Область техники
Описание настоящего изобретения относится к микроорганизму для продуцирования диамина и способу получения диамина с его использованием.
Предшествующий уровень техники
Биогенные амины (ВА) представляют собой азотистые соединения, которые получаются главным образом в результате декарбоксилирования аминокислот или аминирования и трансаминирования альдегидов и кетонов. Эти биогенные амины являются низкомолекулярными соединениями и синтезируются в ходе метаболизма микроорганизмами, растениями и животными, и поэтому биогенные амины известны как компоненты, часто встречающиеся в этих клетках. В частности, биогенные амины представляют собой полиамины, такие как спермидин, спермин, путресцин или 1,4-бутандиамин и кадаверин.
В общем случае, путресцин является важным исходным веществом при получении полиамина найлона-4,6, который образуется при взаимодействии путресцина с адипиновой кислотой. Путресцин обычно получают путем химического синтеза, включающего превращение пропилена в акрилонитрил и в сукцинонитрил.
В качестве способа получения путресцина с использованием микроорганизмов в литературе упомянут способ продуцирования путресцина в высокой концентрации с использованием трансформации Е. coli и Corynebacterium (публикация международной патентной заявки № WO 06/005603; публикация международной патентной заявки № W O 09/125924; Qian ZD et al., Biotechnol. Bioeng., 104: 4, 651-662, 2009; Schneider et al., Appl. Microbiol. Biotechnol., 88: 4, 859-868, 2010; Schneider et al., Appl. Microbiol. Biotechnol., 95, 169-178, 2012). Кроме того, были проведены активные исследования переносчиков путресцина в Е. coli, дрожжах, клетках растений и животных (K. Igarashi, Plant Physiol. Biochem., 48: 506-512, 2010).
Что касается кадаверина, то он представляет собой диаминовое соединение с неприятным запахом, образующееся в результате гидролиза белка при разложении тканей животных. Кадаверин имеет химическую формулу NH2(CH2)5NH2, которая похожа на таковую путресцина.
Кадаверин служит в качестве одного из компонентов таких полимеров, как полиамид или полиуретан, хелатирующих агентов или других вспомогательных веществ. В частности, полиамид с ежегодным оборотом мирового рынка, составляющим 3,5 миллиона тонн, как известно, получают поликонденсацией кадаверина или янтарной кислоты, и поэтому кадаверину уделяется большое внимание как промышленно важному соединению.
Кадаверин представляет собой диамин, обнаруженный в нескольких микроорганизмах (Tabor and Tabor, Microbiol. Rev., 49: 81-99, 1985). Биосинтез кадаверина в грамотрицательной бактерии Е. coli осуществляется из L-лизина под действием L-лизин-декарбоксилазы. Уровень кадаверина в Е. coli регулируется в ходе биосинтеза, деградации, поглощения и секреции кадаверина (Soksawatmaekhin et al., Mol. Microbiol., 51: 1401-1412, 2004).
Техническая проблема
Авторы настоящего изобретения приложили интенсивные усилия для исследования белка, обладающего способностью экспортировать диамин, такой как путресцин или кадаверин, с целью усиления продуцирования диаминов микроорганизмом, обладающим диамин-продуцирующей способностью. В результате они обнаружили, что белок, происходящий из Arcanobacterium haemolyticum, или белок, имеющий с ним высокую гомологию аминокислотной последовательности, обладает диамин-экспортирующей активностью, и этот белок вводят в микроорганизм для продуцирования диамина с целью повышения его эффективности, что приводит к значительному повышению способности экспортировать диамин, такой как путресцин и кадаверин, с решением тем самым задачи настоящего изобретения.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить микроорганизм для продуцирования диамина.
Другой задачей настоящего изобретения является предложение способа получения диамина, включающего стадии (1) культивирования микроорганизма для продуцирования диамина с получением клеточной культуры; и (2) извлечения диамина из подвергнутого культивированию микроорганизма или клеточной культуры.
Наилучший вариант воплощения изобретения
Согласно одному из аспектов настоящего изобретения, для решения указанных выше задач предложен микроорганизм для продуцирования диамина, где активность белка, имеющего аминокислотную последовательность SEQ ID NO: 6, или аминокислотную последовательность, имеющую гомологию последовательности с SEQ ID NO: 6, составляющую 42% или больше, введена или повышена.
Использованный в данном описании термин «диамин» в совокупности относится к соединению, имеющему две аминогруппы, и его конкретные примеры могут включать путресцин и кадаверин. Путресцин представляет собой тетраметилендиамин, который может быть получен из орнитина в качестве предшественника. Кадаверином называют 1,5-пентандиамин или пентаметилендиамин, который может быть получен из лизина в качестве предшественника. Такие диамины используются в промышленности в качестве полезных исходных веществ для синтеза таких полимеров, как полиамин найлон, полиамид или полиуретан.
Использованный в данном описании термин «белок, имеющий аминокислотную последовательность SEQ ID NO: 6» относится к белку, обнаруженному в Arcanobacterium haemolyticum, и также названному ARCH_0271. Было выяснено, что этот белок сохраняет высокую гомологию с мембранным белком коринебактерии, NCgl2522. В одном из воплощений настоящего изобретения белок ARCH_0271 идентифицирован как белок, предположительно вовлеченный в экспорт диаминов в штамме, обладающем диамин-продуцирующей способностью, что тем самым значительно усиливает продуцирование диаминов.
В данной заявке белок ARCH_0271, имеющий аминокислотную последовательность SEQ ID NO: 6, может представлять собой белок, кодируемый нуклеотидной последовательностью SEQ ID NO: 5 или SEQ ID NO: 7. Однако в полинуклеотиде, кодирующем белок ARCH_0271, могут быть выполнены различные модификации в кодирующей области при условии, что они не приводят к изменению аминокислотной последовательности полипептида, экспрессируемого с этой кодирующей области, обусловленные вырожденностью кодонов или соображениями относительно предпочтительных кодонов в микроорганизме, в котором будет экспрессирован белок. Так, белок СЕ2495 может кодироваться различными нуклеотидными последовательностями помимо нуклеотидной последовательности с SEQ ID NO: 5 или SEQ ID NO: 7. Среди них, нуклеотидная последовательность, представленная SEQ ID NO. 7, является последовательностью, полученной посредством оптимизации гена ARCH_0271 (SEQ ID NO: 5) в отношении частоты использования кодонов в Corynebacterium glutamicum, но не ограничивается ею.
Кроме того, белок ARCH_0271 по настоящему изобретению может представлять собой любой белок, имеющий аминокислотную последовательность SEQ ID NO: 6 или имеющий с ним гомологию 40% или больше, предпочтительно 60% или больше, более предпочтительно 80% или больше, еще более предпочтительно 90% или больше, еще более предпочтительно 95% или больше и наиболее предпочтительно 99% или больше, при условии, что данный белок проявляет значительную диамин- экспортирующую активность. Очевидно, что имеющая такую гомологию аминокислотная последовательность, часть которой изменена с использованием делеций, модификаций, замен или вставок, также включена в объем настоящего изобретения, при условии, что белок с полученной в результате этого аминокислотной последовательностью обладает биологической активностью, по существу эквивалентной или соответствующей белку с SEQ ID NO: 6.
Использованный в данном описании термин «белок, имеющий аминокислотную последовательность, имеющую гомологию последовательности с аминокислотной последовательностью SEQ ID NO: 6, составляющую 42% или больше» означает любой белок без ограничения, при условии, что данный белок имеет аминокислотную последовательность, имеющую гомологию последовательности с аминокислотной последовательностью SEQ ID NO: 6, составляющую 42% или больше, и помимо этого он обладает значительной диамин-экспортирующей активностью. Например, белок может представлять собой белок, имеющий аминокислотную последовательность SEQ ID NO: 23 или SEQ ID NO: 26, но этим не ограничивается.
Например, белок, имеющий аминокислотную последовательность SEQ ID NO: 23, представляет собой белок, обнаруженный в Alcaligenes faecalis, подвиде faecalis и также названный QWA_00075. Было выяснено, что этот белок сохраняет 41% гомологии с мембранным белком коринебактерии NCgl2522 и 42% гомологии с ARCH_0271 из Arcanobacterium haemolyticum. В одном из воплощений настоящего изобретения было выяснено, что белок QWA_00075 проявляет диамин-экспортирующую активность в штамме, обладающем диамин-продуцирующей способностью, тем самым значительно усиливая продуцирование диаминов.
