ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к мутантным микроорганизмам, обладающим высокой способностью к продукции путресцина, и способу продукции путресцина с использованием этих микроорганизмов, а конкретнее к мутантным микроорганизмам, обладающим высокой способностью к продукции путресцина, в которых инактивированы или делетированы гены, участвующие в пути деградации или утилизации путресцина, и к способу продуцирования путресцина в большом количестве путем культивирования мутантных микроорганизмов.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Путресцин (также называемый 1,4-бутандиамин) важный исходный материал для продукции полиамида-4,6, в том числе нейлона-4,6, главным образом получают в промышленном масштабе путем гидрирования акрилонитрила, который получают добавлением синильной кислоты к ацетилену. Для известных способов химического синтеза этого соединения требуются невозобновляемые нефтехимические продукты в качестве исходных материалов и относительно жесткие условия реакций, включающие температуру и давление, в многостадийной и многореакторной разработке, а также использование дорогостоящей системы катализаторов. Кроме того, поскольку эти исходные материалы являются высокотоксичными и огнеопасными, известные способы химического синтеза неблагоприятны с экологической точки зрения. Соответственно необходим способ получения путресцина из возобновляемого источника углерода биомассового происхождения, в качестве альтернативы способу химического получения.
Путресцин является разновидностью полиамина, который образуется в ряде организмов, простирающихся от бактерий до животных и растений. Например, известно, что путресцин играет важную роль не только в пролиферации клеток и нормальном росте клеток, но также в защитном механизме против окислительного стресса (Tkachenko et al., Arch. Microbiol., 176: 155-157, 2001). Между тем, внутриклеточное содержание полиаминов тесно контролируется их биосинтезом, деградацией, поглощением и выделением (Igarashi and Kashiwagi et al., J. Bacteriol., 170(7): 3131-3135, 1988). Как это известно в данной области техники, концентрация путресцина в E.coli является очень высокой, составляя приблизительно 2,8 г/л. Также потенциально микроорганизмы обладают хорошей устойчивостью к высоким концентрациям полиаминов. Например, Mimitsuka и др. представили данные, что Corynebacterium glutamicum может расти даже в присутствии более чем 30 г/л кадаверина. Поэтому были продолжены исследования по применению микроорганизмов для продукции полиаминов (путресцина) в высоких концентрациях.
В ЕР 0726240 А1 раскрывается способ продукции путресцина через ферментацию, используя или недорогие промышленные отходы, или материалы, имеющие белок в качестве основной составляющей. Однако поскольку раскрытые материалы являются очень сложными, существует проблема, заключающаяся в том, что необходимо выполнить много стадий очистки для получения продуктов в виде путресцина и кадаверина. Кроме того, в публикации Европейского патента № 1784496 А1 раскрывается способ синтеза путресцина биохимически путем выращивания микроорганизмов в минимальной солевой среде, содержащей глюкозу в качестве источника углерода. В этой публикации Европейского патента для увеличения превращения орнитина в путресцин была увеличена активность орнитиндекарбоксилазы путем сверхэкспрессии гена speC или speF, кодирующего орнитиндекарбоксилазу. Однако когда содержание путресцина увеличивается в результате увеличения активности орнитиндекарбоксилазы, возникают проблемы, заключающиеся в уменьшении биосинтеза путресцина и индуцировании деградации путресцина.
Исследованиями, связанными с деградацией и утилизацией путресцина в микроорганизмах, являются следующие исследования. Bowman и др. представили данные, что спермидинсинтаза, являющаяся продуктом гена speE, активирует биосинтез спермидина из путресцина в E.coli (Bowman et al., J. Biol. Chem., 248: 2480-2486, 1973). Спермидинсинтаза (EC: 2.5.1.16) присутствует в большинстве клеточных систем для синтеза спермидина.
Haywood и др. представили данные, что в дрожжах Candida boidinii N-ацетилтрансфераза ацетилирует путресцин с превращением его в N-ацетилпутресцин. Спермидин-ацетилтрансфераза, являющаяся продуктом гена speG E.coli, обладает высокой степенью гомологии с N-ацетилтрансферазой дрожжей и, следовательно, должна обладать активностью путресцин-ацетитрансферазы (Haywood and Large, Eur. J. Biochem., 148: 277-283, 1985).
Кроме того, Samsonova и др. представили данные, касающиеся другого пути деградации путресцина, в котором спаренное действие путресцин-трансаминазы YgjG и дегидрогеназы YdcW E.coli приводит к превращению путресцина в γ-аминомасляную кислоту без γ-глутамилирования (Samsonova et al., BMC Microbiol., 3:2, 2003; Samsonova et al., FEBS Lett., 579: 4107-4112, 2005).
Кроме того, Kurihara и др. представили данные, что в путь деградации путресцина, «путь c участием puu», включены γ-глутамилированные промежуточные продукты E.coli. Благодаря γ-глутамилированию путресцина в этом пути может быть стабилизирован γ-аминомасляный альдегид, который является промежуточным альдегидным продуктом. γ-Глутамилпутресцинсинтаза, являющаяся продуктом гена puuA, превращает путресцин в γ-глутамил-L-путресцин, активируя первую реакцию этого пути. Также было обнаружено, что этот катаболический путь является основным путем при росте E.coli в питательной среде, содержащей путресцин в качестве единственного источника азота. Кроме того, было обнаружено, что в катаболическом пути участвует импортер путресцина puuC, и он является основным импортером путресцина при росте E.coli в питательной среде, содержащей путресцин в качестве единственного источника азота (Kurihava et al., J. Biol. Chem., 280: 4602-4608, 2005).
Поэтому авторы настоящего изобретения получили мутантные микроорганизмы, в которых инактивирован или делетирован по крайней мере один ген, выбираемый из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина продуцирующих путресцин микроорганизмов. Также авторы настоящего изобретения установили, что при культивировании мутантных микроорганизмов они могут продуцировать путресцин в больших количествах, что делает завершенным настоящее изобретение.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предоставление мутантных микроорганизмов, в которых инактивирован или делетирован по крайней мере один ген, включенный в путь деградации или утилизации путресцина, и которые обладают высокой способностью к продукции путресцина, и способа получения этих мутантных микроорганизмов.
Другой целью настоящего изобретения является предоставление способа продуцирования путресцина в большом количестве путем культивирования мутантных микроорганизмов.
Для достижения вышеприведенных целей настоящим изобретением предоставляется мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина, и способ получения этого мутантного микроорганизма.
Настоящим изобретением также предоставляется мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина, и в котором промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, заменен сильным промотором, и способ получения этого мутантного микроорганизма.
Настоящим изобретением также предоставляется мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина; в котором промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, заменен сильным промотором; и в который введен ген (specC), кодирующий орнитиндекарбоксидазу, или в котором этот ген амплифицирован, и способ получения этого мутантного микроорганизма.
Настоящим изобретением также предоставляется способ продуцирования путресцина, включающий культивирование указанного мутантного микроорганизма для продуцирования путресцина и затем получение путресцина из культуральной жидкости.
Другие характеристики и аспекты настоящего станут понятнее из следующего подробного описания и прилагаемой формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 представляет собой схематическое представление пути синтеза путресцина из глюкозы.
Фиг.2 представляет собой диаграмму в виде графика, демонстрирующую продукцию путресцина из клеток XQ37/pKKSpeC при ферментации в подпитываемой глюкозой культуре.
Фиг.3 представляет собой диаграмму в виде графика, демонстрирующую продукцию путресцина из клеток XQ39 при ферментации в подпитываемой глюкозой культуре.
Фиг.4 представляет собой диаграмму в виде графика, демонстрирующую продукцию путресцина из клеток XQ43 при ферментации в подпитываемой глюкозой культуре.
Фиг.5 представляет собой диаграмму в виде графика, демонстрирующую продукцию путресцина из клеток XQ43/p15SpeC при ферментации в подпитываемой глюкозой культуре.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Подразумевается, что используемый здесь термин «инактивированный» включает мутирование, замену или делетирование части представляющего интерес гена или введение в ген одного или нескольких оснований, для того чтобы снизить активность фермента, который экспрессируется с гена, тем самым блокируя часть, или значительную часть, пути биосинтеза, в котором участвует фермент гена.
Подразумевается, что используемый здесь термин «делетированный» включает мутирование, замену или делетирование части представляющего интерес гена или всего гена или введение в ген одного или нескольких оснований, для того чтобы ген не экспрессировался или не проявлял ферментативную активность, даже если он экспрессируется, тем самым блокируя путь биосинтеза, в котором участвует этот ген.
Подразумевается, что используемый здесь термин «амплифицированный» включает мутирование, замену или делетирование части представляющего интерес гена или введение в ген одного или нескольких оснований, или введение гена другого микробного происхождения, кодирующего тот же самый фермент, для того чтобы усилить активность соответствующего фермента.
