ОБЛАСТЬ ТЕХНИКИ
Настоящее раскрытие относится к микроорганизму, продуцирующему путресцин, и к способу получения путресцина с его применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Путресцин, который представляет собой сырье для нейлона, производят главным образом химическим способом с использованием соединений нефти в качестве сырья. В частности, путресцин получают путем добавления цианистого водорода к акрилонитрилу с получением сукцинонитрила, с последующей гидрогенизацией. Такой химический способ имеет эффективность и экономическую обоснованность, но имеет недостаток в виде нанесения вреда окружающей среде. Следовательно, из-за ужесточения природоохранных правил и приведенного выше, существует потребность в производстве альтернативных веществ посредством биологических путей.
В связи с этим, раскрыты способы получения высоких концентраций путресцина посредством трансформации Е. coli и микроорганизмов рода Corynebacterium (Morris et al., J Biol. Chem. 241: 13, 3129-3135, 1996; WO 06/005603; WO 09/125924; Qian ZD et al., Biotechnol. Bioeng. 104: 4, 651-662, 2009; Schneider et al., Appl. Microbiol. Biotechnol. 88: 4, 859-868, 2010 и Schneider et al., Appl. Microbiol. Biotechnol. 91: 17-30, 2011). Кроме того, известны разные способы получения путресцина с использованием микроорганизмов (US 13/992242, US 14/372000, US 14/373265, ЕР 2236613 В1 и US 8497098 В2).
Среди белков, ассоциированных с путем биосинтеза путресцина, argJ - бифункциональная орнитинацетилтрансфераза/N-ацетилглутаматсинтаза - представляет собой фермент, который может осуществлять реакцию превращения двух веществ: ацетил-КоА и N-ацетилорнитина, и имеет функции как орнитинацетилтрансферазы (L-глутамат-N-ацетилтрансфераза) и N-ацетилглутаматсинтазы, и имеет функции как орнитинацетилтрансферазы, так и N-ацетилглутаматсинтазы. Фермент argJ может уменьшать уровень побочных продуктов и уменьшать бремя продукции путресцина, так как один фермент опосредует две ферментативные реакции. Однако активность argJ может ингибироваться промежуточными метаболитами, такими как орнитин (Sakanyan V, Petrosyan Р, Lecocq М, Boyen A, Legrain С, Demarez М, Hallet J, Glansdorff N: Genes and enzymes of the acetyl cycle of arginine biosynthesis in Corynebacterium glutamicum: enzyme evolution in the early steps of the arginine pathway. Microbiology 1996, 142:99-108), приводя к пониженной биосинтетической эффективности путресцина.
N-ацетилтрансфераза, присутствующая в Corynebacterium glutamicum, имеет способность продуцировать N-ацетил-L-глутамат в присутствии ацетил-КоА и глутамата. Однако сообщалось о том, что данная N-ацетилтрансфераза имеет по меньшей мере в 9,43 раза более высокую удельную активность, чем argJ (A new type of N-acetylglutamate synthase is involved in the first step of arginine biosynthesis in Corynebacterium glutamicum. "BMC genomics 2013(14) c. 713).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Авторы настоящего изобретения предприняли интенсивные усилия по увеличению продукции путресцина в микроорганизмах и, в результате, подтвердили, что введение активности N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активности ацетил орнитинде ацетил азы, происходящей из Е. coli, в микроорганизм рода Corynebacterium приводило к увеличению продукции путресцина и, таким образом, завершили настоящее изобретение.
Техническое решение
Согласно одному аспекту настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина, включающий культивирование в среде микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
Согласно еще одному другому аспекту настоящего раскрытия предложена композиция для получения путресцина, содержащая микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который вводят активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
Полезные эффекты
Микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli, по настоящему изобретению, можно культивировать с получением путресцина с высоким выходом по сравнению с существующим не модифицированным микроорганизмом.
Краткое описание графических материалов
На ФИГ. 1 схематически показан путь биосинтеза путресцина и орнитина в микроорганизме рода Corynebacterium (А) и путь биосинтеза путресцина и орнитина в Е. coli (Б).
На ФИГ. 2 схематически показан путь биосинтеза путресцина и орнитина в микроорганизме рода Corynebacterium с улучшенной способностью продуцировать путресцин и орнитин посредством усиления Ncgl2644 Coryne и введения argE Е. coli.
Подробное описание изобретения
Настоящее изобретение будет конкретно описано следующим образом. Каждое описание и воплощение в настоящем раскрытии, также может применяться к другим описаниям и воплощениям. То есть, все комбинации различных элементов в настоящем изобретении входят в объем настоящего изобретения. Кроме того, объем настоящего изобретения не ограничен конкретным описанием, приведенным ниже.
Согласно одному аспекту настоящего изобретения предложен микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
Термин «путресцин», как он здесь используется, относится к органическому соединению - диамину, состоящему из четырех атомов углерода, которое имеет молекулярную формулу NH2(CH2)4NH2 и также называется 1,4-диаминобутан или бутандиамин.
Термин «N-ацетилтрансфераза», как он здесь используется, относится к ферменту (ЕС 2.3.1.5), который катализирует перенос ацетильной группы от ацетил-КоА на ариламин, арилгидроксиламин и арилгидразин. Данный фермент может катализировать перенос ацетила между ариламинами без КоА и может обладать способностью продуцировать N-ацетил-L-глутамат в присутствии ацетил-КоА и глутамата. Известно, что данный фермент имеет широкую специфичность в отношении ароматических аминов.
В настоящем изобретении N-ацетилтрансфераза может принадлежать к семейству GCN5-родственных N-ацетилтрансфераз (GNAT), но не ограничивается ими.
N-ацетилтрансфераза может иметь аминокислотную последовательность SEQ ID NO: 1, состоять из аминокислотной последовательности SEQ ID NO: 1 или содержать аминокислотную последовательность, как показано в SEQ ID NO: 1, но не ограничивается ими. Последовательность SEQ ID NO: 1 может быть подтверждена в известной базе данных GenBank NCBI (Национальный центр биотехнологической информации США).
В частности, N-ацетилтрансфераза может иметь аминокислотную последовательность SEQ ID NO: 1 и/или аминокислотную последовательность, имеющую по меньшей мере 70%-ную, 80%-ную, 85%-ную, 90%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную или 99%-ную или более гомологию или идентичность с SEQ ID NO: 1. Также очевидно, что даже N-ацетилтрансфераза, имеющая аминокислотную последовательность с делецией, модификацией, заменой или присоединением в ее части, может входить в объем настоящего изобретения, при условии, что данная аминокислотная последовательность имеет такую гомологию или идентичность и демонстрирует функцию, соответствующую N-ацетилтрансферазе. То есть, несмотря на описание как «белок или полипептид, содержащий аминокислотную последовательность, как показано в конкретном номере последовательности» или «белок или полипептид, состоящий из аминокислотной последовательности, как показано в конкретном номере последовательности» в настоящем изобретении, любой белок, состоящий из аминокислотной последовательности с делецией, модификацией, заменой или присоединением в его части, также можно использовать в настоящем изобретении, при условии, что данный белок имеет активность, идентичную или соответствующую активности полипептида, состоящего из аминокислотной последовательности с соответствующим номером последовательности. Например, очевидно, что любой полипептид, который имеет активность, идентичную или соответствующую «полипептиду, содержащему аминокислотную последовательность SEQ ID NO: 1» может попадать в пределы «полипептида, содержащего аминокислотную последовательность SEQ ID NO: 1».
В настоящем раскрытии ген, кодирующий N-ацетилтрансферазу, может происходить из штамма рода Corynebacterium и, в частности, может происходить из Corynebacterium glutamicum, но любой штамм рода Corynebacterium, который может экспрессировать ген, кодирующий N-ацетилтрансферазу, конкретно не ограничен. Данный ген может содержать нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 1, и, более конкретно, может содержать нуклеотидную последовательность SEQ ID NO: 2, но не ограничивается ей. Нуклеотидная последовательность SEQ ID NO: 2 может быть получена из известной базы данных GenBank и названа Ncgl2644. Термин «ген, содержащий нуклеотидную последовательность SEQ ID NO: 2» может использоваться взаимозаменяемо с термином «полинуклеотид, содержащий нуклеотидную последовательность SEQ ID NO: 2», или «ген или полинуклеотид, имеющий нуклеотидную последовательность SEQ ID NO: 2», или «ген или полинуклеотид, состоящий из нуклеотидной последовательности SEQ ID NO: 2».
Термин «полинуклеотид», как он здесь используется, относится к полимеру из нуклеотидов, вытянутому в длину посредством ковалентной связи нуклеотидных мономеров, и, в общем случае, к нити ДНК или РНК определенной длины и, более конкретно, может означать полинуклеотидный фрагмент, кодирующий вариант. Полинуклеотид может быть описан как ген, когда данный полинуклеотид представляет собой совокупность полинуклеотидов, способных выполнять функции. В настоящем изобретении термины «полинуклеотиды» и «гены» могут быть использованы взаимозаменяемо.
В частности, из-за вырожденности кодонов или посредством рассмотрения кодонов, предпочитаемых микроорганизмом, в котором возможна экспрессия данного полипептида, полинуклеотид по настоящему изобретению может быть по-разному модифицирован в его кодирующей области в пределах диапазона, в котором аминокислотная последовательность данного полипептида не изменяется. В частности, может быть включена любая полинуклеотидная последовательность, которая кодирует N-ацетилтрансферазу, состоящую из аминокислотной последовательности SEQ ID NO: 1, без ограничения этим.
Кроме того, может быть включен любой зонд, который можно получить из известной последовательности гена, например может быть включена без ограничения любая последовательность, которая может гибридизоваться с комплементарной последовательностью частью или всей нуклеотидной последовательностью в жестких условиях, чтобы кодировать N-ацетилтрансферазу, состоящую из аминокислотной последовательности SEQ ID NO: 1. Термин «жесткие условия» относится к условиям, которые обеспечивают возможность специфической гибридизации полинуклеотидов. Такие условия хорошо известны в данной области. Например, данные условия могут включать условия, в которых гены, имеющие высокую гомологию или идентичность, такие как гены, имеющие по меньшей мере 40%-ную, в частности, по меньшей мере 90%-ную, более конкретно, по меньшей мере 95%-ную, еще более конкретно, по меньшей мере 97%-ную и еще более конкретно, по меньшей мере 99%-ную гомологию или идентичность, гибридизуются друг с другом, а гены, имеющие меньшую гомологию или идентичность не гибридизуются друг с другом; или типичные условия промывки для саузерн-гибридизации, т.е. условия, когда промывку проводят один раз, более конкретно, два или три раза, с концентрацией соли и температурой, соответствующих 60°С, 1×SSC (раствор хлорида и цитрата натрия), 0,1% SDS (додецилсульфат натрия), конкретно при 60°С, 0,1×SSC и 0,1% SDS, и более конкретно при 68°С, 0,1×SSC, 0,1% SDS.
Гибридизация требует, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя могут быть несоответствия между нуклеотидами, в зависимости от жесткости гибридизации. Термин «комплементарный» используется для описания взаимосвязи между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в отношении ДНК, аденозин является комплементарным тимину, а цитозин является комплементарным гуанину. Следовательно, настоящее изобретение может включать не только по существу аналогичные последовательности нуклеиновой кислоты, но также выделенные фрагменты нуклеиновой кислоты, комплементарные всей последовательности.
В частности, полинуклеотиды, имеющие гомологию или идентичность, могут быть обнаружены с использованием вышеописанных условий гибридизации, включающих стадию гибридизации при значении Tm 55°С. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, но не ограничивается ими, и может быть подходящим образом скорректировано специалистом в данной области согласно цели.
Подходящая степень жесткости для гибридизации полинуклеотидов может зависеть от длин полинуклеотидов и степени комплементарности полинуклеотидов, и ее параметры хорошо известны в данной области (см. Sambrook et al., выше, 9.50-9.51, 11.7-11.8).
Термин «гомология» или «идентичность», как он здесь используется, относится к степени сходства двух данных аминокислотных последовательностей или нуклеотидных последовательностей, и она может быть выражена в виде процента. Термины «гомология» и «идентичность» часто могут использоваться взаимозаменяемо.
Гомологию или идентичность консервативных полинуклеотидов или полипептидов можно определить при помощи стандартных алгоритмов выравнивания, и можно совместно использовать штрафы за пробел по умолчанию, установленные используемой программой. По существу, гомологичные или идентичные последовательности обычно могут гибридизоваться друг с другом целиком или в части в умеренно или очень жестких условиях. Очевидно, что гибридизация также включает гибридизацию с полинуклеотид ом, содержащим обычные кодоны или кодоны с учетом вырожденности кодонов в полинуклеотиде.
Гомологию, сходство или идентичность двух полинуклеотидных или полипептидных последовательностей можно определять с использованием любого известного в данной области компьютерного алгоритма, такого как программа «FASTA», с использованием параметров по умолчанию, раскрытых в Pearson et al (1988) [Proc. Natl. Acad. Sci. USA 85]:2444. В качестве альтернативы, гомологию, сходство или идентичность можно определять с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), как осуществляется в программе Needleman пакета European Molecular Biology Open Software Suite (EMBOSS) (Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или более поздняя) (включая программный пакет GCG (Devereux, J., et al, Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL, J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, и [CARILLO ЕТА/. (1988) SIAM J Applied Math 48: 1073). Например, гомологию, сходство или идентичность можно определять с использованием BLAST базы данных Национального центра биотехнологической информации или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определять путем сравнения информации о последовательности с использованием компьютерной программы GAP, например Needleman et al., (1970), J Mol Biol. 48:443, как известно в Smith и Waterman, Adv. Appl. Math (1981) 2:482. Кратко, программа GAP определяет сходство как число выровненных символов (т.е. нуклеотидов или аминокислот), которые являются аналогичными, поделенное на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) бинарную матрицу сравнений (содержащую значение 1 для идентичности и 0 для неидентичности) и матрицу взвешенных сравнений Gribskov et al. (1986) Nucl. Acids Res. 14:6745, как раскрыто в Schwartz и Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, стр. 353-358 (1979) (или матрица замен EDNAFULL (версия EMBOSS NUC4.4 NCBI); (2) штраф 3,0 за каждый пробел и дополнительный штраф 0,10 за каждый символ в каждом пробеле (или штраф за открытие пробела 10 и штраф за расширение пробела 0,5); и (3) отсутствие штрафа за концевые пробелы.
