Область техники
Настоящее изобретение относится к рекомбинантному микроорганизму для получения путресцина или орнитина и к способу получения путресцина или орнитина с использованием этих микроорганизмов.
Предшествующий уровень техники
Путресцин обнаружен у грамотрицательных бактерий или грибов и присутствует у различных видов в высокой концентрации. Поэтому ожидается, что он играет важные роли в метаболизме микроорганизмов. Как правило, путресцин представляет собой очень важное исходное вещество для синтеза полиамина найлона-4,6, и его получают, главным образом, способом химического синтеза. Способ химического синтеза состоит из 3-стадийного процесса, включающего реакцию каталитического окисления, стадию использования цианидного соединения и реакции гидрогенизации с использованием водорода под высоким давлением. В этом отношении для получения путресцина требуется разработать более экологически безопасный способ с использованием биомассы, который может уменьшать потребление энергии.
В этих обстоятельствах в качестве способов получения путресцина с использованием микроорганизма были раскрыты способы получения путресцина с высоким выходом путем трансформации Е. coli и микроорганизма рода Corynebacterium (публикация международной заявки No. WO 2006/005603; публикация международной заявки No. WO 2009/125924; Qian ZD et al., Biotechnol. Bioeng. 104 (4): 651-662, 2009; Schneider et al., Appl. Microbiol. Biotechnol. 88 (4): 859-868, 2010; Schneider et al., Appl. Microbiol. Biotechnol. 95: 169-178, 2012).
Орнитин представляет собой вещество, широко распространенное у растений, животных и микроорганизмов, и его используют в качестве предшественника в биосинтезе аргинина, пролина и полиаминов. Орнитин играет важную роль в пути экскреции мочевины, продуцируемой из аминокислот или аммиака в орнитиновом цикле в метаболизме высших животных in vivo. Также орнитин используют в качестве пищевых добавок или фармацевтических лекарственных средств в промышленности для улучшения состояния при циррозе печени и функциональных расстройствах печени. Известные способы получения орнитина включают обработку казеина молока пищеварительными ферментами и использование трансформированной Е. coli или микроорганизма рода Corynebacterium (патент Кореи No. 10-1372635; Т. Gotoh et al., Bioprocess Biosyst. Eng., 33: 773-777, 2010).
SugR, представляющий собой регулятор транскрипции в метаболизме сахара (далее SugR), известен как регулятор транскрипции у Corynebacterium, и ранее сообщалось, что SugR ингибирует ген, кодирующий PEP (фосфоенолпируват)-белок фосфотрансферазу системы PTS, и гены, ассоциированные с гликолизом Сахаров (VF Wendisch, et al., J. Bacteriol. 190: 24, 8033-8044, 2008). Цитратсинтаза представляет собой фермент, который первым действует в цикле ТСА (цикл трикарбоновых кислот) и может регулировать его скорость. Сообщалось, что модифицированный штамм Corynebacterium с пониженной активностью GltA увеличивал продуцирование аспартата и лизина (Shiio et al., Agric Biol Chem. 46; 101-107, 1982).
Техническая задача
Авторы настоящего изобретения подтвердили, что манипуляция с sugR, геном, кодирующим SugR, и gltA, геном, кодирующим цитратсинтазу, улучшает продуктивность по путресцину или орнитину, тем самым создав настоящее изобретение.
Решение технической задачи
Задача настоящего изобретения заключается в том, чтобы предложить рекомбинантный микроорганизм, который может продуцировать путресцин или орнитин с высоким выходом.
Другая задача настоящего изобретения заключается в том, чтобы предложить способ получения путресцина или орнитина с использованием вышеупомянутого микроорганизма.
Полезные эффекты изобретения
Авторы настоящего изобретения подтвердили, что одновременное повышение активности цитратсинтазы (далее GltA) при понижении активности SugR у микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, повышает уровень продуцирования путресцина или орнитина. Соответственно, микроорганизм по настоящему изобретению может быть широко использован для промышленного получения путресцина или орнитина, и этот микроорганизм можно широко применять в качестве эффективного и желаемого средства в экономическом аспекте и с точки зрения безопасности окружающей среды для обеспечения исходного вещества для получения различных полимерных продуктов, в которых путресцин или орнитин используются в качестве исходных веществ.
Наилучший способ осуществления изобретения
В одном аспекте настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность регулятора транскрипции метаболизма сахара (SugR) понижена по сравнению с его эндогенной активностью, 2) активность цитратсинтазы (GltA) повышена по сравнению с ее эндогенной активностью, или 3) активность SugR понижена по сравнению с его эндогенной активностью, и активность GltA повышена по сравнению с ее эндогенной активностью.
В примере воплощения по настоящему изобретению предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность SugR понижена по сравнению с его эндогенной активностью, и активность GltA повышена по сравнению с ее эндогенной активностью.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого SugR состоит из аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 3.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого GltA состоит из аминокислотной последовательности SEQ ID NO: 5 или SEQ ID NO: 7.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где микроорганизм рода Corynebacterium выбран из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность орнитиндекарбоксилазы (ODC).
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого ODC состоит из аминокислотной последовательности SEQ ID NO: 17.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность 1) орнитин-карбамоилтрансферазы (ArgF), 2) экспортера глутамата или 3) орнитин-карбамоилтрансферазы и экспортера глутамата дополнительно понижены по сравнению с их эндогенной активностью.
В другом примере воплощения по настоящему изобретению дополнительно предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого орнитин-карбамоилтрансфераза состоит из аминокислотной
последовательности SEQ ID NO: 9 или SEQ ID NO: 11, и экспортер глутамата состоит из аминокислотной последовательности SEQ ID NO: 13 или SEQ ID NO: 15.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность по меньшей мере одного выбранного из группы, состоящей из ацетил-гамма-глутамил-фосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитин-ацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитин-аминотрнасферазы (ArgD), дополнительно повышена по сравнению с его эндогенной активностью.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого ацетил-гамма-глутамил-фосфатредуктаза состоит из аминокислотной последовательности SEQ ID NO: 19 или SEQ ID NO: 21, ацетилглутаматсинтаза или орнитин-ацетилтрансфераза состоят из аминокислотной последовательности SEQ ID NO: 23 или SEQ ID NO: 25, ацетилглутаматкиназа состоит из аминокислотной последовательности SEQ ID NO: 27 или SEQ ID NO: 29, и ацетилорнитин-аминотрнасфераза состоит из аминокислотной последовательности SEQ ID NO: 31 или SEQ ID NO: 33.
В другом примере воплощения по настоящему изобретению дополнительно предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность ацетилтрансферазы дополнительно понижена по сравнению с ее эндогенной активностью.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого ацетилтрансфераза состоит из аминокислотной последовательности SEQ ID NO: 35 или SEQ ID NO: 37.
В другом примере воплощения по настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активность белка, состоящего из SEQ ID NO: 39 или SEQ ID NO: 41, дополнительно повышена по сравнению с его эндогенной активностью.
В другом аспекте настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или из культуральной среды со стадии (1).
В одном примере воплощения по настоящему изобретению предложен способ получения путресцина или орнитина, в котором микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
Далее настоящее изобретение описано более подробно.
Один аспект настоящего изобретения относится к микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин, у которого активность регулятора транскрипции метаболизма сахара (SugR) понижена по сравнению с его эндогенной активностью, 2) активность цитратсинтазы (GltA) повышена по сравнению с ее эндогенной активностью или 3) активность SugR понижена по сравнению с его эндогенной активностью и активность GltA повышена по сравнению с ее эндогенной активностью. В частности, настоящее изобретение относится к микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин, у которого активность регулятора транскрипции метаболизма сахара понижена по сравнению с его эндогенной активностью, и активность цитратсинтазы повышена по сравнению с ее эндогенной активностью.
Как его используют здесь, термин "регулятор транскрипции метаболизма сахара (SugR)" относится к ферменту, который широко задействован в качестве ингибитора в отношении генов, ассоциированных с различными аспектами метаболизма сахара, такими как потребление сахара и фосфотрансферазная система, гликолиз, ферментация, относящаяся к лактатдегидрогеназе, и так далее. В настоящем изобретении SugR включает как эндогенные белки, так и чужеродные белки в микроорганизме рода Corynebacterium, и в частности, SugR, происходящий из микроорганизма рода Corynebacterium.
В настоящем изобретении регулятор транскрипции метаболизма сахара может включать, без ограничения, любой белок, включающий аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 3, или любой белок, включающий аминокислотную последовательность, имеющую гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше, и наиболее точно 99% или больше, с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как регулятор транскрипции метаболизма сахара.