Фиг.1 представляет собой схематическое представление пути синтеза путресцина из глюкозы. Как показано на фиг.1, в настоящем изобретении было установлено, что когда гены (speE, speG, argI и puuP), участвующие в пути деградации или утилизации путресцина микроорганизма, обладающего способностью к продукции путресцина, были инактивированы или делетированы, мутантный микроорганизм не мог продуцировать путресцин в большом количестве. Снижение активностей генов (speE, speG, argI и puuP), участвующих в пути деградации или утилизации путресцина, могло быть подтверждено снижением транскрипционной и трансляционной активности по сравнению с активностями соответствующих генов дикого типа.
В примерах настоящего изобретения авторы настоящего изобретения получили мутантные микроорганизмы, в которых инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего орнитин-карбамоилтрансферазу I, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина. Было установлено, что полученные мутантные микроорганизмы обладают повышенной способностью к продукции путресцина.
Соответственно в одном аспекте настоящее изобретение относится к мутантному микроорганизму, обладающему способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего орнитин-карбамоилтрансферазу I, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина, и к способу получения этого мутантного микроорганизма.
В мутантном микроорганизме настоящего изобретения может быть дополнительно инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (puuA), кодирующего γ-глутамилпутресцинсинтазу, гена (ygjG), кодирующего путресцин-трансаминазу, и гена (argF), кодирующего орнитин-карбамоилтрансферазу F.
Фермент гена (argF), кодирующего орнитин-карбамоилтрансферазу F, является изоферментом гена (argI), кодирующего орнитин-карбамоилтрансферазу I, а ген (puuA), кодирующий γ-глутамилпутресцинсинтазу, является геном, соседствующим с геном (puuP), кодирующим импортер путресцина. Также ген (ygjG), кодирующий путресцин-трансаминазу, является геном, участвующим в деградации путресцина.
В мутантном микроорганизме настоящего изобретения может быть также дополнительно делетирован ген (lacI), кодирующий репрессор оперона lac, вследствие чего усиливается экспрессия генов, кодирующих ферменты, которые участвуют в биосинтезе путресцина. Примеры генов, кодирующих ферменты, которые участвуют в биосинтезе путресцина, включают gdhA, argA, argB, argC, arhD, argE и т.п.
В мутантный микроорганизм настоящего изобретения может быть также дополнительно введен ген (speC), кодирующий орнитиндекарбоксилазу, или этот ген может быть амплифицирован в мутантном микроорганизме. Ген (speC), кодирующий орнитиндекарбоксилазу, вводят в форме экспрессионного вектора, содержащего сильный промотор. Сильный промотор может быть выбран из группы, состоящей из промотора trc, промотора tac, промотора T7, промотора lac и промотора trp.
В качестве микроорганизма в настоящем изобретении можно использовать, без особого ограничения, любой микроорганизм при условии, что он продуцирует путресцин из глюкозы. Примеры микроорганизмов включают Bacillus sp., Corynebacterium sp., Escherichia sp., Pichia sp., Pseudomonas sp., Saccharomyces sp. и т.п.
В настоящем изобретении было установлено, что в случае мутантного микроорганизма, в котором был делетирован ген, участвующий в пути деградации или утилизации путресцина, когда промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, был заменен сильным промотором, получаемый в результате мутантный микроорганизм мог продуцировать путресцин в большем количестве.
В примерах настоящего изобретения, на основе мутантных микроорганизмов, в которых были делетированы гены (speE, speG, argI и puuP), участвующие в пути деградации или утилизации путресцина, и ген (lacI), кодирующий репрессор оперона lac, были получены следующие микроорганизмы: микроорганизм (XQ33), в котором промотор гена (argECBH), кодирующего оперон для биосинтеза аргинина, был заменен сильным промотором (trc); микроорганизм (XQ37), в котором промоторы гена argECBH и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, были заменены сильным промотором trc; мутантный микроорганизм (XQ39), в котором промоторы гена argECBH, гена speF-potE и гена (argD), кодирующего ацетилорнитин-аминотрасферазу, были заменены сильным промотором trc; и мутантный микроорганизм (XQ43), в котором промоторы гена argECBH, гена speF-potE, гена argD и гена (speC), кодирующего орнитиндекарбоксилазу, были заменены сильным промотором (trc). Было установлено, что способности этих микроорганизмов к продукции путресцина были резко усилены.
Соответственно в другом аспекте настоящее изобретение относится к мутантному микроорганизму, обладающему способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего орнитин-карбамоилтрансферазу I, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина, и в котором промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, заменен сильным промотором, и к способу получения этого мутантного микроорганизма.
В настоящем изобретении ген (argECBH), кодирующий оперон для биосинтеза аргинина, является дивергентным опероном, который фланкирован двумя конвергентными промоторами (argEp и argCBHp) и содержит оператор. Два промотора репрессируются аргинином (Charlier and Glansdorff, 2004). Таким образом, когда природный промотор оперона argECBH заменяют сильным промотором, метаболический поток в сторону орнитина может быть увеличен. Ген argE является геном, кодирующим N-ацетилорнитиназу, ген argC представляет собой ген, кодирующий N-ацетилглутамилфосфатредуктазу, ген argB является геном, кодирующим N-ацетилглутаматкиназу, и ген argH является геном, кодирующим аргининосукциназу.
Ген (speF-potE), кодирующий индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, который индуцируется при низком рН, кодирует индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер. Таким образом, когда природный промотор оперона speF-potE заменен сильным промотором, оперон speF-potE может экспрессироваться конститутивно, и, следовательно, способность к продукции путресцина может быть усилена.
Репрессором промотора гена (argD), кодирующего ацетилорнитин-аминотрасферазу, является аргинин (Charlier and Glansdorff, 2004). Таким образом, когда природный промотор оперона argD заменяют сильным промотором, метаболический поток в сторону орнитина может быть увеличен.
Как здесь описано, в мутантном микроорганизме настоящего изобретения может быть дополнительно инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (puuA), кодирующего γ-глутамилпутресцинсинтазу, гена (ygjG), кодирующего путресцин-трансаминазу, и гена (argF), кодирующего орнитин-карбамоилтрансферазу F.
В мутантном микроорганизме, обладающем способностью к продукции путресцина, может быть дополнительно делетирован ген (lacI), кодирующий репрессор оперона lac.
В настоящем изобретении ген (speC), кодирующий орнитиндекарбоксилазу, предпочтительно вводят в форме экспрессионного вектора, содержащего сильный промотор.
В настоящем изобретении сильный промотор, который используют для замены промотора гена и при введении гена (speC), кодирующий орнитиндекарбоксилазу, предпочтительно выбирают из группы, состоящей из промотора trc, промотора tac, промотора T7, промотора lac и промотора trp.
Наиболее предпочтительным примером мутантного микроорганизма в соответствии с настоящим изобретением может быть мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена (speE), кодирующего спермидинсинтазу, гена (speG), кодирующего спермидин-N-ацетилтрансферазу, гена (argI), кодирующего орнитин-карбамоилтрансферазу I, и гена (puuP), кодирующего импортер путресцина, которые участвуют в пути деградации или утилизации путресцина; в котором промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, и гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, заменен сильным промотором; и в который введен ген (specC), кодирующий орнитиндекарбоксилазу, или в котором этот ген амплифицирован.
В еще одном аспекте настоящее изобретение относится к способу продуцирования путресцина, включающему культивирование указанного мутантного микроорганизма для продуцирования путресцина и затем получение путресцина из культуральной жидкости.
В настоящем изобретении процессы культивирования мутантного микроорганизма и получения путресцина из культуральной жидкости можно выполнить, используя обычный способ культивирования (культуру клеток, выращиваемую в замкнутом объеме, или подпитываемую культуру, известные из предшествующих способов ферментации), и известный в данной области техники способ выделения и очистки путресцина.
В настоящем изобретении биосинтетическую продукцию путресцина можно выполнить in vivo или in vitro.
ПРИМЕРЫ
Ниже настоящее изобретение описывается подробнее со ссылкой на примеры. Однако должно быть понятно, что эти примеры приведены только с иллюстративной целью и не должны толковаться как ограничение объема настоящего изобретения.
В частности, хотя в следующих примерах проиллюстрированы только конкретные типы векторов для удаления генов в соответствии с настоящим изобретением и продуцирующие путресцин микроорганизмы Escherichia sp., служащие в качестве клеток-хозяев, квалифицированному в данной области техники специалисту будет также очевидно использование других типов векторов и продуцирующих путресцин микроорганизмов.