В одном воплощении настоящего изобретения вариант по настоящему изобретению может иметь активность фитоендесатуразы. Кроме того, вариант по настоящему изобретению может иметь активность увеличения способности продуцировать 5'-инозинмонофосфат (IMP) по сравнению с полипептидом дикого типа, имеющим активность фитоендесатуразы.
Термин «фитоендесатураза», как он здесь используется, представляет собой полипептид, который превращает бесцветный 15-цис-фитоен в светло-красный ликопина в биохимическом пути, именуемом поли-транс-путь. В частности, термин «фитоендесатураза по настоящему изобретению» может использоваться взаимозаменяемо с фитоендесатуразой или Crt1. В настоящем изобретении последовательность фитоендесатуразы может быть получена из известной базы данных GenBank NCBI. В частности, фитоендесатураза может представлять собой полипептид, имеющий активность фитоендесатуразы, кодируемой crtl, но не ограничивается им.
Термин «ацетилорнитиндеацетилаза», как он здесь используется, относится к ферменту (ЕС 3.5.1.16), который опосредует реакцию, участвующую в продуцировании уксусной кислоты и орнитина, опосредуя гидролиз ацетилорнитина. В настоящем изобретении ацетил op нитинде ацетил аза может представлять собой argE, но не ограничивается этим.
argE может иметь аминокислотную последовательность SEQ ID NO: 3, состоять из аминокислотной последовательности SEQ ID NO: 3 или содержать аминокислотную последовательность, как показано в SEQ ID NO: 3, но не ограничен этим. Последовательность SEQ ID NO: 3 может быть подтверждена в известной базе данных GenBank NCBI.
В настоящем раскрытии ген, кодирующий argE, может происходить из бактерий и, в частности, может происходить из Е. coli, но любой штамм, который может экспрессировать ген, кодирующий argE, конкретно не ограничен. Данный ген может содержать нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 3, и более конкретно, может содержать нуклеотидную последовательность SEQ ID NO: 4, но не ограничен этим. Нуклеотидная последовательность SEQ ID NO: 4 может быть получена из известной базы данных GenBank.
В настоящем раскрытии микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, может иметь ослабленную активность бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (argJ), происходящей из штамма рода Corynebacterium, но не ограничивается этим.
Термин «бифункциональная орнитинацетилтрансфераза/N-ацетилглутаматсинтаза», как он здесь используется, относится к ферменту, который может осуществлять реакции превращения двух веществ: ацетил-КоА и N-ацетилорнитина, и данный фермент имеет функции как орнитинацетилтрансферазы (L-глутамат-N-ацетилтрансфераза (ЕС 2.3.1.35)), так и N-ацетилглутаматсинтазы (ЕС 2.3.1.1). Орнитинацетилтрансфераза превращает N2-ацетил-L-орнитин и L-глутамат в L-орнитин, и N-ацетилглутаматсинтаза катализирует образование N-ацетилглутамата из глутамата и ацетил-КоА. В настоящем изобретении бифункциональная орнитинацетилтрансфераза/N-ацетилглутаматсинтаза может представлять собой argJ, но не ограничена ею.
argJ может уменьшать уровень побочных продуктов и снизить нагрузку при продуцировании путресцина, так как один фермент опосредует две ферментативные реакции. Однако активность argJ может ингибироваться метаболическими промежуточными продуктами, такими как орнитин, что приводит к пониженной эффективности биосинтеза путресцина.
argJ может иметь аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 7, состоять из аминокислотной последовательности SEQ ID NO: 5 или SEQ ID NO: 7, или содержать аминокислотную последовательность AS, как показано в SEQ ID NO: 5 или SEQ ID NO: 7, но не ограничивается этим. Последовательность SEQ ID NO: 5 или SEQ ID NO: 7 может быть подтверждена в известной базе данных GenBank NCBI.
В настоящем раскрытии ген, кодирующий argJ, может происходить из штамма рода Corynebacterium и, в частности, может быть получен из Corynebacterium glutamicum, но любой штамм рода Corynebacterium, который может экспрессировать ген, кодирующий argJ, конкретно не ограничен. В частности, ген, кодирующий аминокислотную последовательность SEQ ID NO: 5, может происходить из Corynebacterium glutamicum АТСС13869, но не ограничивается этим. Данный ген может содержать нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 5, и, более конкретно, может содержать нуклеотидную последовательность SEQ ID NO: 6, но не ограничивается этим. Нуклеотидная последовательность SEQ ID NO: 6 может быть получена из известной базы данных GenBank.
Кроме того, ген, кодирующий аминокислотную последовательность SEQ ID NO: 7, может происходить из Corynebacterium glutamicum АТСС13032, но не ограничен этим. Данный ген может содержать нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 7, и, более конкретно, может содержать нуклеотидную последовательность SEQ ID NO: 8, но не ограничен этим. Нуклеотидная последовательность SEQ ID NO: 8 может быть получена из известной базы данных GenBank.
Термин «микроорганизм, обладающий способностью продуцировать путресцин», как он здесь используется, относится к микроорганизму, имеющему природную способность продуцировать путресцин, или к микроорганизму, полученному путем наделения родительского штамма, не имеющего способности продуцировать путресцин, способностью продуцировать путресцин. В частности, данный микроорганизм представляет собой микроорганизм, продуцирующий путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность argE, происходящей из Е. coli, но не ограничивается ими.
В частности, «микроорганизм, продуцирующий путресцин» включает все из микроорганизмов дикого типа или естественным образом или искусственно генетически модифицированных микроорганизмов. Более конкретно, микроорганизм, продуцирующий путресцин, может представлять собой микроорганизм, в котором ослаблен или усилен специфический механизм из-за вставки экзогенного гена либо из-за усиления или ослабления активности собственного гена, где микроорганизм может иметь генетическую мутацию или усиленную способность продуцировать путресцин для продуцирования целевого путресцина.
В частности, термин «введение» активности означает, что ген, которым исходно не обладал микроорганизм, экспрессируется в данном микроорганизме и, таким образом, данный микроорганизм демонстрирует активность специфического белка или демонстрирует повышенную или усиленную активность соответствующего белка по сравнению с собственной активностью или активностью до модификации. Например, данный термин может указывать на то, что полинуклеотид, кодирующий конкретный белок, вводят в хромосому микроорганизма или вектор, содержащий полинуклеотид, кодирующий конкретный белок, вводят в микроорганизм демонстрируя тем самым активность данного конкретного белка.
Термин «усиление» активности полипептида, как он здесь используется, означает, что активность полипептида увеличена по сравнению с собственной активностью. Термин «усиление» может быть использован взаимозаменяемо с такими терминами, как «активация», «повышающая регуляция», «сверхэкспрессия» и «увеличение». В частности, «активация», «усиление», «повышающая регуляция», «сверхэкспрессия» и «увеличение» могут включать как демонстрирование активности, которой исходно не обладал полипептид, или демонстрирование улучшенной активности по сравнению с собственной активностью, или активностью до модификации. Термин «собственная активность» означает активность конкретного полипептида, которой исходно обладал родительский штамм перед изменением признака или немодифицированный микроорганизм, когда признак изменяется посредством генетической вариации из-за природных или искусственных факторов. Данный термин может использоваться взаимозаменяемо с термином «активность до модификации». Тот факт, что активность полипептида «усилена», «подвергнута повышающей регуляции», «сверхэкспрессируется» или «увеличена» по сравнению с собственной активностью означает, что активность полипептида улучшена по сравнению с активностью и/или концентрацией (уровнем экспрессии) конкретного полипептида, которыми исходного обладал родительский штамм до изменения признака или немодифицированный микроорганизм.
Усиление может быть достигнуто посредством введения экзогенного полипептида или увеличения активности и/или концентрации (уровня экспрессии) эндогенного полипептида. Усиление активности полипептида может быть проверено по увеличению уровня активности или уровня экспрессии соответствующего полипептида, или по увеличению количества продукта, продуцируемого из соответствующего полипептида.
Для усиления активности полипептида можно применять различные способы, хорошо известные в данной области, и способ не ограничен при условии, что активность целевого полипептида может быть усилена по сравнению с микроорганизмом до модификации. В частности, можно использовать генную инженерию и/или белковую инженерию, хорошо известные специалистам в данной области, которые являются традиционными методами молекулярной биологии, но данный способ не ограничивается ими (например, Sitnicka et al., Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol. 2. 1-16; Sambrook et al., Molecular Cloning 2012 и тому подобные).
В частности, на усиление активности полипептида по настоящему изобретению может указывать:
1) увеличение числа внутриклеточных копий полинуклеотида, кодирующего данный полипептид;
2) замена на хромосоме области контроля экспрессии гена, кодирующего данный полипептид, последовательностью, имеющей сильную активность;
3) модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR (5'-нетранслируемая область) область транскрипта гена, кодирующего данный полипептид;
4) модификация аминокислотной последовательности полипептида для усиления активности данного полипептида;
5) модификация полинуклеотидной последовательности, кодирующей данный полипептид, для усиления активности данного полипептида (например модификация полинуклеотидной последовательности гена полипептида так, чтобы кодировать полипептид, модифицированный для усиления активности данного полипептида);
6) введение экзогенного полипептида, демонстрирующего активность данного полипептида, или экзогенного полинуклеотида, кодирующего данный полипептид;
7) оптимизация кодонов полинуклеотида, кодирующего данный полипептид;
8) модификация или химическая модификация экспонированной области, выбранной посредством анализа третичной структуры полипептида; или
9) комбинация двух или более, выбранных из пп. (1) - (8), но конкретно не ограничено этим.
Более конкретно, приведенные выше пункты описываются следующим образом.
Увеличение числа внутриклеточных копий полинуклеотида, кодирующего полипептид в п. (1) выше, может достигаться посредством введения в клетку-хозяина вектора, с которым функциональным образом связан полинуклеотид, кодирующий соответствующий полипептид, и который может реплицироваться и функционировать независимо от хозяина. В качестве альтернативы, увеличение может достигаться посредством введения одной или более копий полинуклеотида, кодирующего соответствующий полипептид, в хромосому в клетке-хозяине. Введение в хромосому может осуществляться посредством введения в клетку-хозяина вектора, способного встраивать полинуклеотид в хромосому в клетке-хозяине, но не ограничено этим. Данный вектор является таким, как описано выше.
Замена на хромосоме области контроля экспрессии гена, кодирующего данный полипептид, последовательностью с сильной активностью в п. 2), может представлять, например, возникновение модификации в данной последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации, или замену последовательностью, имеющей более сильную активность, так, чтобы дополнительно усилить активность области контроля экспрессии. Область контроля экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, последовательность, контролирующую окончание транскрипции и трансляции, и тому подобные, но конкретно не ограничена ими. Например, замена может представлять собой замену исходного промотора сильным промотором, но не ограничивается этим.
Примеры известных сильных промоторов могут включать промоторы CJ1-CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (US 10584338 B2), промотор 02 (US 10273491 B2), промотор tkt, промотор уссА и тому подобные, но не ограничены ими.
Модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR область транскрипта гена, кодирующего полипептид в п. (3), может представлять собой, например, замену нуклеотидной последовательностью, кодирующей, не эндогенный инициирующий кодон, а другой инициирующий кодон, обеспечивающий более высокую скорость экспрессии полипептида, но не ограничивается этим.
Модификация аминокислотной последовательности или полинуклеотидной последовательности в пп. (4) и (5) может представлять собой возникновение модификации в последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации в аминокислотной последовательности полипептида или в полинуклеотидной последовательности, кодирующей данный полипептид, или замены аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной так, чтобы иметь более сильную активность, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной так, чтобы иметь повышенную активность, таким образом, чтобы усиливать активность данного полипептида, но не ограничена ими. В частности, замена может осуществляться путем вставки полинуклеотида в хромосому посредством гомологичной рекомбинации, но не ограничена этим. Использованный вектор может дополнительно включать селективный маркер для проверки вставки в хромосому. Селективный маркер является таким, как описано выше.
Введение экзогенного полипептида, демонстрирующего активность полипептида, в п. (6) может представлять собой введение в клетку-хозяина экзогенного полинуклеотида, кодирующего полипептид, демонстрирующий такую же/аналогичную активность относительно данного полипептида. Экзогенный полинуклеотид не ограничен по его происхождению или последовательности, при условии, что данный экзогенный полинуклеотид демонстрирует идентичную/аналогичную активность относительно полинуклеотида. Введение может быть осуществлено посредством любого известного способа трансформации, подходящим образом выбранного специалистом в данной области, и введенный полинуклеотид экспрессируется в клетке-хозяине, и, таким образом, продуцируется полипептид, и его активность может быть усилена.
Оптимизация кодонов полинуклеотида, кодирующего полипептид в п. (7), может представлять собой оптимизацию кодонов эндогенного полинуклеотида так, чтобы увеличить его транскрипцию или трансляцию в клетке-хозяине, или оптимизацию кодонов экзогенного полинуклеотида так, чтобы осуществлять оптимизированную транскрипцию или трансляцию в клетке-хозяине.
Модификация или химическая модификация экспонированной области, выбранной посредством анализа третичной структуры полипептида, в п. (8) может представлять собой, например, модификацию или химическую модификацию экспонированной области, подлежащей модификации или химической модификации, путем сравнения информации о последовательности полипептида, подлежащего анализу, с базой данных, которая хранит информацию о последовательностях известных белков, для определения шаблонного белка-кандидата согласно сходству последовательностей и идентификации структуры на основе определенного кандидата.
Такое усиление активности полипептида может означать, что активность, или концентрация, или уровень экспрессии соответствующего полипептида увеличены относительно активности или концентрации полипептида, экспрессируемого в микробном штамме дикого типа или микробном штамме перед модификацией; или что увеличивается количество продукта, полученного из соответствующего полипептида, не ограничивается этим.
Модификация части или всего полинуклеотида в микроорганизме по настоящему изобретению может индуцироваться посредством: (а) гомологичной рекомбинации с использованием вектора для вставки в хромосому в микроорганизме или редактирования генома с применением генетически модифицированной нуклеазы (например CRISPR-Cas9) и/или (б) обработки светом, таким как ультрафиолетовый свет и излучение, и/или химическими веществами, но не ограничена этим. Способ модификации части или всего гена может включать способ с использованием технологии рекомбинантной ДНК. Например, посредством введения нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную гену-мишени, в микроорганизм, чтобы вызвать гомологичную рекомбинацию, может быть удалена часть гена или весь ген. Введенная нуклеотидная последовательность или вектор может содержать маркер доминантной селекции, но не ограничивается этим.
