Область техники
Настоящее изобретение относится к рекомбинантному микроорганизму для продуцирования путресцина и способу получения путресцина с его использованием.
Предшествующий уровень техники
Биогенные амины (ВА) представляют собой азотистые соединения, которые получаются главным образом в результате декарбоксилирования аминокислот, аминирования альдегида и кетона и трансаминирования. Такие биогенные амины являются низкомолекулярными соединениями, которые синтезируются в ходе метаболизма микроорганизмами, растениями и животными, и поэтому они известны как компоненты, легко обнаруживаемые в их клетках. В частности, такие полиамины, как спермидин, спермин, путресцин (или 1,4-бутандиамин) и кадаверин, являются веществами, присутствующими в большинстве живых клеток.
Среди них путресцин является важным исходным веществом в синтезе полиамина найлона-4,6, осуществляемом путем взаимодействия с адипиновой кислотой, и его обычно получают путем химического синтеза из пропилена через акрилонитрил и сукцинонитрил.
Помимо этого описан способ получения путресцина в высокой концентрации с использованием трансформации Е. coli и микроорганизмов рода Corynebacterium (международная публикация заявки № WO06/005603; международная публикация заявки № WO09/125924; Qian ZD et al., Biotechnol. Bioeng., 104(4): 651-662, 2009; Schneider et al., Appl. Microbiol. Biotechnol., 88(4): 859-868, 2010; и Schneider et al., Appl. Microbiol. Biotechnol., 95: 169-178, 2012). Кроме того, было проведено активное исследование переносчика путресцина, относящегося к Е. coli, дрожжам, клеткам растений и животных (K. Igarashi, Plant Physiol. Biochem., 48: 506-512, 2010).
В то же время, авторы настоящего изобретения продемонстрировали, что мембранные белки, кодируемые NCgl2522, функционируют в качестве экспортера путресцина в микроорганизме рода Corynebacterium, который имеет путь синтеза путресцина, и подтвердили, что путресцин можно получать с высоким выходом посредством повышения активности NCgl2522 по сравнению с его эндогенной активностью (заявка на патент Кореи (KR) №10-2013-0030020).
Кроме того, ген NCgl2523 представляет собой транскрипционный фактор, связанный с множественной лекарственной устойчивостью, принадлежащий семейству TetR и, как известно, действующий в качестве ингибитора экспрессии NCgl2522 (Hirochi et al., J. Biol. Chem., 280: 46, 38711-38719, 2005).
Авторы настоящего изобретения в течение длительного времени проводили исследования в этом направлении и подтвердили, что продуцирование путресцина усиливается, аналогично эффектам повышения активности NCgl2522, в результате истощения NCgl2523, который входит в состав оперона NCgl2522, с решением тем самым задачи настоящего изобретения.
Описание
Техническая проблема
Задача настоящего изобретения заключается в том, чтобы предложить рекомбинантный микроорганизм, способный продуцировать путресцин с высоким выходом.
Другой задачей настоящего изобретения является предложение способа получения путресцина с высоким выходом с использованием данного микроорганизма.
Техническое решение
Согласно одному из аспектов настоящего изобретения, для решения указанных выше задач предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, в котором инактивирован белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 2.
В одном из типичных воплощений настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, причем в данный микроорганизм дополнительно введена активность орнитиндекарбоксилазы (ODC).
В другом типичном воплощении настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, в котором ODC имеет аминокислотную последовательность, представленную SEQ ID NO: 10.
В еще одном типичном воплощении настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, причем в данном микроорганизме дополнительно инактивирована ацетилтрансферазная активность.
В еще одном воплощении настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, где ацетилтрансфераза содержит аминокислотную последовательность, представленную SEQ ID NO: 15 или 16.
В еще одном воплощении настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, причем данный микроорганизм представляет собой Corynebacterium glutaminicum.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина, включающий:
1) культивирование микроорганизма рода Corynebacterium, обладающего путресцин-продуцирующей способностью, в котором инактивирован белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 2, в культуральной среде; и
2) выделение путресцина из подвергнутого культивированию микроорганизма или культуральной среды, полученных на указанной выше стадии.
В типичном воплощении настоящего изобретения предложен способ получения путресцина, в котором микроорганизм рода Corynebacterium представляет собой Corynebacterium glutaminicum.
Далее, настоящее изобретение будет описано подробно.
Согласно одному из аспектов настоящее изобретение относится к микроорганизму рода Corynebacterium, обладающему путресцин-продуцирующей способностью, в котором инактивирован белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 2, такой как NCgl2523.
Использованный в данном описании термин «NCgl2523» относится к связанному с множественной лекарственной устойчивостью транскрипционному фактору, принадлежащему семейству TetR и, как известно, действующему в качестве ингибитора экспрессии NCgl2522 (Hirochi et al., J. Biol. Chem., 280: 46, 38711-38719, 2005).
В настоящем изобретении NCgl2523 представляет собой белок, имеющий аминокислотную последовательность SEQ ID NO: 2 или аминокислотную последовательность, имеющую гомологию с этой последовательностью, составляющую 70% или более, более конкретно 80% или более, еще конкретнее 90% или более, еще конкретнее 98% или более и наиболее конкретно 99% или более, и не ограничивающуюся таковой, при условии, что он представляет собой белок, обладающей активностью ингибитора экспрессии NCgl2522.
Кроме того, поскольку аминокислотные последовательности белков, демонстрирующих такую активность, могут отличаться в зависимости от вида или штамма микроорганизмов, это не является ограничением. Очевидно, что белок, имеющий аминокислотную последовательность, часть которой изменена с использованием делеций, модификаций, замен или вставок, включен в объем настоящего изобретения, при условии, что белок с последовательностью, имеющей гомологию с данной последовательностью, демонстрирует биологическую активность, по существу эквивалентную или соответствующую активности белка с SEQ ID NO: 2.