Пример 1: Получение мутантного микроорганизма, в котором делетированы гены, участвующие в пути деградации или утилизации путресцина
В настоящем изобретении делецию генов (puuA, ygjG, speE, speG, argF и argI) на хромосомах осуществляли с помощью гомологичной рекомендации при двойном кроссинговере (Datsenko, K.A., & Wanner, B.L. Proc. Natl. Acad. Sci., 97: 6640-6645, 2000). Кассету lox71-маркер устойчивости к хлорамфениколу (CmR)-lox66 получали с помощью ПЦР, используя праймеры, содержащие 50 нуклеотидов, гомологичных 5'- и 3'-участкам гена-мишени. Плазмиду pECmulox (Kim, J.M., Lee, K.H. & Lee. S.Y., FEMS Microbiol. Lett., 278: 78-85, 2008), включающую кассету lox71-CmR-lox66, использовали в качестве матрицы в ПЦР. ПЦР-продукты трансформировали в электрокомпетентные клетки, содержащие рекомбиназу λ. Колонии отбирали на чашках с агаризованной средой Лурия-Бертани (LB), содержащей 34 мкг/мл хлорамфеникола (Cm) (Sambrook, J., Fritsch E.F., & Maniatis, T., Molecular cloning: a laboratory manual, 3rd edition, Cold Spring Harbor Laboratory Press, 2000). Успешную замену гена на CmR подтверждали с помощью ПЦР с непосредственным использованием колоний. Антибиотический маркер удаляли с помощью плазмиды-хелпера pJW168, имеющей термочувствительное начало репликации и экспрессирующей IPTG-индуцируемую рекомбиназу cre (Palmeros et al., Gene, 247(1); 255-264, 2000).
1-1: Получение штамма WL3110
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 1 и 2, получая, таким образом, ПЦР-продукт, в котором был делетирован ген lacI. Затем полученный ПЦР-продукт подвергали электропорации в электрокомпетентный штамм E.coli (W3110), содержащий рекомбиназу λ, получая, таким образом, штамм WL3110 (W3110 ΔlacI).
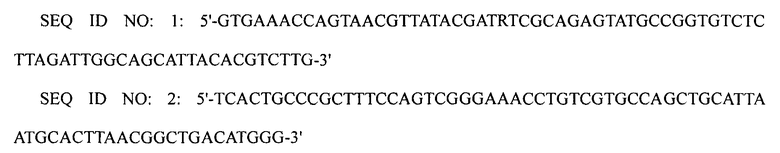
1-2: Получение штамма XQ08
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 3 и 4, получая, таким образом, ПЦР-продукт, в котором был делетирован ген speE. Затем полученный ПЦР-продукт подвергали электропорации в штамм WL3110, приготовленный в примере 1-1, получая, таким образом, штамм XQ08 (W3110 ΔlacI ΔspeE).
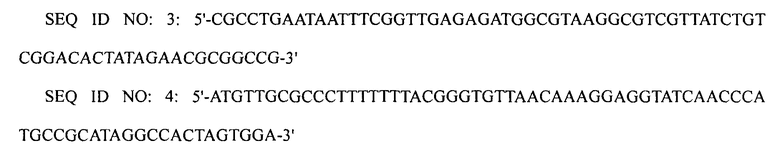
1-3: Получение штамма XQ17
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 5 и 6, получая, таким образом, ПЦР-продукт, в котором был делетирован ген puuA. Затем полученный ПЦР-продукт подвергали электропорации в штамм WL3110, приготовленный в примере 1-1, получая, таким образом, штамм XQ17 (W3110 ΔlacI ΔpuuA).
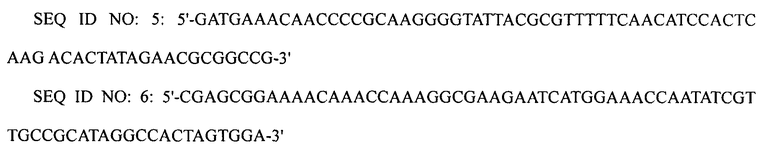
1-4: Получение штамма XQ22
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 6 и 7, получая, таким образом, ПЦР-продукт, в котором был делетирован ген puuP. Затем полученный ПЦР-продукт подвергали электропорации в штамм XQ17 (W3110 ΔlacI ΔpuuA), приготовленный в примере 1-3, получая, таким образом, штамм XQ22 (W3110 ΔlacI ΔpuuP ΔpuuA).
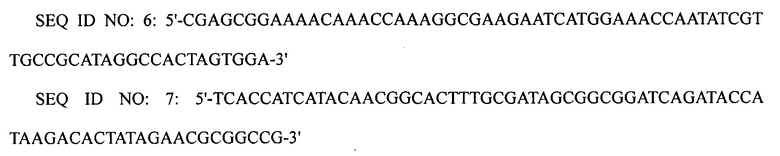
1-5: Получение штамма XQ23
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 8 и 9, получая, таким образом, ПЦР-продукт, в котором был делетирован ген speE. Затем полученный ПЦР-продукт подвергали электропорации в штамм WL3110, приготовленный в примере 1-1, получая, таким образом, штамм XQ23-1 (W3110 ΔlacI ΔspeE).
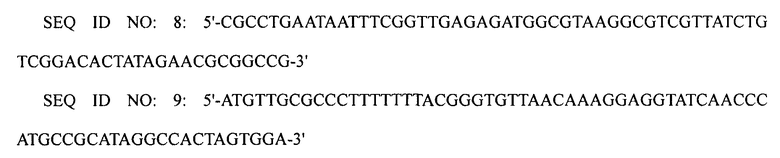
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 10 и 11, получая, таким образом, ПЦР-продукт, в котором был делетирован ген speG. Затем полученный ПЦР-продукт подвергали электропорации в штамм XQ23-1 (W3110 ΔlacI ΔspeE), получая, таким образом, штамм XQ23-2 (W3110 ΔlacI ΔspeE ΔspeG).

ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 12 и 13, получая, таким образом, ПЦР-продукт, в котором был делетирован ген argI. Затем выполняли ПЦР, используя полученный ПЦР-продукт в качестве матрицы и праймеры SEQ ID NO: 14 и 15, получая, таким образом, конечный ПЦР-продукт. Полученный конечный ПЦР-продукт подвергали электропорации в штамм XQ23-2 (W3110 ΔlacI ΔspeE ΔspeG), получая, таким образом, штамм XQ23 (W3110 ΔlacI ΔspeE ΔspeG ΔargI).
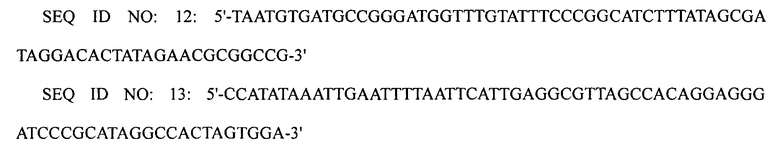
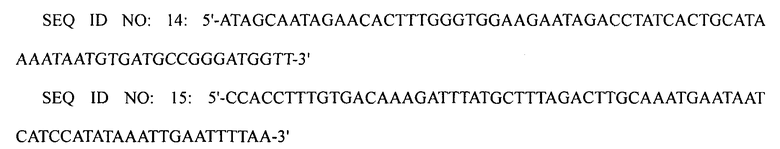
1-6: Получение штамма XQ26
ПЦР-продукт, приготовленный в примере 1-3, в котором был делетирован ген puuA, и ПЦР-продукт, приготовленный в примере 1-4, в котором был делетирован ген puuP, подвергали электропорации в штамм XQ23 (W3110 ΔlacI ΔspeE ΔspeG ΔargI), получая, таким образом, штамм XQ26 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA).
1-7: Получение штамма XQ27
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 16 и 17, получая, таким образом, ПЦР-продукт, в котором был делетирован ген ygjG. Затем полученный ПЦР-продукт подвергали электропорации в штамм XQ23-2 (W3110 ΔlacI ΔspeE ΔspeG), приготовленный в примере 1-5, получая, таким образом, штамм XQ27-1 (W3110 ΔlacI ΔspeE ΔspeG Δ ygjG). Затем ПЦР-продукт, приготовленный в примере 1-3, в котором был делетирован ген puuA, и ПЦР-продукт, приготовленный в примере 1-4, в котором был делетирован ген puuP, подвергали электропорации в штамм XQ27-1 (W3110 ΔlacI ΔspeE ΔspeG ΔygjG), получая, таким образом, штамм XQ27 (W3110 ΔlacI ΔspeE ΔspeG ΔygjG ΔpuuP ΔpuuA).
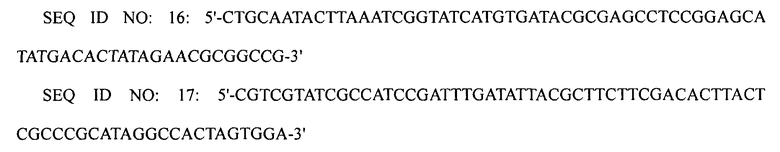
1-8: Получение штамма XQ29
ПЦР-продукт, приготовленный в примере 1-7, в котором был делетирован ген ygjG, подвергали электропорации в штамм XQ26 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA), приготовленный в примере 1-6, получая, таким образом, штамм XQ29 (W3110 ΔlacI ΔspeE ΔspeG ΔygjG ΔargI ΔpuuP ΔpuuA).