Дополнительно, поскольку аминокислотная последовательность белка, кодирующего вышеупомянутую активность, может отличаться в зависимости от вида или штамма микроорганизма, SugR можно не ограничивать в настоящем изобретении в отношении его происхождения, но SugR может иметь происхождение, например, из микроорганизма рода Corynebacterium, и в частности происходить из Corynebacterium glutamicum. Очевидно, что любая аминокислотная последовательность, которая имеет гомологию с вышеупомянутыми последовательностями и обладает биологической активностью по существу такой же как белок, представленный SEQ ID NO: 1 или SEQ ID NO: 3, или соответствующей таковой, также может входить в объем настоящего изобретения, несмотря на то, что аминокислотная последовательность может иметь делецию, модификацию, замену или вставку в части последовательности.
Полинуклеотид, кодирующий регулятор транскрипции метаболизма сахара по настоящему изобретению, при условии, что он обладает активностью, сходной с таковой регулятора транскрипции метаболизма сахара, может включать любой полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 3, или полинуклеотиды, кодирующие белки, имеющие гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше, и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями. Что касается полинуклеотида, кодирующего регулятор транскрипции метаболизма сахара, учитывая кодоны на основе вырожденности кодонов или те, которые предпочтительны для организмов для экспрессии этого регулятора, различные модификации могут быть выполнены в кодирующем участке в пределах объема без изменения аминокислотной последовательности полипептида, и в частности, полинуклеотид может включать полинуклеотидную последовательность SEQ ID NO: 2 или SEQ ID NO: 4, но не ограничиваясь этим.
Как его используют здесь, термин "цитратсинтаза (GltA)" относится к ферменту, вовлеченному в продуцирование различных внутриклеточных промежуточных продуктов биосинтеза и продуцирование восстановленной пуриновой нуклеиновой кислоты. Известно, что GltA действует как посредник в гидролитической конденсации между ацетил-СоА и оксалоацетатом для продуцирования цитрата. В настоящем изобретении GltA включает как эндогенные ферменты, так и чужеродные белки, присутствующие у микроорганизма рода Corynebacterium, и в частности, GltA, имеющий происхождение из микроорганизма рода Corynebacterium.
В настоящем изобретении GltA может включать, без ограничения, белки, имеющие аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 7, и любой белок, который включает аминокислотную последовательность, имеющую гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, и обладает основной активностью посредника в гидролитической конденсации между ацетил-СоА и оксалоацетатом для продуцирования цитрата.
Дополнительно, поскольку аминокислотная последовательность белка, проявляющего активность, может варьировать в зависимости от вида или штамма микроорганизма, GltA может происходить, например, из Corynebacterium, и в частности, Corynebacterium glutamicum, но происхождение GltA не ограничено этим в настоящем изобретении. Очевидно, что любая аминокислотная последовательность, имеющая гомологию с вышеупомянутыми последовательностями и обладающая биологической активностью по существу такой же как белок, представленный SEQ ID NO: 5 или SEQ ID NO: 7, или соответствующей таковой, также может входить в объем настоящего изобретения, несмотря на то что аминокислотная последовательность может иметь делецию, модификацию, замену или вставку в части последовательности.
Полинуклеотид, кодирующий GltA по настоящему изобретению, может включать полинуклеотиды, кодирующие аминокислоту SEQ ID NO: 5 или SEQ ID NO: 7, или полинуклеотиды, кодирующие белки, имеющие гомологию 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями. В отношении полинуклеотида, кодирующего GltA, учитывая кодоны на основе вырожденности кодонов или те, которые предпочтительны для организмов для экспрессии GltA, различные модификации могут быть выполнены в кодирующем участке в пределах объема без изменения аминокислотной последовательности полипептида, и в частности, полинуклеотид может включать полинуклеотидную последовательность SEQ ID NO: 6 или SEQ ID NO: 8, но не ограничиваясь этим.
Как его используют здесь, термин "гомология" относится к степени идентичности при сравнении с данной аминокислотной последовательностью или полинуклеотидной последовательностью и может быть выражена в процентах. В настоящем изобретении гомологичные последовательности, обладающие такой же активностью как данная аминокислотная или полинуклеотидная последовательность или подобной активностью, обозначают в виде "% гомологии." Например, гомологию можно подтвердить с использованием стандартного программного обеспечения для расчета параметров (например, таких параметров как показатель, идентичность и сходство), в частности BLAST 2.0, или сравнения последовательностей путем саузерн-блоттинга в определенных строгих условиях гибридизации, и подходящие условия, подлежащие определению, могут быть определены способом, который входит в уровень техники и хорошо известен специалисту в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
Дополнительно полинуклеотиды, кодирующие SugR и цитратсинтазу по настоящему изобретению, можно гибридизовать в строгих условиях с полинуклеотидными последовательностями SEQ ID NO: 2 или 4, или SEQ ID NO: 6 или 8, или зондами, происходящими из полинуклеотидных последовательностей, соответственно, и могут представлять собой модифицированный тип, кодирующий SugR и цитратсинтазу, которые вовлечены в нормальные функции. Как его используют здесь, термин "строгие условия" относится к условиям, обеспечивающим возможность специфической гибридизации между полинуклеотидами. Например, строгие условия более конкретно описаны в литературе (например, J. Sambrook et al., там же).
В настоящем изобретении предприняли попытки снизить активность SugR, или повысить активность GltA, или применить как снижение активности SugR, так и повышение активности GltA одновременно в микроорганизме рода Corynebacterium, продуцирующем путресцин или орнитин, и в результате было подтверждено, что уровень продуцирования путресцина или орнитина повышался во всех модифицированных штаммах.
В частности, микроорганизм по настоящему изобретению может включать микроорганизмы как дикого типа, так и модифицированного типа при условии, что они могут продуцировать путресцин или орнитин. Например, микроорганизм может принадлежать роду Escherichia, роду Shigella, роду Citrobacter, роду Salmonella, роду Enterobacter, роду Yersinia, роду Klebsiella, роду Erwinia, роду Corynebacterium, роду Brevibacterium, роду Lactobacillus, роду Selenomanas, роду Vibrio, роду Pseudomonas, роду Streptomyces, роду Arcanobacterium и роду Alcaligenes. В частности, микроорганизм по настоящему изобретению может принадлежать роду Corynebacterium и, более конкретно, может быть выбран из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum, и еще более конкретно может представлять собой Corynebacterium glutamicum, но не ограничиваясь этим.
В частности, как его используют здесь, выражение "продуцирующий путресцин или орнитин" относится к микроорганизму, обладающему продуктивностью по путресцину или орнитину, происходящему из родительского штамма, который имеет путресцин или орнитин в природном состоянии или не обладает продуктивностью по путресцину или орнитину.
Дополнительно микроорганизм, продуцирующий путресцин или орнитин, может быть модифицирован для снижения активности орнитин-карбамоилтрансферазы (ArgF), которая вовлечена в синтез аргинина, и/или активность экспортера глутамата (NCgl1221), который представляет собой белок, вовлеченный в экскрецию глутамата, по сравнению с их соответствующими эндогенными активностями.
Кроме того, микроорганизм, обладающий продуктивностью по путресцину, может быть модифицирован для снижения активности ацетилтрансферазы (NCgl1469), которая представляет собой белок, который ацетилирует путресцин, по сравнению с ее эндогенной активностью, и/или для введения активности ODC, которая представляет собой белок, превращающий орнитин в путресцин.
В частности, модификация по повышению или снижению активностей может происходить в процессе, называемом трансформацией в настоящем изобретении. Как его используют здесь, термин "трансформация" относится к процессу введения полинуклеотида, кодирующего определенный белок, или вектора, включающего промоторную последовательность с сильной или слабой активностью, и так далее, в клетку-хозяина, тем самым обеспечивая возможность экспрессии белка, кодируемого полинуклеотидом в клетке-хозяине, или индуцируя модификацию хромосомы клетки-хозяина.
Дополнительно полинуклеотид включает ДНК или РНК, кодирущую белок-мишень. Полинуклеотид может быть вставлен в любой форме при условии, что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все основные элементы, необходимые для самостоятельной экспрессии. Традиционно экспрессионная кассета может включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, домен связывания с рибосомой и сигнал терминации трансляции. Экспрессионная кассета может быть в форме экспрессионного вектора, способного к саморепликации. Дополнительно полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине, но не ограничиваясь этим.
Дополнительно, как его используют здесь, термин "функционально связанный" относится к функциональной связи между промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего белок-мишень, по настоящему изобретению и вышеописанной последовательностью гена.
Как его используют здесь, термин "вектор" относится к любой ДНК-конструкции, которая включает полинуклеотидную последовательность, кодирующую белок-мишень, которая функционально связана с подходящей контрольной последовательностью, способной экспрессировать белок-мишень в подходящей клетке-хозяине. Контрольная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность, способную контролировать транскрипцию, последовательность, кодирующую подходящую мРНК домена связывания с рибосомой, и последовательности, способные контролировать терминацию транскрипции и трансляции. Вектор, после трансформации им подходящей клетки-хозяина, может реплицироваться или действовать независимо от генома хозяина, или может быть интегрирован в геном хозяина самостоятельно.
Вектор для использования в настоящем изобретении может быть конкретно не ограничен, при условии, что вектор обладает способностью реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры традиционно используемого вектора могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее; и в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptll, pGEM, pTZ, pCL, pET и так далее. Вектор для использования в настоящем изобретении можно конкретно не ограничивать, но можно использовать любой известный вектор экспрессии. В частности, можно использовать векторы pDZ, pDZTn, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
Как таковой, полинуклеотид, кодирующий чужеродный белок-мишень, может быть заменен на модифицированный полинуклеотид в хромосоме с помощью вектора для вставки в бактериальную хромосому. Вставка полинуклеотида в хромосому может быть выполнена с использованием любого известного в данной области техники способа, например, путем гомологичной рекомбинации, но не ограниваясь этим. Поскольку вектор по настоящему изобретению может быть вставлен в хромосому путем гомологичной рекомбинации, может быть дополнительно включен селективный маркер для подтверждения вставки в хромосому. Селективный маркер используют для отбора трансформированных клеток, то есть для подтверждения того, был ли вставлен целевой полинуклеотид, и можно использовать маркеры, способные обеспечивать возможность отбора фенотипов, таких как лекарственная устойчивость, потребность в питательных веществах, устойчивость к цитотоксическим агентам и экспрессия поверхностных белков. В условиях обработки селективными агентами только клетки, способные экспрессировать селективные маркеры, могут выживать или проявлять другие фенотипические признаки, и таким образом трансформированные клетки могут быть отобраны.
Как его используют здесь, термин "повышение активности" включает не только достижение более сильного действия, чем исходная функция, вследствие нового введения активности или повышения активности самого белка, но также включает повышение его активности путем повышения активности эндогенного гена, амплификации эндогенного гена под действием внутреннего(их) или внешнего(их) фактора(ов), делеции регуляторного(ых) фактора(ов) для ингибирования экспрессии гена, увеличение числа копий гена, введение гена из другого источника, модификации последовательности контроля экспрессии и, в частности, повышения активности фермента путем замены или модификации промотора и мутации внутри гена и так далее.
В частности, в настоящем изобретении усиление или повышение активности может быть выполнено путем:
1) увеличения числа копий полинуклеотида, кодирующего фермент,
2) модификации последовательности контроля экспрессии для увеличения экспрессии полинуклеотида,
3) модификации полинуклеотидной последовательности на хромосоме для повышения активности фермента, и
4) модификации путем их комбинации, но способ не ограничен этим.
Увеличение числа копий полинуклеотида (способ 1) может быть выполнено в форме, в которой полинуклеотид функционально связан с вектором, или путем вставки полинуклеотида в хромосому клетки-хозяина, хотя способ специально не ограничен этими. В частности, увеличение числа копий полинуклеотида внутри хромосомы клетки-хозяина может быть выполнено путем введения вектора, который может реплицироваться и функционировать независимо от клетки-хозяина и с которым полинуклеотид, кодирующий белок по настоящему изобретению, функционально связан; или может быть выполнен путем введения вектора, который может вставлять полинуклеотид в хромосому клетки-хозяина, и с которым полинуклеотид функционально связан, в клетку-хозяина.
Далее, модификация последовательности контроля экспрессии для увеличения экспрессии полинуклеотида (способ 2) может быть выполнена путем индуцирования модификации в полинуклеотидной последовательности путем делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного повышения активности последовательности контроля экспрессии или путем замены полинуклеотидной последовательности на полинуклеотидную последовательность, имеющую более высокую активность, хотя способ специально не ограничен этим. Последовательность контроля экспрессии включает промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой и последовательность, регулирующую терминацию транскрипции и трансляции.
Сильный экзогенный промотор может быть присоединен вместо исходного промотора к области, лежащей выше по ходу транскрипции от экспрессионной единицы полинуклеотида. Примеры сильного промотора могут представлять собой промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор асеА или асеВ и так далее, и более конкретно, уровень экспрессии может быть улучшен путем функционального связывания с промотором lysCP1, имеющим происхождение из Corynebacterium (WO 2009/096689) или промотором CJ7 (патент Кореи No. 0620092 и WO 2006/065095), но сильный промотор не ограничен этим.
Кроме того, модификация полинуклеотидной последовательности на хромосоме (способ 3) может быть выполнена путем индуцирования модификации в последовательности контроля экспрессии путем делеции, вставки, неконсервативной или консервативной замены полинуклеотидной последовательности или их комбинации для дополнительного повышения активности полинуклеотидной последовательности, или путем замены полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более высокой активностью, хотя способ специально не ограничен этими.
Как его используют здесь, "снижение активности" может быть достигнуто путем делетирования, частично или полностью, полинуклеотида, кодирующего белок, для снижения активности белка, путем модификации последовательности контроля экспрессии для уменьшения экспрессии полинуклеотида, путем модификации полинуклеотидной последовательности на хромосомах для снижения активности белка и способом, выбранным из их комбинации.
В частности, в настоящем изобретении снижение активности может быть достигнуто путем:
1) делетирования, частично или полностью, полинуклеотида, кодирующего белок,
2) модификации последовательности контроля экспрессии для уменьшения экспрессии полинуклеотида,
3) модификации полинуклеотидной последовательности на хромосомах для снижения активности белка, и
4) способа, выбранного из их комбинации, но способ не ограничен этими.
В частности, способ делетирования, частично или полностью, полинуклеотида, кодирующего белок, может быть выполнен путем замены полинуклеотида, кодирующего эндогенный белок-мишень, в хромосоме на полинуклеотид, имеющий частичную делецию в полинуклеотидной последовательности, или ген-маркер с использованием вектора для вставки в хромосому в бактериях. Как его используют здесь, термин "часть" может варьировать в зависимости от типов полинуклеотидов, и в частности он может включать их в количестве от 1 до 300, более конкретно от 1 до 100 и еще более конкретно от 1 до 50.
Дополнительно, способ модификации последовательности контроля экспрессии может быть выполнен путем индуцирования модификации в последовательности контроля экспрессии путем делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного снижения активности последовательности контроля экспрессии или путем замены полинуклеотидной последовательности на полинуклеотидную последовательность, имеющую более слабую активность. Последовательность контроля экспрессии включает промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляции.
Дополнительно, способ модификации полинуклеотидной последовательности на хромосоме может быть выполнен путем индуцирования модификации в полинуклеотидной последовательности путем делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного снижения активности белка или путем замены полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более высокой активностью.
Дополнительно, способ делетирования регуляторного фактора, который ингибирует экспрессию белка с полинуклеотида, может быть выполнен путем замены полинуклеотида для фактора ингибирования экспрессии на полинуклеотид, имеющий частичную делецию в полинуклеотидной последовательности, или ген-маркер. Как его используют здесь, термин "часть" может варьировать с зависимости от типов полинуклеотидов, и в частности он может включать их в количестве от 1 до 300, более конкретно от 1 до 100 и еще более конкретно от 1 до 50.
Как его используют здесь, термин "эндогенная активность" относится к активному состоянию фермента в немодифицированном состоянии, например в природном состоянии, в котором он исходно присутствует у микроорганизма, и фраза "повышение по сравнению с его эндогенной активностью" относится к состоянию повышенной активности белка, которое присутствует у микроорганизма после манипуляции, такой как введение гена, проявляющего активность, или увеличение числа копий соответствующего гена, делеция фактора контроля ингибирования экспрессии гена или модификация последовательности контроля экспрессии, например использование промотора с улучшенной активностью по сравнению с активностью, которой обладал микроорганизм до манипуляции.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium, обладающий продуктивностью по путресцину, в который дополнительно введена активность орнитин-декарбоксилазы (ODC).
Как его используют здесь, термин "орнитин-декарбоксилаза (ODC)" относится к ферменту, способному продуцировать путресцин путем опосредования декарбоксилирования орнитина. Несмотря на то, что микроорганизм рода Corynebacterium не имеет пути биосинтеза путресцина, когда ODC введена из чужеродного источника, путресцин синтезируется и высвобождается наружу из клеток. В настоящем изобретении ODC может состоять из аминокислотной последовательности SEQ ID NO: 17 или может включать, без ограничения, любой белок, который имеет гомологию 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутой аминокислотной последовательностью при условии, что белок обладает по существу такой же активностью ODC.
Дополнительно, поскольку аминокислотная последовательность белка, проявляющего активность, может варьировать в зависимости от вида или штамма микроорганизма, происхождение ODC не ограничено в настоящем изобретении, и в частности, может представлять собой ODC, имеющую происхождение из Е. coli. Очевидно, что любая аминокислотная последовательность, которая имеет гомологию с вышеописанными последовательностями и обладает биологической активностью по существу такой же как белок с SEQ ID NO: 17, или соответствующей таковой, также может входить в объем настоящего изобретения, хотя аминокислотная последовательность может иметь делецию, модификацию, замену или вставку в части последовательности.
