты №№ 6200806; WO 99/20741; WO 01/51616).
D'Amour et al. описывают получение обогащенных культур сформировавшейся эндодермы, производной от человеческих эмбриональных стволовых клеток, в присутствии высокой концентрации активина и низкой концентрации сыворотки (Nature Biotechnology 2005). Трансплантация этих клеток под почечную капсулу мышей приводит к их дифференцировке в более зрелые клетки, обладающие характерными особенностями некоторых эндодермальных органов. Клетки сформировавшейся эндодермы, производные от человеческих эмбриональных стволовых клеток, могут подвергаться дальнейшей дифференцировке в Pdx1-положительные клетки после добавления FGF-10 (US 2005/0266554A1).
В публикации D'Amour et al. (Nature Biotechnology - 24, 1392-1401 (2006)) говорится: «Мы разработали процесс дифференцировки, преобразующий человеческие эмбриональные стволовые клетки (чЭС) в эндокринные клетки, способные синтезировать гормоны поджелудочной железы, инсулин, глюкагон, соматостатин, панкреатический полипептид и грелин. Данный процесс имитирует органогенез поджелудочной железы in vivo, проводя клетки через фазы, напоминающие образование сформировавшейся эндодермы, эндодермы кишечной трубки, панкреатической эндодермы и превращение предшественников эндокринных клеток в клетки, экспрессирующие эндокринные гормоны».
В другом примере, Fisk et al. сообщают о системе для получения островковых клеток поджелудочной железы из человеческих эмбриональных стволовых клеток (заявка на патент US2006/0040387A1). В данном случае процесс дифференцировки был разделен на три стадии. Сначала человеческие эмбриональные стволовые клетки были дифференцированы до эндодермы с помощью сочетания бутирата натрия и активина А. Далее клетки культивировались с антагонистами TGF-β, такими как Noggin, в сочетании с EGF или бета-целлюлином с получением Pdx1-положительных клеток. Окончательная дифференцировка запускалась никотинамидом.
В одном примере, Benvenistry et al. сообщают: «Мы делаем вывод, что сверхэкспрессия Pdx1 увеличивала экспрессию панкреатических обогащенных генов, а для индукции экспрессии инсулина могут требоваться дополнительные сигналы, присутствующие только in vivo» (Benvenistry et al., Stem Cells 2006; 24: 1923-1930).
В другом примере, Odorico et al. сообщается о способах прямой дифференцировки in vitro полипотентных стволовых клеток млекопитающих в клетки панкреатической линии дифференцировки. Данные способы включают культивацию стволовых клеток в присутствии эффективного количества костного морфогенетического белка с индукцией дифференцировки в направлении мезэндодермы. Эти мезэндодермальные клетки далее культивируются до образования эмбриоидных телец (ЭТ), обогащенных клетками, коммитированными в сторону сформировавшейся эндодермы, которые при определенных условиях, в итоге, дифференцируются в клетки панкреатической линии дифференцировки (US20070259423).
В другом примере, Tulachan et al. (Developmental Biology, 305, 2007, страницы 508-521) утверждают: «Ингибирование сигнального пути TGF-B в эмбриональный период может, таким образом, позволить клеткам эпителия поджелудочной железы развиться до эндокринной линии дифференцировки».
Следовательно, острой потребностью по-прежнему остается разработка условий для создания стабильных линий полипотентных стволовых клеток, способных быть размноженными для решения текущих клинических задач и сохраняющих потенциал дифференцировки в эндокринные клетки поджелудочной железы, в клетки, экспрессирующие гормоны поджелудочной железы, или в клетки, секретирующие гормоны поджелудочной железы и обладающие способностью секретировать инсулин в ответ на изменение концентрации глюкозы. Мы использовали альтернативный подход к повышению эффективности дифференцировки человеческих эмбриональных стволовых клеток в сторону эндокринных клеток поджелудочной железы, способных секретировать инсулин в ответ на изменение концентрации глюкозы.
КРАТКОЕ ОПИСАНИЕ
В одном варианте осуществления настоящего изобретения предлагается способ получения клеток, способных секретировать инсулин в ответ на стимуляцию глюкозой, из полипотентных стволовых клеток, включающий следующие стадии:
культивирование полипотентных стволовых клеток,
дифференцировку полипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы,
дифференцировку клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, и
дифференцировку клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы.
В одном варианте осуществления настоящего изобретения предлагается способ активизации стимулируемой глюкозой секреции инсулина в клетках, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы, полученных из полипотентных стволовых клеток, включающий следующие стадии:
культивирование полипотентных стволовых клеток,
дифференцировку полипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы,
дифференцировку клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, и
дифференцировку клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы.
В одном из вариантов осуществления экспрессирующие маркеры, характерные для линии панкреатической эндодермы, дифференцируются из клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, любым из следующих способов:
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, фактором роста фибробластов и ингибитором сигнального пути Hedgehog; затем удаление среды, содержащей фактор роста фибробластов и ингибитор сигнального пути Hedgehog с последующей культивацией клеток в среде, содержащей ретиноевую кислоту, фактор роста фибробластов и ингибитор сигнального пути Hedgehog, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой и по меньшей мере одним из факторов роста фибробластов, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой, ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов и по меньшей мере одним фактором, способным ингибировать костный морфогенетический белок (bone morphogenetic protein, BMP), или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой, ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов и нетрином, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой, ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов, по меньшей мере одним фактором, способным ингибировать BMP и нетрином, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы по меньшей мере одним фактором роста фибробластов и ингибитором фактора «Sonic Hedgehog», затем удаление среды, содержащей по меньшей мере один фактор роста фибробластов и ингибитор фактора «Sonic Hedgehog», с последующей обработкой клеток ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов и ретиноевой кислотой, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы по меньшей мере одним фактором роста фибробластов и ингибитором фактора «Sonic Hedgehog», затем удаление среды, содержащей по меньшей мере один фактор роста фибробластов и ингибитор фактора «Sonic Hedgehog», с последующей обработкой клеток ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов, ретиноевой кислотой и по меньшей мере одним фактором, способным ингибировать BMP, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы по меньшей мере одним фактором роста фибробластов и ингибитором фактора «Sonic Hedgehog», затем удаление среды, содержащей по меньшей мере один фактор роста фибробластов и ингибитор фактора «Sonic Hedgehog», с последующей обработкой клеток ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов, ретиноевой кислотой и нетрином, или
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы по меньшей мере одним фактором роста фибробластов и ингибитором фактора «Sonic Hedgehog», затем удаление среды, содержащей по меньшей мере один фактор роста фибробластов и ингибитор фактора «Sonic Hedgehog», с последующей обработкой клеток ингибитором фактора «Sonic Hedgehog», по меньшей мере одним фактором роста фибробластов, ретиноевой кислотой, по меньшей мере одним фактором, способным ингибировать BMP, и нетрином.
В одном варианте осуществления настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, дифференцируются из клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, любым из следующих способов:
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей ингибитор γ-секретазы и агонист GLP-1, затем удаление среды, содержащей ингибитор γ-секретазы и агонист GLP-1, с последующей культивацией клеток в среде, содержащей агонист GLP-1, IGF-1 и HGF, или
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей агонист GLP-1, затем удаление среды, содержащей агонист GLP-1, с последующей культивацией клеток в среде, содержащей агонист GLP-1, IGF-1 и HGF, или
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей ингибитор γ-секретазы и агонист GLP-1, или
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей агонист GLP-1, или
обработка клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, или
обработка клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь TGF-βR-1, или
обработка клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1, или
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей от приблизительно 10 мМ до приблизительно 20 мМ глюкозы и агонист GLP-1.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана схема протокола дифференцировки, используемого в настоящем изобретении. Панель a) относится к способу, описанному в заявке на патент США № 11/736908, панели b) и c) представляют способы, являющиеся предметом настоящего изобретения.
На фиг.2 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 2. Экспрессия маркеров панкреатической эндодермы (PDX-1, ISL-1) и эндокринных маркеров (NeuroD, инсулин и глюкагон) показана на стадиях с 3 по 5 (S3-S5). На стадиях с 4 по 5 наблюдалось достоверное увеличение экспрессии инсулина и глюкагона.
На фиг.3 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 3. Панкреатические эндокринные маркеры a) инсулин, b) NKX2, c) глюкагон, d) NeuroD и маркер панкреатической эндодермы e) PDX-1 на стадиях с 3 по 5. Влияние ингибиторов различных киназ оценивалось на стадии 3 или 4, либо на стадиях 3 и 4 (+/+ обозначает присутствие данного соединения как на стадии 3, так и на стадии 4, +/- обозначает присутствие данного соединения на стадии 3 и его отсутствие на стадии 4, -/+ обозначает присутствие данного соединения на стадии 4 и его отсутствие на стадии 3).
На фиг.4 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 4. Панкреатические маркеры a) глюкагон и инсулин, b) HAND1 и NeuroD, c) HNF4a и PDX-1, d) NKX2.2 и Sox17 показаны для стадий с 3 по 5. Эффект ингибитора ALK5 II оценивался на стадии 4 или 5, либо на стадиях 4 и 5 (+/+ обозначает присутствие ингибитора на обеих стадиях 4 и 5, +/- обозначает присутствие ингибитора на стадии 4 и его отсутствие на стадии 5, -/+ обозначает присутствие ингибитора на стадии 5 и его отсутствие на стадии 4).
На фиг.5 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 5. Эффект 1-10 мкМ ингибиторов I и II ALK5 на экспрессию: a) глюкагона, b) инсулина, c) PDX-1, d) NeuroD, показан для стадий с 4 по 5. При контрольной обработке ни один из ингибиторов ALK5 в ходе процесса дифференцировки не использовался.
На фиг.6 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 6. Эффект сочетания 1 мкМ ингибитора ALK5 и 0-500 нг/мл рекомбинантного человеческого фактора Noggin на экспрессию a) альбумина, b) CDX2, c) инсулина, d) глюкагона, e) PDX-1, f) NeuroD и g) NKX2.2, показан для стадий с 3 по 5. Ингибитор ALK5 добавлялся на стадиях 4 и 5, а белок Noggin добавлялся на стадии 3.
На фиг.7 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 7. Эффект сочетания 1 мкМ ингибитора ALK5 и 100 нг/мл рекомбинантного человеческого белка Noggin на экспрессию a) инсулина, b) глюкагона, c) ngn3, d) NeuroD, e) CDX-2, f) PDX-1, показан для стадий 4 и 5. Ингибитор ALK5 добавлялся на стадиях 4 и 5, а белок Noggin добавлялся на стадиях 3, 4 или 3 и 4.
На фиг.8 показана морфология клеток, дифференцированных в соответствии со способами, описанными в примере 7: a) 4-кратное фазово-контрастное изображение клеток стадии 5 на 6 день, b) 10-кратное фазово-контрастное изображение клеток стадии 5 на 6 день, c) 4-кратное фазово-контрастное изображение клеток стадии 5 на 12 день, d) 10-кратное фазово-контрастное изображение клеток стадии 5 на 12 день.
На фиг.9 показаны иммунофлуоресцентные изображения клеток, дифференцированных в соответствии со способами, описанными в примере 7. Клетки находятся на стадии 5, день 12. a) Инсулиновое окрашивание единичного кластера, а также в сочетании с b) ядерным красителем DAPI.
На фиг.10 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 7.
На фиг.11 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 8. Демонстрируется эффект сочетания белка Noggin, добавляемого на стадиях 3 и 4, с белком нетрин-4 и/или ингибитором ALK 5, добавляемым на стадии 4, на экспрессию a) ngn3, b) PDX-1, c) NeuroD, d) Pax4, e) инсулина и f) глюкагона.
На фиг.12 показан анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 8. Панель a) инсулин, b) глюкагон, c) ngn3, d) NKX2.2, e) NeuroD и f) PDX-1.
На фиг.13 показана стимулированная глюкозой секреция инсулина (Glucose Stimulated Insulin Secretion, GSIS) in vitro в длительно культивируемых культурах стадии 5. Клетки стадии 5, подготовленные в соответствии со способами, описанными в примере 9, стимулировались глюкозой в день a) 6, b) 12 и c) 8-20 дней в культуре стадии 5.
На фиг.14 показан эффект белка нетрин-1 или нетрин-2 на экспрессию эндокринных маркеров. Анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 10.
На фиг.15 показана индукция появления эндокринных маркеров с использованием альтернативного метода индукции образования сформировавшейся эндодермы. Анализ методом ПЦР в реальном времени линии человеческих эмбриональных стволовых клеток H1, дифференцируемых в соответствии со способами, описываемыми в примере 11. Панель a) инсулин, b) глюкагон, c) NKX2.2, d) Pax4, e) PDX-1 и f) NeuroD.
На фиг.16 показана экспрессия различных маркеров на разных стадиях протокола дифференцировки, приведенного на фиг.1, с. Панель a) показана экспрессия CXCR4, определенная методом FACS (сортировкой флуоресцентно-активированных клеток) в клетках H1 на третий день стадии 1. Панель b) Показана экспрессия маркеров, характерных для линии сформировавшейся эндодермы и внеэмбриональной линии, в клетках на третий день стадии 1, измеренная методом ПЦР в реальном времени. Панели c)-n) демонстрируют экспрессию различных генов в клетках, собранных в конце стадий 2-6, измеренную методом ПЦР в реальном времени.
На фиг.17 показано количество клеток на разных стадиях протокола дифференцировки, приведенного на фиг.1, с.
На фиг.18 показана секреция C-пептида клетками в конце стадии 6 протокола дифференцировки, приведенного на фиг.1, c, в ответ на различные стимулы.
На фиг.19 показано содержание в клетках C-пептида и проинсулина в конце стадии 6 протокола дифференцировки, приведенного на фиг.1, c, в сравнении с панкреатическими островками взрослых людей.
На фиг.20 показана экспрессия инсулина (панель a), синаптофизина (панель b) и ко-экспрессия синаптофизина (ось X) и инсулина (ось Y) (панель c) в клетках в конце стадии 6 протокола дифференцировки, приведенного на фиг.1, c.
На фиг.21 показана экспрессия синаптофизина (панели a и c) и инсулина (панели b и d) в конце стадии 6 протокола дифференцировки, приведенного на фиг.1, c, в клетках, обработанных 100 нг/мл белка Chordin вместо белка Noggin на стадиях 3-4.
На фиг.22 показана экспрессия различных маркеров в клетках линии человеческих эмбриональных стволовых клеток H9, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, c. Приведенные данные представляют собой экспрессию a) HNF4a, b) HNF6, c) CDX2, d) PDX-1, e) NKX6.1, f) Pax4, g) NKX2.2, h) NeuroD, i) NGN3, j) PECAM, k) глюкагона и l) инсулина, измеренную методом ПЦР в реальном времени в клетках, собранных в конце стадий 3-6.
На фиг.23 показана экспрессия различных маркеров в клетках линии человеческих эмбриональных стволовых клеток H1, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, c, где клетки были обработаны с помощью B27 или N2. Приведенные данные представляют собой экспрессию a) CDX2, b) глюкагона, c) инсулина и d) PDX-1, измеренную методом ПЦР в реальном времени в клетках, собранных в конце стадий 3-6.
ПОДРОБНОЕ ОПИСАНИЕ
Для ясности описания, а не для ограничения изобретения, подробное описание изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяющиеся по их способности на уровне единичной клетки как самообновляться, так и дифференцироваться с образованием клеток-потомков, таких как самообновляющиеся предшественники, необновляющиеся предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных линий, происходящих из нескольких зародышевых листков (эндодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от нескольких зародышевых листков, и вносить существенный вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисту.
По потенциалу развития стволовые клетки классифицируются следующим образом: (1) тотипотентные, т.е. способные давать начало всем эмбриональным и внеэмбриональным типам клеток; (2) полипотентные, т.е. способные давать начало всем эмбриональным типам клеток; (3) мультипотентные, т.е. способные давать начало группе клеточных линий, в пределах конкретной ткани, органа или физиологической системы (например, гематопоэтические стволовые клетки (ГСК) могут давать таких потомков, как ГСК (самообновление), олигопотентные предшественники, ограниченные клетками крови, и все типы клеток и клеточных элементов (таких как тромбоциты), являющиеся нормальными компонентами крови); (4) олигопотентные, т.е. способные давать начало более ограниченному набору клеточных линий, чем мультипотентные стволовые клетки; и (5) унипотентные, т.е. способные давать начало единственной клеточной линии (например, сперматогенные стволовые клетки).
Дифференцировка представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или малоспециализированная клетка приобретает свойства специализированной клетки, например, нервной или мышечной. Дифференцированная клетка или клетка с индуцированной дифференцировкой представляет собой клетку, занявшую более специализированную («коммитированную») позицию в пределах клеточной линии дифференцировки. Термин «коммитированная», применительно к процессу дифференцировки, обозначает клетку, дошедшую в ходе дифференцировки до стадии, от которой, в нормальных условиях, она продолжит дифференцироваться до определенного типа клеток или набора типов клеток и не сможет, в нормальных условиях, дифференцироваться в иной тип клеток или вернуться к менее дифференцированному типу. Дедифференцировкой называется процесс, в ходе которого клетка возвращается к менее специализированной (или коммитированной) позиции в линии дифференцировки. В настоящем документе линией дифференцировки клеток называется наследственность клетки, т.е. от каких клеток она происходит и каким клеткам может дать начало. В линии дифференцировки клетка помещается в наследственную схему развития и дифференцировки. Маркером, специфичным для линии дифференцировки, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцировки, которая может использоваться для оценки дифференцировки некоммитированных клеток в данную линию дифференцировки.
«b-клеточной линией дифференцировки» называются клетки, положительные по экспрессии гена транскрипционного фактора PDX-1 и по меньшей мере одного из следующих транскрипционных факторов: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 beta, MAFA, Pax4 и Pax6. К клеткам, экспрессирующим маркеры, характерные для b клеточной линии дифференцировки, относятся b клетки.
Термин «клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы» в настоящем документе обозначает клетки, экспрессирующие по меньшей мере один из следующих маркеров: SOX-17, GATA-4, HNF-3 бета, GSC, Cer1, Nodal, FGF8, Brachyury, Mix homeobox-подобный белок, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF17, GATA-6, CXCR4, C-Kit, CD99 или OTX2. К клеткам, экспрессирующим маркеры, характерные для линии сформировавшейся эндодермы, относятся клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезэндодермы и клетки сформировавшейся эндодермы.
Термин «клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы» в настоящем документе обозначает клетки, экспрессирующие по меньшей мере один из следующих маркеров: PDX-1, HNF-1beta, PTF-1 альфа, HNF-6 или HB9. К клеткам, экспрессирующим маркеры, характерные для линии панкреатической эндодермы, относятся клетки панкреатической эндодермы, клетки первичной кишечной трубки и клетки поздней передней кишки.
Термин «клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы» в настоящем документе обозначает клетки, экспрессирующие по меньшей мере один из следующих маркеров: NGN-3, NeuroD, Islet-1, PDX-1, NKX6.1, Pax-4, Ngn-3 или PTF-1 альфа. К клеткам, экспрессирующим маркеры, характерные для линии эндокринных клеток поджелудочной железы, относятся панкреатические эндокринные клетки, клетки, экспрессирующие гормоны поджелудочной железы, клетки, секретирующие гормоны поджелудочной железы и клетки b-клеточной линии дифференцировки.
«Сформировавшейся эндодермой» в настоящем документе называются клетки, обладающие характерными особенностями клеток, происходящих в ходе гаструляции от эпибласта и формирующих желудочно-кишечный тракт и его производные. Клетки сформировавшейся эндодермы экспрессируют следующие маркеры: HNF-3 beta, GATA-4, SOX-17, Cerberus, OTX2, goosecoid, C-Kit, CD99 и Mixl1.
«Внеэмбриональной эндодермой» в настоящем документе называется популяция клеток, экспрессирующих по меньшей мере один из следующих маркеров: SOX-7, AFP и SPARC.
«Маркерами» в настоящем документе называются молекулы нуклеиновых кислот или полипептидов, дифференциально экспрессируемые в конкретной клетке. В данном контексте, под дифференциальной экспрессией подразумевается повышенный уровень положительного маркера и пониженный уровень отрицательного маркера. Определяемый уровень маркерной аминокислоты или полипептида в интересующих клетках значительно выше или ниже, в сравнении с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из разнообразных способов, известных в данной области.
«Мезэндодермальной клеткой» в настоящем документе называется клетка, экспрессирующая по меньшей мере один из следующих маркеров: CD48, эомезодермин (EOMES), SOX-17, DKK4, HNF-3 beta, GSC, FGF17, GATA-6.
«Панкреатической эндокринной клеткой» или «клеткой, экспрессирующей гормон поджелудочной железы» в настоящем документе называется клетка, экспрессирующая по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
«Панкреатической эндодермальной клеткой» в настоящем документе называется клетка, экспрессирующая по меньшей мере один из следующих маркеров: NGN-3, NeuroD, Islet-1, PDX-1, PAX-4, NKX2.2.