ПЦР выполняли, используя плазмиду pECmulox в качестве матрицы и праймеры SEQ ID NO: 1 и 2, получая, таким образом, ПЦР-продукт, в котором был делетирован ген lacI. Затем полученный ПЦР-продукт подвергали электропорации в электрокомпетентный штамм E.coli (W3110), содержащий рекомбиназу λ, получая, таким образом, штамм WL3110 (W3110 ΔlacI).
Пример 2: Замена промотора
Для усиления способности к продукции путресцина промотор мутантного штамма (XQ26), приготовленного в примере 1, заменяли сильным промотором (trc).
2-1: Получение штамма XQ33
Замену природного промотора оперона argECBH промотором trc осуществляли следующим образом. ДНК-фрагмент, содержащий слияние lox71-антибиотический маркер (маркер устойчивости к хлорамфениколу)-lox66, создавали с помощью первой реакции ПЦР с использованием праймеров SEQ ID NO: 18 и 19, используя плазмиду pECmulox в качестве матрицы.
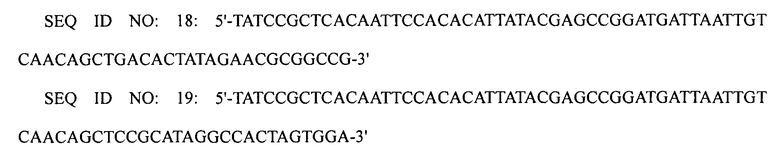
Для того чтобы ввести промотор trc, проводили вторую реакцию ПЦР с использованием праймеров SEQ ID NO: 20 и 21, используя первый ПЦР-продукт в качестве матрицы. Для того чтобы ввести гомологические участки в конечный ПЦР-продукт, проводили третью реакцию ПЦР с использованием праймеров SEQ ID NO: 22 и 23, используя второй ПЦР-продукт в качестве матрицы.
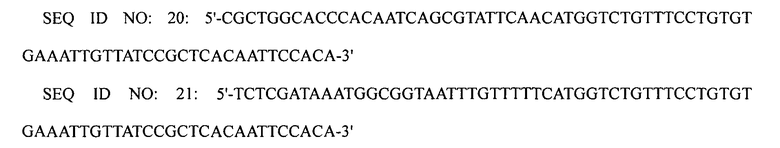
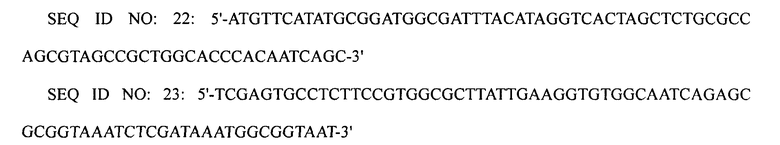
Конечный ПЦР-продукт подвергали электропорации в штамм XQ26 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA), приготовленный в примере 1-1, конструируя, таким образом, рекомбинантный микроорганизм. Затем клетки мутантного микроорганизма культивировали на чашке с агаром, содержащим хлорамфеникол, с отбором на чашке с агаром клеток, в которых произошла двойная гомологичная рекомбинация, получая, таким образом, штамм XQ33 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc). Затем замену промотором trc в полученном штамме подтверждали с помощью анализа последовательности ДНК.
2-2: Получение штамма XQ37
Замену природного промотора оперона speF-potE промотором trc осуществляли следующим образом. Первую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 24, используя плазмиду pECmulox в качестве матрицы.

Вторую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 25 и 26, используя первый ПЦР-продукт в качестве матрицы. Третью реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 27 и 28, используя второй ПЦР-продукт в качестве матрицы.


Конечный ПЦР-продукт подвергали электропорации в штамм XQ33 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc), приготовленный в примере 2-1, конструируя, таким образом, рекомбинантный микроорганизм. Затем клетки рекомбинантного микроорганизма культивировали на чашке с агаром, содержащим хлорамфеникол, с отбором на чашке с агаром клеток, в которых произошла двойная гомологичная рекомбинация, получая, таким образом, штамм XQ37 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc PspeF-potE::Ptrc). Замену промотором trc подтверждали с помощью анализа последовательности ДНК.
2-3: Получение штамма XQ39
Замену природного промотора оперона argD промотором trc осуществляли следующим образом. Первую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 29, используя плазмиду pECmulox в качестве матрицы.
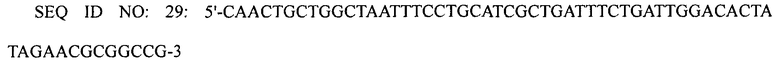
Вторую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 30 и 31, используя первый ПЦР-продукт в качестве матрицы. Третью реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 32 и 33, используя второй ПЦР-продукт в качестве матрицы.
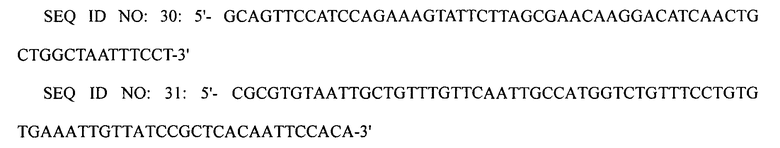
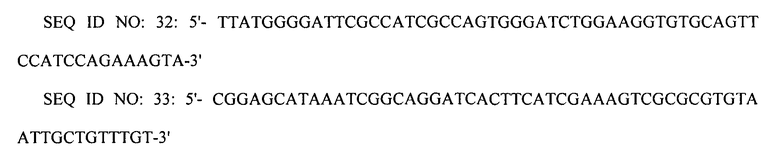
Конечный ПЦР-продукт подвергали электропорации в штамм XQ37 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc PspeF-potE::Ptrc), приготовленный в примере 2-2, получая, таким образом, штамм XQ39 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc PspeF-potE::Ptrc PargD::Ptrc). Затем клетки рекомбинантного микроорганизма культивировали на чашке с агаром, содержащим хлорамфеникол, с отбором на чашке с агаром клеток, в которых произошла двойная гомологичная рекомбинация. Замену промотором trc подтверждали с помощью анализа последовательности ДНК.
2-4: Получение штамма XQ43
Замену природного промотора гена speC промотором trc осуществляли следующим образом. Первую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 36, используя плазмиду pECmulox в качестве матрицы.
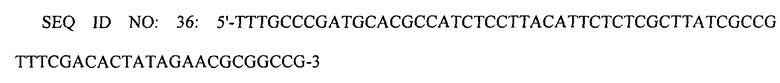
Вторую реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 37 и 38, используя первый ПЦР-продукт в качестве матрицы. Третью реакцию ПЦР проводили с использованием праймеров SEQ ID NO: 39 и 40, используя второй ПЦР-продукт в качестве матрицы.
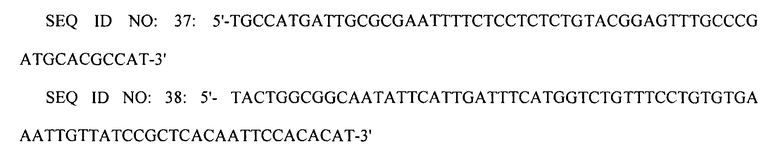
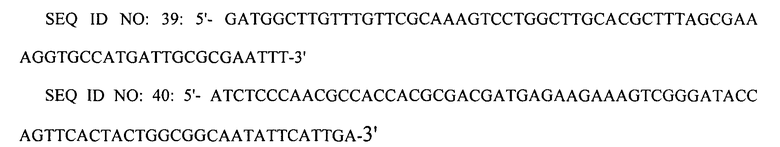
Конечный ПЦР-продукт подвергали электропорации в штамм XQ39 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc PspeF-potE::Ptrc PargD::Ptrc), приготовленный в примере 2-3, получая, таким образом, штамм XQ43 (W3110 ΔlacI ΔspeE ΔspeG ΔargI ΔpuuP ΔpuuA PargECBH::Ptrc PspeF-potE::Ptrc PargD::Ptrc PspeC::Ptrc). Затем клетки рекомбинантного микроорганизма культивировали на чашке с агаром, содержащим хлорамфеникол, с отбором на чашке с агаром клеток, в которых произошла двойная гомологичная рекомбинация. Замену промотором trc подтверждали с помощью анализа последовательности ДНК.
Пример 3: Продукция путресцина, используя мутантные микроорганизмы
Каждый из мутантных штаммов (мутантов E.coli K12 WL3110), представленных в таблице 1, приготовленных в примерах 1 и 2, культивировали в колбе с минимальной средой R, содержащей 4 г/л (NH4)2HPO4, 13,5 г/л KH2PO4, 1,7 г/л лимонной кислоты, 0,7 г/л MgSO4·7H2O и 0,5% (в объемном отношении) раствора сверхредких металлов (Lee, S.Y. & Chang, H.N., Biotechnol. Lett., 15: 971-974, 1993). Раствор сверхредких металлов содержал (на литр) 5 М HCl, 10 г FeSO4·7H2O, 2,25 г ZnSO4·7H2O, 1 г CuSO4·5H2O, 0,5 г MnSO4·5H2O, 0,23 г Na2B4O7·10H2O, 2 г CaCl2·2H2O и 0,1 г (NH4)6Mo7O24. Раствор, содержащий глюкозу (100 г/л), стерилизовали отдельно и добавляли в подвергнутую стерилизации среду до конечной концентрации 10 г/л.
100 мкл каждой клеточной культуры, сформированной в среде LB, инокулировали в минимальную среду и затем культивировали при 30οС при 220 об/мин в течение 24 часов до достижения значения ОП660, составляющего 5. Затем 1 мл культуральной жидкости добавляли в колбу перегородчатой конструкции на 350 мл, содержащую 50 мл такой же среды, и затем культивировали при 30οС при 220 об/мин в течение 15 часов. Клетки отделяли центрифугированием, и супернатант анализировали с помощью HPLC. Содержащиеся в супернатанте амины выявляли по деривации о-фталевого диальдегида (ОРА) в последовательной системе Hewlett Packard 1000 (230 нм), используя колонку С18 для хроматографии с обращенной фазой (буфер А: 45% 0,1М натрия ацетата, рН 7,2; буфер В: метанол). Анализ проводили в следующих условиях: 1-6 мин 100% буфера А для уравновешивания, 6-10 мин линейный градиент от 0 до 30% буфера В, 10-15 мин градиент от 30% до 50% буфера В, 15-19 мин градиент от 50% до 100% буфера В, 19-23 мин градиент до 100% буфера В и 23-25 мин градиент от 100% до 30% буфера В, 25-28 мин от 30% В до 100% А со скоростью потока 0,8 мл/мин). При этом для калибровки использовали стандарт, и определенные концентрации путресцина представлены ниже в таблице 1.
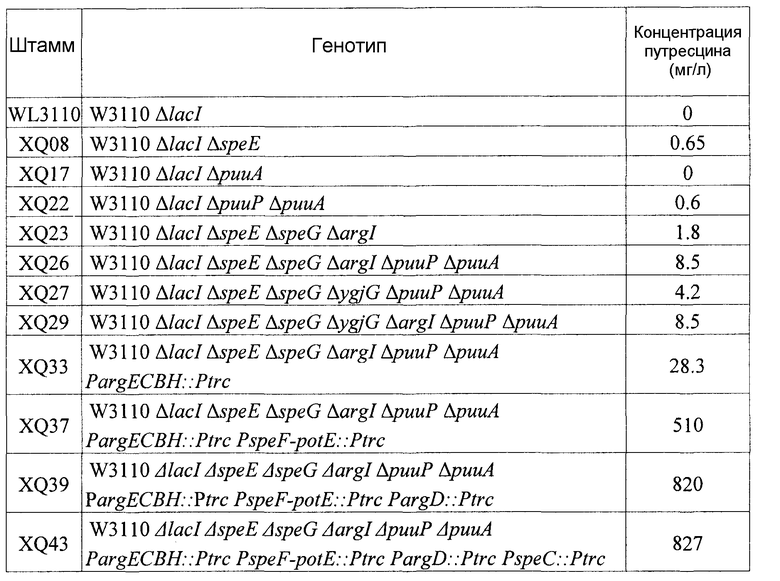
Как видно из таблицы 1, у мутантных микроорганизмов, в которых были делетированы гены (puuP, puuA, speE, speG и argI), участвующие в пути деградации или утилизации путресцина, способности к продукции путресцина были усилены в зависимости от вида и числа делетированных генов. Также можно было наблюдать, что способности мутантных микроорганизмов к продукции путресцина дополнительно усиливаются после замены промоторов гена (argECBH), кодирующего оперон для биосинтеза аргинина, гена (argD), кодирующего ацетилорнитин-аминотрасферазу, гена (speF-potE), кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, и гена (speC), кодирующего орнитиндекарбоксилазу, сильным промотором.
Пример 4: Амплификация гена (speC), кодирующего орнитиндекарбоксилазу
4-1: Приготовление плазмиды pKKSpeC
Кодирующий орнитиндекарбоксилазу ген (speC) E.coli W3110 с ее конститутивным биосинтезом клонировали в экспрессионный вектор pKK223-3 (Pharmacia Biotech, Uppsala, Швеция), в котором для экспрессии гена используется промотор tac. ПЦР выполняли с использованием праймеров SEQ ID NO: 34 и 35, используя геномную ДНК E.coli W3110 (происходящего из E.coli K-12, λ-, F-, прототрофного) в качестве матрицы, получая, таким образом, фрагмент speC (размером 2156 п.о.).

Затем полученный фрагмент speC (размером 2156 п.о.) и плазмиду pKK223-3 обрабатывали ферментами рестрикции (EcoRI и PstI), а затем, используя Т4 ДНК-лигазу, расщепленные ферментами рестрикции фрагмент speC и плазмиду pKK223-3 соединяли друг с другом, получая, таким образом, рекомбинантный плазмидный вектор pKKSpeC с большим числом копий.
4-2: Приготовление плазмиды p15SpeC
Кодирующий орнитиндекарбоксилазу ген (speC) E.coli W3110 с ее конститутивным биосинтезом клонировали в экспрессионный вектор pTac15K (происходящий из p15A, низкой копийности, KmR; продукт KAISTBEL), в котором для экспрессии гена используется промотор tac. ПЦР выполняли с использованием праймеров SEQ ID NO: 34 и 35, используя геномную ДНК E.coli W3110 (происходящего из E.coli K-12, λ-, F-, прототрофного) в качестве матрицы, получая, таким образом, фрагмент speC (размером 2156 п.о.).
Затем полученный фрагмент speC (размером 2156 п.о.) и плазмиду pTac15K обрабатывали ферментами рестрикции (EcoRI и PstI), а затем, используя Т4 ДНК-лигазу, расщепленные ферментами рестрикции фрагмент speC и плазмиду pTac15K соединяли друг с другом, получая, таким образом, рекомбинантный плазмидный вектор p15SpeC с небольшим числом копий.
4-3: Получение штамма WL3110/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм WL3110, приготовленный в примере 1-1, получая, таким образом, штамм WL3110/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-4: Получение штамма XQ17/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ17, приготовленный в примере 1-3, получая, таким образом, штамм XQ17/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-5: Получение штамма XQ22/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ22, приготовленный в примере 1-4, получая, таким образом, штамм XQ22/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-6: Получение штамма XQ26/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ26, приготовленный в примере 1-6, получая, таким образом, штамм XQ26/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-7: Получение штамма XQ33/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ33, приготовленный в примере 2-1, получая, таким образом, штамм XQ33/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-8: Получение штамма XQ37/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ37, приготовленный в примере 2-2, получая, таким образом, штамм XQ37/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-9: Получение штамма XQ39/pKKSpeC
Вектор pKKSpeC, приготовленный в примере 4-1, вводили в штамм XQ39, приготовленный в примере 2-3, получая, таким образом, штамм XQ39/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
4-10: Получение штамма XQ43/pKKSpeC
Вектор p15SpeC, приготовленный в примере 4-2, вводили в штамм XQ43, приготовленный в примере 2-4, получая, таким образом, штамм XQ43/pKKSpeC. Затем полученный штамм культивировали на агаризованной твердой среде, содержащей ампициллин, с отбором рекомбинантных мутантных микроорганизмов.
Пример 5: Продукция путресцина, используя мутантный микроорганизм, у которого амплифицирован ген (speC), кодирующий орнитиндекарбоксилазу
Каждый из мутантных штаммов, приготовленных в примере 4, культивировали в качалочной колбе, содержащей ту же среду, что и среда, описанная в примере 3. 100 мкл каждой клеточной культуры, сформированной в среде LB, инокулировали в минимальную среду и затем культивировали при 30οС при 220 об/мин в течение 30 часов до достижения значения ОП660, составляющего 5. Затем 1 мл культуральной жидкости добавляли в колбу перегородчатой конструкции на 350 мл, содержащую 50 мл такой же среды, и затем культивировали при 30οС при 220 об/мин в течение 27 часов. Клетки отделяли центрифугированием, и супернатант анализировали с помощью HPLC в тех же условиях, что и условия, описанные в примере 3. Результаты анализа представлены в таблице 2.