Полинуклеотид, кодирующий ODC по настоящему изобретению, может включать полинуклеотиды, кодирующие аминокислотную последовательность SEQ ID NO: 17, или полинуклеотиды, кодирующие белки, имеющие гомологию 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше, и наиболее точно 99% или больше с вышеупомянутой аминокислотной последовательностью. В отношении полинуклеотида, кодирующего ODC, учитывая кодоны на основе вырожденности кодонов или те, которые предпочтительны для организмов для экспрессии этого регулятора, различные модификации могут быть выполнены в кодирующем участке в пределах объема без изменения аминокислотной последовательности полипептида.
Микроорганизм рода Corynebacterium может представлять собой микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого активности 1) орнитин-карбамоилтрансферазы (ArgF), 2) экспортера глутамата (NCgl1221) или 3) орнитин-карбамоилтрансферазы и экспортера глутамата дополнительно снижены по сравнению с их эндогенными активностями.
В настоящем изобретении орнитин-карбамоилтрансфераза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 9 или SEQ ID NO: 11, или любой белок, состоящей из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше, и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как орнитин-карбамоилтрансфераза.
Дополнительно, экспортер глутамата по настоящему изобретению может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 13 или SEQ ID NO: 15, или любой белок, включающий аминокислотную последовательность, имеющую гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как экспортер глутамата.
Дополнительно, микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, у которого по меньшей мере одна активность, выбранная из группы, состоящей из ацетил-гамма-глутамил-фосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитин-ацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитин-аминотрансферазы (ArgD), дополнительно повышена по сравнению с их эндогенными активностями.
В настоящем изобретении ацетил-гамма-глутамил-фосфатредуктаза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 19 или SEQ ID NO: 21, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как ацетил-гамма-глутамил-фосфатредуктаза.
Дополнительно, ацетилглутаматсинтаза или орнитин-ацетилтрансфераза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 23 или SEQ ID NO: 25, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как ацетилглутаматсинтаза или орнитин-ацетилтрансфераза.
В настоящем изобретении ацетилглутаматкиназа может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 27 или SEQ ID NO: 29, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как ацетилглутаматкиназа.
Дополнительно, в настоящем изобретении ацетилорнитин-аминотрансфераза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 31 или SEQ ID NO: 33, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как ацетилорнитин-аминотрансфераза.
Более того, микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium, обладающий продуктивностью по путресцину, у которого активность ацетилтрансферазы (NCgl1469) дополнительно понижена по сравнению с ее эндогенной активностью.
В настоящем изобретении ацетилтрансфераза может включать любой белок, который может переносить ацетильную группу на путресцин. Ацетилтрансфераза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 35 или SEQ ID NO: 37, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как ацетилтрансфераза.
Наконец, микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium, обладающий продуктивностью по путресцину, у которого активность NCgl2522 дополнительно повышена по сравнению с его эндогенной активностью.
В настоящем изобретении NCgl2522 представляет собой белок, играющий роль в высвобождении путресцина, и может включать любой белок, который может переносить ацетильную группу на путресцин. Ацетилтрансфераза может включать, без ограничения, любой белок, состоящий из аминокислотной последовательности SEQ ID NO: 39 или SEQ ID NO: 41, или любой белок, состоящий из аминокислотной последовательности, имеющей гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, даже еще более конкретно 98% или больше и наиболее точно 99% или больше с вышеупомянутыми аминокислотными последовательностями, при условии, что белок обладает по существу такой же активностью как NCgl2522.
В другом аспекте настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или из культуры со стадии (1).
В настоящем изобретении микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum.
Микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, по настоящему изобретению, является таким как описано выше.
В вышеизложенном способе микроорганизм можно культивировать путем периодического культивирования, непрерывного культивирования и периодического культивирования с подпиткой, известных в данной области техники, хотя они специально не ограничены этим. В частности, в отношении условий культивирования, подходящий рН (то есть оптимальный рН от 5 до 9, в частности рН от 6 до 8 и более конкретно рН 6,8) можно поддерживать с использованием оснований (например, гидроксида натрия, гидроксида калия или аммиака) или кислот (например, фосфорной кислоты или серной кислоты), хотя они специально не ограничены этим. Дополнительно, аэробные условия можно поддерживать путем добавления кислорода и кислородсодержащей газовой смеси к клеточной культуре. Температуру культуры можно поддерживать в диапазоне от 20°С до 45°С, и в частности от 25°С до 40°С, и микроорганизм можно культивировать в течение периода примерно от 10 часов до 160 часов. Путресцин или орнитин, продуцированные путем вышеописанного культивирования, могут секретироваться в культуральную среду или оставаться внутри клеток.
Дополнительно, источники углерода, такие как сахара и углеводороды (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масла и жиры (например, масло сои, масло семян подсолнечника, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например, глицерин и этанол) и органические кислоты (например, уксусную кислоту) в культуральной среде можно использовать по отдельности или в комбинации, но не ограничиваясь этим; источники азота, такие как азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт, соевая мука и мочевина) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), можно использовать по отдельности или в комбинации, но не ограничиваясь этим; и источники калия, такие как однозамещенный фосфат калия, гидроортофосфат калия, а также соответствующие им натрийсодержащие соли, можно использовать по отдельности или в комбинации, но не ограничиваясь этим. Дополнительно, культуральная среда может содержать другие важные стимулирующие рост вещества, включая соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Способ выделения путресцина или орнитина, продуцируемых во время культивирования, по настоящему изобретению может быть выполнен с использованием подходящего способа культивирования, известного в данной области техники, например периодического культивирования, непрерывного культивирования или периодического культивирования с подпиткой, и таким образом целевое вещество может быть выделено из культуры.
Примеры осуществления изобретения
Далее настоящее изобретение будет раскрыто более подробно со ссылкой на соответствующие примеры воплощений. Однако раскрытые здесь примеры воплощений предназначены исключительно для иллюстративных целей и не предназначены никаким образом ограничивать объем настоящего изобретения.
Пример 1: Получение штаммов, ослабленных по гену sugR, из штаммов, обладающих продуктивностью по путресцину
Авторы настоящего изобретения подтвердили эффект ослабления sugR, который представляет собой ген, кодирующий SugR, у штамма, обладающего продуктивностью по путресцину.
1-1. Получение штаммов, ослабленных по гену sugR, из штамма на основе АТСС13032. обладающего продуктивностью по путресцину
Для подтверждения того, имеет ли отношение ослабление гена sugR к продуктивности по путресцину у штамма на основе Corynebacterium glutamicum АТСС13032, обладающего продуктивностью по путресцину (Публикация заявки на патент Кореи No. 10-2013-0082478), был получен штамм, ослабленный по sugR. Более конкретно, штамм, ослабленный по sugR, получали путем изменения инициирующего кодона гена sugR и замены промотора на Вб-ослабленный промотор (Patek М (2005) Regulation of gene expression. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC, BocaRaton).
Сначала был получен вектор для изменения инициирующего кодона гена sugR. При рассмотрении области вблизи полинуклеотидной последовательности гена, кодирующего SugR, представленной в SEQ ID NO: 2, были получены пара праймеров SEQ ID NO: 43 и 44 для получения фрагментов для гомологичной рекомбинации выше по ходу транскрипции от инициирующего кодона гена sugR и пара праймеров SEQ ID NO: 45, 46 и 47 для получения фрагмента для гомологичной рекомбинации ниже по ходу транскрипции от инициирующего кодона гена sugR. Праймеры, используемые для изменения инициирующих кодонов, приведены в Таблице 1 ниже.

ПЦР проводили с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы наряду с 2 парами праймеров, соответственно, для амплификации участка выше по ходу транскрипции и участка ниже по ходу транскрипции от инициирующего кодона гена sugR, соответственно, и материал, полученный в результате ПЦР, подвергали электрофорезу для получения желаемых фрагментов. В частности, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 30 секунд. Полученные таким образом фрагменты подвергали электрофорезу в 0,8% агарозном геле и материал из полос, соответствующих желаемым размерам, элюировали и очищали.
Вектор pDZ (патент Кореи No. 10-0924065) обрабатывали BamHI и SalI и затем ПЦР-продукты штамма АТСС13032 подвергали клонированию слиянием (fusion cloning). "Клонирование слиянием" выполняли с использованием набора для клонирования In-Fusion® HD Cloning Kit (Clontech). По существу были получены плазмиды pDZ-1'sugR(GTG) и pDZ-1'sugR(TTG).
В случае вектора для замены в В6-ослабленный промотор была получена SEQ ID NO: 48, представленная в Таблице 2 ниже, для получения вектора.