Использованный в данном описании термин «гомология» относится к сходству между заданными аминокислотными последовательностями или нуклеотидными последовательностями, и гомология может быть выражена в процентах. В данной заявке, гомологию последовательности, обладающей идентичной или схожей активностью с заданной аминокислотной последовательностью или нуклеотидной последовательностью, выражают в виде «% гомологии». Например, гомология может быть установлена с использованием традиционного программного обеспечения, с помощью которого рассчитывают относящиеся к ней параметры, такие как балл, идентичность, сходство и т.д., и более конкретно BLAST 2.0, либо путем сравнения последовательностей в эксперименте с использованием гибридизации по Саузерну в условиях определенной жесткости. Определение соответствующих условий гибридизации не выходит за рамки объема данной области техники и может быть выполнено средним специалистом в данной области техники с использованием известных методов (т.е. J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
При том условии, что он кодирует белок, имеющий активность, схожую с активностью белков NCgl2523, полинуклеотид, кодирующий NCgl2523 по настоящему изобретению, может кодировать аминокислотную последовательность SEQ ID NO: 2, или полинуклеотид, кодирующий белок, имеющий гомологию с данной последовательностью, составляющую 70% или более, конкретно 80% или более, более конкретно 90% или более, еще более конкретно, гомологию с ней, составляющую 95% или более, еще более конкретно 98% или более и наиболее конкретно 99% или более, может, в частности, включать нуклеотидную последовательность с SEQ ID NO: 1 или 3.
Кроме того, полинуклеотид, кодирующий NCgl2523 по настоящему изобретению, может гибридизоваться в жестких условиях с зондом с нуклеотидной последовательностью, представленной SEQ ID NO: 1 или 3, или с зондом, происходящим из этой нуклеотидной последовательности, и может представлять собой вариант, кодирующий функционально нормальный NCgl2523. Использованный в данном описании термин «жесткие условия» относится к условиям, в которых возможно осуществление специфической гибридизации между полинуклеотидами. Например, такие жесткие условия описаны подробно в литературе (т.е. J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
В настоящем изобретении было подтверждено, что, когда NCgl2523, который входит в состав оперона NCgl2522, был делетирован в микроорганизме рода Corynebacterium, обладающем путресцин-продуцирующей способностью, продуцирование путресцина усиливалось аналогично тому, как это происходит в случае повышения активности NCgl2522. В связи с этим, согласно настоящему изобретению может быть предложен рекомбинантный микроорганизм, демонстрирующий способность продуцировать путресцин с высоким выходом благодаря ингибирования экспрессии NCgl2522 в результате инактивации активности NCgl2523. Таким образом, как раскрыто в этом воплощении настоящего изобретения, инактивируя соответствующий NCgl2523 и гены, кодирующие схожие с ним аминокислотные последовательности, можно повысить путресцин-продуцирующую способность микроорганизма, в котором имеются аминокислотные последовательности, схожие с последовательностью NCgl2523, где такие аминокислотные последовательности состоят из NCgl2522 и оперона, или повысить путресцин-продуцирующую способность микроорганизма, в котором NCgl2523 действует аналогично модулятору экспрессии NCgl2522.
Использованный в данном описании термин «инактивация» относится к неосуществлению экспрессии гена, кодирующего соответствующий полипептид, к демонстрации некоторого ослабления генной экспрессии, неосуществлению продуцирования соответствующего функционального полипептида, даже в случае его экспрессии.
Помимо этого, термин «инактивация» относится не только к полной инактивации гена, кодирующего соответствующий полипептид, но также к ослаблению или существенному снижению экспрессии по сравнению с диким типом, в результате чего данный ген практически не экспрессируется. Таким образом, инактивация гена может быть полной (нокаут (К/О)) или частичной (например, с получением гипоморфа, для которого продемонстрирован уровень генной экспрессии ниже нормального, или с получением продукта мутантного гена, для которого продемонстрировано частичное снижение активности в результате образования гипоморфа).
В частности, в настоящем изобретении инактивация NCgl2523 может быть вызвана посредством:
1) частичного или полного делетирования полинуклеотида, кодирующего данный белок;
2) модификации регуляторной последовательности с целью ослабления экспрессии этого полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме с целью снижения активности белка; и
4) их комбинации,
но конкретно этим не ограничивается.
1) Частичное или полное делетирование полинуклеотида, кодирующего данный белок, может быть осуществлено путем замены полинуклеотида, кодирующего эндогенные целевые белки, на хромосомы с частично удаленным полинуклеотидом или маркерным геном с использованием вектора для встраивания в хромосому микроорганизма. Термин «частичный» может пониматься по-разному в зависимости от типа полинуклеотидов, но относится, в частности, к 1-300, более конкретно 1-100 и еще конкретнее 1-50 нуклеотидам.
Использованный в данном описании термин «вектор» относится к конструкции на основе ДНК, включающей нуклеотидную последовательность, кодирующую желаемый белок, функционально связанную с соответствующей регулирующей экспрессию последовательностью с целью экспрессии желаемого белка в подходящей клетке хозяина. Такая регуляторная последовательность включает промотор, который может инициировать транскрипцию, возможно последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы с мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После введения вектора посредством трансформации в подходящую клетку хозяина он может реплицироваться или функционировать независимо от генома хозяина и может быть встроен в сам геном.
Вектор, используемый в настоящем изобретении, не имеет конкретных ограничений при условии, что он способен реплицироваться в клетках-хозяинах, и можно использовать любой вектор, известный в данной области техники. Примеры традиционных векторов могут включать существующую в природе или рекомбинантную плазмиду, космиду, вектор на основе вируса и бактериофага. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A, а плазмиды pBR типа, pUC типа, pBluescriptll типа, pGEM типа, pTZ типа, pCL типа и рЕТ типа и т.д. можно использовать в качестве плазмидного вектора. Вектор, который можно использовать в настоящем изобретении, не имеет конкретных ограничений, и можно использовать любой известный экспрессирующий вектор. В частности, может быть использован вектор pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 или pCC1BAC.
Кроме того, полинуклеотид, кодирующий желаемый белок в хромосомах, можно заменить на мутированный полинуклеотид, используя вектор для встраивания в хромосому. Вставка полинуклеотида в хромосому может быть осуществлена любым методом, известным в данной области техники, таким как метод гомологической рекомбинации, но конкретно этим не ограничивается.
Использованный в данном описании термин «трансформация» относится к введению вектора, содержащего полинуклеотид, кодирующий целевые белки, в клетки-хозяева таким образом, чтобы белки, кодируемые этим полинуклеотидом, экспрессировались в клетках-хозяевах. При условии, что введенный посредством трансформации полинуклеотид может экспрессироваться в клетке-хозяине, он относится к любому из двух вариантов, например, он встроен в хромосомы клеток-хозяев или существует экстрахромосомно. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевые белки. Полинуклеотид можно вводить в любой форме, при условии, что он может быть введен в клетки-хозяева и экспрессирован в них. Например, полинуклеотид может быть введен в клетки-хозяева в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую в себя все элементы, необходимые для ее автономной экспрессии. Обычно, экспрессионная кассета включает в себя промотор, функционально связанный с полинуклеотидом, сигналы терминации транскрипции, сайты связывания рибосомы или сигналы терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося экспрессирующего вектора. Помимо этого, полинуклеотид может быть введен в клетки-хозяева в том виде, как он есть, и может быть функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине, но этим не ограничивается.