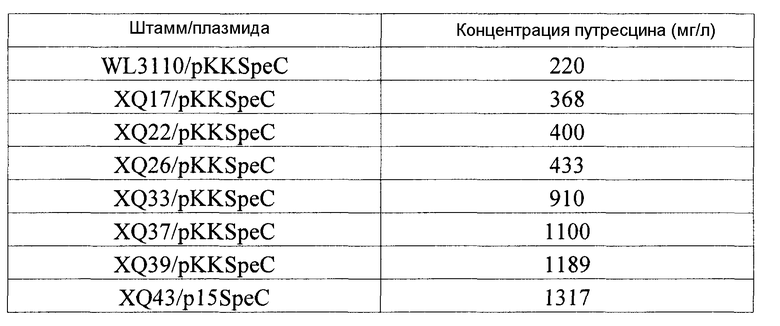
Как видно из таблицы 2, способности к продукции путресцина у мутантных микроорганизмов (WL3110/pKKSpeC, XQ17/pKKSpeC, XQ22/pKKSpeC, XQ26/pKKSpeC, XQ33/pKKSpeC, XQ37/pKKSpeC, XQ39/pKKSpeC и XQ43/p15SpeC), обладающих сниженными активностями в отношении деградации и утилизации путресцина, были значительно усилены по сравнению со способностями мутантных микроорганизмов (WL3110, XQ17, XQ22, XQ26, XQ33, XQ37, XQ39 и XQ43), представленных в таблице 1, в которые не вводили ни pKKSpeC, ни p15SpeC, при коэкспрессии с орнитиндекарбоксилазой speC.
Пример 6: Получение путресцина через подпитываемую культуру штамма XQ37/pKKSpeC
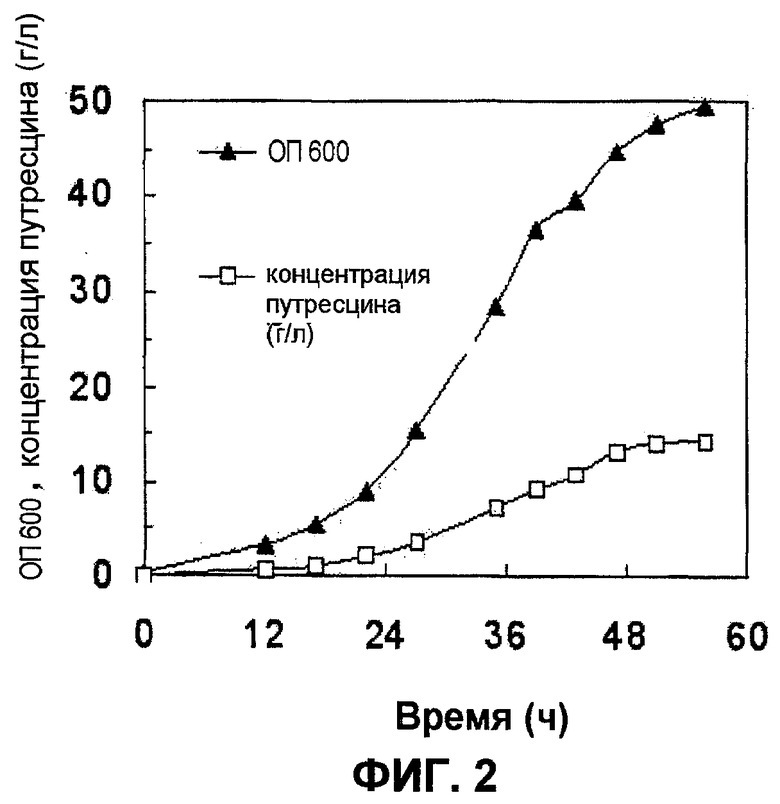
Потенциал сниженной активности в отношении деградации и утилизации путресцина анализировали вместе с декарбоксилазной активностью при ферментации с использованием подпитываемой культуры. Ферментацию с использованием подпитываемой культуры проводили в ферментере на 6,6 литров (Bioflo 3000; New Brunswick Scientific Co., Edison, NJ) после добавления 10 г/л глюкозы к 2 литрам минимальной среды R. 1 мл культуры XQ37/pKKSpeC, сформированной в среде LB, добавляли в колбу перегородчатой конструкции на 350 мл, содержащую 50 мл такой же среды, и затем культивировали при 30οС при 220 об/мин в течение 24 часов до достижения максимального значения ОП660, составляющего 5. 200 мл прекультуры впоследствии использовали для инокуляции в ферментер. Уровень растворенного кислорода в подвергнутой ферментации культуральной жидкости поддерживали с использованием 20%-насыщенного воздуха, автоматически увеличивая скорость перемешивания до 850 об/мин. Когда рН подвергнутой ферментации культуральной жидкости увеличивался на приблизительно 0,2 единицы рН от установленного рН 6,8 в результате истощения глюкозы, автоматически добавляли содержащий глюкозу раствор для увеличения концентрации глюкозы до более чем 3 г/л. Содержащий глюкозу раствор содержал 500 г/л глюкозы и 200 г/л (NH4)2SO4. На протяжении всего периода ферментации, за исключением короткого периода времени, когда рН увеличивался вследствие истощения глюкозы, рН подвергнутой ферментации культуральной жидкости поддерживали на уровне рН, составляющем 6,8, путем добавления 28% (в весовом отношении) раствора аммиака. Отбирали образцы подвергнутой ферментации культуральной жидкости, и их центрифугировали для отделения клеток, и супернатант анализировали с помощью HPLC таким же образом, как это описано в примере 3. Результаты анализов продемонстрированы на фиг.2. Как продемонстрировано на фиг.2, штамм XQ37/pKKSpeC продуцировал 14,3 г/л путресцина за 55,8 часа после инокуляции, и максимальная продукция путресцина составляла 0,28 г л-1 ч-1 через 47 часов после инокуляции.
Пример 7: Получение путресцина через подпитываемую культуру штамма XQ39
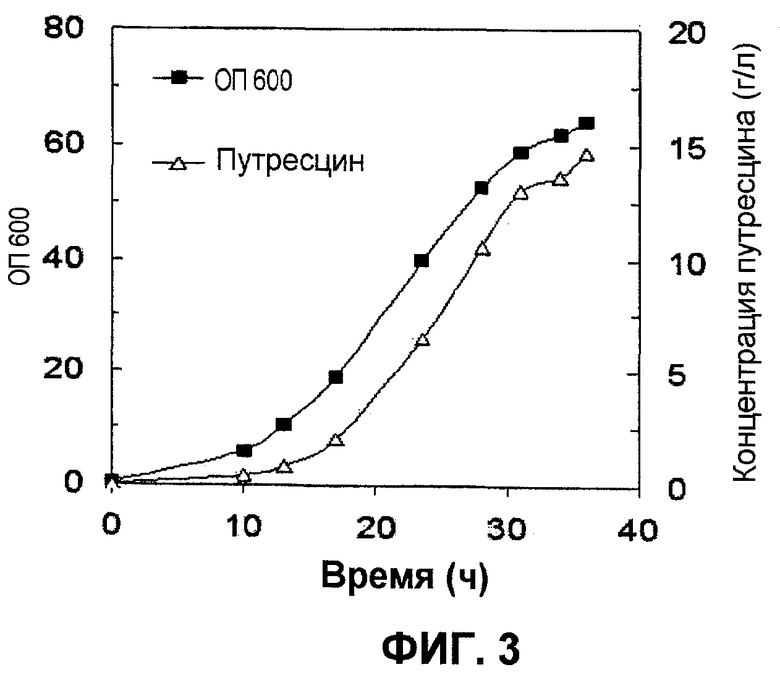
Ферментацию с использованием подпитываемой культуры проводили так же, как это описано в примере 6, за исключением использования штамма XQ37 вместо XQ37/pKKSpeC. Подвергнутую ферментации культуральную жидкость анализировали с помощью HPLC, и результаты анализов продемонстрированы на фиг.3. Как продемонстрировано на фиг.3, штамм XQ39 продуцировал 14,7 г/л путресцина за 36 часов после инокуляции, и максимальная продукция путресцина составляла 0,42 г л-1 ч-1 через 31 час после инокуляции.
Пример 8: Получение путресцина через подпитываемую культуру штамма XQ43
Штамм XQ43 культивировали в колбе с 50 мл минимальной среды R/2 (содержащей 2 г/л (NH4)2HPO4, 6,75 г/л KH2PO4, 0,85 г/л лимонной кислоты, 0,7 г/л MgSO4 .7H2O, 0,5% (в объемном отношении) раствора сверхредких металлов (Qian et al., Biotechnol. and Bioeng, 101(3): 587-601, 2008), дополненной 3 г/л (NH4)2SO4. Раствор сверхредких металлов содержал (на литр) 5 М HCl, 10 г FeSO4·7H2O, 2,25 г ZnSO4·7H2O, 1 г CuSO4·5H2O, 0,5 г MnSO4·5H2O, 0,23 г Na2B4O7·10H2O, 2 г CaCl2·2H2O и 0,1 г (NH4)6Mo7O24. Раствор, содержащий глюкозу (100 г/л), стерилизовали отдельно и добавляли в подвергнутую стерилизации среду до конечной концентрации 10 г/л. 1 мл культуры XQ43, сформированной в среде LB, добавляли в колбу перегородчатой конструкции на 350 мл, содержащую 50 мл описанной выше среды, и затем культивировали при 37οС при 220 об/мин в течение 24 часов до достижения значения ОП660 культуры, составляющего 3,3. 200 мл прекультуры впоследствии использовали для инокуляции в ферментер. Уровень растворенного кислорода в культуре поддерживали с использованием 20%-насыщенного воздуха, автоматически увеличивая скорость перемешивания до 1000 об/мин.