Такое усиление активности белка может означать, что активность или концентрация соответствующего белка увеличивается по сравнению с активностью или концентрацией белка, экспрессируемого в микробном штамме дикого типа или в микробном штамме перед модификацией, но не ограничивается этим. Термин «штамм перед модификацией» или «микроорганизм перед модификацией», как он здесь используется, не исключает штамм, содержащий мутацию, которая может встречаться в природе в микроорганизме, и относится к самому нативному штамму или штамму перед изменением признака из-за генетической мутации, вызванной искусственными факторами. В настоящем изобретении изменение признака может представлять собой введение активности N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активности argE, происходящей из Е. coli. Термины «штамм перед модификацией» или «микроорганизм перед модификацией» могут использоваться взаимозаменяемо с терминами «немутированный штамм», «немодифицированный штамм», «немутированный микроорганизм», «немодифицированный микроорганизм» или «эталонный микроорганизм».
В настоящем изобретении эталонный микроорганизм конкретно не ограничен, при условии, что данный эталонный микроорганизм продуцирует путресцин, и мутированный штамм, обладающий повышенной способностью продуцировать путресцин, по сравнению с микроорганизмом дикого типа, также включен без ограничения. Его примеры могут включать Corynebacterium glutamicum АТСС13032 дикого типа, штамм дикого типа Corynebacterium glutamicum АТСС13869 или штаммы, в которых одна или более чем одна генетическая модификация добавлена в приведенные выше штаммы таким образом, чтобы усилить путь биосинтеза путресцина, но не ограничены ими.
Одна или более чем одна генетическая модификация может представлять собой, например, любую одну или более чем одну генетическую модификацию, выбранную из: усиления гена в пути биосинтеза путресцина; сверхэкспрессии активности путресцинового оперона; улучшения поставки и эффективности предшественника путресцина; улучшение экспорта путресцина; и ослабление активности гена в конкурентном пути, регулятора в направленном пути путресцинового оперона или гена импортера путресцина и генов импортера и лизиса путресцина, но не ограничивается ими.
Генетическая модификация усиления гена в пути биосинтеза путресцина может представлять собой, например, введение гена (speC), кодирующего орнитиндекарбоксилазу (ODC), происходящую из Е. coli W3110 дикого типа, в хромосому, но не ограничивается этим.
Генетическая модификация сверхэкспрессии активности путресцинового оперона может представлять собой, например, замену промотора генного кластера argCJBD, кодирующего ферменты, участвующие в синтезе орнитина из глутамата, но не ограничивается ею.
Генетическая модификация улучшения экспорта путресцина и ослабления активности гена в конкурирующем пути, регулятора в направленном пути путресцинового оперона или гена импортера путресцина и генов импортера и лизиса путресцина, может представлять собой, например, делецию гена (argF), кодирующего орнитинкарбамоилтрансферазу, и гена (NCgl1221), кодирующего экспортер глутамата, участвующий в экспорте глутамата, в хромосоме; или инактивацию активности белка NCgl1469, определенного в качестве гистонацетилтрансферазы НРА2 и родственной ацетилтрансферазы, но не ограничивается ими.
Штамм, имеющий одну или более чем одну генетическую модификацию, может представлять собой, например, штамм СС01-0064, обладающий способностью продуцировать путресцин, в котором: в АТСС 13032 ген (argF), кодирующий орнитинкарбамоилтрансферазу, и ген (NCgl1221), кодирующий экспортер глутамата, участвующий в экспорте глутамата, делетированы; в хромосому введен ген (speC), кодирующий орнитиндекарбоксилазу (ODC), происходящую из Е. coli W3110 дикого типа; и заменен промотор генного кластера argCJBD, кодирующего ферменты, участвующие в синтезе орнитина из глутамата (KCCM11138P, корейская патентная публикация №2012-0064046), или штамм СС01-0063, в котором инактивирована активность белка NCgl1469, определенного как гистонацетилтрансфераза НРА2, и родственной ацетилтрансферазы в штамме СС01-0064 (KCCM11240P, корейская патентная публикация №10-2013-0082478), но не ограничен ими.
В качестве альтернативы, микроорганизм, продуцирующий путресцин, может иметь ослабленную активность argJ, происходящего из штамма рода Corynebacterium, но не ограничивается этим.
Термин «ослабление» белка, как он здесь используется, имеет значение, охватывающее все из уменьшения активности или отсутствия активности по сравнению с собственной активностью. Термин «ослабление» можно использовать взаимозаменяемо с инактивацией, недостаточностью, понижающей регуляцией, снижением, уменьшением, ослаблением или тому подобными.
Ослабление также может включать: случай, когда активность самого полипептида снижена или устранена по сравнению с активностью полипептида, которой обладает исходный микроорганизм из-за мутации или тому подобного полинуклеотида, кодирующего данный полипептид; случай, когда вся активность и/или концентрация (уровень экспрессии) полипептида в клетке ниже по сравнению с нативным штаммом из-за ингибирования экспрессии гена полинуклеотида, кодирующего данный полипептид, или ингибирования трансляции в полипептид; случай, когда экспрессия полинуклеотида не достигается; и/или случай, когда данный полипептид не имеет активности, несмотря на экспрессию полинуклеотида. Термин «собственная активность» относится к активности конкретного полипептида, которой исходно обладал родительский штамм перед модификацией или микроорганизм дикого типа или немодифицированный микроорганизм, когда признак изменяется из-за генетической мутации, вызванной природными или искусственными факторами. Данный термин можно использовать взаимозаменяемо с термином «активность перед модификацией». Термины «инактивация, недостаточность, уменьшение, понижающая регуляция, снижение или ослабление» активности полипепида по сравнению с его собственной активностью означают, что активность данного полипептида снижена по сравнению с активностью конкретного полипептида, которой исходно обладал родительский штамм перед трансформацией или немодифицированный микроорганизм.
Ослабление активности полипептида можно осуществлять любым способом, известным в данной области, без ограничения ими, и оно может быть достигнуто путем применения разных способов, хорошо известных в данной области (например, Nakashima N et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014; 15(2):2773-2793 и Sambrook et al., Molecular Cloning 2012 и т.д.).
В настоящем раскрытии белок, являющийся мишенью ослабления, т.е. белок-мишень, может представлять собой argJ, но не ограничивается им.
1) Делеция части или всего гена, кодирующего данный полипептид;
2) модификация области контроля экспрессии (или последовательности контроля экспрессии) таким образом, чтобы понизить экспрессию гена, кодирующего данный полипептид;
3) модификация аминокислотной последовательности (например делеция/замена/присоединение по меньшей мере одной аминокислоты в данной аминокислотной последовательности), составляющей данный полипептид, таким образом, чтобы устранять или ослаблять активность данного полипептида;
4) модификация последовательности гена, кодирующего полинуклеотид, для устранения или ослабления активности полипептида (например делеция/замена/присоединение по меньшей мере одного нуклеотида нуклеиновой кислоты в нуклеотидной последовательности нуклеиновой кислоты гена полипептида таким образом, чтобы кодировать полипептид, модифицированный для устранения или ослабления активности полипептида);
5) модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR область транскрипта гена, кодирующего полипептид;
6) введение антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего данный полипептид;
7) добавление последовательности, комплементарной последовательности Шайна-Далгарно, перед последовательностью Шайна-Далгарно гена, кодирующего полипептид, таким образом, чтобы образовалась вторичная структура, которая делает невозможным присоединение рибосомы;
8) добавление промотора, который подлежит обратной транскрипции, к 3'-концу открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид (инженерия обратной транскрипции, RTE); или
9) комбинация двух или более чем двух, выбранных из пп. (1) - (8), но конкретно не ограничиваясь ими.
Например, они описываются следующим образом.
Делеция части или всего гена, кодирующего полипептид в п. (1), может представлять собой ликвидацию всего полинуклеотида, кодирующего эндогенный белок-мишень в хромосоме, замену полинуклеотидом с делецией некоторых нуклеотидов или замену маркерным геном.
Модификация области контроля экспрессии (или последовательности контроля экспрессии) в п. (2) может представлять собой возникновение мутации в области контроля экспрессии (или последовательности контроля экспрессии) посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации, или замену последовательностью, имеющей более слабую активность. Область контроля экспрессии включает промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, контролирующую окончание транскрипции и трансляции, но не ограничена ими.
Модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR область транскрипта гена, кодирующего полипептид в п. (5), может представлять собой, например, замену нуклеотидной последовательностью, кодирующей не эндогенный инициирующий кодон, а другой инициирующий кодон, имеющий более высокую скорость экспрессии полипептида, но не ограничивается этим.
Модификация аминокислотной последовательности или последовательности полинуклеотида в пп. (4) и (5) может представлять собой возникновение модификации последовательности посредством делеции, вставки, неконсервативной или консервативной замены, или их комбинации в аминокислотной последовательности полипептида или последовательности полинуклеотида, кодирующей данный полипептид, или замену аминокислотной последовательностью или последовательностью полинуклеотида, модифицированной так, чтобы иметь более слабую активность, или аминокислотной последовательностью или последовательностью полинуклеотида, модифицированной так, чтобы иметь незначительную активность, так, чтобы ослабить активность полипептида, но не ограничивается этим. Например, экспрессия гена может ингибироваться или ослабляться посредством введения мутации в последовательность полинуклеотида с образованием терминирующего кодона.
В отношении введения антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего полипептид в п. (6), можно сослаться, например, на литературу [Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews -Trends in a Genetics, Vol. 1(1) 1986].
Добавление последовательности, комплементарной последовательности Шайна-Далгарно гена, кодирующего данный полипептид, перед последовательностью Шайна-Далгарно таким образом, чтобы образовалась вторичная структура, которая делает невозможным присоединение рибосом, в п. (7) может сделать невозможной трансляцию мРНК или снизить скорость ее трансляции.
Добавление промотора, который подлежит обратной транскрипции, к 3'-концу открытой рамки считывания (ORF) последовательности гена, кодирующего данный полипептид (инженерия обратной транскрипции, RTE), в п. (8) может сделать антисмысловой нуклеотид комплементарным транскрипту гена, кодирующего данный полипептид, ослабляя посредством этого активность полипептида.
Микроорганизм, продуцирующий путресцин, может представлять собой любой микроорганизм, который может продуцировать путресцин посредством введения активности N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активности argE, происходящей из Е. coli, посредством вышеописанного способа. В целях настоящего изобретения микроорганизм, продуцирующий путресцин, представляет собой микроорганизм, обладающий повышенной способностью продуцировать целевой путресцин, в который активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность argE, происходящей из Е. coli, введены посредством вышеописанного способа, и активность argJ, происходящей из штамма рода Corynebacterium, ослаблена, и данный микроорганизм может представлять собой генетически модифицированный микроорганизм или рекомбинантный микроорганизм, но не ограничен им. В настоящем раскрытии термин «микроорганизм, продуцирующий путресцин» может быть использован взаимозаменяемо с термином «путресцин-продуцирующий микроорганизм» или «микроорганизм, обладающий способностью продуцировать путресцин».
Примеры данного микроорганизма могут включать микроорганизмы, принадлежащие к роду Corynebacterium, роду Escherichia, роду Enterbacter, роду Erwinia, роду Serratia, роду Providencia и роду Brevibacterium и, в частности, он может представлять собой микроорганизм рода Corynebacterium.
Более конкретно, микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris, Corynebacterium flavescens или тому подобные, и может представлять собой Corynebacterium glutamicum, и любой микроорганизм, принадлежащий к роду Corynebacterium, может быть включен без ограничения.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина, включающий культивирование в среде микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, включающий культивирование в среде микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli. N-ацетилтрансфераза, ацетилорнитиндеацетилаза, путресцин и микроорганизм являются такими, как описано выше.
Микроорганизм рода Corynebacterium может иметь ослабленную активность бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (argJ), происходящей из штамма рода Corynebacterium, но не ограничивается этим.
Данный способ может быть легко определен специалистом в данной области в оптимизированных условиях культивирования и условиях ферментативной активности, известных в данной области техники. В частности, стадия культивирования микроорганизма конкретно не ограничена, но данную стадию можно проводить известным способом периодического культивирования, способом непрерывного культивирования, способом культивирования с подпиткой или тому подобными. Условия культивирования могут быть конкретно не ограниченными, но корректировка до подходящего рН (например, от рН 5 до рН 9, в частности, от рН 6 до рН 8 и, наиболее конкретно, рН 6,8) может достигаться путем использования щелочного соединения (например гидроксида натрия, гидроксида калия или аммиака) или кислотного соединения (например фосфорной кислоты или серной кислоты), и аэробные условия могут поддерживаться посредством введения в культуру кислорода или кислородсодержащей газовой смеси. Может поддерживаться температура культивирования от 20 до 45°С и, в частности, от 25 до 45°С, и культивирование может проводиться в течение примерно 10-160 часов, но не ограничено этим. Путресцин, полученный путем культивирования, может высвобождаться в среду или может оставаться в клетках.
Кроме того, в среде для культивирования, подлежащей применению, могут использоваться в качестве источника углерода сахара и углеводы (например глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, крахмал и целлюлоза), масла и жиры (например соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например пальмитиновая кислота, стеариновая кислота и линолевая кислота), спирты (например глицерин и этанол), органические кислоты (например уксусная кислота) и тому подобные, отдельно или в комбинации, но источник углерода не ограничивается ими. В качестве источника азота можно использовать азотсодержащие органические соединения (например пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и тому подобные, отдельно или в комбинации, но источник азота не ограничен ими. В качестве источника фосфора можно использовать калия дигидрофосфат, дикалия гидрофосфат, соответствующую им натрийсодержащую соль и тому подобные, отдельно или в смеси, но источники фосфора не ограничены ими. Кроме того, среда также может содержать другие соли металлов (например сульфат магния или сульфат железа) и незаменимые стимулирующие рост вещества, такие как аминокислоты и витамины.
Способ может дополнительно включать выделение путресцина из культуральной среды или микроорганизма после стадии культивирования, но не ограничен этим.