Кроме того, термин «функционально связанный» относится к функциональной связи между полинуклеотидной последовательностью, кодирующей желаемые белки, и последовательностью промотора, с использованием которой инициируется и опосредуется транскрипция данной полинуклеотидной последовательности.
Далее, 2) модификация регуляторной последовательности для ослабления экспрессии полинуклеотида может быть осуществлена путем введения модификации в регуляторную последовательность посредством делеции, вставки, неконсервативной или консервативной замены в нуклеотидной последовательности или их комбинации либо путем замены на нуклеотидную последовательность с более низкой активностью, с целью снижения активности регуляторной последовательности, но конкретно этим не ограничивается. Регуляторная последовательность может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, но этим не ограничивается.
Кроме того, 3) модификация полинуклеотидной последовательности в хромосоме может быть осуществлена путем введения модификации в регуляторную последовательность посредством делеции, вставки, неконсервативной или консервативной замены в нуклеотидной последовательности или их комбинации либо путем замены регуляторной последовательности на нуклеотидную последовательность с более низкой активностью, с целью снижения активности белка, но этим не ограничивается.
Использованный в данном описании термин «микроорганизм, обладающий путресцин-продуцирующей способностью» или «микроорганизм, продуцирующий путресцин» относится к микроорганизму, от природы обладающему путресцин-продуцирующей способностью, или к микроорганизму, полученному путем придания способности продуцировать путресцин родительскому штамму, не обладающему путресцин-продуцирующей способностью.
Микроорганизм, продуцирующий путресцин, может представлять собой, но не ограничивается им, микроорганизм, обладающий улучшенной способностью к продуцированию орнитина, используемому в качестве исходного вещества для биосинтеза путресцина, причем данный микроорганизм модифицирован таким образом, что имеет более высокие активности ацетилглутаматсинтазы, катализирующей превращение глутамата в ацетилглутамат (N-ацетилглутамат), или орнитинацетилтрансферазы (ArgJ), катализирующей превращение ацетилорнитина в орнитин, ацетилглутаматкиназы (ArgB), катализирующей превращение ацетилглутамата в ацетил тутам ил фосфат (N-ацетилглутамилфосфат), ацетил-гамма-глутамил-фосфатредуктазы (ArgC), катализирующей превращение ацетилглутамилфосфата в ацетилглутамат-полуальдегид (N-ацетилглутамат-полуальдегид), или ацетилорнитинаминотрансферазы (ArgD), катализирующей превращение ацетилглутамат-полуальдегида в ацетилорнитин (N-ацетилорнитин), по сравнению с эндогенной активностью с целью усиления пути биосинтеза от глутамата до орнитина глутамата.
Кроме того, данный микроорганизм модифицирован таким образом, чтобы устранить эндогенные активности орнитинкарбамоилтрансферазы (ArgF), вовлеченной в синтез аргинина из орнитина, экспортера глутамата (NCgl1221) и/или ацетилтрансферазы, которая катализирует ацетилирование путресцина, и/или модифицирован с целью введения активности орнитиндекарбоксилазы (ODC).
В данной заявке, орнитинкарбамоилтрансфераза (ArgF), экспортер глутамата (NCgl1221), орнитиндекарбоксилаза (ODC), ацетил-гамма-глутамилфосфатредуктаза (ArgC), ацетилглутаматсинтаза или орнитинацетилтрансфераза (ArgJ), ацетилглутаматкиназа (ArgB) и ацетилорнитинаминотрансфераза (ArgD) могут включать, в частности, аминокислотную последовательность, каждая их которых представлена SEQ ID NO: 8, 9, 10, 11, 12, 13 и 14, или аминокислотную последовательность, имеющую с этой последовательностью гомологию, составляющую 70% или более, более конкретно 80% или более, еще конкретнее 90% или более, но конкретно этим не ограничивается.
Кроме того, ацетилтрансфераза, которая катализирует ацетилирование путресцина, может включать, в частности, аминокислотную последовательность, представленную SEQ ID NO: 15 или 16, или аминокислотную последовательность, имеющую с этой последовательностью гомологию, составляющую 70% или более, более конкретно 80% или более, еще конкретнее 90% или более, но конкретно этим не ограничивается.
В частности, повышение активности в настоящем изобретении может быть осуществлено посредством:
1) увеличения числа копий полинуклеотидов, кодирующих фермент;
2) модификации регуляторной последовательности для усиления экспрессии данного полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме для повышения активности этого фермента; или
4) улучшающей модификации с использованием их комбинации,
но этим не ограничивается.
1) Увеличение числа копий полинуклеотидов может быть осуществлено посредством использования формы, функционально связанной с вектором, или посредством встраивания в хромосомы клеток хозяина, но конкретно этим не ограничивается. В частности, это может быть осуществлено путем введения в клетки-хозяева вектора, с которым функционально связан полинуклеотид, кодирующий фермент по настоящему изобретению, и который может реплицироваться и функционировать независимо от клеток-хозяев, или путем введения в клетки-хозяева вектора, с которым функционально связан полинуклеотид и который способен обеспечить встраивание данного полинуклеотида в хромосомы клеток-хозяев, в результате чего увеличивается число копий полинуклеотидов в хромосомах клеток-хозяев.
Далее, 2) модификация регуляторной последовательности для усиления экспрессии полинуклеотидов может быть осуществлена путем введения модификации в последовательность посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации либо путем замены на нуклеотидную последовательность с повышенной активностью, с целью повышения активности регуляторной последовательности, но конкретно этим не ограничивается. Регуляторная последовательность может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, и т.д., но конкретно этим не ограничивается.
Сильный гетерологичный промотор может быть присоединен вместо исходного промотора в обратном направлении по отношению к полинуклеотид-экспрессирующей единице. Примерами сильного промотора являются промотор CJ7, промотор lysCPI, промотор EF-Tu, промотор groEL, промотор асеА или асеВ и т.д., и более конкретно, могут представлять собой промотор, происходящий из коринебактерий, промотор lysCPI (WO 2009/096689) или промотор CJ7 (патент KR №0620092 и заявка KR WO 2006/065095), функционально связанный с полинуклеотидом, кодирующим фермент, и усиливающий его экспрессию, но ими не ограничиваются.