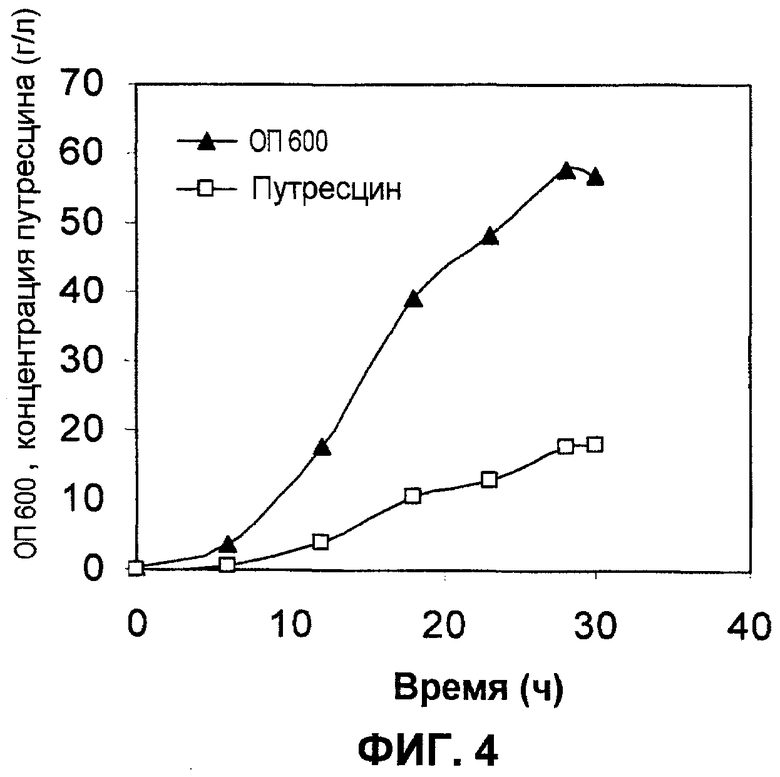
Ферментацию с использованием подпитываемой культуры штамма XQ43 проводили в ферментере на 6,6 литров (Bioflo 3000; New Brunswick Scientific Co., Edison, NJ) после добавления 10 г/л глюкозы. Когда рН подвергнутой ферментации культуральной жидкости увеличивался на приблизительно 0,01 единицы рН от установленного рН 6,8 в результате истощения глюкозы, автоматически добавляли содержащий глюкозу раствор для увеличения концентрации глюкозы до более чем 2 г/л. Содержащий глюкозу раствор содержал 522 г/л глюкозы, 8 г/л MgSO4 и 170 г/л (NH4)2SO4. На протяжении всего периода ферментации, за исключением короткого периода времени, когда рН увеличивался вследствие истощения глюкозы, рН подвергнутой ферментации культуральной жидкости поддерживали на уровне рН, составляющем 6,8, путем добавления 10М раствора KOH. Отбирали образцы подвергнутой ферментации культуральной жидкости, и их центрифугировали для отделения клеток, и супернатант анализировали с помощью HPLC таким же образом, как это описано в примере 3. Результаты анализов продемонстрированы на фиг.4. Как продемонстрировано на фиг.4, штамм XQ43 продуцировал 18 г/л путресцина за 30 часов после инокуляции, и максимальная продукция путресцина составляла 0,63 г л-1 ч-1 через 28 часов после инокуляции.
Пример 9: Получение путресцина через подпитываемую культуру штамма XQ43/p15SpeC
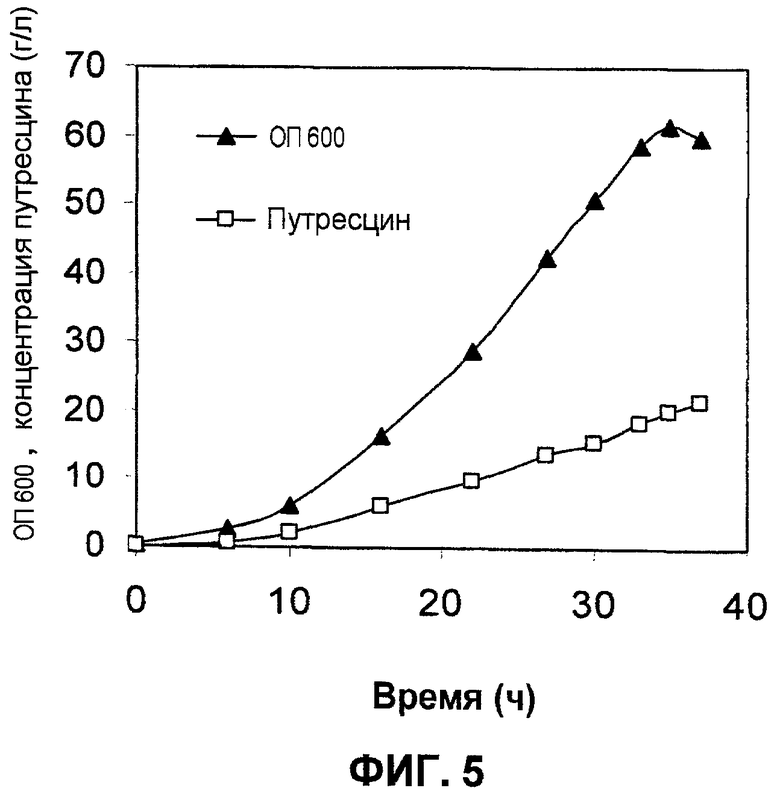
Ферментацию с использованием подпитываемой культуры проводили так же, как это описано в примере 8, за исключением использования штамма XQ43/p15SpeC вместо штамма XQ43. Подвергнутую ферментации культуральную жидкость анализировали с помощью HPLC, и результаты анализов продемонстрированы на фиг.5. Как продемонстрировано на фиг.5, штамм XQ43/p15SpeC продуцировал 21,7 г/л путресцина за 37 часов после инокуляции, и максимальная продукция путресцина составляла 0,58 г л-1 ч-1 через 37 часов после инокуляции.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Как подробно описано выше, настоящим изобретением предоставляются мутантные микроорганизмы, обладающие способностью к продукции путресцина. Эти мутантные микроорганизмы полезны для продукции путресцина в большом количестве, который используется в широком ряде промышленных применений.
Хотя настоящее изобретение было описано со ссылкой на конкретные иллюстративные варианты осуществления, оно не должно ограничиваться этими вариантами осуществления, а должно ограничиваться только прилагаемой формулой изобретения. Должно быть понятно, что квалифицированные в данной области техники специалисты могут изменить или модифицировать эти варианты осуществления, не отступая от объема и сущности настоящего изобретения.








Настоящее изобретение относится к мутантным микроорганизмам, обладающим способностью к повышенной продукции путресцина. Данные микроорганизмы были получены путем разрушения гена SpeE, и, по крайней мере, одного гена, выбранного из SpeG, ArgI или PuuP в микроорганизме, имеющем метаболический путь синтеза путресцина. Кроме того, предлагается способ получения этих микроорганизмов и способ продуцирования путресцина путем их культивирования. Предложенные мутантные микроорганизмы могут использоваться для крупномасштабной продукции путресцина для промышленного применения. 6 н. и 24 з.п. ф-лы, 5 ил., 2 табл.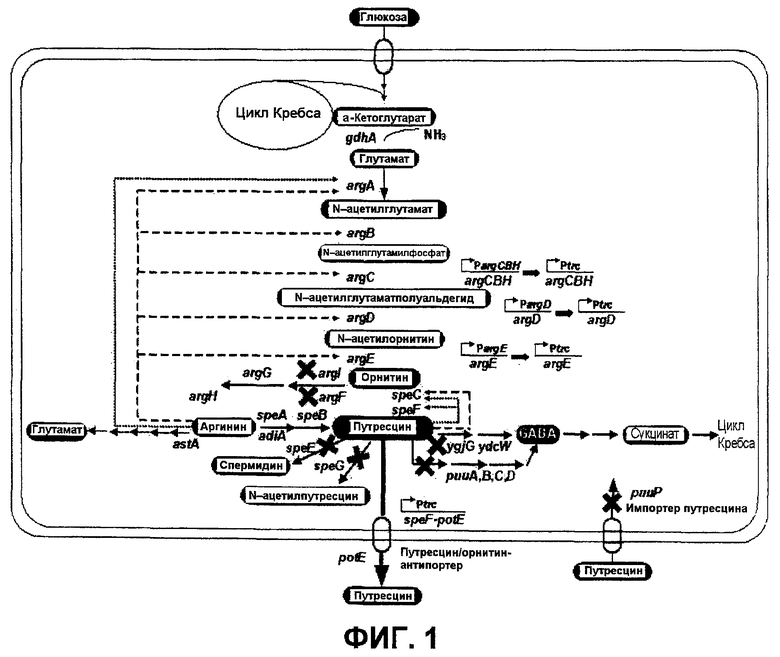
1. Мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором ген speE, кодирующий спермидинсинтазу, участвующую в пути деградации или утилизации путресцина, инактивирован или делетирован, и по крайней мере один ген, выбираемый из группы, состоящей из гена speG, кодирующего спермидин-N-ацетилтрансферазу, которая участвует в пути деградации или утилизации путресцина, гена argI, кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена puuP, кодирующего импортер путресцина, инактивирован или делетирован.
2. Мутантный микроорганизм по п.1, в котором дополнительно инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена puuA, кодирующего γ-глутамилпутресцинсинтазу, гена ygjG, кодирующего путресцин-трансаминазу, и гена argF, кодирующего мономерную цепь F орнитин-карбамоилтрансферазы.
3. Мутантный микроорганизм по п.1, в котором дополнительно делетирован ген lacI, кодирующий репрессор оперона lac, для усиления экспрессии генов, кодирующих ферменты, участвующие в биосинтезе путресцина.
4. Мутантный микроорганизм по п.1, в который дополнительно введен ген speC, кодирующий орнитиндекарбоксилазу, или в котором указанный ген дополнительно амплифицирован.
5. Мутантный микроорганизм по п.4, в который ген speC, кодирующий орнитиндекарбоксилазу, вводят в составе экспрессионного вектора, содержащего сильный промотор.
6. Мутантный микроорганизм по п.5, где сильный промотор выбирают из группы, состоящей из промотора trc, промотора tac, промотора Т7, промотора lac и промотора trp.
7. Мутантный микроорганизм по п.1, который выбирают из группы, состоящей из Bacillus sp., Corynebacterium sp., Escherichia sp., Pichia sp., Pseudomonas sp. и Saccharomyces sp.
8. Мутантный микроорганизм, обладающий способностью к продукции путресцина, в котором ген speE, кодирующий спермидинсинтазу, участвующую в пути деградации или утилизации путресцина, инактивирован или делетирован, и по крайней мере один ген, выбираемый из группы, состоящей из гена speG, кодирующего спермидин-N-ацетилтрансферазу, которая участвует в пути деградации или утилизации путресцина, гена argI, кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена рuuР, кодирующего импортер путресцина, инактивирован или делетирован, и в котором промотор по крайней мере одного гена, выбираемого из группы, состоящей из гена argECBH, кодирующего оперон для биосинтеза аргинина, гена argD, кодирующего ацетилорнитин-аминотрансферазу, и оперона speF-potE, кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, заменен сильным промотором.
9. Мутантный микроорганизм по п.8, в котором дополнительно инактивирован или делетирован по крайней мере один ген, выбираемый из группы, состоящей из гена puuA, кодирующего γ-глутамилпутресцинсинтазу, гена ygjG, кодирующего путресцин-трансаминазу, и гена argF, кодирующего мономерную цепь F орнитин-карбамоилтрансферазы.
10. Мутантный микроорганизм по п.8, в котором дополнительно делетирован ген lacI, кодирующий репрессор оперона lac, для усиления экспрессии генов, кодирующих ферменты, участвующие в биосинтезе путресцина.
11. Мутантный микроорганизм по п.8, в который дополнительно введен ген speC, кодирующий орнитиндекарбоксилазу, или в котором указанный ген дополнительно амплифицирован.
12. Мутантный микроорганизм по п.11, в который ген speC, кодирующий орнитиндекарбоксилазу, вводят в составе экспрессионного вектора, содержащего сильный промотор.
13. Мутантный микроорганизм по п.8 или 12, где сильный промотор выбирают из группы, состоящей из промотора trc, промотора tac, промотора Т7, промотора lac и промотора trp.
14. Мутантный микроорганизм по п.8, который выбирают из группы, состоящей из Bacillus sp., Corynebacterium sp., Escherichia sp., Pichia sp., Pseudomonas sp. и Saccharomyces sp.
15. Способ получения мутантного микроорганизма, обладающего способностью к продукции путресцина, включающий инактивацию или делетирование гена speE, кодирующего спермидинсинтазу, которая участвует в пути деградации или утилизации путресцина, и дополнительную инактивацию или делетирование по крайней мере одного гена, выбираемого из группы, состоящей из гена speG, кодирующего спермидин-N-ацетилтрансферазу, которая участвует в пути деградации или утилизации путресцина, гена argI, кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена рuuР, кодирующего импортер путресцина, в микроорганизме, имеющем путь синтеза путресцина.
16. Способ по п.15, дополнительно включающий инактивацию или делетирование по крайней мере одного гена, выбираемого из группы, состоящей из гена рuuА, кодирующего γ-глутамилпутресцинсинтазу, гена ygjG, кодирующего путресцин-трансаминазу, и гена argF, кодирующего мономерную цепь F орнитин-карбамоилтрансферазы.
17. Способ по п.15, дополнительно включающий делетирование гена lacI, кодирующего репрессор оперона lac, для усиления экспрессии генов, кодирующих ферменты, участвующие в биосинтезе путресцина.
18. Способ по п.15, дополнительно включающий до или после инактивации или делетирования введение или амплификацию гена speC, кодирующего орнитиндекарбоксилазу.
19. Способ по п.18, в котором ген speC, кодирующий орнитиндекарбоксилазу, вводят в составе экспрессионного вектора, содержащего сильный промотор.
20. Способ по п.19, в котором сильный промотор выбирают из группы, состоящей из промотора trc, промотора tac, промотора Т7, промотора lac и промотора trp.
21. Способ по п.15, в котором указанный микроорганизм выбирают из группы, состоящей из Bacillus sp., Corynebacterium sp., Escherichia sp., Pichia sp., Pseudomonas sp. и Saccharomyces sp.
22. Способ получения мутантного микроорганизма, обладающего способностью к продукции путресцина, включающий
a) инактивацию или делетирование гена speE, кодирующего спермидинсинтазу, которая участвует в пути деградации или утилизации путресцина, и дополнительную инактивацию или делетирование по крайней мере одного гена, выбираемого из группы, состоящей из гена speG, кодирующего спермидин-N-ацетилтрансферазу, которая участвует в пути деградации или утилизации путресцина, гена argI, кодирующего мономерную цепь I орнитин-карбамоилтрансферазы, и гена puuP, кодирующего импортер путресцина, в микроорганизме, имеющем путь синтеза путресцина, и
b) замену промотора по крайней мере одного гена, выбираемого из группы, состоящей из гена argECBH, кодирующего оперон для биосинтеза аргинина, гена argD, кодирующего ацетилорнитин-аминотрансферазу, и оперона speF-potE, кодирующего индуцируемую орнитиндекарбоксилазу и путресцин/орнитин-антипортер, сильным промотором.
23. Способ по п.22, в котором на стадии а) дополнительно инактивируют или делетируют по крайней мере один ген, выбираемый из группы, состоящей из гена рuuА, кодирующего γ-глутамилпутресцинсинтазу, гена ygjG, кодирующего путресцин-трансаминазу, и гена argF, кодирующего мономерную цепь F орнитин-карбамоилтрансферазы.
24. Способ по п.22, в котором на стадии а) дополнительно делетируют ген lacI, кодирующий репрессор оперона lac, для усиления экспрессии генов, кодирующих ферменты, участвующие в биосинтезе путресцина.
25. Способ по п.22, дополнительно включающий до стадии а), после стадии b) или между стадиями а) и b) введение или амплификацию гена speC, кодирующего орнитиндекарбоксилазу.
26. Способ по п.25, в котором ген speC, кодирующий орнитиндекарбоксилазу, вводят в составе экспрессионного вектора, содержащего сильный промотор.
27. Способ по п.22 или 26, в котором сильный промотор выбирают из группы, состоящей из промотора trc, промотора tac, промотора Т7, промотора lac и промотора trp.
28. Способ по п.22, в котором указанный микроорганизм выбирают из группы, состоящей из Bacillus sp., Corynebacterium sp., Escherichia sp., Pichia sp., Pseudomonas sp. и Saccharomyces sp.
29. Способ продуцирования путресцина, включающий культивирование мутантного микроорганизма по п.1 для продуцирования путресцина и получение путресцина из культуральной жидкости.
30. Способ продуцирования путресцина, включающий культивирование мутантного микроорганизма по п.8 для продуцирования путресцина и получение путресцина из культуральной жидкости.
| THOLL D | |||
| ET AL.: "Retarded growth of an Escherichia coli mutant deficient in spermidine synthase can be unspecifially repaired by addition of various polyamines", World journal of microbiology and biotechnology, 1998, v.14, p.857-863 | |||
| WO 2006005604, 19.01.2006 | |||
| KASHIWAGI K | |||
| ET AL.: "Coexistence of the genes for putrescine transport protein and |