«Клеткой, продуцирующей гормон поджелудочной железы» в настоящем документе называется клетка, способная производить по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
«Клеткой, секретирующей гормон поджелудочной железы» в настоящем документе называется клетка, способная секретировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
«Поздней клеткой передней кишки» в настоящем документе называется клетка, способная секретировать по меньшей мере один из следующих маркеров: PDX-1, HNF-1, PTF-1A, HNF-6, HB-9, PROX-1.
«Клеткой предпервичной полоски» в настоящем документе называется клетка, способная экспрессировать по меньшей мере один из следующих маркеров: Nodal или FGF8.
«Клеткой первичной кишечной трубки» в настоящем документе называется клетка, способная секретировать по меньшей мере один из следующих маркеров: HNF-1, HNF-4A.
«Клеткой первичной полоски» в настоящем документе называется клетка, способная экспрессировать по меньшей мере один из следующих маркеров: Brachyury, Mix homeobox-подобный белок или FGF4.
ВЫДЕЛЕНИЕ, РАЗМНОЖЕНИЕ И КУЛЬТИВИРОВАНИЕ ПОЛИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК
Характеристика полипотентных стволовых клеток
Полипотентные стволовые клетки могут экспрессировать один или несколько стадийно-специфичных эмбриональных антигенов (stage-specific embryonic antigen, SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282: 1145, 1998). Дифференцировка полипотентных стволовых клеток in vitro приводит к утрате экспрессии SSEA-4, Tra-1-60 и Tra-1-81 (если имеются) и к увеличению экспрессии SSEA-1. В недифференцированных полипотентных стволовых клетках, как правило, активна щелочная фосфатаза, которая может быть обнаружена путем фиксации клеток с помощью 4% параформальдегида, с последующим обнаружением с помощью Vector Red, применяемого в качестве субстрата, в соответствии с инструкциями производителя (Vector Laboratories, Burlingame Calif.) Недифференцированные полипотентные стволовые клетки также, как правило, экспрессируют Oct-4 и TERT, обнаруживаемые методом ОТ-ПЦР.
Другим желательным фенотипическим свойством выращенных полипотентных клеток является потенциал дифференцировки в клетки всех трех зародышевых листков: в эндодермальные, мезодермальные и эктодермальные ткани. Полипотентность полипотентных стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида, и их гистологического исследования для получения доказательств наличия клеточных типов, происходящих от трех зародышевых листков. Как вариант, полипотентность можно определить путем создания эмбриоидных телец и анализа их на предмет присутствия маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии полипотентных стволовых клеток могут быть кариотипированы с применением стандартного метода окрашивания с использованием красителя Гимза (G-banding) и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, причем все человеческие хромосомы должны присутствовать и не иметь видимых изменений.
Источники полипотентных стволовых клеток
К типам полипотентных стволовых клеток, которые можно использовать, относятся стабильные линии полипотентных клеток, получаемых из ткани, образующейся после беременности, в том числе, из преэмбриональной ткани (такой как бластоциста), эмбриональной ткани или ткани плода, взятой в любой момент в ходе беременности, как правило, но не обязательно, до срока приблизительно 10-12 недель беременности. Примерами, не ограничивающими настоящее изобретение, являются стабильные линии человеческих эмбриональных стволовых клеток или человеческих эмбриональных зародышевых клеток, например, клеточные линии человеческих эмбриональных стволовых клеток H1, H7 и H9 (WiCell). Также предусмотрено использование описываемых здесь составов в ходе первоначального установления или стабилизации таких клеток, и в этом случае исходными клетками могут быть первичные полипотентные клетки, взятые напрямую из тканей, служащих источником. Также подходящими являются клетки, взятые из популяции полипотентных стволовых клеток, уже культивируемых в отсутствие питающих клеток. Также подходящими являются мутантные линии человеческих эмбриональных стволовых клеток, например, BG01v (BresaGen, Athens, GA).
В одном из вариантов осуществления, человеческие эмбриональные стволовые клетки подготавливаются, как описано в публикации Thomson et al. (США патент № 5843780; Science 282: 1145, 1998; Curr. Top. Dev. Biol. 38: 133 ff., 1998; Proc. Natl. Acad. Sci. U.S.A. 92: 7844, 1995).
Культивирование полипотентных стволовых клеток
В одном из вариантов осуществления полипотентные стволовые клетки, как правило, культивируются на слое питающих клеток, которые оказывают разностороннюю поддержку полипотентным клеткам. Как вариант, полипотентные стволовые клетки культивируются в культуральной системе, по существу не содержащей питающих клеток, но, тем не менее, поддерживающей пролиферацию полипотентных стволовых клеток и не допускающей существенной дифференцировки. Выращивание полипотентных стволовых клеток в культуре, не содержащей питающих клеток, без дифференцировки, осуществляется при помощи среды, кондиционированной путем предварительного культивирования в ней клеток другого типа. Как вариант, выращивание полипотентных стволовых клеток в культуре, не содержащей питающих клеток, без дифференцировки, осуществляется при помощи среды с определенным химическим составом.
Полипотентные стволовые клетки могут быть нанесены на подходящий культуральный субстрат. В одном из вариантов осуществления подходящим культуральным субстратом является компонент экстраклеточного матрикса, например, полученный из базальной мембраны или тот, который может участвовать в образовании связи между рецептором молекул адгезии и лигандом. В одном из вариантов осуществления подходящим культуральным субстратом является MATRIGEL® (Becton Dickenson). MATRIGEL® - это растворимый препарат из клеток опухоли Engelbreth-Holm-Swarm, который при комнатной температуре представляет собой гель, образующий восстановленную базальную мембрану.
Как вариант, можно использовать другие компоненты экстраклеточного матрикса и смеси компонентов. В зависимости от типа пролиферирующих клеток, это может быть ламинин, фибронектин, протеогликан, энтактин, гепарансульфат и т.п., отдельно или в различных сочетаниях.
Полипотентные стволовые клетки могут наноситься на субстрат с применением подходящего распределения и в присутствии среды, способствующей выживанию, размножению клеток и сохранению ими желательных особенностей. На все эти характеристики оказывает влияние аккуратность распределения клеток при посеве, которая легко может достигаться специалистом в данной области.
Подходящая культуральная среда может состоять из следующих компонентов, например, модифицированная по способу Дульбекко среда Игла (DMEM), Gibco #11965-092; модифицированная по способу Дульбекко среда Игла Knockout (KO DMEM), Gibco #10829-018; базальная среда Хэма F12/50% DMEM; 200 мМ L-глутамина, Gibco #15039-027; раствор заменимых аминокислот, Gibco #11140-050; β-меркаптоэтанол, Sigma #M7522; человеческий рекомбинантный основной фактор роста фибробластов (bFGF), Gibco #13256-029.
Создание клеток, способных секретировать инсулин при стимуляции глюкозой, из полипотентных стволовых клеток
В одном из вариантов осуществления настоящего изобретения предлагается способ получения клеток, способных секретировать инсулин в ответ на стимуляцию глюкозой, из полипотентных стволовых клеток, включающий следующие стадии:
культивирование полипотентных стволовых клеток,
дифференцировку полипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы,
дифференцировку клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, и
дифференцировку клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы.
В одном из вариантов осуществления настоящего изобретения предлагается способ активизации стимулируемой глюкозой секреции инсулина в клетках, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы, полученных из полипотентных стволовых клеток, включающий следующие стадии:
культивирование полипотентных стволовых клеток,
дифференцировку полипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы,
дифференцировку клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, и
дифференцировку клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы.
К полипотентным стволовым клеткам, подходящим для использования в настоящем изобретении, относятся, например, линия человеческих эмбриональных стволовых клеток H9 (код NIH: WA09), линия человеческих эмбриональных стволовых клеток H1 (код NIH: WA01), линия человеческих эмбриональных стволовых клеток H7 (код NIH: WA07) и линия человеческих эмбриональных стволовых клеток SA002 (Cellartis, Sweden). Также подходят для использования в рамках настоящего изобретения клетки, экспрессирующие по меньшей мере один из следующих маркеров, характерных для полипотентных клеток: ABCG2, cripto, CD9, FoxD3, Connexin43, Connexin45, Oct4, Sox2, Nanog, hTERT, UTF-1, ZFP42, SSEA-3, SSEA-4, Tra1-60, Tra1-81.
Маркеры, характерные для линии сформировавшейся эндодермы, выбираются из группы, включающей SOX-17, GATA4, Hnf-3beta, GSC, Cer1, Nodal, FGF8, Brachyury, Mix homeobox-подобный белок, FGF4 CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Подходит для использования в настоящем изобретении клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии сформировавшейся эндодермы. В одном аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для линии сформировавшейся эндодермы, представляет собой клетку-предшественницу первичной полоски. В другом аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для линии сформировавшейся эндодермы, представляет собой мезэндодермальную клетку. В другом аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для линии сформировавшейся эндодермы, представляет собой клетку сформировавшейся эндодермы.
Маркеры, характерные для линии панкреатической эндодермы, выбираются из группы, включающей Pdx1, HNF-1beta, PTF1a, HNF-6, HB9 и PROX1. Подходит для использования в настоящем изобретении клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии панкреатической эндодермы. В одном аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для линии панкреатической эндодермы, представляет собой клетку панкреатической эндодермы.
Маркеры, характерные для линии эндокринных клеток поджелудочной железы, выбираются из группы, включающей NGN-3, NeuroD, Islet-1, Pdx-1, NKX6.1, Pax-4, Ngn-3 и PTF-1 alpha. В одном из вариантов осуществления, панкреатической эндокринной клеткой является клетка, способная экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Подходит для использования в настоящем изобретении клетка, экспрессирующая по меньшей мере один из маркеров, характерных для линии эндокринных клеток поджелудочной железы. В одном аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для линии эндокринных клеток поджелудочной железы, представляет собой панкреатическую эндокринную клетку. Панкреатическая эндокринная клетка может представлять собой клетку, экспрессирующую гормоны поджелудочной железы. Как вариант, панкреатическая эндокринная клетка может представлять собой клетку, секретирующую гормоны поджелудочной железы.
В одном аспекте настоящего изобретения панкреатическая эндокринная клетка представляет собой клетку, экспрессирующую маркеры, характерные для β-клеточной линии. Клетка, экспрессирующая маркеры, характерные для β-клеточной линии, экспрессирует Pdx1 и по меньшей мере один из следующих транскрипционных факторов: NGN-3, Nkx2.2, Nkx6.1, NeuroD, Isl-1, HNF-3 beta, MAFA, Pax4 и Pax6. В одном аспекте настоящего изобретения, клетка, экспрессирующая маркеры, характерные для β-клеточной линии, представляет собой β-клетку.
Создание клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы
Полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, с использованием любого известного специалистам способа или с использованием любого способа, предложенного в настоящем изобретении.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, в соответствии со способами, описанными в публикации D'Amour et al., Nature Biotechnology 23, 1534-1541 (2005).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, в соответствии со способами, описанными в публикации Shinozaki et al., Development 131, 1651-1662 (2004).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, в соответствии со способами, описанными в публикации McLean et al., Stem Cells 25, 29-38 (2007).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, в соответствии со способами, описанными в публикации D'Amour et al., Nature Biotechnology 24, 1392-1401 (2006).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем культивирования полипотентных стволовых клеток в среде, содержащей активин A в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой, а затем культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа описан в публикации Nature Biotechnology 23, 1534-1541 (2005).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем культивирования полипотентных стволовых клеток в среде, содержащей активин А в отсутствие сыворотки, затем культивирования клеток с активином A и сывороткой в другой концентрации. Пример данного способа описан в публикации D'Amour et al., Nature Biotechnology, 2005.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем культивирования полипотентных стволовых клеток в среде, содержащей активин А и лиганд Wnt в отсутствие сыворотки, затем удаления лиганда Wnt и культивирования клеток с активином A и сывороткой. Пример данного способа описан в публикации Nature Biotechnology 24, 1392-1401 (2006).
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 11/736908, принадлежащей LifeScan, Inc.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 11/779311, принадлежащей LifeScan, Inc.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 61/076889.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 61/076900.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 61/076908.
Например, полипотентные стволовые клетки могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, путем обработки полипотентных стволовых клеток в соответствии со способами, изложенными в заявке на патент США серийный № 61/076915.
Выявление клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы
Образование клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, может быть выявлено путем проверки на наличие маркеров до и после выполнения конкретного протокола. Полипотентные стволовые клетки, как правило, не экспрессируют данных маркеров. Таким образом, дифференцировка полипотентных клеток обнаруживается, когда клетки начинают экспрессировать эти маркеры.
Эффективность дифференцировки может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии сформировавшейся эндодермы.
Методы анализа экспрессии маркерных белков и нуклеиновых кислот в культивированных или изолированных клетках являются стандартными для данной области. Сюда относятся количественная ревертазная полимеразная цепная реакция (ОТ-ПЦР), Нозерн-блот, гибридизация in situ (смотрите, например, Current Protocols in Molecular Biology (Ausubel et al., eds. 2001 supplement)), и иммунологические методы, такие как иммуногистохимический анализ среза материала, Вестерн-блот, а для маркеров, доступных в интактных клетках, метод проточной цитометрии (FACS) (смотрите, например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998)).
Характерные особенности полипотентных стволовых клеток хорошо известны специалистам, и дополнительные характеристики еще продолжают выявляться. К маркерам полипотентных стволовых клеток относятся, например, экспрессия одного или нескольких следующих факторов: ABCG2, cripto, FoxD3, Connexin43, Connexin45, Oct4, Sox2, Nanog, hTERT, UTF-1, ZFP42, SSEA-3, SSEA-4, Tra1-60, Tra1-81.
После обработки полипотентных стволовых клеток с применением методов, составляющих предмет изобретения, дифференцированные клетки могут быть выделены путем воздействия на популяцию клеток агентом (например, антителом), специфически распознающим белковый маркер, например, CXCR4, экспрессируемый клетками, экспрессирующими маркеры, характерные для линии сформировавшейся эндодермы.
Создание клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы
Клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, с использованием любого известного специалистам способа или с использованием любого способа, предложенного в настоящем изобретении.
Например, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, описанными в публикации D'Amour et al, Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, фактором роста фибробластов и ингибитором сигнального пути Hedgehog KAAD-циклопамином, затем удаления среды, содержащей фактор роста фибробластов и KAAD-циклопамин с последующим культивированием клеток в среде, содержащей ретиноевую кислоту, фактор роста фибробластов и KAAD-циклопамин. Пример данного способа описан в публикации Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой и по меньшей мере одним фактором роста фибробластов в течение периода времени, соответствующего способам, описанным в заявке на патент США серийный № 11/736908, принадлежащей LifeScan, Inc.
Например, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, ретиноевой кислотой и по меньшей мере одним фактором роста фибробластов в течение периода времени, соответствующего способам, описанным в заявке на патент США серийный № 11/779311, принадлежащей LifeScan, Inc.
В одном из вариантов осуществления настоящего изобретения предлагается способ дифференцировки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, включающий следующие стадии:
культивирование клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, и
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина.
В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина, в течение периода от приблизительно одного до приблизительно шести дней. В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина, в течение приблизительно шести дней.
При использовании этого способа любая клетка, экспрессирующая маркеры, характерные для линии сформировавшейся эндодермы, может дифференцироваться в клетку, экспрессирующую маркеры, характерные для линии панкреатической эндодермы.
В альтернативном варианте осуществления настоящего изобретения предлагается способ дифференцировки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, включающий следующие стадии:
культивирование клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы,
обработка клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты и фактора роста фибробластов, и
удаление по меньшей мере одного фактора, выбранного из группы, состоящей из ретиноевой кислоты и фактора роста фибробластов с последующей обработкой клеток по меньшей мере одним фактором, выбранным из группы, состоящей из ингибитора фактора «Sonic Hedgehog», ретиноевой кислоты, фактора роста фибробластов, фактора, способного ингибировать BMP, и нетрина.
В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты и фактора роста фибробластов, в течение периода от приблизительно одного до приблизительно трех дней. В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты и фактора роста фибробластов, в течение приблизительно трех дней. В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ингибитора фактора «Sonic Hedgehog», ретиноевой кислоты, фактора роста фибробластов, фактора, способного ингибировать BMP, и нетрина, в течение периода от приблизительно одного до приблизительно четырех дней. В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ингибитора фактора «Sonic Hedgehog», ретиноевой кислоты, фактора роста фибробластов, фактора, способного ингибировать BMP, и нетрина, в течение приблизительно четырех дней.
При использовании этого способа любая клетка, экспрессирующая маркеры, характерные для линии сформировавшейся эндодермы, может дифференцироваться в клетку, экспрессирующую маркеры, характерные для линии панкреатической эндодермы.
В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, произведенные при помощи способов, составляющих предмет изобретения, демонстрируют пониженный уровень экспрессии маркеров, ассоциированных с тканями печени и кишечника. В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, произведенные при помощи способов, составляющих предмет изобретения, демонстрируют пониженный уровень экспрессии альбумина и CDX-2.
По меньшей мере один фактор роста фибробластов выбран из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10.
В одном из вариантов осуществления, BMP представляет собой BMP4. В одном из вариантов осуществления по меньшей мере один фактор, способный ингибировать BMP4, представляет собой Noggin.
Нетрин выбран из группы, состоящей из нетрина 1, нетрина 2 и нетрина 4.
Ретиноевая кислота может использоваться в концентрациях от приблизительно 1 нМ до приблизительно 1 мМ. В одном из вариантов осуществления, ретиноевую кислоту используют в концентрации 1 мкМ.
FGF-2 может использоваться в концентрациях от приблизительно 50 пг/мл до приблизительно 50 мкг/мл. В одном из вариантов осуществления, FGF-2 используют в концентрации 50 нг/мл.
FGF-4 может использоваться в концентрациях от приблизительно 50 пг/мл до приблизительно 50 мкг/мл. В одном из вариантов осуществления, FGF-4 используют в концентрации 50 нг/мл.
FGF-7 может использоваться в концентрациях от приблизительно 50 пг/мл до приблизительно 50 мкг/мл. В одном из вариантов осуществления, FGF-7 используют в концентрации 50 нг/мл.
FGF-10 может использоваться в концентрациях от приблизительно 50 пг/мл до приблизительно 50 мкг/мл. В одном из вариантов осуществления, FGF-10 используют в концентрации 50 нг/мл.
Noggin может использоваться в концентрациях от приблизительно 500 нг/мл до приблизительно 500 мкг/мл. В одном из вариантов осуществления, Noggin используют в концентрации 100 нг/мл.
Нетрин 1 может использоваться в концентрациях от приблизительно 500 нг/мл до приблизительно 500 мкг/мл. В одном из вариантов осуществления, нетрин 1 используют в концентрации 100 нг/мл.
Нетрин 2 может использоваться в концентрациях от приблизительно 500 нг/мл до приблизительно 500 мкг/мл. В одном из вариантов осуществления, нетрин 2 используют в концентрации 100 нг/мл.
Нетрин 4 может использоваться в концентрациях от приблизительно 500 нг/мл до приблизительно 500 мкг/мл. В одном из вариантов осуществления, нетрин 4 используют в концентрации 100 нг/мл.
В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним из следующих факторов: ретиноевая кислота, FGF-2, FGF-4, FGF-7, FGF-10, циклопамин, Noggin, нетрин 1, нетрин 2 или нетрин 4.
В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-2, циклопамином и Noggin. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-4, циклопамином и Noggin. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-7, циклопамином и Noggin. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-10, циклопамином и Noggin.
В одном из вариантов осуществления клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-2, циклопамином и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-4, циклопамином и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-7, циклопамином и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-10, циклопамином и нетрином.
Нетрин выбирается из группы, включающей нетрин 1, нетрин 2 и нетрин 4.
В одном из вариантов осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-2, циклопамином, Noggin и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-4, циклопамином, Noggin и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-7, циклопамином, Noggin и нетрином. В альтернативном варианте осуществления, клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой, FGF-10, циклопамином, Noggin и нетрином.
Нетрин выбран из группы, состоящей из нетрина 1, нетрина 2 и нетрина 4.
Клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, могут быть обработаны по меньшей мере одним дополнительным фактором, который может улучшать образование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы. Как вариант, как минимум еще один дополнительный фактор может улучшить пролиферацию клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, полученные с помощью способов, составляющих предмет изобретения. Далее по меньшей мере еще один дополнительный фактор может улучшить способность клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, полученных с помощью способов, которые составляют предмет изобретения, образовывать другие типы клеток или повышать эффективность дополнительных стадий дифференцировки.