Кроме того, 3) модификация полинуклеотидной последовательности в хромосоме может быть осуществлена путем введения модификации в регуляторную последовательность посредством делеции, вставки, неконсервативной или консервативной замены в нуклеотидной последовательности или их комбинации либо путем замены на полинуклеотидную последовательность, модифицированную для приобретения повышенной активности, с целью повышения активности полинуклеотидной последовательности, но конкретно этим не ограничивается.
Кроме того, инактивация орнитинкарбамоилтрансферазы (ArgF), экспортера глутамата и ацетилтрансферазы может быть осуществлена методами инактивации NCgl2523, которые упомянуты выше:
1) частичного или полного делетирования полинуклеотида, кодирующего данный белок;
2) модификации регуляторной последовательности с целью ослабления экспрессии этого полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме с целью снижения активности белка; и
4) их комбинации,
но конкретно этим не ограничивается.
Что касается микроорганизма по настоящему изобретению, то он представляет собой микроорганизм, обладающий путресцин-продуцирующей способностью и включает прокариотические микроорганизмы, экспрессирующие белки, включающие аминокислотную последовательность SEQ ID NO: 2. Примерами таких микроорганизмов являются микроорганизмы Escherichia sp., Shigella sp., Citrobacter sp., Salmonella sp., Enterobacter sp., Yersinia sp., Klebsiella sp., Erwinia sp., Corynebacterium sp., Brevibacterium sp., Lactobacillus sp., Selenomanas sp., V/b/7'0 sp., Pseudomonas sp., Streptomyces sp., Arcanobacterium sp., Alcaligenes sp. и т.д. Микроорганизм по настоящему изобретению может представлять собой, в частности, микроорганизм рода Corynebacterium и более конкретно, Corynebacterium glutamicum, но им не ограничивается.
В одном из типичных воплощений настоящего изобретения используются штаммы, обладающие способностью продуцировать путресцин в высокой концентрации благодаря усилению пути биосинтеза путресцина из глутамата, которые представляют собой микроорганизмы рода Corynebacterium, депонированные с №№ доступа КССМ11138Р (патент KR №2012-0064046) и КССМ11240Р (патент KR 2012-0003634).
В другом типичном воплощении настоящего изобретения используются КССМ11138Р и КССМ11240Р, представляющие собой путресцин-продуцирующие штаммы, происходящие из Corynebacterium glutaminicum АТСС13032, и DAB12-a (патент KR №2013-0082478) и DAB12-D (KR №2013-0082478; DAB12-a ΔNCgl1469), представляющие собой путресцин-продуцирующие штаммы, происходящие из Corynebacterium glutaminicum АТСС13869, при этом все они имеют идентичные генотипы. Штамм АТСС13869 может быть получен из Американской коллекции типовых культур (АТСС).
В частности, авторы настоящего изобретения обозначили микроорганизм рода Corynebacterium, обладающий повышенной путресцин-продуцирующей способностью благодаря инактивации активности NCgl2523 в Corynebacterium glutaminicum КССМ11240Р, который представляет собой путресцин-продуцирующий штамм, как Corynebacterium glutaminicum СС01-0844 и депонировали его в Корейский центр культур микроорганизмов (КССМ) в соответствии с Будапештским договором 25 февраля 2014 г. с номером доступа КССМ11520Р.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина, включающий:
1) культивирование микроорганизма рода Corynebacterium, обладающего повышенной путресцин-продуцирующей способностью, в котором инактивирован белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 2, в культуральной среде; и
2) выделение путресцина из подвергнутого культивированию микроорганизма или культуральной среды, полученных на указанной выше стадии.
В данном способе культивирование микроорганизма может быть осуществлено с использованием известных методов периодического культивирования, методов непрерывного культивирования и методов периодического культивирования с подпиткой и т.д., но конкретно этим не ограничивается. В данной заявке культивирование можно поддерживать в условиях оптимальных значений рН (например, рН 5-9, конкретно рН 6-8 и наиболее конкретно рН 6,8), используя основные соединения (например, гидроксид натрия, гидроксид калия или аммиак) или кислотные соединения (например, фосфорную кислоту или серную кислоту), и в аэробных условиях, добавляя к клеточной культуре кислород или газовую смесь, содержащую кислород, но конкретно этим не ограничиваясь. В процессе культивирования можно поддерживать температуру 20-45°С и конкретно 25-40°С, и культивирование проводить в течение примерно 10-160 часов. Путресцин, образованный в результате культивирования, может быть экспортирован в культуральную среду или может оставаться в клетках.
Кроме того, используемая культуральная среда может включать в себя сахар и углевод (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масло и жир (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирную кислоту (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирт (например, глицерин и этанол) и органическую кислоту (например, уксусную кислоту), по отдельности или в комбинации, в качестве источника углерода; азот-содержащие органические соединения (например, пептон, дрожжевой экстракт, мясной сок, экстракт солода, кукурузный раствор, порошок соевой муки и мочевину) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), по отдельности или в комбинации, в качестве источника азота; дигидрофосфат калия, гидрофосфат калия или соответствующую им натрий-содержащую соль, по отдельности или в комбинации, в качестве источника фосфора; и другие необходимые стимулирующие рост вещества, включая соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины, но этим не ограничивается.
Выделение путресцина, образованного на стадии культивирования по настоящему изобретению, может быть выполнено с использованием подходящего, известного в данной области техники способа, в зависимости от способов культивирования, таких как способы периодического, непрерывного или периодического культивирования с подпиткой и т.д., со сбором, при этом, желаемых аминокислот из культуральной жидкости.
Полезные эффекты
Микроорганизм рода Corynebacterium, обладающий повышенной путресцин-продуцирующей способностью, по настоящему изобретению модифицирован с целью инактивации активности белка, включающего аминокислотную последовательность SEQ ID NO: 2, тем самым индуцируя повышенную активность белка, который, как ожидается, является экспортером путресцина, конкретно NCgl2522, и усиливая экспорт путресцина во внеклеточное пространство, и вследствие этого может эффективно продуцировать путресцин.