Что касается способа выделения путресцина, продуцируемого на стадии культивирования, целевой путресцин можно выделять из культуры, используя подходящий способ, известный в данной области, согласно способу культивирования. Например, можно использовать центрифугирование, фильтрование, анионообменную хроматографию, кристаллизацию, ВЭЖХ (высокоэффективная жидкостная хроматография) и/или тому подобное, и целевой путресцин можно выделять из среды или микроорганизма посредством применения подходящего способа, известного в данной области.
Способ выделения путресцина может дополнительно включать стадию очистки. Стадию очистки можно проводить с использованием подходящего способа, известного в данной области. Следовательно, выделенный путресцин может иметь очищенную форму или может представлять собой ферментированную микроорганизмом жидкость, содержащую путресцин.
Согласно еще одному другому аспекту настоящего раскрытия предложена композиция для продуцирования путресцина, содержащая микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введена активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
N-ацетилтрансфераза, ацетилорнитиндеацетилаза, путресцин и микроорганизм являются такими, как описано выше.
Микроорганизм рода Corynebacterium может иметь ослабленную активность бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (argJ), происходящей из штамма рода Corynebacterium, но не ограничена этим.
Композиция для продуцирования путресцина может включать, без ограничения, элемент, способный вводить активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli. В частности, данный элемент может находиться в форме содержащейся в векторе таким образом, чтобы экспрессировался ген, связанный функциональным образом с клеткой-хозяином, в которую введен данный элемент, и форма является такой, как описано выше.
N-ацетилтрансфераза, происходящая из штамма рода Corynebacterium, может содержать аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней, и это описано выше.
N-ацетилтрансфераза, происходящая из штамма рода Corynebacterium, может содержать аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней, и это описано выше.
Ацетилорнитиндеацетилаза, происходящая от Е. coli, может содержать аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней, и это описано выше.
Композиция по настоящему изобретению может дополнительно содержать любой подходящий эксципиент, который обычно используют в композициях для получения аминокислот, и примеры эксципиента могут представлять собой консервант, увлажнитель, диспергирующий агент, суспендирующий агент, буфер, стабилизатор или изотонический агент, но не ограничены ими.
Согласно еще одному аспекту настоящего раскрытия предложено применение композиции для получения путресцина.
Согласно еще одному другому аспекту настоящего раскрытия предложено применение микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, для получения путресцина, где в данный микроорганизм введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из Е. coli.
Способ осуществления изобретения
Ниже настоящее раскрытие будет подробно описано со ссылкой на типичные воплощения. Однако специалисту в данной области очевидно, что данные типичные воплощения приведены лишь с целью иллюстрации и не предназначены для того, чтобы ограничивать объем настоящего раскрытия.
Пример 1: Способность продуцировать путресцин штамма с усиленным геном argJ, происходящим из Coryne
1-1. Получение штамма, имеющего argJ, происходящий из АТСС13869. усиленный в транспозонном гене штамма, продуцирующего путресцин. на основе АТСС13032
Для усиления гена argJ, кодирующего бифункциональную орнитинацетилтрансферазу/N-ацетилглутаматсинтазу, происходящую из штамма рода Corynebacterium, в продуцирующем путресцин штамме на основе Corynebacterium glutamicum АТСС 13032 ген argJ вводили в транспозонный ген данного штамма.
В качестве трансформирующего вектора, позволяющего вводить ген в хромосому посредством использования области транспозонного гена микроорганизма рода Corynebacterium, использовали pDZTn (WO 2009/125992). Кроме того, и в качестве промотора, использовали промотор lysCPl (WO 2009/096689, SEQ ID NO: 9), полученный модифицированием промотора гена оперона lysC-asd, происходящего из Corynebacterium glutamicum.
В частности, вектор pDZTn конструировали следующим образом. Для получения транспозонного гена, информацию по нуклеотидной последовательности о полной нуклеотидной последовательности транспозонного гена (№ доступа NCBI (Национальный центр биотехнологической информации) NC 003450, NCgI1021), происходящей из Corynebacterium glutamicum АТСС13032, получали из GenBank NIH, и на основе данной информации синтезировали две пары праймеров (Таблица 1, SEQ ID NO: 10-13).
ПЦР (полимеразная цепная реакция) проводили с использованием хромосомной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы, вместе с парой праймеров SEQ ID NO: 10 и 13. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 96°С в течение 30 с, отжиг при 58°С в течение 30 с и удлинение при 72°С в течение 1 мин.

В результате в качестве ПЦР-продуктов получали две пары транспозонных генов (Tn-А, Tn-В), включающих промоторные области из примерно 500 п.н. Tn-А (SEQ ID NO: 14) амплифицировали с использованием SEQ ID NO: 10 и 11 в качестве праймеров, и Tn-B (SEQ ID NO: 15) амплифицировали с использованием SEQ ID NO: 12 и 13 в качестве праймеров. Продукты амплификации клонировали в вектор pDZ, предварительно обработанный рестрикционным ферментом SalI, с использованием набора BD In-Fusion (BD), получая посредством этого вектор pDZTn. Между этими двумя амплифицированными продуктами включали целый ряд сайтов распознавания рестрикционных ферментов, искусственно встроенных во время конструирования праймеров.
Промотор lysCP1 получали следующим образом. ПЦР проводили с использованием в качестве матрицы хромосомной ДНК штамма СА01-0135 (WO 2009-096689, KCCM10919P), полученного трансформацией Corynebacterium glutamicum KFCC10881 вектором, содержащим промотор lysCP1, вместе с приведенными ниже праймерами SEQ ID NO: 16 и 17. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 30 с.
Последовательности использованных праймеров показаны в Таблице 2 ниже.
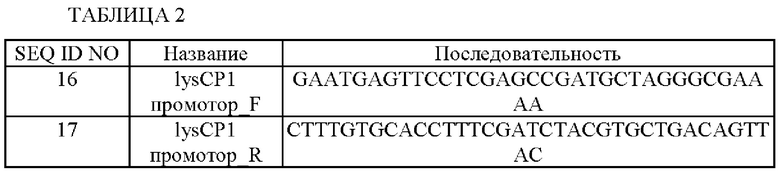
В результате в качестве ПЦР-продукта получали промоторную область lysCP1 (SEQ ID NO: 9).
Для получения гена argJ готовили пару праймеров SEQ ID NO: 18 и 19 для получения гомологичных рекомбинантных фрагментов открытой рамки считывания (ORF) argJ на основе нуклеотидной последовательности (SEQ ID NO: 6) гена argJ, происходящего из Corynebacterium glutamicum АТСС 13869.
ПЦР проводили с использованием хромосомной ДНК Corynebacterium glutamicum АТСС 13869 в качестве матрицы, вместе с парой праймеров SEQ ID NO: 18 и 19. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин и 30 с.
Последовательности использованных праймеров показаны в Таблице 3 ниже.
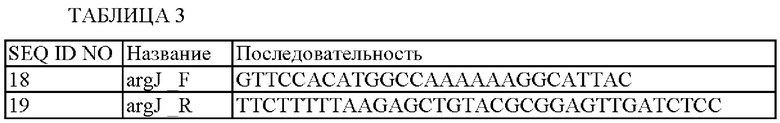
В результате, фрагменты гена argJ примерно 1,2 т.п.н. амплифицировали в качестве ПЦР-продуктов. Данные фрагменты подвергали электрофорезу на 0,8%-ном агарозном геле, и полосы с нужными размерами элюировали и очищали.
Полученный ранее вектор pDZTn обрабатывали рестрикционным ферментом XhoI, и соответствующие ПЦР-продукты (промоторная область lysCP1 и фрагменты гена argJ), полученные выше, клонировали посредством слияния в вектор с использованием набора для клонирования in-Fusion® HD (Clontech), и затем образующуюся плазмиду назвали pDZTn-lysCP1-argJ.
Затем сконструированную плазмиду pDZTn-lysCP1-argJ вводили в штамм СС01-0163 (US 9677099, KCCM11240P) посредством электропорации с получением трансформанта, где штамм был получен путем удаления активности белка NCgl1469, определенного как гистонацетилтрансфераза НРА2, и родственной ацетилтрансферазы в штамме СС01-0064 (US 9890404, KCCM11138P), обладающем способностью продуцировать путресцин, который получали делецией гена, кодирующего орнитинкарбамоилтрансферазу (argF), и гена (NCgl1221), кодирующего экспортер глутамата, участвующий в экспорте глутамата, введением гена (speC), кодирующего орнитиндекарбоксилазу (ODC), происходящую из штамма 3110 Е. coli дикого типа, в хромосому и заменой промотора генного кластера ArgCJBD, кодирующего ферменты, участвующие в синтезе орнитина из глутамата, в Corynebacterium glutamicum АТСС13032 дикого типа. Трансформант сеяли на чашку и культивировали на среде для чашек BHIS (37 г/л сердечно-мозговой вытяжки, 91 г/л сорбита и 2% агара), содержащей 25 мкг/мл канамицина и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), с образованием колоний. Посредством отбора белых колоний из колоний, образованных таким образом, отбирали штаммы трансформантов, в которые вводили плазмиду pDZTn-lysCPl-argJ.
Отобранные штаммы культивировали со встряхиванием в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), говяжий экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л) и рН 6,8) при 30°С в течение 8 часов. Затем каждую культуру клеток серийно разводили от 10-4 до 10-10 и затем сеяли на чашки и культивировали на твердой среде, содержащей X-gal, с образованием колоний. Посредством отбора белых колоний, появляющихся с относительно низкой частотой, среди образованных колоний, окончательно отбирали штамм, в который был введен ген, кодирующий argJ, посредством второго кроссинговера.
Проводили ПЦР на окончательно отобранном штамме посредством использования пары праймеров SEQ ID NO: 18 и 19 для подтверждения того, что был введен ген, кодирующий argJ, и мутантный штамм Corynebacterium glutamicum назвали СС01-0163 Tn:lysCP1-argJ.
1-2. Оценка способности продуцировать путресцин штамма на основе Coryne, усиленного геном argJ, происходящим из Coryne
Для исследования влияния на продуцирование путресцина при усилении гена argJ, происходящего из Corynebacterium, в путресцин-продуцирующем штамме, проводили сравнение способности продуцировать путресцин на мутантном штамме Corynebacterium glutamicum, полученном в Примере 1-1.
В частности, мутантный штамм Corynebacterium glutamicum (СС01-0163 Tn:lysCP1-argJ), полученный в Примере 1-1, сеяли на среду для чашек СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% говяжьего экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50% NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинин, и культивировали при 30°С в течение 24 часов.
Примерно одну платиновую петлю каждого штамма, культивированного таким образом, инокулировали в 25 мл титруемой среды (8% глюкозы, 0,25% соевого белка, 0,50% твердых веществ кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, 100 мкг биотина, 3 мг тиамина гидрохлорида, 3 мг кальций-пантотеновой кислоты, 3 мг никотинамида, 5% СаСО3, на основе 1 л) и затем культивировали со встряхиванием при 30°С и 200 об./мин в течение 98 часов. Затем в среду добавляли 1 мМ аргинин для культивирования каждого штамма. Измеряли концентрацию путресцина, полученного от каждой культуры, и результаты показаны в Таблице 4, приведенной ниже.

В результате, как показано в Таблице 4, продуцирование путресцина возрастало на 6% в мутантном штамме Corynebacterium glutamicum, в котором был усилен argJ, происходящий из Corynebacterium.
Пример 2: Введение argA и argE, происходящих из Е. coli, в путресцин-продуцирующий штамм и его способность продуцировать путресцин
2-1. Получение штамма посредством совместного введения как argA. так и argE. происходящих из Е. coli, в транспозонный ген штамма, продуцирующего путресцин. на основе АТСС 13032
Чтобы исследовать, улучшалась ли способность продуцировать путресцин при вставке argA, кодирующего происходящую из Е. coli N-ацетилглутаматсинтазу, и argE, кодирующего происходящую из Е. coli ацетилорнитиндеацетилазу, в путресцин-продуцирующий штамм на основе Corynebacterium glutamicum АТСС 13032 (СС01-0163), в транспозонный ген данного штамма вводили argA и argE.
С этой целью получали пару праймеров SEQ ID NO: 21 и 22 для получения гомологичных рекомбинантных фрагментов в области ORF argA из нуклеотидной последовательности (SEQ ID NO: 20) гена argA, происходящего из E.coli, и пару праймеров SEQ ID NO: 23 и 24 для получения гомологичных рекомбинантных фрагментов в области ORF argE из нуклеотидной последовательности (SEQ ID NO: 4) гена argE.
Последовательности данных праймеров показаны в Таблице 5.
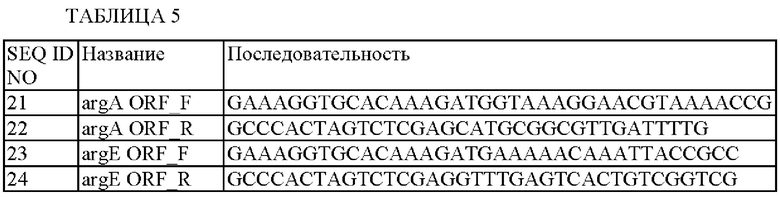
Промоторную область lysCP1 получали посредством ПЦР с использованием хромосомы штамма KCCM10919P в качестве матрицы, вместе с парой праймеров SEQ ID NO: 16 и 17, таким же способом, как и в Примере 1-1.
Для получения гена argA проводили ПЦР с помощью хромосомы штамма W3110 Е. coli в качестве матрицы, вместе с парой праймеров SEQ ID NO: 21 и 22. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин и 30 с.
В результате фрагменты гена argA размером примерно 1,6 т.п.н. амплифицировали в качестве ПЦР продуктов, и данные фрагменты клонировали посредством слияния совместно с промоторной областью lysCP1 в вектор pDZTn, полученный в Примере 1-1, таким же способом, как в Примере 1-1. Образованную плазмиду назвали pDZTn-lysCP1-argA.
Вектор pDZTn, полученный в Примере 1-1, обрабатывали рестрикционным ферментом Xhol, и соответствующие полученные выше ПЦР-продукты (промоторная область lysCP1 и фрагмент гена argA) клонировали посредством слияния в вектор тем же самым способом, что и в Примере 1-1, и затем образующуюся плазмиду назвали pDZTn-lysCP1-argA.
Для получения гена argE проводили ПЦР при помощи хромосомы штамма W3110 Е. coli в качестве матрицы, вместе с парой праймеров SEQ ID NO: 23 и 24, таким же способом, как и выше. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин и 30 с.
В результате фрагменты гена argE размером примерно 1,4 т.п.н. амплифицировали в качестве ПЦР-продуктов, и данные фрагменты клонировали посредством слияния совместно с полученной ранее промоторной областью lysCP1 в вектор pDZTn, полученный в Примере 1-1, таким же способом, как в Примере 1-1. Образовавшуюся плазмиду назвали pDZTn-lysCP1-argE.
Затем плазмиду pDZTn-lysCP1-argA вводили в СС01-0163 посредством электрофореза с получением штаммов трансформантов, и штаммы трансформантов, в которые была введена плазмида pDZTn-lysCP1-argA, отбирали таким же способом, как и в Примере 1-1.
Отобранные штаммы подвергали второму кроссинговеру для окончательного отбора штамма, в который был введен ген, кодирущий argA. На окончательно отобранном штамме проводили ПЦР с использованием пары праймеров SEQ ID NO: 21 и 22 для подтверждения того, что был введен ген, кодирущий argA, и мутантный штамм Corynebacterium glutamicum назвали СС01-0163 Tn:lysCP1-argA.
Для введения argE в полученный мутантный штамм Corynebacterium glutamicum, в который был введен argA, сконструированную ранее pDZTn-lysCPl-argE трансформировали в СС01-0163 Tn:lysCP1-argA таким же способом, как и выше, и проводили ПЦР на отобранном наконец штамме, вместе с парой праймеров SEQ ID NO: 23 и 24, для подтверждения того, что argE был введен в транспозон.
Мутантный штамм Corynebacterium glutamicum, отобранный таким образом, был назван СС01-0163 Tn:lysCP1-argA Tn:lysCPl-argE.
2-2. Оценка способности продуцировать путресцин путресцина-продуцирующего штамма рода Corynebacterium, в который были введены происходящие из Е. coli argA и argE
Для исследования влияния на продуцирование путресцина введения в путресцин-продуцирующий штамм argA и argE, происходящих из Е. coli, проводили сравнение способности продуцировать путресцин на мутантном штамме Corynebacterium glutamicum, полученном в Примере 2-1.
Каждый из мутантного штамма Corynebacterium glutamicum (СС01-0163 Tn:lysCP1-argA Tn:lysCP1-argE), полученного в Примере 2-1, и родительского штамма (СС01-0163) культивировали таким же способом, как и в Примере 1-2, и измеряли концентрацию путресцина, продуцируемого из каждой культуры. Результаты показаны в Таблице 6 ниже.

В результате, как показано в Таблице 6, продукция путресцина увеличивалась на 9,8% в мутантном штамме Corynebacterium glutamicum, в который были введены происходящие из Е. coli гены argA и argE.
Пример 3: Способность продуцировать путресцин штамма, усиленного геном Ncgl2644, происходящим из Coryne
3-1. Получение штамма, в котором Ncgl2644, происходящий из АТСС13869, был введен в транспозонный ген путресцин-продуцирующего штамма на основе АТСС13032
Для исследования того, была ли улучшена способность продуцировать путресцин при усилении гена Ncgl2644, кодирующего N-ацетилтрансферазу семейства родственных GCN5 N-ацетилтрансфераз (GNAT), происходящую из Corynebacterium glutamicum АТСС 13869, в продуцирующем путресцин штамме (СС01-0163) на основе Corynebacterium glutamicum АТСС 13032 (СС01-0163), Ncgl2644 вводили и усиливали в транспозонном гене данного штамма.
С этой целью получали пару праймеров SEQ ID NO: 25 и 26 для получения гомологичных рекомбинантных фрагментов области ORF Ncgl2644 из нуклеотидной последовательности (SEQ ID NO: 2) гена Ncgl2644, происходящего из Corynebacterium.
Последовательности праймеров показаны в Таблице 7.

Для получения гена Ncgl2644 проводили ПЦР при помощи хромосомы штамма АТСС13869 Corynebacterium glutamicum в качестве матрицы, вместе с парой праймеров SEQ ID NO: 25 и 26. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин. В результате, фрагменты гена Ncgl2644 примерно из 1 т.п.н. амплифицировали в качестве ПЦР-продуктов, и данные фрагменты клонировали посредством слияния, вместе с промоторной областью lysCP1, полученной в Примере 2-1, в вектор pDZTn, полученный в Примере 1-1, таким же способом, как и в Примере 1-1. Образовавшуюся плазмиду назвали pDZTn-lysCP1-Ncgl2644.
Затем данную плазмиду pDZTn-lysCP1-Ncgl2644 вводили в СС01-0163 посредством электрофореза с получением штаммов-трансформантов, и штаммы-трансформанты, в которые была введена плазмида pDZTn-lysCP1-Ncgl2644, отбирали таким же способом, что и в Примере 1-1.
Отобранные штаммы подвергали второму кроссинговеру для окончательного отбора штамма, в который был введен ген, кодирующий Ncgl2644. На данном конечном отобранном штамме проводили ПЦР с использованием пары праймеров SEQ ID NO: 25 и 26 для подтверждения того, что был введен ген, кодирующий Ncgl2644, и данный мутантный штамм Corynebacterium glutamicum назвали СС01-0163 Tn:lysCP1-Ncgl2644.
3-2. Оценка способности продуцировать путресцин путресцин-продуцирующего штамма рода Corynebacterium, в котором был усилен Ncgl2644. происходящий из Coryne
Для исследования влияния на продуцирование путресцина при усилении гена Ncgl2644, происходящего из Corynebacterium, в штамме, продуцирующем путресцин, проводили сравнение способности продуцировать путресцин на мутантном штамме Corynebacterium glutamicum, полученном в Примере 3-1.
В частности, каждый из мутантного штамма Corynebacterium glutamicum (СС01-0163), полученного в Примере 3-1, и родительского штамма (СС01-0163) культивировали таким же способом, как и в Примере 1-2, и измеряли концентрацию путресцина, полученного из каждой культуры. Результаты показаны в Таблице 8 ниже.

В результате, как показано в Таблице 8, способность продуцировать путресцин снижалась при усилении Ncgl2644, происходящего из Corynebacterium. Это может происходить из-за появления бутылочного горлышка в данном биосинтетическом пути, вызванного увеличением концентрации ацетилглутамата - промежуточного метаболита в пути продукции путресцина. Следовательно, влияние Ncgl2644 на продуцирование путресцина оценивали посредством дополнительных примеров, приведенных ниже.
Пример 4: Способность продуцировать путресцин штамма с делецией гена argJ и усилением Ncgl2644
4-1. Получение штамма, в котором делетирован argJ в продуцирующем путресцин штамме на основе АТСС13032
Получали штамм, в котором ген argJ, происходящий из Corynebacterium glutamicum АТСС13032, был делетирован в продуцирующем путресцин штамме на основе Corynebacterium glutamicum АТСС13032 (СС01-0163).
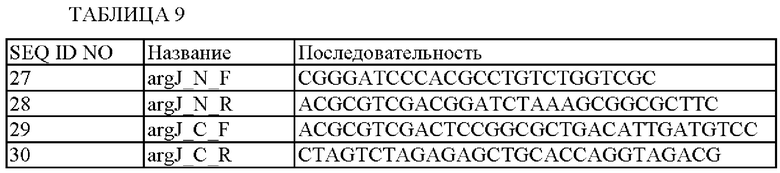
С этой целью получали пару праймеров SEQ ID NO: 27 и 28 для получения гомологичных рекомбинантных фрагментов N-концевой области argJ и пару праймеров SEQ ID NO: 29 и 30 для получения гомологичных рекомбинантных фрагментов С-концевой области argJ из нуклеотидной последовательности (SEQ ID NO: 8) гена argJ, происходящего из Corynebacterium glutamicum АТСС13032.
Последовательности данных праймеров показаны в Таблице 9.
Для получения гомологичных рекомбинантных фрагментов С-концевой области и N-концевой области argJ проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы, вместе с парой праймеров SEQ ID NO: 27 и 28, и парой праймеров SEQ ID NO: 29 и 30. В качестве полимеразы использовали ДНК-полимеразу высокой точности PfuUltra™ (Stratagene), и устанавливали следующие условия ПЦР: 30 циклов денатурации при 95°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 30 с. В результате, ПЦР-фрагменты N-концевой области и С-концевой области амплифицировали в виде ПЦР-продуктов соответственно, и данные фрагменты подвергали электрофорезу на 0,8%-ном агарозном геле, и полосы с нужными размерами элюировали и очищали.
Полученные фрагменты N-концевой области обрабатывали рестрикционными ферментами BamHI и SalI, а полученные фрагменты С-концевой области обрабатывали рестрикционными ферментами SalI и XbaI. Обработанные фрагменты клонировали в вектор pDZ, обработанный рестрикционными ферментами BamHI и XbaI, для конструирования плазмиды pDZ-1'argJ(K/O).
Сконструированную плазмиду pDZ-1'argJ(K/O) вводили в Corynebacterium glutamicum СС01-0163 посредством электрофореза с получением трансформанта, и штаммы с введенным pDZ-1'NCglargJ(K/O) отбирали таким же способом, как и в Примере 1-1.
Из отобранных штаммов окончательно отбирали штамм Corynebacterium glutamicum, имеющий ослабленную продуктивность по путресцину, посредством делеции гена, кодирующего argJ, и мутантный штамм Corynebacterium glutamicum с ослабленной способностью к экспорту путресцина назвали СС01-0163 ΔargJ.
Ген Ncgl2644 трансформировали в полученный штамм СС01-0163 ΔargJ таким же способом, как и в Примере 3-1, с получением штамма СС01-0163 ΔargJ Tn:lysCP1-Ncgl2644.
4-2. Оценка способности продуцировать путресцин путресцин-продуцирующего щтамма рода Corynebacterium, в котором был делетирован argJ. происходящий из Coryne, и усилен Ncgl2644
Для исследования влияния на продуцирование путресцина при делеции гена argJ, происходящего из Corynebacterium, и усиления гена Ncgl2644, происходящего из Corynebacterium, в путресцин-продуцирующем штамме, проводили сравнение способности продуцировать путресцин на мутантном штамме Corynebacterium glutamicum, полученном в Примере 4-1.
В частности, каждый из мутантного штамма Corynebacterium glutamicum (СС01-0163 ΔargJ Tn:lysCP1-Ncgl2644) и родительского штамма (СС01-0163 ΔargJ) культивировали таким же способом, как и в Примере 1-2, и измеряли концентрацию путресцина, продуцированного от каждой культуры. Результаты показаны в Таблице 10 ниже.

В результате, как показано в Таблице 10, штамм с делецией argJ имел сложность со способностью продуцировать путресцин, и даже несмотря на то, что был введен Ncgl2644, способный генерировать ацетилглутамат, продуцирование путресцина было низким из-за отсутствия фермента, который может деацетилировать ацетилорнитин промежуточный метаболит, генерируемый после введения. Следовательно, штамм, с совместно усиленными Ncgl2644 и argE, оценивали на способность продуцировать путресцин посредством дополнителных рабочих примеров.
Пример 5: Способность продуцировать путресцин штаммов, усиленных геном Ncgl2644, происходящим из Coryne, и с введением гена argE, происходящего из Е. coli
5-1. Получение штаммов, усиленных геном Ncgl2644. из штаммов, продуцирующих путресцин. на основе АТСС 13032
В штамм СС01-0163 Tn:lysCP1-Ncgl2644, полученный в Примере 3-1, и штамм СС01-0163 ΔargJ Tn:lysCP1-Ncgl2644, полученный в Примере 4-1, вводили argE, кодирующий ацетил орнитинде ацетил азу, способную деацетилировать ацетилорнитин, происходящую из Е. coli W3110, argE.
В частности, плазмиду pDZTn-lysCP1-argE, сконструированную в Примере 2-1, трансформировали в штамм СС01-0163 Tn:lysCP1-Ncgl2644 и штамм СС01-0163 ΔargJ Tn:lysCP1-Ncgl2644 таким же способом, как и в Примере 2-1, с получением трансформантов, и штаммы трансформантов, в которые была введена плазмида pDZTn-lysCP1-argE, отбирали таким же способом, как и в Примере 1-1.
Из отобранных штаммов окончательно отбирали штаммы Corynebacterium glutamicum с введенным argE, и получали подготовленные штаммы Corynebacterium glutamicum СС01-0163 Tn:lysCP1-Ncgl2644 Tn: lysCP1-argE и CC01-0163 ΔargJ Tn:lysCP1-Ncgl2644 Tn:lysCP1-argE. CC01-0163 ΔargJ Tn:lysCPl-Ncgl2644 Tn:lysCP1-argE назвали Corynebacterium glutamicum CC01-1425. Corynebacterium glutamicum CC01-1425 (CC01-0163 ΔargJ Tn:lysCP1-Ncgl2644 Tn:lysCP1-argE) депонировали (KCCM12774P) 24 июля 2020 г.
5-2. Оценка способности продуцировать путресцин путресцин-продуцирующего штамма рода Corynebacterium, в котором были усилены ген Ncgl2644 и argE
Для исследования влияния на продуцирование путресцина при усилении Ncgl2644, происходящего из Corynebacterium, и введении argE, происходящего из Corynebacterium, в штамм, продуцирующий путресцин, проводили сравнение способности продуцировать путресцин на мутантном штамме Corynebacterium glutamicum, полученном в Примере 5-1.
В частности, каждый из мутантных штаммов Corynebacterium glutamicum (СС01-0163 Tn:lysCP1-Ncgl2644 Tn:lysCP1-argE и CC01-0163 AargJ Tn:lysCPl-Ncgl2644 Tn:lysCP1-argE) и родительский штамм (СС01-0163) культивировали таким же способом, как и в Примере 1-2, и измеряли концентрацию путресцина, продуцированного из каждой культуры. Данные результаты показаны в Таблице 11 ниже.

Как показано в Таблице 11, совместное введение Ncgl2644 и argE приводило к значительно улучшенной продукции путресцина по сравнению со штаммом СС01-0163. В частности, штамм СС01-0163 ΔargJ Tn:lysCPl-Ncgl2644 Tn: lysCP1-argE с устраненной активностью argJ демонстрировал способность продуцировать путресцин, улучшенную на 36,6% по сравнению со штаммом СС01-0163, и штамм СС01-0163 Tn:lysCP1-Ncgl2644 Tn:lysCP1-argE с оставшейся активностью argJ демонстрировал низкую способность продуцировать путресцин по сравнению со штаммом с устраненной активностью argJ, но обладал способностью продуцировать путресцин, улучшенную только на 30,8% по сравнению со штаммом СС01-0163. Кроме того, штамм СС01-0163 Tn:lysCP1-Ncgl2644 Tn:lysCP1-argE с оставшейся активностью argJ демонстрировал значимо более высокую продукцию путресцина по сравнению с мутантным штаммом Corynebacterium glutamicum, в который были совместно вводили argA и argE, происходящие из Е. coli, демонстрирующие аналогичные функции Ncgl2644 (Таблица 6).
Результаты настоящих примеров подтвердили, что делеция argJ и совместная экспрессия Ncgl2644 и argE приводили к более значительной способности продуцировать путресцин по сравнению с отсутствием делеции argJ или делецией argJ и введением argA и argE, происходящих из Е. coli, демонстрирующих аналогичные функции, в Ncgl2644.
Как изложено выше, специалист в области, к которой относится настоящее раскрытие, может понять, что настоящее изобретение может быть воплощено в других конкретных формах без отступления от его технической сущности или существенных характеристик. Следовательно, описанные выше типичные воплощения следует истолковывать как типичные и не ограничивающие настоящее изобретение. Объем настоящего изобретения следует понимать таким образом, что все изменения или модификации, производные от определений и объема формулы изобретения, и их эквиваленты входят в объем данного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> МИКРООРГАНИЗМ ДЛЯ ПРОДУЦИРОВАНИЯ ПУТРЕСЦИНА И СПОСОБ ПОЛУЧЕНИЯ
ПУТРЕСЦИНА С ЕГО ПРИМЕНЕНИЕМ
<130> OPA21055
<150> KR 10-2020-0101894
<151> 2020-08-13
<160> 30
<170> KoPatentIn 3.0
<210> 1
<211> 335
<212> PRT
<213> Unknown
<220>
<223> N-ацетилтрансфераза семейства GNAT C. glutamicum
<400> 1
Met Thr Pro Ser Leu Pro Arg Phe Arg Ser Gln Lys Pro Ala Val Gly
1 5 10 15
Asp Arg Val Val Ala Arg Arg Arg Ile Pro Gly Ala Asn Val His Trp
20 25 30
Thr Asp Ala Ile Gly His Val Ile Gly Val Asp Pro Leu Val Val Arg
35 40 45
Pro Gln Ser Val Gly Gly Met Pro Ser Asp Ala Glu Glu Ile Val Ile
50 55 60
Pro Asp Asp Gln Leu Glu Val Ile Lys Ile Leu Ser Pro Arg Thr Ile
65 70 75 80
Arg Asn Ser Asp Ile Arg Ala Val Glu Val Ala Thr Ala Lys Ala Phe
85 90 95
Pro Gly Leu Val Asn Glu Trp His Asp Gly Trp Leu Leu Arg Ala Gly
100 105 110
Asp Gly Ile Ala Glu Arg Ser Asn Ser Ala Ser Pro Leu Gly Pro Ser
115 120 125
Val Gly Phe Glu Pro Val Pro Met Glu Asp Ile Ser Arg Phe Tyr Ala
130 135 140
Arg His Asp Leu Pro Val Lys Leu His Ile Pro Glu Arg Ile Gly Arg
145 150 155 160
Pro Ala Gln Lys Val Ile Asp Ala Asp Pro Gln Lys Trp Val Met Gly
165 170 175
Pro Glu Ile Leu Val Met Thr Lys Ser Leu Asp His Val Glu Ser His
180 185 190
Glu Leu Pro Gly Gly Leu Glu Phe Ser Val Asp Lys Gln Pro Asp Gln
195 200 205
Glu Trp Leu Gly Met Tyr His Phe Arg Gly Gln Ala Leu Pro Ala His
210 215 220
Ala Leu Glu Leu Leu Arg Thr Gln Ile Glu Gly Arg Met Gly Phe Gly
225 230 235 240
Arg Leu Thr Thr Pro Ala Gly Gln Thr Val Ala Ile Thr Arg Ala Thr
245 250 255
Ile Thr Ala Ala Glu Glu Arg Ile Phe Leu Gly Tyr Ser Ala Val Glu
260 265 270
Val Asp Pro Ala Phe Arg Arg Gln Gly Leu Gly Thr Ala Leu Gly Ser
275 280 285
Arg Ile Gln Glu Trp Gly Ala Glu Gln His Ala Gln Glu Ala Tyr Leu
290 295 300
Gln Val Val Ala His Asn Glu Ala Gly Ile Gly Leu Tyr Gln Lys Leu
305 310 315 320
Gly Phe Ser Glu His His Arg His Arg Tyr Ala Glu Arg Lys Phe
325 330 335
<210> 2
<211> 1008
<212> DNA
<213> Unknown
<220>
<223> N-ацетилтрансфераза (Ncgl2644) семейства GNAT C. glutamicum
<400> 2
atgacgccta gtcttccccg tttccgcagc cagaaacctg ccgtcggcga tcgtgttgtt 60
gcacgtcgcc ggattcctgg tgccaatgtg cattggacag atgccattgg ccatgtgatt 120
ggggtggatc cgttggtggt tcgcccgcag tcggttggtg ggatgccgtc ggatgcggaa 180
gaaattgtca ttcctgatga tcagcttgag gtgattaaga ttttgtcgcc gcgcaccatt 240
aggaattcgg atattcgtgc ggtggaggtt gccacggcga aggcctttcc ggggctggtc 300
aatgagtggc atgatggttg gctgctgcgt gccggtgatg gcattgcgga gcgttctaat 360
tctgcgtcgc cactcggccc aagtgttggt tttgagccgg taccgatgga ggatatttcg 420
cggttttacg caaggcacga tctccccgtg aagctgcaca ttccggagcg gattggtcgg 480
cctgcgcaga aagtcattga cgccgatccc cagaaatggg tgatgggccc ggagattttg 540
gtgatgacga aatctttgga ccatgtggag tcgcacgagt tgcccggtgg cctagaattt 600
agcgtcgata agcagcctga ccaggagtgg ctgggcatgt accatttccg cggacaggcg 660
ttgcccgctc acgcccttga gcttttgcgc acgcaaatcg agggccgcat ggggttcggg 720
cgcctgacca cgccggcggg gcaaaccgtc gcgatcacgc gcgccaccat cacggctgcg 780
gaggagcgca tatttttggg ctattcagcg gtcgaggtgg atcctgcttt tcgacgtcag 840
gggctgggca ccgcgctcgg ctcgcgcatc caggagtggg gcgccgagca acacgcacaa 900
gaggcatatc tccaggttgt cgcccataat gaagcaggta tcggcctgta tcaaaagctc 960
gggttcagtg aacaccaccg acaccggtac gccgaacgga aattctaa 1008
<210> 3
<211> 408
<212> PRT
<213> Unknown
<220>
<223> argE E.coli
<400> 3
Met Asp Lys Leu Leu Glu Arg Phe Leu Asn Tyr Val Ser Leu Asp Thr
1 5 10 15
Gln Ser Lys Ala Gly Val Arg Gln Val Pro Ser Thr Glu Gly Gln Trp
20 25 30
Lys Leu Leu His Leu Leu Lys Glu Gln Leu Glu Glu Met Gly Leu Ile
35 40 45
Asn Val Thr Leu Ser Glu Lys Gly Thr Leu Met Ala Thr Leu Pro Ala
50 55 60
Asn Val Pro Gly Asp Ile Pro Ala Ile Gly Phe Ile Ser His Val Asp
65 70 75 80
Thr Ser Pro Asp Cys Ser Gly Lys Asn Val Asn Pro Gln Ile Val Glu
85 90 95
Asn Tyr Arg Gly Gly Asp Ile Ala Leu Gly Ile Gly Asp Glu Val Leu
100 105 110
Ser Pro Val Met Phe Pro Val Leu His Gln Leu Leu Gly Gln Thr Leu
115 120 125
Ile Thr Thr Asp Gly Lys Thr Leu Leu Gly Ala Asp Asp Lys Ala Gly
130 135 140
Ile Ala Glu Ile Met Thr Ala Leu Ala Val Leu Gln Gln Lys Lys Ile
145 150 155 160
Pro His Gly Asp Ile Arg Val Ala Phe Thr Pro Asp Glu Glu Val Gly
165 170 175
Lys Gly Ala Lys His Phe Asp Val Asp Ala Phe Asp Ala Arg Trp Ala
180 185 190
Tyr Thr Val Asp Gly Gly Gly Val Gly Glu Leu Glu Phe Glu Asn Phe
195 200 205
Asn Ala Ala Ser Val Asn Ile Lys Ile Val Gly Asn Asn Val His Pro
210 215 220
Gly Thr Ala Lys Gly Val Met Val Asn Ala Leu Ser Leu Ala Ala Arg
225 230 235 240
Ile His Ala Glu Val Pro Ala Asp Glu Ser Pro Glu Met Thr Glu Gly
245 250 255
Tyr Glu Gly Phe Tyr His Leu Ala Ser Met Lys Gly Thr Val Glu Arg
260 265 270
Ala Asp Met His Tyr Ile Ile Arg Asp Phe Asp Arg Lys Gln Phe Glu
275 280 285
Ala Arg Lys Arg Lys Met Met Glu Ile Ala Lys Lys Val Gly Lys Gly
290 295 300
Leu His Pro Asp Cys Tyr Ile Glu Leu Val Ile Glu Asp Ser Tyr Tyr
305 310 315 320
Asn Met Arg Glu Lys Val Val Glu His Pro His Ile Leu Asp Ile Ala
325 330 335
Gln Gln Ala Met Arg Asp Cys Asp Ile Glu Pro Glu Leu Lys Pro Ile
340 345 350
Arg Gly Gly Thr Asp Gly Ala Gln Leu Ser Phe Met Gly Leu Pro Cys
355 360 365
Pro Asn Leu Phe Thr Gly Gly Tyr Asn Tyr His Gly Lys His Glu Phe
370 375 380
Val Thr Leu Glu Gly Met Glu Lys Ala Val Gln Val Ile Val Arg Ile
385 390 395 400
Ala Glu Leu Thr Ala Gln Arg Lys
405
<210> 4
<211> 1227
<212> DNA
<213> Unknown
<220>
<223> argE E.coli
<400> 4
atggataaac tacttgagcg atttttgaac tacgtgtctc tggataccca atcaaaagca 60
ggggtgagac aggttcccag cacggaaggc caatggaagt tattgcatct gctgaaagag 120
cagctcgaag agatggggct tatcaatgtg accttaagtg agaagggcac tttgatggcg 180
acgttaccgg ctaacgtccc tggcgatatc ccggcgattg gctttatttc tcatgtggat 240
acctcaccgg attgcagcgg caaaaatgtg aatccgcaaa ttgttgaaaa ctatcgcggt 300
ggcgatattg cgctgggtat cggcgatgaa gttttatcac cggttatgtt cccggtgctg 360
catcagctac tgggtcagac gctgattacc accgatggta aaaccttgtt aggtgccgat 420
gacaaagcag gtattgcaga aatcatgacc gcgctggcgg tattgcaaca gaaaaaaatt 480
ccgcatggtg atattcgcgt cgcctttacc ccggatgaag aagtgggcaa aggggcgaaa 540
cattttgatg ttgacgcctt cgatgcccgc tgggcttaca ctgttgatgg tggtggcgta 600
ggcgaactgg agtttgaaaa cttcaacgcc gcgtcggtca atatcaaaat tgtcggtaac 660
aatgttcatc cgggcacggc gaaaggagtg atggtaaatg cgctgtcgct ggcggcacgt 720
attcatgcgg aagttccggc ggatgaaagc ccggaaatga cagaaggcta tgaaggtttc 780
tatcatctgg cgagcatgaa aggcaccgtt gaacgggccg atatgcacta catcatccgt 840
gatttcgacc gtaaacagtt tgaagcgcgt aaacgtaaaa tgatggagat cgccaaaaaa 900
gtgggcaaag ggttacatcc tgattgctac attgaactgg tgattgaaga cagttactac 960
aatatgcgcg agaaagtggt tgagcatccg catattctcg atatcgccca gcaggcgatg 1020
cgcgattgcg atattgaacc ggaactgaaa ccgatccgcg gtggtaccga cggcgcgcag 1080
ttgtcgttta tgggattacc gtgcccgaac ctgttcactg gcggttacaa ctatcatggt 1140
aagcatgagt ttgtgactct ggaaggtatg gaaaaagcgg tgcaggtgat cgtccgtatt 1200
gccgagttaa cggcgcaacg gaagtaa 1227
<210> 5
<211> 388
<212> PRT
<213> Unknown
<220>
<223> argJ C. glutamicum 13869
<400> 5
Met Ala Lys Lys Gly Ile Thr Ala Pro Lys Gly Phe Val Ala Ser Ala
1 5 10 15
Thr Thr Ala Gly Ile Lys Ala Ser Gly Asn Pro Asp Met Ala Leu Val
20 25 30
Val Asn Gln Gly Pro Glu Phe Ser Ala Ala Ala Val Phe Thr Arg Asn
35 40 45
Arg Val Phe Ala Ala Pro Val Lys Val Ser Arg Glu Asn Val Ala Asp
50 55 60
Gly Gln Ile Arg Ala Val Leu Tyr Asn Ala Gly Asn Ala Asn Ala Cys
65 70 75 80
Asn Gly Leu Gln Gly Glu Lys Asp Ala Arg Glu Ser Val Ser His Leu
85 90 95
Ala Gln Asn Leu Gly Leu Glu Asp Ser Asp Ile Gly Val Cys Ser Thr
100 105 110
Gly Leu Ile Gly Glu Leu Leu Pro Met Asp Lys Leu Asn Thr Gly Ile
115 120 125
Asp Gln Leu Thr Ala Glu Gly Ala Leu Gly Asp Asn Gly Ala Ala Ala
130 135 140
Ala Lys Ala Ile Met Thr Thr Asp Thr Val Asp Lys Glu Thr Val Val
145 150 155 160
Phe Ala Asp Gly Trp Thr Val Gly Gly Met Gly Lys Gly Val Gly Met
165 170 175
Met Ala Pro Ser Leu Ala Thr Met Leu Val Cys Leu Thr Thr Asp Ala
180 185 190
Ser Val Thr Gln Glu Met Ala Gln Ile Ala Leu Ala Asn Ala Thr Ala
195 200 205
Val Thr Phe Asp Thr Leu Asp Ile Asp Gly Ser Thr Ser Thr Asn Asp
210 215 220
Thr Val Phe Leu Leu Ala Ser Gly Ala Ser Gly Ile Thr Pro Thr Gln
225 230 235 240
Asp Glu Leu Asn Asp Ala Val Tyr Ala Ala Cys Ser Asp Ile Ala Ala
245 250 255
Lys Leu Gln Ala Asp Ala Glu Gly Val Thr Lys Arg Val Ala Val Thr
260 265 270
Val Val Gly Thr Thr Asn Asn Glu Gln Ala Ile Asn Ala Ala Arg Thr
275 280 285
Val Ala Arg Asp Asn Leu Phe Lys Cys Ala Met Phe Gly Ser Asp Pro
290 295 300
Asn Trp Gly Arg Val Leu Ala Ala Val Gly Met Ala Asp Ala Asp Met
305 310 315 320
Glu Pro Glu Lys Ile Ser Val Phe Phe Asn Asp Gln Ala Val Cys Leu
325 330 335
Asp Ser Thr Gly Ala Pro Gly Ala Arg Glu Val Asp Leu Ser Gly Ala
340 345 350
Asp Ile Asp Val Arg Ile Asp Leu Gly Thr Ser Gly Glu Gly Gln Ala
355 360 365
Thr Val Arg Thr Thr Asp Leu Ser Phe Ser Tyr Val Glu Ile Asn Ser
370 375 380
Ala Tyr Ser Ser
385
<210> 6
<211> 1167
<212> DNA
<213> Unknown
<220>
<223> argJ C. glutamicum 13869
<400> 6
atggccaaaa aaggcattac cgcgccgaaa ggcttcgttg cttctgcaac gaccgcgggt 60
attaaagctt ctggcaatcc tgacatggcg ttggtggtta accagggtcc agagttttcc 120
gcagcggccg tgtttacacg caaccgagtt ttcgcagcgc ctgtgaaggt gagccgggag 180
aacgttgctg atggccagat cagggctgtt ttgtacaacg ctggtaatgc taatgcgtgt 240
aatggtctgc agggtgagaa ggatgctcgt gagtctgttt ctcatctagc tcaaaatttg 300
ggcttggagg attccgatat tggtgtgtgt tccactggtc ttattggtga gttgcttccg 360
atggataagc tcaatacagg tattgatcag ctgaccgctg agggcgcttt gggtgacaat 420
ggtgcagctg ctgccaaggc gatcatgacc actgacacgg tggataagga aaccgtcgtg 480
tttgctgatg gttggactgt cggcggaatg ggcaagggcg tgggcatgat ggcgccgtct 540
cttgccacca tgctggtctg cttgaccact gatgcatccg ttactcagga aatggctcag 600
attgcgctgg ctaatgctac ggccgttacg tttgacaccc tggatattga tggatcaacc 660
tccaccaatg acaccgtgtt cctgctggca tctggcgcta gcggaatcac cccaactcag 720
gatgaactca acgatgcggt gtacgcagct tgttctgata tcgcagcgaa gcttcaggct 780
gatgcagagg gggtgaccaa gcgcgttgct gtgacagtgg tgggaaccac caacaacgag 840
caggcgatca atgcggctcg cacggttgct cgtgacaatt tgttcaagtg cgcaatgttt 900
ggatctgatc caaactgggg tcgcgtgttg gctgcagtcg gcatggctga tgctgatatg 960
gaaccagaga agatttctgt gttcttcaat gatcaagcag tatgccttga ttccactggc 1020
gctcctggtg ctcgtgaggt ggatctttcc ggcgctgaca ttgatgtccg aattgatttg 1080
ggcaccagtg gggaaggcca ggcaacagtt cgaaccactg acctgagctt ctcctacgtg 1140
gagatcaact ccgcgtacag ctcttaa 1167
<210> 7
<211> 388
<212> PRT
<213> Unknown
<220>
<223> argJ C. glutamicum 13032
<400> 7
Met Ala Glu Lys Gly Ile Thr Ala Pro Lys Gly Phe Val Ala Ser Ala
1 5 10 15
Thr Thr Ala Gly Ile Lys Ala Ser Gly Asn Pro Asp Met Ala Leu Val
20 25 30
Val Asn Gln Gly Pro Glu Phe Ser Ala Ala Ala Val Phe Thr Arg Asn
35 40 45
Arg Val Phe Ala Ala Pro Val Lys Val Ser Arg Glu Asn Val Ala Asp
50 55 60
Gly Gln Ile Arg Ala Val Leu Tyr Asn Ala Gly Asn Ala Asn Ala Cys
65 70 75 80
Asn Gly Leu Gln Gly Glu Lys Asp Ala Arg Glu Ser Val Ser His Leu
85 90 95
Ala Gln Asn Leu Gly Leu Glu Asp Ser Asp Ile Gly Val Cys Ser Thr
100 105 110
Gly Leu Ile Gly Glu Leu Leu Pro Met Asp Lys Leu Asn Ala Gly Ile
115 120 125
Asp Gln Leu Thr Ala Glu Gly Ala Leu Gly Asp Asn Gly Ala Ala Ala
130 135 140
Ala Lys Ala Ile Met Thr Thr Asp Thr Val Asp Lys Glu Thr Val Val
145 150 155 160
Phe Ala Asp Gly Trp Thr Val Gly Gly Met Gly Lys Gly Val Gly Met
165 170 175
Met Ala Pro Ser Leu Ala Thr Met Leu Val Cys Leu Thr Thr Asp Ala
180 185 190
Ser Val Thr Gln Glu Met Ala Gln Ile Ala Leu Ala Asn Ala Thr Ala
195 200 205
Val Thr Phe Asp Thr Leu Asp Ile Asp Gly Ser Thr Ser Thr Asn Asp
210 215 220
Thr Val Phe Leu Leu Ala Ser Gly Ala Ser Gly Ile Thr Pro Thr Gln
225 230 235 240
Asp Glu Leu Asn Asp Ala Val Tyr Ala Ala Cys Ser Asp Ile Ala Ala
245 250 255
Lys Leu Gln Ala Asp Ala Glu Gly Val Thr Lys Arg Val Ala Val Thr
260 265 270
Val Val Gly Thr Thr Asn Asn Glu Gln Ala Ile Asn Ala Ala Arg Thr
275 280 285
Val Ala Arg Asp Asn Leu Phe Lys Cys Ala Met Phe Gly Ser Asp Pro
290 295 300
Asn Trp Gly Arg Val Leu Ala Ala Val Gly Met Ala Asp Ala Asp Met
305 310 315 320
Glu Pro Glu Lys Ile Ser Val Phe Phe Asn Gly Gln Ala Val Cys Leu
325 330 335
Asp Ser Thr Gly Ala Pro Gly Ala Arg Glu Val Asp Leu Ser Gly Ala
340 345 350
Asp Ile Asp Val Arg Ile Asp Leu Gly Thr Ser Gly Glu Gly Gln Ala
355 360 365
Thr Val Arg Thr Thr Asp Leu Ser Phe Ser Tyr Val Glu Ile Asn Ser
370 375 380
Ala Tyr Ser Ser
385
<210> 8
<211> 1167
<212> DNA
<213> Unknown
<220>
<223> argJ C. glutamicum 13032
<400> 8
atggcagaaa aaggcattac cgcgccgaaa ggcttcgttg cttctgcaac gaccgcgggt 60
attaaagctt ctggcaatcc tgacatggcg ttggtggtta accagggtcc agagttttcc 120
gcagcggccg tgtttacacg taaccgagtt ttcgcagcgc ctgtgaaggt gagccgagag 180
aacgttgctg atggccagat cagggctgtt ttgtacaacg ctggtaatgc taatgcgtgt 240
aatggtctgc agggtgagaa ggatgctcgt gagtctgttt ctcatctagc tcaaaatttg 300
ggcttggagg attccgatat tggtgtgtgt tccactggtc ttattggtga gttgcttccg 360
atggataagc tcaatgcagg tattgatcag ctgaccgctg agggcgcttt gggtgacaat 420
ggtgcagctg ctgccaaggc gatcatgacc actgacacgg tggataagga aaccgtcgtg 480
tttgctgatg gttggactgt cggcggaatg ggcaagggcg tgggcatgat ggcgccgtct 540
cttgccacca tgctggtctg cttgaccact gatgcatccg ttactcagga aatggctcag 600
atcgcgctgg ctaatgctac ggccgttacg tttgacaccc tggatattga tggatcaacc 660
tccaccaatg acaccgtgtt cctgctggca tctggcgcta gcggaatcac cccaactcag 720
gatgaactca acgatgcggt gtacgcagct tgttctgata tcgcagcgaa gcttcaggct 780
gatgcagagg gtgtgaccaa gcgcgttgct gtgacagtgg tgggaaccac caacaacgag 840
caggcgatta atgcggctcg cactgttgct cgtgacaatt tgttcaagtg cgcaatgttt 900
ggatctgatc caaactgggg tcgcgtgttg gctgcagtcg gcatggctga tgctgatatg 960
gaaccagaga agatttctgt gttcttcaat ggtcaagcag tatgccttga ttccactggc 1020
gctcctggtg ctcgtgaggt ggatctttcc ggcgctgaca ttgatgtccg aattgatttg 1080
ggcaccagtg gggaaggcca ggcaacagtt cgaaccactg acctgagctt ctcctacgtg 1140
gagatcaact ccgcgtacag ctcttaa 1167
<210> 9
<211> 289
<212> DNA
<213> Unknown
<220>
<223> lysCP1
<400> 9
ccgatgctag ggcgaaaagc acggcgagca gattgctttg cacttgattc agggtagttg 60
actaaagagt tgctcgcgaa gtagcacctg tcacttttgt ctcaaatatt aaatcgaata 120
tcaatatatg gtctgtttat tggaacgcgt cccagtggct gagacgcatc cgctaaagcc 180
ccaggaaccc tgtgcagaaa gaaaacactc ctctggctag gtagacacag tttattgtgg 240
tagagttgag cgggtaactg tcagcacgta gatcgaaagg tgcacaaag 289
<210> 10
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-A-F
<400> 10
atcctctaga gtcgaccatc gctgacacca tctgcc 36
<210> 11
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-A-R
<400> 11
gggcccacta gtctcgagtt caccgcggga gccaagcc 38
<210> 12
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-B-F
<400> 12
ctcgagacta gtgggccctg gattccaagg ctacgcc 37
<210> 13
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Tn-B-R
<400> 13
atgcctgcag gtcgaccctg aatggataag gcaccg 36
<210> 14
<211> 510
<212> DNA
<213> Unknown
<220>
<223> Tn-A
<400> 14
gcagacgcac tcgactacac ctccacctgc ccagaatgct cccaacctgg ggtgtttcgt 60
catcacaccc accggatgct cattgattta cccatcgtcg ggtttcccac caaactgttt 120
atccgtctac ctcgctaccg ctgcaccaac cccacatgta agcaaaagta tttccaagca 180
gaactaagct gcgctgacca cggtaaaaag gtcacccacc gggtcacccg ctggatttta 240
caacgccttg ctattgaccg gatgagtgtt cacgcaaccg cgaaagcact tgggctaggg 300
tgggatttaa cctgccaact agccctcgat atgtgccgtg agctggtcta taacgatcct 360
caccatcttg atggagtgta tgtcattggg gtggatgagc ataagtggtc acataatagg 420
gctaagcatg gtgatgggtt tgtcaccgtg attgtcgata tgaccgggca tcggtatgac 480
tcacggtgtc ctgcccggtt attagatgtc 510
<210> 15
<211> 479
<212> DNA
<213> Unknown
<220>
<223> Tn-B
<400> 15
aactcattcc ttctgctcgt cgcgtgatgg atccattcca tgttgtgcgg cttgctggtg 60
acaagctcac cgcctgccgg caacgcctcc agcgggagaa ataccagcgt cgtggtttaa 120
gccaggatcc gttgtataaa aaccggaaga ccttgttgac cacgcacaag tggttgagtc 180
ctcgtcagca agaaagcttg gagcagttgt gggcgtatga caaagactac ggggcgttaa 240
agcttgcgtg gcttgcgtat caggcgatta ttgattgtta tcagatgggt aataagcgtg 300
aagcgaagaa gaaaatgcgg accattattg atcagcttcg ggtgttgaag gggccgaata 360
aggaactcgc gcagttgggt cgtagtttgt ttaaacgact tggtgatgtg ttggcgtatt 420
tcgatgttgg tgtctccaac ggtccggtcg aagcgatcaa cggacggttg gagcatttg 479
<210> 16
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> lysCP1 promotor_F
<400> 16
gaatgagttc ctcgagccga tgctagggcg aaaa 34
<210> 17
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> lysCP1 promotor_R
<400> 17
ctttgtgcac ctttcgatct acgtgctgac agttac 36
<210> 18
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ _F
<400> 18
gttccacatg gccaaaaaag gcattac 27
<210> 19
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ _R
<400> 19
ttctttttaa gagctgtacg cggagttgat ctcc 34
<210> 20
<211> 1332
<212> DNA
<213> Unknown
<220>
<223> arg A E.coli
<400> 20
gtggtaaagg aacgtaaaac cgagttggtc gagggattcc gccattcggt tccctatatc 60
aatacccacc ggggaaaaac gtttgtcatc atgctcggcg gtgaagccat tgagcatgag 120
aatttctcca gtatcgttaa tgatatcggg ttgttgcaca gcctcggcat ccgtctggtg 180
gtggtctatg gcgcacgtcc gcagatcgac gcaaatctgg ctgcgcatca ccacgaaccg 240
ctgtatcaca agaatatacg tgtgaccgac gccaaaacac tggaactggt gaagcaggct 300
gcgggaacat tgcaactgga tattactgct cgcctgtcga tgagtctcaa taacacgccg 360
ctgcagggcg cgcatatcaa cgtcgtcagt ggcaatttta ttattgccca gccgctgggc 420
gtcgatgacg gcgtggatta ctgccatagc gggcgtatcc ggcggattga tgaagacgcg 480
atccatcgtc aactggacag cggtgcaata gtgctaatgg ggccggtcgc tgtttcagtc 540
actggcgaga gctttaacct gacctcggaa gagattgcca ctcaactggc catcaaactg 600
aaagctgaaa agatgattgg tttttgctct tcccagggcg tcactaatga cgacggtgat 660
attgtctccg aacttttccc taacgaagcg caagcgcggg tagaagccca ggaagagaaa 720
ggcgattaca actccggtac ggtgcgcttt ttgcgtggcg cagtgaaagc ctgccgcagc 780
ggcgtgcgtc gctgtcattt aatcagttat caggaagatg gcgcgctgtt gcaagagttg 840
ttctcacgcg acggtatcgg tacgcagatt gtgatggaaa gcgccgagca gattcgtcgc 900
gcaacaatca acgatattgg cggtattctg gagttgattc gcccactgga gcagcaaggt 960
attctggtac gccgttctcg cgagcagctg gagatggaaa tcgacaaatt caccattatt 1020
cagcgcgata acacgactat tgcctgcgcc gcgctctatc cgttcccgga agagaagatt 1080
ggggaaatgg cctgtgtggc agttcacccg gattaccgca gttcatcaag gggtgaagtt 1140
ctgctggaac gcattgccgc tcaggcgaag cagagcggct taagcaaatt gtttgtgctg 1200
accacgcgca gtattcactg gttccaggaa cgtggattta ccccagtgga tattgattta 1260
ctgcccgaga gcaaaaagca gttgtacaac taccagcgta aatccaaagt gttgatggcg 1320
gatttagggt aa 1332
<210> 21
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> argA ORF_F
<400> 21
gaaaggtgca caaagatggt aaaggaacgt aaaaccg 37
<210> 22
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> argA ORF_R
<400> 22
gcccactagt ctcgagcatg cggcgttgat tttg 34
<210> 23
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> argE ORF_F
<400> 23
gaaaggtgca caaagatgaa aaacaaatta ccgcc 35
<210> 24
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> argE ORF_R
<400> 24
gcccactagt ctcgaggttt gagtcactgt cggtcg 36
<210> 25
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> Ncgl2644 ORF_F
<400> 25
gaggagatca aaacacatat gacgcctagt cttccccg 38
<210> 26
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Ncgl2644 ORF_R
<400> 26
ttagaatttc cgttcggcgt acc 23
<210> 27
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ_N_F
<400> 27
cgggatccca cgcctgtctg gtcgc 25
<210> 28
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ_N_R
<400> 28
acgcgtcgac ggatctaaag cggcgcttc 29
<210> 29
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ_C_F
<400> 29
acgcgtcgac tccggcgctg acattgatgt cc 32
<210> 30
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> argJ_C_R
<400> 30
ctagtctaga gagctgcacc aggtagacg 29
<---
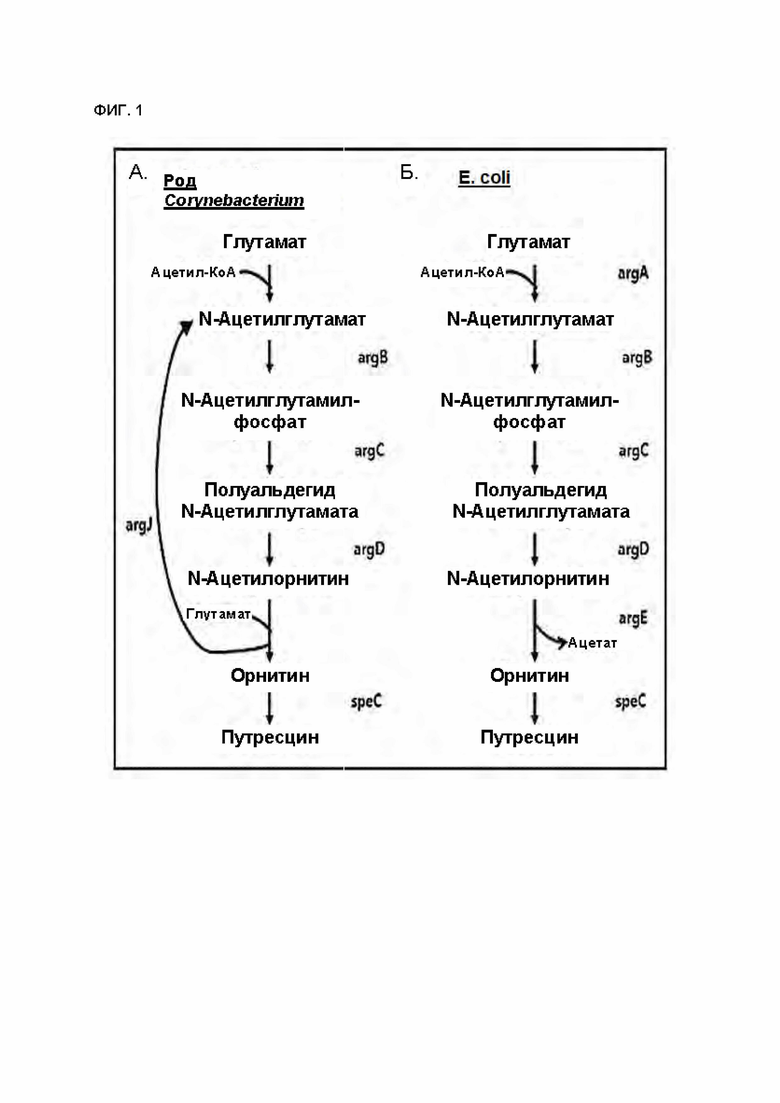
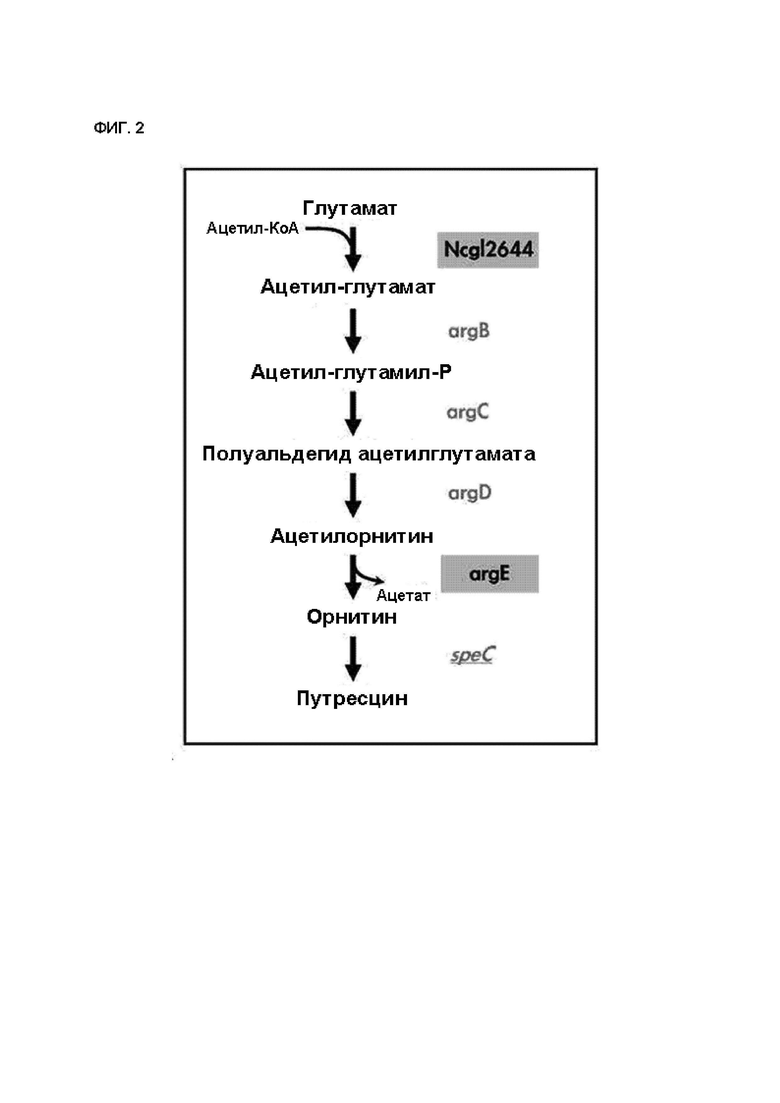
Изобретение относится к биотехнологии. Предложен микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из E. coli. Также предложены способ получения путресцина с использованием указанного микроорганизма, композиция для получения путресцина, содержащая указанный микроорганизм, и применение указанного микроорганизма для получения путресцина. Изобретение обеспечивает получение путресцина с высоким выходом. 4 н. и 9 з.п. ф-лы, 2 ил., 11 табл., 5 пр.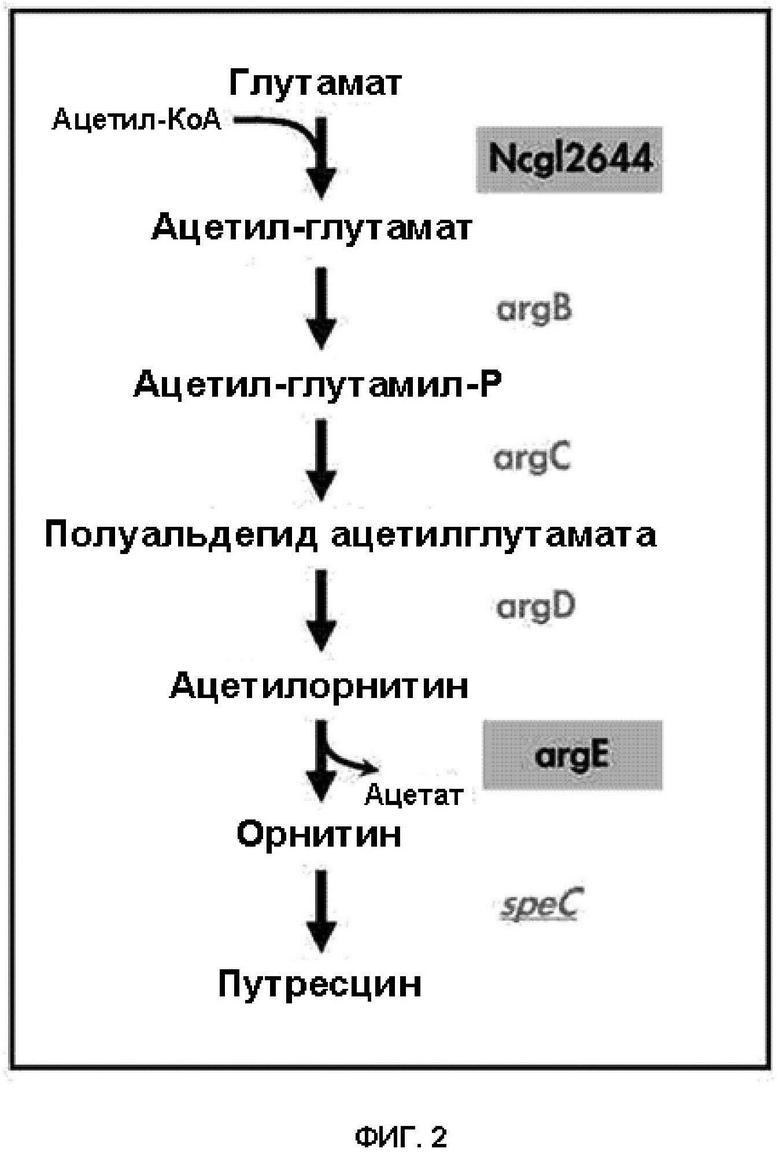
1. Микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из E. coli.
2. Микроорганизм рода Corynebacterium по п. 1, где N-ацетилтрансфераза, происходящая из штамма рода Corynebacterium, содержит аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней.
3. Микроорганизм рода Corynebacterium по п. 1, где ацетилорнитиндеацетилаза (argE), происходящая из E. coli, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней.
4. Микроорганизм рода Corynebacterium по п. 1, который дополнительно имеет ослабленную активность бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (argJ), происходящей из штамма рода Corynebacterium, где ослабленная активность получена посредством делеции гена argJ.
5. Микроорганизм рода Corynebacterium по п. 1, который представляет собой Corynebacterium glutamicum.
6. Способ получения путресцина, включающий культивирование в среде микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из E. coli.
7. Способ по п. 6, дополнительно включающий выделение путресцина из культуральной среды или микроорганизма.
8. Способ по п. 6, где N-ацетилтрансфераза, происходящая из штамма рода Corynebacterium, имеет аминокислотную последовательность SEQ ID NO: 1.
9. Способ по п. 6, где ацетилорнитиндеацетилаза, происходящая из E. coli, имеет аминокислотную последовательность SEQ ID NO: 3.
10. Композиция для получения путресцина, содержащая микроорганизм рода Corynebacterium, обладающий способностью продуцировать путресцин, в который введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из E. coli, и эксципиент.
11. Композиция по п. 10, где N-ацетилтрансфераза, происходящая из штамма рода Corynebacterium, содержит аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней.
12. Композиция по п. 10, где ацетилорнитиндеацетилаза, происходящая из E. coli, содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, имеющую по меньшей мере 90%-ную идентичность последовательности с ней.
13. Применение микроорганизма рода Corynebacterium, обладающего способностью продуцировать путресцин, для получения путресцина, где в данный микроорганизм введены активность N-ацетилтрансферазы, происходящей из штамма рода Corynebacterium, и активность ацетилорнитиндеацетилазы (argE), происходящей из E. coli.
| WO 2017014532 A1, 26.01.2017 | |||
| US 8481293 B2, 09.07.2013 | |||
| NGUYEN A.Q | |||
| et al | |||
| Elimination of polyamine N-acetylation and regulatory engineering improved putrescine production by Corynebacterium glutamicum | |||
| J Biotechnol | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ ПОВЫШЕННОЙ СПОСОБНОСТЬЮ ПРОДУЦИРОВАТЬ ПУТРЕСЦИН, И СПОСОБ ПОЛУЧЕНИЯ ПУТРЕСЦИНА С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2604806C2 |