По меньшей мере одним дополнительным фактором могут быть, например, никотинамид, члены семейства TGF-β, включая TGF-β1, 2 и 3, сывороточный альбумин, члены семейства факторов роста фибробластов, тромбоцитарные факторы роста -AA, и -BB, плазма, богатая тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор роста и дифференцировки (GDF-5, -6, -8, -10, 11), глюкагонподобный пептид-I и II (GLP-I и II), миметические антитела GLP-1 и GLP-2, экзендин-4, ретиноевая кислота, паратиреоидный гормон, инсулин, прогестерон, апротинин, гидрокортизон, этаноламин, бета-меркаптоэтанол, эпидермальный фактор роста (EGF), гастрин I и II, хелатирующие агенты на основе меди, например, триэтиленпентамин, форсколин, Na-Butyrate, активин, бета-целлюлин, ITS, Noggin, фактор роста нейритов, Nodal, вальпроевая кислота, трихостатин A, натрий бутират, фактор роста гепатоцитов (HGF), сфингозин-1, VEGF, MG132 (EMD, CA), добавки N2 и B27 (Gibco, CA), стероидные алкалоиды, например, циклопамин (EMD, CA), фактор роста кератиноцитов (KGF), белки семейства Dickkopf, экстракт бычьего гипофиза, белок, ассоциированный с неогенезом островков (islet neogenesis-associated protein, INGAP), Indian Hedgehog, Sonic Hedgehog, протеосомальные ингибиторы, ингибиторы сигнального пути Notch, ингибиторы Sonic Hedgehog и их сочетания.
По меньшей мере один дополнительный фактор может поступать из кондиционированной среды, полученной от линий панкреатических клеток, таких как PANC-1 (ATCC № CRL-1469), CAPAN-1 (ATCC № HTB-79), BxPC-3 (ATCC № CRL-1687), HPAF-II (ATCC № CRL-1997), линий клеток печени, таких как HepG2 (ATCC № HTB-8065), линий клеток кишечника, таких как FHs 74 (ATCC № CCL-241).
Обнаружение клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы
Маркеры, характерные для линии панкреатической эндодермы, хорошо известны специалистам в данной сфере, и дополнительные маркеры, характерные для линии панкреатической эндодермы, продолжают выявляться. Данные маркеры могут использоваться для подтверждения того, что клетки, прошедшие обработку в соответствии с настоящим изобретением, дифференцировались и приобрели характерные особенности линии панкреатической эндодермы. К маркерам, специфическим для линии панкреатической эндодермы, относится экспрессия одного или нескольких факторов транскрипции, таких как, например, Hlxb9, PTF-1a, PDX-1, HNF-6, HNF-1beta.
Эффективность дифференцировки может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии панкреатической эндодермы.
Способы анализа экспрессии маркерных белков и нуклеиновых кислот в культивированных или изолированных клетках являются стандартными для данной области. Сюда относятся количественная ревертазная полимеразная цепная реакция (ОТ-ПЦР), Нозерн-блот, гибридизация in situ (смотрите, например, Current Protocols in Molecular Biology (Ausubel et al., eds. 2001 supplement)), и иммунологические методы, такие как иммуногистохимический анализ среза материала, Вестерн-блот, а для маркеров, доступных в интактных клетках, метод проточной цитометрии (FACS) (смотрите, например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998)).
Создание клеток, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы
Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, с использованием любого известного специалистам способа или с использованием любого способа, описанного в настоящем изобретении.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, могут быть дифференцированы в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, в соответствии со способами, описанными в публикации D'Amour et al., Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, путем культивирования клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей DAPT и экзендин-4, затем удаления среды, содержащей DAPT и экзендин-4 с последующим культивированием клеток в среде, содержащей экзендин-1, IGF-1 и HGF. Пример данного способа описан в публикации Nature Biotechnology 24, 1392-1401 (2006).
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы путем культивирования клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей экзендин-4, затем удаления среды, содержащей экзендин-4 с последующим культивированием клеток в среде, содержащей экзендин-1, IGF-1 и HGF. Пример данного способа описан в публикации D'Amour et al, Nature Biotechnology, 2006.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы путем культивирования клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей DAPT и экзендин-4. Пример данного способа описан в публикации D'Amour et al, Nature Biotechnology, 2006.
Например, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы путем культивирования клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в среде, содержащей экзендин-4. Пример данного способа описан в публикации D'Amour et al., Nature Biotechnology, 2006.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, в соответствии с способами, описанными в заявке на патент США серийный № 11/736908, принадлежащей LifeScan, Inc.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, в соответствии со способами, описанными в заявке на патент США серийный № 11/77311, принадлежащей LifeScan, Inc.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, в соответствии с способами, описанными в заявке на патент США серийный № 60/953178, принадлежащей LifeScan, Inc.
В одном аспекте настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, далее дифференцируются в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь TGF-βR-1. Фактором, ингибирующим сигнальный путь TGF-βR-1, может быть антагонист рецептора-1 TGF-β на поверхности клеток. Как вариант, этот фактор может ингибировать биологическую активность рецептора TGF-βR-1. Как вариант, этот фактор может ингибировать или являться антагонистом элемента пути сигнальной трансдукции TGF-βR-1 в клетке.
Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь TGF-βR-1 в течение периода от приблизительно одного до приблизительно двенадцати дней. Как вариант, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь TGF-βR-1 в течение периода от приблизительно пяти до приблизительно двенадцати дней. Как вариант, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь TGF-βR-1 в течение приблизительно двенадцати дней.
При использовании этого способа любая клетка, экспрессирующая маркеры, характерные для линии панкреатической эндодермы, может дифференцироваться в клетку, экспрессирующую маркеры, характерные для линии эндокринных панкреатических клеток.
В одном из вариантов осуществления настоящего изобретения предлагается способ дифференцировки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, включающий следующие стадии:
культивирование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, и
обработка клеток фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1.
Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь Notch и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от приблизительно одного до приблизительно двенадцати дней. Как вариант, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от приблизительно пяти до приблизительно двенадцати дней. Как вариант, клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатываются фактором, ингибирующим сигнальный путь Notch и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение приблизительно двенадцати дней.
При использовании этого способа любая клетка, экспрессирующая маркеры, характерные для линии панкреатической эндодермы, может дифференцироваться в клетку, экспрессирующую маркеры, характерные для линии эндокринных панкреатических клеток.
В одном из вариантов осуществления, фактор, ингибирующий сигнальный путь Notch, представляет собой ингибитор γ-секретазы. В одном из вариантов осуществления, ингибитор γ-секретазы представляет собой 1S-бензил-4R-[1-(1S-карбамоил-2-фенэтилкарбамоил)-1S-3-метилбутилкарбамоил]-2R-гидрози-5-фенилпентил] карбаминовой кислоты трет-бутиловый эфир, также называемый L-685458.
L-685458 может использоваться в концентрации от приблизительно 0,1 мкМ до приблизительно 100 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 90 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 80 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 70 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 60 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 50 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 40 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 30 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 20 мкМ. В одном из вариантов осуществления L-685458 используют в концентрации 10 мкМ.
В одном из вариантов осуществления фактор, ингибирующий сигнальный путь TGF-βR-1, представляет собой ингибитор TGF-βR-1-киназы. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы представляет собой (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин). В другой реализации ингибитор TGF-βR-1-киназы представляет собой [3-(пиридин-2-ил)-4-(4-хинонил)]-1H-пиразол.
Ингибитор TGF-βR-1-киназы может использоваться в концентрации от приблизительно 0,1 мкМ до приблизительно 100 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 90 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 80 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 70 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 60 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 50 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 40 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 30 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 20 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 10 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 1 мкМ. В одном из вариантов осуществления, ингибитор TGF-βR-1-киназы используют в концентрации приблизительно 0,1 мкМ.
Клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, могут быть обработаны по меньшей мере одним дополнительным фактором, который может улучшать образование клеток, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы. Как вариант, по меньшей мере еще один дополнительный фактор может улучшить пролиферацию клеток, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы, полученных с помощью способов, составляющих предмет настоящего изобретения. Далее по меньшей мере еще один дополнительный фактор может улучшить способность клеток, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы, полученных с помощью способов, которые составляют предмет настоящего изобретения, образовывать другие типы клеток или повышать эффективность дополнительных стадий дифференцировки.
По меньшей мере одним дополнительным фактором могут быть, например, никотинамид, члены семейства TGF-β, включая TGF-β1, 2 и 3, сывороточный альбумин, члены семейства факторов роста фибробластов, тромбоцитарные факторы роста -AA, и -BB, плазма, богатая тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор роста и дифференцировки (GDF-5, -6, -8, -10, 11), глюкагонподобный пептид-I и II (GLP-I и II), миметические антитела GLP-1 и GLP-2, экзендин-4, ретиноевая кислота, паратиреоидный гормон, инсулин, прогестерон, апротинин, гидрокортизон, этаноламин, бета-меркаптоэтанол, эпидермальный фактор роста (EGF), гастрин I и II, хелатирующие агенты на основе меди, например, триэтиленпентамин, форсколин, бутират натрия, активин, бетацеллюлин, ITS, Noggin, фактор роста нейритов, Nodal, вальпроевая кислота, трихостатин A, натрий бутират, фактор роста гепатоцитов (HGF), сфингозин-1, VEGF, MG132 (EMD, CA), добавки N2 и B27 (Gibco, CA), стероидные алкалоиды, например, циклопамин (EMD, CA), фактор роста кератиноцитов (KGF), белки семейства Dickkopf, экстракт бычьего гипофиза, белок, ассоциированный с неогенезом островков (islet neogenesis-associated protein, INGAP), Indian Hedgehog, Sonic Hedgehog, протеосомальные ингибиторы, ингибиторы сигнального пути Notch, ингибиторы Sonic Hedgehog и их сочетания.
По меньшей мере один дополнительный фактор может поступать из кондиционированной среды, полученной от линий панкреатических клеток, таких как PANC-1 (ATCC № CRL-1469), CAPAN-1 (ATCC № HTB-79), BxPC-3 (ATCC № CRL-1687), HPAF-II (ATCC № CRL-1997), линий клеток печени, таких как HepG2 (ATCC № HTB-8065), линий клеток кишечника, таких как FHs 74 (ATCC № CCL-241).
Обнаружение клеток, экспрессирующих маркеры, характерные для линии эндокринных клеток поджелудочной железы
Маркеры, характерные для линии эндокринных клеток поджелудочной железы, хорошо известны специалистам в данной сфере, и дополнительные маркеры, характерные для линии эндокринных клеток поджелудочной железы, продолжают выявляться. Данные маркеры могут использоваться для подтверждения того, что клетки, прошедшие обработку в соответствии с настоящим изобретением, дифференцировались и приобрели характерные особенности линии эндокринных клеток поджелудочной железы. К маркерам, специфическим для линии эндокринных клеток поджелудочной железы, относится экспрессия одного или нескольких факторов транскрипции, таких как, например, NGN-3, NeuroD, Islet-1.
Маркеры, характерные для линии β-клеток, хорошо известны специалистам в данной сфере, и дополнительные маркеры, характерные для линии β-клеток, продолжают выявляться. Эти маркеры могут использоваться для подтверждения того, что клетки, прошедшие обработку в соответствии с настоящим изобретением, дифференцировались и приобрели свойства, характерные для линии β-клеток. К специфическим особенностям линии β-клеток относится экспрессия одного или нескольких факторов транскрипции, в том числе, таких как Pdx1 (pancreatic and duodenal homeobox gene-1), Nkx2.2, Nkx6.1, Isl1, Pax6, Pax4, NeuroD, Hnf1b, Hnf-6, Hnf-3beta и MafA. Эти транскрипционные факторы хорошо известны в данной сфере в плане идентификации эндокринных клеток. Смотрите, например, Edlund (Nature Reviews Genetics 3: 524-632 (2002)).
Эффективность дифференцировки может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии эндокринных клеток поджелудочной железы. Как вариант, эффективность дифференцировки может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для линии β-клеток.
Способы анализа экспрессии маркерных белков и нуклеиновых кислот в культивированных или изолированных клетках являются стандартными для данной области. Сюда относятся количественная ревертазная полимеразная цепная реакция (ОТ-ПЦР), Нозерн-блот, гибридизация in situ (смотрите, например, Current Protocols in Molecular Biology (Ausubel et al., eds. 2001 supplement)), и иммунологические способы, такие как иммуногистохимический анализ среза материала, Вестерн-блот, а для маркеров, доступных в интактных клетках, способ проточной цитометрии (FACS) (смотрите, например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998)).
В одном аспекте настоящего изобретения эффективность дифференцировки определялась путем измерения процентной доли инсулин-положительных клеток в данной клеточной культуре после обработки. В одном из вариантов осуществления, способы настоящего изобретения давали приблизительно 100% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 90% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 80% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 70% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 60% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 50% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 40% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 30% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 20% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 10% инсулин-положительных клеток в данной культуре. В альтернативном варианте осуществления, способы настоящего изобретения давали приблизительно 5% инсулин-положительных клеток в данной культуре.
В одном аспекте настоящего изобретения эффективность дифференцировки определялась путем измерения стимулированной глюкозой секреции инсулина, определявшейся по количеству С-пептида, секретируемого клетками. В одном из вариантов осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 1000 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 900 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 800 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 700 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 600 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 500 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 400 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 500 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 400 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 300 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 200 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 100 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 90 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 80 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 70 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 60 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 50 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 40 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 30 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 20 нг C-пептида/пг ДНК. В альтернативном варианте осуществления, клетки, полученные способами настоящего изобретения, производили приблизительно 10 нг C-пептида/пг ДНК.
Способы лечения
В одном аспекте настоящего изобретения предлагается способ лечения пациентов, страдающих от диабета 1 типа или подвергающихся риску развития этого заболевания. Данный способ включает культивирование полипотентных стволовых клеток, дифференцировку полипотентных стволовых клеток in vitro в линию β-клеток и имплантирование клеток β-линии пациенту
В еще одном аспекте в настоящем изобретении предлагается способ лечения пациентов, страдающих от диабета 2 типа или подвергающихся риску развития этого заболевания. Данный способ включает культивирование полипотентных стволовых клеток, дифференцировку полипотентных культивированных стволовых клеток in vitro в линию β-клеток и имплантирование клеток β-линии пациенту.
При необходимости пациент далее может получать лечение фармакологическими агентами или биологически активными веществами, улучшающими выживаемость и функционирование трансплантированных клеток. К этим агентам могут относиться, например, в том числе, инсулин, члены семейства TGF-β, включая TGF-β1, 2 и 3, костные морфогенетические белки (BMP-2, -3, -4, -5, -6, -7, -11, -12 и -13), факторы роста фибробластов-1 и -2, тромбоцитарный фактор роста -AA и -BB, плазма, богатая тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор роста и дифференцировки (GDF-5, -6, -7, -8, -10, -15), фактор роста из клеток эндотелия сосудов (VEGF), плейотропин, эндотелин. К другим фармакологическим веществам могут относиться, например, никотинамид, глюкагонподобный пептид-I (GLP-1) и II, миметические антитела GLP-1 и 2, экзендин-4, ретиноевая кислота, паратиреоидный гормон, ингибиторы MAPK, такие, например, как описаны в опубликованной патентной заявке США 2004/0209901 и опубликованной патентной заявке США 2004/0132729.
Полипотентные стволовые клетки могут быть дифференцированы в инсулин-продуцирующие клетки до трансплантации реципиенту. В специфической реализации полипотентные стволовые клетки являются полностью дифференцированными в β-клетки до трансплантации реципиенту. Как вариант, полипотентные стволовые клетки могут быть трансплантированы реципиенту в недифференцированном или частично дифференцированном состоянии. Дальнейшая дифференцировка может происходить в организме реципиента.
Клетки сформировавшейся эндодермы, или, как вариант, клетки панкреатической эндодермы, или, как вариант, β-клетки могут имплантироваться в форме дисперсных клеток или клеток, образовавших кластеры, которые способом инфузии могут вводиться в воротную вену печени. Как вариант, клетки могут вводиться в биологически-совместимых разрушающихся полимерных вспомогательных материалах, в пористых неразрушающихся приспособлениях или в инкапсулированном виде для защиты от иммунного ответа организма-хозяина. Клетки могут имплантироваться в подходящее место организма реципиента. К местам для имплантации относятся, например, печень, естественная поджелудочная железа, пространство под почечной капсулой, сальник, брюшная полость, субсерозное пространство, кишечник, желудок или подкожный карман.
Для облегчения дальнейшей дифференцировки, выживания и активности имплантированных клеток, предварительно, одновременно или после имплантации клеток могут вводиться дополнительные факторы, такие как факторы роста, антиоксиданты или противовоспалительные агенты. В некоторых вариантах осуществления, факторы роста применяются для дифференцировки введенных клеток in vivo. Данные факторы могут секретироваться эндогенными клетками и воздействовать на внедренные клетки in situ. Дифференцировку имплантированных клеток можно индуцировать любым сочетанием эндогенных или введенных экзогенно факторов роста, известных специалистам.
Количество клеток, используемое при имплантации, зависит от ряда различных факторов, в том числе, от состояния пациента и его реакции на лечение, и оно может быть определено специалистом в данной сфере.
В одном аспекте в настоящем изобретении предлагается способ лечения пациентов, страдающих от диабета или подвергающихся риску развития этого заболевания. Данный способ включает культивирование полипотентных стволовых клеток, дифференцировку культивированных стволовых клеток in vitro в линию β-клеток и заключение клеток в опорный материал с трехмерной структурой. Клетки могут поддерживаться in vitro на этом опорном материале до имплантации пациенту. Как вариант, опорный материал, содержащий клетки, может напрямую имплантироваться в организм пациента без дополнительного культивирования in vitro. В опорный материал может быть заключен по меньшей мере один фармакологический агент, улучшающий выживание и функционирование трансплантированных клеток.
К опорным материалам, подходящим для применения для целей настоящего изобретения, относятся тканевые шаблоны, каналы, перегородки и резервуары, применяемые для лечения тканей. В частности, для практического применения способов настоящего изобретения подходят синтетические и природные материалы в форме пеноматериалов, губок, гелей, гидрогелей, тканных и нетканных структур, применяемых in vitro и in vivo для реконструкции и регенерации биологической ткани, а также для доставки хемотаксических агентов, индуцирующих рост ткани. Смотрите, например, материалы, описанные в патенте США 5770417, патенте США 6022743, патенте США 5567612, патенте США 5759830, патенте США 6626950, патенте США 6534084, патенте США 6306424, патенте США 6365149, патенте США 6599323, патенте США 6656488, патентной заявке США 2004/0062753 A1, патенте США 4557264 и патенте США 6333029.
Чтобы создать опорный материал с включенным фармакологическим агентом, фармакологический агент можно смешать с раствором полимера до изготовления опорной структуры. Как вариант, фармакологический агент можно нанести в виде покрытия на изготовленную опорную структуру, предпочтительно, в присутствии фармакологического носителя. Фармакологический агент может иметь форму жидкости, мелкодисперсного твердого вещества или любую другую подходящую физическую форму. Как вариант, в опорный материал могут быть добавлены инертные наполнители для изменения скорости высвобождения фармакологического агента. В альтернативном варианте осуществления, в опорный материал включается по меньшей мере одно фармакологическое соединение, являющееся противовоспалительным средством, как например, описанные в патенте США 6509369.
В опорный материал может включаться по меньшей мере одно фармакологическое соединение, являющееся антиапоптозным средством, как например, соединения, описанные в патенте США 6793945.
В опорный материал также может включаться по меньшей мере одно фармакологическое соединение, являющееся ингибитором фиброза, как например, соединения, описанные в патенте США 6331298.
В опорный материал также может включаться по меньшей мере одно фармакологическое соединение, являющееся стимулятором ангиогенеза, как например, соединения, описанные в Опубликованной патентной заявке США 2004/0220393 и в Опубликованной патентной заявке США 2004/0209901.
В опорный материал также может включаться по меньшей мере одно фармакологическое соединение, являющееся иммуносупрессором, как например, соединения, описанные в Опубликованной патентной заявке США 2004/0171623.
В опорный материал также может включаться по меньшей мере одно фармакологическое вещество, представляющее собой фактор роста, например, члены семейства TGF-β, включая TGF-β1, 2 и 3, костные морфогенетические белки (BMP-2, -3, -4, -5, -6, -7, -11, -12 и -13), факторы роста фибробластов-1 и -2, тромбоцитарный фактор роста -AA и -BB, плазма, богатая тромбоцитами, инсулиноподобный фактор роста (IGF-I, II), фактор роста и дифференцировки (GDF-5, -6, -8, -10, -15), фактор роста из клеток эндотелия сосудов (VEGF), плейотропин, эндотелин. К другим фармакологическим соединениям могут относиться, например, никотинамид, индуцированный гипоксией фактор 1-альфа, глюкагонподобный пептид-I (GLP-1), миметические антитела GLP-1 и GLP-2, и II, экзендин-4, Nodal, Noggin, NGF, ретиноевая кислота, паратиреоидный гормон, тенасцин-C, тропоэластин, пептиды тромбинового происхождения, кателицидины, дефензины, ламинин, биологические пептиды, содержащие связывающие клетки и гепарин домены протеинов адгезивного экстраклеточного матрикса, таких как фибронектин и витронектин, ингибиторы MAPK, такие как соединения, описанные в Опубликованной патентной заявке США 2004/0209901 и Опубликованной патентной заявке США 2004/0132729.