Описание графических материалов
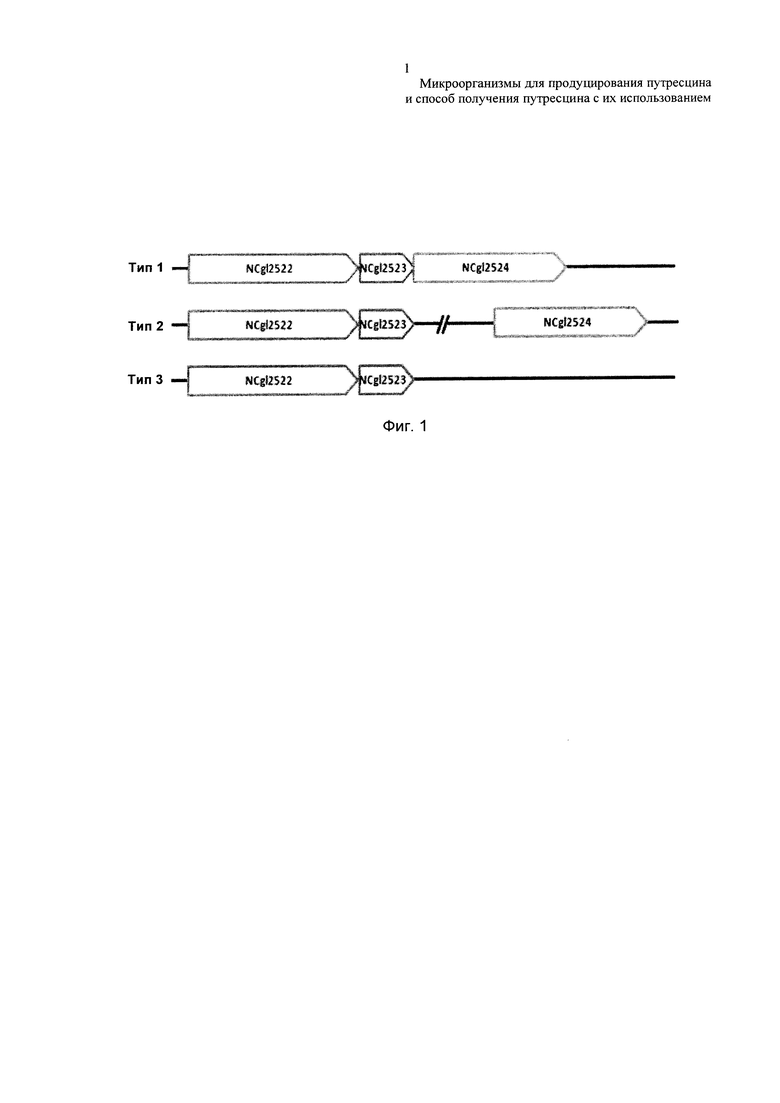
Фиг. 1 представляет собой схематическую диаграмму, показывающую комбинированные структуры из NCgl2523 и соседних единиц. В частности, на Фиг. 1 представлена схематическая диаграмма, показывающая комбинированную структуру NCgl2522-NCgl2523-NCgl2524, представляющую собой комбинированную структуру соседних генов микроорганизма, включающую NCgl2523 (тип 1); структуру, составленную из комбинированных NCgl2522-NCgl2523 и расположенного отдельно NCgl2524 (тип 2); или комбинированную структуру NCgl2522-NCgl2523 (тип 3).
Способ осуществления изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако эти примеры приведены только в целях иллюстрации, и не подразумевается, что данное изобретение ограничено этими примерами.
Пример 1. Получение штаммов с делецией NCgl2523 и проверка их путресцин-продуцирующей способности
1-1. Получение штаммов с делецией NCgl2523 на основе путресцин-продуцирующих штаммов, происходящих из АТСС13032
Для проверки того, оказывает ли делеция NCgl2523, происходящего из Corynebacterium glutaminicum АТСС13032, влияние на путресцин-продуцирующую способность, готовили векторы для делеции гена, кодирующего NCgl2523.
В частности, с учетом нуклеотидной последовательности гена, кодирующего NCgl2523, представленной SEQ ID NO: 1, конструировали пару праймеров с SEQ ID NO: 4 и 5 для получения гомологичных рекомбинантных фрагментов, соответствующих N-концевому участку NCgl2523, и пару праймеров с SEQ ID NO: 6 и 7 для получения гомологичных рекомбинантных фрагментов, соответствующих С-концевому участку NCgl2523, которые приведены ниже в Таблице 1.
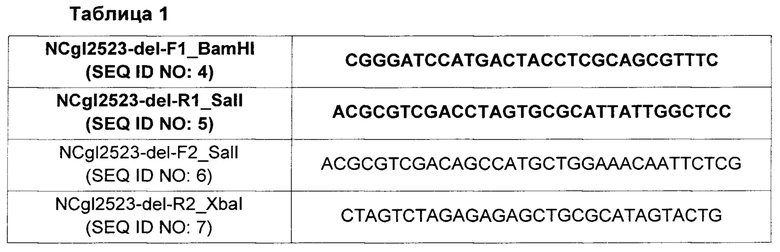
Проводили полимеразную цепную реакцию (ПЦР), используя геномную ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы и две пары праймеров, амплифицируя тем самым ПЦР-фрагменты гена NCgl2523, соответствующие N-концевому и С-концевому участкам. С использованием электрофореза этих ПЦР-фрагментов получали желаемые фрагменты. В данной заявке ПЦР реакцию повторяли, применяя 30 циклов, включающих в себя денатурацию в течение 30 секунд при 95°С, отжиг в течение 30 секунд при 55°С и удлинение в течение 30 секунд при 72°С. Полученные таким образом фрагменты, соответствующие N-концевому и С-концевому участкам, обрабатывали ферментами рестрикции BamHI и SaiI и ферментами рестрикции SalI и XbaI, соответственно. Обработанные фрагменты клонировали в векторы pDZ, обработанные ферментами рестрикции BamHI и XbaI, конструируя тем самым плазмиды pDZ-1'NCgl2523(K/O).
Плазмиды pDZ-1'NCgl2523(K/O) вводили путем трансформации в каждый из штаммов КССМ11138Р (патент KR №10-1348461) и КССМ11240Р (патент KR №2013-0082478), используя электропорацию, с получением трансформантов. Для образования колоний трансформанты высевали и культивировали на чашках с BHIS (дополненной сердечно-мозговым экстрактом) средой (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащей канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид). Среди образованных колоний отбирали голубые колонии, как колонии штаммов, в которые введены плазмиды pDZ-1'NCgl2523(K/O).
Отобранные штаммы культивируют в СМ среде (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л) и рН 6,8) при 30°С в течение 8 часов. После проведения серийного разведения в диапазоне от 10-4 до 10-10 для каждой клеточной культуры разведенные образцы высевали на чашки и культивировали на X-gal-содержащей твердой среде для образования колоний. Среди образованных колоний окончательно отбирали белые колонии, которые появлялись с относительно низкой частотой, в качестве колоний штаммов с делецией гена, кодирующего NCgl2523, в результате вторичного кроссинговера.
В окончательно отобранных штаммах проводили ПЦР, используя пару праймеров с SEQ ID NO: 4 и 7, для подтверждения делеции гена, кодирующего NCgl2523. Каждый мутант Corynebacterium glutamicum обозначали как КССМ11138Р ΔNCgl2523 и КССМ11240Р ΔNCgl2523.
1-2. Получение штаммов с делецией NCgl2523 на основе путресцин-продуцирующих штаммов, происходящих из АТСС13869
Штаммы с делецией NCgl2523 конструировали на основе DAB12-a (патент KR №2013-0082478) и DAB12-D (патент KR №2013-0082478; DAB12-a ANCgl1469), которые представляют собой путресцин-продуцирующие штаммы, происходящие из Corynebacterium glutaminicum АТСС13869.
В частности, для проверки последовательности гена NCgl2523 и экспрессированного на его основе белка, происходящего из Corynebacterium glutaminicum АТСС13869, проводили ПЦР, используя геномную ДНК Corynebacterium glutaminicum АТСС13869 в качестве матрицы и пару праймеров с SEQ ID NO: 4 и 7. В данной заявке ПЦР реакцию повторяли, применяя 30 циклов, включающих денатурацию в течение 30 секунд при 95°С, отжиг в течение 30 секунд при 55°С и удлинение в течение 1 минуты 30 секунд при 72°С.
В результате разделения посредством электрофореза полученных таким образом продуктов ПЦР и анализа путем секвенирования обнаруживали, что ген, кодирующий NCgl2523, происходящий из Corynebacterium glutaminicum АТСС13869, включает нуклеотидную последовательность, представленную SEQ ID NO: 3. Кроме того, сравнивали аминокислотную последовательность белков, кодируемых этим геном, с аминокислотной последовательностью NCgl2523 (SEQ ID NO: 2), происходящего из Corynebacterium glutaminicum ATCC13032, и в результате показано 100% гомологии.
Чтобы делетировать ген, кодирующий NCgl2523, происходящий из Corynebacterium glutaminicum АТСС 13869, проводили ПЦР, используя геномную ДНК Corynebacterium glutaminicum АТСС 13869 в качестве матрицы и две пары праймеров, описанных в Таблице 1, приведенной в примере 1-1, и амплифицировали ПЦР-фрагменты гена NCgl2523, соответствующие N-концевому и С-концевому участкам, получая в результате электрофореза желаемые фрагменты. В данной заявке ПЦР реакцию повторяли, применяя 30 циклов, включающих денатурацию в течение 30 секунд при 95°С, отжиг в течение 30 секунд при 55°С и удлинение в течение 30 секунд при 72°С. Полученные фрагменты, соответствующие N-концевому и С-концевому участкам, обрабатывали ферментами рестрикции BamHI и SalI и ферментами рестрикции SalI и XbaI, соответственно. Обработанные фрагменты клонировали в векторы pDZ, обработанные ферментами рестрикции BamHI и XbaI, конструируя таким образом плазмиды pDZ-2'NCgl2523(K/O).
После введения путем трансформации pDZ-2'NCgl2523(K/O) в каждый из штаммов Corynebacterium glutaminicum DAB12-a и DAB12-b аналогично тому, как описано в примере 1-1, отбирали штаммы с делецией гена, кодирующего NCgl2523. Отобранные мутанты Corynebacterium glutaminicum обозначали как DAB12-a ΔNCgl2523 и DAB12-D ΔNCgl2523.
1-3. Оценка путресцин-продуцирующей способности штаммов с делецией NCgl2523
Чтобы проверить влияние делеции NCgl2523 на продуцирование путресцина путресцин-продуцирующими штаммами, сравнивали мутанты Corynebacterium glutaminicum, сконструированные, как описано в примере 1-1 и 1-2, в отношении путресцин-продуцирующей способности.
В частности, каждый из 4 разных типов мутантов Corynebacterium glutamicum (КССМ11138Р ΔNCgl2523, КССМ11240Р ΔNCgl2523, DAB12-a ΔNCgl2523 и DAB12-D ΔNCgl2523) и 4 разных типов родительских штаммов (КССМ11138Р, КССМ11240Р, DAB12-a и DAB12-b) высевали на содержащую 1 мМ аргинин СМ среду для чашек (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-ного раствора NaOH и 2% агара, рН 6,8; из расчета на 1 л) и культивировали при 30°С в течение 24 часов. После инокулирования в количестве 1 платиновой петли каждым из штаммов, подвергнутых культивированию, 25 мл среды для титрования (8% глюкозы, 0,25% соевых белков, 0,50% твердых веществ кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, 100 мкг биотина, 3 мг тиамина гидрохлорида, 3 мг кальциевой соли пантотеновой кислоты, 3 мг никотинамида и 5% CaCO3; из расчета на 1 л) проводили культивирование со встряхиванием при 30°С и 200 об./мин в течение 98 часов. В среды для культивирования всех штаммов добавляли 1 мМ аргинин. Измеряли концентрацию путресцина, образованного в каждой культуральной жидкости, и результаты показаны в Таблице 2.


Как показано в Таблице 2, продуцирование путресцина усиливалось у 4-х типов мутантов Corynebacterium glutamicum с делецией NCgl2523.
Пример 2. Оценка концентрации внутриклеточного путресцина для штаммов с делецией NCgl2523
Чтобы исследовать изменения концентрации внутриклеточного путресцина для мутантов Corynebacterium glutaminicum с устраненной активностью NCgl2523, измеряли концентрации внутриклеточного путресцина для мутанта Corynebacterium glutaminicum КССМ11240Р ΔNCgl2523 и родительского штамма КССМ11240Р, используя экстракцию с применением органического растворителя. Анализ внутриклеточных метаболитов проводили в соответствии с методом, описанным в литературе (Nakamura J. et al., Appl. Environ. Microbiol., 73(14): 4491-4498, 2007).