Белок QWA_00075, имеющий аминокислотную последовательность SEQ ID NO: 23, может представлять собой белок, кодируемый нуклеотидной последовательностью SEQ ID NO: 22 или 24. Однако в полинуклеотиде, кодирующем этот белок, могут быть выполнены различные модификации в кодирующей области при условии, что они не приводят к изменению аминокислотной последовательности полипептида, экспрессируемого с этой кодирующей области, обусловленные вырожденностью кодонов или соображениями относительно предпочтительных кодонов в микроорганизме, в котором будет экспрессирован белок. Так, этот белок может кодироваться различными нуклеотидными последовательностями помимо нуклеотидной последовательности SEQ ID NO: 22 или SEQ ID NO: 24. Среди них, нуклеотидная последовательность, представленная SEQ ID NO: 24, является последовательностью, полученной посредством оптимизации гена QWA_00075 (SEQ ID NO: 22) в отношении частоты использования кодонов в Corynebacterium glutamicum, но не ограничивается ею.
Кроме того, белок, имеющий аминокислотную последовательность SEQ ID NO: 26, представляет собой белок, обнаруженный в Stenotrophomonas maltophilia и также названный SMD_2351. Было выяснено, что этот белок сохраняет 52% гомологии с мембранным белком коринебактерии NCgl2522 и 47% гомологии с ARCH_0271 из Arcanobacterium haemolyticum. В одном из воплощений настоящего изобретения было выяснено, что белок SMD_2351 проявляет диамин-экспортирующую активность в штамме, обладающем диамин-продуцирующей способностью, тем самым значительно усиливая продуцирование диаминов.
Белок SMD_2351, имеющий аминокислотную последовательность SEQ ID NO: 26, может представлять собой белок, кодируемый нуклеотидной последовательностью SEQ ID NO: 25 или 27. Однако, в полинуклеотиде, кодирующем этот белок, могут быть выполнены различные модификации в кодирующей области при условии, что они не приводят к изменению аминокислотной последовательности полипептида, экспрессируемого с этой кодирующей области, обусловленные вырожденностью кодонов или соображениями относительно предпочтительных кодонов в микроорганизме, в котором будет экспрессирован белок. Так, этот белок может кодироваться различными нуклеотидными последовательностями помимо нуклеотидной последовательности SEQ ID NO: 25 или SEQ ID NO: 27. Среди них, нуклеотидная последовательность, представленная SEQ ID NO. 27, является последовательностью, полученной посредством оптимизации гена SMD_2351 (SEQ ID NO: 25) в отношении частоты использования кодонов в Corynebacterium glutamicum, но не ограничивается ею.
Термин «гомология», использованный в данном описании в отношении последовательности, относится к идентичности с заданной аминокислотной последовательностью или нуклеотидной последовательностью, и гомология может быть выражена в процентах. В настоящем изобретении, гомологию последовательности, обладающей идентичной или схожей активностью с заданной аминокислотной последовательностью или нуклеотидной последовательностью, выражают в виде «% гомологии». Например, гомология может быть установлена с использованием стандартной программы из системы программного обеспечения, с помощью которой рассчитывают параметры оценки, идентичность и сходство, главным образом BLAST 2.0, либо путем сравнения последовательностей в эксперименте с использованием гибридизации по Саузерну в жестких условиях, как указано. Определение соответствующих условий гибридизации находится в пределах компетенции специалистов в данной области техники (например, см. Sambrook et al., 1989, ниже), и их определяют способом, известным специалистам в данной области техники.
Использованный в данном описании термин «микроорганизм для продуцирования диамина» относится к микроорганизму, полученному путем придания способности продуцировать диамины родительскому штамму, не обладающему диамин-продуцирующей способностью, или к микроорганизму, обладающему эндогенной диамин-продуцирующей способностью. Конкретно, микроорганизмом, обладающим диамин-продуцирующей способностью, может быть микроорганизм, обладающий путресцин- или кадаверин-продуцирующей способностью.
«Микроорганизм, обладающий путресцин-продуцирующей способностью», может представлять собой, но не ограничивается им, микроорганизм, в котором активности ацетилглутаматсинтазы, катализирующей превращение глутамата в N-ацетилглутамат, орнитинацетилтрансферазы (ArgJ), катализирующей превращение ацетилорнитина в орнитин, ацетилглутаматкиназы (ArgB), катализирующей превращение ацетилглутамата в N-ацетилглутамилфосфат, ацетил-гамма-глутамил-фосфатредуктазы (ArgC), катализирующей превращение ацетилглутамилфосфата в N-ацетилглутамат-полуальдегид, или ацетилорнитинаминотрансферазы (ArgD), катализирующей превращение ацетилглутамат-полуальдегида в N-ацетилорнитин, повышены по сравнению с их эндогенной активностью с целью усиления пути биосинтеза от глутамата до орнитина, и в котором способность к продуцированию орнитина, использующегося в качестве предшественника для биосинтеза путресцина, повышена, но этим не ограничивается.
Кроме того, микроорганизмом, обладающим путресцин-продуцирующей способностью, может быть микроорганизм, который модифицирован с целью изменения активностей орнитинкарбамоилтрансферазы (ArgF), вовлеченной в синтез аргинина из орнитина, белка (NCgl1221), вовлеченного в экспорт глутамата, и/или белка (NCgl469), вовлеченного в ацетилирование путресцина, в сторону ослабления_по сравнению с их эндогенной активностью, и/или который модифицирован с целью введения активности орнитиндекарбоксилазы (ODC).
В данной заявке, в качестве неограничивающих примеров, ацетил-гамма- глутамилфосфатредуктаза (ArgC) может иметь аминокислотную последовательность SEQ ID NO: 15, ацетилглутаматсинтаза или орнитинацетилтрансфераза (ArgJ) может иметь аминокислотную последовательность SEQ ID NO: 16, ацетилглутаматкиназа (ArgB) может иметь аминокислотную последовательность SEQ ID NO: 17, а ацетилорнитинаминотрансфераза (ArgD) может иметь аминокислотную последовательность SEQ ID NO: 18. Однако аминокислотные последовательности соответствующих ферментов-белков конкретно не ограничены указанными последовательностями, и такие ферменты могут представлять собой белки, имеющие аминокислотные последовательности, имеющие с ними гомологию 80% или больше, предпочтительно 90% или больше либо более предпочтительно 95% или больше, при условии, что они обладают активностями соответствующих ферментов.
Кроме того, в качестве неограничивающих примеров, орнитинкарбамоилтрансфераза (ArgF) может иметь аминокислотную последовательность, представленную SEQ ID NO: 19, белок, вовлеченный в экспорт глутамата, может иметь аминокислотную последовательность, представленную SEQ ID NO: 20, а орнитиндекарбоксилаза (ODC) может иметь аминокислотную последовательность, представленную SEQ ID NO: 21. Однако аминокислотные последовательности соответствующих ферментов-белков конкретно не ограничены указанными последовательностями, и такие ферменты могут содержать аминокислотные последовательности, имеющие с ними гомологию 80% или больше, предпочтительно 90% или больше, более предпочтительно 95% или больше либо особенно предпочтительно 97% или больше, при условии, что они обладают активностями соответствующих ферментов.
Что касается «микроорганизма, обладающего кадаверин-продуцирующей способностью», то он может представлять собой, но не ограничиваться им, микроорганизм, полученный путем дополнительного введения лизиндекарбоксилазы (LDC) в микроорганизм или повышения ее активности в микроорганизме, обладающем лизин-продуцирующей способностью. Например, данным микроорганизмом может быть микроорганизм, обладающий повышенной лизин-продуцирующей способностью, с целью усиления продуцирования кадаверина. Способ повышения лизин-продуцирующей способности может быть осуществлен известным способом, очевидным специалисту в данной области техники.
Лизиндекарбоксилаза представляет собой фермент, катализирующий превращение лизина в кадаверин, и его вводят или повышают его активность, вследствие чего эффективно продуцируется кадаверин.
Лизиндекарбоксилаза может иметь аминокислотную последовательность SEQ ID NO: 33, но конкретно не ограничена ею. Фермент может иметь аминокислотную последовательность, имеющую с ней гомологию 80% или больше, предпочтительно 90% или больше либо более предпочтительно 95% или больше, при условии, что он обладает указанной выше активностью.
Использованный в данном описании термин «продуцирование» относится к понятию, включающему в себя высвобождение диамина во внеклеточное пространство, например, высвобождение диамина в культуральную среду, а также продуцирование диамина внутри микроорганизма.
Что касается термина «введение активности белка», как он использован в данном описании, то он означает, что в микроорганизм, не имеющий такого эндогенного белка, вводят извне активный белок, и это может быть осуществлено, например, посредством введения чужеродного гена. Помимо этого, термин «повышение активности белка» означает, что активное состояние белка, имеющегося в микроорганизме или введенного в микроорганизм, повышено по сравнению со свойственным ему активным состоянием.
Неограничивающие примеры введения или повышения активности белка могут включать улучшение активности самого белка, имеющегося в микроорганизме, обусловленное мутацией, проведенной с целью достижения эффектов, выходящих за рамки эндогенных функций, и/или улучшение активности эндогенного гена, кодирующего белок, присутствующий в микроорганизме, амплификацию эндогенного гена с использованием внутренних или внешних факторов, увеличение числа копий гена, повышение активности посредством дополнительного введения чужеродного гена либо замены или модификации промотора, но не ограничиваются этим.