Включение клеток, составляющих предмет настоящего изобретения, в опорный каркас может осуществляться путем простого нанесения клеток на опорный каркас. Клетки могут входить внутрь каркаса путем простой диффузии (J. Pediatr. Surg. 23 (1 Pt 2): 3-9 (1988)). Было разработано несколько других подходов для повышения эффективности рассевания клеток. Например, для нанесения хондроцитов на каркас из полигликолевой кислоты используются центрифужные пробирки (Biotechnol. Prog. 14(2): 193-202 (1998)). Другой подход к нанесению клеток представляет собой использование центрифугирования, создающего минимальный стресс для рассеваемых клеток и повышающий эффективность нанесения. Например, в публикации Yang et al. разработан способ нанесения клеток (J. Biomed. Mater. Res. 55(3): 379-86 (2001)), названный центрифужной иммобилизацией клеток (Centrifugational Cell Immobilization, CCI).
Настоящее изобретение иллюстрируют, но не ограничивают, следующие примеры.
ПРИМЕРЫ
Пример 1
Культура человеческих эмбриональных стволовых клеток
Линии человеческих эмбриональных стволовых клеток H1, H7 и H9 были получены в WiCell Research Institute, Inc., (Madison, WI) и культивировались в соответствии с инструкциями, предоставленными указанным выше институтом. Вкратце, клетки культивировались на питающих клетках, представляющих собой мышиные эмбриональные фибробласты (МЭФ) в среде для эмбриональных стволовых клеток, содержащей DMEM/F12 (Invitrogen/GIBCO), с добавкой 20% заменителя сыворотки Knockout, 100 нМ раствора заменимых аминокислот MEM, 0,5 мМ бетамеркаптоэтанола, 2 нМ L-глутамина с 4 нг/мл человеческого основного фактора роста фибробластов (bFGF) (все производства Invitrogen/GIBCO). МЭФ-клетки из мышиных эмбрионов E13 -13.5, были приобретены в Charles River. МЭФ-клетки выращивались в среде DMEM с добавлением 10% эмбриональной телячьей сыворотки (ЭТС) (Hyclone), 2 мМ глутамина и 100 мМ заменимых аминокислот MEM. Субконфлюэнтные культуры МЭФ-клеток были обработаны раствором 10 мкг/мл митомицина C (Sigma, St. Louis, MO) в течение 3 ч для остановки деления клеток, затем обработаны трипсином и рассеяны по 2×104/см2 на покрытые 0,1% бычьим желатином чашки. В качестве слоя питающих клеток использовались МЭФ-клетки пассажей со второго по четвертый. Человеческие эмбриональные стволовые клетки, нанесенные на слой питающих МЭФ-клеток, культивировались при 37°C в атмосфере, содержащей 5% CO2 в инкубаторе с увлажнением для тканевых культур. По достижении конфлюэнтного роста (приблизительно, через 5-7 дней после нанесения), человеческие эмбриональные стволовые клетки обрабатывались раствором коллагенозы типа IV с концентрацией 1 мг/мл (Invitrogen/GIBCO) в течение 5-10 мин, а затем мягко счищались с поверхности при помощи пипетки на 5 мл. Клетки осаждались центрифугированием при 900 об/мин в течение 5 мин, осадок ресуспендировался и снова высевался с соотношением клеток от 1:3 до 1:4 в свежую культуральную среду.
Пример 2
Дифференцировка человеческих эмбриональных стволовых клеток до эндокринных клеток поджелудочной железы на субстрате тканевой культуры, покрытом MATRIGEL TM
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 45, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще трех дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN). Общая схема процедуры изображена на фиг.1, a. Образцы РНК отбирались из культуры на различных стадиях дифференцировки. На фиг.2 показаны данные анализа методом ПЦР в реальном времени, полученные на клетках, собранных на стадиях с 3 по 5. Видно достоверное увеличение экспрессии эндокринных маркеров, таких как инсулин и глюкагон, наблюдаемое в клетках стадий 4 и 5, в сочетании с увеличением экспрессии NeuroD.
Пример 3
Влияние различных соединений на экспрессию маркеров, характерных для линии панкреатических клеток, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, a
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 51 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN). Некоторые из культур были обработаны приведенными ниже соединениями в концентрации 1 мкм на стадии 3, стадии 4, или на стадиях 3+4: ингибитор MEK/MAPK (2′-амино-3′-метоксифлавон) (PD98059, Calbiochem, CA), ингибитор RAF-киназы (5-йодо-3-[(3,5-дибромо-4-гидроксифенил)метилен]-2-индолинон) (№553008, Calbiochem, CA), ингибитор SMAD3 (6,7-диметил-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-ил-проп-2-еноил))-1,2,3,4-тетрагидроизохинолин) (№ 566405, Calbiochem, CA), ингибитор AKT (1L6-гидроксиметил-хироинозитол-2-(R)-2-O-метил-3-O-октадецил-sn-глицерокарбонат) (№124005, Calbiochem, CA), ингибитор MEK (№444937 Calbiochem, CA), и ингибитор рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) (ингибирует активиновый рецептор-подобную киназу 5, № 616452, Calbiochem, CA).
На фиг.3, a-e показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадий с 3 по 5, обработанных в соответствии с указанными условиями. Следует обратить внимание, что добавление ингибитора киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) к клеткам на стадии 4 или на стадии 3 и 4 существенно увеличивало экспрессию инсулина, глюкагона, NeuroD и NKX2.2 и, при этом, незначительно влияло на экспрессию PDX-1 в конце стадии 5. Добавление ингибитора киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) к клеткам только на стадии 3 незначительно увеличивало экспрессию эндокринных маркеров в клетках в конце стадии 5.
Пример 4
Влияние добавления ингибитора киназы рецептора TGF-b I на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, a
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 44 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN). Некоторые культуры были обработаны ингибитором киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) в концентрации 1 мкм (ингибитор активиновый рецепторподобной киназы 5, № 616452, Calbiochem, CA) на стадии 4, на стадии 5 или на стадиях 4 и 5.
На фиг.4, a-e показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадий с 3 по 5, при наличии/отсутствии ингибитора киназы на стадиях 4-5. Следует обратить внимание, что добавление ингибитора киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) к клеткам на стадии 4 или на стадии 4 и 5 существенно увеличивало экспрессию инсулина, глюкагона, NeuroD и NKX2.2 и, при этом, незначительно влияло на экспрессию PDX-1 в конце стадии 5. Добавление ингибитора киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) к клеткам только на стадии 5 незначительно увеличивало экспрессию эндокринных маркеров в клетках в конце стадии 5.
Пример 5
Влияние различных ингибиторов киназы рецептора TGF-b I на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, a
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 41 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN). Некоторые культуры были обработаны ингибитором киназы рецептора TGF-B I (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин) в концентрации 1-10 мкм (ингибитор II ALK5) (ингибитор активиновый рецепторподобной киназы 5, № 616452, Calbiochem, CA) или ингибитором I TGF-B рецептора I (ингибитор I ALK5) ([3-(пиридин-2-ил)-4-(4-хинонил)]- 1H-пиразол) (№ 616451, Calbiochem, CA) на стадиях 4 и 5.
На фиг.5, a-e показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадий с 4-5, при наличии/отсутствии ингибитора ALK5 I или II. Следует обратить внимание, что добавление ингибитора ALK5 I или II в концентрации 1-10 мкм на стадиях 4 и 5 существенно увеличивало экспрессию инсулина, глюкагона, PDX-1 и NeuroD в конце стадий 4-5 в сравнении с контролем (стандартной обработкой). Все образцы анализировались трехкратно.
Пример 6
Влияние ингибиторов Noggin и ALK5 на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, a
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 41 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO)+0-500 нг/мл Noggin (R&D Systems, MN).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, Ca)+1 мкм ингибитора II ALK5 (Calbiochem, Ca) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN)+1 мкм ингибитора II ALK5.
На фиг. 6,a-g показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадий 3-5, при наличии/отсутствии 0-100 нг/мл Noggin. Добавление Noggin в концентрации 100 нг/мл на стадии 3 незначительно увеличивало экспрессию инсулина и глюкагона в конце стадии 4 и, при этом, существенно подавляло экспрессию альбумина и CDX2 в сравнении с необработанными образцами. Добавление Noggin в концентрации 500 нг/мл не влияло на экспрессию PDX-1, но существенно снижало экспрессию эндокринных маркеров, а также альбумина и CDX2. Альбумин является маркером клеток-предшественников печеночных клеток, тогда как CDX2 является маркером клеток кишечника.
Пример 7
Влияние добавления Noggin на стадии 3 и ингибиторов ALK5 на стадиях 4 и 5 на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 44 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO)+100 нг/мл Noggin (R&D Systems, MN).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, Ca)+1 мкм ингибитора II ALK5 (Calbiochem, Ca)+/-100 нг/мл Noggin в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN)+1 мкм ингибитора II ALK5.
На фиг.7, a-f показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадий 4-5, при наличии/отсутствии 100 нг/мл Noggin. Добавление 100 нг/мл Noggin на стадиях 3+4, в сочетании с добавлением ингибитора II ALK5, значительно повышало экспрессию инсулина и глюкагонов на стадиях 4-5. В частности, добавление Noggin на обеих стадиях 3 и 4 значительно повышало экспрессию NGN3, маркера предшественников эндокринных клеток и, при этом, существенно подавляло экспрессию альбумина и CDX2 по сравнению с необработанными образцами.
На фиг.8, a-d показано фазово-контрастное изображение культур, соответствующих приведенному выше протоколу. В некоторых культурах стадия 5 продлялась на срок от 3 до 21 дня. К 10-12 дню культивирования в среде стадии 5 по всей культуральной чашке наблюдались отдельные кластеры клеток, напоминающие островки у человека. На панелях a и b показаны изображения культур стадии 5 на 6 день, а на панелях показаны изображения культур стадии 5 на 12 день. Некоторые кластеры были вручную сняты, рассеяны на планшеты, покрытые 1:30 MATRIGEL, со средой, соответствующей стадии 5. После 2 дней инкубации кластеры окрашивались для определения инсулина и глюкагона. Большинство клеток в кластерах, как показано на фиг.9, a-b, являлись положительными в отношении продукции человеческого инсулина. Для дальнейшей оптимизации дозы ингибитора BMP, Noggin, культуры на стадиях 2-4 проходили обработку Noggin в концентрации 0, 10, 50 или 100 нг/мл. На фиг.10 показана экспрессия NGN3 на стадии 4 в культурах, которые были обработаны различными дозами Noggin на стадиях 2-4. Представляется, что концентрация 50-100 нг/мл Noggin приводит к максимальной экспрессии NGN3 на стадии 4.
Пример 8
Влияние добавления Noggin на стадиях 3-4, нетрина на стадии 4 и ингибиторов ALK5 на стадии 4 на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 51 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 2% БСА с жирными кислотами и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% БСА и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO)+100 нг/мл Noggin (R&D Systems, MN).
Далее клетки культивировались в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA)+1 мкм ингибитора II ALK5 (Calbiochem, Ca)+/-100 нг/мл Noggin+100 нг/мл нетрина-4 в течение трех дней.
На фиг.11, a-f показаны данные ПЦР в реальном времени, полученные на клетках, собранных в конце стадии 4, при наличии/отсутствии 100 нг/мл нетрина-4 на стадии 4. Добавление 100 нг/мл Noggin на стадии 3 и 4, в сочетании с добавлением ингибитора II ALK5 и 100 нг/мл нетрина-4 на стадии 4, существенно увеличивало экспрессию NGN3, NeuroD и Pax4 в клетках, собранных в конце стадии 5.
В некоторых культурах стадии 4 Noggin не использовался, тогда как добавлялся нетрин-4 в сочетании с ингибитором Alk5. На фиг.12, a-d показано, что отсутствие Noggin на стадии 4 существенно уменьшало экспрессию NGN3, NKX2.2 и NeuroD, и оказывало умеренное влияние на экспрессию инсулина и глюкагона в клетках, собранных в конце стадии 5.
Пример 9
Стимулируемая глюкозой секреция инсулина (GSIS) in vitro полипотентными стволовыми клетками, обработанными в соответствии с протоколом дифференцировки, приведенном на фиг.1, b
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 51 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 2% БСА с жирными кислотами и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% БСА и 100 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO)+100 нг/мл Noggin (R&D Systems, MN).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, Ca)+1 мкм ингибитора II ALK5 (Calbiochem, Ca)+/-100 нг/мл Noggin+100 нг/мл нетрин-4 в течение шести дней с последующей дополнительной инкубацией в течение 3-21 дней в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN)+1 мкм ингибитора II ALK5.
Клетки культуры стадии 5 в день 6, 8, 12 и 20 тестировались на GSIS путем отмывки в течение 1 ч в буфере Кребса (раствор Кребса-Рингера: 0,1% БСА, 10 мМ HEPES, 5 мМ NaHCO3, 129 мМ NaCl, 4,8 мМ KCl, 2,5 мМ CaCl2, 1,2 мМ KH2PO4, 1,2 мМ MgSO4, 5 мМ NaHCO3) при 37°C, с последующей инкубацией в течение 1 часа в буфере Кребса с добавлением 2 мМ D-глюкозы, и дополнительной инкубацией в течение 1 часа в буфере Кребса с добавлением 20 мМ D-глюкозы. После каждой инкубации супернатант отбирался и замораживался при -20°C для последующего анализа. Концентрации секретированного С-пептида анализировались с помощью сверхчувствительного твердофазного иммуноферментного анализа (ELISA) на С-пептид (Alpco Diagnostics, Sweden). На фиг.13, a-c показаны уровни секретированного человеческого С-пептида в ответ на стимуляцию глюкозой in vitro. Начальный и промежуточный период инкубации на стадии 5 не характеризовался высоким коэффициентом стимуляции (отношением количества секретированного C-пептида при высокой концентрации глюкозы к количеству секретированного С-пептида при низкой концентрации глюкозы), тогда как образцы, инкубированные в среде стадии 5 в течение 20 дней, демонстрировали высокую стимуляцию в ответ на глюкозу. Это показывает, что длительное воздействие среды стадии 5 приводит к созреванию кластеров, которые демонстрируют признаки GSIS.
Пример 10
Влияние добавления нетрина-1 или нетрина-2 на экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы, в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, b
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 44, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 44 нг/мл Activin-A (AA) в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO).
Далее клетки культивировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA) в течение шести дней с последующей дополнительной трехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+50 нг/мл IGF (Peprotech, NJ)+50 нг/мл HGF (R&D Systems, MN). К некоторым культурам на стадиях 2-5 добавлялось 50 нг/мл нетрина-1 или нетрина-2 (R&D Systems, MN). На фиг.14 показаны данные ПЦР в реальном времени, полученные на клетках, собранных на стадии 5, при наличии/отсутствии 50 нг/мл нетрина-1 нетрина-2 на стадиях со 2 по 5. Добавление и нетрина-1, и нетрина-2 значительно увеличивает экспрессию глюкагона и инсулина в клетках, собранных в конце стадии 5.
Пример 11
Альтернативный способ индукции экспрессии маркеров, характерных для линии сформировавшейся эндодермы
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 48, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, либо следующим путем:
культивирование в среде RPMI с добавлением 1% B27 (Invitrogen, Ca) и 50 нг/мл Activin-A (R&D Systems, MN), плюс 1 мМ Na Butyrate (Sigma, MO) в течение одного дня с последующей обработкой средой RPMI с добавлением 1% B27 (Invitrogen, Ca) и 50 нг/мл Activin-A (R&D Systems, MN), плюс 0,5 мМ Na Butyrate (Sigma, MO) в течение дополнительных трех дней, либо
в среде DMEM/F12 с добавлением 0,5% ЭТС и 100 нг/мл Activin-A (R&D Systems, MN) плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), в течение двух дней, с последующей обработкой средой DMEM/F12, с добавлением 2% ЭТС и 100 нг/мл Activin-A (AA) в течение еще двух дней.
Далее культуры обрабатывались DMEM/F12+2% ЭТС+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение трех дней, с последующей четырехдневной инкубацией в среде DMEM/F12+1% B27 (Invitrogen, CA)+20 нг/мл FGF7+0,25 мкм Cyclopamine-KAAD+2 мкм ретиноевой кислоты (RA) (Sigma, MO). Далее клетки инкубировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл Exendin-4 (Sigma, MO)+1 мкм DAPT (Calbiochem, CA)+1 мкм ингибитора II ALK5 (Calbiochem, Ca) в течение трех дней.
Индуцирование дифференцировки культур в сформировавшуяся эндодерму с использованием альтернативного способа а), приведенного выше, приводила к 78% экспрессии CXCR4, измеренной методом FACS, в сравнении с 56% экспрессией CXCR4 при использовании альтернативного способа b), приведенного выше. CXCR4 рассматривается как маркер клеток сформировавшейся эндодермы.
На фиг.15, a-f показаны данные ПЦР в реальном времени, полученные на клетках, собранных на стадии 5, при использовании альтернативного способа a) или b). Представляется, что альтернативный способ a) также может применяться для индуцирования экспрессии маркеров панкреатической эндодермы и линии эндокринных клеток.
Пример 12
Дифференцировка человеческих эмбриональных стволовых клеток, культивируемых в отсутствие сыворотки, на субстрате для тканевой культуры с покрытием MATRIGEL TM , в панкреатические эндокринные клетки
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 52, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и подвергались воздействию среды RPMI с добавлением 2% БСА (№ по каталогу 152401, MP Biomedical, Ohio) и 100 нг/мл Activin A (R&D Systems, MN), плюс 20 нг/мл WNT-3a (№ по каталогу 1324-WN-002, R&D Systems, MN), плюс 8 нг/мл bFGF (№ по каталогу 100-18B, PeproTech, NJ), в течение одного дня, с последующей обработкой средой RPMI с добавлением 2% БСА и 100 нг/мл Activin A, плюс 8 нг/мл bFGF в течение еще двух дней. Далее культуры обрабатывались DMEM/F12+2% БСА+50 нг/мл FGF7+0,25 мкМ Cyclopamine-KAAD (№ 239804, Calbiochem, CA) в течение двух дней, с последующей четырехдневной инкубацией в DMEM/F12+1% B27 (Invitrogen, CA)+50 нг/мл FGF7+0,25 мкМ Cyclopamine-KAAD+2 мкМ ретиноевой кислоты (RA) (Sigma, MO)+100 нг/мл Noggin (R&D Systems, MN). Далее клетки инкубировались в среде DMEM/F12+1% B27 (Invitrogen, CA)+100 нг/мл Noggin+1 мкМ DAPT (№ по каталогу 565784, Calbiochem, CA)+1 мкМ ингибитора II ALK5 (№ по каталогу 616452, Calbiochem, Ca)+100 нг/мл нетрина-4 (R&D Systems, MN) в течение трех дней, с последующей дополнительной инкубацией в течение семи дней в среде DMEM/F12+1% B27 (Invitrogen, CA)+1 мкМ ингибитора II ALK5 (Calbiochem, Ca). Для дальнейшего созревания эндокринных культур была добавлена последняя стадия, состоящая в семидневной обработке средой DMEM/F12+1% B27 (Invitrogen, CA). За исключением последней стадии, все прочие стадии были связаны с ежедневной сменой среды. Общая схема процедуры изображена на фиг.1, с. На каждой стадии количество клеток подсчитывалось с помощью гемоцитометра, и для ПЦР-анализа отбирались образцы РНК. Все образцы отбирались трехкратно.
На фиг.16, a-b показана экспрессия CXCR4, измеренная методом FACS, и данные ПЦР в реальном времени для стадии 1 день 3. Виден многократный рост экспрессии относительно недифференцированных эмбриональных стволовых клеток H1. На фиг.16, c-n показаны данные ПЦР в реальном времени для основных панкреатических и эндокринных маркеров в клетках, собранных на стадиях 2-6. На фиг.17 показано количество клеток в конце стадий 1-6.
Пример 13
Содержание C-пептида и проинсулина в полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, c
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 42, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в соответствии с протоколом, приведенным на фиг.1, с, как описано в приведенном выше примере 12.
Клетки отмывались в буфере Кребса-Рингера (129 мМ NaCl, 4,8 мМ KCl, 1,2 мМ NaH2PO4, 1,2 мМ MgSO4, 2,5 мМ CaCl2, 5 мМ NaHCO3, 10 мМ HEPES, 0,3% (масса/объем) БСА) и инкубировались в буфере Кребса в течение 15 минут с последующей инкубацией в буфере Кребса с добавлением 2 мМ D-глюкозы при 37°C, 5% CO2, 95% воздуха. Отбирался объем 0,5 мл супернатанта, оставшаяся среда удалялась и заменялась буфером Кребса с добавлением одного из следующих стимуляторов: 20 мМ D-глюкозы (HG), 30 мМ KCl, 100 мкМ толбутамида (tol), 100 мМ IBMX, 2 мкМ BAY K8644, или 10 мМ кетоизокапроновой кислоты (KIC) в течение 45 минут при 37°C, 5% CO2, 95% воздуха. Отбирался объем 0,5 мл супернатанта, а оставшаяся среда отбрасывалась. Супернатанты анализировались на содержание человеческого С-пептида с использованием комплекта для твердофазного иммуноферментного анализа ELISA на С-пептид (№ по каталогу 80-CPTHU-E01, Alpco Diagnostics, NH). Образцы стандартно разводились в 5-10 раз, чтобы обеспечить соответствие линейному диапазону стандартов, входящих в комплект.
На фиг.18 показана секреция С-пептида после стимуляции различными стимулами. На диаграмме для каждого агента также показан коэффициент стимуляции (частное от деления концентрации C-пептида в супернатанте после стимуляции на концентрацию С-пептида в основном супернатанте, содержащем 2 мМ глюкозы).
Для измерения содержания C-пептида, проинсулина и ДНК в культурах шестой стадии, 6-луночные планшеты, содержащие культуры стадии 6, обрабатывались 500 мкл лизирующего клетки буфера (10 мМ Tris-HCL+1 мМ EDTA, pH 7,5) с последующей ультразвуковой обработкой в течение 30 секунд. Содержание C-пептида и проинсулина измерялись с использованием комплектов для твердофазного иммуноферментного анализа (ELISA) на С-пептид (№ по каталогу 80-CPTHU-E01, Alpco Diagnostics, NH) и проинсулин (№ по каталогу 11-1118-01, Alpco Diagnostics, NH), соответственно. Количественный анализ содержания ДНК в лизатах проводился с помощью комплекта Quant-IT™ для ДНК (№ по каталогу P7589, Invitrogen, Ca). Образцы стандартно разводились в 500-1000 раз, чтобы обеспечить соответствие линейному диапазону стандартов, входящих в комплект. На фиг.19 показано содержание C-пептида и проинсулина, нормализованное к ДНК, для трех разных лунок 6-луночного планшета, содержащей культуры стадии 6. Для сравнения, также было измерено содержание C-пептида и проинсулина в трех образцах человеческих трупных островков поджелудочной железы. Следует учитывать, что данные отражают содержание инсулина во всей культуре, а не только в кластерах, присутствующих в культуре.
Пример 14
Анализ методом FASC полипотентных стволовых клетках, обработанных в соответствии с протоколом дифференцировки, приведенным на фиг.1, c
Клетки линии человеческих эмбриональных стволовых клеток H1 культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в соответствии с протоколом, приведенным на фиг.1, с, как описано в приведенном выше примере 12. Клетки подвергались диссоциации из монослойных культур на одиночные клетки с использованием TrypLE Express (Invitrogen, Carlsbad, CA) и отмывались в холодном PBS. Для фиксации клетки ресуспендировались в 200-300 мкл буфера Cytofix/Cytoperm (BD 554722, BD, Ca) и инкубировались в течение 30 мин при 4°C. Клетки дважды отмывались в 1 мл буферного раствора Perm/Wash (BD 554723) и ресуспендировались в 100 мкл раствора для окраски/блокировки, содержащем 2% нормальной козьей сыворотки в буфере Perm/Wash. Для анализа методом проточной цитометрии клетки окрашивались следующими первичными антителами: антиинсулиновые (кроличьи мАт, Cell Signaling № C27C9; разведение 1:100), антиглюкагоновые (мышиные мАт, Sigma № G2654, 1:100); антисинаптофизиновые (кроличьи поликлональные антитела, DakoCytomation № A0010, 1:50); Клетки инкубировались в течение 30 мин при 4°C с последующей двукратной отмывкой в буфере Perm/Wash и дальнейшей инкубацией в течение 30 мин в следующих подходящих вторичных антителах: козьи антикроличьи Alexa 647 (Invitrogen № A21246) или козьи антимышиные 647 (Invitrogen № A21235); козьи антикроличьи R-PE (BioSource № ALI4407). Все вторичные антитела применялись в разведении 1:200. Клетки отмывались по меньшей мере один раз в буфере Perm/Wash и анализировались с применением BD FACSArray. Для анализа фиксировалось не менее 10000 событий. В качестве контроля использовались недифференцированные клетки H1 и клеточная линия b-TC (ATCC, VA). На фиг.20, a-c показана процентная доля инсулин-положительных и синаптофизин-положительных клеток в культурах стадии 6.
Пример 15
Добавление хордина на стадиях 3 и 4 протокола дифференцировки, указанного на фиг.1, c, повышает экспрессию маркеров, характерных для линии эндокринных клеток поджелудочной железы
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 52, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в соответствии с протоколом, приведенным на фиг.1, с, за исключением того, что вместо Noggin на стадиях 3-4 добавлялось 50-100 нг/мл хордина (R&D Systems, MN). Как и Noggin, также является известным ингибитором сигнального пути BMP. Анализ культур стадии 6 методом FACS (фиг.21, a-b) выявил экспрессию инсулина и синаптофизина, очень сходную с экспрессией, наблюдавшейся при добавлении Noggin на стадиях 3-4.
Пример 16
Дифференцировка человеческих эмбриональных стволовых клеток, культивируемых в отсутствие сыворотки, на субстрате для тканевой культуры с покрытием MATRIGEL TM , в панкреатические эндокринные клетки
Клетки линии человеческих эмбриональных стволовых клеток H9, пассаж 39, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в соответствии с протоколом, приведенным на фиг.1, с, как описано в приведенном выше примере 12. РНК отбиралась в конце стадий 1-6. На фиг.22, a-l показаны данные ПЦР в реальном времени для основных панкреатических и эндокринных маркеров в клетках, собранных в конце стадий 3-6. По аналогии с результатами, полученными на линии H1, клетки H9 были способны экспрессировать маркеры, характерные для линии эндокринных клеток поджелудочной железы.
Пример 17
Влияние добавок для культуральной среды на способность полипотентных стволовых клеток дифференцироваться в панкреатические эндокринные клетки
Клетки линии человеческих эмбриональных стволовых клеток H1, пассаж 39, культивировались на чашках с покрытием MATRIGEL™ (разведение 1:30) и дифференцировались в соответствии с протоколом, приведенным на фиг.1, с, за исключением того, что в некоторых культурах в базовую среду DMEM/F12 добавлялись 0,25-1% B27 или 1% N2 (Invitrogen, Ca)+2% БСА. Все прочие компоненты протокола дифференцировки сохранялись такими же, как описано в примере 12. Образцы отбирались трехкратно в конце стадий 3, 5 и 6, и анализировались методом ПЦР в реальном времени. На фиг.23, a-d показаны данные ПЦР в реальном времени для основных панкреатических и эндокринных маркеров в клетках, собранных в конце стадий 3, 5 и 6. Использование N2/БСА вместо B27 давало очень сходную экспрессию ключевых маркеров эндокринных клеток поджелудочной железы. Более того, концентрацию B27 можно было снизить до 0,25% без существенного влияния на экспрессию ключевых маркеров эндокринных клеток поджелудочной железы.
Публикации, цитируемые в данном документе, полностью включаются в настоящий документ в форме ссылок. Хотя различные аспекты изобретения иллюстрируются выше ссылками на примеры и предпочтительные варианты осуществления, подразумевается, что область изобретения ограничивается не упомянутым выше описанием, а следующими пунктами формулы изобретения, составленными в соответствии с принципами патентного законодательства.
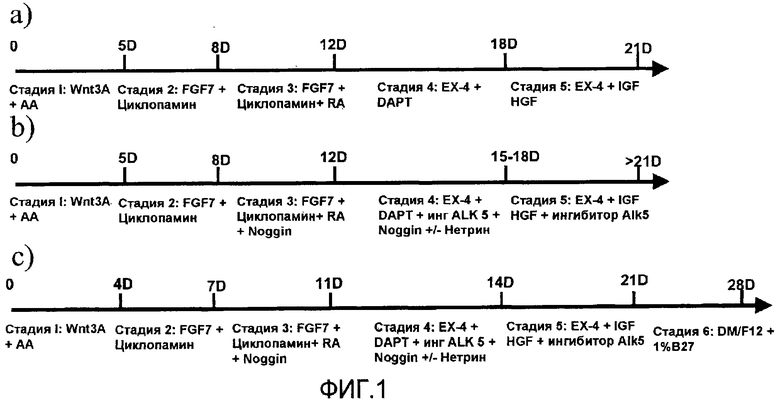
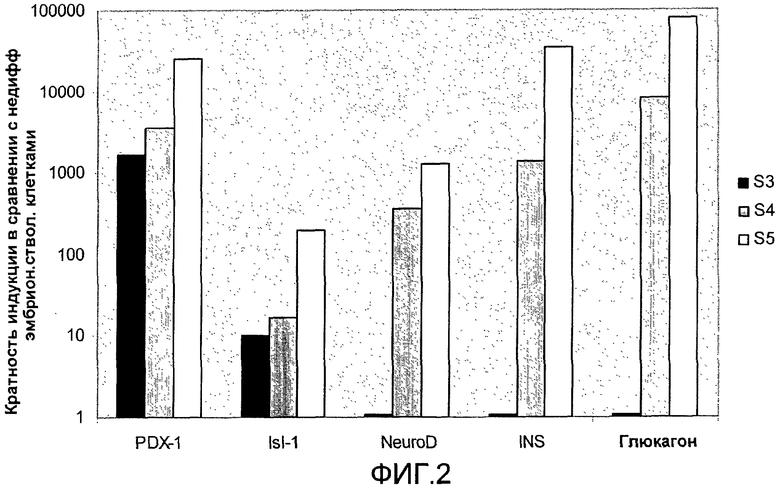
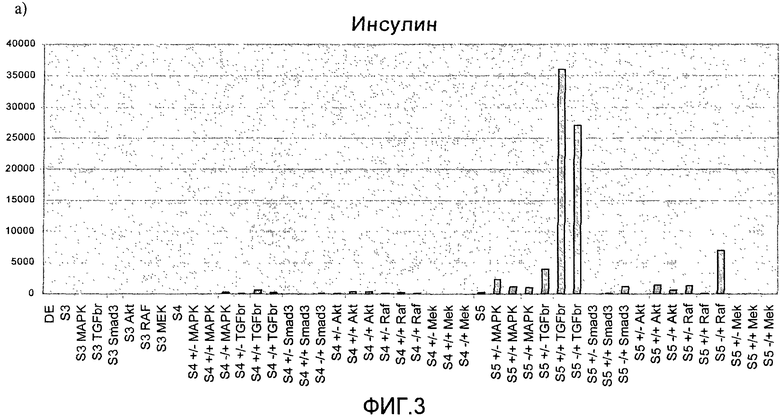
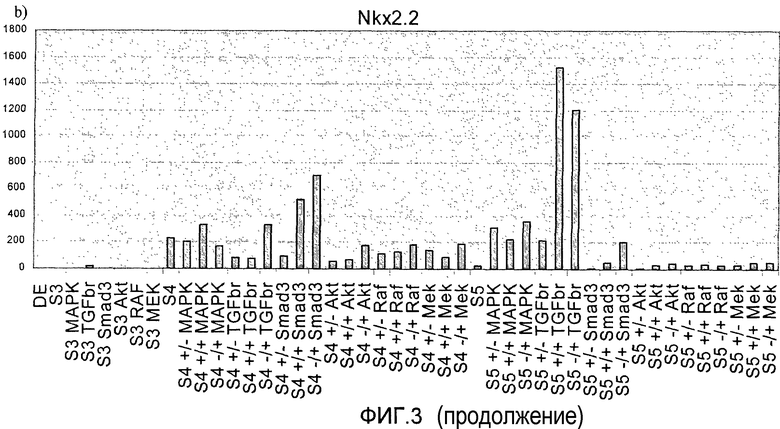
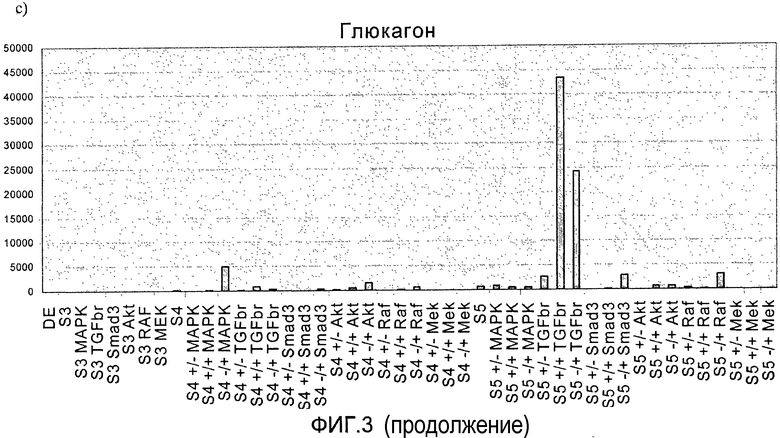
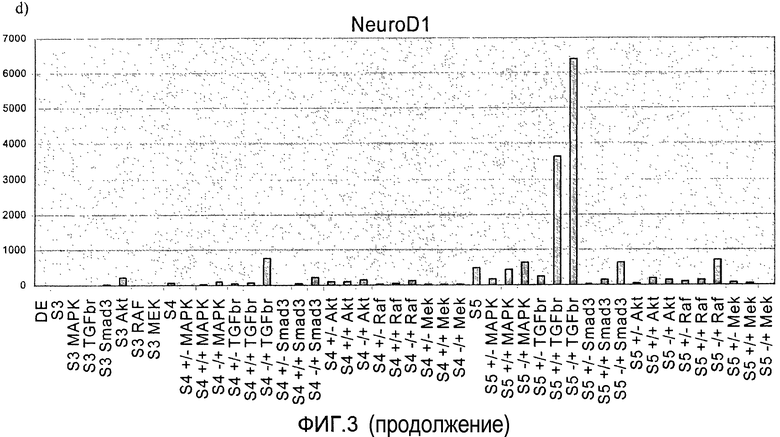
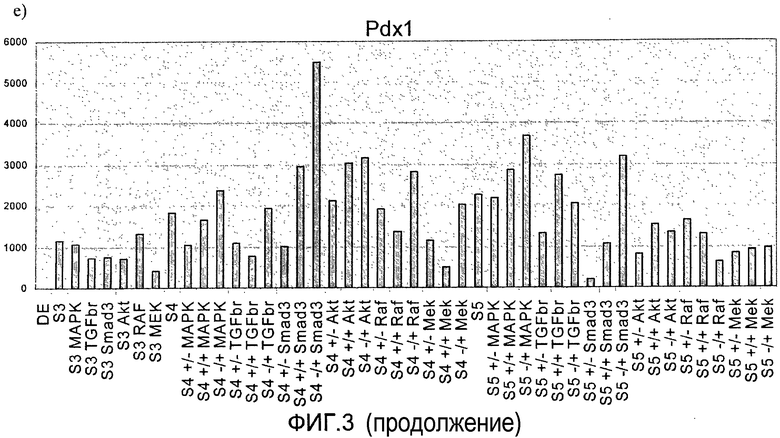
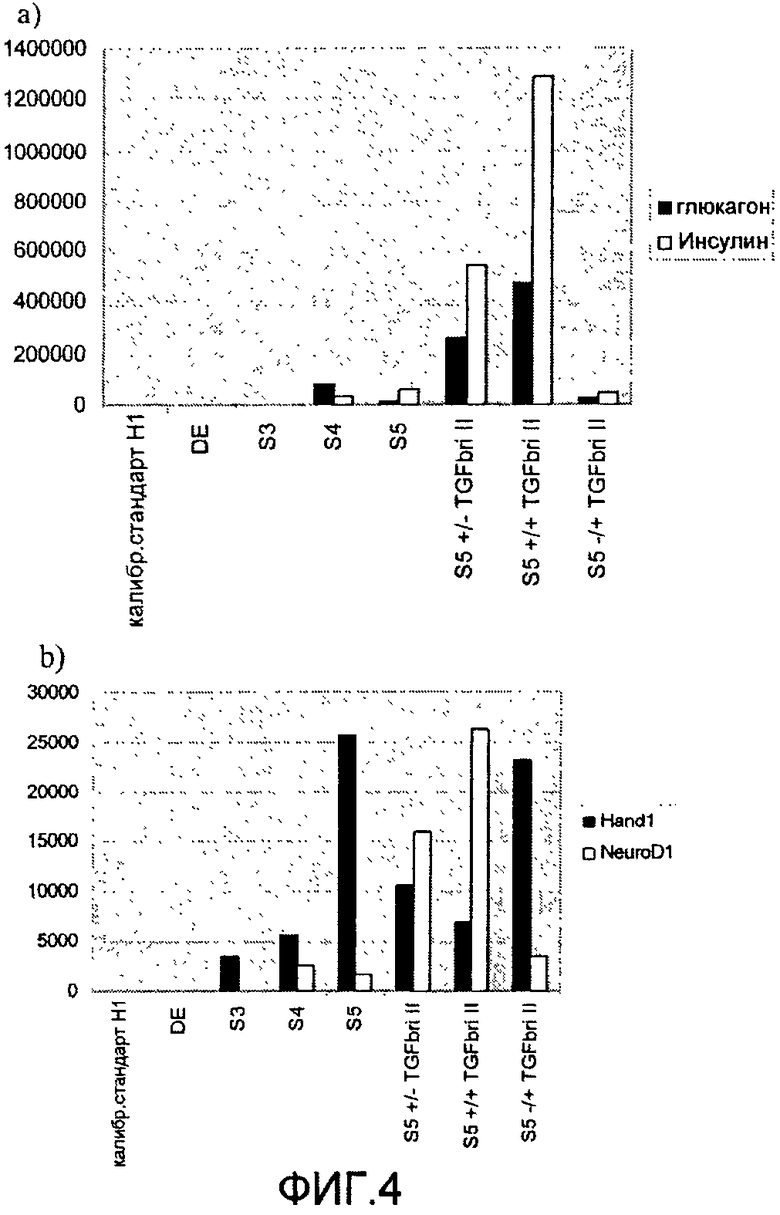
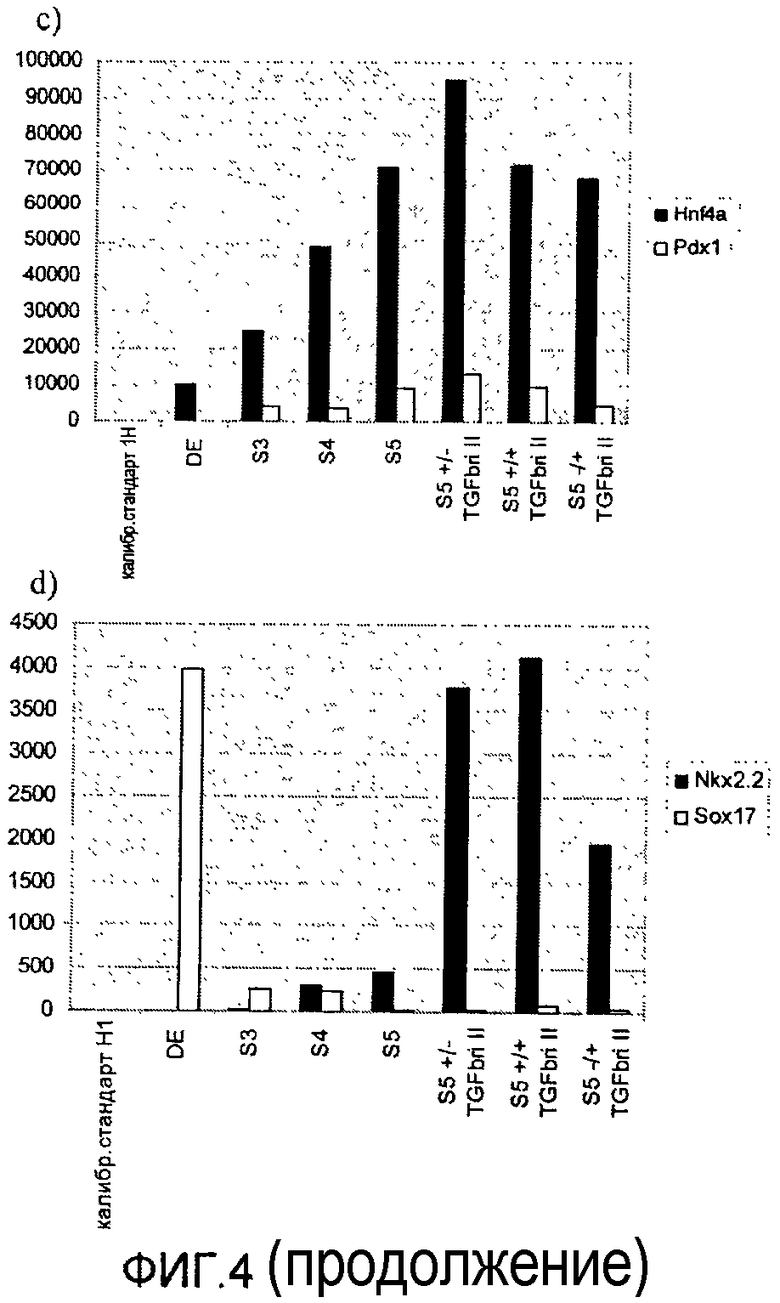
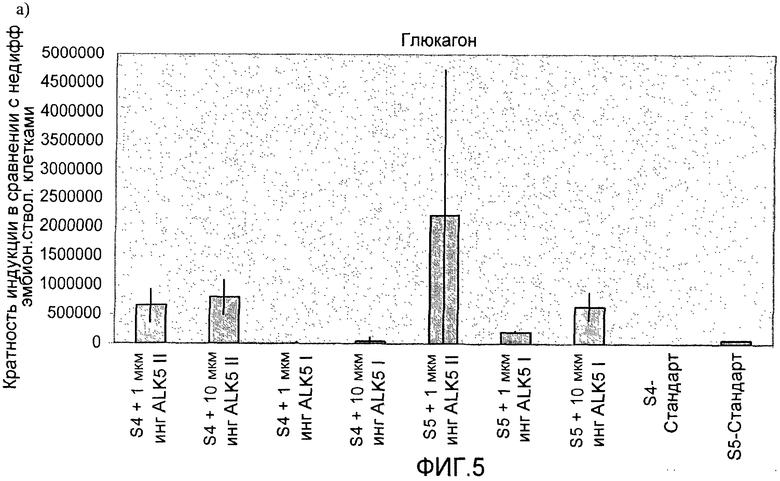
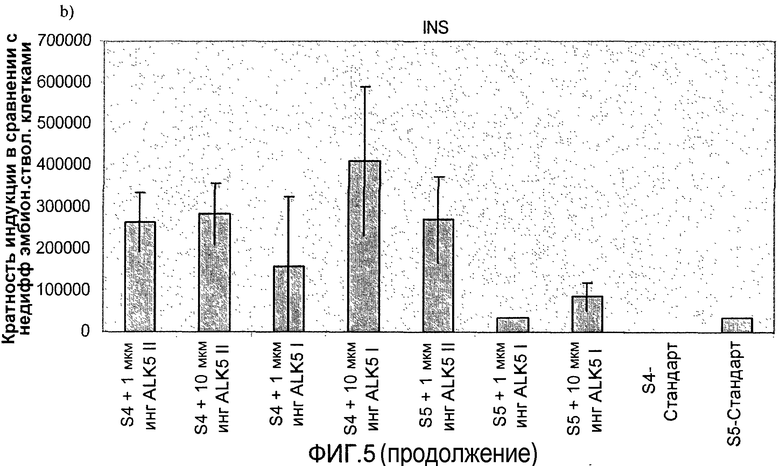
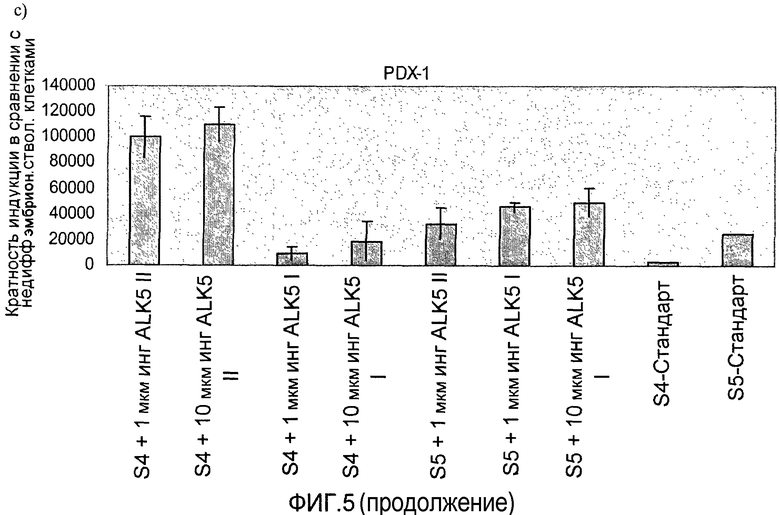
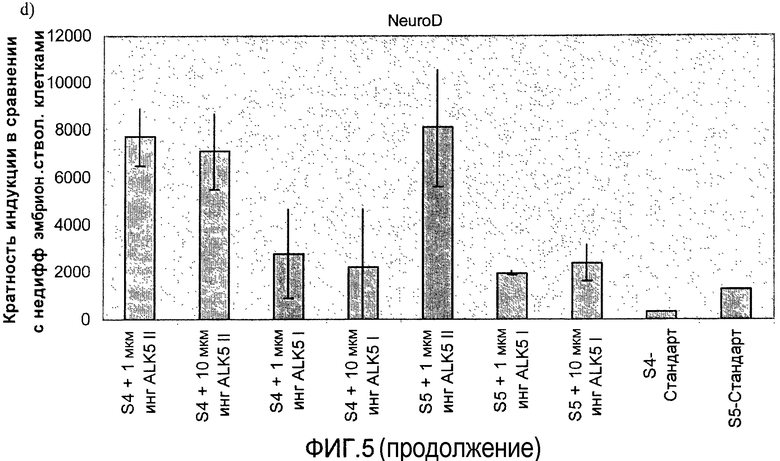
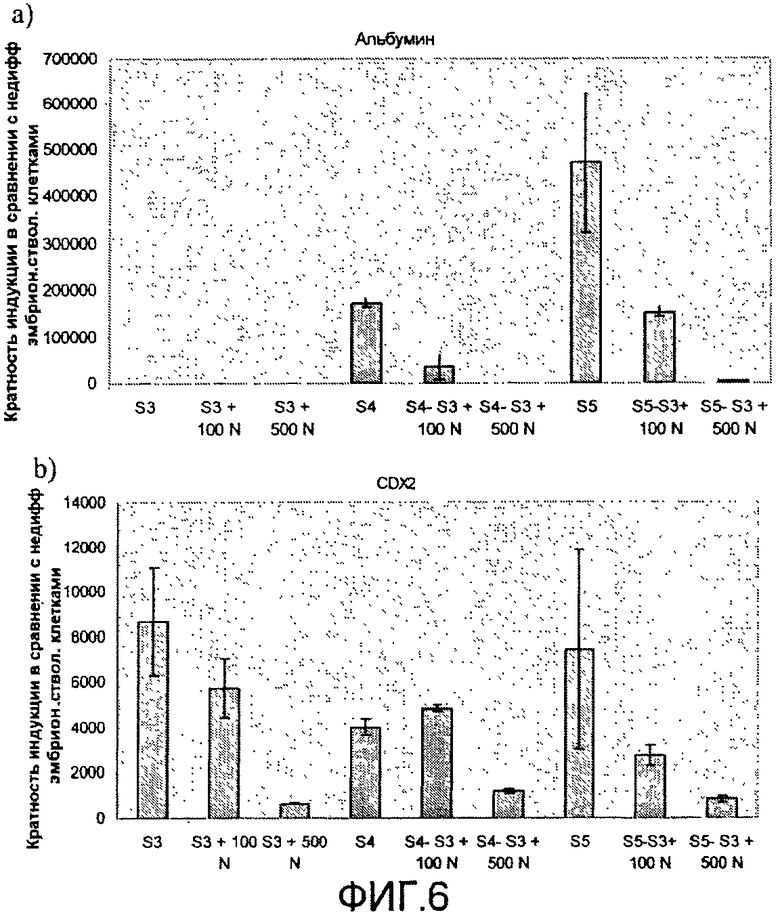
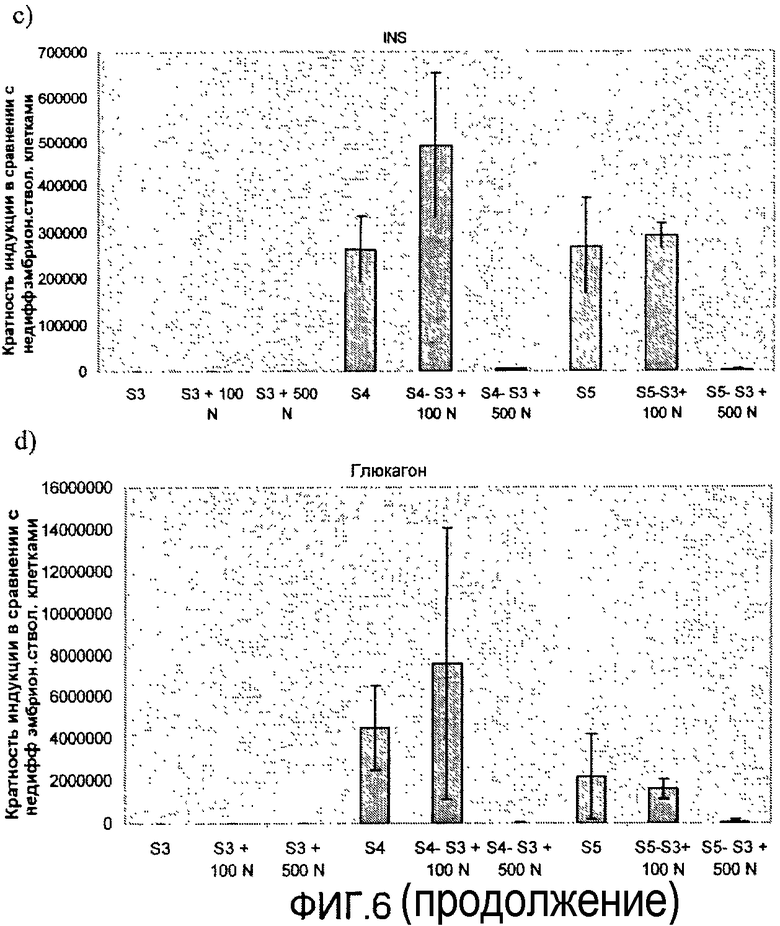
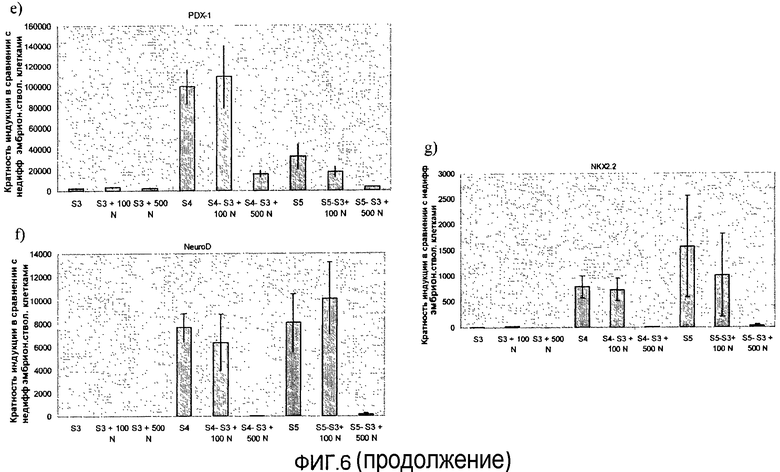
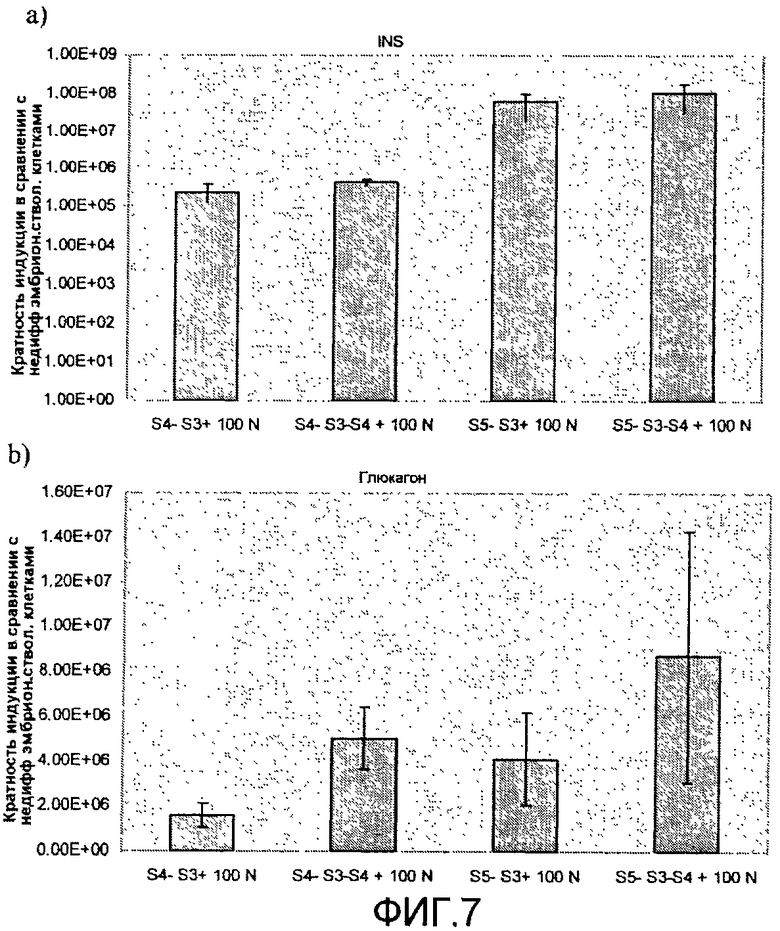
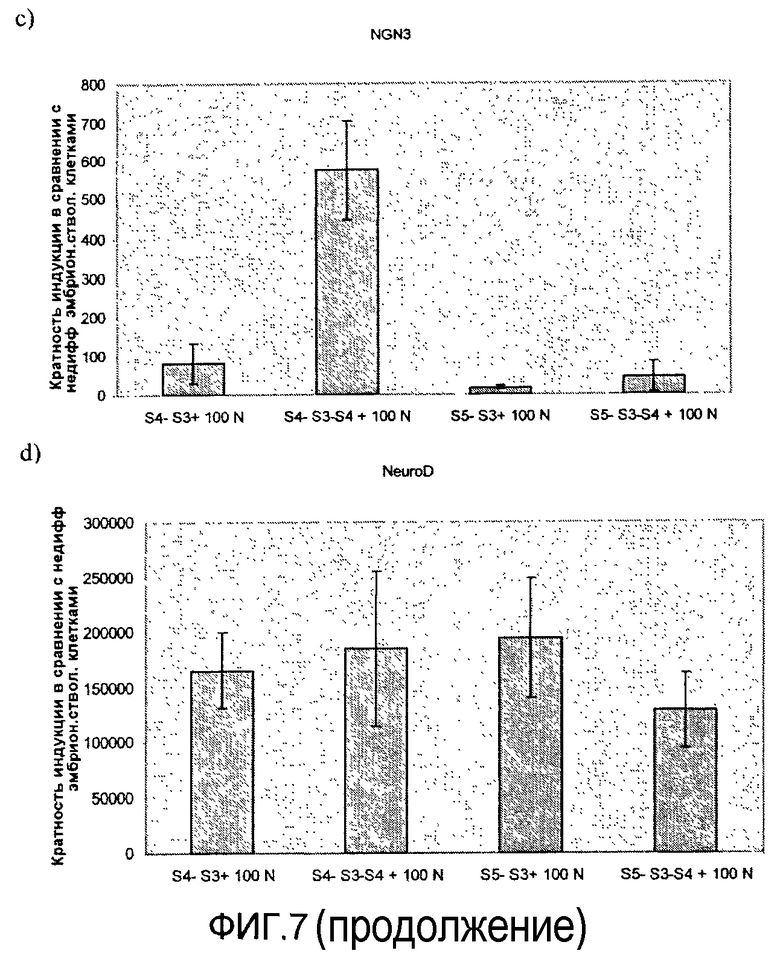

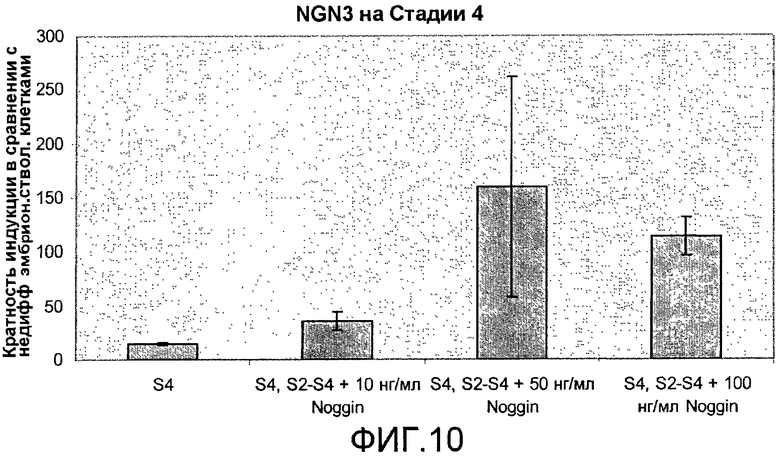
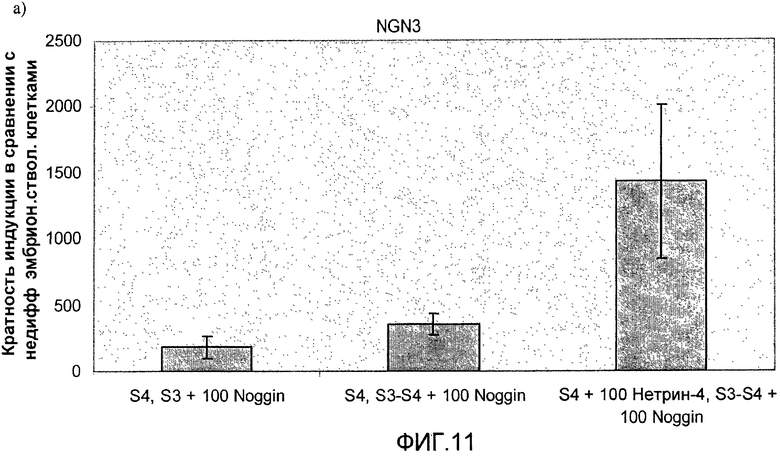
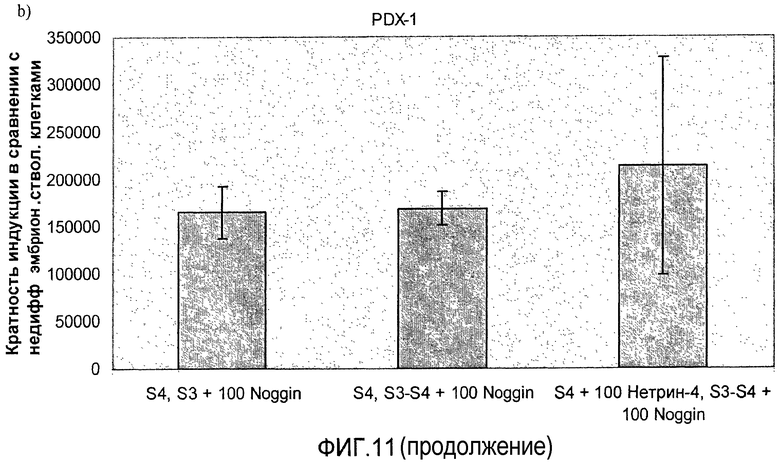
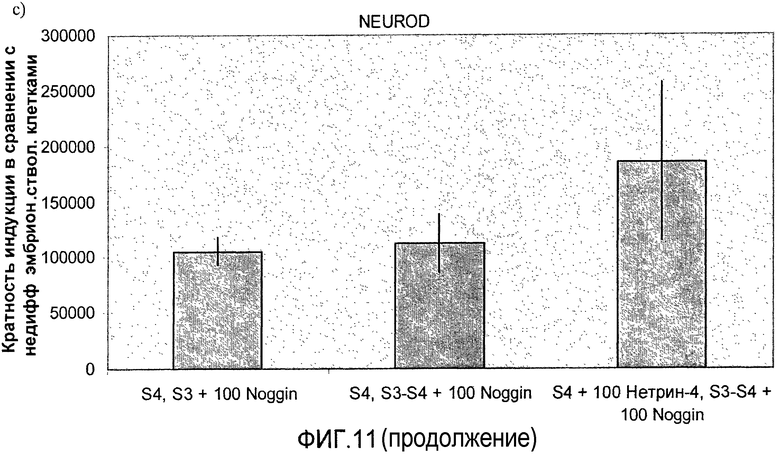
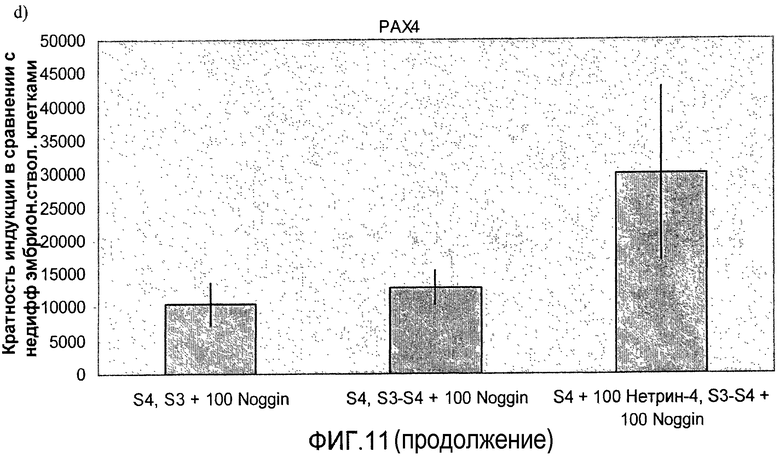
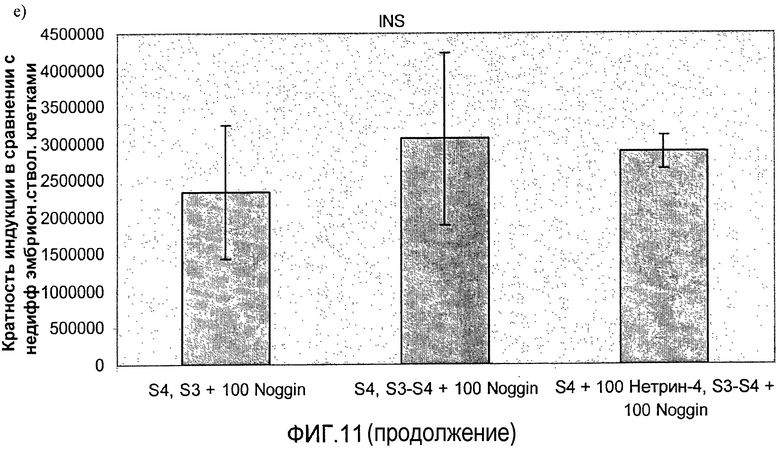
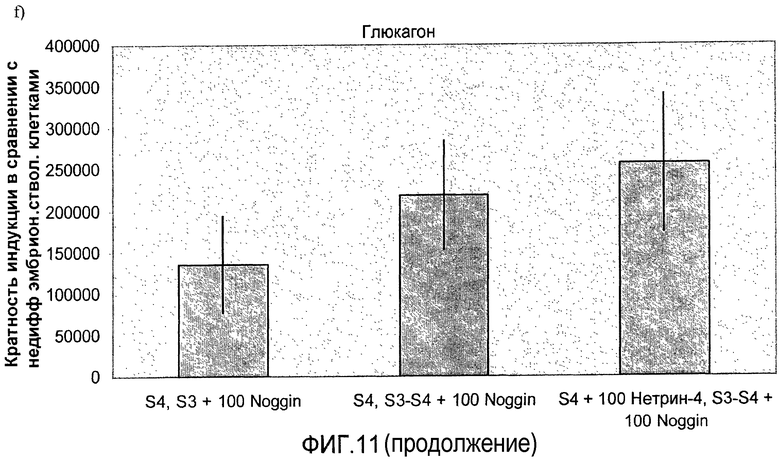
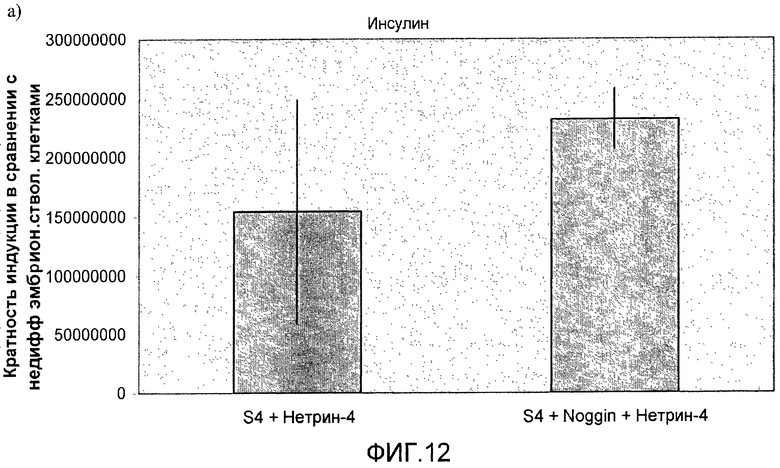
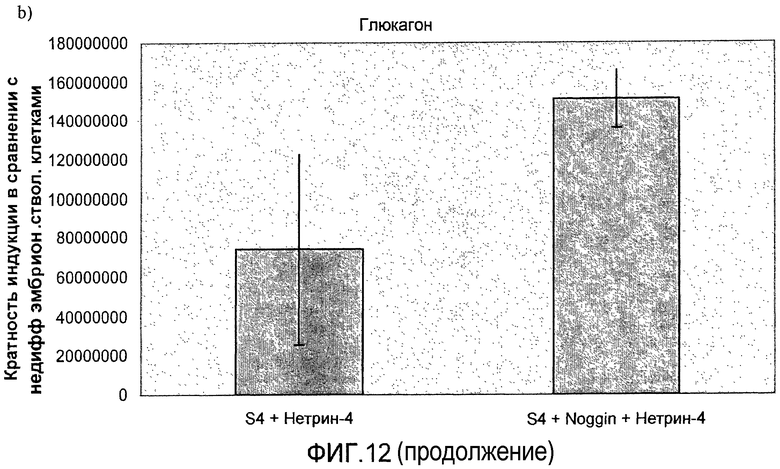
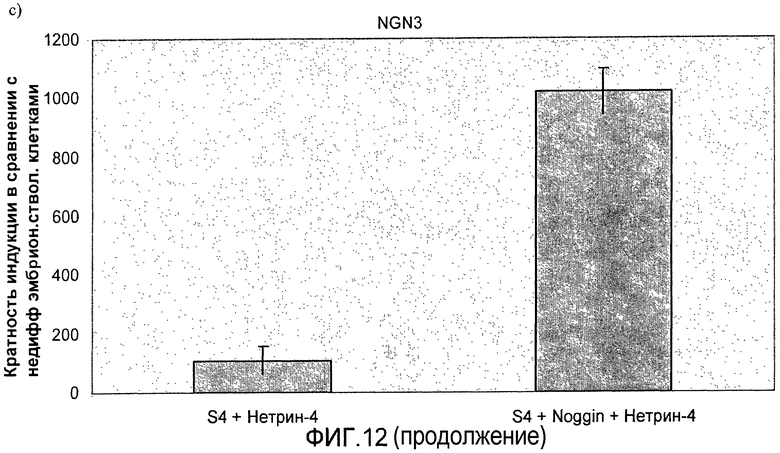
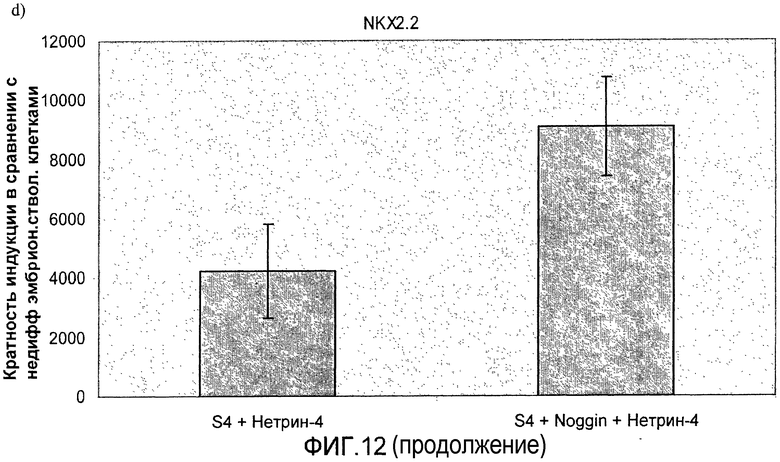
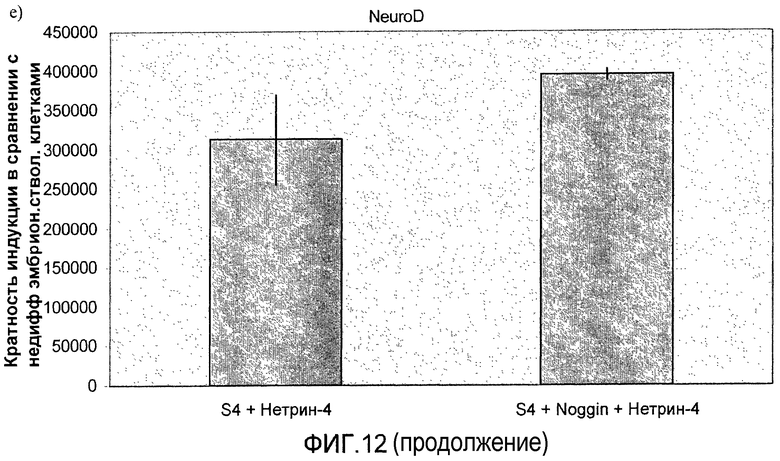
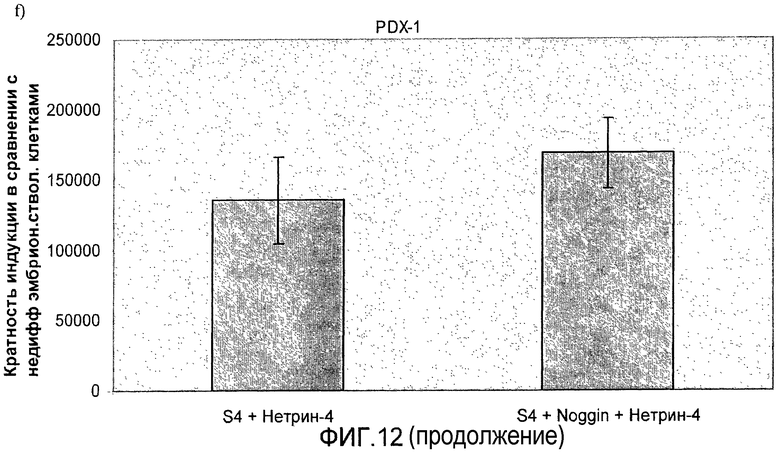
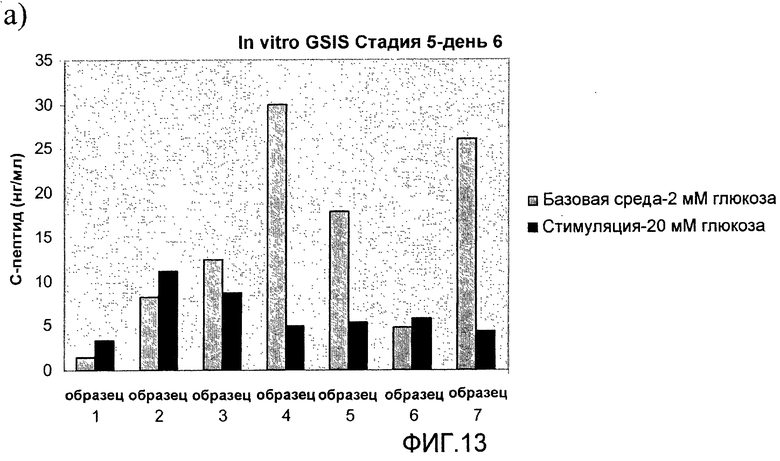
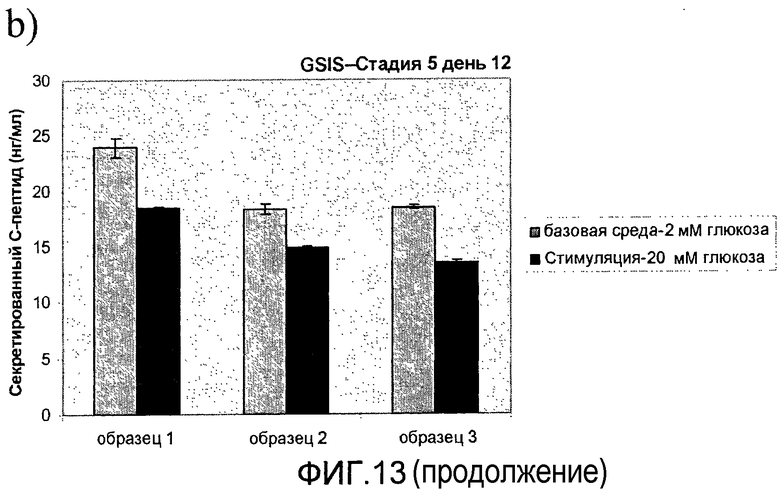
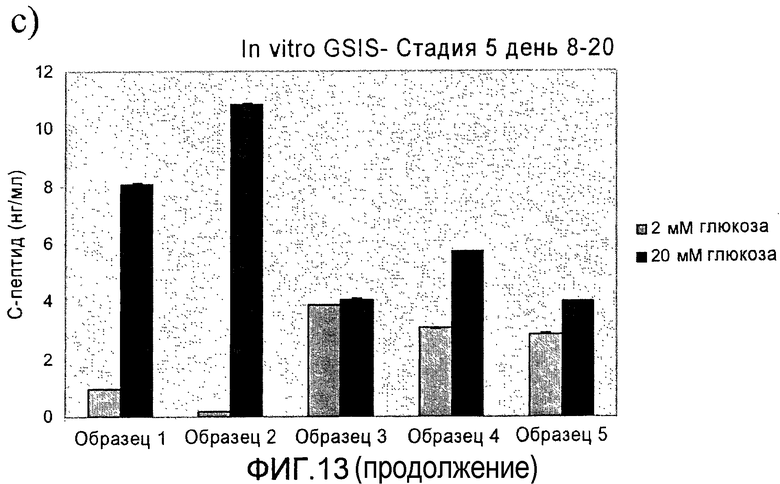
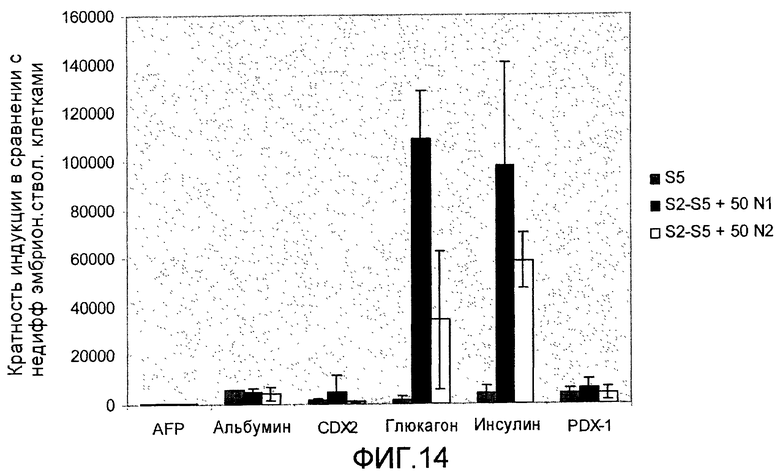
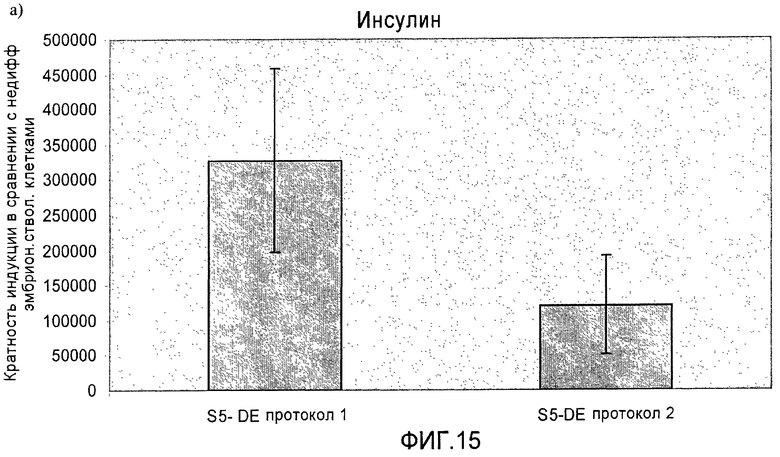
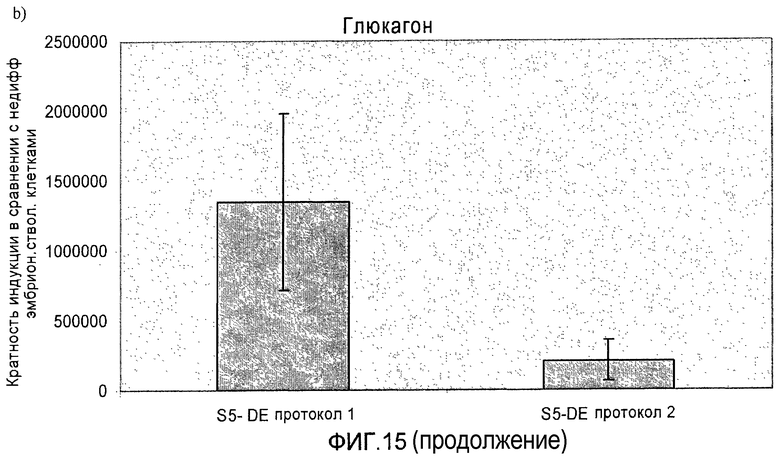
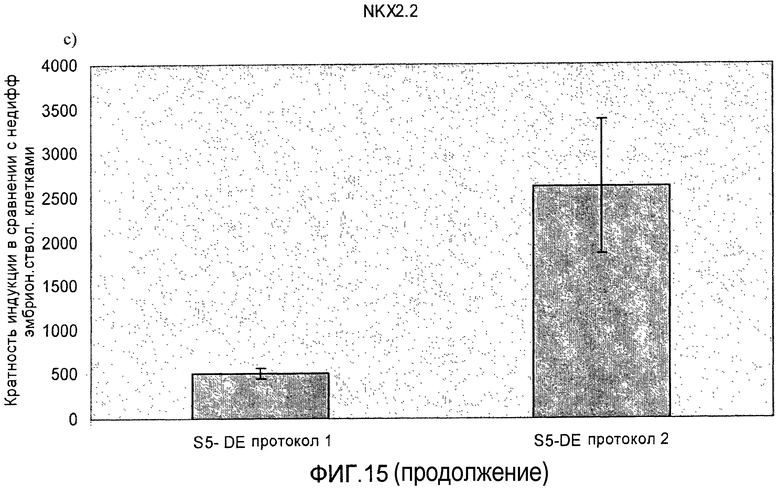
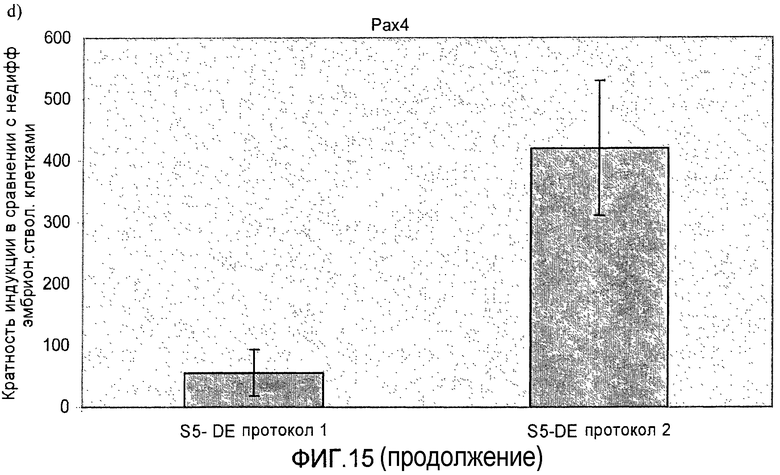
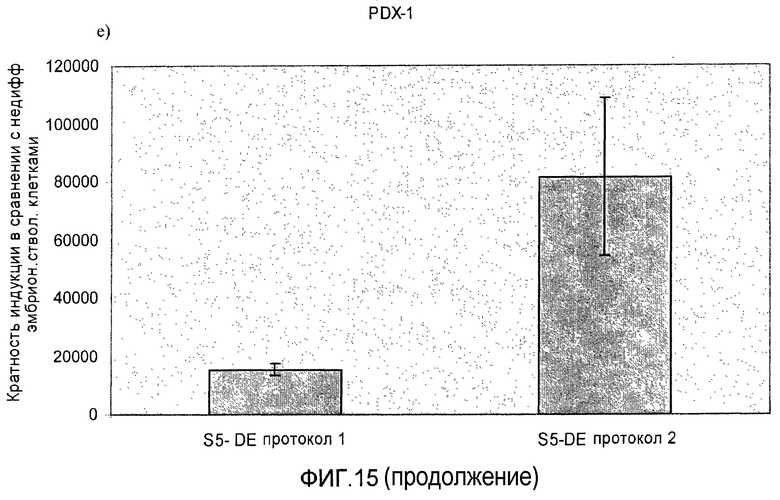
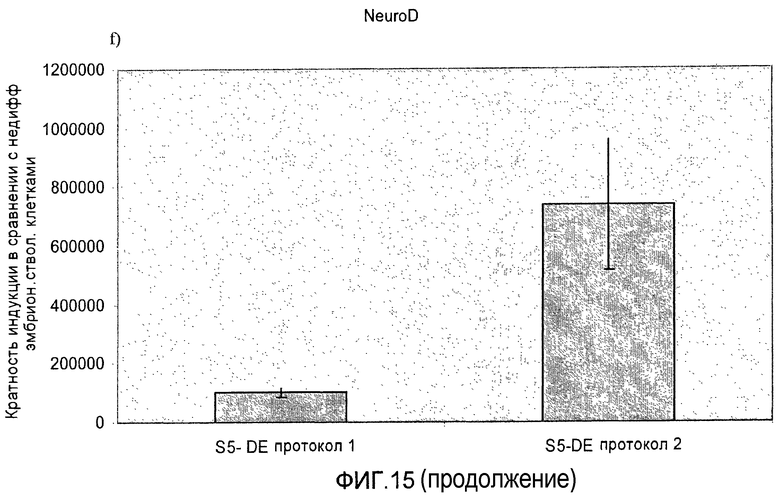
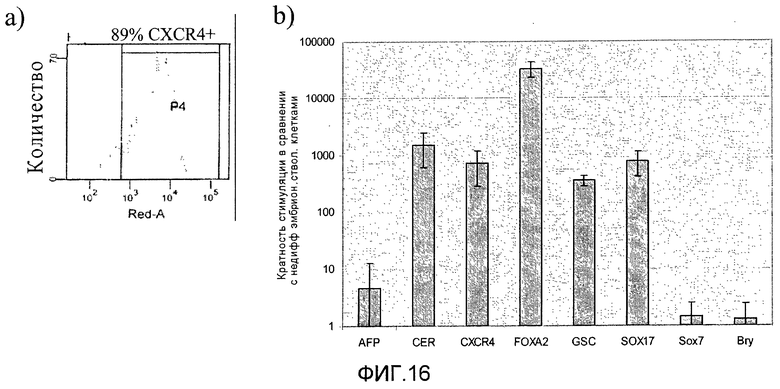
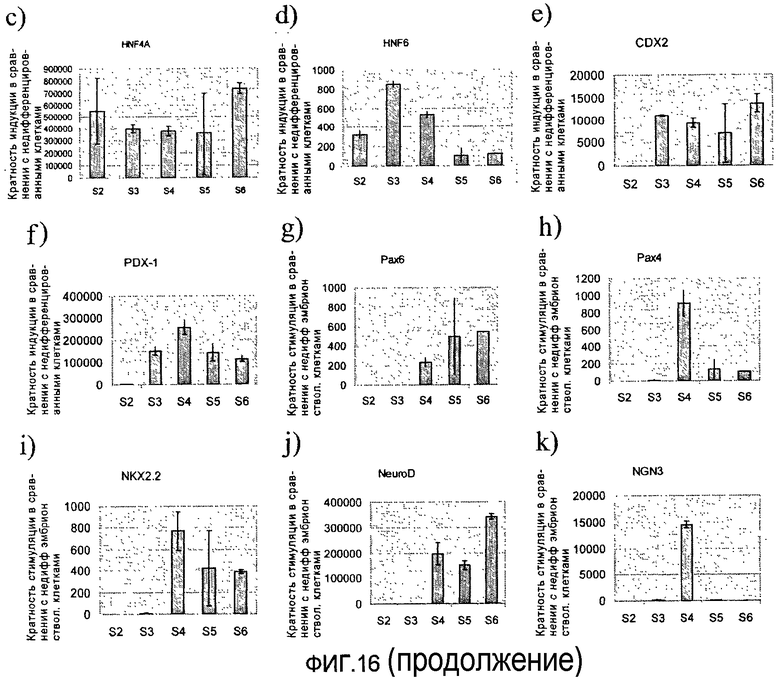
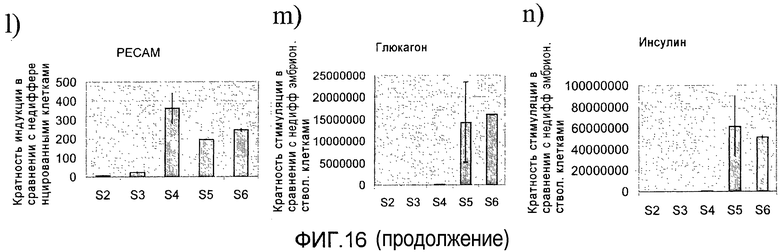
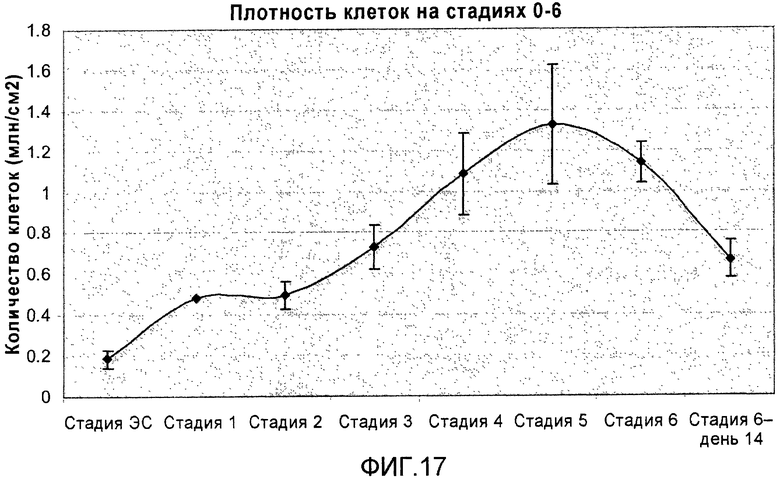
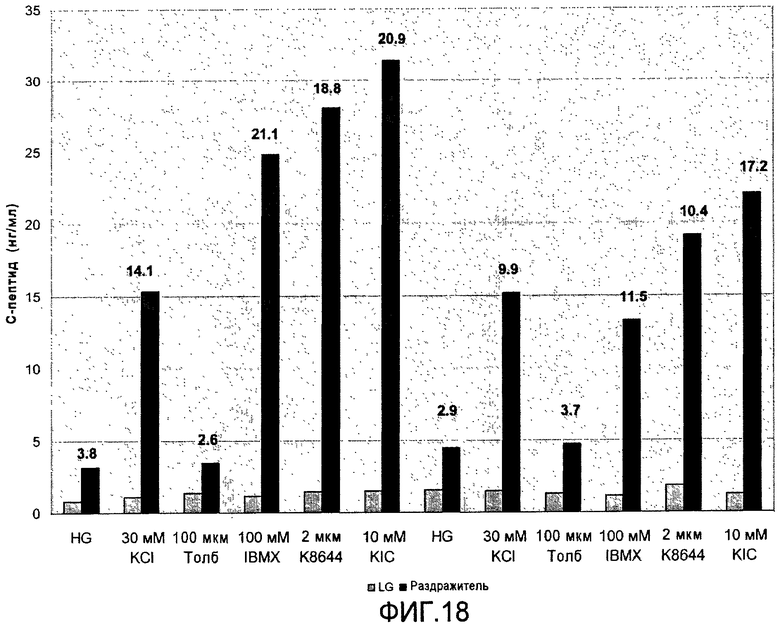
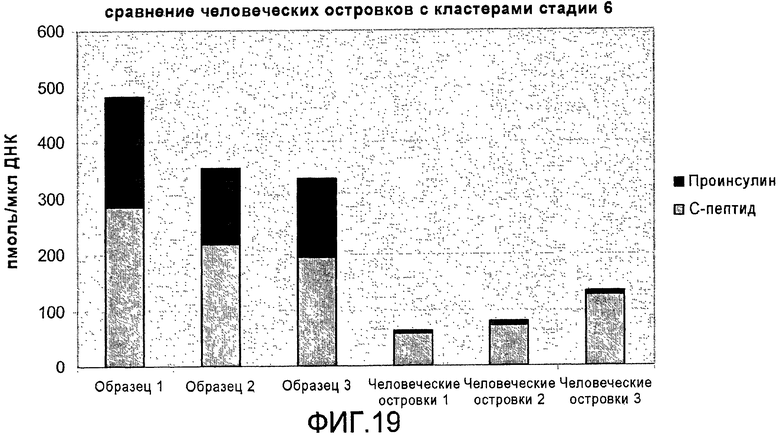

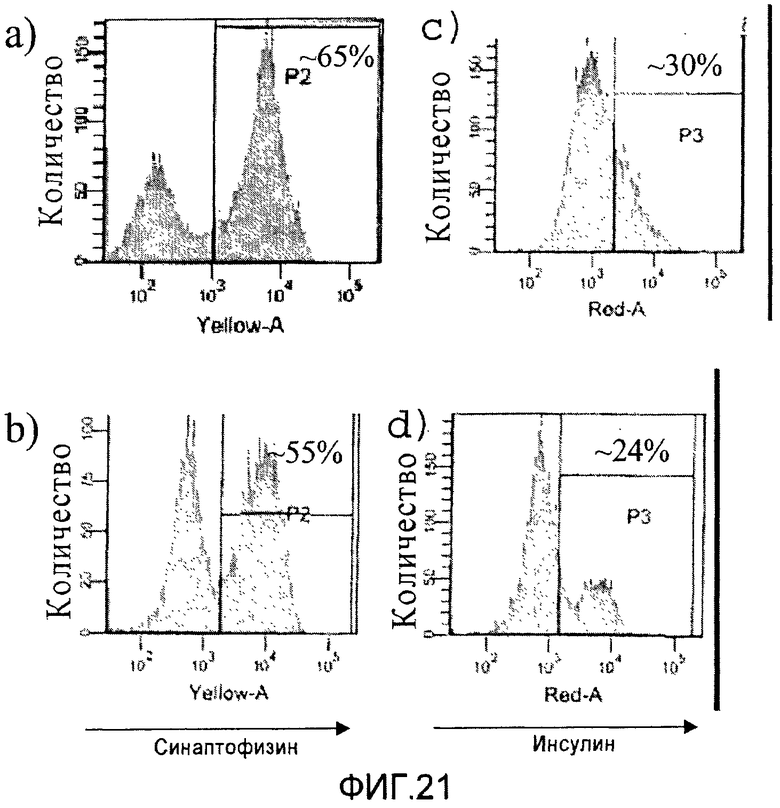
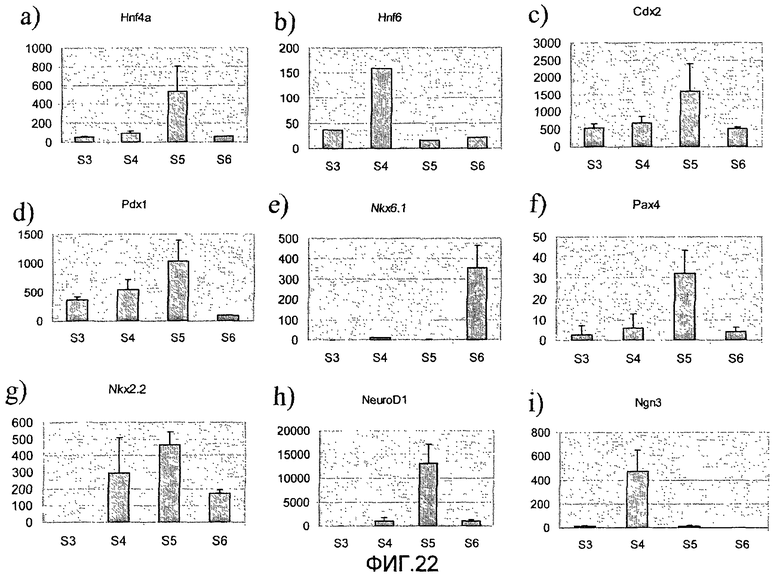
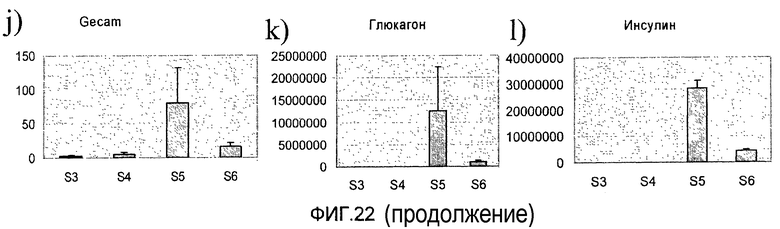
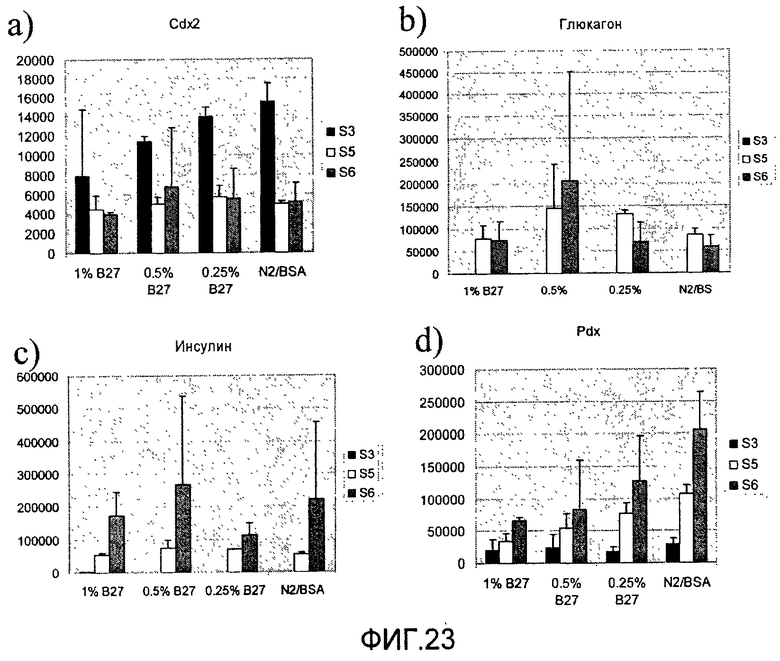
Изобретение относится к области молекулярной биологии и клеточной инженерии. Предложен способ получения инсулинпродуцирующих клеток, полученных из полипотентных стволовых клеток и способных секретировать инсулин без использования слоя питающих клеток. Изобретение может быть использовано в фармакологической промышленности для получения инсулина. 71 з.п. ф-лы, 23 ил., 17 пр.
1. Способ получения клеток, способных секретировать инсулин в ответ на стимуляцию глюкозой, из полипотентных стволовых клеток человека, включающий следующие стадии:
a. культивирование полипотентных стволовых клеток человека,
b. дифференцировку полипотентных стволовых клеток человека в клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы,
c. дифференцировку клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, и
d. дифференцировку клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, способные секретировать инсулин в ответ на стимуляцию глюкозой, путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь TGF-βR-1.
2. Способ по п.1, где клетки, экспрессирующие маркеры, характерные для линии клеток панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от одного до двенадцати дней.
3. Способ по п.1, где клетки, экспрессирующие маркеры, характерные для линии клеток панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от пяти до двенадцати дней.
4. Способ по п.1, где клетки, экспрессирующие маркеры, характерные для линии клеток панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь TGF-βR-1, в течение двенадцати дней.
5. Способ по п.1, где фактор, ингибирующий путь TGF-βR-1, представляет собой ингибитор киназы TGF-βR-1.
6. Способ по п.5, где фактор, ингибирующий сигнальный путь TGF-βR-1, представляет собой ингибитор киназы TGF-βR-1.
7. Способ по п.6, где киназу TGF-βR-1 используют в концентрации от 0,1 мкМ до 100 мкМ.
8. Способ по п.6, где ингибитор киназы TGF-βR-1 представляет собой 2-(3-(6-метилпиридин-2-ил)-1Н-пиразол-4-ил)-1,5-нафтиридин.
9. Способ по п.6, где ингибитор киназы TGF-βR-1 представляет собой [3-(пиридин-2-ил)-4-(4-хинонил)]-1Н-пиразол.
10. Способ по п.1, где клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, также обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из фактора, способного ингибировать BMP, и нетрина.
11. Способ по п.1, где стадию дифференцировки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в клетки, экспрессирующие маркеры, характерные для линии эндокринных клеток поджелудочной железы, проводят путем обработки клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1.
12. Способ по п.11, где клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от одного до двенадцати дней.
13. Способ по п.11, где клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение периода от пяти до двенадцати дней.
14. Способ по п.11, где клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, обрабатывают фактором, ингибирующим сигнальный путь Notch, и фактором, ингибирующим сигнальный путь TGF-βR-1, в течение двенадцати дней.
15. Способ по п.11, где фактор, ингибирующий сигнальный путь Notch, представляет собой ингибитор γ-секретазы.
16. Способ по п.15, где ингибитор γ-секретазы представляет собой L-685458.
17. Способ по п.15, где L-685458 используют в концентрации от 0,1 мкМ до 100 мкМ.
18. Способ по п.1, где клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, также обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из фактора, способного ингибировать BMP, и нетрина.
19. Способ по п.1, где стадию дифференцировки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, проводят путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина.
20. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина, в течение от одного до шести дней.
21. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина, в течение шести дней.
22. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до четырех дней.
23. Способ по п.22, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
24. Способ по п.22, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение четырех дней.
25. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение от одного до четырех дней.
26. Способ по п.25, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
27. Способ по п.25, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7, FGF-10 в течение четырех дней.
28. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, нетрином, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до четырех дней.
29. Способ по п.28, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
30. Способ по п.28, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, нетрином, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7, FGF-10, в течение четырех дней.
31. Способ по п.19, где фактор, который выбран из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, представляет собой FGF-7.
32. Способ по п.31, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают FGF-7 в концентрации от 50 пг/мл до 50 мкг/мл.
33. Способ по п.31, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают FGF-7 в концентрации 20 нг/мл.
34. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой в концентрации от 1 нМ до 1 мМ.
35. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой в концентрации 1 мкМ.
36. Способ по п.19, где ингибитор фактора «Sonic Hedgehog» представляет собой циклопамин.
37. Способ по п.36, где циклопамин используют в концентрации от 0,1 мкМ до 10 мкМ.
38. Способ по п.36, где циклопамин используют в концентрации, равной 0,25 мкМ.
39. Способ по п.19, где фактор, способный ингибировать BMP, представляет собой ингибитор ВМР4.
40. Способ по п.39, где ингибитор ВМР4 представляет собой Noggin.
41. Способ по п.39, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают Noggin в концентрации от 500 нг/мл до 100 мкг/мл.
42. Способ по п.39, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают Noggin в концентрации 100 нг/мл.
43. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают нетрином в концентрации от 500 нг/мл до 100 мкг/мл.
44. Способ по п.19, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают нетрином в концентрации 100 нг/мл.
45. Способ по п.19, где нетрин выбран из группы, состоящей из нетрина 1, нетрина 2 и нетрина 4.
46. Способ по п.1, где стадию дифференцировки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы, в клетки, экспрессирующие маркеры, характерные для линии панкреатической эндодермы, проводят путем обработки клеток, экспрессирующих маркеры, характерные для линии сформировавшейся эндодермы по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина.
47. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина в течение периода от одного до шести дней.
48. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают по меньшей мере одним фактором, выбранным из группы, состоящей из ретиноевой кислоты, FGF-2, FGF-4, FGF-7, FGF-10, ингибитора фактора «Sonic Hedgehog», фактора, способного ингибировать BMP, и нетрина в течение шести дней.
49. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до четырех дней.
50. Способ по п.49, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
51. Способ по п.49, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение четырех дней.
52. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором по меньшей мере фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до четырех дней.
53. Способ по п.52, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
54. Способ по п.52, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7, FGF-10, в течение четырех дней.
55. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение периода от одного до трех дней, а затем клетки обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, нетрином, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF- 7 и FGF-10, в течение периода от одного до четырех дней.
56. Способ по п.55, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog» и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, в течение трех дней.
57. Способ по п.55, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ингибитором фактора «Sonic Hedgehog», фактором, способным ингибировать BMP, нетрином, ретиноевой кислотой и по меньшей мере одним фактором, выбранным из группы, состоящей из FGF-2, FGF-4, FGF-7, FGF-10, в течение четырех дней.
58. Способ по п.46, где фактор, который выбран из группы, состоящей из FGF-2, FGF-4, FGF-7 и FGF-10, представляет собой FGF-7.
59. Способ по п.58, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают FGF-7 в концентрации от 50 пг/мл до 50 мкг/мл.
60. Способ по п.58, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают FGF-7 в концентрации 20 нг/мл.
61. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой в концентрации от 1 нМ до 1 мМ.
62. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают ретиноевой кислотой в концентрации 1 мкМ.
63. Способ по п.46, где ингибитор фактора «Sonic Hedgehog» представляет собой циклопамин.
64. Способ по п.63, где циклопамин используют в концентрации от 0,1 мкМ до 10 мкМ.
65. Способ по п.63, где циклопамин используют в концентрации, равной 0,25 мкМ.
66. Способ по п.46, где фактор, способный ингибировать BMP, представляет собой ингибитор ВМР4.
67. Способ по п.66, где ингибитор ВМР4 представляет собой Noggin.
68. Способ по п.66, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают Noggin в концентрации от 500 нг/мл до 100 мкг/мл.
69. Способ по п.66, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают Noggin в концентрации 100 нг/мл.
70. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают нетрином в концентрации от 500 нг/мл до 100 мкг/мл.
71. Способ по п.46, где клетки, экспрессирующие маркеры, характерные для линии сформировавшейся эндодермы, обрабатывают нетрином в концентрации 100 нг/мл.
72. Способ по п.46, где нетрин выбран из группы, состоящей из нетрина 1, нетрина 2 и нетрина 4.
| D′AMOUR KA, et al | |||
| Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells | |||
| Nat Biotechnol | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| SKOUDY A, et al | |||
| Transforming growth factor (TGF)beta, fibroblast growth factor (FGF) and retinoid signalling pathways promote pancreatic exocrine gene expression in mouse | |||