Сначала, после инокулирования каждым штаммом из мутанта Corynebacterium glutaminicum КССМ11240Р ΔNCgl2523 и родительского штамма КССМ11240Р, 25 мл жидкой среды СМ, содержащей 1 мМ аргинин (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины и 100 мкл 50%-ного раствора NaOH, при рН 6,8; из расчета на 1 л), проводили культивирование со встряхиванием при 30°С и 200 об./мин. Когда в процессе культивирования клеточный рост достигал экспоненциальной фазы, клетки выделяли из культуральных сред посредством быстрой вакуумной фильтрации (Durapore HV; 0,45 мкм; Millipore, Billerica, MA). Фильтр с адсорбированными на нем клетками промывали дважды 10 мл охлажденной воды и экстрагировали в метанол-содержащей 5 М морфолин-этансульфоновой кислоте и 5 М метионинсульфоне в течение 10 минут. Жидкость, полученную посредством экстрагирования, хорошо перемешивали с равным объемом хлороформа и 0,4-кратным объемом воды. Для удаления белковых примесей на спин-колонку наносили только водную фазу. Отфильтрованную жидкость после экстрагирования анализировали с использованием капиллярного электрофореза в сочетании с масс-спектрометрией, и результаты показаны в Таблице 3.

Как показано в Таблице 3, концентрация внутриклеточного путресцина значительно уменьшалась для мутантов Corynebacterium glutaminicum с устраненной активностью NCgl2523 по сравнению с активным родительским штаммом КССМ11240Р.
Можно предположить, что, когда NCgl2523 делетирован у мутанта Corynebacterium glutaminicum КССМ11240Р, ингибирование экспрессии NCgl2522 прекращается, а активность NCgl2522 повышается, в результате чего повышается путресцин-экспортирующая способность, и соответственно экспорт путресцина, продуцируемого внутри клетки, во внеклеточное пространство протекает эффективно.
Пример 3. Поиск ортологов гена NCgl2523 и анализ геномного контекста
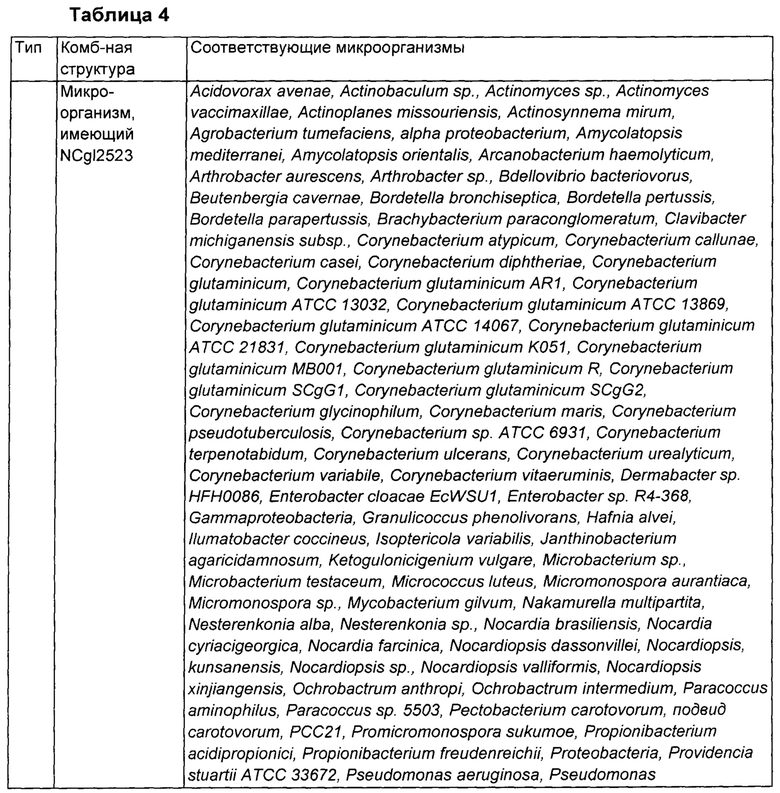
Проводили поиск ортологов для NCgl2523, происходящего из Corynebacterium glutaminicum АТСС13032, используя базу данных из KEGG (Энциклопедия генов и геномов Киото), MetaCyc и программу BlastP от Национального центра биотехнологической информации (NCBI). С использованием анализа кластеров генов изучали комбинированную структуру на основе NCgl2523 с NCgl2522 и NCgl2524, которые входят в состав оперона в геноме каждого из микроорганизмов.
Ген белка NCgl2522, который относится к пермеазе из суперсемейства основных переносчиков, принадлежащей семейству DHA2, обозначают как cgmA, а ген белка NCgl2523, который представляет собой регулятор транскрипции из TetR-семейства, обозначают как cgmR. Функция NCgl2524 пока еще не описана, тем не менее он представляет собой пермеазу из суперсемейства основных переносчиков, принадлежащую семейству UMF1 (неизвестный основной переносчик 1).
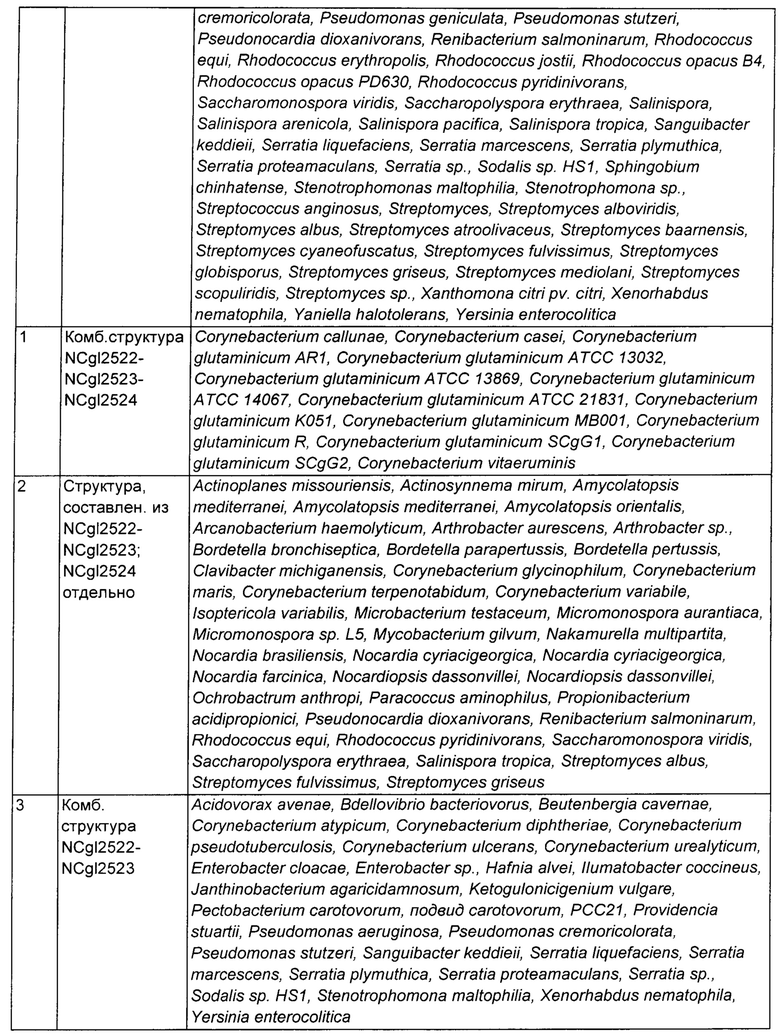
Согласно результатам анализа, в то время, как NCgl2522 и NCgl2523 главным образом входят в состав одного и того же оперона, NCgl2524, в зависимости от микроорганизмов, может присутствовать в этих же оперонах (тип 1), присутствовать в другом положении в геноме (тип 2) или не присутствовать в геноме (тип 3).