Увеличение числа копий гена может быть осуществлено, но конкретно этим не ограничивается, посредством функционального связывания данного гена с вектором или посредством встраивания в геном клетки хозяина. В частности, число копий полинуклеотида в геноме клетки хозяина можно увеличить путем введения в клетку хозяина вектора, который функционально связан с полинуклеотидом, кодирующим белок по настоящему изобретению, и реплицируется и функционирует независимо от клетки хозяина, или путем введения в клетку хозяина вектора, который функционально связан с полинуклеотидом и способен обеспечить встраивание данного полинуклеотида в геном клетки хозяина.
Как использовано в данном описании, «модификация регулирующей экспрессию последовательности для усиления экспрессии полинуклеотида» может быть осуществлена, но конкретно этим не ограничивается, путем введения модификации в регулирующую экспрессию последовательность посредством делеции, вставки, неконсервативной или консервативной замены в нуклеотидной последовательности или их комбинации с целью дополнительного повышения активности регулирующей экспрессию последовательности либо путем замены регулирующей экспрессию последовательности на нуклеотидную последовательность, имеющую более высокую активность. Регулирующая экспрессию последовательность включает, но конкретно этим не ограничивается, промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции.
Как использовано в данном описании, замена или модификация промотора, хотя конкретно этим не ограничивается, может быть осуществлена путем замены на более сильный промотор или модификации с получением более сильного промотора по сравнению с исходным промотором. Сильный гетерологичный промотор, используемый вместо исходного промотора, может быть присоединен в положении вверх по течению по отношению к полинуклеотид-экспрессирующей единице, и примеры сильного промотора могут включать промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор асеА или асеВ и особенно промотор, происходящий из коринебактерий, промотор lysCP1 или промотор CJ7, функционально связанный с полинуклеотидом, кодирующим фермент, с тем, чтобы можно было повысить эффективность его экспрессии. В данной заявке промотор lysCP1 представляет собой промотор, улучшенный посредством замены нуклеотидной последовательности в промоторной области полинуклеотида, кодирующего аспартаткиназу и аспартат-полуальдегид-дегидрогеназу (WO 2009/096689). Помимо этого, промотор CJ7 является сильным промотором, происходящим из Corynebacterium ammoniagenes (патент Кореи №0620092 и заявка WO 2006/065095).
Кроме того, модификация полинуклеотидной последовательности в хромосоме, хотя конкретно этим не ограничивается, может быть осуществлена путем внесения мутации в регулирующую экспрессию последовательность посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации с целью дополнительного повышения активности полинуклеотидной последовательности, либо путем замены последовательности на полинуклеотидную последовательность, модифицированную с целью достижения более высокой активности.
Использованный в данном описании термин «вектор» относится к конструкции на основе ДНК, включающей нуклеотидную последовательность, кодирующую желаемый белок, функционально связанную с соответствующей регулирующей экспрессию последовательностью с целью экспрессии желаемого белка в подходящей клетке-хозяине. Такая регуляторная последовательность может включать промотор, который может инициировать транскрипцию, возможно последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы с мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После введения вектора в подходящую клетку- хозяина он может реплицироваться или функционировать независимо от генома хозяина и может быть встроен в сам геном.
Вектор, используемый в настоящем изобретении, не имеет конкретных ограничений при условии, что он способен реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры традиционных векторов могут включать существующую в природе или рекомбинантную плазмиду, космиду, вектор на основе вируса и бактериофага. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, λMBL3, λMBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A и Charon21A. Плазмиды pBR типа, pUC типа, pBluescriptll типа, pGEM типа, pTZ типа, pCL типа и рЕТ типа можно использовать в качестве плазмидного вектора. Вектор, который можно использовать в настоящем изобретении, не имеет конкретных ограничений, и можно использовать любой известный экспрессирующий вектор. Предпочтительно может быть использован вектор pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 или pCCIBAC.
Кроме того, полинуклеотид, кодирующий желаемый эндогенный белок в хромосоме, можно заменить на мутированный полинуклеотид, используя вектор для встраивания в бактериальную хромосому. Вставка полинуклеотида в хромосому может быть осуществлена любым методом, известным в данной области техники, например, посредством гомологической рекомбинации. Поскольку вектор по настоящему изобретению может быть встроен в хромосому посредством гомологической рекомбинации, он может дополнительно включать селективный маркер для подтверждения встраивания в хромосому. Селективный маркер необходим для отбора клеток, трансформированных вектором, то есть для подтверждения встраивания желаемого полинуклеотида, и селективный маркер может включать маркеры, обеспечивающие приобретение селектируемых фенотипов, таких как лекарственная устойчивость, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия белка на поверхности. Только клетки, экспрессирующие селективный маркер, способны выживать или демонстрировать разные фенотипы в среде, обработанной агентом для селекции, и таким образом можно отобрать трансформированные клетки.
Использованный в данном описании термин «трансформация» означает введение вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина таким образом, чтобы белок, кодируемый этим полинуклеотидом, экспрессировался в клетке хозяина. При условии, что введенный посредством трансформации полинуклеотид может экспрессироваться в клетке-хозяине, он может быть либо встроен и помещен в хромосому клетки-хозяина, либо может существовать экстрахромосомально. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид можно вводить в любой форме, при условии, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую в себя все элементы, необходимые для ее автономной экспрессии. Обычно, экспрессионная кассета включает в себя промотор, функционально связанный с полинуклеотидом, сигналы терминации транскрипции, сайты связывания рибосомы или сигналы терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося экспрессирующего вектора. Помимо этого, полинуклеотид может быть введен в клетку-хозяина сам по себе и может быть функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине.
Кроме того, использованный в данном описании термин «функционально связанный» означает функциональную связь между полинуклеотидной последовательностью, кодирующей желаемый белок по настоящему изобретению, и последовательностью промотора, с использованием которой инициируется и опосредуется транскрипция данной полинуклеотидной последовательности.
Кроме того, микроорганизмом, обладающим диамин-продуцирующей способностью, может быть микроорганизм, в котором диаминацетилтрансферазная активность снижена по сравнению с эндогенной активностью с целью усиления продуцирования диаминов.
Использованный в данном описании термин «диаминацетилтрансфераза» относится к ферменту, катализирующему перенос ацетильной группы от ацетил-КоА к диамину, и примером его может быть NCgl1469 из Corynebacterium glutamicum или SpeG из Е. coli, но его название может меняться в зависимости от вида микроорганизма, обладающего диамин-продуцирующей способностью. NCgl1469 может иметь аминокислотную последовательность SEQ ID NO: 12 или 13, a SpeG может иметь аминокислотную последовательность SEQ ID NO: 14, но последовательности могут отличаться в зависимости от вида микроорганизма. Данный белок может иметь аминокислотную последовательность, имеющую с ними гомологию, составляющую 80% или больше, предпочтительно 90% или больше, либо более предпочтительно 95% или больше, или особенно предпочтительно 97% или больше, при условии, что он обладает диаминацетилтрансферазной активностью.
Поскольку диаминацетилтрансфераза катализирует превращение диамина в ацетилдиамин (например, в N-Ac-путресцин или N-Ac-кадаверин), диамин-продуцирующая способность может быть повышена путем снижения ее активности по сравнению с эндогенной активностью.
Использованный в данном описании термин «эндогенная активность» относится к активности белка, который имеется у исходного микроорганизма в его нативном или не подвергнутом изменению естественных свойств состоянии, а «модифицированный с целью снижения активности по сравнению с эндогенной активностью» означает, что активность данного белка дополнительно снижена по сравнению с активностью соответствующего белка, которой обладает исходный микроорганизм в его нативном или не подвергнутом изменению естественных свойств состоянии.
Снижение активности белка означает, что активность белка снижена по сравнению с немодифицированным штаммом или что активность устранена. Для снижения активности белка можно применить способ, хорошо известный в данной области техники.
Примеры данного способа могут включать метод замены гена, кодирующего этот белок в хромосоме, на ген, мутированный с целью уменьшения ферментативной активности или с целью устранения активности белка, метод внесения мутации в регулирующую экспрессию последовательность гена, кодирующего белок в хромосоме, метод замены регулирующей экспрессию последовательности в гене, кодирующем белок, на последовательность, имеющую более слабую активность, метод делетирования части или всего гена, кодирующего белок в хромосоме, метод введения антисмыслового олигонуклеотида, который комплементарно связывается с транскриптом данного гена в хромосоме, с целью ингибирования трансляции мРНК в белок, метод искусственного добавления последовательности, комплементарной последовательности SD (Шайна-Дальгарно), в положении против хода транскрипции по отношению к SD-последовательности гена, кодирующего белок, с целью образования вторичной структуры с предотвращением тем самым доступа субъединиц рибосомы, и метод конструирования обратной транскрипции (RTE), осуществляемый путем добавления промотора для обратной транскрипции на 3'-конец открытой рамки считывания (ORF) соответствующей последовательности, и их комбинации, но конкретно ими не ограничены.
Более подробно, частичное или полное делетирование гена, кодирующего белок, может быть осуществлено путем введения в микроорганизм вектора для встраивания в хромосому, благодаря чему полинуклеотид, кодирующий эндогенный целевой белок в хромосоме, заменяется на полинуклеотид, имеющий частичную делецию или маркерный ген. Термин «частичный» может варьироваться в зависимости от типа полинуклеотида, но относится, например, к 1-300, предпочтительно 1-100 и более предпочтительно 1-50 нуклеотидам.
Что касается микроорганизма по настоящему изобретению, то он представляет собой микроорганизм, обладающий диамин-продуцирующей способностью, и включает прокариотический микроорганизм, экспрессирующий белок, имеющий аминокислотную последовательность SEQ ID NO: 6, и примеры его могут включать микроорганизмы, принадлежащие Escherichia sp., Shigella sp., Citrobacter sp., Salmonella sp., Enterobacter sp., Yersinia sp., Klebsiella sp., Erwinia sp., Corynebacterium sp., Brevibacterium sp., Lactobacillus sp., Selenomanas sp., Vibrio sp., Pseudomonas sp., Streptomyces sp., Arcanobacterium sp., Alcaligenes sp. или тому подобное, но ими не ограничиваются. Микроорганизм по настоящему изобретению представляет собой, в частности, микроорганизм, принадлежащий Corynebacterium sp. или Escherichia sp. и более конкретно, Corynebacterium glutamicum или Escherichia coli, но ими не ограничивается.
Конкретным примером может быть микроорганизм, полученный путем удаления NCgl2522, представляющего собой белок, обладающий путресцин-экспортирующей активностью, из путресцин-продуцирующего штамма KCCM11240P, полученного на основе Corynebacterium glutamicum АТСС13032 (Американская коллекция типовых культур) (публикация заявки на патент Кореи №2013-0082478), и затем введения ARCH_0271 в транспозонный ген. Таким образом, данный микроорганизм KCCM11240P ΔNCgl2522 Tn:P(cj7)-ARCH_0271 обозначен как СС01-0758 и депонирован в соответствии с Будапештским договором в Корейский центр культур микроорганизмов (KCCM) 15 ноября 2013 г. с номером доступа KCCM11476P.
Согласно другому аспекту настоящего изобретения предложен способ получения диамина, включающий: (1) культивирование микроорганизма, продуцирующего диамин путресцин, где активность белка имеющего аминокислотную последовательность SEQ ID NO: 6 или имеющего с ней гомологию последовательности, составляющую 42% или более, введена или повышена, с получением клеточной культуры; и (2) извлечение диамина из подвергнутого культивированию микроорганизма или клеточной культуры.
Понятия диамин; белок, имеющий аминокислотную последовательность SEQ ID NO: 6, или белок, имеющий аминокислотную последовательность, имеющую с ней гомологию последовательности, составляющую 42% или более; введение активности белка; повышение активности белка; диамин и микроорганизм, обладающий диамин-продуцирующей способностью, являются такими же, как описано выше.
В данном способе стадия культивирования микроорганизма может быть предпочтительно осуществлена, хотя конкретно не ограничена этим, с использованием методов периодического культивирования, непрерывного культивирования и периодического культивирования с подпиткой, известных в данной области техники. В этом смысле, условия культивирования не имеют конкретных ограничений, тем не менее оптимальное значение рН (например, рН 5-9, предпочтительно рН 6-8 и наиболее предпочтительно рН 6,8) можно поддерживать путем использования основного химического реагента (например, гидроксида натрия, гидроксида калия или аммиака) или кислотного химического реагента (например, фосфорной кислоты или серной кислоты). Кроме того, аэробные условия можно поддерживать путем добавления к клеточной культуре кислорода или газовой смеси, содержащей кислород. В процессе культивирования можно поддерживать температуру 20-45°C и предпочтительно 25-40°C, и культивирование можно проводить в течение примерно 10-160 часов.
Кроме того, среда, которая будет использована для культивирования, может включать в себя сахар и углевод (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масло и жир (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирную кислоту (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирт (например, глицерин и этанол) и органическую кислоту (например, уксусную кислоту), по отдельности или в комбинации, в качестве источника углерода; азот-содержащее органическое соединение (например, пептон, дрожжевой экстракт, мясной сок, экстракт солода, кукурузный раствор, порошок соевой муки и мочевину) или неорганическое соединение (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), по отдельности или в комбинации, в качестве источника азота; дигидрофосфат калия, гидрофосфат калия или соответствующую им натрий-содержащую соль, по отдельности или в комбинации, в качестве источника фосфора; другие необходимые стимулирующие рост вещества, включая соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины. В настоящем изобретении термин «среда» можно использовать как синоним термина «культуральная жидкость».
Использованный в данном описании термин «клеточная культура» относится к материалу, полученному посредством культивирования микроорганизма и включающему среду, культивируемый микроорганизм и вещества, высвободившиеся из культивируемого микроорганизма. Например, может быть включен источник питательных веществ, необходимых для культивирования клеток, таких как минеральные вещества, аминокислоты, витамины, нуклеиновые кислоты и/или другие компоненты, обычно содержащиеся в культуральной среде (или культуральной жидкости) помимо источника углерода и источника азота. Кроме того, могут быть включены желаемое вещество или фермент, продуцируемый/секретируемый клетками.
Поскольку диамин, продуцируемый культурой, может быть секретирован в среду или может оставаться в клетках, клеточная культура может включать в себя диамин, который продуцируется культивируемым микроорганизмом.
Способ извлечения путресцина, образованного на стадии культивирования по настоящему изобретению, может быть осуществлен, например, с использованием подходящего, известного в данной области техники способа, соответствующего способу культивирования, например, периодического культивирования, непрерывного культивирования или периодического культивирования с подпиткой, со сбором, при этом, желаемых аминокислот из культуральной жидкости.
Полезные эффекты
В настоящем изобретении продемонстрировано, что белок ARCH_0271, происходящий из Arcanobacterium haemolyticum, представляет собой белок, обладающий диамин-экспортирующей активностью, и путресцин-экспортирующая активность может быть повышена посредством введения белка с такой активностью в микроорганизм Corynebacterium sp., у которого имеется путь синтеза путресцина, но низкая путресцин-экспортирующая активность. Также продемонстрировано, что эффективность продуцирования путресцина и кадаверина может быть одновременно повышена посредством введения белка с такой активностью в бактерию E.coli, у которой имеются пути синтеза путресцина и кадаверина. Соответственно, диамин может эффективно продуцироваться в результате использования белка ARCH_0271, происходящего из Arcanobacterium haemolyticum, в микроорганизме, обладающем диамин-продуцирующей способностью.
Способ осуществления изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако, эти примеры приведены только в целях иллюстрации, и не подразумевается, что данное изобретение ограничено этими примерами.
Пример сравнения 1. Получение микроорганизма Corynebacterium sp., обладающего путресцин-продуцирующей способностью
Было подтверждено, что продуцирование путресцина снижалось, когда NCgl2522, пермеаза, принадлежащая суперсемейству главных переносчиков (MFS), была удалена в путресцин-продуцирующем штамме KCCM11240P, полученном на основе Corynebacterium glutamicum АТСС13032 (публикация заявки на патент Кореи №2013-0082478), и в путресцин-продуцирующем штамме DAB12-a ΔNCgl1469, полученном на основе Corynebacterium glutamicum АТСС13869 (делеция argF, делеция NCgl1221, введение speC Е. coli, замена промотора arg-оперона, делеция NCgl1469; обозначение штамма DAB12-b; публикация заявки на патент Кореи №2013-0082478) как микроорганизмов Corynebacterium sp., обладающих путресцин-продуцирующей способностью.
Также было подтверждено, что путресцин продуцировался с высоким выходом в штаммах Corynebacterium glutamicum, полученных путем дополнительного введения гена NCgl2522 в транспозон в KCCM11240P или DAB12-D либо полученных путем замены промотора NCgl2522 в хромосоме на промотор cj7 с целью повышения активности NCgl2522. Помимо этого, измеряли количество путресцина внутри клеток в штамме с усиленной экспрессией NCgl2522, и в результате определяли меньшее количество путресцина по сравнению с контрольной группой. Это указывает на то, что NCgl2522 обладает способностью экспортировать путресцин.
Более подробно, с учетом нуклеотидной последовательности гена, кодирующего NCgl2522 в Corynebacterium glutamicum АТСС13032, использовали пару праймеров с SEQ ID NO: 1 и 2 для получения гомологичного рекомбинантного фрагмента, соответствующего N-концевому участку NCgl2522, и пару праймеров с SEQ ID NO: 3 и 4 для получения гомологичного рекомбинантного фрагмента, соответствующего С-концевому участку NCgl2522, которые приведены ниже в Таблице 1.

Проводили полимеразную цепную реакцию (ПЦР), используя геномную ДНК Corynebacterium glutamicum ATTC13032 в качестве матрицы и две пары праймеров, чтобы амплифицировать ПЦР-фрагменты, соответствующие N-концевому и C-концевому участкам, соответственно. Эти ПЦР-фрагменты подвергали электрофорезу, получая желаемые фрагменты. На данном этапе ПЦР проводили, применяя 30 циклов с денатурацией в течение 30 секунд при 95°C, отжигом в течение 30 секунд при 55°C и удлинением в течение 30 секунд при 72°C. Полученный таким образом фрагмент, соответствующий N-концевому участку, обрабатывали ферментами рестрикции BamHI и SalI, а полученный таким образом фрагмент C-концевого участка обрабатывали ферментами рестрикции SalI и XbaI. Обработанные таким образом фрагменты клонировали в вектор pDZ, обработанный ферментами рестрикции BamHI и XbaI, чтобы сконструировать плазмиду pDZ-1'NCgl2522(K/O) (нокаут).
Плазмиду pDZ-1'NCgl2522(K/O) вводили в Corynebacterium glutamicum KCCM11240P посредством электропорации, чтобы получить трансформанты. Затем трансформанты высевали и культивировали на чашках с BHIS (дополненной сердечно-мозговым экстрактом) средой (сердечно-мозговой экстракт 37 г/л, сорбит 91 г/л и агар 2%), содержащей канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид) для образования колоний. Среди образованных таким образом колоний отбирали окрашенные голубым цветом колонии, как колонии штамма, в который введена плазмида pDZ-1'NCgl2522(K/O).
Отобранные штаммы культивировали со встряхиванием в СМ среде (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л и мочевина 2 г/л, рН 6,8) при 30°C в течение 8 часов. После этого готовили серийные разведения в диапазоне от 10-4 до 10-10 для каждой клеточной культуры. Затем эти разведенные образцы высевали на чашки и культивировали на X-gal-содержащей твердой среде для образования колоний. Среди образованных таким образом колоний отбирали белые колонии, появляющиеся с относительно низкой частотой, окончательно получая штамм Corynebacterium glutamicum, в котором был делетирован ген, кодирующий NCgl2522, и снижена путресцин-продуцирующая способность. Штамм Corynebacterium glutamicum с пониженной путресцин-экспортирующей активностью обозначали как KCCM11240P ΔNCgl2522.
Аналогичным образом проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы и двух пар праймеров, приведенных в Таблице 1, чтобы сконструировать плазмиду pDZ-2'NCgl2522(K/O) описанным выше способом. Конструировали штамм Corynebacterium glutamicum, в котором был делетирован ген, кодирующий NCgl2522 из штамма DAB12-D, используя вектор в соответствии с описанным выше способом снижения путресцин-продуцирующей способности. Этот штамм Corynebacterium glutamicum с пониженной путресцин-экспортирующей активностью обозначали как DAB12-D ΔNCgl2522.
Пример 1. Отбор ARCH_0271 из Arcanobacterium haemolyticum
Как подтверждено в примере сравнения 1, оказалось, что мембранный белок NCgl2522 функционирует в качестве экспортера путресцина. Ввиду этого, на основании аминокислотной последовательности продукта гена NCgl2522, авторы настоящего изобретения с использованием программы BlastP Национального центра биотехнологической информации (NCBI, www.ncbi.nlm.nih.gov) рассмотрели гены, имеющие с ним гомологию, отличные от генов Corynebacterium sp., и в результате этого они обнаружили нуклеотидную последовательность (SEQ ID NO: 5) и аминокислотную последовательность (SEQ ID NO: 6) ARCH_0271 из Arcanobacterium haemolyticum DSM 20595, которая имеет с ним гомологию 56%. Среди мембранных белков, имеющих гомологию аминокислотной последовательности с NCgl2522, которые обнаружены в других видах, нежели Corynebacterium sp., аминокислотная последовательность ARCH_0271 является самой филогенетически близкой к аминокислотной последовательности NCgl2522.
Аналогичным образом были установлены нуклеотидная последовательность (SEQ ID NO: 22) и аминокислотная последовательность (SEQ ID NO: 23) QWA_00075 из Alcaligenes faecalis, подвида faecalis, NCIB 8687, которая демонстрирует 41% гомологии с аминокислотной последовательностью NCgl2522, и нуклеотидная последовательность (SEQ ID NO: 25) и аминокислотная последовательность (SEQ ID NO: 26) SMD_2351 из Stenotrophomonas maltophilia D457, которая демонстрирует 52% гомологии с аминокислотной последовательностью NCgl2522. Аминокислотная последовательность QWA_00075 и аминокислотная последовательность SMD_2351 демонстрируют 42% и 47% гомологии с аминокислотной последовательностью ARCH_0271, соответственно, как показано в приведенной ниже Таблице 2.

При этом ниже в Таблице 3 приведены микроорганизмы, имеющие гены, демонстрирующие гомологию с NCgl2522, и степень их гомологии.
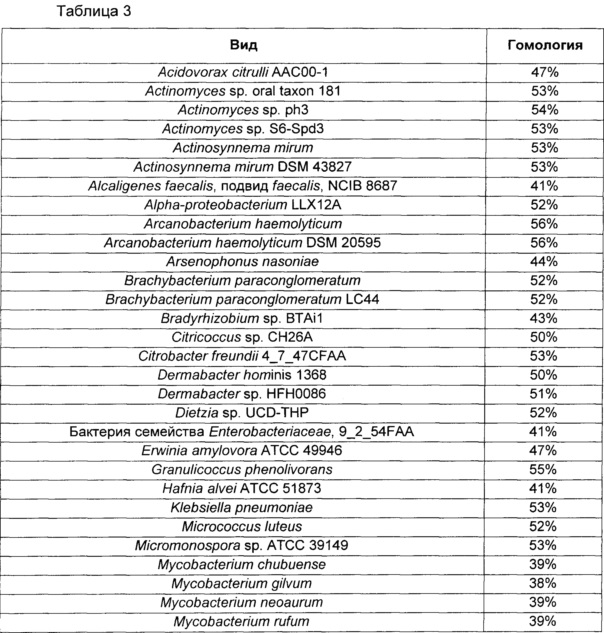
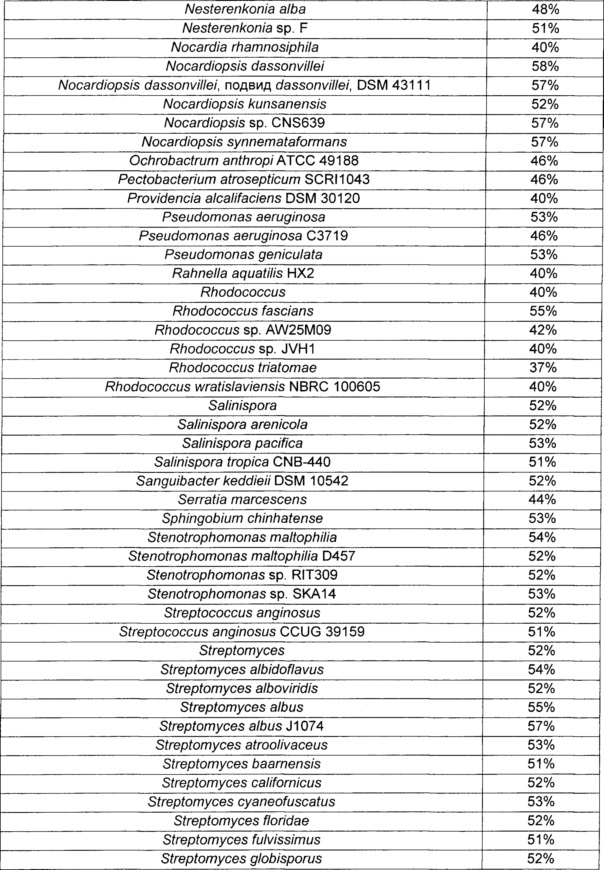
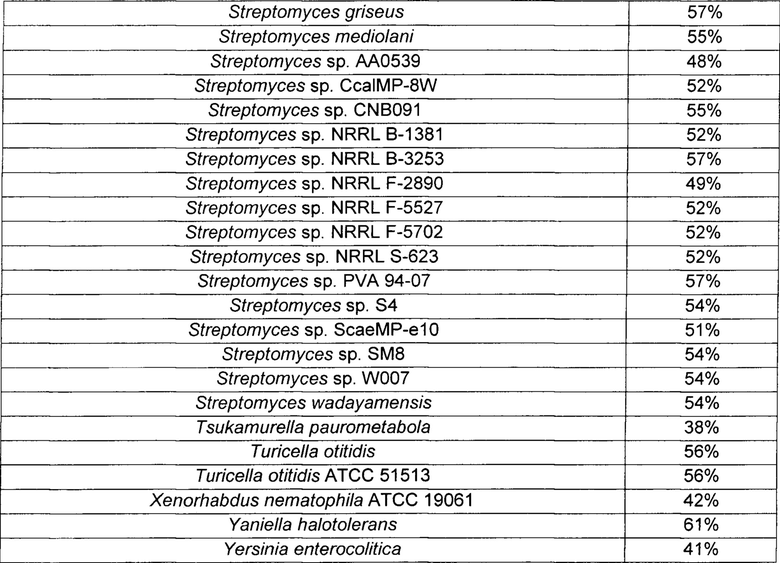
Пример 2. Получение путресцина посредством ферментации путресцин-продуцирующего штамма, происходящего из Corynebacterium sp., после введения в него белка, обладающего путресцин-экспортирующей активностью
2-1. Введение ARCH_0271, QWA_00075 или SMD_2351 в транспозонный ген в хромосому путресцин-продуцирующего штамма, полученного на основе штамма АТСС13032
Для изучения того, будет ли встраивание генов ARCH_0271, QWA_00075 или SMD_2351 в хромосому влиять на экспорт путресцина в штамме КССМ11240Р ΔNCgl2522, обладающем сниженной путресцин-экспортирующей активностью, который получали как в примере сравнения 1, в транспозонный ген вводили ARCH_0271, QWA_00075 или SMD_2351 приведенным далее способом.
В качестве вектора для трансформации, дающего возможность осуществить встраивание гена в хромосому с использованием транспозонного гена микроорганизма Corynebacterium sp., применяли pDZTn (WO 2009/125992), а в качестве промотора использовали cj7 (WO 2006/65095).
На основании нуклеотидной последовательности Arcanobacterium haemolyticum DSM 20595, синтезировали ген ARCH_0271 посредством оптимизации нуклеотидной последовательности (SEQ ID NO: 7) в отношении частоты использования кодонов в Corynebacterium glutamicum. Аналогичным образом также синтезировали QWA_00075, происходящий из Alcaligenes faecalis, подвида faecalis, NCIB 8687, и SMD_2351, происходящий из Stenotrophomonas maltophilia D457, посредством оптимизации нуклеотидных последовательностей (SEQ ID NO: 24 и SEQ ID NO: 27) в отношении частоты использования кодонов в Corynebacterium glutamicum, соответственно. Синтезированные гены получали в виде генов, клонированных в pGEM В1.
Полученные таким образом плазмиды обозначали как pGEM В1-ARCH_0271, pGEM B1-QWA_00075 и pGEM B1-SMD_2351, соответственно.
Фрагмент гена размером примерно 1,51 тысяч пар нуклеотидов (т.п.н.), 1,53 т.п.н. или 1,53 т.п.н. амплифицировали, используя плазмиду pGEM В1-ARCH_0271, pGEM B1-QWA_00075 или pGEM B1-SMD_2351 в качестве матрицы и пару праймеров с SEQ ID NO: 10 и 11, SEQ ID NO: 28 и 29 или SEQ ID NO: 30 и 31 (см. Таблицу 4), соответственно. На данном этапе ПЦР проводили, применяя 30 циклов с денатурацией в течение 30 секунд при 95°C, отжигом в течение 30 секунд при 55°C и удлинением в течение 1 минуты при 72°C.Далее эти продукты ПЦР подвергали электрофорезу в 0,8%-ном агарозном геле для элюирования и очистки каждой зоны с фрагментом желаемого размера.
Кроме того, получали участок cj7 промотора, проводя ПЦР с применением 30 циклов с денатурацией в течение 30 секунд при 95°C, отжигом в течение 30 секунд при 55°C и удлинением в течение 30 секунд при 72°C, используя р117-Pcj7-gfp в качестве матрицы и пару праймеров с SEQ ID NO: 8 и 9 (см. Таблицу 4). Фрагмент cj7 промотора гена подвергали электрофорезу в 0,8%-ном агарозном геле для элюирования и очистки зоны с фрагментом желаемого размера.

Вектор pDZTn обрабатывали, используя XhoI, и осуществляли клонирование с образованием слитой конструкции для каждого полученного выше продукта ПЦР. При проведении клонирования с образованием слитой конструкции использовали набор для клонирования ln-Fusion HD (Clontech). Полученные плазмиды обозначали как pDZTn-P(cj7)-ARCH_0271, pDZTn-P(cj7)- QWA_00075 и pDZTn-P(cj7)-SMD_2351, соответственно.
HD (Clontech). Полученные плазмиды обозначали как pDZTn-P(cj7)-ARCH_0271, pDZTn-P(cj7)- QWA_00075 и pDZTn-P(cj7)-SMD_2351, соответственно.
Каждую из трех плазмид pDZTn-P(cj7)-ARCH_0271, pDZTn-P(cj7)-QWA_00075 и pDZTn-P(cj7)-SMD_2351 вводили посредством электропорации в штамм Corynebacterium glutamicum KCCM11240P ΔNCgl2522 со сниженной путресцин-экспортирующей активностью, описанный в примере сравнения 1, получая трансформанты. Трансформанты культивировали со встряхиванием в СМ среде (глюкоза 10 г/л, полипептон 10 г/л, дрожжевой экстракт 5 г/л, мясной экстракт 5 г/л, NaCl 2,5 г/л и мочевина 2 г/л, рН 6,8) (30°C в течение 8 часов). После этого готовили серийные разведения в диапазоне от 10-4 до 10-10 для каждой клеточной культуры. Затем эти разведенные образцы высевали на чашки и культивировали на X-gal-содержащей твердой среде для образования колоний.
Среди образованных колоний отбирали белые колонии, появляющиеся с относительно низкой частотой, окончательно получая штаммы, в которых ген, кодирующий ARCH_0271, QWA_00075 или SMD_2351, был введен в результате вторичного кроссинговера. Окончательно отобранные штаммы подвергали ПЦР, используя пару праймеров с SEQ ID NO: 8 и 11, SEQ ID NO: 8 и 29 или SEQ ID NO: 8 и 31, для подтверждения введения гена, кодирующего ARCH_0271, QWA_00075 или SMD_2351. Эти мутантные штаммы Corynebacterium glutamicum обозначали как KCCM11240P ΔNCgl2522 Tn:P(cj7)-ARCH_0271, KCCM11240P ΔNCgl2522 Tn:P(cj7)-QWA_00075 и KCCM11240P ΔNCgl2522 Tn:P(cj7)-SMD_2351, соответственно.
2-2. Введение ARCH_0271, QWA_00075 или SMD_2351 в транспозонный ген в хромосому путресцин-продуцирующего штамма, полученного на основе штамма АТСС13869
Для изучения того, будет ли встраивание генов ARCH_0271 в хромосому влиять на экспорт путресцина в штамме DAB12-b ΔNCgl2522, обладающем сниженной путресцин-экспортирующей активностью, который получали как в примере сравнения 1, полученные выше pDZTn-P(cj7)-ARCH_0271, pDZTn-P(cj7)-QWA_00075 или pDZTn-P(cj7)-SMD_2351 вводили в Corynebacterium glutamicum DAB12-D ΔNCgl2522 и подтверждали введение ARCH_0271, QWA_00075 или SMD_2351 в транспозонный ген этих штаммов аналогично тому, как это выполняли в примере 2-1.
Отобранные таким образом мутантные штаммы Corynebacterium glutamicum обозначали как DAB12-D ΔNCgl2522 Tn:P(cj7)-ARCH_0271, DAB12-b ΔNCgl2522 Tn:P(cj7)-QWA_00075 и DAB12-b ΔNCgl2522 Tn:P(cj7)-SMD_2351, соответственно.
2-3. Оценка путресцин-продуцирующей способности происходящего из Corynebacterium sp.путресцин-продуцирующего штамма с введенным в него ARCH_0271, QWA_00075 или SMD_2351
Чтобы подтвердить эффект введения ARCH_0271 на путресцин- продуцирующую способность путресцин-продуцирующего штамма, сравнивали путресцин-продуцирующие способности мутантных штаммов Corynebacterium glutamicum, полученных как в примерах 2-1 и 2-2.
Более подробно, 10 типов мутантов Corynebacterium glutamicum (KCCM11240P; KCCM11240P ΔNCgl2522; KCCM11240P ΔNCgl2522 Tn:P(cj7)-ARCH_0271; KCCM11240P ΔNCgl2522 Tn:P(cj7)-QWA_00075; KCCM11240P ΔNCgl2522 Tn:P(cj7)-SMD_2351; DAB12-b; DAB12-b ΔNCgl2522; DAB12-b ΔNCgl2522 Tn:P(cj7)-ARCH_0271; DAB12-b ΔNCgl2522 Tn:P(cj7)-QWA_00075 и DAB12-b ΔNCgl2522 Tn:P(cj7)-SMD_2351) высевали на содержащую 1 мМ аргинин СМ среду для чашек (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-ного раствора NaOH и 2% агара, рН 6,8, из расчета на 1 л) и культивировали при 30°C в течение 24 часов, соответственно. Каждым штаммом, подвергнутым такому культивированию, в количестве 1 платиновой петли инокулировали 25 мл среды для титрования (8% глюкозы, 0,25% соевого белка, 0,50% твердых веществ кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, 100 мкг биотина, 3 мг тиамина гидрохлорида, 3 мг кальциевой соли пантотеновой кислоты, 3 мг никотинамида и 5% СаСО3, рН 7,0; из расчета на 1 л) и затем культивировали со встряхиванием при 30°C и 200 об./мин в течение 98 часов. Во все среды для культивирования штаммов добавляли 1 мМ аргинин. В каждой клеточной культуре измеряли концентрацию путресцина, и результаты показаны в приведенной ниже Таблице 5.
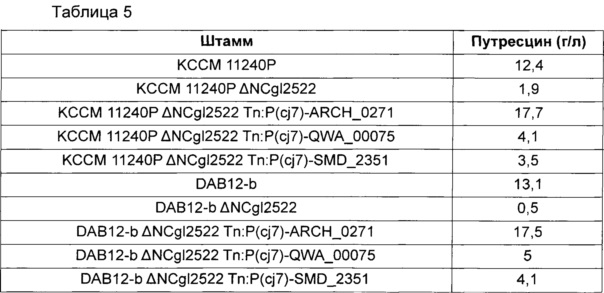
Оказалось, как показано в Таблице 5, что продуцирование путресцина усиливалось в обоих из этих 2-х типов мутантных штаммов Corynebacterium glutamicum с введенным ARCH_0271. Кроме того, было обнаружено, что по сравнению с родительским штаммом KCCM 11240Р ΔNCgl2522 или DAB12-b ΔNCgl2522 продуцирование путресцина усиливалось в штамме, в который был введен QWA_00075 или SMD_2351. Это указывает на то, что QWA_00075 или SMD_2351 обладает путресцин-экспортирующей активностью.
Пример 3. Получение кадаверина посредством ферментации лизин-продуцирующего штамма, происходящего из Corynebacterium sp., после введения в него ARCH_0271 и экспрессии лизиндекарбоксилазы
3-1. Введение ARCH_0271 в транспозонный ген в хромосому L-лизин-продуцирующего штамма Corynebacterium glutamicum KCCM11016P
Чтобы подтвердить наличие кадаверин-экспортирующей активности у белка ARCH_0271, ген ARCH_0271 вводили в хромосому лизин-продуцирующего штамма KCCM11016P (этот микроорганизм был депонирован в Корейский центр культур микроорганизмов 18 декабря 1995 г. с номером доступа KFCC10881 и затем депонирован в Международный орган по депонированию в соответствии с Будапештским договором с номером доступа KCCM11016P; патент Кореи №10-0159812). Полученную выше плазмиду pDZTn-P(cj7)-ARCH_0271 вводили в Corynebacterium glutamicum KCCM11016P, и подтверждали введение ARCH_0271 в транспозон штамма аналогично тому, как это выполняли в примере 2-1.
Отобранный таким образом мутантный штамм Corynebacterium glutamicum обозначали как KCCM11016P Tn:P(cj7)-ARCH_0271.
3-2. Введение происходящего из E.coli гена лизиндекарбоксилазы в L-лизин-продуцирующий штамм с введенным в него ARCH_0271
В L-лизин-продуцирующий штамм с введенным в него ARCH_0271, KCCM11016P Tn:P(cj7)-ARCH_0271, полученный как в примере 3-1, вводили происходящий из E.coli ген лизиндекарбоксилазы в форме плазмиды с целью продуцирования кадаверина. Нуклеотидную последовательность (SEQ ID NO: 32) и аминокислотную последовательность (SEQ ID NO: 33) лизиндекарбоксилазы IdcC из Е. coli получали из базы данных NCBI.
Фрагмент гена IdcC размером примерно 2,1 т.п.н. получали посредством проведения ПЦР, применяя 30 циклов с денатурацией в течение 30 секунд при 95°C, отжигом в течение 30 секунд при 55°C и удлинением в течение 2 минут при 72°C, используя хромосому штамма W3110 E.coli в качестве матрицы и пару праймеров с SEQ ID NO: 36 и 37 (см. Таблицу 6). Этот продукт обрабатывали HindIII и XbaI и затем подвергали электрофорезу в 0,8%-ном агарозном геле для элюирования и очистки зоны с фрагментом желаемого размера.
Кроме того, получали участок cj7 промотора, проводя ПЦР с применением 30 циклов с денатурацией в течение 30 секунд при 95°C, отжигом в течение 30 секунд при 55°C и удлинением в течение 30 секунд при 72°C, используя р117-Pcj7-gfp в качестве матрицы и пару праймеров с SEQ ID NO: 34 и 35 (см. Таблицу 6). Фрагмент cj7 промотора гена обрабатывали KpnI и HindII и затем подвергали электрофорезу в 0,8%-ном агарозном геле для элюирования и очистки зоны с фрагментом желаемого размера.

Фрагмент гена, который получали, проводя электрофорез обработанного KpnI и XbaI вектора pECCG117 (Biotechnology letters, vol. 13, №10, p.721-726 (1991)) в 0,8%-ом агарозном геле и затем элюируя и очищая зону с фрагментом желаемого размера, фрагмент cj7 промотора гена, обработанный KpnI и HindIII, и фрагмент гена лизиндекарбоксилазы IdcC, обработанный HindIII и XbaI, клонировали, используя Т4 ДНК-лигазу (NEB). Плазмиду, кодирующую IdcC из Е. coli, полученную согласно приведенному выше эксперименту, обозначали как pECCG117-Pcj7-IdcC.
Полученный вектор pECCGI17-Pcj7-IdcC или вектор pECCG117 вводили в KCCM11016P и KCCM11016P Tn:P(cj7)-ARCH_0271 посредством электропорации, соответственно. Трансформанты высевали на чашки с BHIS средой, содержащей канамицин (25 мкг/мл) для проведения отбора. Отобранные штаммы обозначали как KCCM11016P pECCG117, KCCM11016P pECCG117-Pcj7-IdcC, KCCM11016P Tn:P(cj7)-ARCH_0271 pECCG117 и KCCM11016P Tn:P(cj7)-ARCH_0271 pECCGI 17-Pcj7-IdcC, соответственно.
3-3. Оценка кадаверин-продуцирующей способности происходящего из Corynebacterium sp.лизин-продуцирующего штамма со встроенным в хромосому ARCH_0271 и с геном лизиндекарбоксилазы в составе плазмиды
Чтобы проверить, влияет ли введение ARCH_0271 в кадаверин- продуцирующий штамм на продуцирование кадаверина, проводили сравнение мутантных штаммов Corynebacterium glutamicum, полученных как в примере 3-2, в отношении кадаверин-продуцирующей способности.
Более подробно, 4 типа мутантных штаммов Corynebacterium glutamicum (KCCM11016P pECCG117; KCCM11016P pECCG117-Pcj7-IdcC; KCCM11016P Tn:P(cj7)-ARCH_0271 pECCG117 и KCCM11016P Tn:P(cj7)-ARCH_0271 pECCGI 17-Pcj7-IdcC) культивировали, используя приведенный далее метод, и проводили их сравнение в отношении кадаверин-продуцирующей способности.
Соответствующие мутантные штаммы высевали на чашки с СМ средой (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-ного раствора NaOH и 2% агара, рН 6,8; из расчета на 1 л) и культивировали при 30°C в течение 24 часов. Каждый из подвергнутых культивированию штаммов использовали для инокулирования конической колбы с перегородками емкостью 250 мл, содержащей 25 мл среды для затравки (2% глюкозы, 1% пептона, 0,5% дрожжевого экстракта, 0,15% мочевины, 0,4% KH2PO4, 0,8% K2HPO4, 0,05% MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг кальциевой соли пантотеновой кислоты и 2000 мкг никотинамида, рН 7,0; из расчета на 1 л) и культивировали со встряхиванием при 30°C и 200 об./мин в течение 20 часов.
Затем 1 мл этой затравочной культуры использовали для инокулирования конической колбы с перегородками емкостью 250 мл, содержащей 24 мл среды для продуцирования (4% глюкозы, 2% (NH4)2SO4, 2,5% соевого белка, 5% твердых веществ кукурузного экстракта, 0,3% мочевины, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина гидрохлорида, 2000 мкг кальциевой соли пантотеновой кислоты, 3000 мкг никотинамида, 0,2 г лейцина, 0,1 г треонина, 0,1 г метионина и 5% СаСО3, рН 7,0; из расчета на 1 л) и затем культивировали со встряхиванием при 30°C и 200 об./мин в течение 72 часов.
После проведения культивирования оценивали кадаверин-продуцирующую способность, используя высокоэффективную жидкостную хроматографию (HPLC). Концентрации кадаверина в клеточной культуре
каждого штамма приведены ниже в Таблице 7.
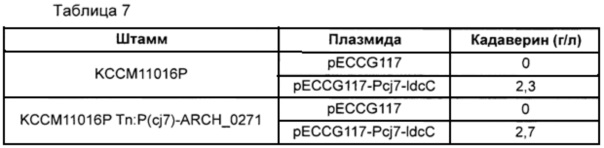
Как показано в Таблице 7, продуцирование кадаверина усиливалось более чем на 17% в мутантных штаммах Corynebacterium glutamicum с введенным ARCH_0271.
Пример 4. Получение диамина посредством ферментации Е. coli с введенным белком, обладающим диамин-экспортирующей активностью
4-1. Получение штамма посредством введения ARCH_0271, QWA_00075 или SMD_2351 в W3110
Чтобы проверить, будет ли экспрессия ARCH_0271, происходящего из Arcanobacterium haemolyticum DSM 20595, QWA_00075, происходящего из Alcaligenes faecalis, или SMD_2351, происходящего из Stenotrophomonas maltophilia, усиливать продуцирование путресцина и кадаверина в штамме дикого типа W3110 Е. coli, в котором имеется путь биосинтеза путресцина и кадаверина, в W3110 вводили, в составе челночного вектора для коринебактерий и Е. coli, pDZTn-P(cj7)-ARCH_0271, pDZTn-P(cj7)-QWA_00075 или pDZTn-P(cj7)-SMD_2351, соответственно.
2Х TSS раствор (раствор для трансформации и хранения) (Epicentre) использовали для трансформации Е. coli, и трансформанты высевали и культивировали на чашках со средой LB (Лурия-Бертани) (10 г триптона, 5 г дрожжевого экстракта, 10 г NaCl и 2% агара; из расчета на 1 л), содержащей канамицин (50 мкг/мл), для образования колоний. Образовавшиеся в результате этого колонии обозначали как колонии штаммов W3110 pDZTn-P(cj7)-ARCH_0271, W3110 pDZTn-P(cj7)-QWA_00075 и W3110 pDZTn-P(cj7)- SMD_2351, соответственно.
4-2. Сравнение диамин-продуцирующей способности E.coli с введенными ARCH_0271, QWA_00075 или SMD_2351
Исследовали путресцин- и кадаверин-продуцирующие способности полученных выше штаммов.
Более подробно, W3110 и W3110 pDZTn-P(cj7)-ARCH_0271, W3110 pDZTn-P(cj7)-QWA_00075 или W3110 pDZTn-P(cj7)-SMD_2351 культивировали на твердой LB среде при 37°C в течение 24 часов.
Затем каждый из них культивировали в 25 мл среды для титрования (2 г (NH4)2PO4, 6,75 г KH2PO4, 0,85 г лимонной кислоты, 0,7 г MgSO4⋅7H2O, 0,5% (об./об.) микроэлементов, 10 г глюкозы, 3 г AMS (сульфат аммония) и 30 г СаСО3; из расчета на 1 л) при 37°C в течение 24 часов. Раствор микроэлементов металлов в 5 М HCl содержал: 10 г FeSO4⋅7H2O; 2,25 г ZnSO4⋅7H2O; 1 г CuSO4⋅5H2O; 0,5 г MnSO4⋅5H2O; 0,23 г Na2B4O7⋅10H2O; 2 г CaCl2⋅2H2O и 0,1 г (NH4)6Mo7O2⋅4H2O на 1 литр.
Измеряли концентрации путресцина и кадаверина, продуцируемых каждой клеточной культурой, и результаты приведены ниже в Таблице 8.
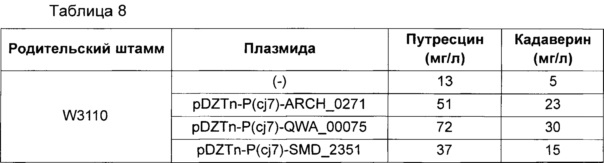
Как показано в Таблице 8, по сравнению с родительским штаммом W3110, продуцирование путресцина и кадаверина значительно усиливалось в штаммах W3110 pDZTn-P(cj7)-ARCH_0271, W3110 pDZTn-P(cj7)-QWA_00075 или W3110 pDZTn-P(cj7)-SMD_2351, в которые вводили ARCH_0271, QWA_00075 или SMD_2351, соответственно.
То есть, установлено, что количество диамина, продуцируемого в клеточной культуре, значительно возрастало в результате повышения активности белков ARCH_0271, QWA_00075 или SMD_2351, имеющих 56%, 41% или 52% гомологии последовательности с аминокислотной последовательностью NCgl2522, и это подтверждает, что способность экспортировать такой диамин, как путресцин и кадаверин, может быть улучшена путем повышения активности СЕ2495 или белка, имеющего с ним гомологию последовательности, составляющую 42% или больше.
По существу, авторы настоящего изобретения продемонстрировали, что микроорганизм Corynebacterium glutamicum, обладающий повышенной активностью ARCH_0271, приобретенной посредством введения ARCH_0271 в транспозон микроорганизма Corynebacterium sp. KCCM11240P ΔNCgl2522, в котором имеется путь синтеза путресцина, но снижена путресцин-экспортирующая активность, обладает повышенной путресцин-экспортирующей активностью с получением тем самым путресцина с высоким выходом.
Таким образом, этот штамм KCCM11240P ΔNCgl2522 Tn:P(cj7)- ARCH_0271 обозначен как СС01-0758 и депонирован в соответствии с Будапештским договором в Корейский центр культур микроорганизмов (KCCM) 15 ноября 2013 г. с номером доступа KCCM11476P.




















Группа изобретений относится к микроорганизму для продуцирования диамина и способу получения диамина с его использованием. Предложен микроорганизм, в котором активность белка, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 23 и SEQ ID NO: 26 или белка, обладающего диамин-экспортирующей активностью и имеющего аминокислотную последовательность, имеющую по меньшей мере 95% гомологии последовательности с SEQ ID NO: 6, SEQ ID NO: 23 или SEQ ID NO: 26 введена или повышена по сравнению с эндогенной активностью. Предложен также способ получения диамина, включающий культивирование указанного микроорганизма с получением клеточной культуры и извлечение диамина из подвергнутого культивированию микроорганизма или клеточной культуры. 2 н. и 5 з.п. ф-лы, 8 табл., 4 пр.
1. Микроорганизм для продуцирования диамина, где активность белка, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 23 и SEQ ID NO: 26 или белка, обладающего диамин-экспортирующей активностью и имеющего аминокислотную последовательность, имеющую по меньшей мере 95% гомологии последовательности с SEQ ID NO: 6, SEQ ID NO: 23 или SEQ ID NO: 26, в микроорганизме, способном продуцировать диамин, введена или повышена по сравнению с эндогенной активностью.
2. Микроорганизм по п.1, где диамин-ацетилтрансферазная активность дополнительно снижена по сравнению с эндогенной активностью.
3. Микроорганизм по п.2, где диамин-ацетилтрансфераза имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12, 13 и 14.
4. Микроорганизм по п.1, где диамин представляет собой путресцин или кадаверин.
5. Микроорганизм по п.1, где микроорганизм представляет собой микроорганизм, принадлежащий роду Corynebacterium или роду Escherichia.
6. Способ получения диамина, включающий:
(1) культивирование микроорганизма по любому из пп.1-5 с получением клеточной культуры; и
(2) извлечение диамина из подвергнутого культивированию микроорганизма или клеточной культуры.
7. Способ по п.6, где диамин представляет собой путресцин или кадаверин.
| WO 2013093737 A1, 27.06.2013 | |||
| KIND S | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ ПУТРЕСЦИНА В ВЫСОКОЙ КОНЦЕНТРАЦИИ, СПОСОБ ПОЛУЧЕНИЯ ЭТОГО МИКРООРГАНИЗМА И СПОСОБ ПРОДУКЦИИ ПУТРЕСЦИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2009 |
|
RU2433180C2 |
| Биохимия: Учебник / Под ред | |||
| Е.С | |||
| Северина | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| - М.: ГЭОТАР-МЕД, 2004 | |||
| Способ замораживания пищевых веществ | 1916 |
|
SU784A1 |
| - (Серия "XXI век"), см | |||
| с | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |