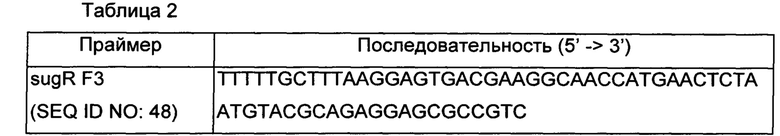
ПЦР проводили с использованием пары праймеров SEQ ID NO: 43 и 44 для получения фрагмента для гомологичной рекомбинации фрагмента выше по ходу транскрипции от инициирующего кодона гена sugR и пары праймеров SEQ ID NO: 48 и 47 для получения фрагмента для гомологичной рекомбинации ниже по ходу транскрипции от инициирующего кодона гена sugR, которые были получены при рассмотрении области вблизи полинуклеотидной последовательности гена, кодирующего SugR, представленной в SEQ ID NO: 2, и область выше по ходу транскрипции и область ниже по ходу транскрипции от инициирующего кодона гена sugR амплифицировали, соответственно, и материал, полученный в результате ПЦР, подвергали электрофорезу для получения желаемых фрагментов. Полученные таким образом фрагменты подвергали электрофорезу в 0,8% агарозном геле и материал из полос, соответствующих желаемым размерам, элюировали и очищали. Вектор pDZ обрабатывали BamHI и SalI и затем ПЦР-продукты штамма АТСС13032 подвергали клонированию слиянием. Клонирование слиянием выполняли с использованием набора для клонирования In-Fusion® HD Cloning Kit (Clontech). По существу была получена плазмида pDZ-1'sugR(B6).
Плазмиды pDZ-1'sugR(GTG), pDZ-1'sugR(TTG) и pDZ-1'sugR(B6) вводили в микроорганизм рода Corynebacterium КССМ11240Р (Публикация заявки на патент Кореи No. 10-2013-0082478) путем электропорации для получения трансформантов и трансформанты высевали на чашку со средой BHIS (бульон с сердечно-мозговым экстрактом 37 г/л, сорбит 91 г/л и агар 2%), содержащей канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали для получения колоний. Среди этих колоний отбирали голубые колонии, и таким образом были отобраны штаммы, в которые были введены плазмиды pDZ-1'sugR(GTG), pDZ-1'sugR(TTG) и pDZ-1'sugR(B6).
Выбранные штаммы культивировали при встряхивании (30°С, 8 часов) в среде СМ (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л, мочевина 2 г/л, рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний.
Среди образованных таким образом колоний отбирали белые колонии, которые появлялись в относительно небольшом количестве, и в итоге выбирали штаммы, у которых инициирующий кодон sugR был заменен на GTG или TTG путем вторичного кроссинговера, или штаммы, у которых промотор был заменен на В6. В отношении отобранных в итоге штаммов проводили ПЦР с использованием пары праймеров SEQ ID NO: 43 и 47 и подтвердили, что инициирующий кодон sugR был заменен на GTG или TTG либо промотор был преобразован в В6-ослабленный промотор, и модифицированные штаммы Corynebacterium glutamicum получили название КССМ11240Р sugR (GTG), КССМ11240Р sugR (TTG), KCCM11240P sugR (B6).
1-2. Получение штамма, ослабленного по гену sugR, из штамма на основе АТСС13869. обладающего продуктивностью по путресцину
DAB12-a ANCgl1469 (Публикация заявки на патент Кореи No. 10-2014-0115244), который представляет собой штамм на основе Corynebacterium glutamicum АТСС13869, обладающий продуктивностью по путресцину, получил название DAB12-b, и штамм, ослабленный по sugR, был получен на основе штамма DAB12-b.
В частности, для подтверждения последовательностей гена, кодирующего SugR, происходящих из Corynebacterium glutamicum АТСС13869, и белка, экспрессируемого на их основе, проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы наряду с парой праймеров SEQ ID NO: 43 и SEQ ID NO: 49.
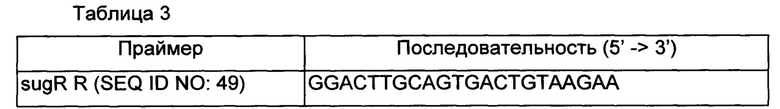
Более конкретно, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 1 минуты и 30 секунд.
Полученные таким образом ПЦР-продукты разделяли путем электрофореза и проводили анализ последовательностей, и в результате было подтверждено, что ген, кодирующий SugR, происходящий из Corynebacterium glutamicum АТСС13869, включает полинуклеотидную последовательность, представленную в SEQ ID NO: 4. Сравнение белковой последовательности, кодируемой на ее основе, и аминокислотной последовательности SugR, происходящего из Corynebacterium glutamicum АТСС13032 (SEQ ID NO: 1), выявило, что их гомология составила 99%.
Для изменения инициирующего кодона sugR, происходящего из Corynebacterium glutamicum АТСС13869, и замены В6-ослабленного промотора проводили ПЦР как в Примере 1-1 с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы наряду с праймерами, описанными в Таблицах 1 и 2 выше, и ПЦР-фрагменты области выше по ходу транскрипции и области ниже по ходу транскрипции от инициирующего кодона sugR амплифицировали, соответственно, и затем подвергали электрофорезу для получения желаемых фрагментов. В частности, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 30 секунд. Полученные таким образом фрагменты подвергали электрофорезу в 0,8% агарозном геле и материал из полос, соответствующих желаемому размеру, элюировали и очищали.
Вектор pDZ обрабатывали BamHI и SalI и затем ПЦР-продукты штамма АТСС13032 подвергали клонированию слиянием. Клонирование слиянием выполняли с использованием набора для клонирования In-Fusion® HD Cloning Kit (Clontech). По существу были получены плазмиды pDZ-2'sugR(GTG), pDZ-2'sugR(TTG) и pDZ-2'sugR(B6).
Плазмидами pDZ-2'sugR(GTG), pDZ-2'sugR(TTG) и pDZ-2'sugR(B6) трансформировали Corynebacterium glutamicum DAB12-b таким же образом, как в Примере 1-1, и отбирали штаммы, у которых инициирующий кодон sugR был заменен и/или промотор был преобразован в В6-ослабленный промотор. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия DAB12-b sugR(GTG), DAB12-b sugR(TTG) и DAB12-b sugR(B6), соответственно.
Пример 2: Получение штамма, усиленного по gltA. из штамма, обладающего продуктивностью по путресцину
Для подтверждения действия по усилению активности gltA, который представляет собой цитратсинтазу, у штамма, обладающего продуктивностью по путресцину, был получен модифицированный штамм, у которого ген gltA был введен в ген транспозона в хромосоме штамма, обладающего продуктивностью по путресцину. Использовали вектор pDZTn (WO 2009/125992) для трансформации, который может вводить ген в хромосому, и область гена транспозона микроорганизма рода Corynebacterium.
2-1. Получение штамма, усиленного по gltA, из штамма на основе АТСС13032, обладающего продуктивностью по путресцину
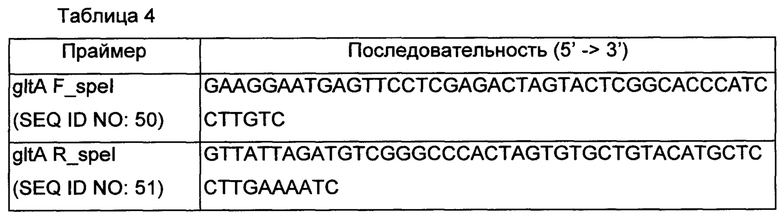
Фрагменты гена gltA амплифицировали с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы наряду с праймерами SEQ ID NO: 50 и 51 (Таблица 4). В частности, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 30 секунд или 1 минуты и 30 секунд. Полученные таким образом фрагменты подвергали электрофорезу в 0,8% агарозном геле, и материал из полос, соответствующих желаемым размерам, элюировали и очищали.
Вектор pDZTn обрабатывали Spel и затем ПЦР-продукты подвергали клонированию слиянием, соответственно. Клонирование слиянием выполняли с использованием набора для клонирования In-Fusion® HD Cloning Kit (Clontech). Полученная таким образом плазмида получила название pDZTn1'-gltA.
Полученную таким образом плазмиду вводили в штамм КССМ11240Р путем электропорации для получения трансформанта и трансформант культивировали при встряхивании (30°С, 8 часов) в среде СМ (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л, мочевина 2 г/л, рН 6,8), последовательно разведенный от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний.
Среди образованных таким образом колоний отбирали белые колонии, которые появлялись в относительно небольшом количестве, и в итоге был отобран штамм, которому вторичным кроссинговером был введен ген, кодирующий gltA. В отношении отобранных в итоге штаммов, было подтверждено, что ПЦР была выполнена с использованием пары праймеров SEQ ID NO: 50 и 51, и что ген, кодирующий gltA, был введен в них, и этот модифицированный штамм Corynebacterium glutamicum получил название КССМ11240Р Tn::gltA.
2-2. Получение штамма, усиленного по gltA, из штамма на основе АТСС13869. обладающего продуктивностью по путресцину
В отношении штамма DAB12-b, используемого в Примере 1-2, был получен штамм, усиленный по gltA.
В частности, для подтверждения последовательностей гена, кодирующего gltA, имеющий происхождение от Corynebacterium glutamicum АТСС13869, и белка, экспрессируемого на их основе, проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы наряду с парой праймеров SEQ ID NO: 50 и SEQ ID NO: 51.
В частности, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 1 минуты и 30 секунд. Полученные таким образом ПЦР-продукты разделяли электрофорезом и подвергали анализу последовательности, и в результате было подтверждено, что ген, кодирующий gltA, имеющий происхождение из Corynebacterium glutamicum АТСС13869, включает полинуклеотидную последовательность, представленную в SEQ ID NO: 8. Сравнение кодируемой ею белковой последовательности и аминокислотной последовательности gltA, имеющей происхождение от Corynebacterium glutamicum АТСС13032 (SEQ ID NO: 5), выявило, что гомология между ними составляет 99%.
Для усиления gltA, имеющего происхождение из Corynebacterium glutamicum АТСС13869, ПЦР проводили как в Примере 2-1 с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы наряду с праймерами SEQ ID NO: 50 и 51 для амплификации фрагментов гена. В частности, ПЦР проводили в течение 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения цепи при 72°С в течение 30 секунд или 1 минуты 30 секунд. Полученные таким образом ПЦР-фрагменты подвергали электрофорезу в 0,8% агарозном геле и материал из полос, соответствующих желаемым размерам, элюировали и очищали.
Вектор pDZTn обрабатывали SpeI и затем ПЦР-продукты подвергали клонированию слиянием, соответственно. Клонирование слиянием выполняли с использованием набора для клонирования In-Fusion® HD Cloning Kit (Clontech). Полученная таким образом плазмида получила название pDZTn2'-gltA. Плазмидой pDZTn2'-gltA трансформировали штамм Corynebacterium glutamicum DAB12-b также как в Примере 2-1 и таким образом отбирали штамм, у которого gltA был усилен. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum получил название DAB12-b Tn:gltA.
Пример 3: Получение штаммов, обладающих продуктивностью по путресцину. с объединением ослабления по sugR и усиления gltA и подтверждение продуктивности этих штаммов по путресцину
Для подтверждения улучшения продуктивности по путресцину штаммов, ослабленных по sugR, полученных в Примерах 1-1 и 1-2 путем вставки гена gltA, ген gltA вводили в ген транспозона. В частности, использовали векторы pDZTn1'-gltA и pDZTn2'-gltA, полученные в Примерах 2-1 и 2-2.
В частности, плазмидой pDZTn1'-gltA трансформировали Corynebacterium glutamicum КССМ11240Р sugR(GTG), -КССМ11240Р sugR(TTG) и -KCCM11240P sugR(B6) таким же образом как в Примере 2-1 для получения штаммов, усиленных по gltA. Полученные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия КССМ11240Р sugR(GTG) Tn::gltA, КССМ11240Р sugR(TTG) Tn::gltA и KCCM11240P sugR(B6) Tn::gltA, соответственно, и среди них КССМ11240Р sugR(TTG) Tn::gltA (Corynebacterium glutamicum CC01-1147) был депонирован в Корейском центре культур микроорганизмов (КССМ) 28 ноября 2014 года под номером доступа КССМ11615Р.
Дополнительно, плазмидой pDZTn2'-gltA трансформировали Corynebacterium glutamicum DAB12-b sugR(GTG), -DAB12-b sugR(TTG) и -DAB12-b sugR(B6) таким же образом, как в Примере 2-2, для получения штаммов, усиленных по gltA. Полученные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия DAB12-b sugR(GTG) Tn::gltA, DAB12-b sugR(TTG) Tn::gltA и DAB12-b sugR(B6) Tn::gltA, соответственно.
Пример 4: Оценка продуктивности по путресцину штаммов, обладающих продуктивностью по путресцину. с объединением ослабления sugR и усиления gltA
Для подтверждения эффекта ослабления sugR и усиления gltA у штаммов, обладающих продуктивностью по путресцину, в отношении продуцирования путресцина сравнивали продуктивность по путресцину модифицированных штаммов Corynebacterium glutamicum, обладающих продуктивностью по путресцину, полученных в Примерах 1, 2 и 3.
Более конкретно, 6 различных типов модифицированных штаммов Corynebacterium glutamicum (а именно КССМ11240Р sugR (GTG) Tn::gltA/ КССМ11240Р sugR (TTG) Tn::gltA/ KCCM11240P sugR (B6) Tn::gltA/ DAB12-b sugR (GTG) Tn::gltA/ DAB12-b sugR (TTG) Tn::gltA и DAB12-b sugR (B6) Tn::gltA)) и 2 различных типа родительских штаммов (а именно КССМ11240Р и DAB12-b) высевали соответственно на чашки со средой СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50% NaOH, 2% агара, рН 6,8, в расчете на 1 л), содержащей 1 мМ аргинин, и культивировали при 30°С в течение 24 часов.
Каждый из культивируемых таким образом штаммов в количестве примерно одной платиновой петли инокулировали в 25 мл среды для титрования (8% глюкозы, 0,25% соевого белка, 0,50% твердой фазы кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, 100 г биотина, 3 мг тиамина HCl, 3 мг кальций-пантотеновой кислоты, 3 мг никотинамида, 5% СаСО3, в расчете на 1 л), и культивировали при 30°С со встряхиванием со скоростью 200 об/мин в течение 50 часов.
В среду ко всем культивируемым штаммам добавляли 1 мМ аргинин. После завершения культивирования измеряли концентрацию путресцина, полученного в каждой культуре, и результаты представлены в Таблице 5 ниже. 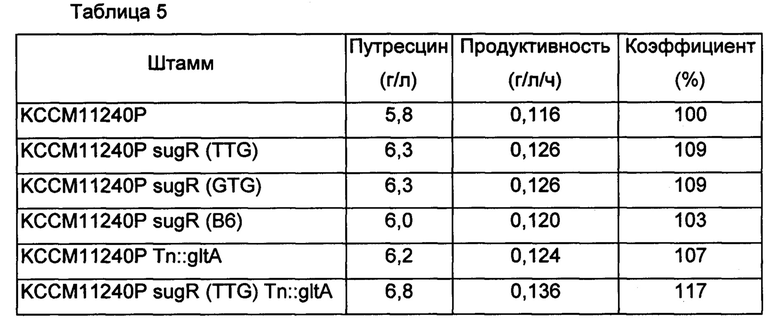
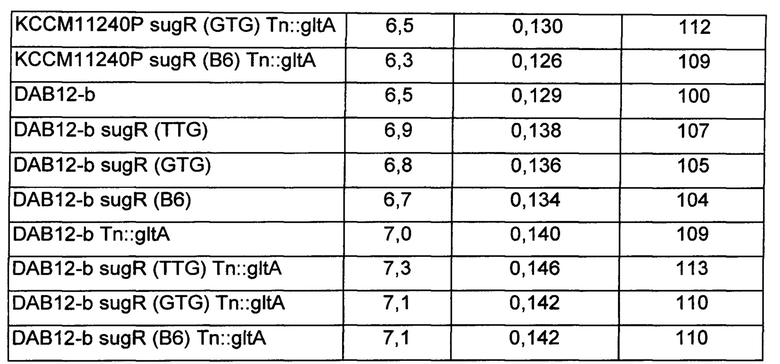
Как показано в Таблице 5 выше, модифицированные штаммы Corynebacterium glutamicum с ослабленным sugR или усиленным gltA демонстрировали увеличение продуктивности по путресцину по сравнению с немодифицированным штаммом КССМ11240Р на величину от 3% до 9%, и также модифицированные штаммы Corynebacterium glutamicum с одновременным ослабленным sugR и усиленным gltA демонстрировали увеличение продуктивности по путресцину на величину от 9% до 17%.
Кроме того, модифицированные варианты штамма DAB12-b с ослабленным sugR или усиленным gltA демонстрировали увеличение продуктивности по путресцину по сравнению с немодифицированным штаммом на величину от 4% до 9%, и также модифицированные варианты штамма DAB12-b с одновременно ослабленным sugR и усиленным gltA демонстрировали увеличение продуктивности по путресцину на величину от 10% до 13%.
Пример 5: Получение штаммов с повышенной способностью к секреции путресцина на основе штаммов, обладающих продуктивностью по путресцину, с объединением ослабления sugR и усиления gltA, и подтверждение продуктивности этих штаммов по путресцину.
5-1. Получение штаммов с повышенной способностью к секреции путресцина на основе штаммов с объединением ослабления sugR и усиления gltA
Для подтверждения того, можно ли у штамма КССМ11401Р с повышенной способностью к секреции путресцина (Публикация заявки на патент Кореи No. 10-2014-0115244) улучшить продуктивность по путресцину путем ослабления активности гена sugR и усиления активности гена gltA, были получены модифицированные штаммы.
Более конкретно, сначала плазмидами pDZ-1'sugR(GTG), pDZ-1'sugR(TTG) и pDZ-1'sugR(B6), полученными в Примере 1-1, трансформировали Corynebacterium glutamicum КССМ 11401Р, и из них были выбраны штаммы, у которых инициирующий кодон sugR был преобразован в TTG, что в результате приводило к ослаблению sugR. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия КССМ11401Р sugR(GTG), КССМ11401Р sugR(TTG) и КССМ11401Р sugR(B6), соответственно.
Затем, для подтверждения того, может ли продуктивность по путресцину быть улучшена путем повышения активности гена gltA, ген gltA вводили в ген транспозона штаммов с ослабленным геном sugR, полученных выше. В частности, использовали вектор pDZTn1'-gltA, полученный в Примере 2-1.
Более конкретно, плазмидой pDZTn1'-gltA, полученной в Примере 2-1, трансформировали КССМ11401Р sugR(GTG), КССМ11401Р sugR(TTG) и КССМ11401Р sugR(B6), и отбирали штаммы, усиленные по gltA. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия КССМ11401Р sugR(GTG) Tn::gltA, КССМ11401Р sugR(TTG) Tn::gltA и KCCM11401P sugR(B6) Tn::gltA.
5-2. Оценка штаммов с повышенной способностью к секреции путресцина на основе штаммов, обладающих продуктивностью по путресцину. с объединением ослабления sugR и усиления gltA в отношении продуктивности по путресцину
Для подтверждения эффекта ослабления sugR и усиления gltA у штаммов, обладающих продуктивностью по путресцину, в отношении их продуцирования путресцина сравнивали продуктивность по путресцину у модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 5-1.
Более конкретно, 7 различных типов модифицированных штаммов Corynebacterium glutamicum (то есть, КССМ11401Р sugR(GTG), КССМ11401Р sugR(TTG), КССМ11401Р sugR(B6), КССМ11401Р Tn::gltA, KCCM11401P sugR(GTG) Tn::gltA, KCCM11401P sugR(TTG) Tn::gltA и KCCM11401P sugR(B6) Tn::gltA) и один родительский штамм (КССМ11401Р) высевали, соответственно, на чашку со средой СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50% NaOH, 2% агара, рН 6,8, в расчете на 1 л), содержащей 1 мМ аргинин, и культивировали при 30°С в течение 24 часов.
Каждый из культивируемых таким образом штаммов в количестве примерно одной платиновой петли инокулировали в 25 мл среды для титрования (8% глюкозы, 0,25% соевого белка, 0,50% твердой фазы кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4-7H2O, 0,15% мочевины, 100 г биотина, 3 мг тиамина HCl, 3 мг кальций-пантотеновой кислоты, 3 мг никотинамида, 5% СаСО3 1 л, в расчете на 1 л), и культивировали при 30°С со встряхиванием со скоростью 200 об/мин в течение 50 часов.
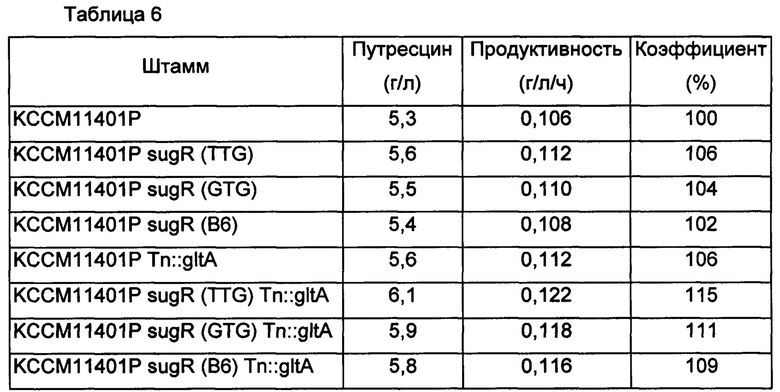
Как показано в Таблице 6 выше, модифицированные штаммы Corynebacterium glutamicum с ослабленным sugR или усиленным gltA демонстрировали повышение продуктивности по путресцину по сравнению с немодифицированным штаммом КССМ11401Р на величину от 2% до 6%, и также модифицированные штаммы Corynebacterium glutamicum с одновременно ослабленным sugR и усиленным gltA демонстрировали повышение продуктивности по путресцину на величину от 9% до 15% по продуктивности по путресцину. Было подтверждено, что эти результаты согласуются с интерпретацией результатов, представленных в Таблице 5.
Пример 6. Получение штаммов, ослабленных по sugR. из штамма, обладающего продуктивностью по орнитину
Для подтверждения того, оказывает ли ослабление sugR, происходящего из Corynebacterium glutamicum АТСС13032, эффект в отношении продуктивности по орнитину, были получены модифицированные штаммы с использованием векторов, полученных в Примере 1-1.
Плазмиды, полученные в Примере 1-1, то есть pDZ-1'sugRfGrGJ, pDZ-1'sugR(TTG) и pDZ-1'sugR(B6), вводили в штамм КССМ11137Р (Патент Кореи No. 10-1372635), который был получен с использованием Corynebacterium glutamicum АТСС13032 в качестве родительского штамма, путем электропорации для получения трансформантов и трансформанты высевали на чашку со средой BHIS (бульон с сердечно-мозговым экстрактом 37 г/л, сорбит 91 г/л и агар 2%), содержащей канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с получением колоний. Среди колоний отбирали голубые колонии, и таким образом были отобраны штаммы, в которые были введены плазмиды pDZ-1'sugR(GTG), pDZ-1'sugR(TTG) и pDZ-1'sugR(B6).
Выбранные штаммы культивировали при встряхивании (30°С, 8 часов) в среде СМ (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л, мочевина 2 г/л, рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Среди колоний, образованных таким образом, отбирали белые колонии, которые появлялись в относительно небольшом количестве, и в итоге отбирали штаммы, у которых инициирующий кодон sugR был заменен на GTG или TTG путем вторичного кроссинговера, или штаммы, у которых промотор был заменен на В6. В отношении отобранных в итоге штаммов проводили ПЦР с использованием пары праймеров SEQ ID NO: 43 и 47 и затем подтверждали, что инициирующий кодон sugR был заменен на GTG или TTG. Полученные модифицированные штаммы Corynebacterium glutamicum получили названия КССМ11137Р sugR(GTG), КССМ11137Р sugR(TTG) и КССМ11137Р sugR(B6), соответственно.
Пример 7. Получение штаммов, усиленных по gltA. из штаммов, обладающих продуктивностью по орнитину Для подтверждения эффекта усиления гена gltA у штамма, обладающего продуктивностью по орнитину, на его продуцировние орнитина, был получен модифицированный штамм путем вставки гена gltA в хромосому штамма, обладающего продуктивностью по орнитину, с использованием векторов, полученных в Примере 2-1.
Более конкретно, векторы, полученные в Примере 2-1, вводили в штамм КССМ11137Р (Патент Кореи No. 10-1372635) путем электропорации для получения трансформантов и трансформанты культивировали при встряхивании (30°С, 8 часов) в среде СМ (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л, мочевина 2 г/л, рН 6,8), последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний.
Среди колоний, образованных таким образом, отбирали белые колонии, которые появлялись в относительно небольшом количестве, и в итоге отбирали штаммы, которым был введен ген, кодирующий gltA, путем вторичного кроссинговера. В отношении отобранных в итоге штаммов проводили ПЦР с использованием пары праймеров SEQ ID NO: 50 и 51 и подтверждали, что им был введен ген, кодирующий gltA, и модифицированный штамм Corynebacterium glutamicum получил название КССМ11137Р Tn::gltA.
Пример 8. Получение штаммов с объединением ослабления sugR и усиления gltA и подтверждение продуктивности этих штаммов по путресцину
8-1. Получение штаммов на основе АТСС13032, обладающих продуктивностью по орнитину, с объединением ослабления sugR и усиления gltA
Для подтверждения действия по усилению активности продуктивности по орнитину у ослабленных по sugR КССМ11137Р sugR(GTG), КССМ11137Р sugR(TTG) и КССМ11137Р sugR(B6), полученных в Примере 6 путем вставки гена gltA в хромосому, ген gltA вводили в ген транспозона. В частности, использовали вектор pDZTn1'-gltA, полученный в Примере 2-1.
Плазмидой pDZTn1'-gltA трансформировали Corynebacterium glutamicum КССМ11137Р sugR TTG таким же образом как в Примере 2-1 и отбирали штаммы, усиленные по gltA. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum получили названия КССМ11137Р sugR(GTG) Tn::gltA, KCCM11137P sugR(TTG) Tn::gltA и KCCM11137P sugR(B6) Tn::gltA, соответственно.
8-2. Оценка штаммов с объединением ослабления sugR и усиления gltA в отношении продуктивности по орнитину
Для подтверждения эффекта ослабления sugR и усиления gltA у штаммов, обладающих продуктивностью по орнитину, на их продуцирование орнитина, сравнивали продуктивность по орнитину у модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 8-1.
Более конкретно, 7 различных типов модифицированных штаммов Corynebacterium glutamicum (то есть, КССМ11137Р sugR(GTG), КССМ11137Р sugR(TTG), КССМ11137Р sugR(B6), КССМ11137Р Tn::gltA, KCCM11137P sugR(GTG) Tn::gltA, KCCM11137P sugR(TTG) Tn::gltA и KCCM11137P sugR(B6) Tn::gltA) и один родительский штамм (КССМ11137Р), соответственно, высевали на чашку со средой СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50% NaOH, 2% агара, рН 6,8, в расчете на 1 л), содержащей 1 мМ аргинин, и культивировали при 30°С в течение 24 часов.
Каждый из культивированных таким образом штаммов в количестве примерно одной платиновой петли инокулировали в 25 мл среды для титрования (8% глюкозы, 0,25% соевого белка, 0,50% твердой фазы кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4-7H2O, 0,15% мочевины, 100 г биотина, 3 мг тиамина HCl, 3 мг кальций-пантотеновой кислоты, 3 мг никотинамида, 5% СаСО3, в расчете на 1 л), и культивировали со встряхиванием со скоростью 200 об/мин при 30°С в течение 50 часов. В среду ко всем культурам штаммов добавляли 1 мМ аргинин. По завершении культивирования измеряли концентрацию орнитина, продуцированного в каждой культуре, и результаты представлены в Таблице 7 ниже. 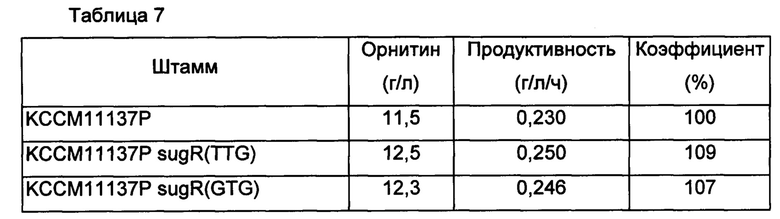
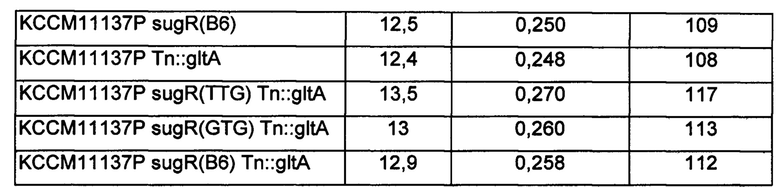
Как показано в Таблице 7 выше, модифицированные штаммы Corynebacterium glutamicum с ослабленным sugR или усиленным gltA показали увеличение продуктивности по орнитину по сравнению с немодифицированным штаммом КССМ11137Р на величину от 7% до 9%, и также модифицированные штаммы Corynebacterium glutamicum с одновременно ослабленным sugR и усиленным gltA демонстрировали увеличение продуктивности по орнитину на величину от 12% до 17% по продуктивности по орнитину.
В заключение, было подтверждено, что у штамма Corynebacterium, продуцирующего путресцин или орнитин, продуцирование путресцина и орнитина может быть увеличено путем ослабления sugR или усиления gltA, и когда gltA был усилен при одновременном ослаблении sugR, продуцирование путресцина и орнитина увеличивалось в большей степени.
На основании вышеизложенного специалист в области техники, к которой принадлежит данное изобретение, понимает, что настоящее изобретение может быть воплощено в других конкретных формах без отступления от технических идей или существенных признаков настоящего изобретения. В этом отношении примеры воплощений, раскрытые здесь, служат исключительно для иллюстративных целей и не предназначены ограничивать объем настоящего изобретения. С другой стороны, настоящее изобретение предназначено охватывать не только примеры воплощений, но также различные варианты, модификации, эквиваленты и другие воплощения, которые могут быть включены в пределах идеи и объема настоящего изобретения как определено в прилагаемой формуле.
Группа изобретений относится к области биотехнологии. Предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин, с пониженной активностью регулятора транскрипции метаболизма сахара (SugR) по сравнению с его эндогенной активностью и повышенной активностью цитратсинтазы (GltA) по сравнению с ее эндогенной активностью. Предложен способ получения путресцина с использованием указанного микроорганизма. Группа изобретений позволяет увеличить продуктивность указанного модифицированного микроорганизма по путресцину по сравнению с немодифицированным микроорганизмом. 2 н. и 13 з. п. ф-лы, 7 табл., 8 пр.
1. Модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин, с пониженной активностью регулятора транскрипции метаболизма сахара (SugR) по сравнению с его эндогенной активностью и повышенной активностью цитратсинтазы (GltA) по сравнению с ее эндогенной активностью.
2. Микроорганизм по п. 1, где регулятор транскрипции метаболизма сахара состоит из аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 3.
3. Микроорганизм по п. 1, где цитратсинтаза состоит из аминокислотной последовательности SEQ ID NO: 5 или SEQ ID NO: 7.
4. Микроорганизм по п. 1, где микроорганизм рода Corynebacterium выбран из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum.
5. Микроорганизм по п. 1, где дополнительно введена активность орнитин-декарбоксилазы (ODC).
6. Микроорганизм по п. 5, где орнитин-декарбоксилаза состоит из аминокислотной последовательности SEQ ID NO: 17.
7. Микроорганизм по п. 1, где активность 1) орнитин-карбамоилтрансферазы (ArgF), 2) экспортера глутамата или 3) орнитин-карбамоилтрансферазы и экспортера глутамата дополнительно понижена по сравнению с их эндогенной активностью.
8. Микроорганизм по п. 7, где орнитин-карбамоилтрансфераза состоит из аминокислотной последовательности SEQ ID NO: 9 или SEQ ID NO: 11, и экспортер глутамата состоит из аминокислотной последовательности SEQ ID NO: 13 или SEQ ID NO: 15.
9. Микроорганизм по п. 1, где активность по меньшей мере одного выбранного из группы, состоящей из ацетил-гамма-глутамил-фосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитин-ацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитин-аминотрансферазы (ArgD), дополнительно повышена по сравнению с его эндогенной активностью.
10. Микроорганизм по п. 9, где ацетил-гамма-глутамил-фосфатредуктаза состоит из аминокислотной последовательности SEQ ID NO: 19 или SEQ ID NO: 21, ацетилглутаматсинтаза или орнитин-ацетилтрансфераза состоит из аминокислотной последовательности SEQ ID NO: 23 или SEQ ID NO: 25, ацетилглутаматкиназа состоит из аминокислотной последовательности SEQ ID NO: 27 или SEQ ID NO: 29, и ацетилорнитин-аминотрансфераза состоит из аминокислотной последовательности SEQ ID NO: 31 или SEQ ID NO: 33.
11. Микроорганизм по п. 1, где активность ацетилтрансферазы дополнительно понижена по сравнению с ее эндогенной активностью.
12. Микроорганизм по п. 11, где ацетилтрансфераза состоит из аминокислотной последовательности SEQ ID NO: 35 или SEQ ID NO: 37.
13. Микроорганизм по п. 1, где активность белка, состоящего из SEQ ID NO: 39 или SEQ ID NO: 41, дополнительно повышена по сравнению с его эндогенной активностью.
14. Способ получения путресцина, включающий:
(1) культивирование микроорганизма рода Corynebacterium по любому из пп. 1-13 в среде; и
(2) выделение путресцина из культивированного микроорганизма или из культуральной среды.
15. Способ по п. 14, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
| JP 2009254323 A, 05.11.2009 | |||
| KR 101372635 B1, 13.03.2014 | |||
| NCBI, NCBI Reference Sequence: WP_011014761.1, 18.07.2013 | |||
| NCBI, NCBI Reference Sequence: WP_011013914.1, 22.03.2015 | |||
| KR 20130082478 A, 19.07.2013 | |||
| NCBI, NCBI Reference Sequence: WP_011015248.1, 18.07.2013 | |||
| BLOMBACH B | |||
| et al., L-Valine Production during Growth of Pyruvate Dehydrogenase ComplexDeficient Corynebacterium glutamicum in the Presence of Ethanol or by Inactivation of the Transcriptional Regulator SugR, APPLIED AND ENVIRONMENTAL MICROBIOLOGY Vol | |||
| Фальцовая черепица | 0 |
|
SU75A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Паровой котел с перегревателем | 1925 |
|
SU1197A1 |
| RU 2013131033 A, 20.01.2015 | |||
| RU 2013131039 A, 20.01.2015. | |||