В связи с этим, каждый из анализируемых микроорганизмов классифицировали на 3 типа в соответствии с комбинированной структурой соседних с NCgl2523 генов. В частности, микроорганизмы, которые, как ожидается, имеют NCgl2523, классифицировали как имеющие: комбинированную структуру NCgl2522-NCgl2523-NCgl2524 (тип 1); структуру, составленную из NCgl2522-NCgl2523 и расположенного отдельно NCgl2524 (тип 2); или комбинированную структуру NCgl2522-NCgl2523 (тип 3). (Фиг. 1). Результаты классификации показаны в Таблице 4.
Исходя из этих результатов ожидается, что путресцин-продуцирующую способность можно повысить, как в настоящем изобретении, посредством инактивации генов, кодирующих NCgl2523 и аминокислотную последовательность, схожую с ним, у микроорганизмов, у которых имеется аминокислотная последовательность, включая последовательность, схожую с NCgl2523, и входящих в состав оперона, при этом ожидается, что белок представляет собой экспортер путресцина, такой как NCgl2522, как в данной классификации.
На основании приведенного выше описания обычному специалисту в данной области техники следует понимать, что в осуществлении настоящего изобретения могут быть использованы и другие конкретные воплощения без отклонения от технической идеи или существенных признаков настоящего изобретения. В связи с этим, описанные выше примеры приведены только в целях иллюстрации, и не подразумевается, что данное изобретение ограничено этими примерами. Следует понимать, что объем настоящего изобретения включает в себя все модификации или модифицированную форму, проистекающие из смысла и объема следующей далее формулы изобретения или ее эквивалентных понятий, а не приведенного выше подробного описания.
Классификация объекта депонирования
Орган по депонированию: Корейский центр культур микроорганизмов.
№ доступа: КССМ11520Р.
Дата депонирования: 2014.02.25.










Группа изобретений относится к микроорганизму рода Corynebacterium, обладающему повышенной путресцин-продуцирующей способностью, и способу получения путресцина с его использованием. В указанному микроорганизме активность белка, имеющего аминокислотную последовательность, представленную SEQ ID NO: 2, инактивирована. Предложен также способ получения путресцина с использованием указанного микроорганизма, включающий культивирование микроорганизма и выделение путресцина из подвергнутого культивированию микроорганизма или культуральной среды, полученных на указанной выше стадии. Группа изобретений обеспечивает получение путресцина с высоким выходом. 2 н. и 6 з.п. ф-лы, 1 ил., 4 табл., 3 пр.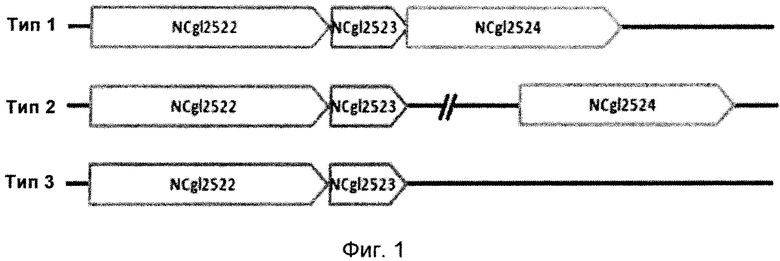
1. Микроорганизм рода Corynebacterium, обладающий повышенной путресцин-продуцирующей способностью, где активность белка, имеющего аминокислотную последовательность, представленную SEQ ID NO: 2, в указанном микроорганизме рода Corynebacterium, способном продуцировать путресцин, инактивирована.
2. Микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, по п. 1, в который дополнительно введена активность орнитиндекарбоксилазы (ODC).
3. Микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, по п. 2, где ODC имеет аминокислотную последовательность, представленную SEQ ID NO: 10.
4. Микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, по п. 1, в котором дополнительно инактивирована ацетилтрансферазная активность.
5. Микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, по п. 4, где ацетилтрансфераза содержит аминокислотную последовательность, представленную SEQ ID NO: 15 или 16.
6. Микроорганизм рода Corynebacterium, обладающий путресцин-продуцирующей способностью, по п. 1, представляющий собой Corynebacterium glutamicum.
7. Способ получения путресцина, включающий: культивирование микроорганизма по п. 1 и
выделение путресцина из подвергнутого культивированию микроорганизма или культуральной среды, полученных на указанной выше стадии.
8. Способ получения путресцина по п. 7, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
| WO 2013105827 A2, 18.07.2013 | |||
| WO 2012077995 A2, 14.06.2012 | |||
| МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ ПУТРЕСЦИНА В ВЫСОКОЙ КОНЦЕНТРАЦИИ, СПОСОБ ПОЛУЧЕНИЯ ЭТОГО МИКРООРГАНИЗМА И СПОСОБ ПРОДУКЦИИ ПУТРЕСЦИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2009 |
|
RU2433180C2 |
| база данных NCBI: WP_011015249.1, 15.05.2013 | |||
| ITOU Н | |||
| ET AL | |||
| Crystal Structures of the Multidrug Binding Repressor Corynebacterium glutamicum CgmR in Complex with Inducers and with an Operator // J | |||
| Mol | |||
| Biol | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |

